
Interhemispheric Communication via Direct Connections for Alternative Meanings of Ambiguous Words
20
Brain and Language 80, 77–96 (2002) doi:10.1006/brln.2001.2582, available online at http://www.idealibrary.com on Interhemispheric Communication via Direct Connections for Alternative Meanings of Ambiguous Words Marjorie Collins Murdoch University, Perth, Western Australia, Australia A priming experiment was used to investigate Burgess and Simpson’s (1988) claim that interhemispheric cooperation plays an essential role in the interpretation of ambiguous text. In doing so, the merits of two models of interhemispheric cooperation, the homotopic inhibi- tion theory (Cook, 1986) and the direct connections model (Collins & Coney, 1998), were examined. Priming of alternative meanings of ambiguous words was measured using homo- graphs and their dominant (e.g., BARK–DOG) and subordinate meanings (e.g., BARK– TREE) as related pairs in a lexical decision task, with normal university students as subjects. Stimulus pairs were temporally separated by stimulus onset asynchronies (SOAs) of 180 and 350 ms and were independently projected to the left or right visual fields (LVF or RVF). At the shorter SOA, priming was restricted to LVF–RVF presentations, with homograph primes directed to the LVF equally facilitating responses to RVF targets which were associated with their dominant and subordinate meanings. This suggests that within 180 ms, a homograph projected to the right hemisphere activates a range of alternative meanings in the left hemi- sphere. At an SOA of 350 ms, LVF–RVF priming was obtained along with RVF–LVF and RVF–RVF priming. Evidently at this stage of processing, an ambiguous word directed to either hemisphere activates a range of alternative meanings in the contralateral hemisphere, while RVF primes also activate subordinate, but not dominant meanings in the left hemisphere. A homograph directed to the LVF did not activate dominant or subordinate meanings within the right hemisphere at either SOA. Generally, ambiguous words directed to either hemisphere activated a more extensive array of meanings in the contralateral hemisphere than in the hemi- sphere to which the prime was directed. This confirms the importance of interhemispheric cooperation in generating alternate meanings of ambiguous words. Strong support was found for the direct connections model (Collins & Coney, 1998), but no support for the homotopic inhibition theory (Cook, 1986). 2002 Elsevier Science (USA) Key Words: cerebral hemispheres; lexical ambiguity; priming; dominant and subordinate meanings; interhemispheric communication; direct connections; homotopic inhibition. INTRODUCTION The left cerebral hemisphere has traditionally been attributed with superior lan- guage abilities, although recent investigations suggest that the right hemisphere also makes a significant contribution to the comprehension of language (e.g., Collins, 1999; Collins & Frew, 2001; (Collins) Abernethy & Coney, 1993; Brownell, 1988; Zaidel, White, Sakurai, & Banks, 1988). Evidence derived from clinical studies sug- This research was supported by an Australian Research Council grant (File number: 03-20-198-193). I thank Ms. Adele Summers for collecting the data, and Dr. Robert Kane for statistical advice. Address correspondence and reprint requests to Marjorie Collins, School of Psychology, Murdoch University, Murdoch, Perth, Western Australia, 6150. E-mail: [email protected]. 77 0093-934X/02 $35.00 2002 Elsevier Science (USA) All rights reserved.
-
Upload
marjorie-collins -
Category
Documents
-
view
212 -
download
0
Transcript of Interhemispheric Communication via Direct Connections for Alternative Meanings of Ambiguous Words

Brain and Language 80, 77–96 (2002)doi:10.1006/brln.2001.2582, available online at http://www.idealibrary.com on
Interhemispheric Communication via DirectConnections for Alternative Meanings of
Ambiguous Words
Marjorie Collins
Murdoch University, Perth, Western Australia, Australia
A priming experiment was used to investigate Burgess and Simpson’s (1988) claim thatinterhemispheric cooperation plays an essential role in the interpretation of ambiguous text.In doing so, the merits of two models of interhemispheric cooperation, the homotopic inhibi-tion theory (Cook, 1986) and the direct connections model (Collins & Coney, 1998), wereexamined. Priming of alternative meanings of ambiguous words was measured using homo-graphs and their dominant (e.g., BARK–DOG) and subordinate meanings (e.g., BARK–TREE) as related pairs in a lexical decision task, with normal university students as subjects.Stimulus pairs were temporally separated by stimulus onset asynchronies (SOAs) of 180 and350 ms and were independently projected to the left or right visual fields (LVF or RVF). Atthe shorter SOA, priming was restricted to LVF–RVF presentations, with homograph primesdirected to the LVF equally facilitating responses to RVF targets which were associated withtheir dominant and subordinate meanings. This suggests that within 180 ms, a homographprojected to the right hemisphere activates a range of alternative meanings in the left hemi-sphere. At an SOA of 350 ms, LVF–RVF priming was obtained along with RVF–LVF andRVF–RVF priming. Evidently at this stage of processing, an ambiguous word directed toeither hemisphere activates a range of alternative meanings in the contralateral hemisphere,while RVF primes also activate subordinate, but not dominant meanings in the left hemisphere.A homograph directed to the LVF did not activate dominant or subordinate meanings withinthe right hemisphere at either SOA. Generally, ambiguous words directed to either hemisphereactivated a more extensive array of meanings in the contralateral hemisphere than in the hemi-sphere to which the prime was directed. This confirms the importance of interhemisphericcooperation in generating alternate meanings of ambiguous words. Strong support was foundfor the direct connections model (Collins & Coney, 1998), but no support for the homotopicinhibition theory (Cook, 1986). 2002 Elsevier Science (USA)
Key Words: cerebral hemispheres; lexical ambiguity; priming; dominant and subordinatemeanings; interhemispheric communication; direct connections; homotopic inhibition.
INTRODUCTION
The left cerebral hemisphere has traditionally been attributed with superior lan-guage abilities, although recent investigations suggest that the right hemisphere alsomakes a significant contribution to the comprehension of language (e.g., Collins,1999; Collins & Frew, 2001; (Collins) Abernethy & Coney, 1993; Brownell, 1988;Zaidel, White, Sakurai, & Banks, 1988). Evidence derived from clinical studies sug-
This research was supported by an Australian Research Council grant (File number: 03-20-198-193).I thank Ms. Adele Summers for collecting the data, and Dr. Robert Kane for statistical advice.
Address correspondence and reprint requests to Marjorie Collins, School of Psychology, MurdochUniversity, Murdoch, Perth, Western Australia, 6150. E-mail: [email protected].
770093-934X/02 $35.00
2002 Elsevier Science (USA)All rights reserved.

78 MARJORIE COLLINS
gests that one area of language comprehension in which the right hemisphere playsa vital role is the interpretation of ambiguous text (e.g., Brownell, Potter, & Mi-chelow, 1984; Weylman, Brownell, Roman, & Gardner, 1989; Kaplan, Brownell,Jacobs, & Gardner, 1990). There is also growing clinical evidence that cooperationbetween the cerebral hemispheres is essential to derive the full import of an ambigu-ous linguistic message (e.g., Brownell, 1988; Brownell, Simson, Bihrle, Potter &Gardner, 1990; Brownell et al., 1984).
Burgess and Simpson (1988) have confirmed clinical observations of the righthemisphere’s contribution to the interpretation of ambiguous text in a priming studywith normal subjects. They used homographs as ambiguous primes because they havemore than one meaning and so are usually interpreted within context. These ambigu-ous primes (e.g., BANK) were presented in central vision and followed by targetsassociated with the prime’s dominant (e.g., MONEY) or subordinate meaning (e.g.,RIVER). The targets were projected to the left (LVF) or right visual fields (RVF)either 35 or 750 ms after the onset of the prime. For the dominant meanings, Burgessand Simpson found equivalent priming for LVF and RVF target presentations atboth of these stimulus onset asynchronies (SOAs). This suggests that after centralpresentation of a homograph, dominant meanings were activated in both hemisphereswithin 35 ms and remained active for at least 750 ms. In contrast, for subordinatemeanings there was a different pattern of activation in each hemisphere. When targetswere projected to the RVF within 35 ms, priming was obtained when these targetswere associated with the subordinate meaning of the prime. However, no RVF prim-ing was evident for these same targets when they were presented after an intervalof 750 ms. Instead, responses to these targets were considerably slower (by 46 ms)when preceded by a prime pertaining to the subordinate meaning than when precededby an unrelated prime. Hence, it seems that although dominant meanings of ambigu-ous words were still active in the left hemisphere at the longer SOA, subordinatemeanings were inhibited. The pattern of results was quite different when targets wereprojected to the right hemisphere via the LVF. When a target related to the prime’ssubordinate meaning was projected to the LVF, no priming was evident at the shorterSOA. However, at the longer SOA (750 ms), a 34 ms priming effect was present.This suggests that subordinate meanings took longer to become activated in the righthemisphere relative to the left hemisphere, but remained active for longer.
These differing patterns of priming for alternative meanings presented to the LVFand RVF provide an insight to the relative contributions each hemisphere makes tothe interpretation of ambiguous words. It appears that the left hemisphere may quicklyselect information pertaining to a single interpretation of ambiguous text, while theright hemisphere maintains multiple interpretations. This is consistent with Taylor’sview (1988) that the right hemisphere proposes and the left hemisphere disposesinterpretations. Burgess and Simpson (1988) take this idea further in suggesting thatwhen a subordinate meaning is required to disambiguate text, but the relevant infor-mation has been suppressed in the left hemisphere, the latter may call upon the righthemisphere to access memory information and provide an appropriate interpretation.If this is the case, interhemispheric communication must play an integral role in theinterpretation of ambiguous text. The present study aims to investigate whether thisis the case and to explore the mechanism underlying any such cross-hemisphere com-munication.
To date, limited attention has been directed to the issue of interhemispheric cooper-ation in linguistic processing. Cook’s (1986) homotopic inhibition theory is one ex-ception, and is particularly relevant here, as it specifies the mechanism underlyinginterhemispheric cooperation and makes clear predictions about the complementaryprocesses of activation and inhibition in each hemisphere when deriving meaning

INTERHEMISPHERIC COMMUNICATION 79
from linguistic stimuli. Cook proposes, somewhat controversially, that similar infor-mation is represented in homologous areas of each hemisphere, with these areas con-nected through the corpus callosum. When a cortical area in one hemisphere is acti-vated, neighboring areas within that hemisphere are inhibited. In the contralateralhemisphere, however, the homologous area is inhibited via the corpus callosum, andthis acts to disinhibit ‘‘surround’’ areas. Consequently, a word directed to one hemi-sphere will activate closely related meanings therein, while peripheral meanings areinhibited. Simultaneously, peripheral meanings will be activated in the contralateralhemisphere while direct meanings are suppressed.
Cook’s model (1986) provides a neat account of recent clinical observations ofthe complementary specialization of each hemisphere for linguistic processing, wherethe left hemisphere has been shown to predominate in extracting the literal, denotativeaspects of language, while the right hemisphere appears more sensitive to peripheraland connotative meanings (e.g., Brownell et al., 1990; 1984; Tompkins, 1990; Bee-man, 1993). Rodel, Cook, Regard, and Landis (1992) have also found support forthis model in a study using normal subjects who were asked to judge whether wordpairs were semantically related. They presented one word of each pair centrally, whilethe other was simultaneously presented to the RVF or LVF. When one word wasdirected to the LVF, they found that subjects were more likely to judge distant associ-ates as semantically related. In contrast, words were less likely to be judged as seman-tically related when one word was directed to the RVF. Rodel et al. viewed theseresults as consistent with Cook’s proposal that left hemisphere activation of a wordinhibits identical processing in the right hemisphere and thereby activates more dis-tant associates in that hemisphere.
However, Chiarello, Maxfield and Kahan (1995) question the viability of the ho-motopic inhibition theory as a general account of interhemispheric interaction on thebasis of their failure to find any support for the model’s predictions in a study whichused a priming task where homograph primes were paired with targets which wereassociated with their dominant and subordinate meanings. These primes were pre-sented centrally, while targets appeared laterally 80, 130, and 200 ms after the onsetof the prime. At each of these SOAs, they found equivalent priming in each hemi-sphere for dominant and subordinate meanings. This is contrary to the predictionsof Cook’s model (1986) that a centrally presented ambiguous prime will activatedominant meanings, and thereby suppress subordinate meanings, in the left hemi-sphere. It is also contrary to his prediction that a converse pattern of activation andsuppression will simultaneously occur in the right hemisphere, with priming of subor-dinate meanings and suppression of dominant meanings. In the current context, it isinteresting that the pattern of priming observed in their study was also inconsistentwith Burgess and Simpson’s (1988) proposal that subordinate meanings become ac-tive in the right hemisphere only after they have been suppressed in the left hemi-sphere. This led Chiarello et al. to conclude that left hemisphere meaning selectionis not the mechanism by which subordinate meanings are activated in the right hemi-sphere. More generally, the findings from this study indicate that interhemisphericcooperation during linguistic processing is not achieved by the mechanisms proposedby either Cook (1986) or Burgess and Simpson (1988).
The ‘‘direct connections model,’’ recently developed by Collins and Coney (1998),may more successfully account for interhemispheric communication during linguisticprocessing. A fundamental premise of this model, for which there is ample evidence,is that both hemispheres can make lexical decisions without the necessity for callosalrelay of information, and words are primarily encoded in the hemisphere to whichthey are initially projected (e.g., (Collins) Abernethy & Coney, 1990; Chiarello, 1988;Zaidel et al., 1988). From this premise, the direct connections model proposes that

80 MARJORIE COLLINS
direct callosal connections exist between concepts in one hemisphere and particularrelated concepts in the opposite hemisphere, where interhemispheric communicationat the sematic level can occur via activation spreading through these direct connec-tions, rather than by transference of a coded form of the prime from one hemisphereto the other. Moreover, concepts directed to a particular hemisphere may activaterelated concepts in the contralateral hemisphere without activating those conceptswithin the directly stimulated hemisphere. Consequently in the priming procedure,a prime projected to one hemisphere may directly activate related meanings in thecontralateral hemisphere, and this occurs independently of any activation of thesemeanings within the hemisphere initially receiving the prime. In this model, inter-hemispheric cooperation is integral to the extraction of meaning during linguisticprocessing.
Support for the direct connections model can be garnered from studies which haveindependently projected prime–target pairs to the same and different hemispheresand have found cross-hemisphere facilitation without concomitant within-hemispherefacilitation. Several studies have observed this unusual pattern of priming. (Collins)Abernethy and Coney (1993) found cross-hemisphere facilitation when a prime pro-jected to the LVF was followed by an associated RVF target, even though there wasno concomitant within-hemisphere facilitation for LVF–LVF presentations of thesepairs. We also found LVF–RVF facilitation without LVF–LVF facilitation for cate-gory exemplars confounded by association ((Collins) Abernethy & Coney, 1990).The findings from these two studies are consistent with a tenet of the direct connec-tions model that the processing of a word within the right hemisphere can activateits associates in the left hemisphere independently of any activation of those associ-ates within the right hemisphere itself. Even so, the most striking evidence in favorof the direct connections model comes from a study we designed to test the predic-tions of the direct connections model relative to two other spreading activation mod-els (Collins & Coney, 1998). An unusually large priming facilitation, of 58 ms, wasfound for RVF–LVF presentations of low imagery primes and their concrete associ-ates, but there was no accompanying LVF–LVF or RVF–RVF priming, with RTsfor the related and baseline conditions differing by only 2 ms. The pattern of primingobserved in that study ruled out the possibility that cross-hemisphere facilitation wasaccomplished by transference of a coded form of the prime from one hemisphere tothe other. Rather, consistent with the direct connections model, the evidence wasconsistent with the conclusion that low imagery words projected to the left hemi-sphere directly activated their concrete associates in the right hemisphere withoutactivating these associates within the left hemisphere itself. Hence, the findings fromthese three studies clearly demonstrate that cross-hemisphere priming can occur inde-pendently of within-hemisphere priming, with a word directed to one hemispheredirectly activating related concepts in the contralateral hemisphere without activatingthese concepts in the hemisphere initially receiving, and semantically encoding, theprime. This raises the possibility that cross-hemisphere cooperation in the generationof alternative meanings of ambiguous text occurs in accordance with the principlesof the direct connections model, where semantic encoding of ambiguous text directedto one hemisphere initiates the activation of a range of alternative meanings in thecontralateral hemisphere via direct callosal connections.
To elucidate the mechanism underlying cross-hemisphere cooperation in the ex-traction of alternative meanings during linguistic processing, the current study exam-ined the veracity of the differing predictions of the direct connections and homotopicinhibition theories. An effective procedure to test these predictions is the primingtask, where prime and target pairs are independently projected to either the RVF orLVF. Both within- and cross-hemisphere priming conditions were included in the

INTERHEMISPHERIC COMMUNICATION 81
design for this purpose. To assess the activation and suppression of alternative mean-ings of ambiguous words in each hemisphere, the relationship between prime–targetpairs was also manipulated. Some pairs were related via the dominant meaning ofhomograph primes (e.g., BARK–DOG) and others were related via the prime’s subor-dinate meaning (e.g., BARK–TREE). As the profile of priming for alternative mean-ings differs in each hemisphere over time (Burgess & Simpson, 1988) the temporalinterval between these pairs was manipulated. To this end, and to restrict the focusof this study to automatic processes, prime–target pairs were temporally separatedby SOAs of 180 and 350 ms.
The direct connections and homotopic inhibition theories both assume that thegeneration of meaning during linguistic processing entails true hemispheric coopera-tion, without duplication of processes in each hemisphere. However, the two modelspostulate a different mechanism by which interhemispheric communication occurs,so they make differing predictions about the pattern of priming expected under theexperimental conditions of the current study. According to the homotopic inhibitiontheory (Cook, 1986), a prime projected to one hemisphere will activate dominantmeanings therein, thereby suppressing subordinate meanings in that hemisphere. Si-multaneously, subordinate meanings will be activated in the contralateral hemispherewhile direct meanings are suppressed. Hence, the pattern of priming in each hemi-sphere will be complementary, and determined by the hemisphere initially receivingthe prime, rather than hemispheric specialization per se. In contrast, the direct connec-tions model (Collins & Coney, 1998) assumes that a prime projected to one hemi-sphere can directly activate alternative meanings in the contralateral hemisphere with-out activating these meanings in the hemisphere initially receiving the prime. Hence,priming for cross-hemisphere presentations is possible without concomitant within-hemisphere priming. As the right hemisphere appears to supply alternative meaningsof ambiguous text to the left hemisphere (Burgess & Simpson, 1988), the direct con-nections model holds that the processing of an ambiguous prime in the right hemi-sphere activates alternative meanings in the left hemisphere via direct callosal con-nections. In this context, support for the direct connections model would be foundif priming for LVF–RVF presentations occurs without any concomitant LVF–LVFpriming.
METHOD
Subjects
Thirty-two undergraduate psychology students acted as subjects. Data for 3 of these subjects werediscarded as their error rates exceeded 30% in either the word or nonword condition in the first or secondexperimental session. Fifteen female and 13 male subjects remained, their mean age being 22.6 years(SD 5 6.08). All were predominantly right handed according to Bryden’s (1982) hand preference ques-tionnaire (mean handedness quotient: 10.67; SD 5 0.25) and had normal or corrected-to-normal visionand English was their first language.
Apparatus
Subjects were tested in a well-lit room containing an IBM personal computer system which controlledrandomization of trial sequencing, stimulus presentation, timing, and data collection. The onset and offsetof all stimuli was controlled by a program code that queried the hardware and synchronized writes tovideo memory such that each complete write occurred only during that part of a raster cycle when theraster scan was not utilizing the relevant memory locations. Stimuli were printed on a high-resolution,nonglare monitor in white, enlarged uppercase letters against a dark background. Screen intensity wasset at a low level to minimize phosphor persistence. An adjustable chin rest was used to stabilize eachsubject’s head in the correct central position and distance from the screen, such that all stimuli were

82 MARJORIE COLLINS
presented with their innermost boundary, whether to the left or right of center, exactly 2° of visual anglefrom the central fixation. Reaction time (RT) was measured to a resolution of 1 ms via a centrallypositioned microswitch response box connected to the user port of the microcomputer. Eye movementswere monitored by a Sanyo video camera connected to a video monitor, which provided a magnified viewof the subject’s eyes. EMH12 High Performance earmuffs were used to minimize any noise interference.
Design
Reaction time (RT) was the principal dependent variable, with errors also recorded and analyzed. Alexical decision task was used, requiring subjects to discriminate words from nonwords using a GO–NOGO response procedure. Four experimental variables were manipulated: type of stimulus pair, stimu-lus list, SOA, and visual field of presentation. Repeated measures were taken on all variables. The firstexperimental variable, type of stimulus pair, incorporated a total of 1200 prime–target pairs, which wereequally divided into two stimulus lists carefully matched on all relevant variables (see details below).Each list comprised related, neutral, and unrelated word pairs as well as word–nonword and neutral–nonword pairs. These lists were presented, in counterbalanced order, in two experimental sessions. Thesecond experimental variable was stimulus list, where the related pairs in one stimulus list were associatedthrough the homograph prime’s dominant meaning, while related pairs in the other list were associatedvia the prime’s subordinate meaning.
The third experimental variable, visual field of presentation, comprised four levels: (i) RVF primeand target; (ii) LVF prime, RVF target; (iii) LVF prime and target; and (iv) RVF prime, LVF target.Stimuli were presented at 2° of visual angle from a central fixation and subtended a horizontal visualangle of between 2° and 6°. SOA was the fourth experimental variable, where the temporal intervalbetween the onset of prime and target presentation was manipulated. In one experimental session theSOA was 180 ms, while in the other session the SOA was 350 ms. SOA conditions were presented incounterbalanced order. Stimulus pairs were presented in one visual field condition per experimentalsession, but were presented again in the second session. Selection of visual field for each pair wasrandomly determined both within and across experimental sessions, as was order of presentation of wordpairs in each condition. Each subject was exposed to a unique distribution of pairs in each visual fieldcondition, and a different sequence of prime–target conditions within and across testing sessions.
Stimulus Materials
Two stimulus lists were presented, each comprised of 600 stimulus pairs. Of these, 100 were related-word pairs, with 100 homographs as primes. Each prime was paired with a target associated with itsdominant (e.g., BARK–TREE) and subordinate meaning (e.g., BARK–DOG) and placed into differentstimulus lists. Hence, identical primes were used in each list, but the targets differed. All related pairswere drawn from homograph norms (Twilley, Dixon, Taylor, & Clark, 1994; Nelson, McEvoy, Wall-ing, & Wheeler, 1980). The 100 unrelated word pairs were formed by repairing primes and targets fromthe related set, with the restriction that the resulting pairs featured no orthographic, phonemic, categoricalor associative relationship (e.g., BARK–CUP). The 100 neutral pairs were formed by pairing the primeBLANK with each of the targets in the related set (e.g., BLANK–CUP).
For the word–nonword pairs, 100 additional homographs were selected from the same homographnorms (Twilley et al., 1994; Nelson et al., 1980) and used as primes. These word primes were selectedto match the primes in the related-word condition in relation to grammatical category, length, and wordfrequency [t (198) 5 .268; p 5 .789]. Each of these primes was paired with an orthographically legal,pronounceable nonword target. None of the resultant pairs were orthographically or phonologically simi-lar. Nonwords were generated by unsystematically changing between one and three letters of the targetsin the positive set. To equate the number of repetitions of these stimuli with those in the positive set,each of these additional word primes was re-paired with a different nonword target from the same list,giving 200 word–nonword pairs per list (e.g., AIR–CORMS and AIR–PELOT). Last, to form 100 neutralcontrol pairs which mirrored the neutral condition in the positive set, each nonword target was alsopaired with the word BLANK (e.g., BLANK–CORMS).
All pairs were phonemically and orthographically dissimilar, and none began with the same first letter.All words were nouns or adjectives, and all stimuli were between three and eight letters in length, with95.5% between three and six letters. Mean frequency of primes in the positive set was 62.01 wpm (SD5 115.6), while primes in the negative set had a mean frequency of 57.42 wpm (SD 5 126.33: Kucera &Francis, 1967). There was no significant difference in the frequencies for these two sets [t (198) 5 .268;p 5 .789]. Mean word frequency of targets in the dominant meanings set was 90.32 wpm (SD 5 174.83),while targets in the subordinate set were 95.21 wpm (SD 5 109.53: Kucera & Francis, 1967). Again,frequencies in these two sets were matched [t (198) 5 .237; p 5 .813].

INTERHEMISPHERIC COMMUNICATION 83
Procedure
Prior to the first testing session, subjects were given instructions outlining their task as one of distin-guishing target words from nonwords. No allusion was made to the presence of related pairs in thestimulus list, and subjects were advised that they could choose to ignore the prime. The necessity ofmaintaining fixation on the central cross during presentation of all stimuli was emphasized. Subjectswere positioned in front of the display monitor with their heads positioned by a chin-rest directly infront of, and 60 cm distant from, the center of the screen. They were given 72 practice trials with thesame structure as the experimental trials. No stimuli from the experimental set were used. Feedbackrelating to accuracy was provided after each practice trial. If the video monitor revealed any deviationof eyes from fixation during these trials, subjects were reminded of the importance of maintaining fixationat all times.
Each trial began with a central fixation cross which remained on throughout the trial. After 600 ms,a prime word was displayed in the LVF or RVF for 120 ms. Either 50 ms or 230 ms after the primedisappeared, the target was presented in the LVF or RVF for 150 ms. The shorter interval was employedin the SOA 180 ms condition, and the longer interval in the SOA 350 ms condition. To minimize maskingeffects, the target appeared one line beneath the display location of the prime. The central fixation endureduntil the target was erased from the screen. The entire screen then remained blank for 1500 ms, duringwhich the subject signaled a response. Randomization of trials ensured that subjects were unable topredict the visual field in which either the prime or target would appear.
Subjects responded in accordance with a GO–NOGO procedure. When the target was a word, theywere required to respond by simultaneously depressing two centrally positioned microswitches with bothindex fingers, the faster of the two responses being taken as RT for that trial. When the target was nota word, they were required to withhold their response. Subjects were permitted 1500 ms after erasureof the target to respond. Failure to respond within 1500 ms was treated as a NOGO response. Followingtrials in which an incorrect response was made, the word ERROR appeared directly above the centralfixation for 32 ms. Subjects were permitted to rest after each block of 72 trials and were given feedbackon their overall speed and accuracy for the preceding block(s). They were also encouraged to maintainan error rate of less than eight per block. Subjects completed two experimental sessions which differedin the SOA between prime and target. Both stimulus lists were presented, in counterbalanced order, ineach session. Hence, each stimulus pair was presented once per experimental session and twice acrossthe two experimental sessions, but each time in a different SOA condition. Each session was separatedby a minimum of 2 days.
RESULTS
Reaction Time Analyses
Statistical analyses were performed on mean correct reaction time (RT) for thepositive set. A four-way ANOVA (2 3 2 3 3 3 4) was computed on SOA, stimuluslist, type of stimulus pair, and visual field. The first step in interpreting these datawas to ascertain whether responses differed as a function of stimulus list. After all,targets differed in each list, even though the primes were identical. So, although carewas taken to ensure that the targets in each stimulus list were carefully matched, itis important to ascertain whether responses differed as a function of stimulus list,rather the relationship between prime and target. There was no such confound in thedata. There was no main effect for stimulus list [F(1, 27) 5 1.232; p 5 .277], withresponses to targets in the dominant meanings list a mere 4 ms faster than responsesto targets in the subordinate list. Stimulus list did not interact with SOA [F(1,27) 5.41; p 5 .528], which indicates that the pattern of RTs to targets in the two stimuluslists was similar at the two SOAs. Nor did stimulus list interact with visual field[F(3, 81) 5 1.747; p 5 .164] or type of stimulus pair [F(2, 54) 5 .051; p 5 .95],where RTs followed the same rank order for each visual field in the dominant andsubordinate lists, and there was no difference in the pattern of responses to related,neutral, and unrelated targets in the two lists. Taken together, these results confirmthat the two stimulus lists were well matched, as responses to targets did not differas a function of stimulus list. Hence, priming effects for dominant and subordinatemeanings can be interpreted without qualification.

84 MARJORIE COLLINS
In relation to the remaining variables, all three main effects were significant. Re-sponses were significantly faster (by 24 ms) when prime–target pairs were separatedby an SOA of 350 ms relative to 180 ms [F(1, 27) 5 15.705; p , .001]. Apparentlya performance advantage was gained when additional processing time was availableto generate lexical decisions. The main effect for stimulus pair [F(2,54) 5 30.45; p, .001] reflects an overall facilitation when targets were preceded by related primes(508 ms), relative to responses when these targets were preceded by neutral or unre-lated primes (521 ms and 519 ms respectively). The main effect for visual field [F(3,81) 5 34.514; p , .001] reflects the faster responses when prime–target pairs wereprojected to the same visual field, relative to the conditions where they were projectedto different visual fields (refer to Figs. 1–4). Responses were fastest when both primeand target were projected to the RVF (at 482 ms) and slowest when a RVF primewas followed by a LVF target (at 555 ms). Although my research commonly findsfaster (and more accurate) responses for LVF–RVF than LVF–LVF presentationsof prime–target pairs (e.g., (Collins) Abernethy & Coney, 1996; Collins & Coney,1998), the current experiment found the converse, with faster RTs for LVF–LVFthan LVF–RVF presentations (at 497 and 532 ms respectively). Even so, the visualfield advantage obtained in the current experiment was consistent across both SOAsand stimulus list (and errors), which indicates that this effect was reliable.
There were two two-way interactions. Although RTs were faster in all visual fieldconditions at the longer SOA, the interaction between SOA and visual field [F(3, 81)5 9.005; p , .001] reflects the greater benefit for crossed–visual field presentationsseparated by an SOA of 350 ms relative to 180 ms (35.5 ms) relative to the benefitfor same-visual field presentations (13 ms). It appears that an additional 170 ms be-tween the onset of prime and target assisted the generation of responses in all condi-tions. However, this benefit was greater when prime and target were projected todifferent visual fields. Perhaps additional time assisted the integration of stimulusinformation across the hemispheres. Visual field also interacted with type of stimuluspair [F(6, 162) 5 3.458; p 5 .003]. Here, the magnitude of priming was greater forcrossed- than within-visual field presentations, with facilitation averaging 20 and 7.5ms respectively. Reaction times to targets in the neutral and unrelated conditionswere similar in all visual field conditions (with an average 3 ms difference) exceptthe LVF–RVF condition, where responses to unrelated targets were 12 ms faster thanto neutral targets.
SOA and visual field also entered into a three-way interaction with stimulus list[F(3, 81) 5 4.182; p 5 .008]. There were no other three-way interactions, nor wasthere a four-way interaction [F(6, 162) 5 1.315; p 5 .253]. To identify the sourceof the interaction between SOA, visual field and stimulus list, separate two-way AN-OVAs (2 3 2) were calculated for each visual field condition, with SOA and stimuluslist as the variables. There were no significant effects for the LVF–LVF condition.For the remaining three visual field conditions, there was a main effect for SOA[LVF–RVF: F(1, 27) 5 18.95; p , .001; RVF–RVF: F(1, 27) 5 6.5; p 5 .017;RVF–LVF: F(1, 27) 5 19.574; p , .001]. No other effects were significant. Thisindicates that responses were significantly faster when prime and target were sepa-rated by the longer SOA in all visual field conditions except the LVF–LVF condition.Moreover, this RT advantage was consistent for the dominant and subordinate sets.
The homotopic inhibition and direct connections theories make different predic-tions about the priming of dominant and subordinate meanings in each visual fieldcondition. To examine this, related samples t tests were used to carry out plannedcomparisons between each related condition and its neutral baseline. Type I errorwas controlled by adjusting the critical p to .0469 using a modified Bonferroni test(Keppel, 1991). When a homograph prime was followed after 180 ms by a target

INTERHEMISPHERIC COMMUNICATION 85
pertaining to its dominant meaning, significant priming facilitation (of 13 ms) waspresent for LVF–RVF presentations [t(27) 5 2.462; p 5 .02]. However, there wasno priming for the remaining three visual field conditions [RVF–RVF: t(27) 5 .769;p 5 .449; LVF–LVF: t(27) 5 1.78; p 5 .086; RVF–LVF: t (27) 5 2.036; p 5 .052].(Refer to Fig. 1.) When the SOA between these stimulus pairs was increased to 350ms, significant priming facilitation was present when prime and target were projectedto different visual fields [LVF–RVF: t (27) 5 4.352; p , .001, 29 ms; and RVF–LVF: t (27) 5 3.677; p 5 .001, 17 ms]. In contrast, there was no priming when thesepairs were projected to the same visual field [RVF–RVF: t(27) 5 .896; p 5 .378;LVF–LVF: t(27) 5 .155; p 5 .878]. (See Fig. 2.) When a homograph prime wasfollowed 180 ms later by a target pertaining to its subordinate meaning, the patternof priming was identical to that for dominant meanings (see Fig. 3). That is, signifi-cant priming facilitation (of 23 ms) was present for LVF–RVF presentations [t(27)5 4.16; p , .001], but the response advantage for the related condition did not reachsignificance for the remaining three visual field conditions [RVF–RVF: t(27) 51.999; p 5 .056; LVF–LVF: t (27) 5 1.227; p 5 .23; RVF–LVF: t (27) 5 1.569; p5 .128]. However, when the SOA between the prime and its subordinate meaningwas increased to 350 ms, significant priming facilitation was present for LVF–RVF[t (27) 5 3.11; p 5 .004, 20 ms], RVF–LVF [t (27) 5 3.651; p 5 .001, 25 ms] andRVF–RVF presentations [t(27) 5 2.476; p 5 .02, 16 ms]. Nonetheless, there wasno facilitation for LVF–LVF presentations at this SOA [t (27) 5 .148; p 5 .883]with responses to related pairs 1 ms slower than the neutral condition (see Fig. 4).
Hence, these results illustrate that priming of alternative meanings of ambiguouswords differs in each hemisphere as a function of time as well as dominance of theassociation between prime and target. At both SOAs, projection of a homograph tothe right hemisphere facilitated responses to RVF targets which were associated with
FIG. 1. Mean correct RT for dominantly related, neutral, and unrelated word pairs presented in theleft and right visual fields at an SOA of 180 ms.

86 MARJORIE COLLINS
FIG. 2. Mean correct RT for dominantly related, neutral, and unrelated word pairs presented in theleft and right visual fields at an SOA of 350 ms.
FIG. 3. Mean correct RT for subordinately related, neutral, and unrelated word pairs presented inthe left and right visual fields at an SOA of 180 ms.
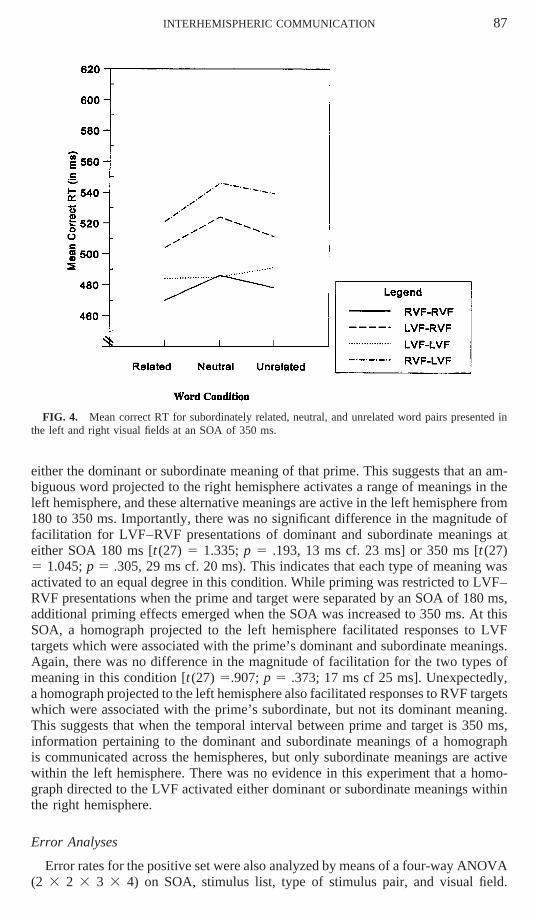
INTERHEMISPHERIC COMMUNICATION 87
FIG. 4. Mean correct RT for subordinately related, neutral, and unrelated word pairs presented inthe left and right visual fields at an SOA of 350 ms.
either the dominant or subordinate meaning of that prime. This suggests that an am-biguous word projected to the right hemisphere activates a range of meanings in theleft hemisphere, and these alternative meanings are active in the left hemisphere from180 to 350 ms. Importantly, there was no significant difference in the magnitude offacilitation for LVF–RVF presentations of dominant and subordinate meanings ateither SOA 180 ms [t (27) 5 1.335; p 5 .193, 13 ms cf. 23 ms] or 350 ms [t (27)5 1.045; p 5 .305, 29 ms cf. 20 ms). This indicates that each type of meaning wasactivated to an equal degree in this condition. While priming was restricted to LVF–RVF presentations when the prime and target were separated by an SOA of 180 ms,additional priming effects emerged when the SOA was increased to 350 ms. At thisSOA, a homograph projected to the left hemisphere facilitated responses to LVFtargets which were associated with the prime’s dominant and subordinate meanings.Again, there was no difference in the magnitude of facilitation for the two types ofmeaning in this condition [t(27) 5.907; p 5 .373; 17 ms cf 25 ms]. Unexpectedly,a homograph projected to the left hemisphere also facilitated responses to RVF targetswhich were associated with the prime’s subordinate, but not its dominant meaning.This suggests that when the temporal interval between prime and target is 350 ms,information pertaining to the dominant and subordinate meanings of a homographis communicated across the hemispheres, but only subordinate meanings are activewithin the left hemisphere. There was no evidence in this experiment that a homo-graph directed to the LVF activated either dominant or subordinate meanings withinthe right hemisphere.
Error Analyses
Error rates for the positive set were also analyzed by means of a four-way ANOVA(2 3 2 3 3 3 4) on SOA, stimulus list, type of stimulus pair, and visual field.

88 MARJORIE COLLINS
Generally, the outcomes were consistent with the RT analyses. Like the RT data, theerror data confirmed that the two stimulus lists were well matched. There was nomain effect for stimulus list [F(1, 27) 5 1.328; p 5 .259], with error rates for thedominant and subordinate lists differing by only 0.61%. Furthermore, stimulus listdid not interact with SOA [F(1, 27) 5 .191; p 5 .665], visual field [F(3, 81) 5 2.099;p 5 .107], or stimulus type [F(2, 54) 5 .523; p 5 .596]. This indicates that bothspeed and accuracy of responses were similar for both stimulus lists.
Like the RT data, there were main effects for the remaining three variables. Themain effect for visual field [F(3, 81) 5 9.243; p , .001] reflected greater accuracywhen prime and target were both projected to the same relative to different visualfields (refer to Table 1). The identical rankings for each visual field condition in boththe accuracy and RT data indicates that there were no speed–accuracy trade-offs.The main effect for type of stimulus pair [F(2, 54) 5 9.37; p , .001] reflected thegreater accuracy in responding to the targets when they were preceded by a relatedprime (6.8% errors) relative to responses when these targets were preceded by unre-lated or neutral primes (7.64 and 8.7% errors respectively). Last, the main effect forSOA [F(1, 27) 5 46.418; p , .001] indicates that responses were faster and moreaccurate, by 4.9%, when prime and target were separated by an SOA of 350 ms,relative to the shorter SOA of 180 ms.
Three interactions were significant. Like the RT data, the two-way interaction be-tween SOA and visual field [F(3, 81) 5 11.437; p , .001] reflected the greater benefit
TABLE 1Percentage of Accurate Responses for Word Pairs Presented in Each Visual Field for the
Dominant and Subordinate Stimulus Lists at SOAs 180 ms and 350 ms
Visual field of presentation
RVF–RVF LVF–RVF RVF–LVF LVF–LVF
Subordinate listSOA 180 ms
Word pair relationshipRelated 97.1 86.6 87.7 90Neutral 93.9 85.4 80.2 93.2Unrelated 95.9 87.3 82.3 93.8Mean 95.6 86.4 83.4 92.3
SOA 350 msWord pair relationship
Related 97.1 96.3 93.2 93.2Neutral 96.9 94.6 89.6 95.4Unrelated 97.5 93.6 93.6 93Mean 97.2 94.8 92.1 93.9
Dominant ListSOA 180 ms
Word pair relationshipRelated 96.1 91.1 87.5 93.4Neutral 94.8 82.7 83.4 94.1Unrelated 95 87.5 85.2 92.1Mean 95.3 87.1 85.4 93.2
SOA 350 msWord pair relationship
Related 96.9 95.7 94.5 94.8Neutral 95.7 92.7 94.1 94.1Unrelated 95.7 94.6 95.2 95.4Mean 96.1 94.3 94.6 94.8

INTERHEMISPHERIC COMMUNICATION 89
when prime and target appeared in different visual fields, relative to the same visualfield, at the longer SOA. While there were less errors for RVF–RVF and LVF–LVFpresentations at an SOA of 350 ms relative to 180 ms (1.19 and 1.55% respectively),this difference was considerably larger for LVF–RVF and RVF–LVF presentations(7.82 and 8.98% respectively). The interaction between type of stimulus pair andvisual field [F(6, 162) 5 4.328; p , .001] reflected the greater response advantagewhen related word pairs were presented in different visual fields relative to the same-visual field conditions. Unlike the RT data, these two variables also entered into athree-way interaction with stimulus list [F(6, 162) 5 2.572; p 5 .021]. This interac-tion was analyzed with two separate ANOVAs (3 3 4) for the dominant and subordi-nate lists, with type of stimulus pair and visual field as variables. For the subordinatelist, all effects were significant. The main effect for stimulus pair [F(2, 54) 5 3.213;p 5 .048] reflected greater accuracy in response to targets preceded by related primes(92.66%) relative to neutral and unrelated primes (91.16 and 92.12% respectively).The main effect for visual field [F(11.69; p , .001] reflected greater response accu-racy when prime–target pairs were projected to the same hemisphere (96.43% forRVF–RVF and 93.09% for LVF–LVF presentations) relative to pairs appearing indifferent hemispheres (90.62% for LVF–RVF and 87.77% for RVF–LVF). Therewas also an interaction between these variables [F(6, 162) 5 5.026; p , .001]. Re-sponses were more accurate for related pairs in all visual field conditions exceptLVF–LVF presentations, where responses to related targets were less accurate thanthe neutral condition by 1.8% (see Table 2). The ANOVA for the dominant list alsoreturned all significant effects. The main effects for stimulus type [F(2, 54) 5 7.165;p 5 .002] and visual field [F(3, 81) 5 5.306; p 5 .002] reflected the same trendsas the subordinate set: responses were more accurate when the target was preceded bya related prime (93.75%) relative to neutral and unrelated primes (91.45 and 92.59%respectively). Accuracy was also higher when prime and target were projected to thesame visual field. The interaction between these two variables [F(6, 162) 5 2.273;p 5 .039] reflects the response advantage for the related condition in all visual fieldconditions except the LVF–LVF condition, where accuracy was equal for relatedand neutral pairs. The largest priming effects were obtained for cross-visual fieldpresentations (see Table 2). These differences are consistent with the RT data.
TABLE 2Percentage of Accurate Responses for Word Targets in Each Visual Field for the
Dominant and Subordinate Stimulus Lists
Visual field of presentation
RVF–RVF LVF–RVF RVF–LVF LVF–LVF
Subordinate listWord pair relationship
Related 97.1 91.4 90.4 91.6Neutral 96.7 90.4 87.9 93.4Unrelated 95.4 90 84.9 94.3Priming 10.4% 11.0% 12.5% 21.8%
Dominant listWord pair relationship
Related 96.5 93.4 90.9 94.1Neutral 95.4 91.1 90.2 93.8Unrelated 95.3 87.7 88.7 94.1Priming 11.1% 12.3% 10.7% 0.0

90 MARJORIE COLLINS
DISCUSSION
Before interpreting these results, it is important to establish the hemispheric locusof initial processing of these stimuli. There is considerable evidence that in primingtasks using lexical decision, stimuli directly access the lexicon in the hemisphere towhich they are initially projected, and are processed therein (e.g., Collins & Coney,1998; (Collins) Abernethy & Coney, 1996; and see Chiarello, 1988, for a review ofthe evidence). Hardyck, Chiarello, Dronkers, and Simpson (1985) specifically investi-gated this question and concluded that lexical decisions occur in the hemisphere ofinput and do not require shared hemispheric resources. Similarly, Rodel et al. (1992)as well as (Collins) Abernethy and Coney (1990) concluded that their results areinconsistent with the idea that hemispheric differences in performance on the linguis-tic tasks they employed are due simply to degradation through callosal relay. Con-firmation for such findings can be found in clinical studies, where it has been demon-strated that split brain patients are able to make lexical decisions for stimuli presentedto either hemisphere (e.g., Zaidel, 1983). In the same vein, Zaidel et al. (1988) con-cluded that evidence from aphasics, commissurotomy, and normal subjects all sug-gest that the right hemisphere is able to make lexical decisions and that both hemi-spheres have a lexicon. Therefore, consistent with previous research in this area (e.g.,Koivisto, 1999; Rodel et al., 1992; Chiarello, Burgess, Richards, & Pollock, 1990),it will be assumed that primes and targets were initially processed in the hemisphereto which they were projected.
The results of the current experiment provide clear support for the direct connec-tions model (Collins & Coney, 1998) and no support for the homotopic inhibitiontheory (Cook, 1986). According to the latter theory, the activation of dominant mean-ings within a given hemisphere initiates a complementary pattern of activation andsuppression in homologous areas of the contralateral hemisphere. Hence, the patternof priming is determined by the hemisphere to which the prime is directed ratherthan hemispheric specialization per se. Consequently, under the experimental condi-tions of the current experiment, priming effects arising from the projection of a primeto the left hemisphere should be identical to those initiated by right hemisphereprimes. There was no support for this assumption in the current experiment. Instead,the pattern of facilitation initiated by left hemisphere primes differed from the facilita-tion observed after projection of these primes to the right hemisphere. This is particu-larly evident when prime and target were separated by an SOA of 350 ms. Whileambiguous primes directed to the RVF facilitated responses to subordinate meaningsprojected to the RVF 350 ms later, there was no priming when these primes and theirsubordinate meanings were both projected to the LVF.
Furthermore, a comparison of the priming effects for within- and cross-hemispherepresentations provides no support for the homotopic inhibition theory’s assumptionthat projection of a prime to a given hemisphere will activate dominant meaningstherein and simultaneously activate subordinate meanings in the contralateral hemi-sphere. Rather, in the current experiment, priming of dominant meanings was exclu-sively associated with cross-hemisphere presentations. That is, projection of an am-biguous prime to the RVF failed to facilitate responses to dominant meaningsprojected to the left hemisphere after 180 or 350 ms. Nevertheless, RVF primes didfacilitate responses to subordinate meanings directed to the left hemisphere after 350ms. The activation of subordinate, but not dominant meanings in the left hemisphereafter the appearance of an ambiguous prime in the RVF, is in direct contrast to theassumptions of the homotopic inhibition theory (Cook, 1986). Further contrary evi-dence is found in the priming for RVF–LVF presentations. The theory predicts thatambiguous primes directed to the left hemisphere will activate subordinate, but not

INTERHEMISPHERIC COMMUNICATION 91
dominant meanings in the right hemisphere. In the current experiment, a homographprojected to the RVF did facilitate responses to subordinate meanings which appearedin the LVF 350 ms later. However, equivalent RVF–LVF priming was also foundfor dominant meanings at this SOA. Hence, the pattern of priming obtained whenhomograph primes were directed to the left hemisphere is altogether inconsistent withthe predictions of the homotopic inhibition theory.
The homotopic inhibition theory does not fare any better when right hemispherepriming is considered. The theory assumes that the activation of dominant meaningswithin a given hemisphere initiates a complementary pattern of activation and sup-pression in homologous areas of the contralateral hemisphere. As there was no prim-ing in the current experiment when an ambiguous prime and its dominant meaningwere both projected to the LVF, simultaneous activation of subordinate meanings inthe left hemisphere is precluded. Despite this, LVF primes facilitated responses toRVF targets which were associated with the subordinate meanings of these primes,and this occurred irrespective of the temporal interval separating the prime–targetpairs. Furthermore, LVF primes also facilitated responses to RVF targets pertainingto the prime’s dominant meaning and to the same degree as subordinate meanings.The homotopic inhibition theory cannot account for equal activation of direct andperipheral meanings occurring simultaneously in a given hemisphere. Hence, like thefindings of Chiarello et al. (1995), the pattern of priming found in the current studyis inconsistent with the predictions of Cook’s homotopic inhibition theory.
In contrast, the results of the current study provide strong support for the directconnections model. This support is provided by the cross-hemisphere facilitationwhich occurred independently of within-hemisphere facilitation in both hemispheres.First, LVF–RVF priming occurred without LVF–LVF priming at SOAs 180 and 350ms and for both types of meaning. That is, homographs directed to the right hemi-sphere facilitated responses to both dominant and subordinate meanings projected tothe left hemisphere 180 and 350 ms later, but did not facilitate responses to eithertype of meaning when they were projected directly to the right hemisphere. Hence,an ambiguous word in the LVF did not activate either its dominant or subordinatemeanings within the right hemisphere. Despite this, these LVF primes activated bothtypes of meaning in the left hemisphere. Importantly, when stimulus pairs were sepa-rated by an SOA of 180 ms, LVF–RVF facilitation occurred without concomitantfacilitation for any other visual field condition. The absence of facilitation for alterna-tive meanings projected directly to the left hemisphere at this SOA mitigates againstthe conclusion that the source of this priming effect was the left hemisphere andoccurred subsequent to callosal transfer of LVF primes to this hemisphere. If thathad been the case, priming would also be expected for RVF–RVF presentations. Thisdid not occur for either type of meaning. Instead, this profile of priming is entirelyconsistent with the direct connections model, which assumes that within- and cross-hemisphere facilitation are independent, as a prime projected to one hemisphere candirectly activate related concepts in the opposite hemisphere via direct callosal con-nections and can do so without activating those words within the hemisphere initiallyreceiving the prime. In this instance, it appears that an ambiguous word projected tothe right hemisphere directly activated a range of its alternative meanings in the lefthemisphere, without activating these meanings within the right hemisphere itself.LVF–RVF facilitation has also been found without LVF–LVF facilitation for cate-gory exemplars ((Collins) Abernethy & Coney, 1990) and associates ((Collins) Aber-nethy & Coney, 1993). Taken together, these findings indicate that a word directedto the right hemisphere can activate a range of meanings in the left hemisphere inde-pendently of the activation of these aspects of meaning in the right hemisphere itself.
Further evidence consistent with the direct connections model lies in the indepen-

92 MARJORIE COLLINS
dence of cross- and within- hemisphere facilitation when the prime was directed tothe left hemisphere. A homograph projected to the left hemisphere facilitated re-sponses to dominant meanings projected to the right hemisphere 350 ms later, butdid not facilitate responses to these meanings when they were projected directly tothe left hemisphere. The same trend was evident when these pairs were separated byan SOA of 180 ms, with the 22 ms facilitation for RVF–LVF presentations almostreaching significance (p 5 .052), unlike the accompanying RVF–RVF condition,where responses to targets preceded by related and neutral primes differed by only5 ms. In view of the absence of facilitation for LVF–LVF presentations of thesepairs, and the general left hemisphere superiority in performing this task, it is unlikelythat this facilitation arose through callosal transfer of the prime from left to righthemisphere (see Collins & Coney, 1998, for further discussion of this issue). Rather,this profile of priming is consistent with the direct connections model and suggeststhat an ambiguous word presented to the left hemisphere activates its dominant mean-ings in the right hemisphere via direct callosal connections, with this cross-hemi-sphere activation occurring independently of any activation within the left hemisphereitself.
Cross-hemisphere facilitation was also evident for RVF–LVF presentations of sub-ordinate meanings, although this was accompanied by RVF–RVF facilitation. How-ever, facilitation for these two conditions occurred when prime–target pairs wereseparated by an SOA of 350 ms, but not by 180 ms. The difference in priming effectsat these two intervals is consistent with previous research which indicates that theactivation of various meanings differs within and between each hemisphere over time(Collins, 1999; Collins & Coney, 1998; (Collins) Abernethy & Coney, 1993, 1996;Koivisto, 1997; Nakagawa, 1991) and encapsulates the dynamic nature of informa-tion processing in the cerebral hemispheres. No firm conclusions can be made aboutthe source of this cross-hemisphere priming for subordinate meanings separated byan SOA of 350 ms, as it was accompanied by facilitation for RVF–RVF presenta-tions. Even so, it is unlikely that the mechanism by which the left hemisphere acti-vates subordinate meanings in the right hemisphere differs from that identified abovefor dominant meanings. Hence, it seems reasonable to conclude that ambiguousprimes directed to the left hemisphere activate a range of alternative meanings in theright hemisphere via direct callosal connections. This is consistent with our previousstudy which found RVF–LVF facilitation without RVF–RVF facilitation for lowimagery primes and their concrete associates (Collins & Coney, 1998). Hence, it alsoappears that a word directed to the left hemisphere can activate a range of associatedmeanings in the right hemisphere independently of any activation within the lefthemisphere itself.
Generally, the present results indicate that interhemispheric cooperation plays acentral role in the interpretation of ambiguous text. The predominance of facilitationeffects in the cross-hemisphere conditions combined with the paucity of facilitationwhen stimulus pairs were projected to the same hemisphere warrant such a conclu-sion. The only facilitation for within-hemisphere presentations occurred when homo-graph primes and their subordinate meanings were both directed to the left hemi-sphere within 350 ms, while cross-hemisphere priming was found for all but twoof the conditions examined here. Moreover, while the LVF–RVF priming confirmsBurgess and Simpson’s (1988) proposal that the left hemisphere calls upon the righthemisphere to access memory information pertaining to subordinate meanings, thecurrent results indicate that a broader conclusion is warranted. The cross-hemispherepriming effects in the current study indicate that when processing an ambiguous word,both hemispheres have the capacity to make use of memory representations of alterna-tive meanings in the contralateral hemisphere. This conclusion is based on the obser-

INTERHEMISPHERIC COMMUNICATION 93
vation that an ambiguous word directed to the right hemisphere activated a range ofalternative meanings in the left hemisphere within 180 ms, and this activation wasmaintained at 350 ms. Likewise, a homograph prime directed to the left hemisphereactivated both dominant and subordinate meanings in the right hemisphere, althoughthis did not manifest until the interval between prime and target was 350 ms. Impor-tantly, an ambiguous word directed to either hemisphere activated a more extensivearray of meanings in the contralateral hemisphere than in the hemisphere to whichthe prime was directed. Furthermore, the magnitude of facilitation for LVF–RVFpresentations was equivalent for dominant and subordinate meanings at both SOAs,just as it was for RVF–LVF presentations. This suggests that meaning dominancedid not mediate the level of activation of alternate meanings in the contralateral hemi-sphere. Rather, it appears that the processing of ambiguous text by a given hemisphereentails equivalent activation of a range of alternate meanings in the contralateralhemisphere.
Although the current experiment provides support for Burgess and Simpson’s(1988) suggestion that one aspect of linguistic processing in the left hemisphere en-tails the activation of a range of alternative meanings in the right hemisphere, likeChiarello et al. (1995), there is no support for their proposal that subordinate mean-ings become active in the right hemisphere only after they have been suppressed inthe left hemisphere. At an SOA of 350 ms, concurrent facilitation was found forsubordinate meanings presented in the RVF–RVF and RVF–LVF conditions. Evi-dently, the processing of an ambiguous word by the left hemisphere involves concur-rent activation of its subordinate meanings in both hemispheres. Instead, this experi-ment provides strong support for the view of interhemispheric communication positedby the direct connections model, where the processing of ambiguous text involvesthe activation of a range of alternative meanings in the contralateral hemisphere viadirect callosal connections. This indicates that interhemispheric cooperation is essen-tial for successful interpretation of ambiguous text. Therefore, an interruption of com-munication between the hemispheres may underlie the linguistic deficits noted inpatients who have sustained damage to the right hemisphere (Brownell, 1988;Brownell et al., 1984; Weylman et al., 1989; Kaplan et al., 1990; Beeman, 1993)rather than interference in right hemisphere processing per se.
The suggestion that interhemispheric cooperation is integral to the interpretationof ambiguous text is consistent with a recent study conducted by Hasbrooke andChiarello (1998). They used a lexical decision task with centrally positioned homo-graph primes, and targets which were associated with the prime’s dominant or subor-dinate meaning. The impact of hemispheric cooperation was measured by the inclu-sion of bilateral and unilateral visual field presentations where, after an SOA of 750ms, targets were projected to one visual field or redundant targets were simulta-neously presented in both visual fields. The pattern of priming for unilateral andbilateral presentations indicated that the two hemispheres cooperate in processingambiguous words, with the left hemisphere making a greater contribution. On unilat-eral trials, both LVF and RVF priming was found for dominant meanings, whilepriming for subordinate meanings was restricted to RVF presentations. Bilateral vi-sual field trials produced priming effects which were similar to those observed forRVF presentations, although responses were generally faster and more accurate thanresponses to unilateral presentations. Unlike Burgess and Simpson’s (1988) study,subordinate priming was restricted to the left hemisphere. On the basis of these find-ings, Hasbrooke and Chiarello concluded that bilateral exhaustive activation of am-biguous word meanings is followed by a yoking of left hemisphere meaning selectionwith the availability of alternative meanings in the right hemisphere. The results ofthe current experiment indicate that this exhaustive bilateral activation of alternative

94 MARJORIE COLLINS
meanings involves cross-hemisphere activation via direct callosal connections ratherthan activation of the full range of meanings within a particular hemisphere. Thisconclusion is based upon the cross-hemisphere priming effects found at an SOA of350 ms for LVF–RVF and RVF–LVF presentations of dominant and subordinatemeanings, in contrast to within-hemisphere priming, which was restricted to RVF–RVF presentations of homograph primes and their subordinate meanings.
The importance of cross-hemisphere communication in generating alternativemeanings of ambiguous text has not been evident in studies which have lateralizedthe target, but presented the prime centrally (i.e. Chiarello et al., 1995; Burgess &Simpson, 1988). The findings of the current study differ from these studies, althoughthis is to be expected as previous research clearly demonstrates that priming arisingfrom central primes differs markedly from priming associated with lateral presenta-tion of both prime and target. For instance, Chiarello et al. (1990) found symmetricalpriming in each hemisphere when primes were presented centrally, but hemisphericdifferences emerged when primes and targets were lateralized. Similarly, Koivistoand Laine (2000) found equivalent categorical priming in each hemisphere whenprimes were presented centrally, while hemispheric differences emerged when bothprime and target were presented laterally. One explanation for these differences isthat lateral presentation stalls processing of the prime and thereby stalls the activationof associated concepts. It is also possible that a stimulus directed to one hemisphereinitiates a pattern of activation that differs from the activation associated with projec-tion of the same stimulus to the contralateral hemisphere. The differences in cross-and within-hemisphere facilitation in the current experiment support this suggestion.Zaidel et al.’s (1988) conclusion that the use of central primes is problematic, as theleft hemisphere may ‘‘take over in some individuals’’ (p. 87), is also consistent withthis view. Their caution must also be applied when interpreting priming effects de-rived from a naming task. It is generally acknowledged that the naming response isgenerated by the left hemisphere (Koivisto, 1999). Hence, even when the focus ofactivation lies within the right hemisphere, the naming response is generated by theleft hemisphere, which is likely to obscure the unique contributions of each hemi-sphere in performing this task. It is interesting in this context, that Chiarello et al.(1995) found equivalent facilitation for dominant and subordinate meanings in eachhemisphere at SOAs ranging from 80 to 200 ms and used a task that involved centralprime presentation and a naming response. In view of the current findings of asym-metrical priming for alternative meanings of ambiguous words, it seems likely that thecombination of central primes with a naming task obscures the unique contribution ofeach hemisphere to the generation of alternative meanings. The differences in primingobserved by Hasbrooke and Chiarello (1998) for unilateral and bilateral presentationsis consistent with this suggestion, particularly as the pattern of priming in their studyindicated that interhemispheric interaction plays a role in processing ambiguous wordmeanings.
In conclusion, this study has confirmed Hasbrooke and Chiarello’s (1998) observa-tion that interhemispheric communication plays an integral part in the interpretationof ambiguous text. It has also produced evidence consistent with the model of inter-hemispheric communication postulated by the direct connections model (Collins &Coney, 1998) and failed to find any support for the homotopic inhibition theory(Cook, 1986). It seems that an ambiguous word directed to either hemisphere acti-vates a range of alternative meanings in the contralateral hemisphere, and this occursindependently of any activation of alternative meanings in the directly stimulatedhemisphere. This parallels previous findings of cross-hemisphere cooperation in gen-erating categorical and associative aspects of word meanings (Collins & Coney, 1998;(Collins) Abernethy & Coney, 1990, 1993; Collins & Frew, 2001). Taken together,

INTERHEMISPHERIC COMMUNICATION 95
these findings suggest that interhemispheric cooperation plays an essential role in theextraction of meaning in linguistic processing in general.
REFERENCES
Beeman, M. (1993). Semantic processing in the right hemisphere may contribute to drawing inferencesfrom discourse. Brain and Language, 44, 80–120.
Brownell, H. (1988). Appreciation of metaphoric and connotative word meanings by brain damagedpatients. In C. Chiarello (Ed.), Right hemisphere contributions to lexical semantics. Berlin: Springer-Verlag.
Brownell, H., Potter, H., & Michelow, D. (1984). Sensitivity to lexical denotation and connotation inbrain-damaged patients: A double dissociation? Brain and Language, 22, 253–265.
Brownell, H., Simson, T., Bihrle, A., Potter, H., & Gardner, H. (1990). Appreciation of metaphoricalternative word meanings by left and right brain-damaged patients. Neuropsychologia, 28, 375–383.
Bryden, M. P. (1982). Laterality: Functional asymmetry in the intact brain. New York: Academic Press.
Burgess, C., & Simpson, G. (1988). Cerebral hemispheric mechanisms in the retrieval of ambiguousword meanings. Brain and Language, 33, 86–103.
Chiarello, C. (1988). Lateralization of lexical processes in the normal brain: A review of visual half-field research. In H. Whitaker (Ed.), Contemporary reviews in neuropsychology. Berlin: Springer-Verlag.
Chiarello, C., Burgess, C., Richards, L., & Pollock, A. (1990). Semantic and associative priming in thecerebral hemispheres: Some words do, some words don’t . . . sometimes, some places. Brain andLanguage, 38, 75–104.
Chiarello, C., Maxfield, L., & Kahan, T. (1995). Initial right hemisphere activation of subordinate wordmeanings is not due to homotopic callosal inhibition. Psychonomic Bulletin & Review, 2, 375–380.
Collins, M. (1999). Differences in semantic category priming in the left and right cerebral hemispheresunder automatic and controlled processing conditions. Neuropsychologia, 37, 1071–1085.
(Collins) Abernethy, M., & Coney, J. (1990). Semantic and phonemic priming in the cerebral hemi-spheres. Neuropsychologia, 28, 933–945.
(Collins) Abernethy, M., & Coney, J. (1993). Associative priming in the hemispheres as a function ofSOA. Neuropsychologia, 31, 1397–1409.
(Collins) Abernethy, M., & Coney, J. (1996). Semantic category priming in the left cerebral hemisphere.Neuropsychologia, 34, 339–350.
Collins, M., & Coney, J. (1998). Interhemispheric communication is via direct connections. Brain andLanguage, 64, 28–52.
Collins, M., & Frew, A. (2001). Right hemisphere contributions to the comprehension of low imagerywords. Laterality, 6, 111–132.
Cook, N. (1986). The brain code. London: Methuen.
Hardyck, C., Chiarello, C., Dronkers, N., & Simpson, G. (1985). Orienting attention within visual fields:How efficient is interhemispheric transfer? Journal of Experimental Psychology: Human Perceptionand Performance, 11, 650–666.
Hasbrooke, R., & Chiarello, C. (1998). Bihemispheric processing of redundant bilateral lexical informa-tion. Neuropsychology, 12, 78–94.
Kaplan, J., Brownell, H., Jacobs, J., & Gardner, H. (1990). The effects of right hemisphere damage onthe pragmatic interpretation of conversational remarks. Brain and Language, 38, 315–333.
Keppel, G. (1991). Design and analysis: A researcher’s handbook. Englewood Cliffs, NJ: Prentice-Hall.
Koivisto, M. (1997). Time course of semantic activation in the cerebral hemispheres. Neuropsychologia,35(4), 497–504.
Koivisto, M. (1999). Backward priming and postlexical processing in the right hemisphere. Laterality,3(1), 21–40.
Koivisto, M., & Laine, M. (2000). Hemispheric asymmetries in activation and integration of categoricalinformation. Laterality, 5, 1–21.
Kucera H., & Francis, W. (1967). Computational analysis of present-day American English. Providence,RI: Brown Univ. Press.

96 MARJORIE COLLINS
Nakagawa, A. (1991). Role of anterior and posterior attention networks in hemispheric asymmetriesduring lexical decisions. Journal of Cognitive Neuroscience, 3, 313–321.
Nelson, D., McEvoy, C., Walling, J., & Wheeler, J. (1980). The University of South Florida homographnorms. Behaviour, Research Methods and Instrumentation, 12, 16–37.
Rodel, M., Cook, N., Regard, M., & Landis, T. (1992). Hemispheric dissociation in judging semanticrelations: Complementarity for close and distant associates. Brain and Language, 43, 448–459.
Taylor, M. (1988). The bilateral cooperative model of reading. In D. de Kerckhove & C. Lumsden (Eds.),The alphabet and the brain. Berlin: Springer-Verlag.
Tompkins, C. (1990). Knowledge and strategies for processing lexical metaphor after right or left hemi-sphere brain damage. Journal of Speech and Hearing Research, 33, 307–316.
Twilley, L., Dixon, P., Taylor, D., & Clark, K. (1994). University of Alberta norms of relative meaningfrequency for 566 homographs. Memory & Cognition, 22, 111–126.
Weylman, S., Brownell, H., Roman, M., & Gardner, H. (1989). Appreciation of indirect requests byleft- and right-brain damaged patients: The effects of verbal context and conventionality of wording.Brain and Language, 36, 580–591.
Zaidel, E. (1983). Disconnection syndrome as a model for laterality effects in the normal brain. InJ. Hellige (Ed.), Cerebral hemisphere asymmetry: Method, theory, and application. New York:Praeger.
Zaidel, E., White, H., Sakurai, E., & Banks, W. (1988). Hemispheric locus of lexical congruity effects:Neuropsychological reinterpretation of psycholinguistic results. In C. Chiarello (Ed.), Right hemi-sphere contributions to lexical semantics. Berlin: Springer-Verlag.








![Falx and Interhemispheric Fissure on Axial CT: I. falx cerebri and interhemispheric fissure, although recognized early on axial CT [1], received little attention in the literature.](https://static.fdocuments.in/doc/165x107/5d35b31788c993ee5c8c0e1d/falx-and-interhemispheric-fissure-on-axial-ct-i-falx-cerebri-and-interhemispheric.jpg)









