
Intestinal Motility in response to EPI, MCh, ADP, and...
23
1 Intestinal Motility in response to EPI, MCh, ADP, and Ca free Ringer’s solution By: Hee Joo Kim
Transcript of Intestinal Motility in response to EPI, MCh, ADP, and...

1
Intestinal Motility in response to EPI, MCh, ADP, and Ca free Ringer’s solution
By: Hee Joo Kim

2
Introduction
The digestive system in humans involves the mouth, pharynx, esophagus, stomach, small
intestine, large intestine, and gastric juices. Each part helps with the digestive processes;
motility, digestion, secretion, and absorption. (Sherwood, 2010, pg 589). Through this, the food
humans eat can be processed and used as energy.
The digestive tract wall is further divided up into layers. The four major ones, from the
innermost layer to the outer most layer, are mucosa, submucosa, muscularis externa, and serosa.
The mucosa is used to protect the lining with mucous membrane. The submucosa gives
elasticity so the digestive tract can stretch when food comes in. The muscularis externa mixes
and propels food throughout by longitudinal and circular movements. And the serosa layer
prevents friction between the digestive organs with the rest of the body. (Sherwood, 2010, pg
591-592).
The digestive system reacts to parasympathetic and sympathetic stimulations. During a
sympathetic stimulation, it will decrease digestion, and during a parasympathetic stimulation, it
will increase digestion. (Sherwood, 2010, pg 242). Different factors can effect digestion due to
these stimulations.
In this lab we used a segment of a rabbit’s intestine to see the effect of digestive motility
in response to epinephrine (EPI), methacholine (MCh), adenosine-5-diphosphate (ADP), and Ca-
free ringer-tyrode’s solution. This was done by measuring the difference in tension, amplitude,
and frequency each time a stimulant was added. We expected to see a decrease in tension and
amplitude, and no change in frequency for EPI. For MCh, we expected to see an increase in
tension, a decrease in amplitude, and no change in frequency. For ADP, we expected to see a

3
decrease in tension and amplitude, and no change in frequency. For Ca-free ringer’s, we
expected to see a decrease in all three tension, amplitude, and frequency.
Materials and Methods
A 2 cm segment of rabbit’s intestine was looped with some string and attached to a
transducer. It was also immersed in a circulating water bath cuvette filled with normal ringer’s
solution to maintain a temperature of 38 degrees Celsius. Air bubbles were also supplied at 1-3
bubbles per second inside the cuvette with the intestine. Activity of this intestine was measured
for 10 minutes. The gut was rinsed with normal ringer’s solution and put into a clean cuvette
with ringer’s solution after each new addition of stimulants. 3 drops of EPI were dropped into
the cuvette and activity was measured for another 10 minutes. After rinsing, equilibrating, and
recording its baseline for 5 minutes, 1 drop of MCh was added and recorded for 2 minutes, then
3 more drops were added and recorded for 10 minutes. After rinsing, equilibrating, and
recording its baseline, 10 drops of ADP were added and recorded for 10 minutes. An adjacent
cuvette was rinsed and filled with Ca-free ringer’s solution, and fter rinsing, equilibrating, and
recording its baseline activity, the gut was moved to the Ca-free ringer’s solution and its activity
was recorded for 5 minutes. It was then moved back into normal ringer solution and its activity
was recorded for another 5 minutes. For a more detailed Method and Materials, refer to
experiment 8 of NPB 101L Physiology Lab Manual (Bautista, 2009, pgs 76-81).
4
Results
In each part of the lab, 10 cycles were chosen from what seemed to be the best representation of
the intestinal activity before and after a stimulus. These 10 cycles were then averaged out to
show on the graphs. Although represented by the same Y-graph, tension and amplitude is
measured in grams (g) while frequency is measured in hertz (Hz).
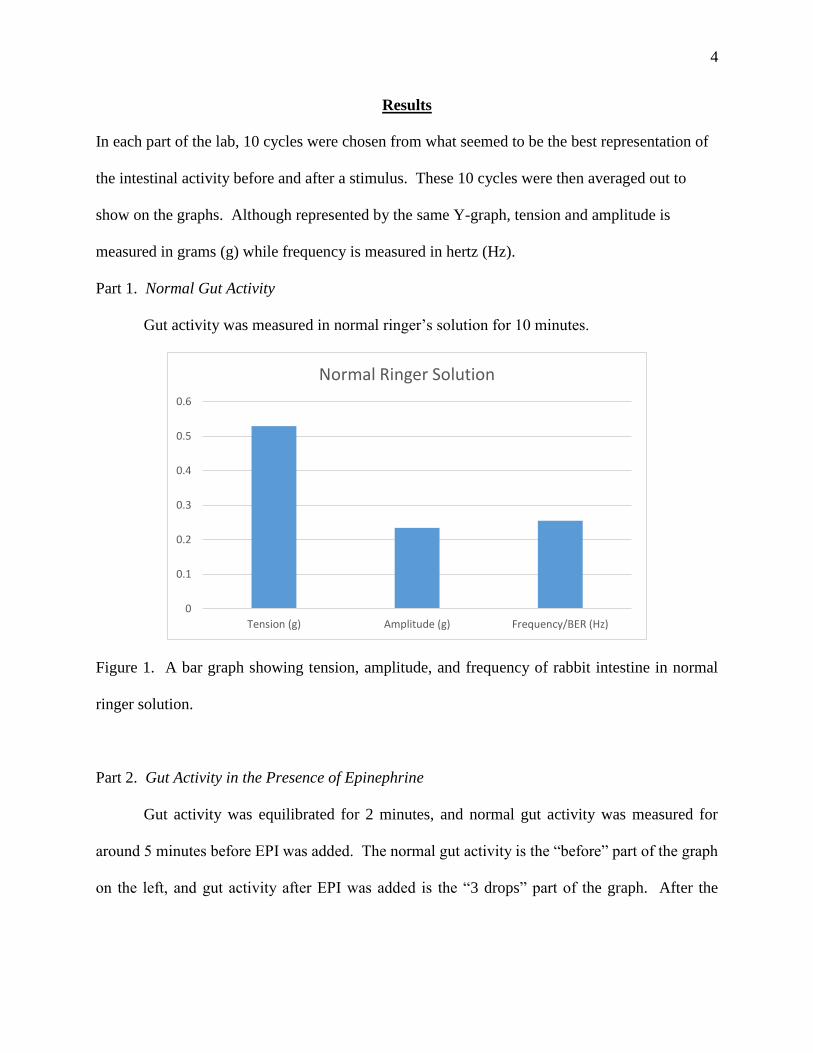
Part 1. Normal Gut Activity
Gut activity was measured in normal ringer’s solution for 10 minutes.
Figure 1. A bar graph showing tension, amplitude, and frequency of rabbit intestine in normal
ringer solution.
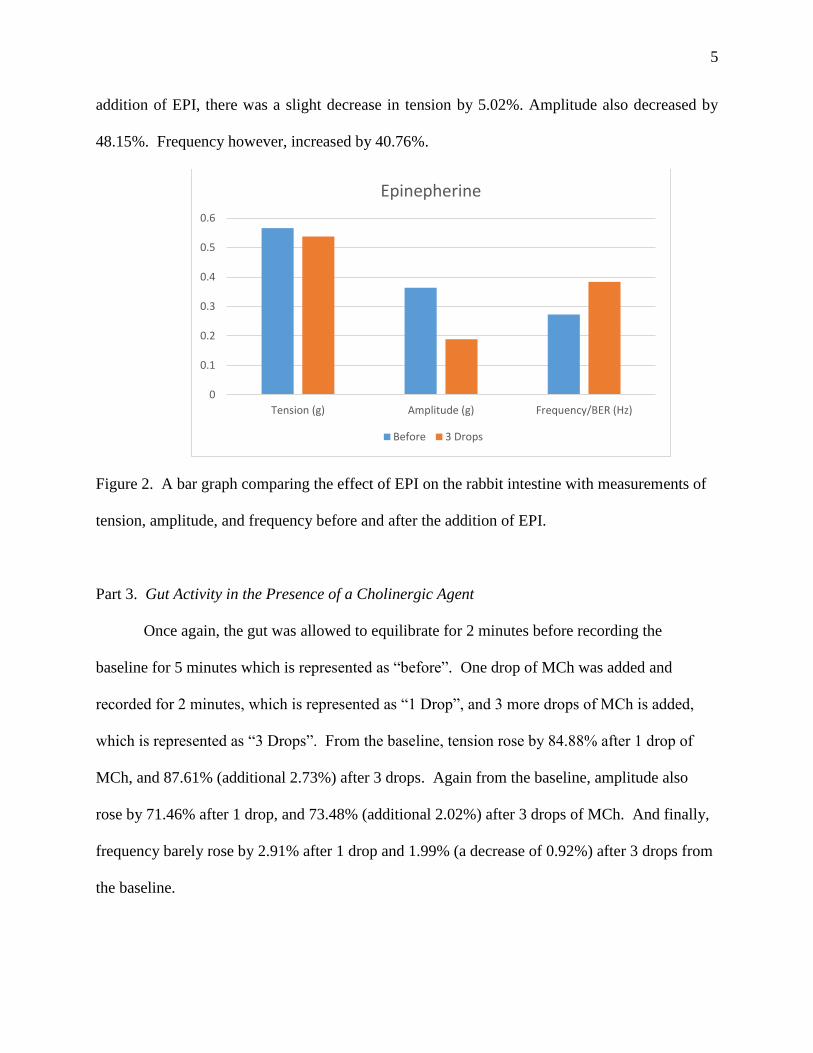
Part 2. Gut Activity in the Presence of Epinephrine
Gut activity was equilibrated for 2 minutes, and normal gut activity was measured for
around 5 minutes before EPI was added. The normal gut activity is the “before” part of the graph
on the left, and gut activity after EPI was added is the “3 drops” part of the graph. After the
0
0.1
0.2
0.3
0.4
0.5
0.6
Tension (g) Amplitude (g) Frequency/BER (Hz)
Normal Ringer Solution
5
addition of EPI, there was a slight decrease in tension by 5.02%. Amplitude also decreased by
48.15%. Frequency however, increased by 40.76%.
Figure 2. A bar graph comparing the effect of EPI on the rabbit intestine with measurements of
tension, amplitude, and frequency before and after the addition of EPI.
Part 3. Gut Activity in the Presence of a Cholinergic Agent
Once again, the gut was allowed to equilibrate for 2 minutes before recording the
baseline for 5 minutes which is represented as “before”. One drop of MCh was added and
recorded for 2 minutes, which is represented as “1 Drop”, and 3 more drops of MCh is added,
which is represented as “3 Drops”. From the baseline, tension rose by 84.88% after 1 drop of
MCh, and 87.61% (additional 2.73%) after 3 drops. Again from the baseline, amplitude also
rose by 71.46% after 1 drop, and 73.48% (additional 2.02%) after 3 drops of MCh. And finally,
frequency barely rose by 2.91% after 1 drop and 1.99% (a decrease of 0.92%) after 3 drops from
the baseline.
0
0.1
0.2
0.3
0.4
0.5
0.6
Tension (g) Amplitude (g) Frequency/BER (Hz)
Epinepherine
Before 3 Drops

6
Figure 3. A bar graph comparing the effect of MCh on the rabbit intestine with measurements of
tension, amplitude, and frequency before and after the addition of 1 drop of MCh and 3 drops of
MCh.
Part 4. Gut Activity in the Presence of Purinergic Agent
Gut was allowed to equilibrate for 2 minutes before recording the baseline for 5 minutes
which is represented as “before”. 10 drops of ADP was added to give results of after, presented
as “10 Drops”. There was an overall decrease after adding ADP in all three categories measured.
Tension decreased by 19.88%, amplitude by 20.66%, and frequency slightly by 8.62%.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Tension (g) Amplitude (g) Frequency (Hz)
MCh
Before 1 Drop 3 Drops

7
Figure 4. A bar graph comparing the effect of ADP on the rabbit intestine with measurements of
tension, amplitude, and frequency before and after the addition of 10 drops of ADP.
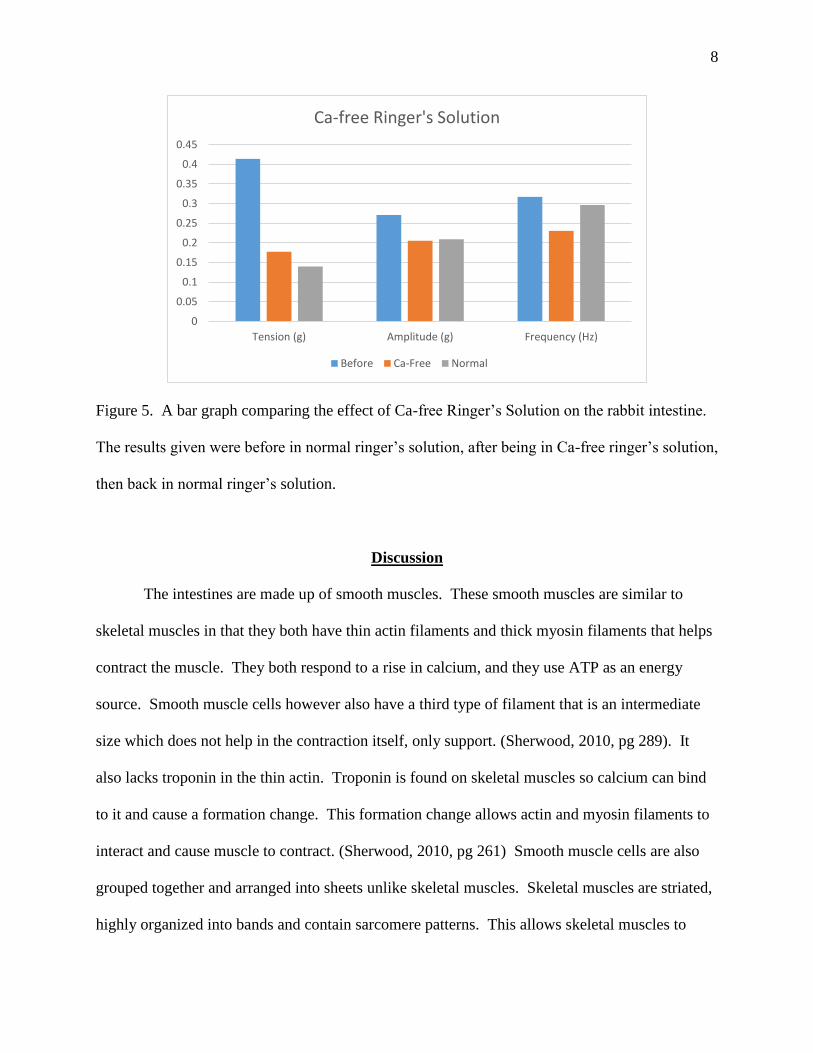
Part 5. Gut Activity in Ca-free Ringer-Tyrode’s Solution
After equilibration the gut activity was measured it was recorded for 5 minutes,
represented on the graph as “before”. The gut was then dunked in a Ca-free ringer’s solution for
5 minutes, represented on the graph as “Ca-Free”, then dunked back into the normal solution for
another 5 minutes which is represented on the graph as “Normal”. All three decreased from the
baseline with a decreasing trend in tension, while signs of recuperation for amplitude and
frequency. From the baseline, tension decreased by 57.16% when dunked in Ca-Free, and
66.26% (a decrease by 9.1%) when dunked back in normal solution. From the baseline,
amplitude also decreased by 24.32% when in Ca-Free, and 22.78% (an increase of 1.54%) when
back in normal solution. And finally, for frequency, a decrease of 27.25% was seen in Ca-Free
solution, and a decrease of 6.52% (an increase by 20.73%) was seen when back in normal
solution.
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
Tension (g) Amplitude (g) Frequency/BER (Hz)
ADP
Before After 10 Drops
8
Figure 5. A bar graph comparing the effect of Ca-free Ringer’s Solution on the rabbit intestine.
The results given were before in normal ringer’s solution, after being in Ca-free ringer’s solution,
then back in normal ringer’s solution.
Discussion
The intestines are made up of smooth muscles. These smooth muscles are similar to
skeletal muscles in that they both have thin actin filaments and thick myosin filaments that helps
contract the muscle. They both respond to a rise in calcium, and they use ATP as an energy
source. Smooth muscle cells however also have a third type of filament that is an intermediate
size which does not help in the contraction itself, only support. (Sherwood, 2010, pg 289). It
also lacks troponin in the thin actin. Troponin is found on skeletal muscles so calcium can bind
to it and cause a formation change. This formation change allows actin and myosin filaments to
interact and cause muscle to contract. (Sherwood, 2010, pg 261) Smooth muscle cells are also
grouped together and arranged into sheets unlike skeletal muscles. Skeletal muscles are striated,
highly organized into bands and contain sarcomere patterns. This allows skeletal muscles to
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
Tension (g) Amplitude (g) Frequency (Hz)
Ca-free Ringer's Solution
Before Ca-Free Normal

9
extend the full length of the muscle needed, unlike the smooth muscle sheets. Smooth muscle
cell is spindle shaped with a single nucleus and much smaller when compared to skeletal
muscles. The size of smooth muscles is 2-10 micrometers in diameter and 50-400 micrometers
long. (Sherwood, 2010, pg 289). The size of the cylindrical shaped skeletal muscle is 10-100
micrometers in diameter and 750,000 micrometers long. (Sherwood, 2010, pg 258). The smooth
muscle cells are involuntary, and involves the autonomic nervous system. (Sherwood, 2010, pg
290).
The way smooth muscles contract is also different from skeletal muscle. Instead of a
physical change by calcium binding to troponin in skeletal muscle, a chemical change occurs for
binding in smooth muscle. Calcium binds to calmodulin in the smooth muscle to form a Ca-
calmodulin complex. This then binds to and activates a myosin light chain kinase (MLCK). The
MLCK uses a phosphate on ATP to phosphorylate the myosin light chain (MLC). The myosin
light chain can now bind with the actin so cross-bridge cycle can begin and muscle contraction
can happen. (Sherwood, 2010, pgs 292-293).
As mentioned before, there are 4 primary layers of the digestive tract wall. The inner
most layer called the mucosa, lines the surface of the digestive tract. This layer is further divided
up into 3 layers; the mucous membrane (inner layer), lamina propria (middle layer), and
muscularis mucosa (outer layer). The mucous membrane protects the surface from digestive
juices with mucous. The lamina propria contains the gut-associated lymphoid tissue (GALT)
which is an important factor to fight against bacteria in the intestines that can cause diseases.
The mucosa is highly folded to increase surface area, especially in the small intestine. This is
done to promote absorption.

10
The second layer, submucosa, is a thick layer that provides flexibility for the digestive
tract. This can be important since there are a lot of gut activity in the third layer, muscularis
externa, which helps mix and propel food forward. It is further divided up into two layers called
the inner circular layer and outer longitudinal layer. The inner circular layer can decrease in
diameter, even constricting a point in the tube, when contracted. The outer longitudinal layer can
shorten the tube when contracted. The movement of decreasing the diameter and shortening the
tube can mix and propel food.
The fourth layer, serosa, is the most outer layer of the digestive tract. This layer secretes
a serous fluid which is slippery to the digestive tract so when the digestive tract moves, there is
less friction between the digestive organs and the body. It is also attached to mesentery, which
attaches the digestive system to the abdominal cavity so it can swing around while mixing, but
still stabilizes it in place. (Sherwood, 2010, pgs591-593).
The movements to digest and absorb food is regulated by four factors; autonomous
smooth muscle function, intrinsic nerve plexuses, extrinsic nerves, and gastrointestinal
hormones. The autonomous smooth muscle function is self-induced by slow-wave potentials, or
the basic electrical rhythm (BER). The pacemakers are the interstitial cells of Cajal, the non-
contractile cells that lie between the longitudinal and circular smooth muscle cell layers. They
pacemakers are rhythmic, cyclically bringing the membranes either closer or farther away from
threshold potential, unlike action potentials. The slow waves will continue to oscillate until a
mechanical, neural, or hormonal facts influence it to reach threshold and depolarize. (Sherwood,
2010, pgs 593-594).
The intrinsic nerve plexus, or enteric nervous system, are two nerve fibers; the
submucosal plexus and the myeneric plexus. These two plexus are found throughout the

11
digestive tract wall can give the digestive system self-regulation. The intrinsic nerve plexus have
various neurons in its system that will coordinate with local activity and respond to it by
secreting digestive juices, hormones, excitatory neurons, inhibitory neurons, or whatever it needs
to. (Sherwood, 2010, pgs 594-595)
The extrinsic nerves helps coordinate the different parts of the digestive tract’s activities.
These nerve fibers are the ones that starts on the outside of the digestive system and supply the
digestive system with nerves. It uses the autonomic nerves, the parasympathetic and the
sympathetic systems to control the digestibility. The sympathetic system, or “fight or flight”
system inhibits the digestion. The parasympathetic system, or the “rest and digest” system
encourage digestion. And gastrointestinal hormones from the endocrine gland in the mucosa
layer are released into the blood to the digestive tract. These hormones either promote or inhibit
digestion by exerting excitatory or inhibitory influences on the smooth muscle. (Sherwood,
2010, pg 595).
In response to epinephrine on the rabbit’s intestine, tension and amplitude both
decreased, while frequency increased. The decrease in tension corresponded to a different
experiment done on a toad’s digestive tract. Upon administering adrenaline (epinenpherine), the
piece of digestive tract relaxed, which would lead to a decrease in tension. (Gruber, 1922, pg
321). In another study, epinephrine was given to a dog’s intestine, before the administration of
cocaine. The results were congruent to our results in that both tension and amplitude decreased.
(Gruber, 1936, pg 349). This decrease in amplitude and tension correlates with sympathetic
nervous system because the sympathetic nervous system inhibits digestion. Epinephrine can
bind to beta2 receptors in the smooth muscles of the digestive tract by traveling through blood
and bring out sympathetic activity. (Sherwood, 2010, pg 706). The increase in frequency is not

12
expected however, since rate is controlled by the pacemaker (interstitial cells of Cajal). The
pacemaker generate the BER which can reach threshold for contraction. (Kim, 2002, pg 797).
When threshold happens, the voltage gated calcium channels are activated and calcium can go
into the smooth muscle cells. This allows for the rising phase of the action potential (calcium
influx) and the falling phase (potassium efflux) which can allow the muscle to contract. How
frequently this muscles contract is dependent on the concentration of calcium. (Sherwood, 2010,
pg 594). Since epinephrine was the only stimulant added, there should have been no change in
the frequency of the result, however, there was a significant increase. This increase in frequency
could be due to an error involving the movement of the table, or the gas bubbles not delivering 1-
3 bubbles per second.
With the addition of MCh, which was used to mimic the effect of ACh on the intestinal
segment, we saw opposite results. Tension and amplitude increased significantly while
frequency increased very slightly (around 2% compared to an increase by around 40% with the
addition of epinephrine). In a study done with various animals, similar results occurred. The
study was done on the intestines of a mouse, cat, rabbit, and dog. With the exception of the cat’s
upper jejunum, the intestine showed an increase in contraction (tension). (Bernheim, 1934, pg
65).
ACh will bind to the cholinoceptors, which can be nicotinic or muscarinic. The
muscarinic receptor, M3- choline receptor which occurs mainly in the smooth muscles, will then
activate the PLC in the postganglionic neuron via the G-protein. The inositol tri-phosphate (IP3)
and diacylglyceral (DAG) can be released, and an influx of calcium will occur. The increase in
calcium will allow for more calcium-calmodulin binding to occur, promoting more cross
bridging and eventually increasing tension. (Silbernagl, 2009, pg 82). The IP3 would normally

13
open calcium channels of the sarcoplasmic reticulum, which will lead to an increase in calcium,
and a decrease in amplitude. However, both data from the rabbit intestine and Bernheim’s data
showed an increase in amplitude. Bernheim’s data could give these results because epinephrine
was also administered to the intestine. MCh was administered after adding EPI for the rabbit’s
intestine as well. The rabbit’s intestine may not have been thoroughly rinsed before adding MCh
into the cuvette. The epinephrine residual could cause the increase in amplitude despite
administering ACh.
Next, ADP was added to the rabbit’s intestine to mimic the effect of ATP. The results
showed that tension, amplitude, and frequency all went down after the administration of ADP.
Frequency, as mentioned before should not be affected by the addition of ADP, however since
the value was very low (around 8%), it could be due to small errors in the machine. Tension and
amplitude corresponded to the addition of ADP. ATP stimulates the Gq protein which allows
PIP2 to turn into IP3 and DAG. IP3 allows the efflux of calcium stores from the intracellular
stores to the cytosol by opening calcium channels. DAG can activate protein kinase C (PKC),
and with the calcium freed by IP3 can be transferred to the intracellular membrane from the
cytosol. And this phosphorylates MLC at sites that MLCK does not act upon. (Silbernagl, 2009,
pg 278-279). MLCK, as previously mentioned, helps phosphorylate MLC, which leads to cross-
bridging and muscles contraction. (Sherwood, 2010, pg 292). With MLC phosphorylated not
through MLCK, there is a decrease in contraction and amplitude.
And finally, when the rabbit intestine was exposed to a calcium free ringer’s solution,
tension, amplitude, and frequency all decreased significantly. As mentioned in previous texts,
calcium plays a huge role in smooth muscle contraction. When it is not supplied externally in
normal ringer’s solution, the only calcium source is what is left in the intestine. Without

14
calcium, there will be no calcium-calmodulin complex which will help with the contraction of
muscle. Without muscle contraction, tension and amplitude will decrease. The influx of calcium
that opens calcium channels for depolarization and repolarization due to efflux of potassium will
not happen. The interstitial cells of Cajal (pacemakers) will slow down and frequency will
decrease as well.
In this lab, we used EPI, MCh, ADP, and Ca-free Ringer’s solution to mimic the body’s
internal control system on the intestine. We saw that EPI, part of the sympathetic system,
reduced the motility of the intestine. Another part of the autonomic nervous system, the Ach
(MCh in this lab) which is a part of the parasympathetic system, increased intestinal motility.
ADP also reduced the motility of the intestine and without Calcium, an important part of smooth
muscle contraction, the intestine motility was also reduced.

15
Citations/References
Bautista E, Korber J. NPB101L: Physiology Lab Manual. Second Edition. Ohio: Cengage
Learning, 2009.
Bernheim Frederick. Interaction of acetylcholine and epinephrine on the isolated small intestines
of various animals. Journal of Pharmacology and Experimental Therapeutics.1934;
51(1):59-67.
Gruber Charles M. The action of epinephrine, tyramine and ephedrine on the small intestine of
the unanesthetized dog, before and following the administration of cocaine. Journal of
Pharmacology and Experimental Therapeutics. 1936; 57:347-360
Gruber Charles M. The Effect of Epinephrine on excised strips of frog’s digestive tracts.
Journal of Pharmacology and Experimental Therapeutics. 1922; 20(5):321-357.
Kim Young Chul, Koh Sand Don, and Sanders Kenton M. Voltage-dependent inward currents
of interstitial cells of Cajal from murine colon and small intestine. The Journal of
Physiology. 2002; 541:797-810.
Silbernagl Stefan and Despopoulos Agamemnon. Color Atlas of Physiology. 6th ed. New
York: Thieme, 2009. P. 82
Sherwood, Lauralee. Human Physiology: From Cells to Systems. 7th ed. California: Brooks/Cole,
Cengage Learning, 2010. P. 242, 258, 261, 289-290, 292-293, 589, 591-595, 706.

16
Appendices
Raw Data:
Column1 Tension (g) Amplitude (g) Frequency (Hz)
Normal 0.5291621 0.2342853 0.2552444
Before EPI 0.565936 0.3634597 0.2726
EPI 0.5375181 0.1884575 0.3837063
Before MCh 0.383611 0.414607 0.288228
1 Drop MCh 0.709224 0.710904 0.296605
3 Drops MCh 0.719694 0.71928 0.293962
Before ADP 0.252738 0.176825 0.299961
After ADP 0.202482 0.140296 0.274096
Before Ca-Free 0.413974 0.27117 0.316921
Ca-Free 0.177355 0.20521 0.230555
Normal Ringer's 0.139663 0.209397 0.296232
ECG of Normal:
208.96 250.75 292.54 334.33seconds
0.00
0.29
0.58
0.87
gra
ms
Forc
e

17
ECG Before EPI:
ECG After EPI:
141.98 165.64 189.30 212.97seconds
0.34
0.51
0.69
0.86
gra
ms
Forc
e
378.61 402.27 425.93 449.60seconds
0.34
0.51
0.69
0.86
gra
ms
Forc
e

18
ECG Before MCh:
ECG After 1 Drop MCh and 3 Drops MCh:
start baseline, 11:01:18 AM
232.30 261.34 290.38 319.42seconds
0.36
0.54
0.72
0.90
gra
ms
Forc
e
Mch 1 drop, 11:06:39 AM mch 1, 11:08:48 AM
mch 2ch3, 11:08:52 AM
637.89 708.77 779.65 850.52seconds
0.37
0.74
1.10
1.47
gra
ms
Forc
e

19
ECG Before ADP:
ECG After ADP:
173.62 202.56 231.49 260.43seconds
0.21
0.31
0.42
0.52
gra
ms
Forc
e
add adp, 11:26:22 AM finish adp, 11:26:33 AM
306.42 334.28 362.14 389.99
seconds
0.21
0.31
0.41
0.52gra
ms
Forc
e
20
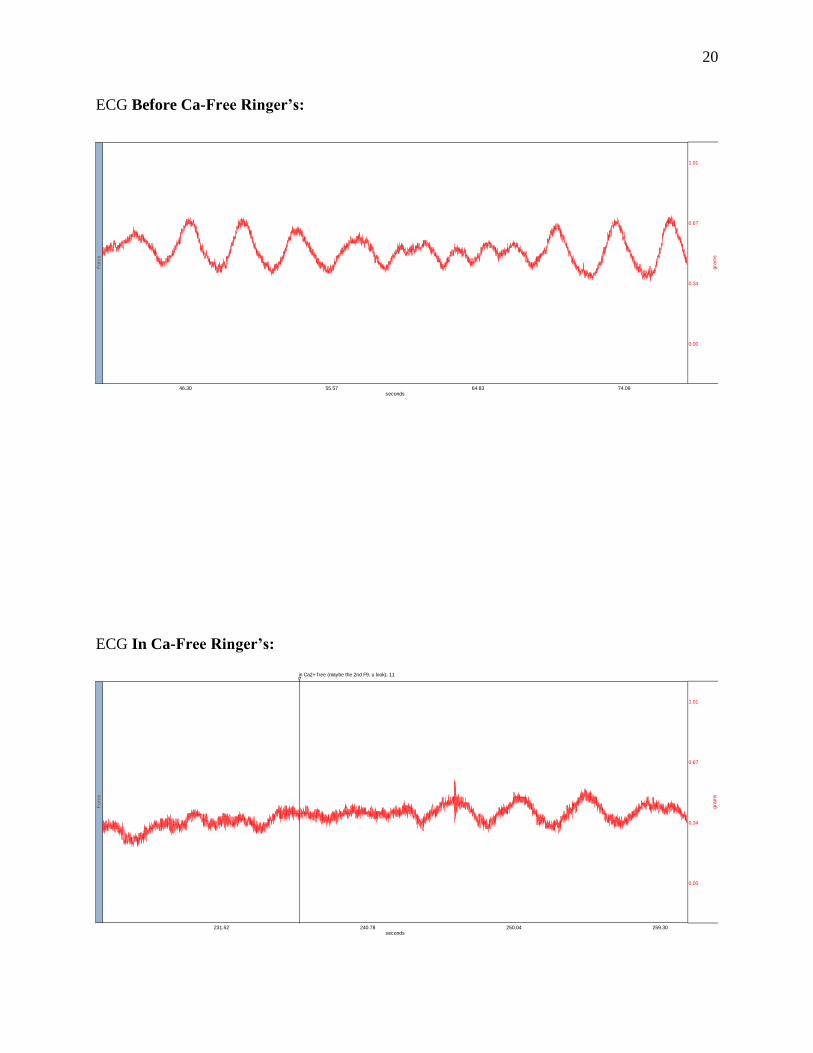
ECG Before Ca-Free Ringer’s:
ECG In Ca-Free Ringer’s:
46.30 55.57 64.83 74.09seconds
0.00
0.34
0.67
1.01
gra
ms
Forc
e
in Ca2+ free (maybe the 2nd F9, u look), 11
231.52 240.78 250.04 259.30seconds
0.00
0.34
0.67
1.01
gra
ms
Forc
e
21
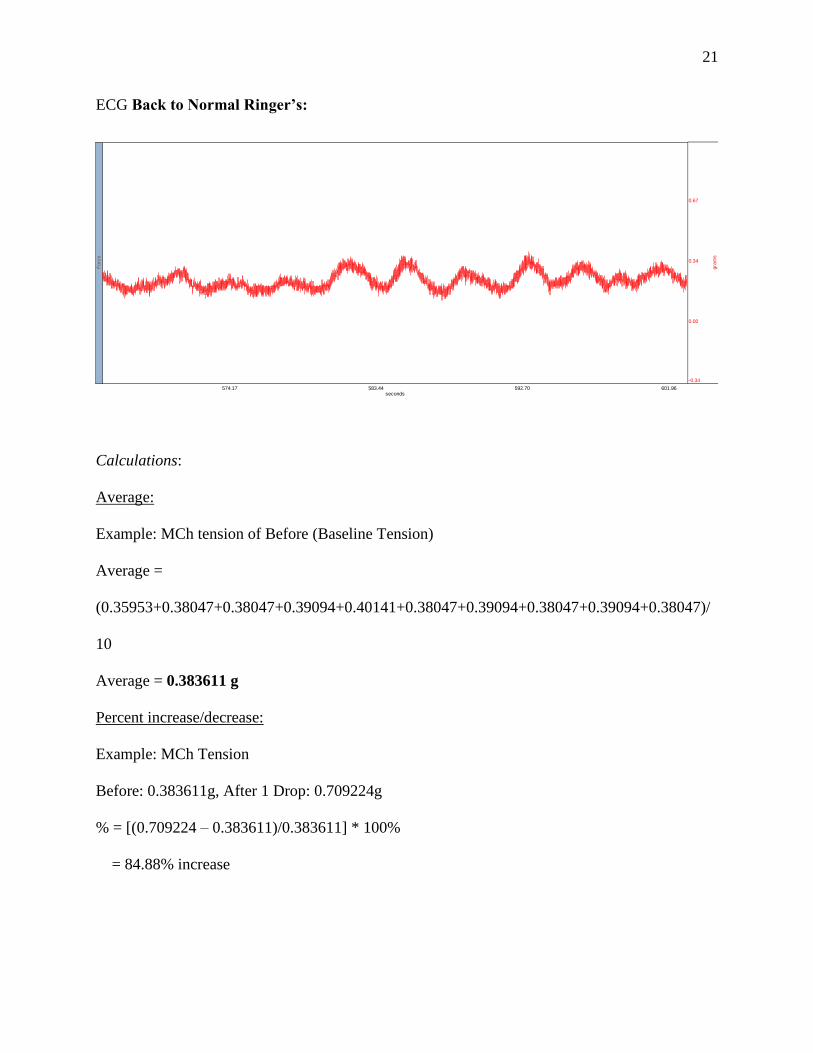
ECG Back to Normal Ringer’s:
Calculations:
Average:
Example: MCh tension of Before (Baseline Tension)
Average =
(0.35953+0.38047+0.38047+0.39094+0.40141+0.38047+0.39094+0.38047+0.39094+0.38047)/
10
Average = 0.383611 g
Percent increase/decrease:
Example: MCh Tension
Before: 0.383611g, After 1 Drop: 0.709224g
% = [(0.709224 – 0.383611)/0.383611] * 100%
= 84.88% increase
574.17 583.44 592.70 601.96seconds
-0.34
0.00
0.34
0.67
gra
ms
Forc
e

22
References:
“FREDERICK BERNHEIM
all the cases studied except for the upper jejunum of the cat, acetyicholine will produce a
sustained contraction indicating that the intestine contains no esterase. Having obtained the
sustained contraction, the effect of epinephrine on it could be studied. The differences then seen
in the intestines of the various animals ought not to be interpreted as simply a difference in the
way epinephrine acts but as a difference in the interaction of acetyicholine and epinephrmne and
therefore in all probability the parasympathetic and sympathetic systems. On the one extreme
there are the dog and the guinea pig on intestines of which epinephrine has very little effect after
acetylcholine although it has a marked effect after other drugs…..”
“ACTION OF EPINEPHRINE ON INTESTINE
In the twelve animals used in these experiments, epinephrine was injected separately, 24 times,
tyramine 17 times and ephe- drine 10 times, before the administration of cocaine, and in every
instance there was a marked decrease in the general tonus and in the amplitude of the rhythmical
contractions. The duration of the decreased tonus was, as a rule, longer for either tyramine or
ephedrine, in the doses used, than it was for epinephrine (see fig. 3). In some cases with
ephedrine and tyramine the decreased tonus and amplitude of the contractions lasted for over
forty…”
“THE EFFECT OF EPINEPHRINE ON EXCISED STRIPS OF FROGS’ DIGESTIVE
TRACTS
CHARLES M. GRUBER
In studying the effect of epinephrine on the smooth muscle of the esophagus and stomach in cold
blooded animals a variety of methods has been used by numerous observers with varying results.

23
Bottazzi and Torretta (1) and Bottazzi (2) used solutions of adrenalin’ ranging in strength from
1:1000 to 1:150,000. They found that solutions of 1:20,000 to 1: 50,000 produced relaxation of
the longitudinal coat of the toad’s esophagus (Bufo vulgaris) but that a solution of 1:120,000 had
no effect. Bottazzi also noted that if the adrenalin solution was not too strong nor applied too
often this relaxation was in some cases followed by increased tonus….”
“KIM YC, KOH SD, SANDERS KM
“Electrical slow waves in gastrointestinal (GI) muscles are generated by pacemaker cells, known
as interstitial cells of Cajal (ICC). The pacemaker conductance is regulated by periodic release of
Ca2+ from inositol 1,4,5-trisphosphate (IP(3)) receptor-operated stores, but little is known about
how slow waves are actively propagated….”














![Small Intestine Prof. K. Sivapalan.. 2013Small Intestine2 Small Intestinal Motility Segmental movement [mixing 7/min in ileum] Peristalsis [propagation.](https://static.fdocuments.in/doc/165x107/56649c915503460f9494c7b5/small-intestine-prof-k-sivapalan-2013small-intestine2-small-intestinal.jpg)



