
Interleukin 6 enhancement of interleukin 3-dependent proliferation

Interleukin-3 promotes hemangioblast development in mouse aorta-gonad-mesonephros region
by Wen-Yan He, Yu Lan, Hui-Yu Yao, Zhuan Li, Xiao-Yan Wang, Xiu-Sen Li,Ji-Yan Zhang, Yu Zhang, Bing Liu, and Ning Mao
Haematologica 2009 [Epub ahead of print]
doi:10.3324/haematol.2009.014241
Publisher's Disclaimer. E-publishing ahead of print is increasingly important for the rapid dissemination of science.Haematologica is, therefore, E-publishing PDF files of an early version of manuscripts thathave completed a regular peer review and have been accepted for publication. E-publishingof this PDF file has been approved by the authors. After having E-published Ahead of Print,manuscripts will then undergo technical and English editing, typesetting, proof correction andbe presented for the authors' final approval; the final version of the manuscript will then appe-ar in print on a regular issue of the journal. All legal disclaimers that apply to the journal alsopertain to this production process.
Haematologica (pISSN: 0390-6078, eISSN: 1592-8721, NLM ID: 0417435, www.haemato-logica.org) publishes peer-reviewed papers across all areas of experimental and clinicalhematology. The journal is owned by the Ferrata Storti Foundation, a non-profit organiza-tion, and serves the scientific community with strict adherence to the principles of openaccess publishing (www.doaj.org). In addition, the journal makes every paper publishedimmediately available in PubMed Central (PMC), the US National Institutes of Health (NIH)free digital archive of biomedical and life sciences journal literature.
Official Organ of the European Hematology AssociationPublished by the Ferrata Storti Foundation, Pavia, Italy
www.haematologica.org
Early Release Paper
Support Haematologica and Open Access Publishing by becoming a member of the European Hematology Association (EHA)and enjoying the benefits of this membership, which include free participation in the online CME program
Copyright 2009 Ferrata Storti Foundation.Published Ahead of Print on December 8, 2009, as doi:10.3324/haematol.2009.014241.

DOI: 10.3324/haematol.2009.014241
1
Interleukin-3 promotes hemangioblast development in mouse
aorta-gonad-mesonephros region
Short title: Promoting AGM hemangioblast development by IL-3
Wen-Yan He1, 2, #
, Yu Lan3, #
, Hui-Yu Yao1, #
, Zhuan Li2, Xiao-Yan Wang
1, Xiu-Sen Li
1,
Ji-Yan Zhang4, Yu Zhang
5, Bing Liu
2, *, Ning Mao
1, *
1Department of Cell Biology, Institute of Basic Medical Sciences, Beijing 100850, China
2Laboratory of Oncology, Affiliated Hospital of Academy of Military Medical Sciences,
Beijing 100071, China; 3Genetic Laboratory of Development and Diseases, State Key
Laboratory of Proteomics, Institute of Biotechnology, Beijing 100071, China; 4Department
of Molecular Immunology, Institute of Basic Medical Sciences, Beijing 100850, China;
5Department of Immunology, Peking University Health Science Center, Beijing 100191,
China
*Corresponding authors: Ning Mao, Department of Cell Biology, Institute of Basic
Medical Sciences, Tai Ping Road 27, Beijing 100850, China, E-mail:
[email protected]; Bing L iu , Laboratory of Oncology, Affiliated Hospital of
Academy of Military Medical Sciences, Dongdajie 8, Beijing 100071, China, E-mail:
# These authors contributed equally to this study.
Key words: interleukin-3, hemangioblast, AGM, definitive hematopoiesis, hematopoietic
stem cells.

DOI: 10.3324/haematol.2009.014241
2
Background Hemangioblast represents bi-potential precursor with hematopoietic and
vascular capacities. In the mouse E7.0–7.5 embryos, the hemangioblast is identified by
clonal blast colony-forming cell (BL-CFC) assay or single cell OP9 co-culture. However,
ontogeny of hemangioblast in mid-gestation embryos is poorly defined.
Design and Methods The BL-CFC assay and the OP9 system were combined to
illustrate the hemangioblast with lymphomyeloid and vascular potential in the mouse
aorta-gonad-mesonephros (AGM) region. The colony-forming assay, RT-PCR,
immunostaining and flow cytometry were used to identify the hematopoietic potential, and
the Matrigel- or OP9-based methods were employed to evaluate the endothelial
progenitor activity.
Results Functionally, the AGM-derived BL-CFCs produced erythroid/myeloid
progenitors, CD19+ B lymphocytes, and CD3
+TCR_
+ T lymphocytes. Meanwhile, the BL-
CFC-derived adherent cells generated CD31+ tube-like structures on OP9 stromal cells,
validating the endothelial progenitor potential. The AGM-derived hemangioblast was
greatly enriched in the CD31+, Endomucin
+ and CD105
+ subpopulations, which
collectively pinpoints the endothelial layer as the main location. Interestingly, the BL-CFC
was not detected in yolk sac, placenta, fetal liver or embryonic circulation. Screening of
candidate cytokines revealed that interleukin-3 (IL-3) was remarkable in expanding the
BL-CFC in a dose-dependent manner through JAK2/STAT5 and MAPK/ERK pathway.
Neutralizing IL-3 in the AGM region resulted in reduced number of BL-CFC, reflective of
the physiological requirement. Both hematopoietic and endothelial differentiation potential
were significantly increased in IL-3-treated BL-CFC, suggesting a persistent positive
influence. Intriguingly, IL-3 markedly amplified primitive erythroid and macrophage
precursors in E7.5 embryos. Quantitative RT-PCR demonstrated declined Flk-1 and
elevated Scl and vWF transcription upon IL-3 stimulation, indicating accelerated
hemangiopoiesis.
Conclusions The hemangioblast with lymphomyeloid potential is one of the precursors
of definitive hematopoiesis in the mouse AGM region. Moreover, the regulatory role of IL-
3 is extended to the dual-potential hemangioblast concerning both number and capacity.

DOI: 10.3324/haematol.2009.014241
3
Introduction
There are generally two stages regarding early embryonic hematopoiesis in the mouse.
The first stage, the primitive hematopoiesis, is characterized by de novo generation of
nucleated erythrocytes (large and production of embryonic hemoglobin) in the extra-
embryonic yolk sac (YS). The second stage, the definitive hematopoiesis, is marked by
autonomous formation of adult type hematopoietic stem cells (HSCs) in E10.5 major
vessels or highly vascularized structures, i.e. the aorta-gonad-mesonephros (AGM)
region, umbilical/vitelline arteries, YS, and placenta (1-8). Both stages, though largely
differential in hematopoietic activities, closely intertwine with vascular lineage in two
speculated forms: the hemangioblast (a common precursor for hematopoietic and
endothelial lineages) and the hemogenic endothelium (dedifferentiation of mature
endothelium into hematopoietic lineage) (9-12).
Eighty years ago, the term of hemangioblast was proposed by the finding that, in
the YS, the aggregated mesodermal cells simultaneously gave rise to central primitive
erythrocytes and peripheral endothelial cells in close proximity, forming a distinct
structure called blood island. It was until 1998 that two independent groups provided the
first convincing evidence of hemangioblast by in vitro differentiation assay of mouse
embryonic stem cells (ES cells). Nishikawa et al. utilized collagen-based 2 dimensional
differentiation system with OP9 feeder layer to visualize the blood budding and the tube
formation from single Flk-1+CD144
+ cells (13). Alternatively, Choi et al. defined the clonal
blast colony-forming cells (BL-CFCs) with remarkable VEGF responsiveness in early
embryoid bodies (EB) (14). Recently, they revealed that the comparable BL-CFC
(Brachyury+Flk-1
+) predominantly arose at the posterior primitive streak of E7.5 embryos,
contrary to the presumed YS (15). Such a finding suggests that 1) the precursor
represents a transient mesoderm population undergoing hematopoietic and vascular
specification, and 2) hematopoietic commitment initiates prior to the establishment of YS
blood island. It is believed that the ES cell-derived hematopoietic differentiation
recapitulates the YS-stage hematopoiesis (16). Up to date, the ES cell/hemangioblast
models are instrumental to understand complicated molecular mechanisms underlying

DOI: 10.3324/haematol.2009.014241
4
the mesoderm-hemangioblast-hematopoietic commitment cascade. In comparison, the
hemangioblastic origin and molecular modulation of intra-embryonic definitive
hematopoiesis remain poorly understood. Because the AGM region-derived HSCs
possess nearly all the immunophenotypic features of endothelium (LDL receptor+, CD31
+,
CD144+, CD34
+ and Tie2
+), it is accepted that the hemogenic endothelium serves as the
direct precursor of HSC (17-20). Recently, we found that the high proliferation potential
precursors in the mouse AGM region could be categorized into 2 types: one restricted to
hematopoietic lineage, and the other with hemangioblastic properties (refereed to be
HPP-HA) (21). Of note, whether hemangioblasts (from either ES cells or embryos)
possess lymphoid potential remains unknown.
Here, we combine the BL-CFC and the OP9 systems to quantify hemangioblast with
B and T lymphoid potential in the mouse AGM region. Screening of candidate cytokines
reveals interleukin-3 (IL-3) a positive regulator of hemangioblast development in the AGM
region.

DOI: 10.3324/haematol.2009.014241
5
Design and methods
Embryo dissections
C57BL/6 mice were mated overnight. Noon of the day when a vaginal plug was detected
was designated as E0.5. At the time point as indicated, mice were killed, and embryos
were dissected free of decidual tissues and Reichter’s membrane. Somite-stage embryos
were staged by counting somite numbers. The embryonic tissues were treated with 0.1%
type I collagenase (Sigma, St. Louis, MO) at 37°C for 1 hour (for the AGM region,
placenta and YS) or 1-1.5 hours (for E7.5 embryos), and then dispersed into single-cell
suspension. Cell counts and viability were estimated by trypan blue exclusion.
BL-CFC and hematopoietic CFC assay
BL-CFC media was composed of 0.9% methylcellulose (Sigma), 2 mM glutamine
(Hyclone, Logan, Utah), 100U/ml;100!g/ml penicillin/streptomycin (Hyclone), 5% protein-
free hybridoma medium II (Gibco, Grand Island, NY), 200 !g/ml iron-saturated transferrin
(Sigma), 1% BSA (Stem Cell Technologies, Vancouver, Canada), 0.45 mM
monothioglycerol (MTG, Sigma), 15% FBS (Hyclone) and a cytokine cocktail including
rhbFGF (10 ng/ml), rmSCF (50 ng/ml), rhIL-6 (5 ng/ml), rhIGF-I (50 ng/ml), rmLIF (103
U/ml) and rhVEGF-165 (5 ng/ml). Following 4-5 days of inoculation, blast colonies were
scored. For hematopoietic CFC assay, cytokine cocktail included rmSCF (50 ng/ml),
rmIL-3 (10 ng/ml), rhIL-6 (5 ng/ml), rhIL-11 (10 ng/ml), rmGM-CSF (10 ng/ml), rhTpo (20
ng/ml), rhEpo (3 U/ml). Primitive hematopoietic colonies were scored at day 4 to 7 of
culture, whereas definitive erythroid colonies were counted after 3 to 5 days of
incubation. CFU-GM, BFU-E and CFU-Mix were counted after 7 to 10 days of culture.
In vitro tube formation assay
Individual blast colonies were plucked and transferred to single wells of 96-well plates
containing IMDM with 10% FBS, 10% horse serum, rhbFGF (10 ng/ml), rhVEGF-165 (5
ng/ml), rhIGF-I (10 ng/ml), rmSCF (50 ng/ml), rmIL-3 (10 ng/ml), rhIL-11 (10 ng/ml),
rhEpo (3 U/ml) ,ECGS(100!g/ml), L-glutamine (2 mM) and 0.45 mM MTG. After 4 days of
incubation, nonadherent cells were removed and the adherent cells were inoculated with
endothelial growth medium-2 (EGM2, Cambrex, Walkersville, MD) for further 6 days.

DOI: 10.3324/haematol.2009.014241
6
They were then trypsinized and replated into single wells of 48-well plates pre-coated
with Matrigel (Biosciences BD, Bedford, MA). Tube-like and network structures were
documented after 12-24 hours of culture. Measurement was performed in four
independent fields. For LDL uptake assay, the adherent cells were cultured in the
presence of 10 _g/ml DiI-Ac-LDL (Biomedical Technologies, Inc., Stoughton, MA) for 4
hours. Tube-like structures were fixed in 4% paraformaldehyde for 30 minutes, and were
incubated with primary antibodies against _-SMA (Sigma) or Calponin (Sigma) overnight
at 4°C. Then they were washed 3 times and incubated with FITC-conjugated goat anti-
mouse IgG (Southern Biotechnology Associates, Inc.) for an additional 60 minutes at
room temperature.
OP9-based hematopoietic and endothelial progenitor potential assay
Individual blast colonies were plated on the OP9 stromal cells in the presence of VEGF
(200 ng/ml), FL (20 ng/ml) and IL-7 (20 ng/ml). After 10 days of culture, they were fixed in
4% paraformaldehyde for 30 minutes, and were incubated with monoclonal antibody
against CD45 (PE-conjugated, eBioscience, San Diego, CA) overnight at 4°C. Thereafter,
a two-step staining protocol was used to identify the endothelial development. Following
a primary anti-CD31 antibody (BD Pharmingen) incubation, the culture was visualized by
the indirect immunoperoxidase method using horseradish peroxidase–conjugated anti-rat
IgG (Zymed, South San Francisco, CA, USA). The peroxidase activity was visualized
using 3,38-diaminobenzidine (Zymed).
To determine B lymphoid potential, blast colonies were replated on fresh OP9
stromal cells in the presence of FL (20 ng/ml) and IL-7 (20 ng/ml). After 7-10 days of
culture, B lymphocyte differentiation was assessed.
To generate T lymphocytes, blast colonies were replated onto OP9-DL1 stromal
cells supplemented with FL (5 ng/ml), IL-7 (5 ng/ml) and TPO (5 ng/ml). After 4 days of
culture, TPO were withdrawn. T lymphocyte differentiation was analyzed after further 11-
16 days of culture.
For flow cytometry analysis, the non-adherent cells in the co-culture were harvested
and stained for 30 minutes at 4°C with fluorescein isothilcyanate (FITC)-conjugated or

DOI: 10.3324/haematol.2009.014241
7
phycoerythrin (PE)-conjugated monoclonal antibodies. Antibodies recognizing CD19,
B220, IgM, CD3, CD4 and CD8 were purchased from eBioscience (San Diego, CA,
USA). Antibodies against TCR ! chain and NK1.1 were purchased from BD Biosciences
(PharMingen, Franklin Lakes, NJ, USA). Flow cytometry was performed using a
FACScalibur instrument (BD Biosci-ences, San Diego, CA, USA).
Magnetic sorting
Single-cell suspension from AGM region was labeled with purified anti-mouse CD31 (BD
Pharmingen), Endomucin, biotin-conjugated CD105 (Endoglin) or PDGFRß monoclonal
antibodies (eBioscience), followed by incubating with anti-rat or anti-biotin microbeads.
Magnetic sorting was performed according to the manufacture’s protocol and processed
through an MS-separation column attached to a Mini-MACS separation unit (Miltenyi
Biotech).
Nested RT-PCR, RT-PCR and real-time PCR
Nested RT-PCR for individual colonies was performed as described previously (21). For
RT-PCR analysis, total RNA from E7.5 embryos or E10.5 AGM region were extracted
using TRIZOL reagent (Invitrogen) and reverse transcribed using mRNA Selective PCR
Kit (TaKaRa). Blast colonies were harvested and analyzed by quantitative real-time PCR
as described previously (22). The specific primer sequences and amplified product size
are listed in Table S1 to S3 respectively.
Immunofluorescence staining
Blast colony cells were stained with monoclonal antibodies against Sca-1 (PE-
conjugated), Ter-119 (PE-conjugated), or F4/80 (FITC-conjugated, eBioscience) in PBS
containing 0.1% BSA at 4°C for 30 minutes. For adherent cells, they were fixed in 4%
paraformaldehyde for 10 minutes, and were incubated with primary antibodies against
CD45 (eBioscience) overnight at 4°C. Then they were washed 3 times and incubated
with F4/80 (FITC-conjugated, eBioscience), FITC-conjugated goat anti-rat IgG (Southern
Biotechnology Associates, Inc. Birmingham, Alabama) for an additional 60 minutes at
room temperature.

DOI: 10.3324/haematol.2009.014241
8
Immunohistochemistry
Immunohistochemistry was performed as described previously (22). The primary
antibodies used were as follows: anti-CD31, anti-Endomucin, and anti-PDGFR! (Cell
Singaling).
Western blotting
Total proteins of AGM cells were harvested. 60 micrograms of proteins were loaded on
12% SDS-PAGE. Immunoblotting was performed using anti-Stat5, anti-Phospho-Stat5,
anti-Erk1/2, anti-Phospho-Erk1/2 and anti-Actin antibodies (Cell Signaling Technology,
Danvers, MA) according to the manufacturer’s instructions.
Other reagents
Oncostatin M (OSM) was obtained from R&D Systems (Minneapolis, MN, USA). IL-3-
blocking antibody and IgG1 were purchased from BD PharMingen (San Diego, CA,
USA). The MEK1/2 inhibitor PD98059 was purchased from Promega (Madison, WI, USA)
and the JAK2 inhibitor AG490 from Sigma.
Phase contrast and fluorescence imaging
Phase contrast images were collected by using a Nikon TE2000-U (Tokyo, Japan)
inverted microscope with Nikon 4x/0.13 NA, 10_/0.30 NA, 20x/0.45 NA Plan Fluor
objective lenses. Images were acquired by using a digital camera (Cool SNAP 5.0; Roper
Scientific, Tucson, AZ, USA). Fluorescence and confocal images were collected by a
zeiss microscope (LSM 510 META system, Germany). Composite images were
assembled in Adobe Photoshop version 7.0 (Adobe Systems, Munich, Germany)
Statistical analysis
The results were expressed as the means ± s.e.m. Significance is determined using the
Student’s t- test: *, P<0.05; **, P<0.01, compared with the control data.

DOI: 10.3324/haematol.2009.014241
9
Results
Characterization of BL-CFC in the mouse AGM region
To fulfill clonal BL-CFC analysis, single-cell suspension from mouse E10.5 AGM region
(0.5-1x105 cells/ml) was plated in methylcellulose media containing bFGF, SCF, VEGF,
LIF, IGF-I and IL-6. After 4-5 days of incubation, the typical blast colonies with unique
and identifiable morphology emerged. The blast colony was composed of a compact core
and a loose peripheral layer with clear intercellular boundaries (Fig. 1A). Under such a
condition, the typical hematopoietic colonies were rare and could be easily identified. The
blast colony predominantly consisted of immature cells with a high nuclear/plasma ratio
as demonstrated by Wright-Giemsa staining (data not shown). Nested RT-PCR assay of
individually plucked colonies revealed that most of them harbored the hemangioblast-like
molecular pattern -- Brachyury-, Flk-1
+, and Runx1
+ (Fig. 1B). Immunofluorescence
staining demonstrated that the colonies contained Sca-1+, F4/80
+ and Ter119
+ cells (Fig.
1C). The data suggested that the AGM region-derived blast colonies progressed beyond
the mesoderm and underwent hemangioblastic commitment.
Then we attempted to determine the critical cytokines for generating the blast
colonies from the AGM region. By eliminating a cytokine from the original formula
(SCF+bFGF+IL-6+IGF-I+LIF+VEGF) one by one, we found that bFGF and SCF had the
strongest effect on colony growth (p<0.01). The removal of IL-6 had a modest effect on
the number of colonies (p<0.05) (Fig. S1).
To validate the hematopoietic potential, the blast colonies were individually
harvested and replated into hematopoietic CFC system containing SCF, IL-3, IL-6, IL-11,
Epo, TPO and GM-CSF. As demonstrated in Fig. 1D, they gave rise to CFU-E, CFU-GM,
CFU-Mix, and BFU-E. Of 12 representative colonies, all could generate hematopoietic
CFCs (Fig. 1E). On average, a single BL-CFC yielded 5.7 CFU-E, 1.9 CFU-GM, and 5.7
CFU-Mix.
To further identify the vascular potential, a single blast colony was cultured in a
liquid expansion system with hematopoietic and endothelial cytokines (bFGF, VEGF,
SCF, IL-3, IL-11, Epo, IGF-I and ECGS). After 24h of incubation, the adherent cells were

DOI: 10.3324/haematol.2009.014241
10
evident in over 97% (64/66) colonies. In a couple of days, they amplified significantly
(Fig. 1F). Meanwhile, non- or semi-adherent hematopoietic clusters developed in the
upper layer and retained hematopoietic colony-forming capacities (data not shown). At
day 4, the non-adherent hematopoietic population was depleted and the culture was
replaced with the EGM-2 to support vascular differentiation and maturation. At day 10,
the adherent cells with cobble-stone or spindle-like morphology were predominant in the
culture (Fig. 1G). Most of them incorporated DiI-Ac-LDL, but occasionally expressed
CD45 and F4/80 (Fig. 1H, I). It indicated that the endothelial lineage (other than
hematopoietic/monocytic cells) prevailed in the culture. At the same time, the _-SMA+
cells appeared in the adherent layer (Fig. 1J). Next, in vitro tube forming assay was used
to evaluate whether the adherent population was functionally equivalent to the vascular
cells. As shown in Fig. 1K, the adherent cells from 45 of a total of 60 colonies were
capable of forming tube-like structures in Matrigel. Alternatively, the adherent cells were
labelled with DiI-Ac-LDL prior to replating. The participation of elongated LDL-
incorporating endothelial cells and _-SMA+ or Calponin
+ cells in the capillary formation
was visualized (Fig. 1L, M). Of note, under appropriate stimuli, the blast colonies failed to
generate osteogenic and adipogenic lineages in vitro (data not shown).
OP9, the bone marrow stromal cell line, is known to provide appropriate
microenvironment for endothelial and lymphocyte differentiation (13, 23, 24). To further
validate the hemangioblastic activity, 50 blast colonies were individually co-cultured with
OP9 cells for 10 days. All of them demonstrated both hematopoietic and endothelial
progenitor potential. As shown in Fig. 2A, 4 representative colonies simultaneously
generated abundant CD45+ hematopoietic cells and CD31
+ tube-like structures on OP9
stromal cells.
To determine B lymphoid potential, blast colonies were incubated with OP9 in the
presence of FL and IL-7 for 7-10 days. After 6 days of culture, significant proliferation of
round non-adherent cells was detected (Fig. 2B). B220 (CD45R) is expressed at all
stages of B-cell ontogeny. However, CD19 remains the most reliable marker for B-
lineage commitment. As shown in Figure 2C, abundant B220+ and CD19
+ populations

DOI: 10.3324/haematol.2009.014241
11
were generated from blast colonies. Mature B cells were detected in approximately 1.5%
of total cells by analyzing surface expression of IgM.
To further assay T lymphoid potential, blast colonies were co-cultured with OP9-
DL1 cells (proven to support T lymphocyte development from hematopoietic stem
progenitor cells and ES cells) (24, 25). After 8 days of culture, round non-adherent cells
proliferated abundantly (Fig. 2D). After 15-20 days, the CD3+TCR_
+ and CD4
+/CD8
+ T
lymphocytes were generated (Fig. 2E). Also, NK1.1+ natural killer cells emerged in the
culture. Furthermore, 6 blast colonies were individually expanded and tested for the B
and T potential. All demonstrated both B (CD19+) and T (CD3
+TCR_
+) lymphocyte
potential (Table S4). Compared with the OP9 co-culture, differentiation of CD19+ B
lymphocytes was greatly suppressed in the OP9-DL1 system (data not shown).
Therefore, the blast colonies contained hematopoietic precursors with both myeloid and
lymphoid potential, indicating the definitive hematopoietic identity.
As previously described (14, 15), two steps were taken to validate the clonality of
the AGM-derived blast colonies. First, cells from the AGM region were plated at different
concentrations. As shown in Fig. 3A, the number of blast colonies was directly
proportional to that of AGM cells input, indicating that a single cell corresponded to the
growth of a colony. Second, the cell mixing analysis was carried out (Fig. 3B). An equal
number of cells from the AGM region of male wild type (Sry+GFP
-) and female GFP
transgenic (Sry-GFP
+) mice were mixed at the density of 0.5-1x10
5 cells/ml. After 4 days
of culture, the blast colonies were picked out and expanded individually to determine the
content of genomic DNA. Among 20 representative colonies analyzed, 10 displayed
exclusively Sry+GFP
- genotype and the rest 10 Sry
-GFP
+ (Fig. 3B). Representative blast
colonies from wild type and GFP transgenic AGM region in the cell mixing analysis were
shown in Fig. 3C. Therefore, the experiments suggested that the blast colony originated
from a single cell.

DOI: 10.3324/haematol.2009.014241
12
Spatiotemporal distribution and surface markers of BL-CFC in the mouse AGM
region
To characterize the developmental kinetics of BL-CFC in the caudal half (E9.5) or AGM
region (E10.5-12.5), the tissues were analyzed daily between E9.5 and E12.5. As shown
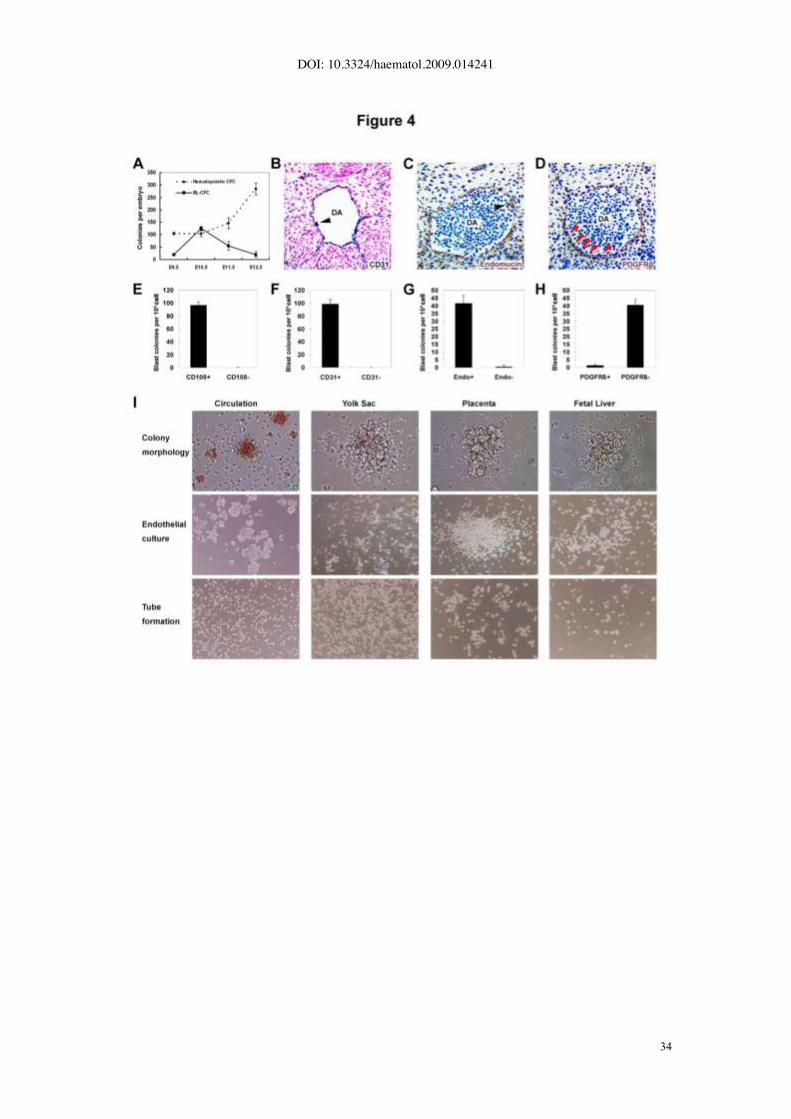
in Figure 4A, E9.5 caudal half contained 19.3±2.1 BL-CFC on average, and the number
achieved maximum of 125.0±8.0 at E10.5, followed by a significant decline to 54.8±17.6
at E11.5 and 20.0±10.6 at E12.5. The kinetics of BL-CFC was different from that of
hematopoietic CFC peaking in number at E12.5. A further separation of dorsal aorta with
surrounding mesenchyme (AoM) from gonad and mesonephros (GM) in the E11.5 AGM
region revealed that over 95% of BL-CFCs were concentrated in the former subregion
(data not shown).
Next, four molecules were selected to determine the location of BL-CFC in the AGM
region. As shown in Fig. 4B, C and previous reports (26, 27), CD31, Endomucin, and
CD105 had similar expression pattern (restricted to the endothelial layers and intro-aortic
hematopoietic clusters). In contrast, PDGFR_ is expressed by mural cells (Fig. 4D), and
its response to PDGF secreted by endothelial cells results in migration and maturation of
mural cells (28). As demonstrated in Fig. 4E-G, the E10.5 AGM-derived BL-CFCs were
enriched by CD31, Endomucin, and CD105 exclusively. Together with the finding that
most of BL-CFCs were negative for PDGFR_ (Fig. 4H), we concluded that the
hemangioblasts were mainly located within the endothelium lining the vessel wall of
dorsal aorta.
Of note, the blast colonies were not readily detected in cells from yolk sac (E9.5-
E11.5), placenta (E9.5-E11.5), fetal liver (E10.5-E11.5) and embryonic circulation (E10.5-
E12.5) (Fig. 4I).

DOI: 10.3324/haematol.2009.014241
13
Positive regulation of AGM-derived BL-CFC by IL-3
As the BL-CFC model was quantitative, a set of hematopoietic/vascular cytokines (i.e.,
bFGF, SCF, GM-CSF, IL-6, OSM, VEGF, TPO, IL-3, EPO) were screened for their
modulation on the hemangioblast development in the mouse AGM region. Initially, single-
cell suspension of the AGM region was incubated with indicated cytokines for 12h, and
then collected for the BL-CFC assay. As anticipated, a significant expansion of BL-CFC
was detected in the presence of bFGF (10 ng/ml). However, among other cytokines, only
IL-3 (50 ng/ml) could remarkably amplify the BL-CFC (148.0±22.5 Vs 46.7±8.6, p<0.01)
and the effect appeared more profound than bFGF (Fig. 5A). Interestingly, IL-3-
stimulated BL-CFCs formed typical blast colonies at day 2.5 of culture, 12h earlier than
the control. Furthermore, the effect was dose dependent (Fig. 5B). As shown in Fig. 5C,
without IL-3 treated, the number of BL-CFC gradually dropped after in vitro incubation. In
contrast, a significant expansion was detected after 6h- (165.7±21.0 Vs control of
93.3±23.5, p<0.01) and 12h-incubation with IL-3 (210.3±41.2 Vs control of 93.3±23.5,
p<0.01). However, a prolonged culture to 24h resulted in a dramatic decrease
(103.2±31.7), indicating that IL-3 failed to support hemangioblast expansion in vitro for a
long duration. To determine whether IL-3 was physiologically required, the IL-3
neutralizing antibody and the isotype control (IgG1) were used. Compared with the IgG1,
neutralization of tissue-derived IL-3 reduced the number of BL-CFC (31.3±4.5 Vs
67.7±1.5, P<0.01) (Fig. 5D). Of note, the result was obtained by continuous exposure to
neutralizing antibody from the AGM incubation to the BL-CFC assay. It implied that the
cytokines in the BL-CFC assay could compensate for a transient absence of endogenous
IL-3 signals during 12h incubation.
IL-3 signaling pathway has been well characterized (29). Following IL-3 stimulation,
an immediate response of cells is the tyrosine phosphorylation of Janus kinase-2
(JAK2)/STAT5 proteins as well as the activation of Ras, Raf, MEK and ERK.
Subsequently, these signals are transduced to the nucleus, where they bind specific DNA
elements to activate transcription of target genes. As demonstrated in Fig. 5E,
supplement of IL-3 enhanced the phosphorylation of STAT5 and ERK1/2 in the AGM

DOI: 10.3324/haematol.2009.014241
14
cells. To further determine the IL-3-mediated signaling involved in hemangioblast
expansion, the AGM cells were stimulated with IL-3 for 12h in the presence of
pharmacological inhibitors or vehicle control (DMSO). The result revealed >50% inhibition
of BL-CFC amplification (P<0.01) when cells were incubated with AG490 (inhibitor of
JAK2, 100 !M). The use of PD98059 (inhibitor of MEK1/2, 50 !M) demonstrated similar
result (P<0.01). Thus, IL-3 expanded BL-CFC at least partially through the JAK2/STAT5
and MAPK/ERK signaling (Fig. 5F).
Increased hematopoietic and vascular potential of IL-3-treated BL-CFC
We carried out differentiation assay to determine the regulatory roles of IL-3 in the
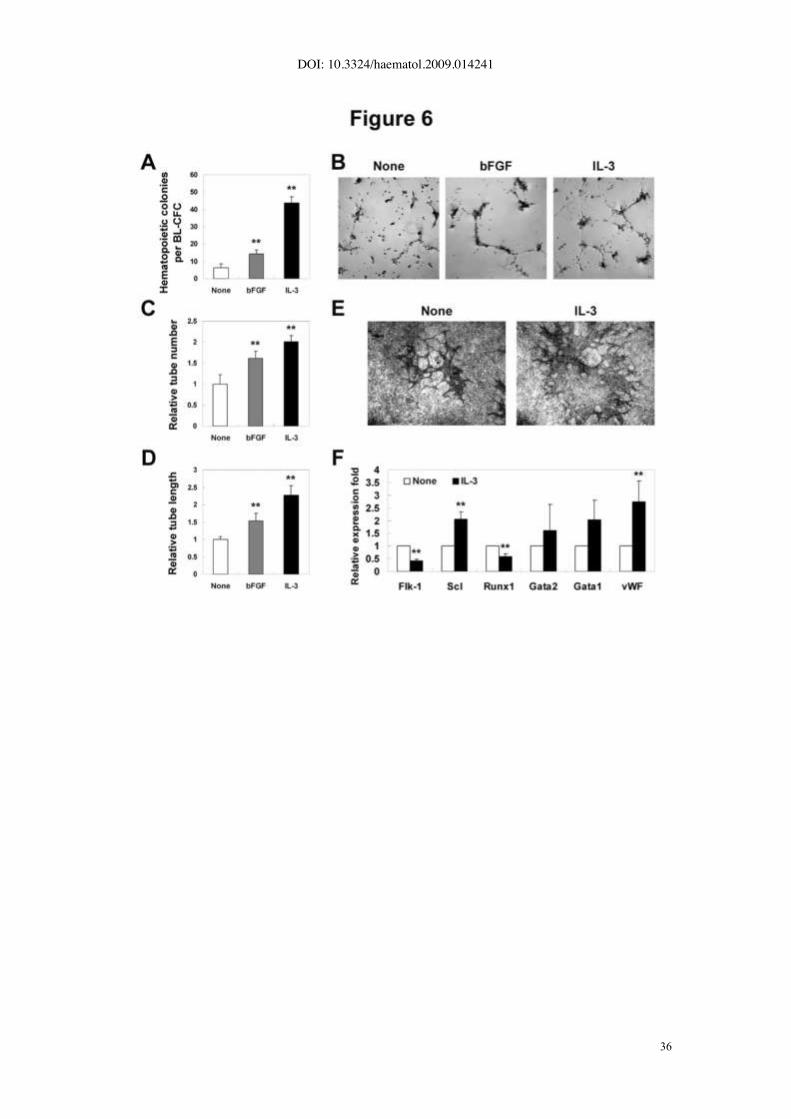
hematopoietic/vascular potential of BL-CFCs. As shown in Fig. 6A, the IL-3-stimulated
BL-CFC gave rise to increased number of CFCs (43.7±3.5 Vs 6.4±2.1 per blast colony,
p<0.01), more profound than bFGF-treated group (p<0.01). Furthermore, quantification of
in vitro tube-forming capacity on Matrigel revealed that IL-3-stimulated BL-CFCs
displayed significant increase in the tube number (2.0±0.1 fold, p<0.01) as well as the
tube length (2.3±0.3 fold, p<0.01) (Fig. 6B-D). Alternatively, IL-3 was added during the
formation of blast colonies. As shown in Fig.6E, the IL-3-treated blast colonies generated
increased number of CD31+ tubes on OP9 stromal cells, confirming the enhanced
endothelial progenitor capacity.
Subsequently, several hematopoietic- and vascular-related genes were selected to
define the IL-3-mediated molecular changes in the AGM cells. Upon stimulation, up-
regulated expression of Scl and vWF were detected, corroborating the enhanced
hematopoiesis and endothelial differentiation in BL-CFC assay (Fig. 6F). Of interest,
expression of Runx1 was significantly decreased, presumably reflecting its complicated
and dose-dependent involvement in regulating embryonic hematopoiesis.
These data collectively indicated that incubation with IL-3 led to elevated number as
well as intrinsic hemangiopoietic commitment of the hemangioblast.

DOI: 10.3324/haematol.2009.014241
15
IL-3 promoted primitive hematopoiesis in E7.5 embryos
The first wave of hemangioblast (BL-CFC) is detected mainly in posterior primitive streak
of E7.5 embryos. Of note, the number of BL-CFC in this stage is small, and the hypoxia
culture condition is critical. We only detected typical hematopoietic other than BL-CFC
colonies in E7.5 embryos. Once transferred to the liquid expansion media, these
hematopoietic-like colonies remained non-adherent and failed to expand as adherent
population. As shown in Fig. 7A, expression of two types of IL-3 receptors (IL-3R_/ßC, IL-
3R_/ßIL-3) was detected by RT-PCR in both E10.5 AGM and E7.5 embryos. After
incubation with IL-3 for 12h, single-cell suspension of E7.5 embryos was subjected to the
CFC assay. Hematopoietic colonies were classified as 3 types of primitive hematopoietic
progenitors, i.e. CFU-EryP, CFU-Mac and CFU-EM (Figure 7C). As shown in Fig. 7B, D,
IL-3 could significantly amplify CFU-EryP (23.7±3.1 Vs 6.3±2.5, p<0.01), CFU-M
(17.0±4.0 Vs 3.0±1.0, p<0.01) and CFU-EM (11.3±2.5 Vs 2.0±0, p<0.01). Quantitative
RT-PCR revealed that, upon IL-3 stimulation, transcriptional changes of the
hematopoietic- and vascular-related genes were similar to those of E10.5 AGM, including
down-regulation of Flk-1 and up-regulation of Scl and vWF. In contrast, expression of
Runx1 and GATA1 was significantly elevated, demonstrating somewhat different
mechanism of IL-3 on regulating primitive and definitive hematopoiesis (Fig. 7E).
Collectively, these data indicated that IL-3 was a positive modulator of the first wave of
e m b r y o n i c hematopoiesis.

DOI: 10.3324/haematol.2009.014241
16
Discussion
It remains ill defined whether hemangioblast serves as the precursor of intra-embryonic
definitive hematopoiesis. Here, we show that the AGM-derived BL-CFCs are capable of
generating various kinds of lymphomyeloid progenitors and vascular lineages in vitro,
representing a canonical definitive hemangioblast. Based on the model, the novel role of
IL-3 in governing ontogeny and differentiation of hemangioblast is demonstrated.
Characterization of hemangioblast with lymphomyeloid potential in the mouse
AGM region
The precursor of definitive hematopoiesis in the AGM region has been supposed as
hemogenic endothelium or differentiated mesoderm cells, i.e., hemangioblast and
mesenchyme. Thus, the site may harbor multiple hematopoiesis-related precursors, with
their sequential maturation and migration under dynamical but stringent control.
The existence of hemangioblast in the AGM region is initially implied by a bulk
culture system, where hematopoietic clusters are budded from proliferating endothelial
layers in the presence of bFGF, SCF and OSM (30). Recently, the HPP-HA model has
been used to quantify the hemangioblasts with high proliferating potential in the AGM
region (21). In comparison, the BL-CFC assay revealed more progenitors with
hemangioblastic activity in the AGM region (E10.5: 125 BL-CFC Vs 8 HPP-HA per
tissue). More importantly, it demonstrated the lymphoid capacity of hemangioblast in
vitro.
Based on the analysis of 4 membrane proteins and the knowledge regarding their
distribution in the AGM region, we concluded that the endothelial layer of dorsal aorta
(CD31+, Endomucin
+, CD105
+ and PDGFR!
-) was the main location for BL-CFCs. In
particular, the identity of the AGM-derived BL-CFC as authentic hemangioblast other than
more primitive precursor was reinforced by its failure to generate additional mesoderm
progenies such as bone and fat. Interestingly, the BL-CFC was not detected in yolk sac
and placenta. It may reflect different nature of hematopoietic precursors in various blood-
forming tissues. Alternatively, the AGM region (a hemogenic other than hematopoietic

DOI: 10.3324/haematol.2009.014241
17
site) may provide a unique niche to maintain the hematopoietic precursors in a primitive
undifferentiated state. The absence of BL-CFC in circulation and fetal liver suggests that
the AGM-derived hemangioblast may differentiate and become undetected upon
intravasation.
Several lines of evidence support the hypothesis that hemogenic endothelium may
be the direct precursor of definitive hematopoiesis in the AGM region. Firstly,
morphological analysis has shown that the cells in the hematopoietic clusters form tight
junction with the endothelial cells of the dorsal aorta, and appear to be budding from the
endothelium (31, 32). Furthermore, these cells express either endothelial or
hematopoietic markers, i.e. CD144, CD31, CD34 and CD45, indicative of their ongoing
immunophenotypical transition from endothelium to blood. Of interest, North et al.
pinpoint that, unlike normal embryos (33), the mesenchymal cells (CD45-CD34
-) can
function as HSC in the Runx1+/-
embryos. Most recently, using in vitro ES cell
differentiation system, Christophe Lancrin et al. demonstrate that the hemangioblast
generates hematopoietic cells through a hemogenic endothelium stage, providing the first
direct link between the two types of precursors (34). Thus, it will be intriguing to
determine whether the BL-CFC in the AGM region is capable of generating
hematopoietic lineages through the hemogenic endothelium or other unknown
intermediates. Virtually, in close association with dorsal aorta, the mesoangioblast is
identified as a common precursor for vascular and extra-vascular mesodermal
derivatives. Compared with the BL-CFC, the mesoangioblast emerges at a higher
frequency, unable to proliferate in semisolid culture, and fails to generate typical
hematopoietic colonies (35, 36). Therefore, it is challenging to illustrate relationships
between hemogenic endothelium, hemangioblast and mesoangioblast in the AGM
region.

DOI: 10.3324/haematol.2009.014241
18
IL-3-mediated positive regulation of hemangioblast development in the mouse
AGM region
Up to date, in vitro differentiation of ES cells has provided substantial understanding
of hemangioblast regulation by intrinsic transcriptional factors or microenvironment cues
(9-12, 37). Particularly, a precise and stage-specific control of the differentiation cascade
(Brachyury+ meseoderm / Flk-1
+ hemangioblast / CD41
+ or CD45
+ hematopoietic cells)
has been linked to multiple molecules including BMP4, bFGF, Activin A, Wnt, Runx1, Scl,
Smad5, Smad1, CD105, Hex and GATA2 et al (38-56). In striking contrast, little is known
regarding the modulation of embryonic hemangioblast development. Among currently
tested cytokines, IL-3 appeared to be the most potent stimulator of the AGM
hemangiopoiesis. In line with this, Robin et al. reveal that IL-3 can remarkably amplify
HSCs with long-term reconstitution capacity in the AGM region up to 30 folds (57). They
attempt to define which target gene can rescue the defective HSC ontogeny of Runx1-/-
embryos, and find IL-3 other than GM-CSF can fulfill such a rescue by promoting survival
and proliferation of HSCs in the AGM region. Likewise, 12 h of IL-3 treatment was
capable of preventing apoptosis of CD31+ cells in the AGM region (data not shown).
Together with the temporal analysis of BL-CFC development in vitro after IL-3 stimulation,
we concluded that IL-3 was more of a survival than an expanding factor on
hemangioblast. Most currently, Taoudi S et al. report that the cytokine cocktail containing
IL-3, SCF and FL can promote HSC generation within a novel reaggregate composed of
CD144+CD45
+ cells from the AGM region (58). While addition of IL-3 to the explant
culture of E10 AGM leads to precocious HSC activity (57), the IL-3-treated AGM cells
forms the blast colonies 12h earlier than the control. The similarity suggests that IL-3 can
fuel the intrinsic hematopoietic potential of pre-hematopoietic precursors in addition to
increase the number of hemangioblast. In comparison, IL-3-induced expansion of HSC
appears more drastic than that of hemangioblast. The discrepancy may be ascribed to a
longer duration of IL-3 incubation (3 days Vs 12 h) and more importantly, to the 3-
dimentional organ culture advantageous for maintaining supportive microenvironment
cues. Of note, bFGF fails to promote HSC expansion, in striking contrast to its

DOI: 10.3324/haematol.2009.014241
19
unambiguous function on avian and mouse hemangioblast (21, 42, 59). It may reflect
that, unlike IL-3, bFGF-induced BL-CFC can produce more hematopoietic progenitors
other than HSCs. In previous report, a bulk AGM culture reveals that OSM can promote
hemogenesis from endothelial cells (30). However, OSM expands neither HSC nor BL-
CFC, suggesting that it may predominantly accelerate the generation of hematopoietic
progenitors from hemogenic endothelium or hemangioblast (57). Thus, the multi-step
control of embryonic hemangiopoiesis by candidate molecules can be accurately
analyzed by the BL-CFC assay.
Albeit dispensable, the regulatory roles of IL-3 in the AGM hematopoiesis are
physiologically important. The expression of IL-3 is detected mainly in the cells lining the
lumen or attaching to the endothelium of dorsal aorta by in situ hybridization (57). Thus,
in the mouse AGM region, the HSC and BL-CFC with endothelial immunophenotype
(CD31+Endomucin
+) are directly regulated by the autocrine or paracrine IL-3.
Consistently, the use of IL-3-blocking antibody leads to decreased activity of HSC as well
as BL-CFC (57). Direct transplantation of the E11 IL-3-/-
AGM cells into irradiated
recipients results in undetectable donor-derived HSC activity (57). Indeed, IL-3 acts as a
target gene of Runx1, and can rescue HSC defect in the Runx1-/-
AGM region (57). Here,
upon IL-3 stimulation, the expression of Runx1 in the AGM cells is significantly down-
regulated, implying a possible negative feedback. In line with our data, the hemangioblast
and hematopoietic specification from mesodermal precursors is accelerated in the Runx1
heterozygous EBs (60). Within Runx1+/-
embryos, HSC activity is reduced in the AGM
region, contrasting its increase in the YS and the placenta. Therefore, the gene dosage
of Runx1 markedly affects spatiotemporal distribution of HSC in the mid-gestation blood-
forming tissues (33, 61).
Another novel finding is the positive role of IL-3 in the stage of primitive
hematopoiesis. The dramatic amplification of primitive hematopoietic precursors by short-
time stimulation of IL-3 may be largely ascribed to a rapid hemogenesis from preexisting
precursors, such as Flk-1+ mesodermal cells or hemangioblasts. In agreement with this,
expression of Flk-1 is declined in the presence of IL-3, with elevated transcription of a

DOI: 10.3324/haematol.2009.014241
20
panel of transcriptional factors such as Scl and GATA1. As defective primitive
hematopoiesis is undetected in the gastrulating IL-3 mutants, the data may reveal a
cytokine redundancy.
Most recently, Zeoli et al. report that IL-3 is capable of expanding angiogenic cells
from adult CD45+ hematopoietic progenitors via the STAT5 signaling and promoting
arterial specification (62). Taken together, these data suggest that IL-3 can exert
functions on the hemangiopoietic system from active embryogenesis to homeostatic
adulthood.
Authorship and Disclosures
Wen-Yan He: performed research, analyzed data, wrote the paper; Yu Lan: performed
research, analyzed data, wrote the paper; Hui-Yu Yao: performed research, analyzed
data; Zhuan li: performed research; Xiao-Yan Wang: performed research; Xiu-Sen Li:
performed research; Ji-Yan Zhang: analyzed data; Yu Zhang: designed research,
analyzed data; Bing Liu: designed research, analyzed data, wrote the paper: Ning Mao:
designed research, analyzed data, wrote the paper.
The authors declare no competing financial interests.
Funding
This study was supported by the National Natural Science Foundation (no. 30730043,
30770921, 30871437, 30800427), the National Key Basic Research Program of China
(no. 2005CB522705, 2005CB522506 and 2006CB943601), the National 863 program
(no. 2007AA021109), the Beijing Natural Science Foundation (no. 5092024), and the
S ta te K e y Laboratory of Proteomics_ gran t N o . SKLP-Y200807.

DOI: 10.3324/haematol.2009.014241
21
References
1. Ferkowicz MJ, Yoder MC. Blood island formation: longstanding observations and modern
interpretations. Experimental hematology. 2005 Sep;33(9):1041-7.
2. Gekas C, Dieterlen-Lievre F, Orkin SH, Mikkola HK. The placenta is a niche for
hematopoietic stem cells. Developmental cell. 2005 Mar;8(3):365-75.
3. McGrath KE, Palis J. Hematopoiesis in the yolk sac: more than meets the eye.
Experimental hematology. 2005 Sep;33(9):1021-8.
4. Ottersbach K, Dzierzak E. The murine placenta contains hematopoietic stem cells within
the vascular labyrinth region. Developmental cell. 2005 Mar;8(3):377-87.
5. Cumano A, Godin I. Ontogeny of the hematopoietic system. Annual review of
immunology. 2007;25:745-85.
6. Samokhvalov IM, Samokhvalova NI, Nishikawa S. Cell tracing shows the contribution of
the yolk sac to adult haematopoiesis. Nature. 2007 Apr 26;446(7139):1056-61.
7. Dzierzak E, Speck NA. Of lineage and legacy: the development of mammalian
hematopoietic stem cells. Nature immunology. 2008 Feb;9(2):129-36.
8. Rhodes KE, Gekas C, Wang Y, Lux CT, Francis CS, Chan DN, et al. The emergence of
hematopoietic stem cells is initiated in the placental vasculature in the absence of circulation.
Cell stem cell. 2008 Mar 6;2(3):252-63.
9. Lacaud G, Robertson S, Palis J, Kennedy M, Keller G. Regulation of hemangioblast
development. Annals of the New York Academy of Sciences. 2001 Jun;938:96-107;
discussion 8.
10. Nishikawa SI. A complex linkage in the developmental pathway of endothelial and
hematopoietic cells. Current opinion in cell biology. 2001 Dec;13(6):673-8.
11. Jaffredo T, Nottingham W, Liddiard K, Bollerot K, Pouget C, de Bruijn M. From
hemangioblast to hematopoietic stem cell: an endothelial connection? Experimental
hematology. 2005 Sep;33(9):1029-40.
12. Park C, Ma YD, Choi K. Evidence for the hemangioblast. Experimental hematology. 2005
Sep;33(9):965-70.
13. Nishikawa SI, Nishikawa S, Hirashima M, Matsuyoshi N, Kodama H. Progressive lineage
analysis by cell sorting and culture identifies FLK1+VE-cadherin+ cells at a diverging point of

DOI: 10.3324/haematol.2009.014241
22
endothelial and hemopoietic lineages. Development (Cambridge, England). 1998
May;125(9):1747-57.
14. Choi K, Kennedy M, Kazarov A, Papadimitriou JC, Keller G. A common precursor for
hematopoietic and endothelial cells. Development (Cambridge, England). 1998
Feb;125(4):725-32.
15. Huber TL, Kouskoff V, Fehling HJ, Palis J, Keller G. Haemangioblast commitment is
initiated in the primitive streak of the mouse embryo. Nature. 2004 Dec 2;432(7017):625-30.
16. Keller G. Embryonic stem cell differentiation: emergence of a new era in biology and
medicine. Genes & development. 2005 May 15;19(10):1129-55.
17. Chen MJ, Yokomizo T, Zeigler BM, Dzierzak E, Speck NA. Runx1 is required for the
endothelial to haematopoietic cell transition but not thereafter. Nature. 2009 Feb
12;457(7231):887-91.
18. de Bruijn MF, Ma X, Robin C, Ottersbach K, Sanchez MJ, Dzierzak E. Hematopoietic
stem cells localize to the endothelial cell layer in the midgestation mouse aorta. Immunity.
2002 May;16(5):673-83.
19. Kim I, Yilmaz OH, Morrison SJ. CD144 (VE-cadherin) is transiently expressed by fetal
liver hematopoietic stem cells. Blood. 2005 Aug 1;106(3):903-5.
20. Taoudi S, Morrison AM, Inoue H, Gribi R, Ure J, Medvinsky A. Progressive divergence of
definitive haematopoietic stem cells from the endothelial compartment does not depend on
contact with the foetal liver. Development (Cambridge, England). 2005 Sep;132(18):4179-91.
21. Yao H, Liu B, Wang X, Lan Y, Hou N, Yang X, et al. Identification of high proliferative
potential precursors with hemangioblastic activity in the mouse aorta-gonad- mesonephros
region. Stem cells (Dayton, Ohio). 2007 Jun;25(6):1423-30.
22. Lan Y, Liu B, Yao H, Li F, Weng T, Yang G, et al. Essential role of endothelial Smad4 in
vascular remodeling and integrity. Molecular and cellular biology. 2007 Nov;27(21):7683-92.
23. Nakano T, Kodama H, Honjo T. Generation of lymphohematopoietic cells from embryonic
stem cells in culture. Science (New York, NY. 1994 Aug 19;265(5175):1098-101.
24. Schmitt TM, Zuniga-Pflucker JC. Induction of T cell development from hematopoietic
progenitor cells by delta-like-1 in vitro. Immunity. 2002 Dec;17(6):749-56.
25. Schmitt TM, de Pooter RF, Gronski MA, Cho SK, Ohashi PS, Zuniga-Pflucker JC.

DOI: 10.3324/haematol.2009.014241
23
Induction of T cell development and establishment of T cell competence from embryonic stem
cells differentiated in vitro. Nature immunology. 2004 Apr;5(4):410-7.
26. Pimanda JE, Chan WY, Wilson NK, Smith AM, Kinston S, Knezevic K, et al. Endoglin
expression in blood and endothelium is differentially regulated by modular assembly of the
Ets/Gata hemangioblast code. Blood. 2008 Dec 1;112(12):4512-22.
27. Brachtendorf G, Kuhn A, Samulowitz U, Knorr R, Gustafsson E, Potocnik AJ, et al. Early
expression of endomucin on endothelium of the mouse embryo and on putative hematopoietic
clusters in the dorsal aorta. Dev Dyn. 2001 Nov;222(3):410-9.
28. Armulik A, Abramsson A, Betsholtz C. Endothelial/pericyte interactions. Circulation
research. 2005 Sep 16;97(6):512-23.
29. Blalock WL, Weinstein-Oppenheimer C, Chang F, Hoyle PE, Wang XY, Algate PA, et al.
Signal transduction, cell cycle regulatory, and anti-apoptotic pathways regulated by IL-3 in
hematopoietic cells: possible sites for intervention with anti-neoplastic drugs. Leukemia. 1999
Aug;13(8):1109-66.
30. Mukouyama Y, Hara T, Xu M, Tamura K, Donovan PJ, Kim H, et al. In vitro expansion of
murine multipotential hematopoietic progenitors from the embryonic aorta-gonad-
mesonephros region. Immunity. 1998 Jan;8(1):105-14.
31. Garcia-Porrero JA, Godin IE, Dieterlen-Lievre F. Potential intraembryonic hemogenic
sites at pre-liver stages in the mouse. Anatomy and embryology. 1995 Nov;192(5):425-35.
32. North T, Gu TL, Stacy T, Wang Q, Howard L, Binder M, et al. Cbfa2 is required for the
formation of intra-aortic hematopoietic clusters. Development (Cambridge, England). 1999
Jun;126(11):2563-75.
33. North TE, de Bruijn MF, Stacy T, Talebian L, Lind E, Robin C, et al. Runx1 expression
marks long-term repopulating hematopoietic stem cells in the midgestation mouse embryo.
Immunity. 2002 May;16(5):661-72.
34. Lancrin C, Sroczynska P, Stephenson C, Allen T, Kouskoff V, Lacaud G. The
haemangioblast generates haematopoietic cells through a haemogenic endothelium stage.
Nature. 2009 Feb 12;457(7231):892-5.
35. Minasi MG, Riminucci M, De Angelis L, Borello U, Berarducci B, Innocenzi A, et al. The
meso-angioblast: a multipotent, self-renewing cell that originates from the dorsal aorta and

DOI: 10.3324/haematol.2009.014241
24
differentiates into most mesodermal tissues. Development (Cambridge, England). 2002
Jun;129(11):2773-83.
36. Cossu G, Bianco P. Mesoangioblasts--vascular progenitors for extravascular
mesodermal tissues. Current opinion in genetics & development. 2003 Oct;13(5):537-42.
37. Murry CE, Keller G. Differentiation of embryonic stem cells to clinically relevant
populations: lessons from embryonic development. Cell. 2008 Feb 22;132(4):661-80.
38. Faloon P, Arentson E, Kazarov A, Deng CX, Porcher C, Orkin S, et al. Basic fibroblast
growth factor positively regulates hematopoietic development. Development (Cambridge,
England). 2000 May;127(9):1931-41.
39. Robertson SM, Kennedy M, Shannon JM, Keller G. A transitional stage in the
commitment of mesoderm to hematopoiesis requiring the transcription factor SCL/tal-1.
Development (Cambridge, England). 2000 Jun;127(11):2447-59.
40. Endoh M, Ogawa M, Orkin S, Nishikawa S. SCL/tal-1-dependent process determines a
competence to select the definitive hematopoietic lineage prior to endothelial differentiation.
The EMBO journal. 2002 Dec 16;21(24):6700-8.
41. Filippi MD, Porteu F, Le Pesteur F, Schiavon V, Millot GA, Vainchenker W, et al.
Requirement for mitogen-activated protein kinase activation in the response of embryonic
stem cell-derived hematopoietic cells to thrombopoietin in vitro. Blood. 2002 Feb
15;99(4):1174-82.
42. Lacaud G, Gore L, Kennedy M, Kouskoff V, Kingsley P, Hogan C, et al. Runx1 is
essential for hematopoietic commitment at the hemangioblast stage of development in vitro.
Blood. 2002 Jul 15;100(2):458-66.
43. Chan RJ, Johnson SA, Li Y, Yoder MC, Feng GS. A definitive role of Shp-2 tyrosine
phosphatase in mediating embryonic stem cell differentiation and hematopoiesis. Blood. 2003
Sep 15;102(6):2074-80.
44. Ema M, Faloon P, Zhang WJ, Hirashima M, Reid T, Stanford WL, et al. Combinatorial
effects of Flk1 and Tal1 on vascular and hematopoietic development in the mouse. Genes &
development. 2003 Feb 1;17(3):380-93.
45. Liu B, Sun Y, Jiang F, Zhang S, Wu Y, Lan Y, et al. Disruption of Smad5 gene leads to
enhanced proliferation of high-proliferative potential precursors during embryonic

DOI: 10.3324/haematol.2009.014241
25
hematopoiesis. Blood. 2003 Jan 1;101(1):124-33.
46. Park C, Afrikanova I, Chung YS, Zhang WJ, Arentson E, Fong Gh G, et al. A hierarchical
order of factors in the generation of FLK1- and SCL-expressing hematopoietic and endothelial
progenitors from embryonic stem cells. Development (Cambridge, England). 2004
Jun;131(11):2749-62.
47. Ramirez-Bergeron DL, Runge A, Dahl KD, Fehling HJ, Keller G, Simon MC. Hypoxia
affects mesoderm and enhances hemangioblast specification during early development.
Development (Cambridge, England). 2004 Sep;131(18):4623-34.
48. D'Souza SL, Elefanty AG, Keller G. SCL/Tal-1 is essential for hematopoietic commitment
of the hemangioblast but not for its development. Blood. 2005 May 15;105(10):3862-70.
49. Hirai H, Samokhvalov IM, Fujimoto T, Nishikawa S, Imanishi J, Nishikawa S. Involvement
of Runx1 in the down-regulation of fetal liver kinase-1 expression during transition of
endothelial cells to hematopoietic cells. Blood. 2005 Sep 15;106(6):1948-55.
50. Kubo A, Chen V, Kennedy M, Zahradka E, Daley GQ, Keller G. The homeobox gene
HEX regulates proliferation and differentiation of hemangioblasts and endothelial cells during
ES cell differentiation. Blood. 2005 Jun 15;105(12):4590-7.
51. Willey S, Ayuso-Sacido A, Zhang H, Fraser ST, Sahr KE, Adlam MJ, et al. Acceleration
of mesoderm development and expansion of hematopoietic progenitors in differentiating ES
cells by the mouse Mix-like homeodomain transcription factor. Blood. 2006 Apr
15;107(8):3122-30.
52. Lugus JJ, Chung YS, Mills JC, Kim SI, Grass J, Kyba M, et al. GATA2 functions at
multiple steps in hemangioblast development and differentiation. Development (Cambridge,
England). 2007 Jan;134(2):393-405.
53. Perlingeiro RC. Endoglin is required for hemangioblast and early hematopoietic
development. Development (Cambridge, England). 2007 Aug;134(16):3041-8.
54. Wang C, Faloon PW, Tan Z, Lv Y, Zhang P, Ge Y, et al. Mouse lysocardiolipin
acyltransferase controls the development of hematopoietic and endothelial lineages during in
vitro embryonic stem-cell differentiation. Blood. 2007 Nov 15;110(10):3601-9.
55. Zafonte BT, Liu S, Lynch-Kattman M, Torregroza I, Benvenuto L, Kennedy M, et al.
Smad1 expands the hemangioblast population within a limited developmental window. Blood.

DOI: 10.3324/haematol.2009.014241
26
2007 Jan 15;109(2):516-23.
56. Pearson S, Sroczynska P, Lacaud G, Kouskoff V. The stepwise specification of
embryonic stem cells to hematopoietic fate is driven by sequential exposure to Bmp4, activin
A, bFGF and VEGF. Development (Cambridge, England). 2008 Apr;135(8):1525-35.
57. Robin C, Ottersbach K, Durand C, Peeters M, Vanes L, Tybulewicz V, et al. An
unexpected role for IL-3 in the embryonic development of hematopoietic stem cells.
Developmental cell. 2006 Aug;11(2):171-80.
58. Taoudi S, Gonneau C, Moore K, Sheridan JM, Blackburn CC, Taylor E, et al. Extensive
hematopoietic stem cell generation in the AGM region via maturation of VE-cadherin+CD45+
pre-definitive HSCs. Cell stem cell. 2008 Jul 3;3(1):99-108.
59. Pardanaud L, Dieterlen-Lievre F. Manipulation of the angiopoietic/hemangiopoietic
commitment in the avian embryo. Development (Cambridge, England). 1999 Feb;126(4):617-
27.
60. Lacaud G, Kouskoff V, Trumble A, Schwantz S, Keller G. Haploinsufficiency of Runx1
results in the acceleration of mesodermal development and hemangioblast specification upon
in vitro differentiation of ES cells. Blood. 2004 Feb 1;103(3):886-9.
61. Cai Z, de Bruijn M, Ma X, Dortland B, Luteijn T, Downing RJ, et al. Haploinsufficiency of
AML1 affects the temporal and spatial generation of hematopoietic stem cells in the mouse
embryo. Immunity. 2000 Oct;13(4):423-31.
62. Zeoli A, Dentelli P, Rosso A, Togliatto G, Trombetta A, Damiano L, et al. Interleukin-3
promotes expansion of hemopoietic-derived CD45+ angiogenic cells and their arterial
commitment via STAT5 activation. Blood. 2008 Jul 15;112(2):350-61.

DOI: 10.3324/haematol.2009.014241
27
Figure legends
Figure 1. Molecular characteristics, hematopoietic and vascular potential of BL-
CFCs derived from E10.5 mouse AGM region. (A) Morphology of a typical blast colony.
(B) Expression of Brachyury, Flk-1 and Runx1 in 7 representative blast colonies by
nested RT-PCR. (C) Immunofluorescence staining of Sca-1, F4/80 and Ter119 in blast
colonies. (D) CFU-E, CFU-GM, CFU-mix and BFU-E derived from blast colonies. (E)
Quantification of hematopoietic colonies generated from 12 representative blast colonies.
Morphology of individually plucked blast colonies in liquid expansion system after 48h (F)
and 10d (G) of incubation. (H-I) DiI-Ac-LDL incorporation (red) combined with CD45 or
F4/80 staining (green) of the adherent cells. Arrowheads, CD45 positive in H or F4/80
positive in I. (J) _-SMA+ staining (green) of the adherent cells. (K) Tube-like structures in
Matrigel of the adherent cells. (L-M) DiI-Ac-LDL incorporation (red) combined with _-SMA
or Calponin staining (green) of the tube-like structures. Inserts show the corresponding
bright fields. Original magnification: _40 (K), _100 (D: upper panels; F, G), _200 (A, C, H-
J, L, M), and _1000 (D: lower panels).
Figure 2. Identification of hematopoietic and endothelial potential of BL-CFCs by
OP9-based system. (A) Individual blast colonies are co-cultured with OP9 for 10 days in
48-well plate. In four representative wells, both CD45+ cells (upper panel) and CD31
+
tube-like structures are detected. (B) Morphology of blast colonies co-cultured with OP9
in B lymphocyte induction system for 6 days, showing significant proliferation of round
non-adherent cells. (C ) Flow cytometry analysis of B lymphocyte markers of blast
colonies differentiated on OP9 stroma for 10 days. (D) Morphology of blast colonies co-
cultured with OP9-DL1 for 8 days, showing significant proliferation of round non-adherent
cells. (E) Flow cytometry analysis of T lymphocyte markers and NK1.1 of blast colonies
differentiated on OP9-DL1 stroma for 15 days. Representative data from three
independent experiments are shown. Original magnification: _40 (A: lower panel), _100
(A: upper panel; B, D).

DOI: 10.3324/haematol.2009.014241
28
Figure 3. Clonality of the AGM-derived blast colonies. (A) The number of colonies is
directly proportional to that of AGM cells input, indicating that a single cell corresponds to
the growth of a colony. (B) Cell mixing analysis. An equal number of cells from the AGM
region of male wild type (Sry+GFP
-) and female GFP transgenic (Sry
-GFP
+) mice are
mixed, and after 4 days of culture, colonies are picked out and expanded individually to
determine their genomic DNA content. Among 20 representative colonies analyzed, 10
display exclusively Sry+GFP
- genotype and the rest 10 Sry
-GFP
+. (C) Representative
blast colonies from wild type (top) and GFP transgenic AGM region (bottom).
Fluorescence (left), bright field (right).
Figure 4. Development kinetics, surface markers and tissue distribution of BL-
CFCs. (A) BL-CFCs and hematopoietic CFCs derived from caudal half (E9.5) or AGM
region (E10.5-12.5) are analyzed in parallel, showing different development kinetics. (B-
D) Immunohistochemical analysis of E10 embryos. Transverse sections of dorsal aortas
(DA) are stained with anti-CD31 (B), anti-Endomucin (C) and anti-PDGFR_ (D)
antibodies. The dorsal aspect is upward. Endothelial layer is CD31 and Endomucin
positive but PDGFR_ negative (red arrowheads in D). CD31 and Endomucin are also
expressed in hematopoietic cell clusters (arrowheads in B and C) associated with
endothelium. (E-H) The frequency of BL-CFCs in CD105 (E), CD31 (F), Endomucin (G),
and PDGFR_ (H) positive cells sorted from E10.5 AGM region. The results are
representative data expressed as means ± s.e.m from 3-4 independent experiments,
respectively. ( I ) Identification of BL-CFCs in various blood-forming tissues of mid-
gestation embryos, including circulation, yolk sac, placenta and fetal liver. Single-cell
suspensions from these tissues yield only canonical hematopoietic colonies, which fail to
expand as adherent layers and are incapable of forming tube-like structures in Matrigel.
Original magnifications: x100 (I, middle and bottom panel); x200 (I, top panel).

DOI: 10.3324/haematol.2009.014241
29
Figure 5. Influence of BL-CFC development in E10.5 AGM region by IL-3-mediated
signaling. (A) Among a panel of cytokines, IL-3 and bFGF significantly amplify the BL-
CFC in E10.5 AGM region. IL-3-mediated effect is dose dependent (B) and temporally
regulated (C). Neutralizing tissue-derived IL-3 markedly reduces the number of BL-CFC
in the AGM region. Neu denotes IL-3 neutralizing antibody (D). Phosphorylation of STAT5
and ERK1/2 in AGM cells are enhanced by IL-3 (E ), and the corresponding
pharmacological inhibitors abrogate IL-3-mediated BL-CFC expansion (F). The results
represent the means ± s.e.m. Significance is determined using the Student’s t- test: *,
P<0.05; **, P<0.01, compared with the control data.
Figure 6. Increased hematopoietic and vascular potential of IL-3-treated BL-CFC
from E10.5 AGM region. (A) IL-3- and bFGF-stimulated BL-CFCs give rise to increased
number of hematopoietic CFC. (B-D) Matrigel-based test reveals IL-3- and bFGF-
stimulated BL-CFCs have enhanced tube-forming capacity both in number and in length.
(E) CD31 staining of blast colonies co-cultured with OP9 for 10 days. Increased tube
formation is detected when IL-3 is added during blast colony formation. (F) Real-time
PCR analysis of BL-CFC cultures to examine IL-3-mediated molecular changes. Original
magnification: _40 (B, E). The results represent the means ± s.e.m. Significance is
determined using the Student’s t- test: **, P<0.01, compared with the control data.
Figure 7. IL-3 promotes primitive hematopoiesis in E7.5 embryos. (A) Transcripts of
2 types of IL-3 receptors (IL-3R_/ßC, IL-3R_/ßIL-3) are detected by RT-PCR in E7.5
embryos and E10.5 AGM region. (B) Morphology of hematopoietic colonies derived from
single-cell suspension of E7.5 embryos with or without IL-3 treatment for 12h. (C)
General (upper) and cellular (lower) morphology of CFU-EryP, CFU-M and CFU-EM
derived from E7.5 embryos. CFU-EryP indicates colonies of primitive erythrocytes; CFU-
Mac, colonies of macrophages; CFU-EM, mixed primitive erythroid/macrophage colonies.
(D) Number of hematopoietic colonies per E7.5 embryos with or without IL-3 treatment
for 12h. (E ) Real-time PCR analysis of CFC cultures derived from E7.5 embryos to

DOI: 10.3324/haematol.2009.014241
30
examine IL-3-mediated molecular changes. Original magnification: _100 (B; C, upper
panel), _1000 (C, lower panel). The results represent the means ± s.e.m. Significance
is determined using the Student’s t- test: *, P<0.05; **, P<0.01, compared with the control
data.

DOI: 10.3324/haematol.2009.014241
31

DOI: 10.3324/haematol.2009.014241
32

DOI: 10.3324/haematol.2009.014241
33
DOI: 10.3324/haematol.2009.014241
34

DOI: 10.3324/haematol.2009.014241
35
DOI: 10.3324/haematol.2009.014241
36

DOI: 10.3324/haematol.2009.014241
37

DOI: 10.3324/haematol.2009.014241
Supplemental DataTable S1. Oligonucleotide primers used for nested RT-PCR
Genes Primer sequence Tm (_)Size
(bp)
first set1 5'-TGCTGCCTGTGAGTCATAAC-3'first set2 5'-CAGATTGTCTTTGGCTACTT-3'
55 883brachyury
nest set1 5'-CTCTAGCCAGTATCCCAGTC-3'
nest set2 5'-TATCCATACAGTTGACTTCC-3'
60 812
first set1 5'-ACTTCCTCTGCTCCGTGCTA-3'first set2 5'-GTCCACTGTGATTTTGATGGC-3'
57 307Runx1
nest set1 5'-ACTCACTGGCGCTGCAACAA-3'
nest set2 5'-AAGCTCTTGCCTCTACCGCT -3'
57 208
first set1 5'-CTTGATTTCACCTGGCACTCT-3'first set2 5'-TACTTCACAGGGATTCGGACT-3'
56 275Flk-1
nest set1 5'-CACCTGGCACTCTCCACCTTC- 3'
nest set2 5'-GATTTCATCCCACTACCGAAAG-3'
61 239
first set1 5'-GGGGGCTATAAGTTCTTTGC-3'first set2 5'-TCCAACACTTCGAGAGGTCC-3'
56 313HPRT
nest set1 5'-GTTCTTTGCTGACCTGCTGG-3'
nest set2 5'-TGGGGCTGTACTGCTTAACC-3'
57 247

DOI: 10.3324/haematol.2009.014241
Table S2. Oligonucleotide primers used for RT-PCR
Genes Primer sequence Tm (_)Size(bp)
IL-3R_C Sense 5’-TGAGCTATAACCACATCCAGA-3’
Anti-sense 5’-GGAGTTGGTCCCTCTACTTGA-3’
56 727
IL-3R_ Sense 5’-CCTCCAATCCTCAACCAGACC-3’Anti-sense 5’-CATTGGGAAGTTGCTCTGTGG-3’
58 676
IL-3R_ Sense 5’-CAACTTCACCGTCTTCCTG-3’Anti-sense 5’-CTCTGACCTCGACTTGACC-3’
59 649
HPRT Sense 5'-GTTCTTTGCTGACCTGCTGG-3'
Anti-sense 5'-TGGGGCTGTACTGCTTAACC-3'
57 247

DOI: 10.3324/haematol.2009.014241
Table S3. Oligonucleotide primers used for real-time PCR
Genes Primer sequence Tm (_)Size(bp)
HPRTSense 5'-GCTGGTGAAAAGGAC CTCT-3' Anti-
sense 5'-CACAGGACTAGAACACCTGC-3'60 249
Flk-1Sense 5'-CAGAACACCAAAAGAGAGGAACG-3'Anti-sense 5'-TGACAGGAGTGGAGATAGAGGAA-3'
60 193
SclSense 5'-GGCAAGAAACAGGAGTGAGAA G-3' Anti-sense 5'-GCAAGGAAGACACAGAGGAAGA-3'
60 162
Runx1Sense 5'-ACTCACTGGCGCTGCAACAA-3'
Anti-sense 5'-AAGCTCTTGCCTCTACCGCT -3'60 208
Gata2Sense 5'-GCG AAAACCAAACTGCATAAGC-3' Anti-sense 5'-CTGTCTCCCAGAAACCAAGAGC-3'
60 235
Gata1Sense 5'-AGCCTATTCTTC CCCCAAGTT T-3' Anti-sense 5'-GGTTCCTCGTCTGGATTCCATC-3'
60 371
vWFSense 5'-GTGTACCACGAGGTCATCAACG-3'Anti-sense 5'-GAGGGCACAAAGGTCAGGAG-3'
60 164

DOI: 10.3324/haematol.2009.014241
Table S4. Lymphoid potential of blast colony from E10.5 mouse AGM region.
CD19+_%_ CD3
+/TCR_
+_%_
1 + 8.31
2 14.09 5.74
3 5.04 5.87
4 + 3.92
5 + 4.92
6 13.1 1.2
+ indicates that the CD19+ cells are determined by fluorescent microscope other than
flow cytometry.

DOI: 10.3324/haematol.2009.014241
Figure S1
Figure S1. Roles of cytokines in growth of blast colonies. One cytokine is eliminated
from original formula in a sequential fashion to determine the roles of cytokines in colony
formation. SCF and bFGF have the most dramatic effect on colony growth (p<0.01). The
removal of IL-6 has a modest effect on the number of colonies (p<0.05). The results
represent the means ± s.e.m. Significance is determined using the Student’s t- test: *,
P<0.05; **, P<0.01, compared with the control data.


















