Interplays between sumoylation, SUMO-targeted Ubiquitin ...20Bonne%20K%F8hler.pdf(DOCX)...
194
FACULTY OF SCIENCE UNIVERSITY OF COPENHAGEN PhD thesis Julie Bonne Køhler Interplays between sumoylation, SUMO-targeted Ubiquitin Ligases, and the ubiquitin-adaptor protein Ufd1 in fission yeast Academic advisor: Geneviève Thon Submitted: 16/07/14
Transcript of Interplays between sumoylation, SUMO-targeted Ubiquitin ...20Bonne%20K%F8hler.pdf(DOCX)...

F A C U L T Y O F S C I E N C E U N I V E R S I T Y O F C O P E N H A G E N
PhD thesis Julie Bonne Køhler
Interplays between sumoylation, SUMO-targeted Ubiquitin Ligases, and the ubiquitin-adaptor protein Ufd1 in fission yeast
Academic advisor: Geneviève Thon
Submitted: 16/07/14

1
Contents Acknowledgements............................................................................................................................... 3
Summary ................................................................................................................................................. 4
Introduction ........................................................................................................................................... 5
The ubiquitin-like protein family..................................................................................................... 5
The SUMO modification system ............................................................................................. 6
-The SUMO family ................................................................................................................................ 6
-The mammalian SUMO proteins ......................................................................................................... 7
-SUMO conjugation .............................................................................................................................. 8
-The SUMO E3s .................................................................................................................................... 9
-SUMO E3s in fission yeast ..................................................................................................................11
-Substrate selection ............................................................................................................................11
-SUMO chains .....................................................................................................................................12
-SUMO deconjugation .........................................................................................................................13
-Function of SUMO chains ...................................................................................................................16
-SUMO substrates ...............................................................................................................................17
-Protein group modification ................................................................................................................17
-Identification of SUMO substrates .....................................................................................................18
The ubiquitin modification system .................................................................................................21
-Covalent modification by ubiquitin ....................................................................................................22
-The ubiquitin-proteasome system (UPS) ............................................................................................23
-The Cdc48/p97 ubiquitin-selective ATPase .........................................................................................24
-The Cdc48-Ufd1-Npl4 complex ...........................................................................................................25
-Cdc48/p97 as a “molecular gearbox” .................................................................................................27
-Cdc48/p97 binding motifs ..................................................................................................................27
-Proteasomal delivery .........................................................................................................................28
Crosstalks between the SUMO and ubiquitin systems ...............................................................30
The SUMO-targeted ubiquitin ligases (STUbLs)........................................................................31
-RNF4 STUbL homologs .......................................................................................................................31
-STUbL targeting/substrate recognition ...............................................................................................32
-STUbL functions .................................................................................................................................34

2
-Genetic relationship between STUbLs and Ulp2 .................................................................................38
-Other types of STUbLs ........................................................................................................................39
The DNA damage response .............................................................................................................40
-The DNA-damage checkpoints ...........................................................................................................40
-Postreplicative repair (PRR) ................................................................................................................41
-DNA double strand break (DSB) repair ...............................................................................................43
-Homologous recombination (HR) repair .............................................................................................44
Project Aim ...........................................................................................................................................46
Results .....................................................................................................................................................47
Manuscript: Concerted Action of the Ubiquitin-Fusion Degradation Protein 1 (Ufd1) and SUMO-Targeted Ubiquitin Ligases (STUbLs) in the DNA-Damage Response...................47
Investigation of a putative ”VIM” (VCP-interacting) motif in Slx8 .......................................69
Manuscript: Quantitative Identification of Sumoylation Sites in Fission Yeast Wild-type Cells, Ufd1 and SUMO-targeted ubiquitin Ligase (STUbL) Mutants ....................................73
A final discussion ............................................................................................................................... 148
References ............................................................................................................................................ 169
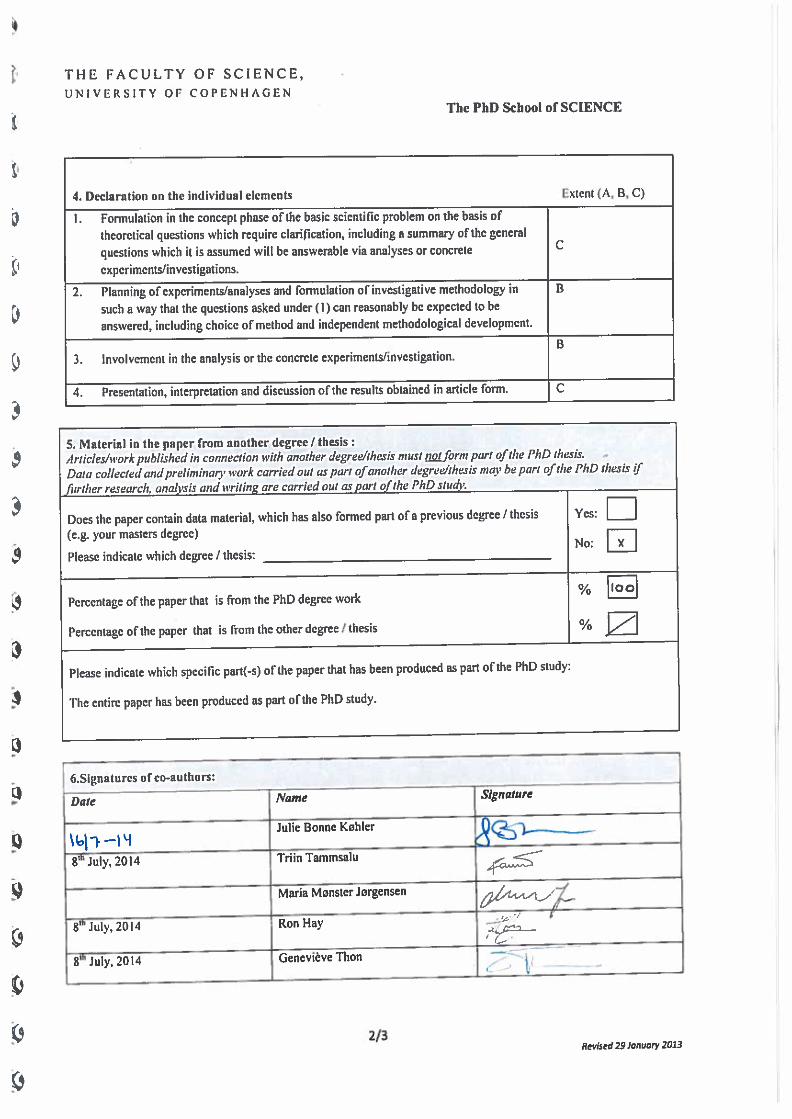
Co-author declarements .................................................................................................................. 187

3
Acknowledgements I would first of all like to thank my supervisor Geneviève Thon for all her tremendous support over
the years. She has been an extremely motivating and inspirational scientific guide and I am very
grateful for her always countable help and encouragement in any situation. Being in her lab has
been a very nice and learningfull experience. Secondly, I would like to thank Janne Verhein-Hansen
who has been a great technical support and a very pleasant colleague. I have been very grateful for
all her help. Similarly, I would like to thank all the current people of the lab and all the people who
have passed through the lab over the years and made it an enjoyable place to work. Special thanks
to Lærke Rebekka Holm, Maria Mønster Jørgensen and Tadas Jakociunas which have been my
fellow PhD students through most of the period, to Michael Thorsen and Gabriele Beinoraitė with
whom I collaborated on parts of my project and to Bethany, Dennis and Emil who have more
recently joined the lab. Also a big special thanks to all my lovely officemates; Irene, Sonia, Gjedre,
Louise and David, and to all the nice people in the corridor and building who makes the Biocenter a
great place to work. Lastly, but not least, thanks to my family and friends for all their support.

4
Summary Posttranslational modification by the ubiquitin or SUMO (small ubiquitin-like modifier)
polypeptides represents essential as well as evolutionary conserved ways of regulating the
proteomes of eukaryotic cells. Both modifiers generally change the function of their targets by
altering their conformation or interactions with other macromolecules. Though, whereas the
downstream consequence of ubiquitin conjugation is often protein degradation, the functional
outcomes of sumoylation are less unifiable. A class of ubiquitin E3 ligases able to target sumoylated
proteins for degradation by the 26S proteasome mediates direct cross-talk between the two
modification systems. By contributing to the dynamic turnover of SUMO conjugated species these
SUMO-targeted ubiquitin ligases (STUbLs) fulfills essential roles in both yeast and man. However,
the specific sumoylated proteins affected by STUbL activity and the specific molecular interactions
and sequence of events linking sumoylation, ubiquitylation and substrate degradation, has been
largely uncovered.
Using the fission yeast model organism I here present evidence for a role of the Ufd1 (ubiquitin-
fusion degradation 1) protein, and by extension of the Cdc48-Ufd1-Npl4 complex, in the STUbL
pathway. Cdc48-Ufd1-Npl4 forms a highly conserved molecular chaperone. By coupling ubiquitin-
selective binding with Cdc48 ATPase activity, Cdc48-Ufd1-Npl4 enables the mobilization of
ubiquitylated proteins from higher order complexes to promote their degradation or potential other
downstream fates. My work provides insight into how Cdc48-Ufd1-Npl4 also contributes to the
processing of SUMO conjugates and suggests that at least some of these activities are coordinated
with STUbL function. To gain insight into the sumoylated species regulated by Ufd1 and/or by
STUbLs, I made use of a newly developed two-step purification strategy for isolating sumoylated
species which allow their site-specific identification by mass spectrometry. In combination with
SILAC-based quantitative proteomics I compared sumoylation levels between wild type cells and
mutant strains deficient either in STUbL or Ufd1 function. In addition to identifying more than 900
unique sumoylated sites, these efforts revealed a number of proteins with upregulated sumoylation
either in STUbL and/or Ufd1 mutant cells. These findings propose specific candidate substrates
through which STUbL and Cdc48-Ufd1-Npl4 activities are coupled to dynamically regulate cellular
processes.

5
Introduction The ubiquitin-like protein family Cellular proteins are dynamically regulated by a variety of covalent modifications. These so-called
posttranslational modifications (PTMs) include covalent attachment of small chemical groups,
lipids, sugars or of other small proteins. By modulating the properties of proteins in often reversible
ways, PTMs diversify protein function and help drive biological processes. Ubiquitin and the
ubiquitin-like proteins (Ubls) are a family of posttranslational modifiers that are themselves small
proteins (Hochstrasser, 2009). The members of this family are unified by sharing a characteristic
three-dimensional structure, known as a β-grasp or ubiquitin-fold. In addition, Ubls share a similar
mechanism of protein attachment as they are always bound covalently to substrate proteins through
the carboxyl group of a C-terminal glycine residue. Substrate attachment is an energy-dependent
process with related, yet distinct, enzymatic pathways being dedicated to catalyzing conjugation by
the different Ubls. Ubl modifications are furthermore often transient as they are reversed by another
group of proteolytic enzymes that also display specificity for a given modifier. Despite these
structural and mechanistic similarities, it is clear that the different ubiquitin-like family members
affect protein function in distinct ways (Geiss-Friedlander & Melchior, 2007; Gill, 2004; Kerscher
et al, 2006; Komander, 2009; van der Veen & Ploegh, 2012).
Close to a dozen different Ubl proteins have been identified so far. Beside the founding member,
ubiquitin, they include (among others) the small ubiquitin-like modifiers (SUMOs), Ned8, the lipid
modifier Atg8 and the interferon-induced ISG15, which is only found in higher eukaryotes (van der
Veen & Ploegh, 2012). Of these, ubiquitin and SUMO are the ones which seemingly have the most
cellular targets (Golebiowski et al, 2009; Wagner et al, 2011; Wohlschlegel et al, 2004). Their
conjugation onto other proteins (referred to as ubiquitylation and sumoylation) plays both essential
and extremely widespread regulatory roles in all eukaryotes examined to date (Flotho & Melchior,
2013; Park & Ryu, 2014; Tanaka et al, 1999). Their most frequently used target site within proteins
is the ɛ-amino group of a lysine side chain. As both SUMO and ubiquitin possess the ability to self-
conjugate via internal lysine residues, they can modify proteins either as monomers or as polymeric
chains. Such polymeric chains can be of variable length and linkage type and thus lead to a variety
of modification signals (Kerscher, 2007; Komander & Rape, 2012). Ubiquitin is probably best
known for its role in targeting proteins for degradation by the proteasome when attached in the form

6
of a K48-linked chain (Hochstrasser, 1996b). Yet this is just one out of many signaling functions
mediated by this modifier. Similarly, conjugation by SUMO can have diverse downstream
consequences that are generally target-specific. Nevertheless, at the molecular level, both SUMO
and ubiquitin can be said to change the function of their targets by altering their interactions with
other macromolecules.
The SUMO modification system The SUMO family The small ubiquitin like modifiers (SUMOs) form a family of highly conserved eukaryotic proteins
each of around 100 amino acids in length. With a molecular weight of ~12 kDa, they are slightly
bigger than their ubiquitin cousin (8.5 kDa). Although the SUMOs and ubiquitin share very similar
three dimensional structures, they are only ~18% similar at the sequence level. One unique feature
of SUMO is a flexible N-terminal tail of ~20 amino acids which, as discussed later, is engaged in
SUMO polymerization (Bayer et al, 1998; Skilton et al, 2009). In S. pombe this N-tail is further
extended and phosphorylation of serine residues at the very N-terminus is proposed to contribute to
the stability of the SUMO molecule (Skilton et al, 2009).
Figure 1 (Adapted from Gill 2004). Ribbon diagrams of human SUMO-1 and ubiquitin. SUMO and ubiquitin share
very similar three-dimensional structures. Unique to SUMO is an elongated and flexible N-terminal tail. Secondary
structure elements are color coded with sheets represented in green and helices in red.

7
The SUMO proteins were first discovered in the mid-1990s. As they were identified in different
contexts, they were given a variety of names, such as Smt3C (also known as Ubl1, sentrin, Pic1 and
Gmp1), Smt3A and Smt3B. Today these are known as mammalian SUMO-1, -2 and -3,
respectively (Boddy et al, 1996; Lapenta et al, 1997; Mannen et al, 1996; Matunis et al, 1996;
Meluh & Koshland, 1995; Okura et al, 1996; Shen et al, 1996). Whereas mammals contain three
conjugatable SUMO isoforms, other eukaryotes like yeast, worms and flies express only a single
SUMO protein. At the other end of the spectrum, plants contain up to eight different SUMO forms
(Park & Yun, 2013).
The mammalian SUMO proteins Of the mammalian SUMO proteins, SUMO-1 shares about 50 % sequence identity with SUMO-2
and SUMO-3 that on the other hand are more than 95% alike. As SUMO-2 and -3 function in a
largely redundant manner, they are often referred to as one. In contrast, more functional divergences
exist between SUMO-2/3 and SUMO-1. For instance, although the two types of paralogues share
many substrates, they are also found to modify distinct sets of targets (Rosas-Acosta et al, 2005;
Vertegaal et al, 2006). Another difference is their ability to form chains, as these are more easily
formed by SUMO-2/3 (Tatham et al, 2001). It is also noteworthy that mammalian cells express
significantly more of SUMO-2/3 than of SUMO-1 (Saitoh & Hinchey, 2000). Compared to SUMO-
1, which is primarily detected in its conjugated form, the preponderance of SUMO-2/3 exists freely
in the cell. This large pool of free SUMO-2/3 is however seen to engage in conjugation upon
specific stimuli such as cellular stress. It has hence been suggested that SUMO-2/3 functions as a
SUMO reserve that can be activated upon demand (Saitoh & Hinchey, 2000). Despite functional
differences, the observation that SUMO-1 is not essential in mice suggests that at least the essential
functions of SUMO can be carried out by SUMO-2 and -3 (Evdokimov et al, 2008). It should also
be mentioned that a fourth SUMO protein (SUMO-4) exists in humans. Though, SUMO-4 seems to
lack the ability to be covalently attached to proteins and its cellular roles are at the moment still
relatively unclear (Owerbach et al, 2005).
SUMO in yeast The single SUMO gene of the budding yeast Saccharomyces cerevisiae was originally discovered
as a high-copy suppressor of mutations in mif2 (a gene encoding a centromeric protein) and was

8
consequently named smt3 (suppressor of mif2, clone 3) (Meluh & Koshland, 1995). The fission
yeast (Schizosaccharomyces pombe) homolog, pmt3, was later isolated in a two-hybrid screen with
PCNA (proliferating cell nuclear antigen) (Tanaka et al, 1999).The yeast SUMO proteins are most
identical to SUMO-1 with which they share ~50% sequence identity. The conservation of the
SUMO genes through evolution is underscored by the fact that human SUMO-1 is able to rescue
smt3∆ in budding yeast (Takahashi et al, 1999). Notably, while the smt3 gene is essential, S. pombe
pmt3∆ cells are viable. However, they are severely growth inhibited and extremely sensitive to
cellular stress. Another rare example of an organism in which the SUMO gene can be deleted is the
filamentous fungus, Aspergillus nidulans (Wong et al, 2008).
SUMO conjugation SUMO conjugation is stimulated by a SUMO-specific enzymatic cascade involving an E1
activating, an E2 conjugating and an E3 ligating enzyme. The SUMO pathway makes use of unique
E1 and E2 enzymes and only a few E3s. This is in contrast to the ubiquitin system where more than
one E1, several E2 and many hundred E3 enzymes exist (Komander, 2009).
Before SUMO can be recognized by the SUMO conjugating enzymes, it needs to be processed at its
extreme C-terminus to expose a diglycine motif used in substrate conjugation. This SUMO
maturation step is carried out by a SUMO-specific protease (see below). SUMO can then be
activated in an ATP-dependent reaction by the E1. The SUMO E1 is made up of two subunits,
SUMO-activating enzyme 1 and 2 (Sae1/Sae2), also known as Aos1/Uba2 in humans or
Rad31/Fub2 in S. pombe . In the E1 activation step a high-energy thioester bond is formed between
the SUMO C-terminus and the active-site cysteine of the Sae2 subunit. Through interactions
between the E1 and E2, SUMO is then transferred onto another catalytic cysteine of the E2 enzyme.
The E2, which in most organisms is known as Ubc9 (or Hus5 in S. pombe) (al-Khodairy et al, 1995;
Desterro et al, 1997; Ho et al, 2001; Johnson & Blobel, 1997), forms another intermediate thioester
with SUMO before an E3 ligase catalyzes the transfer of SUMO onto a substrate. The net result is
the formation of an isopeptide bond between the carboxyl group of the C-terminal SUMO glycine
and the ɛ-amino group of a lysine residue within the substrate (Figure 2) (Flotho & Melchior, 2013;
Geiss-Friedlander & Melchior, 2007). The E3 ligases mediate this last reaction step by two overall
means. Firstly, by binding both the substrate and the E2, the E3 functions as a bridge that helps
bring the substrate and E2~SUMO into close enough proximity for conjugation to occur. Secondly,

9
the E3 helps orient the E2~SUMO thioester in a configuration that is optimal for nucleophilic attack
by an incoming substrate lysine (Reverter & Lima, 2005; Yunus & Lima, 2009).
Figure 2 (Adapted from Geiss‑Friedlander and Melchior, 2007). Schematic depiction of the SUMO conjugation cycle.
Before SUMO can be conjugated onto substrate proteins it first needs to be maturated to expose a C-terminal diglycine
motif. This is stimulated by a SUMO-specific protease (SENP). Then an E1 activating enzyme (Aos1/Uba2) forms an
intermediate thioester bond with SUMO before transferring it to the SUMO E2 (Ubc9). In a final step, an E3 acts to
stimulate substrate conjugation onto a lysine (K) residue. Sumoylation is a reversible process due to the SUMO
proteases which are responsible for cleaving SUMO of its targets.
The SUMO E3s The SUMO E3 ligases that have been described to date can be classified into two overall groups;
ligases belonging to the zinc-coordinating SP-RING family, and ligases without a RING motif. The
SP-RING domain E3s make up the largest group. They include the mammalian PIAS proteins
(Protein-Inhibitors of Activated STAT) (Sachdev et al, 2001; Schmidt & Muller, 2002) and their
yeast PIAS homologs Siz1 and Siz2 in S. cerevisae (Johnson & Gupta, 2001; Takahashi et al, 2001)
and Pli1 in S. pombe (Xhemalce et al, 2004). Non-PIAS SP-RING E3s include the conserved
Smc5/6 complex component Mms21, also known as Nse2 in S. pombe (Andrews et al, 2005; Potts
& Yu, 2005). The SP-RING motif of these enzymes is structurally very similar to the RING and
RING-like U-box domains of many ubiquitin E3s and these domains are thought to function in
similar ways (Plechanovova et al, 2012; Yunus & Lima, 2009). The SP-RING is responsible for
binding E2 and is thus essential for the E3 function (Sachdev et al, 2001; Schmidt & Muller, 2002;

10
Takahashi et al, 2001). In addition, structural studies of the Siz1 (PIAS) ligase have revealed acidic
residues C-terminal to the SP-RING which by interacting with a basic patch on SUMO help
stabilize the E2~SUMO thioester in its catalytic active conformation (Yunus & Lima, 2009).
Notably, in this E3 RING:E2~SUMO intermediate complex there is never any covalent interaction
formed between the E3 and SUMO. This is in contrast to the HECT-domain E3s found in the
ubiquitin pathway where an intermediate thioester bond is formed between the ligase and ubiquitin
(Huibregtse et al, 1995). Beside the domains mediating E2~SUMO interactions, other parts of the
E3s are involved in recruiting substrates, as shown for the N-terminal PINIT domain of Siz1 (Yunus
& Lima, 2009). Due to variations in these interaction surfaces, the various E3s exhibit distinct
substrate preferences, even though they may also share some substrate redundancy (Makhnevych et
al, 2009; Reindle et al, 2006; Watts et al, 2007). The different E3s may also localize to differential
subcellular regions, explaining some of their observed functional divergences (Johnson & Gupta,
2001; Psakhye & Jentsch, 2012; Reindle et al, 2006).
The vertebrate-specific nucleoporin RanBP2 is the best established non-RING protein exhibiting
SUMO E3 activity (Pichler et al, 2002). RanBP2 is associated with the cytosolic side of nuclear
pores in interphase cells and with kinetochores and the mitotic spindle during mitosis. Its E3 active
region shows no homology to other known ubiquitin-like E3 ligases (Pichler et al, 2004). RanBP2
forms a stable complex with Ubc9 and sumoylated RanGAP1 (Ran GTPase-activating protein). It
was recently shown that this complex in fact constitutes the actual ligase and thus works as a multi-
subunit ligase similar to some composite E3s found in the ubiquitin pathway (Werner et al, 2012).
Although RanBP2 does not contain a RING motif, its way of catalyzing SUMO transfer is thought
to be similar to the SP-RING E3s, namely by affecting the E2~SUMO thioester through non-
covalent interactions (Reverter & Lima, 2005; Werner et al, 2012).
Other types of proteins have also been described in the literature to function as SUMO E3s. These
include among others the human polycomb protein Pc2, Topors (topoisomerase I-binding RING
finger protein) and the transcription factor Krox20. However, whether these proteins are function as
“true” enzymatic E3s, or more as scaffolds enhancing sumoylation of specific interaction partners,
is still debated (Flotho & Melchior, 2013).

11
SUMO E3s in fission yeast In yeast only the SP-RING type of SUMO E3s are known to exist. These, which in S. pombe
include Pli1 and Nse2, are seen to mediate largely individual cellular roles. While Pli1 is
responsible for the vast majority of detectable sumoylation in the cell, Nse2-mediated sumoylation
has a special function during the response to DNA damage (Watts et al, 2007). Genetic data suggest
that such differential Pli1 and Nse2-stimulated sumoylation is regulated by distinct non-covalent
partner choices of Ubc9 (Prudden et al, 2011). For example, a non-covalent complex formed of
Ubc9 and SUMO (Capili & Lima, 2007; Knipscheer et al, 2007) stimulates bulk sumoylation
together with Pli1. Alternatively, the same residues of Ubc9 may engage in a complex with the
SUMO-like domain of the Rad60 protein (human NFATC2IP), which by interacting with the
Smc5/6 complex can in turn recruit Ubc9 to Nse2-dependent sumoylation processes (Prudden et al,
2011). Yet, the two ligases may also be able to compensate for each other and they may share
common substrates (Watts et al, 2007). Whereas pli1∆ and a sumoylation deficient nse2-SA mutant
do not by themselves reconstitute the phenotypic abnormalities observed for the pmt3∆ deletion, the
double mutant is barely viable, consistent with redundancy between the activities of the two
proteins.
Substrate selection A common target site for SUMO within substrate proteins are lysine residues lying within a
sequence context of ψKXE/D (where ψ is a large hydrophobic amino acid and X any residue)
(Rodriguez et al, 2001), as this motif can be directly recognized by Ubc9 (Bernier-Villamor et al,
2002). An unbiased proteomic study found at least half of all SUMO targets to contain this SUMO
consensus site (Matic et al, 2010). Occasionally this motif is expanded to include a phosphorylated
serine or a small patch of negatively charged residues situated a few amino acids downstream
(Hietakangas et al, 2006; Yang et al, 2006), making up what is known as an PDSM
(phosphorylation-dependent SUMOylation motif) or an NDSM (negatively charged amino acid-
dependent SUMOylation motif), respectively. A positively charged basic patch in Ubc9 can interact
with these negatively charged residues and thus further increase substrate recognition (Yang et al,
2006). Other variations, like inverted (E/DXKψ) or hydrophobic (ψψψKXE) consensus sites have
also been reported (Matic et al, 2010). Intriguingly, modification of consensus lysines can often be
stimulated in the absence of an E3 in vitro, if the Ubc9 concentration is high enough (Yunus &

12
Lima, 2005). However, even though Ubc9 has an important role in target selection, assistance of an
E3 enzyme is still crucial for substrate modification in vivo, especially for lysines not conforming to
the ψKXE/D motif. By interacting with alternative regions of the substrates they provide an extra
layer of specificity to the reaction and they enhance conjugation also to consensus sites (Yunus &
Lima, 2009). For example, although SUMO conjugation to a consensus lysine (K127) in the
proliferating cell nuclear antigen (PCNA) can be stimulated by Ubc9 alone, its efficient in vivo
modification requires Siz1. Furthermore, modification of the PCNA non-consensus lysine (K164) is
strictly Siz1-dependent (Yunus & Lima, 2009).
Another molecular feature that can help direct SUMO conjugation is the presence of SUMO-
interaction motifs (SIMs, see below) within target proteins. These motifs interact non-covalently
with SUMO and can by recruiting SUMO-loaded Ubc9 stimulate sumoylation of nearby lysines.
SIM-directed sumoylation can be considered a rather promiscuous event in that any proximal lysine
seems to be able to serve as a target (Flotho & Melchior, 2013; Zhu et al, 2008).
SUMO chains The SUMO proteins of both yeast and mammals contain lysine residues within their unstructured
N-terminal domains that can function as acceptor sites for conjugation by other SUMO molecules,
leading to the formation of SUMO chains (Bylebyl et al, 2003; Matic et al, 2008; Tatham et al,
2001; Ulrich, 2008). In S. pombe, SUMO chains are primarily formed through the two non-
consensus lysines, K14 and K30 (Prudden et al, 2011; Skilton et al, 2009). Structural studies have
revealed a non-covalent interaction formed between SUMO and a surface of Ubc9 opposite its
active site that is required during SUMO polymerization. This Ubc9:SUMO interaction is predicted
to help orient an accepting SUMO molecule in a way that allows its N-terminus to engage in
conjugation by another Ubc9~thioester linked SUMO donor (Capili & Lima, 2007; Knipscheer et
al, 2007). Consistent with this model, a SUMO mutation which specifically disrupts the non-
covalent Ubc9:SUMO interface abolishes the detection of SUMO chains in S. pombe cells (Prudden
et al, 2011). Although formation of SUMO chains can be stimulated in vitro by either of the two
fission yeast SUMO ligases (Skilton et al, 2009), Pli1 has been suggested to be the major ligase
responsible for polysumoylation in vivo (Prudden et al, 2011). The Siz1/Siz2 homologs similarly
seem to stimulate SUMO chain formation in budding yeast (Bylebyl et al, 2003).
In addition to the situation where a chain of SUMO moieties is attached to a single substrate lysine,
several mono SUMOs may be simultaneously attached to distinct lysine residues within the same

13
protein, leading to multisite modification (Tammsalu et al, 2014). The distinction between these two
phenomenons, poly- versus multiple monosumolyation, may be difficult to determine as both result
in the slowed migration of modified proteins in denaturing gels (Bekes et al, 2011; Tatham et al,
2011; Ulrich, 2008)
SUMO deconjugation Sumoylation is, as for modification with other ubiquitin-like proteins, a reversible process.
Desumoylation is carried out by a specific class of proteases capable of hydrolyzing SUMO
isopeptide bonds. By cleaving SUMO of its targets, these enzymes allow proteins to return to their
unmodified state and SUMO to re-enter its unconjugated pool for use in other modification events.
The SUMO-specific proteases where first defined in budding yeast by the Ulp1 protein (Li &
Hochstrasser, 1999). The Ulp1 S. pombe homolog was characterized later by Taylor et al., 2002. In
addition to mediating SUMO deconjugation, Ulp1 is also required for processing inactive SUMO
into its mature form. Another SUMO protease in yeast is Ulp2 (Li & Hochstrasser, 2000). Ulp2 is
not engaged in SUMO maturation but instead has a special role in depolymerizing SUMO chains.
Though, Ulp2 can also cleave linkages between SUMO and substrates (Bylebyl et al, 2003). The
mammalian homologs of the Ulps are the so-called SENPs (sentrin-specific proteases) of which six
different types exist (Hickey et al, 2012). The Ulps/SENP family all share related core cysteine
protease domains in their C-termini (Hickey et al, 2012; Mukhopadhyay & Dasso, 2007). Their N-
termini contain regions responsible for targeting the proteins to specific subcellular compartments
(Kroetz et al, 2009; Li & Hochstrasser, 2003). While Ulp1 (SENP1 and -2) localizes to the inner
surface of the nuclear pore complex (except during mitosis in S. pombe) (Schwienhorst et al, 2000;
Taylor et al, 2002), Ulp2 (SENP6 and -7) is scattered throughout the nucleoplasm (Li &
Hochstrasser, 2000; Mukhopadhyay et al, 2006). These restricted localization patterns are major
determinants of which substrates the enzymes are able to de-modify (Li & Hochstrasser, 2003). In
line with individual roles for the different SUMO proteases, the two Ulp enzymes in yeast are
unable to complement for each other. While Ulp1 is essential in budding yeast (Li & Hochstrasser,
1999), ulp2 deleted cells are viable but grow slowly, are sensitive to various kinds of stress and they
are defective in chromosome segregation and recovery from checkpoint-induced cell cycle arrest
(Bylebyl et al, 2003; Li & Hochstrasser, 2000; Prudden et al, 2011; Schwartz et al, 2007). The
essential function of Ulp1 is due to its role in SUMO C-terminal processing as supplying cells with
maturate SUMO (SUMO-GG) restores viability ((Li & Hochstrasser, 1999). Though even with this,

14
ulp1∆ cells are still extremely sick. In contrast to budding yeast, ulp1∆ is not lethal in S. pombe.
This is in line with SUMO itself not being required for viability of this organism (Taylor et al,
2002).
Other more recently recognized SUMO-specific proteases (also of the cysteine type) are the
mammalian proteins DESI1, DESI2 and USPL1 (Hickey et al, 2012). However, these seem to play
more restricted roles than the Ulp/SENP proteins and they are likely to only affect very specific
substrates (Flotho & Melchior, 2013). Yeast may also contain other types of SUMO deconjugases
as has been demonstrated in S. cerevisae for the metalloprotease Wss1 (Mullen et al, 2010).
Table 1 (Adapted from Jentsch and Psakhye 2013). Overview of the SUMO pathway enzymes in S. cerevisae, S. pombe
and humans, respectively. Mms21a/Nse2a homologs form part of the evolutionary-conserved Smc5/6 complex
(structural maintenance of chromosomes). PIAS2b is also known as PIASxβ. In addition shorter isoform of PIAS2 with
truncated C-terminus are known as PIASxα.

15
Molecular effects of SUMO conjugation
SUMO generally changes the function of its targets by modulating their conformation or interaction
with other macromolecules. SUMO may in this way alter the activity, stability, localization or
binding partner affinity of substrate proteins. In particular, SUMO attachment can add a new
binding surface to modified proteins, allowing them to interact with a different set of downstream
factors. Accordingly, many downstream interactors of sumoylated proteins contain specialized
SUMO-interaction motifs (SIMs) (Hannich et al, 2005; Lin et al, 2006; Makhnevych et al, 2009;
Pfander et al, 2005) which enables them to form non-covalent interactions with SUMO or SUMO-
modified proteins (Kerscher, 2007). SIMs are short linear motifs characterized by a hydrophobic
core often flanked by some acidic residues. They are typically found in unstructured regions and
adopts into a β strand conformation upon binding to a hydrophobic groove present in SUMO. The
classically defined SIMs have a core consensus of typically (V/I)-X-(V/I)-(V/I) or (V/I)-(V/I)-X-
(V/I) (Hecker et al, 2006; Song et al, 2004). Another subgroup of higher affinity SIMs have also
been described; (V/I/L/F/Y)-(V/I)-DLT (Sun & Hunter, 2012). Though, similar to other PTM
sensing motifs (Husnjak & Dikic, 2012), SIMs generally mediate relatively low affinity interactions
(Escobar-Cabrera et al, 2011; Keusekotten et al, 2014; Tatham et al, 2008). In some cases SUMO-
SIM interactions are complemented by additional associations mediated directly by the sumoylated
target and the SIM containing protein. This is for example the case for the Srs2 helicase which is
recruited to sumoylated PCNA both via a SIM and via PCNA binding module (Armstrong et al,
2012). Some proteins contain multiple SIMs and can in turn specifically bind to and mediate the
effects of poly-SUMO chains (Sun & Hunter, 2012; Tatham et al, 2008; Ulrich, 2008). Interactions
with SIMs thus represent a key way in which SUMO is able to modulate protein-protein
interactions.
An alternative mode by which SUMO may alter protein function is by masking pre-existing
interaction sites within a substrate, in this way preventing rather than enhancing specific
interactions. For example, SUMO may block for other posttranslational modifications such as
ubiquitination or phosphorylation and thereby antagonize their effects (Desterro et al, 1998; Ulrich,
2005). Finally, SUMO attachment may cause conformational changes in its targets. Such changes
may also be induced by SIMs as has been exemplified for the thymidine DNA glycosylase (TDG)
16
where an intramolecular SUMO-SIM interaction regulates its conformation and hence enzymatic
activity (Steinacher & Schar, 2005).
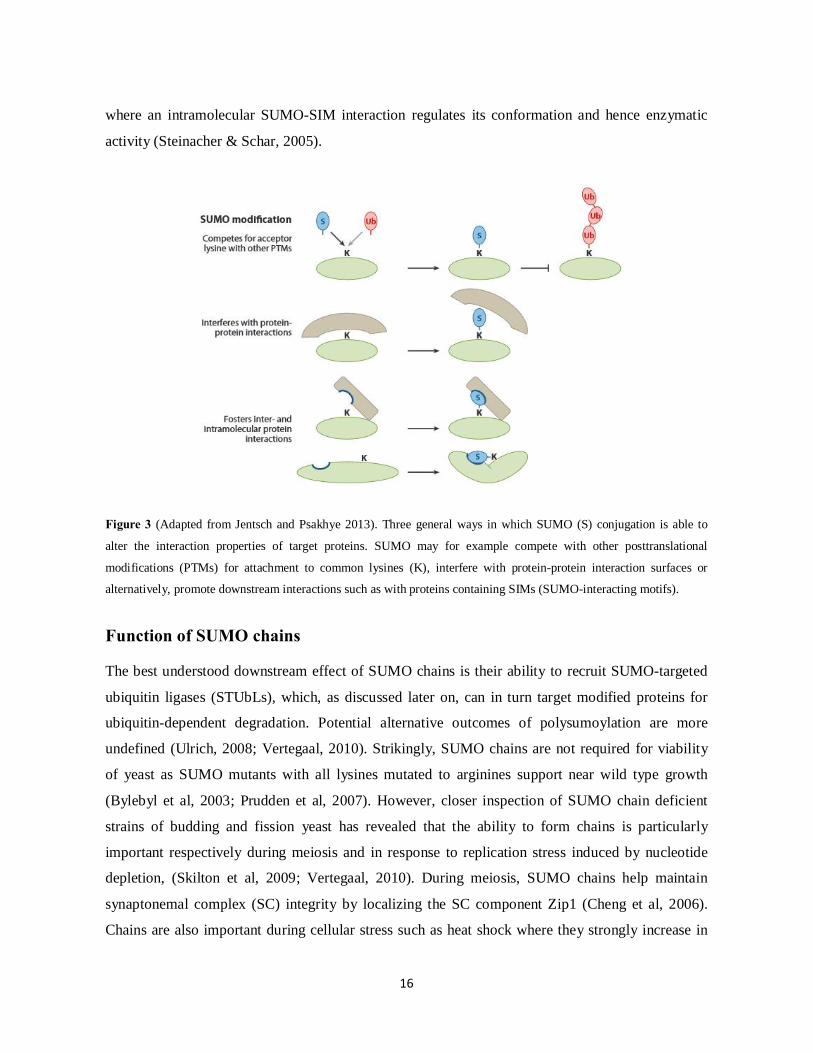
Figure 3 (Adapted from Jentsch and Psakhye 2013). Three general ways in which SUMO (S) conjugation is able to
alter the interaction properties of target proteins. SUMO may for example compete with other posttranslational
modifications (PTMs) for attachment to common lysines (K), interfere with protein-protein interaction surfaces or
alternatively, promote downstream interactions such as with proteins containing SIMs (SUMO-interacting motifs).
Function of SUMO chains The best understood downstream effect of SUMO chains is their ability to recruit SUMO-targeted
ubiquitin ligases (STUbLs), which, as discussed later on, can in turn target modified proteins for
ubiquitin-dependent degradation. Potential alternative outcomes of polysumoylation are more
undefined (Ulrich, 2008; Vertegaal, 2010). Strikingly, SUMO chains are not required for viability
of yeast as SUMO mutants with all lysines mutated to arginines support near wild type growth
(Bylebyl et al, 2003; Prudden et al, 2007). However, closer inspection of SUMO chain deficient
strains of budding and fission yeast has revealed that the ability to form chains is particularly
important respectively during meiosis and in response to replication stress induced by nucleotide
depletion, (Skilton et al, 2009; Vertegaal, 2010). During meiosis, SUMO chains help maintain
synaptonemal complex (SC) integrity by localizing the SC component Zip1 (Cheng et al, 2006).
Chains are also important during cellular stress such as heat shock where they strongly increase in

17
abundance (Golebiowski et al, 2009). A proteomic study enriching for polysumoylated proteins in
human cells after heat shock identified more than 300 poly-modified proteins with a high
representation of factors involved in mRNA splicing, chromatin architecture and transcription as
well as DNA replication and repair (Bruderer et al, 2011). More recently, a detailed study in
budding yeast analyzing a chain-deficient SUMO mutant (smt3KallR), revealed a general role for
SUMO chains in the maintenance of higher-order chromatin structure. More specifically, these
authors found the smt3KallR strain to show a general defect both in chromosome compaction and
organization which likely caused the derepression of otherwise inactive genes (Srikumar et al,
2013).
SUMO substrates In line with nuclear enrichment of SUMO and SUMO pathway enzymes, a major proportion of
identified SUMO targets are nuclear proteins (Golebiowski et al, 2009; Hay, 2013; Tanaka et al,
1999; Taylor et al, 2002; Watts, 2013). Accordingly, a major focus has been on the roles played by
SUMO in multiple nuclear events such as transcription, chromatin structure and dynamics, nucleo-
cytoplasmic transport and DNA replication and repair (Cubenas-Potts & Matunis, 2013; Seeler et
al, 2007; Ulrich, 2014). However, SUMO targets also include cytoplasmic proteins (Flotho &
Melchior, 2013; Manning Fox et al, 2012; Zunino et al, 2007). Thus the repertoire of SUMO
substrates is diverse and the breadth of cellular pathways reported to be influenced by SUMO is
continuously expanding (Hannich et al, 2005; Makhnevych et al, 2009; Tatham et al, 2011;
Westman et al, 2010; Wohlschlegel et al, 2004).
Protein group modification It is seen that SUMO often targets multiple components of physical and/or functional complexes
(Golebiowski et al, 2009; Psakhye & Jentsch, 2012; Wohlschlegel et al, 2004). One emerging theme
is that concerted targeting of SUMO to networks of interacting proteins work to stabilize physical
complexes through multiple SUMO-SIM interactions, a concept known as “protein group
modification” (Jentsch & Psakhye, 2013). Collective protein group modification may be triggered
by a specific internal or external stimulus to assist a coordinated cellular response (Johnson &
Blobel, 1999; Psakhye & Jentsch, 2012; Schimmel et al, 2014). Protein group sumoylation has been
exemplified during the response to DNA damage in S. cerevisae cells. Here, DNA damage induced

18
generation of single-stranded DNA stimulates recruitment of the Siz2 SUMO ligase to trigger a
“wave” of SUMO modifications with similar kinetics on multiple repair components. Interestingly,
substrate specificity seemed in this case to be conferred by the damage induced enrichment of the
Siz2 SUMO ligase at the damage site. During this so-called “SUMO wave”, single modifications
were seen to work synergistically and/or redundantly, as only abrogation of multiple sumoylation
events led to obvious repair defects (Psakhye & Jentsch, 2012). Although such functional
redundancy in sumoylation accounts for some SUMO targets, single target modifications may still
mediate more explicit downstream molecular effects (Miyagawa et al, 2014; Papouli et al, 2005;
Pfander et al, 2005).
Identification of SUMO substrates Insight into the specific proteins modified by SUMO is central to understanding how SUMO
functions. However, SUMO substrate identification has traditionally been difficult due to the very
low abundance of sumoylated species in the cell and the high activity of SUMO proteases in cell
lysates. In particular, often only a small fraction of the total amount of a protein is modified by
SUMO at any given time (Hay, 2005; Sacher et al, 2005; Tatham et al, 2011). To meet these
obstacles, methods have gradually been developed which together have allowed identification of
many hundred putative SUMO substrates in different cell types (Barysch et al, 2014; Golebiowski
et al, 2009; Hannich et al, 2005; Miller et al, 2010; Vertegaal et al, 2004; Wohlschlegel et al, 2004;
Zhao et al, 2004). Two overall approaches have been applied, namely candidate-based approaches,
where suspected SUMO substrates are tested individually, and large-scale approaches aiming at
detecting sumoylated proteins in an unbiased manner. In the candidate-based approaches, candidate
proteins are often selected based on their presence of SUMO consensus sites. Though, as it is
becoming increasingly clear that SUMO also often modifies lysines lying outside any consensus
context (Matic et al, 2010; Xu et al, 2008), such strategy is clearly inadequate in predicting the
entire SUMO proteome.

19
In vitro identification
A method that has been widely used to test candidate SUMO substrates is by in vitro sumoylation
assays (Desterro et al, 1999). This strategy circumvents the in vivo problems of detecting low
modification levels (Gocke & Yu, 2009). In in vitro assays, a candidate substrate is typically mixed
with recombinant versions of SUMO and the SUMO pathway enzymes (E1, E2 and E3) in the
presence of ATP (Flotho et al, 2012). Detection of sumoylated reaction products can afterwards be
analyzed by SDS-PAGE/Western blotting. With an aim to unbiasedly identify SUMO substrates,
laboratories have furthermore adapted the system to even screen a whole pool of in vitro translated
proteins from cDNA libraries (Gocke et al, 2005). One problem with in vitro modification reactions
is, however, that they generally contain non-physiological high concentrations of the different
reagents which make them prone to catalyzing unspecific sumoylation products. Misfolding of
recombinant proteins may similarly stimulate their aberrant modification. Thus, despite the
usefulness of in vitro assays in establishing whether a protein at least in theory can serve as a
SUMO target, evidence for in vivo modification is necessary to demonstrate a biological relevance
(Ulrich & Davies, 2009).
In vivo identification Due to the fact that SUMO-modified forms of proteins are often only present in sub-stoichiometric
amounts, any effort aimed at their identification in vivo relies on the ability to efficiently enrich for
them. A range of affinity-based enrichment strategies have been employed to detect sumoylated
forms of proteins either from purifications of specific candidate proteins or from enrichments of
SUMO itself (Cremona et al, 2012; Da Silva-Ferrada et al, 2013; Hoege et al, 2002; Vertegaal et al,
2004). In many cases SUMO or candidate target proteins have been overexpressed from ectopic
promoters to increase their detection level. Fusion to the metal-binding hexahistidine (His6) tag is
commonly used in SUMO protocols as it allows purifications to be carried out under denaturing
conditions which abolish non-covalent interactions and greatly help preserve modifications by
inactivating the SUMO proteases (Ulrich & Davies, 2009). Alternatively, SUMO-protease
inhibitors can be added to native lysis buffers (Sacher et al, 2005). Sumoylated species derived
from in vivo purifications can either be identified by Western blotting (Cremona et al, 2012; Ulrich
& Davies, 2009) or with higher sensitivity, by mass spectrometry (Filosa et al, 2013).

20
Mass-spectrometry (MS)-based proteomics Development of mass-spectrometry (MS)-based proteomics methods to investigate protein
posttranslational modifications (PTMs) such as sumoylation represents a major breakthrough in the
attempts to unbiasedly identify PTM targets at a global scale (Filosa et al, 2013; Wilson & Heaton,
2008). In a typical MS procedure, protein mixtures, derived either from total lysates or from
enriched samples, are digested with a protease to generate smaller peptides which are more easily
analyzed by MS (Cox & Mann, 2011). This approach where peptides are analyzed to infer
information of the proteins in the sample is also known as peptide-based or “bottom-up”
proteomics. A frequently used enzyme in proteomic studies is trypsin which cleaves at the C-
terminal side of both lysine and arginine residues. Digested peptides are then separated by reversed-
phase liquid chromatography and ionized in gas phase before injection into the mass spectrometer.
Inside the mass spectrometer, ionized peptides are first separated based on their mass-to-charge
(m/z) ratio. The most intense peptidic ions are then selected and fragmented through collision-
induced dissociation (CID) to generate smaller ions with characteristic m/z ratios. This event is also
known as tandem MS (MS/MS) (Altelaar et al, 2013; Filosa et al, 2013). The measured m/z ratios of
precursor peptides and their fragmented ions are used to reconstruct peptide sequences using search
engines able to match them with known protein sequences (Steen & Mann, 2004). Identification of
PTMs is enabled by the training of these programs to search for deviations in mass values due to
specified modifications. MS analysis has been applied on samples derived from various kinds of
SUMO-conjugate enrichment strategies and has in many cases been able to identify hundreds of
putative SUMO substrates in a single experiment (Bruderer et al, 2011; Golebiowski et al, 2009;
Rosas-Acosta et al, 2005; Wohlschlegel et al, 2004). The application of MS in combination with
quantitative approaches such as SILAC (stable isotope labeling of amino acids in cell culture) is
furthermore extremely useful for assessing relative changes in PTM abundance during different
experimental conditions (Choudhary & Mann, 2010). Quantitative proteomics has for example been
used to quantify global changes in sumoylation during the cell cycle (Schimmel et al, 2014) and in
response to cellular stress such as heat shock (Golebiowski et al, 2009), proteasomal inhibition
(Schimmel et al, 2008; Tatham et al, 2011) or DNA damage (Psakhye & Jentsch, 2012; Yin et al,
2012).
Although numerous putative SUMO targets have been identified by MS analysis of various protein
preparations, the incomplete stringency of many enrichment strategies result in co-purification of
unspecific targets which may lead to false-positives. Thus to gain definite proof of SUMO

21
modifications, the specific SUMO acceptor sites needs to be determined (Filosa et al, 2013). Such
identification is furthermore crucial in order to examine the specific effects of individual
sumoylation events, for example by enabling their functional characterization through site-specific
mutagenesis. However, information of SUMO acceptor sites is generally difficult to extract from
complex sample mixtures such as derived from protein-level purifications of total SUMO species.
Site analysis is further complicated by the long peptide remnant left of SUMO on modified lysines
after trypsin cleavage. These fragments comprise for instance 19 and 32 amino acids for SUMO-1
and SUMO-2/3, respectively, and 23 residues for the S. pombe SUMO protein. The result is
branched peptides that generate MS/MS fragmentation spectra which are difficult to interpret.
Strategies to overcome these obstacles have been developed. They generally involve the
introduction of protease cleavage sites into the C-terminus of SUMO which allows the SUMO
remnant on modified peptides to be shortened to a size that can be predicted by MS (Knuesel et al,
2005; Matic et al, 2010; Wohlschlegel et al, 2006). Though, despite these various efforts in
identifying sumoylation sites, they have only been moderately successful in identifying sites at a
large scale (Matic et al, 2010). By contrast, methods applied within the ubiquitylation field have
enabled identification of more than 10.000 ubiquitylation sites within a single experiment (Wagner
et al, 2011; Wagner et al, 2012). This success in ubiquitin site identification has been empowered
by the development of antibodies directed against the digly-remnant left of ubiquitin on acceptor
lysines after trypsination (Xu et al, 2010)(Xu et al., 2010), highlighting the power of peptide level
enrichment strategies for efficient PTM identification (Choudhary et al, 2009; Olsen et al, 2006).

22
The ubiquitin modification system Covalent modification by ubiquitin Covalent conjugation of the 76 amino acid ubiquitin polypeptide to lysine residues of cellular
proteins is one of the most common posttranslational modifications in eukaryotic cells. As for
SUMO, ubiquitin attachment is brought about by a hierarchal E1, E2 and E3 pathway, involving
related enzymatic steps. Protein deubiquitylation can similarly be stimulated by specialized
ubiquitin proteases. Though compared to sumoylation, the ubiquitylation process makes use of a
much greater repertoire of conjugating and deconjugating enzymes, generally displaying a very
high degree of substrate specificity. For instance, in the mammalian ubiquitin cascade, at least two
E1s, approximately 40 E2s and more than 600 different E3s are known to exist (Kerscher et al,
2006; Komander, 2009). Ubiquitin is able to modify proteins either as a monomer or in the form of
polymeric chains. Ubiquitin contains seven lysines of which all can engage in ubiquitin
polymerization, leading to various chain topologies (Peng et al, 2003; Xu et al, 2009). In addition,
linear fusions may occur via the free amine of the ubiquitin N-terminus (Tokunaga et al, 2009). The
length and linkage type of conjugated ubiquitin molecules largely determine the downstream
consequences of the modification (Komander & Rape, 2012; Zhao & Ulrich, 2010). One major
effect of ubiquitylation is the targeting of proteins for degradation by the 26S proteasome. However,
ubiquitin is also seen to mediate many non-proteolytic functions. Non-degradative roles for
ubiquitin have in particular been elucidated in the contexts of cellular signaling, gene regulation,
endocytosis and DNA repair (Hochstrasser, 1996a; Polo, 2012; Ulrich, 2014; Zhao et al, 2014). The
differential functional outcomes of ubiquitylation are mediated through downstream effectors
containing a variety of non-covalent ubiquitin-binding modules able to recognize particular types of
ubiquitin modifications. More than twenty different types of ubiquitin-binding domains (UBDs)
have been described to date which diverges both in their structure and in their mode of ubiquitin
interaction (Dikic et al, 2009; Hanzelmann et al, 2012; Komander & Rape, 2012).

23
The ubiquitin-proteasome system (UPS) The ubiquitin-protesome system (UPS) represents the major pathway for selectively degrading
proteins in eukaryotic cells, affecting both nuclear and cytoplasmic proteins (Schmidt & Finley,
2014; Wojcik & DeMartino, 2003). In addition to eliminating misfolded and dysfunctional proteins
(Matsuo et al, 2011; Sontag et al, 2014), the UPS system provides a mechanism for irreversibly
controlling protein activity during a wide variety of cellular processes, including cell cycle
progression, differentiation, signal transduction and DNA repair (Koepp, 2014; Laney &
Hochstrasser, 2004; Motegi et al, 2009). Such proteolytic control can be very effective as it ensures
that the function performed by a specific protein is completely arrested. Consistent with the
widespread implications for UPS-mediated degradation in cellular pathways, defects in the system
is associated with the development of various human diseases (Cuanalo-Contreras et al, 2013;
Devoy et al, 2005; Ding et al, 2014). The degradation of a protein is a highly regulated event
initiated by its marking by the ubiquitylation machinery. The specific proteolytic signal is a
ubiquitin chain of at least four molecules generally linked through lysine 48 (K48) (Thrower et al,
2000). However, alternatively linked ubiquitin chains have also been suggested to stimulate
proteasomal targeting (Dammer et al, 2011; Johnson et al, 1995; Xu et al, 2009). Once at the 26S
proteasome, substrates are first unfolded and deubiquitinated at the 19S regulatory particle and then
passed into the 20S catalytic core for final hydrolysis (Hochstrasser, 1996b; Schrader et al, 2009).
Essential to the degradation process is the selective docking of the poly-ubiqutinated protein onto
the proteasome. While some substrates are directly recognized by components of the 19S
regulatory particle (Elsasser et al, 2002; Husnjak et al, 2008), other substrates require additional
factors to assist their proteasome delivery. Several such factors working in-between the
ubiquitination machinery and the proteasome have been identified, greatly expanding the regulative
potential for the degradation process and suggesting that various routes to the proteasome are likely
to exist (Elsasser et al, 2004; Finley et al, 2012). The recognition of ubiquitin-marked proteins by
both proteosomal and extra-proteasomal adaptor proteins is mediated through different types of
ubiquitin-binding domains (UBDs), of which the ubiquitin-associated (UBA) domain and the
ubiquitin-interacting (UIM) motif are especially common in the ubiquitin-proteasome system (Dikic
et al, 2009).

24
The Cdc48/p97 ubiquitin-selective ATPase An important effector in the ubiquitin-proteasome pathway is the ubiquitin-selective Cdc48 ATPase
(also known as p97 or VCP in mammals) of the AAA family (ATPases associated with various
cellular activities) which is a highly conserved and distant relative of the 19S proteasome particle.
Although originally identified in a screen for genes required for the cell division cycle (cdc) of S.
cerevisiae (Moir et al, 1982), Cdc48/p97 is now known to be implicated in an extremely wide range
of ubiquitin-dependent cellular processes, often involving its ability to promote UPS-mediated
proteolysis (Baek et al, 2013; Franz et al, 2014; Ghislain et al, 1996; Meyer et al, 2012). These
include (among many others) cell-cycle control, endoplasmic reticulum-associated degradation
(ERAD), homo-typic membrane fusion and transcription factor activation (Bays et al, 2001; Cao et
al, 2003; Fu et al, 2003; Hetzer et al, 2001; Hitchcock et al, 2001; Ramadan et al, 2007). In addition,
roles for Cdc48/p97 as a regulator of lysosomal proteolysis through autophagic and endosomal
pathways have also been described (Bug & Meyer, 2012). However, Cdc48/p97 activity may also
mediate non-proteolytic downstream effects (Hetzer et al, 2001; Latterich et al, 1995; Wilcox &
Laney, 2009). Consistent with its wide-ranging cellular roles, Cdc48/p97 is an essential as well as a
highly abundant protein in eukaryotic cells, proposed to account for up to 1% of total cellular
proteins (Peters et al, 1990). It consists of two AAA ATPase domains (D1 and D2) and a terminal N
domain which self-assembles into a homo-hexameric ring with the N-termini projecting outwards
(DeLaBarre & Brunger, 2003). These serve as major docking sites for a number of regulatory co-
factors. A prominent feature of the Cdc48/p97 macromolecule is its ability to convert chemical
energy derived from ATP hydrolysis into conformational changes of its homo-hexameric barrel
(Rouiller et al, 2002). These structural changes are believed to generate mechanical tension that can
facilitate the disassembly/extraction of protein substrates from their molecular contexts. This so-
called “segregase” activity seems to lie at the core of the various Cdc48/p97 functions by enabling
the mobilization of ubiquitylated proteins, such as from larger protein complexes or lipid
membranes, to allow their downstream proteasomal targeting or potential non-proteolytic fates
(Nakatsukasa et al, 2013; Ramadan et al, 2007; Stolz et al, 2011; Wilcox & Laney, 2009).
Cdc48/p97 ATPase activity may also contribute to the unfolding of proteins prior to their
proteasomeal processing (Beskow et al, 2009; Pye et al, 2006).

25
The Cdc48-Ufd1-Npl4 complex Engagement of Cdc48/p97 in its various cellular activates is facilitated through a variety of
substrate-binding cofactors. Although Cdc48/p97 is seen to weakly bind ubiquitin by itself in vitro,
the assistance of ubiquitin-binding co-factors greatly contribute to substrate targeting in vivo, with
different interaction partners being responsible for guiding Cdc48/p97 to different cellular tasks.
One of its most prevalent substrate-binding adaptors is a heterodimeric complex made up of the
Ufd1 (~40 kDa) and Npl4 (~60 kDa) proteins. While Ufd1 was first isolated in a screen for mutants
incompetent in degrading an artificially ubiquitin-fused protein (giving rise to the name ubiquitin
fusion degradation 1), Npl4 was initially described as a factor required for nuclear import
(DeHoratius & Silver, 1996; Johnson et al, 1995). Ufd1 and Npl4 were later co-purified from rat
liver cytosol and shown to cooperatively associate with Cdc48/p97 in both mammals and yeast
(Meyer et al, 2000; Rape et al, 2001). Although gel filtration experiments has suggested that a
significant fraction of Ufd1-Npl4 sub-complex exists in the cell in a Cdc48/p97 unbound state
(Meyer et al, 2000), another human study indicates that Ufd1-Npl4 is actually predominantly
associated with Cdc48/p97 (Alexandru et al, 2008). Insight into the structural organization of the
Cdc48/p97-Ufd1-Npl4 complex has revealed that both subunits of the Ufd1-Npl4 dimer bind to the
Cdc48/p97 N-terminus with high affinity via bipartite cooperation (Bruderer et al, 2004; Richly et
al, 2005). It has further been seen that only one Ufd1-Npl4 sub-complex is able to bind per
Cdc48/p97 hexamer, suggesting that Cdc48/p97 is able to simultaneously engage in other
interactions (Pye et al, 2007). Binding of Ufd1 to Cdc48/p97 is mediated through a short
hydrophobic stretch (BS1/SHP-box domain) within its C-terminus. Npl4 on the other hand
associates with Cdc48/p97 through an N-terminal portion predicted to form a ubiquitin fold domain
(UBD) (Bruderer et al, 2004; Pye et al, 2007). While all three components of the Cdc48/p97-Ufd1-
Npl4 complex are individually capable of binding ubiquitin chains, binding is synergistically
enhanced upon complex formation (Ye et al, 2003). The ubiquitin binding portion of Ufd1 is
situated at its N-terminus and resembles the ubiquitin-binding domain of Cdc48/p97 and other AAA
ATPases. Although it contains distinct binding sites for both mono- and poly-ubiquitin, it seems to
display a preference for binding poly-ubiquitin (Park et al, 2005). The association of Npl4 (at least
of the mammalian protein) with ubiquitin is mediated through its C-terminal zinc-finger (NZF)
region (Meyer et al, 2002). In contrast to the ubiquitin-binding domains of Cdc48/p97 and Ufd1

26
which bind ubiquitin chains linked through K48, the NZF domain is able to interact with K63-type
ubiquitin chains (Ye et al, 2003).
Figure 4 (Modified from Park et al., 2005) Simplified representation of the Cdc48p97-Ufd1-Npl4 core complex. The
Ufd1-Npl4 sub-complex binds the Cdc48/p97 N-terminal domain in a cooperative manner with only Ufd1-Npl4 dimer
binding per Cdc48/p97 hexamer (Pye et al, 2007).
The Cdc48/p97-Ufd1-Npl4 core complex has been implicated in a myriad of cellular functions. One
of the best understood examples of Cdc48/p97-Ufd1-Npl4 function is in endoplasmic reticulum-
associated degradation (ERAD). Here, Cdc48/p97-Ufd1-Npl4 promotes the translocation of
misfolded ubiquitylated proteins from the ER membrane back into the cytosol for proteasomal
degradation (Ye et al, 2003). Cdc48/p97-Ufd1-Npl4 is similarly seen to enable the mobilization of
some ER membrane-bound transcription factors in a process coupled to their proteolytic processing
(Hitchcock et al, 2001; Rape et al, 2001). More recent studies find Cdc48/p97-Ufd1-Npl4 to
execute key roles at chromatin; again presumably by coupling ubiquitin binding to ATPase driven
protein remodeling (Ramadan et al., 2007; Meyer et al., 2010; Raman et al., 2011; Franz et al.,
2011; Acs et al., 2011). For instance in Xenopus egg extracts, Cdc48/p97-Ufd1-Npl4 is seen to
enable the removal of Aurora B from post-mitotic chromosomes to inhibit its activity and allow
chromosome decondensation and nuclear envelope reformation (Ramadan et al, 2007). In addition,
human Cdc48/p97 has been seen to localize at DNA double strand breaks in a Ufd1-Npl4 and K48-
linked ubiquitin-dependent manner. By promoting the turnover of K48-linked ubiquitin conjugates
at damage sites, Cdc48/p97 contributes to efficient repair progression (Meerang et al, 2011).

27
Cdc48/p97 as a “molecular gearbox” Besides the Ufd1-Npl4 sub-complex, a plethora of other Cdc48/p97 co-factors are known to exist
(Alexandru et al, 2008; Schuberth & Buchberger, 2008). These can be functionally sub-divided into
substrate-recruiting and substrate–processing factors. The latter comprise ubiquitin conjugating and
deconjugating enzymes (Buchberger, 2010; Richly et al, 2005; Uchiyama et al, 2002; Ye, 2006). It
has consequently been suggested that Cdc48/p97 forms a kind of “molecular gearbox” (Jentsch &
Rumpf, 2007) that helps regulate the fate of substrate proteins by integrating different ubiquitination
and deubiquitinating activities. The balance between such activities may determine whether a
substrate is passed on to the proteasome or rather recycled (Richly et al, 2005; Rumpf & Jentsch,
2006). Detailed studies in S. cerevisiae has proposed a model for such ubiquitin substrate regulation
(Richly et al, 2005). In this model, oligo-ubiquitinated substrates are first recognized the
Cdc48/p97-Ufd1-Npl4 complex and then further processed by the ubiquitin “E4” enzyme Ufd2,
which associates with the Cdc48/p97 C-terminus. By extending the oligo-ubiquitin tag, Ufd2 works
to increase the affinity of the ubiquitin chain for the ubiquitin-recognizing parts of the proteasome.
Alternatively, deubiquitinating enzymes, also recruited by Cdc48/p97, may antagonize such chain
elongation and in this way prevent substrate proteolysis.
Cdc48/p97 binding motifs The various Cdc48/p97 co-factors may bind to Cdc48/p97 in more or less mutually exclusive
manners to make up functionally distinct sub-complexes (Alexandru et al, 2008; Meyer et al, 2000;
Rumpf & Jentsch, 2006; Schuberth & Buchberger, 2008). Different modes of Cdc48/p97 binding
have been described (Buchberger, 2010; Yeung et al, 2008). One of the most common Cdc48/p97
binding modules is the ubiquitin regulatory X (UBX) domain or UBX-like domains which binds the
Cdc48/p97 N-terminus (Alexandru et al, 2008; Schuberth et al, 2004). Other co-factors contain the
so-called PUB [PNGase (peptide-N-glycosidase)/ ubiquitin associated] and PUL [PLAP
(phospholipase A2-Activating protein), Ufd3 and Lub1] domains which stand out by binding the
Cdc48/p97 C-terminal unstructured region. While the UBX, PUB and PUL domain are structural
domains, a number of shorter linear motifs are also able to mediate Cdc48/p97 binding. These
include the N-terminal binding BS1/SHP box motif (as present in Ufd1) (Bruderer et al, 2004) and
the VCP-binding (VBM) and VCP-interacting (VIM) motifs (Ballar et al, 2006; Boeddrich et al,
2006). Some proteins contain a combination of more than one Cdc48/p97 binding site (Stapf et al,

28
2011), suggesting some flexibility in their Cdc48/p97 binding mode that may allow them to adapt to
different Cdc48/p97 functions (Yeung et al, 2008).
Proteasomal delivery In addition to substrate-recruiting and –processing co-factors, another set of Cdc48/p97 adaptor
proteins, characterized by possessing both ubiquitin-binding (UBA) and ubiquitin-like (UBL or
UBX) domains are able to mediate the final transport of some Cdc48/p97-bound substrates to the
proteasome (Richly et al, 2005). These include the UBA-UBL domain proteins Rad23 and Dsk2
(Rhp23 and Dhp1 in S. pombe). These factors preferentially bind poly-ubiquitin chains of more than
two ubiquitin moieties and in yeast work together with the chain elongating enzyme Ufd2 to
facilitate substrate proteasomal delivery (Hanzelmann et al, 2012; Richly et al, 2005). Though in
contrast to Cdc48/p97-Ufd1-Npl4 complex factors, neither Ufd2 nor the Rad23 or Dsk2 proteins are
essential for viability in either budding and fission yeast, highlighting the potential diversity and/or
redundancy in Cdc48/p97-Ufd1-Npl4-mediated downstream events (Elsasser & Finley, 2005;
Schiestl & Prakash, 1989)
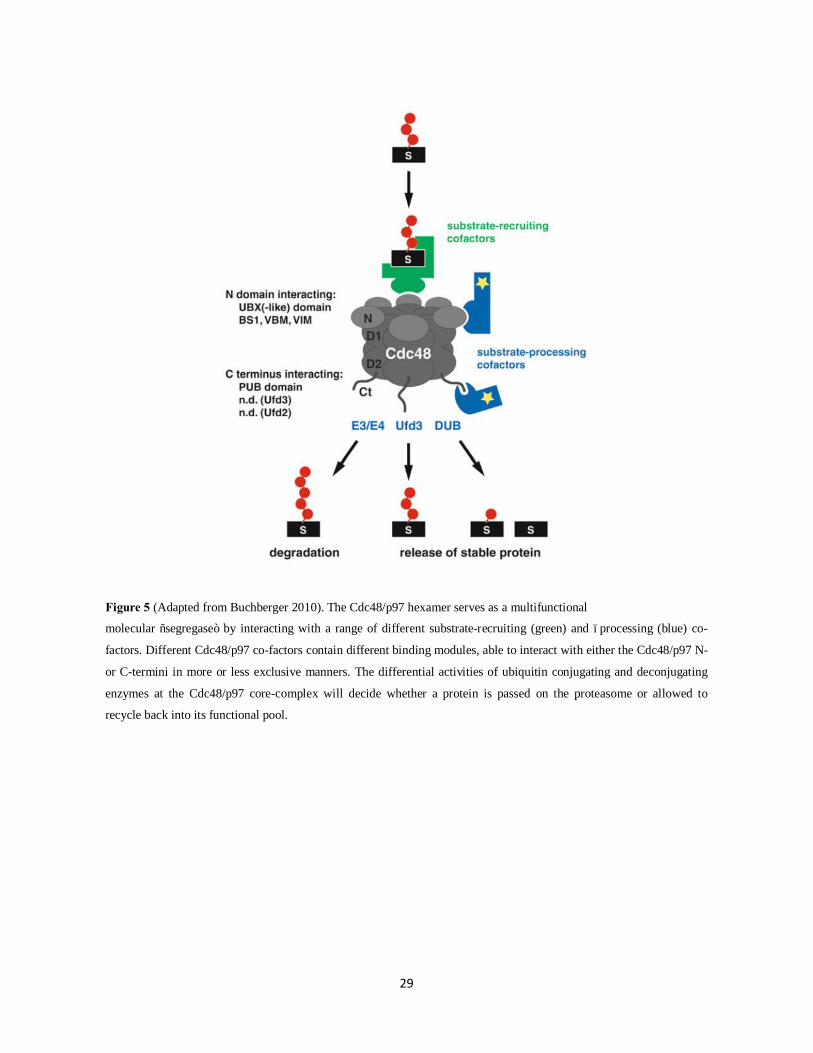
29
Figure 5 (Adapted from Buchberger 2010). The Cdc48/p97 hexamer serves as a multifunctional
molecular “segregase” by interacting with a range of different substrate-recruiting (green) and –processing (blue) co-
factors. Different Cdc48/p97 co-factors contain different binding modules, able to interact with either the Cdc48/p97 N-
or C-termini in more or less exclusive manners. The differential activities of ubiquitin conjugating and deconjugating
enzymes at the Cdc48/p97 core-complex will decide whether a protein is passed on the proteasome or allowed to
recycle back into its functional pool.

30
Crosstalks between the SUMO and ubiquitin systems Although conjugation to SUMO and ubiquitin constitute separate modification pathways, the two
pathways intersect in multiple ways (Denuc & Marfany, 2010; Praefcke et al, 2012; Ulrich, 2005).
Interplays occur both at the level of individual substrate proteins (Desterro et al, 1998) and through
direct regulation of each other’s enzymatic machineries (Buschmann et al, 2000; Pichler et al, 2005;
Watts, 2013). Due to the fact that all ubiquitin-like proteins are conjugated to lysine residues, there
are e.g. cases where the same lysine can be modified by either SUMO or ubiquitin. Such direct
crosstalk on a single lysine has been well established for IκBα, an inhibitor of the NF-κB (nuclear
factor κB) transcriptional activator (Desterro et al, 1998). While sumoylation of IκBα has a
stabilizing effect, modification by ubiquitin stimulates IκBα proteasomal degradation and,
consequently, activation of NF-κB-mediated transcriptional events. Thus, in this specific case,
SUMO and ubiquitin can be viewed to regulate protein function in an antagonistic manner. In
another case, SUMO and ubiquitin operate in a coordinated manner to regulate the activity of the
IκB kinase (IKK) complex through modification of the IKK regulatory subunit NEMO (NF-κB
essential modulator). By stimulating the sequential shuttling of NEMO in and out of the nucleus,
respectively, modification by SUMO or ubiquitin contribute to the formation of a functionally
active kinase within the cytosol in response to genotoxic stress (Huang et al, 2003). As activated
IKK can in turn phosphorylate IκBα to prime its ubiquitylation and thus proteasomal degradation,
the NF-κB signaling pathway represents a case where SUMO and ubiquitin communicate at several
steps to regulate a functional response. SUMO and ubiquitin crosstalks are also common in other
aspects of the DNA damage response (Galanty et al, 2009; Pinder et al, 2013; Ulrich, 2014). For
instance, as will be touched upon later, differential modification of a common lysine on the
proliferating cell nuclear antigen (PCNA) coordinates the repair response to replication stalling
lesions, as well delineated in budding yeast (Hoege et al, 2002). From these and many other
examples, it is clear that numerous cellular pathways involve a combinatorial regulation by both
SUMO and ubiquitin (as well as other PTMs) and that this kind of crosstalk can result in diverse
functional outcomes (Yang, 2005).

31
The SUMO-targeted ubiquitin ligases (STUbLs) The discovery of a class of ubiquitin ligases containing SUMO-interaction motifs (SIMs) has
provided an example for how SUMO and ubiquitin activities can be directly coupled. By combining
SIMs and ubiquitin E3 RING domains, these so-called SUMO-targeted ubiquitin ligases (or
STUbLs) are specifically recruited to sumoylated proteins to stimulate their ubiquitylation
(Heideker et al, 2009; Prudden et al, 2007; Tatham et al, 2008; Uzunova et al, 2007). As this can
target modified proteins for degradation by the proteasome, STUbLs represent a proteolytic
pathway of downregulating SUMO-modified proteins (Lallemand-Breitenbach et al, 2008; Tatham
et al, 2008; Uzunova et al, 2007).
RNF4 STUbL homologs The STUbLs were first defined in fission yeast by the Rfp1/Slx8 and Rfp2/Slx8 dimers (Kosoy et
al, 2007; Prudden et al, 2007; Sun et al, 2007), and in budding yeast by the Slx5/Slx8 dimer (also
known as Hex3/Slx8 or Uls2) and monomeric Uls1 (also known as Ris1) (Mullen et al, 2001;
Uzunova et al, 2007; Xie et al, 2007). Direct biochemical evidence that Uls1 functions as a STUbL
is still lacking (Sriramachandran & Dohmen, 2014), but functional and genetic evidence strongly
suggests that Uls1 is indeed a STUbL similar to Slx5/Slx8 (Uzunova et al, 2007). In S. pombe, Rfp1
and Rfp2 are functionally redundant and thus may be referred to collectively as Rfp (Prudden et al,
2007). The STUbL enzymes all contain C-terminal RING domains and one or several SIMs placed
more N-terminally. While Slx5/Slx8 and Rfp/Slx8 dimerizes through their RING domains, only the
Slx8 subunits possess functional RING E3 activity (Prudden et al, 2007; Xie et al, 2007). Though,
the Slx5 and Rfp subunits are likely to contribute to E3 activity in vivo by presenting interaction
surfaces that help stabilize and hence activate E2~ubiquitin thioesters (Plechanovova et al, 2012).
Moreover, as the Slx5 and Rfp subunits harbor the primary SUMO binding capacity of the
heterodimers, they are thus responsible for SUMO substrate targeting (Prudden et al, 2007;
Uzunova et al, 2007; Xie et al, 2007). The mammalian homolog to the yeast STUbLs is RNF4
which only functions as homodimer (Plechanovova et al, 2012; Tatham et al, 2008).

32
STUbL targeting/substrate recognition
As suggested by their multiple and tandemly placed SIMs, STUbLs are preferentially targeted to
poly-sumoylated species. This was demonstrated through detailed binding studies of RNF4 showing
that the RNF4 tandem of four SIMs binds only poorly to mono-SUMO but with high affinity to
SUMO chains of extended length (Keusekotten et al, 2014; Tatham et al, 2008). A recent study
found that whereas two of the SIMs are necessary and sufficient for interaction with di-SUMO, the
cooperative binding by additional SIMs in each RNF4 monomer mediates an even stronger affinity
for longer SUMO chains. Consistently, various RNF4 SIM mutants are impaired in modifying
SUMO substrates in vivo (Keusekotten et al, 2014). Compared to RNF4, S. pombe STUbL proteins
contain fewer SIMs. As seen in Figure 6, each subunit contains only one or two putative SIMs.
However, upon dimerization these may allow cooperative binding to longer SUMO polymers as is
the case for the four SIMs of RNF4 (Keusekotten et al, 2014). Mechanistic insight into RNF4
function was recently provided by the discovery that RNF4 activation is directly linked to the
availability of its poly-SUMO substrate. While the in vivo concentration of RNF4 is too low to
promote dimerization, binding to poly-SUMO chains helps increase the local concentration of
RNF4 to force dimerization and hence activation (Rojas-Fernandez et al, 2014). Thus, interestingly,
the spatial and temporal activity of STUbLs seems to be directly governed by the growth of SUMO
chains (Keusekotten et al, 2014; Rojas-Fernandez et al, 2014). Even though STUbLs are effectively
recruited to poly-SUMO, it is possible that they may also be able to bind proteins modified by
several mono SUMOs on distinct sites.

33
Figure 6 (Modified from Sriramachandran and Dohmen 2014): Depiction of RNF4 STUbL homologs displaying their
RING and SIM domains. Different SIM consensus sequences are shown as blue, green or purple bars. While RNF4
function as a homodimer, the budding yeast Slx5 and Slx8 and the fission yeast Rfp and Slx8 STUbLs function as
heterodimers. Rfp1 and -2 in S. pombe are functionally redundant. Even though Slx8 fails to interact with SUMO in
two-hybrid systems (Prudden et al, 2007; Uzunova et al, 2007), it is likely that Slx8 contributes to poly-SUMO binding
once in complex with its STUbL partners (Keusekotten et al, 2014). RNF4 contains four predicted SIMs; only the three
SIMs that contribute to SUMO binding are represented here (Keusekotten et al, 2014). Budding yeast Uls1 is a
monomeric STUbL but recent studies suggest that it may in some cases compete with Slx8 for Slx5 dimerization (Tan
et al, 2013). Special about Uls1 is also its Swi2/Snf2-like translocase domain.

34
Figure 7 (Adapted from Rojas Fernandez et al., 2014). Model for RNF4 STUbL activation: The binding of RNF4 N-
terminal SIMs (SUMO-interacting motifs) to polymeric SUMO chains increases the local concentration of the ligase to
force its dimerization (via its C-terminal RING domain) and hence activation.
STUbL functions Consistent with STUbLs targeting poly-sumoylated proteins for degradation by the proteasome,
both STUbL dysfunction and proteasomal inhibition individually lead to the accumulation of high
molecular weight SUMO conjugates (Bailey & O'Hare, 2005; Kosoy et al, 2007; Prudden et al,
2007; Schimmel et al, 2008; Uzunova et al, 2007; Wang et al, 2006). Furthermore, proteasome
inhibition and RNF4 depletion induce the accumulation of proteins that are simultaneously
modified by SUMO and ubiquitin (Schimmel et al, 2008; Uzunova et al, 2007). Co-modifications
are difficult to detect under normal conditions, suggesting that they are usually transient (Bailey &
O'Hare, 2005; Prudden et al, 2007). In vitro studies of STUbLs have found that STUbLs attach
ubiquitin mainly to SUMO chains, but also to some extent to the SUMO-modified protein itself
(Mullen & Brill, 2008; Tatham et al, 2008).
Indicating that STUbL-mediated regulation of sumoylated proteins is fundamental to the cell,
STUbL mutations in yeast are either lethal or cause severe growth defects and genomic instability,
(Heideker et al, 2009; Prudden et al, 2007; Wang et al, 2006; Zhang et al, 2006). Ectopic expression
of human RNF4 complements the yeast mutations, highlighting the evolutionary conservation of the
STUbL pathway (Sun et al, 2007; Uzunova et al, 2007). Specific evidence that the essential
function of STUbLs is through their downregulation of poly-sumoylated proteins comes from

35
studies in S. pombe showing that reducing SUMO conjugation in the cell by either deleting the
major E3 ligase Pli1 or overexpressing the SUMO chain depolymerization enzyme Ulp2 suppresses
the lethality and DNA damage sensitivity of slx8∆ (or slx8-1) and rfp1∆ rfp2∆ mutants, respectively
(Kosoy et al, 2007; Prudden et al, 2007). Similarly, preventing chain formation by mutation of the
major lysine acceptor sites in SUMO has a rescuing effect (Prudden et al, 2011).
It should be mentioned that modifications by the RNF4 STUbL homologs may not exclusively lead
to proteasomal degradation. For instance, RNF4 can catalyze the formation of ubiquitin chains
linked through K11, K48 or K63 in vitro (Tatham et al, 2008). Of these, K48-linked (and K11-
linked) chains function as the major proteolytic signal while K63-polyubiquitin is primarily
involved in other types of cellular signaling (Komander & Rape, 2012; Zhao & Ulrich, 2010).
Moreover, the discovery of proteins containing composite SUMO and ubiquitin recognition motifs
has led to suggest that these may work as effectors of SUMO-ubiquitin hybrid chains, such as
produced by STUbLs, and potentially stimulate non-proteolytic outcomes (Guzzo & Matunis,
2013).
STUbLs are required for genome integrity Similar to SUMO and the SUMO pathway enzymes, STUbLs are primarily enriched in the nucleus
where they contribute to several aspects of genome integrity (Burgess et al, 2007; Cook et al, 2009;
Nagai et al, 2008; Prudden et al, 2007; van de Pasch et al, 2013; Yin et al, 2012). The importance of
STUbLs in genome maintenance pathways was recognized early on, as the budding yeast slx5 and
slx8 STUbL genes were first identified in a screen for genes required in cells lacking the Sgs1
helicase (BLM in humans and Rqh1 in S. pombe) (Mullen et al, 2001). Mutations of the BLM
family helicases cause several phenotypes of genome instability, including hyper-recombination
and chromosome missegregation. As Sgs1 deletion leads to the accumulation of high molecular
weight SUMO conjugates, Slx5/Slx8 are likely required for the efficient turnover of these species
(Mullen & Brill, 2008). STUbL mutants in yeast are generally sensitive to replication stress and
DNA damaging agents, they accumulate spontaneous Rad52 (Rad22 in S. pombe) foci and they
display negative genetic interactions with homologous recombination (HR) repair proteins (Burgess
et al, 2007; Kosoy et al, 2007; Prudden et al, 2007). STUbL mutants also suffer from elevated
recombination rates and increased chromosomal rearrangements (Nagai et al, 2008; Zhang et al,
2006). In S. pombe, STUbLs have been proposed to locally counteract unscheduled recombination
at the programmed replication fork barrier RTS1, through removal of Top1 (topoisomerase 1)-

36
SUMO adducts (Steinacher et al, 2013) and to generally contribute to spontaneous Top1-mediated
DNA damage tolerance (Heideker et al, 2011). In accord with a direct function of STUbLs in the
suppression of genome instability, the STUbLs of budding yeast localize to sites of active
replication (Burgess et al, 2007) and to DNA repair foci (Cook et al, 2009). Moreover, Slx5 and
Slx8 associate with nuclear pore complexes (NPC) where they participate in the repair of collapsed
replication forks that have been moved to the nuclear periphery (Nagai et al, 2008).
Further insight into STUbL function at sites of DNA damage has been obtained for RNF4 in
mammalian cells (Galanty et al, 2012; Luo et al, 2012; Yin et al, 2012). Here, RNF4 is recruited to
DNA double strand breaks (DSBs) in a SUMO and SIM-dependent manner where it facilitates the
removal of DNA repair factors, including MDC1 (mediator of DNA-damage checkpoint 1). MDC1
sumoylation is induced by DNA damage, and sumoylated MDC1 is in turn able to recruit RNF4
(Luo et al, 2012; Yin et al, 2012). Notably, RNF4 depletion impairs the loading of Rad51 onto
single-stranded DNA, consistent with the general DSB repair defects observed in RNF4-depleted
cells (Galanty et al, 2012). The precise molecular mechanisms for the reduced Rad51 loading have
not been clarified but they are likely linked to impaired DSB end resection due to an inability to
effectively recruit the CtIP nuclease complex (Yin et al, 2012). As suggested by the significant
RNF4-dependent increase in ubiquitin adducts at DSB sites, RNF4 may modify several factors
participating in the repair process. Suggesting that at least some of the RNF4 stimulated
ubiquitylation events are coupled to substrate proteolysis, proteasomal proteins accumulate at DSBs
in an RFN4-dependent manner. Sumoylated MDC1 may represent such an RNF4-targeted substrate
whose proteolytic removal facilitates recruitment of downstream repair factors(Galanty et al, 2012)
(Luo et al, 2012). However, RNF4 may also mediate non-proteolytic effects at sites of damage (Yin
et al, 2012). For example, RNF4-catalyzed SUMO-ubiquitin hybrid chains can work as targeting
signals for the BRCA1 associated protein Rap80 (receptor-associated protein 80) which hold
bipartite SUMO and ubiquitin binding domains, in this way enabling BRCA1 recruitment (Guzzo &
Matunis, 2013).
Together these various studies suggest that STUbLs play several important roles during the
response to DNA damage. In particular they seem to directly integrate SUMO- and ubiquitin-
mediated signaling at damage sites to ensure coordinated repair progression. Depending on the
specific context, and on the type of ubiquitin chain formed, such regulation may involve either
proteolytic or non-proteolytic events. Despite the striking consequences of STUbL dysfunction on

37
genome integrity, insight into the specific targets affected by STUbL activity in these pathways is
still lacking. The numerous DNA replication and repair proteins sumoylated in the DNA damage
response (Cremona et al, 2012; Psakhye & Jentsch, 2012; Yin et al, 2012) represent candidate
substrates whose STUbL-mediated turnover may help terminate SUMO-induced repair events.
Mitotic roles for STUbLs There is also increasing evidence that STUbLs fulfill essential roles at centromeres. A recent study
in S. cerevisae revealed that Slx5 is stably associated with centromeres (as seen by chromatin
immunoprecipitation) and that both slx5∆ and slx8∆ mutants display severe mitotic defects
including chromosome missegregation and aberrant spindle dynamics (van de Pasch et al, 2013).
Similar mitotic defects were observed in RNF4-depleted human cells (van de Pasch et al., 2013).
From these observations the authors speculated that mitotic segregation errors may be the source of
some of the increased levels of DNA damage detected in STUbL mutant cells (Burgess et al, 2007;
Prudden et al, 2007). Conversely, it is also possible that damage arising during S-phase remains
unrepaired in mitosis, causing faulty segregation (van de Pasch et al, 2013). However, supporting
that STUbLs do indeed play direct roles at kinetochores, RNF4 targets the kinetochore protein
CENP-I for proteasomal degradation (Mukhopadhyay et al, 2010). The association of CENP-I at
kinetochores is controlled by the balanced activities of RNF4 and the SUMO-chain depolymerizing
protease SENP6 (Ulp2 homolog). While RNF4 causes CENP-I destabilization, SENP6 prevents this
fate by antagonizing CENP-I polysumoylation and thus RNF4 recognition. The balance of these
two activities is believed to help coordinate proper assembly of the inner kinetochore plate
(Mukhopadhyay et al, 2010). A role for STUbLs during mitosis is in line with the many proposed
roles for sumoylation in the regulation of mitotic spindles and chromosome segregation (Bachant et
al, 2002; Meluh & Koshland, 1995; Tanaka et al, 1999; Watts, 2007). In particular, the spatio-
temporal timing of sumoylation and desumoylation events seems critical for normal mitotic
progression (Cubenas-Potts et al, 2013; Wan et al, 2012). By facilitating dynamic turnover of some
of these sumoylated products, STUbLs are likely to contribute to this regulation in the course of
chromosome segregation (van de Pasch et al, 2013).

38
STUbL-mediated regulation of PML bodies The most studied physiological STUbL substrate is the mammalian PML (promyelocytic leukemia)
protein (Lallemand-Breitenbach et al, 2008; Tatham et al, 2008; Weisshaar et al, 2008). This protein
is the major constituent of PML nuclear bodies (NBs) whose assembly depends on the effective
sumoylation of its components (Seeler & Dejean, 2001). Within the cell PML form discrete foci of
which at least some are closely associated with chromatin. Besides PML these bodies are enriched
in a variety of proteins including factors involved in transcription, apoptosis and DNA repair.
Consequently, PML NBs have been implicated in the regulation of diverse cellular processes
(Lallemand-Breitenbach & de The, 2010; Seeler et al, 1998). RNF4 localizes to PML bodies and it
stimulates the ubiquitin-dependent turnover of PML as well as the oncogenic PML fusion protein–
PML-RARα (retinoic acid receptor α) in a SUMO-dependent manner (Geoffroy et al, 2010). As
arsenic trioxide induces the polysumoylation of PML (and PML-RARα), this drug can be
effectively used as a treatment for acute promyelocytic leukemia (APL) by causing the
destabilization of the toxic PML-RARα fusion protein (Lallemand-Breitenbach et al, 2001). Given
that many other PML NB components are also modified by SUMO (Seeler & Dejean, 2001), it is
possible that these are similarly regulated by RNF4 activity.
Genetic relationship between STUbLs and Ulp2 It has been observed that the synthetic lethality of the slx5∆ sgs1∆ and slx8∆ sgs1∆ budding yeast
double mutants can be rescued by deleting the major chain-editing desumoylase Ulp2 and thus by
stabilizing SUMO chains (Mullen et al, 2011). Similarly, ulp2∆ in S. pombe is supposed to rescue
the lethality of slx8∆ (N. Boddy, personal communication). This mutual rescue of STUbL and Ulp2
mutations is counter-intuitive to the idea that the species causing toxicity in both mutant
backgrounds are accumulated high molecular weight poly-SUMO conjugates (Bylebyl et al, 2003)
(Prudden et al, 2007). As an explanation to this paradox it has been suggested that in the absence of
both STUbL and Ulp2 activities, sumoylated species are allowed to be eliminated through yet an
alternative pathway requiring extended SUMO chains normally antagonized in the single mutants
(Mullen et al, 2011). Consistent with this model, Ulp2 overexpression is inhibitory to growth of S.
cerevisae slx5∆ cells. Though in S. pombe, Ulp2 overexpression is reported to suppress at least
some rfp1∆ rfp2∆ STUbL mutant phenotypes (Kosoy et al, 2007). Whether these two studies of
Ulp2 overexpression reflects true differences between the two yeast species or rather experimental

39
differences would be interesting to clarify. Nevertheless, it seems, at least for S. pombe, that both an
excess and a deficiency of Ulp2 is able to rescue STUbL mutant phenotypes; the former by
antagonizing polysumoylation altogether and the latter by stabilizing SUMO chains. The exact
mechanism governing the tolerance to such stabilized SUMO chains remains to be clarified. Yet a
screen for genes required for slx5∆ sgs1∆ ulp2∆ S. cerevisae viability has suggested that such a
tolerance pathway involves the less-well characterized SUMO protease Wss1 (Mullen et al, 2010;
Mullen et al, 2011).
Other types of STUbLs Apart from the RNF4 STUbL homologs, other ubiquitin ligases have also been reported to contain
SIMs and to be recruited to their substrates in a SUMO-SIM stimulated manner (Sriramachandran
& Dohmen, 2014). One example is Rad18 in budding yeast which is recruited to sumoylated
PCNA via a single SIM to stimulate PCNA mono-ubiquitylation during postreplicative repair
(PRR). In this case though, Rad18 is recruited to its substrate not only through its SUMO-SIM
interaction but also through interactions with PCNA itself (Parker & Ulrich, 2012). Identification of
the SUMO-SIM interaction between PCNA and its ubiquitin ligase revealed a direct mechanism for
how the switch from PCNA sumoylation to damage-induced ubiquitylation occurs. This discovery,
as well as other examples (Erker et al, 2013; Poulsen et al, 2013), indicate that the concept of
STUbLs is more general than originally perceived, and that SUMO-SIM mediated targeting may be
a feature employed by several types of ubiquitin E3s to conduct different kinds of cellular functions.

40
The DNA damage response The genomes of cells are continuously challenged by both endogenous and exogenous source of
DNA damage. If not properly dealt with, such genomic stress can lead to cell death or to the
development of cellular abnormalities such as cancer. To overcome these threats cells have evolved
a variety of tolerance mechanisms, including DNA damage checkpoints and repair pathways, which
act in an integrated manner to ensure the maintenance of genome stability through cellular
generations (Harper & Elledge, 2007; Jackson & Bartek, 2009).
The DNA-damage checkpoints A primary response to genotoxic stress is the activation of complex signaling pathways, termed
DNA damage checkpoints, which serve to coordinate DNA repair with progression through the cell
cycle (Latif et al, 2001). Key initiators of these pathways are the PI3-related protein kinases ATM
(ataxia-telangiectasia-mutated) and ATR (ATM and Rad3-related) kinases (Tel1 and Rad3 in S.
pombe, respectively)(Savitsky et al, 1995) (Seaton et al, 1992). Though, before these kinases can be
activated, the damage must first be detected. A major DNA damage “sensor” is the RPA protein
complex which binds to exposed single-stranded DNA (ssDNA) resulting from genomic insult
(Iftode et al, 1999; Seroussi & Lavi, 1993) (Rouse & Jackson, 2002). RPA-coated ssDNA in turn
recruits several complexes of checkpoint proteins. These include ATR via its regulatory partner
ATRIP (Zou & Elledge, 2003), and the PCNA-related Rad9-Rad1-Hus1 (9-1-1) clamp complex
(Parrilla-Castellar et al, 2004). In the case of a DNA double-strand break (DSB), no ssDNA will
initially be exposed. Instead DSBs are first recognized by the Mre11-Rad50-Nbs1 (MRN) complex.
MRN interacts with ATM to stimulate ATM mediated checkpoint signaling (Nakada et al, 2003).
The nuclease Mre11 component of MRN is further responsible for initiating 5’-3’ resection of the
DSB which together with other nucleases such as CtIP (Sae2 in S. cerevisae and Ctp1 in S. pombe)
generate large regions of ssDNA that can attract RPA and eventually ATR. Recruitment of the
ATM and ATR kinases results in signaling cascades that ultimately leads to cell-cycle arrest and
regulation of DNA repair (Gobbini et al, 2013). Among the ATR and ATM substrates are cell-
cycle-specific effectors such as the Chk1 (Cds1 in S. pombe) and Chk2 (Chk1 in S. pombe) kinases,
which are required for mediating S-phase and G2/M checkpoint signaling, respectively (Boddy et
al, 1998; Limbo et al, 2011).

41
Postreplicative repair (PRR) Postreplicative repair (PRR) is a collective term for pathways operating to tolerate DNA damage
encountered during DNA replication. By allowing cells to overcome replication-stalling lesions,
these mechanisms contribute greatly to the resistance of single-stranded damage accumulating into
S-phase. PRR can essentially be sub-divided into two overall branches; an error-prone branch
involving replication through the lesion by translesion synthesis (TLS) and an error-free branch
which is thought to involve a switch in template to the undamaged newly synthesized sister strand.
Both branches are controlled by genes of the Rad6 epistasis group, of which many encode ubiquitin
conjugating or ligating enzymes. These include the Rad6 (Rhp6 in S. pombe) and Ubc13/Mms2
ubiquitin E2s and the DNA-binding Rad18 and Rad5 (Rhp18 and Rad8 in S. pombe) ubiquitin
RING-type E3s (Broomfield et al, 1998; Brown et al, 2002; Prakash, 1981; Ulrich & Jentsch, 2000;
Xiao et al, 2000). By stimulating different kinds of ubiquitin modifications on the proliferating-cell
nuclear antigen (PCNA) these enzymes guide differential PRR responses (Frampton et al, 2006;
Hoege et al, 2002). PCNA forms a homotrimeric ring-shaped structure which functions as a
processivity clamp for replicative DNA polymerases and platform for various factors involved in
DNA replication and repair. PCNA mono-ubiquitination on the highly conserved lysine K164 as
catalyzed by the Rad6-Rad18 complex promotes the TLS pathway by recruiting low-fidelity TLS
polymerases such as Polη, Polκ, Polζ or Polι (Stelter & Ulrich, 2003; Ulrich, 2009). By containing
intrinsic ubiquitin-binding domains, the TLS polymerases are specifically recruited to mono-
ubiquitylated PCNA (Kannouche et al, 2004; Watanabe et al, 2004). Replication through a damaged
area occurs as the TLS polymerases are able to accommodate bulky DNA lesions in their active
site. Though, this happens at the cost of accuracy, making the TLS pathway a potential mutagenic
process. By contrast, extension of the mono-ubiquitin tag on PCNA into K63-linked poly-ubiquitin
chain guides a process of error-free repair. This is stimulated by the Ubc13/Mms2 and Rad5
enzymes which are recruited to stalled forks through interactions between Rad5 and Rad18 (Hoege
et al, 2002). The specific molecular outcomes of PCNA poly-ubiquitylation are less well defined
but the poly-ubiquitin signal is basically thought to displace the TLS polymerases and instead
initiate damage bypass through a pathway involving template switching. The exact mechanism of
template switching is also not clear but two overall models have been suggested (Branzei, 2011). In
one model, helicase-driven fork reversal promotes annealing of the two newly synthesized strands
as well as re-annealing of the parental strands to provide a lesion-free template for damage repair

42
and fork progression (Blastyak et al, 2007). Alternatively, template switching may involve a
recombination-based strand exchange between the damaged strand and the newly synthesized sister
chromatid in a process dependent on the homologous recombination protein Rad51 (Branzei et al,
2008; Liberi et al, 2005; Petermann et al, 2010). In contrast to the fork reversal model which is
restricted to occur at the fork, such recombination-based repair could in theory happen either at or
behind the fork. Indeed studies suggest that fork the PRR tolerance mechanism largely operates on
gaps behind restarted replication forks (Karras & Jentsch, 2010). Of note, although the
postreplicative repair proteins seem to be highly conserved, there might exists some mechanistic
differences between different organisms in how the PRR response is regulated. For instance,
whereas DNA damage is seen to induce both mono- and poly-ubiquitylation on PCNA in both
budding and fission yeast, PCNA has so far only been detected in its mono-ubiquitylated form in
humans (Frampton et al, 2006; Ulrich, 2009).
Regulation of PRR by sumoylation Studies from S. cerevisae have revealed mechanistic insights into the control of PRR pathways by
sumoylation (Branzei et al, 2006; Branzei et al, 2008; Hoege et al, 2002; Papouli et al, 2005; Watts,
2006). In particular, the same site on PCNA that is ubiquitylated (K164) in response to replication
stress can also be sumoylated, (in addition to another site, K127) by the Siz1 ligase. PCNA
sumoylation is observed constitutively during S-phase. Sumoylated PCNA is in turn able to recruit
the Srs2 helicase through a SUMO-SIM interaction, which by disrupting Rad51-nucleofilament
formation prevents inappropriate recombination to occur at transiently stalled forks (Papouli et al,
2005; Pfander et al, 2005). In response to DNA damage, PCNA sumoylation is furthermore
proposed to channel repair through a Rad18-dependent recombination-based error-free repair
pathway (Branzei et al, 2008). Even though PCNA SUMOylation has not been detected either in
humans or in fission yeast (Frampton et al, 2006), genetic data suggests that Pli1–stimulated
sumoylation of an unknown factor(s) also promotes damage bypass through a Rhp18-dependent
error-free mode in S. pombe cells in a process involving checkpoint-mediated phosphorylation of
the checkpoint clamp protein Rad9 (at Thr225) (Kai et al, 2007). More recently, budding yeast
Rad18 was shown to possess a preference for binding sumoylated PCNA via an internal SIM motif,
providing a mechanism for how sumoylation of PCNA can help guide a Rad18-dependent PRR
response (Parker & Ulrich, 2012).

43
Figure 8 (Adapted from Watts 2006). An overview of the different repair outcomes in response to replication stalling
lesions in S. cerevisae. (A) PCNA mono-ubiquitylation stimulates TLS through recruitment of error-prone polymerases.
Extension of the ubiquitin tag on PCNA promotes damage bypass by template switching, either by fork regression (B)
or by a recombination-dependent strand invasion process (C) to allow error-free repair. Rad51-dependent recombination
events at undamaged replication forks are inhibited by PCNA sumoylation and Srs2 recruitment.
DNA double strand break (DSB) repair DNA double strand breaks (DSBs) represents potential lethal DNA lesions as they may lead to
chromosomal rearrangements or loss of genetic material. In addition to being induced by external
sources such as chemicals and ionizing radiation, DSBs may arise during DNA replication as forks
encounter a damaged template (Haber, 1999; Raji & Hartsuiker, 2006). On the other hand, besides
being potentially toxic species, DSBs induced by the cell during meiosis promotes genetic
recombination and thus genetic diversity. Two overall modes of DSB repair employed by cells
include non-homologous end-joining (NHEJ) and repair by homologous recombination (HR)
(Kakarougkas & Jeggo, 2014). NHEJ, where the two free DNA ends are basically ligated back

44
together, may be error-prone as due to loss of base-pairs at the break site. By contrast HR is
generally considered as more error-free strategy of DSB repair. The HR process involves the
invading of a region of homologous sequence (either in a sister chromatid or in a homologous
chromosome) to be used as a template for copying the sequence disrupted by the DNA lesion (Raji
& Hartsuiker, 2006; San Filippo et al, 2008). In addition to repairing DSBs, HR may also be used
to repair single-stranded breaks (SSBs) and to allow restart stalled and broken replication forks, as
mentioned above. In yeast, DSB repair is preferentially repaired by HR which is mediated by
proteins of the Rad52 (Rad22 in S. pombe) epistasis group (Doe et al, 2004; Krogh & Symington,
2004).
Homologous recombination (HR) repair The HR process is initiated by resection of the 5’ DNA ends at both sides of the break, generating
3’ single-stranded (ss) overhangs that can be coated by RPA. DNA resection is stimulated by the
exonucleolytic activity of the MRN complex component Mre11 (Rad32 in S. pombe) (as well as by
other exonucleases with 5’to 3’activity) (Li & Heyer, 2008). RPA will eventually be dislodged from
the ssDNA by the RecA homolog, Rad51 (Rhp51 in S. pombe), to form a nucleoprotein filament
capable of mediating strand invasion (Candelli et al, 2013; Liu et al, 2011; Muris et al, 1993).
Though, the loading of Rad51 needs often assistance by special mediator proteins to allow efficient
RPA displacement. In yeast, these mediators include Rad52 (Rad22 in S. pombe), which seems
required for most, if not all, recombination events in the cell (Benson et al, 1998; Doe et al, 2004;
Krogh & Symington, 2004). Additional Rad51-mediators include Rad54, Rad55 and Rad57 (Krogh
& Symington, 2004). Establishment of a proper Rad51-DNA filament allows it to invade a region
of homologous DNA. By displacing the non-complementary strand in the target DNA molecular it
forms a so-called D-loop. The invading 3’ end can then prime DNA synthesis, using the undamaged
homologous strand as a template to replicate the missing sequence (Li & Heyer, 2008). While the
D-loop may subsequently simply be displaced in a process known as synthesis-dependent strand
annealing (SDSA), the junction between the two crossing strands of the recombining DNA
molecules may also mature into a four-way structure called a Holliday junction (HJ). The invading
strand may furthermore pair with its complementary strand at the other end of the break (second end
capture) to form a double HJ. The differential resolution of these structures leads to different
genetic outcomes by either generating crossover or non-crossover products (Krogh & Symington,
2004). The processing of these different HR structures is catalyzed by specialized nucleases and

45
helicases, able to cleave or unwind DNA, respectively. Such enzymes include the Mus81-Eme1
structure-specific endonuclease and the RecQ helicase homologs (Rqh1 in S. pombe, Sgs1 in S.
cerevisiae, BLM, WRN, RECQ1,-4 and -5 in humans) (Ashton & Hickson, 2010; Boddy et al,
2001; Doe et al, 2004; Osman & Whitby, 2007).

46
Project Aim Proteins interacting with the HP1 homolog Swi6 were sought through two-hybrid screens to
improve our understanding of heterochromatin dynamics in the fission yeast Schizosaccharomyces
pombe (G. Thon, unpublished). The SUMO E3 ligase Pli1 and the STUbL subunit Rfp1, repeatedly
identified in these screens, were in turn used for further screens that led to the identification of the
Ufd1 protein (or its C-terminal half encoding the last 199 amino acids) as an interactor of both Pli1
and Rfp1. STUbLs are essential to the SUMO pathway homeostasis by targeting sumoylated
proteins for proteasomal degradation (Kosoy et al, 2007; Prudden et al, 2007; Sun et al, 2007;
Uzunova et al, 2007). However, little was known about the relevant STUbL targets and the
molecular pathway(s) linking protein sumoylation, ubiquitylation and proteolytic targeting. Given
that Ufd1 (together with Npl4) make up a well-defined ubiquitin-binding co-factor of the Cdc48
chaperone, we were encouraged to further investigate a potential involvement of the Cdc48-Ufd1-
Npl4 complex in SUMO and STUbL substrate processing. Our findings are presented in the
following manuscript “Concerted Action of the Ubiquitin-Fusion Degradation Protein 1 (Ufd1) and
SUMO Targeted Ubiquitin Ligases (STUbLs) in the DNA-Damage Response “. In parallel to our
studies were reports from other laboratories proposing roles for Cdc48-Ufd1-Npl4 in the SUMO
and/or STUbL pathway (Bergink et al, 2013; Nie et al, 2012). These findings are discussed in the
context of ours.
Another part of my project, presented in the second manuscript “Quantitative identification of
sumoylation sites in fission yeast wild-type cells, Ufd1 and SUMO-targeted ubiquitin ligase
(STUbL) mutants”, has been to use a global and unbiased approach to investigate the sumoylated
proteome of fission yeast. More specifically, we have used a novel strategy to enrich for total
sumoylated species from complex protein mixtures which allow site-specific determination of
SUMO substrates by mass spectrometry (Tammsalu et al, 2014). In combination with quantitative
proteomics this method has furthermore allowed insight into the sumoylated proteins potentially
regulated through STUbL and/or Ufd1 activities.

Concerted Action of the Ubiquitin-Fusion DegradationProtein 1 (Ufd1) and Sumo-Targeted Ubiquitin Ligases(STUbLs) in the DNA-Damage ResponseJulie Bonne Køhler, Maria Louise Mønster Jørgensen, Gabriele Beinoraité, Michael Thorsen, GenevièveThon*
Department of Biology, University of Copenhagen, Copenhagen, Denmark
Abstract
In eukaryotes many players in the DNA-damage response (DDR) catalyze protein sumoylation or ubiquitylation.Emphasis has been placed on how these modifications orchestrate the sequential recruitment of repair factors tosites of DNA damage or stalled replication forks. Here, we shed light on a pathway in which sumoylated factors areeliminated through the coupled action of Sumo-targeted ubiquitin ligases (STUbLs) and the ubiquitin-fusiondegradation protein 1 (Ufd1). Ufd1 is a subunit of the Cdc48-Ufd1-Npl4 complex implicated in the sorting ofubiquitylated substrates for degradation by the proteasome. We find that in fission yeast, Ufd1 interacts physicallyand functionally with the Sumo-targeted ubiquitin ligase (STUbL) Rfp1, homologous to human RNF4, and with theSumo E3 ligase Pli1, homologous to human PIAS1. Deleting a C-terminal domain of Ufd1 that mediates theinteraction of Ufd1 with Rfp1, Pli1, and Sumo (ufd1ΔCt213-342) lead to an accumulation of high-molecular-weight Sumoconjugates and caused severe genomic instabilities. The spectrum of sensitivity of ufd1ΔCt213-342 cells to genotoxins,the epistatic relationships of ufd1ΔCt213-342 with mutations in DNA repair factors, and the localization of the repairfactor Rad22 in ufd1ΔCt213-342 cells point to ufd1ΔCt213-342 cells accumulating aberrant structures during replication thatrequire homologous recombination (HR) for their repair. We present evidence that HR is however often notsuccessful in ufd1ΔCt213-342 cells and we identify Rad22 as one of the high-molecular-weight conjugates accumulatingin the ufd1ΔCt213-342 mutant consistent with Rad22 being a STUbL/Ufd1 substrate. Suggesting a direct role of Ufd1 inthe processing of Sumo-conjugates, Ufd1 formed nuclear foci colocalizing with Sumo during the DDR, and Sumo-conjugates accumulated in foci in the ufd1ΔCt213-342 mutant. Broader functional relationships between Ufd1 andSTUbLs conceivably affect numerous cellular processes beyond the DDR.
Citation: Køhler JB, Jørgensen MLM, Beinoraité G, Thorsen M, Thon G (2013) Concerted Action of the Ubiquitin-Fusion Degradation Protein 1 (Ufd1) andSumo-Targeted Ubiquitin Ligases (STUbLs) in the DNA-Damage Response. PLoS ONE 8(11): e80442. doi:10.1371/journal.pone.0080442
Editor: Anja-Katrin Bielinsky, University of Minnesota, United States of America
Received June 22, 2013; Accepted October 2, 2013; Published November 12, 2013
Copyright: © 2013 Køhler et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permitsunrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: The authors' research was supported by the Danish Research Council (http://fivu.dk/en/; grant 09-064284 to GT), the Novo Nordisk Foundation(http://www.novonordiskfonden.dk/en) and the University of Copenhagen Center of Excellence MolPhysX (http://www.ku.dk/). The funders had no role instudy design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing interests: The authors have declared that no competing interests exist.
* E-mail: [email protected]
Introduction
The small modifiers SUMO and ubiquitin are effectors ofmany regulatory changes occurring in eukaryotic cells. Inreactions catalyzed by E1, E2 and E3 enzymes that act in acascade, SUMO or ubiquitin can each be conjugated to lysineresidues of target proteins. Conjugation often results in achange in the interaction properties of the target. In contrast tothe ubiquitin pathway where substrate selection is mediated bylarge sets of E2 and E3 enzymes, sumoylation appearsrestricted to the use of very few E2 and E3 enzymes in allorganisms examined to date [1-3]. In fission yeast the onlyknown SUMO E3 ligases are Nse2 and the PIAS familymember Pli1. Nse2 and Pli1 are both of the SP-RING type,
functioning together with a single E2 enzyme, Ubc9 (also calledHus5; [4-6]).
Among the many processes affected by sumoylation orubiquitylation a special focus has been on the roles played bythese modifications in DNA replication and repair. Many repairenzymes catalyze ubiquitylation or sumoylation and manyfactors acting at stalled replication forks or other DNA lesionsare conditionally ubiquitylated or sumoylated [5,7-20]. Theirmodification can affect downstream repair events or repairpathway choices. The molecular mechanisms through whichthe modifications operate are known in a few cases. One of thebest-understood examples is modification of the proliferatingcell nuclear antigen (PCNA) sliding-clamp by either mono- orpoly-ubiquitin which signals distinct bypass strategies in the
PLOS ONE | www.plosone.org 1 November 2013 | Volume 8 | Issue 11 | e80442

face of replication-stalling lesions [21-23]. Another extensivelystudied, albeit less understood, event is sumoylation of the HRfactor Rad52. S. cerevisiae Rad52 and its fission yeasthomolog Rad22 are established sumoylation targets [7,8].Sumoylation of Rad52 has been proposed to influence theefficiency with which HR proceeds by altering protein stabilityand by reducing the affinity of Rad52 for DNA [8,24]. In yetanother example, in mammalian cells, the RNF8 E3 ubiquitinligase mediates DSB repair. An RNF8-initiated ubiquitylationcascade centered on lesion-flanking histones H2A and H2AXdrives the sequential recruitment of repair factors in a mannerpartly dependent on concurrent sumoylation events[18,12,13,25]. Much emphasis is currently being placed on howcross talks between the ubiquitylation and sumoylationpathways affect repair factors and DNA transactions.
Providing mechanistic insight for how sumoylation andubiquitylation events might be coupled, some ubiquitin E3ligases of the RING family can recognize sumoylated proteinsvia SUMO-interacting motifs (SIMs). These SUMO-targetedubiquitin ligases or STUbLs were defined by the Rfp1/Slx8 andRfp2/Slx8 dimers in S. pombe, the Slx5/Slx8 dimer in S.cerevisiae and human RNF4 [26-30]. Modification by STUbLscan lead to substrate degradation by the proteasome [30], andit might also mediate non-proteolytic functions. The impact ofSTUbLs on cellular SUMO homeostasis is revealed byincreased sumoylation levels and accumulation of poly-SUMOchains in STUbL mutants [26-29,31]. The imbalance in SUMOdynamics in these mutants is associated with genomicinstabilities and slow-growth phenotypes. Consistent with directroles in genome maintenance, both human and S. cerevisaeSTUbLs have been seen at DSBs or sites of active replication[9,32-35]. Other classes of ubiquitin ligases might also functionas STUbLs. This was shown in a recent study for the S.cerevisiae Rad18 ubiquitin ligase [36].
The molecular recognitions and sequence of events linkingsumoylation, ubiquitylation and substrate degradation, thesubstrate specificities and interaction partners of STUbLsremain largely unknown. We attempted to shed light on someof these issues by searching for proteins interacting with thefission yeast SUMO E3 ligase Pli1 or the STUbL Rfp1. Ufd1was identified in both searches. Ufd1 makes up together withNpl4 a major substrate-recruiting co-factor of the homo-hexameric Cdc48 AAA+ ATPase (p97 in mammals) [37], whichis implicated in various ubiquitin-dependent processes in thecell [38,39]. We found that a mutant with a C-terminaltruncation of Ufd1 that removes the interaction domain with Pli1and Rfp1 (ufd1ΔCt213-342) displayed many phenotypicsimilarities with STUbL mutants. Like STUbL mutants,ufd1ΔCt213-342 cells accumulated sumoylated proteins, theywere hypersensitive to DNA-damaging agents and theydisplayed frequent spontaneous Rad22 foci. Also, the epistaticrelationship of ufd1ΔCt213-342 to mutations in HR factors was thesame as for STUbL mutants. Combined with localizationstudies of Ufd1 and SUMO in cells subjected to genotoxicstress, these phenotypes lead us to refine a proposed functionfor Ufd1 in genome maintenance and in the STUbL pathway.
Results
Identification of physical and functional interactionsbetween Pli1, Rfp1, and Ufd1
We conducted large-scale two-hybrid screens usingrespectively the fission yeast SUMO E3 ligase Pli1 and theSTUbL protein Rfp1 as baits to identify factors acting in concertwith these proteins (Figure 1). An interaction between Pli1 andRfp1 and interactions of the two proteins with sumoylationfactors (SUMO and Ubc9) were observed in the screens,confirming previous reports ([26,28]; Figure 1A). Yet-undescribed interactions were also revealed by both screens.In particular, both Pli1 and Rfp1 interacted with a C-terminaldomain of Ufd1 (Figure 1A). The interactions were confirmed inin vitro GST pull-downs (Figure 1B), indicating the proteinsmake direct contact with each other.
Many reports document the role of Ufd1 in endoplasmicreticulum associated degradation (ERAD) where Ufd1 functionsin complex with the Cdc48/p97 ATPase and Npl4 to sort andextract proteins destined for degradation [40,41]. Other studieshave suggested broader roles for the Cdc48-Ufd1-Npl4complex in ubiquitin-dependent processes [42,43,38,39], with arecent focus on events occurring in chromatin [44-54]. Here,the ability of Ufd1 to interact with both Pli1 and Rfp1 suggestedthat Ufd1 might facilitate the degradation of STUbL substrates.Further suggesting a direct physical relationship between theSTUbL pathway and the Cdc48-Ufd1-Npl4 complex, Slx8 (thecatalytic subunit of the Slx8/Rfp1 STUbL dimer) co-purified withCdc48 in immunoprecipitations of Cdc48 from S. pombe cellextracts (Figure 1D). Together these observations motivated afurther investigation of a possible functional relationshipbetween Cdc48-Ufd1-Npl4 and STUbLs. We created a mutantlacking the C-terminal domain of Ufd1 (ufd1ΔCt213-342 encodingUfd1 with a deletion of aa 213-342) which includes the partresponsible for the two-hybrid interactions of Ufd1 with Pli1 andRfp1. According to structural data, Ufd1 binds ubiquitin via anN-terminal domain while the C-terminal portion of the proteininteracts with Cdc48 and Npl4 [40,55-57]. Some aspects ofCdc48/Ufd1/Npl4 complex formation might be affected inufd1ΔCt213-342 cells but remaining interactions bridged byubiquitin or other shared interaction partners possibly accountfor the fact that the ufd1ΔCt213-342 mutant is viable in contrast tothe full ufd1 deletion [58].
As can be seen in Figure 1E, the ufd1ΔCt213-342 mutantaccumulated sumoylated species that migrated slowly indenaturing polyacrylamide gels. This is similar to what isobserved when STUbL function is compromised ([26-28];Figure 1E). The slowly-migrating species might be high-molecular weight, poly-modified proteins, or they might containbranched molecules whose migration would be retarded ingels. The elevated level of SUMO conjugates in ufd1ΔCt213-342
cells suggests that Ufd1 normally participates in the down-regulation of these conjugates.
SUMO accumulates in nuclear foci when Ufd1 functionis impaired
We set up to determine the intracellular localization of theSUMO conjugates that accumulate in ufd1ΔCt213-342 cells. In S.
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 2 November 2013 | Volume 8 | Issue 11 | e80442

Figure 1. Physical and functional interactions between S. pombe Ufd1/Cdc48, the STUbL Rfp1/Slx8 and the SUMO E3ligase Pli1. (A) Yeast two-hybrid interactions. 10-fold dilution series of S. cerevisae strain PJ69-4A expressing a C-terminal domainof Ufd1 (aa 248-342) fused to the Gal4 DNA activation domain (GAD) together with either Pli1 or Rfp1 fused to the Gal4 DNA-binding domain (GBD) were spotted on the indicated media. Protein interactions result in activation of the ADE2 reporter gene in thetester strain and growth on SC-leu-trp-ade. -: empty vectors. (B) Ufd1Ct interacts with Pli1 and Rfp1 in vitro. GST pull-downexperiment with GST, GST- Pli1 or GST-Rfp1 incubated with in vitro-translated, 35S-labeled Ufd1 (aa 110-342). (C) Summary of two-hybrid interactions. Red arrows indicate novel interactions identified in this study; black arrows, previously reported interactions thatwere (solid arrows) or not (dashed arrows) also identified in our screens. (D) Co-purification of Cdc48 and Slx8. Cdc48-GFP waspurified on a GFP affinity matrix from cells expressing Myc-tagged Slx8 or an untagged control. Cdc48-GFP and Slx8-Myc weredetected by Western blotting with respectively GFP and Myc antibodies. (E) Accumulation of sumoylated proteins in ufd1ΔCt213-342
cells. Whole-cell extracts of wild type and indicated mutants were probed with an anti-SUMO antibody. The slx8-1 and mts3-1mutants were shifted from 30°C to 37°C for 4 hr prior to harvesting; all other strains were propagated at 30°C. The strains were,from left to right, JK8; JK9; JK10; JK11; NBY1008; PI131; Δsph2.doi: 10.1371/journal.pone.0080442.g001
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 3 November 2013 | Volume 8 | Issue 11 | e80442

pombe, SUMO detected by immunofluorescence or GFP-tagging produces a somewhat diffuse nuclear signal,punctuated by more intense nuclear foci [59]. The most intenseSUMO focus colocalizes with the clustered centromeres at thespindle-pole body [59]. In our fluorescence microscopy images,GFP-SUMO foci appeared strikingly brighter in theufd1ΔCt213-342 mutant than in wild-type cells imaged in parallelfor comparison (Figure 2A). Figure 2B plots the voxels of
highest intensity in ufd1ΔCt213-342 and wild-type nuclei. Despitea certain degree of variation between cells, higher intensitieswere consistently measured in the ufd1ΔCt213-342 mutantconfirming simple visual inspection. Thus, the high molecular-weight SUMO-containing species detected by Western blottingin ufd1ΔCt213-342 cells appear to accumulate preferentially atspecific subnuclear loci.
Figure 2. Increased SUMO foci intensity in ufd1ΔCt213-342 cells. (A) Fluorescence imaging of GFP-SUMO in wild-type andufd1ΔCt213-342 cells propagated in the presence or absence of Zeocin (350 µg/ml for 2 1/2 hr). (B) Quantification of GFP-SUMO fociintensity. The intensity of the brightest GFP voxel was measured in > 140 nuclei for each of the strains and conditions shown in (A).Bar graphs in the upper panel display the mean intensities and standard deviations measured in the experiment; histograms in thetwo lower panels show the distribution of GFP-SUMO intensities. Statistically-significant differences (p ˂ 0.0001) betweenufd1ΔCt213-342 cells and wild type, and between ufd1ΔCt213-342 cells before and after Zeocin treatment were observed in thisexperiment and for two replicates of the experiment (not shown).doi: 10.1371/journal.pone.0080442.g002
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 4 November 2013 | Volume 8 | Issue 11 | e80442

Colocalization of SUMO and Ufd1 during the DNA-damage response
Cdc48 and Ufd1 are both predominantly nuclear in fissionyeast ([60,61]; Figure 3A). We observed that a Ufd1-YFPfusion protein formed nuclear foci at low frequency in a wild-type background. The DSB-inducing drug Zeocin increased thefrequency of Ufd1-YFP foci so that approximately 30% of cellsdisplayed at least one focus after 3-4 hrs of exposure to Zeocin(Figure 3B). In the vast majority of cases, a single focus wasdetected. This focus was at the nuclear periphery andcolocalized with the most intense CFP-SUMO spot in the samenuclei (Figure 3C). Colocalization of Ufd1 and SUMO indicatesthat Ufd1 participates in the turn-over of SUMO-conjugates in adirect manner. Ufd1-YFP foci would be induced by increasedsumoylation activities occurring as part of the DNA damageresponse whose products need to be turned-over by Ufd1.Consistent with a need for Ufd1 for this turn-over, GFP-SUMOfoci increased in intensity in the ufd1ΔCt213-342 background afterZeocin treatment (Figure 2B). GFP-SUMO foci did not increasein intensity in wild-type cells after Zeocin treatment (Figure 2B)
indicating that sumoylation events induced by DNA damageare efficiently turned-over by Ufd1 in wild type.
Genome instability in ufd1ΔCt 213-342 mutant cellsSTUbL mutants are sensitive to genotoxic stress [26-28]. We
tested whether the ufd1ΔCt213-342 mutant might be similarlysensitive to DNA-damaging agents by exposing cells tocamptothecin (CPT), hydroxyurea (HU) or UV irradiation. Wefound that ufd1ΔCt213-342 cells were not particularly sensitive tolow doses of UV irradiation (50 J/m2) but they werehypersensitive to higher doses as well as to respectively HUand CPT (Figure 4A). The toxic effect of CPT is thought tooccur during DNA replication when incoming replication forkscollide with topoisomerase 1-DNA complexes stabilized byCPT, resulting in fork collapse [62]. High doses of UV light orchronic exposure to HU can lead to similar types of damage[63,64]. Hence the spectrum of sensitivities of ufd1ΔCt213-342
cells indicates that the mutant might fail to repair aberrant DNAstructures that arise from replicative stress, such as collapsedreplication forks and resulting double-strand breaks (DSBs).
Figure 3. Ufd1 forms foci upon DNA damage which colocalize with CFP-SUMO. (A) Fluorescence imaging of cells expressingUfd1-YFP propagated in the presence or absence of Zeocin (350 µg/ml for 3 1/2 hrs). (B) Bars represent the percentage of cellswith at least one Ufd1-YFP focus before and after Zeocin treatment averaged from three independent experiments. (C)Colocalization of Ufd1-YFP with CFP-SUMO after Zeocin treatment.doi: 10.1371/journal.pone.0080442.g003
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 5 November 2013 | Volume 8 | Issue 11 | e80442

Figure 4. Ufd1 is required for the maintenance of genome integrity. (A) ufd1ΔCt213-342 cells are sensitive to DNA damage: 10-fold dilution series of the indicated strains were spotted onto YES plates containing camptothecin (CPT) or hydroxyurea (HU), orsubsequently exposed to UV irradiation as indicated. (B) Fluorescence imaging of wild-type and ufd1ΔCt213-342 asynchronouscultures expressing Rad22-YFP. (C) ufd1ΔCt213-342 cells display an increased frequency of spontaneous Rad22 foci: Nuclei wereclassified according to their position in the cell cycle based on cell morphology. Bars represent the percentage of nuclei with at leastone Rad22-YFP focus averaged from three independent experiments. Error bars correspond to the standard deviations for thecombined data and p-values were calculated with a Fishers exact test. More than 300 cells of each strain were counted in eachexperiment, the numbers are reported in Table S2. Only very few cells were counted in the ‘multiple nuclei/septa’ category, givingrise to the large standard deviations in this category. (D) ufd1ΔCt213-342 cells are unable to recover from Zeocin-induced damage:Rad22-YFP foci were quantified after 1 hr of growth in Zeocin-containing medium (300 µg/ml) and again 13 hr after Zeocin had beenremoved from the media. Between 50 and 100 cells were counted for each strain at each time point; error bars indicate exactbinomial 95% confidence intervals. Indicated p-values were obtained with a Fishers exact test. Fluorescence images are shown inFigure S3. (E) and (F) Rad22-YFP accumulation in ufd1ΔCt213-342 cells: (E) Rad22-YFP from whole cell extracts of the indicatedstrains was detected by an anti-GFP antibody. (F) Rad22-YFP was affinity-purified from a wild-type or ufd1ΔCt213-342 geneticbackground and detected by anti-GFP immunoblotting. HMW indicates higher molecular weight species of Rad22-YFPaccumulating in ufd1ΔCt213-342. * indicates a crossreacting species.doi: 10.1371/journal.pone.0080442.g004
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 6 November 2013 | Volume 8 | Issue 11 | e80442

Consistent with the hypersensitivity of the ufd1ΔCt213-342
mutant to DNA stress, we observed more spontaneous Rad22-YFP foci in ufd1ΔCt213-342 cells than in wild type (Figure 4B andC). In addition, the Rad22-YFP foci often appeared moreintense in the ufd1ΔCt213-342 mutant. As a central player in HR,Rad22 forms foci at DSBs and other regions with exposedssDNA where HR engages in repair [65]. Analysis of focidistribution along the cell cycle in ufd1ΔCt213-342 cells found thatRad22 foci appeared during S-phase and persisted into G2.This is in contrast to the wild-type situation in whichrecombination-based repair of damage occurring during S-phase is delayed until the end of replication, leading to atemporary accumulation of Rad22 foci only in late S-phase/G2cells ([66,67]; Figure 4C). The greater number of S-phase cellscontaining Rad22 foci thus suggest that the ufd1ΔCt213-342
mutant suffers more damage during replication such ascollapsed replication forks that present substrates for HRproteins. The persistence of foci during G2 further suggeststhat ufd1ΔCt213-342 cells are also impaired in the recovery fromDNA damage through HR. Consistently, ufd1ΔCt213-342 cellsrecovered poorly from Zeocin treatment and cells arrested withbright Rad22 foci indicating incomplete HR events (Figure 4D).Thus as for STUbLs [26,28], impaired Ufd1 function decreasesthe resistance to various forms of genotoxic insults includingreplicative stress. Moreover, as for STUbL mutants [26,27], theG2/M checkpoint appeared functional in the ufd1ΔCt213-342
mutant as Rad22 foci were not observed in mitosis, indicatingthat cells are prevented from dividing with unrepaired DNA.Furthermore, ufd1ΔCt213-342 cells were in general elongatedsuggesting delayed cell cycle progression due to checkpointactivation and they did not display a ‘cut’ phenotype.
Rad22 is a known target for sumoylation in S. pombe [7]. Itssumoylation is catalyzed by Pli1 [4]. We examined themodification status of Rad22 in wild-type and ufd1ΔCt213-342
cells by Western blot. Corroborating our microscopy results weobserved that Rad22 protein levels were increased inufd1ΔCt213-342 cells compared to wild type as seen in whole cellextracts and by pull-down using the Rad22-YFP construct(Figure 4E and F). Furthermore, immunoprecipitation of Rad22-YFP revealed that slowly-migrating forms of Rad22 wereenriched in ufd1ΔCt213-342 cells (Figure 4F), suggesting Rad22is a substrate in the STUbL/Ufd1 pathway. The observedincrease in Rad22 amounts in the ufd1ΔCt213-342 mutant couldbe due either to increased protein synthesis or to proteinstabilization. A recent report proposes Rad22 protein levels arecontrolled by a proteasomal pathway involving the proteasome-associated factor Bag101. Consistent with this the authorsdetected more Rad22 in proteasome mutants [68]. Yet anotherstudy, in S. cerevisae, found that mutations in proteasomalsubunits influence Rad52 levels rather through increasedtranscription [69]. However, as we did not observe any changein rad22 mRNA expression in ufd1ΔCt213-342 cells as judged byquantitative PCR (Figure S3), our data suggest that Rad22may instead accumulate in the ufd1ΔCt213-342 background dueto stabilization.
Epistasis with Pli1Reducing SUMO-conjugate formation by either deleting the
major SUMO E3 ligase Pli1 (pli1∆) or overexpressing thedeSUMOylase Ulp2 suppresses the growth defects andgenotoxin sensitivity of STUbL mutants [26,27]. In contrast,pli1∆ failed to rescue the growth defect or damage sensitivity ofufd1ΔCt213-342 cells; if anything the ufd1ΔCt213-342 pli1∆ doublemutant grew slightly more poorly than the single ufd1ΔCt213-342
mutant (Figure 5A). This shows that the growth impairment andDNA-damage sensitivity in ufd1ΔCt213-342 cells are not merelycaused by deregulation of Pli1-dependent SUMO conjugates,as inferred for STUbL mutants.
Epistasis analysis with Rad22, Rhp51, Rqh1 and Rhp18The relationship between Ufd1 and the DNA-damage
response was investigated further through epistasis analyses.Synthetic growth defects and increased sensitivity to DNA-damaging agents were observed when the ufd1ΔCt213-342
mutation was combined with the deletion of factors required forHR, respectively rad22∆, rhp51∆ and rqh1∆ (Figure 5B). Thesephenotypes are consistent with the ufd1ΔCt213-342 mutantaccumulating more damage needing HR for repair than wild-type cells as suggested by the increase in spontaneous Rad22foci seen in ufd1ΔCt213-342 cells (Figure 4B). A similardependency on recombination factors was previously reportedfor STUbL mutants [26].
The genetic interactions between ufd1 and a gene central topost-replication repair (PRR), rhp18, were more complex.Rhp18 stimulates mono-ubiquitylation of PCNA as part of thePRR response to tolerate replication-stalling lesionsencountered during S-phase [21,23]. This mono-ubiquitylationleads to error-prone replication past the lesion by TLSpolymerases. PCNA can also be further modified by a differentset of enzymes and this stimulates a more error-freemechanism of damage bypass thought to involve a switch intemplate to the newly synthesized sister strand [21-23,70,71].We found that the ufd1ΔCt213-342 mutation was epistatic torhp18∆ for growth and HU sensitivity and the double mutantshowed a cumulative sensitivity to UV irradiation (Figure 5C).These phenotypes are consistent with Ufd1 and Rhp18operating in separate pathways. They are consistent with Ufd1mediating aspects of HR necessary to overcome replication-fork damage such as caused by prolonged HU treatment, andwith Rhp18-mediated PRR being primarily required for thetolerance to single-stranded damage, such as photoproducts[23]. Since HR is essential for UV-tolerance when the PRRpathways are inactivated [23], the cumulative sensitivity to UVirradiation in the double ufd1ΔCt rhp18∆ mutant supports thenotion that Ufd1 takes part in HR. The sensitivity ofufd1ΔCt213-342 cells to CPT however was slightly suppressed byrhp18∆ (Figure 5B). The rhp18∆ single mutant is notparticularly sensitive to CPT, at least not at the doses testedhere (concentrations ≤10µM), consistent with HR being apreferred mode of CPT-induced damage tolerance at thesedoses. Suppression of the CPT-sensitivity of ufd1ΔCt213-342 byrhp18∆ suggests that in addition to performing Rhp18-independent functions, Ufd1 might be necessary either toprevent inappropriate entry into the Rhp18 PRR pathway, or to
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 7 November 2013 | Volume 8 | Issue 11 | e80442

complete events initiated by Rhp18. To further investigatewhich branch of PRR is detrimental in the ufd1ΔCt213-342
background, the epistatic relationship between ufd1 and agene controlling entry into the “error-free” pathway of PRR,
Figure 5. Epistasis analysis of the ufd1ΔCt213-342 mutation with mutations in DNA repair or STUbL pathway. (A, B, C) 10-foldserial dilutions of the indicated strains were spotted onto YES plates with or without DNA-damaging agents as indicated, or ontoplates subsequently exposed to UV irradiation. (A) Epistatic relationship of ufd1ΔCt213-342 with pli1Δ indicates non-overlappingfunctions of Ufd1 and the STUbL pathway. (B) Negative genetic interactions between ufd1ΔCt213-342 and mutations of HR repairproteins. (C) Partial suppression of ufd1ΔCt213-342 CPT sensitivity by rhp18Δ and ubc13Δ.doi: 10.1371/journal.pone.0080442.g005
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 8 November 2013 | Volume 8 | Issue 11 | e80442

ubc13, was examined. Ubc13 forms part of an E2 heterodimer(Ubc13/Mms2) that stimulates PCNA poly-ubiquitylation,promoting repair by template switching. Interestingly we foundthat similar to rhp18∆, the ubc13∆ deletion partly suppressedthe ufd1ΔCt213-342 mutant’s sensitivity to CPT (Figure 5C). Thus,entry into the Ubc13-dependent sub-pathway of PRR inresponse to CPT-induced damage appears deleterious in theabsence of functional Ufd1. In response to UV and HU, thesame epistatic patterns were observed when combiningubc13∆ with ufd1ΔCt213-342 as for rhp18∆ (data not shown).
No suppression of the sensitivity to CPT was observed whencombining slx8-1 and rhp18∆. Instead a cumulative effect wasobserved (Figure 5C), suggesting that a function for Ufd1 incontrolling Rhp18-mediated responses represents an Slx8-independent role.
Discussion
Ufd1 mediates the interactions of the Cdc48/Ufd1/Npl4complex with ubiquitylated proteins, permitting the extraction ofthese proteins from higher-order complexes in an energy-driven process catalyzed by the Cdc48 ATPase [38,39]. Oncemobilized, the ubiquitylated proteins can either be degraded orallowed to perform new tasks. The specificity of targeting andits outcomes are governed through exchangeable interactionpartners of Cdc48/Ufd1/Npl4, including both ubiquitin-bindingand -processing co-factors [43,72,73]. Our observations leadus to propose that Ufd1 operates in the STUbL pathway to
decide the fate of nuclear proteins sumoylated as part of theDDR.
Both physical and functional overlaps between Ufd1-mediated protein processing and the STUbL pathway wererevealed by our experiments. First, a C-terminal domain ofUfd1 bound both the SUMO E3 ligase Pli1 and the STUbL Rfp1suggesting Cdc48/Ufd1/Npl4 has a function in the turnover ofsumoylated proteins (Figure 1A and B). Second, sumoylatedspecies of high molecular weight accumulated in a ufd1 mutantwith a C-terminal truncation of the protein, similar to whenSTUbL function is compromised (Figure 1E). Given the abilityof Ufd1 to mediate the association of the Cdc48/Ufd1/Npl4complex with ubiquitylated proteins we imagine that Cdc48/Ufd1/Npl4 operates downstream of STUbLs to promote furtherprocessing of STUbL substrates and possibly their proteasomaldegradation (Figure 6). Third, supporting a model where Ufd1affects the turnover of SUMO-conjugates in a direct manner,foci formed by Ufd1 in response to DNA damage colocalizedwith a major SUMO focus at the nuclear periphery (Figure 3C).In the absence of DNA damage, nuclear SUMO foci were moreintense in ufd1ΔCt213-342 cells than in wild type and upon DNAdamage the intensity of these foci increased even further inufd1ΔCt213-342 cells (Figure 2B). As many proteins aresumoylated during DDR responses [5,7-16] these foci mightcontain sumoylated repair factors that are normally processedthrough Ufd1.
The nature of the physical interactions between Ufd1 andSTUbLs is of particular interest. A recent study identified a SIMnear the C-terminus of S. pombe Ufd1 [74]. This indicated to
Figure 6. Model depicting a role for Ufd1 (Cdc48/Ufd1/Npl4) in the processing of STUbL substrates. In addition to directrecognition of STUbL modified substrates, interactions between Cdc48/Ufd1/Npl4 and the modifying enzymes (Pli1 and Rfp1/Slx8)could facilitate Cdc48/Ufd1/Npl4 recruitment and a concerted action in the regulation of protein fate.doi: 10.1371/journal.pone.0080442.g006
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 9 November 2013 | Volume 8 | Issue 11 | e80442

the authors that Ufd1 might be a STUbL effector since Ufd1would have the potential of being recruited cooperatively toSTUbL substrates co-modified by SUMO and ubiquitin. Here,our observations suggest that Ufd1 might not be solelyrecruited to STUbL substrates through dual SUMO/ubiquitinrecognition of the substrates but by additional interactionsbetween Ufd1 and the modifying enzymes Pli1 and Rfp1. Wefound that the last 95 aa of Ufd1 are sufficient for theseinteractions. Mutations in the Ufd1 SIM, which is located at thevery C-terminus of Ufd1, abrogated the two-hybrid interactionsof Ufd1 with both Pli1 and Rfp1 (Figure S2). This indicates thatthe interactions are enhanced by SUMO moieties bound to theSIM of Ufd1, or that, in some other way, the same portion ofUfd1 is required for its interactions with SUMO, Pli1 and Rfp1(Figure S2). Our in vitro GST pull-downs suggested that Ufd1has intrinsic affinity for both Pli1 and Rfp1 (Figure 1B). We alsofound that the Pli1 SIM is neither necessary nor sufficient forthe two-hybrid interaction between Pli1 and Ufd1 in S.cerevisiae, supporting the conclusion that the two proteinsestablish direct contact with each other rather than – or inaddition to being bridged by SUMO moieties (Figure S1).According to earlier studies Ufd1 and Cdc48 are sumoylated inS. cerevisiae [75,76]. Hannich et al. (2005) [75] also identified aputative SIM in Cdc48 and they proposed that functionalinteractions between Cdc48 and Ufd1 might be regulated bysumoylation. Interactions between the sumoylation pathwayand Cdc48 in S. cerevisiae were also detected in other large-scale studies [77,78]. Collectively, these and our observationslead to the view that a combination of direct interactions andindirect associations through SUMO and ubiquitin modulate theformation of higher-order complexes comprising Cdc48/Ufd1/Npl4, Pli1, STUbLs and their substrates to enable a concertedaction in the regulation of protein fate.
The slow growth and genome instability of STUbL mutantshave been attributed to an accumulation of one or several poly-sumoylated species since deleting Pli1, overexpressing thedesumoylase Ulp2, or preventing chain formation by mutatingmajor lysine acceptor sites in SUMO suppresses thesephenotypes [26,27,31,74]. The observed genome instability ofthe ufd1ΔCt213-342 mutant is unlikely to be explained solely by anaccumulation of the same SUMO conjugates as deleting Pli1did not suppress the ufd1ΔCt213-342 mutation. This is consistentwith the emerging picture that Cdc48/Ufd1/Npl4 is required forseveral aspects of genome maintenance [44-54].
Recent studies have proposed several functions for Cdc48/Ufd1/Npl4 in ubiquitin-mediated maintenance of genomeintegrity. In metazoans, Cdc48/Ufd1/Npl4 deficient cells fail todegrade the replication-licensing factor Cdt1 and arrest in S-phase with reduced DNA content and Rad51 foci [46-49].Another proposed function for Cdc48/Ufd1/Npl4, in mammaliancells, is in the response to DSBs where ubiquitin-dependentrecruitment of Cdc48/Ufd1/Npl4 permits the removal of K48-linked ubiquitylated species from the sites of lesion and therecruitment of downstream repair factors including 53BP1 [51].Mechanistically, recruitment of 53BP1 is believed to depend onCdc48/p97 dissociating the L3MBTL1 polycomb group proteinfrom histone H4K20me2, a binding site common to both 53BP1and L3MBTL1 [52]. Although this mechanism might not be
conserved in yeast as no homolog of L3MBTL1 has beenfound, the S. pombe ufd1ΔCt213-342 mutant was clearly defectivein some aspects of HR. The ufd1ΔCt213-342 mutant was sensitiveto DSB-inducing agents; ufd1ΔCt213-342 cells treated with Zeocinarrested with bright Rad22 foci; and Rad22 accumulated inhigh-molecular-weight forms in ufd1ΔCt213-342 cells (Figure 4F).Hence, our observations permit to add Rad22 to the short list ofknown factors whose processing depends on Cdc48/Ufd1/Npl4. Evidence for S. cerevisae Rad52 also being a target ofCdc48-Ufd1-Npl4 was recently provided by Bergink et al. [79].These authors proposed that Cdc48-Ufd1-Npl4 recognizessumoylated Rad52 through the Ufd1 SIM and mediates thedisassociation of Rad52 and Rad51 from each other and fromDNA. Whether this specific pathway involves STUbL activitywas not investigated. Even though the study indicates thatsumoylation is sufficient for the association of Cdc48-Ufd1-Npl4with Rad52, STUbL-mediated ubiquitylation of Rad52 mightstrengthen this interaction in vivo or alternatively operatedownstream to regulate Rad52 fate. Consistent with thisproposition, S. cerevisae Rad52 is an in vitro STUbL substrate[80,81].
PRR mediates tolerance to DNA damage encounteredduring S-phase [71]. Entry into the PRR pathway is controlledby the ubiquitin E3 ligase Rhp18. Rhp18-mediated mono-ubiquitylation of PCNA [21,23] stimulates translesion synthesisby recruiting TLS polymerases able to replicate across fork-stalling lesions [22,82,83]. Further elongation of the mono-ubiquitin tag on PCNA by Ubc13/Mms2 stimulates an error-freemode of damage bypass known as template switching[21,70,84]. Here we observed that deleting rhp18 or ubc13partially suppressed the CPT sensitivity of ufd1ΔCt213-342 cells(Figure 5C). These data indicate that entry into the templateswitch branch of PRR is detrimental in the ufd1ΔCt213-342
background, at least when dealing with CPT-induced damage.The residual CPT-sensitivity of the ufd1ΔCt213-342 rhp18∆ doublemutant indicates additional role(s) for Ufd1 in repair of CPT-induced damage. Also consistent with Ufd1 and Rhp18 actingseparately, the ufd1ΔCt213-342 rhp18∆ double mutant showedsynergistic sensitivity to UV irradiation, which creates damagethat critically depend on HR when PRR is compromised [23].
Our observations strengthen and refine the notion thatCdc48-Ufd1-Npl4 is essential to the maintenance of genomeintegrity by acting in several pathways including the STUbLpathway. We propose that Ufd1 acts downstream and/or inconcert with SUMO ligases and STUbLs to remove or recyclesumoylated species. Given the widespread use of sumoylationin repair complexes this action might be necessary to thedynamics of repair at multiple points in the DDR. A broaderrelationship between sumoylation and the Cdc48-Ufd1-Npl4complex might affect cellular processes other than the DDR.
Materials and Methods
Strains and mediaGenotypes are listed in Table S1. S. pombe strains were
propagated in yeast extract medium or in Edinburgh minimalmedium (EMM2) with indicated supplements. S. cerevisiae waspropagated in YPD or SC drop-out media.
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 10 November 2013 | Volume 8 | Issue 11 | e80442

Two-hybrid screensThe Pli1 and Rfp1 ORFs were cloned into pGBKT7
(Clontech) as in-frame fusions with the ORF for the Gal4 DNA-binding domain (GBD) and each plasmid was transformed intoPJ69-4A (MATa trp1-901 leu2-3,112 ura3-52 his3-200 gal4Δgal80Δ LYS2::GAL1-HIS3 GAL2-ADE2 met2::GAL7-lacZ; [85]),selecting for Trp+. The resulting strains were transformed with atwo-hybrid library in pGAD-GH in which S. pombe cDNAs werefused to the sequence coding for the Gal4 transcription-activating domain (Clontech). More than 5 million Leu+ Trp+
transformants were obtained for Rfp1 and more than 10 millionfor Pli1. Separate selections were applied for activation of theADE2 reporter gene (by plating on SC-leu-trp-ade) and for theactivation of the HIS3 reporter gene (by plating on SC-leu-trp-his+3 mM 3-aminotriazol, or 1.5 mM 3-aminotriazol). Ade+
transformants were subsequently tested for expression of HIS3and, vice versa, His+ transformants were tested for theexpression of ADE2. Plasmids were extracted from Ade+ His+
candidates, retested by co-transformation with bait-encodingplasmids or empty vectors, and partially sequenced. Theclones presented in Figure 1 represent only a fraction of theclones capable of activating both the ADE2 and HIS3 reportergenes in a bait-dependent manner identified in the screens.
Construction of ufd1∆Ct213-342 strainsTo produce Ufd1 lacking amino acids 213-343, a plasmid
was prepared as follows: part of the ufd1 ORF (nucleotide26-969) was amplified by PCR on S. pombe genomic DNAusing as primers GTO-336(CCATCCCGGGGTACGTTGACTTACTCACGTATTAG,introducing an XmaI site, underlined) and GTO-337(GTATGGCGCGCCTCATAACCGATGGGGGGGATCAAAATC, introducing an AscI site, underlined). GTO-337 was designedwith an extra G in a row of six guanines, which upon genomicintegration of the amplified sequence should create a singlenucleotide insertion in the ufd1 ORF at nucleotide 953 causinga frameshift and a resulting stop at codon 213. ufd1 3’flankingDNA (933 bp downstream of the ORF) was amplified withGTO-338(CCATGAGCTCTGTTAATCGTCTCAAGTTATTACTTG,introducing a SacI site, underlined) and GTO-339(GTCTACTAGTGAGGAGCTTGACGGCGTCTGCGAGG,introducing an SpeI site, underlined). Coding and 3’fragmentswere cloned into pFA6a-hphMX6 [86] using the restriction sitesintroduced in the primers and the resulting plasmid wasdigested with XmaI-SpeI before transformation into a diploidstrain, selecting for hygromycin-resistance. ufd1∆Ct213-342
hygromycin-resistant haploid progeny were obtained by tetraddissection of diploid JK5. Correct integration of the constructwas confirmed by PCR analysis using the primers GTO-336(see above) and GTO-340(GGGCAGCGTTCTTAGCACGAGCTTC) or GTO-336 andGTO-342 (CGCTATACTGCTGTCGATTCG) and by Southernblotting. The hph1 gene was subsequently replaced with thenourseothricin resistance gene (nat1) by transformation of theufd1∆Ct213-342 strain JK9 with the pCR2.1-nat plasmid digestedwith EcoRI [86].
GST pull-downsPli1 and Rfp1 were expressed as GST-fusions from the
pGEX-KG vector in the bacterial strain BL21 (DE3) pLysS cellsand purified under native conditions (1x PBS, 0.1% Triton, 1mM DTT, 1x Complete protease inhibitor Cocktail EDTA-freefrom Roche) on a glutathione–Sepharose column (Amersham)using standard protocols. For 35S-methionine-labeling of Ufd1,a PCR product amplified from genomic DNA with the primersGTO-483(GCATGGATCCATGATGACTACACTTAGCCTTGAGCC,introducing a BamHI site, underlined) and GTO-484(CGATGAATTCTTAAGCATCAATATCGATTGGGTC,introducing an EcoRI site, underlined) was cloned into pING14[87] and used to produce a C-terminal fragment correspondingto amino acids 110-342 of Ufd1 by in vitro translation using thethe SP6 TNT-coupled reticulocyte lysate kit (Promega) and 35S-methionine (Perkin Elmer). ~20 µg of GST or GST-fusionproteins were incubated with the in vitro translation productovernight at 4πC in 1x PBS, 0.1% NP-40, 5% glycerol, 1 mMDTT. Glutathione–Sepharose beads were added for 3 hr andwashed 5 times. Bound proteins were released by boiling in 1xLaemmli sample buffer. Released proteins were resolved bygel electrophoresis and Ufd1 was detectedby phosphorimaging.
Anti-SUMO Western blottingWhole-cell extracts for immunoblot analysis of total SUMO
species were prepared from cells growing exponentially insupplemented EMM2. Cells were lysed in TNET buffer (200mM NaCl, 0.1% Triton, 0.01% SDS, 50 mM Tris-HCl pH8.0, 50mM EDTA, 10 mM N-ethylmaleimide, 1x Complete proteaseinhibitor Cocktail EDTA-free from Roche) with glass beads in aFastprep® Instrument kept at 4 °C. Equal amounts of proteinextracts boiled in 1x Laemmli sample buffer were separated ona 4-20% Tris-glycine gradient gel (Lonza), transferred to anitrocellulose/MCE membrane (Advantec), and probed with ananti-SUMO rabbit antibody ([4]; kindly provided by J. Seeler)followed by a horseradish peroxidase (HRP)-conjugated swineanti-rabbit IgG secondary antibody (Dako). Detection was withan ECL plus kit (GE Healthcare).
GFP immunoprecipitationsFor immunoprecipitations of Rad22-YFP, 300 ml of cultures
growing exponentially in YEL were harvested at OD600~0.4,resuspended in 800 µl lysis buffer (10 mM Tris pH 7.5, 150 mMNaCl, 0.5 mM EDTA, 10% glycerol, 0.5% NP40, 10 mM N-ethylmaleimide, 1 mM PMSF, 1x Complete protease inhibitorCocktail EDTA-free from Roche, 1µg/µl DNaseI and 2.5 mMMgCl2) and lyzed by the glass bead method. Proteinconcentrations were determined using a NanodropSpectrophotometer (280 nm). Equal amounts of total protein(~54 mg per sample) were diluted 2x in binding buffer (same aslysis buffer but without NP-40, DNaseI and MgCl2) to a finalvolume of ~1.5 ml and incubated with 30 µl pre-equilibratedGFP-Trap®_M beads (Chromotek) for 3 hr at 4°C underconstant mixing. Beads were washed 3 times in binding buffer.Bound proteins were released by incubation at 90°C for 10 minin 2x Laemmli buffer, separated on a 4-20% Tris-glycine
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 11 November 2013 | Volume 8 | Issue 11 | e80442

gradient gel (Lonza), and transferred to a nitrocellulose/MCEmembrane (Advantec). GFP-trapped proteins were detectedusing an anti-GFP mouse antibody (Roche) in combination with(HRP)-conjugated goat anti-mouse from Thermo Scientific andan ECL plus kit (GE Healthcare).
For Cdc48-GFP co-IP experiment, 200 ml of cultures weregrown in YEL to an OD600~0.9. Cells were lysed in 1.4 ml lysisbuffer (25 mM Tris pH 7.5, 50 mM NaCl, 10% glycerol, 0.1%Triton 100-X, 2 mM DTT, 1 mM PMSF and protease inhibitors)with glass beads and ~60 mg of total protein per sample wasmixed with ~130 µl pre-equilibrated GFP-Trap®_M beads(Chromotek) for 1-2 hr at 4°C. Beads were washed 4 times withan increased salt concentration (300 mM NaCl) before elutionof bound material by boiling in Laemmli buffer. Cdc48-GFP wasidentified by the anti-GFP mouse antibody from Roche and co-precipitated Slx8-Myc was identified with a Myc-tag mouseantibody from Cell Signaling Technology.
Fluorescence microscopyStrains used for fluorescence microscopy were propagated
overnight at 30°C in supplemented EMM2, to early exponentialphase. Rad22-YFP was expressed as a replacement of therad22 ORF [88]. Ufd1-YFP was expressed from the nmt1promoter [61] in cells with a full deletion of the endogenousufd1 gene. The nmt1-ufd1-YFP construct integrated at leu1fully complemented the full deletion of ufd1 in media lackingthiamine in which the nmt1 promoter is active. For treatmentwith Zeocin, cells were incubated with 350 µg/ml Zeocin(Invitrogen) for 1-5 hr prior to microscopy as indicated. For theZeocin-recovery experiment, cells were pelleted after a 1 hrtreatment with 350 µg/ml Zeocin and they were allowed to growin fresh supplemented EMM2 medium for ~13 hr in a shaking30°C incubator. Images were obtained with a Zeiss AxioImager Z.I (Carl Zeiss) microscope linked to an Orca-ER CCDcamera (Hamamatsu). Images were analyzed using Volocity(version 5, Improvision).
Quantification of GFP-SUMO focus intensityThe intensity of the brightest GFP-SUMO focus was
measured in wild-type and ufd1ΔCt213-342 cells before and afterZeocin treatment using the Volocity analysis module (version 5,Improvision). The Magic Wand ROI tool was used to selectentire nuclei in 3-D. A value for the voxel of highest intensitywas obtained for each nucleus. Three independentexperiments were performed, producing very similar results.Values obtained in one experiment were compiled to preparethe graphs shown in Figure 2.
RNA extraction and real-time PCRCells were propagated in YEL medium to an OD600~ 0.2
and RNA was extracted as described in Lyne et al., 2003 [89]. Three independent cultures were processed for each strain.The following primers were used for transcript detection byreal-time PCR: rad22+, GTO-551 (5’GACAATCAAAGATGGTGCCTATC 3’) and GTO-552 (5’CATCTGTAGTGCCCTCTTTCTTG 3’); act1+, TJO-55 (5’CTGTTTTGTCTTTGTATGCC 3’) and TJO-58 (5’TAAGGTAGTCAGTCAAGTCA 3’). Real-time PCR was
performed on a BioRad CFX96 system, using a QuantiTectSYBR Green PCR Kit from Qiagen according to themanufacturer instructions. The reverse-transcription step wasperformed at 50°C for 30 min. Following reverse transcription,the samples were heated at 95°C for 15 min, and subjected to39 cycles of (95°C for 15 s, 55°C for 30 s and 72°C for 30 s). All reactions were set up in triplicates and the melting curve ofall PCR products was determined after amplification. Ten-folddilution series of genomic DNA were used to determine primerefficiencies and the exponential range of amplification for eachprimer pair. Mean normalized expression (MNE) values fortranscript levels were calculated according to the equationMNE= (Eref)CTref,mean / (Etarget)CTtarget,mean [90].
Supporting Information
Figure S1. Interactions of Pli1 subclones with Ufd1 andSUMO. (A) Schematic representation of Pli1 domains and Pli1subclones used for the two-hybrid assays displayed in (B). (B)Yeast two-hybrid interactions. 10-fold dilution series of S.cerevisae strain PJ69-4A expressing the indicated fusion toGAD or GBP were spotted on the indicated media. GAD-Ufd1expresses the C-terminal domain of Ufd1 (aa 248-342) fused tothe Gal4 DNA activation domain (GAD). GAD-Pmt3 expressesS. pombe SUMO fused to GAD. Protein interactions result inactivation of the ADE2 and HIS3 reporter genes in the testerstrain and growth on SC-leu-trp-ade and SC-leu-trp-his+3mM3-AT. Blue arrows point to Pli1 subclones that do notinteract with SUMO, yet interact with Ufd1. The red arrowpoints to a Pli1 subclone interacting strongly with SUMO butnot Ufd1. Globally, the interactions of Pli1 with SUMO requirethe Pli1 SIM or SP-RING domain whereas the interactions ofPli1 with Ufd1 occur in the absence of these two domains.(TIF)
Figure S2. Mutations in the Ufd1 SIM abrogate the two-hybrid interactions of Ufd1 with Pli1 and Rfp1. (A)Representation of the Ufd1 C-terminal portion used for two-hybrid assays, annotating a SIM motif in the last seven aminoacids that was either deleted (Ufd1Ct∆SIM) or mutated(Ufd1CtDAADADA) for the interaction tests shown below. (B)Transformants of S. cerevisiae strain PJ69-4A expressing theindicated GAD- and GBD-fusion proteins were spotted ontoselective media to test for interactions.(TIF)
Figure S3. Supplemental data to Figure 4. (A) Fluorescenceimaging of Rad22-YFP in wild type and ufd1ΔCt213-342 mutantbefore and after treatment with Zeocin (300 µg/ml for 1 hr.) (B)rad22 transcript levels are similar in wild-type and ufd1ΔCt213-342
mutant cells. Real-time RT-PCR analysis of rad22 RNAisolated from wild-type or ufd1ΔCt213-342 cells. rad22 RNAquantities were measured relative to actin. Error bars indicatethe standard deviation obtained from three independentbiological isolates.(EPS)
Table S1. Strain Table.
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 12 November 2013 | Volume 8 | Issue 11 | e80442

(DOC)
Table S2. Number of Rad22-YFP foci and nuclei countedto produce the bar graphs shown in Figure 4C. Numberswere combined from three independent experiments.(DOCX)
Acknowledgements
We thank Jacob Seeler for the anti-Pmt3 antibody, and KatsuTanaka, Nick Boddy, Colin Gordon, Eishi Noguchi and the
Riken Bioresource Center for strains. We are also very gratefulto Janne Verhein-Hansen for excellent technical assistance.
Author Contributions
Conceived and designed the experiments: JBK MLMJ GB MTGT. Performed the experiments: JBK MLMJ GB MT GT.Analyzed the data: JBK MLMJ GB MT GT. Contributedreagents/materials/analysis tools: GT. Wrote the manuscript:JBK GT.
References
1. Gareau JR, Lima CD (2010) The SUMO pathway: emergingmechanisms that shape specificity, conjugation and recognition. NatRev Mol Cell Biol 11: 861-871. doi:10.1038/nrm3011. PubMed:21102611.
2. Wilkinson KA, Henley JM (2010) Mechanisms, regulation andconsequences of protein SUMOylation. Biochem J 428: 133-145. doi:10.1042/BJ20100158. PubMed: 20462400.
3. Hochstrasser M (2009) Origin and function of ubiquitin-like proteins.Nature 458: 422-429. doi:10.1038/nature07958. PubMed: 19325621.
4. Xhemalce B, Seeler JS, Thon G, Dejean A, Arcangioli B (2004) Role ofthe fission yeast SUMO E3 ligase Pli1p in centromere and telomeremaintenance. EMBO J 23: 3844-3853. doi:10.1038/sj.emboj.7600394.PubMed: 15359282.
5. Andrews EA, Palecek J, Sergeant J, Taylor E, Lehmann AR et al.(2005) Nse2, a component of the Smc5-6 complex, is a SUMO ligaserequired for the response to DNA damage. Mol Cell Biol 25: 185-196.doi:10.1128/MCB.25.1.185-196.2005. PubMed: 15601841.
6. Watts FZ, Skilton A, Ho JC, Boyd LK, Trickey MA et al. (2007) The roleof Schizosaccharomyces pombe SUMO ligases in genome stability.Biochem Soc Trans 35: 1379-1384. doi:10.1042/BST0351379.PubMed: 18031226.
7. Ho JC, Warr NJ, Shimizu H, Watts FZ (2001) SUMO modification ofRad22, the Schizosaccharomyces pombe homologue of therecombination protein Rad52. Nucleic Acids Res 29: 4179-4186. doi:10.1093/nar/29.20.4179. PubMed: 11600706.
8. Sacher M, Pfander B, Hoege C, Jentsch S (2006) Control of Rad52recombination activity by double-strand break-induced SUMOmodification. Nat Cell Biol 8: 1284-1290. doi:10.1038/ncb1488.PubMed: 17013376.
9. Burgess RC, Rahman S, Lisby M, Rothstein R, Zhao X (2007) TheSlx5-Slx8 complex affects sumoylation of DNA repair proteins andnegatively regulates recombination. Mol Cell Biol 27: 6153-6162. doi:10.1128/MCB.00787-07. PubMed: 17591698.
10. Ohuchi T, Seki M, Kugou K, Tada S, Ohta K et al. (2009) Accumulationof sumoylated Rad52 in checkpoint mutants perturbed in DNAreplication. DNA Repair (Amst) 8: 690-696. doi:10.1016/j.dnarep.2009.01.018. PubMed: 19261547.
11. Branzei D, Sollier J, Liberi G, Zhao X, Maeda D et al. (2006) Ubc9- andmms21-mediated sumoylation counteracts recombinogenic events atdamaged replication forks. Cell 127: 509-522. doi:10.1016/j.cell.2006.08.050. PubMed: 17081974.
12. Morris JR, Boutell C, Keppler M, Densham R, Weekes D et al. (2009)The SUMO modification pathway is involved in the BRCA1 response togenotoxic stress. Nature 462: 886-890. doi:10.1038/nature08593.PubMed: 20016594.
13. Galanty Y, Belotserkovskaya R, Coates J, Polo S, Miller KM et al.(2009) Mammalian SUMO E3-ligases PIAS1 and PIAS4 promoteresponses to DNA double-strand breaks. Nature 462: 935-939. doi:10.1038/nature08657. PubMed: 20016603.
14. Dou H, Huang C, Singh M, Carpenter PB, Yeh ET (2010) Regulation ofDNA repair through deSUMOylation and SUMOylation of replicationprotein A complex. Mol Cell 39: 333-345. doi:10.1016/j.molcel.2010.07.021. PubMed: 20705237.
15. Cremona CA, Sarangi P, Yang Y, Hang LE, Rahman S et al. (2012)Extensive DNA damage-induced sumoylation contributes to replicationand repair and acts in addition to the mec1 checkpoint. Mol Cell 45:422-432. doi:10.1016/j.molcel.2011.11.028. PubMed: 22285753.
16. Psakhye I, Jentsch S (2012) Protein group modification and synergy inthe SUMO pathway as exemplified in DNA repair. Cell 151: 807-820.doi:10.1016/j.cell.2012.10.021. PubMed: 23122649.
17. Hardeland U, Steinacher R, Jiricny J, Schär P (2002) Modification ofthe human thymine-DNA glycosylase by ubiquitin-like proteinsfacilitates enzymatic turnover. EMBO J 21: 1456-1464. doi:10.1093/emboj/21.6.1456. PubMed: 11889051.
18. Mailand N, Bekker-Jensen S, Faustrup H, Melander F, Bartek J et al.(2007) RNF8 ubiquitylates histones at DNA double-strand breaks andpromotes assembly of repair proteins. Cell 131: 887-900. doi:10.1016/j.cell.2007.09.040. PubMed: 18001824.
19. Ulrich HD (2012) Ubiquitin and SUMO in DNA repair at a glance. J CellSci 125: 249-254. doi:10.1242/jcs.091801. PubMed: 22357966.
20. Bekker-Jensen S, Mailand N (2011) The ubiquitin- and SUMO-dependent signaling response to DNA double-strand breaks. FEBS Lett585: 2914-2919. doi:10.1016/j.febslet.2011.05.056. PubMed:21664912.
21. Hoege C, Pfander B, Moldovan GL, Pyrowolakis G, Jentsch S (2002)RAD6-dependent DNA repair is linked to modification of PCNA byubiquitin and SUMO. Nature 419: 135-141. doi:10.1038/nature00991.PubMed: 12226657.
22. Stelter P, Ulrich HD (2003) Control of spontaneous and damage-induced mutagenesis by SUMO and ubiquitin conjugation. Nature 425:188-191. doi:10.1038/nature01965. PubMed: 12968183.
23. Frampton J, Irmisch A, Green CM, Neiss A, Trickey M et al. (2006)Postreplication repair and PCNA modification in Schizosaccharomycespombe. Mol Cell Biol 17: 2976-2985. doi:10.1091/mbc.E05-11-1008.PubMed: 16641370.
24. Altmannova V, Eckert-Boulet N, Arneric M, Kolesar P, Chaloupkova Ret al. (2010) Rad52 SUMOylation affects the efficiency of the DNArepair. Nucleic Acids Res 38: 4708-4721. doi:10.1093/nar/gkq195.PubMed: 20371517.
25. Danielsen JR, Povlsen LK, Villumsen BH, Streicher W, Nilsson J et al.(2012) DNA damage-inducible SUMOylation of HERC2 promotesRNF8 binding via a novel SUMO-binding Zinc finger. J Cell Biol 197:179-187. doi:10.1083/jcb.201106152. PubMed: 22508508.
26. Prudden J, Pebernard S, Raffa G, Slavin DA, Perry JJ et al. (2007)SUMO-targeted ubiquitin ligases in genome stability. EMBO J 26:4089-4101. doi:10.1038/sj.emboj.7601838. PubMed: 17762865.
27. Kosoy A, Calonge TM, Outwin EA, O'Connell MJ (2007) Fission yeastRnf4 homologs are required for DNA repair. J Biol Chem 282:20388-20394. doi:10.1074/jbc.M702652200. PubMed: 17502373.
28. Sun H, Leverson JD, Hunter T (2007) Conserved function of RNF4family proteins in eukaryotes: targeting a ubiquitin ligase toSUMOylated proteins. EMBO J 26: 4102-4112. doi:10.1038/sj.emboj.7601839. PubMed: 17762864.
29. Uzunova K, Göttsche K, Miteva M, Weisshaar SR, Glanemann C et al.(2007) Ubiquitin-dependent proteolytic control of SUMO conjugates. JBiol Chem 282: 34167-34175. doi:10.1074/jbc.M706505200. PubMed:17728242.
30. Tatham MH, Geoffroy MC, Shen L, Plechanovova A, Hattersley N et al.(2008) RNF4 is a poly-SUMO-specific E3 ubiquitin ligase required forarsenic-induced PML degradation. Nat Cell Biol 10: 538-546. doi:10.1038/ncb1716. PubMed: 18408734.
31. Prudden J, Perry JJ, Nie M, Vashisht AA, Arvai AS et al. (2011) DNArepair and global sumoylation are regulated by distinct Ubc9noncovalent complexes. Mol Cell Biol 31: 2299-2310. doi:10.1128/MCB.05188-11. PubMed: 21444718.
32. Nagai S, Dubrana K, Tsai-Pflugfelder M, Davidson MB, Roberts TM etal. (2008) Functional targeting of DNA damage to a nuclear pore-associated SUMO-dependent ubiquitin ligase. Science 322: 597-602.doi:10.1126/science.1162790. PubMed: 18948542.
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 13 November 2013 | Volume 8 | Issue 11 | e80442

33. Cook CE, Hochstrasser M, Kerscher O (2009) The SUMO-targetedubiquitin ligase subunit Slx5 resides in nuclear foci and at sites of DNAbreaks. Cell Cycle 8: 1080-1089. doi:10.4161/cc.8.7.8123. PubMed:19270524.
34. Galanty Y, Belotserkovskaya R, Coates J, Jackson SP (2012) RNF4, aSUMO-targeted ubiquitin E3 ligase, promotes DNA double-strand breakrepair. Genes Dev 26: 1179-1195. doi:10.1101/gad.188284.112.PubMed: 22661229.
35. Yin Y, Seifert A, Chua JS, Maure JF, Golebiowski F et al. (2012)SUMO-targeted ubiquitin E3 ligase RNF4 is required for the responseof human cells to DNA damage. Genes Dev 26: 1196-1208. doi:10.1101/gad.189274.112. PubMed: 22661230.
36. Parker JL, Ulrich HD (2012) A SUMO-interacting motif activatesbudding yeast ubiquitin ligase Rad18 towards SUMO-modified PCNA.Nucleic Acids Res 40: 11380-11388. doi:10.1093/nar/gks892. PubMed:23034805.
37. Meyer HH, Shorter JG, Seemann J, Pappin D, Warren G (2000) Acomplex of mammalian ufd1 and npl4 links the AAA-ATPase, p97, toubiquitin and nuclear transport pathways. EMBO J 19: 2181-2192. doi:10.1093/emboj/19.10.2181. PubMed: 10811609.
38. Meyer H, Bug M, Bremer S (2012) Emerging functions of the VCP/p97AAA-ATPase in the ubiquitin system. Nat Cell Biol 14: 117-123. doi:10.1038/ncb2407. PubMed: 22298039.
39. Stolz A, Hilt W, Buchberger A, Wolf DH (2011) Cdc48: a powermachine in protein degradation. Trends Biochem Sci 36: 515-523. doi:10.1016/j.tibs.2011.06.001. PubMed: 21741246.
40. Ye Y, Meyer HH, Rapoport TA (2003) Function of the p97-Ufd1-Npl4complex in retrotranslocation from the ER to the cytosol: dualrecognition of nonubiquitinated polypeptide segments and polyubiquitinchains. J Cell Biol 162: 71-84. doi:10.1083/jcb.200302169. PubMed:12847084.
41. Wolf DH, Stolz A (2012) The Cdc48 machine in endoplasmic reticulumassociated protein degradation. Biochim Biophys Acta 1823: 117-124.doi:10.1016/j.bbamcr.2011.09.002. PubMed: 21945179.
42. Moir D, Stewart SE, Osmond BC, Botstein D (1982) Cold-sensitive cell-division-cycle mutants of yeast: isolation, properties, andpseudoreversion studies. Genetics 100: 547-563. PubMed: 6749598.
43. Alexandru G, Graumann J, Smith GT, Kolawa NJ, Fang R et al. (2008)UBXD7 binds multiple ubiquitin ligases and implicates p97 in HIF1alphaturnover. Cell 134: 804-816. doi:10.1016/j.cell.2008.06.048. PubMed:18775313.
44. Vong QP, Cao K, Li HY, Iglesias PA, Zheng Y (2005) Chromosomealignment and segregation regulated by ubiquitination of survivin.Science 310: 1499-1504. doi:10.1126/science.1120160. PubMed:16322459.
45. Ramadan K, Bruderer R, Spiga FM, Popp O, Baur T et al. (2007)Cdc48/p97 promotes reformation of the nucleus by extracting thekinase Aurora B from chromatin. Nature 450: 1258-1262. doi:10.1038/nature06388. PubMed: 18097415.
46. Mouysset J, Deichsel A, Moser S, Hoege C, Hyman AA et al. (2008)Cell cycle progression requires the CDC-48UFD-1/NPL-4 complex forefficient DNA replication. Proc Natl Acad Sci U S A 105: 12879-12884.doi:10.1073/pnas.0805944105. PubMed: 18728180.
47. Deichsel A, Mouysset J, Hoppe T (2009) The ubiquitin-selectivechaperone CDC-48/p97, a new player in DNA replication. Cell Cycle 8:185-190. doi:10.4161/cc.8.2.7356. PubMed: 19158489.
48. Raman M, Havens CG, Walter JC, Harper JW (2011) A genome-widescreen identifies p97 as an essential regulator of DNA damage-dependent CDT1 destruction. Mol Cell 44: 72-84. doi:10.1016/j.molcel.2011.06.036. PubMed: 21981919.
49. Franz A, Orth M, Pirson PA, Sonneville R, Blow JJ et al. (2011)CDC-48/p97 coordinates CDT-1 degradation with GINS chromatindissociation to ensure faithful DNA replication. Mol Cell 44: 85-96. doi:10.1016/j.molcel.2011.08.028. PubMed: 21981920.
50. Verma R, Oania R, Fang R, Smith GT, Deshaies RJ (2011) Cdc48/p97mediates UV-dependent turnover of RNA Pol II. Mol Cell 41: 82-92. doi:10.1016/j.molcel.2010.12.017. PubMed: 21211725.
51. Meerang M, Ritz D, Paliwal S, Garajova Z, Bosshard M et al. (2011)The ubiquitin-selective segregase VCP/p97 orchestrates the responseto DNA double-strand breaks. Nat Cell Biol 13: 1376-1382. doi:10.1038/ncb2367. PubMed: 22020440.
52. Acs K, Luijsterburg MS, Ackermann L, Salomons FA, Hoppe T et al.(2011) The AAA-ATPase VCP/p97 promotes 53BP1 recruitment byremoving L3MBTL1 from DNA double-strand breaks. Nat Struct MolBiol 18: 1345-1350. doi:10.1038/nsmb.2188. PubMed: 22120668.
53. Dantuma NP, Hoppe T (2012) Growing sphere of influence: Cdc48/p97orchestrates ubiquitin-dependent extraction from chromatin. TrendsCell Biol 22: 483-491. doi:10.1016/j.tcb.2012.06.003. PubMed:22818974.
54. Ramadan K (2012) p97/VCP- and Lys48-linked polyubiquitination forma new signaling pathway in DNA damage response. Cell Cycle 11:1062-1069. doi:10.4161/cc.11.6.19446. PubMed: 22391235.
55. Hetzer M, Meyer HH, Walther TC, Bilbao-Cortes D, Warren G et al.(2001) Distinct AAA-ATPase p97 complexes function in discrete stepsof nuclear assembly. Nat Cell Biol 3: 1086-1091. doi:10.1038/ncb1201-1086. PubMed: 11781570.
56. Bruderer RM, Brasseur C, Meyer HH (2004) The AAA ATPasep97/VCP interacts with its alternative co-factors, Ufd1-Npl4 and p47,through a common bipartite binding mechanism. J Biol Chem 279:49609-49616. doi:10.1074/jbc.M408695200. PubMed: 15371428.
57. Park S, Isaacson R, Kim HT, Silver PA, Wagner G (2005) Ufd1 exhibitsthe AAA-ATPase fold with two distinct ubiquitin interaction sites.Structure 13: 995-1005. doi:10.1016/j.str.2005.04.013. PubMed:16004872.
58. Naula N, Hilti N, Schweingruber AM, Schweingruber ME (2003)Cordycepin in Schizosaccharomyces pombe: effects on the wild typeand phenotypes of mutants resistant to the drug. Curr Genet 43:400-406. doi:10.1007/s00294-003-0413-4. PubMed: 12827445.
59. Tanaka K, Nishide J, Okazaki K, Kato H, Niwa O et al. (1999)Characterization of a fission yeast SUMO-1 homologue, pmt3p,required for multiple nuclear events, including the control of telomerelength and chromosome segregation. Mol Cell Biol 19: 8660-8672.PubMed: 10567589.
60. Yuasa T, Hayashi T, Ikai N, Katayama T, Aoki K et al. (2004) Aninteractive gene network for securin-separase, condensin, cohesin,Dis1/Mtc1 and histones constructed by mass transformation. GenesCells 9: 1069-1082. doi:10.1111/j.1365-2443.2004.00790.x. PubMed:15507118.
61. Matsuyama A, Arai R, Yashiroda Y, Shirai A, Kamata A et al. (2006)ORFeome cloning and global analysis of protein localization in thefission yeast Schizosaccharomyces pombe. Nat Biotechnol 24:841-847. doi:10.1038/nbt1222. PubMed: 16823372.
62. Wan S, Capasso H, Walworth NC (1999) The topoisomerase I poisoncamptothecin generates a Chk1-dependent DNA damage checkpointsignal in fission yeast. Yeast 15: 821-828. doi:10.1002/(SICI)1097-0061(199907)15:10A. PubMed: 10407262.
63. Saintigny Y, Delacôte F, Varès G, Petitot F, Lambert S et al. (2001)Characterization of homologous recombination induced by replicationinhibition in mammalian cells. EMBO J 20: 3861-3870. doi:10.1093/emboj/20.14.3861. PubMed: 11447127.
64. Petermann E, Orta ML, Issaeva N, Schultz N, Helleday T (2010)Hydroxyurea-stalled replication forks become progressively inactivatedand require two different RAD51-mediated pathways for restart andrepair. Mol Cell 37: 492-502. doi:10.1016/j.molcel.2010.01.021.PubMed: 20188668.
65. Kim WJ, Lee S, Park MS, Jang YK, Kim JB et al. (2000) Rad22 protein,a rad52 homologue in Schizosaccharomyces pombe, binds to DNAdouble-strand breaks. J Biol Chem 275: 35607-35611. doi:10.1074/jbc.M007060200. PubMed: 10956666.
66. Noguchi E, Noguchi C, McDonald WH, Yates JR 3rd, Russell P (2004)Swi1 and Swi3 are components of a replication fork protection complexin fission yeast. Mol Cell Biol 24: 8342-8355. doi:10.1128/MCB.24.19.8342-8355.2004. PubMed: 15367656.
67. Meister P, Taddei A, Vernis L, Poidevin M, Gasser SM et al. (2005)Temporal separation of replication and recombination requires theintra-S checkpoint. J Cell Biol 168: 537-544. doi:10.1083/jcb.200410006. PubMed: 15716375.
68. Saito Y, Takeda J, Okada M, Kobayashi J, Kato A et al. (2013) Theproteasome factor Bag101 binds to Rad22 and suppresseshomologous recombination. Sci Rep 3: 2022. PubMed: 23779158.
69. Asleson EN, Livingston DM (2003) Investigation of the stability of yeastrad52 mutant proteins uncovers post-translational and transcriptionalregulation of Rad52p. Genetics 163: 91-101. PubMed: 12586699.
70. Branzei D, Vanoli F, Foiani M (2008) SUMOylation regulates Rad18-mediated template switch. Nature 456: 915-920. doi:10.1038/nature07587. PubMed: 19092928.
71. Sale JE (2012) Competition, collaboration and coordination-determininghow cells bypass DNA damage. J Cell Sci 125: 1633-1643. doi:10.1242/jcs.094748. PubMed: 22499669.
72. Richly H, Rape M, Braun S, Rumpf S, Hoege C et al. (2005) A series ofubiquitin binding factors connects CDC48/p97 to substratemultiubiquitylation and proteasomal targeting. Cell 120: 73-84. doi:10.1016/j.cell.2004.11.013. PubMed: 15652483.
73. Schuberth C, Buchberger A (2008) UBX domain proteins: majorregulators of the AAA ATPase Cdc48/p97. Cell Mol Life Sci 65:2360-2371. doi:10.1007/s00018-008-8072-8. PubMed: 18438607.
74. Nie M, Aslanian A, Prudden J, Heideker J, Vashisht AA et al. (2012)Dual recruitment of Cdc48 (p97)-Ufd1-Npl4 ubiquitin-selective
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 14 November 2013 | Volume 8 | Issue 11 | e80442

segregase by small ubiquitin-like modifier protein (SUMO) and ubiquitinin SUMO-targeted ubiquitin ligase-mediated genome stability functions.J Biol Chem 287: 29610-29619. doi:10.1074/jbc.M112.379768.PubMed: 22730331.
75. Hannich JT, Lewis A, Kroetz MB, Li SJ, Heide H et al. (2005) Definingthe SUMO-modified proteome by multiple approaches inSaccharomyces cerevisiae. J Biol Chem 280: 4102-4110. PubMed:15590687.
76. Makhnevych T, Sydorskyy Y, Xin X, Srikumar T, Vizeacoumar FJ et al.(2009) Global map of SUMO function revealed by protein-proteininteraction and genetic networks. Mol Cell 33: 124-135. doi:10.1016/j.molcel.2008.12.025. PubMed: 19150434.
77. Zhou W, Ryan JJ, Zhou H (2004) Global analyses of sumoylatedproteins in Saccharomyces cerevisiae. Induction of protein sumoylationby cellular stresses. J Biol Chem 279: 32262-32268. doi:10.1074/jbc.M404173200. PubMed: 15166219.
78. Wohlschlegel JA, Johnson ES, Reed SI, Yates JR 3rd (2004) Globalanalysis of protein sumoylation in Saccharomyces cerevisiae. J BiolChem 279: 45662-45668. doi:10.1074/jbc.M409203200. PubMed:15326169.
79. Bergink S, Ammon T, Kern M, Schermelleh L, Leonhardt H et al. (2013)Role of Cdc48/p97 as a SUMO-targeted segregase curbing Rad51-Rad52 interaction. Nat Cell Biol 15: 526-532. doi:10.1038/ncb2729.PubMed: 23624404.
80. Xie Y, Kerscher O, Kroetz MB, McConchie HF, Sung P et al. (2007)The yeast Hex3.Slx8 heterodimer is a ubiquitin ligase stimulated bysubstrate sumoylation. J Biol Chem 282: 34176-34184. doi:10.1074/jbc.M706025200. PubMed: 17848550.
81. Ii T, Fung J, Mullen JR, Brill SJ (2007) The yeast Slx5-Slx8 DNAintegrity complex displays ubiquitin ligase activity. Cell Cycle 6:2800-9.80. Kalocsay M, Hiller NJ. PubMed: 18032921
82. Kannouche PL, Wing J, Lehmann AR (2004) Interaction of human DNApolymerase eta with monoubiquitinated PCNA: a possible mechanismfor the polymerase switch in response to DNA damage. Mol Cell 14:491-500. doi:10.1016/S1097-2765(04)00259-X. PubMed: 15149598.
83. Watanabe K, Tateishi S, Kawasuji M, Tsurimoto T, Inoue H et al.(2004) Rad18 guides poleta to replication stalling sites through physical
interaction and PCNA monoubiquitination. EMBO J 23: 3886-3896. doi:10.1038/sj.emboj.7600383. PubMed: 15359278.
84. Brown M, Zhu Y, Hemmingsen SM, Xiao W (2002) Structural andfunctional conservation of error-free DNA postreplication repair inSchizosaccharomyces pombe. DNA Repair (Amst) 1: 869-880. doi:10.1016/S1568-7864(02)00111-8. PubMed: 12531016.
85. James P, Halladay J, Craig EA (1996) Genomic libraries and a hoststrain designed for highly efficient two-hybrid selection in yeast.Genetics 144: 1425-1436. PubMed: 8978031.
86. Sato M, Dhut S, Toda T (2005) New drug-resistant cassettes for genedisruption and epitope tagging in Schizosaccharomyces pombe. Yeast22: 583-591. doi:10.1002/yea.1233. PubMed: 15942936.
87. Bannister AJ, Cook A, Kouzarides T (1991) In vitro DNA binding activityof Fos/Jun and BZLF1 but not C/EBP is affected by redox changes.Oncogene 6: 1243-1250. PubMed: 1907361.
88. Noguchi C, Noguchi E (2007) Sap1 promotes the association of thereplication fork protection complex with chromatin and is involved in thereplication checkpoint in Schizosaccharomyces pombe. Genetics 175:553-566. doi:10.1534/genetics.106.065334. PubMed: 17151242.
89. Lyne R, Burns G, Mata J, Penkett CJ, Rustici G et al. (2003) Whole-genome microarrays of fission yeast: characteristics, accuracy,reproducibility, and processing of array data. BMC Genomics 4: 27. doi:10.1186/1471-2164-4-27. PubMed: 12854975.
90. Simon P (2003) Q-Gene: processing quantitative real-time RT-PCRdata. Bioinformatics 19: 1439-1440. doi:10.1093/bioinformatics/btg157.PubMed: 12874059.
91. Xhemalce B, Riising EM, Baumann P, Dejean A, Arcangioli B et al.(2007) Role of SUMO in the dynamics of telomere maintenance infission yeast. Retrieved onpublished at whilst December year 1111from Proc Natl Acad Sci. U S A. 104:893-8
92. Thon G, Bjerling P, Bünner CM, Verhein-Hansen J (2002) Expression-state boundaries in the mating-type region in fission yeast. Genetics161:611-22
93. Nielsen IS, Nielsen O, Murray JM, Thon G (2002) The fission yeastubiquitin-conjugating enzymes UbcP3, Ubc15 and Rhp6 affecttranscriptional silencing of the mating-type region. Eukaryot Cell1:613-25
Concerted Action of Ufd1 and STUbLs in the DDR
PLOS ONE | www.plosone.org 15 November 2013 | Volume 8 | Issue 11 | e80442

Gen
Typewritten Text
Figure S1
Gen
Typewritten Text
16

Gen
Typewritten Text
Figure S2
Gen
Typewritten Text
17

A+Zeocin
DIC
Rad22-YFP
No drug
ufd1∆Ct213-342
wild type
+ZeocinNo drug
DIC
Rad22-YFP
0
0,01
0,02
0,03
0,04
0,05 wild typeufd1∆Ct213-342
Rad22-YFP
Rad
22 m
RN
A re
lativ
e to
act
in
Rad22
B
Gen
Typewritten Text
Gen
Typewritten Text
Figure S3
Gen
Typewritten Text
18
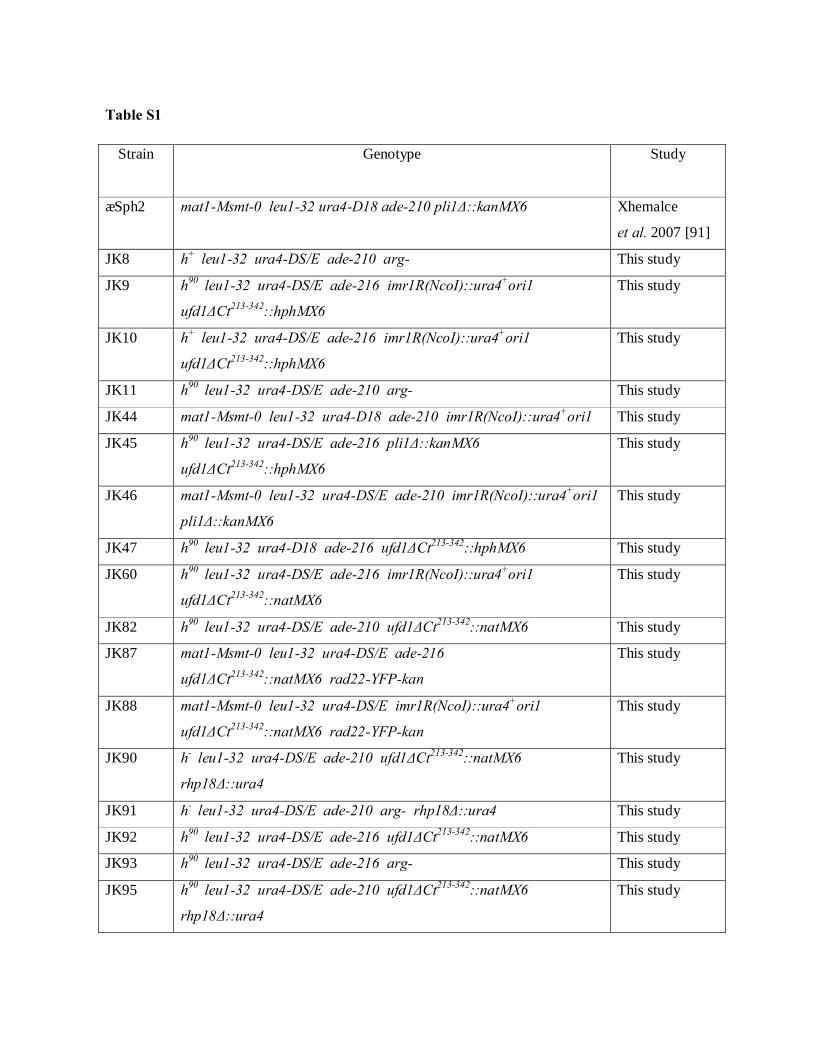
Table S1
Strain
Genotype
Study
∆Sph2 mat1-Msmt-0 leu1-32 ura4-D18 ade-210 pli1Δ::kanMX6
Xhemalce
et al. 2007 [91]
JK8 h+ leu1-32 ura4-DS/E ade-210 arg- This study
JK9 h90 leu1-32 ura4-DS/E ade-216 imr1R(NcoI)::ura4+ori1
ufd1ΔCt213-342::hphMX6
This study
JK10 h+ leu1-32 ura4-DS/E ade-216 imr1R(NcoI)::ura4+ori1
ufd1ΔCt213-342::hphMX6
This study
JK11 h90 leu1-32 ura4-DS/E ade-210 arg- This study
JK44 mat1-Msmt-0 leu1-32 ura4-D18 ade-210 imr1R(NcoI)::ura4+ori1 This study
JK45 h90 leu1-32 ura4-DS/E ade-216 pli1Δ::kanMX6
ufd1ΔCt213-342::hphMX6
This study
JK46 mat1-Msmt-0 leu1-32 ura4-DS/E ade-210 imr1R(NcoI)::ura4+ori1
pli1Δ::kanMX6
This study
JK47 h90 leu1-32 ura4-D18 ade-216 ufd1ΔCt213-342::hphMX6 This study
JK60 h90 leu1-32 ura4-DS/E ade-216 imr1R(NcoI)::ura4+ori1
ufd1ΔCt213-342::natMX6
This study
JK82 h90 leu1-32 ura4-DS/E ade-210 ufd1ΔCt213-342::natMX6 This study
JK87 mat1-Msmt-0 leu1-32 ura4-DS/E ade-216
ufd1ΔCt213-342::natMX6 rad22-YFP-kan
This study
JK88 mat1-Msmt-0 leu1-32 ura4-DS/E imr1R(NcoI)::ura4+ori1
ufd1ΔCt213-342::natMX6 rad22-YFP-kan
This study
JK90 h- leu1-32 ura4-DS/E ade-210 ufd1ΔCt213-342::natMX6
rhp18Δ::ura4
This study
JK91 h- leu1-32 ura4-DS/E ade-210 arg- rhp18Δ::ura4 This study
JK92 h90 leu1-32 ura4-DS/E ade-216 ufd1ΔCt213-342::natMX6 This study
JK93 h90 leu1-32 ura4-DS/E ade-216 arg- This study
JK95 h90 leu1-32 ura4-DS/E ade-210 ufd1ΔCt213-342::natMX6
rhp18Δ::ura4
This study
Gen
Typewritten Text
19
JK97 h90 leu1-32 ura4-DS/E ade-210 ufd1ΔCt213-342::natMX6
rhp18Δ::ura4
This study
JK98 h90 leu1-32 ura4-DS/E ade-216 slx8-1:myc:kanMX6
ufd1ΔCt213-342::natMX6
This study
JK109 h+ ade-210 his3-1 ufd1ΔCt213-342::natMX6 rhp51::his3 This study
JK110 h90 leu1-32 ade-216 his3-1 ufd1ΔCt213-342::natMX6 This study
JK111 h90 leu1-32 ura4-DS/E ade-210 his3-1 This study
JK112 h+ ura4-DS/E ade-216 his3-1 rhp51::his3 This study
JK113 h- leu1-32 ura4-DS/E ade-216 ufd1ΔCt213-342::natMX6 This study
JK114 h90 leu1-32 ura4-DS/E ade-210 arg- This study
JK115 h90 leu1-32 ura4-DS/E ade-210 arg- rqh1Δ::ura4 This study
JK116 h- leu1-32 ura4-DS/E ade-216 ufd1ΔCt213-342::natMX6
rqh1Δ::ura4
This study
JK124 h90 leu1-32 ura4-D18 ade-210/216 pli1Δ::kanMX6
ufd1ΔCt213-342::hphMX6
This study
JK295 h90 leu1-32 ura4 slx8:myc:: natMX6 This study
JK296 h- leu1-32 ura4-D18 slx8-1:myc:kanMX6 rhp18Δ::ura4 This study
JK298 h90 leu1-32 ura4-D18 slx8-1:myc:kanMX6 rhp18Δ::ura4 This study
JK299 h90 leu1-32 ura4-D18 ade-DN/N slx8-1:myc::kanMX6
rhp18Δ::ura4
This study
JK309 h90 leu1-32 ura4 slx8:myc:: natMX6 cdc48-GFP:kanR This study
JK332 h90 ura4-DS/E ade-216 arg- leu1::ufd1-YFP-FLAG2-His6
ufd1Δ::hphMX6
This study
JK333 h90 ura4-DS/E ade-210 arg- leu1::ufd1-YFP-FLAG2-His6
ufd1Δ::hphMX6
This study
JK334 h90 leu1-32 ura4-D18 ade-704 ufd1ΔCt213-342::natMX6
pmt3:pYC11(Leu+)-GFP-pmt3
This study
JK336 h90 leu1-32 ura4-D18 ade-704 pmt3:pYC11(Leu+)-GFP-pmt3 This study
JK339 h90 ura4-DS/E ade-210 arg- pmt3:nat-CFP-pmt3
leu1+::ufd1-YFP-FLAG2-His6 ufd1Δ::hphMX6
This study
JK346 h90 leu1-32 ura4 ade6 ufd1ΔCt213-342::natMX6 ubc13Δ:: ura4 This study
Gen
Typewritten Text
20

JK347 h90 leu1-32 ura4 ade6 ufd1ΔCt213-342::natMX6 ubc13Δ:: ura4 This study
JK349 h90 leu1-32 ura4 ade6 ubc13Δ:: ura4 This study
JK380 mat1-Msmt-0 leu1-32 ura4-DS/E ade-216 ufd1ΔCt213-342::natMX6 This study
FY17243 h- leu1-32 cdc48-GFP:kanR M. Yanagida
NBY1008 h- leu1-32 ura4-D18 slx8-1:myc:kanMX6 Prudden
et al. 2007 [26]
PG1636 h90 leu1-32 ura4-DS/E ade-210 arg- Thon et al. 2002
[92]
PG1637 h90 leu1-32 ura4-DS/E ade-216 arg- Thon et al. 2002
[92]
PG3453 mat1-Msmt-0 leu1-32 ura4-DS/E ade-216 This study
PI131 h90 mat3-M(EcoRV)::ura4 leu1-32 ura4-D18 ade-216 mts3-1 Nielsen et al.
2002 [93]
Y1928 mat1-Msmt-0 leu1-32 ura4-D18 rad22-YFP-kan Noguchi
et al. 2007 [88]
Gen
Typewritten Text
21

22
Table S2 Wild type ufd1∆Ct213-342 Cell cycle stage Number of foci Total nuclei Number of foci Total nuclei S 18 182 41 200 S/G2 33 111 60 187 G2 42 458 127 559 M/G1 0 62 2 36 Cells with 2 or more septa 1 8 9 44 Total all phases 94 821 239 1026

69
Investigation of a putative ”VIM” (VCP-interacting) motif in Slx8 With an interest in the specific physical interactions responsible for proposed coordinated roles of
Cdc48-Ufd1-Npl4 and STUbLs we “searched” for potential binding modules within the S. pombe
STUbL proteins that may contribute to Cdc48-Ufd1-Npl4 targeting. Cdc48/p97 homologs are
known to associate with various co-factors generally characterized by containing one (or sometimes
a few) defined Cdc48/p97 binding motifs (Buchberger, 2010; Yeung et al, 2008). Intriguingly,
visual inspection of the Slx8 STUbL subunit revealed a short amino acid stretch within its N-
terminus conforming to the previously reported minimal “VIM” (VCP-interacting) consensus motif
able to interact with the Cdc48/p97 N-domain (Figure 1A) (Hanzelmann & Schindelin, 2011; Stapf
et al, 2011). Functional and bioinformatic studies have proposed the existence of two general
minimum VIM-subtypes; i.e. VIMs with two flanking arginine’s and two central alanines arranged
with the fixed spacing -RX5AAX2R- and a shorter VIM signature which lacks the N-terminal Arg
(AAX2R). The “RX5AAX2R” VIM type is proposed to mediate a stronger Cdc48 binding affinity
than the “AAX2R” motif. The VIMs are predicted to form α-helical structures thought to bind into a
hydrophobic subdomain grove of the Cdc48/p97 N-terminus (Hanzelmann & Schindelin, 2011;
Stapf et al, 2011). The conserved arginine and alanines making up the VIM consensus motif are
required for Cdc48/p97 binding, at least in investigated cases (Stapf et al, 2011).Thus we sought to
investigate further a putative Slx8 “VIM”-Cdc48 interaction. Inspired by Stapf et al., 2011 we
specifically mutated the two central alanines to leucines (AA32,33LL) and the N-terminal Arg to
alanine (R26A). We have in Køhler et al., 2013 demonstrated the co-purification of Slx8-Myc with
Cdc48-GFP from S. pombe extracts. In line with the predicted “RX5AAX2R” VIM sequence
mediating at least part of this interaction, co-immunoprecipitation of Slx8AA32,33LL and a triple
Slx8R26A; AA32,33LL mutant protein with GFP-tagged Cdc48 was significantly reduced compared to
the wild-type protein (Figure 1B). Though, as residual interaction could still be observed, it
suggests that other undefined binding modules within Slx8, or possibly indirect associations
mediated through other common interactors, are sufficient for some Cdc48 binding. It is also
possible that the Slx8R26A; AA32,33LL mutations may not be sufficient for complete abrogation of the
VIM binding surface. For instance it has been proposed that although VIM containing proteins
bind to the same surface on Cdc48/p97, variations in their amino acid composition mediates slightly
different kinds and strengths of Cdc48/p97 binding (Hanzelmann & Schindelin, 2011). Further
investigation of the slx8AA32,33LL and slx8R26A; AA32,33LL mutants did not reveal any obvious growth
inhibition nor sensitivity to increased temperature or DNA damaging stress (data not shown). Thus

70
the observed reduced physical association of Slx8”VIM” mutants with Cdc48 does not seem to
compromise overall Slx8 and/or Cdc48 function. Whether the “RX5AAX2R” sequence is directly
involved in complex formation with Cdc48 will need further direct physical evidence. In any case,
we speculate that the Slx8 “VIM” domain may be enhancing associations of the Slx8/Rfp STUbL
dimer with the Cdc48-Ufd1-Npl4 complex to ensure a dynamic regulation of STUbL targets. The
Ufd1-Npl4 sub-complex also associates with a Cdc48/p97 N-terminal region. Notably, even though
only one Ufd1-Npl4 hetero-dimer is proposed to bind per Cdc48/p97 hexamer (Pye et al, 2007), the
association of Ufd1-Npl4 with Cdc48/p97 seems to be mutually exclusive with some VIM-
containing proteins (Ballar et al, 2006; Heo et al, 2010). The molecular reason for this remains
unknown but it may suggest that a putative Slx8”VIM” -Cdc48 interaction may also be mutually
exclusive with Ufd1-Npl4 binding. It could be speculated that flexible and dynamic associations of
the Ufd1-Npl4 dimer and STUbLs with the same Cdc48 hexamer may be underlying a coordinated
role in the processing of SUMO targets. More specifically, it could be speculated that the
Slx8”VIM” may facilitate initiate Cdc48 recruitment to relevant STUbL substrates. The subsequent
exchange of Slx8 with the Ufd1-Npl4 dimer on Cdc48 could then allow efficient Cdc48
“anchoring” of STUbL co-modified species via the Ufd1 SUMO and ubiquitin binding domains
(Nie et al, 2012). An alternative order of events is also conceivable. A dynamic intermediate in this
pathway may be interactions between the Ufd1-Npl4 dimer and STUbLs themselves. In particular
we detected interactions between the Ufd1 C-terminus and the Rfp1 STUbL subunit both in two-
hybrid screens and in in vitro GST pulldown assays (Kohler et al, 2013). The two-hybrid interaction
was found to be dependent on the Ufd1 SIM (SUMO-interaction motif) (Kohler et al, 2013; Nie et
al, 2012), indicating that it is indirect through SUMO. It could be speculated that such indirect
Ufd1-STUbL interaction through SUMO may be implicated in dynamic substrate “hand-over” in
vivo. Certainly, deeper insight into the interaction surfaces mediating proposed cooperative roles of
Cdc48-Ufd1-Npl4 and STUbLs should elucidate how such roles are coordinated.
Visual inspection of STUbL homologs in a few other organisms did not indicate conservation of the
Slx8 “VIM”. Though, the closely related Schizosaccharomyces japonicas Slx8 protein did display a
short “VIM” version AAX2R, also in its N-terminus (from amino acid position 88-92 “AAEMR”),
suggesting putative conservation within the Schizosaccharomyces lineage. Although functionally
similar, the STUbLs of fission and budding yeast and mammals diverge significantly in their amino
acid sequence (Sun et al, 2007) and thus it may not be surprising not to find conservation of the

71
small putative Slx8 “VIM” motif (Neduva & Russell, 2005). Though, it is possible that alternative
binding modes may have evolved in other organisms to allow coupling of STUbL and Cdc48/p97
activities.
Figure 1. Mutation of predicted Slx8 “VIM” residues reduces the physical association between Slx8 and Cdc48 in S.
pombe cells. (A) A minimum core VIM (VCP-interacting) motif of “RX5AAX2R” can be predicted in the S. pombe
Slx8 N-terminus (position A26 to R36). The Slx8 N-terminal sequence (amino acid 1 to 50) is shown. (B).
Immunoprecipitation of Cdc48-GFP with either Slx8-Myc wild type or “RX5AAX2R” mutated proteins.
Immunoprecipitated material was analyzed by Western blotting either with an anti-GFP or an anti-Myc antibody. The
Slx8-Myc inputs are shown. Some degradation of Slx8-Myc tagged proteins may result in the detection of lower
molecular weight Slx8-Myc species.

72
Construction of slx8 “VIM” mutant strains slx8-Myc tagged “VIM” mutants were constructed using standard genetic procedures. slx8+ 5’ and
3’ regions were amplified in two separate PCR reactions from genomic DNA of an slx8-
Myc:KanMX6 tagged strain where the KanMX6 resistance cassette had been changed into NatMX6
(Prudden et al, 2007). Primers were designed to perform specific point mutations into the slx8 N-
terminus. For amplification of the slx8+ 5’ region the primers GTO-536
(5’CGATATGCATCAATAATGTCG GTATCCATGC TTCC 3’) and GTO-537 were used (5’
CATTTCCAGCTGCGACTCTAGCGGAT 3’). To introduce the slx8AA32,33LL mutation, a forward
primer (GTO-540) was used changing the two alanine codons (GCA GCA) to leucines (TTA CTT)
(5’CTAGAGTCGCAGCTGGA AATTTACTTATAAATA 3’) together with a the reverse primer
(5’ TGTTTACATGTTGGAATACAAGAC 3’) located in the slx8+ 3’ region. The 5’ and 3’
fragments were cloned into the pJET1.2 blunt end vector and ligated together using an internal
PvuII site in the slx8+ 5’ region (to yield plasmid pJBK135). To also generate the R26A mutation,
the slx8+ 5’ region was amplified with the same GTO-536 primer as above together with the primer
GTO-538 (5’ CATTTCCAGCTGCGACTGCAGCGGAT 3’), changing the arginine codon “AGA”
to alanine “GCA”. The resulting fragment was ligated together with the 3’ slx8+ fragment stated
above (plasmid pJBK136). Resulting plasmids were linearized with PciI + NsiI and transformed
into S. pombe cells. Integration of point mutations were verified by PCR and sequencing.
GFP-immunoprecipitations of Cdc48-GFP from Slx8 wild type and “VIM” mutant Myc-tagged
strains were essentially performed as described in Køhler et al., 2013.
Strain table
Strain name Genotype
JK360 h90 leu32-1 ura4-DS/E slx8:myc:natMX6
JK362 h90 leu32-1 ura4-DS/E slx8(AA32,33LL):myc:natMX6
JK364 h90 leu32-1 ura4-DS/E slx8:myc:natMX6 cdc48-GFP:kanR
JK371 h90 leu32-1 ura4-DS/E slx8(R26A;AA32,33LL):myc:natMX6
JK372 h90 leu32-1 ura4-DS/E slx8(R26A;AA32,33LL):myc:natMX6 cdc48-GFP:kanR
JK375 h90 leu32-1 ura4-DS/E slx8(AA32,33LL):myc:natMX6 cdc48-GFP:kanR

73
Manuscript: Quantitative Identification of Sumoylation Sites in Fission Yeast Wild-type Cells, Ufd1 and SUMO-targeted ubiquitin Ligase (STUbL) Mutants Julie Bonne Køhler1*, Triin Tammsalu2*, Maria Mønster Jørgensen1, Ron Hay2, Geneviève Thon1
1 Department of Biology, University of Copenhagen, DK-2200 Copenhagen, Denmark 2 Centre for Gene Regulation and Expression, Sir Jamens Black Centre, College of Life Sciences, University
of Dundee, Dow Street, Dundee DD1 5EH, UK
The authors with * contributed equally to this work

74
ABSTRACT The posttranslational modification of proteins by the small ubiquitin-like modifier SUMO regulates
a wide range of processes in all eukaryotic cells. In these regulations, SUMO conjugates can be
desumoylated or they can be directed for degradation by SUMO-targeted ubiquitin ligases
(STUbLs). A recently identified player in their processing is the ubiquitin-selective chaperone
Cdc48/p97-Ufd1-Npl4, which might act in STUbL-dependent or independent pathways. While
proteomic analyses have identified hundreds of sumoylated proteins, pinpointing the effects of
sumoylation at the molecular level remains a major challenge to our understanding of SUMO
functions. Here, we report a comprehensive, proteome-wide, identification of sumoylated residues
in fission yeast. Their identification exploited a genetically engineered SUMO variant (His6-
SUMOL109K) expressed at endogenous levels and amenable to a newly developed two-step
purification strategy followed by mass spectrometry. This novel purification scheme was combined
with SILAC-based quantitative proteomics to compare endogenous sumoylation at the resolution of
single residues in wild-type and mutant strains deficient in either STUbL (slx8-1) or Ufd1
(ufd1ΔCt213-342) function. Over 900 sumoylated lysines were identified in a total of 431 proteins, of
which 178 distinctly modified peptides were quantified in both mutant backgrounds. This
quantification revealed a number of proteins whose sumoylation was increased either specifically in
the slx8-1 or ufd1ΔCt213-342 mutant or in a coordinated manner in both mutants. Hence, in addition
to uncovering STUbL and Ufd1 candidate substrates, our findings shed light on potential common
SUMO substrates on which STUbL and Ufd1 (Cdc48-Ufd1-Npl4) activities may impinge.
Together the datasets we present constitute a rich, comprehensive resource for guiding future
endeavors in understanding SUMO function and dynamics.

75
Introduction The small ubiquitin-like modifier SUMO can be conjugated to hundreds of protein species in
eukaryotic cells (Flotho & Melchior, 2013; Wilkinson & Henley, 2010). SUMO is thereby believed
to participate in numerous regulations, even though the molecular details of the regulatory
mechanisms remain in most cases unknown. Prior to conjugation, SUMO is processed at its C-
terminus to expose a conserved diglycine motif (diGly). Substrate conjugation is then catalyzed by
the sequential action of an E1 activating, an E2 conjugating and an E3 ligating enzyme which
ultimately leads to the formation of an isopeptide bond between the glycine carboxyl group of
SUMO and the ɛ-amino-group of a substrate lysine. Self-modification of SUMO through internal
lysine residues can furthermore result in the formation of polymeric SUMO chains (Skilton et al,
2009; Tatham et al, 2001). The fission yeast sumoylating enzymes comprise the E1 heterodimer
Rad31/Fub2 (Aos1/Uba2), the E2 enzyme Ubc9 (also known as Hus5) and two identified E3s of the
SP-RING family; i.e. the PIAS homolog Pli1 and the Mms21 homolog Nse2 (Andrews et al, 2005;
Ho et al, 2001; Watts et al, 2007; Xhemalce et al, 2004). A fundamental aspect of sumoylation is its
reversible nature. Desumoylation is carried out by SUMO specific proteases (Ulp/SENP proteins)
that by hydrolyzing SUMO isopeptide bonds allow proteins to return to their non-sumoylated form
(Hickey et al, 2012).
An alternative to desumoylation is the elimination of sumoylated proteins through the STUbL-
pathway. STUbLs (SUMO-targeted ubiquitin ligases), as represented by the Rfp1/Slx8 and
Rfp2/Slx8 dimers in S. pombe, the Slx5/Slx8 dimer in S. cerevisiae, and homo-dimeric RNF4 in
mammals, are conserved ubiquitin E3 enzymes that can target sumoylated proteins for degradation
by the proteasome (Kosoy et al, 2007; Prudden et al, 2007; Sun et al, 2007; Tatham et al, 2008;
Uzunova et al, 2007). The activity of STUbLs is determined by a C-terminal RING domain and
several N-terminal SUMO-interaction motifs (SIMs) that bind poly-sumoylated species
(Keusekotten et al, 2014; Rojas-Fernandez et al, 2014; Tatham et al, 2008). The importance of
RNF4 STUbL homologues in maintaining SUMO homeostasis is revealed by the accumulation of
high molecular weight SUMO conjugates in STUbL deficient cells - a phenotype associated with
genomic instability and general cellular dysfunction (Kosoy et al, 2007; Prudden et al, 2007; Yin et
al, 2012). The best described STUbL substrate to date is the mammalian PML protein (Lallemand-
Breitenbach et al, 2008; Tatham et al, 2008). Other reported STUbL substrates include the
transcriptional regulator Mot1(Wang & Prelich, 2009), the inner kinetochore protein CENP1

76
(Mukhopadhyay et al, 2010), the DNA damage protein MDC1 (Galanty et al, 2012; Yin et al,
2012), the cohesin α-kleisin subunit Mcd1 (D'Ambrosio & Lavoie, 2014) and the SUMO ligase
Siz1 (Westerbeck et al, 2014). Though given the extensive number of SUMO targets in the cell,
many more proteins are likely to be affected by STUbL activity. Indeed, STUbLs have been
detected at centromeres, nuclear pores, replication forks and at sites of DNA damage (Burgess et al,
2007; Cook et al, 2009; Nagai et al, 2008; van de Pasch et al, 2013; Yin et al, 2012).
A recently identified factor also participating in the processing of sumoylated proteins is the
conserved Cdc48-Ufd1-Npl4 segregase complex (Bergink et al, 2013; Kohler et al, 2013; Nie et al,
2012). Cdc48-Ufd1-Npl4 has well-documented roles in several ubiquitin-related processes. By
coupling the ubiquitin binding properties of Ufd1-Npl4 with the ATPase activity of Cdc48 (p97 in
mammals), Cdc48-Ufd1-Npl4 is believed to aid in the extraction of ubiquitylated proteins from
higher order complexes (Meyer et al, 2012); (Baek et al, 2013). Identification of a SIM motif in
both fission and budding yeast Ufd1 revealed that Cdc48-Ufd1-Npl4 is in addition able to recognize
sumoylated proteins (Bergink et al, 2013; Nie et al, 2012), suggesting the complex might bind
protein species co-modified by SUMO and ubiquitin, such as produced by STUbLs (Nie et al,
2012). The existence of both physical and functional interactions between Cdc48, Ufd1 and
STUbLs supports the idea that they perform coordinated tasks in a shared pathway (Bergink et al,
2013; Kohler et al, 2013). Cdc48-Ufd1-Npl4 may help mobilize STUbL substrates to promote their
further processing, either by the proteasome or, alternatively, by demodifying enzymes. Consistent
with this model, Ufd1 mutants, similar to STUbL mutants, accumulate sumoylated proteins that
coalesce in sub-nuclear foci (Kohler et al, 2013; Nie et al, 2012). However, despite evidence
suggesting a cooperative function, details of the substrates affected by a common STUbL/Cdc48-
Ufd1-Npl4 pathway and the extent to which the two activities are coupled remain to be determined.
Mass spectrometry (MS)-based proteomics has become a widely used tool to study protein
sumoylation (Filosa et al, 2013). However, while hundreds of putative SUMO substrates have been
identified by various proteomic studies (Becker et al, 2013; Bruderer et al, 2011; Golebiowski et al,
2009; Hannich et al, 2005; Matic et al, 2010; Schimmel et al, 2014; Tatham et al, 2011;
Wohlschlegel et al, 2004), mapping of the specific modification sites at a large scale has been much
less efficient. Such information is nonetheless crucial in order to address the biological outcomes of
individual sumoylation events. Identification of sumoylation sites has traditionally been hampered
by an inability to enrich for SUMO modifications at a peptide level. Instead, protein level

77
purifications produces highly complex peptide mixtures from which site-specific information is
hard to extract. Moreover, the long SUMO branch left on modified peptides after trypsin cleavage
gives rise to MS/MS spectra that are difficult to interpret (Wohlschlegel et al, 2006). Here, we made
use of a newly designed two-step enrichment strategy that overcomes these obstacles and permits
site-specific identification of SUMO targets at a proteome-wide scale (Tammsalu et al, 2014). The
method relies on a His6-tagged version of SUMO engineered to contain a lysine residue
immediately upstream its C-terminal diGly. Digestion of Ni2+-purified His6-SUMOL109K-conjugates
with the lysine-specific endoproteinase Lys-C produces diGly-modified peptides that can be
isolated with a diGly-Lys-specific antibody (Tammsalu et al, 2014; Xu et al, 2010). In combination
with quantitative proteomics, this strategy enabled us to identify more than 900 sumoylated lysines
and to compare sumoylation levels for a subset of these between fission yeast wild type and mutant
strains deficient either in STUbL (slx8-1) or Ufd1 (ufd1ΔCt213-342) function.
Results and Discussion Genetic engineering of a His6-SUMOL109K variant for the identification of sumoylation sites from complex protein mixtures The identification of sumoylated proteins, and in particular of their modification sites, is challenged
by the relatively low abundance of these species in cell lysates and by the complexity of peptide
mixtures derived from protein level purifications of SUMO conjugates. In order to specifically
enrich sumoylated peptides from S. pombe cells, we replaced the endogenous SUMO gene (pmt3)
with a His6-tagged version in which the leucine at position 109 was changed to a lysine (His6-
SUMOL109K allele). A lysine at position 109 will upon digestion with the endoproteinase Lys-C
leave a diGly-remnant on SUMO acceptor sites suitable for the purification of modified peptides
with a diGly-Lys specific antibody (anti-k-ε-GG) (Tammsalu et al, 2014). The diGly-Lys-antibody
was originally developed for the enrichment of ubiquitylated peptides for which a diGly-remnant is
left upon trypsin digest (Wagner et al, 2011; Xu et al, 2010). Importantly, as ubiquitin contains an
arginine at the position upstream its diglycine sequence, Lys-C will not cleave at this site. Similarly,
Lys-C digestion of proteins conjugated to other ubiquitin-like proteins (Ubls) generates long
peptide-branches not recognized by the diGly-Lys-antibody (Figure 1A). Thus, Lys-C digestion of
Ni2+-purified His6-SUMOL109K-conjugates allows specific enrichment of SUMO diGly-modified
peptides to facilitate MS analysis of the diGly-modified lysines based on their unique mass-to-
charge ratio. This strategy has recently been successful in identifying more than a thousand

78
sumoylation sites in heat shock treated human cells using a similar C-terminal SUMO mutant (His6-
SUMOThr90K) (Tammsalu et al, 2014).
Examinaton of the His6-SUMOL109K strain revealed cells with a wild-type morphology and a
generation time similar to an isogenic wild-type strain (Figure 1B). The His6-SUMOL109K mutation
was crossed into a slx8-1 STUbL mutant background (Prudden et al, 2007) and into a ufd1ΔCt213-342
mutant background where the C-terminal half of Ufd1 is deleted (Kohler et al, 2013). There again,
growth analysis showed that slx8-1 and ufd1ΔCt213-342 His6-SUMOL109K cells were phenotypically
similar to their wild-type SUMO counterparts (Figure S1). We next analyzed the effect of the
SUMOL109K mutation on cellular SUMO conjugates by Western blotting (Figure 1C). Compared to
the conjugates produced by wild-type SUMO we detected a similar pattern of bands in the His6-
SUMOL109K strains, suggesting that the same spectrum of proteins is modified by the two SUMO
versions . The overall conjugation levels for the His6-SUMOL109K strains was though slightly
reduced, suggesting that the His6-SUMOL109K mutant is somewhat less efficiently conjugated to its
targets than wild-type SUMO, as proposed for the analogous S. cerevisae SUMOI96K mutant
(Wohlschlegel et al, 2006). Though, as the SUMOL109K mutation did not seem to affect overall
sumoylation patterns, or compromise cell growth, we sought to use the His6-SUMOL109K mutant to
globally map the sumoylation sites of fission yeast wild-type and mutant cells.
Comparative proteome-wide identification of sumoylation sites in wild-type cells, Ufd1 and STUbL mutants, using the His6-SUMOL109K variant and SILAC Sumoylated proteins accumulate in Ufd1 mutants (Kohler et al, 2013; Nie et al, 2012). To start
identifying these species we first isolated total His6-SUMOL109K-modified peptides from ufd1ΔCt213-
342 cells. Cells were lysed under denaturing conditions and His6-SUMOL109K conjugates were
isolated by Ni2+-chromatography. The purified material was digested with Lys-C, or Lys-C and
Glu-C to generate shorter peptides, and diGly-Lys modified peptides were immunoprecipitated
(IP’ed) with an anti-K-ε-GG antibody prior to MS analysis (Figure 2A). This workflow identified a
total of 749 unique diGly-modified lysines mapped with a localization probability of >0.9. Four
additional sites were identified with localization probabilities below 0.9. Together these sumoylated
sites were distributed between a total of 376 proteins (Table S1).
We next set out to quantitatively compare sumoylation in wild type and mutant strains deficient
either in Ufd1 or STUbL (Slx8) function. For this we employed His6-SUMOL109K variant in
conjunction with SILAC-based (stable isotope labeling by amino acids in cell culture) proteomics

79
(Mann, 2006) to analyze site-specific differences in sumoylation between wild type, slx8-1 and
ufd1ΔCt213-342 backgrounds. His6-SUMOL109K lysine auxotrophic strains were grown in medium
supplemented with either light (Lys0) or heavy (Lys8) lysine. Two parallel SILAC experiments
were performed in which wild-type and ufd1ΔCt213-342 strains, each grown in light (L) lysine
conditions, were individually compared to heavily (H) labeled slx8-1 cultures (Figure 2B). The
parallel experiments were repeated three times starting from isolated colonies. Heavy and light
cultures propagated at 30°C were shifted to 33°C for 12 hr to induce the slx8-1 temperature-
sensitive phenotype at the semi-permissive temperature of slx8-1 (Prudden et al, 2007). The slx8-1
cultures were split in two and mixed in 1:1 ratios with wild type and ufd1ΔCt213-342, respectively.
Small aliquots of cell lysate obtained from each mixture were subjected to in-solution digestion
with Lys-C for total proteome analysis, while His6-SUMOL109K conjugated peptides were isolated as
above from the remaining material. Combined, these SILAC experiments detected a total of 564
different diGly-Lys modified peptides, representing 538 distinct sumoylation sites in 256 proteins.
351 of these sites (or 201 proteins) had also been identified in the ufd1ΔCt213-342 His6-SUMOL109K
pilot IP performed under non-SILAC conditions, indicating a high degree of experimental
reproducibility (Figure 2C). Even though approximately the same amount of starting cell material
was used in both experiments, fewer sites were identified in each SILAC IP than in the non-SILAC
experiment. This may be a consequence of the higher complexity of SILAC peptide samples where
very intense SILAC pairs can hinder detection of lower abundant ones. Together the SILAC and
non-SILAC experiments produced a comprehensive list of a total of 937 distinct sumoylation sites
covering 422 unique proteins (Table S1).
In agreement with proteins commonly displaying more than one sumoylation site (Tammsalu et al,
2014), we found that around half of the proteins in the list were modified at multiple sites by the
SUMOL109K moiety. Some proteins contained extensive numbers of sites with for example whole 22
different sites identified in Top2. The clustering of some of these sites suggests that some protein
regions are more susceptible to modification and/or that some lysines can be redundantly modified
(Tammsalu et al, 2014). This is in line with the lack of phenotypes often observed for single SUMO
site mutants (McAleenan et al, 2012; Psakhye & Jentsch, 2012). Demonstrating that at least in some
cases proteins are simultaneously modified by several SUMOs on distinct lysines, we were able to
detect a number of doubly or even, in a few cases, triply diGly-Lys modified peptides. Such SUMO
co-modification was detected for 18 different proteins suggesting that SUMO multisite modification
may be fairly common. In addition to diGly-doubly modified peptides we were also able to detect

80
sumoylated peptides co-modified with other posttranslational modifications. These included
peptides of SUMO itself revealing phosphorylation on Thr20 and Thr24. Phosphorylation of other
SUMO residues in this part of the protein has previously been reported (Skilton et al, 2009),
suggesting that the fission yeast SUMO N-terminus is subject to profound phosphor-regulation. S.
pombe SUMO primarily forms chains via the two lysines K14 and K30, although other lysines can
also serve as acceptors (Prudden et al, 2011; Skilton et al, 2009).Various SUMO-SUMO linkages
were confirmed in our experiments. Notably, we also detected SUMO modification of ubiquitin,
both on K48 and K63. Conjugation of SUMO to ubiquitin has similarly been detected for the
human wild-type SUMO-2 protein (Tammsalu et al, 2014). These findings reveal the existence of
so far unexplored types of SUMO and ubiquitin polymeric species.
A sequence motif analysis was performed for all digly-Lys-containing peptides identified in the
SILAC experiments, using the pLogo software (O'Shea et al, 2013). Similar to a recent study
(Tammsalu et al, 2014) this analysis revealed sumoylation sites to preferentially conform to either
the forward (ᴪKxD/E) (Rodriguez et al, 2001) or to the reversed consensus motifs (D/ExKᴪ) (Matic
et al, 2010). For forward sites, we detected a strong preference for a Glu at the second position C-
terminal to the modified Lys (+2 position), and of a large hydrophobic residue, especially a Val, and
to a lesser extent an Ile, at position -1 (Figure 2D). Sumoylation sites lying within the inverted
consensus motif did not display any preference for a hydrophobic residue at their +1 position but
they were significantly prone to containing either a Glu or Asp at position -2, with a slight
preference for Glu.
Functional clustering of SUMO targets Gene Ontology (GO) analysis of the ensemble of identified SUMO targets revealed significant
enrichment for proteins involved in biological processes connected to chromosome organization
and segregation, chromatin modification and transcription as well as DNA recombination and
repair. Also enriched were proteins associated with ribosome biogenesis and nucleo-cytoplasmic
transport (Figure 3A). These observations are generally consistent with the literature on SUMO
(Golebiowski et al, 2009; Makhnevych et al, 2009; Wohlschlegel et al, 2004) and with the
predominant nuclear localization of SUMO (Tanaka et al, 1999).
Expanding on previous large-scale studies, the total list of SUMO substrates (Table S1) identified in
our experiments revealed clusters of proteins forming parts of common macromolecular complexes
or biological pathways (Golebiowski et al, 2009; Psakhye & Jentsch, 2012; Tammsalu et al, 2014;

81
Wohlschlegel et al, 2004). For example, we found significant enrichments of components of the
SWI/SNF chromatin-remodeling complex, of the histone acetylase SAGA complex and of the
TFIID transcription factor and RNA polymerase core complexes, supporting a conserved role for
SUMO in the regulation of these protein groups (Wohlschlegel et al, 2004). Other complexes that
appeared to be highly influenced by sumoylation were the core cohesion and condensin complexes,
with the latter being modified on all five components and holding a total of 16 sumoylation sites.
The regulation of chromosome cohesion and condensation by SUMO is also in line with the
literature (Watts, 2007). Chromatin immunoprecipiation of SUMO in S. pombe cells has revealed
local enrichment of SUMO in centromeric and telomeric regions (Watts, 2007). As this enrichment
is dependent on a functional Ubc9, SUMO is believed to concentrate in these regions in a
conjugated form. Enrichment of SUMO at centromeres throughout the cell cycle can also be
observed by fluorescence microscopy (Tanaka et al, 1999). Our experiments provide an exquisitely
detailed view of these modifications at the molecular level, revealing extensive numbers of
sumoylated sites in telomere sheltherin factors, core centromere and kinetochore proteins as well as
in proteins involved in the anchoring of these chromosome domains to the nuclear envelope (NE).
The abundance of sumoylation events at telomeric and centromeric regions is consistent with these
domains undergoing dynamic changes throughout the cell cycle, mediated by a plethora of
regulated protein interactions (Fujita et al, 2012; Funabiki et al, 1993; Hediger et al, 2002; Hou et
al, 2013). Centromeres and telomeres are packaged into heterochromatin characterized by specific
histone modifications and associated proteins. Of interest, we mapped multiple sumoylation sites in
histone-modification writers and readers that had not been identified in a previous study (Shin et al,
2005).
Another nuclear region in which SUMO has been proposed to play pivotal roles is the nucleolus
which forms the compartment where ribosomes are pre-assembled. Here, SUMO is proposed to
contribute to several aspects of nucleolar functions, including rRNA processing, ribosome assembly
and overall rDNA organization (Finkbeiner et al, 2011; Panse et al, 2006; Srikumar et al, 2013a;
Srikumar et al, 2013b). In agreement with these views and with other high-throughput studies our
list of SUMO targets includes a number of nucleolar proteins, in particular multiple components of
the U3 sno-RNA-containing complex (Westman et al, 2010). Together these findings highlight the
extensive regulation of higher order protein complexes by SUMO and the widespread links between
SUMO and chromosome transactions.

82
Shared effects of Ufd1 and STUbLs on the sumoylated proteome Of all the diGly-Lys-containing peptides identified in the SILAC experiments, 547 were quantified
in at least one biological replicate based on a minimum of two SILAC ratio counts while a few were
only detected in either heavy or light conditions. However, only peptides for which SILAC ratios
were measured in at least two biological replicates were used for further quantification. The same
criterion was applied to the total proteome, resulting in the quantification of 1507 different proteins
(Table S3). SILAC ratios for “crude” proteins were normalized using the MaxQuant software (Cox
& Mann, 2008), while the mean of their non-normalized ratios was used to normalize the ratios
from the purified material (Tatham et al, 2011). Normalized SILAC ratios from at least two
replicates were averaged yielding the quantification of 248 and 267 diGly-Lys modified peptides in
the slx8-1/wild type and slx8-1/ufd1ΔCt213-342 IPs, respectively (Table S2). As 178 of these peptides
were quantified in the two parallel experiments their relative abundance between ufd1ΔCt213-342 and
wild type could also be calculated. Normalized ratios were transformed to log2 values and log2
ratios obtained for slx8-1/wild type and ufd1ΔCt213-342/ wild type were plotted against each other to
provide a triple SILAC map (tsMAP) (Figure 4B) (Golebiowski et al, 2009). Peptides that were
only quantified in one of the parallel SILAC IP experiments did not give rise to a coordinate on the
tsMAP. With a significance criterion of a log2 ratio ≥1.0 (i.e. an at least two-fold change), we
identified 57 sites (in 33 proteins) of increased sumoylation in the slx8-1 mutant and 30 upregulated
sites (in 24 proteins) in the ufd1ΔCt213-342 mutant, representing proteins involved in different cellular
pathways. Applying a regression analysis on the two data sets we found that they correlated with a
Pearson coefficient of r=0.48. Even with a great degree of variation, this supports some common
tendencies in sumoylation dynamics in the two strains. In total, 17 sites distributed between 11
different proteins were found to be significantly upregulated in both mutant backgrounds.
Conversely, approximately 20 sites were significantly downregulated (log2≤-1) in each of the
mutants, with some being downregulated in both. Thus, while there is a general tendency for protein
sumoylation to increase in slx8-1 and ufd1ΔCt213-342 backgrounds (Kohler et al, 2013; Prudden et al,
2007) this study), this seems to be accompanied by a decrease in the modification state of other
proteins, perhaps due to competition for a limited pool of free SUMO. Proteins with sites that
showed an at least two-fold upregulation in both mutant backgrounds are highlighted in grey in
Table S2. In accord with STUbLs promoting the turnover of polysumoylated proteins (Rojas-
Fernandez et al, 2014), diGly-modified peptides of SUMO itself constituted the most upregulated
species in the slx8-1 strain (up to more than a ten-fold increase). SUMO-SUMO linkages were also

83
upregulated in ufd1ΔCt213-342, albeit to a lesser extent, supporting the notion that Ufd1 contributes to
their downregulation (Nie et al, 2012). Interestingly, one of the most upregulated SUMO peptides in
slx8-1 and ufd1ΔCt213-342 cells was one co-modified on both K14 and K30. DiGly-modification of
alternative SUMO chain acceptors K39, K51, K60 and K63 were also generally upregulated in the
two mutants, indicating that they accumulate diverse kinds of SUMO branches.
Notably, among the proteins with increased sumoylation in both mutant strains was the
transcriptional regulator Mot1 (TATA-binding protein associated factor), co-modified on K101 and
K105 (log2=1.2 in ufd1ΔCt213-342 or log2= 0.9 in slx8-1). As S. cerevisae Mot1 has previously been
established to undergo protein quality control through a SUMO and STUbL-dependent proteolytic
pathway (Wang & Prelich, 2009), this seems to support the validity of our experimental design.
Sumoylation of the kinetochore associated proteins Mis17 and Ask1 was also increased in both the
slx8-1 and ufd1ΔCt213-342 mutants. Mis17 associates with the inner kinetochore plate together with
the Mis6-Mal2-Sim4 complex (CENP-I/-O/-K homologs) throughout the cell cycle (Shiroiwa et al,
2011) while Ask1 forms part the DASH complex that binds the outer kinetochore only during
mitosis (Liu et al, 2005; Sanchez-Perez et al, 2005). Another DASH component, Dam1, was also
increasingly sumoylated in ufd1ΔCt213-342 (log2 ratio=1.2) and to a lesser degree in slx8-1 (log2
ratio=0.9). The idea that STUbLs function at centromeres/kinetochores was suggested by studies in
S. cerevisae, showing local enrichment of the Slx5 STUbL subunit at the core centromeric region
(van de Pasch et al, 2013). Moreover, in human cells, RNF4-mediated turnover of the inner
kinetochore homolog CENP-I appears to help coordinate proper formation of the inner kinetochore
plate (Mukhopadhyay et al, 2010). Our finding of elevated Mis17 sumoylation in STUbL and Ufd1
mutant cells could indicate a conserved role for STUbLs and Ufd1 in regulating inner kinetochore
structures. STUbL/Ufd1-mediated turnover of outer kinetochore components may similarly
contribute to normal spindle dynamics, possibly explaining some of the mitotic defects observed in
STUbL mutant cells (van de Pasch et al, 2013).
Telomere-associated proteins constituted other potential common Slx8/Ufd1 substrates identified in
our SILAC screen. In particular, sumoylation of the telomere sheltherin proteins Rap1 (on K344)
and Ccq1 (on K56, K78, K491) were increased in the range of 3 to 6 fold in the mutants. Telomeric
shelterin protects telomeric ends and it regulates their replication by telomerase. In S. cerevisae,
polysumoylated Rap1 is downregulated by Uls1- a dual STUbL and DNA-dependent
ATPase/translocase. By clearing non-functional polysumoylated versions of Rap1 from telomeres,
Uls1 helps sustain Rap1-dependent inhibition of telomeric end-to-end fusions (Lescasse et al,

84
2013). No direct Uls1 STUbL homolog exists in S. pombe. It is thus tempting to speculate that a
similar function could be carried out by the combination of Slx8/Rfp STUbL and Cdc48-Ufd1-
Npl4-dependent APTase activity.
Important for proper telomere function and regulation is their localized clustering at the nuclear
periphery in interphase cells (Chikashige et al, 2009; Ferreira et al, 2011; Hediger et al, 2002;
Schober et al, 2009). During mitosis these associations need to be released in order to allow faithful
chromosome segregation. In S. pombe the association of telomeres with the nuclear envelope (NE)
involves interactions between telomeric sheltherin (Taz1 and Rap1) proteins and the inner nuclear
membrane factors Bqt3 and Bqt4 (Chikashige et al, 2009; Fujita et al, 2012). Other factors more
recently implicated in telomere-NE anchoring include the distantly related integral nuclear
membrane proteins of the animal LEM (Lap2/Emerin/Man1) subfamily of lamin–associated
proteins, Lem2 and Man1 (Gonzalez et al, 2012; Steglich et al, 2012). Interestingly, we detected a
greater than 4 fold increase in the sumoylation of Lem2 at K121, a lysine situated in the chromatin
binding Helix-Extension-Helix (HEH) domain of Lem2, in the ufd1ΔCt213-342 background. Lem2
K121 sumoylation was not increased in slx8-1 (log2= -0.8). Although quantitative information was
only obtained for K121, we detected an additional five Lem2 sumoylation sites (all located within
the HEH region), specific to the ufd1ΔCt213-342 strain, from all our experiments combined (Table
S1). In addition, a total of nine sumoylation sites were identified in Bqt4, also solely detected in
ufd1ΔCt213-342. Telomere anchoring to the nuclear periphery has been shown to depend on
sumoylation in S. cerevisae and C. elegans but the targets of sumoylation have only been poorly
defined (Ferreira et al, 2011; Ferreira et al, 2013). Our findings propose undiscovered modes of
telomere-NE regulation through sumoylation of inner membrane proteins. Such regulation is likely
to also apply to other heterochromatic regions associating with the NE, such as centromeres. For
instance, Lem2 is highly enriched at the centromere/spindle pole body region (Gonzalez et al, 2012)
and we detected several sumoylation sites on the two centromere NE anchoring proteins Sad1 (SUN
domain protein) and Csi1. Again, Sad1 and Csi1 sumoylated peptides were almost exclusively
detected in the ufd1ΔCt213-342 background. Since the Cdc48-Ufd1-Npl4 ATPase driven translocation
of membrane bound proteins has been well established at the endoplasmic reticulum (ER) one may
speculate similar roles for Cdc48-Ufd1-Npl4 of in the turnover of NE associated proteins, possibly
via Ufd1SIM –SUMO interactions.

85
Finally to be mentioned among the factors whose sumoylation increased in both the slx8-1 and
ufd1ΔCt213-342 mutant was the methyltransferase Clr4, responsible for histone H3K9 methylation in
S. pombe. The main points of action of Clr4 are in subtelomeric and pericentric chromosomal
regions, in close physical proximity to both the shelterin and kinetochore complexes.
Heterochromatin is subject to profound changes along the cell cycle (Chen et al, 2008; Kloc et al,
2008), that might be coordinated with concomitant changes in kinetochores or centromeres by
STUbLs and Ufd1.
Separate effects of Ufd1 and STUbLs on the sumoylated proteome Some SUMO substrates were more specifically upregulated in only one mutant background. For
instance, while the sumoylation of the DNA-directed RNA polymerase I and III complex subunits
and of the Cnd2, Cut3 and Cut14 condensin proteins was generally increased in slx8-1, their
modification status seemed rather down in ufd1ΔCt213-342. A similar trend was detected for the
nucleolar protein Dnt1. Strikingly, Dnt1 showed 2 to 6 fold upregulated sumoylation on 8 different
sites in slx8-1 with the most upregulated peptide being one doubly modified on both K89 and K90.
In contrast, the sumoylation of other nucleolar proteins showed a general tendency to decrease in
slx8-1. It is possible that local sequestering of SUMO on a single substrate could lead to the
reduction in other modification events within the same subcellular compartment. The S. cerevisae
Dnt1 homolog, Tof2, is an established SUMO target. Tof2 is part of a network of proteins involved
in the tethering of the rDNA to the inner nuclear membrane and sumoylation may modulate these
interactions (Srikumar et al, 2013b). Proper localization of the rDNA at the nuclear periphery is
required for retaining rDNA structure and silencing. Consequently, loss of sumoylation leads to
decreased rDNA silencing (Watts et al, 2007) and an inability to form SUMO chains has been
linked to gross defects in rDNA compaction/organization (Srikumar et al, 2013a). Moreover,
reduced rDNA silencing can also be observed in STUbL deficient budding yeast cells (Darst et al,
2008), suggesting that a pathway involving both polysumoylation and STUbL activity is required
for normal rDNA function. The identification of Dnt1 as a highly upregulated SUMO target in slx8-
1 cells suggests that Dnt1 may constitute a substrate for such a pathway. Future studies should
elucidate this possibility.
slx8-1 upregulated SUMO targets also included the Psm1 and Psm3 core cohesion proteins. This
finding is interesting in light of a recent study conducted in S. cerevisae. Here, the other core
cohesion subunit Mcd1 (Rad21 homolog) was shown to be targeted for STUbL-mediated

86
degradation in the absence of the cohesin-associated factor Pds5, leading to premature sister
chromatid separation (D'Ambrosio & Lavoie, 2014). By promoting cohesin proteolysis, STUbLs
may equally allow efficient chromosome segregation during anaphase in wild-type cells
(D'Ambrosio & Lavoie, 2014) and/or they might regulate cohesion functions outside mitosis such
as during DNA repair where the sumoylation of cohesins plays important, yet incompletely defined,
roles (McAleenan et al, 2012).
In summary, our quantitative approach succeeded in identifying a number of proteins whose
sumoylation is increased in the slx8-1 and/or ufd1ΔCt213-342 backgrounds, of which only some were
discussed above. Whether these represent direct targets of STUbLs or Ufd1, or rather SUMO
conjugates whose abundance increases through indirect effects of the slx8-1 or ufd1ΔCt213-342
mutations clearly needs to be established through more specific experiments. Nonetheless, we
believe that the list of proteins identified in our SILAC approach is likely to include bona fide
STUbL substrates. This is based on the finding of an already known STUbL target (Mot1) in the
list, and of proteins localizing to subcellular structures or protein complexes previously shown to be
affected by STUbL activity.
Comparison of sumoylation and total protein levels If proteins with increased sumoylation in slx8-1 and/or ufd1ΔCt213-342 represent proteins normally
targeted for degradation by STUbL-mediated ubiquitylation, these proteins may be stabilized in the
mutant backgrounds. We could quantify protein levels for a subset of the proteins with quantified
sumoylation sites (62 and 50 proteins were quantified in both crude and diGly-Lys IP’ed samples
for slx8-1/wild type and ufd1ΔCt213-342/ wild type, respectively). To compare relative changes in
sumoylation with potential changes in protein abundance, log2 ratios for quantified sumoylated
peptides were plotted against crude peptide ratios (Figure 5). As revealed by these maps, changes in
protein sumoylation were not generally correlated with a change in protein abundance, at least not
for the fraction of proteins we were able to compare. Importantly, the protein levels of SUMO itself
were unchanged in the mutants excluding the possibility that protein sumoylation is increased in the
slx8-1 or ufd1ΔCt213-342 mutants due to an elevation in the amount of SUMO. One exception was
Slt1, an uncharacterized orphan protein, which displayed a nearly two-fold increase in ufd1ΔCt213-
342 cells compared to a three-fold increase in sumoylation. Although Slt1 sumoylation was also
prominently increased in slx8-1, the total levels did not change. Further investigation of Slt1

87
sumoylation may provide exciting insight into the biological function of this protein. A moderate
increase in the protein levels of ubiquitin and of Cdc48 could also be detected specifically in
ufd1ΔCt213-342 (log2 ratios of 0.59 and 0.66, respectively). Increased ubiquitin levels could be due to
the stabilization of ubiquitylated proteins normally turned-over through Ufd1. Similarly, indirect
effects on Cdc48 stability and/or expression levels may be induced in the absence of its major Ufd1
co-factor. Of note, increased sumoylation of ubiquitin and Cdc48, as detected in the ufd1ΔCt213-342
strain, may thus partly be explained by increased protein levels. This notwithstanding, the lack of
correlation between changes in sumoylation status and protein amount, as determined for at least a
subset of slx8-1 and/or ufd1ΔCt213-342 regulated SUMO substrates, is in line with the notion that
only a minor fraction of a protein is typically modified by SUMO at any given time (Hay, 2005;
Tatham et al, 2011). Consequently, the total pool of a protein likely to be subjected to
STUbL/Ufd1-mediated regulation is proportionally low and any change may thus go undetected.
Conclusion Using a novel strategy to purify peptides modified by SUMO we have analyzed the endogenous
sumoylated proteome of fission yeast wild-type and mutant cells. More specifically, we have
isolated His6SUMOL109K-modified peptides from a ufd1ΔCt213-342 mutant background which
displays increased levels of SUMO conjugates, and we have used SILAC-based quantitative
proteomics to compare sumoylation levels in a ufd1ΔCt213-342 mutant, an slx8-1 STUbL mutant, and
wild-type. These experiments combined identify over 900 unique sumoylated sites within 431
different proteins. They constitute the first proteome-wide investigation of sumoylation in fission
yeast, an extensively studied model organism to which powerful genetic and biochemical analyses
can be applied to test the molecular mechanisms suggested by our findings. More broadly, the
work reported here provides a precious resource to the entire SUMO research community by
offering specific information about the lysine residues to which SUMO is conjugated. Furthermore,
through quantitative comparison we shed light on potential SUMO substrates regulated by STUbL
and/or Cdc48-Ufd1-Npl4-mediated activities. Our findings should allow for a deeper understanding
of the dynamic regulation of SUMO targets through these highly conserved factors.

88
Experimental procedures Strain construction All strains used in this study are listed in the table below. The endogenous S. pombe SUMO gene
was engineered to contain an N-terminal His6-tag and a single amino acid substitution changing the
leucine at position 109, immediately upstream the C-terminal diGly, into a lysine (His6-
SUMOL109K). This was performed using a “loop-in-loop out” strategy (Gao et al, 2014) where a
plasmid containing His6-SUMOL109K, pmt3+ 5’ and 3’ flanking regions and an ura4+ gene
(pJBK142) was integrated into the pmt3-flanking region by homologous recombination in a ura-
D18 strain. “Loop-outs” where His6-SUMOL109K remained as the single pmt3 genomic copy were
subsequently selected by 5'-fluoro-orotic acid (FOA) resistance and they were verified by PCR
sequencing. To produce the pJBK142 “loop-in” plasmid, the S. pombe SUMO (pmt3) ORF and its
flanking regions were amplified in two separate PCR reactions: The ORF and its 5’ region were
amplified with the forward primer GTO-492 (5’GCTCCACAATTCTCACAAGCACCC 3’) and
reverse primer GTO-548 (5’ CGATGGATCCCTAAAGGCATAGATGGGTGCAAC
CACCTTTCTGTTCTAAG 3’), changing the leucine codon for amino acid 109 (TTA) codon to a
lysine codon (TTT) , as underlined. The pmt3 downstream region was amplified with GTO-549(5’
GCATGGATCCAGTACAAGTATTTTTAAGCTGTTTC 3’) and GTO-550
(5’GCATCCATGGCGCTCAGTCAAGGTATGCTTCCATAAGCAG 3’). Prior to chromosomal
integration the plasmid was digested with SphI. After His6-SUMOL109K allele replacements had been
identified, an ura4+ gene was inserted at a tightly linked location 5’ of pmt3 to follow the allele
through subsequent crosses. The int::pJBK111(ura4+)-His6-SUMOL109K allele was crossed into slx8-
1 and ufd1ΔCt213-342 mutant backgrounds and combined the lys3-27 allele using standard genetic
crosses techniques.
Strain table
Strain name Genotype JK408 h- ura-D18 lys3-27 int::pJBK111(ura4+)-His6-SUMO(L109K)
JK414 h- ura-D18 lys3-27 int::pJBK111(ura4+)-His6-SUMO(L109K) ufd1-ΔC::nat
JK418 h- ura-D18 lys3-27 int::pJBK111(ura4+)-His6-SUMO(L109K) slx8-1:myc:kanMX6
JK424 h- ura-D18 lys3-27 int::pJBK111(ura4+)-SUMO
JK425 h- ura-D18 lys3-27 int::pJBK111(ura4+)-SUMO; ufd1-ΔC::nat
JK426 h- ura-D18 lys3-27 int::pJBK111(ura4+)-SUMO; slx8-1:myc:kanMX6

89
Anti-SUMO Western blot The indicated strain were grown in yeast-extract (YEL) medium at 30°C and harvested at an OD600
~0.5. Cells were lysed with the glass bead method in a denaturing buffer containing 8M UREA; 100
mM sodium phosphate pH 7.5; 50 mM Tris-HCl pH 7.5; 150 mM NaCl and 20 mM NEM (N-
etylmaleimide) using a fast prep instrument. Equal amounts of protein lysates were separated on a
SDS-page 4-20% gradient gel (Lonza), transferred to a nitrocellulose/MCE membrane (Advantec)
and immnoblotted using an anti-SUMO rabbit antibody ((Xhemalce et al, 2004); kindly provided by
J. Seeler). Following incubation with a horseradish peroxidase (HRP)-conjugated swine anti-rabbit
IgG secondary antibody (Dako), Sumoylated species were detected with an ECL plus kit (GE
Healthcare). Parallel Western blots were performed with an anti-tubulin antibody (kindly provided
by R. Hartmann-Petersen).
Propagation and harvesting of ufd1ΔCt213-342 His6-SUMOL109K cells for His6-SUMOL109K SUMO site identification A ufd1ΔCt213-342 His6-SUMOL109K expressing strain was grown in Edinburgh minimal medium
(EMM2) medium supplemented with 30 mg/L lysine at 30°C. Approximately 2.5 L of culture was
harvested from exponentially growing cells at an OD600 of 0.9.
Culture conditions for SILAC Strains were grown in Edinburgh minimal medium (EMM2) supplemented with 30 mg/L of either
“light” (12C614N2, Lys0) or “heavy” L-lysine (13C6
15N2, Lys8) (Cambridge Isotope Laboratories).
The wild-type and ufd1ΔCt213-342 strains were each grown in light lysine conditions and individually
compared with the slx8-1 grown in parallel in heavy lysine. The experiments were performed in
triplicates starting from independent biological isolates. To ensure full incorporation of heavy
lysine, the slx8-1 strain was propagated in SILAC medium for at least 25 generations after which
the incorporation efficiency was determined to be more than 98%. The cultures were propagated at
30°C, except for the last 12 hr where they were propagated at 33°C to induce the slx8-1
temperature-sensitive phenotype at a semi-permissive temperature (Prudden et al, 2007).
Approximately 2 L of culture at an OD600 ~1 was harvested per strain per experiment. Cell pellets
were washed in ice-cold PBS and frozen in liquid nitrogen.

90
Protein extraction and nickel affinity purification Protein extraction and Ni2+ purification of His6-SUMOL109K conjugates from both ufd1ΔCt213-342 un-
labeled cultures and from SILAC labeled mixed cells were performed using similar protocols. The
SILAC experiments, comparing slx8-1 with wild type and slx8-1 with ufd1ΔCt213-342, were
processed in parallel. Heavy and light labeled cells were mixed in a 1:1 ratio based on the weight of
cell pellets prior to lysis. Mixed pellets were resuspended in a total 50 ml of ice-cold water and
lysed by adding an equal volume of NaOH and β-mercaptoethanol to a final concentration of 1.85
M and 1.85 %, respectively (~12 ml lysis buffer per 1 g of cell pellet). After incubation on ice for
30 min, proteins were precipitated with trichloroacetic acid (TCA), washed with ice-cold acetone
and resuspended in 60 ml of binding buffer (6M Guanidinium-HCl; 100 mM sodium phosphate, pH
8.0; 10 mM Tris-HCl, pH 8.0; 20 mM Imidazole; 5 mM β-mercaptoethanol). Proteins were allowed
to go in solution in binding buffer on a roller mixer for up to 1 hour before lysates were finally
clarified by high-speed ultracentrifugation (30.000 G for 20 min). In all cases, the total protein yield
was ~1.5 g as determined by Nanodrop analysis (A280). ~15 µg of cell lysates was saved for
complete proteome analysis while the remaining material was incubated with 5 ml of pre-washed
Ni2+-NTA agarose beads (Qiagen) at 4°C overnight. Beads were washed on gravity columns with
50 ml volumes as follows: binding buffer, wash buffer pH 8.0 (8 M urea; 100 mM sodium
phosphate buffer, pH 8.0; 10 mM Tris-HCl, pH 8.0; 20 mM Imidazole; 5 mM β-mercaptoethanol),
wash buffer pH 6.3 (8 M urea; 100 mM sodium phosphate buffer, pH 6.3; 10 mM Tris-HCl, pH 6.3;
20 mM Imidazole; 5 mM β-mercaptoethanol) and once more with wash buffer pH 8.0. Proteins
were eluted in three sequencial steps with 5 ml of wash buffer pH 8.0 containing 200 mM imidazole
and pulled for further processing.
Filter-aided sample preparation and protein digestion His6-SUMOL109K conjugated proteins (~300 µg as determined by Nanodrop analysis) were digested
on 30 kD cut-off filter-units (Sartorius) essentially as described in Tammsalu et al., 2014. 15 ml of
Ni2+ purified sample was concentrated onto three filters. Each filter-unit was washed twice with 200
µl UA buffer (8M urea, 100 mM Tris-HCl pH7.5) and treated with 50 mM chloroacetamide in UA
buffer for 20 min in the dark. After two additional washes with UA buffer, the samples were
equilibrated with 3x 200 µl volumes of immunoprecipitation (IP) buffer (50 mM MOPS-NaOH pH
7.2; 10 mM Na2HPO4; 50 mM NaCl). Proteins were digested with Lys-C (Wako) in 50 µl IP buffer
for 16 hours at 37°C at a 1:50 enzyme-to-protein ratio. The digested peptides were collected by

91
centrifugation and filter-units were washed with 50 µl of IP buffer to increase the yield. To recover
longer peptides retained on filters, another digestion step was performed with Glu-C (1:100
enzyme-to-protein ratio) in 50 µl IP buffer for 16 hours at 20°C. Peptides collected after Lys-C or
Lys-C and Glu-C digestions were kept separated. For complete proteome analysis, 15 µg of protein
in binding buffer was diluted ten-fold into 8M urea, 100 mM Tris-HCl, pH7.5 and treated with 50
mM chloroacetamide in the dark at room temperature for 1.5 hours. The samples were then diluted
4-fold with 50 mM ammonium bicarbonate to reduce the urea concentration and they were digested
in solution with Lys-C (1:50 ratio) 16 hours at 20°C.
DiGly-Lys-specific immunoprecipitation An anti-KεGG antibody coupled to protein A beads (PTMScan, Cell Signaling Technology) was
used for immunoprecipitation of diGly-Lys containing peptides. Prior to use, coupled beads were
washed twice with 20 bead volumes of conjugation buffer (20 mM sodium phosphate; 150 mM
NaCl, pH 9.0) and crosslinked in 25 bead volumes of 5 mM Bis(sulfosuccinimidyl)suberate (BS3)
for 30 min at room temperature while rotating. The reaction was quenched with 1M Tris-HCl (pH
7.5) for 15 min at room temperature and beads were washed three times with 20 bead volumes of
cold IP buffer (50 mM MOPS-NaOH, pH 7.2; 10 mM Na2HPO4; 50 mM NaCl). 3 µl of beads
cross-linked to 19 µg of anti-KεGG antibody were added to Lys-C and Lys-C plus Glu-C digested
peptide mixtures in IP buffer and incubated at 4°C overnight. The beads were washed twice with
500 µl of ice-cold PBS and peptides were eluted in three sequential steps with 50 µl of 0.1%
trifluoroacetic acid (TFA). Eluted samples were pulled.
Mass Spectrometry analysis All peptide samples were desalted and concentrated with home-made reverse-phase C18 (Empore)
Stop and Go extraction Tips (Rappsilber et al, 2007), resuspended in 0.1% TFA and analyzed by
liquid chromatography-tandem mass spectrometry (LC-MS/MS). An EASY-nLC 1000 liquid
chromatography system (Thermo Scientific) coupled to a Q Exactive mass spectrometer (Thermo
Scientific) through an EASY-Spray ion source (Thermo Scientific) was used and purified peptides
were separated on an EASY-Spray column (75 µm x 500 mm) with a 90 min linear gradient of 5 %
to 40% of acetonitrile in 0.1% formic acid at a flow rate of 250 nl/min. MS and MS/MS spectra of
the eluting peptides were analysed online by Q Exactive mass spectrometer operated in a data-
dependent acquisition mode. Precursor ion full-scan MS spectra were acquired at a target value of 1

92
000 000 ions (maximum injection time 20 ms) over a scan range of 300 Th to 1600 Th with a
resolution of 70 000 at m/z 400. Up to one data-dependent Higher-Collisional Dissociation (HCD)
tandem mass spectrum (MSMS) was acquired at a target value of 500 000 ions (maximum injection
time 1000 ms) with a resolution 35 000 at m/z 400 and normalized collision energy of 30%. Singly
and highly (>8) charged ions, or ions with unassigned charge states were rejected and the intensity
threshold was set to 2.0 x 104 U. M/z values of acquired ions were added to the exclusion list for 40
s and peptide match option was set to preferred.
Data analysis Raw mass spectrometric data files were processed with MaxQuant software (version 1.3.0.5) (Cox
& Mann, 2008; Cox et al, 2011a) and searched against UniprotKB S. pombe reference protein
database containing canonical and isoform sequences (downloaded on 14th December
2013).Protease specificity was set for Lys-C or Lys-C and Glu-C permitting up to three or five
missed cleavages, respectively. Carbamidomethylation of Cys residues was added as a fixed
modification and oxidation of Met, acetylation of protein N-termini and diGly adduction to internal
Lys in the peptide sequence were set as variable modifications. In addition, phosphorylation of Ser,
Thr and Tyr or acetylation of Lys were added as variable modifications and the minimum peptide
length was set to seven amino acids. For SILAC samples, multiplicity was set to 2, and Lys8 and
was selected as a heavy labeled counterpart. The data set was filtered by posterior error probability
to achieve a false discovery rate of 1% at both, protein and peptide level.
Analysis of His6-SUMOL109K modified sites Sequence motifs were visualised using pLogo (O'Shea et al, 2013). A list of 13 amino acid
sequence window of all SUMO-modified lysines identified from SILAC experiments was used as
an input and N- or C-terminal sequences that did not cover the residue window were omitted from
the output. UniprotKB S. pombe reference protein database (downloaded on 14.12.2013) containing
145132 lysines was used as a background data set and residues were scaled relative to their
Bonferroni-corrected statistical significance.

93
Figure Legends
Figure 1. Construction and phenotypic examination of a His6-SUMOL109K S. pombe SUMO variant.
(A) Depiction of the carboxyl termini of S. pombe Ubiquitin-like proteins (Ubls) possessing diGly
motifs. C-terminal lysine residues recognized by endo-Lys-C are highlighted in red to indicate the
branched peptide left after Lys-C digestion. (B) Growth analysis of His6-SUMOL109K wild type,
slx8-1 and ufd1ΔCt213-342 strains. (Left) 10-fold dilution series of the indicated strains were spotted
onto YES plates and incubated at 33°C. (Right) Growth curves of isogenic wild-type and His6-
SUMOL109K cells, portraying three independent isolates of each strain. Growth curves of slx8-1 and
ufd1ΔCt213-342 His6-SUMOL109K expressing cells are shown in Figure S1. Cells were propagated at
30°C in EMM2 medium with required supplements and culture absorbance at 600 nm was recorded
at fixed time intervals, as depicted on graphs. (C) Western blot analysis of wild-type SUMO and
His6-SUMOL109K conjugates. Whole-cell extracts of the indicated strains grown at 30°C were
probed with an anti-SUMO antibody. An anti-tubulin antibody was used as a loading control.
Figure 2. Identification of His6-SUMOL109K modified residues in the proteomes of S. pombe wild
type cells, Ufd1 and STUbL mutants. (A) Two-step enrichment workflow to identify His6-
SUMOL109K modified sites using the ufd1ΔCt213-342 mutant background. (B) Depiction of
quantitative proteomic experiments comparing His6-SUMOL109K modification levels between wild-
type, slx8-1 and ufd1ΔCt213-342 cells. Parallel experiments were performed in which wild-type and
ufd1ΔCt213-342 cultures in “light” lysine (Lys0) medium, were individually mixed 1:1 with slx8-1
“heavy” lysine (Lys8) cultures. ~15 µg of mixed cell lysates were kept for total proteome analysis
while His6-SUMOL109K conjugates were isolated and analyzed as in (A). The parallel SILAC
experiments were performed in triplicate with independent biological isolates. (C) Overview of total
sites identified using the non-SILAC and SILAC approaches. Of the 749 sites identified from the
un-labeled ufd1ΔCt213-342 His6-SUMOL109K IP workflow, 351 were also identified in at least one of
the SILAC experiments. Combined from all experiments 937 distinct His6-SUMOL109K modified
sites were identified distributed between among 422 different proteins. (D) Sequence analysis of the
amino acid context of His6-SUMOL109K targeted lysines. Four pLogo graphs are shown displaying
the consensus context of all lysines targeted by His6-SUMOL109K, of KxE/KxD consensus site
lysines, of lysines within the ExK/DxK inverted consensus site or of sites not conforming to any of
these (residual sites). The numbers in parentheses above graphs each indicate the number of sites

94
with a full 13-amino acid sequence compared to the total number of lysines conforming to that
motif. The log-odds of the binomial probability (π) are represented by the y-axis. Significance
threshold values of 3.68 (P<0.05) are shown in red.
Figure 3. Functional analysis of the S. pombe sumoylated proteome. (A) Gene Ontology (GO)-slim
“biological process” analysis on the ensemble of His6-SUMOL109K -modified protein identified in all
experiments combined. The graphs show the most significantly enriched “process terms” (P <
0.025) of His6-SUMOL109K modified proteins compared to the predicted S. pombe proteome. GO-
slim analysis was performed using the GOTermMapper online tool at Princeton and P-values were
calculated with a hypergeometric probability test. (B) GO enrichment analysis of “cellular
compartment” terms for His6-SUMOL109K targets using the AmiGo online tool. Selected enriched
GO-categories (P < 0.01) are shown as bar-graphs arranged according to their degree of
significance. (C) Examples of interacting protein complexes heavily influenced by sumoylation
localizing at S. pombe centromeres or telomeres. Only proteins with identified sumoylation sites are
displayed. S. pombe centromeres are clustered at the spindle pole body (SPB) in interphase cells
linked through inner and outer nuclear membrane (INM and ONM) proteins. Dashed lines indicate
proteins associating with kinetochores in a dynamic manner only during mitosis.
Figure 4. Comparison of His6-SUMOL109K –modified sites in wild type cells, slx8-1 and ufd1ΔCt213-
342 mutants. (A) Overview of peptides quantified in the triplicate SILAC experiments. Some sites
were quantified both as singly diGly-Lys modified and as peptides simultaneously modified on
more than one site. Only diGly-Lys-modified peptides for which SILAC ratios had been obtained in
at least two biological replicates (based on a minimum of two SILAC ratio counts) were used for
quantification. In total, 178 distinct diGly-Lys- modified peptides were quantified in both slx8-
1/wild type and slx8-1/ufd1ΔCt213-342 IP experiments, which enabled us to calculate their relative
abundance also between ufd1ΔCt213-342 and wild type. (B) The normalized log2 ratios of quantified
diGly-Lys-modified peptides for ufd1ΔCt213-342/ wild type and for slx8-1/wild type are represented
on a single scatter plot (tsMAP) (Golebiowski et al, 2009), giving an overview of the relative
change in abundance of individual peptides (sites) between each mutant background and wild type.
Note that peptides that were only quantified in one of the slx8-1/wild type and slx8-1/ufd1ΔCt213-342
IP experiments did not give rise to a coordinate on the tsMAP. The two data sets are found to
correlate with a Pearson coefficient of r=0.48. DiGly-Lys modified sites upregulated more than

95
two-fold (log2≥1) in both slx8-1 and ufd1ΔCt213-342 mutant backgrounds are displayed on the chart.
SUMO K14;30 annotates a doubly modified SUMO peptide. Also depicted on the plot is a Mot1
peptide doubly-modified on K101;K105 which was upregulated in both mutant strains, although
slightly below the log2≥1 significance criterion in slx8-1, and several peptides of the nucleolar
protein Dnt1 which were specifically enriched in slx8-1.
Figure 5. Comparison of relative changes in His6-SUMOL109K sumoylation with total protein
abundance. Of proteins with quantified sumoylation sites, 62 and 50 were also quantified from the
“crude” preparations for slx8-1 /wild type and ufd1ΔCt213-342 / wild type, respectively. Log2 SILAC
ratios for sumoylated peptides were plotted against quantified ratios obtained for these proteins,
yielding the maps in (A) and (B). Changes in protein sumoylation were not generally correlated
with changes in their total amount in either the slx8-1 or ufd1ΔCt213-342 backgrounds.
Figure S1. Growth curves of slx8-1 and ufd1ΔCt213-342 His6-SUMOL109K expressing cells. Cells
were propagated as in Figure 1.
Tabel S1. List of all His6-SUMOL109K -modified sites from our combined experiments (Figure 2A
and B). Sites identified in the ufd1∆Ct213-342 un-labeled and in the SILAC experiments are indicated
in separate columns. Sites are arranged in alphabetic order according to gene name. Multiplicity
refers to whether a site was identified as part of a singly, a doubly or a triply diGly-Lys modified
peptide. The MaxQuant calculated score values (Andromeda) are displayed for individual sites
(Cox et al, 2011b). At the bottom of table are shown four diGly-Lys modified sites for which the
location probability was below 0.9.
Table S2. List of all quantified His6-SUMOL109K-modified sites from the SILAC experiments
(Figure 2B). Sites are arranged in alphabetic order according to gene name. Some sites are
quantified as part of doubly or triply-modified peptides as indicated by a semicolon between lysine
positions. Sites that were significantly upregulated (log2 ratios ≥1.0) in both slx8-1 and in
ufd1∆Ct213-342 are highlighted in grey.
Table S3. List of all quantified proteins from crude SILAC lysates.

96
References Andrews EA, Palecek J, Sergeant J, Taylor E, Lehmann AR, Watts FZ (2005) Nse2, a component of the
Smc5-6 complex, is a SUMO ligase required for the response to DNA damage. Molecular and cellular
biology 25: 185-196
Baek GH, Cheng H, Choe V, Bao X, Shao J, Luo S, Rao H (2013) Cdc48: a swiss army knife of cell biology.
Journal of amino acids 2013: 183421
Becker J, Barysch SV, Karaca S, Dittner C, Hsiao HH, Berriel Diaz M, Herzig S, Urlaub H, Melchior F
(2013) Detecting endogenous SUMO targets in mammalian cells and tissues. Nature structural & molecular
biology 20: 525-531
Bergink S, Ammon T, Kern M, Schermelleh L, Leonhardt H, Jentsch S (2013) Role of Cdc48/p97 as a
SUMO-targeted segregase curbing Rad51-Rad52 interaction. Nature cell biology 15: 526-532
Bruderer R, Tatham MH, Plechanovova A, Matic I, Garg AK, Hay RT (2011) Purification and identification
of endogenous polySUMO conjugates. EMBO reports 12: 142-148
Burgess RC, Rahman S, Lisby M, Rothstein R, Zhao X (2007) The Slx5-Slx8 complex affects sumoylation
of DNA repair proteins and negatively regulates recombination. Molecular and cellular biology 27: 6153-
6162
Chen ES, Zhang K, Nicolas E, Cam HP, Zofall M, Grewal SI (2008) Cell cycle control of centromeric repeat
transcription and heterochromatin assembly. Nature 451: 734-737
Chikashige Y, Yamane M, Okamasa K, Tsutsumi C, Kojidani T, Sato M, Haraguchi T, Hiraoka Y (2009)
Membrane proteins Bqt3 and -4 anchor telomeres to the nuclear envelope to ensure chromosomal bouquet
formation. The Journal of cell biology 187: 413-427
Cook CE, Hochstrasser M, Kerscher O (2009) The SUMO-targeted ubiquitin ligase subunit Slx5 resides in
nuclear foci and at sites of DNA breaks. Cell cycle 8: 1080-1089
Cox J, Mann M (2008) MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass
accuracies and proteome-wide protein quantification. Nature biotechnology 26: 1367-1372
Cox J, Michalski A, Mann M (2011a) Software lock mass by two-dimensional minimization of peptide mass
errors. Journal of the American Society for Mass Spectrometry 22: 1373-1380
Cox J, Neuhauser N, Michalski A, Scheltema RA, Olsen JV, Mann M (2011b) Andromeda: a peptide search
engine integrated into the MaxQuant environment. Journal of proteome research 10: 1794-1805
D'Ambrosio LM, Lavoie BD (2014) Pds5 prevents the PolySUMO-dependent separation of sister
chromatids. Current biology : CB 24: 361-371
Darst RP, Garcia SN, Koch MR, Pillus L (2008) Slx5 promotes transcriptional silencing and is required for
robust growth in the absence of Sir2. Molecular and cellular biology 28: 1361-1372

97
Ferreira HC, Luke B, Schober H, Kalck V, Lingner J, Gasser SM (2011) The PIAS homologue Siz2
regulates perinuclear telomere position and telomerase activity in budding yeast. Nature cell biology 13:
867-874
Ferreira HC, Towbin BD, Jegou T, Gasser SM (2013) The shelterin protein POT-1 anchors Caenorhabditis
elegans telomeres through SUN-1 at the nuclear periphery. The Journal of cell biology 203: 727-735
Filosa G, Barabino SM, Bachi A (2013) Proteomics strategies to identify SUMO targets and acceptor sites: a
survey of RNA-binding proteins SUMOylation. Neuromolecular medicine 15: 661-676
Finkbeiner E, Haindl M, Raman N, Muller S (2011) SUMO routes ribosome maturation. Nucleus 2: 527-532
Flotho A, Melchior F (2013) Sumoylation: a regulatory protein modification in health and disease. Annual
review of biochemistry 82: 357-385
Fujita I, Nishihara Y, Tanaka M, Tsujii H, Chikashige Y, Watanabe Y, Saito M, Ishikawa F, Hiraoka Y,
Kanoh J (2012) Telomere-nuclear envelope dissociation promoted by Rap1 phosphorylation ensures faithful
chromosome segregation. Current biology : CB 22: 1932-1937
Funabiki H, Hagan I, Uzawa S, Yanagida M (1993) Cell cycle-dependent specific positioning and clustering
of centromeres and telomeres in fission yeast. The Journal of cell biology 121: 961-976
Galanty Y, Belotserkovskaya R, Coates J, Jackson SP (2012) RNF4, a SUMO-targeted ubiquitin E3 ligase,
promotes DNA double-strand break repair. Genes & development 26: 1179-1195
Gao J, Kan F, Wagnon JL, Storey AJ, Protacio RU, Davidson MK, Wahls WP (2014) Rapid, efficient and
precise allele replacement in the fission yeast Schizosaccharomyces pombe. Current genetics 60: 109-119
Golebiowski F, Matic I, Tatham MH, Cole C, Yin Y, Nakamura A, Cox J, Barton GJ, Mann M, Hay RT
(2009) System-wide changes to SUMO modifications in response to heat shock. Science signaling 2: ra24
Gonzalez Y, Saito A, Sazer S (2012) Fission yeast Lem2 and Man1 perform fundamental functions of the
animal cell nuclear lamina. Nucleus 3: 60-76
Hannich JT, Lewis A, Kroetz MB, Li SJ, Heide H, Emili A, Hochstrasser M (2005) Defining the SUMO-
modified proteome by multiple approaches in Saccharomyces cerevisiae. The Journal of biological chemistry
280: 4102-4110
Hay RT (2005) SUMO: a history of modification. Molecular cell 18: 1-12
Hediger F, Neumann FR, Van Houwe G, Dubrana K, Gasser SM (2002) Live imaging of telomeres: yKu and
Sir proteins define redundant telomere-anchoring pathways in yeast. Current biology : CB 12: 2076-2089
Hickey CM, Wilson NR, Hochstrasser M (2012) Function and regulation of SUMO proteases. Nature
reviews Molecular cell biology 13: 755-766
Ho JC, Warr NJ, Shimizu H, Watts FZ (2001) SUMO modification of Rad22, the Schizosaccharomyces
pombe homologue of the recombination protein Rad52. Nucleic acids research 29: 4179-4186
Hou H, Kallgren SP, Jia S (2013) Csi1 illuminates the mechanism and function of Rabl configuration.
Nucleus 4: 176-181

98
Keusekotten K, Bade VN, Meyer-Teschendorf K, Sriramachandran AM, Fischer-Schrader K, Krause A,
Horst C, Schwarz G, Hofmann K, Dohmen RJ, Praefcke GJ (2014) Multivalent interactions of the SUMO-
interaction motifs in RING finger protein 4 determine the specificity for chains of the SUMO. The
Biochemical journal 457: 207-214
Kloc A, Zaratiegui M, Nora E, Martienssen R (2008) RNA interference guides histone modification during
the S phase of chromosomal replication. Current biology : CB 18: 490-495
Kohler JB, Jorgensen ML, Beinoraite G, Thorsen M, Thon G (2013) Concerted action of the ubiquitin-fusion
degradation protein 1 (Ufd1) and Sumo-targeted ubiquitin ligases (STUbLs) in the DNA-damage response.
PloS one 8: e80442
Kosoy A, Calonge TM, Outwin EA, O'Connell MJ (2007) Fission yeast Rnf4 homologs are required for
DNA repair. The Journal of biological chemistry 282: 20388-20394
Lallemand-Breitenbach V, Jeanne M, Benhenda S, Nasr R, Lei M, Peres L, Zhou J, Zhu J, Raught B, de The
H (2008) Arsenic degrades PML or PML-RARalpha through a SUMO-triggered RNF4/ubiquitin-mediated
pathway. Nature cell biology 10: 547-555
Lescasse R, Pobiega S, Callebaut I, Marcand S (2013) End-joining inhibition at telomeres requires the
translocase and polySUMO-dependent ubiquitin ligase Uls1. The EMBO journal 32: 805-815
Liu X, McLeod I, Anderson S, Yates JR, 3rd, He X (2005) Molecular analysis of kinetochore architecture in
fission yeast. The EMBO journal 24: 2919-2930
Makhnevych T, Sydorskyy Y, Xin X, Srikumar T, Vizeacoumar FJ, Jeram SM, Li Z, Bahr S, Andrews BJ,
Boone C, Raught B (2009) Global map of SUMO function revealed by protein-protein interaction and
genetic networks. Molecular cell 33: 124-135
Mann M (2006) Functional and quantitative proteomics using SILAC. Nature reviews Molecular cell biology
7: 952-958
Matic I, Schimmel J, Hendriks IA, van Santen MA, van de Rijke F, van Dam H, Gnad F, Mann M, Vertegaal
AC (2010) Site-specific identification of SUMO-2 targets in cells reveals an inverted SUMOylation motif
and a hydrophobic cluster SUMOylation motif. Molecular cell 39: 641-652
McAleenan A, Cordon-Preciado V, Clemente-Blanco A, Liu IC, Sen N, Leonard J, Jarmuz A, Aragon L
(2012) SUMOylation of the alpha-kleisin subunit of cohesin is required for DNA damage-induced cohesion.
Current biology : CB 22: 1564-1575
Meyer H, Bug M, Bremer S (2012) Emerging functions of the VCP/p97 AAA-ATPase in the ubiquitin
system. Nature cell biology 14: 117-123
Mukhopadhyay D, Arnaoutov A, Dasso M (2010) The SUMO protease SENP6 is essential for inner
kinetochore assembly. The Journal of cell biology 188: 681-692

99
Nagai S, Dubrana K, Tsai-Pflugfelder M, Davidson MB, Roberts TM, Brown GW, Varela E, Hediger F,
Gasser SM, Krogan NJ (2008) Functional targeting of DNA damage to a nuclear pore-associated SUMO-
dependent ubiquitin ligase. Science 322: 597-602
Nie M, Aslanian A, Prudden J, Heideker J, Vashisht AA, Wohlschlegel JA, Yates JR, 3rd, Boddy MN (2012)
Dual recruitment of Cdc48 (p97)-Ufd1-Npl4 ubiquitin-selective segregase by small ubiquitin-like modifier
protein (SUMO) and ubiquitin in SUMO-targeted ubiquitin ligase-mediated genome stability functions. The
Journal of biological chemistry 287: 29610-29619
O'Shea JP, Chou MF, Quader SA, Ryan JK, Church GM, Schwartz D (2013) pLogo: a probabilistic approach
to visualizing sequence motifs. Nature methods 10: 1211-1212
Panse VG, Kressler D, Pauli A, Petfalski E, Gnadig M, Tollervey D, Hurt E (2006) Formation and nuclear
export of preribosomes are functionally linked to the small-ubiquitin-related modifier pathway. Traffic 7:
1311-1321
Prudden J, Pebernard S, Raffa G, Slavin DA, Perry JJ, Tainer JA, McGowan CH, Boddy MN (2007) SUMO-
targeted ubiquitin ligases in genome stability. The EMBO journal 26: 4089-4101
Prudden J, Perry JJ, Nie M, Vashisht AA, Arvai AS, Hitomi C, Guenther G, Wohlschlegel JA, Tainer JA,
Boddy MN (2011) DNA repair and global sumoylation are regulated by distinct Ubc9 noncovalent
complexes. Molecular and cellular biology 31: 2299-2310
Psakhye I, Jentsch S (2012) Protein group modification and synergy in the SUMO pathway as exemplified in
DNA repair. Cell 151: 807-820
Rappsilber J, Mann M, Ishihama Y (2007) Protocol for micro-purification, enrichment, pre-fractionation and
storage of peptides for proteomics using StageTips. Nature protocols 2: 1896-1906
Rodriguez MS, Dargemont C, Hay RT (2001) SUMO-1 conjugation in vivo requires both a consensus
modification motif and nuclear targeting. The Journal of biological chemistry 276: 12654-12659
Rojas-Fernandez A, Plechanovova A, Hattersley N, Jaffray E, Tatham MH, Hay RT (2014) SUMO chain-
induced dimerization activates RNF4. Molecular cell 53: 880-892
Sanchez-Perez I, Renwick SJ, Crawley K, Karig I, Buck V, Meadows JC, Franco-Sanchez A, Fleig U, Toda
T, Millar JB (2005) The DASH complex and Klp5/Klp6 kinesin coordinate bipolar chromosome attachment
in fission yeast. The EMBO journal 24: 2931-2943
Schimmel J, Eifler K, Sigurethsson JO, Cuijpers SA, Hendriks IA, Verlaan-de Vries M, Kelstrup CD,
Francavilla C, Medema RH, Olsen JV, Vertegaal AC (2014) Uncovering SUMOylation dynamics during
cell-cycle progression reveals FoxM1 as a key mitotic SUMO target protein. Molecular cell 53: 1053-1066
Schober H, Ferreira H, Kalck V, Gehlen LR, Gasser SM (2009) Yeast telomerase and the SUN domain
protein Mps3 anchor telomeres and repress subtelomeric recombination. Genes & development 23: 928-938
Shin JA, Choi ES, Kim HS, Ho JC, Watts FZ, Park SD, Jang YK (2005) SUMO modification is involved in
the maintenance of heterochromatin stability in fission yeast. Molecular cell 19: 817-828

100
Shiroiwa Y, Hayashi T, Fujita Y, Villar-Briones A, Ikai N, Takeda K, Ebe M, Yanagida M (2011) Mis17 is a
regulatory module of the Mis6-Mal2-Sim4 centromere complex that is required for the recruitment of
CenH3/CENP-A in fission yeast. PloS one 6: e17761
Skilton A, Ho JC, Mercer B, Outwin E, Watts FZ (2009) SUMO chain formation is required for response to
replication arrest in S. pombe. PloS one 4: e6750
Srikumar T, Lewicki MC, Costanzo M, Tkach JM, van Bakel H, Tsui K, Johnson ES, Brown GW, Andrews
BJ, Boone C, Giaever G, Nislow C, Raught B (2013a) Global analysis of SUMO chain function reveals
multiple roles in chromatin regulation. The Journal of cell biology 201: 145-163
Srikumar T, Lewicki MC, Raught B (2013b) A global S. cerevisiae small ubiquitin-related modifier (SUMO)
system interactome. Molecular systems biology 9: 668
Steglich B, Filion GJ, van Steensel B, Ekwall K (2012) The inner nuclear membrane proteins Man1 and
Ima1 link to two different types of chromatin at the nuclear periphery in S. pombe. Nucleus 3: 77-87
Sun H, Leverson JD, Hunter T (2007) Conserved function of RNF4 family proteins in eukaryotes: targeting a
ubiquitin ligase to SUMOylated proteins. The EMBO journal 26: 4102-4112
Tammsalu T, Matic I, Jaffray EG, Ibrahim AF, Tatham MH, Hay RT (2014) Proteome-wide identification of
SUMO2 modification sites. Science signaling 7: rs2
Tanaka K, Nishide J, Okazaki K, Kato H, Niwa O, Nakagawa T, Matsuda H, Kawamukai M, Murakami Y
(1999) Characterization of a fission yeast SUMO-1 homologue, pmt3p, required for multiple nuclear events,
including the control of telomere length and chromosome segregation. Molecular and cellular biology 19:
8660-8672
Tatham MH, Geoffroy MC, Shen L, Plechanovova A, Hattersley N, Jaffray EG, Palvimo JJ, Hay RT (2008)
RNF4 is a poly-SUMO-specific E3 ubiquitin ligase required for arsenic-induced PML degradation. Nature
cell biology 10: 538-546
Tatham MH, Jaffray E, Vaughan OA, Desterro JM, Botting CH, Naismith JH, Hay RT (2001) Polymeric
chains of SUMO-2 and SUMO-3 are conjugated to protein substrates by SAE1/SAE2 and Ubc9. The Journal
of biological chemistry 276: 35368-35374
Tatham MH, Matic I, Mann M, Hay RT (2011) Comparative proteomic analysis identifies a role for SUMO
in protein quality control. Science signaling 4: rs4
Uzunova K, Gottsche K, Miteva M, Weisshaar SR, Glanemann C, Schnellhardt M, Niessen M, Scheel H,
Hofmann K, Johnson ES, Praefcke GJ, Dohmen RJ (2007) Ubiquitin-dependent proteolytic control of
SUMO conjugates. The Journal of biological chemistry 282: 34167-34175
van de Pasch LA, Miles AJ, Nijenhuis W, Brabers NA, van Leenen D, Lijnzaad P, Brown MK, Ouellet J,
Barral Y, Kops GJ, Holstege FC (2013) Centromere binding and a conserved role in chromosome stability
for SUMO-dependent ubiquitin ligases. PloS one 8: e65628

101
Wagner SA, Beli P, Weinert BT, Nielsen ML, Cox J, Mann M, Choudhary C (2011) A proteome-wide,
quantitative survey of in vivo ubiquitylation sites reveals widespread regulatory roles. Molecular & cellular
proteomics : MCP 10: M111 013284
Wang Z, Prelich G (2009) Quality control of a transcriptional regulator by SUMO-targeted degradation.
Molecular and cellular biology 29: 1694-1706
Watts FZ (2007) The role of SUMO in chromosome segregation. Chromosoma 116: 15-20
Watts FZ, Skilton A, Ho JC, Boyd LK, Trickey MA, Gardner L, Ogi FX, Outwin EA (2007) The role of
Schizosaccharomyces pombe SUMO ligases in genome stability. Biochemical Society transactions 35: 1379-
1384
Westerbeck JW, Pasupala N, Guillotte M, Szymanski E, Matson BC, Esteban C, Kerscher O (2014) A
SUMO-targeted ubiquitin ligase is involved in the degradation of the nuclear pool of the SUMO E3 ligase
Siz1. Molecular biology of the cell 25: 1-16
Westman BJ, Verheggen C, Hutten S, Lam YW, Bertrand E, Lamond AI (2010) A proteomic screen for
nucleolar SUMO targets shows SUMOylation modulates the function of Nop5/Nop58. Molecular cell 39:
618-631
Wilkinson KA, Henley JM (2010) Mechanisms, regulation and consequences of protein SUMOylation. The
Biochemical journal 428: 133-145
Wohlschlegel JA, Johnson ES, Reed SI, Yates JR, 3rd (2004) Global analysis of protein sumoylation in
Saccharomyces cerevisiae. The Journal of biological chemistry 279: 45662-45668
Wohlschlegel JA, Johnson ES, Reed SI, Yates JR, 3rd (2006) Improved identification of SUMO attachment
sites using C-terminal SUMO mutants and tailored protease digestion strategies. Journal of proteome
research 5: 761-770
Xhemalce B, Seeler JS, Thon G, Dejean A, Arcangioli B (2004) Role of the fission yeast SUMO E3 ligase
Pli1p in centromere and telomere maintenance. The EMBO journal 23: 3844-3853
Xu G, Paige JS, Jaffrey SR (2010) Global analysis of lysine ubiquitination by ubiquitin remnant
immunoaffinity profiling. Nature biotechnology 28: 868-873
Yin Y, Seifert A, Chua JS, Maure JF, Golebiowski F, Hay RT (2012) SUMO-targeted ubiquitin E3 ligase
RNF4 is required for the response of human cells to DNA damage. Genes & development 26: 1196-1208
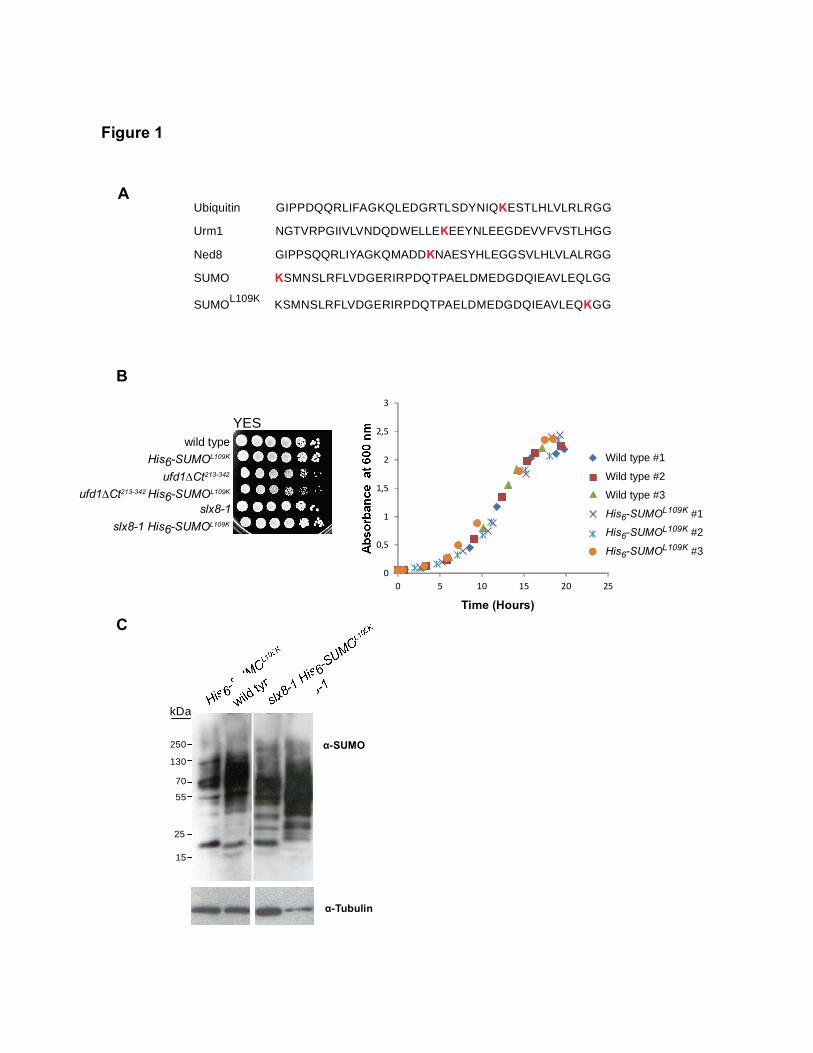
KSMNSLRFLVDGERIRPDQTPAELDMEDGDQIEAVLEQKGGSUMOL109K
Ubiquitin GIPPDQQRLIFAGKQLEDGRTLSDYNIQKESTLHLVLRLRGG
SUMO KSMNSLRFLVDGERIRPDQTPAELDMEDGDQIEAVLEQLGG
Urm1 NGTVRPGIIVLVNDQDWELLEKEEYNLEEGDEVVFVSTLHGG
Ned8 GIPPSQQRLIYAGKQMADDKNAESYHLEGGSVLHLVLALRGG
0
0,5
1
1,5
2
2,5
3
0 5 10 15 20 25
Time (Hours)
Wild type #1
Wild type #2
Wild type #3
His6-SUMOL109K #1
His6-SUMOL109K #2
His6-SUMOL109K #3
YES
ufd1∆Ct213-342 His6-SUMOL109K
His6-SUMOL109K
ufd1∆Ct213-342
wild type
slx8-1 His6-SUMOL109K
slx8-1
A
B
α-Tubulin
α-SUMO
kDa
15
25
55
70
130
250
C
Figure 1
Gen
Typewritten Text
102

ufd1∆Ct213-342 His6-SUMOL109K
Ni2+-purification
Lys-C or Lys-C + Glu-C digestion
DiGly-Lys immunoprecipitation (IP)
LC-MS/MS analysis
A B
wild type
Lys0
Light (L)
Lys0
ufd1∆Ct213-342
Light (L)
Lys8
slx8-1
Heavy (H)
1:1 1:1
LC-MS/MS analysis
398 351 187
Total His6-SUMOL109Ksites identifiedin ufd1∆Ct213-342 non-SILAC experiment
Total His6-SUMOL109K sites identifiedin SILAC experiments
C
D
All sites (518/547) KxE/KxD (188/200)
0.000.00+3.68
-3.68
+3.68
-3.68
+62.69
-62.69
+159.43
-159.43
ExK/DxK (127/131) Residual sites (243/258)
0.000.00+3.66
-3.66
+3.68
-3.68
+87.77
-87.77
+22.17
-22.17
Total proteome analysis
Figure 2
(749) (538)
103

0
1
2
3
4
5
6
7
8
9
0
10
20
30
40
50
A
B
Spc7Fta4Mis17
Ndc80
SPB
Outer kinetochore
Inner kinetochore
Centromericcentral core
Pericentric heterochromatin
Dam1Ask1
Nuf2
DASH complex
Sgo2Cnp3
Pcp1Gamma tubulin complex
Alp4
Bub1
ONMINM
SPB/Kinetochores/Centromeric chromatin
Sad1
Scm3
Mal2Mis13
Clr4Swi6 Epe1
Clr1Pst1
Cnl2
Cbh1Cbh2
Mde4
Monopolin complex
Bqt4 Lem2
Tpz1 Ccq1Rif1Pot1Poz1
Taz1
Rap1
Telomeres/Telomere anchorage to INM
Nuclear telomere cap complex
ONMINM
Figure 3
C
Gen
Typewritten Text
104

Normalized Log2(ufd1∆Ct213-342 / wild type)
Alm1 K505, K639Ccq1 K56, K78
Ask1 K128
Cdc48 K11Mis17 K148
Nse2 K109Rap1 K344Slt1 K383
SUMO K14, K39, K51 and K14;30
Thi2 K38
Adn3 K904
A
B
-4
-3
-2
-1
0
1
2
3
4
-4 -3 -2 -1 0 1 2 3 4
Mot1 K101;105
Dnt1 K89, K101, K270, K282
70178
slx8-1 / ufd1∆Ct213-342 slx8-1 / wild type
DiGly-Lys-modified peptides quantified in at least two replicates
Figure 4
(248)
89
(267)
Gen
Typewritten Text
105

B
-4
-3
-2
-1
0
1
2
3
4
-4 -3 -2 -1 0 1 2 3 4
Crude lysates
Mean normalized log2(ufd1∆Ct213-342/wild type)
Slt1
Ubiquitin
Cdc48SUMO
Crude lysates-4
-3
-2
-1
0
1
2
3
4
-4 -3 -2 -1 0 1 2 3 4
Mean normalized log2(slx8-1/wild type)
SUMO
A
Figure 5
Gen
Typewritten Text
Gen
Typewritten Text
106

0
0,5
1
1,5
2
2,5
3
0 5 10 15 20 25
Time (Hours)
slx8-1 #1slx8-1 #2slx8-1 #3slx8-1 His6-SUMOL109K #1slx8-1 His6-SUMOL109K #2slx8-1 His6-SUMOL109K #3
0
0,5
1
1,5
2
2,5
3
0 10 20 30 40 50
Time (Hours)
ufd1∆Ct213-342 His6-SUMOL109K #1ufd1∆Ct213-342 His6-SUMOL109K #2ufd1∆Ct213-342 His6-SUMOL109K #3
ufd1∆Ct213-342 #1ufd1∆Ct213-342 #2ufd1∆Ct213-342 #3
Figure S1
Gen
Typewritten Text
107

Table S1. List of all His6-SUMOL109K-modified sites identified in this study
108
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp.
Multiplicity Score Multiplicity Score
Q9USY4 ARS-binding protein 2 abp2 95 1 175,45 1 196,97
O14258 Metallothionein expression activator ace2 283 1 107,74
O14258 Metallothionein expression activator ace2 317 1 92,307
O14258 Metallothionein expression activator ace2 376 1 156,11
P10989 Actin act1 326 1 95,428 P10989 Actin act1 61 1 126,19 Q9P7J7 Transcriptional adapter 2 ada2 316 1 99,566
Q9UUB4 Phosphoribosylaminoimidazole-succinocarboxamide synthase ade7 5 1 88,552
P00332 Alcohol dehydrogenase adh1 22 1 100,04 1 68,173 P00332 Alcohol dehydrogenase adh1 253 1 79,467 1 40,622 P00332 Alcohol dehydrogenase adh1 86 1 145,55 P00332 Alcohol dehydrogenase adh1 317 1 44,426 O94619 Adhesion defective protein 2 adn2 19 1 120,33 O74522 Adhesion defective protein 3 adn3 904 1 193,8 1 192,16 O74522 Adhesion defective protein 3 adn3 915 1 155,25 1 160,36
Q9UTK5 Abnormal long morphology protein 1 alm1 275 1 142,26 1 120,11
Q9UTK5 Abnormal long morphology protein 1 alm1 358 1 167,63 1 159,75
Q9UTK5 Abnormal long morphology protein 1 alm1 388 1 96,012 1 123,55
Q9UTK5 Abnormal long morphology protein 1 alm1 505 1 96,993 1 120,66
Q9UTK5 Abnormal long morphology protein 1 alm1 602 1 180,49 1 178,03
Q9UTK5 Abnormal long morphology protein 1 alm1 639 1 62,891 1 113,25
Q9UTK5 Abnormal long morphology protein 1 alm1 668 1 67,385 1 63,091
Q9UTK5 Abnormal long morphology protein 1 alm1 709 1 96,317 1 87,098
Q9UTK5 Abnormal long morphology protein 1 alm1 765 1 45,723 1 128,9
Q9UTK5 Abnormal long morphology protein 1 alm1 1100 1 103,7 1 114,21
Q9UTK5 Abnormal long morphology protein 1 alm1 1114 1 58,92 1 153,1
Q9UTK5 Abnormal long morphology protein 1 alm1 355 1 61,732
Q9UTK5 Abnormal long morphology protein 1 alm1 976 1 134,37
Q9UTK5 Abnormal long morphology protein 1 alm1 1095 1 101,53
Q9UTK5 Abnormal long morphology protein 1 alm1 265 1 138,09
Q9Y705 Spindle pole body component alp4 alp4 11 1 74,698 Q9Y705 Spindle pole body component alp4 alp4 529 1 142,76 Q9P7X7 Actin-related protein 4 alp5 300 1 80,522 Q9P7E8 Protein app1 app1 308 1 69,522
O74548 Probable acetylornithine aminotransferase, mitochondrial arg1 41 1 186,16
Q9Y7X8 Actin-like protein arp5 arp5 348 1 35,118
Q9UTQ7 SWI/SNF and RSC complexes subunit arp9 arp9 64 1 119,56 1 150,59
Q9UTQ7 SWI/SNF and RSC complexes subunit arp9 arp9 69 1 135,68 1 220,23
Q9UTQ7 SWI/SNF and RSC complexes subunit arp9 arp9 287 1 123,14 1 148,89
Q9P6S5 DASH complex subunit ask1 ask1 128 1 90,193 1 97,253
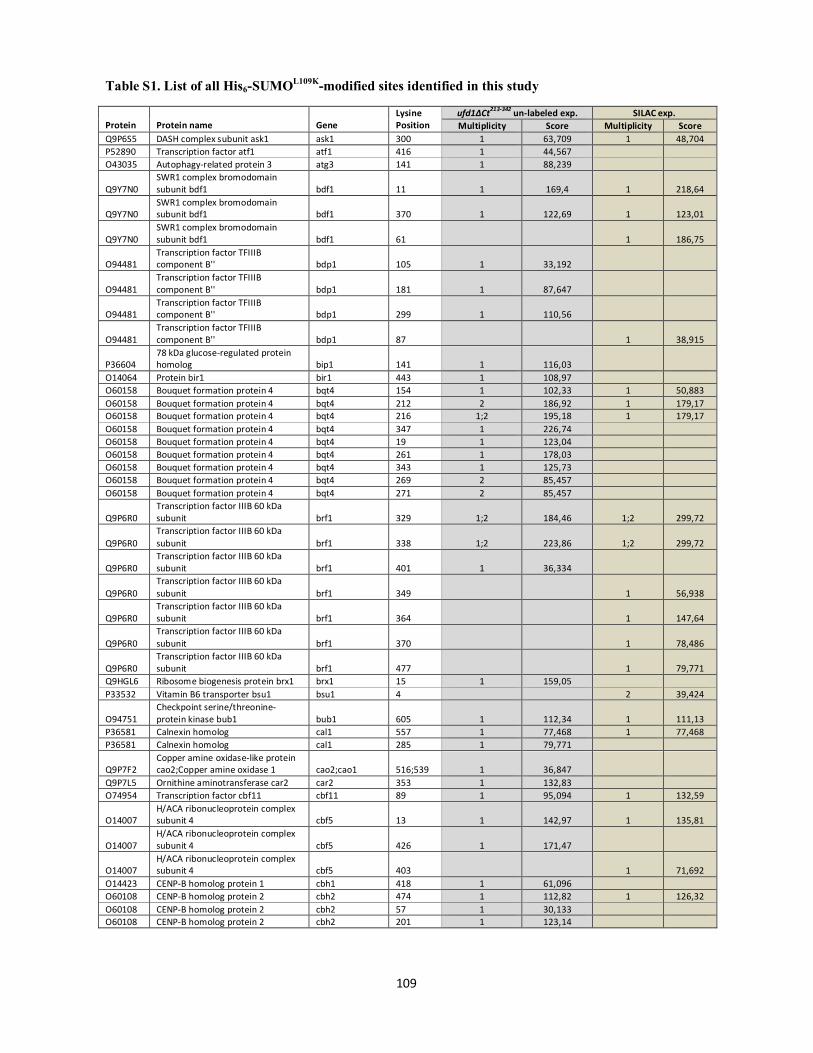
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
109
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
Q9P6S5 DASH complex subunit ask1 ask1 300 1 63,709 1 48,704 P52890 Transcription factor atf1 atf1 416 1 44,567 O43035 Autophagy-related protein 3 atg3 141 1 88,239
Q9Y7N0 SWR1 complex bromodomain subunit bdf1 bdf1 11 1 169,4 1 218,64
Q9Y7N0 SWR1 complex bromodomain subunit bdf1 bdf1 370 1 122,69 1 123,01
Q9Y7N0 SWR1 complex bromodomain subunit bdf1 bdf1 61 1 186,75
O94481 Transcription factor TFIIIB component B'' bdp1 105 1 33,192
O94481 Transcription factor TFIIIB component B'' bdp1 181 1 87,647
O94481 Transcription factor TFIIIB component B'' bdp1 299 1 110,56
O94481 Transcription factor TFIIIB component B'' bdp1 87 1 38,915
P36604 78 kDa glucose-regulated protein homolog bip1 141 1 116,03
O14064 Protein bir1 bir1 443 1 108,97 O60158 Bouquet formation protein 4 bqt4 154 1 102,33 1 50,883 O60158 Bouquet formation protein 4 bqt4 212 2 186,92 1 179,17 O60158 Bouquet formation protein 4 bqt4 216 1;2 195,18 1 179,17 O60158 Bouquet formation protein 4 bqt4 347 1 226,74 O60158 Bouquet formation protein 4 bqt4 19 1 123,04 O60158 Bouquet formation protein 4 bqt4 261 1 178,03 O60158 Bouquet formation protein 4 bqt4 343 1 125,73 O60158 Bouquet formation protein 4 bqt4 269 2 85,457 O60158 Bouquet formation protein 4 bqt4 271 2 85,457
Q9P6R0 Transcription factor IIIB 60 kDa subunit brf1 329 1;2 184,46 1;2 299,72
Q9P6R0 Transcription factor IIIB 60 kDa subunit brf1 338 1;2 223,86 1;2 299,72
Q9P6R0 Transcription factor IIIB 60 kDa subunit brf1 401 1 36,334
Q9P6R0 Transcription factor IIIB 60 kDa subunit brf1 349 1 56,938
Q9P6R0 Transcription factor IIIB 60 kDa subunit brf1 364 1 147,64
Q9P6R0 Transcription factor IIIB 60 kDa subunit brf1 370 1 78,486
Q9P6R0 Transcription factor IIIB 60 kDa subunit brf1 477 1 79,771
Q9HGL6 Ribosome biogenesis protein brx1 brx1 15 1 159,05 P33532 Vitamin B6 transporter bsu1 bsu1 4 2 39,424
O94751 Checkpoint serine/threonine-protein kinase bub1 bub1 605 1 112,34 1 111,13
P36581 Calnexin homolog cal1 557 1 77,468 1 77,468 P36581 Calnexin homolog cal1 285 1 79,771
Q9P7F2 Copper amine oxidase-like protein cao2;Copper amine oxidase 1 cao2;cao1 516;539 1 36,847
Q9P7L5 Ornithine aminotransferase car2 car2 353 1 132,83 O74954 Transcription factor cbf11 cbf11 89 1 95,094 1 132,59
O14007 H/ACA ribonucleoprotein complex subunit 4 cbf5 13 1 142,97 1 135,81
O14007 H/ACA ribonucleoprotein complex subunit 4 cbf5 426 1 171,47
O14007 H/ACA ribonucleoprotein complex subunit 4 cbf5 403 1 71,692
O14423 CENP-B homolog protein 1 cbh1 418 1 61,096 O60108 CENP-B homolog protein 2 cbh2 474 1 112,82 1 126,32 O60108 CENP-B homolog protein 2 cbh2 57 1 30,133 O60108 CENP-B homolog protein 2 cbh2 201 1 123,14

Table S1. List of all His6-SUMOL109K-modified sites identified in this study
110
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 56 1 81,224 1 129,2
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 128 1 119,16 1 120,23
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 491 1 233,49 1 239,58
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 78 1 194,13
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 456 1 81,847
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 584 1 101,01
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 625 1 205,62
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 641 1 113,42
O74874
Glucose-repressible alcohol dehydrogenase transcriptional effector ccr4 677 1 34,695
P36602 Ribonucleoside-diphosphate reductase large chain cdc22 785 1 162,59
P30261 DNA polymerase delta subunit 3 cdc27 317 1 59,35 Q9P3A7 Cell division cycle protein 48 cdc48 11 1 177,12 1 155,22 Q9P3A7 Cell division cycle protein 48 cdc48 156 1 56,432 Q9UUE7 Cell division control protein 73 cdc73 18 1 67,095 Q02088 Tropomyosin cdc8 65 1 262,77 1 235,73 Q02088 Tropomyosin cdc8 3 1 74,424 O74740 Cleavage factor two protein 2 cft2 684 1 131,45 O59732 Chitin synthase regulatory factor 1 chr1 34 1 127,83 1 112,85
Q10306 Probable citrate synthase, mitochondrial cit1 477 1 30,331
O74808 Cryptic loci regulator protein 1 clr1 286 1 43,791
O60016
Histone-lysine N-methyltransferase, H3 lysine-9 specific clr4 4 1 173,56 1 142,84
O60016
Histone-lysine N-methyltransferase, H3 lysine-9 specific clr4 109 1 72,793 1 98,804
O60016
Histone-lysine N-methyltransferase, H3 lysine-9 specific clr4 174 1 105,6
O59702 Histone deacetylase clr6 clr6 397 1 178,15 1 62,162 O59702 Histone deacetylase clr6 clr6 392 1 162,46 O94679 Condensin complex subunit 1 cnd1 460 1 93,152 Q9Y7R3 Condensin complex subunit 2 cnd2 318 1 101,3 1 141,1 Q9Y7R3 Condensin complex subunit 2 cnd2 553 1 137,78 1 74,392 Q9Y7R3 Condensin complex subunit 2 cnd2 689 1 107,07 1 159,31 Q9Y7R3 Condensin complex subunit 2 cnd2 379 1 60,178 Q9Y7R3 Condensin complex subunit 2 cnd2 168 1 129,15 Q9Y7R3 Condensin complex subunit 2 cnd2 309 1 69,812 Q10429 Condensin complex subunit 3 cnd3 870 1 99,855 O13952 Centromere-localized protein 2 cnl2 85 1 36,856 Q9USR9 Centromere protein 3 cnp3 481 1 49,436 1 56,746 Q9USR9 Centromere protein 3 cnp3 37 1 113,37 Q9USR9 Centromere protein 3 cnp3 226 1 83,088 O13849 Carboxypeptidase Y cpy1 487 1 58,04 O13849 Carboxypeptidase Y cpy1 577 1 81,904 Q9UUE2 Protein csx2 csx2 67 1 108,7 1 57,348
Q1MTR4 Putative histone deacetylase complex subunit cti6 cti6 207 1 178,68
Q1MTR4 Putative histone deacetylase complex subunit cti6 cti6 263 1 109,62

Table S1. List of all His6-SUMOL109K-modified sites identified in this study
111
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
O74986 DNA endonuclease ctp1 ctp1 137 1 158,01
Q09728 Metal-binding regulatory protein cuf1 cuf1 160 1 130,21 1 122,75
Q09728 Metal-binding regulatory protein cuf1 cuf1 373 1 51,348
P41003 Structural maintenance of chromosomes protein 2 cut14 824 1 93,614 1 127,56
P41003 Structural maintenance of chromosomes protein 2 cut14 852 1 68,016 1 76,679
P41003 Structural maintenance of chromosomes protein 2 cut14 321 1 107,15
P41003 Structural maintenance of chromosomes protein 2 cut14 349 1 73,781
P41003 Structural maintenance of chromosomes protein 2 cut14 413 1 61,862
P41004 Structural maintenance of chromosomes protein 4 cut3 47 1 84,498 1 239,47
P41004 Structural maintenance of chromosomes protein 4 cut3 89 1 62,055 1 62,861
P41004 Structural maintenance of chromosomes protein 4 cut3 340 1 138,87 1 162,37
P24339 Kinesin-like protein cut7 cut7 955 1 120,24 P24339 Kinesin-like protein cut7 cut7 1004 1 99,139 O94316 Pre-mRNA-splicing factor cwf10 cwf10 75 1 101,38 Q9UU80 Pre-mRNA-splicing factor cwf18 cwf18 70 1 34,755 Q9P6R9 Pre-mRNA-splicing factor cwf22 cwf22 609 1 33,542 Q9USV3 Pre-mRNA-splicing factor cwf7 cwf7 42 1 61,167 Q9HDZ6 DASH complex subunit dam1 dam1 19 1 90,853 1 150,11 Q09747 ATP-dependent RNA helicase dbp5 dbp5 43 1 53,565 Q76PD3 ATP-dependent RNA helicase dbp6 dbp6 100 1 72,089 O74354 Nucleolar protein dnt1 dnt1 89 1 163,78 1;2 150,06 O74354 Nucleolar protein dnt1 dnt1 101 1 79,875 1 110,56 O74354 Nucleolar protein dnt1 dnt1 200 1 158,29 1 178,03 O74354 Nucleolar protein dnt1 dnt1 270 1 200,07 1;2 176,27 O74354 Nucleolar protein dnt1 dnt1 282 1 109,85 1;2 162,07 O74354 Nucleolar protein dnt1 dnt1 293 1 154,82 1 154,84 O74354 Nucleolar protein dnt1 dnt1 394 1 50,079 1 57,173 O74354 Nucleolar protein dnt1 dnt1 99 2 88,101 1;2 94,059 O74354 Nucleolar protein dnt1 dnt1 90 1;2 222,34 1;2 136,81 O74354 Nucleolar protein dnt1 dnt1 512 1 47,198 O74354 Nucleolar protein dnt1 dnt1 525 1 103,9 Q10315 DNA polymerase epsilon subunit C dpb3 161 1 292,47
O94752 Enhancer of mRNA-decapping protein 3 edc3 152 1 58,172
O14460 Elongation factor 2 eft201 42 1 62,765 O14460 Elongation factor 2 eft201 829 1 40,085
P87147 Nascent polypeptide-associated complex subunit alpha egd2 10 1 83,881
P87147 Nascent polypeptide-associated complex subunit alpha egd2 99 1 114,15
P87147 Nascent polypeptide-associated complex subunit alpha egd2 118 1 134,31
P87147 Nascent polypeptide-associated complex subunit alpha egd2 152 1 154,11
P40370 Enolase 1-1 eno101 79 1 221,25 1 240,71 P40370 Enolase 1-1 eno101 88 1 111,92 1 131,51 P40370 Enolase 1-1 eno101 96 1 84,946 1 87,519 P40370 Enolase 1-1 eno101 263 1 124,39 1 97,779 P40370 Enolase 1-1 eno101 287 1 34,498 1 115,31 P40370 Enolase 1-1 eno101 337 1 104,01 P40370 Enolase 1-1 eno101 64 1 142,43 P40370 Enolase 1-1 eno101 199 1 59,793 O74399 Ribosome biogenesis protein erb1 erb1 52 1 140,64
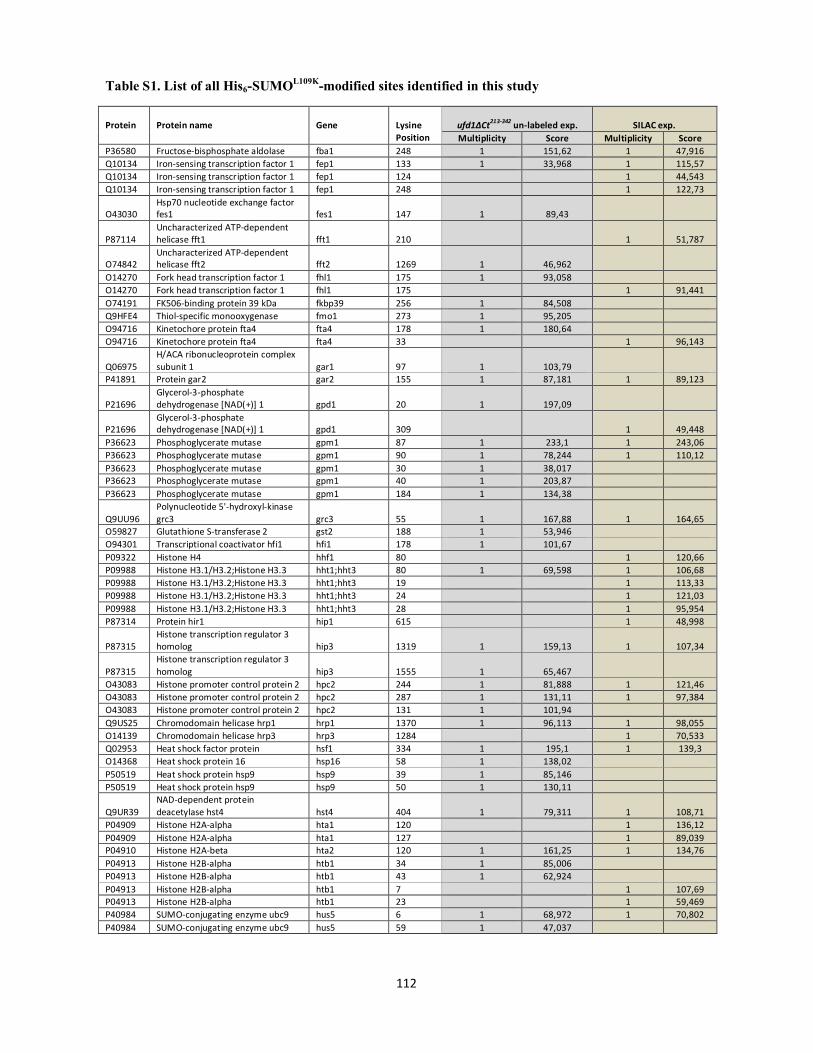
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
112
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
P36580 Fructose-bisphosphate aldolase fba1 248 1 151,62 1 47,916 Q10134 Iron-sensing transcription factor 1 fep1 133 1 33,968 1 115,57 Q10134 Iron-sensing transcription factor 1 fep1 124 1 44,543 Q10134 Iron-sensing transcription factor 1 fep1 248 1 122,73
O43030 Hsp70 nucleotide exchange factor fes1 fes1 147 1 89,43
P87114 Uncharacterized ATP-dependent helicase fft1 fft1 210 1 51,787
O74842 Uncharacterized ATP-dependent helicase fft2 fft2 1269 1 46,962
O14270 Fork head transcription factor 1 fhl1 175 1 93,058 O14270 Fork head transcription factor 1 fhl1 175 1 91,441 O74191 FK506-binding protein 39 kDa fkbp39 256 1 84,508 Q9HFE4 Thiol-specific monooxygenase fmo1 273 1 95,205 O94716 Kinetochore protein fta4 fta4 178 1 180,64 O94716 Kinetochore protein fta4 fta4 33 1 96,143
Q06975 H/ACA ribonucleoprotein complex subunit 1 gar1 97 1 103,79
P41891 Protein gar2 gar2 155 1 87,181 1 89,123
P21696 Glycerol-3-phosphate dehydrogenase [NAD(+)] 1 gpd1 20 1 197,09
P21696 Glycerol-3-phosphate dehydrogenase [NAD(+)] 1 gpd1 309 1 49,448
P36623 Phosphoglycerate mutase gpm1 87 1 233,1 1 243,06 P36623 Phosphoglycerate mutase gpm1 90 1 78,244 1 110,12 P36623 Phosphoglycerate mutase gpm1 30 1 38,017 P36623 Phosphoglycerate mutase gpm1 40 1 203,87 P36623 Phosphoglycerate mutase gpm1 184 1 134,38
Q9UU96 Polynucleotide 5'-hydroxyl-kinase grc3 grc3 55 1 167,88 1 164,65
O59827 Glutathione S-transferase 2 gst2 188 1 53,946 O94301 Transcriptional coactivator hfi1 hfi1 178 1 101,67 P09322 Histone H4 hhf1 80 1 120,66 P09988 Histone H3.1/H3.2;Histone H3.3 hht1;hht3 80 1 69,598 1 106,68 P09988 Histone H3.1/H3.2;Histone H3.3 hht1;hht3 19 1 113,33 P09988 Histone H3.1/H3.2;Histone H3.3 hht1;hht3 24 1 121,03 P09988 Histone H3.1/H3.2;Histone H3.3 hht1;hht3 28 1 95,954 P87314 Protein hir1 hip1 615 1 48,998
P87315 Histone transcription regulator 3 homolog hip3 1319 1 159,13 1 107,34
P87315 Histone transcription regulator 3 homolog hip3 1555 1 65,467
O43083 Histone promoter control protein 2 hpc2 244 1 81,888 1 121,46 O43083 Histone promoter control protein 2 hpc2 287 1 131,11 1 97,384 O43083 Histone promoter control protein 2 hpc2 131 1 101,94 Q9US25 Chromodomain helicase hrp1 hrp1 1370 1 96,113 1 98,055 O14139 Chromodomain helicase hrp3 hrp3 1284 1 70,533 Q02953 Heat shock factor protein hsf1 334 1 195,1 1 139,3 O14368 Heat shock protein 16 hsp16 58 1 138,02 P50519 Heat shock protein hsp9 hsp9 39 1 85,146 P50519 Heat shock protein hsp9 hsp9 50 1 130,11
Q9UR39 NAD-dependent protein deacetylase hst4 hst4 404 1 79,311 1 108,71
P04909 Histone H2A-alpha hta1 120 1 136,12 P04909 Histone H2A-alpha hta1 127 1 89,039 P04910 Histone H2A-beta hta2 120 1 161,25 1 134,76 P04913 Histone H2B-alpha htb1 34 1 85,006 P04913 Histone H2B-alpha htb1 43 1 62,924 P04913 Histone H2B-alpha htb1 7 1 107,69 P04913 Histone H2B-alpha htb1 23 1 59,469 P40984 SUMO-conjugating enzyme ubc9 hus5 6 1 68,972 1 70,802 P40984 SUMO-conjugating enzyme ubc9 hus5 59 1 47,037
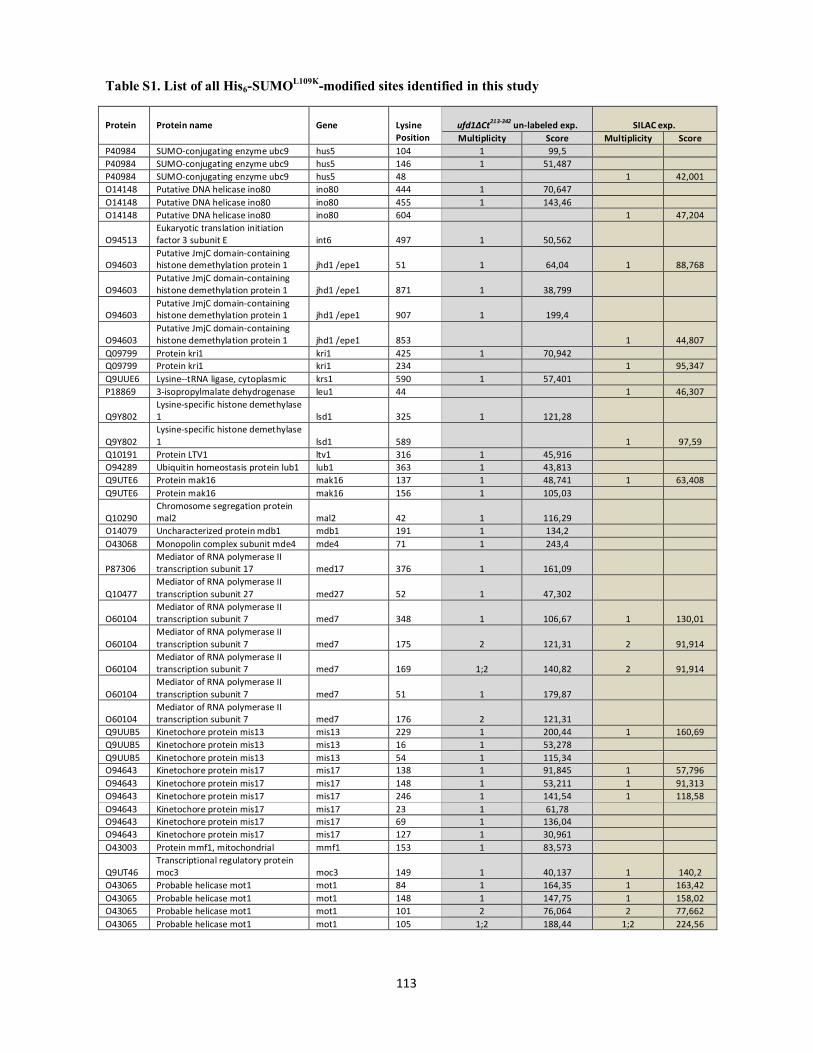
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
113
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
P40984 SUMO-conjugating enzyme ubc9 hus5 104 1 99,5 P40984 SUMO-conjugating enzyme ubc9 hus5 146 1 51,487 P40984 SUMO-conjugating enzyme ubc9 hus5 48 1 42,001 O14148 Putative DNA helicase ino80 ino80 444 1 70,647 O14148 Putative DNA helicase ino80 ino80 455 1 143,46 O14148 Putative DNA helicase ino80 ino80 604 1 47,204
O94513 Eukaryotic translation initiation factor 3 subunit E int6 497 1 50,562
O94603 Putative JmjC domain-containing histone demethylation protein 1 jhd1 /epe1 51 1 64,04 1 88,768
O94603 Putative JmjC domain-containing histone demethylation protein 1 jhd1 /epe1 871 1 38,799
O94603 Putative JmjC domain-containing histone demethylation protein 1 jhd1 /epe1 907 1 199,4
O94603 Putative JmjC domain-containing histone demethylation protein 1 jhd1 /epe1 853 1 44,807
Q09799 Protein kri1 kri1 425 1 70,942 Q09799 Protein kri1 kri1 234 1 95,347 Q9UUE6 Lysine--tRNA ligase, cytoplasmic krs1 590 1 57,401 P18869 3-isopropylmalate dehydrogenase leu1 44 1 46,307
Q9Y802 Lysine-specific histone demethylase 1 lsd1 325 1 121,28
Q9Y802 Lysine-specific histone demethylase 1 lsd1 589 1 97,59
Q10191 Protein LTV1 ltv1 316 1 45,916 O94289 Ubiquitin homeostasis protein lub1 lub1 363 1 43,813 Q9UTE6 Protein mak16 mak16 137 1 48,741 1 63,408 Q9UTE6 Protein mak16 mak16 156 1 105,03
Q10290 Chromosome segregation protein mal2 mal2 42 1 116,29
O14079 Uncharacterized protein mdb1 mdb1 191 1 134,2 O43068 Monopolin complex subunit mde4 mde4 71 1 243,4
P87306 Mediator of RNA polymerase II transcription subunit 17 med17 376 1 161,09
Q10477 Mediator of RNA polymerase II transcription subunit 27 med27 52 1 47,302
O60104 Mediator of RNA polymerase II transcription subunit 7 med7 348 1 106,67 1 130,01
O60104 Mediator of RNA polymerase II transcription subunit 7 med7 175 2 121,31 2 91,914
O60104 Mediator of RNA polymerase II transcription subunit 7 med7 169 1;2 140,82 2 91,914
O60104 Mediator of RNA polymerase II transcription subunit 7 med7 51 1 179,87
O60104 Mediator of RNA polymerase II transcription subunit 7 med7 176 2 121,31
Q9UUB5 Kinetochore protein mis13 mis13 229 1 200,44 1 160,69 Q9UUB5 Kinetochore protein mis13 mis13 16 1 53,278 Q9UUB5 Kinetochore protein mis13 mis13 54 1 115,34 O94643 Kinetochore protein mis17 mis17 138 1 91,845 1 57,796 O94643 Kinetochore protein mis17 mis17 148 1 53,211 1 91,313 O94643 Kinetochore protein mis17 mis17 246 1 141,54 1 118,58 O94643 Kinetochore protein mis17 mis17 23 1 61,78 O94643 Kinetochore protein mis17 mis17 69 1 136,04 O94643 Kinetochore protein mis17 mis17 127 1 30,961 O43003 Protein mmf1, mitochondrial mmf1 153 1 83,573
Q9UT46 Transcriptional regulatory protein moc3 moc3 149 1 40,137 1 140,2
O43065 Probable helicase mot1 mot1 84 1 164,35 1 163,42 O43065 Probable helicase mot1 mot1 148 1 147,75 1 158,02 O43065 Probable helicase mot1 mot1 101 2 76,064 2 77,662 O43065 Probable helicase mot1 mot1 105 1;2 188,44 1;2 224,56
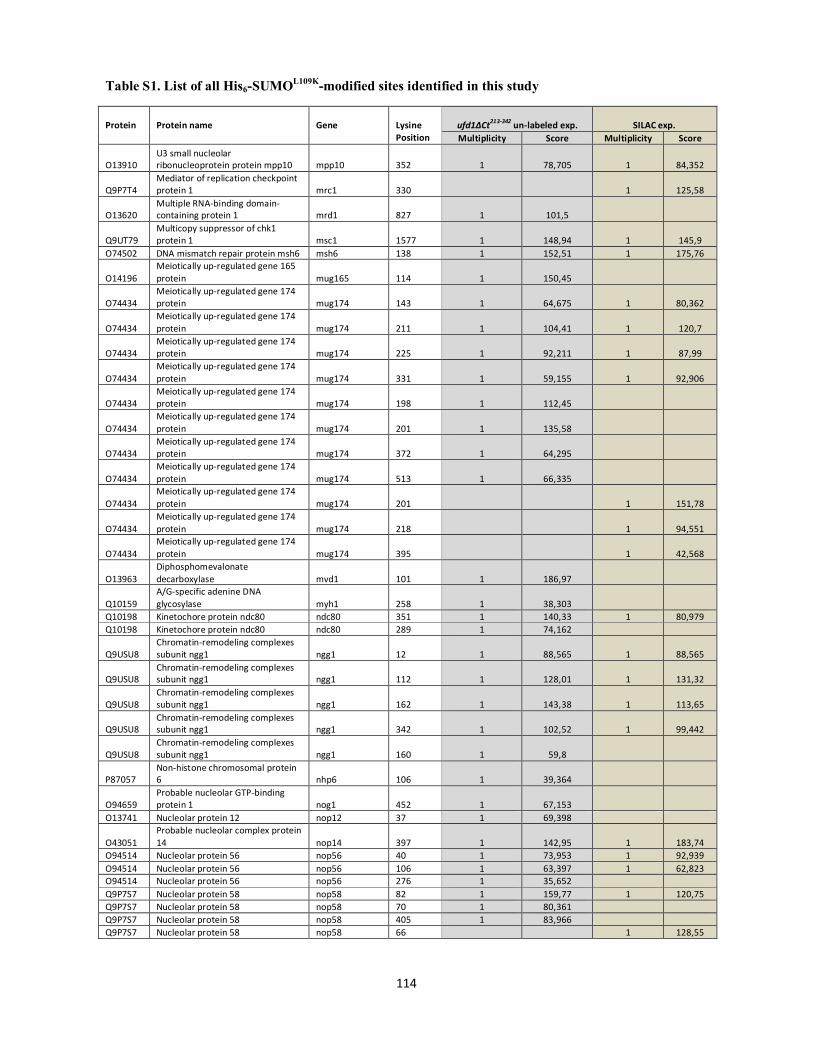
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
114
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
O13910 U3 small nucleolar ribonucleoprotein protein mpp10 mpp10 352 1 78,705 1 84,352
Q9P7T4 Mediator of replication checkpoint protein 1 mrc1 330 1 125,58
O13620 Multiple RNA-binding domain-containing protein 1 mrd1 827 1 101,5
Q9UT79 Multicopy suppressor of chk1 protein 1 msc1 1577 1 148,94 1 145,9
O74502 DNA mismatch repair protein msh6 msh6 138 1 152,51 1 175,76
O14196 Meiotically up-regulated gene 165 protein mug165 114 1 150,45
O74434 Meiotically up-regulated gene 174 protein mug174 143 1 64,675 1 80,362
O74434 Meiotically up-regulated gene 174 protein mug174 211 1 104,41 1 120,7
O74434 Meiotically up-regulated gene 174 protein mug174 225 1 92,211 1 87,99
O74434 Meiotically up-regulated gene 174 protein mug174 331 1 59,155 1 92,906
O74434 Meiotically up-regulated gene 174 protein mug174 198 1 112,45
O74434 Meiotically up-regulated gene 174 protein mug174 201 1 135,58
O74434 Meiotically up-regulated gene 174 protein mug174 372 1 64,295
O74434 Meiotically up-regulated gene 174 protein mug174 513 1 66,335
O74434 Meiotically up-regulated gene 174 protein mug174 201 1 151,78
O74434 Meiotically up-regulated gene 174 protein mug174 218 1 94,551
O74434 Meiotically up-regulated gene 174 protein mug174 395 1 42,568
O13963 Diphosphomevalonate decarboxylase mvd1 101 1 186,97
Q10159 A/G-specific adenine DNA glycosylase myh1 258 1 38,303
Q10198 Kinetochore protein ndc80 ndc80 351 1 140,33 1 80,979 Q10198 Kinetochore protein ndc80 ndc80 289 1 74,162
Q9USU8 Chromatin-remodeling complexes subunit ngg1 ngg1 12 1 88,565 1 88,565
Q9USU8 Chromatin-remodeling complexes subunit ngg1 ngg1 112 1 128,01 1 131,32
Q9USU8 Chromatin-remodeling complexes subunit ngg1 ngg1 162 1 143,38 1 113,65
Q9USU8 Chromatin-remodeling complexes subunit ngg1 ngg1 342 1 102,52 1 99,442
Q9USU8 Chromatin-remodeling complexes subunit ngg1 ngg1 160 1 59,8
P87057 Non-histone chromosomal protein 6 nhp6 106 1 39,364
O94659 Probable nucleolar GTP-binding protein 1 nog1 452 1 67,153
O13741 Nucleolar protein 12 nop12 37 1 69,398
O43051 Probable nucleolar complex protein 14 nop14 397 1 142,95 1 183,74
O94514 Nucleolar protein 56 nop56 40 1 73,953 1 92,939 O94514 Nucleolar protein 56 nop56 106 1 63,397 1 62,823 O94514 Nucleolar protein 56 nop56 276 1 35,652 Q9P7S7 Nucleolar protein 58 nop58 82 1 159,77 1 120,75 Q9P7S7 Nucleolar protein 58 nop58 70 1 80,361 Q9P7S7 Nucleolar protein 58 nop58 405 1 83,966 Q9P7S7 Nucleolar protein 58 nop58 66 1 128,55

Table S1. List of all His6-SUMOL109K-modified sites identified in this study
115
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
O94698 Ribosome biogenesis protein nsa1 nsa1 351 1 138,97 1;2 238,68 O94698 Ribosome biogenesis protein nsa1 nsa1 379 2 102,48 2 169,84 O94698 Ribosome biogenesis protein nsa1 nsa1 380 2 102,48 2 169,84 O94698 Ribosome biogenesis protein nsa1 nsa1 214 1 122,42 O94698 Ribosome biogenesis protein nsa1 nsa1 214 1 151,98 O94698 Ribosome biogenesis protein nsa1 nsa1 352 2 157,27 Q4PIR3 E3 SUMO-protein ligase nse2 nse2 109 1 185,37 1 154,74
Q9Y7U4 Non-structural maintenance of chromosome element 3 nse3 9 1 146,5
Q09907 Endonuclease III homolog nth1 280 2 41,724 Q09907 Endonuclease III homolog nth1 284 1;2 87,729 Q10173 Kinetochore protein nuf2 nuf2 161 1 82,705 Q10173 Kinetochore protein nuf2 nuf2 245 1 53,001 Q10173 Kinetochore protein nuf2 nuf2 338 1 150,69 Q9UTK4 Nucleoporin nup189 nup189 1000 1 73,546 O74424 Nucleoporin nup211 nup211 1490 1 121,61 1 126,95 O74424 Nucleoporin nup211 nup211 397 1 60,345 O74424 Nucleoporin nup211 nup211 532 1 33,685 O74424 Nucleoporin nup211 nup211 589 1 127,22 O74500 Nucleoporin nup60 nup60 344 1 94,281 1 102,33 O74500 Nucleoporin nup60 nup60 139 1 43,306 O74500 Nucleoporin nup60 nup60 732 1 166,62 Q9USL4 Nucleoporin nup61 nup61 40 1 35,757 P30821 P25 protein obr1 35 1 98,132
Q9Y794 Origin recognition complex subunit 4 orc4 211 1 144,3 1 106
O43114 Origin recognition complex subunit 5 orc5 265 1 92,29
Q01663 AP-1-like transcription factor pap1 18 1 194,25 1 260,82 Q92351 Spindle pole body protein pcp1 pcp1 33 1 143,57 1 160,13 Q92351 Spindle pole body protein pcp1 pcp1 79 1 203,99 1 109,42 Q92351 Spindle pole body protein pcp1 pcp1 188 1 138,39 1 128,9 Q92351 Spindle pole body protein pcp1 pcp1 14 1;2 171,98 1;2 155,79 Q92351 Spindle pole body protein pcp1 pcp1 30 1;2 156,92 2 60,133 Q92351 Spindle pole body protein pcp1 pcp1 91 1 50,029 Q92351 Spindle pole body protein pcp1 pcp1 106 1 44,788 Q92351 Spindle pole body protein pcp1 pcp1 126 1 117,36 Q92351 Spindle pole body protein pcp1 pcp1 234 1 156,49 Q92351 Spindle pole body protein pcp1 pcp1 276 1 64,394 Q92351 Spindle pole body protein pcp1 pcp1 535 1 64,64
Q9P6Q6 mRNA-capping enzyme subunit beta pct1 4 1 102,89
O94516 Peroxisomal membrane protein PEX16 pex16 80 1 61,691
O60101 Phosphoglycerate kinase pgk1 6 1 125,53 1 150,12 O60101 Phosphoglycerate kinase pgk1 272 1 105,9 1 100,77 O60101 Phosphoglycerate kinase pgk1 15 1 107,11 O60101 Phosphoglycerate kinase pgk1 139 1 60,474 O60101 Phosphoglycerate kinase pgk1 86 1 151,56
Q09908 SWM histone demethylase complex subunit phf2 phf2 290 1 145,75
P28745 Protein pim1 pim1 528 1 54,524 O13959 RING-box protein pip1 pip1 9 1 41,598 1 100,55
O94395 ATP-dependent DNA helicase II subunit 1 pku70 528 1 124,45
O94395 ATP-dependent DNA helicase II subunit 1 pku70 548 1 52,265
O94395 ATP-dependent DNA helicase II subunit 1 pku70 605 1 109,07
Q10295 Poly(A) polymerase pla1 pla1 449 1 56,407 O94451 E3 SUMO-protein ligase pli1 pli1 15 1 89,959 O94451 E3 SUMO-protein ligase pli1 pli1 472 1 42,338
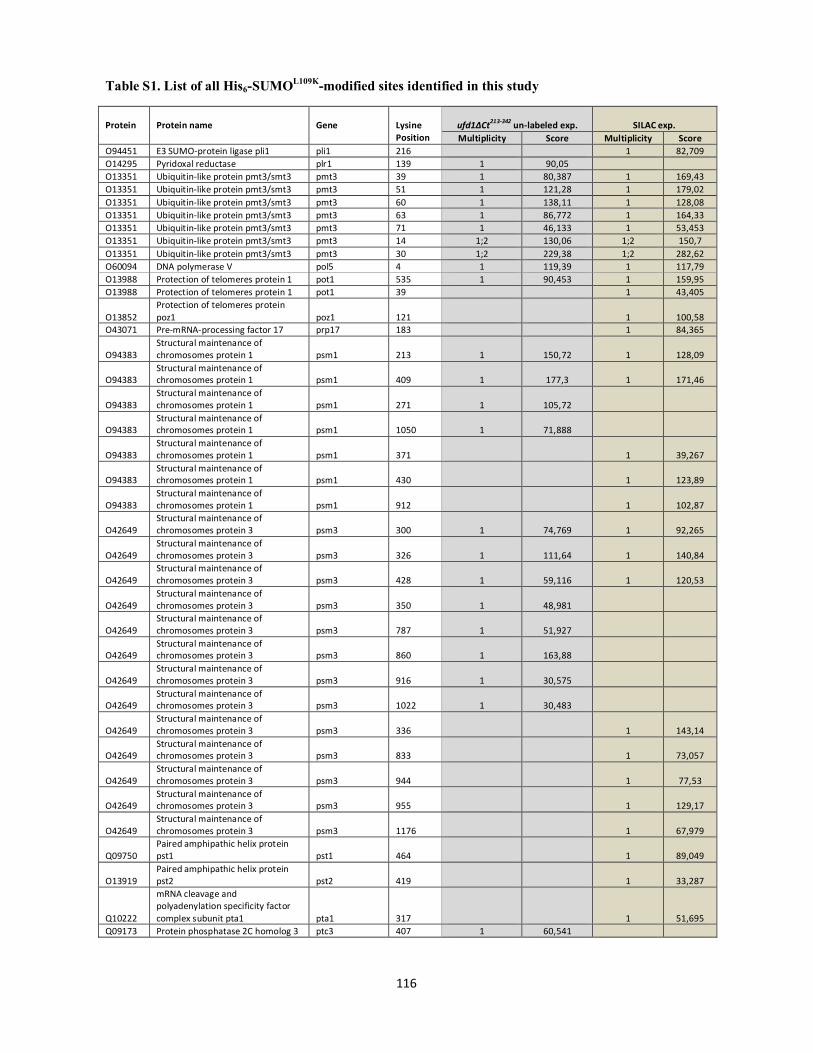
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
116
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
O94451 E3 SUMO-protein ligase pli1 pli1 216 1 82,709 O14295 Pyridoxal reductase plr1 139 1 90,05 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 39 1 80,387 1 169,43 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 51 1 121,28 1 179,02 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 60 1 138,11 1 128,08 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 63 1 86,772 1 164,33 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 71 1 46,133 1 53,453 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 14 1;2 130,06 1;2 150,7 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 30 1;2 229,38 1;2 282,62 O60094 DNA polymerase V pol5 4 1 119,39 1 117,79 O13988 Protection of telomeres protein 1 pot1 535 1 90,453 1 159,95 O13988 Protection of telomeres protein 1 pot1 39 1 43,405
O13852 Protection of telomeres protein poz1 poz1 121 1 100,58
O43071 Pre-mRNA-processing factor 17 prp17 183 1 84,365
O94383 Structural maintenance of chromosomes protein 1 psm1 213 1 150,72 1 128,09
O94383 Structural maintenance of chromosomes protein 1 psm1 409 1 177,3 1 171,46
O94383 Structural maintenance of chromosomes protein 1 psm1 271 1 105,72
O94383 Structural maintenance of chromosomes protein 1 psm1 1050 1 71,888
O94383 Structural maintenance of chromosomes protein 1 psm1 371 1 39,267
O94383 Structural maintenance of chromosomes protein 1 psm1 430 1 123,89
O94383 Structural maintenance of chromosomes protein 1 psm1 912 1 102,87
O42649 Structural maintenance of chromosomes protein 3 psm3 300 1 74,769 1 92,265
O42649 Structural maintenance of chromosomes protein 3 psm3 326 1 111,64 1 140,84
O42649 Structural maintenance of chromosomes protein 3 psm3 428 1 59,116 1 120,53
O42649 Structural maintenance of chromosomes protein 3 psm3 350 1 48,981
O42649 Structural maintenance of chromosomes protein 3 psm3 787 1 51,927
O42649 Structural maintenance of chromosomes protein 3 psm3 860 1 163,88
O42649 Structural maintenance of chromosomes protein 3 psm3 916 1 30,575
O42649 Structural maintenance of chromosomes protein 3 psm3 1022 1 30,483
O42649 Structural maintenance of chromosomes protein 3 psm3 336 1 143,14
O42649 Structural maintenance of chromosomes protein 3 psm3 833 1 73,057
O42649 Structural maintenance of chromosomes protein 3 psm3 944 1 77,53
O42649 Structural maintenance of chromosomes protein 3 psm3 955 1 129,17
O42649 Structural maintenance of chromosomes protein 3 psm3 1176 1 67,979
Q09750 Paired amphipathic helix protein pst1 pst1 464 1 89,049
O13919 Paired amphipathic helix protein pst2 pst2 419 1 33,287
Q10222
mRNA cleavage and polyadenylation specificity factor complex subunit pta1 pta1 317 1 51,695
Q09173 Protein phosphatase 2C homolog 3 ptc3 407 1 60,541
Gen
Typewritten Text
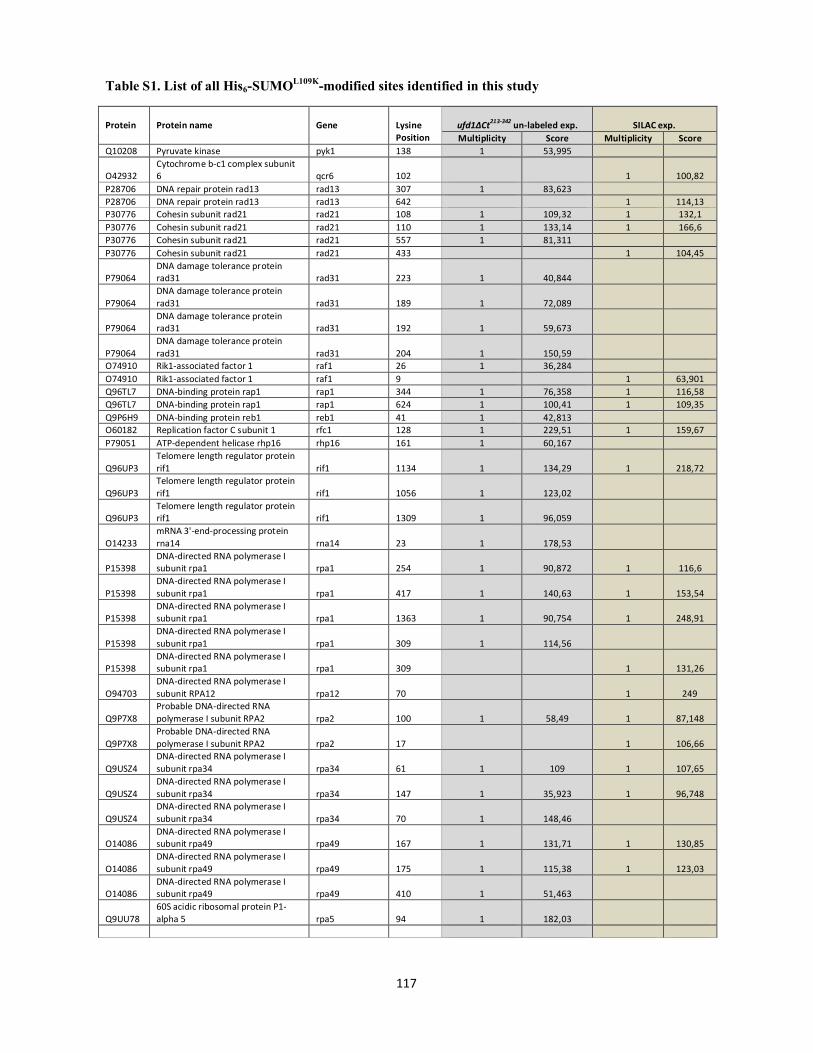
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
117
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
Q10208 Pyruvate kinase pyk1 138 1 53,995
O42932 Cytochrome b-c1 complex subunit 6 qcr6 102 1 100,82
P28706 DNA repair protein rad13 rad13 307 1 83,623 P28706 DNA repair protein rad13 rad13 642 1 114,13 P30776 Cohesin subunit rad21 rad21 108 1 109,32 1 132,1 P30776 Cohesin subunit rad21 rad21 110 1 133,14 1 166,6 P30776 Cohesin subunit rad21 rad21 557 1 81,311 P30776 Cohesin subunit rad21 rad21 433 1 104,45
P79064 DNA damage tolerance protein rad31 rad31 223 1 40,844
P79064 DNA damage tolerance protein rad31 rad31 189 1 72,089
P79064 DNA damage tolerance protein rad31 rad31 192 1 59,673
P79064 DNA damage tolerance protein rad31 rad31 204 1 150,59
O74910 Rik1-associated factor 1 raf1 26 1 36,284 O74910 Rik1-associated factor 1 raf1 9 1 63,901 Q96TL7 DNA-binding protein rap1 rap1 344 1 76,358 1 116,58 Q96TL7 DNA-binding protein rap1 rap1 624 1 100,41 1 109,35 Q9P6H9 DNA-binding protein reb1 reb1 41 1 42,813 O60182 Replication factor C subunit 1 rfc1 128 1 229,51 1 159,67 P79051 ATP-dependent helicase rhp16 rhp16 161 1 60,167
Q96UP3 Telomere length regulator protein rif1 rif1 1134 1 134,29 1 218,72
Q96UP3 Telomere length regulator protein rif1 rif1 1056 1 123,02
Q96UP3 Telomere length regulator protein rif1 rif1 1309 1 96,059
O14233 mRNA 3'-end-processing protein rna14 rna14 23 1 178,53
P15398 DNA-directed RNA polymerase I subunit rpa1 rpa1 254 1 90,872 1 116,6
P15398 DNA-directed RNA polymerase I subunit rpa1 rpa1 417 1 140,63 1 153,54
P15398 DNA-directed RNA polymerase I subunit rpa1 rpa1 1363 1 90,754 1 248,91
P15398 DNA-directed RNA polymerase I subunit rpa1 rpa1 309 1 114,56
P15398 DNA-directed RNA polymerase I subunit rpa1 rpa1 309 1 131,26
O94703 DNA-directed RNA polymerase I subunit RPA12 rpa12 70 1 249
Q9P7X8 Probable DNA-directed RNA polymerase I subunit RPA2 rpa2 100 1 58,49 1 87,148
Q9P7X8 Probable DNA-directed RNA polymerase I subunit RPA2 rpa2 17 1 106,66
Q9USZ4 DNA-directed RNA polymerase I subunit rpa34 rpa34 61 1 109 1 107,65
Q9USZ4 DNA-directed RNA polymerase I subunit rpa34 rpa34 147 1 35,923 1 96,748
Q9USZ4 DNA-directed RNA polymerase I subunit rpa34 rpa34 70 1 148,46
O14086 DNA-directed RNA polymerase I subunit rpa49 rpa49 167 1 131,71 1 130,85
O14086 DNA-directed RNA polymerase I subunit rpa49 rpa49 175 1 115,38 1 123,03
O14086 DNA-directed RNA polymerase I subunit rpa49 rpa49 410 1 51,463
Q9UU78 60S acidic ribosomal protein P1-alpha 5 rpa5 94 1 182,03
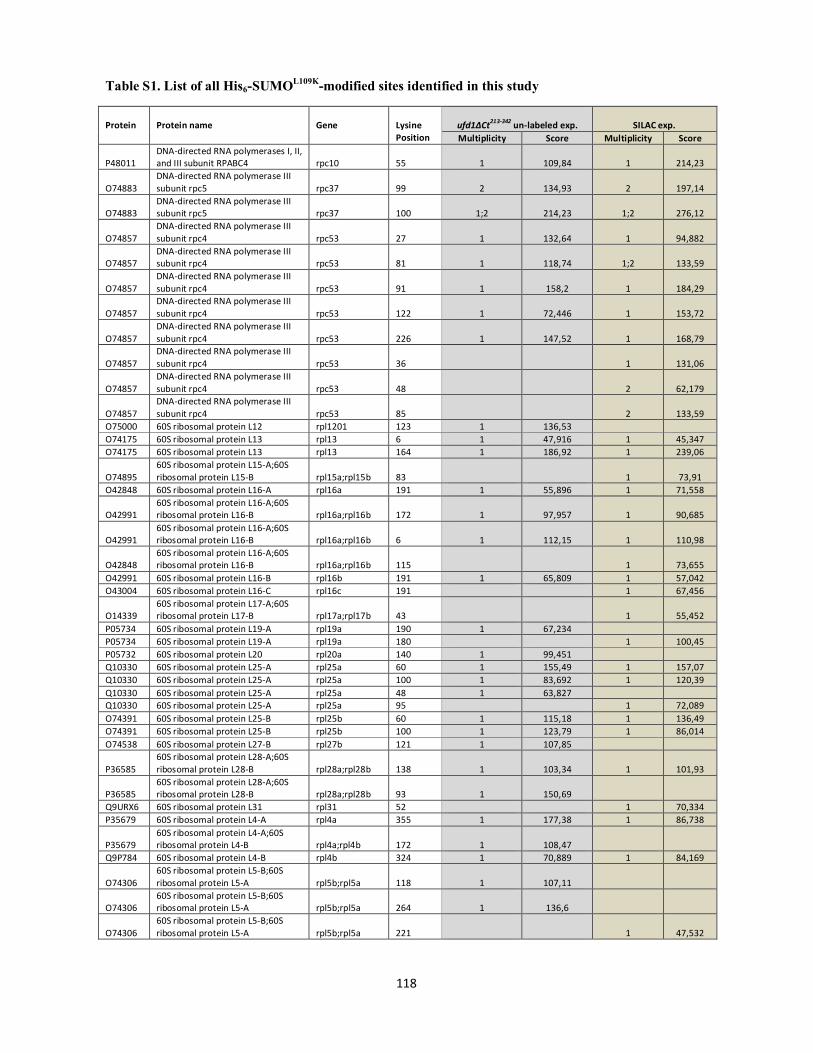
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
118
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
P48011 DNA-directed RNA polymerases I, II, and III subunit RPABC4 rpc10 55 1 109,84 1 214,23
O74883 DNA-directed RNA polymerase III subunit rpc5 rpc37 99 2 134,93 2 197,14
O74883 DNA-directed RNA polymerase III subunit rpc5 rpc37 100 1;2 214,23 1;2 276,12
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 27 1 132,64 1 94,882
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 81 1 118,74 1;2 133,59
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 91 1 158,2 1 184,29
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 122 1 72,446 1 153,72
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 226 1 147,52 1 168,79
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 36 1 131,06
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 48 2 62,179
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 85 2 133,59
O75000 60S ribosomal protein L12 rpl1201 123 1 136,53 O74175 60S ribosomal protein L13 rpl13 6 1 47,916 1 45,347 O74175 60S ribosomal protein L13 rpl13 164 1 186,92 1 239,06
O74895 60S ribosomal protein L15-A;60S ribosomal protein L15-B rpl15a;rpl15b 83 1 73,91
O42848 60S ribosomal protein L16-A rpl16a 191 1 55,896 1 71,558
O42991 60S ribosomal protein L16-A;60S ribosomal protein L16-B rpl16a;rpl16b 172 1 97,957 1 90,685
O42991 60S ribosomal protein L16-A;60S ribosomal protein L16-B rpl16a;rpl16b 6 1 112,15 1 110,98
O42848 60S ribosomal protein L16-A;60S ribosomal protein L16-B rpl16a;rpl16b 115 1 73,655
O42991 60S ribosomal protein L16-B rpl16b 191 1 65,809 1 57,042 O43004 60S ribosomal protein L16-C rpl16c 191 1 67,456
O14339 60S ribosomal protein L17-A;60S ribosomal protein L17-B rpl17a;rpl17b 43 1 55,452
P05734 60S ribosomal protein L19-A rpl19a 190 1 67,234 P05734 60S ribosomal protein L19-A rpl19a 180 1 100,45 P05732 60S ribosomal protein L20 rpl20a 140 1 99,451 Q10330 60S ribosomal protein L25-A rpl25a 60 1 155,49 1 157,07 Q10330 60S ribosomal protein L25-A rpl25a 100 1 83,692 1 120,39 Q10330 60S ribosomal protein L25-A rpl25a 48 1 63,827 Q10330 60S ribosomal protein L25-A rpl25a 95 1 72,089 O74391 60S ribosomal protein L25-B rpl25b 60 1 115,18 1 136,49 O74391 60S ribosomal protein L25-B rpl25b 100 1 123,79 1 86,014 O74538 60S ribosomal protein L27-B rpl27b 121 1 107,85
P36585 60S ribosomal protein L28-A;60S ribosomal protein L28-B rpl28a;rpl28b 138 1 103,34 1 101,93
P36585 60S ribosomal protein L28-A;60S ribosomal protein L28-B rpl28a;rpl28b 93 1 150,69
Q9URX6 60S ribosomal protein L31 rpl31 52 1 70,334 P35679 60S ribosomal protein L4-A rpl4a 355 1 177,38 1 86,738
P35679 60S ribosomal protein L4-A;60S ribosomal protein L4-B rpl4a;rpl4b 172 1 108,47
Q9P784 60S ribosomal protein L4-B rpl4b 324 1 70,889 1 84,169
O74306 60S ribosomal protein L5-B;60S ribosomal protein L5-A rpl5b;rpl5a 118 1 107,11
O74306 60S ribosomal protein L5-B;60S ribosomal protein L5-A rpl5b;rpl5a 264 1 136,6
O74306 60S ribosomal protein L5-B;60S ribosomal protein L5-A rpl5b;rpl5a 221 1 47,532
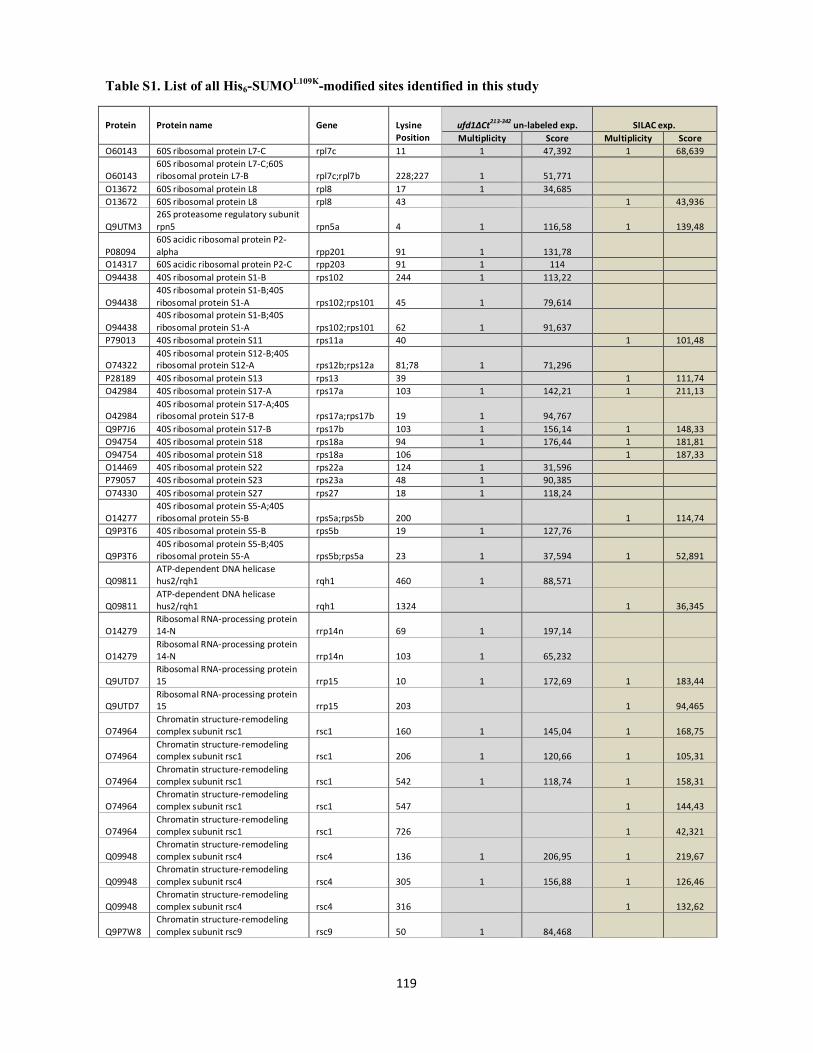
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
119
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
O60143 60S ribosomal protein L7-C rpl7c 11 1 47,392 1 68,639
O60143 60S ribosomal protein L7-C;60S ribosomal protein L7-B rpl7c;rpl7b 228;227 1 51,771
O13672 60S ribosomal protein L8 rpl8 17 1 34,685 O13672 60S ribosomal protein L8 rpl8 43 1 43,936
Q9UTM3 26S proteasome regulatory subunit rpn5 rpn5a 4 1 116,58 1 139,48
P08094 60S acidic ribosomal protein P2-alpha rpp201 91 1 131,78
O14317 60S acidic ribosomal protein P2-C rpp203 91 1 114 O94438 40S ribosomal protein S1-B rps102 244 1 113,22
O94438 40S ribosomal protein S1-B;40S ribosomal protein S1-A rps102;rps101 45 1 79,614
O94438 40S ribosomal protein S1-B;40S ribosomal protein S1-A rps102;rps101 62 1 91,637
P79013 40S ribosomal protein S11 rps11a 40 1 101,48
O74322 40S ribosomal protein S12-B;40S ribosomal protein S12-A rps12b;rps12a 81;78 1 71,296
P28189 40S ribosomal protein S13 rps13 39 1 111,74 O42984 40S ribosomal protein S17-A rps17a 103 1 142,21 1 211,13
O42984 40S ribosomal protein S17-A;40S ribosomal protein S17-B rps17a;rps17b 19 1 94,767
Q9P7J6 40S ribosomal protein S17-B rps17b 103 1 156,14 1 148,33 O94754 40S ribosomal protein S18 rps18a 94 1 176,44 1 181,81 O94754 40S ribosomal protein S18 rps18a 106 1 187,33 O14469 40S ribosomal protein S22 rps22a 124 1 31,596 P79057 40S ribosomal protein S23 rps23a 48 1 90,385 O74330 40S ribosomal protein S27 rps27 18 1 118,24
O14277 40S ribosomal protein S5-A;40S ribosomal protein S5-B rps5a;rps5b 200 1 114,74
Q9P3T6 40S ribosomal protein S5-B rps5b 19 1 127,76
Q9P3T6 40S ribosomal protein S5-B;40S ribosomal protein S5-A rps5b;rps5a 23 1 37,594 1 52,891
Q09811 ATP-dependent DNA helicase hus2/rqh1 rqh1 460 1 88,571
Q09811 ATP-dependent DNA helicase hus2/rqh1 rqh1 1324 1 36,345
O14279 Ribosomal RNA-processing protein 14-N rrp14n 69 1 197,14
O14279 Ribosomal RNA-processing protein 14-N rrp14n 103 1 65,232
Q9UTD7 Ribosomal RNA-processing protein 15 rrp15 10 1 172,69 1 183,44
Q9UTD7 Ribosomal RNA-processing protein 15 rrp15 203 1 94,465
O74964 Chromatin structure-remodeling complex subunit rsc1 rsc1 160 1 145,04 1 168,75
O74964 Chromatin structure-remodeling complex subunit rsc1 rsc1 206 1 120,66 1 105,31
O74964 Chromatin structure-remodeling complex subunit rsc1 rsc1 542 1 118,74 1 158,31
O74964 Chromatin structure-remodeling complex subunit rsc1 rsc1 547 1 144,43
O74964 Chromatin structure-remodeling complex subunit rsc1 rsc1 726 1 42,321
Q09948 Chromatin structure-remodeling complex subunit rsc4 rsc4 136 1 206,95 1 219,67
Q09948 Chromatin structure-remodeling complex subunit rsc4 rsc4 305 1 156,88 1 126,46
Q09948 Chromatin structure-remodeling complex subunit rsc4 rsc4 316 1 132,62
Q9P7W8 Chromatin structure-remodeling complex subunit rsc9 rsc9 50 1 84,468

Table S1. List of all His6-SUMOL109K-modified sites identified in this study
120
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
Q9P7W8 Chromatin structure-remodeling complex subunit rsc9 rsc9 63 1 51,211
O94506 mRNA export factor rsm1 rsm1 113 1 159,73 O94692 RuvB-like helicase 2 rvb2 35 1 55,453
Q09825 Spindle pole body-associated protein sad1 sad1 10 1 180,18 1 165,58
Q09825 Spindle pole body-associated protein sad1 sad1 49 1 254,29 1 88,427
P40847 Switch-activating protein 1 sap1 5 1 39,873 1 74,533 P40847 Switch-activating protein 1 sap1 172 1 93,716 1 164,93 P40847 Switch-activating protein 1 sap1 196 1 162,43 1 192,63 P40847 Switch-activating protein 1 sap1 26 1 177,47 P40847 Switch-activating protein 1 sap1 152 1 153,09
Q09717 Ran-specific GTPase-activating protein 1 sbp1 44 1 163,51 1 83,53
Q09717 Ran-specific GTPase-activating protein 1 sbp1 18 1 62,377
Q09717 Ran-specific GTPase-activating protein 1 sbp1 29 1 103,61
Q10342 Protein sda1 sda1 512 1 55,37 O13817 Protein transport protein sec73 sec73 984 1 116,98 Q10099 Nucleoporin seh1 seh1 278 1 72,846
Q9Y7R4
Histone-lysine N-methyltransferase, H3 lysine-4 specific set1 412 1 60,278 1 136,89
Q9UST7 Transcription factor tau subunit sfc3 sfc3 319 1 130,15 1 136,7
Q9UST7 Transcription factor tau subunit sfc3 sfc3 573 1 87,72
Q9UST7 Transcription factor tau subunit sfc3 sfc3 776 1 74,12
Q9UST7 Transcription factor tau subunit sfc3 sfc3 784 1 61,097
O74458 Transcription factor tau subunit sfc4 sfc4 52 1 88,561 1 140,27
O94397 SAGA-associated factor 73 sgf73 251 1 139,08 O13734 Shugoshin-2 sgo2 3 1 156,6 1 120,24 O13734 Shugoshin-2 sgo2 439 1 129,36 1 145,52 O13734 Shugoshin-2 sgo2 170 1 93,226 O13734 Shugoshin-2 sgo2 186 1 108,17 O13734 Shugoshin-2 sgo2 189 1 169,39 O13734 Shugoshin-2 sgo2 205 1 144,92 O13734 Shugoshin-2 sgo2 223 1 237,19 O13734 Shugoshin-2 sgo2 232 1 44,587 O13734 Shugoshin-2 sgo2 242 1 158,89 O13734 Shugoshin-2 sgo2 433 1 143,38 O13734 Shugoshin-2 sgo2 525 1 69,229 O13734 Shugoshin-2 sgo2 603 1 73,91 Q10284 Heat shock protein sks2 sks2 596 1 70,412 P87058 La protein homolog sla1 16 1 60,401 1 72,175 O13791 Protein slt1 slt1 383 1 194,24 1 217,82
O13710 Structural maintenance of chromosomes protein 5 smc5 352 1 58,889
O13710 Structural maintenance of chromosomes protein 5 smc5 849 1 119,75
P53692 Structural maintenance of chromosomes protein 6 smc6 41 1 31,138 1 63,302
P53692 Structural maintenance of chromosomes protein 6 smc6 351 1 60,157 1 114,74
P53692 Structural maintenance of chromosomes protein 6 smc6 393 1 69,451
P53692 Structural maintenance of chromosomes protein 6 smc6 384 1 119,45
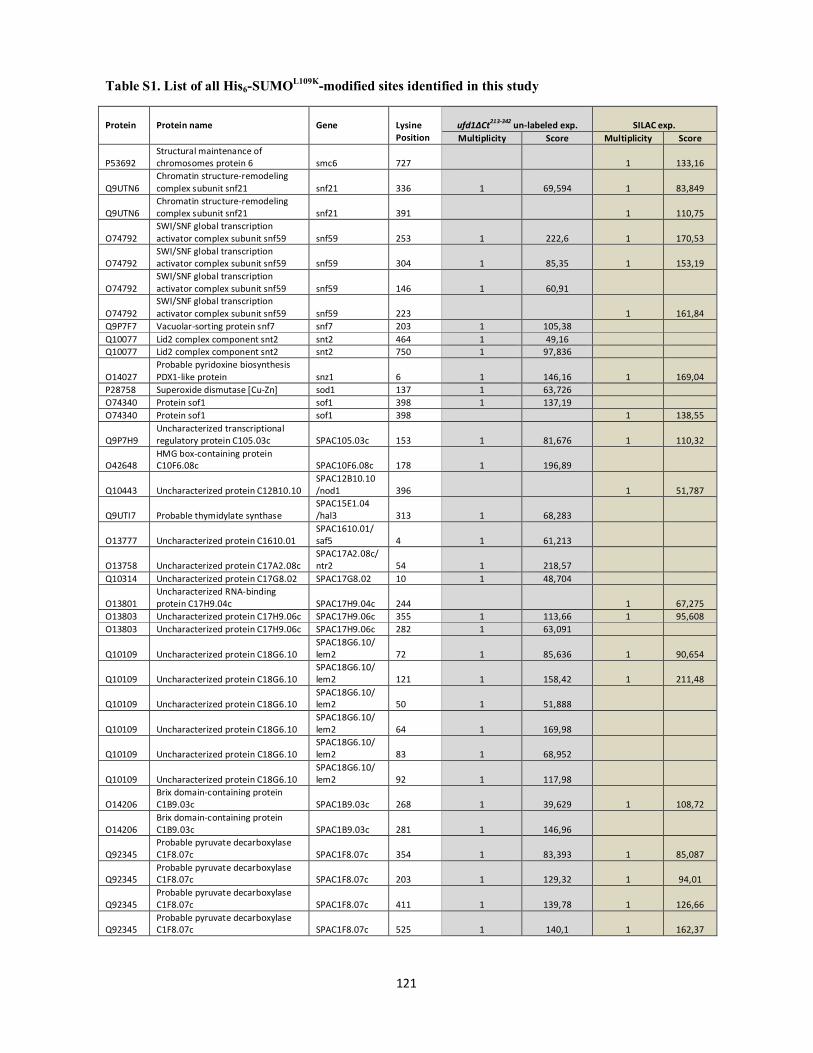
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
121
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
P53692 Structural maintenance of chromosomes protein 6 smc6 727 1 133,16
Q9UTN6 Chromatin structure-remodeling complex subunit snf21 snf21 336 1 69,594 1 83,849
Q9UTN6 Chromatin structure-remodeling complex subunit snf21 snf21 391 1 110,75
O74792 SWI/SNF global transcription activator complex subunit snf59 snf59 253 1 222,6 1 170,53
O74792 SWI/SNF global transcription activator complex subunit snf59 snf59 304 1 85,35 1 153,19
O74792 SWI/SNF global transcription activator complex subunit snf59 snf59 146 1 60,91
O74792 SWI/SNF global transcription activator complex subunit snf59 snf59 223 1 161,84
Q9P7F7 Vacuolar-sorting protein snf7 snf7 203 1 105,38 Q10077 Lid2 complex component snt2 snt2 464 1 49,16 Q10077 Lid2 complex component snt2 snt2 750 1 97,836
O14027 Probable pyridoxine biosynthesis PDX1-like protein snz1 6 1 146,16 1 169,04
P28758 Superoxide dismutase [Cu-Zn] sod1 137 1 63,726 O74340 Protein sof1 sof1 398 1 137,19 O74340 Protein sof1 sof1 398 1 138,55
Q9P7H9 Uncharacterized transcriptional regulatory protein C105.03c SPAC105.03c 153 1 81,676 1 110,32
O42648 HMG box-containing protein C10F6.08c SPAC10F6.08c 178 1 196,89
Q10443 Uncharacterized protein C12B10.10 SPAC12B10.10 /nod1 396 1 51,787
Q9UTI7 Probable thymidylate synthase SPAC15E1.04 /hal3 313 1 68,283
O13777 Uncharacterized protein C1610.01 SPAC1610.01/ saf5 4 1 61,213
O13758 Uncharacterized protein C17A2.08c SPAC17A2.08c/ntr2 54 1 218,57
Q10314 Uncharacterized protein C17G8.02 SPAC17G8.02 10 1 48,704
O13801 Uncharacterized RNA-binding protein C17H9.04c SPAC17H9.04c 244 1 67,275
O13803 Uncharacterized protein C17H9.06c SPAC17H9.06c 355 1 113,66 1 95,608 O13803 Uncharacterized protein C17H9.06c SPAC17H9.06c 282 1 63,091
Q10109 Uncharacterized protein C18G6.10 SPAC18G6.10/ lem2 72 1 85,636 1 90,654
Q10109 Uncharacterized protein C18G6.10 SPAC18G6.10/ lem2 121 1 158,42 1 211,48
Q10109 Uncharacterized protein C18G6.10 SPAC18G6.10/ lem2 50 1 51,888
Q10109 Uncharacterized protein C18G6.10 SPAC18G6.10/ lem2 64 1 169,98
Q10109 Uncharacterized protein C18G6.10 SPAC18G6.10/ lem2 83 1 68,952
Q10109 Uncharacterized protein C18G6.10 SPAC18G6.10/ lem2 92 1 117,98
O14206 Brix domain-containing protein C1B9.03c SPAC1B9.03c 268 1 39,629 1 108,72
O14206 Brix domain-containing protein C1B9.03c SPAC1B9.03c 281 1 146,96
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 354 1 83,393 1 85,087
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 203 1 129,32 1 94,01
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 411 1 139,78 1 126,66
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 525 1 140,1 1 162,37

Table S1. List of all His6-SUMOL109K-modified sites identified in this study
122
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 145 1 33,685
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 214 1 177,1
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 217 1 54,65
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 368 1 53,252
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 515 1 169,43
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 363 1 79,652
P87115 UPF0202 protein C20G8.09c SPAC20G8.09c/ nat10 925 1 149,31
Q10369 Uncharacterized protein C22E12.19 SPAC22E12.19/ snt1 474 1 141,36
Q09801 Pre-mRNA polyadenylation factor fip1
SPAC22G7.10/ iss1 89 1 135,01 1 127,5
Q10304 Uncharacterized protein C22H10.11c SPAC22H10.11c 328 1 38,839 1 41,227
O13964 Uncharacterized protein C24C9.04 SPAC24C9.04 104 1 171,29 O13964 Uncharacterized protein C24C9.04 SPAC24C9.04 104 1 246,96
O13972
Probable serine hydroxymethyltransferase, cytosolic SPAC24C9.12c 13 1 83
O13972
Probable serine hydroxymethyltransferase, cytosolic SPAC24C9.12c 265 1 100,22
Q9UTA8 Uncharacterized methyltransferase-like C25B8.10 SPAC25B8.10 8 1 54,471
Q9UTA6 Uncharacterized hydrolase C25B8.12c SPAC25B8.12c 196 1 114,84
Q9UTA6 Uncharacterized hydrolase C25B8.12c SPAC25B8.12c 169 1 124,42
Q10425 Eukaryotic translation initiation factor 3 subunit B SPAC25G10.08 16 1 147,56 1 122,57
Q10494 Uncharacterized oxidoreductase C26F1.07 SPAC26F1.07 240 1 84,188
O13987 Ankyrin and IPT/TIG repeat-containing protein C26H5.05
SPAC26H5.05/ mga2 615 1 48,112 1 100,38
Q9P7C0 Uncharacterized WD repeat-containing protein C2E1P5.05
SPAC2E1P5.05/ rrp9 83 1 60,436 1 138,49
Q09813 Putative transcription initiation factor TFIID 111 kDa subunit
SPAC2G11.14/ taf111 122 1 126,19
Q9P6K3 UPF0662 protein C30C2.08 SPAC30C2.08 215 1 47,743 O14105 Ribulose-phosphate 3-epimerase SPAC31G5.05c 5 1 79,511
O14114 Uncharacterized AAA domain-containing protein C31G5.19
SPAC31G5.19/ abo1 981 1 30,065
O14114 Uncharacterized AAA domain-containing protein C31G5.19
SPAC31G5.19/ abo1 66 1 207,67
O14114 Uncharacterized AAA domain-containing protein C31G5.19
SPAC31G5.19/ abo1 957 1 133,47
Q9P3W6 Uncharacterized protein C458.02c SPAC458.02c 338 1 120,79 1 111,57
O14174 Uncharacterized protein C4D7.10c SPAC4D7.10c/ spt20 167 1 65,784 1 159,7
O14174 Uncharacterized protein C4D7.10c SPAC4D7.10c/ spt20 27 1 93,572
O14174 Uncharacterized protein C4D7.10c SPAC4D7.10c/ spt20 340 1 99,836
O14174 Uncharacterized protein C4D7.10c SPAC4D7.10c/ spt20 395 1 127,46
O36021 Uncharacterized protein C4F10.09c SPAC4F10.09c 539 1 92,977 1 81,317 O36021 Uncharacterized protein C4F10.09c SPAC4F10.09c 734 1 117,25 1 122,69

Table S1. List of all His6-SUMOL109K-modified sites identified in this study
123
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
P87137 Uncharacterized protein C57A7.06 SPAC57A7.06 475 1 186,85
P87142 Heat shock protein 70 homolog C57A7.12 SPAC57A7.12 487 1 65,879
Q09671 Putative NADPH dehydrogenase C5H10.10 SPAC5H10.10 12 1 35,204
Q9HGP4 Bromodomain-containing protein C631.02
SPAC631.02/ bdf2 183 1 149,04 1 247,04
Q9P6Q5 Probable deoxyuridine 5'-triphosphate nucleotidohydrolase
SPAC644.05c/ dut1 7 1 114,83 1 136,99
O14210 Uncharacterized protein C6B12.05c SPAC6B12.05c/ ies2 31 1 32,682
A6X972 Putative uncharacterized protein C6B12.18
SPAC6B12.18/ gon7 69 1 42,21
O14269 Uncharacterized protein C7D4.14c SPAC7D4.14c/ iss10 313 1 84,515
Q9P6N2 Uncharacterized phosphatase C823.14
SPAC823.14/ ptf1 224 1 46,462
Q9UT36 Probable hydroxyacylglutathione hydrolase C824.07 SPAC824.07 46 1 57,989
O14273 Meiotically up-regulated protein C8C9.04 SPAC8C9.04 240 1 31,818
O14293 Putative aldehyde dehydrogenase-like protein C9E9.09c
SPAC9E9.09c/ atd1 4 1 104,44 1 99,569
O14297 Uncharacterized protein C9E9.15 SPAC9E9.15 124 1 86,467 1 65,5
Q9HDY7 Uncharacterized protein PB1A10.02 SPAPB1A10.02/ scm3 48 1 118,87
Q9HDY7 Uncharacterized protein PB1A10.02 SPAPB1A10.02/ scm3 58 1 65,092
Q9USZ6 mRNA turnover protein 4 homolog SPBC11G11.03/ mrt4 91 1 84,829
Q9P6R2 Uncharacterized protein C13E7.08c SPBC13E7.08c/ leo1 369 1 88,264
Q9P6R2 Uncharacterized protein C13E7.08c SPBC13E7.08c/ leo1 386 1 70,438
Q9Y808 Uncharacterized protein C146.01 SPBC146.01/ med15 813 1 47,568
O74324 Uncharacterized protein C1685.04 SPBC1685.04 2 1 74,475
O42909 Zinc-type alcohol dehydrogenase-like protein C16A3.02c SPBC16A3.02c 6 1 129,54 1 204,35
O42914 Uncharacterized protein C16A3.08c SPBC16A3.08c/ oga1 6 1 97,253
O42914 Uncharacterized protein C16A3.08c SPBC16A3.08c/ oga1 217 1 59,185
O14325 Uncharacterized AAA domain-containing protein C16E9.10c SPBC16E9.10c 409 1 54,004 1 86,367
O14325 Uncharacterized AAA domain-containing protein C16E9.10c SPBC16E9.10c 132 2 126,26
O14325 Uncharacterized AAA domain-containing protein C16E9.10c SPBC16E9.10c 133 2 126,26
Q9P785 LisH domain-containing protein C1711.05 SPBC1711.05 32 1 43,791 1 109,35
Q9P785 LisH domain-containing protein C1711.05 SPBC1711.05 55 1 95,292
O94561 Peroxiredoxin C1773.02c SPBC1773.02c/ bcp1 21 1 35,834
Q9USY2 Uncharacterized RNA-binding protein C1861.04c
SPBC1861.04c/ prp24 25 1 60,301
Q9P7K6 SDO1-like protein C21C3.19 SPBC21C3.19 86 1 86,291
O60177 Uncharacterized ATP-dependent helicase C23E6.02
SPBC23E6.02/ rrp2 319 1 123,32 1 102,07
O60177 Uncharacterized ATP-dependent helicase C23E6.02
SPBC23E6.02/ rrp2 104 1 74,325
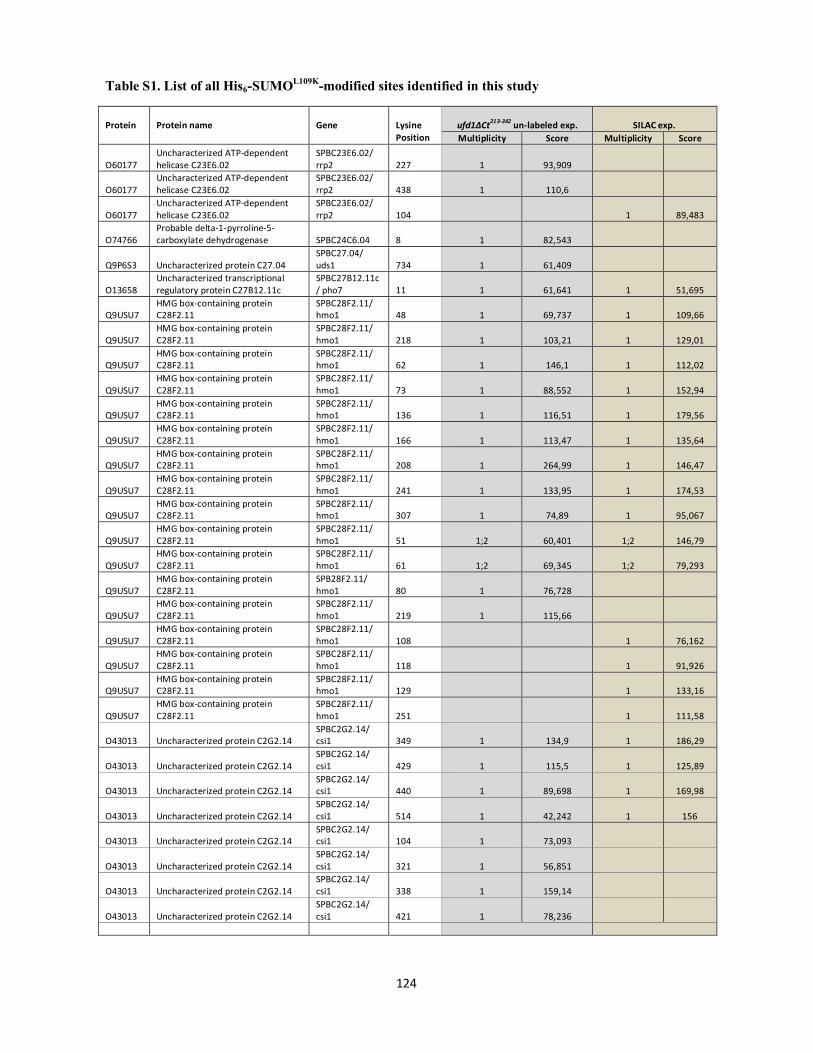
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
124
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
O60177 Uncharacterized ATP-dependent helicase C23E6.02
SPBC23E6.02/ rrp2 227 1 93,909
O60177 Uncharacterized ATP-dependent helicase C23E6.02
SPBC23E6.02/ rrp2 438 1 110,6
O60177 Uncharacterized ATP-dependent helicase C23E6.02
SPBC23E6.02/ rrp2 104 1 89,483
O74766 Probable delta-1-pyrroline-5-carboxylate dehydrogenase SPBC24C6.04 8 1 82,543
Q9P6S3 Uncharacterized protein C27.04 SPBC27.04/ uds1 734 1 61,409
O13658 Uncharacterized transcriptional regulatory protein C27B12.11c
SPBC27B12.11c/ pho7 11 1 61,641 1 51,695
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 48 1 69,737 1 109,66
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 218 1 103,21 1 129,01
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 62 1 146,1 1 112,02
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 73 1 88,552 1 152,94
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 136 1 116,51 1 179,56
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 166 1 113,47 1 135,64
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 208 1 264,99 1 146,47
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 241 1 133,95 1 174,53
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 307 1 74,89 1 95,067
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 51 1;2 60,401 1;2 146,79
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 61 1;2 69,345 1;2 79,293
Q9USU7 HMG box-containing protein C28F2.11
SPB28F2.11/ hmo1 80 1 76,728
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 219 1 115,66
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 108 1 76,162
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 118 1 91,926
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 129 1 133,16
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 251 1 111,58
O43013 Uncharacterized protein C2G2.14 SPBC2G2.14/ csi1 349 1 134,9 1 186,29
O43013 Uncharacterized protein C2G2.14 SPBC2G2.14/ csi1 429 1 115,5 1 125,89
O43013 Uncharacterized protein C2G2.14 SPBC2G2.14/ csi1 440 1 89,698 1 169,98
O43013 Uncharacterized protein C2G2.14 SPBC2G2.14/ csi1 514 1 42,242 1 156
O43013 Uncharacterized protein C2G2.14 SPBC2G2.14/ csi1 104 1 73,093
O43013 Uncharacterized protein C2G2.14 SPBC2G2.14/ csi1 321 1 56,851
O43013 Uncharacterized protein C2G2.14 SPBC2G2.14/ csi1 338 1 159,14
O43013 Uncharacterized protein C2G2.14 SPBC2G2.14/ csi1 421 1 78,236

Table S1. List of all His6-SUMOL109K-modified sites identified in this study
125
Protein Protein name Gene Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
O43013 Uncharacterized protein C2G2.14 SPBC2G2.14/ csi1 519 1 145,14
O43013 Uncharacterized protein C2G2.14 SPBC2G2.14/ csi1 321 1 143,28
Q9URM2 Probable transketolase SPBC2G5.05 586 1 71,933
O14362 KNR4/SMI1 homolog SPBC30D10.17c/ smi1 502 1 74,533 1 194,12
P87177 Uncharacterized WD repeat-containing protein C3D6.12
SPBC3D6.12/ dip2 724 1 50,608 1 34,094
P87177 Uncharacterized WD repeat-containing protein C3D6.12
SPBC3D6.12/ dip2 326 1 33,875
O94358 Uncharacterized protein C428.10 SPBC428.10 510 1 82,482
O74398 LIM domain-containing protein C4F6.12
SPBC4F6.12/ pxl1 115 1 51,147
P78812 6-phosphogluconate dehydrogenase, decarboxylating SPBC660.16 53 1 61,815
O13651 Isoleucine--tRNA ligase, cytoplasmic
SPBC8D2.06/ irs1 949 1 117,82
O13639 Adenosylhomocysteinase SPBC8D2.18c 7 1 180,93 1 100,69 O59757 Kinetochore protein spc7 spc7 778 1 79,511 O94523 Probable acyl-CoA desaturase SPCC1281.06c 452 1 42,913
O94720 GATA zinc finger domain-containing protein C1393.08 SPCC1393.08 237 1 190,81 1 185,71
O94720 GATA zinc finger domain-containing protein C1393.08 SPCC1393.08 344 1 87,216 1 108,34
O94720 GATA zinc finger domain-containing protein C1393.08 SPCC1393.08 483 1 211,89 1 266,78
O94720 GATA zinc finger domain-containing protein C1393.08 SPCC1393.08 545 2 182,54 2 227,96
O94720 GATA zinc finger domain-containing protein C1393.08 SPCC1393.08 544 1;2 222,74 1;2 227,96
O94577 Reduced meiotic recombination protein C1442.04c SPCC1442.04c 93 1 85,937
O94577 Reduced meiotic recombination protein C1442.04c SPCC1442.04c 347 1 59,57
Q9Y7N1 Uncharacterized protein C1450.03 SPCC1450.03 10 1 260,53 1 280,36
O94419 Putative GTPase-activating protein C1620.12c SPCC1620.12c 20 1 178,77
O14053 Uncharacterized WD repeat-containing protein C1672.07 SPCC1672.07 749 1 108,77
Q9UU99 CRAL-TRIO domain-containing protein C23B6.04c SPCC23B6.04c 182 1 38,511
Q9UU99 CRAL-TRIO domain-containing protein C23B6.04c SPCC23B6.04c 186 1 158,38
Q9P7J4 Uncharacterized protein SPCC24B10.11c
SPCC24B10.11c/ mft1 111 1 110,84 1 93,106
Q9P7I7 Uncharacterized protein C24B10.19c SPCC24B10.19c 286 1 46,319
Q9Y7R7 Putative ribosome biogenesis protein C306.07c SPCC306.07c 20 1 123,84
O74989 Uncharacterized protein C338.12 SPCC338.12/ pbi2 4 1 47,894
P87243 Uncharacterized protein C4G3.18 SPCC4G3.18/ rix1 270 1 42,599
O74504 UPF0590 protein C594.01 SPCC594.01 455 1 192,58 1 158,69 O94305 Putative monoglyceride lipase SPCC5E4.05c 281 1 121,82
Q9Y7T9 Uncharacterized J domain-containing protein C63.13 SPCC63.13 14 1 67,645
Q9Y7U0 Uncharacterized protein C63.14 SPCC63.14 525 1 120,55 Q9URT2 Uncharacterized protein P25A2.03 SPCP25A2.03 5 1 64,42
Q96WV0 Uncharacterized urease accessory protein ureG-like SPCPB16A4.05c 235 1 114,55
Q96WV0 Uncharacterized urease accessory protein ureG-like SPCPB16A4.05c 71 1 76,151
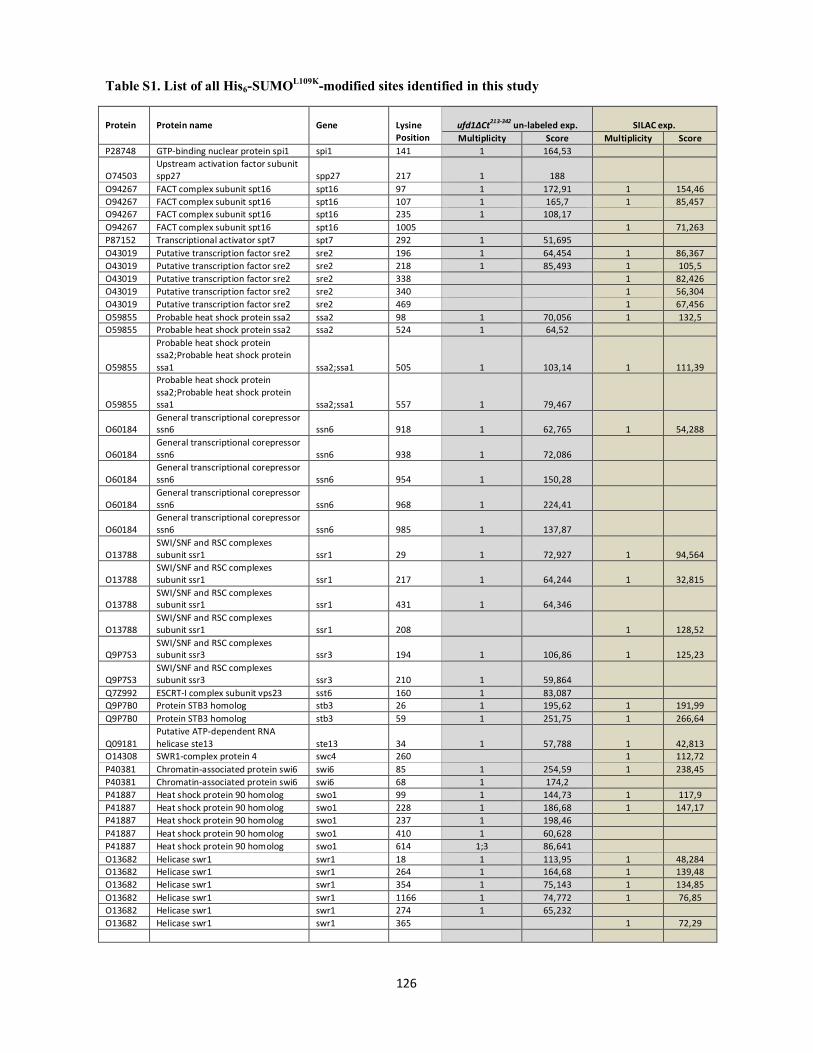
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
126
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
P28748 GTP-binding nuclear protein spi1 spi1 141 1 164,53
O74503 Upstream activation factor subunit spp27 spp27 217 1 188
O94267 FACT complex subunit spt16 spt16 97 1 172,91 1 154,46 O94267 FACT complex subunit spt16 spt16 107 1 165,7 1 85,457 O94267 FACT complex subunit spt16 spt16 235 1 108,17 O94267 FACT complex subunit spt16 spt16 1005 1 71,263 P87152 Transcriptional activator spt7 spt7 292 1 51,695 O43019 Putative transcription factor sre2 sre2 196 1 64,454 1 86,367 O43019 Putative transcription factor sre2 sre2 218 1 85,493 1 105,5 O43019 Putative transcription factor sre2 sre2 338 1 82,426 O43019 Putative transcription factor sre2 sre2 340 1 56,304 O43019 Putative transcription factor sre2 sre2 469 1 67,456 O59855 Probable heat shock protein ssa2 ssa2 98 1 70,056 1 132,5 O59855 Probable heat shock protein ssa2 ssa2 524 1 64,52
O59855
Probable heat shock protein ssa2;Probable heat shock protein ssa1 ssa2;ssa1 505 1 103,14 1 111,39
O59855
Probable heat shock protein ssa2;Probable heat shock protein ssa1 ssa2;ssa1 557 1 79,467
O60184 General transcriptional corepressor ssn6 ssn6 918 1 62,765 1 54,288
O60184 General transcriptional corepressor ssn6 ssn6 938 1 72,086
O60184 General transcriptional corepressor ssn6 ssn6 954 1 150,28
O60184 General transcriptional corepressor ssn6 ssn6 968 1 224,41
O60184 General transcriptional corepressor ssn6 ssn6 985 1 137,87
O13788 SWI/SNF and RSC complexes subunit ssr1 ssr1 29 1 72,927 1 94,564
O13788 SWI/SNF and RSC complexes subunit ssr1 ssr1 217 1 64,244 1 32,815
O13788 SWI/SNF and RSC complexes subunit ssr1 ssr1 431 1 64,346
O13788 SWI/SNF and RSC complexes subunit ssr1 ssr1 208 1 128,52
Q9P7S3 SWI/SNF and RSC complexes subunit ssr3 ssr3 194 1 106,86 1 125,23
Q9P7S3 SWI/SNF and RSC complexes subunit ssr3 ssr3 210 1 59,864
Q7Z992 ESCRT-I complex subunit vps23 sst6 160 1 83,087 Q9P7B0 Protein STB3 homolog stb3 26 1 195,62 1 191,99 Q9P7B0 Protein STB3 homolog stb3 59 1 251,75 1 266,64
Q09181 Putative ATP-dependent RNA helicase ste13 ste13 34 1 57,788 1 42,813
O14308 SWR1-complex protein 4 swc4 260 1 112,72 P40381 Chromatin-associated protein swi6 swi6 85 1 254,59 1 238,45 P40381 Chromatin-associated protein swi6 swi6 68 1 174,2 P41887 Heat shock protein 90 homolog swo1 99 1 144,73 1 117,9 P41887 Heat shock protein 90 homolog swo1 228 1 186,68 1 147,17 P41887 Heat shock protein 90 homolog swo1 237 1 198,46 P41887 Heat shock protein 90 homolog swo1 410 1 60,628 P41887 Heat shock protein 90 homolog swo1 614 1;3 86,641 O13682 Helicase swr1 swr1 18 1 113,95 1 48,284 O13682 Helicase swr1 swr1 264 1 164,68 1 139,48 O13682 Helicase swr1 swr1 354 1 75,143 1 134,85 O13682 Helicase swr1 swr1 1166 1 74,772 1 76,85 O13682 Helicase swr1 swr1 274 1 65,232 O13682 Helicase swr1 swr1 365 1 72,29
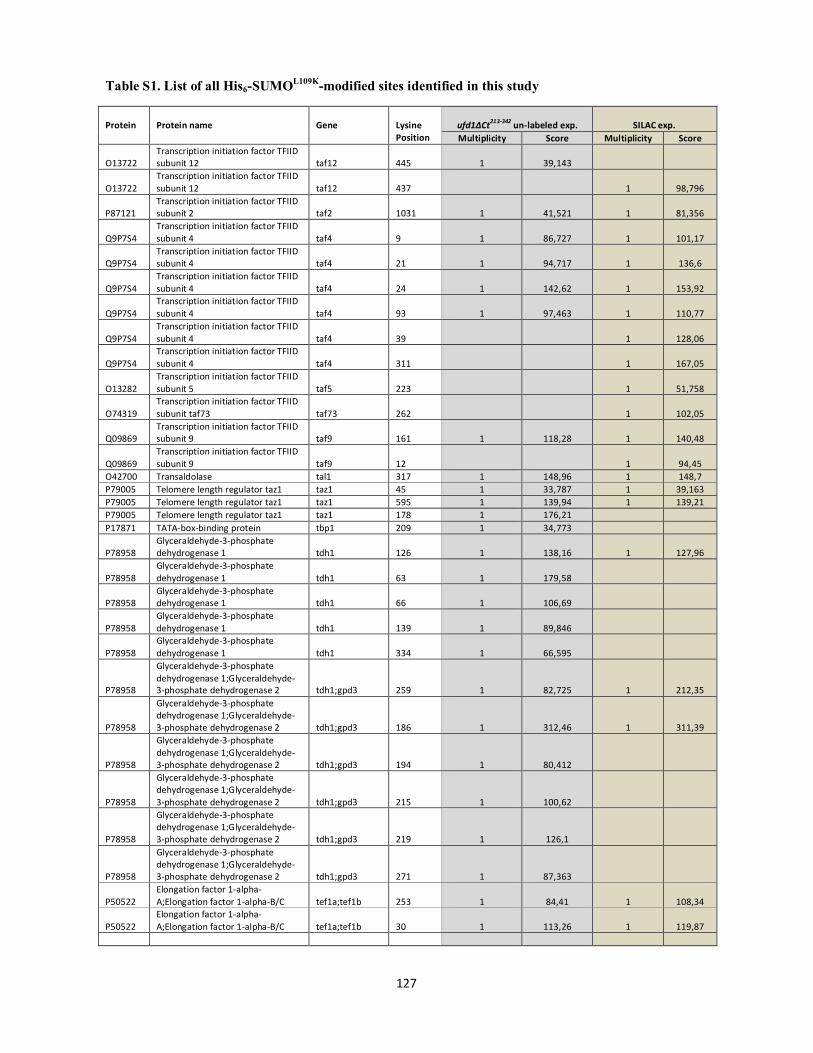
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
127
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
O13722 Transcription initiation factor TFIID subunit 12 taf12 445 1 39,143
O13722 Transcription initiation factor TFIID subunit 12 taf12 437 1 98,796
P87121 Transcription initiation factor TFIID subunit 2 taf2 1031 1 41,521 1 81,356
Q9P7S4 Transcription initiation factor TFIID subunit 4 taf4 9 1 86,727 1 101,17
Q9P7S4 Transcription initiation factor TFIID subunit 4 taf4 21 1 94,717 1 136,6
Q9P7S4 Transcription initiation factor TFIID subunit 4 taf4 24 1 142,62 1 153,92
Q9P7S4 Transcription initiation factor TFIID subunit 4 taf4 93 1 97,463 1 110,77
Q9P7S4 Transcription initiation factor TFIID subunit 4 taf4 39 1 128,06
Q9P7S4 Transcription initiation factor TFIID subunit 4 taf4 311 1 167,05
O13282 Transcription initiation factor TFIID subunit 5 taf5 223 1 51,758
O74319 Transcription initiation factor TFIID subunit taf73 taf73 262 1 102,05
Q09869 Transcription initiation factor TFIID subunit 9 taf9 161 1 118,28 1 140,48
Q09869 Transcription initiation factor TFIID subunit 9 taf9 12 1 94,45
O42700 Transaldolase tal1 317 1 148,96 1 148,7 P79005 Telomere length regulator taz1 taz1 45 1 33,787 1 39,163 P79005 Telomere length regulator taz1 taz1 595 1 139,94 1 139,21 P79005 Telomere length regulator taz1 taz1 178 1 176,21 P17871 TATA-box-binding protein tbp1 209 1 34,773
P78958 Glyceraldehyde-3-phosphate dehydrogenase 1 tdh1 126 1 138,16 1 127,96
P78958 Glyceraldehyde-3-phosphate dehydrogenase 1 tdh1 63 1 179,58
P78958 Glyceraldehyde-3-phosphate dehydrogenase 1 tdh1 66 1 106,69
P78958 Glyceraldehyde-3-phosphate dehydrogenase 1 tdh1 139 1 89,846
P78958 Glyceraldehyde-3-phosphate dehydrogenase 1 tdh1 334 1 66,595
P78958
Glyceraldehyde-3-phosphate dehydrogenase 1;Glyceraldehyde-3-phosphate dehydrogenase 2 tdh1;gpd3 259 1 82,725 1 212,35
P78958
Glyceraldehyde-3-phosphate dehydrogenase 1;Glyceraldehyde-3-phosphate dehydrogenase 2 tdh1;gpd3 186 1 312,46 1 311,39
P78958
Glyceraldehyde-3-phosphate dehydrogenase 1;Glyceraldehyde-3-phosphate dehydrogenase 2 tdh1;gpd3 194 1 80,412
P78958
Glyceraldehyde-3-phosphate dehydrogenase 1;Glyceraldehyde-3-phosphate dehydrogenase 2 tdh1;gpd3 215 1 100,62
P78958
Glyceraldehyde-3-phosphate dehydrogenase 1;Glyceraldehyde-3-phosphate dehydrogenase 2 tdh1;gpd3 219 1 126,1
P78958
Glyceraldehyde-3-phosphate dehydrogenase 1;Glyceraldehyde-3-phosphate dehydrogenase 2 tdh1;gpd3 271 1 87,363
P50522 Elongation factor 1-alpha-A;Elongation factor 1-alpha-B/C tef1a;tef1b 253 1 84,41 1 108,34
P50522 Elongation factor 1-alpha-A;Elongation factor 1-alpha-B/C tef1a;tef1b 30 1 113,26 1 119,87
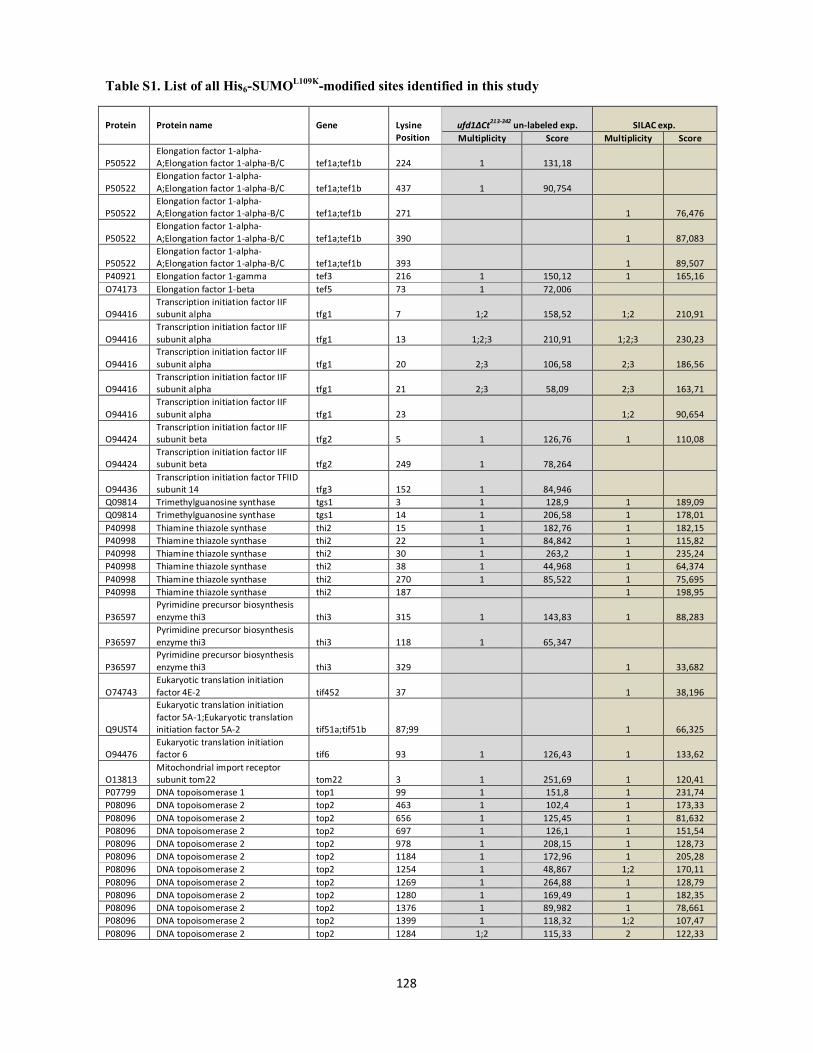
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
128
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
P50522 Elongation factor 1-alpha-A;Elongation factor 1-alpha-B/C tef1a;tef1b 224 1 131,18
P50522 Elongation factor 1-alpha-A;Elongation factor 1-alpha-B/C tef1a;tef1b 437 1 90,754
P50522 Elongation factor 1-alpha-A;Elongation factor 1-alpha-B/C tef1a;tef1b 271 1 76,476
P50522 Elongation factor 1-alpha-A;Elongation factor 1-alpha-B/C tef1a;tef1b 390 1 87,083
P50522 Elongation factor 1-alpha-A;Elongation factor 1-alpha-B/C tef1a;tef1b 393 1 89,507
P40921 Elongation factor 1-gamma tef3 216 1 150,12 1 165,16 O74173 Elongation factor 1-beta tef5 73 1 72,006
O94416 Transcription initiation factor IIF subunit alpha tfg1 7 1;2 158,52 1;2 210,91
O94416 Transcription initiation factor IIF subunit alpha tfg1 13 1;2;3 210,91 1;2;3 230,23
O94416 Transcription initiation factor IIF subunit alpha tfg1 20 2;3 106,58 2;3 186,56
O94416 Transcription initiation factor IIF subunit alpha tfg1 21 2;3 58,09 2;3 163,71
O94416 Transcription initiation factor IIF subunit alpha tfg1 23 1;2 90,654
O94424 Transcription initiation factor IIF subunit beta tfg2 5 1 126,76 1 110,08
O94424 Transcription initiation factor IIF subunit beta tfg2 249 1 78,264
O94436 Transcription initiation factor TFIID subunit 14 tfg3 152 1 84,946
Q09814 Trimethylguanosine synthase tgs1 3 1 128,9 1 189,09 Q09814 Trimethylguanosine synthase tgs1 14 1 206,58 1 178,01 P40998 Thiamine thiazole synthase thi2 15 1 182,76 1 182,15 P40998 Thiamine thiazole synthase thi2 22 1 84,842 1 115,82 P40998 Thiamine thiazole synthase thi2 30 1 263,2 1 235,24 P40998 Thiamine thiazole synthase thi2 38 1 44,968 1 64,374 P40998 Thiamine thiazole synthase thi2 270 1 85,522 1 75,695 P40998 Thiamine thiazole synthase thi2 187 1 198,95
P36597 Pyrimidine precursor biosynthesis enzyme thi3 thi3 315 1 143,83 1 88,283
P36597 Pyrimidine precursor biosynthesis enzyme thi3 thi3 118 1 65,347
P36597 Pyrimidine precursor biosynthesis enzyme thi3 thi3 329 1 33,682
O74743 Eukaryotic translation initiation factor 4E-2 tif452 37 1 38,196
Q9UST4
Eukaryotic translation initiation factor 5A-1;Eukaryotic translation initiation factor 5A-2 tif51a;tif51b 87;99 1 66,325
O94476 Eukaryotic translation initiation factor 6 tif6 93 1 126,43 1 133,62
O13813 Mitochondrial import receptor subunit tom22 tom22 3 1 251,69 1 120,41
P07799 DNA topoisomerase 1 top1 99 1 151,8 1 231,74 P08096 DNA topoisomerase 2 top2 463 1 102,4 1 173,33 P08096 DNA topoisomerase 2 top2 656 1 125,45 1 81,632 P08096 DNA topoisomerase 2 top2 697 1 126,1 1 151,54 P08096 DNA topoisomerase 2 top2 978 1 208,15 1 128,73 P08096 DNA topoisomerase 2 top2 1184 1 172,96 1 205,28 P08096 DNA topoisomerase 2 top2 1254 1 48,867 1;2 170,11 P08096 DNA topoisomerase 2 top2 1269 1 264,88 1 128,79 P08096 DNA topoisomerase 2 top2 1280 1 169,49 1 182,35 P08096 DNA topoisomerase 2 top2 1376 1 89,982 1 78,661 P08096 DNA topoisomerase 2 top2 1399 1 118,32 1;2 107,47 P08096 DNA topoisomerase 2 top2 1284 1;2 115,33 2 122,33
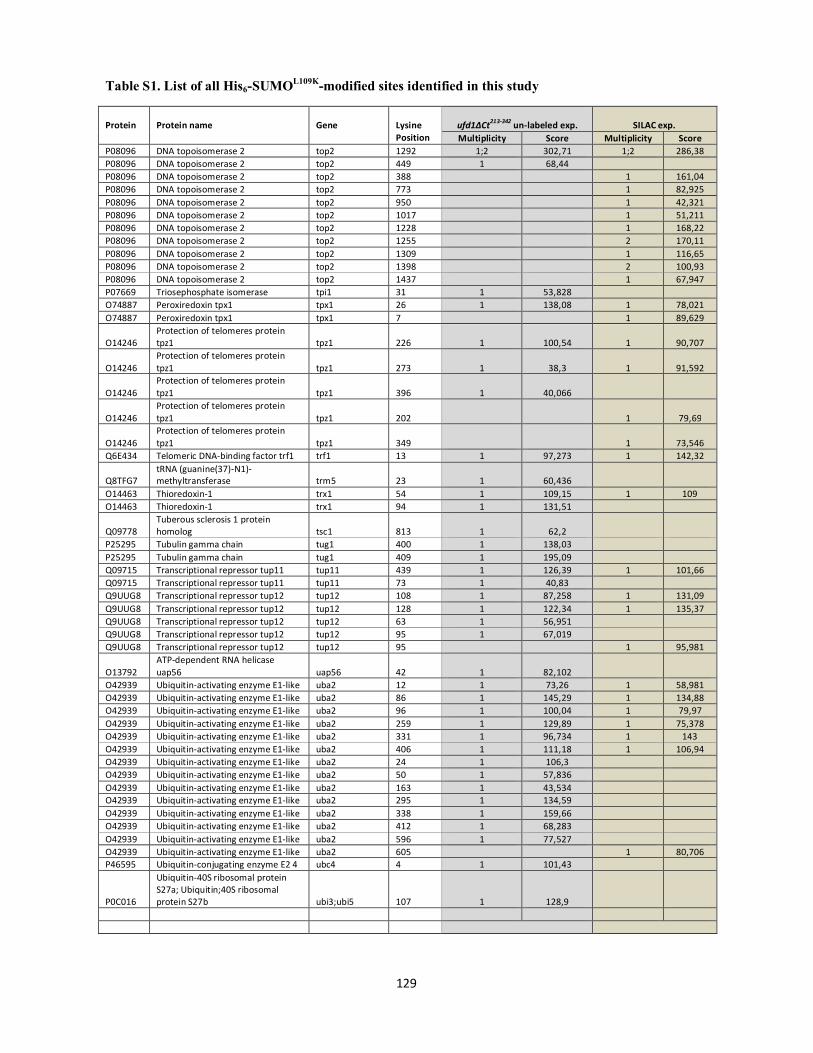
Table S1. List of all His6-SUMOL109K-modified sites identified in this study
129
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
P08096 DNA topoisomerase 2 top2 1292 1;2 302,71 1;2 286,38 P08096 DNA topoisomerase 2 top2 449 1 68,44 P08096 DNA topoisomerase 2 top2 388 1 161,04 P08096 DNA topoisomerase 2 top2 773 1 82,925 P08096 DNA topoisomerase 2 top2 950 1 42,321 P08096 DNA topoisomerase 2 top2 1017 1 51,211 P08096 DNA topoisomerase 2 top2 1228 1 168,22 P08096 DNA topoisomerase 2 top2 1255 2 170,11 P08096 DNA topoisomerase 2 top2 1309 1 116,65 P08096 DNA topoisomerase 2 top2 1398 2 100,93 P08096 DNA topoisomerase 2 top2 1437 1 67,947 P07669 Triosephosphate isomerase tpi1 31 1 53,828 O74887 Peroxiredoxin tpx1 tpx1 26 1 138,08 1 78,021 O74887 Peroxiredoxin tpx1 tpx1 7 1 89,629
O14246 Protection of telomeres protein tpz1 tpz1 226 1 100,54 1 90,707
O14246 Protection of telomeres protein tpz1 tpz1 273 1 38,3 1 91,592
O14246 Protection of telomeres protein tpz1 tpz1 396 1 40,066
O14246 Protection of telomeres protein tpz1 tpz1 202 1 79,69
O14246 Protection of telomeres protein tpz1 tpz1 349 1 73,546
Q6E434 Telomeric DNA-binding factor trf1 trf1 13 1 97,273 1 142,32
Q8TFG7 tRNA (guanine(37)-N1)-methyltransferase trm5 23 1 60,436
O14463 Thioredoxin-1 trx1 54 1 109,15 1 109 O14463 Thioredoxin-1 trx1 94 1 131,51
Q09778 Tuberous sclerosis 1 protein homolog tsc1 813 1 62,2
P25295 Tubulin gamma chain tug1 400 1 138,03 P25295 Tubulin gamma chain tug1 409 1 195,09 Q09715 Transcriptional repressor tup11 tup11 439 1 126,39 1 101,66 Q09715 Transcriptional repressor tup11 tup11 73 1 40,83 Q9UUG8 Transcriptional repressor tup12 tup12 108 1 87,258 1 131,09 Q9UUG8 Transcriptional repressor tup12 tup12 128 1 122,34 1 135,37 Q9UUG8 Transcriptional repressor tup12 tup12 63 1 56,951 Q9UUG8 Transcriptional repressor tup12 tup12 95 1 67,019 Q9UUG8 Transcriptional repressor tup12 tup12 95 1 95,981
O13792 ATP-dependent RNA helicase uap56 uap56 42 1 82,102
O42939 Ubiquitin-activating enzyme E1-like uba2 12 1 73,26 1 58,981 O42939 Ubiquitin-activating enzyme E1-like uba2 86 1 145,29 1 134,88 O42939 Ubiquitin-activating enzyme E1-like uba2 96 1 100,04 1 79,97 O42939 Ubiquitin-activating enzyme E1-like uba2 259 1 129,89 1 75,378 O42939 Ubiquitin-activating enzyme E1-like uba2 331 1 96,734 1 143 O42939 Ubiquitin-activating enzyme E1-like uba2 406 1 111,18 1 106,94 O42939 Ubiquitin-activating enzyme E1-like uba2 24 1 106,3 O42939 Ubiquitin-activating enzyme E1-like uba2 50 1 57,836 O42939 Ubiquitin-activating enzyme E1-like uba2 163 1 43,534 O42939 Ubiquitin-activating enzyme E1-like uba2 295 1 134,59 O42939 Ubiquitin-activating enzyme E1-like uba2 338 1 159,66 O42939 Ubiquitin-activating enzyme E1-like uba2 412 1 68,283 O42939 Ubiquitin-activating enzyme E1-like uba2 596 1 77,527 O42939 Ubiquitin-activating enzyme E1-like uba2 605 1 80,706 P46595 Ubiquitin-conjugating enzyme E2 4 ubc4 4 1 101,43
P0C016
Ubiquitin-40S ribosomal protein S27a; Ubiquitin;40S ribosomal protein S27b ubi3;ubi5 107 1 128,9

Table S1. List of all His6-SUMOL109K-modified sites identified in this study
130
Protein Protein name Gene Lysine Position
ufd1∆Ct213-342 un-labeled exp. SILAC exp. Multiplicity Score Multiplicity Score
P0C016
Ubiquitin-40S ribosomal protein S27a; Ubiquitin;40S ribosomal protein;S27b;Polyubiquitin;Ubiquitin;Ubiquitin-60S ribosomal protein L40
ubi3;ubi5;ubi4;ubi1;ubi2 11 1 200,39 1 244,46
P0C016
Ubiquitin-40S ribosomal protein S27a; Ubiquitin;40S ribosomal protein;S27b;Polyubiquitin;Ubiquitin;Ubiquitin-60S ribosomal protein L40
ubi3;ubi5;ubi4;ubi1;ubi2 48 1 117,73 1 180,88
P0C016
Ubiquitin-40S ribosomal protein S27a;Ubiquitin;40S ribosomal protein;S27b;Polyubiquitin;Ubiquitin;Ubiquitin-60S ribosomal protein L40
ubi3;ubi5;ubi4;ubi1;ubi2 63 1 141,37 1 139,86
P0C016
Ubiqitin-40S ribosomal protein S27a; Ubiquitin;40S ribosomal protein;S27b;Polyubiquitin;Ubiquitin;Ubiquitin-60S ribosomal protein L40
ubi3;ubi5;ubi4;ubi1;ubi2 27 1 158,64
P0C016
Ubiquitin-40S ribosomal protein S27a; Ubiquitin;40S ribosomal protein;S27b;Polyubiquitin;Ubiquitin;Ubiquitin-60S ribosomal protein L40
ubi3;ubi5;ubi4;ubi1;ubi2 6 1 142,07
O42957 Ubiquitin-like-specific protease 1 ulp1 309 1 230,06 1 210,91 O42957 Ubiquitin-like-specific protease 1 ulp1 311 1 99,021 1 155,11 O42957 Ubiquitin-like-specific protease 1 ulp1 328 1 87,568 1 122,97 O42957 Ubiquitin-like-specific protease 1 ulp1 75 1 76,713 O42957 Ubiquitin-like-specific protease 1 ulp1 264 1 208,68 O42957 Ubiquitin-like-specific protease 1 ulp1 299 1 116,79 O42957 Ubiquitin-like-specific protease 1 ulp1 299 1 162,88
O14082 Uncharacterized protein UNK4.17
Uncharacterized protein UNK4.17 6 1 106,62
O94259 Probable U3 small nucleolar RNA-associated protein 16 utp16 144 1 122,51 1 115,29
Q9HE11 U3 small nucleolar RNA-associated protein 5 utp5 637 1 71,806
Q9P4X3 Probable U3 small nucleolar RNA-associated protein 7 utp7 5 1 115,1
P87216 Protein vip1 vip1 254 1 48,659
Q9UTI5 Vacuolar protein sorting-associated protein 29 vps29 81 1 58,952 1 43,175
Q11004 40 kDa peptidyl-prolyl cis-trans isomerase wis2 213 1 125,86
Q11118 Protein wos2 wos2 90 1 258,12 O13794 Endoribonuclease ysh1 ysh1 733 1 98,507 2 82,707 O13794 Endoribonuclease ysh1 ysh1 723 1;2 224,42 1 196,58 O13794 Endoribonuclease ysh1 ysh1 729 1;2 158,77 2 82,707 O13794 Endoribonuclease ysh1 ysh1 752 2 39,424 O13794 Endoribonuclease ysh1 ysh1 753 2 39,424 Q9URY0 Ribosome biogenesis protein ytm1 ytm1 184 1 89,471 Q9URY0 Ribosome biogenesis protein ytm1 ytm1 438 1 51,643 Q10424 Transcription factor zip1 zip1 90 1 127,87 1 171,34 Q10424 Transcription factor zip1 zip1 150 1 126,84 Q10424 Transcription factor zip1 zip1 15 1 139,4 O13724 Zinc finger protein zpr1 zpr1 448 1 100,02 O94639 Zinc-regulated transporter 1 zrt1 224 1 172,57

Table S1. List of all His6-SUMOL109K-modified sites identified in this study
131
Protein
Protein name
Gene
Lysine Position
ufd1∆Ct213-342 un-labeled exp.
Location prob. Multiplicity Score Q9UUB5 Kinetochore protein mis13 mis13 2 or 4 0,5 1 47,545
Q9UU78 60S acidic ribosomal protein P1-alpha 5 rpa5 90 0,60278 1 64,488
Q10169 Deubiquitination-protection protein dph1 dph1 63 0,759861 1 36,424
O42700 Transaldolase tal1 266 0,88371 1 69,935
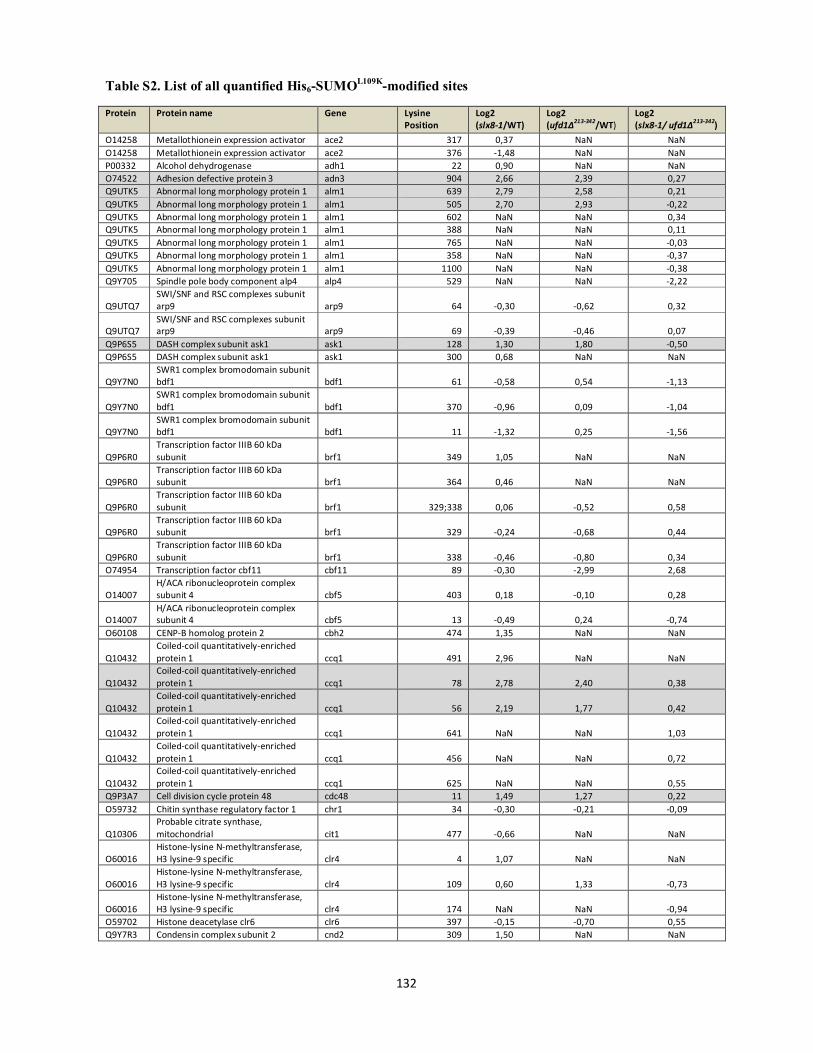
Table S2. List of all quantified His6-SUMOL109K-modified sites
132
Protein Protein name Gene Lysine Position
Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O14258 Metallothionein expression activator ace2 317 0,37 NaN NaN O14258 Metallothionein expression activator ace2 376 -1,48 NaN NaN P00332 Alcohol dehydrogenase adh1 22 0,90 NaN NaN O74522 Adhesion defective protein 3 adn3 904 2,66 2,39 0,27 Q9UTK5 Abnormal long morphology protein 1 alm1 639 2,79 2,58 0,21 Q9UTK5 Abnormal long morphology protein 1 alm1 505 2,70 2,93 -0,22 Q9UTK5 Abnormal long morphology protein 1 alm1 602 NaN NaN 0,34 Q9UTK5 Abnormal long morphology protein 1 alm1 388 NaN NaN 0,11 Q9UTK5 Abnormal long morphology protein 1 alm1 765 NaN NaN -0,03 Q9UTK5 Abnormal long morphology protein 1 alm1 358 NaN NaN -0,37 Q9UTK5 Abnormal long morphology protein 1 alm1 1100 NaN NaN -0,38 Q9Y705 Spindle pole body component alp4 alp4 529 NaN NaN -2,22
Q9UTQ7 SWI/SNF and RSC complexes subunit arp9 arp9 64 -0,30 -0,62 0,32
Q9UTQ7 SWI/SNF and RSC complexes subunit arp9 arp9 69 -0,39 -0,46 0,07
Q9P6S5 DASH complex subunit ask1 ask1 128 1,30 1,80 -0,50 Q9P6S5 DASH complex subunit ask1 ask1 300 0,68 NaN NaN
Q9Y7N0 SWR1 complex bromodomain subunit bdf1 bdf1 61 -0,58 0,54 -1,13
Q9Y7N0 SWR1 complex bromodomain subunit bdf1 bdf1 370 -0,96 0,09 -1,04
Q9Y7N0 SWR1 complex bromodomain subunit bdf1 bdf1 11 -1,32 0,25 -1,56
Q9P6R0 Transcription factor IIIB 60 kDa subunit brf1 349 1,05 NaN NaN
Q9P6R0 Transcription factor IIIB 60 kDa subunit brf1 364 0,46 NaN NaN
Q9P6R0 Transcription factor IIIB 60 kDa subunit brf1 329;338 0,06 -0,52 0,58
Q9P6R0 Transcription factor IIIB 60 kDa subunit brf1 329 -0,24 -0,68 0,44
Q9P6R0 Transcription factor IIIB 60 kDa subunit brf1 338 -0,46 -0,80 0,34
O74954 Transcription factor cbf11 cbf11 89 -0,30 -2,99 2,68
O14007 H/ACA ribonucleoprotein complex subunit 4 cbf5 403 0,18 -0,10 0,28
O14007 H/ACA ribonucleoprotein complex subunit 4 cbf5 13 -0,49 0,24 -0,74
O60108 CENP-B homolog protein 2 cbh2 474 1,35 NaN NaN
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 491 2,96 NaN NaN
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 78 2,78 2,40 0,38
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 56 2,19 1,77 0,42
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 641 NaN NaN 1,03
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 456 NaN NaN 0,72
Q10432 Coiled-coil quantitatively-enriched protein 1 ccq1 625 NaN NaN 0,55
Q9P3A7 Cell division cycle protein 48 cdc48 11 1,49 1,27 0,22 O59732 Chitin synthase regulatory factor 1 chr1 34 -0,30 -0,21 -0,09
Q10306 Probable citrate synthase, mitochondrial cit1 477 -0,66 NaN NaN
O60016 Histone-lysine N-methyltransferase, H3 lysine-9 specific clr4 4 1,07 NaN NaN
O60016 Histone-lysine N-methyltransferase, H3 lysine-9 specific clr4 109 0,60 1,33 -0,73
O60016 Histone-lysine N-methyltransferase, H3 lysine-9 specific clr4 174 NaN NaN -0,94
O59702 Histone deacetylase clr6 clr6 397 -0,15 -0,70 0,55 Q9Y7R3 Condensin complex subunit 2 cnd2 309 1,50 NaN NaN
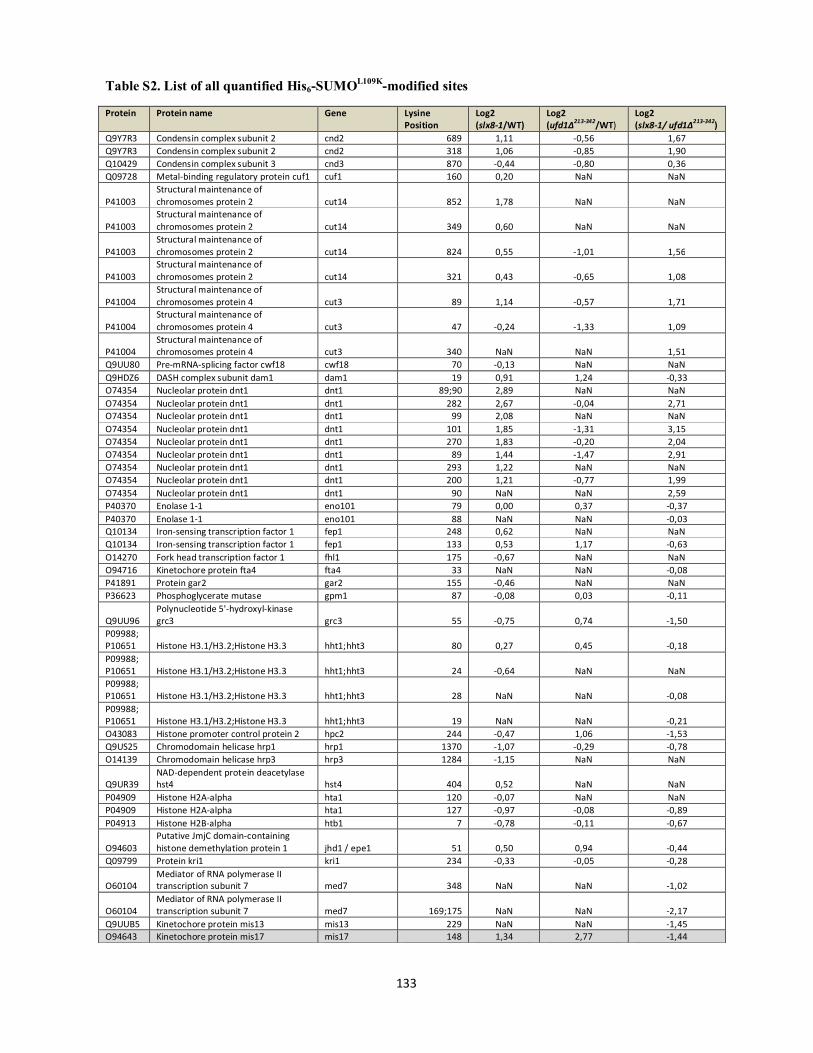
Table S2. List of all quantified His6-SUMOL109K-modified sites
133
Protein Protein name Gene Lysine Position
Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
Q9Y7R3 Condensin complex subunit 2 cnd2 689 1,11 -0,56 1,67 Q9Y7R3 Condensin complex subunit 2 cnd2 318 1,06 -0,85 1,90 Q10429 Condensin complex subunit 3 cnd3 870 -0,44 -0,80 0,36 Q09728 Metal-binding regulatory protein cuf1 cuf1 160 0,20 NaN NaN
P41003 Structural maintenance of chromosomes protein 2 cut14 852 1,78 NaN NaN
P41003 Structural maintenance of chromosomes protein 2 cut14 349 0,60 NaN NaN
P41003 Structural maintenance of chromosomes protein 2 cut14 824 0,55 -1,01 1,56
P41003 Structural maintenance of chromosomes protein 2 cut14 321 0,43 -0,65 1,08
P41004 Structural maintenance of chromosomes protein 4 cut3 89 1,14 -0,57 1,71
P41004 Structural maintenance of chromosomes protein 4 cut3 47 -0,24 -1,33 1,09
P41004 Structural maintenance of chromosomes protein 4 cut3 340 NaN NaN 1,51
Q9UU80 Pre-mRNA-splicing factor cwf18 cwf18 70 -0,13 NaN NaN Q9HDZ6 DASH complex subunit dam1 dam1 19 0,91 1,24 -0,33 O74354 Nucleolar protein dnt1 dnt1 89;90 2,89 NaN NaN O74354 Nucleolar protein dnt1 dnt1 282 2,67 -0,04 2,71 O74354 Nucleolar protein dnt1 dnt1 99 2,08 NaN NaN O74354 Nucleolar protein dnt1 dnt1 101 1,85 -1,31 3,15 O74354 Nucleolar protein dnt1 dnt1 270 1,83 -0,20 2,04 O74354 Nucleolar protein dnt1 dnt1 89 1,44 -1,47 2,91 O74354 Nucleolar protein dnt1 dnt1 293 1,22 NaN NaN O74354 Nucleolar protein dnt1 dnt1 200 1,21 -0,77 1,99 O74354 Nucleolar protein dnt1 dnt1 90 NaN NaN 2,59 P40370 Enolase 1-1 eno101 79 0,00 0,37 -0,37 P40370 Enolase 1-1 eno101 88 NaN NaN -0,03 Q10134 Iron-sensing transcription factor 1 fep1 248 0,62 NaN NaN Q10134 Iron-sensing transcription factor 1 fep1 133 0,53 1,17 -0,63 O14270 Fork head transcription factor 1 fhl1 175 -0,67 NaN NaN O94716 Kinetochore protein fta4 fta4 33 NaN NaN -0,08 P41891 Protein gar2 gar2 155 -0,46 NaN NaN P36623 Phosphoglycerate mutase gpm1 87 -0,08 0,03 -0,11
Q9UU96 Polynucleotide 5'-hydroxyl-kinase grc3 grc3 55 -0,75 0,74 -1,50
P09988; P10651 Histone H3.1/H3.2;Histone H3.3 hht1;hht3 80 0,27 0,45 -0,18 P09988; P10651 Histone H3.1/H3.2;Histone H3.3 hht1;hht3 24 -0,64 NaN NaN P09988; P10651 Histone H3.1/H3.2;Histone H3.3 hht1;hht3 28 NaN NaN -0,08 P09988; P10651 Histone H3.1/H3.2;Histone H3.3 hht1;hht3 19 NaN NaN -0,21 O43083 Histone promoter control protein 2 hpc2 244 -0,47 1,06 -1,53 Q9US25 Chromodomain helicase hrp1 hrp1 1370 -1,07 -0,29 -0,78 O14139 Chromodomain helicase hrp3 hrp3 1284 -1,15 NaN NaN
Q9UR39 NAD-dependent protein deacetylase hst4 hst4 404 0,52 NaN NaN
P04909 Histone H2A-alpha hta1 120 -0,07 NaN NaN P04909 Histone H2A-alpha hta1 127 -0,97 -0,08 -0,89 P04913 Histone H2B-alpha htb1 7 -0,78 -0,11 -0,67
O94603 Putative JmjC domain-containing histone demethylation protein 1 jhd1 / epe1 51 0,50 0,94 -0,44
Q09799 Protein kri1 kri1 234 -0,33 -0,05 -0,28
O60104 Mediator of RNA polymerase II transcription subunit 7 med7 348 NaN NaN -1,02
O60104 Mediator of RNA polymerase II transcription subunit 7 med7 169;175 NaN NaN -2,17
Q9UUB5 Kinetochore protein mis13 mis13 229 NaN NaN -1,45 O94643 Kinetochore protein mis17 mis17 148 1,34 2,77 -1,44
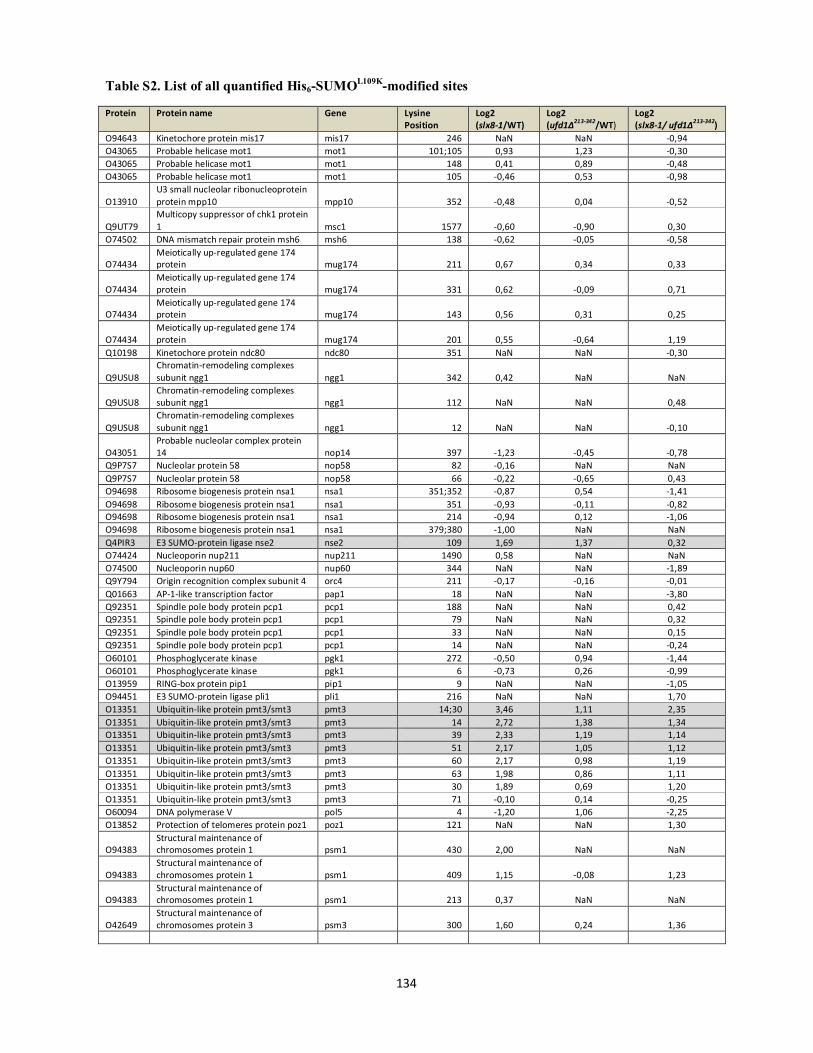
Table S2. List of all quantified His6-SUMOL109K-modified sites
134
Protein Protein name Gene Lysine Position
Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O94643 Kinetochore protein mis17 mis17 246 NaN NaN -0,94 O43065 Probable helicase mot1 mot1 101;105 0,93 1,23 -0,30 O43065 Probable helicase mot1 mot1 148 0,41 0,89 -0,48 O43065 Probable helicase mot1 mot1 105 -0,46 0,53 -0,98
O13910 U3 small nucleolar ribonucleoprotein protein mpp10 mpp10 352 -0,48 0,04 -0,52
Q9UT79 Multicopy suppressor of chk1 protein 1 msc1 1577 -0,60 -0,90 0,30
O74502 DNA mismatch repair protein msh6 msh6 138 -0,62 -0,05 -0,58
O74434 Meiotically up-regulated gene 174 protein mug174 211 0,67 0,34 0,33
O74434 Meiotically up-regulated gene 174 protein mug174 331 0,62 -0,09 0,71
O74434 Meiotically up-regulated gene 174 protein mug174 143 0,56 0,31 0,25
O74434 Meiotically up-regulated gene 174 protein mug174 201 0,55 -0,64 1,19
Q10198 Kinetochore protein ndc80 ndc80 351 NaN NaN -0,30
Q9USU8 Chromatin-remodeling complexes subunit ngg1 ngg1 342 0,42 NaN NaN
Q9USU8 Chromatin-remodeling complexes subunit ngg1 ngg1 112 NaN NaN 0,48
Q9USU8 Chromatin-remodeling complexes subunit ngg1 ngg1 12 NaN NaN -0,10
O43051 Probable nucleolar complex protein 14 nop14 397 -1,23 -0,45 -0,78
Q9P7S7 Nucleolar protein 58 nop58 82 -0,16 NaN NaN Q9P7S7 Nucleolar protein 58 nop58 66 -0,22 -0,65 0,43 O94698 Ribosome biogenesis protein nsa1 nsa1 351;352 -0,87 0,54 -1,41 O94698 Ribosome biogenesis protein nsa1 nsa1 351 -0,93 -0,11 -0,82 O94698 Ribosome biogenesis protein nsa1 nsa1 214 -0,94 0,12 -1,06 O94698 Ribosome biogenesis protein nsa1 nsa1 379;380 -1,00 NaN NaN Q4PIR3 E3 SUMO-protein ligase nse2 nse2 109 1,69 1,37 0,32 O74424 Nucleoporin nup211 nup211 1490 0,58 NaN NaN O74500 Nucleoporin nup60 nup60 344 NaN NaN -1,89 Q9Y794 Origin recognition complex subunit 4 orc4 211 -0,17 -0,16 -0,01 Q01663 AP-1-like transcription factor pap1 18 NaN NaN -3,80 Q92351 Spindle pole body protein pcp1 pcp1 188 NaN NaN 0,42 Q92351 Spindle pole body protein pcp1 pcp1 79 NaN NaN 0,32 Q92351 Spindle pole body protein pcp1 pcp1 33 NaN NaN 0,15 Q92351 Spindle pole body protein pcp1 pcp1 14 NaN NaN -0,24 O60101 Phosphoglycerate kinase pgk1 272 -0,50 0,94 -1,44 O60101 Phosphoglycerate kinase pgk1 6 -0,73 0,26 -0,99 O13959 RING-box protein pip1 pip1 9 NaN NaN -1,05 O94451 E3 SUMO-protein ligase pli1 pli1 216 NaN NaN 1,70 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 14;30 3,46 1,11 2,35 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 14 2,72 1,38 1,34 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 39 2,33 1,19 1,14 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 51 2,17 1,05 1,12 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 60 2,17 0,98 1,19 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 63 1,98 0,86 1,11 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 30 1,89 0,69 1,20 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 71 -0,10 0,14 -0,25 O60094 DNA polymerase V pol5 4 -1,20 1,06 -2,25 O13852 Protection of telomeres protein poz1 poz1 121 NaN NaN 1,30
O94383 Structural maintenance of chromosomes protein 1 psm1 430 2,00 NaN NaN
O94383 Structural maintenance of chromosomes protein 1 psm1 409 1,15 -0,08 1,23
O94383 Structural maintenance of chromosomes protein 1 psm1 213 0,37 NaN NaN
O42649 Structural maintenance of chromosomes protein 3 psm3 300 1,60 0,24 1,36

Table S2. List of all quantified His6-SUMOL109K-modified sites
135
Protein Protein name Gene Lysine Position
Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O42649 Structural maintenance of chromosomes protein 3 psm3 336 1,29 0,25 1,04
O42649 Structural maintenance of chromosomes protein 3 psm3 428 NaN NaN 1,62
O42649 Structural maintenance of chromosomes protein 3 psm3 833 NaN NaN 1,40
O42649 Structural maintenance of chromosomes protein 3 psm3 955 NaN NaN 1,02
Q09750 Paired amphipathic helix protein pst1 pst1 464 NaN NaN 0,37 P28706 DNA repair protein rad13 rad13 642 1,12 NaN NaN P30776 Cohesin subunit rad21 rad21 110 0,57 -0,30 0,87 Q96TL7 DNA-binding protein rap1 rap1 344 1,78 1,93 -0,15 Q96TL7 DNA-binding protein rap1 rap1 624 NaN NaN -0,21
Q96UP3 Telomere length regulator protein rif1 rif1 1134 NaN NaN -0,75
P15398 DNA-directed RNA polymerase I subunit rpa1 rpa1 254 2,82 NaN NaN
P15398 DNA-directed RNA polymerase I subunit rpa1 rpa1 309 1,06 NaN NaN
P15398 DNA-directed RNA polymerase I subunit rpa1 rpa1 417 0,80 -1,01 1,81
P15398 DNA-directed RNA polymerase I subunit rpa1 rpa1 1363 -0,11 -0,86 0,75
Q9P7X8 Probable DNA-directed RNA polymerase I subunit RPA2 rpa2 100 -0,40 -0,53 0,13
Q9USZ4 DNA-directed RNA polymerase I subunit rpa34 rpa34 147 0,28 -0,62 0,90
Q9USZ4 DNA-directed RNA polymerase I subunit rpa34 rpa34 61 NaN NaN 0,59
O14086 DNA-directed RNA polymerase I subunit rpa49 rpa49 175 1,28 NaN NaN
O14086 DNA-directed RNA polymerase I subunit rpa49 rpa49 167 NaN NaN 0,50
P48011 DNA-directed RNA polymerases I, II, and III subunit RPABC4 rpc10 55 0,25 -0,65 0,90
O74883 DNA-directed RNA polymerase III subunit rpc5 rpc37 99;100 1,36 0,22 1,14
O74883 DNA-directed RNA polymerase III subunit rpc5 rpc37 100 0,07 -1,08 1,15
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 91 1,10 0,32 0,78
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 122 0,85 -1,00 1,86
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 85 0,84 -1,04 1,88
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 81 0,84 -1,02 1,86
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 226 0,49 -1,00 1,49
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 27 0,08 NaN NaN
O74857 DNA-directed RNA polymerase III subunit rpc4 rpc53 81 -0,07 -0,86 0,79
O74175 60S ribosomal protein L13 rpl13 6 -0,29 NaN NaN O74175 60S ribosomal protein L13 rpl13 164 -0,72 -1,07 0,35 O42848 60S ribosomal protein L16-A rpl16a 191 -0,53 -0,70 0,17 O42848;O42991
60S ribosomal protein L16-A;60S ribosomal protein L16-B
rpl16a; rpl16b 172 -0,27 -0,47 0,20
O42848;O42991
60S ribosomal protein L16-A;60S ribosomal protein L16-B
rpl16a; rpl16b 6 -0,70 -0,45 -0,25
O42991 60S ribosomal protein L16-B rpl16b 191 -0,52 -0,68 0,15 O43004 60S ribosomal protein L16-C rpl16c 191 -0,02 NaN NaN Q10330 60S ribosomal protein L25-A rpl25a 60 -0,30 -0,31 0,00 Q10330 60S ribosomal protein L25-A rpl25a 100 -0,49 NaN NaN

Table S2. List of all quantified His6-SUMOL109K-modified sites
136
Protein Protein name Gene Lysine Position
Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O74391 60S ribosomal protein L25-B rpl25b 60 -0,25 -0,31 0,05 O74391 60S ribosomal protein L25-B rpl25b 100 -0,55 NaN NaN P36585;P57728
60S ribosomal protein L28-A;60S ribosomal protein L28-B
rpl28a; rpl28b 138 -0,33 NaN NaN
Q9URX6 60S ribosomal protein L31 rpl31 52 NaN NaN 0,13 P35679 60S ribosomal protein L4-A rpl4a 355 -0,47 NaN NaN P35679 60S ribosomal protein L4-A rpl4a 324 NaN NaN -0,36
Q9UTM3 26S proteasome regulatory subunit rpn5 rpn5a 4 2,17 0,38 1,79
P79013 40S ribosomal protein S11 rps11a 40 -0,28 NaN NaN P28189 40S ribosomal protein S13 rps13 39 -0,79 NaN NaN O42984 40S ribosomal protein S17-A rps17a 103 -0,33 -0,69 0,36 Q9P7J6 40S ribosomal protein S17-B rps17b 103 -0,35 -0,67 0,32 O94754 40S ribosomal protein S18 rps18a 106 -0,03 NaN NaN O94754 40S ribosomal protein S18 rps18a 94 -0,30 0,1 -0,44 O14277;Q9P3T6
40S ribosomal protein S5-A;40S ribosomal protein S5-B
rps5a; rps5b 200 NaN NaN 0,26
O74964 Chromatin structure-remodeling complex subunit rsc1 rsc1 542 -0,39 NaN NaN
O74964 Chromatin structure-remodeling complex subunit rsc1 rsc1 206 -0,47 NaN NaN
O74964 Chromatin structure-remodeling complex subunit rsc1 rsc1 160 -0,61 -0,76 0,15
Q09948 Chromatin structure-remodeling complex subunit rsc4 rsc4 316 -0,15 NaN NaN
Q09948 Chromatin structure-remodeling complex subunit rsc4 rsc4 305 -0,39 NaN NaN
Q09948 Chromatin structure-remodeling complex subunit rsc4 rsc4 136 -0,49 0,70 -1,19
Q9P7W8 Chromatin structure-remodeling complex subunit rsc9 rsc9 63 0,37 NaN NaN
Q9UST7 Transcription factor tau subunit sfc3 sfc3 319 -0,08 0,32 -0,40 O74458 Transcription factor tau subunit sfc4 sfc4 52 0,04 0,47 -0,43 O13734 Shugoshin-2 sgo2 439 -0,62 1,99 -2,61 O13791 Protein slt1 slt1 383 2,22 1,64 0,58
O13710 Structural maintenance of chromosomes protein 5 smc5 849 1,11 0,81 0,29
O13710 Structural maintenance of chromosomes protein 5 smc5 352 NaN NaN 1,13
P53692 Structural maintenance of chromosomes protein 6 smc6 384 NaN NaN 0,65
Q9UTN6 Chromatin structure-remodeling complex subunit snf21 snf21 391 0,21 NaN NaN
Q9UTN6 Chromatin structure-remodeling complex subunit snf21 snf21 336 -0,04 -0,51 0,47
O74792 SWI/SNF global transcription activator complex subunit snf59 snf59 304 1,93 0,80 1,13
O14027 Probable pyridoxine biosynthesis PDX1-like protein snz1 6 -0,36 1,10 -1,46
O74340 Protein sof1 sof1 398 NaN NaN -0,17
Q9P7H9 Uncharacterized transcriptional regulatory protein C105.03c SPAC105.03c 153 -0,78 -0,41 -0,37
O13801 Uncharacterized RNA-binding protein C17H9.04c SPAC17H9.04c 244 NaN NaN -1,02
O13803 Uncharacterized protein C17H9.06c SPAC17H9.06c 355 NaN NaN -0,41
Q10109 Uncharacterized protein C18G6.10 SPAC18G6.10/ lem2 121 -0,80 2,26 -3,06
O14206 Brix domain-containing protein C1B9.03c SPAC1B9.03c 268 -1,59 0,00 -1,59
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 411 NaN NaN 0,11
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 203 NaN NaN -0,21
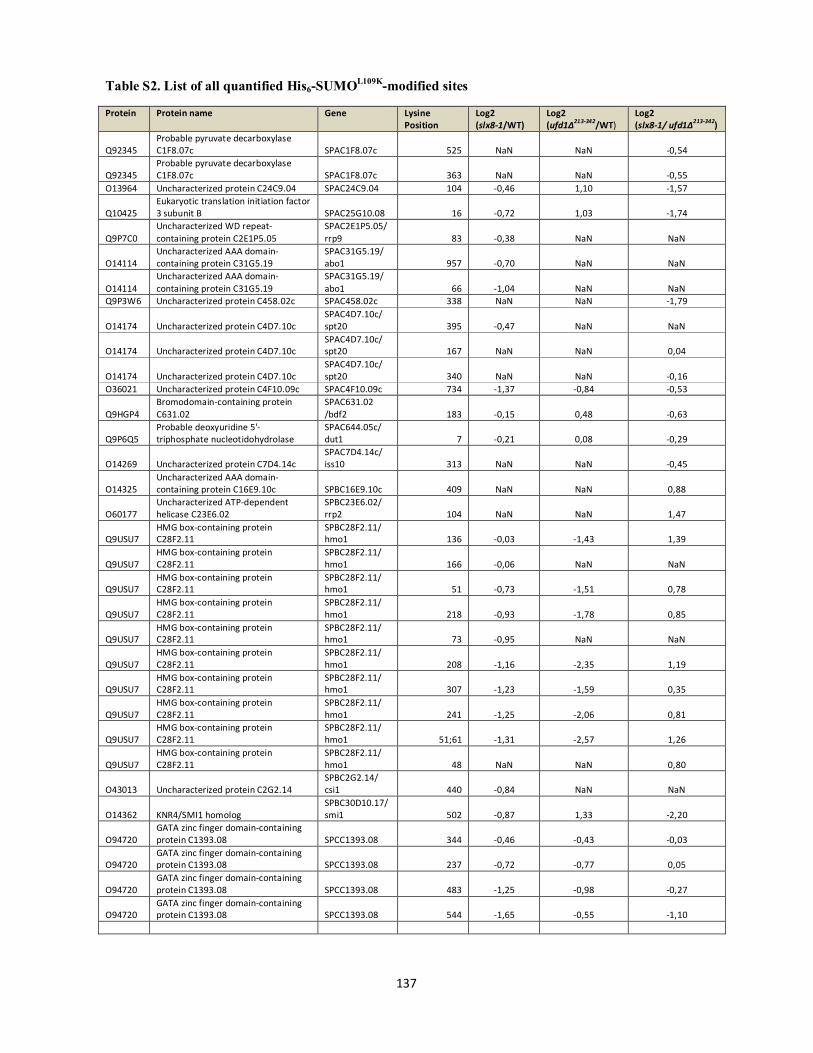
Table S2. List of all quantified His6-SUMOL109K-modified sites
137
Protein Protein name Gene Lysine Position
Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 525 NaN NaN -0,54
Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 363 NaN NaN -0,55
O13964 Uncharacterized protein C24C9.04 SPAC24C9.04 104 -0,46 1,10 -1,57
Q10425 Eukaryotic translation initiation factor 3 subunit B SPAC25G10.08 16 -0,72 1,03 -1,74
Q9P7C0 Uncharacterized WD repeat-containing protein C2E1P5.05
SPAC2E1P5.05/ rrp9 83 -0,38 NaN NaN
O14114 Uncharacterized AAA domain-containing protein C31G5.19
SPAC31G5.19/ abo1 957 -0,70 NaN NaN
O14114 Uncharacterized AAA domain-containing protein C31G5.19
SPAC31G5.19/ abo1 66 -1,04 NaN NaN
Q9P3W6 Uncharacterized protein C458.02c SPAC458.02c 338 NaN NaN -1,79
O14174 Uncharacterized protein C4D7.10c SPAC4D7.10c/ spt20 395 -0,47 NaN NaN
O14174 Uncharacterized protein C4D7.10c SPAC4D7.10c/ spt20 167 NaN NaN 0,04
O14174 Uncharacterized protein C4D7.10c SPAC4D7.10c/ spt20 340 NaN NaN -0,16
O36021 Uncharacterized protein C4F10.09c SPAC4F10.09c 734 -1,37 -0,84 -0,53
Q9HGP4 Bromodomain-containing protein C631.02
SPAC631.02 /bdf2 183 -0,15 0,48 -0,63
Q9P6Q5 Probable deoxyuridine 5'-triphosphate nucleotidohydrolase
SPAC644.05c/ dut1 7 -0,21 0,08 -0,29
O14269 Uncharacterized protein C7D4.14c SPAC7D4.14c/ iss10 313 NaN NaN -0,45
O14325 Uncharacterized AAA domain-containing protein C16E9.10c SPBC16E9.10c 409 NaN NaN 0,88
O60177 Uncharacterized ATP-dependent helicase C23E6.02
SPBC23E6.02/ rrp2 104 NaN NaN 1,47
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 136 -0,03 -1,43 1,39
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 166 -0,06 NaN NaN
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 51 -0,73 -1,51 0,78
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 218 -0,93 -1,78 0,85
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 73 -0,95 NaN NaN
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 208 -1,16 -2,35 1,19
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 307 -1,23 -1,59 0,35
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 241 -1,25 -2,06 0,81
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 51;61 -1,31 -2,57 1,26
Q9USU7 HMG box-containing protein C28F2.11
SPBC28F2.11/ hmo1 48 NaN NaN 0,80
O43013 Uncharacterized protein C2G2.14 SPBC2G2.14/ csi1 440 -0,84 NaN NaN
O14362 KNR4/SMI1 homolog SPBC30D10.17/ smi1 502 -0,87 1,33 -2,20
O94720 GATA zinc finger domain-containing protein C1393.08 SPCC1393.08 344 -0,46 -0,43 -0,03
O94720 GATA zinc finger domain-containing protein C1393.08 SPCC1393.08 237 -0,72 -0,77 0,05
O94720 GATA zinc finger domain-containing protein C1393.08 SPCC1393.08 483 -1,25 -0,98 -0,27
O94720 GATA zinc finger domain-containing protein C1393.08 SPCC1393.08 544 -1,65 -0,55 -1,10
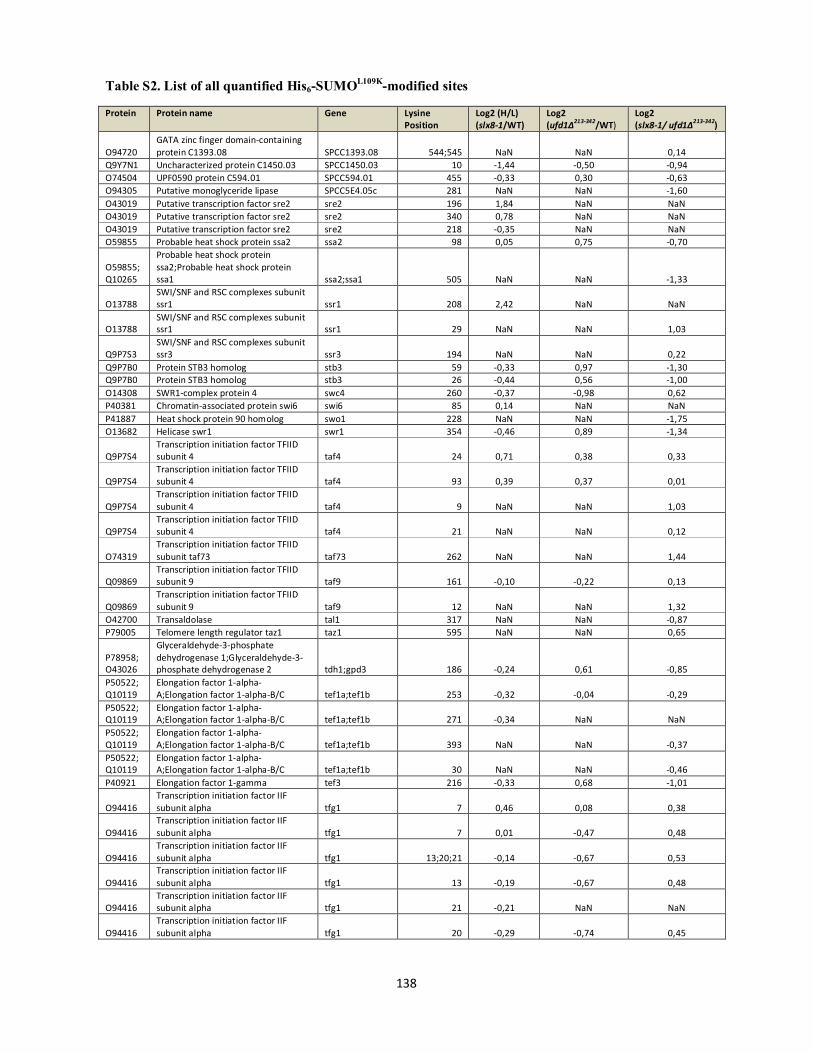
Table S2. List of all quantified His6-SUMOL109K-modified sites
138
Protein Protein name Gene Lysine Position
Log2 (H/L) (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O94720 GATA zinc finger domain-containing protein C1393.08 SPCC1393.08 544;545 NaN NaN 0,14
Q9Y7N1 Uncharacterized protein C1450.03 SPCC1450.03 10 -1,44 -0,50 -0,94 O74504 UPF0590 protein C594.01 SPCC594.01 455 -0,33 0,30 -0,63 O94305 Putative monoglyceride lipase SPCC5E4.05c 281 NaN NaN -1,60 O43019 Putative transcription factor sre2 sre2 196 1,84 NaN NaN O43019 Putative transcription factor sre2 sre2 340 0,78 NaN NaN O43019 Putative transcription factor sre2 sre2 218 -0,35 NaN NaN O59855 Probable heat shock protein ssa2 ssa2 98 0,05 0,75 -0,70
O59855;Q10265
Probable heat shock protein ssa2;Probable heat shock protein ssa1 ssa2;ssa1 505 NaN NaN -1,33
O13788 SWI/SNF and RSC complexes subunit ssr1 ssr1 208 2,42 NaN NaN
O13788 SWI/SNF and RSC complexes subunit ssr1 ssr1 29 NaN NaN 1,03
Q9P7S3 SWI/SNF and RSC complexes subunit ssr3 ssr3 194 NaN NaN 0,22
Q9P7B0 Protein STB3 homolog stb3 59 -0,33 0,97 -1,30 Q9P7B0 Protein STB3 homolog stb3 26 -0,44 0,56 -1,00 O14308 SWR1-complex protein 4 swc4 260 -0,37 -0,98 0,62 P40381 Chromatin-associated protein swi6 swi6 85 0,14 NaN NaN P41887 Heat shock protein 90 homolog swo1 228 NaN NaN -1,75 O13682 Helicase swr1 swr1 354 -0,46 0,89 -1,34
Q9P7S4 Transcription initiation factor TFIID subunit 4 taf4 24 0,71 0,38 0,33
Q9P7S4 Transcription initiation factor TFIID subunit 4 taf4 93 0,39 0,37 0,01
Q9P7S4 Transcription initiation factor TFIID subunit 4 taf4 9 NaN NaN 1,03
Q9P7S4 Transcription initiation factor TFIID subunit 4 taf4 21 NaN NaN 0,12
O74319 Transcription initiation factor TFIID subunit taf73 taf73 262 NaN NaN 1,44
Q09869 Transcription initiation factor TFIID subunit 9 taf9 161 -0,10 -0,22 0,13
Q09869 Transcription initiation factor TFIID subunit 9 taf9 12 NaN NaN 1,32
O42700 Transaldolase tal1 317 NaN NaN -0,87 P79005 Telomere length regulator taz1 taz1 595 NaN NaN 0,65
P78958;O43026
Glyceraldehyde-3-phosphate dehydrogenase 1;Glyceraldehyde-3-phosphate dehydrogenase 2 tdh1;gpd3 186 -0,24 0,61 -0,85
P50522;Q10119
Elongation factor 1-alpha-A;Elongation factor 1-alpha-B/C tef1a;tef1b 253 -0,32 -0,04 -0,29
P50522;Q10119
Elongation factor 1-alpha-A;Elongation factor 1-alpha-B/C tef1a;tef1b 271 -0,34 NaN NaN
P50522;Q10119
Elongation factor 1-alpha-A;Elongation factor 1-alpha-B/C tef1a;tef1b 393 NaN NaN -0,37
P50522;Q10119
Elongation factor 1-alpha-A;Elongation factor 1-alpha-B/C tef1a;tef1b 30 NaN NaN -0,46
P40921 Elongation factor 1-gamma tef3 216 -0,33 0,68 -1,01
O94416 Transcription initiation factor IIF subunit alpha tfg1 7 0,46 0,08 0,38
O94416 Transcription initiation factor IIF subunit alpha tfg1 7 0,01 -0,47 0,48
O94416 Transcription initiation factor IIF subunit alpha tfg1 13;20;21 -0,14 -0,67 0,53
O94416 Transcription initiation factor IIF subunit alpha tfg1 13 -0,19 -0,67 0,48
O94416 Transcription initiation factor IIF subunit alpha tfg1 21 -0,21 NaN NaN
O94416 Transcription initiation factor IIF subunit alpha tfg1 20 -0,29 -0,74 0,45
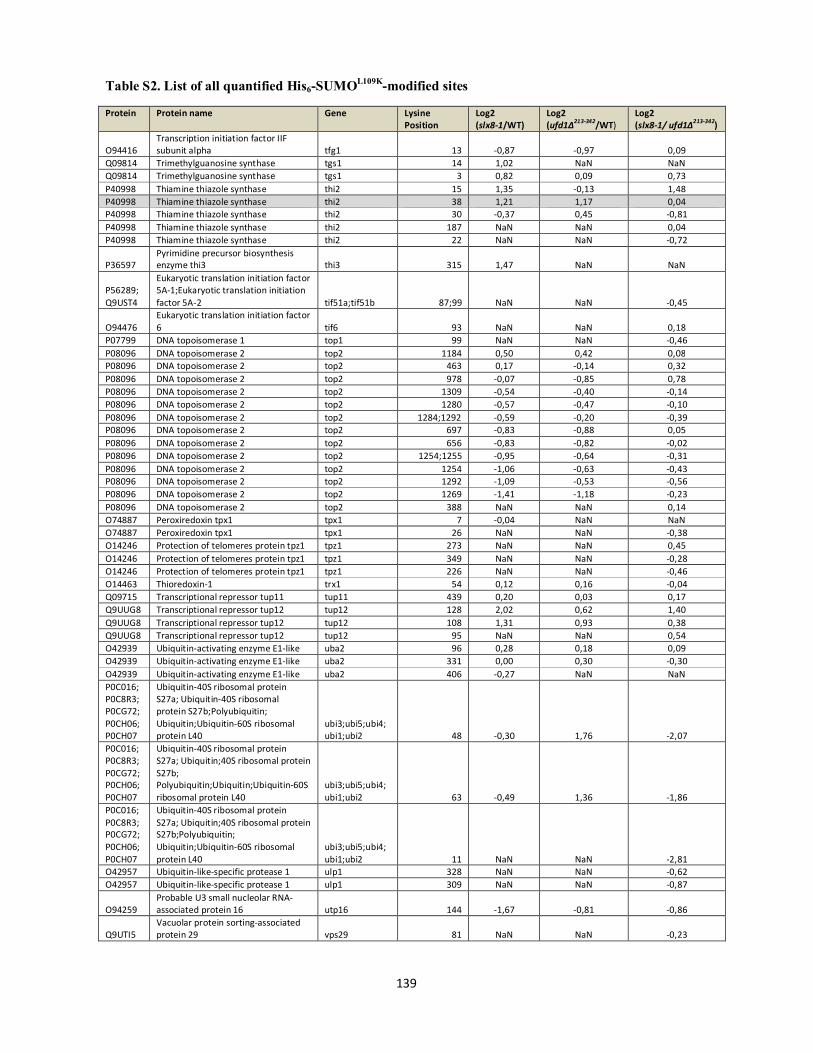
Table S2. List of all quantified His6-SUMOL109K-modified sites
139
Protein Protein name Gene Lysine Position
Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O94416 Transcription initiation factor IIF subunit alpha tfg1 13 -0,87 -0,97 0,09
Q09814 Trimethylguanosine synthase tgs1 14 1,02 NaN NaN Q09814 Trimethylguanosine synthase tgs1 3 0,82 0,09 0,73 P40998 Thiamine thiazole synthase thi2 15 1,35 -0,13 1,48 P40998 Thiamine thiazole synthase thi2 38 1,21 1,17 0,04 P40998 Thiamine thiazole synthase thi2 30 -0,37 0,45 -0,81 P40998 Thiamine thiazole synthase thi2 187 NaN NaN 0,04 P40998 Thiamine thiazole synthase thi2 22 NaN NaN -0,72
P36597 Pyrimidine precursor biosynthesis enzyme thi3 thi3 315 1,47 NaN NaN
P56289;Q9UST4
Eukaryotic translation initiation factor 5A-1;Eukaryotic translation initiation factor 5A-2 tif51a;tif51b 87;99 NaN NaN -0,45
O94476 Eukaryotic translation initiation factor 6 tif6 93 NaN NaN 0,18
P07799 DNA topoisomerase 1 top1 99 NaN NaN -0,46 P08096 DNA topoisomerase 2 top2 1184 0,50 0,42 0,08 P08096 DNA topoisomerase 2 top2 463 0,17 -0,14 0,32 P08096 DNA topoisomerase 2 top2 978 -0,07 -0,85 0,78 P08096 DNA topoisomerase 2 top2 1309 -0,54 -0,40 -0,14 P08096 DNA topoisomerase 2 top2 1280 -0,57 -0,47 -0,10 P08096 DNA topoisomerase 2 top2 1284;1292 -0,59 -0,20 -0,39 P08096 DNA topoisomerase 2 top2 697 -0,83 -0,88 0,05 P08096 DNA topoisomerase 2 top2 656 -0,83 -0,82 -0,02 P08096 DNA topoisomerase 2 top2 1254;1255 -0,95 -0,64 -0,31 P08096 DNA topoisomerase 2 top2 1254 -1,06 -0,63 -0,43 P08096 DNA topoisomerase 2 top2 1292 -1,09 -0,53 -0,56 P08096 DNA topoisomerase 2 top2 1269 -1,41 -1,18 -0,23 P08096 DNA topoisomerase 2 top2 388 NaN NaN 0,14 O74887 Peroxiredoxin tpx1 tpx1 7 -0,04 NaN NaN O74887 Peroxiredoxin tpx1 tpx1 26 NaN NaN -0,38 O14246 Protection of telomeres protein tpz1 tpz1 273 NaN NaN 0,45 O14246 Protection of telomeres protein tpz1 tpz1 349 NaN NaN -0,28 O14246 Protection of telomeres protein tpz1 tpz1 226 NaN NaN -0,46 O14463 Thioredoxin-1 trx1 54 0,12 0,16 -0,04 Q09715 Transcriptional repressor tup11 tup11 439 0,20 0,03 0,17 Q9UUG8 Transcriptional repressor tup12 tup12 128 2,02 0,62 1,40 Q9UUG8 Transcriptional repressor tup12 tup12 108 1,31 0,93 0,38 Q9UUG8 Transcriptional repressor tup12 tup12 95 NaN NaN 0,54 O42939 Ubiquitin-activating enzyme E1-like uba2 96 0,28 0,18 0,09 O42939 Ubiquitin-activating enzyme E1-like uba2 331 0,00 0,30 -0,30 O42939 Ubiquitin-activating enzyme E1-like uba2 406 -0,27 NaN NaN P0C016;P0C8R3;P0CG72;P0CH06;P0CH07
Ubiquitin-40S ribosomal protein S27a; Ubiquitin-40S ribosomal protein S27b;Polyubiquitin; Ubiquitin;Ubiquitin-60S ribosomal protein L40
ubi3;ubi5;ubi4;ubi1;ubi2 48 -0,30 1,76 -2,07
P0C016;P0C8R3;P0CG72;P0CH06;P0CH07
Ubiquitin-40S ribosomal protein S27a; Ubiquitin;40S ribosomal protein S27b; Polyubiquitin;Ubiquitin;Ubiquitin-60S ribosomal protein L40
ubi3;ubi5;ubi4;ubi1;ubi2 63 -0,49 1,36 -1,86
P0C016;P0C8R3;P0CG72;P0CH06;P0CH07
Ubiquitin-40S ribosomal protein S27a; Ubiquitin;40S ribosomal protein S27b;Polyubiquitin; Ubiquitin;Ubiquitin-60S ribosomal protein L40
ubi3;ubi5;ubi4;ubi1;ubi2 11 NaN NaN -2,81
O42957 Ubiquitin-like-specific protease 1 ulp1 328 NaN NaN -0,62 O42957 Ubiquitin-like-specific protease 1 ulp1 309 NaN NaN -0,87
O94259 Probable U3 small nucleolar RNA-associated protein 16 utp16 144 -1,67 -0,81 -0,86
Q9UTI5 Vacuolar protein sorting-associated protein 29 vps29 81 NaN NaN -0,23
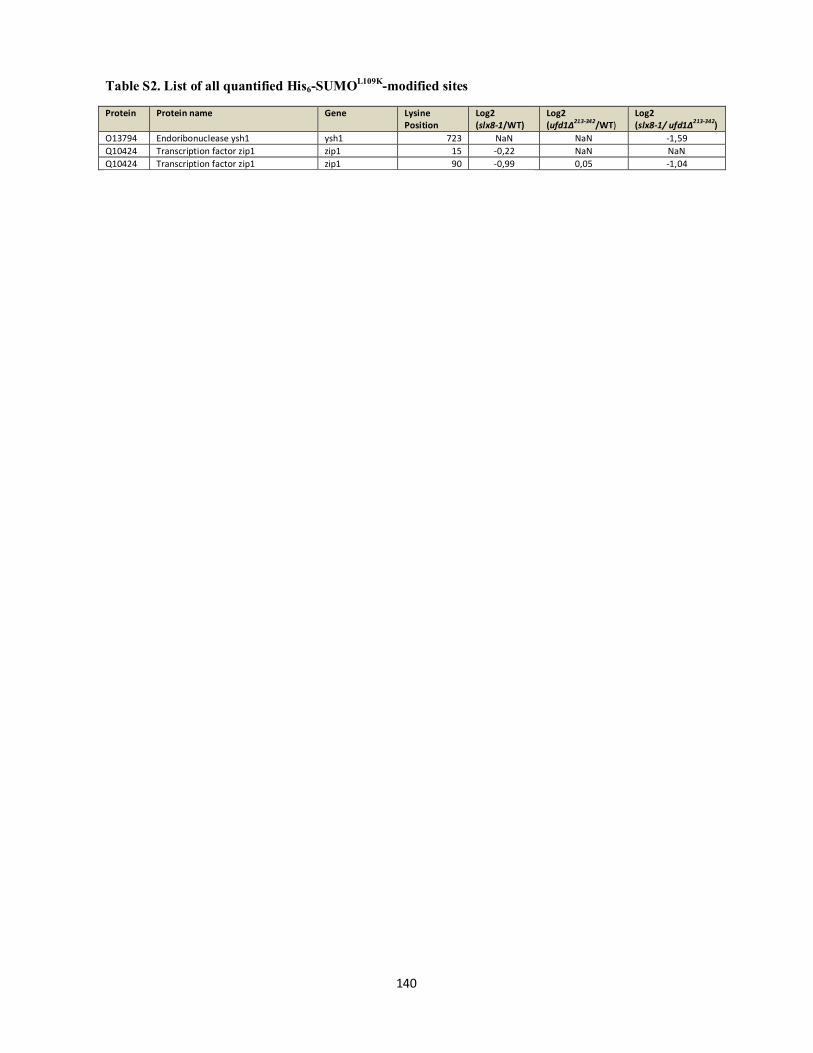
Table S2. List of all quantified His6-SUMOL109K-modified sites
140
Protein Protein name Gene Lysine Position
Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O13794 Endoribonuclease ysh1 ysh1 723 NaN NaN -1,59 Q10424 Transcription factor zip1 zip1 15 -0,22 NaN NaN Q10424 Transcription factor zip1 zip1 90 -0,99 0,05 -1,04

Table S3. List of all quantified proteins from crude SILAC lysates
141
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
B5BP46 Putative glutathione S-transferase C1183.02 SPBC460.02c -0,18 -0,21 0,03 G2TRP7 Mitochondrial import receptor subunit tom5 tom5 0,21 -0,01 0,21 G2TRQ0 Uncharacterized protein tam13 tam13 0,61 0,54 0,07
G2TRT6 NADH dehydrogenase [ubiquinone] 1 alpha subcomplex subunit 4 homolog SPCC417.16 -0,02 -0,63 0,61
O00084 Urease ure1 0,25 0,07 0,18 O00087 Dihydrolipoyl dehydrogenase, mitochondrial dld1 -0,09 0,11 -0,20 O00091 Glucose-6-phosphate 1-dehydrogenase zwf1 -0,07 0,35 -0,42 O00102 Ubiquitin-conjugating enzyme E2-18 kDa ubc7 0,04 0,14 -0,11 O13282 Transcription initiation factor TFIID subunit 5 taf5 0,16 0,38 -0,22 O13326 O-acetylhomoserine (thiol)-lyase SPBC428.11 -0,36 0,22 -0,58 O13351 Ubiquitin-like protein pmt3/smt3 pmt3 0,04 -0,02 0,07 O13370 ATP-dependent RNA helicase ded1 ded1 -0,01 -0,09 0,08 O13614 40S ribosomal protein S10-B rps10b -0,06 -0,35 0,30 O13620 Multiple RNA-binding domain-containing protein 1 mrd1 -0,43 -0,15 -0,28 O13633 Uncharacterized J domain-containing protein C17A3.05c SPB C17A3.05c -0,12 0,70 -0,81
O13636 Mitochondrial import inner membrane translocase subunit tim50 tim50 -0,16 -0,33 0,17
O13637 Protein transport protein sec31 sec31 0,08 0,12 -0,03 O13639 Adenosylhomocysteinase 0,06 -0,03 0,08 O13640 Uncharacterized alpha-1,2-galactosyltransferase C8D2.17 SPB C8D2.17 -0,02 0,35 -0,37 O13648 Ribosomal protein arginine N-methytransferase rmt3 rmt3 -0,04 -0,06 0,02 O13651 Isoleucine--tRNA ligase, cytoplasmic -0,17 -0,24 0,07 O13656 Probable mitochondrial import receptor subunit tom40 tom40 0,02 -0,05 0,07 O13666 C-5 sterol desaturase erg32 erg32 -0,02 0,30 -0,33 O13671 Importin-alpha re-exporter kap109 0,10 0,15 -0,04 O13672 60S ribosomal protein L8 rpl8 0,00 -0,26 0,26 O13685 Ubiquitin-conjugating enzyme E2 13 ubc13 0,14 0,27 -0,13 O13687 V-type proton ATPase subunit E vma4 0,10 0,06 0,04 O13696 Isocitrate dehydrogenase [NAD] subunit 1, mitochondrial idh1 -0,06 0,46 -0,52 O13704 Thioredoxin domain-containing protein C13F5.05, mitochondrial SPAC13F5.05 -0,43 0,52 -0,95 O13711 Uncharacterized protein C14C4.04 SPAC14C4.04 0,12 0,34 -0,21 O13718 Vacuolar transporter chaperone 2 vtc2 0,12 0,32 -0,20 O13724 Zinc finger protein zpr1 zpr1 -0,04 0,34 -0,39 O13727 Uncharacterized protein C15A10.07 SPAC15A10.07 -0,01 0,31 -0,32 O13736 Actin cytoskeleton-regulatory complex protein sla1 sla1 0,14 -0,01 0,15 O13739 Probable chorismate mutase SPAC16E8.04c -0,04 0,03 -0,07 O13741 Nucleolar protein 12 nop12 0,04 0,33 -0,30
O13750 Probable succinyl-CoA ligase [ADP-forming] subunit alpha, mitochondrial SPAC16E8.17c 0,03 -0,03 0,06
O13753 V-type proton ATPase subunit d vma6 0,22 0,31 -0,10 O13754 Hsp70/Hsp90 co-chaperone cns1 cns1 0,06 0,14 -0,08 O13755 Putative fumarate reductase SPAC17A2.05 0,20 -0,24 0,45 O13770 Uncharacterized membrane protein C17A5.08 SPAC17A5.08 0,05 0,24 -0,18 O13774 GTP cyclohydrolase 1 SPAC17A5.13 -0,18 -0,31 0,13 O13775 Probable glutamate--tRNA ligase, cytoplasmic SPAC17A5.15c 0,01 -0,04 0,05 O13777 Uncharacterized protein C1610.01 -0,24 0,79 -1,03 O13783 Vacuolar protein-sorting protein bro1 bro1 0,05 0,16 -0,11 O13784 40S ribosomal protein S24-A rps24a 0,01 -0,31 0,32 O13787 Translocation protein sec62 sec62 -0,08 0,28 -0,35 O13788 SWI/SNF and RSC complexes subunit ssr1 ssr1 -0,07 0,21 -0,28
O13789 Uncharacterized beta-glucan synthesis-associated protein C17G6.11c SPAC17G6.11c -0,04 0,40 -0,44
O13791 Protein slt1 slt1 0,06 0,99 -0,93 O13792 ATP-dependent RNA helicase uap56 uap56 0,07 -0,02 0,08 O13793 Probable mitochondrial transport protein fsf1 fsf1 0,03 0,09 -0,06
O13797 Small glutamine-rich tetratricopeptide repeat-containing protein 2 sgt2 0,08 0,04 0,04
O13801 Uncharacterized RNA-binding protein C17H9.04c SPAC17H9.04c -0,13 -0,13 -0,01 O13802 Probable rRNA-processing protein ebp2 ebp2 -0,09 0,00 -0,10 O13804 Uncharacterized protein C17H9.07 SPAC17H9.07 -0,15 -0,06 -0,09 O13808 Uncharacterized protein C17H9.11 SPAC17H9.11 -0,03 0,19 -0,22 O13809 Uncharacterized FAD-binding protein C17H9.12c SPAC17H9.12c 0,08 0,02 0,06 O13810 Probable glutamate 5-kinase SPAC17H9.13c 0,03 0,10 -0,07 O13811 Protein disulfide-isomerase C17H9.14c SPAC17H9.14c 0,04 0,23 -0,19 O13813 Mitochondrial import receptor subunit tom22 tom22 -0,03 -0,06 0,03 O13820 Cytochrome P450 61 erg5 0,11 0,32 -0,21 O13831 Tryptophan synthase trp2 -0,12 -0,11 -0,01 O13844 Uncharacterized mitochondrial carrier C19G12.05 SPAC19G12.05 -0,07 -0,09 0,02
O13846 Ceramide very long chain fatty acid hydroxylase-like protein C19G12.08 SPAC19G12.08 -0,05 0,22 -0,27
O13848 NAD/NADP-dependent indole-3-acetaldehyde reductase SPAC19G12.09 0,01 1,33 -1,32

Table S3. List of all quantified proteins from crude SILAC lysates
142
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O13849 Carboxypeptidase Y cpy1 0,19 0,57 -0,38 O13857 Putative lysophospholipase C1A6.03c SPAC1A6.03c 0,01 0,07 -0,05 O13861 Uncharacterized protein C1A6.10 0,33 0,30 0,03 O13864 Importin subunit beta-1 kap95 0,05 -0,02 0,07 O13867 Uracil phosphoribosyltransferase 1 SPAC1B3.01c 0,24 -0,19 0,43 O13870 General negative regulator of transcription subunit 3 not3 -0,11 0,04 -0,15 O13871 Uncharacterized methyltransferase C1B3.06c SPAC1B3.06c 0,20 0,56 -0,35 O13878 U3 small nucleolar RNA-associated protein 17 utp17 0,08 0,36 -0,28 O13880 Vitamin H transporter 1 vht1 -0,21 -0,03 -0,18 O13892 Uncharacterized protein C20G4.08 0,06 -0,23 0,29 O13898 Dolichyl-phosphate-mannose--protein mannosyltransferase 1 ogm1 -0,04 -0,14 0,10
O13899 Uncharacterized CDP-alcohol phosphatidyltransferase class-I family protein C22A12.08c
SPAC22A12.08c 0,19 0,18 0,01
O13902 Dihydroxyacetone kinase 1 dak1 -0,04 0,27 -0,31 O13907 Probable ATP-citrate synthase subunit 2 SPAC22A12.16 -0,06 -0,14 0,08 O13910 U3 small nucleolar ribonucleoprotein protein mpp10 mpp10 -0,05 0,30 -0,35 O13914 Alanine--tRNA ligase ala1 -0,01 -0,07 0,05
O13917 Uncharacterized protein C23C11.13c SPAC23C11.13c -0,06 -0,06 0,00
O13920 Uncharacterized protein C23C11.17 SPAC23C11.17 0,08 0,36 -0,28 O13923 Coronin-like protein crn1 crn1 0,05 -0,06 0,11 O13928 GTP-binding protein rho3 rho3 0,16 0,11 0,05 O13929 Uncharacterized protein C23C4.09c SPAC23C4.09c 0,00 -0,18 0,19 O13936 Transcription elongation factor spt5 spt5 0,11 0,20 -0,09 O13940 Probable low-specificity L-threonine aldolase gly1 -0,23 0,73 -0,96 O13946 Endoplasmic reticulum vesicle protein 25 erv25 0,07 0,20 -0,13 O13953 Chromatin modification-related protein eaf3 alp13 0,00 0,22 -0,22 O13956 Ribosome biogenesis protein tsr1 homolog SPAC23H4.15 -0,13 -0,11 -0,01 O13963 Diphosphomevalonate decarboxylase mvd1 -0,01 -0,16 0,15 O13966 Aconitate hydratase, mitochondrial SPAC24C9.06c 0,17 0,39 -0,22 O13968 Uncharacterized carboxypeptidase C24C9.08 SPAC24C9.08 0,23 0,00 0,23 O13972 Probable serine hydroxymethyltransferase, cytosolic SPAC24C9.12c 0,14 -0,23 0,37 O13981 ER membrane protein complex subunit 1 emc1 -0,03 0,49 -0,52 O13986 Uncharacterized protein C26H5.04 SPAC26H5.04 0,27 1,01 -0,75 O13989 Uncharacterized protein C26H5.07c SPAC26H5.07c -0,01 -0,26 0,24 O13991 Uncharacterized oxidoreductase C26H5.09c SPAC26H5.09c 0,43 -0,53 0,97 O13998 Uncharacterized GTP-binding protein C27E2.03c SPAC27E2.03c -0,01 -0,16 0,14 O14003 Replication factor C subunit 3 rfc3 -0,08 -0,02 -0,06 O14006 37S ribosomal protein S9, mitochondrial mrps9 0,09 -0,20 0,29 O14007 H/ACA ribonucleoprotein complex subunit 4 cbf5 0,06 0,03 0,02 O14008 Myosin 1 light chain cam2 cam2 0,01 0,14 -0,12 O14011 Pre-mRNA-splicing factor cwf8 cwf8 0,07 0,05 0,02 O14018 Serine--tRNA ligase, cytoplasmic SPAC29A4.15 -0,01 0,01 -0,02 O14019 Serine/threonine-protein kinase hal4 hal4 -0,10 0,05 -0,15 O14023 Probable elongator complex protein 3 elp3 -0,18 0,03 -0,21 O14027 Probable pyridoxine biosynthesis PDX1-like protein snz1 -0,29 0,25 -0,54 O14029 COPII coat assembly protein sec16 sec16 -0,75 -0,27 -0,48 O14037 UPF0538 protein C2C4.04c SPAC2C4.04c 0,00 0,64 -0,63 O14044 Pre-rRNA-processing protein pno1 rbp28 -0,12 -0,24 0,12 O14047 Serine/threonine-protein kinase ppk11 ppk11 -0,09 -0,15 0,06 O14049;Q9P7B2 40S ribosomal protein S8-A;40S ribosomal protein S8-B rps8a;rps8b 0,01 -0,25 0,26 O14053 Uncharacterized WD repeat-containing protein C1672.07 SPCC1672.07 -0,11 0,10 -0,21 O14055 Tyrosine--tRNA ligase, cytoplasmic tys1 -0,01 -0,15 0,14 O14057 Probable guanine deaminase SPCC1672.03c -0,13 -0,32 0,18 O14062 40S ribosomal protein S12-A rps12a 0,03 -0,18 0,20 O14063 Importin subunit alpha-1 cut15 -0,02 0,09 -0,11 O14065 Uncharacterized protein C962.01 0,08 0,26 -0,18 O14069 Probable 60S ribosomal protein L28e rpl28e 0,04 -0,34 0,38 O14072 Cation-transporting ATPase 4 cta4 0,20 -0,09 0,29 O14075 Putative 2-hydroxyacid dehydrogenase UNK4.10 0,17 -0,07 0,23 O14077 Putative zinc protease mug138 mug138 -0,13 -0,05 -0,08 O14080 Uncharacterized protein UNK4,15 0,15 0,64 -0,49
O14081 Putative alpha,alpha-trehalose-phosphate synthase [UDP-forming] 106 kDa subunit 0,02 0,59 -0,57
O14085 Translocation protein sec72 sec72 -0,12 0,14 -0,26 O14086 DNA-directed RNA polymerase I subunit rpa49 rpa49 -0,14 -0,04 -0,10 O14087 Single-stranded DNA-binding protein rim1, mitochondrial rim1 0,08 0,13 -0,05 O14088 Uncharacterized oxidoreductase C2F3.05c SPAC2F3.05c 0,51 1,00 -0,49 O14089 Importin subunit beta-2 kap104 -0,19 -0,14 -0,05 O14092 5-aminolevulinate synthase, mitochondrial hem1 -0,19 -0,14 -0,05 O14094 Putative exopolyphosphatase SPAC2F3.11 0,03 0,35 -0,32

Table S3. List of all quantified proteins from crude SILAC lysates
143
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O14096 Queuine tRNA-ribosyltransferase-like protein SPAC2F3.13c -0,09 -0,11 0,02 O14104 Probable homoisocitrate dehydrogenase lys12 -0,27 -0,32 0,05 O14105 Ribulose-phosphate 3-epimerase SPAC31G5.05c 0,10 0,20 -0,10 O14110 Probable aminomethyltransferase, mitochondrial gcv1 0,20 -0,44 0,64 O14112 40S ribosomal protein S10-A rps10a 0,02 -0,21 0,23
O14121 Probable NADH-ubiquinone oxidoreductase C3A11.07, mitochondrial SPAC3A11.07 0,01 -0,42 0,43
O14124 Uncharacterized dipeptidase C3A11.10c SPAC3A11.10c 0,15 0,83 -0,67 O14126 26S protease regulatory subunit 6A tbp1 -0,01 0,32 -0,33 O14134 mRNA export factor elf1 elf1 0,01 -0,19 0,20 O14139 Chromodomain helicase hrp3 hrp3 -0,04 0,04 -0,08 O14141 Maf-like protein C3G6.03c SPAC3G6.03c -0,07 0,09 -0,16 O14148 Putative DNA helicase ino80 ino80 -0,50 -0,41 -0,09 O14150 40S ribosomal protein S14 rps14a 0,01 -0,26 0,26 O14155 UPF0047 protein C4A8.02c SPAC4A8.02c 0,13 0,32 -0,20 O14156 Protein phosphatase 2C homolog 4 ptc4 -0,07 -0,24 0,17 O14159 Sphingoid long chain base kinase 4 lcb4 0,21 0,41 -0,20 O14164 Eukaryotic translation initiation factor 3 subunit C nip1 -0,03 -0,04 0,01 O14165 Uncharacterized protein C4C5.01 SPAC4C5.01 -0,12 -0,26 0,14 O14179 Ribosome maturation protein sdo1 sdo1 -0,05 0,02 -0,07 O14187 Pre-mRNA-splicing factor spp42 spp42 0,17 0,38 -0,21 O14192 Aromatic amino acid aminotransferase C56E4.03 SPAC56E4.03 0,05 0,13 -0,08 O14209 Uncharacterized aminotransferase C6B12.04c SPAC6B12.04c 0,09 0,13 -0,04 O14217 Probable mitochondrial import receptor subunit tom70 tom70 0,02 -0,06 0,08 O14222 Syntaxin-like protein fsv1 fsv1 -0,19 0,11 -0,30 O14224 Rho GDP-dissociation inhibitor SPAC6F12.06 0,11 0,26 -0,15 O14225 Mitochondrial import receptor subunit tom20 tom20 0,17 -0,06 0,22 O14228 Probable phosphoribosylformylglycinamidine synthase ade3 -0,07 -0,26 0,19 O14230 Farnesyl pyrophosphate synthase fps1 -0,03 -0,09 0,06 O14236 Nucleolar GTP-binding protein 2 nog2 -0,34 0,00 -0,34 O14241 Actin-related protein 2/3 complex subunit 2 arc2 -0,01 0,15 -0,16 O14242 Putative pyridoxal kinase C6F6.11c SPAC6F6.11c 0,07 0,32 -0,25 O14250 Probable proteasome subunit alpha type-1 SPAC6G10.04c 0,02 0,33 -0,31 O14254 Probable isocitrate dehydrogenase [NADP], mitochondrial idp1 0,00 -0,57 0,57 O14255 Probable mannosyl-oligosaccharide glucosidase SPAC6G10.09 -0,06 0,04 -0,10 O14265 V-type proton ATPase subunit H vma13 0,04 0,17 -0,14 O14273 Meiotically up-regulated protein C8C9.04 SPAC8C9.04 0,09 0,27 -0,18 O14274 D-tyrosyl-tRNA(Tyr) deacylase dtd1 -0,13 0,38 -0,51 O14289 3-isopropylmalate dehydratase leu2 0,04 -0,07 0,11 O14290 Uncharacterized protein C9E9.04 SPAC9E9.04 -0,10 0,32 -0,43 O14293 Putative aldehyde dehydrogenase-like protein C9E9.09c SPAC9E9.09c -0,07 0,18 -0,25 O14295 Pyridoxal reductase plr1 -0,07 1,31 -1,37 O14301 Uncharacterized WD repeat-containing protein C9G1.05 SPAC9G1.05 0,08 0,18 -0,11
O14307 tRNA (adenine(58)-N(1))-methyltransferase catalytic subunit trm61 cpd1 0,07 0,09 -0,02
O14310 Nucleoporin npp106 npp106 0,35 0,37 -0,02 O14313 Putative peroxiredoxin pmp20 pmp20 0,05 0,11 -0,06 O14317 60S acidic ribosomal protein P2-C rpp203 0,06 -0,33 0,39 O14319 CUE domain-containing protein 5 SPBC16E9.02c 0,08 0,09 -0,01 O14321 Sterol 24-C-methyltransferase erg6 erg6 0,10 -0,01 0,11 O14324 Uncharacterized membrane protein C16E9.09c SPBC16E9.09c 0,20 0,43 -0,23 O14327 Polyadenylate-binding protein 2 pab2 0,04 0,01 0,03 O14339 60S ribosomal protein L17-A rpl17a 0,03 -0,31 0,34 O14340 Oxysterol-binding protein homolog C2F12.05c SPBC2F12.05c 0,16 0,45 -0,28 O14344 Inosine-5-monophosphate dehydrogenase gua1 -0,06 0,14 -0,21 O14349 IMP-specific 5-nucleotidase 1 isn1 0,04 0,10 -0,06
O14351 Uncharacterized oxidoreductase C30D10.05c SPBC30D10.05c -0,04 -0,06 0,01
O14352 Probable U6 snRNA-associated Sm-like protein LSm4 lsm4 0,09 0,03 0,06
O14353
Biotin--protein ligase;Biotin--[methylmalonyl-CoA-carboxytransferase] ligase;Biotin--[propionyl-CoA-carboxylase [ATP-hydrolyzing]] ligase;Biotin--[methylcrotonoyl-CoA-carboxylase] ligase;Biotin--[acetyl-CoA-carboxylase] ligase bpl1 -0,23 -0,17 -0,07
O14359 Uncharacterized AIM2 family protein C30D10.14 SPBC30D10.14 0,07 0,43 -0,36 O14361 Putative prephenate dehydratase pha2 -0,27 -0,09 -0,18 O14363 60S ribosomal protein L1-A rpl1a 0,10 -0,09 0,19 O14368 Heat shock protein 16 hsp16 0,15 3,23 -3,07 O14369 Probable RNA-binding protein sce3 sce3 -0,10 -0,22 0,12 O14370 Branched-chain-amino-acid aminotransferase, mitochondrial eca39 -0,11 -0,10 -0,01 O14388 60S ribosomal protein L27-A rpl27a 0,01 -0,20 0,22 O14450 Probable prefoldin subunit 6 -0,03 0,08 -0,11 O14459 DNA-directed RNA polymerase II subunit rpb7 rpb7 0,09 -0,10 0,19

Table S3. List of all quantified proteins from crude SILAC lysates
144
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O14460 Elongation factor 2 eft201 -0,05 -0,11 0,06 O14463 Thioredoxin-1 trx1 -0,08 0,12 -0,20 O14466 Dolichol-phosphate mannosyltransferase dpm1 0,10 0,01 0,10 O14469 40S ribosomal protein S22 rps22a -0,01 -0,26 0,25 O14470 SWI/SNF and RSC complexes subunit ssr2 ssr2 0,00 0,11 -0,11 O36014 Aspartyl aminopeptidase 1 aap1 0,21 0,35 -0,14 O36021 Uncharacterized protein C4F10.09c SPAC4F10.09c 0,00 0,00 -0,01 O36022 Mannan polymerase complex subunit mnn9 mnn9 0,04 0,08 -0,05 O36023 Septin homolog spn1 spn1 0,21 0,30 -0,09 O36025 GYF domain-containing protein mpd2 mpd2 0,08 -0,01 0,09 O36032 Glutaredoxin-1 grx1 0,18 0,89 -0,71 O42644 CTP synthase ura7 -0,05 -0,04 -0,01 O42649 Structural maintenance of chromosomes protein 3 psm3 -0,05 0,39 -0,45 O42652 Aspartate aminotransferase, cytoplasmic aat2 -0,07 0,06 -0,13 O42661 Small nuclear ribonucleoprotein Sm D1 smd1 0,05 0,13 -0,08 O42668 Tetratricopeptide repeat protein 1 tpr1 -0,04 -0,30 0,25 O42699 60S ribosomal protein L19-B rpl19b 0,03 -0,27 0,29 O42700 Transaldolase tal1 0,00 0,34 -0,35 O42707 L-type lectin-like domain-containing protein C4F6.05c SPBC4F6.05c 0,13 0,06 0,07 O42842 Adenine phosphoribosyltransferase apt1 -0,15 -0,05 -0,10 O42846 60S ribosomal protein L34-A rpl34a -0,01 -0,30 0,28 O42848;O42991 60S ribosomal protein L16-A;60S ribosomal protein L16-B rpl16a;rpl16b 0,01 -0,27 0,28 O42849 Phenylalanine--tRNA ligase beta subunit SPAC23A1.12c -0,08 -0,12 0,04 O42851 Uncharacterized trans-sulfuration enzyme C23A1.14c SPAC23A1.14c 0,03 0,42 -0,39 O42854 SH3 domain-containing protein C23A1.17 SPAC23A1.17 0,11 0,22 -0,11 O42861 Uncharacterized ATP-dependent helicase C25A8.01c SPAC25A8.01c -0,10 0,03 -0,13 O42867 60S ribosomal protein L23 rpl23a 0,00 -0,34 0,34 O42870 Phenylalanine--tRNA ligase alpha subunit SPAC3G9.06 -0,03 -0,03 0,01 O42881 Acyl-protein thioesterase 1 SPAC8E11.04c 0,15 0,48 -0,34 O42885 Putative inorganic phosphate transporter C8E4.01c 0,03 -0,58 0,62 O42888 Uncharacterized oxidoreductase C8E4.04 SPBC8E4.04 0,22 0,06 0,15 O42893 Neutral trehalase ntp1 0,18 1,17 -0,98 O42897 Probable 26S proteasome regulatory subunit rpn3 rpn3 -0,03 0,16 -0,19 O42898 Probable catechol O-methyltransferase 1 SPBC119.03 0,03 1,93 -1,90 O42899 Protein sco1 sco1 0,67 0,28 0,39 O42908 Mitochondrial presequence protease cym1 -0,02 0,12 -0,14 O42909 Zinc-type alcohol dehydrogenase-like protein C16A3.02c SPBC16A3.02c 0,10 1,12 -1,02 O42914 Uncharacterized protein C16A3.08c SPBC16A3.08c -0,05 -0,10 0,05 O42923 RNA-binding post-transcriptional regulator cip1 cip1 0,15 0,54 -0,39 O42929 Translation machinery-associated protein 22 tma22 -0,09 0,23 -0,32 O42931 26S protease regulatory subunit 7 homolog rpt1 0,03 0,35 -0,32 O42932 Cytochrome b-c1 complex subunit 6 qcr6 0,02 -0,23 0,25 O42933 Dolichyl-phosphate-mannose--protein mannosyltransferase 4 ogm4 0,67 0,43 0,24 O42935 60S ribosomal protein L32-B rpl32b 0,02 -0,27 0,29 O42937 Probable coatomer subunit beta sec27 -0,01 0,00 -0,01 O42938 6-phosphofructokinase pfk1 -0,04 0,16 -0,20 O42940 Inorganic phosphate transport protein pho88 pho88 0,01 -0,18 0,19 O42943 Uncharacterized ABC transporter ATP-binding protein C16H5.08c SPBC16H5.08c 0,00 0,07 -0,07
O42945 Probable pre-mRNA-splicing factor ATP-dependent RNA helicase prp43 prp43 -0,04 0,05 -0,09
O42947 Uncharacterized protein C16H5.12c SPBC16H5.12c 0,16 0,33 -0,17 O42952 40S ribosomal protein S30 rps30a 0,00 -0,29 0,29 O42958 DNA topoisomerase 2-associated protein pat1 SPBC19G7.10c -0,04 0,15 -0,20 O42964 Transcription factor iws1 iws1 -0,10 0,22 -0,31 O42965 Uncharacterized protein C19G7.17 0,05 -0,35 0,40 O42975 UPF0614 protein C20F10.05 SPBC20F10.05 0,16 0,37 -0,20 O42978 U6 snRNA-associated Sm-like protein LSm5 lsm5 0,09 0,08 0,00 O42984 40S ribosomal protein S17-A rps17a 0,04 -0,21 0,25 O42993 FK506-binding protein 0,06 0,35 -0,29 O42998 Uncharacterized protein C27B12.08 0,09 -0,22 0,31 O43001 Inositol-1,4,5-trisphosphate 5-phosphatase 1 syj1 -0,05 -0,24 0,19 O43002 Protein transport protein sec61 subunit beta sbh1 0,02 -0,19 0,21 O43003 Protein mmf1, mitochondrial mmf1 0,05 0,03 0,02
O43007
C-1-tetrahydrofolate synthase, mitochondrial;Methylenetetrahydrofolate dehydrogenase;Methenyltetrahydrofolate cyclohydrolase;Formyltetrahydrofolate synthetase ade9 -0,02 0,05 -0,07
O43010 Glycylpeptide N-tetradecanoyltransferase nmt1 -0,04 0,08 -0,12 O43011 Histidine--tRNA ligase hts1 -0,02 0,03 -0,05 O43014 Mannose-6-phosphate isomerase pmi40 0,04 0,05 -0,01 O43021 Oxysterol-binding protein homolog C354.07c SPBC354.07c 0,05 0,19 -0,13

Table S3. List of all quantified proteins from crude SILAC lysates
145
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O43024 CUE domain-containing protein C354.10 SPBC354.10 -0,08 0,08 -0,16 O43026 Glyceraldehyde-3-phosphate dehydrogenase 2 gpd3 0,32 2,53 -2,21 O43028 Vacuolar protein 8 vac8 -0,02 0,28 -0,30 O43030 Hsp70 nucleotide exchange factor fes1 fes1 0,05 0,81 -0,76 O43036 DNA-directed RNA polymerase I subunit rpa43 rpa43 -0,02 -0,15 0,12 O43042 54S ribosomal protein L3, mitochondrial mrpl3 0,02 0,20 -0,18 O43044 Nucleoporin nup120 nup120 0,02 0,16 -0,14 O43046 V-type proton ATPase subunit F vma7 0,13 0,10 0,02 O43047 GrpE protein homolog, mitochondrial mge1 -0,17 0,15 -0,32 O43049 Serine/threonine-protein phosphatase T ppt1 0,01 -0,10 0,12 O43050 Sterol-4-alpha-carboxylate 3-dehydrogenase, decarboxylating erg26 0,13 0,22 -0,08 O43056 Probable homoserine kinase SPBC4C3.03 -0,05 0,01 -0,06 O43059 Cytoskeletal protein syp1 syp1 -0,21 0,03 -0,24 O43060 Eukaryotic translation initiation factor 3 subunit F eif3f -0,09 0,13 -0,22 O43063 Probable proteasome subunit beta type-6 SPBC4C3.10c 0,03 0,28 -0,25 O43065 Probable helicase mot1 mot1 0,12 -0,10 0,22 O43078 Protein sur2 sur2 -0,01 0,48 -0,49 O43079 AP-1 complex subunit beta-1 apl2 0,20 0,03 0,17
O43090 Probable NADH-ubiquinone oxidoreductase C947.15c, mitochondrial SPBC947.15c 0,03 -0,09 0,12
O59667 Histidine biosynthesis bifunctional protein his7;Phosphoribosyl-AMP cyclohydrolase;Phosphoribosyl-ATP pyrophosphatase his7 -0,05 0,02 -0,07
O59672 Uncharacterized ABC transporter ATP-binding protein C29A3.09c SPBC29A3.09c -0,02 0,01 -0,04 O59673 ATP synthase subunit H, mitochondrial atp14 0,09 -0,33 0,42 O59675 40S ribosomal protein S9-B rps9b 0,02 -0,25 0,26 O59677 37S ribosomal protein S23, mitochondrial rsm23 0,18 -0,25 0,43 O59678 Ribosome biogenesis regulatory protein homolog SPBC29A3.16 -0,02 -0,06 0,03 O59680 Cytochrome c1, heme protein, mitochondrial cyt1 0,12 -0,28 0,40 O59681 Uncharacterized protein C18E5.01 0,12 0,23 -0,11 O59682 Serine palmitoyltransferase 1 lcb1 0,11 0,21 -0,09 O59701 Cysteine synthase 1 cys11 0,16 -0,11 0,26 O59704 Elongator complex protein 1 iki3 0,92 0,94 -0,02 O59711 Saccharopine dehydrogenase [NADP(+), L-glutamate-forming] lys9 -0,05 0,00 -0,05 O59725 Formation of crista junctions protein C3E7.05c SPBC3E7.05c -0,05 -0,22 0,18 O59727 UPF0368 protein C3E7.07c SPBC3E7.07c 0,08 -0,01 0,09 O59730 Probable methionine aminopeptidase 1 SPBC3E7.10 0,37 0,27 0,10 O59734 Small nuclear ribonucleoprotein F smf1 -0,14 0,00 -0,14 O59736 2-isopropylmalate synthase leu3 -0,30 -0,11 -0,19 O59739 BAG family molecular chaperone regulator 1B bag102 0,17 0,23 -0,06 O59743 Uncharacterized protein C530.07c SPBC530.07c -0,02 -0,10 0,08 O59760 Putative uncharacterized hydrolase C1020.07 SPCC1020.07 -0,02 -0,02 0,00 O59770 Probable proteasome subunit alpha type-3 SPCC1795.04c 0,04 0,30 -0,27 O59771 Probable uridylate kinase SPCC1795.05c -0,09 -0,05 -0,05 O59778 Biotin synthase bio2 -0,09 -0,51 0,42 O59779 Uncharacterized protein C1235.01 0,24 -0,21 0,45 O59787 Signal recognition particle subunit srp72 srp72 -0,08 0,05 -0,12 O59791 Serine racemase SRR 1,24 -0,21 1,45 O59794 60S ribosomal protein L17-B rpl17b -0,04 -0,21 0,17 O59797 Putative nucleosome assembly protein C364.06 SPCC364.06 0,02 -0,16 0,19 O59804 10 kDa heat shock protein, mitochondrial hsp10 0,07 0,54 -0,47 O59809 Probable importin c550.11 SPCC550.11 -0,13 -0,12 -0,01 O59810 KH domain-containing protein C550.14 SPCC550.14 0,02 -0,21 0,23 O59811 Zinc finger protein C550.15c SPCC550.15c -0,17 0,16 -0,33 O59813 Uncharacterized amino-acid permease C794.03 SPCC794.03 -0,33 -0,38 0,05
O59816 Dihydrolipoyllysine-residue acetyltransferase component of pyruvate dehydrogenase complex, mitochondrial SPCC794.07 -0,06 -0,11 0,05
O59817 Protein efr3 efr3 0,11 -0,09 0,19 O59823 V-type proton ATPase subunit D vma8 0,08 0,12 -0,04 O59826 Putative voltage-gated potassium channel subunit beta SPCC965.06 0,71 -0,15 0,85 O59827 Glutathione S-transferase 2 gst2 0,15 2,27 -2,11 O59834 Probable cytosine deaminase SPCC965.14c 0,60 0,11 0,49 O59838 Heat shock protein homolog pss1 pss1 0,04 0,34 -0,30 O59855 Probable heat shock protein ssa2 ssa2 0,05 0,33 -0,29 O59858 Glutathione peroxidase gpx1 0,09 0,84 -0,75 O59865 40S ribosomal protein S24-B rps24b -0,03 -0,22 0,19
O59866 Dolichyl-diphosphooligosaccharide--protein glycosyltransferase 48 kDa subunit SPCC338.15 0,02 0,03 -0,01
O59945 Fimbrin fim1 0,14 0,07 0,08 O60059 Pumilio domain-containing protein C56F2.08c SPBC56F2.08c 0,06 0,02 0,04 O60060 Carbamoyl-phosphate synthase arginine-specific small chain arg5 0,04 -0,26 0,30 O60062 Homoserine O-acetyltransferase met6 0,39 -0,01 0,40 O60064 Probable mannose-1-phosphate guanyltransferase SPBC13G1.02 0,06 0,09 -0,04

Table S3. List of all quantified proteins from crude SILAC lysates
146
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O60071 Uncharacterized protein C13G1.09 SPBC13G1.09 -0,20 -0,18 -0,01 O60073 Synaptobrevin homolog ykt6 ykt6 0,17 0,05 0,12 O60079 Probable ubiquitin carboxyl-terminal hydrolase 12 ubp12 0,03 0,05 -0,01 O60080 ATP-dependent RNA helicase dbp9 dbp9 -0,12 -0,04 -0,08
O60084 Mitochondrial import inner membrane translocase subunit tim44 tim44 -0,44 -0,13 -0,31
O60085 Methionine aminopeptidase 2 fma2 -0,09 0,07 -0,16 O60086 Probable acetolactate synthase small subunit SPBC14C8.04 -0,08 -0,02 -0,05 O60094 DNA polymerase V pol5 -0,21 0,13 -0,35 O60096 37S ribosomal protein S35, mitochondrial bot1 -0,21 -0,30 0,09 O60100 Probable importin subunit beta-4 kap123 -0,02 0,02 -0,04 O60101 Phosphoglycerate kinase pgk1 -0,04 0,18 -0,22 O60102 Uncharacterized ABC transporter ATP-binding protein C14F5.06 SPBC14F5.06 0,12 0,18 -0,06 O60105 Adenylosuccinate lyase ade8 -0,05 -0,15 0,10 O60116 Putative ribokinase SPBC16G5.02c 0,05 0,45 -0,39 O60118 54S ribosomal protein L23, mitochondrial mrpl23 -0,20 0,20 -0,40
O60119 Vesicle-associated membrane protein-associated protein C16G5.05c SPBC16G5.05c 0,04 -0,09 0,13
O60121 Uncharacterized protein C16G5.07c SPBC16G5.07c -0,23 0,30 -0,54 O60122 Anthranilate phosphoribosyltransferase trp4 0,07 0,25 -0,18 O60125 BAG family molecular chaperone regulator 1A bag101 0,04 0,26 -0,22 O60128 40S ribosomal protein S3 rps3 -0,01 -0,23 0,22 O60135 Long-chain-fatty-acid--CoA ligase 1 lcf1 0,07 -1,04 1,11 O60143 60S ribosomal protein L7-C rpl7c 0,03 -0,20 0,23 O60144 40S ribosomal protein S16 rps16a 0,02 -0,24 0,26
O60150 Uncharacterized protein C18H10.20c SPBC18H10.20c -0,19 -0,12 -0,07
O60155 Putative proline--tRNA ligase C19C7.06 SPBC19C7.06 0,03 -0,21 0,24 O60158 Bouquet formation protein 4 bqt4 0,01 0,21 -0,20 O60162 Uncharacterized protein C19F5.03 SPBC19F5.03 0,08 0,13 -0,05 O60163 Probable aspartokinase SPBC19F5.04 -0,19 -0,06 -0,12 O60164 Pescadillo homolog ppp1 0,04 0,17 -0,13 O60166 Nuclear movement protein nudc nudc -0,02 -0,07 0,05 O60171 Transcription initiation factor TFIID subunit 10 taf10 0,02 0,27 -0,24 O60179 U3 small nucleolar RNA-associated protein 10 utp10 -0,04 -0,06 0,02 O60180 Probable metalloprotease arx1 arx1 0,13 0,09 0,03 O60181 3,4-dihydroxy-2-butanone 4-phosphate synthase SPBC23E6.06c -0,02 0,44 -0,45 O60184 General transcriptional corepressor ssn6 ssn6 0,11 -0,15 0,26 O60185 Methylthioribose-1-phosphate isomerase mri1 0,09 -0,04 0,12 O60198 S-adenosylmethionine synthase sam1 0,11 -0,06 0,17 O74173 Elongation factor 1-beta tef5 0,00 0,03 -0,04 O74174 V-type proton ATPase subunit G vma10 0,02 -0,06 0,08 O74175 60S ribosomal protein L13 rpl13 -0,01 -0,24 0,23 O74178 Protein kes1 kes1 0,05 -0,08 0,13 O74191 FK506-binding protein 39 kDa fkbp39 -0,11 0,08 -0,20 O74306;P52822 60S ribosomal protein L5-B;60S ribosomal protein L5-A rpl5b;rpl5a 0,00 -0,26 0,26 O74314 Probable cystathionine gamma-synthase SPBC15D4.09c 0,13 0,07 0,06 O74322 40S ribosomal protein S12-B rps12b -0,08 -0,34 0,26 O74323 Signal peptidase complex catalytic subunit sec11 sec11 -0,11 0,36 -0,48 O74329 40S ribosomal protein S29 rps29 0,02 -0,20 0,22 O74330 40S ribosomal protein S27 rps27 0,01 -0,25 0,27 O74341 T-complex protein 1 subunit gamma cct3 -0,03 0,13 -0,16
O74349 E3 ubiquitin-protein ligase listerin SPBC21D10.09c 0,11 0,08 0,03
O74351 Probable cysteine desulfurase, mitochondrial SPBC21D10.11c 0,13 0,10 0,03
O74352 Protein hob1 hob1 0,17 0,13 0,04 O74354 Nucleolar protein dnt1 dnt1 0,34 0,31 0,03 O74374 Probable phosphoglucomutase SPBC32F12.10 -0,05 0,32 -0,37 O74375 Golgi apparatus membrane protein tvp18 tvp18 0,18 0,12 0,06 O74378 2-oxoglutarate dehydrogenase, mitochondrial kgd1 0,12 0,43 -0,31 O74382 Probable phosphoserine phosphatase SPBC3H7.07c -0,08 -0,16 0,08 O74391 60S ribosomal protein L25-B rpl25b 0,01 -0,33 0,34 O74399 Ribosome biogenesis protein erb1 erb1 -0,13 0,06 -0,19 O74400 Uncharacterized RNA-binding protein C4F6.14 SPBC4F6.14 0,09 0,25 -0,16 O74401 ERO1-like protein 1 ero11 0,27 0,80 -0,53 O74402 Heat shock protein 78, mitochondrial hsp78 0,22 1,44 -1,22 O74407 Aspartate--tRNA ligase, cytoplasmic dps1 -0,14 -0,20 0,07 O74409 Uricase SPCC1223.09 1,40 0,08 1,33 O74413 Chorismate synthase -0,16 -0,19 0,03 O74417 Uncharacterized protein C14G10.04 SPCC14G10.04 0,17 0,56 -0,39

Table S3. List of all quantified proteins from crude SILAC lysates
147
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O74419 Uncharacterized protein C162.02c SPCC162.02c -0,07 -0,42 0,34 O74423 Epsin-1 ent1 0,20 -0,15 0,35 O74427 Uridine kinase urk1 0,15 0,09 0,06
O74429 Inositol hexakisphosphate and diphosphoinositol-pentakisphosphate kinase asp1 0,06 0,08 -0,01
O74432 Golgi to ER traffic protein 4 get4 0,06 0,21 -0,15 O74433 Cytochrome b-c1 complex subunit 9 qcr9 0,40 0,10 0,30 O74436 Signal recognition particle subunit srp68 srp68 0,07 0,09 -0,02 O74439 Uncharacterized mitochondrial carrier C1682.09c SPCC1682.09c 0,40 -0,33 0,73 O74440 26S proteasome regulatory subunit rpn8 rpn8 -0,01 0,07 -0,09 O74445 Probable 26S protease subunit rpt4 rpt4 -0,01 0,36 -0,37 O74448 Peptidyl-prolyl cis-trans isomerase pin1 pin1 0,18 0,46 -0,28 O74453 Shk1 kinase-binding protein 15 skb15 -0,10 0,27 -0,37 O74455 Probable 6-phosphogluconolactonase SPCC16C4.10 0,07 0,31 -0,24 O74456 Serine/threonine-protein kinase pef1 pef1 0,14 0,33 -0,19 O74471 Cytochrome c oxidase subunit 6A, mitochondrial cox13 0,23 -0,42 0,66 O74472 Meiotically up-regulated gene 33 protein mug33 0,07 0,15 -0,09 O74475 1,3-beta-glucan synthase component bgs4 bgs4 -0,08 0,13 -0,22 O74476 Importin subunit beta-3 sal3 -0,04 -0,09 0,05 O74478 Phosphoglucomutase-2 SPCC1840.05c 0,00 -0,03 0,03 O74479 ATP synthase subunit 5, mitochondrial atp5 0,06 -0,38 0,45 O74482 Uncharacterized protein C1840.09 SPCC1840.09 0,04 0,17 -0,13 O74484 Mannose-1-phosphate guanyltransferase mpg1 -0,01 0,00 -0,01 O74489 Probable quinone oxidoreductase zta1 0,01 0,72 -0,72 O74499 U6 snRNA-associated Sm-like protein LSm7 lsm7 0,09 0,03 0,06 O74500 Nucleoporin nup60 nup60 -0,23 0,09 -0,31 O74503 Upstream activation factor subunit spp27 spp27 0,01 -0,22 0,23 O74504 UPF0590 protein C594.01 0,08 0,09 -0,01 O74511 Extragenic suppressor of kinetochore protein 1 ekc1 0,03 -0,01 0,04 O74515 Histone chaperone cia1 cia1 0,10 0,22 -0,12 O74519 Uncharacterized N-acetyltransferase C663.13c SPCC663.13c -0,18 -0,30 0,11 O74521 Uncharacterized protein C663.15c SPCC663.15c 0,11 0,58 -0,47 O74524 Probable proline dehydrogenase, mitochondrial SPCC70.03c 0,40 -0,37 0,77 O74533 Cytochrome b-c1 complex subunit 7 qcr7 0,12 -0,28 0,40 O74534 Protein sly1 sly1 -0,07 0,11 -0,18 O74536 SNF1-like protein kinase ssp2 ssp2 0,31 0,35 -0,04 O74538 60S ribosomal protein L27-B rpl27b -0,02 -0,41 0,39 O74545 Putative hydrolase C777.06c SPCC777.06c 0,09 0,37 -0,28 O74548 Probable acetylornithine aminotransferase, mitochondrial arg1 0,16 0,00 0,16 O74549 NEDD8-conjugating enzyme ubc12 ubc12 0,18 0,22 -0,03 O74551 Uncharacterized protein C777.12c SPCC777.12c 0,12 0,31 -0,19 O74555 Branchpoint-bridging protein bpb1 0,15 0,31 -0,16 O74557 NADH-cytochrome b5 reductase 1 cbr1 0,01 -0,03 0,04 O74628 Uncharacterized oxidoreductase C162.03 SPCC162.03 0,18 0,02 0,16 O74653 Protein pob1 pob1 -0,02 -0,02 0,01 O74690 13 kDa ribonucleoprotein-associated protein snu13 0,03 0,15 -0,12 O74718 Eukaryotic peptide chain release factor GTP-binding subunit sup35 -0,04 -0,04 0,00 O74729 Peptidyl-prolyl cis-trans isomerase cyp3 cyp3 -0,15 0,30 -0,45 O74732 3-keto-steroid reductase erg27 -0,43 -0,12 -0,31 O74733 Protein cft1 cft1 -0,31 0,06 -0,37 O74734 Putative ribosomal recycling factor, mitochondrial rrf1 0,04 0,42 -0,38 O74735 Metal homeostasis factor atx1 atx1 0,08 0,19 -0,11 O74742 Probable folylpolyglutamate synthase met7 -0,02 0,06 -0,08 O74743 Eukaryotic translation initiation factor 4E-2 tif452 0,01 0,07 -0,05 O74745 Mannan polymerase II complex anp1 subunit anp1 0,04 -0,06 0,11 O74752 Mitochondrial protein import protein mas5 SPBC1734.11 0,04 0,63 -0,59 O74754 ATP synthase subunit gamma, mitochondrial atp3 0,13 -0,30 0,43 O74760 Eukaryotic translation initiation factor 3 subunit A tif32 0,01 -0,01 0,03 O74762 26S proteasome regulatory subunit rpn2 rpn2 -0,03 0,07 -0,10 O74763 Uncharacterized WD repeat-containing protein C17D11.08 SPBC17D11.08 0,12 0,54 -0,42 O74766 Probable delta-1-pyrroline-5-carboxylate dehydrogenase SPBC24C6.04 0,03 -0,10 0,13 O74767 Probable coatomer subunit epsilon sec28 0,05 0,18 -0,13 O74774 Elongation factor 1 alpha-like protein hbs1 0,17 0,77 -0,60 O74777 KRR1 small subunit processome component homolog mis3 -0,07 -0,33 0,26 O74781 Probable arginine--tRNA ligase, cytoplasmic SPBC25B2.09c -0,05 -0,15 0,10 O74787 Phosphatidylethanolamine N-methyltransferase cho2 0,18 0,11 0,07 O74790 Monothiol glutaredoxin-4 grx4 -0,13 0,22 -0,35 O74791 GTPase grn1 grn1 -0,11 -0,15 0,04 O74793 Uncharacterized protein C26H8.11c SPBC26H8.11c 0,34 0,43 -0,10 O74794 Putative cytochrome c heme lyase SPBC26H8.12 -0,08 0,07 -0,15 O74797 UPF0587 protein C2D10.03c SPBC2D10.03c 0,10 0,20 -0,10 O74799 Glucan 1,3-beta-glucosidase 3 exg3 0,12 0,38 -0,26

Table S3. List of all quantified proteins from crude SILAC lysates
148
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O74802 3-hydroxyisobutyryl-CoA hydrolase, mitochondrial ehd3 0,08 0,17 -0,09 O74803 UV excision repair protein rhp23 rhp23 -0,01 0,06 -0,07 O74814 UPF0400 protein C337.03 SPBC337.03 -0,24 -0,09 -0,15 O74818 Uncharacterized carboxypeptidase C337.07c SPBC337.07c 0,01 0,26 -0,26 O74821 Fe-S cluster assembly protein dre2 dre2 0,04 0,01 0,03 O74828 Pre-rRNA-processing protein esf1 esf1 -0,13 -0,11 -0,02 O74830 Putative glutathione S-transferase C1183.02 SPCC1183.02 -0,06 0,18 -0,24 O74835 rRNA biogenesis protein rrp5 rrp5 -0,06 -0,09 0,04 O74836 60S ribosomal protein L1-B rpl1b -0,01 -0,30 0,29 O74850 Diacylglycerol O-acyltransferase 1 dga1 -0,10 0,05 -0,15 O74861 Set1 complex component sdc1 sdc1 0,07 -0,01 0,08 O74862 rRNA-processing protein utp23 utp23 0,00 0,11 -0,11 O74863 tRNA (guanine-N(7)-)-methyltransferase subunit trm82 trm82 0,10 0,33 -0,24 O74864 60S acidic ribosomal protein P0 rpp0 0,02 -0,26 0,29 O74865 WD repeat-containing protein C18.15 SPCC18.15 0,23 -0,14 0,37 O74866 Riboflavin kinase fmn1 0,34 0,74 -0,40
O74871 Uncharacterized protein C31H12.03c SPCC31H12.03c -0,03 -0,02 -0,01
O74873 Protein transport protein sec23-1 sec231 0,22 0,05 0,17 O74879 Ribosome biogenesis protein enp2 homolog enp2 0,23 0,28 -0,05 O74883 DNA-directed RNA polymerase III subunit rpc5 rpc37 -0,13 0,03 -0,16 O74884;Q92354 60S ribosomal protein L24-B;60S ribosomal protein L24-A rpl24b;rpl24a 0,02 -0,30 0,31 O74887 Peroxiredoxin tpx1 tpx1 0,01 0,05 -0,04 O74891 Probable coatomer subunit zeta ret3 0,06 0,15 -0,10 O74892 40S ribosomal protein S2 rps2 0,00 -0,24 0,24 O74893 40S ribosomal protein S20 rps20 0,01 -0,28 0,29 O74894 26S protease regulatory subunit 6B homolog rpt3 0,02 0,29 -0,26 O74898 Diphthine synthase dph5 -0,21 -0,07 -0,14 O74904 60S ribosomal protein L35 rpl35 0,03 -0,30 0,33 O74905 60S ribosomal protein L9-B rpl9b -0,07 -0,36 0,29 O74907 Protein bcp1 bcp1 -0,12 0,05 -0,17 O74919 RNA-binding protein rnc1 rnc1 0,04 -0,04 0,08 O74927 Methylenetetrahydrofolate reductase 2 met11 0,13 -0,13 0,26
O74928
Bifunctional purine biosynthesis protein ade10;Phosphoribosylaminoimidazolecarboxamide formyltransferase;IMP cyclohydrolase ade10 -0,01 -0,33 0,31
O74940 Putative glutamine-dependent NAD(+) synthetase SPCC553.02 -0,05 0,27 -0,32 O74943 Uncharacterized protein C553.06 SPCC553.06 0,04 0,07 -0,03 O74945 Ribosome assembly protein 1 ria1 0,04 0,00 0,04 O74959 Uncharacterized oxidoreductase C736.13 SPCC736.13 -0,09 0,24 -0,33
O74960 Probable sphingolipid long chain base-responsive protein C736.15 SPCC736.15 0,06 0,43 -0,37
O74965 Eukaryotic translation initiation factor 2A SPBC4B4.04 -0,01 0,02 -0,02 O74970 Pre-mRNA-processing factor 39 prp39 0,23 0,14 0,09 O74975 Probable choline-phosphate cytidylyltransferase SPCC1827.02c 0,04 -0,08 0,12 O74976 Putative peroxisomal-coenzyme A synthetase SPCC1827.03c 0,07 0,59 -0,53 O74978 Uncharacterized RNA-binding protein C1827.05c SPCC1827.05c -0,01 0,01 -0,02 O74982 Caffeine-induced death protein 2 cid2 -0,26 0,05 -0,32 O74983 Ubiquitin-conjugating enzyme spm2 spm2 -0,04 0,18 -0,22 O74985 N-terminal acetyltransferase A complex subunit nat1 nat1 0,00 -0,04 0,04 O74988 Cytochrome c oxidase polypeptide 5, mitochondrial cox5 0,18 -0,37 0,55 O74989 Uncharacterized protein C338.12 SPCC338.12 0,38 0,66 -0,28 O75000 60S ribosomal protein L12 rpl1201 -0,01 -0,22 0,22 O75002 Nuclear import protein mog1 mog1 -0,18 0,13 -0,32 O75005 Valine--tRNA ligase vas2 -0,01 -0,26 0,25 O94236 Eukaryotic translation initiation factor 3 subunit D moe1 0,02 0,04 -0,02 O94238 60S ribosomal protein L14 rpl14 0,00 -0,29 0,29 O94246 Putative glutamate--cysteine ligase regulatory subunit SPCC737.06c -0,06 0,13 -0,18 O94255 Carbonic anhydrase SPBP8B7.05c 0,26 -0,14 0,40 O94258 Exportin-T los1 -0,15 -0,41 0,27 O94260 Putative G3BP-like protein nxt3 -0,06 -0,15 0,08
O94265 Putative hydroxymethylpyrimidine/phosphomethylpyrimidine kinase 1 SPBP8B7.17c -0,26 0,25 -0,50
O94266 Putative hydroxymethylpyrimidine/phosphomethylpyrimidine kinase 2 SPBP8B7.18c 0,04 -0,01 0,05
O94267 FACT complex subunit spt16 spt16 0,00 0,24 -0,24 O94268 Putative ribosomal RNA methyltransferase nop2 SPBP8B7.20c -0,01 0,05 -0,06 O94273 Peptidyl-prolyl cis-trans isomerase B cyp4 -0,04 0,20 -0,24 O94274 Uncharacterized protein P8B7.26 SPBP8B7.26 -0,15 -0,19 0,04 O94280 UPF0674 endoplasmic reticulum membrane protein C2G5.01 SPBC2G5.01 0,37 0,22 0,14 O94284 Sulfide:quinone oxidoreductase, mitochondrial hmt2 0,01 -0,01 0,02

Table S3. List of all quantified proteins from crude SILAC lysates
149
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O94286 1,2-dihydroxy-3-keto-5-methylthiopentene dioxygenase adi1 -0,05 -0,21 0,16 O94289 Ubiquitin homeostasis protein lub1 lub1 -0,03 0,35 -0,38
O94303 Imidazole glycerol phosphate synthase hisHF;Glutamine amidotransferase;Cyclase his4 -0,30 -0,16 -0,14
O94305 Putative monoglyceride lipase SPCC5E4.05c 0,02 0,33 -0,30 O94311 UPF0743 protein C215.06c SPBC215.06c -0,23 0,05 -0,28 O94312 PWWP domain-containing protein2 pdp2 0,05 0,26 -0,20 O94313 Carbamoyl-phosphate synthase arginine-specific large chain arg4 0,05 -0,21 0,26 O94314 Uncharacterized hydrolase C215.10 SPBC215.10 -0,05 0,04 -0,09 O94315 Uncharacterized oxidoreductase C215.11c SPBC215.11c 0,07 0,44 -0,37 O94316 Pre-mRNA-splicing factor cwf10 cwf10 0,06 0,25 -0,19 O94319 Protein transport protein sec13 sec13 0,05 0,15 -0,10 O94320 Aspartate aminotransferase, mitochondrial SPBC725.01 0,10 -0,14 0,23 O94326 Zinc finger protein C725.08 SPBC725.08 0,08 0,20 -0,12 O94331 Orotate phosphoribosyltransferase ura5 0,08 -0,05 0,12 O94337 Probable deoxyhypusine synthase SPBC1271.04c 0,03 0,14 -0,11 O94340 Uncharacterized N-acetyltransferase C1271.07c SPBC1271.07c -0,31 -0,35 0,04
O94346
Arginine biosynthesis bifunctional protein ArgJ, mitochondrial;Glutamate N-acetyltransferase;Amino-acid acetyltransferase SPBC1271.14 0,42 -0,04 0,46
O94347 ATP-dependent (S)-NAD(P)H-hydrate dehydratase SPCC61.03 0,02 0,86 -0,84 O94350 Cystathionine beta-lyase str3 0,11 -0,01 0,12 O94354 Argininosuccinate synthase arg12 0,15 0,00 0,15 O94362 Uncharacterized GTP-binding protein C428.15 SPBC428.15 -0,03 0,05 -0,07 O94365 U3 small nucleolar RNA-associated protein 15 utp15 -0,14 -0,01 -0,13 O94370 Uncharacterized mitochondrial carrier C1604.04 SPBC1604.04 -0,12 0,04 -0,15 O94373 ATP synthase subunit 4, mitochondrial atp4 0,13 -0,35 0,48 O94374 Importin subunit alpha-2 imp1 0,05 0,40 -0,35 O94377 ATP synthase subunit f, mitochondrial atp17 -0,28 -0,61 0,32 O94378 Uncharacterized protein C1604.12 SPBC1604.12 0,07 0,42 -0,36 O94390 ATP synthase subunit d, mitochondrial atp7 0,07 -0,34 0,41
O94391 Probable cytochrome b5 1 SPBC29A10.16c -0,04 -0,43 0,39
O94396 tRNA pseudouridine synthase 1 pus1 -0,08 -0,18 0,10 O94399 Twinfilin twf1 -0,03 0,22 -0,26 O94401 L-type lectin-like domain-containing protein C126.08c SPCC126.08c 0,39 0,30 0,09 O94403 Uncharacterized RNA-binding protein C126.11c SPCC126.11c 0,04 -0,11 0,14 O94404 Protein NIF3 homolog SPCC126.12 0,21 0,23 -0,02 O94407 Signal recognition particle sec65 subunit sec65 -0,03 -0,05 0,02 O94413 Ribose-phosphate pyrophosphokinase 2 SPCC1620.06c 0,10 0,06 0,04
O94415 Probable succinyl-CoA ligase [GDP-forming] subunit beta, mitochondrial SPCC1620.08 0,02 -0,06 0,08
O94416 Transcription initiation factor IIF subunit alpha tfg1 -0,05 0,46 -0,51 O94424 Transcription initiation factor IIF subunit beta tfg2 -0,09 0,34 -0,43 O94431 Uncharacterized aminotransferase C660.12c SPBC660.12c 0,23 0,26 -0,03 O94432 Uncharacterized RNA-binding protein C660.15 SPBC660.15 -0,32 -0,52 0,20 O94435 Meiotically up-regulated gene 35 protein mug35 0,08 0,63 -0,55 O94436 Transcription initiation factor TFIID subunit 14 tfg3 0,20 0,30 -0,10 O94438 40S ribosomal protein S1-B rps102 -0,02 -0,26 0,25 O94442 Cytosolic Fe-S cluster assembly factor nbp35 nbp35 0,26 0,36 -0,10 O94444 26S proteasome regulatory subunit rpn10 rpn10 -0,03 0,17 -0,19 O94447 Uncharacterized PH domain-containing protein C637.13c SPAC637.13c -0,12 -0,09 -0,02 O94460 Probable queuine tRNA-ribosyltransferase SPAC1687.19c 0,01 1,32 -1,30 O94461 Probable phosphoglycerate mutase C1687.21 0,01 0,14 -0,13 O94465 Protein cms1 cms1 -0,11 0,14 -0,25 O94466 Probable Rho-GTPase-activating protein 7 rga7 0,19 0,40 -0,21 O94476 Eukaryotic translation initiation factor 6 tif6 -0,05 0,16 -0,21 O94489 Elongation factor 3 tef3 -0,03 -0,05 0,02 O94497 Uncharacterized serine-rich protein C18E5.07 SPBC18E5.07 0,06 0,32 -0,26 O94501 T-complex protein 1 subunit alpha cct1 -0,01 0,05 -0,06 O94504 Thioredoxin-2, mitochondrial trx2 -0,01 0,65 -0,66 O94505 3(2),5-bisphosphate nucleotidase tol1 0,05 0,08 -0,03 O94511 Putative enoyl reductase C646.07c SPBC646.07c 0,11 -0,37 0,48 O94512 Oxysterol-binding protein-like protein 1 obp1 0,09 0,27 -0,18 O94513 Eukaryotic translation initiation factor 3 subunit E int6 -0,05 0,00 -0,05 O94514 Nucleolar protein 56 nop56 -0,02 -0,01 -0,01 O94515 T-complex protein 1 subunit zeta cct6 -0,02 0,03 -0,05 O94517 Probable proteasome subunit alpha type-6 SPBC646.16 0,07 0,29 -0,21 O94523 Probable acyl-CoA desaturase SPCC1281.06c 0,15 -0,17 0,32 O94529 FACT complex subunit pob3 pob3 0,16 0,19 -0,02 O94530 tRNA threonylcarbamoyladenosine biosynthesis protein sua5 sua5 -0,07 0,09 -0,17 O94544 Leukotriene A-4 hydrolase homolog SPCC1322.05c -0,01 0,21 -0,21
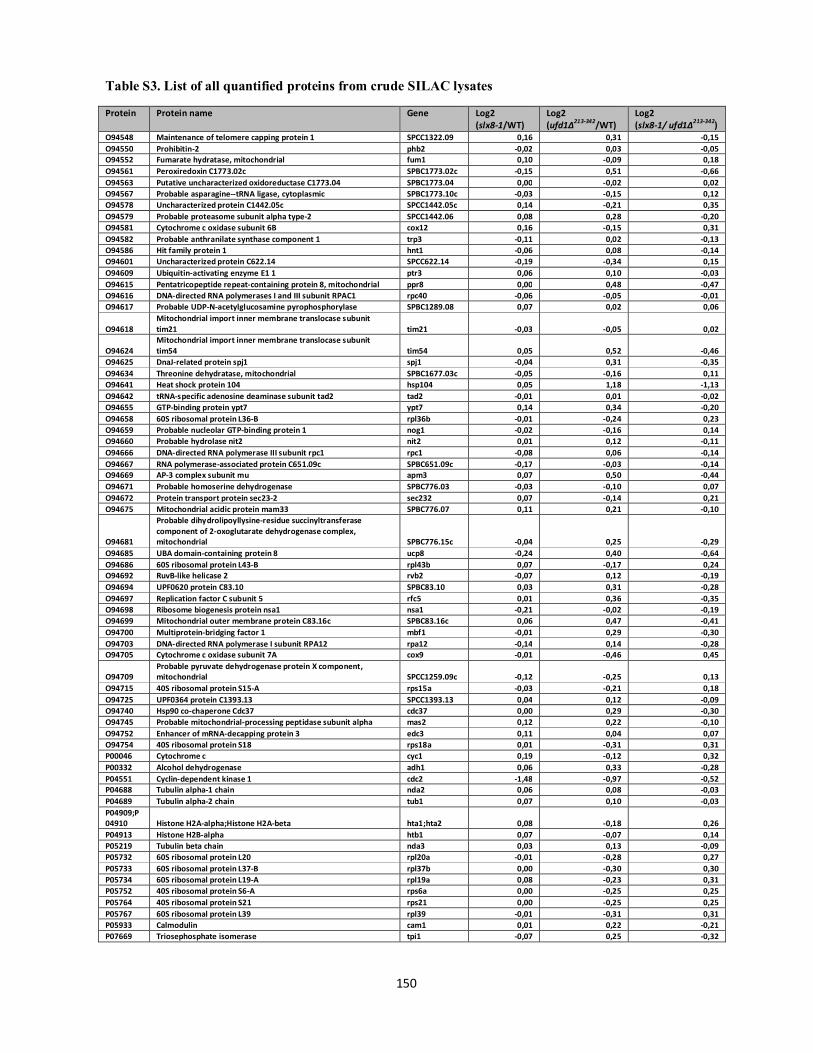
Table S3. List of all quantified proteins from crude SILAC lysates
150
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
O94548 Maintenance of telomere capping protein 1 SPCC1322.09 0,16 0,31 -0,15 O94550 Prohibitin-2 phb2 -0,02 0,03 -0,05 O94552 Fumarate hydratase, mitochondrial fum1 0,10 -0,09 0,18 O94561 Peroxiredoxin C1773.02c SPBC1773.02c -0,15 0,51 -0,66 O94563 Putative uncharacterized oxidoreductase C1773.04 SPBC1773.04 0,00 -0,02 0,02 O94567 Probable asparagine--tRNA ligase, cytoplasmic SPBC1773.10c -0,03 -0,15 0,12 O94578 Uncharacterized protein C1442.05c SPCC1442.05c 0,14 -0,21 0,35 O94579 Probable proteasome subunit alpha type-2 SPCC1442.06 0,08 0,28 -0,20 O94581 Cytochrome c oxidase subunit 6B cox12 0,16 -0,15 0,31 O94582 Probable anthranilate synthase component 1 trp3 -0,11 0,02 -0,13 O94586 Hit family protein 1 hnt1 -0,06 0,08 -0,14 O94601 Uncharacterized protein C622.14 SPCC622.14 -0,19 -0,34 0,15 O94609 Ubiquitin-activating enzyme E1 1 ptr3 0,06 0,10 -0,03 O94615 Pentatricopeptide repeat-containing protein 8, mitochondrial ppr8 0,00 0,48 -0,47 O94616 DNA-directed RNA polymerases I and III subunit RPAC1 rpc40 -0,06 -0,05 -0,01 O94617 Probable UDP-N-acetylglucosamine pyrophosphorylase SPBC1289.08 0,07 0,02 0,06
O94618 Mitochondrial import inner membrane translocase subunit tim21 tim21 -0,03 -0,05 0,02
O94624 Mitochondrial import inner membrane translocase subunit tim54 tim54 0,05 0,52 -0,46
O94625 DnaJ-related protein spj1 spj1 -0,04 0,31 -0,35 O94634 Threonine dehydratase, mitochondrial SPBC1677.03c -0,05 -0,16 0,11 O94641 Heat shock protein 104 hsp104 0,05 1,18 -1,13 O94642 tRNA-specific adenosine deaminase subunit tad2 tad2 -0,01 0,01 -0,02 O94655 GTP-binding protein ypt7 ypt7 0,14 0,34 -0,20 O94658 60S ribosomal protein L36-B rpl36b -0,01 -0,24 0,23 O94659 Probable nucleolar GTP-binding protein 1 nog1 -0,02 -0,16 0,14 O94660 Probable hydrolase nit2 nit2 0,01 0,12 -0,11 O94666 DNA-directed RNA polymerase III subunit rpc1 rpc1 -0,08 0,06 -0,14 O94667 RNA polymerase-associated protein C651.09c SPBC651.09c -0,17 -0,03 -0,14 O94669 AP-3 complex subunit mu apm3 0,07 0,50 -0,44 O94671 Probable homoserine dehydrogenase SPBC776.03 -0,03 -0,10 0,07 O94672 Protein transport protein sec23-2 sec232 0,07 -0,14 0,21 O94675 Mitochondrial acidic protein mam33 SPBC776.07 0,11 0,21 -0,10
O94681
Probable dihydrolipoyllysine-residue succinyltransferase component of 2-oxoglutarate dehydrogenase complex, mitochondrial SPBC776.15c -0,04 0,25 -0,29
O94685 UBA domain-containing protein 8 ucp8 -0,24 0,40 -0,64 O94686 60S ribosomal protein L43-B rpl43b 0,07 -0,17 0,24 O94692 RuvB-like helicase 2 rvb2 -0,07 0,12 -0,19 O94694 UPF0620 protein C83.10 SPBC83.10 0,03 0,31 -0,28 O94697 Replication factor C subunit 5 rfc5 0,01 0,36 -0,35 O94698 Ribosome biogenesis protein nsa1 nsa1 -0,21 -0,02 -0,19 O94699 Mitochondrial outer membrane protein C83.16c SPBC83.16c 0,06 0,47 -0,41 O94700 Multiprotein-bridging factor 1 mbf1 -0,01 0,29 -0,30 O94703 DNA-directed RNA polymerase I subunit RPA12 rpa12 -0,14 0,14 -0,28 O94705 Cytochrome c oxidase subunit 7A cox9 -0,01 -0,46 0,45
O94709 Probable pyruvate dehydrogenase protein X component, mitochondrial SPCC1259.09c -0,12 -0,25 0,13
O94715 40S ribosomal protein S15-A rps15a -0,03 -0,21 0,18 O94725 UPF0364 protein C1393.13 SPCC1393.13 0,04 0,12 -0,09 O94740 Hsp90 co-chaperone Cdc37 cdc37 0,00 0,29 -0,30 O94745 Probable mitochondrial-processing peptidase subunit alpha mas2 0,12 0,22 -0,10 O94752 Enhancer of mRNA-decapping protein 3 edc3 0,11 0,04 0,07 O94754 40S ribosomal protein S18 rps18a 0,01 -0,31 0,31 P00046 Cytochrome c cyc1 0,19 -0,12 0,32 P00332 Alcohol dehydrogenase adh1 0,06 0,33 -0,28 P04551 Cyclin-dependent kinase 1 cdc2 -1,48 -0,97 -0,52 P04688 Tubulin alpha-1 chain nda2 0,06 0,08 -0,03 P04689 Tubulin alpha-2 chain tub1 0,07 0,10 -0,03 P04909;P04910 Histone H2A-alpha;Histone H2A-beta hta1;hta2 0,08 -0,18 0,26 P04913 Histone H2B-alpha htb1 0,07 -0,07 0,14 P05219 Tubulin beta chain nda3 0,03 0,13 -0,09 P05732 60S ribosomal protein L20 rpl20a -0,01 -0,28 0,27 P05733 60S ribosomal protein L37-B rpl37b 0,00 -0,30 0,30 P05734 60S ribosomal protein L19-A rpl19a 0,08 -0,23 0,31 P05752 40S ribosomal protein S6-A rps6a 0,00 -0,25 0,25 P05764 40S ribosomal protein S21 rps21 0,00 -0,25 0,25 P05767 60S ribosomal protein L39 rpl39 -0,01 -0,31 0,31 P05933 Calmodulin cam1 0,01 0,22 -0,21 P07669 Triosephosphate isomerase tpi1 -0,07 0,25 -0,32

Table S3. List of all quantified proteins from crude SILAC lysates
151
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
P08091 Acid phosphatase pho1 0,17 0,42 -0,25 P08093 60S ribosomal protein L2 rpl2a 0,00 -0,28 0,27 P08094 60S acidic ribosomal protein P2-alpha rpp201 -0,06 -0,35 0,30 P08096 DNA topoisomerase 2 top2 -0,04 0,19 -0,23 P08647 Ras-like protein 1 ras1 0,08 0,18 -0,10 P09322 Histone H4 hhf1 0,06 -0,21 0,27 P09627 Plasma membrane ATPase 1 pma1 0,10 -0,10 0,20 P09988;P10651 Histone H3.1/H3.2;Histone H3.3 hht1;hht3 0,06 -0,19 0,25
P0C016;P0C8R3
Ubiquitin-40S ribosomal protein S27a;Ubiquitin;40S ribosomal protein S27a;Ubiquitin-40S ribosomal protein S27b;Ubiquitin;40S ribosomal protein S27b ubi3;ubi5 0,06 0,59 -0,52
P10989 Actin act1 0,06 0,22 -0,16 P11620 GTP-binding protein ypt1 ypt1 0,12 0,05 0,08 P13681 Serine/threonine-protein phosphatase PP1-1 dis2 -0,05 0,33 -0,38 P14068 Exportin-1 xpo1 0,07 0,21 -0,14 P14965 Orotidine 5-phosphate decarboxylase ura4 -0,05 0,01 -0,06 P15398 DNA-directed RNA polymerase I subunit rpa1 rpa1 -0,05 0,03 -0,09 P15567 Phosphoribosylaminoimidazole carboxylase ade6 0,04 -0,07 0,10 P17476 60S acidic ribosomal protein P1-alpha 1 rpa1 -0,10 -0,29 0,19 P17477 60S acidic ribosomal protein P1-alpha 3 rpa3 -0,06 -0,22 0,16 P17478 60S acidic ribosomal protein P2-beta rpp202 -0,42 -0,26 -0,16 P17608 GTP-binding protein ryh1 ryh1 0,10 -0,10 0,20 P17609 GTP-binding protein ypt2 ypt2 0,07 0,09 -0,02 P17610 GTP-binding protein ypt3 ypt3 0,06 -0,20 0,26 P17937 60S ribosomal protein L7-A rpl7a 0,07 0,02 0,05 P18253 Peptidyl-prolyl cis-trans isomerase ppi1 0,05 0,27 -0,22 P18869 3-isopropylmalate dehydrogenase leu1 -0,09 -0,11 0,03 P19117 Inorganic pyrophosphatase ppa1 0,02 0,03 -0,01
P20772
Bifunctional purine biosynthetic protein ADE1;Phosphoribosylamine--glycine ligase;Phosphoribosylformylglycinamidine cyclo-ligase ade1 -0,03 -0,37 0,34
P21565 Signal recognition particle 54 kDa protein homolog srp54 -0,06 0,06 -0,12 P21696 Glycerol-3-phosphate dehydrogenase [NAD(+)] 1 gpd1 -0,15 0,71 -0,86
P21911 Succinate dehydrogenase [ubiquinone] iron-sulfur subunit, mitochondrial sdh2 0,23 0,24 -0,01
P22068 ATP synthase subunit beta, mitochondrial atp2 0,04 -0,19 0,24 P22192 Double-strand-specific pac1 ribonuclease pac1 -0,06 0,50 -0,56 P22774;P22774-2 Heat shock 70 kDa protein, mitochondrial ssp1 -0,06 0,35 -0,40 P23566 Ubiquitin-conjugating enzyme E2 2 rhp6 -0,03 0,28 -0,31 P24487 ATP synthase subunit alpha, mitochondrial atp1 0,05 -0,26 0,31 P24782 ATP-dependent RNA helicase dbp2 dbp2 0,05 -0,03 0,08 P25457 60S ribosomal protein L7-B rpl7b -0,02 -0,50 0,47 P28189 40S ribosomal protein S13 rps13 0,01 -0,29 0,30 P28745 Protein pim1 pim1 0,12 0,26 -0,14 P28748 GTP-binding nuclear protein spi1 spi1 -0,04 0,21 -0,25 P28758 Superoxide dismutase [Cu-Zn] sod1 0,13 -0,13 0,27 P29458 DNA replication licensing factor mcm4 mcm4 -0,14 -0,02 -0,13 P30316 DNA polymerase delta catalytic subunit pol3 0,56 1,05 -0,49 P30655 Proteasome subunit beta type-5 pts1 0,13 0,33 -0,20 P30666 DNA replication licensing factor mcm3 mcm3 -0,02 0,29 -0,31 P30821 P25 protein obr1 -0,45 1,74 -2,19 P31209 Polyadenylate-binding protein, cytoplasmic and nuclear pab1 -0,01 -0,08 0,06 P31317 Ornithine carbamoyltransferase, mitochondrial arg3 0,21 0,20 0,02
P31318 Protein arg11, mitochondrial;N-acetyl-gamma-glutamyl-phosphate reductase;Acetylglutamate kinase arg11 0,19 0,02 0,17
P31406 V-type proton ATPase catalytic subunit A vma1 0,07 -0,06 0,13 P31411 V-type proton ATPase subunit B vma2 0,00 0,08 -0,08 P32235 GTP-binding protein 1 gtp1 0,03 0,09 -0,07 P32390 Actin-related protein 3 act2 0,02 0,11 -0,09 P32747 Dihydroorotate dehydrogenase (quinone), mitochondrial ura3 -0,10 0,13 -0,24 P33075 Adenylate kinase 1 adk1 0,01 -0,17 0,18 P33532 Vitamin B6 transporter bsu1 bsu1 -0,34 -0,54 0,20 P35551 rRNA 2-O-methyltransferase fibrillarin fib1 0,00 -0,02 0,02 P35669 Glutathione synthetase large chain gsa1 -0,11 0,20 -0,30 P35679 60S ribosomal protein L4-A rpl4a 0,00 -0,33 0,33 P36579 ADP-ribosylation factor 1 arf1 0,08 0,06 0,02 P36580 Fructose-bisphosphate aldolase fba1 -0,06 0,05 -0,11 P36581 Calnexin homolog cal1 0,08 0,42 -0,34 P36582 Protein kinase C-like 1 pck1 0,04 0,37 -0,33

Table S3. List of all quantified proteins from crude SILAC lysates
152
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
P36584;P40372 60S ribosomal protein L3-B;60S ribosomal protein L3-A rpl3b;rpl3a 0,00 -0,24 0,24 P36585 60S ribosomal protein L28-A rpl28a 0,03 -0,31 0,34 P36586 GTP-binding protein ypt5 ypt5 0,12 0,00 0,11 P36587 NADPH--cytochrome P450 reductase ccr1 0,01 0,19 -0,18 P36591 Dihydrofolate reductase dfr1 0,16 0,07 0,09 P36594 DNA-directed RNA polymerase II subunit rpb1 rpb1 -0,04 0,32 -0,36 P36595 DNA-directed RNA polymerases I, II, and III subunit RPABC2 rpb6 -0,04 0,03 -0,07 P36596 Squalene synthase erg9 -0,01 -0,01 0,01 P36597 Pyrimidine precursor biosynthesis enzyme thi3 thi3 0,21 -0,34 0,55 P36602 Ribonucleoside-diphosphate reductase large chain cdc22 -0,02 0,19 -0,21 P36603 Ribonucleoside-diphosphate reductase small chain suc22 0,07 0,10 -0,03 P36604 78 kDa glucose-regulated protein homolog bip1 0,07 0,97 -0,90 P36605 Histidinol-phosphate aminotransferase his3 -0,04 -0,03 -0,01 P36612 26S protease regulatory subunit 4 homolog mts2 0,06 0,34 -0,28 P36614 Serine/threonine-protein phosphatase ppe1 ppe1 0,04 0,01 0,03 P36620 Acetolactate synthase, mitochondrial ilv1 -0,13 -0,12 0,00 P36621 Adenylyl cyclase-associated protein cap1 0,05 0,29 -0,24 P36623 Phosphoglycerate mutase gpm1 -0,10 0,10 -0,20 P36627 Cellular nucleic acid-binding protein homolog byr3 0,03 -0,38 0,41 P36629 Splicing factor U2AF 59 kDa subunit prp2 -0,02 0,11 -0,12 P37202 Exosome complex exonuclease dis3 dis3 0,15 0,19 -0,04 P39825 Profilin cdc3 0,07 0,15 -0,08 P40231 Casein kinase II subunit alpha cka1 -0,13 -0,05 -0,07 P40232 Casein kinase II subunit beta ckb1 -0,09 -0,09 0,00 P40233 Casein kinase I homolog 1 cki1 0,83 0,49 0,33 P40235 Casein kinase I homolog hhp1 hhp1 0,17 0,16 0,01 P40370 Enolase 1-1 eno101 -0,03 0,09 -0,13 P40373 ATP phosphoribosyltransferase his1 0,01 -0,32 0,33 P40374 Imidazoleglycerol-phosphate dehydratase his5 0,08 -0,05 0,13 P40375 NAD-dependent malic enzyme mae2 0,06 0,30 -0,24 P40383 5-3 exoribonuclease 1 exo2 0,04 -0,10 0,14
P40386 Probable thiamine biosynthetic bifunctional enzyme;Thiamine-phosphate synthase;Hydroxyethylthiazole kinase thi4 -0,01 0,04 -0,05
P40387 Alpha,alpha-trehalose-phosphate synthase [UDP-forming] tps1 0,07 1,07 -1,01 P40388 UV-induced protein uvi15 uvi15 -0,03 -0,64 0,60 P40847 Switch-activating protein 1 sap1 0,03 -0,10 0,13 P40848 5-3 exoribonuclease 2 dhp1 0,16 0,28 -0,12
P40903 Sexual differentiation process putative subtilase-type proteinase isp6 isp6 0,89 0,63 0,25
P40921 Elongation factor 1-gamma tef3 0,00 0,11 -0,11 P40976 L-aminoadipate-semialdehyde dehydrogenase lys1 -0,15 -0,33 0,18 P40998 Thiamine thiazole synthase thi2 -0,04 -0,10 0,06 P41003 Structural maintenance of chromosomes protein 2 cut14 -0,05 -0,09 0,05 P41390 Amidophosphoribosyltransferase ade4 0,12 0,02 0,10 P41391 Ran GTPase-activating protein 1 rna1 -0,08 0,10 -0,17 P41820 Brefeldin A resistance protein bfr1 -0,33 0,05 -0,38 P41831 Ribose-phosphate pyrophosphokinase 1 SPAC4A8.14 0,01 0,08 -0,07 P41836 26S protease regulatory subunit 8 homolog let1 -0,04 0,18 -0,22 P41838 Poly(A)+ RNA export protein rae1 0,12 0,17 -0,05 P41878 26S proteasome regulatory subunit rpn11 rpn11 0,12 0,28 -0,17 P41887 Heat shock protein 90 homolog swo1 0,06 0,32 -0,25 P41891 Protein gar2 gar2 0,00 -0,11 0,10 P41893 Low molecular weight phosphotyrosine protein phosphatase stp1 -0,02 0,19 -0,20 P42656 DNA damage checkpoint protein rad24 rad24 -0,04 0,19 -0,23 P42657 DNA damage checkpoint protein rad25 rad25 0,10 0,34 -0,24 P46595 Ubiquitin-conjugating enzyme E2 4 ubc4 0,05 0,44 -0,39 P47943 ATP-dependent RNA helicase eIF4A tif1 0,02 -0,01 0,04 P48003 Histone H2A.Z pht1 -0,01 -0,09 0,09 P48008 Septin homolog spn3 spn3 0,89 0,43 0,46 P48009 Septin homolog spn4 spn4 0,10 0,20 -0,10 P49373 Transcription elongation factor S-II tfs1 0,03 -0,13 0,16 P49740 Nucleoside diphosphate kinase ndk1 0,18 0,55 -0,38 P49776 Bis(5-nucleosyl)-tetraphosphatase [asymmetrical] aph1 0,13 0,37 -0,24 P50514 Probable argininosuccinate lyase argx 0,18 -0,34 0,52 P50519 Heat shock protein hsp9 hsp9 0,10 1,53 -1,43 P50521 Hexokinase-2 hxk2 -0,12 0,22 -0,35 P50523 Cytochrome b-c1 complex subunit 8 qcr8 0,23 -0,30 0,53 P50524 26S proteasome regulatory subunit rpn12 rpn12 -0,05 0,10 -0,15 P50998 AMP deaminase ada1 0,07 0,12 -0,05 P50999 T-complex protein 1 subunit delta cct4 -0,01 0,05 -0,06 P52808 60S ribosomal protein L30-1 rpl30a 0,04 -0,28 0,31

Table S3. List of all quantified proteins from crude SILAC lysates
153
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
P53694 Reticulon-like protein 1 rtn1 0,20 0,41 -0,21 P54069 Protein bem46 bem46 -0,03 -0,11 0,08 P54874 Hydroxymethylglutaryl-CoA synthase hcs1 0,07 -0,08 0,15 P55306 Catalase cta1 0,51 1,28 -0,77 P55877 Eukaryotic translation initiation factor 1A tif11 0,06 0,17 -0,11 P56286 Eukaryotic translation initiation factor 2 subunit alpha tif211 -0,01 0,11 -0,11 P56287 Probable translation initiation factor eIF-2B subunit epsilon tif225 0,19 0,41 -0,22 P56288 Probable translation initiation factor eIF-2B subunit gamma tif223 -0,02 0,08 -0,09 P56289 Eukaryotic translation initiation factor 5A-1 tif51a 0,00 0,00 -0,01 P56329 Probable eukaryotic translation initiation factor 2 subunit beta tif212 -0,04 -0,06 0,02 P57728 60S ribosomal protein L28-B rpl28b 0,02 -0,11 0,13 P58234 40S ribosomal protein S19-A rps19a 0,00 -0,15 0,15 P62506 Probable 54S ribosomal protein L44, mitochondrial SPBC56F2.14 0,19 -0,46 0,65 P78750 Probable U3 small nucleolar RNA-associated protein 18 SPBC29A3.06 -0,13 0,21 -0,34 P78753 Probable asparagine synthetase [glutamine-hydrolyzing] asn1 -0,16 0,07 -0,23 P78761 Cytochrome b-c1 complex subunit 2, mitochondrial qcr2 0,00 -0,37 0,38 P78763 Uncharacterized mitochondrial carrier C83.13 SPBC83.13 0,29 -0,47 0,77 P78773 Probable acetyl-coenzyme A synthetase 0,05 -0,23 0,28 P78774 Actin-related protein 2/3 complex subunit 1 arc1 0,09 0,23 -0,14 P78780 Probable aspartate-semialdehyde dehydrogenase SPCC1827.06c -0,33 0,00 -0,33 P78789 Uncharacterized protein C1919.03c SPCC1919.03c 0,29 0,18 0,12
P78790 Probable electron transfer flavoprotein subunit alpha, mitochondrial SPAC27D7.06 0,00 0,10 -0,10
P78795 Eukaryotic translation initiation factor 3 subunit G tif35 -0,04 0,04 -0,09 P78804 NADP-specific glutamate dehydrogenase gdh1 0,06 -0,01 0,07 P78810 Vacuolar transporter chaperone 4 vtc4 0,18 0,03 0,14 P78811 Probable UTP--glucose-1-phosphate uridylyltransferase SPCC1322.04 0,05 0,36 -0,31 P78812 6-phosphogluconate dehydrogenase, decarboxylating SPBC660.16 -0,08 -0,02 -0,06 P78813 ENTH domain-containing protein C794.11c SPCC794.11c 0,06 0,01 0,05 P78814 Pre-mRNA-splicing factor srp2 srp2 0,11 -0,05 0,16 P78820 Acetyl-CoA carboxylase;Biotin carboxylase cut6 0,16 -0,80 0,96 P78825 Adenosine kinase ado1 0,01 -0,14 0,14 P78827 Probable ketol-acid reductoisomerase, mitochondrial ilv5 -0,09 -0,19 0,09 P78831 High-affinity glucose transporter ght5 ght5 1,23 1,44 -0,21
P78833 Uncharacterized protein C21B10.08c SPBC21B10.08c 0,04 0,49 -0,46
P78854 Lysophospholipase 1 plb1 0,10 0,26 -0,15 P78870 Probable S-(hydroxymethyl)glutathione dehydrogenase 1 SPBC1539.07c 0,16 -0,09 0,25 P78875 Trehalose-phosphatase tpp1 -0,05 0,51 -0,56 P78917 Glucose-6-phosphate isomerase pgi1 -0,02 0,15 -0,17 P78920 Putative nucleosome assembly protein C2D10.11C SPBC2D10.11c 0,00 0,20 -0,20 P78921 Probable T-complex protein 1 subunit theta cct8 -0,03 0,05 -0,08 P78929 Cofilin cof1 0,08 0,08 0,00 P78937 Sulfate adenylyltransferase sua1 0,08 -0,06 0,13 P78946 60S ribosomal protein L26 rpl26 0,01 -0,27 0,28 P78954 Eukaryotic translation initiation factor 4E-1 tif451 0,02 0,06 -0,03 P78958 Glyceraldehyde-3-phosphate dehydrogenase 1 tdh1 -0,13 -0,07 -0,05 P78963 Protein arginine N-methyltransferase skb1 skb1 -0,08 0,04 -0,12 P78965 Glutathione reductase pgr1 -0,06 0,01 -0,07 P78974 Delta-aminolevulinic acid dehydratase hem2 -0,15 0,08 -0,24 P79009 40S ribosomal protein S25-A rps25a 0,00 -0,21 0,21 P79010 Cytochrome c oxidase subunit 4, mitochondrial cox4 0,03 -0,32 0,34 P79013 40S ribosomal protein S11 rps11a -0,01 -0,24 0,23 P79015 60S ribosomal protein L32-A rpl32a 0,08 -0,32 0,40 P79016 40S ribosomal protein S19-B rps19b -0,05 -0,11 0,06 P79057 40S ribosomal protein S23 rps23a -0,02 -0,24 0,23 P79058 Uncharacterized protein C10F6.16 SPAC10F6.16 -0,04 0,38 -0,42 P79060 Protein translation factor sui1 sui1 -0,07 -0,09 0,01 P79063 Eukaryotic peptide chain release factor subunit 1 sup45 0,04 0,00 0,05 P79065 Tip elongation protein 1 tip1 0,01 0,36 -0,35 P79071 60S ribosomal protein L6 rpl6 0,02 -0,26 0,28 P79083 Eukaryotic translation initiation factor 3 subunit I tif34 -0,02 0,05 -0,07 P79088 Protein transport protein sec61 subunit alpha sec61 0,01 -0,18 0,19 P87048 26S proteasome regulatory subunit rpn1 rpn1 0,03 0,20 -0,17 P87057 Non-histone chromosomal protein 6 nhp6 0,23 0,28 -0,05 P87058 La protein homolog sla1 -0,10 0,09 -0,19
P87059 Mitochondrial intermembrane space import and assembly protein 40 mia40 -0,32 0,60 -0,92
P87111 Probable electron transfer flavoprotein-ubiquinone oxidoreductase, mitochondrial SPAC20G8.04c 0,21 0,42 -0,21
P87112 General negative regulator of transcription subunit 1 not1 0,03 0,15 -0,13 P87115 UPF0202 protein C20G8.09c SPAC20G8.09c -0,02 0,09 -0,11

Table S3. List of all quantified proteins from crude SILAC lysates
154
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
P87128 Probable eukaryotic translation initiation factor 3 subunit J SPAC3A12.13c -0,14 -0,06 -0,08 P87140 Probable coatomer subunit gamma sec21 0,02 0,16 -0,15 P87142 Heat shock protein 70 homolog C57A7.12 SPAC57A7.12 -0,02 0,04 -0,06 P87144 Threonine--tRNA ligase, cytoplasmic ths1 -0,04 -0,25 0,21 P87147 Nascent polypeptide-associated complex subunit alpha egd2 0,01 -0,18 0,19 P87153 Probable T-complex protein 1 subunit eta cct7 -0,06 0,04 -0,10 P87157 Probable ADP-ribosylation factor-binding protein C25H2.16c SPBC25H2.16c -0,01 -0,01 0,00 P87158 40S ribosomal protein S4-A rps4a 0,02 -0,35 0,36 P87167 Uba3-binding protein but2 but2 0,18 0,05 0,14 P87171 Ribose-phosphate pyrophosphokinase 5 SPBC3D6.06c -0,02 -0,06 0,04 P87177 Uncharacterized WD repeat-containing protein C3D6.12 SPBC3D6.12 -0,24 -0,10 -0,14 P87178 Uncharacterized protein C3D6.13c SPBC3D6.13c 0,00 -0,21 0,21 P87216 Protein vip1 vip1 0,06 0,51 -0,45 P87228 Putative D-3-phosphoglycerate dehydrogenase -0,42 -0,03 -0,39
P87230 2-methoxy-6-polyprenyl-1,4-benzoquinol methylase, mitochondrial coq5 -0,09 0,00 -0,09
P87238 CUE domain-containing protein 4, mitochondrial SPCC4G3.13c -0,02 0,61 -0,64 P87239 DnaJ homolog 1, mitochondrial mdj1 -0,07 0,49 -0,56 P87240 General negative regulator of transcription subunit 2 not2 0,05 0,22 -0,17 P87243 Uncharacterized protein C4G3.18 SPCC4G3.18 -0,05 0,01 -0,06 P87308 Protein lsb5 lsb5 0,31 0,11 0,20 P87313 Translation machinery-associated protein 20 tma20 -0,02 0,10 -0,12 P87316 Putative ATP synthase subunit epsilon, mitochondrial atp15 0,15 -0,32 0,47 P87317 Uncharacterized protein C31F10.16 SPBC31F10.16 -0,02 0,19 -0,21 Q00472 4-nitrophenylphosphatase pho2 0,00 0,22 -0,23 Q01112 Cell division control protein 42 homolog cdc42 0,07 0,07 0,00 Q01475 Small COPII coat GTPase sar1 sar1 0,07 0,16 -0,09 Q01663 AP-1-like transcription factor pap1 -0,19 0,60 -0,80 Q01682 Thiamine-repressible acid phosphatase pho4 pho4 0,07 -0,04 0,11 Q02061 DNA-directed RNA polymerase II subunit RPB2 rpb2 0,01 0,36 -0,35 Q02088 Tropomyosin cdc8 0,07 0,06 0,01 Q02592 Heavy metal tolerance protein hmt1 -0,12 0,13 -0,25 Q02787 Adenylosuccinate synthetase ade2 -0,04 -0,13 0,09 Q03392 Proliferating cell nuclear antigen pcn1 0,02 0,23 -0,21 Q06975 H/ACA ribonucleoprotein complex subunit 1 gar1 0,02 -0,03 0,05 Q09116 Septin homolog spn2 spn2 -0,11 0,37 -0,49 Q09127 60S ribosomal protein L10-A rpl10a 0,00 -0,29 0,29 Q09130 Eukaryotic translation initiation factor 2 subunit gamma tif213 0,00 0,04 -0,04 Q09140 UDP-glucose:glycoprotein glucosyltransferase gpt1 -0,04 0,07 -0,11 Q09146 Brefeldin A resistance protein hba1 -0,04 0,23 -0,26
Q09171 Pyruvate dehydrogenase E1 component subunit beta, mitochondrial pdb1 -0,04 -0,11 0,07
Q09172 Protein phosphatase 2C homolog 2 ptc2 -0,09 0,23 -0,32 Q09173 Protein phosphatase 2C homolog 3 ptc3 0,10 0,05 0,05 Q09174 Alpha-1,2-galactosyltransferase gma12 -0,12 -0,06 -0,06 Q09176 Splicing factor U2AF 23 kDa subunit SPAP8A3.06 0,13 0,20 -0,07 Q09177 DNA-directed RNA polymerases I and III subunit RPAC2 rpc19 -0,06 -0,06 0,00 Q09179 Glutamine synthetase gln1 0,48 -0,11 0,59 Q09181 Putative ATP-dependent RNA helicase ste13 ste13 0,04 0,12 -0,08 Q09184 Curved DNA-binding protein cdb4 -0,07 -0,16 0,09 Q09188 ADP,ATP carrier protein anc1 0,03 -0,03 0,06 Q09191 DNA-directed RNA polymerases I, II, and III subunit RPABC1 rpb5 -0,08 -0,01 -0,07 Q09196 Myosin regulatory light chain cdc4 cdc4 0,50 0,07 0,43 Q09330 mRNA export protein mlo3 mlo3 -0,02 -0,07 0,05 Q09668 60S ribosomal protein L22 rpl22 0,01 -0,27 0,28 Q09669 Alcohol dehydrogenase 4 adh4 -1,24 -1,46 0,22 Q09671 Putative NADPH dehydrogenase C5H10.10 SPAC5H10.10 -0,51 0,23 -0,73 Q09673 Pantoate--beta-alanine ligase pan6 0,11 -0,07 0,18 Q09674 UPF0317 protein C5H10.01 SPAC5H10.01 1,80 -1,12 2,93 Q09676 Uncharacterized protein C5H10.03 SPAC5H10.03 0,02 0,11 -0,10 Q09681 Probable alpha-1,2-galactosyltransferase gmh2 gmh2 -0,08 -0,16 0,08 Q09682 Probable proteasome subunit alpha type-4 -0,02 0,36 -0,38 Q09686 Putative glutamine amidotransferase-like protein C13C5.04 SPAC13C5.04 0,19 0,83 -0,64 Q09689 Probable eukaryotic translation initiation factor 5 SPAC2F7.05c -0,23 -0,33 0,10 Q09692 Tryptophan--tRNA ligase, cytoplasmic wrs1 0,00 -0,10 0,10 Q09694 Saccharopine dehydrogenase [NAD(+), L-lysine-forming] lys3 -0,26 0,03 -0,29 Q09697 Rho-GTPase-activating protein 8 rga8 0,09 0,21 -0,12 Q09702 Negative regulator of differentiation 1 nrd1 0,06 0,04 0,02 Q09704 Exosome complex component rrp4 rrp4 -0,03 0,03 -0,06 Q09715 Transcriptional repressor tup11 tup11 0,01 -0,13 0,14 Q09717 Ran-specific GTPase-activating protein 1 sbp1 -0,03 0,16 -0,19 Q09720 Probable proteasome subunit beta type-2 SPAC31A2.04c 0,25 0,85 -0,59
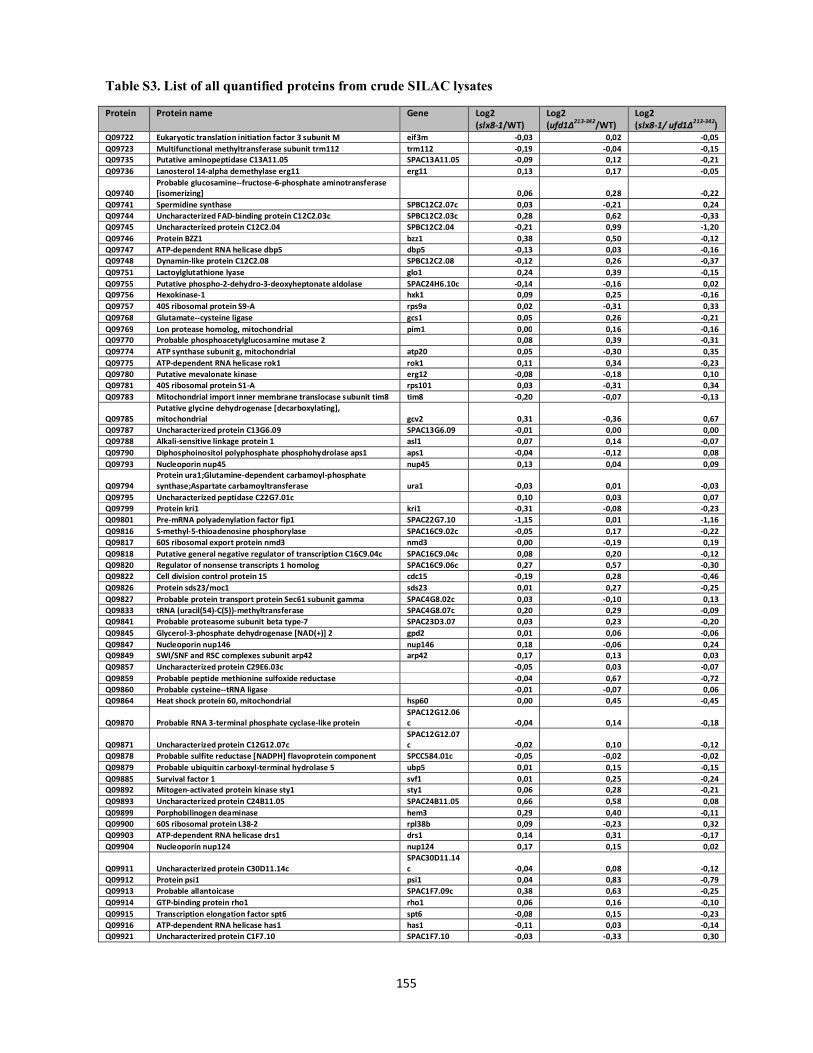
Table S3. List of all quantified proteins from crude SILAC lysates
155
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
Q09722 Eukaryotic translation initiation factor 3 subunit M eif3m -0,03 0,02 -0,05 Q09723 Multifunctional methyltransferase subunit trm112 trm112 -0,19 -0,04 -0,15 Q09735 Putative aminopeptidase C13A11.05 SPAC13A11.05 -0,09 0,12 -0,21 Q09736 Lanosterol 14-alpha demethylase erg11 erg11 0,13 0,17 -0,05
Q09740 Probable glucosamine--fructose-6-phosphate aminotransferase [isomerizing] 0,06 0,28 -0,22
Q09741 Spermidine synthase SPBC12C2.07c 0,03 -0,21 0,24 Q09744 Uncharacterized FAD-binding protein C12C2.03c SPBC12C2.03c 0,28 0,62 -0,33 Q09745 Uncharacterized protein C12C2.04 SPBC12C2.04 -0,21 0,99 -1,20 Q09746 Protein BZZ1 bzz1 0,38 0,50 -0,12 Q09747 ATP-dependent RNA helicase dbp5 dbp5 -0,13 0,03 -0,16 Q09748 Dynamin-like protein C12C2.08 SPBC12C2.08 -0,12 0,26 -0,37 Q09751 Lactoylglutathione lyase glo1 0,24 0,39 -0,15 Q09755 Putative phospho-2-dehydro-3-deoxyheptonate aldolase SPAC24H6.10c -0,14 -0,16 0,02 Q09756 Hexokinase-1 hxk1 0,09 0,25 -0,16 Q09757 40S ribosomal protein S9-A rps9a 0,02 -0,31 0,33 Q09768 Glutamate--cysteine ligase gcs1 0,05 0,26 -0,21 Q09769 Lon protease homolog, mitochondrial pim1 0,00 0,16 -0,16 Q09770 Probable phosphoacetylglucosamine mutase 2 0,08 0,39 -0,31 Q09774 ATP synthase subunit g, mitochondrial atp20 0,05 -0,30 0,35 Q09775 ATP-dependent RNA helicase rok1 rok1 0,11 0,34 -0,23 Q09780 Putative mevalonate kinase erg12 -0,08 -0,18 0,10 Q09781 40S ribosomal protein S1-A rps101 0,03 -0,31 0,34 Q09783 Mitochondrial import inner membrane translocase subunit tim8 tim8 -0,20 -0,07 -0,13
Q09785 Putative glycine dehydrogenase [decarboxylating], mitochondrial gcv2 0,31 -0,36 0,67
Q09787 Uncharacterized protein C13G6.09 SPAC13G6.09 -0,01 0,00 0,00 Q09788 Alkali-sensitive linkage protein 1 asl1 0,07 0,14 -0,07 Q09790 Diphosphoinositol polyphosphate phosphohydrolase aps1 aps1 -0,04 -0,12 0,08 Q09793 Nucleoporin nup45 nup45 0,13 0,04 0,09
Q09794 Protein ura1;Glutamine-dependent carbamoyl-phosphate synthase;Aspartate carbamoyltransferase ura1 -0,03 0,01 -0,03
Q09795 Uncharacterized peptidase C22G7.01c 0,10 0,03 0,07 Q09799 Protein kri1 kri1 -0,31 -0,08 -0,23 Q09801 Pre-mRNA polyadenylation factor fip1 SPAC22G7.10 -1,15 0,01 -1,16 Q09816 S-methyl-5-thioadenosine phosphorylase SPAC16C9.02c -0,05 0,17 -0,22 Q09817 60S ribosomal export protein nmd3 nmd3 0,00 -0,19 0,19 Q09818 Putative general negative regulator of transcription C16C9.04c SPAC16C9.04c 0,08 0,20 -0,12 Q09820 Regulator of nonsense transcripts 1 homolog SPAC16C9.06c 0,27 0,57 -0,30 Q09822 Cell division control protein 15 cdc15 -0,19 0,28 -0,46 Q09826 Protein sds23/moc1 sds23 0,01 0,27 -0,25 Q09827 Probable protein transport protein Sec61 subunit gamma SPAC4G8.02c 0,03 -0,10 0,13 Q09833 tRNA (uracil(54)-C(5))-methyltransferase SPAC4G8.07c 0,20 0,29 -0,09 Q09841 Probable proteasome subunit beta type-7 SPAC23D3.07 0,03 0,23 -0,20 Q09845 Glycerol-3-phosphate dehydrogenase [NAD(+)] 2 gpd2 0,01 0,06 -0,06 Q09847 Nucleoporin nup146 nup146 0,18 -0,06 0,24 Q09849 SWI/SNF and RSC complexes subunit arp42 arp42 0,17 0,13 0,03 Q09857 Uncharacterized protein C29E6.03c -0,05 0,03 -0,07 Q09859 Probable peptide methionine sulfoxide reductase -0,04 0,67 -0,72 Q09860 Probable cysteine--tRNA ligase -0,01 -0,07 0,06 Q09864 Heat shock protein 60, mitochondrial hsp60 0,00 0,45 -0,45
Q09870 Probable RNA 3-terminal phosphate cyclase-like protein SPAC12G12.06c -0,04 0,14 -0,18
Q09871 Uncharacterized protein C12G12.07c SPAC12G12.07c -0,02 0,10 -0,12
Q09878 Probable sulfite reductase [NADPH] flavoprotein component SPCC584.01c -0,05 -0,02 -0,02 Q09879 Probable ubiquitin carboxyl-terminal hydrolase 5 ubp5 0,01 0,15 -0,15 Q09885 Survival factor 1 svf1 0,01 0,25 -0,24 Q09892 Mitogen-activated protein kinase sty1 sty1 0,06 0,28 -0,21 Q09893 Uncharacterized protein C24B11.05 SPAC24B11.05 0,66 0,58 0,08 Q09899 Porphobilinogen deaminase hem3 0,29 0,40 -0,11 Q09900 60S ribosomal protein L38-2 rpl38b 0,09 -0,23 0,32 Q09903 ATP-dependent RNA helicase drs1 drs1 0,14 0,31 -0,17 Q09904 Nucleoporin nup124 nup124 0,17 0,15 0,02
Q09911 Uncharacterized protein C30D11.14c SPAC30D11.14c -0,04 0,08 -0,12
Q09912 Protein psi1 psi1 0,04 0,83 -0,79 Q09913 Probable allantoicase SPAC1F7.09c 0,38 0,63 -0,25 Q09914 GTP-binding protein rho1 rho1 0,06 0,16 -0,10 Q09915 Transcription elongation factor spt6 spt6 -0,08 0,15 -0,23 Q09916 ATP-dependent RNA helicase has1 has1 -0,11 0,03 -0,14 Q09921 Uncharacterized protein C1F7.10 SPAC1F7.10 -0,03 -0,33 0,30

Table S3. List of all quantified proteins from crude SILAC lysates
156
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
Q09923 Aldo-keto reductase yakc [NADP(+)] yakc 0,10 0,21 -0,11 Q09924 Probable translation initiation factor eIF-2B subunit delta tif224 -0,08 0,01 -0,08 Q09925 Serine palmitoyltransferase 2 lcb2 0,00 0,08 -0,08 Q10055 ATP-dependent RNA helicase fal1 tif412 -0,36 0,02 -0,38 Q10057 Putative protein disulfide-isomerase C1F5.02 SPAC1F5.02 0,08 0,09 -0,02 Q10058 Putative oxidoreductase C1F5.03c SPAC1F5.03c 0,06 0,22 -0,16 Q10061 Heat shock protein 70 homolog SPAC1F5.06 0,01 0,01 0,00 Q10066 Arginase aru1 -0,15 0,83 -0,97 Q10071 Probable CAAX prenyl protease 1 SPAC3H1.05 0,15 0,28 -0,14 Q10081 UPF0039 protein C11D3.02c SPAC11D3.02c -0,33 -0,14 -0,19 Q10083 Uncharacterized protein C11D3.04c SPAC11D3.04c 0,26 -0,67 0,93 Q10094 Uncharacterized protein C11D3.15 SPAC11D3.15 1,05 -0,44 1,49 Q10099 Nucleoporin seh1 seh1 -0,10 0,20 -0,30 Q10101 40S ribosomal protein S7 rps7 0,00 -0,29 0,29 Q10104 Serine hydroxymethyltransferase, mitochondrial shm2 0,06 -0,11 0,17 Q10105 Translational activator gcn1 gcn1 0,01 0,07 -0,06 Q10107 Probable ribosomal RNA small subunit methyltransferase mra1 mra1 -0,10 0,03 -0,14 Q10113 Microtubule integrity protein mal3 mal3 0,12 0,42 -0,30 Q10119 Elongation factor 1-alpha-B/C tef1b 0,04 -0,01 0,05 Q10132 Isopentenyl-diphosphate Delta-isomerase idi1 0,19 -0,10 0,29 Q10137 Sec14 cytosolic factor sec14 0,00 0,03 -0,03 Q10138 CRAL-TRIO domain-containing protein C3H8.02 SPAC3H8.02 0,06 0,30 -0,25 Q10147 Probable T-complex protein 1 subunit beta cct2 -0,07 0,06 -0,13 Q10149 Uncharacterized protein C1D4.02c SPAC1D4.02c -0,03 0,49 -0,52 Q10157 60S ribosomal protein L11 rpl11a 0,02 -0,25 0,27 Q10161 Probable clathrin heavy chain chc1 0,09 0,02 0,07 Q10163 Small nuclear ribonucleoprotein-associated protein B smb1 0,02 -0,07 0,10 Q10166 UPF0012 hydrolase C26A3.11 SPAC26A3.11 0,14 0,03 0,10 Q10168 Nucleoporin nsp1 nsp1 0,08 0,06 0,02 Q10169 Deubiquitination-protection protein dph1 dph1 0,05 0,03 0,01 Q10170 Arginine N-methyltransferase 2 rmt2 -0,07 0,03 -0,09 Q10172 Actin cytoskeleton-regulatory complex protein pan1 pan1 0,12 0,01 0,11 Q10175 Probable peptidyl-prolyl cis-trans isomerase C27F1.06c SPAC27F1.06c -0,04 0,01 -0,05
Q10176 Dolichyl-diphosphooligosaccharide--protein glycosyltransferase subunit 1 ost1 0,08 0,03 0,04
Q10179 Putative glycine--tRNA ligase SPAC3F10.03 -0,06 -0,19 0,13
Q10184
1-(5-phosphoribosyl)-5-[(5-phosphoribosylamino)methylideneamino] imidazole-4-carboxamide isomerase SPAC3F10.09 0,09 0,07 0,01
Q10190 Large subunit GTPase 1 lsg1 0,48 0,37 0,11 Q10192 60S ribosomal protein L18-A rpl18a -0,02 -0,34 0,32
Q10195 Uncharacterized protein C11C11.06c SPBC11C11.06c 0,01 0,57 -0,56
Q10199 Septation protein imp2 imp2 0,00 0,33 -0,32 Q10202 ATP-dependent RNA helicase dbp3 dbp3 -0,14 -0,17 0,02 Q10205 Exosome complex component rrp43 rrp43 -0,10 -0,02 -0,08 Q10208 Pyruvate kinase pyk1 0,04 -0,14 0,17 Q10209 Uncharacterized J domain-containing protein C4H3.01 SPAC4H3.01 -0,13 0,00 -0,13 Q10215 Putative thiosulfate sulfurtransferase, mitochondrial SPAC4H3.07c 0,08 -0,13 0,21 Q10217 Putative acyl carrier protein, mitochondrial SPAC4H3.09 0,18 0,26 -0,08 Q10224 Uncharacterized protein C13D6.03c SPAC13D6.03c 0,11 0,00 0,11 Q10232 60S ribosomal protein L9-A rpl9a 0,05 -0,17 0,22 Q10234 Probable 37S ribosomal protein S5, mitochondrial mrps5 0,17 0,04 0,13 Q10235 Cell polarity protein alp11 alp11 0,04 0,34 -0,30 Q10241 Mismatch-binding protein cmb1 cmb1 0,10 0,57 -0,47 Q10242 Probable gluconokinase SPAC4G9.12 0,06 0,25 -0,18 Q10245 3-ketoacyl-CoA reductase SPAC4G9.15 0,20 -0,05 0,24 Q10251 Eukaryotic translation initiation factor 5B SPAC56F8.03 0,01 -0,03 0,04 Q10253 Meiotically up-regulated gene 64 protein mug64 -0,16 -0,20 0,04 Q10256 UBA domain-containing protein mud1 mud1 0,13 0,17 -0,04 Q10258 Methylenetetrahydrofolate reductase 1 met9 0,10 0,12 -0,02 Q10265 Probable heat shock protein ssa1 ssa1 0,10 2,02 -1,91 Q10270 Probable phosphoadenosine phosphosulfate reductase SPAC13G7.06 -0,04 0,39 -0,44 Q10276 Putative choline kinase SPAC13G7.12c -0,29 -0,27 -0,02 Q10281 Guanine nucleotide-binding protein subunit beta-like protein rkp1 -0,02 -0,12 0,10 Q10283 3-hydroxy-3-methylglutaryl-coenzyme A reductase hmg1 -0,06 -0,35 0,29 Q10284 Heat shock protein sks2 sks2 -0,05 -0,14 0,09
Q10289
Fatty acid synthase subunit alpha;Acyl carrier;3-oxoacyl-[acyl-carrier-protein] reductase;3-oxoacyl-[acyl-carrier-protein] synthase fas2 0,13 -0,44 0,57
Q10294 cAMP-independent regulatory protein pac2 pac2 -0,19 -0,08 -0,10 Q10295 Poly(A) polymerase pla1 pla1 -0,17 -0,05 -0,12

Table S3. List of all quantified proteins from crude SILAC lysates
157
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
Q10297 Importin subunit beta-5 kap114 0,07 -0,12 0,18
Q10299 Protein clp1 SPAC22H10.05c -0,10 -0,03 -0,07
Q10305 Probable secretory pathway GDP dissociation inhibitor 1 gdi1 -0,03 0,31 -0,34 Q10306 Probable citrate synthase, mitochondrial cit1 -0,15 0,39 -0,54 Q10311 Ankyrin repeat-containing protein C6C3.08 SPAC6C3.08 0,02 0,30 -0,29 Q10316 Actin-related protein 2/3 complex subunit 5 arc5 -0,16 0,21 -0,37 Q10318 Putative dihydroxy-acid dehydratase, mitochondrial SPAC17G8.06c -0,13 -0,15 0,02 Q10329 Probable proteasome subunit alpha type-7 SPBC106.16 0,01 0,29 -0,29 Q10330 60S ribosomal protein L25-A rpl25a 0,02 -0,30 0,33 Q10333 Uncharacterized protein C582.09 SPBC582.09 0,02 0,05 -0,04 Q10334 Putative alanine aminotransferase SPBC582.08 -0,10 -0,10 0,00 Q10335 Probable 26S proteasome regulatory subunit rpn7 rpn7 0,08 0,27 -0,19 Q10341 Probable serine-O-acetyltransferase cys2 cys2 -0,19 0,42 -0,61 Q10344 Translationally-controlled tumor protein homolog p23fy 0,03 -0,13 0,16 Q10346 Uncharacterized protein C1F12.04c SPAC1F12.04c -0,02 0,09 -0,11 Q10349 Putative phosphoserine aminotransferase SPAC1F12.07 -0,15 0,03 -0,18 Q10352 Uncharacterized protein C1F12.10c SPAC1F12.10c -0,12 -0,07 -0,05
Q10356 Uncharacterized protein C22E12.03c SPAC22E12.03c 0,02 0,45 -0,43
Q10357 Superoxide dismutase 1 copper chaperone ccs1 -0,01 0,35 -0,37 Q10367 ADP-ribosylation factor GTPase-activating protein glo3 glo3 -0,05 0,14 -0,19 Q10410 Probable ADP-ribosylation factor-binding protein C1F3.05 SPAC1F3.05 0,06 0,13 -0,07 Q10411 Sporulation-specific protein 15 spo15 0,03 0,24 -0,21 Q10414 CWF19-like protein mug161 mug161 -0,03 0,25 -0,28 Q10421 40S ribosomal protein S28 rps28a 0,02 -0,23 0,25 Q10425 Eukaryotic translation initiation factor 3 subunit B SPAC25G10.08 -0,02 0,06 -0,08
Q10428 Serine/threonine-protein phosphatase 2A 56 kDa regulatory subunit delta 1 isoform par1 0,19 0,36 -0,17
Q10434 F-actin-capping protein subunit alpha acp1 0,06 0,11 -0,06
Q10436 Uncharacterized protein C12B10.02c SPAC12B10.02c 0,24 0,42 -0,18
Q10437 UBP9-binding protein bun62 bun62 -0,35 0,08 -0,43 Q10452 Protein kinase gsk3 gsk3 0,07 0,26 -0,19 Q10475 Eukaryotic translation initiation factor 4 gamma tif471 -0,02 0,03 -0,05 Q10478 SAM50-like protein SpAC17C9.06 SPAC17C9.06 -0,02 0,48 -0,50 Q10480 Nuclease 1, mitochondrial pnu1 -0,30 0,01 -0,31
Q10481 Mitochondrial import inner membrane translocase subunit tim13 tim13 0,06 0,31 -0,26
Q10484 Uncharacterized protein C17C9.12 SPAC17C9.12 -0,06 0,12 -0,18 Q10488 Probable trans-2-enoyl-CoA reductase, mitochondrial etr1 -0,06 1,06 -1,12
Q10489 Pyruvate dehydrogenase E1 component subunit alpha, mitochondrial pda1 -0,01 -0,07 0,06
Q10490 Putative leucine--tRNA ligase, cytoplasmic SPAC26F1.13c -0,13 -0,31 0,18 Q10494 Uncharacterized oxidoreductase C26F1.07 SPAC26F1.07 0,10 0,36 -0,26 Q10498 FAM203 family protein C1020.12c SPAC26F1.12c 0,00 0,16 -0,16 Q10499 Apoptosis-inducing factor 1 aif1 0,35 1,22 -0,87 Q10659 Membrane glycoprotein spo14 spo14 -0,04 0,21 -0,26 Q10668 Mating-type switching protein swi2 swi2 -0,12 -0,19 0,07 Q11004 40 kDa peptidyl-prolyl cis-trans isomerase wis2 0,11 0,69 -0,58 Q11118 Protein wos2 wos2 -0,10 0,29 -0,39 Q11119 Ubiquitin carboxyl-terminal hydrolase 14 ubp14 0,11 0,29 -0,17 Q11120 PKHD-type hydroxylase ofd1 ofd1 0,36 -0,20 0,56 Q12381 Pre-mRNA-splicing factor prp1 prp1 -0,01 -0,12 0,11 Q12702 Protein phosphatase PP2A regulatory subunit B pab1 -0,07 0,18 -0,25 Q1K9B6 Uncharacterized protein C19D5.02c SPAC19D5.02c -0,10 0,09 -0,19 Q1K9C2 Sulfite reductase [NADPH] subunit beta sir1 0,03 -0,16 0,19 Q1MTQ9 60S ribosome subunit biogenesis protein nip7 nip7 -0,06 0,06 -0,12 Q1MTR3 Vacuolar import and degradation protein 27 vid27 0,25 0,15 0,10 Q1MTS0 Uncharacterized protein C3B8.09 SPBC3B8.09 -0,34 0,02 -0,36 Q42598 Threonine synthase thrc -0,08 -0,09 0,01 Q7Z991 Uncharacterized ABC transporter ATP-binding protein C20G4.01 SPAC20G4.01 0,05 0,04 0,00 Q7Z9I2 Uncharacterized oxidoreductase C663.09c SPCC663.09c 0,20 0,35 -0,15 Q7Z9I4 Uncharacterized oxidoreductase C663.06c SPCC663.06c 0,21 0,32 -0,11 Q8NIL3 Antisense-enhancing sequence 1 aes1 -0,52 0,23 -0,76
Q8TFH0 Probable 6-phosphofructo-2-kinase PB17E12.14c SPAPB17E12.14c -0,42 -0,19 -0,23
Q8TFH1 60S ribosomal protein L18-B rpl18b 0,01 -0,24 0,25
Q8TFH3 Uncharacterized protein PB17E12.11 SPAPB17E12.11 0,09 0,14 -0,05
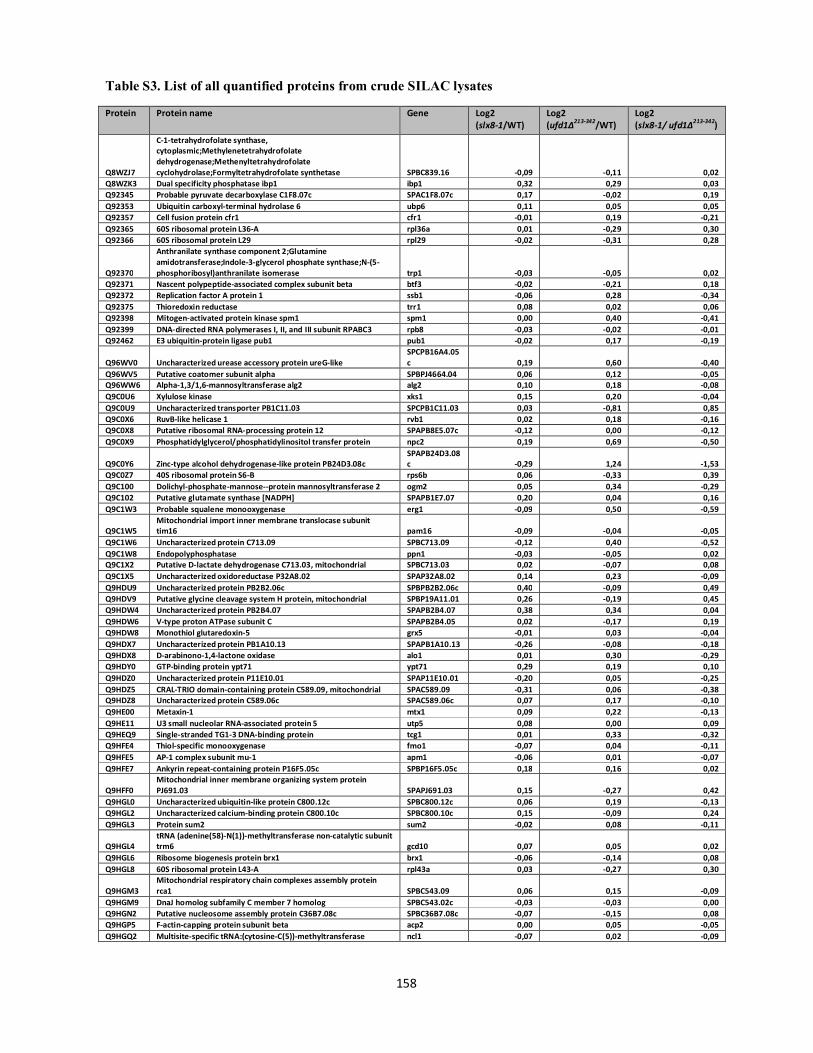
Table S3. List of all quantified proteins from crude SILAC lysates
158
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
Q8WZJ7
C-1-tetrahydrofolate synthase, cytoplasmic;Methylenetetrahydrofolate dehydrogenase;Methenyltetrahydrofolate cyclohydrolase;Formyltetrahydrofolate synthetase SPBC839.16 -0,09 -0,11 0,02
Q8WZK3 Dual specificity phosphatase ibp1 ibp1 0,32 0,29 0,03 Q92345 Probable pyruvate decarboxylase C1F8.07c SPAC1F8.07c 0,17 -0,02 0,19 Q92353 Ubiquitin carboxyl-terminal hydrolase 6 ubp6 0,11 0,05 0,05 Q92357 Cell fusion protein cfr1 cfr1 -0,01 0,19 -0,21 Q92365 60S ribosomal protein L36-A rpl36a 0,01 -0,29 0,30 Q92366 60S ribosomal protein L29 rpl29 -0,02 -0,31 0,28
Q92370
Anthranilate synthase component 2;Glutamine amidotransferase;Indole-3-glycerol phosphate synthase;N-(5-phosphoribosyl)anthranilate isomerase trp1 -0,03 -0,05 0,02
Q92371 Nascent polypeptide-associated complex subunit beta btf3 -0,02 -0,21 0,18 Q92372 Replication factor A protein 1 ssb1 -0,06 0,28 -0,34 Q92375 Thioredoxin reductase trr1 0,08 0,02 0,06 Q92398 Mitogen-activated protein kinase spm1 spm1 0,00 0,40 -0,41 Q92399 DNA-directed RNA polymerases I, II, and III subunit RPABC3 rpb8 -0,03 -0,02 -0,01 Q92462 E3 ubiquitin-protein ligase pub1 pub1 -0,02 0,17 -0,19
Q96WV0 Uncharacterized urease accessory protein ureG-like SPCPB16A4.05c 0,19 0,60 -0,40
Q96WV5 Putative coatomer subunit alpha SPBPJ4664.04 0,06 0,12 -0,05 Q96WW6 Alpha-1,3/1,6-mannosyltransferase alg2 alg2 0,10 0,18 -0,08 Q9C0U6 Xylulose kinase xks1 0,15 0,20 -0,04 Q9C0U9 Uncharacterized transporter PB1C11.03 SPCPB1C11.03 0,03 -0,81 0,85 Q9C0X6 RuvB-like helicase 1 rvb1 0,02 0,18 -0,16 Q9C0X8 Putative ribosomal RNA-processing protein 12 SPAPB8E5.07c -0,12 0,00 -0,12 Q9C0X9 Phosphatidylglycerol/phosphatidylinositol transfer protein npc2 0,19 0,69 -0,50
Q9C0Y6 Zinc-type alcohol dehydrogenase-like protein PB24D3.08c SPAPB24D3.08c -0,29 1,24 -1,53
Q9C0Z7 40S ribosomal protein S6-B rps6b 0,06 -0,33 0,39 Q9C100 Dolichyl-phosphate-mannose--protein mannosyltransferase 2 ogm2 0,05 0,34 -0,29 Q9C102 Putative glutamate synthase [NADPH] SPAPB1E7.07 0,20 0,04 0,16 Q9C1W3 Probable squalene monooxygenase erg1 -0,09 0,50 -0,59
Q9C1W5 Mitochondrial import inner membrane translocase subunit tim16 pam16 -0,09 -0,04 -0,05
Q9C1W6 Uncharacterized protein C713.09 SPBC713.09 -0,12 0,40 -0,52 Q9C1W8 Endopolyphosphatase ppn1 -0,03 -0,05 0,02 Q9C1X2 Putative D-lactate dehydrogenase C713.03, mitochondrial SPBC713.03 0,02 -0,07 0,08 Q9C1X5 Uncharacterized oxidoreductase P32A8.02 SPAP32A8.02 0,14 0,23 -0,09 Q9HDU9 Uncharacterized protein PB2B2.06c SPBPB2B2.06c 0,40 -0,09 0,49 Q9HDV9 Putative glycine cleavage system H protein, mitochondrial SPBP19A11.01 0,26 -0,19 0,45 Q9HDW4 Uncharacterized protein PB2B4.07 SPAPB2B4.07 0,38 0,34 0,04 Q9HDW6 V-type proton ATPase subunit C SPAPB2B4.05 0,02 -0,17 0,19 Q9HDW8 Monothiol glutaredoxin-5 grx5 -0,01 0,03 -0,04 Q9HDX7 Uncharacterized protein PB1A10.13 SPAPB1A10.13 -0,26 -0,08 -0,18 Q9HDX8 D-arabinono-1,4-lactone oxidase alo1 0,01 0,30 -0,29 Q9HDY0 GTP-binding protein ypt71 ypt71 0,29 0,19 0,10 Q9HDZ0 Uncharacterized protein P11E10.01 SPAP11E10.01 -0,20 0,05 -0,25 Q9HDZ5 CRAL-TRIO domain-containing protein C589.09, mitochondrial SPAC589.09 -0,31 0,06 -0,38 Q9HDZ8 Uncharacterized protein C589.06c SPAC589.06c 0,07 0,17 -0,10 Q9HE00 Metaxin-1 mtx1 0,09 0,22 -0,13 Q9HE11 U3 small nucleolar RNA-associated protein 5 utp5 0,08 0,00 0,09 Q9HEQ9 Single-stranded TG1-3 DNA-binding protein tcg1 0,01 0,33 -0,32 Q9HFE4 Thiol-specific monooxygenase fmo1 -0,07 0,04 -0,11 Q9HFE5 AP-1 complex subunit mu-1 apm1 -0,06 0,01 -0,07 Q9HFE7 Ankyrin repeat-containing protein P16F5.05c SPBP16F5.05c 0,18 0,16 0,02
Q9HFF0 Mitochondrial inner membrane organizing system protein PJ691.03 SPAPJ691.03 0,15 -0,27 0,42
Q9HGL0 Uncharacterized ubiquitin-like protein C800.12c SPBC800.12c 0,06 0,19 -0,13 Q9HGL2 Uncharacterized calcium-binding protein C800.10c SPBC800.10c 0,15 -0,09 0,24 Q9HGL3 Protein sum2 sum2 -0,02 0,08 -0,11
Q9HGL4 tRNA (adenine(58)-N(1))-methyltransferase non-catalytic subunit trm6 gcd10 0,07 0,05 0,02
Q9HGL6 Ribosome biogenesis protein brx1 brx1 -0,06 -0,14 0,08 Q9HGL8 60S ribosomal protein L43-A rpl43a 0,03 -0,27 0,30
Q9HGM3 Mitochondrial respiratory chain complexes assembly protein rca1 SPBC543.09 0,06 0,15 -0,09
Q9HGM9 DnaJ homolog subfamily C member 7 homolog SPBC543.02c -0,03 -0,03 0,00 Q9HGN2 Putative nucleosome assembly protein C36B7.08c SPBC36B7.08c -0,07 -0,15 0,08 Q9HGP5 F-actin-capping protein subunit beta acp2 0,00 0,05 -0,05 Q9HGQ2 Multisite-specific tRNA:(cytosine-C(5))-methyltransferase ncl1 -0,07 0,02 -0,09

Table S3. List of all quantified proteins from crude SILAC lysates
159
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
Q9P376 RNA polymerase II subunit A C-terminal domain phosphatase fcp1 0,19 0,36 -0,17 Q9P382 Nucleoporin nup82 nup82 -0,06 0,03 -0,09 Q9P383 Nuclear cap-binding protein subunit 2 cbc2 -0,09 0,09 -0,18 Q9P3A7 Cell division cycle protein 48 cdc48 0,05 0,66 -0,61 Q9P3A9 Uncharacterized protein C1565.05 SPAC1565.05 -0,14 0,02 -0,16 Q9P3B2 Altered inheritance rate of mitochondria protein 38 homolog aim38 0,32 -0,91 1,22 Q9P3T6 40S ribosomal protein S5-B rps5b 0,01 -0,17 0,18 Q9P3U0 Probable ubiquitin carboxyl-terminal hydrolase 2 ubp2 -0,29 0,22 -0,51 Q9P3W6 Uncharacterized protein C458.02c SPAC458.02c 0,01 -0,06 0,07 Q9P4X3 Probable U3 small nucleolar RNA-associated protein 7 utp7 -0,13 0,03 -0,16 Q9P4X4 Probable vesicular-fusion protein sec17 homolog sec17 0,09 -0,03 0,12 Q9P544 Probable mitochondrial outer membrane protein porin SPAC1635.01 0,12 0,47 -0,35 Q9P546 40S ribosomal protein S0-B rps0b 0,02 -0,20 0,22 Q9P5N2 Amino acid transporter 1 aat1 0,03 0,86 -0,83 Q9P6I2 Cys-Gly metallodipeptidase dug1 dug1 0,11 -0,07 0,18 Q9P6I5 Guanylate kinase SPBC1198.05 -0,03 -0,01 -0,02 Q9P6K0 Uncharacterized HTH La-type RNA-binding protein C1527.03 SPAC1527.03 -0,25 0,06 -0,31 Q9P6K3 UPF0662 protein C30C2.08 SPAC30C2.08 0,06 0,36 -0,30 Q9P6K7 Uncharacterized tRNA-binding protein C30C2.04 SPAC30C2.04 0,04 0,02 0,02 Q9P6K9 Deoxyhypusine hydroxylase lia1 0,08 -0,46 0,53 Q9P6L5 Endocytosis protein end4 end4 0,06 -0,03 0,09 Q9P6M1 Glutathione S-transferase 3 gst3 0,06 0,42 -0,36 Q9P6N2 Uncharacterized phosphatase C823.14 SPAC823.14 -0,08 -0,19 0,11 Q9P6N8 ATP-dependent rRNA helicase rrp3 rrp3 -0,03 -0,22 0,19 Q9P6Q1 UPF0001 protein C644.09 SPAC644.09 0,14 -0,04 0,18 Q9P6Q2 Enolase-phosphatase E1 utr4 0,05 0,24 -0,19 Q9P6Q5 Probable deoxyuridine 5-triphosphate nucleotidohydrolase SPAC644.05c 0,08 0,20 -0,12 Q9P6R1 Verprolin vrp1 -0,44 -0,44 0,01 Q9P6R2 Uncharacterized protein C13E7.08c SPBC13E7.08c -0,02 -0,06 0,04 Q9P6R3 Uncharacterized protein C13E7.07 SPBC13E7.07 0,03 0,14 -0,11 Q9P6R6 ATP synthase subunit delta, mitochondrial atp16 -0,03 -0,26 0,23 Q9P768 Uncharacterized amino-acid permease P7G5.06 SPAP7G5.06 1,48 0,19 1,29 Q9P772 GMP synthase [glutamine-hydrolyzing] gua1 -0,09 -0,13 0,04 Q9P775 Uncharacterized WD repeat-containing protein C17D11.16 SPBC1711.16 -0,05 0,00 -0,05 Q9P777 Histidinol dehydrogenase his2 -0,23 -0,07 -0,16 Q9P780 Nuclear protein localization protein 4 npl4 0,05 0,09 -0,04 Q9P782 Uncharacterized protein C1711.08 SPBC1711.08 0,08 0,40 -0,31 Q9P783 Ribosome assembly protein rrb1 rrb1 -0,13 0,03 -0,15 Q9P784 60S ribosomal protein L4-B rpl4b 0,00 -0,26 0,26 Q9P785 LisH domain-containing protein C1711.05 SPBC1711.05 0,10 0,03 0,07 Q9P786 Methylenetetrahydrofolate dehydrogenase [NAD(+)] mtd1 0,09 0,03 0,06 Q9P789 Pumilio domain-containing protein P35G2.14 SPBP35G2.14 -0,11 -0,01 -0,10 Q9P791 Uncharacterized Nudix hydrolase P35G2.12 SPBP35G2.12 -0,09 0,35 -0,44 Q9P7A1 PITH domain-containing protein P35G2.02 SPBP35G2.02 -0,08 0,73 -0,81 Q9P7B4 Uncharacterized oxidoreductase C521.03 SPAC521.03 0,44 0,26 0,18 Q9P7C9 m7GpppX diphosphatase nhm1 0,12 0,14 -0,02 Q9P7D4 Probable aconitate hydratase 2 SPBP4H10.15 -0,07 -0,21 0,14 Q9P7D6 UPF0303 protein P4H10.12 SPBP4H10.12 -0,01 0,30 -0,31 Q9P7D7 Long-chain-fatty-acid--CoA ligase 2 SPBP4H10.11c 0,11 -0,51 0,62 Q9P7E0 Cytochrome b-c1 complex subunit 10 qcr10 0,21 -0,28 0,49
Q9P7E3
S-adenosylmethionine decarboxylase proenzyme;S-adenosylmethionine decarboxylase alpha chain;S-adenosylmethionine decarboxylase beta chain spe2 -0,30 0,04 -0,33
Q9P7E8 Protein app1 app1 -0,07 0,19 -0,27 Q9P7F2 Copper amine oxidase 1 cao1 0,38 -0,07 0,44 Q9P7F8 ATPase get3 get3 0,06 0,05 0,01 Q9P7G0 Uncharacterized NOC2 family protein C1142.04 SPAC1142.04 -0,09 0,02 -0,10 Q9P7G9 Adenylyl-sulfate kinase SPAC1782.11 0,01 -0,25 0,27 Q9P7H0 H/ACA ribonucleoprotein complex subunit 2 nhp2 -0,04 -0,04 0,00 Q9P7H3 Prohibitin-1 phb1 -0,05 0,01 -0,06 Q9P7J6 40S ribosomal protein S17-B rps17b 0,06 -0,17 0,24 Q9P7J8 Serine/threonine-protein kinase gad8 gad8 -0,11 0,21 -0,32 Q9P7K6 SDO1-like protein C21C3.19 SPBC21C3.19 -0,08 1,08 -1,16 Q9P7K9 Putative aldehyde dehydrogenase-like protein C21C3 SPBC21C3.15c 0,06 -0,07 0,13 Q9P7L0 Uncharacterized protein C21C3.14c SPBC21C3.14c -0,07 0,04 -0,11 Q9P7L1 Thioredoxin domain-containing protein C21C3.12c SPBC21C3.12c -0,04 0,17 -0,21 Q9P7L2 Uncharacterized protein C21C3.11 SPBC21C3.11 -0,20 0,31 -0,50 Q9P7L4 Uncharacterized hydrolase C21C3.09c SPBC21C3.09c -0,04 0,12 -0,16 Q9P7L5 Ornithine aminotransferase car2 car2 -0,19 0,07 -0,26 Q9P7M5 H/ACA ribonucleoprotein complex subunit 3 nop10 -0,02 0,04 -0,06 Q9P7M8 Nucleoporin nup184 nup184 0,00 -0,20 0,20 Q9P7M9 Probable GTP cyclohydrolase-2 rib1 -0,02 0,34 -0,35

Table S3. List of all quantified proteins from crude SILAC lysates
160
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
Q9P7Q4 Vesicular-fusion protein sec18 sec18 0,09 0,41 -0,31
Q9P7R0
Pentafunctional AROM polypeptide;3-dehydroquinate synthase;3-phosphoshikimate 1-carboxyvinyltransferase;Shikimate kinase;3-dehydroquinate dehydratase;Shikimate dehydrogenase aro1 -0,12 -0,23 0,11
Q9P7R4 Ubiquitin-conjugating enzyme E2 8 ubc8 -0,04 0,56 -0,60 Q9P7S1 PDZ domain-containing protein C23G3.12c SPAC23G3.12c -0,04 -0,08 0,04 Q9P7S2 Probable 26S proteasome regulatory subunit rpn6 rpn6 0,01 0,26 -0,25 Q9P7S3 SWI/SNF and RSC complexes subunit ssr3 ssr3 0,21 0,27 -0,07 Q9P7S7 Nucleolar protein 58 nop58 -0,03 0,05 -0,07 Q9P7T0 L-ornithine 5-monooxygenase sib2 -0,18 0,26 -0,44 Q9P7T6 UPF0160 protein C694.04c SPAC694.04c 0,10 -0,23 0,32 Q9P7T8 Uncharacterized helicase C694.02 SPAC694.02 0,00 -0,09 0,09 Q9P7U2 Putative aryl-alcohol dehydrogenase C977.14c SPAC977.14c -0,30 1,85 -2,14 Q9P7V5 Protein transport protein sec72 sec72 -0,24 -0,33 0,10 Q9P7V8 Probable mitochondrial phosphate carrier protein SPBC1703.13c 0,05 -0,46 0,51 Q9P7W3 Probable ATP-citrate synthase subunit 1 SPBC1703.07 -0,03 -0,18 0,14 Q9P7W7 Uncharacterized ARM-like repeat-containing protein C1703.03c SPBC1703.03c -0,20 0,06 -0,26 Q9P7X1 Probable mitochondrial-processing peptidase subunit beta qcr1 0,05 -0,30 0,36 Q9P7X7 Actin-related protein 4 alp5 0,53 0,21 0,32 Q9P7X8 Probable DNA-directed RNA polymerase I subunit RPA2 rpa2 0,05 0,09 -0,03 Q9P7Y7 Pyrroline-5-carboxylate reductase pro3 -0,16 -0,14 -0,01 Q9P7Z7 Cytochrome c oxidase copper chaperone cox17 0,97 0,79 0,18 Q9UQW6 Acetyl-CoA acetyltransferase erg10 -0,01 -0,03 0,03 Q9UQW9 Ornithine decarboxylase spe1 -1,24 -0,72 -0,52 Q9UQX0 Superoxide dismutase [Mn], mitochondrial sod2 0,16 0,46 -0,31 Q9UQY2 Probable proteasome subunit beta type-1 pam1 0,00 0,34 -0,34 Q9UR06 Protein mmf2, mitochondrial mmf2 1,86 -0,23 2,09 Q9UR09 Cell wall synthesis protein psu1 psu1 0,00 0,18 -0,18 Q9UR17 Vacuolar transporter chaperone 1 nrf1 0,09 -0,09 0,17 Q9URM2 Probable transketolase SPBC2G5.05 0,14 0,01 0,13 Q9URT8 60S ribosomal protein L34-B rpl34b -0,02 -0,07 0,05 Q9URT9 Protein kinase gsk31 gsk31 0,00 0,30 -0,30 Q9URV8 UPF0744 protein C106.03 SPBC106.03 0,03 0,36 -0,34 Q9URW6 SH3 domain-containing protein PJ696.02 SPAPJ696.02 0,02 0,18 -0,16 Q9URW7 Vacuolar protein sorting-associated protein 17 vps17 -0,02 0,22 -0,24 Q9URW9 Putative aldehyde dehydrogenase-like protein C922.07c SPAC922.07c 0,77 -0,25 1,02 Q9URX2 Uncharacterized protein C922.04 SPAC922.04 0,21 0,14 0,07 Q9URX6 60S ribosomal protein L31 rpl31 0,01 -0,28 0,28 Q9URX7 Probable protein arginine N-methyltransferase SPAC890.07c -0,12 0,10 -0,22 Q9URX8 Probable nucleoporin C890.06 SPAC890.06 0,30 0,13 0,17 Q9URY0 Ribosome biogenesis protein ytm1 ytm1 0,00 0,18 -0,18 Q9URZ5 Vacuolar protein sorting-associated protein 1 vps1 0,06 0,30 -0,24 Q9URZ7 Probable fructose-2,6-bisphosphatase C732.02c SPAC732.02c 1,51 0,97 0,54 Q9US06 RNA polymerase II-associated protein 1 homolog SPAC664.03 -0,08 -0,20 0,12 Q9US07 Probable actin-related protein 8 SPAC664.02c 0,03 0,13 -0,10 Q9US13 Probable 26S proteasome regulatory subunit rpn9 rpn9 0,02 0,25 -0,24 Q9US20 Uncharacterized acyltransferase C1851.02 SPAC1851.02 0,04 0,09 -0,04 Q9US22 60S ribosomal protein L15-B rpl15b 0,00 -0,23 0,23 Q9US25 Chromodomain helicase hrp1 hrp1 0,07 0,18 -0,11 Q9US35 D-serine dehydratase SPAC1039.06 1,30 0,08 1,23 Q9US39 Uncharacterized protein C1039.02 SPAC1039.02 0,71 -0,62 1,34 Q9US42 Protein urg3 urg3 0,22 0,08 0,14
Q9US47 Putative succinate-semialdehyde dehydrogenase C1002.12c [NADP(+)] SPAC1002.12c 0,45 0,55 -0,10
Q9US55 Glucosidase 2 subunit alpha gls2 -0,01 0,03 -0,04 Q9USG6 60S ribosomal protein L33-B rpl33b -0,11 -0,48 0,37 Q9USH8 Glucosidase 2 subunit beta gtb1 -0,11 0,25 -0,36 Q9USH9 Uncharacterized ABC transporter ATP-binding protein C825.01 SPCC825.01 0,01 0,07 -0,05 Q9USI5 Heat shock protein sti1 homolog sti1 0,01 0,40 -0,39 Q9USJ1 Putative 3-mercaptopyruvate sulfurtransferase 0,06 0,44 -0,38 Q9USJ5 Uroporphyrinogen decarboxylase hem12 0,00 0,15 -0,15 Q9USJ6 NAD(P)H-dependent FMN reductase C4B3.06c SPCC4B3.06c 0,16 0,54 -0,38 Q9USJ7 Negative regulator of ofd1 nro1 0,10 -0,13 0,23 Q9USJ9 54S ribosomal protein L12, mitochondrial mnp1 -0,16 -0,07 -0,09 Q9USK0 Inositol-pentakisphosphate 2-kinase ipk1 -0,37 0,37 -0,75 Q9USK8 Cell wall alpha-1,3-glucan synthase ags1 ags1 0,03 0,15 -0,12 Q9USL4 Nucleoporin nup61 nup61 -0,05 0,05 -0,09 Q9USM3 Chromatin structure-remodeling complex subunit sfh1 sfh1 0,07 -0,05 0,12
Q9USM6 Probable cytochrome b5 2 SPCC16A11.10c 0,17 -0,19 0,36

Table S3. List of all quantified proteins from crude SILAC lysates
161
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
Q9USM7 Mitochondrial import inner membrane translocase subunit tim23 tim23 0,35 0,30 0,05
Q9USN3 Probable U3 small nucleolar RNA-associated protein 13 utp13 -0,05 0,12 -0,17 Q9USN5 Putative NAD-specific glutamate dehydrogenase SPCC132.04c 0,46 1,05 -0,58 Q9USN8 Uncharacterized protein C132.01c 0,04 0,13 -0,09 Q9USP0 Translation initiation factor eIF-2B subunit alpha tif221 0,10 0,05 0,05 Q9USP6 Clathrin light chain clc1 0,05 0,08 -0,04 Q9USP8 Isocitrate dehydrogenase [NAD] subunit 2, mitochondrial idh2 -0,03 0,40 -0,42 Q9USP9 Uncharacterized RNA-binding protein C902.04 SPBC902.04 0,04 0,19 -0,15 Q9USQ4 NASP-related protein sim3 sim3 -0,32 0,38 -0,70 Q9USQ9 Probable proteasome subunit beta type-4 SPBC577.10 0,04 0,32 -0,28 Q9USR1 Thioredoxin-like protein 1 txl1 0,03 0,35 -0,32 Q9USR7 60S ribosomal protein L38-1 rpl38a 0,03 -0,27 0,30 Q9USS2 Serine/threonine-protein kinase ppk29 ppk29 0,07 0,16 -0,09 Q9USS6 Probable acid phosphatase SPBC4.06 SPBC4.06 0,11 0,00 0,10 Q9USS7 Uncharacterized protein C4.03c SPBC4.03c 0,08 0,01 0,07 Q9UST4 Eukaryotic translation initiation factor 5A-2 tif51b -0,04 -0,08 0,04 Q9USU2 Dimethyladenosine transferase dim1 -0,28 -0,24 -0,04 Q9USU5 1,3-beta-glucanosyltransferase gas2 gas2 -0,44 0,75 -1,19 Q9USU7 HMG box-containing protein C28F2.11 SPBC28F2.11 -0,03 -0,28 0,25 Q9USV4 mRNA export protein 33 mep33 0,02 0,21 -0,19 Q9USW0 Uncharacterized protein C21B10.02 SPBC21B10.02 -0,08 0,29 -0,37
Q9USW1 Uncharacterized protein C21B10.03c SPBC21B10.03c -0,09 0,01 -0,11
Q9USW5 40S ribosomal protein S4-B rps4b 0,02 -0,26 0,28 Q9USX1 Aminopeptidase 1 ape1 0,17 0,23 -0,06 Q9USX4 60S ribosomal protein L33-A rpl33a 0,01 -0,24 0,25 Q9USZ1 Elongation factor G, mitochondrial mef1 0,06 -0,35 0,42 Q9USZ2 Uncharacterized protein C11G11.07 0,03 -0,12 0,14 Q9USZ4 DNA-directed RNA polymerase I subunit rpa34 rpa34 -0,02 0,00 -0,02 Q9USZ6 mRNA turnover protein 4 homolog SPBC11G11.03 0,10 0,03 0,07 Q9USZ7 Actin cytoskeleton-regulatory complex protein end3 end3 0,03 0,03 0,01
Q9UT00 Uncharacterized protein PYUK71.03c SPAPYUK71.03c -0,17 0,12 -0,29
Q9UT05 Tripeptidyl-peptidase 2 homolog SPAP8A3.12c 0,10 0,32 -0,22 Q9UT08 Protein phosphatase PP2A regulatory subunit A paa1 -0,03 0,07 -0,10
Q9UT09 Phospho-2-dehydro-3-deoxyheptonate aldolase, tyrosine-inhibited aro4 -0,21 -0,15 -0,06
Q9UT11 Uncharacterized zinc transporter P8A3.03 SPAP8A3.03 0,14 0,40 -0,26 Q9UT18 Uncharacterized amino-acid permease C9.10 SPAC9.10 0,02 0,04 -0,02
Q9UT19 Probable 5-methyltetrahydropteroyltriglutamate--homocysteine methyltransferase SPAC9.09 -0,03 -0,52 0,49
Q9UT21 Uncharacterized GTP-binding protein C9.07c SPAC9.07c -0,09 0,19 -0,28 Q9UT24 Pre-mRNA-splicing factor brr2 brr2 0,19 0,14 0,05 Q9UT25 Uncharacterized N-acetyltransferase C9.02c SPAC9.02c 0,02 0,15 -0,13 Q9UT27 Pyruvyl transferase 1 pvg1 -0,13 -1,07 0,94 Q9UT28 Putative nicotinamide N-methyltransferase nnt1 0,05 0,14 -0,09 Q9UT32 Putative ribosome biogenesis protein C8F11.04 SPAC8F11.04 -0,12 -0,05 -0,07 Q9UT36 Probable hydroxyacylglutathione hydrolase C824.07 SPAC824.07 0,06 0,55 -0,48
Q9UT37 Mitochondrial import inner membrane translocase subunit tim14 pam18 -0,04 0,01 -0,04
Q9UT44 Probable gamma-glutamyl phosphate reductase pro1 -0,09 0,07 -0,16 Q9UT48 Eukaryotic translation initiation factor 3 subunit H eif3h -0,08 0,04 -0,12 Q9UT55 UPF0364 protein C806.04c SPAC806.04c -0,03 -0,04 0,01 Q9UT56 40S ribosomal protein S26-A rps26a -0,05 -0,29 0,24 Q9UT57 Probable cytosolic Fe-S cluster assembly factor C806.02c SPAC806.02c 0,10 0,23 -0,14 Q9UT59 Putative uncharacterized oxidoreductase C513.07 SPAC513.07 0,08 1,59 -1,51 Q9UT74 Homoaconitase, mitochondrial lys2 0,04 -0,20 0,24 Q9UT76 Probable translation initiation factor eIF-2B subunit beta tif222 -0,17 0,14 -0,31 Q9UT81 UBX domain-containing protein 3 ubx3 -0,16 0,83 -0,99 Q9UT85 Uncharacterized WD repeat-containing protein C343.04c SPAC343.04c 0,12 0,41 -0,29 Q9UT87 54S ribosomal protein subunit img1, mitochondrial img1 -0,01 0,02 -0,03 Q9UT88 Probable phosphomevalonate kinase erg8 -0,05 0,05 -0,10 Q9UT95 Uncharacterized ABC transporter ATP-binding protein C323.04 SPAC323.04 -0,19 0,41 -0,60 Q9UT97 Probable proteasome subunit alpha type-5 SPAC323.02c -0,24 0,16 -0,41 Q9UTA6 Uncharacterized hydrolase C25B8.12c SPAC25B8.12c -0,12 0,06 -0,18 Q9UTA8 Uncharacterized methyltransferase-like C25B8.10 SPAC25B8.10 0,05 0,29 -0,24 Q9UTA9 Uncharacterized methyltransferase C25B8.09 SPAC25B8.09 0,38 0,37 0,01 Q9UTC6 Iron sulfur cluster assembly protein 1, mitochondrial isu1 -0,16 0,27 -0,43 Q9UTC9 Probable prefoldin subunit 2 SPAC227.10 0,23 0,28 -0,05 Q9UTD7 Ribosomal RNA-processing protein 15 rrp15 -0,18 0,13 -0,30
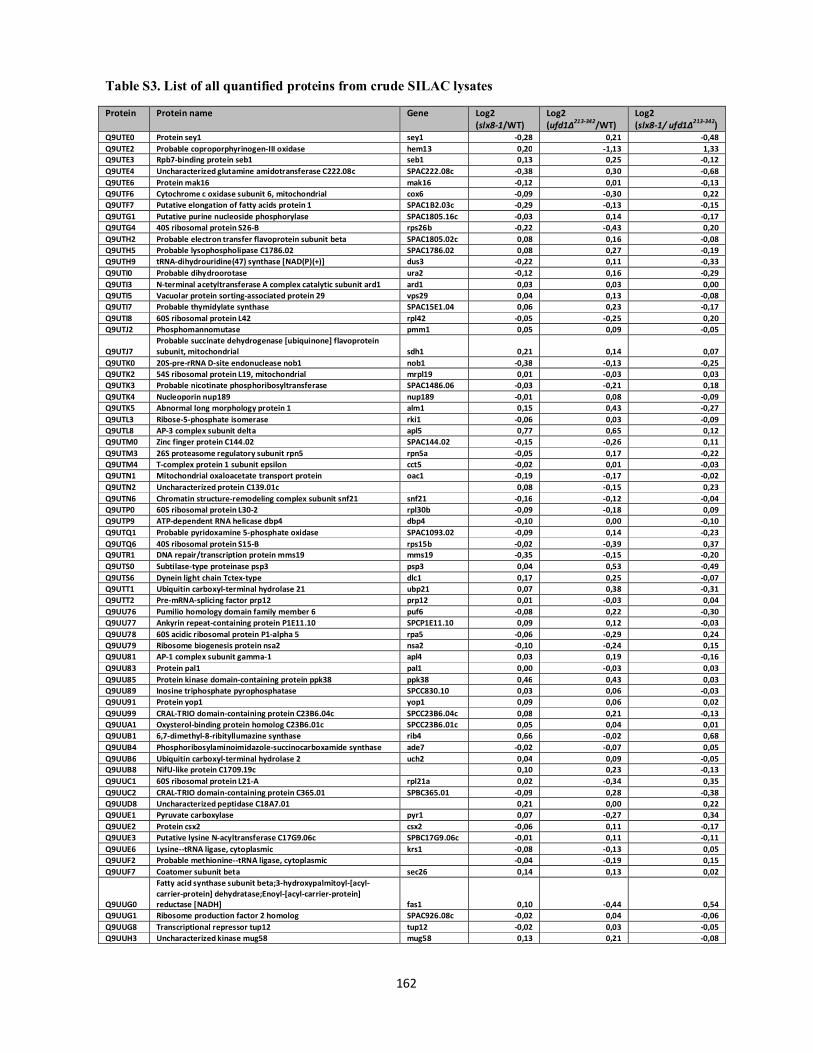
Table S3. List of all quantified proteins from crude SILAC lysates
162
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
Q9UTE0 Protein sey1 sey1 -0,28 0,21 -0,48 Q9UTE2 Probable coproporphyrinogen-III oxidase hem13 0,20 -1,13 1,33 Q9UTE3 Rpb7-binding protein seb1 seb1 0,13 0,25 -0,12 Q9UTE4 Uncharacterized glutamine amidotransferase C222.08c SPAC222.08c -0,38 0,30 -0,68 Q9UTE6 Protein mak16 mak16 -0,12 0,01 -0,13 Q9UTF6 Cytochrome c oxidase subunit 6, mitochondrial cox6 -0,09 -0,30 0,22 Q9UTF7 Putative elongation of fatty acids protein 1 SPAC1B2.03c -0,29 -0,13 -0,15 Q9UTG1 Putative purine nucleoside phosphorylase SPAC1805.16c -0,03 0,14 -0,17 Q9UTG4 40S ribosomal protein S26-B rps26b -0,22 -0,43 0,20 Q9UTH2 Probable electron transfer flavoprotein subunit beta SPAC1805.02c 0,08 0,16 -0,08 Q9UTH5 Probable lysophospholipase C1786.02 SPAC1786.02 0,08 0,27 -0,19 Q9UTH9 tRNA-dihydrouridine(47) synthase [NAD(P)(+)] dus3 -0,22 0,11 -0,33 Q9UTI0 Probable dihydroorotase ura2 -0,12 0,16 -0,29 Q9UTI3 N-terminal acetyltransferase A complex catalytic subunit ard1 ard1 0,03 0,03 0,00 Q9UTI5 Vacuolar protein sorting-associated protein 29 vps29 0,04 0,13 -0,08 Q9UTI7 Probable thymidylate synthase SPAC15E1.04 0,06 0,23 -0,17 Q9UTI8 60S ribosomal protein L42 rpl42 -0,05 -0,25 0,20 Q9UTJ2 Phosphomannomutase pmm1 0,05 0,09 -0,05
Q9UTJ7 Probable succinate dehydrogenase [ubiquinone] flavoprotein subunit, mitochondrial sdh1 0,21 0,14 0,07
Q9UTK0 20S-pre-rRNA D-site endonuclease nob1 nob1 -0,38 -0,13 -0,25 Q9UTK2 54S ribosomal protein L19, mitochondrial mrpl19 0,01 -0,03 0,03 Q9UTK3 Probable nicotinate phosphoribosyltransferase SPAC1486.06 -0,03 -0,21 0,18 Q9UTK4 Nucleoporin nup189 nup189 -0,01 0,08 -0,09 Q9UTK5 Abnormal long morphology protein 1 alm1 0,15 0,43 -0,27 Q9UTL3 Ribose-5-phosphate isomerase rki1 -0,06 0,03 -0,09 Q9UTL8 AP-3 complex subunit delta apl5 0,77 0,65 0,12 Q9UTM0 Zinc finger protein C144.02 SPAC144.02 -0,15 -0,26 0,11 Q9UTM3 26S proteasome regulatory subunit rpn5 rpn5a -0,05 0,17 -0,22 Q9UTM4 T-complex protein 1 subunit epsilon cct5 -0,02 0,01 -0,03 Q9UTN1 Mitochondrial oxaloacetate transport protein oac1 -0,19 -0,17 -0,02 Q9UTN2 Uncharacterized protein C139.01c 0,08 -0,15 0,23 Q9UTN6 Chromatin structure-remodeling complex subunit snf21 snf21 -0,16 -0,12 -0,04 Q9UTP0 60S ribosomal protein L30-2 rpl30b -0,09 -0,18 0,09 Q9UTP9 ATP-dependent RNA helicase dbp4 dbp4 -0,10 0,00 -0,10 Q9UTQ1 Probable pyridoxamine 5-phosphate oxidase SPAC1093.02 -0,09 0,14 -0,23 Q9UTQ6 40S ribosomal protein S15-B rps15b -0,02 -0,39 0,37 Q9UTR1 DNA repair/transcription protein mms19 mms19 -0,35 -0,15 -0,20 Q9UTS0 Subtilase-type proteinase psp3 psp3 0,04 0,53 -0,49 Q9UTS6 Dynein light chain Tctex-type dlc1 0,17 0,25 -0,07 Q9UTT1 Ubiquitin carboxyl-terminal hydrolase 21 ubp21 0,07 0,38 -0,31 Q9UTT2 Pre-mRNA-splicing factor prp12 prp12 0,01 -0,03 0,04 Q9UU76 Pumilio homology domain family member 6 puf6 -0,08 0,22 -0,30 Q9UU77 Ankyrin repeat-containing protein P1E11.10 SPCP1E11.10 0,09 0,12 -0,03 Q9UU78 60S acidic ribosomal protein P1-alpha 5 rpa5 -0,06 -0,29 0,24 Q9UU79 Ribosome biogenesis protein nsa2 nsa2 -0,10 -0,24 0,15 Q9UU81 AP-1 complex subunit gamma-1 apl4 0,03 0,19 -0,16 Q9UU83 Protein pal1 pal1 0,00 -0,03 0,03 Q9UU85 Protein kinase domain-containing protein ppk38 ppk38 0,46 0,43 0,03 Q9UU89 Inosine triphosphate pyrophosphatase SPCC830.10 0,03 0,06 -0,03 Q9UU91 Protein yop1 yop1 0,09 0,06 0,02 Q9UU99 CRAL-TRIO domain-containing protein C23B6.04c SPCC23B6.04c 0,08 0,21 -0,13 Q9UUA1 Oxysterol-binding protein homolog C23B6.01c SPCC23B6.01c 0,05 0,04 0,01 Q9UUB1 6,7-dimethyl-8-ribityllumazine synthase rib4 0,66 -0,02 0,68 Q9UUB4 Phosphoribosylaminoimidazole-succinocarboxamide synthase ade7 -0,02 -0,07 0,05 Q9UUB6 Ubiquitin carboxyl-terminal hydrolase 2 uch2 0,04 0,09 -0,05 Q9UUB8 NifU-like protein C1709.19c 0,10 0,23 -0,13 Q9UUC1 60S ribosomal protein L21-A rpl21a 0,02 -0,34 0,35 Q9UUC2 CRAL-TRIO domain-containing protein C365.01 SPBC365.01 -0,09 0,28 -0,38 Q9UUD8 Uncharacterized peptidase C18A7.01 0,21 0,00 0,22 Q9UUE1 Pyruvate carboxylase pyr1 0,07 -0,27 0,34 Q9UUE2 Protein csx2 csx2 -0,06 0,11 -0,17 Q9UUE3 Putative lysine N-acyltransferase C17G9.06c SPBC17G9.06c -0,01 0,11 -0,11 Q9UUE6 Lysine--tRNA ligase, cytoplasmic krs1 -0,08 -0,13 0,05 Q9UUF2 Probable methionine--tRNA ligase, cytoplasmic -0,04 -0,19 0,15 Q9UUF7 Coatomer subunit beta sec26 0,14 0,13 0,02
Q9UUG0
Fatty acid synthase subunit beta;3-hydroxypalmitoyl-[acyl-carrier-protein] dehydratase;Enoyl-[acyl-carrier-protein] reductase [NADH] fas1 0,10 -0,44 0,54
Q9UUG1 Ribosome production factor 2 homolog SPAC926.08c -0,02 0,04 -0,06 Q9UUG8 Transcriptional repressor tup12 tup12 -0,02 0,03 -0,05 Q9UUH3 Uncharacterized kinase mug58 mug58 0,13 0,21 -0,08
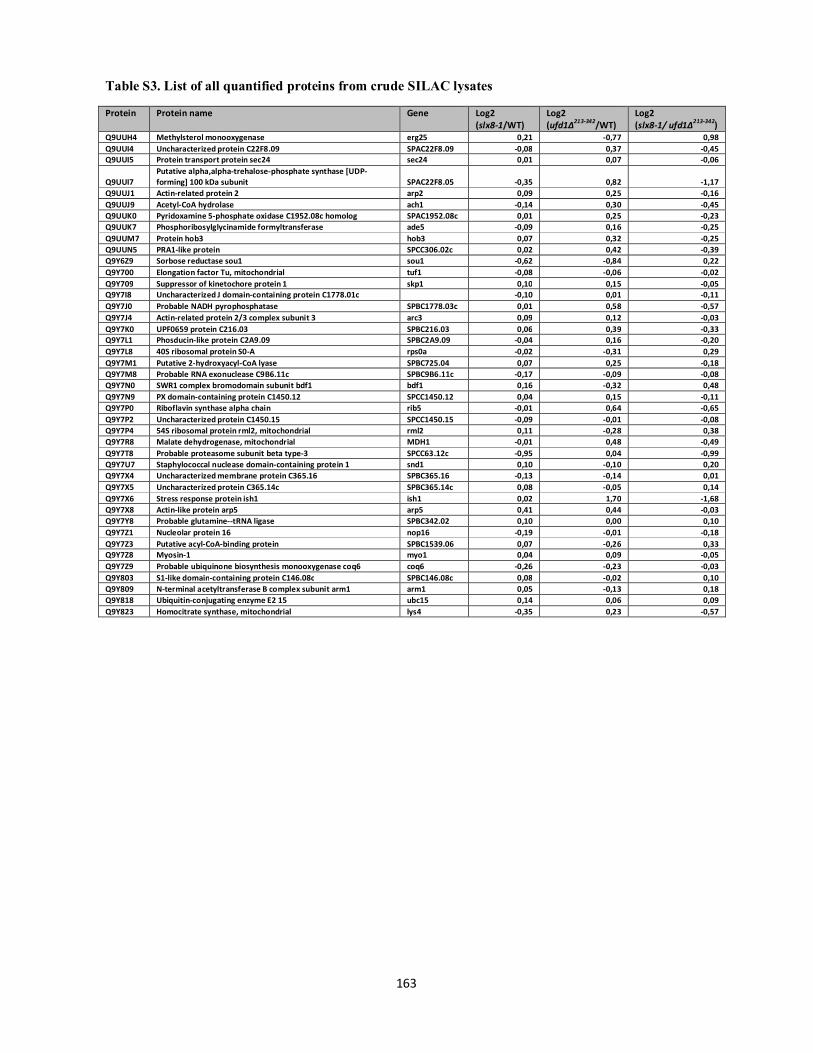
Table S3. List of all quantified proteins from crude SILAC lysates
163
Protein Protein name Gene Log2 (slx8-1/WT)
Log2 (ufd1∆213-342/WT)
Log2 (slx8-1/ ufd1∆213-342)
Q9UUH4 Methylsterol monooxygenase erg25 0,21 -0,77 0,98 Q9UUI4 Uncharacterized protein C22F8.09 SPAC22F8.09 -0,08 0,37 -0,45 Q9UUI5 Protein transport protein sec24 sec24 0,01 0,07 -0,06
Q9UUI7 Putative alpha,alpha-trehalose-phosphate synthase [UDP-forming] 100 kDa subunit SPAC22F8.05 -0,35 0,82 -1,17
Q9UUJ1 Actin-related protein 2 arp2 0,09 0,25 -0,16 Q9UUJ9 Acetyl-CoA hydrolase ach1 -0,14 0,30 -0,45 Q9UUK0 Pyridoxamine 5-phosphate oxidase C1952.08c homolog SPAC1952.08c 0,01 0,25 -0,23 Q9UUK7 Phosphoribosylglycinamide formyltransferase ade5 -0,09 0,16 -0,25 Q9UUM7 Protein hob3 hob3 0,07 0,32 -0,25 Q9UUN5 PRA1-like protein SPCC306.02c 0,02 0,42 -0,39 Q9Y6Z9 Sorbose reductase sou1 sou1 -0,62 -0,84 0,22 Q9Y700 Elongation factor Tu, mitochondrial tuf1 -0,08 -0,06 -0,02 Q9Y709 Suppressor of kinetochore protein 1 skp1 0,10 0,15 -0,05 Q9Y7I8 Uncharacterized J domain-containing protein C1778.01c -0,10 0,01 -0,11 Q9Y7J0 Probable NADH pyrophosphatase SPBC1778.03c 0,01 0,58 -0,57 Q9Y7J4 Actin-related protein 2/3 complex subunit 3 arc3 0,09 0,12 -0,03 Q9Y7K0 UPF0659 protein C216.03 SPBC216.03 0,06 0,39 -0,33 Q9Y7L1 Phosducin-like protein C2A9.09 SPBC2A9.09 -0,04 0,16 -0,20 Q9Y7L8 40S ribosomal protein S0-A rps0a -0,02 -0,31 0,29 Q9Y7M1 Putative 2-hydroxyacyl-CoA lyase SPBC725.04 0,07 0,25 -0,18 Q9Y7M8 Probable RNA exonuclease C9B6.11c SPBC9B6.11c -0,17 -0,09 -0,08 Q9Y7N0 SWR1 complex bromodomain subunit bdf1 bdf1 0,16 -0,32 0,48 Q9Y7N9 PX domain-containing protein C1450.12 SPCC1450.12 0,04 0,15 -0,11 Q9Y7P0 Riboflavin synthase alpha chain rib5 -0,01 0,64 -0,65 Q9Y7P2 Uncharacterized protein C1450.15 SPCC1450.15 -0,09 -0,01 -0,08 Q9Y7P4 54S ribosomal protein rml2, mitochondrial rml2 0,11 -0,28 0,38 Q9Y7R8 Malate dehydrogenase, mitochondrial MDH1 -0,01 0,48 -0,49 Q9Y7T8 Probable proteasome subunit beta type-3 SPCC63.12c -0,95 0,04 -0,99 Q9Y7U7 Staphylococcal nuclease domain-containing protein 1 snd1 0,10 -0,10 0,20 Q9Y7X4 Uncharacterized membrane protein C365.16 SPBC365.16 -0,13 -0,14 0,01 Q9Y7X5 Uncharacterized protein C365.14c SPBC365.14c 0,08 -0,05 0,14 Q9Y7X6 Stress response protein ish1 ish1 0,02 1,70 -1,68 Q9Y7X8 Actin-like protein arp5 arp5 0,41 0,44 -0,03 Q9Y7Y8 Probable glutamine--tRNA ligase SPBC342.02 0,10 0,00 0,10 Q9Y7Z1 Nucleolar protein 16 nop16 -0,19 -0,01 -0,18 Q9Y7Z3 Putative acyl-CoA-binding protein SPBC1539.06 0,07 -0,26 0,33 Q9Y7Z8 Myosin-1 myo1 0,04 0,09 -0,05 Q9Y7Z9 Probable ubiquinone biosynthesis monooxygenase coq6 coq6 -0,26 -0,23 -0,03 Q9Y803 S1-like domain-containing protein C146.08c SPBC146.08c 0,08 -0,02 0,10 Q9Y809 N-terminal acetyltransferase B complex subunit arm1 arm1 0,05 -0,13 0,18 Q9Y818 Ubiquitin-conjugating enzyme E2 15 ubc15 0,14 0,06 0,09 Q9Y823 Homocitrate synthase, mitochondrial lys4 -0,35 0,23 -0,57

164
A final discussion
Covalent attachment of SUMO to cellular proteins constitutes a widespread mechanism for rapidly
and reversibly regulating protein function in response to a changing cellular environment. Thus,
gaining insight into the triggers and downstream consequences of sumoylation is important for
understanding many cellular functions. Similarly, gaining insight into the mechanisms responsible
for reversing SUMO modifications lies at the core of our understanding of the dynamics of SUMO
signaling events.
Within this work we have provided evidence for a role of the Ufd1 protein, and by extension of the
Cdc48-Ufd1-Npl4 complex, in the downstream processing of sumoylated proteins. In particular, our
work has revealed both physical and functional links between Ufd1 and the STUbL enzymes, which
by ubiquitylating sumoylated proteins mediate their proteolytic turnover (Praefcke et al, 2012;
Sriramachandran & Dohmen, 2014). Ufd1 is a well-described ubiquitin-binding protein that
together with Npl4 forms an essential co-factor to the Cdc48/p97 ATPase. Originally best
understood for its role in ERAD (endoplasmic reticulum-associated degradation), the Cdc48-Ufd1-
Npl4 complex has later been implicated much more globally in the regulation of ubiquitin-related
processes by coupling ubiquitin-binding with ATPase-driven molecular tension. Cdc48-Ufd1-Npl4
activity is in this way believed to enable the mobilization of ubiquitylated proteins destined for
degradation or possibly other fates (Meyer et al, 2012; Stolz et al, 2011). In parallel with our studies
were reports identifying a SIM motif close to the C-terminus of both fission and budding yeast
Ufd1, revealing the ability of Cdc48-Ufd1-Npl4 to also act on sumoylated proteins (Bergink et al,
2013; Nie et al, 2012). Cdc48 SIMs can also be identified, although a direct SUMO interaction has
so-far only been demonstrated in S. cerevisae (Bergink et al, 2013; Hannich et al, 2005). Indeed
budding yeast Cdc48 and Ufd1 have both been shown to interact with the homologous
recombination protein Rad52 in a sumoylation-dependent manner and to dissociate Rad52 from its
Rad51 binding partner and from DNA (Bergink et al, 2013). Although not investigated in the
Bergnik et al. study, the presence of both SUMO and ubiquitin-binding motifs within Ufd1 suggests
that these domains may mediate cooperative binding of Cdc48-Ufd1-Npl4 complex to SUMO-
ubiquitin co-modified STUbL substrates (Nie et al, 2012). More specific physical studies should
elucidate such a potential dual binding mode. Nevertheless, genetic analysis of the fission yeast
Ufd1 SIM found that while it is not required for growth under normal circumstances, the ability of
Ufd1 to interact with SUMO becomes essential in the absence of STUbL activity (slx8-29) (Nie et
al, 2012). This could support the idea that Ufd1 is redundantly targeted to STUbL substrates via

165
bipartite SUMO and ubiquitin recognition; i.e. in the absence of target ubiquitylation, the SUMO-
SIM interaction becomes critical. Another interpretation of the slx8-29 ufd1SIM∆ genetic
relationship would be that the two proteins operate in separate pathways to downregulate SUMO
conjugates. Cdc48-Ufd1-Npl4 may simply disassemble sumoylated proteins from protein
complexes to allow their subsequent demodification or potential proteolytic targeting through
alternative Cdc48-associated ubiquitin ligases (Jentsch & Psakhye, 2013). The physical associations
between STUbL and Cdc48-Ufd1-Npl4 complex components identified in this study support at
least some cooperative functions. Coordinated roles of STUbL and Cdc48-Ufd1-Npl4 are likely to
also involve other factors. This may for example include the SUMO specific isopeptidase Wss1
which associates with Cdc48 through both VIM and SHP box motifs (Hitt & Wolf, 2004; Stapf et
al, 2011). Interestingly, Wss1 also associates with proteasomal subunits and it is able to deconjugate
SUMO-ubiquitin hybrid species (Mullen et al, 2010). Together, these observations single out Wss1
as a potential regulator of STUbL substrate fate that may determine whether STUbL targets are
channeled to the proteasome or allowed to escape. In addition, Wss1 might downregulate poly-
sumoylated species in parallel to the STUbL pathway (Mullen et al, 2011). Future studies should
elucidate how STUbL and Wss1 activities may be integrated through Cdc48-Ufd1-Npl4. As Wss1
has been found required for the viability of slx5∆ sgs1∆ ulp2∆ S. cervisae cells, possibly by
allowing the turnover of the poly-sumoylated species accumulating in this background (Mullen et
al, 2011), it should be interesting to investigate whether this pathway also involves Cdc48-Ufd1-
Npl4 via the Ufd1 SIM.
In line with accumulating evidence that the Cdc48-Ufd1-Npl4 complex mediates specific processes
in chromatin, we found that the fission yeast ufd1ΔCt213-342 mutant displays several phenotypes
associated with genomic instability. While these phenotypes are likely to reflect an abrogation of
several Cdc48-Ufd1-Npl4 mediated cellular effects, our data suggest the HR protein Rad22 (Rad52
homologue) as one potential Cdc48-Ufd1-Npl4 candidate substrate accumulating in the ufd1ΔCt213-
342 background. As proposed by Bergnik et al., Cdc48-Ufd1-Npl4 may “curb” the SUMO-SIM
interaction between sumoylated Rad52 and Rad51 to antagonize HR events (Bergink et al, 2013).
Evidence from S. pombe has furthermore shown that Rad22 levels and hence HR repair are directly
downregulated through a proteasomal pathway involving the proteasome associated Bag101 protein
(Saito et al, 2013). Interestingly, Bag101 has been identified in a screen for proteins interacting
non-covalently with SUMO (Nie et al, 2012), though the functional implications of this interaction
have not been further investigated. Nevertheless, together these findings highlight different modes

166
of controlling the association of Rad22 with DNA that may possibly be connected to fine-tune the
specific pool of Rad22/Rad52 engaged in HR repair. Involvement of STUbLs in such Rad22/Rad52
regulation is also possible since, Rad52 is an in vitro STUbL substrate (Ii et al, 2007; Xie et al,
2007).
To unbiasedly shed light on the sumoylated proteins whose processing depends on STUbL and/or
Ufd1 activities we analyzed site-specific changes in the sumoylated proteome between wild type
cells, ufd1ΔCt213-342and slx8-1 STUbL mutants using SILAC-based quantitative proteomics. This
screen revealed a number of putative STUbL and Ufd1 candidate substrates, including SUMO
targets that may be regulated through a common STUbL/Ufd1 pathway. The power of our approach
was underscored by the fact that we identified an already known STUbL target as well as proteins
forming part of complexes previously shown to be affected by STUbL activity. In particular, our
findings indicate potential common roles for STUbLs and Ufd1 at centromeres and telomeres. More
detailed investigations of these SUMO substrates may provide exciting insight into how STUbL and
Cdc48-Ufd1-Npl4 activities are coordinated to ensure proper centromere and telomere function.
Interestingly, our screen identified the Mot1 transcriptional regulator as an upregulated SUMO
target in both slx8-1 and ufd1ΔCt213-342. A temperature sensitive version of the Mot1 protein (Mot-
301) has previously been shown to undergo accelerated proteolysis through a SUMO and STUbL-
dependent pathway, suggesting a protein quality control mechanism (Wang & Prelich, 2009).
Sumoylated forms of even the wild type Mot1 protein accumulates in STUbL mutant backgrounds,
indicating some wild type Mot1 products also being subject to STUbL regulation. These could
represent some misfolded or damaged forms of the proteins as suggested by Wang et al., 2009. Our
data suggests that Ufd1 may participate in the Mot1 regulatory pathway. Several lines of evidence
have suggested roles for sumoylation in protein quality control. In particular, the levels of SUMO
conjugates are seen to dramatically increase in response to protein damaging stress such as heat or
H2O2 (Golebiowski et al, 2009; Saitoh & Hinchey, 2000). Sumoylation levels also increase in
response to proteasomal inhibition. As blocking protein synthesis abolishes this increase in
sumoylated species, it has been suggested that the sumoylation response to proteasomal stress is due
to an accumulation of newly synthesized misfolded proteins (Castoralova et al, 2012; Tatham et al,
2011). It seems that at least some of the stress-responsive SUMO targets are misfolded proteins
themselves (Castoralova et al, 2012). Protein damage such as caused by external stress or intrinsic
translation/folding errors could be imagined to cause an increase in the sumoylation of some targets
due to the exposure of modification sites normally inaccessible for the conjugation enzymes. It is

167
possible that such increased sumoylation levels may in turn stimulate their recognition and
degradation through the STUbL enzymes. Protein quality control pathways play pivotal roles in
ensuring protein homeostasis both in the cytosol and in the nucleus involving different ubiquitin
ligases and molecular chaperones (Amm et al, 2014; Nielsen et al, 2014). It has been shown that
newly synthesized proteins are more fragile to proteotoxic stress presumably until they have
obtained a more stable conformation or been stably bound to interaction partners (Medicherla &
Goldberg, 2008). The Cdc48-Ufd1-Npl4 complex has previously been implicated in the ubiquitin-
proteasome mediated turn-over of some newly synthesized misfolded proteins (Medicherla &
Goldberg, 2008). If such species are allowed to accumulate in ufd1DCt cells, this could potentially
trigger a sumoylation response. Clearly future experiments should clarify the functional
implications of the increased sumoylation levels observed in the slx8-1 and ufd1ΔCt213-342
backgrounds. Moreover it should be interesting to gain deeper insight into the relationship between
protein sumoylation, proteasomal targeting and misfolded protein responses. In the case for Mot1
for example, it could be that in addition to targeting the protein for degradation, sumoylation would
under normal “undamaged” conditions work to regulate other aspects of its function. For instance
some evidence suggests that its modification may be specifically stimulated upon binding to its
TBP (TATA-binding protein) interaction partner (Wang & Prelich, 2009). This could indicate that
Mot1 sumoylation may work to regulate some aspects of TBP-mediated transcription.
Among other proteins with upregulated sumoylation in both slx8-1 and ufd1ΔCt213-342, which are
poorly discussed in the accompanying manuscript, was the Ufd1 interactor Cdc48. Cdc48 is a
previously reported SUMO substrate (Makhnevych et al, 2009). We speculate that an increase in
Cdc48 sumoylation could reflect enhanced recruitment of Cdc48 to the sumoylated substrate whose
processing is defective in the slx8-1 and ufd1ΔCt213-342backgrounds. Recruitment of Cdc48 to sites
of active sumoylation may result in elevated modification of Cdc48 itself or alternatively, Cdc48
sumoylation may work to regulate its sub-cellular recruitment to SUMO targets irrespective of its
Ufd1 co-factor.

168
Of note, Rad22 was not identified in our mass-spectrometry screen for sumoylated proteins. As
Rad22 sumoylation seems to be specifically induced upon DNA damage (Ho et al, 2001; Sacher et
al, 2006), its sumoylated levels in unperturbed cells are likely to be too low to be detected among
more abundant SUMO substrates. Moreover, Rad22 is by itself a low-abundance protein (Gunaratne
et al, 2013). Future studies should seek SUMO conjugates potentially regulated by
STUbL/Ufd1during the DNA damage response by investigating quantitative difference in
sumoylation between wild-type and mutant strains after DNA damage. Other stresses and mutant
backgrounds will also be amenable to large-scale analyses through the method we employed.

169
References al-Khodairy F, Enoch T, Hagan IM, Carr AM (1995) The Schizosaccharomyces pombe hus5 gene encodes a ubiquitin conjugating enzyme required for normal mitosis. Journal of cell science 108 ( Pt 2): 475-486 Alexandru G, Graumann J, Smith GT, Kolawa NJ, Fang R, Deshaies RJ (2008) UBXD7 binds multiple ubiquitin ligases and implicates p97 in HIF1alpha turnover. Cell 134: 804-816 Altelaar AF, Munoz J, Heck AJ (2013) Next-generation proteomics: towards an integrative view of proteome dynamics. Nature reviews Genetics 14: 35-48 Amm I, Sommer T, Wolf DH (2014) Protein quality control and elimination of protein waste: the role of the ubiquitin-proteasome system. Biochimica et biophysica acta 1843: 182-196 Andrews EA, Palecek J, Sergeant J, Taylor E, Lehmann AR, Watts FZ (2005) Nse2, a component of the Smc5-6 complex, is a SUMO ligase required for the response to DNA damage. Molecular and cellular biology 25: 185-196 Armstrong AA, Mohideen F, Lima CD (2012) Recognition of SUMO-modified PCNA requires tandem receptor motifs in Srs2. Nature 483: 59-63 Ashton TM, Hickson ID (2010) Yeast as a model system to study RecQ helicase function. DNA repair 9: 303-314 Bachant J, Alcasabas A, Blat Y, Kleckner N, Elledge SJ (2002) The SUMO-1 isopeptidase Smt4 is linked to centromeric cohesion through SUMO-1 modification of DNA topoisomerase II. Molecular cell 9: 1169-1182 Baek GH, Cheng H, Choe V, Bao X, Shao J, Luo S, Rao H (2013) Cdc48: a swiss army knife of cell biology. Journal of amino acids 2013: 183421 Bailey D, O'Hare P (2005) Comparison of the SUMO1 and ubiquitin conjugation pathways during the inhibition of proteasome activity with evidence of SUMO1 recycling. The Biochemical journal 392: 271-281 Ballar P, Shen Y, Yang H, Fang S (2006) The role of a novel p97/valosin-containing protein-interacting motif of gp78 in endoplasmic reticulum-associated degradation. The Journal of biological chemistry 281: 35359-35368 Barysch SV, Dittner C, Flotho A, Becker J, Melchior F (2014) Identification and analysis of endogenous SUMO1 and SUMO2/3 targets in mammalian cells and tissues using monoclonal antibodies. Nature protocols 9: 896-909 Bayer P, Arndt A, Metzger S, Mahajan R, Melchior F, Jaenicke R, Becker J (1998) Structure determination of the small ubiquitin-related modifier SUMO-1. Journal of molecular biology 280: 275-286 Bays NW, Wilhovsky SK, Goradia A, Hodgkiss-Harlow K, Hampton RY (2001) HRD4/NPL4 is required for the proteasomal processing of ubiquitinated ER proteins. Molecular biology of the cell 12: 4114-4128 Bekes M, Prudden J, Srikumar T, Raught B, Boddy MN, Salvesen GS (2011) The dynamics and mechanism of SUMO chain deconjugation by SUMO-specific proteases. The Journal of biological chemistry 286: 10238-10247 Benson FE, Baumann P, West SC (1998) Synergistic actions of Rad51 and Rad52 in recombination and DNA repair. Nature 391: 401-404 Bergink S, Ammon T, Kern M, Schermelleh L, Leonhardt H, Jentsch S (2013) Role of Cdc48/p97 as a SUMO-targeted segregase curbing Rad51-Rad52 interaction. Nature cell biology 15: 526-532 Bernier-Villamor V, Sampson DA, Matunis MJ, Lima CD (2002) Structural basis for E2-mediated SUMO conjugation revealed by a complex between ubiquitin-conjugating enzyme Ubc9 and RanGAP1. Cell 108: 345-356 Beskow A, Grimberg KB, Bott LC, Salomons FA, Dantuma NP, Young P (2009) A conserved unfoldase activity for the p97 AAA-ATPase in proteasomal degradation. Journal of molecular biology 394: 732-746

170
Blastyak A, Pinter L, Unk I, Prakash L, Prakash S, Haracska L (2007) Yeast Rad5 protein required for postreplication repair has a DNA helicase activity specific for replication fork regression. Molecular cell 28: 167-175 Boddy MN, Furnari B, Mondesert O, Russell P (1998) Replication checkpoint enforced by kinases Cds1 and Chk1. Science (New York, NY 280: 909-912 Boddy MN, Gaillard PH, McDonald WH, Shanahan P, Yates JR, 3rd, Russell P (2001) Mus81-Eme1 are essential components of a Holliday junction resolvase. Cell 107: 537-548 Boddy MN, Howe K, Etkin LD, Solomon E, Freemont PS (1996) PIC 1, a novel ubiquitin-like protein which interacts with the PML component of a multiprotein complex that is disrupted in acute promyelocytic leukaemia. Oncogene 13: 971-982 Boeddrich A, Gaumer S, Haacke A, Tzvetkov N, Albrecht M, Evert BO, Muller EC, Lurz R, Breuer P, Schugardt N, Plassmann S, Xu K, Warrick JM, Suopanki J, Wullner U, Frank R, Hartl UF, Bonini NM, Wanker EE (2006) An arginine/lysine-rich motif is crucial for VCP/p97-mediated modulation of ataxin-3 fibrillogenesis. The EMBO journal 25: 1547-1558 Branzei D (2011) Ubiquitin family modifications and template switching. FEBS letters 585: 2810-2817 Branzei D, Sollier J, Liberi G, Zhao X, Maeda D, Seki M, Enomoto T, Ohta K, Foiani M (2006) Ubc9- and mms21-mediated sumoylation counteracts recombinogenic events at damaged replication forks. Cell 127: 509-522 Branzei D, Vanoli F, Foiani M (2008) SUMOylation regulates Rad18-mediated template switch. Nature 456: 915-920 Broomfield S, Chow BL, Xiao W (1998) MMS2, encoding a ubiquitin-conjugating-enzyme-like protein, is a member of the yeast error-free postreplication repair pathway. Proceedings of the National Academy of Sciences of the United States of America 95: 5678-5683 Brown M, Zhu Y, Hemmingsen SM, Xiao W (2002) Structural and functional conservation of error-free DNA postreplication repair in Schizosaccharomyces pombe. DNA repair 1: 869-880 Bruderer R, Tatham MH, Plechanovova A, Matic I, Garg AK, Hay RT (2011) Purification and identification of endogenous polySUMO conjugates. EMBO reports 12: 142-148 Bruderer RM, Brasseur C, Meyer HH (2004) The AAA ATPase p97/VCP interacts with its alternative co-factors, Ufd1-Npl4 and p47, through a common bipartite binding mechanism. The Journal of biological chemistry 279: 49609-49616 Buchberger A (2010) Control of ubiquitin conjugation by cdc48 and its cofactors. Sub-cellular biochemistry 54: 17-30 Bug M, Meyer H (2012) Expanding into new markets--VCP/p97 in endocytosis and autophagy. Journal of structural biology 179: 78-82 Burgess RC, Rahman S, Lisby M, Rothstein R, Zhao X (2007) The Slx5-Slx8 complex affects sumoylation of DNA repair proteins and negatively regulates recombination. Molecular and cellular biology 27: 6153-6162 Buschmann T, Fuchs SY, Lee CG, Pan ZQ, Ronai Z (2000) SUMO-1 modification of Mdm2 prevents its self-ubiquitination and increases Mdm2 ability to ubiquitinate p53. Cell 101: 753-762 Bylebyl GR, Belichenko I, Johnson ES (2003) The SUMO isopeptidase Ulp2 prevents accumulation of SUMO chains in yeast. The Journal of biological chemistry 278: 44113-44120 Candelli A, Modesti M, Peterman EJ, Wuite GJ (2013) Single-molecule views on homologous recombination. Quarterly reviews of biophysics 46: 323-348 Cao K, Nakajima R, Meyer HH, Zheng Y (2003) The AAA-ATPase Cdc48/p97 regulates spindle disassembly at the end of mitosis. Cell 115: 355-367

171
Capili AD, Lima CD (2007) Structure and analysis of a complex between SUMO and Ubc9 illustrates features of a conserved E2-Ubl interaction. Journal of molecular biology 369: 608-618 Castoralova M, Brezinova D, Sveda M, Lipov J, Ruml T, Knejzlik Z (2012) SUMO-2/3 conjugates accumulating under heat shock or MG132 treatment result largely from new protein synthesis. Biochimica et biophysica acta 1823: 911-919 Cheng CH, Lo YH, Liang SS, Ti SC, Lin FM, Yeh CH, Huang HY, Wang TF (2006) SUMO modifications control assembly of synaptonemal complex and polycomplex in meiosis of Saccharomyces cerevisiae. Genes & development 20: 2067-2081 Choudhary C, Kumar C, Gnad F, Nielsen ML, Rehman M, Walther TC, Olsen JV, Mann M (2009) Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science (New York, NY 325: 834-840 Choudhary C, Mann M (2010) Decoding signalling networks by mass spectrometry-based proteomics. Nature reviews 11: 427-439 Cook CE, Hochstrasser M, Kerscher O (2009) The SUMO-targeted ubiquitin ligase subunit Slx5 resides in nuclear foci and at sites of DNA breaks. Cell cycle (Georgetown, Tex 8: 1080-1089 Cox J, Mann M (2011) Quantitative, high-resolution proteomics for data-driven systems biology. Annual review of biochemistry 80: 273-299 Cremona CA, Sarangi P, Yang Y, Hang LE, Rahman S, Zhao X (2012) Extensive DNA damage-induced sumoylation contributes to replication and repair and acts in addition to the mec1 checkpoint. Molecular cell 45: 422-432 Cuanalo-Contreras K, Mukherjee A, Soto C (2013) Role of protein misfolding and proteostasis deficiency in protein misfolding diseases and aging. International journal of cell biology 2013: 638083 Cubenas-Potts C, Goeres JD, Matunis MJ (2013) SENP1 and SENP2 affect spatial and temporal control of sumoylation in mitosis. Molecular biology of the cell 24: 3483-3495 Cubenas-Potts C, Matunis MJ (2013) SUMO: a multifaceted modifier of chromatin structure and function. Developmental cell 24: 1-12 Da Silva-Ferrada E, Xolalpa W, Lang V, Aillet F, Martin-Ruiz I, de la Cruz-Herrera CF, Lopitz-Otsoa F, Carracedo A, Goldenberg SJ, Rivas C, England P, Rodriguez MS (2013) Analysis of SUMOylated proteins using SUMO-traps. Scientific reports 3: 1690 Dammer EB, Na CH, Xu P, Seyfried NT, Duong DM, Cheng D, Gearing M, Rees H, Lah JJ, Levey AI, Rush J, Peng J (2011) Polyubiquitin linkage profiles in three models of proteolytic stress suggest the etiology of Alzheimer disease. The Journal of biological chemistry 286: 10457-10465 DeHoratius C, Silver PA (1996) Nuclear transport defects and nuclear envelope alterations are associated with mutation of the Saccharomyces cerevisiae NPL4 gene. Molecular biology of the cell 7: 1835-1855 DeLaBarre B, Brunger AT (2003) Complete structure of p97/valosin-containing protein reveals communication between nucleotide domains. Nature structural biology 10: 856-863 Denuc A, Marfany G (2010) SUMO and ubiquitin paths converge. Biochemical Society transactions 38: 34-39 Desterro JM, Rodriguez MS, Hay RT (1998) SUMO-1 modification of IkappaBalpha inhibits NF-kappaB activation. Molecular cell 2: 233-239 Desterro JM, Rodriguez MS, Kemp GD, Hay RT (1999) Identification of the enzyme required for activation of the small ubiquitin-like protein SUMO-1. The Journal of biological chemistry 274: 10618-10624 Desterro JM, Thomson J, Hay RT (1997) Ubch9 conjugates SUMO but not ubiquitin. FEBS letters 417: 297-300

172
Devoy A, Soane T, Welchman R, Mayer RJ (2005) The ubiquitin-proteasome system and cancer. Essays in biochemistry 41: 187-203 Dikic I, Wakatsuki S, Walters KJ (2009) Ubiquitin-binding domains - from structures to functions. Nature reviews 10: 659-671 Ding F, Xiao H, Wang M, Xie X, Hu F (2014) The role of the ubiquitin-proteasome pathway in cancer development and treatment. Frontiers in bioscience 19: 886-895 Doe CL, Osman F, Dixon J, Whitby MC (2004) DNA repair by a Rad22-Mus81-dependent pathway that is independent of Rhp51. Nucleic acids research 32: 5570-5581 Elsasser S, Chandler-Militello D, Muller B, Hanna J, Finley D (2004) Rad23 and Rpn10 serve as alternative ubiquitin receptors for the proteasome. The Journal of biological chemistry 279: 26817-26822 Elsasser S, Finley D (2005) Delivery of ubiquitinated substrates to protein-unfolding machines. Nature cell biology 7: 742-749 Elsasser S, Gali RR, Schwickart M, Larsen CN, Leggett DS, Muller B, Feng MT, Tubing F, Dittmar GA, Finley D (2002) Proteasome subunit Rpn1 binds ubiquitin-like protein domains. Nature cell biology 4: 725-730 Erker Y, Neyret-Kahn H, Seeler JS, Dejean A, Atfi A, Levy L (2013) Arkadia, a novel SUMO-targeted ubiquitin ligase involved in PML degradation. Molecular and cellular biology 33: 2163-2177 Escobar-Cabrera E, Okon M, Lau DK, Dart CF, Bonvin AM, McIntosh LP (2011) Characterizing the N- and C-terminal Small ubiquitin-like modifier (SUMO)-interacting motifs of the scaffold protein DAXX. The Journal of biological chemistry 286: 19816-19829 Evdokimov E, Sharma P, Lockett SJ, Lualdi M, Kuehn MR (2008) Loss of SUMO1 in mice affects RanGAP1 localization and formation of PML nuclear bodies, but is not lethal as it can be compensated by SUMO2 or SUMO3. Journal of cell science 121: 4106-4113 Filosa G, Barabino SM, Bachi A (2013) Proteomics strategies to identify SUMO targets and acceptor sites: a survey of RNA-binding proteins SUMOylation. Neuromolecular medicine 15: 661-676 Finley D, Ulrich HD, Sommer T, Kaiser P (2012) The ubiquitin-proteasome system of Saccharomyces cerevisiae. Genetics 192: 319-360 Flotho A, Melchior F (2013) Sumoylation: a regulatory protein modification in health and disease. Annual review of biochemistry 82: 357-385 Flotho A, Werner A, Winter T, Frank AS, Ehret H, Melchior F (2012) Recombinant reconstitution of sumoylation reactions in vitro. Methods in molecular biology 832: 93-110 Frampton J, Irmisch A, Green CM, Neiss A, Trickey M, Ulrich HD, Furuya K, Watts FZ, Carr AM, Lehmann AR (2006) Postreplication repair and PCNA modification in Schizosaccharomyces pombe. Molecular biology of the cell 17: 2976-2985 Franz A, Ackermann L, Hoppe T (2014) Create and preserve: proteostasis in development and aging is governed by Cdc48/p97/VCP. Biochimica et biophysica acta 1843: 205-215 Fu X, Ng C, Feng D, Liang C (2003) Cdc48p is required for the cell cycle commitment point at Start via degradation of the G1-CDK inhibitor Far1p. The Journal of cell biology 163: 21-26 Galanty Y, Belotserkovskaya R, Coates J, Jackson SP (2012) RNF4, a SUMO-targeted ubiquitin E3 ligase, promotes DNA double-strand break repair. Genes & development 26: 1179-1195

173
Galanty Y, Belotserkovskaya R, Coates J, Polo S, Miller KM, Jackson SP (2009) Mammalian SUMO E3-ligases PIAS1 and PIAS4 promote responses to DNA double-strand breaks. Nature 462: 935-939 Geiss-Friedlander R, Melchior F (2007) Concepts in sumoylation: a decade on. Nature reviews 8: 947-956 Geoffroy MC, Jaffray EG, Walker KJ, Hay RT (2010) Arsenic-induced SUMO-dependent recruitment of RNF4 into PML nuclear bodies. Molecular biology of the cell 21: 4227-4239 Ghislain M, Dohmen RJ, Levy F, Varshavsky A (1996) Cdc48p interacts with Ufd3p, a WD repeat protein required for ubiquitin-mediated proteolysis in Saccharomyces cerevisiae. The EMBO journal 15: 4884-4899 Gill G (2004) SUMO and ubiquitin in the nucleus: different functions, similar mechanisms? Genes & development 18: 2046-2059 Gobbini E, Cesena D, Galbiati A, Lockhart A, Longhese MP (2013) Interplays between ATM/Tel1 and ATR/Mec1 in sensing and signaling DNA double-strand breaks. DNA repair 12: 791-799 Gocke CB, Yu H (2009) Identification of SUMO targets through in vitro expression cloning. Methods in molecular biology 497: 51-61 Gocke CB, Yu H, Kang J (2005) Systematic identification and analysis of mammalian small ubiquitin-like modifier substrates. The Journal of biological chemistry 280: 5004-5012 Golebiowski F, Matic I, Tatham MH, Cole C, Yin Y, Nakamura A, Cox J, Barton GJ, Mann M, Hay RT (2009) System-wide changes to SUMO modifications in response to heat shock. Science signaling 2: ra24 Gunaratne J, Schmidt A, Quandt A, Neo SP, Sarac OS, Gracia T, Loguercio S, Ahrne E, Xia RL, Tan KH, Lossner C, Bahler J, Beyer A, Blackstock W, Aebersold R (2013) Extensive mass spectrometry-based analysis of the fission yeast proteome: the Schizosaccharomyces pombe PeptideAtlas. Molecular & cellular proteomics : MCP 12: 1741-1751 Guzzo CM, Matunis MJ (2013) Expanding SUMO and ubiquitin-mediated signaling through hybrid SUMO-ubiquitin chains and their receptors. Cell cycle (Georgetown, Tex 12: 1015-1017 Haber JE (1999) DNA repair. Gatekeepers of recombination. Nature 398: 665, 667 Hannich JT, Lewis A, Kroetz MB, Li SJ, Heide H, Emili A, Hochstrasser M (2005) Defining the SUMO-modified proteome by multiple approaches in Saccharomyces cerevisiae. The Journal of biological chemistry 280: 4102-4110 Hanzelmann P, Schafer A, Voller D, Schindelin H (2012) Structural insights into functional modes of proteins involved in ubiquitin family pathways. Methods in molecular biology 832: 547-576 Hanzelmann P, Schindelin H (2011) The structural and functional basis of the p97/valosin-containing protein (VCP)-interacting motif (VIM): mutually exclusive binding of cofactors to the N-terminal domain of p97. The Journal of biological chemistry 286: 38679-38690 Harper JW, Elledge SJ (2007) The DNA damage response: ten years after. Molecular cell 28: 739-745 Hay RT (2005) SUMO: a history of modification. Molecular cell 18: 1-12 Hay RT (2013) Decoding the SUMO signal. Biochemical Society transactions 41: 463-473 Hecker CM, Rabiller M, Haglund K, Bayer P, Dikic I (2006) Specification of SUMO1- and SUMO2-interacting motifs. The Journal of biological chemistry 281: 16117-16127 Heideker J, Perry JJ, Boddy MN (2009) Genome stability roles of SUMO-targeted ubiquitin ligases. DNA repair 8: 517-524

174
Heideker J, Prudden J, Perry JJ, Tainer JA, Boddy MN (2011) SUMO-targeted ubiquitin ligase, Rad60, and Nse2 SUMO ligase suppress spontaneous Top1-mediated DNA damage and genome instability. PLoS genetics 7: e1001320 Heo JM, Livnat-Levanon N, Taylor EB, Jones KT, Dephoure N, Ring J, Xie J, Brodsky JL, Madeo F, Gygi SP, Ashrafi K, Glickman MH, Rutter J (2010) A stress-responsive system for mitochondrial protein degradation. Molecular cell 40: 465-480 Hetzer M, Meyer HH, Walther TC, Bilbao-Cortes D, Warren G, Mattaj IW (2001) Distinct AAA-ATPase p97 complexes function in discrete steps of nuclear assembly. Nature cell biology 3: 1086-1091 Hickey CM, Wilson NR, Hochstrasser M (2012) Function and regulation of SUMO proteases. Nature reviews 13: 755-766 Hietakangas V, Anckar J, Blomster HA, Fujimoto M, Palvimo JJ, Nakai A, Sistonen L (2006) PDSM, a motif for phosphorylation-dependent SUMO modification. Proceedings of the National Academy of Sciences of the United States of America 103: 45-50 Hitchcock AL, Krebber H, Frietze S, Lin A, Latterich M, Silver PA (2001) The conserved npl4 protein complex mediates proteasome-dependent membrane-bound transcription factor activation. Molecular biology of the cell 12: 3226-3241 Hitt R, Wolf DH (2004) Der1p, a protein required for degradation of malfolded soluble proteins of the endoplasmic reticulum: topology and Der1-like proteins. FEMS yeast research 4: 721-729 Ho JC, Warr NJ, Shimizu H, Watts FZ (2001) SUMO modification of Rad22, the Schizosaccharomyces pombe homologue of the recombination protein Rad52. Nucleic acids research 29: 4179-4186 Hochstrasser M (1996a) Protein degradation or regulation: Ub the judge. Cell 84: 813-815 Hochstrasser M (1996b) Ubiquitin-dependent protein degradation. Annu Rev Genet 30: 405-439 Hochstrasser M (2009) Origin and function of ubiquitin-like proteins. Nature 458: 422-429 Hoege C, Pfander B, Moldovan GL, Pyrowolakis G, Jentsch S (2002) RAD6-dependent DNA repair is linked to modification of PCNA by ubiquitin and SUMO. Nature 419: 135-141 Huang TT, Wuerzberger-Davis SM, Wu ZH, Miyamoto S (2003) Sequential modification of NEMO/IKKgamma by SUMO-1 and ubiquitin mediates NF-kappaB activation by genotoxic stress. Cell 115: 565-576 Huibregtse JM, Scheffner M, Beaudenon S, Howley PM (1995) A family of proteins structurally and functionally related to the E6-AP ubiquitin-protein ligase. Proceedings of the National Academy of Sciences of the United States of America 92: 5249 Hurley JH, Lee S, Prag G (2006) Ubiquitin-binding domains. The Biochemical journal 399: 361-372 Husnjak K, Dikic I (2012) Ubiquitin-binding proteins: decoders of ubiquitin-mediated cellular functions. Annual review of biochemistry 81: 291-322 Husnjak K, Elsasser S, Zhang N, Chen X, Randles L, Shi Y, Hofmann K, Walters KJ, Finley D, Dikic I (2008) Proteasome subunit Rpn13 is a novel ubiquitin receptor. Nature 453: 481-488 Iftode C, Daniely Y, Borowiec JA (1999) Replication protein A (RPA): the eukaryotic SSB. Critical reviews in biochemistry and molecular biology 34: 141-180 Ii T, Fung J, Mullen JR, Brill SJ (2007) The yeast Slx5-Slx8 DNA integrity complex displays ubiquitin ligase activity. Cell cycle (Georgetown, Tex 6: 2800-2809 Jackson SP, Bartek J (2009) The DNA-damage response in human biology and disease. Nature 461: 1071-1078

175
Jentsch S, Psakhye I (2013) Control of nuclear activities by substrate-selective and protein-group SUMOylation. Annual review of genetics 47: 167-186 Jentsch S, Rumpf S (2007) Cdc48 (p97): a "molecular gearbox" in the ubiquitin pathway? Trends in biochemical sciences 32: 6-11 Johnson ES, Blobel G (1997) Ubc9p is the conjugating enzyme for the ubiquitin-like protein Smt3p. The Journal of biological chemistry 272: 26799-26802 Johnson ES, Blobel G (1999) Cell cycle-regulated attachment of the ubiquitin-related protein SUMO to the yeast septins. The Journal of cell biology 147: 981-994 Johnson ES, Gupta AA (2001) An E3-like factor that promotes SUMO conjugation to the yeast septins. Cell 106: 735-744 Johnson ES, Ma PC, Ota IM, Varshavsky A (1995) A proteolytic pathway that recognizes ubiquitin as a degradation signal. The Journal of biological chemistry 270: 17442-17456 Kai M, Furuya K, Paderi F, Carr AM, Wang TS (2007) Rad3-dependent phosphorylation of the checkpoint clamp regulates repair-pathway choice. Nature cell biology 9: 691-697 Kakarougkas A, Jeggo PA (2014) DNA DSB repair pathway choice: an orchestrated handover mechanism. The British journal of radiology 87: 20130685 Kannouche PL, Wing J, Lehmann AR (2004) Interaction of human DNA polymerase eta with monoubiquitinated PCNA: a possible mechanism for the polymerase switch in response to DNA damage. Molecular cell 14: 491-500 Karras GI, Jentsch S (2010) The RAD6 DNA damage tolerance pathway operates uncoupled from the replication fork and is functional beyond S phase. Cell 141: 255-267 Kerscher O (2007) SUMO junction-what's your function? New insights through SUMO-interacting motifs. EMBO reports 8: 550-555 Kerscher O, Felberbaum R, Hochstrasser M (2006) Modification of proteins by ubiquitin and ubiquitin-like proteins. Annual review of cell and developmental biology 22: 159-180 Keusekotten K, Bade VN, Meyer-Teschendorf K, Sriramachandran AM, Fischer-Schrader K, Krause A, Horst C, Schwarz G, Hofmann K, Dohmen RJ, Praefcke GJ (2014) Multivalent interactions of the SUMO-interaction motifs in RING finger protein 4 determine the specificity for chains of the SUMO. The Biochemical journal 457: 207-214 Knipscheer P, van Dijk WJ, Olsen JV, Mann M, Sixma TK (2007) Noncovalent interaction between Ubc9 and SUMO promotes SUMO chain formation. The EMBO journal 26: 2797-2807 Knuesel M, Cheung HT, Hamady M, Barthel KK, Liu X (2005) A method of mapping protein sumoylation sites by mass spectrometry using a modified small ubiquitin-like modifier 1 (SUMO-1) and a computational program. Molecular & cellular proteomics : MCP 4: 1626-1636 Koepp DM (2014) Cell cycle regulation by protein degradation. Methods in molecular biology 1170: 61-73 Kohler JB, Jorgensen ML, Beinoraite G, Thorsen M, Thon G (2013) Concerted action of the ubiquitin-fusion degradation protein 1 (Ufd1) and Sumo-targeted ubiquitin ligases (STUbLs) in the DNA-damage response. PloS one 8: e80442 Komander D (2009) The emerging complexity of protein ubiquitination. Biochemical Society transactions 37: 937-953 Komander D, Rape M (2012) The ubiquitin code. Annual review of biochemistry 81: 203-229

176
Kosoy A, Calonge TM, Outwin EA, O'Connell MJ (2007) Fission yeast Rnf4 homologs are required for DNA repair. The Journal of biological chemistry 282: 20388-20394 Kroetz MB, Su D, Hochstrasser M (2009) Essential role of nuclear localization for yeast Ulp2 SUMO protease function. Molecular biology of the cell 20: 2196-2206 Krogh BO, Symington LS (2004) Recombination proteins in yeast. Annual review of genetics 38: 233-271 Lallemand-Breitenbach V, de The H (2010) PML nuclear bodies. Cold Spring Harbor perspectives in biology 2: a000661 Lallemand-Breitenbach V, Jeanne M, Benhenda S, Nasr R, Lei M, Peres L, Zhou J, Zhu J, Raught B, de The H (2008) Arsenic degrades PML or PML-RARalpha through a SUMO-triggered RNF4/ubiquitin-mediated pathway. Nature cell biology 10: 547-555 Lallemand-Breitenbach V, Zhu J, Puvion F, Koken M, Honore N, Doubeikovsky A, Duprez E, Pandolfi PP, Puvion E, Freemont P, de The H (2001) Role of promyelocytic leukemia (PML) sumolation in nuclear body formation, 11S proteasome recruitment, and As2O3-induced PML or PML/retinoic acid receptor alpha degradation. The Journal of experimental medicine 193: 1361-1371 Laney JD, Hochstrasser M (2004) Ubiquitin-dependent control of development in Saccharomyces cerevisiae. Current opinion in microbiology 7: 647-654 Lapenta V, Chiurazzi P, van der Spek P, Pizzuti A, Hanaoka F, Brahe C (1997) SMT3A, a human homologue of the S. cerevisiae SMT3 gene, maps to chromosome 21qter and defines a novel gene family. Genomics 40: 362-366 Latif C, Harvey SH, O'Connell MJ (2001) Ensuring the stability of the genome: DNA damage checkpoints. TheScientificWorldJournal 1: 684-702 Latterich M, Frohlich KU, Schekman R (1995) Membrane fusion and the cell cycle: Cdc48p participates in the fusion of ER membranes. Cell 82: 885-893 Li SJ, Hochstrasser M (1999) A new protease required for cell-cycle progression in yeast. Nature 398: 246-251 Li SJ, Hochstrasser M (2000) The yeast ULP2 (SMT4) gene encodes a novel protease specific for the ubiquitin-like Smt3 protein. Molecular and cellular biology 20: 2367-2377 Li SJ, Hochstrasser M (2003) The Ulp1 SUMO isopeptidase: distinct domains required for viability, nuclear envelope localization, and substrate specificity. The Journal of cell biology 160: 1069-1081 Li X, Heyer WD (2008) Homologous recombination in DNA repair and DNA damage tolerance. Cell Res 18: 99-113 Liberi G, Maffioletti G, Lucca C, Chiolo I, Baryshnikova A, Cotta-Ramusino C, Lopes M, Pellicioli A, Haber JE, Foiani M (2005) Rad51-dependent DNA structures accumulate at damaged replication forks in sgs1 mutants defective in the yeast ortholog of BLM RecQ helicase. Genes & development 19: 339-350 Limbo O, Porter-Goff ME, Rhind N, Russell P (2011) Mre11 nuclease activity and Ctp1 regulate Chk1 activation by Rad3ATR and Tel1ATM checkpoint kinases at double-strand breaks. Molecular and cellular biology 31: 573-583 Lin DY, Huang YS, Jeng JC, Kuo HY, Chang CC, Chao TT, Ho CC, Chen YC, Lin TP, Fang HI, Hung CC, Suen CS, Hwang MJ, Chang KS, Maul GG, Shih HM (2006) Role of SUMO-interacting motif in Daxx SUMO modification, subnuclear localization, and repression of sumoylated transcription factors. Molecular cell 24: 341-354 Liu J, Renault L, Veaute X, Fabre F, Stahlberg H, Heyer WD (2011) Rad51 paralogues Rad55-Rad57 balance the antirecombinase Srs2 in Rad51 filament formation. Nature 479: 245-248 Luo K, Zhang H, Wang L, Yuan J, Lou Z (2012) Sumoylation of MDC1 is important for proper DNA damage response. The EMBO journal 31: 3008-3019

177
Makhnevych T, Sydorskyy Y, Xin X, Srikumar T, Vizeacoumar FJ, Jeram SM, Li Z, Bahr S, Andrews BJ, Boone C, Raught B (2009) Global map of SUMO function revealed by protein-protein interaction and genetic networks. Molecular cell 33: 124-135 Mannen H, Tseng HM, Cho CL, Li SS (1996) Cloning and expression of human homolog HSMT3 to yeast SMT3 suppressor of MIF2 mutations in a centromere protein gene. Biochemical and biophysical research communications 222: 178-180 Manning Fox JE, Hajmrle C, Macdonald PE (2012) Novel roles of SUMO in pancreatic beta-cells: thinking outside the nucleus. Canadian journal of physiology and pharmacology 90: 765-770 Matic I, Schimmel J, Hendriks IA, van Santen MA, van de Rijke F, van Dam H, Gnad F, Mann M, Vertegaal AC (2010) Site-specific identification of SUMO-2 targets in cells reveals an inverted SUMOylation motif and a hydrophobic cluster SUMOylation motif. Molecular cell 39: 641-652 Matic I, van Hagen M, Schimmel J, Macek B, Ogg SC, Tatham MH, Hay RT, Lamond AI, Mann M, Vertegaal AC (2008) In vivo identification of human small ubiquitin-like modifier polymerization sites by high accuracy mass spectrometry and an in vitro to in vivo strategy. Molecular & cellular proteomics : MCP 7: 132-144 Matsuo Y, Kishimoto H, Tanae K, Kitamura K, Katayama S, Kawamukai M (2011) Nuclear protein quality is regulated by the ubiquitin-proteasome system through the activity of Ubc4 and San1 in fission yeast. The Journal of biological chemistry 286: 13775-13790 Matunis MJ, Coutavas E, Blobel G (1996) A novel ubiquitin-like modification modulates the partitioning of the Ran-GTPase-activating protein RanGAP1 between the cytosol and the nuclear pore complex. The Journal of cell biology 135: 1457-1470 Medicherla B, Goldberg AL (2008) Heat shock and oxygen radicals stimulate ubiquitin-dependent degradation mainly of newly synthesized proteins. The Journal of cell biology 182: 663-673 Meerang M, Ritz D, Paliwal S, Garajova Z, Bosshard M, Mailand N, Janscak P, Hubscher U, Meyer H, Ramadan K (2011) The ubiquitin-selective segregase VCP/p97 orchestrates the response to DNA double-strand breaks. Nature cell biology 13: 1376-1382 Meluh PB, Koshland D (1995) Evidence that the MIF2 gene of Saccharomyces cerevisiae encodes a centromere protein with homology to the mammalian centromere protein CENP-C. Molecular biology of the cell 6: 793-807 Meyer H, Bug M, Bremer S (2012) Emerging functions of the VCP/p97 AAA-ATPase in the ubiquitin system. Nature cell biology 14: 117-123 Meyer HH, Shorter JG, Seemann J, Pappin D, Warren G (2000) A complex of mammalian ufd1 and npl4 links the AAA-ATPase, p97, to ubiquitin and nuclear transport pathways. The EMBO journal 19: 2181-2192 Meyer HH, Wang Y, Warren G (2002) Direct binding of ubiquitin conjugates by the mammalian p97 adaptor complexes, p47 and Ufd1-Npl4. The EMBO journal 21: 5645-5652 Miller MJ, Barrett-Wilt GA, Hua Z, Vierstra RD (2010) Proteomic analyses identify a diverse array of nuclear processes affected by small ubiquitin-like modifier conjugation in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America 107: 16512-16517 Miyagawa K, Low RS, Santosa V, Tsuji H, Moser BA, Fujisawa S, Harland JL, Raguimova ON, Go A, Ueno M, Matsuyama A, Yoshida M, Nakamura TM, Tanaka K (2014) SUMOylation regulates telomere length by targeting the shelterin subunit Tpz1(Tpp1) to modulate shelterin-Stn1 interaction in fission yeast. Proceedings of the National Academy of Sciences of the United States of America 111: 5950-5955 Moir D, Stewart SE, Osmond BC, Botstein D (1982) Cold-sensitive cell-division-cycle mutants of yeast: isolation, properties, and pseudoreversion studies. Genetics 100: 547-563

178
Motegi A, Murakawa Y, Takeda S (2009) The vital link between the ubiquitin-proteasome pathway and DNA repair: impact on cancer therapy. Cancer letters 283: 1-9 Mukhopadhyay D, Arnaoutov A, Dasso M (2010) The SUMO protease SENP6 is essential for inner kinetochore assembly. The Journal of cell biology 188: 681-692 Mukhopadhyay D, Ayaydin F, Kolli N, Tan SH, Anan T, Kametaka A, Azuma Y, Wilkinson KD, Dasso M (2006) SUSP1 antagonizes formation of highly SUMO2/3-conjugated species. The Journal of cell biology 174: 939-949 Mukhopadhyay D, Dasso M (2007) Modification in reverse: the SUMO proteases. Trends in biochemical sciences 32: 286-295 Mullen JR, Brill SJ (2008) Activation of the Slx5-Slx8 ubiquitin ligase by poly-small ubiquitin-like modifier conjugates. The Journal of biological chemistry 283: 19912-19921 Mullen JR, Chen CF, Brill SJ (2010) Wss1 is a SUMO-dependent isopeptidase that interacts genetically with the Slx5-Slx8 SUMO-targeted ubiquitin ligase. Molecular and cellular biology 30: 3737-3748 Mullen JR, Das M, Brill SJ (2011) Genetic evidence that polysumoylation bypasses the need for a SUMO-targeted Ub ligase. Genetics 187: 73-87 Mullen JR, Kaliraman V, Ibrahim SS, Brill SJ (2001) Requirement for three novel protein complexes in the absence of the Sgs1 DNA helicase in Saccharomyces cerevisiae. Genetics 157: 103-118 Muris DF, Vreeken K, Carr AM, Broughton BC, Lehmann AR, Lohman PH, Pastink A (1993) Cloning the RAD51 homologue of Schizosaccharomyces pombe. Nucleic acids research 21: 4586-4591 Nagai S, Dubrana K, Tsai-Pflugfelder M, Davidson MB, Roberts TM, Brown GW, Varela E, Hediger F, Gasser SM, Krogan NJ (2008) Functional targeting of DNA damage to a nuclear pore-associated SUMO-dependent ubiquitin ligase. Science (New York, NY 322: 597-602 Nakada D, Shimomura T, Matsumoto K, Sugimoto K (2003) The ATM-related Tel1 protein of Saccharomyces cerevisiae controls a checkpoint response following phleomycin treatment. Nucleic acids research 31: 1715-1724 Nakatsukasa K, Brodsky JL, Kamura T (2013) A stalled retrotranslocation complex reveals physical linkage between substrate recognition and proteasomal degradation during ER-associated degradation. Molecular biology of the cell 24: 1765-1775, S1761-1768 Neduva V, Russell RB (2005) Linear motifs: evolutionary interaction switches. FEBS letters 579: 3342-3345 Nie M, Aslanian A, Prudden J, Heideker J, Vashisht AA, Wohlschlegel JA, Yates JR, 3rd, Boddy MN (2012) Dual recruitment of Cdc48 (p97)-Ufd1-Npl4 ubiquitin-selective segregase by small ubiquitin-like modifier protein (SUMO) and ubiquitin in SUMO-targeted ubiquitin ligase-mediated genome stability functions. The Journal of biological chemistry 287: 29610-29619 Nielsen SV, Poulsen EG, Rebula CA, Hartmann-Petersen R (2014) Protein quality control in the nucleus. Biomolecules 4: 646-661 Okura T, Gong L, Kamitani T, Wada T, Okura I, Wei CF, Chang HM, Yeh ET (1996) Protection against Fas/APO-1- and tumor necrosis factor-mediated cell death by a novel protein, sentrin. Journal of immunology 157: 4277-4281 Olsen JV, Blagoev B, Gnad F, Macek B, Kumar C, Mortensen P, Mann M (2006) Global, in vivo, and site-specific phosphorylation dynamics in signaling networks. Cell 127: 635-648 Osman F, Whitby MC (2007) Exploring the roles of Mus81-Eme1/Mms4 at perturbed replication forks. DNA repair 6: 1004-1017

179
Owerbach D, McKay EM, Yeh ET, Gabbay KH, Bohren KM (2005) A proline-90 residue unique to SUMO-4 prevents maturation and sumoylation. Biochemical and biophysical research communications 337: 517-520 Papouli E, Chen S, Davies AA, Huttner D, Krejci L, Sung P, Ulrich HD (2005) Crosstalk between SUMO and ubiquitin on PCNA is mediated by recruitment of the helicase Srs2p. Molecular cell 19: 123-133 Park CW, Ryu KY (2014) Cellular Ubiquitin Pool Dynamics and Homeostasis. BMB reports Park HJ, Yun DJ (2013) New insights into the role of the small ubiquitin-like modifier (SUMO) in plants. International review of cell and molecular biology 300: 161-209 Park S, Isaacson R, Kim HT, Silver PA, Wagner G (2005) Ufd1 exhibits the AAA-ATPase fold with two distinct ubiquitin interaction sites. Structure 13: 995-1005 Parker JL, Ulrich HD (2012) A SUMO-interacting motif activates budding yeast ubiquitin ligase Rad18 towards SUMO-modified PCNA. Nucleic acids research 40: 11380-11388 Parrilla-Castellar ER, Arlander SJ, Karnitz L (2004) Dial 9-1-1 for DNA damage: the Rad9-Hus1-Rad1 (9-1-1) clamp complex. DNA repair 3: 1009-1014 Peng J, Schwartz D, Elias JE, Thoreen CC, Cheng D, Marsischky G, Roelofs J, Finley D, Gygi SP (2003) A proteomics approach to understanding protein ubiquitination. Nature biotechnology 21: 921-926 Petermann E, Orta ML, Issaeva N, Schultz N, Helleday T (2010) Hydroxyurea-stalled replication forks become progressively inactivated and require two different RAD51-mediated pathways for restart and repair. Molecular cell 37: 492-502 Peters JM, Walsh MJ, Franke WW (1990) An abundant and ubiquitous homo-oligomeric ring-shaped ATPase particle related to the putative vesicle fusion proteins Sec18p and NSF. The EMBO journal 9: 1757-1767 Pfander B, Moldovan GL, Sacher M, Hoege C, Jentsch S (2005) SUMO-modified PCNA recruits Srs2 to prevent recombination during S phase. Nature 436: 428-433 Pichler A, Gast A, Seeler JS, Dejean A, Melchior F (2002) The nucleoporin RanBP2 has SUMO1 E3 ligase activity. Cell 108: 109-120 Pichler A, Knipscheer P, Oberhofer E, van Dijk WJ, Korner R, Olsen JV, Jentsch S, Melchior F, Sixma TK (2005) SUMO modification of the ubiquitin-conjugating enzyme E2-25K. Nature structural & molecular biology 12: 264-269 Pichler A, Knipscheer P, Saitoh H, Sixma TK, Melchior F (2004) The RanBP2 SUMO E3 ligase is neither HECT- nor RING-type. Nature structural & molecular biology 11: 984-991 Pinder JB, Attwood KM, Dellaire G (2013) Reading, writing, and repair: the role of ubiquitin and the ubiquitin-like proteins in DNA damage signaling and repair. Frontiers in genetics 4: 45 Plechanovova A, Jaffray EG, Tatham MH, Naismith JH, Hay RT (2012) Structure of a RING E3 ligase and ubiquitin-loaded E2 primed for catalysis. Nature 489: 115-120 Polo S (2012) Signaling-mediated control of ubiquitin ligases in endocytosis. BMC biology 10: 25 Potts PR, Yu H (2005) Human MMS21/NSE2 is a SUMO ligase required for DNA repair. Molecular and cellular biology 25: 7021-7032 Poulsen SL, Hansen RK, Wagner SA, van Cuijk L, van Belle GJ, Streicher W, Wikstrom M, Choudhary C, Houtsmuller AB, Marteijn JA, Bekker-Jensen S, Mailand N (2013) RNF111/Arkadia is a SUMO-targeted ubiquitin ligase that facilitates the DNA damage response. The Journal of cell biology 201: 797-807

180
Praefcke GJ, Hofmann K, Dohmen RJ (2012) SUMO playing tag with ubiquitin. Trends in biochemical sciences 37: 23-31 Prakash L (1981) Characterization of postreplication repair in Saccharomyces cerevisiae and effects of rad6, rad18, rev3 and rad52 mutations. Molecular & general genetics : MGG 184: 471-478 Prudden J, Pebernard S, Raffa G, Slavin DA, Perry JJ, Tainer JA, McGowan CH, Boddy MN (2007) SUMO-targeted ubiquitin ligases in genome stability. The EMBO journal 26: 4089-4101 Prudden J, Perry JJ, Nie M, Vashisht AA, Arvai AS, Hitomi C, Guenther G, Wohlschlegel JA, Tainer JA, Boddy MN (2011) DNA repair and global sumoylation are regulated by distinct Ubc9 noncovalent complexes. Molecular and cellular biology 31: 2299-2310 Psakhye I, Jentsch S (2012) Protein group modification and synergy in the SUMO pathway as exemplified in DNA repair. Cell 151: 807-820 Pye VE, Beuron F, Keetch CA, McKeown C, Robinson CV, Meyer HH, Zhang X, Freemont PS (2007) Structural insights into the p97-Ufd1-Npl4 complex. Proceedings of the National Academy of Sciences of the United States of America 104: 467-472 Pye VE, Dreveny I, Briggs LC, Sands C, Beuron F, Zhang X, Freemont PS (2006) Going through the motions: the ATPase cycle of p97. Journal of structural biology 156: 12-28 Raji H, Hartsuiker E (2006) Double-strand break repair and homologous recombination in Schizosaccharomyces pombe. Yeast (Chichester, England) 23: 963-976 Ramadan K, Bruderer R, Spiga FM, Popp O, Baur T, Gotta M, Meyer HH (2007) Cdc48/p97 promotes reformation of the nucleus by extracting the kinase Aurora B from chromatin. Nature 450: 1258-1262 Rape M, Hoppe T, Gorr I, Kalocay M, Richly H, Jentsch S (2001) Mobilization of processed, membrane-tethered SPT23 transcription factor by CDC48(UFD1/NPL4), a ubiquitin-selective chaperone. Cell 107: 667-677 Reindle A, Belichenko I, Bylebyl GR, Chen XL, Gandhi N, Johnson ES (2006) Multiple domains in Siz SUMO ligases contribute to substrate selectivity. Journal of cell science 119: 4749-4757 Reverter D, Lima CD (2005) Insights into E3 ligase activity revealed by a SUMO-RanGAP1-Ubc9-Nup358 complex. Nature 435: 687-692 Richly H, Rape M, Braun S, Rumpf S, Hoege C, Jentsch S (2005) A series of ubiquitin binding factors connects CDC48/p97 to substrate multiubiquitylation and proteasomal targeting. Cell 120: 73-84 Rodriguez MS, Dargemont C, Hay RT (2001) SUMO-1 conjugation in vivo requires both a consensus modification motif and nuclear targeting. The Journal of biological chemistry 276: 12654-12659 Rojas-Fernandez A, Plechanovova A, Hattersley N, Jaffray E, Tatham MH, Hay RT (2014) SUMO chain-induced dimerization activates RNF4. Molecular cell 53: 880-892 Rosas-Acosta G, Russell WK, Deyrieux A, Russell DH, Wilson VG (2005) A universal strategy for proteomic studies of SUMO and other ubiquitin-like modifiers. Molecular & cellular proteomics : MCP 4: 56-72 Rouiller I, DeLaBarre B, May AP, Weis WI, Brunger AT, Milligan RA, Wilson-Kubalek EM (2002) Conformational changes of the multifunction p97 AAA ATPase during its ATPase cycle. Nature structural biology 9: 950-957 Rouse J, Jackson SP (2002) Interfaces between the detection, signaling, and repair of DNA damage. Science (New York, NY 297: 547-551 Rumpf S, Jentsch S (2006) Functional division of substrate processing cofactors of the ubiquitin-selective Cdc48 chaperone. Molecular cell 21: 261-269

181
Sachdev S, Bruhn L, Sieber H, Pichler A, Melchior F, Grosschedl R (2001) PIASy, a nuclear matrix-associated SUMO E3 ligase, represses LEF1 activity by sequestration into nuclear bodies. Genes & development 15: 3088-3103 Sacher M, Pfander B, Hoege C, Jentsch S (2006) Control of Rad52 recombination activity by double-strand break-induced SUMO modification. Nature cell biology 8: 1284-1290 Sacher M, Pfander B, Jentsch S (2005) Identification of SUMO-protein conjugates. Methods in enzymology 399: 392-404 Saito Y, Takeda J, Okada M, Kobayashi J, Kato A, Hirota K, Taoka M, Matsumoto T, Komatsu K, Isobe T (2013) The proteasome factor Bag101 binds to Rad22 and suppresses homologous recombination. Scientific reports 3: 2022 Saitoh H, Hinchey J (2000) Functional heterogeneity of small ubiquitin-related protein modifiers SUMO-1 versus SUMO-2/3. The Journal of biological chemistry 275: 6252-6258 San Filippo J, Sung P, Klein H (2008) Mechanism of eukaryotic homologous recombination. Annual review of biochemistry 77: 229-257 Savitsky K, Sfez S, Tagle DA, Ziv Y, Sartiel A, Collins FS, Shiloh Y, Rotman G (1995) The complete sequence of the coding region of the ATM gene reveals similarity to cell cycle regulators in different species. Human molecular genetics 4: 2025-2032 Schiestl RH, Prakash S (1989) Interactions of the RAD7 and RAD23 excision repair genes of Saccharomyces cerevisiae with DNA repair genes in different epistasis groups. Current genetics 16: 219-223 Schimmel J, Eifler K, Sigurethsson JO, Cuijpers SA, Hendriks IA, Verlaan-de Vries M, Kelstrup CD, Francavilla C, Medema RH, Olsen JV, Vertegaal AC (2014) Uncovering SUMOylation dynamics during cell-cycle progression reveals FoxM1 as a key mitotic SUMO target protein. Molecular cell 53: 1053-1066 Schimmel J, Larsen KM, Matic I, van Hagen M, Cox J, Mann M, Andersen JS, Vertegaal AC (2008) The ubiquitin-proteasome system is a key component of the SUMO-2/3 cycle. Molecular & cellular proteomics : MCP 7: 2107-2122 Schmidt D, Muller S (2002) Members of the PIAS family act as SUMO ligases for c-Jun and p53 and repress p53 activity. Proceedings of the National Academy of Sciences of the United States of America 99: 2872-2877 Schmidt M, Finley D (2014) Regulation of proteasome activity in health and disease. Biochimica et biophysica acta 1843: 13-25 Schrader EK, Harstad KG, Matouschek A (2009) Targeting proteins for degradation. Nature chemical biology 5: 815-822 Schuberth C, Buchberger A (2008) UBX domain proteins: major regulators of the AAA ATPase Cdc48/p97. Cell Mol Life Sci 65: 2360-2371 Schuberth C, Richly H, Rumpf S, Buchberger A (2004) Shp1 and Ubx2 are adaptors of Cdc48 involved in ubiquitin-dependent protein degradation. EMBO reports 5: 818-824 Schwartz DC, Felberbaum R, Hochstrasser M (2007) The Ulp2 SUMO protease is required for cell division following termination of the DNA damage checkpoint. Molecular and cellular biology 27: 6948-6961 Schwienhorst I, Johnson ES, Dohmen RJ (2000) SUMO conjugation and deconjugation. Molecular & general genetics : MGG 263: 771-786 Seaton BL, Yucel J, Sunnerhagen P, Subramani S (1992) Isolation and characterization of the Schizosaccharomyces pombe rad3 gene, involved in the DNA damage and DNA synthesis checkpoints. Gene 119: 83-89

182
Seeler JS, Bischof O, Nacerddine K, Dejean A (2007) SUMO, the three Rs and cancer. Current topics in microbiology and immunology 313: 49-71 Seeler JS, Dejean A (2001) SUMO: of branched proteins and nuclear bodies. Oncogene 20: 7243-7249 Seeler JS, Marchio A, Sitterlin D, Transy C, Dejean A (1998) Interaction of SP100 with HP1 proteins: a link between the promyelocytic leukemia-associated nuclear bodies and the chromatin compartment. Proceedings of the National Academy of Sciences of the United States of America 95: 7316-7321 Seroussi E, Lavi S (1993) Replication protein A is the major single-stranded DNA binding protein detected in mammalian cell extracts by gel retardation assays and UV cross-linking of long and short single-stranded DNA molecules. The Journal of biological chemistry 268: 7147-7154 Shen Z, Pardington-Purtymun PE, Comeaux JC, Moyzis RK, Chen DJ (1996) UBL1, a human ubiquitin-like protein associating with human RAD51/RAD52 proteins. Genomics 36: 271-279 Skilton A, Ho JC, Mercer B, Outwin E, Watts FZ (2009) SUMO chain formation is required for response to replication arrest in S. pombe. PloS one 4: e6750 Song J, Durrin LK, Wilkinson TA, Krontiris TG, Chen Y (2004) Identification of a SUMO-binding motif that recognizes SUMO-modified proteins. Proceedings of the National Academy of Sciences of the United States of America 101: 14373-14378 Sontag EM, Vonk WI, Frydman J (2014) Sorting out the trash: the spatial nature of eukaryotic protein quality control. Current opinion in cell biology 26: 139-146 Srikumar T, Lewicki MC, Costanzo M, Tkach JM, van Bakel H, Tsui K, Johnson ES, Brown GW, Andrews BJ, Boone C, Giaever G, Nislow C, Raught B (2013) Global analysis of SUMO chain function reveals multiple roles in chromatin regulation. The Journal of cell biology 201: 145-163 Sriramachandran AM, Dohmen RJ (2014) SUMO-targeted ubiquitin ligases. Biochimica et biophysica acta 1843: 75-85 Stapf C, Cartwright E, Bycroft M, Hofmann K, Buchberger A (2011) The general definition of the p97/valosin-containing protein (VCP)-interacting motif (VIM) delineates a new family of p97 cofactors. The Journal of biological chemistry 286: 38670-38678 Steen H, Mann M (2004) The ABC's (and XYZ's) of peptide sequencing. Nature reviews 5: 699-711 Steinacher R, Osman F, Lorenz A, Bryer C, Whitby MC (2013) Slx8 removes Pli1-dependent protein-SUMO conjugates including SUMOylated topoisomerase I to promote genome stability. PloS one 8: e71960 Steinacher R, Schar P (2005) Functionality of human thymine DNA glycosylase requires SUMO-regulated changes in protein conformation. Current biology : CB 15: 616-623 Stelter P, Ulrich HD (2003) Control of spontaneous and damage-induced mutagenesis by SUMO and ubiquitin conjugation. Nature 425: 188-191 Stolz A, Hilt W, Buchberger A, Wolf DH (2011) Cdc48: a power machine in protein degradation. Trends in biochemical sciences 36: 515-523 Sun H, Hunter T (2012) Poly-small ubiquitin-like modifier (PolySUMO)-binding proteins identified through a string search. The Journal of biological chemistry 287: 42071-42083 Sun H, Leverson JD, Hunter T (2007) Conserved function of RNF4 family proteins in eukaryotes: targeting a ubiquitin ligase to SUMOylated proteins. The EMBO journal 26: 4102-4112

183
Takahashi Y, Iwase M, Konishi M, Tanaka M, Toh-e A, Kikuchi Y (1999) Smt3, a SUMO-1 homolog, is conjugated to Cdc3, a component of septin rings at the mother-bud neck in budding yeast. Biochemical and biophysical research communications 259: 582-587 Takahashi Y, Kahyo T, Toh EA, Yasuda H, Kikuchi Y (2001) Yeast Ull1/Siz1 is a novel SUMO1/Smt3 ligase for septin components and functions as an adaptor between conjugating enzyme and substrates. The Journal of biological chemistry 276: 48973-48977 Tammsalu T, Matic I, Jaffray EG, Ibrahim AF, Tatham MH, Hay RT (2014) Proteome-wide identification of SUMO2 modification sites. Science signaling 7: rs2 Tan W, Wang Z, Prelich G (2013) Physical and Genetic Interactions Between Uls1 and the Slx5-Slx8 SUMO-Targeted Ubiquitin Ligase. G3 Tanaka K, Nishide J, Okazaki K, Kato H, Niwa O, Nakagawa T, Matsuda H, Kawamukai M, Murakami Y (1999) Characterization of a fission yeast SUMO-1 homologue, pmt3p, required for multiple nuclear events, including the control of telomere length and chromosome segregation. Molecular and cellular biology 19: 8660-8672 Tatham MH, Geoffroy MC, Shen L, Plechanovova A, Hattersley N, Jaffray EG, Palvimo JJ, Hay RT (2008) RNF4 is a poly-SUMO-specific E3 ubiquitin ligase required for arsenic-induced PML degradation. Nature cell biology 10: 538-546 Tatham MH, Jaffray E, Vaughan OA, Desterro JM, Botting CH, Naismith JH, Hay RT (2001) Polymeric chains of SUMO-2 and SUMO-3 are conjugated to protein substrates by SAE1/SAE2 and Ubc9. The Journal of biological chemistry 276: 35368-35374 Tatham MH, Matic I, Mann M, Hay RT (2011) Comparative proteomic analysis identifies a role for SUMO in protein quality control. Science signaling 4: rs4 Taylor DL, Ho JC, Oliver A, Watts FZ (2002) Cell-cycle-dependent localisation of Ulp1, a Schizosaccharomyces pombe Pmt3 (SUMO)-specific protease. Journal of cell science 115: 1113-1122 Thrower JS, Hoffman L, Rechsteiner M, Pickart CM (2000) Recognition of the polyubiquitin proteolytic signal. The EMBO journal 19: 94-102 Tokunaga F, Sakata S, Saeki Y, Satomi Y, Kirisako T, Kamei K, Nakagawa T, Kato M, Murata S, Yamaoka S, Yamamoto M, Akira S, Takao T, Tanaka K, Iwai K (2009) Involvement of linear polyubiquitylation of NEMO in NF-kappaB activation. Nature cell biology 11: 123-132 Uchiyama K, Jokitalo E, Kano F, Murata M, Zhang X, Canas B, Newman R, Rabouille C, Pappin D, Freemont P, Kondo H (2002) VCIP135, a novel essential factor for p97/p47-mediated membrane fusion, is required for Golgi and ER assembly in vivo. The Journal of cell biology 159: 855-866 Ulrich HD (2005) Mutual interactions between the SUMO and ubiquitin systems: a plea of no contest. Trends in cell biology 15: 525-532 Ulrich HD (2008) The fast-growing business of SUMO chains. Molecular cell 32: 301-305 Ulrich HD (2009) Regulating post-translational modifications of the eukaryotic replication clamp PCNA. DNA repair 8: 461-469 Ulrich HD (2014) Two-way communications between ubiquitin-like modifiers and DNA. Nature structural & molecular biology 21: 317-324 Ulrich HD, Davies AA (2009) In vivo detection and characterization of sumoylation targets in Saccharomyces cerevisiae. Methods in molecular biology 497: 81-103

184
Ulrich HD, Jentsch S (2000) Two RING finger proteins mediate cooperation between ubiquitin-conjugating enzymes in DNA repair. The EMBO journal 19: 3388-3397 Uzunova K, Gottsche K, Miteva M, Weisshaar SR, Glanemann C, Schnellhardt M, Niessen M, Scheel H, Hofmann K, Johnson ES, Praefcke GJ, Dohmen RJ (2007) Ubiquitin-dependent proteolytic control of SUMO conjugates. The Journal of biological chemistry 282: 34167-34175 van de Pasch LA, Miles AJ, Nijenhuis W, Brabers NA, van Leenen D, Lijnzaad P, Brown MK, Ouellet J, Barral Y, Kops GJ, Holstege FC (2013) Centromere binding and a conserved role in chromosome stability for SUMO-dependent ubiquitin ligases. PloS one 8: e65628 van der Veen AG, Ploegh HL (2012) Ubiquitin-like proteins. Annual review of biochemistry 81: 323-357 Vertegaal AC. (2010) SUMO chains: polymeric signals. Biochem Soc Trans, Vol. 38, pp. 46-49. Vertegaal AC, Andersen JS, Ogg SC, Hay RT, Mann M, Lamond AI (2006) Distinct and overlapping sets of SUMO-1 and SUMO-2 target proteins revealed by quantitative proteomics. Molecular & cellular proteomics : MCP 5: 2298-2310 Vertegaal AC, Ogg SC, Jaffray E, Rodriguez MS, Hay RT, Andersen JS, Mann M, Lamond AI (2004) A proteomic study of SUMO-2 target proteins. The Journal of biological chemistry 279: 33791-33798 Wagner SA, Beli P, Weinert BT, Nielsen ML, Cox J, Mann M, Choudhary C (2011) A proteome-wide, quantitative survey of in vivo ubiquitylation sites reveals widespread regulatory roles. Molecular & cellular proteomics : MCP 10: M111 013284 Wagner SA, Beli P, Weinert BT, Scholz C, Kelstrup CD, Young C, Nielsen ML, Olsen JV, Brakebusch C, Choudhary C (2012) Proteomic analyses reveal divergent ubiquitylation site patterns in murine tissues. Molecular & cellular proteomics : MCP 11: 1578-1585 Wan J, Subramonian D, Zhang XD (2012) SUMOylation in control of accurate chromosome segregation during mitosis. Current protein & peptide science 13: 467-481 Wang Z, Jones GM, Prelich G (2006) Genetic analysis connects SLX5 and SLX8 to the SUMO pathway in Saccharomyces cerevisiae. Genetics 172: 1499-1509 Wang Z, Prelich G (2009) Quality control of a transcriptional regulator by SUMO-targeted degradation. Molecular and cellular biology 29: 1694-1706 Watanabe K, Tateishi S, Kawasuji M, Tsurimoto T, Inoue H, Yamaizumi M (2004) Rad18 guides poleta to replication stalling sites through physical interaction and PCNA monoubiquitination. The EMBO journal 23: 3886-3896 Watts FZ (2006) Sumoylation of PCNA: Wrestling with recombination at stalled replication forks. DNA repair 5: 399-403 Watts FZ (2007) The role of SUMO in chromosome segregation. Chromosoma 116: 15-20 Watts FZ (2013) Starting and stopping SUMOylation. What regulates the regulator? Chromosoma 122: 451-463 Watts FZ, Skilton A, Ho JC, Boyd LK, Trickey MA, Gardner L, Ogi FX, Outwin EA (2007) The role of Schizosaccharomyces pombe SUMO ligases in genome stability. Biochemical Society transactions 35: 1379-1384 Weisshaar SR, Keusekotten K, Krause A, Horst C, Springer HM, Gottsche K, Dohmen RJ, Praefcke GJ (2008) Arsenic trioxide stimulates SUMO-2/3 modification leading to RNF4-dependent proteolytic targeting of PML. FEBS letters 582: 3174-3178 Werner A, Flotho A, Melchior F (2012) The RanBP2/RanGAP1*SUMO1/Ubc9 complex is a multisubunit SUMO E3 ligase. Molecular cell 46: 287-298

185
Westman BJ, Verheggen C, Hutten S, Lam YW, Bertrand E, Lamond AI (2010) A proteomic screen for nucleolar SUMO targets shows SUMOylation modulates the function of Nop5/Nop58. Molecular cell 39: 618-631 Wilcox AJ, Laney JD (2009) A ubiquitin-selective AAA-ATPase mediates transcriptional switching by remodelling a repressor-promoter DNA complex. Nature cell biology 11: 1481-1486 Wilson VG, Heaton PR (2008) Ubiquitin proteolytic system: focus on SUMO. Expert review of proteomics 5: 121-135 Wohlschlegel JA, Johnson ES, Reed SI, Yates JR, 3rd (2004) Global analysis of protein sumoylation in Saccharomyces cerevisiae. The Journal of biological chemistry 279: 45662-45668 Wohlschlegel JA, Johnson ES, Reed SI, Yates JR, 3rd (2006) Improved identification of SUMO attachment sites using C-terminal SUMO mutants and tailored protease digestion strategies. Journal of proteome research 5: 761-770 Wojcik C, DeMartino GN (2003) Intracellular localization of proteasomes. The international journal of biochemistry & cell biology 35: 579-589 Wong KH, Todd RB, Oakley BR, Oakley CE, Hynes MJ, Davis MA (2008) Sumoylation in Aspergillus nidulans: sumO inactivation, overexpression and live-cell imaging. Fungal genetics and biology : FG & B 45: 728-737 Xhemalce B, Seeler JS, Thon G, Dejean A, Arcangioli B (2004) Role of the fission yeast SUMO E3 ligase Pli1p in centromere and telomere maintenance. The EMBO journal 23: 3844-3853 Xiao W, Chow BL, Broomfield S, Hanna M (2000) The Saccharomyces cerevisiae RAD6 group is composed of an error-prone and two error-free postreplication repair pathways. Genetics 155: 1633-1641 Xie Y, Kerscher O, Kroetz MB, McConchie HF, Sung P, Hochstrasser M (2007) The yeast Hex3.Slx8 heterodimer is a ubiquitin ligase stimulated by substrate sumoylation. The Journal of biological chemistry 282: 34176-34184 Xu G, Paige JS, Jaffrey SR (2010) Global analysis of lysine ubiquitination by ubiquitin remnant immunoaffinity profiling. Nature biotechnology 28: 868-873 Xu J, He Y, Qiang B, Yuan J, Peng X, Pan XM (2008) A novel method for high accuracy sumoylation site prediction from protein sequences. BMC bioinformatics 9: 8 Xu P, Duong DM, Seyfried NT, Cheng D, Xie Y, Robert J, Rush J, Hochstrasser M, Finley D, Peng J (2009) Quantitative proteomics reveals the function of unconventional ubiquitin chains in proteasomal degradation. Cell 137: 133-145 Yang SH, Galanis A, Witty J, Sharrocks AD (2006) An extended consensus motif enhances the specificity of substrate modification by SUMO. The EMBO journal 25: 5083-5093 Yang XJ (2005) Multisite protein modification and intramolecular signaling. Oncogene 24: 1653-1662 Ye Y (2006) Diverse functions with a common regulator: ubiquitin takes command of an AAA ATPase. Journal of structural biology 156: 29-40 Ye Y, Meyer HH, Rapoport TA (2003) Function of the p97-Ufd1-Npl4 complex in retrotranslocation from the ER to the cytosol: dual recognition of nonubiquitinated polypeptide segments and polyubiquitin chains. The Journal of cell biology 162: 71-84 Yeung HO, Kloppsteck P, Niwa H, Isaacson RL, Matthews S, Zhang X, Freemont PS (2008) Insights into adaptor binding to the AAA protein p97. Biochemical Society transactions 36: 62-67 Yin Y, Seifert A, Chua JS, Maure JF, Golebiowski F, Hay RT (2012) SUMO-targeted ubiquitin E3 ligase RNF4 is required for the response of human cells to DNA damage. Genes & development 26: 1196-1208

186
Yunus AA, Lima CD (2005) Purification and activity assays for Ubc9, the ubiquitin-conjugating enzyme for the small ubiquitin-like modifier SUMO. Methods in enzymology 398: 74-87 Yunus AA, Lima CD (2009) Structure of the Siz/PIAS SUMO E3 ligase Siz1 and determinants required for SUMO modification of PCNA. Molecular cell 35: 669-682 Zhang C, Roberts TM, Yang J, Desai R, Brown GW (2006) Suppression of genomic instability by SLX5 and SLX8 in Saccharomyces cerevisiae. DNA repair 5: 336-346 Zhao S, Ulrich HD (2010) Distinct consequences of posttranslational modification by linear versus K63-linked polyubiquitin chains. Proceedings of the National Academy of Sciences of the United States of America 107: 7704-7709 Zhao Y, Brickner JR, Majid MC, Mosammaparast N (2014) Crosstalk between ubiquitin and other post-translational modifications on chromatin during double-strand break repair. Trends in cell biology 24: 426-434 Zhao Y, Kwon SW, Anselmo A, Kaur K, White MA (2004) Broad spectrum identification of cellular small ubiquitin-related modifier (SUMO) substrate proteins. The Journal of biological chemistry 279: 20999-21002 Zhu J, Zhu S, Guzzo CM, Ellis NA, Sung KS, Choi CY, Matunis MJ (2008) Small ubiquitin-related modifier (SUMO) binding determines substrate recognition and paralog-selective SUMO modification. The Journal of biological chemistry 283: 29405-29415 Zou L, Elledge SJ (2003) Sensing DNA damage through ATRIP recognition of RPA-ssDNA complexes. Science (New York, NY 300: 1542-1548 Zunino R, Schauss A, Rippstein P, Andrade-Navarro M, McBride HM (2007) The SUMO protease SENP5 is required to maintain mitochondrial morphology and function. Journal of cell science 120: 1178-1188

T H E F A C U L T Y O F S C I E N C E , U N I V E R S I T Y O F C O P E N H A G E N The PhD School of SCIENCE
1/3 Revised 29 January 2013
3A. Co-authorship statement
All papers/manuscripts with multiple authors enclosed as annexes to a PhD thesis synopsis should contain a co-author statement, stating the PhD student’s contribution to the paper.
1.General information
PhD student
Name Julie Bonne Køhler Civ.reg.no. (If not applicable, then birth date) 2604841952 E-mail [email protected] Department Department of biology
Please mark PhD Scheme
x 5+3 Scheme 4+4 Scheme Industrial PhD
Principal supervisor
Name Geneviève Thon E-mail [email protected]
2.Title of PhD thesis
Interplays between sumoylation, SUMO-targeted Ubiquitin Ligases, and the ubiquitin-adaptor protein Ufd1in fission yeast
3.This co-authorship declaration applies to the following paper
“Concerted Action of the Ubiquitin-Fusion Degradation Protein 1 (Ufd1) and Sumo-Targeted Ubiquitin Ligases (STUbLs) in the DNA-Damage Response” Julie Bonne Køhler, Maria Mønster Jørgensen, Gabriele Beinoraité, Michael Thorsen, Geneviève Thon
The extent of the PhD student’s contribution to the article is assessed on the following scale
A. has contributed to the work (0-33%)
B. has made a substantial contribution (34-66%)
C. did the majority of the work independently (67-100%).



T H E F A C U L T Y O F S C I E N C E , U N I V E R S I T Y O F C O P E N H A G E N The PhD School of SCIENCE
1/3 Revised 29 January 2013
3A. Co-authorship statement
All papers/manuscripts with multiple authors enclosed as annexes to a PhD thesis synopsis should contain a co-author statement, stating the PhD student’s contribution to the paper.
1. General information
PhD student
Name Julie Bonne Køhler Civ.reg.no. (If not applicable, then birth date) 2604841952 E-mail [email protected] Department Department of biology
Please mark PhD Scheme
x 5+3 Scheme 4+4 Scheme Industrial PhD
Principal supervisor
Name Genevieve Thon E-mail [email protected]
2.Title of PhD thesis
Interplays between sumoylation, SUMO-targeted Ubiquitin Ligases, and the ubiquitin-adaptor protein Ufd1in fission yeast
3.This co-authorship declaration applies to the following paper
Quantitative identification of sumoylation sites in fission yeast wild-type cells, Ufd1 and SUMO-targeted ubiquitin ligase (STUbL) mutants Julie Bonne Køhler, Triin Tammsalu, Maria Mønster Jørgensen, Ron Hay, Geneviève Thon
The extent of the PhD student’s contribution to the article is assessed on the following scale
A. has contributed to the work (0-33%)
B. has made a substantial contribution (34-66%)
C. did the majority of the work independently (67-100%).