
Interactive Effects of Prey and Weather on Golden Eagle...
13
Journalof Animal Interactive effects of prey and weather on golden eagle Ecology1997, 66, 350-362 reproduction KAREN STEENHOF, MICHAEL N. KOCHERT and TRENT L. MCDONALD* US National Biological Service, Raptor Research and Technical Assistance Center, 970 Lusk Street, Boise, ID 83706, USA; and * Western EcoSystems Technology, 2003 Central Avenue, Cheyenne, Wyoming 82001 USA Summary 1. The reproduction of the golden eagle Aquila chrysaetos was studied in southwestern Idaho for 23 years, and the relationship between eagle reproduction and jackrabbit Lepus californicus abundance, weather factors, and their interactions, was modelled using general linear models. Backward elimination procedures were used to arrive at parsimonious models. 2. The number of golden eagle pairs occupying nesting territories each year showed a significant decline through time that was unrelated to either annual rabbit abundance or winter severity. However, eagle hatching dates were significantly related to both winter severity and jackrabbit abundance. Eagles hatched earlier when jackrabbits were abundant, and they hatched later after severe winters. 3. Jackrabbit abundance influenced the proportion of pairs that laid eggs, the pro- portion of pairs that were successful, mean brood size at fledging, and the number of young fledged per pair. Weather interacted with prey to influence eagle reproductive rates. 4. Both jackrabbit abundance and winter severity were important in predicting the percentage of eagle pairs that laid eggs. Percentage laying was related positively to jackrabbit abundance and inversely related to winter severity. 5. The variables most useful in predicting percentage of laying pairs successful were rabbit abundance and the number of extremely hot days during brood-rearing. The number of hot days and rabbit abundance were also significant in a model predicting eagle brood size at fledging. Both success and brood size were positively related to jackrabbit abundance and inversely related to the frequency of hot days in spring. 6. Eagle reproduction was limited by rabbit abundance during approximately two- thirds of the years studied. Weather influenced how severely eagle reproduction declined in those years. 7. This study demonstrates that prey and weather can interact to limit a large raptor population's productivity. Smaller raptors could be affected more strongly, especially in colder or wetter climates. Key-words: Aquila chrysaetos, black-tailed jackrabbits, Idaho, Lepus californicus, modelling. Journal of Animal Ecology (1997) 66, 350-362 Introduction breeding areas (Galushin 1974; Hamerstrom 1979; Smithet al. 1981; Korpimaki & Hongell 1986; Kor- Numerous studies of raptors have emphasized the pimaki, Lagerstr6m & Saurola 1987;Korpimaki & close relationship between raptorreproduction and Norrdahl 1991). In other species, most individuals the densities of principal preyspecies (e.g. Woffinden remain on nesting territories but do not lay eggs dur- & Murphy 1977; Adamcik, Todd& Keith 1978; Smith ing periods of low prey abundance (Southern 1970; & Murphy1979; Smith,Murphy & Woffinden 1981; Adamcik et al. 1978; Village1981). Stillother raptors ? 1997 British Korpimaki 1984, 1992). In some species, most indi- lay fewer eggs and/or experience increased egg or nes- Ecological Society viduals respond to prey declines by moving to other tling mortality when preypopulations are low (Cave 350
Transcript of Interactive Effects of Prey and Weather on Golden Eagle...

Journalof Animal Interactive effects of prey and weather on golden eagle Ecology 1997, 66, 350-362 reproduction
KAREN STEENHOF, MICHAEL N. KOCHERT and TRENT L. MCDONALD* US National Biological Service, Raptor Research and Technical Assistance Center, 970 Lusk Street, Boise, ID
83706, USA; and * Western EcoSystems Technology, 2003 Central Avenue, Cheyenne, Wyoming 82001 USA
Summary
1. The reproduction of the golden eagle Aquila chrysaetos was studied in southwestern Idaho for 23 years, and the relationship between eagle reproduction and jackrabbit Lepus californicus abundance, weather factors, and their interactions, was modelled
using general linear models. Backward elimination procedures were used to arrive at
parsimonious models. 2. The number of golden eagle pairs occupying nesting territories each year showed a significant decline through time that was unrelated to either annual rabbit abundance or winter severity. However, eagle hatching dates were significantly related to both winter severity and jackrabbit abundance. Eagles hatched earlier when jackrabbits were abundant, and they hatched later after severe winters. 3. Jackrabbit abundance influenced the proportion of pairs that laid eggs, the pro- portion of pairs that were successful, mean brood size at fledging, and the number of
young fledged per pair. Weather interacted with prey to influence eagle reproductive rates. 4. Both jackrabbit abundance and winter severity were important in predicting the
percentage of eagle pairs that laid eggs. Percentage laying was related positively to
jackrabbit abundance and inversely related to winter severity. 5. The variables most useful in predicting percentage of laying pairs successful were rabbit abundance and the number of extremely hot days during brood-rearing. The number of hot days and rabbit abundance were also significant in a model predicting eagle brood size at fledging. Both success and brood size were positively related to
jackrabbit abundance and inversely related to the frequency of hot days in spring. 6. Eagle reproduction was limited by rabbit abundance during approximately two- thirds of the years studied. Weather influenced how severely eagle reproduction declined in those years. 7. This study demonstrates that prey and weather can interact to limit a large raptor population's productivity. Smaller raptors could be affected more strongly, especially in colder or wetter climates.
Key-words: Aquila chrysaetos, black-tailed jackrabbits, Idaho, Lepus californicus, modelling.
Journal of Animal Ecology (1997) 66, 350-362
Introduction breeding areas (Galushin 1974; Hamerstrom 1979; Smith et al. 1981; Korpimaki & Hongell 1986; Kor-
Numerous studies of raptors have emphasized the pimaki, Lagerstr6m & Saurola 1987; Korpimaki & close relationship between raptor reproduction and Norrdahl 1991). In other species, most individuals the densities of principal prey species (e.g. Woffinden remain on nesting territories but do not lay eggs dur- & Murphy 1977; Adamcik, Todd & Keith 1978; Smith ing periods of low prey abundance (Southern 1970; & Murphy 1979; Smith, Murphy & Woffinden 1981; Adamcik et al. 1978; Village 1981). Still other raptors
? 1997 British Korpimaki 1984, 1992). In some species, most indi- lay fewer eggs and/or experience increased egg or nes- Ecological Society viduals respond to prey declines by moving to other tling mortality when prey populations are low (Cave
350

1968; Smith & Murphy 1979; Village 1981; Korpimaki 1984, 1987, 1992).
Few studies have documented weather as a limiting factor for raptor reproduction. Weather can affect
raptors through its effect on prey or on hunting behav-
iour, and adverse weather also can directly affect the survival of both adults and young. Rainfall may influence foraging success and thereby affect either a female's body reserves prior to egg-laying or a male's
ability to deliver prey to an incubating female (Hirons 1982; Village 1986). European kestrel Falco tin- nunculus L. laying dates are influenced by winter and
spring precipitation and spring temperatures (Cave 1968), and nesting failures correlate with the amount of spring rainfall (Village 1986). Annual nesting suc- cess rates of African black eagles Aquila verreauxi Lesson in Rhodesia (Gargett 1977) and honey buz- zards Pernis apivorus L. in Europe (Kostrzewa 1989) are also related inversely to rainfall. Red-tailed hawk Buteo jamaicensis Gmelin nestling mortality in Alberta is related to the biomass of food delivered to the nest as well as the frequency of rain (Adamcik, Todd & Keith 1979). Cold and rain are associated with mortality of small nestlings of both northern
goshawks Accipiter gentilis L. and common buzzards Buteo buteo L. in Europe (Kostrzewa & Kostrzewa
1990). In Wyoming, prairie falcon Falco mexicanus
Schlegel pairs abandoned their nesting attempts dur-
ing a spring snowstorm (Squires, Anderson & Oakleaf
1991). Bald eagle Haliaeetus leucocephalus L. repro- ductive success is negatively correlated with the sever-
ity of spring weather (Swenson, Alt & Eng 1986). Severe winter weather depleted nesting populations
of common barn-owls (Tyto alba Scopoli) and reduced their productivity the following spring (Marti &
Wagner 1985). In Germany, the density of kestrel
pairs and the percentage of pairs laying eggs are
closely related to winter temperatures and snow cover as well as vole abundance (Kostrzewa & Kostrzewa
1991). Kestrels are more likely to starve during severe
winters, and surviving kestrels are less likely to lay eggs because of their reduced mass. Winter severity, however, does not affect density or laying rates of the larger common buzzards and northern goshawks (Kostrzewa & Kostrzewa 1991), and Tengmalm's owl
Aegolius funereus L. laying dates are unaffected by winter temperatures and snow depth (Korpimaki & Hakkarainen 1991).
Golden eagle Aquila chrysaetos L. reproductive rates are known to fluctuate with prey densities (Smith & Murphy 1979; Tjernberg 1983; Watson & Langslow 1989; Bates & Moretti 1994). Failure of eagles to lay eggs during periods of low prey abundance has been documented in both Sweden (Tjernberg 1983) and the western United States (Smith & Murphy 1979). However, some evidence suggests that this response may be mitigated by mild winter weather (Tjernberg 1983). Spring weather also may affect survival of
golden eagle nestlings. Mosher & White (1976) con-
cluded that golden eagle young are susceptible to ther- mal stress during the first 6 weeks after hatching, particularly between 3 and 6 weeks of age. Beecham & Kochert (1975) found that heat stress was a significant mortality factor for nestling eagles in Idaho.
In this paper we examine 23 years of data on golden eagle reproductive rates in relation to both weather conditions and prey abundance. Golden eagles that nest in southwestern Idaho are year-round residents
(Steenhof, Kochert & Moritsch 1984), and black- tailed jackrabbits Lepus californicus Gray are their
principal prey (Steenhof & Kochert 1988). We pre- dicted that rabbit abundance would influence whether female eagles laid eggs and whether pairs would suc-
cessfully raise young. We also predicted that these effects would be modified by weather. Winter severity should influence the proportion of females that lay eggs, particularly when increased energy demands occur during periods of decreased prey availability. Spring weather should influence nesting success of
pairs that lay eggs: cold, wet weather during incu- bation and early brood-rearing could result in chilling of eggs or small young; extremely hot weather could cause increased nestling mortality.
Study area
The study area was 243 000 ha of public and private lands within the Snake River Birds of Prey National Conservation Area (NCA) in southwestern Idaho, USA (42?', 115?'W). The principal physiographic fea- ture of the study area is the Snake River Canyon, with basalt cliffs ranging from 2 to 125m in height. Topography above the canyon is generally flat or
slightly rolling with a few isolated buttes. Elevation
ranges from 770 m in the canyon bottom to 1000 m at the rim. Native vegetation in the area is characteristic of a shrub steppe community, with big sagebrush Art- emisia tridentata Nutt., shadscale Atriplex con-
fertifolia [Torr. and Frem.] Wats. and winterfat Cera- toides lanata [Pursh] J.T. Howell vegetation associations (US Department of the Interior 1979). Wildfires destroyed approximately 50% of the shrub cover between 1980 and 1985 (Kochert & Pellant
1986), resulting in large stands of exotic annual grasses
(primarily Bromus tectorum L.) and forbs. Annual
precipitation averages 20cm and occurs mainly in
winter; summers are hot and dry.
Methods
EAGLE NESTING DENSITY AND REPRODUCTION
From 1971 to 1994, the entire NCA was searched for
nesting golden eagles. Nearly all golden eagle nesting territories had been identified by previous research when our surveys began (Beecham 1970; Kochert
1972). We located occupied territories by observing
351 K. Steenhof, M.N. Kochert & T.L. McDonald
? 1997 British Ecological Society Journal of Animal Ecology, 66, 350-362

territorial activity, courtship, brood-rearing activity, eggs, young, or any other conspicuous field sign (e.g. whitewash at a roost). Any area of cliff or power line where nests were found in successive years and where no more than one eagle pair bred at one time was considered a nesting territory (Newton & Marquiss 1982). In each year, we considered the number of
nesting pairs to be equivalent to the number of occu-
pied territories. From 1971 to 1994 we monitored reproductive
activity at all known eagle nesting territories. We sur-
veyed each territory at least twice each year from foot, boat, helicopter or fixed-wing aircraft. Most surveys prior to 1984 were from the ground; most surveys from 1984 to 1994 were by helicopter. We used fixed-
wing aircraft only in 1971. Between 1971 and 1981, we entered some nests as often as every 4 days to assess nestling diet (Steenhof & Kochert 1988). To minimize stress to eggs or young, we did not enter nests during inclement weather. From 1985 to 1994, we conducted at least two helicopter surveys each
year. The first survey in March determined if ter- ritories were occupied and if adults were incubating. We rechecked territories with incubating birds during a second flight in late May or early June to assess
productivity. All territories for which the breeding outcome could not be determined from the helicopter were surveyed from the ground.
We categorized eagle pairs that showed no evidence of egg laying after repeated observations or after
climbing into and examining potential nests as 'non-
laying.' We confirmed 'laying' if an occupied territory contained an incubating adult, eggs, young, or any other indication that eggs were laid (e.g. fresh eggshell fragments in fresh nesting material). A pair was 'suc- cessful' if it produced > 1 young that reached 80% of normal fledging age [i.e. 51 days (Steenhof 1987)]. A small number of eagle nests (< 10) inspected after
young had fledged were considered successful if: (i) a nest platform decorated the same year was worn flat and contained fresh prey remains; (ii) fresh faecal matter covered the back and extended over the nest's
edge; and (iii) no dead young birds were found within a 50-m radius of the nest (Steenhof & Kochert 1982).
Our analysis of nesting success and productivity excluded nesting attempts where research personnel had treated diseased young, installed shade devices, radioed adults, or transferred young in or out of nests. These activities, which may have influenced the out- come of a nesting attempt (M. Kochert, unpublished data), occurred at fewer than 15% of the nesting attempts over all years. We also excluded nests where
investigators were known to have caused egg or chick mortality (n = 6).
We assigned median hatching dates to all nests where we observed young prior to fledging (n = 873). We used a photographic ageing key (Hoechlin 1976) to age young. We were unable to determine hatching dates at nests where young died or fledged before we
could age them, and we were unable to assign laying dates to pairs that failed during incubation.
We considered the following reproductive par- ameters in our analyses: percentage of territorial pairs laying eggs; percentage of laying pairs that were suc- cessful; brood size at fledging; and number of young fledged per territorial pair. Brood size at fledging was based on counts of young in successful nests at or after 80% of fledging age (51 days). It includes infor- mation on clutch sizes and egg and nestling survival rates at successful nests, which we did not measure
directly. Number of young fledged per territorial pair was a derived variable that described overall pro- ductivity of the population. It was calculated as the
product of percentage of pairs laying, percentage of
laying pairs successful, and brood size at fledging.
PREY ABUNDANCE
Each year from 1977 to 1991, black-tailed jackrabbit abundance was assessed by spotlighting from a vehicle
along line transects in May and June (Smith & Nyd- egger 1985). Jackrabbit densities for the NCA were estimated using the program DISTANCE (Buckland et al. 1993). In 1971, 1973 and 1975-1983 the average number of jackrabbits seen per day from late March to July by a 2-person raptor survey crew was tabulated from field notes. Jackrabbits per crew-day and jack- rabbit densities from spotlighting were correlated
(r = 0-86) from 1977 to 1983, and the relationship (y = 0-078 + 0-063x, where y = predicted density andx = jackrabbits seen per crew-day) was used to estimate pre-1977 jackrabbit densities. We used the mean of previous and current spring densities ('mid- point' or estimated winter density) in our analyses. This measure incorporates information about rabbits from both years that could influence eagle repro- duction. We assumed that the midpoint density would best approximate rabbit density during egg-laying in late winter and incubation in early spring.
WEATHER
We obtained temperature and precipitation data from US Department of Commerce climatological sum- maries for three weather stations (Bruneau, Grand View and Swan Falls) in or near the NCA. We obtained data on snow and heating degree days from summaries for the Boise Airport. We analysed five variables that described the severity of winter immedi-
ately preceding the breeding season: number of days when the temperature never rose above 20 ?F (-7 ?C); number of days when the temperature fell below 0 ?F
(-18 C); number of days with snow cover; the last date of continuous snow cover on the ground; and heating degree days. One heating degree day is accumulated for each whole Fahrenheit degree that the mean daily temperature was below 65 ?F (18 3 ?C). We tallied heating degree days for the entire winter
352 Golden eagle reproduction
? 1997 British Ecological Society Journal of Animal Ecology, 66, 350-362

(November to February) and for the two coldest win- ter months (December and January). Other variables were summarized for the entire winter as well as for the period just prior to egg-laying (15 January to 15
February). Extreme heat was measured from 15 May to 15 June each year by counting the number of days when temperatures exceeded 32 ?C and 35 ?C. We also summarized the amount of precipitation during incu- bation (24 February to 10 April), brood-rearing (11 April to 15 June), and the entire nesting period (24 February to 15 June).
ANALYSIS
We undertook a model building process to investigate how prey and weather interact to affect golden eagle abundance, breeding chronology and reproduction. We first examined a SYSTAT (Wilkinson 1989) Pearson correlation matrix of data from all years to reduce the number of variables by eliminating non-independent or highly correlated variables. This reduced the pool of 19 weather and jackrabbit variables to 7. The pool of explanatory variables available in the model build-
ing process for each dependent variable was further reduced for two reasons. First, we did not use a poten- tial explanatory variable if the connection between it and a certain dependent variable did not make sense
biologically. For example, it did not make sense to use May/June temperatures to explain variation in
hatching dates because hatching occurred before May. Secondly, we kept the number of terms low in the models for which only 19-21 data points were avail- able because we did not want to overfit the data for certain dependent variables. As a result, the pool of variables available as explanatory variables differed in each model building and are listed separately for each dependent variable in the subsections below. Dis- tributional assumptions also differed for each variable due to the different nature of dependent variables. All of our modelling fits under the framework of 'general linear modeling.' We did not build a model for one
variable, number fledged per pair, because our best estimator for this variable was a function of three
dependent variables that we modelled, and conse-
quently its distributional properties were unknown.
Nesting abundance and breeding chronology
To assess factors influencing the number of nesting pairs and breeding chronology, we modelled number of nesting pairs and hatching date using regular nor- mal theory multiple regression and performed back- ward elimination of explanatory variables to reduce the number of terms. Terms in the full model were estimated winter jackrabbit densities per square kilo- metre (JRW), winter heating degree days (HDD), year, and the two-way interactions (JRW:YEAR, JRW:HDD and HDD:YEAR). Terms were elim- inated based on the magnitude of the individual term's
F statistic, which for normal theory models is roughly equivalent to the Akaike's information criterion (AIC) used below. Backward elimination stopped when elim- ination of additional terms drastically reduced the
predictive power of the model, as measured by the residuals and the F statistic.
Reproduction
We modelled the relationships between three depen- dent variables that characterize reproduction: per- centage of pairs laying eggs, percentage of laying pairs successful, and brood size at fledging. The explanatory variables used in the modelling process were estimated winter jackrabbit density (JRW) and the following weather variables: total precipitation (PRECIP), pre- cipitation during brood rearing (PRECIPB), number of days in January and February below-18 ?C (LT18), number of days in May and June above 32 ?C (GT32), number of days with snow cover (SNOW), and winter
heating degree days (HDD). A generalized linear
response surface model was fitted by backwards elim- ination with a forward look at each step (Weisberg 1985, chapter 8). The backward elimination was car- ried out using the S + (? 1995; MathSoft Inc., ver- sion 3-3 for Windows) routine step.glm. At each step in the elimination process, single terms were deleted from the model, and the AIC statistic (Akaike 1973, 1985) of the reduced model was calculated. If deleting a term improved the AIC statistic (i.e. lowered AIC), the term that lowered AIC the most upon deletion was eliminated. The forward look at each step involved
adding single terms eliminated in previous steps. If the AIC statistic was reduced upon addition of a term, the term that reduced AIC the most was added back to the model (Statistical Sciences 1995). Models simi- lar to the final model selected via backwards elim- ination were investigated to see if any further improve- ment in the model could be achieved. Statistical
significance of terms in the final model was assigned by dropping each in turn and performing a likelihood ratio test using the 'full' and 'reduced' likelihoods.
Percentage of pairs laying eggs. The initial logistic response surface model in the backwards elimination
process for percentage of laying pairs included JRW, HDD, SNOW, JRW2, HDD2, SNOW2, and the two-
way interactions JRW:HDD, JRW:SNOW and HDD:SNOW. The logistic response surface model assumed the number of pairs laying eggs each year to be a binomial random variable with index mk (mk = total pairs in year k) and probability Pk. Under the binomial assumption, the logistic model related log- it(pk) = lg(pk/(l-pk)) to a linear function of the
explanatory variables and obtained coefficient esti- mates by maximizing the binomial likelihood func- tion. All hypotheses about model coefficients were tested using likelihood ratio tests. The predictive power of other explanatory variables, relative to the
353 K. Steenhof, M.N. Kochert & T.L. McDonald
? 1997 British Ecological Society Journal of Animal Ecology, 66, 350-362

final model selected in the backwards elimination pro- cess, was investigated. Additional variables con- sidered after backwards elimination were LT18, PRE-
CIP, current year jackrabbit densities, previous year jackrabbit densities, and the interaction of previous year and current year jackrabbit densities. The wea- ther variables of PRECIP and LT18 were investigated by first dropping any weather variables from the final model and then adding PRECIP and LT18 one at a time to assess their predictive power. Current year jackrabbit density, previous year jackrabbit density, and the interaction were investigated by first dropping all jackrabbit variables from the model and then
adding all three to assess their predictive power.
Percentage of laying pairs successful. The initial logis- tic response surface model in the backward elim- ination procedure for percentage of laying pairs that
successfully raised young included JRW, GT32, HDD JRW2, GT322, HDD2 and the interactions
JRW:GT32, JRW:HDD and GT32:HDD. The logis- tic response surface model assumed the number of successful pairs each year to be a binomial random variable with index mk (mk = total pairs in year k) and
probability Pk The logistic model related logit(pk) =
log(p/(l-pk)) to a linear function of the explanatory variables and obtained coefficient estimates by max-
imizing the binomial likelihood function. All hypoth- eses were tested using likelihood ratio tests. The additional predictive power of other explanatory vari- ables was investigated, relative to the final model selec- ted in the backwards elimination process. Additional variables considered after backwards elimination were
PRECIP, LT18, SNOW, current year jackrabbit den-
sities, previous year jackrabbit densities, and the inter- action of previous year and current year jackrabbit densities. The weather variables of PRECIP, LT18 and SNOW were investigated by first dropping any weather variables from the final model and then
adding each in turn to assess their predictive power over current year jackrabbit density, previous year jackrabbit density, and the interaction were inves-
tigated by first dropping all jackrabbit variables from the model and then adding all three to assess their
predictive power.
Brood size at fledging. The initial model in the back- wards elimination procedure for brood size at fledging included JRW, HDD, GT32, PRECIPB, JRW2, HDD2, GT322, PRECIPB2, and all two-factor inter- actions among linear terms. The response surface model used for brood size assumed that the average brood size in each year was a normal random variable with variance proportional to the inverse of sample size in each year. Coefficients in the linear model were estimated through weighted least squares techniques with weights equal to the sample size in each year (Table 1).
Results
GOLDEN EAGLE NESTING POPULATIONS
An average of 31 golden eagle pairs occupied nesting territories in the NCA each year from 1971 to 1994
(range = 28-35, SD = 2-1; Table 1). The percentage of pairs that laid eggs each year (range: 38-100%, mean = 70%, SD = 15 5; Table 1) was relatively more variable than the number of pairs occupying ter- ritories (cv = 19-6 vs. 6-6; t = 4-0444, P < 0-001). The
percentage of laying pairs that successfully raised
young also showed considerable variation among years (range: 32-80%, mean = 60%, SD = 13-9, cv = 23-2). Brood size at fledging, in contrast, remained fairly stable across the years (range = 1 00-
2-00, mean = 1-56, SD = 0-22, cv = 14-0). Estimated number of young produced per pair reflected the vari- ation in percentage of pairs laying and percentage of
laying pairs successful (range = 0-16-1-38, mean = 0 79, SD = 0 35, cv = 44 3), as did total num- ber of young produced by the population (range = 5- 41,mean = 25, SD = 10-3, cv = 41-2). Hatching dates
ranged from 17 March to 18 May (mean = 13 April, SD = 10 days). The earliest mean annual hatching date was 5 April, and the latest was 21 April.
JACKRABBIT POPULATIONS
Estimated jackrabbit densities ranged from 1 3 to 54 8
per km2 between 1971 and 1994. Jackrabbit densities were highest in 1971, but they decreased sharply in 1973 and remained low through to 1978 (Fig. 1). Num- bers were high from 1979 to 1981, but the peak reached in 1979 was lower than that attained in 1971. Estimated densities declined again between 1981 and
1982, and the estimated population size between 1984 and 1986 was even lower than in the mid-1970s. In
1987, populations began rebuilding to a third peak in 1992, decreased sharply in 1993, and rebounded
slightly in 1994. Jackrabbit populations had peaked at successively lower densities from 1971, to 1979-81, and 1990-92. Knick & Dyer (1997) distinguished two
jackrabbit population phases (high and low) in the NCA. During our 23-year study, we identified 8 years when jackrabbit populations were at high phases and 15 years when they were in low phases of their popu- lation cycle (Fig. 1). Jackrabbit abundance was not correlated with any of the weather variables measured
(r's < 0-4; P's > 0-10).
EAGLE-JACKRABBIT-WEATHER RELATIONSHIPS
Nesting abundance and breeding chronology
Regression analyses showed that the number of eagle pairs occupying nesting territories each year was unre- lated to rabbit density and winter severity. Year, how-
ever, was the single significant variable in the final model for the number of nesting pairs. The number of
354 Golden eagle reproduction
? 1997 British Ecological Society Journal of Animal Ecology, 66, 350-362

Table 1. Nesting success and productivity of golden eagles in the NCA, 1971-94. Sample sizes are in parentheses
Year Number of occupied % of pairs nesting territories laying
1971 34 1972 34 1973 35 1974 35 1975 33 1976 35 1977 34 1978 32 1979 30 1980 31 1981 30 1982 30 1983 28 1984 31 1985 32 1986 29 1987 32 1988 32 1989 30 1990 30 1991 29 1992 30 1993 29 1994 30 Mean 31-5 SD 2-1 CV 6-6
100% (31)
65% (34) 67% (33) 75% (32) 68% (34) 82% (34) 75% (32) 97% (30) 87% (31)
100% (30) 87% (30) 96% (27)
38% (32) 55% (29) 78% (32) 74% (31) 80% (30) 87% (30) 90% (29) 86% (29) 90% (29) 63% (30) 79% 15-5 19-6
% of laying pairs Brood size at Number fledged Total successful fledging per pair* fledged
62% (21)
44% (18) 53% (17) 56% (18) 50% (16) 59% (17) 77% (13) 63% (24) 76% (21) 73% (22) 79% (24) 67% (18) 59% (17) 42% (12) 40% (14) 32% (25) 52% (23) 63% (19) 80% (25) 74% (23) 56% (16) 80% (15) 50% (12) 60% 13-9 23-2
1-89 1 64 1-38 1-50 1-45 1-60 1-50 1 70 1 40 1-78 1 74 2-00 1-57 1-56 1 00 1 20 1-38 1-54 1-36 1 65 1-59 1 73 1-71 1-71 1-57 0-22
14-0
(19) 1-17 (11) -
(8) 0.40 (10) 0.53 (11) 0-61 (10) 0.54 (10) 0.73 (10) 0.98 (15) 0-86 (18) 1-18 (19) 1-27 (18) 1-38 (14) 1.01 (9) 0-64t (4) 0-16 (5) 0-26 (8) 0.34 (13) 0-59 (14) 0-69 (20) 1-15 (17) 1-06 (11) 0-83 (14) 1-23 (7) 0-54
0-79 0-35 44.3
40
14 19 20 19 25 31 26 37 38 41 28 20 5 8
11 19 21 34 31 25 36 16 25 10-3 41.2
*Based on percentage of pairs laying, percentage of laying pairs successful, and brood size at fledging. tBased on percentage of pairs successful and brood size at fledging.
60
50 -
E 4 40 - High
30
30 -
.o20- L-rl z
10
Low [ , i I I
i
I I I I I I I I I I I I I I I I 0 I I
i i I i , i i , i i
i i i i i i i
i i I I , .
71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94
Fig. 1. Black-tailed jackrabbit densities (n km-2) in the Snake River Birds of Prey National Conservation Area, 1971-94. The shaded line separates 'high' and 'low' phases of the population cycle.
eagle pairs in the NCA showed a slight but significant primarily to estimated winter jackrabbit densities decrease during the 24-year study. Year explained (JRW) and also to winter severity, as measured by 57% of the variation in number of pairs (P < 0-001) HDD. Eagles hatched earlier in years when jack-
? 1997 British according to the linear regression: number of pairs rabbits were abundant, and they hatched later after
Ecological Society = 50'4-0-23 x YEAR. severe winters. The final model relating hatching dates Journal of Animal Golden eagle hatching dates, in contrast, showed no to explanatory variables was: hatching date = 95-2- Ecology, 66, 350-362 significant trends over time, and instead were related 0-236 x JRW + 0-002 x HDD. The three-dimen-
355 K. Steenhof, M.N. Kochert & T.L. McDonald
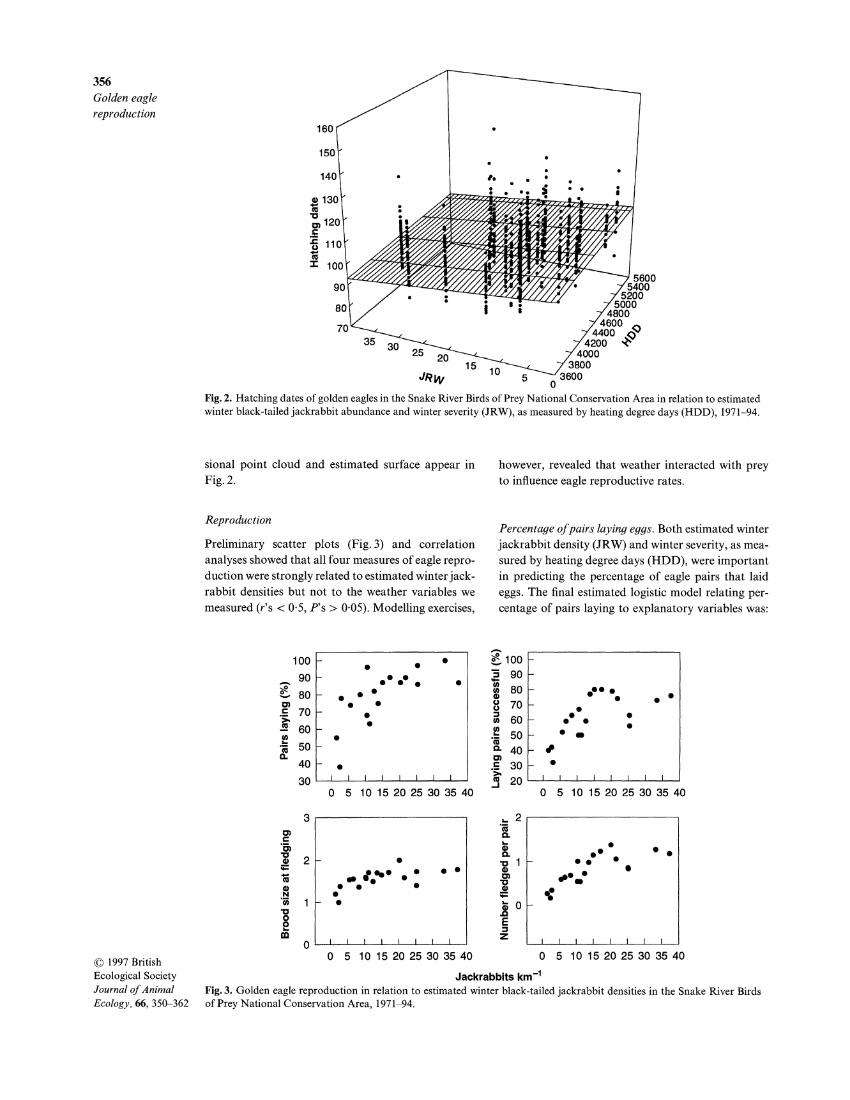
356 Golden eagle reproduction
v 2 0 ' "
'-7. %J%J 25 @ ;_ , 4000
25 20 3800 JRW 5 3600
Fig. 2. Hatching dates of golden eagles in the Snake River Birds of Prey National Conservation Area in relation to estimated winter black-tailed jackrabbit abundance and winter severity (JRW), as measured by heating degree days (HDD), 1971-94.
sional point cloud and estimated surface appear in
Fig. 2.
Reproduction
Preliminary scatter plots (Fig. 3) and correlation
analyses showed that all four measures of eagle repro- duction were strongly related to estimated winter jack- rabbit densities but not to the weather variables we measured (r's < 0-5, P's > 0-05). Modelling exercises,
however, revealed that weather interacted with prey to influence eagle reproductive rates.
Percentage of pairs laying eggs. Both estimated winter
jackrabbit density (JRW) and winter severity, as mea- sured by heating degree days (HDD), were important in predicting the percentage of eagle pairs that laid
eggs. The final estimated logistic model relating per- centage of pairs laying to explanatory variables was:
100 90
80 0) ., 70
- 60 0
? 50 a.
40 30
3
c la 0)
m
'o
*0 0
L. 0
2
1
0
? 1997 British Ecological Society Journal of Animal Ecology, 66, 350-362
@0. 0 . ?
0 0
0
0 5 10 15 202530 35 40
a s~- . - * 0 ? @
* 0 0 - 0
0 5 10 15 20 2530 35 40
lo ioo 3s 90 0 0 80 0
o 70 o 60 0
50 0- 40 cm r 30 t3 20
.
0 0.
0 U) 0 vc 1
0 lm
.0 1- o
E z
- 0-00 0 0_
0 0 - 05 0 0
0 a
-_ I II I I5 3 I I
0 5 10 15 20 25 30 35 40
?
400 *
_ a
0
I I I I I I I
0 5 10 15 202530 35 40
Jackrabbits km-1 Fig. 3. Golden eagle reproduction in relationon to estimated winter black-tailed jackrabbit densities in the Snake River Birds of Prey National Conservation Area, 1971-94.
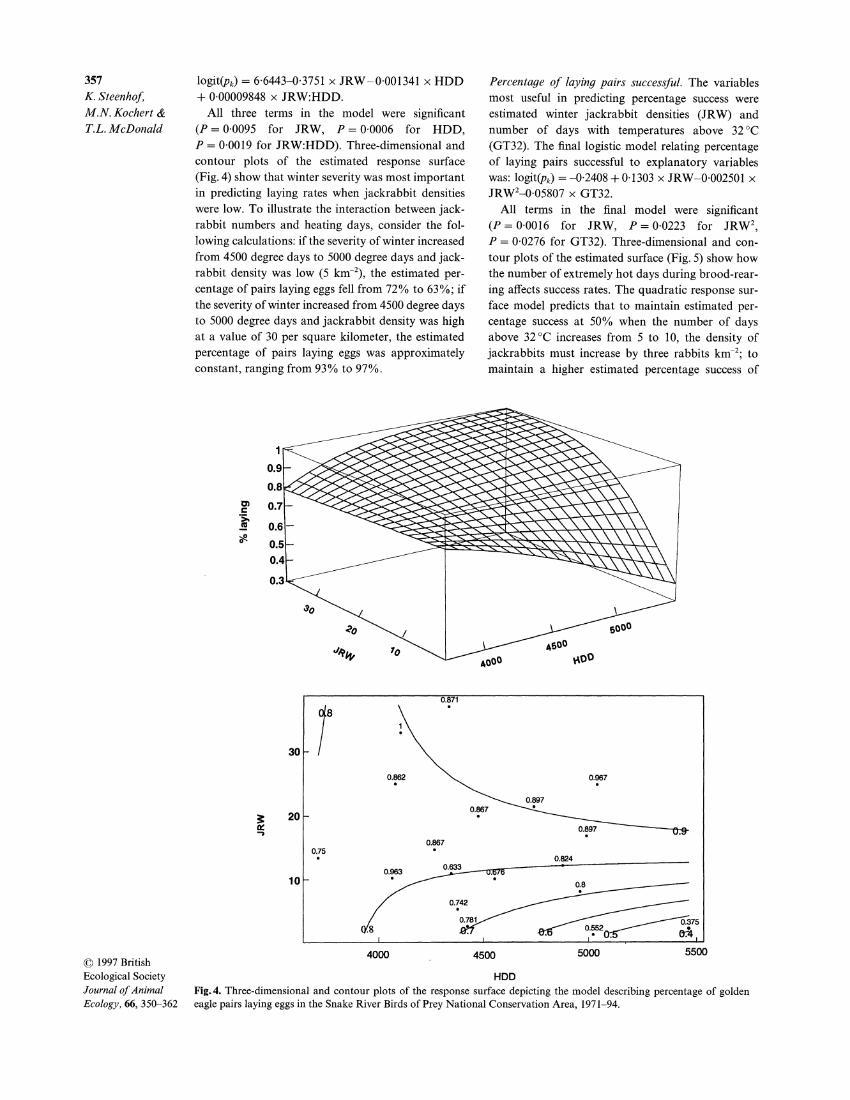
logit(pk) = 66443-0-3751 x JRW-0-001341 x HDD + 0-00009848 x JRW:HDD.
All three terms in the model were significant (P = 00095 for JRW, P = 0-0006 for HDD, P = 0-0019 for JRW:HDD). Three-dimensional and contour plots of the estimated response surface
(Fig. 4) show that winter severity was most important in predicting laying rates when jackrabbit densities were low. To illustrate the interaction between jack- rabbit numbers and heating days, consider the fol-
lowing calculations: if the severity of winter increased from 4500 degree days to 5000 degree days and jack- rabbit density was low (5 km-2), the estimated per- centage of pairs laying eggs fell from 72% to 63%; if the severity of winter increased from 4500 degree days to 5000 degree days and jackrabbit density was high at a value of 30 per square kilometer, the estimated
percentage of pairs laying eggs was approximately constant, ranging from 93% to 97%.
0.9 0.8
CD 0.7-
0.6 -
n. "^^
Percentage of laying pairs successful. The variables most useful in predicting percentage success were estimated winter jackrabbit densities (JRW) and number of days with temperatures above 32?C
(GT32). The final logistic model relating percentage of laying pairs successful to explanatory variables was: logit(pk) = -042408 + 0.1303 x JRW-0-002501 x JRW2-0-05807 x GT32.
All terms in the final model were significant (P= 00016 for JRW, P= 0-0223 for JRW2, P = 0-0276 for GT32). Three-dimensional and con- tour plots of the estimated surface (Fig. 5) show how the number of extremely hot days during brood-rear-
ing affects success rates. The quadratic response sur- face model predicts that to maintain estimated per- centage success at 50% when the number of days above 32 C increases from 5 to 10, the density of
jackrabbits must increase by three rabbits km-2; to maintain a higher estimated percentage success of
8
30
0.862
20 -
0.967 ?
0.867 a
0.867 0.75 ?~~~~~~~~~~~* ~~0.824
0.633 0.963 u.b/6 / *
0.8
0./ 742
0.781 .375 .~~~8 t ~~~0.552
1 1 1 1~~~~~~~~~~~~~~~~~~~O'
? 1997 British Ecological Society Journal of Animal Ecology, 66, 350-362
4000 4500 5000 5500
HDD Fig. 4. Three-dimensional and contour plots of the response surface depicting the model describing percentage of golden eagle pairs laying eggs in the Snake River Birds of Prey National Conservation Area, 1971-94.
357 K. Steenhof, M.N. Kochert & T.L. McDonald
10 -

358 Golden eagle reproduction
0
0
C0) 010
20-
0.8
0.7
GT32 Fig. 5. Three-dimensional and contour plots of the response surface depicting the model describing percentage of laying golden eagle pairs successful in the Snake River Birds of Prey National Conservation Area, 1971-94.
70% when the number of days above 32 ?C increases from 5 to 10, the density of jackrabbits must increase by 8-5 rabbits km-2.
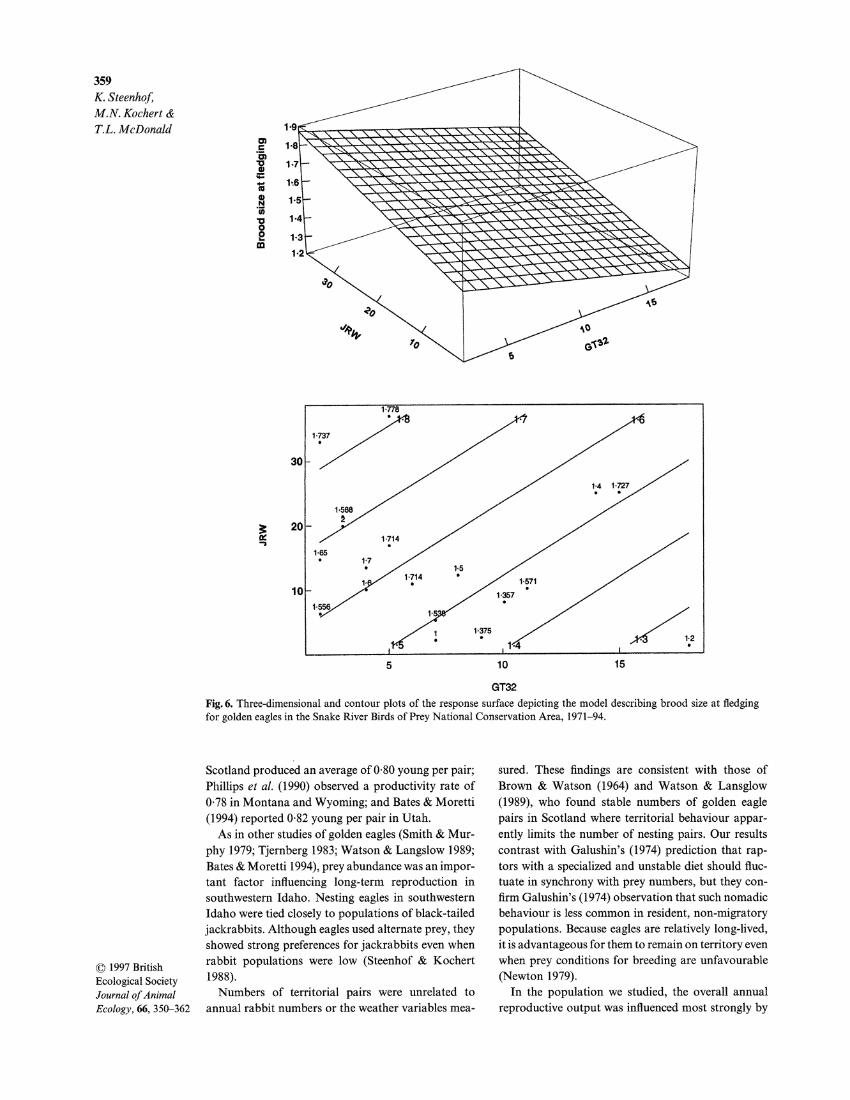
Brood size at fledging. Both winter rabbit densities (JRW) and the number of hot days during brood- rearing (GT32) also were significant (P = 0-024 for JRW, P = 0-029 for GT32) in the final model relating brood size at fledging to explanatory variables: (mean brood size) = 1-587 + 0-008702 x JRW - 0-01939 x GT32.
Three-dimensional and contour plots of the esti- mated surface are shown in Fig. 6. The magnitude of the estimated coefficients was relatively small, and the practical importance of the model for predicting future changes in brood size should be considered.
Under the least favourable conditions within the range of observed data, jackrabbit density was 1-5 km2, GT32 was 18, and the estimated average brood size was 1-25 chicks per nest. In the most favourable situ- ation within the range of observed data, jackrabbit density was 37 km2, GT32 was 2, and estimated aver- age brood size was 1-87 chicks per nest. All other estimated average brood sizes were between 1-25 and 1-87.
Discussion
Long-term productivity of golden eagles in south- western Idaho (0-79 young per pair) was similar to that of golden eagle populations in other parts of the world. Watson (1957) reported that golden eagles in
? 1997 British Ecological Society Journal of Animal Ecology, 66, 350-362
359 K. Steenhof, M.N. Kochert & T.L. McDonald
CD
cu lal 0
40
(I) 0 0 la 0 2~
30
20 -
I0--
1-2 ?
5 10 15
GT32
Fig. 6. Three-dimensional and contour plots of the response surfae depicting the model describing brood size at fledging for golden eagles in the Snake River Birds of Prey National Conservation Area, 1971-94.
Scotland produced an average of 0'80 young per pair; Phillips et al. (1990) observed a productivity rate of 0-78 in Montana and Wyoming; and Bates & Moretti
(1994) reported 0-82 young per pair in Utah. As in other studies of golden eagles (Smith & Mur-
phy 1979; Tjernberg 1983; Watson & Langslow 1989; Bates & Moretti 1994), prey abundance was an impor- tant factor influencing long-term reproduction in southwestern Idaho. Nesting eagles in southwestern Idaho were tied closely to populations of black-tailed
jackrabbits. Although eagles used alternate prey, they showed strong preferences for jackrabbits even when rabbit populations were low (Steenhof & Kochert
1988). Numbers of territorial pairs were unrelated to
annual rabbit numbers or the weather variables mea-
sured. These findings are consistent with those of Brown & Watson (1964) and Watson & Lansglow (1989), who found stable numbers of golden eagle pairs in Scotland where territorial behaviour appar- ently limits the number of nesting pairs. Our results contrast with Galushin's (1974) prediction that rap- tors with a specialized and unstable diet should fluc- tuate in synchrony with prey numbers, but they con- firm Galushin's (1974) observation that such nomadic behaviour is less common in resident, non-migratory populations. Because eagles are relatively long-lived, it is advantageous for them to remain on territory even when prey conditions for breeding are unfavourable (Newton 1979).
In the population we studied, the overall annual
reproductive output was influenced most strongly by
? 1997 British Ecological Society Journal of Animal Ecology, 66, 350-362

the proportion of pairs that laid eggs. As we predicted, golden eagle laying rates were related to food and weather prior to the nesting season. Both hatching dates and the percentage of pairs laying eggs were related to estimated winter jackrabbit densities and the severity of the winter preceding the nesting season. In Scotland, prey abundance was the most important factor influencing barn owl laying dates (Taylor 1994; p. 128), but owl laying dates also correlated closely with temperatures immediately before breeding. When Idaho jackrabbits were in the low phase of their
population cycle and after cold winters, eagles either laid eggs later or early clutches failed before hatching. Fewer golden eagles laid eggs in Idaho when jack- rabbits were scarce, and fewer laid eggs after severe winters. The severity of winter influenced the per- centage of pairs laying eggs primarily at low phases of the jackrabbit population cycle. During peaks in the jackrabbit cycle, the percentage of pairs laying eggs was relatively high regardless of how cold the winter was. Well-fed eagles can probably withstand cold better than food-stressed birds (Stalmaster & Gessaman 1984). Our results are consistent with
Tjernberg's (1983) finding that eagle reproduction is related to prey and weather prior to egg laying.
Weather during the nesting season also influenced
eagle reproduction in our study. The number of
extremely hot days during brood-rearing was impor- tant in explaining the variability in both percentage of laying pairs successful and mean brood size at
fledging. Both variables were related positively to
jackrabbit abundance and inversely to the number of hot spring days. Tomback & Murphy (1981) found that underfed ferruginous hawk Buteo regalis Gray nestlings were especially vulnerable to heat stress and that the combined effects of inadequate food and high temperatures caused increased nestling mortality in
years when prey populations were low. The number of golden eagle young fledged per pair
was the product of all three independent measures of
reproduction (percentage of pairs laying, percentage of laying pairs successful, and brood size at fledging), and it reflected the patterns described above. Jack- rabbit abundance influenced the number of young fledged per pair in all years. In peak rabbit years, number of young fledged per pair was related inversely to the number of hot days in May and June. During low phases of the jackrabbit population cycle, number
fledged per pair was related primarily (and inversely) to HDD and secondarily (and inversely) to the num- ber of days > 32 ?C in May and June.
Weather can affect raptors directly, and it also can influence them indirectly by affecting prey densities or
prey behaviour. Reproduction of Aquila and other
raptor populations in the southern hemisphere decreased following drought that reduced prey den- sities (Ridpath & Brooker 1986; Hustler & Howells
1990). However, in our study, both cold and heat affected eagles independently of their effects on the
abundance of the eagle's principal prey. Weather con- ditions were similar during low and high phases of the
jackrabbit population cycle. There was no evidence that rabbit populations were influenced by winter
severity or spring heat. Instead, rabbit abundance in our study area was unrelated to weather conditions and fluctuated cyclically, as has been documented in other parts of western North America (Gross, Stod- dart & Wagner 1974; Smith 1987). Weather, however, may have affected rabbit behaviour and their vul-
nerability to eagles. Weather also may have affected the availability of alternative prey due to either behav- iour or population changes that we did not measure.
Our study demonstrated how prey populations and weather can interact to limit a raptor population's productivity. Other non-migratory populations of
raptors that depend on cyclic prey in arctic and tem-
perate environments are likely to show similar pat- terns [e.g. kestrels (Kostrzewa & Kostrzewa 1991); snowy owls (Wiklund & Stigh 1986); and barn owls
(Taylor 1994)]. The shrub steppe environment we studied may be unique in that both extreme cold and extreme heat affected nesting populations. Rainfall, however, may be a more important factor for raptors in higher precipitation zones (Cave 1968; Adamcik et al. 1979; Village 1986). Precipitation was not a
significant factor in our southwestern Idaho study area, where rainfall rarely exceeded 5 cm during the
nesting season. Variations in prey and weather should have more serious consequences for smaller, short- lived raptors, because the relative costs of egg for- mation are higher for smaller species (Newton 1979). Kostrzewa & Kostrzewa (1991) found that, unlike
kestrels, northern goshawks and common buzzards are able to withstand severe winters in Germany because of their large masses, their relatively low
energy demands, and their consequent ability to ther-
moregulate effectively. Our results, however, show that even larger raptors can be influenced by cold weather at more southern latitudes.
Our results re-emphasize the importance of long- term studies. Shorter investigations would have been unable to identify the effects of weather on eagles because weather effects were overshadowed by prey effects and were only apparent in some years. Mod-
elling can be an effective approach to understand how weather and prey factors interact. Previous studies had documented short-term responses of raptors to severe weather conditions, but our study is one of the few that has examined how more subtle differences in weather and prey interact over the long term to influ- ence populations. Our results support Wiens' (1977) theory of variable environments in which resources are limiting in some, but not all, years. With golden eagles in southwestern Idaho, years when prey limit
reproduction were approximately twice as frequent as
years when prey were superabundant. Weather influ- enced how severe the effect of decreased prey would be.
360 Golden eagle reproduction
? 1997 British Ecological Society Journal of Animal Ecology, 66, 350-362

Acknowledgements
This paper is a contribution of the National Biological Service's Raptor Research and Technical Assistance Center (formerly the US Bureau of Land Management [BLM] Snake River Birds of Prey Research Project). The BLM provided funds from 1972 to 1994. From 1984 to 1989 Pacific Power and Light Company pro- vided funds for aerial surveys. The Idaho Army National Guard (IDARNG) assisted with funding from 1990 to 1994 as part of the Cooperative BLM/ IDARNG Research Project.
The Idaho Cooperative Wildlife Research Unit pro- vided the 1971 golden eagle data. J. H. Doremus and A. R. Bammann supervised the collection of nesting data between 1976 and 1983, and J. H. Doremus assisted with data collection from 1983 to 1990. R. N.
Lehman, L. B. Carpenter, and their crews helped to collect data from 1990 to 1994. The study could not have been completed without the assistance of numer- ous field technicians and dedicated volunteers, especially J.R. Oakley, T.E. Hamer, O.R. Fremming and M. W. Collopy.
Data on jackrabbit abundance were provided by M. L. Wolfe and L. O. Oftedahl, Department of Wildlife
Science, Utah State University (BLM contract 52500-
CT5-1003), G. W. Smith, D. J. Johnson, N. C. Nyd- egger, Department of Biology, University of Idaho
(BLM contract 52500-CT5-1002), M. L. McHenry (Idaho Army National Guard), J.H. Doremus (BLM Bruneau Resource Area), S. T. Knick and S. E. Watts
(National Biological Service). M. W. Collopy, E. Kor-
pimaki, J. M. Marzluff, J. T. Rotenberry, and anony- mous referees critically reviewed drafts of the manu-
script. L. McDonald provided valuable input on statistical modelling. D. M. Parrish tabulated and automated most of the data, and A. Teutsch prepared the graphics.
References
Adamcik, R.S., Todd, A.W. & Keith, L.B. (1978) Demo- graphic and dietary responses of great horned owls during a snowshoe hare decline. Canadian Field-Naturalist, 92, 156-166.
Adamcik, R.S., Todd, A.W. & Keith, L.B. (1979) Demo- graphic and dietary responses of red-tailed hawks during a snowshoe hare decline. Canadian Field-Naturalist, 93, 16-27.
Akaike, H. (1973) Information theory as an extension of the maximum likelihood principle. Second International Symposium on Information Theory (eds B. Petrov & F. Csaki), pp. 267-281. Akademiai Kiado, Budapest, Hun- gary.
Akaike, H. (1985) Prediction and entropy. A Celebration of Statistics (eds A. Atkinson & S. Fienberg), pp. 1-24. Springer, New York.
Bates, J.W. & Moretti, M.O. (1994) Golden eagle (Aquila chrysaetos) population ecology in eastern Utah. Great Basin Naturalist, 54, 248-255.
Beecham, J.J. Jr (1970) Nesting ecology of the golden eagle in southwestern Idaho. MSc thesis, University of Idaho, Moscow.
Beecham, J.J. Jr & Kochert, M.N. (1975) Breeding biology of the golden eagle in southwestern Idaho. Wilson Bulletin, 87, 506-513.
Brown, L.H. & Watson, A. (1964) The golden eagle in relation to its food supply. Ibis, 106, 78-100.
Buckland, S.T., Anderson, D.R., Burnham, K.P. & Laake, J.L. (1993) Distance Sampling: Estimating Abundance of Biological Populations. Chapman and Hall, New York.
Cave, A.J. (1968) The breeding of the kestrel, Falco tinnun- culus, L., in the reclaimed area Oostelijk Flevoland. Netherlands Journal of Zoology, 18, 313-407.
Galushin, V.M. (1974) Synchronous fluctuations in popu- lations of some raptors and their prey. Ibis, 116, 210-213.
Gargett, V. (1977) A 13-year population study of the black eagles in the Matopos, Rhodesia, 1964-76. Ostrich, 48, 17-27.
Gross, J.E., Stoddart, L.C. & Wagner, F.H. (1974) Demo- graphic analysis of a northern Utah jackrabbit population. Wildlife Monographs, 40, 71 pp.
Hamerstrom, F. (1979) Effect of prey on predator: voles and harriers. Auk, 96, 370-374.
Hirons, G.J.M. (1982) The effects of fluctuations in rodent numbers on breeding success in the tawny owl Strix aluco. Mammal Review, 14, 155-157.
Hoechlin, D.R. (1976) Development of golden eaglets in southern California. Western Birds, 7, 137-152.
Hustler, K. & Howells, W.W. (1990) The influence of pri- mary production on a raptor community in Hwange National Park, Zimbabwe. Journal of Tropical Ecology, 6, 343-354.
Knick, S.T. & Dyer, D.L. (1997) Distribution of black-tailed jackrabbit habitat determined by GIS in southwestern Idaho. Journal of Wildlife Management, 61, 75-86.
Kochert, M.N. (1972) Population status and chemical con- tamination of golden eagles in southwestern Idaho. MSc thesis, University of Idaho, Moscow.
Kochert, M.N. & Pellant, M. (1986) Multiple use in the Snake River Birds of Prey Area. Rangelands, 8, 217-220.
Korpimaki, E. (1984) Population dynamics of birds of prey in relation to fluctuations in small mammal populations in western Finland. Annales Zooligici Fennici, 21,287-293.
Korpimiki, E. (1987) Clutch size, breeding success and brood size experiments in Tengmalm's owl Aegolius funereus: a test of hypotheses. Ornis Scandinavica, 18, 277- 284.
Korpimaki, E. (1992) Diet composition, prey choice, and breeding success of long-eared owls: effects of multiannual fluctuations in food abundance. Canadian Journal of Zoology, 70, 2373-2381.
Korpimaki, E. & Hakkarainen, H. (1991) Fluctuating food supply affects the clutch size of Tengmalm's owl inde- pendent of laying date. Oecologia, 85, 543-552.
Korpimiki, E. & Hongell, H. (1986) Partial migration as an adaptation to nest-site scarcity and vole cycles in Tengmalm's Owl Aegoliusfunereus. Vdr Figelvdrld, Sup- plement, 11, 85-92.
Korpimaki, E. & Norrdahl, K. (1991) Numerical and func- tional responses of kestrels, short-eared owls, and long- eared owls to vole densities. Ecology, 72, 814-826.
Korpimaki, E., Lagerstr6m, M. & Saurola, P. (1987) Field evidence for nomadism in Tegmalm's Owl Aegolius funereus. Ornis Scandinavica, 18, 1-4.
Kostrzewa, A. (1989) The effect of weather on density and reproduction success in honey buzzards Pernis apivorus. Raptors in the Modern World (eds B. -U. Meyburg & R. D. Chancellor), pp. 187-191. World Working Group on Birds of Prey, Berlin, London & Paris.
Kostrzewa, R. & Kostrzewa, A. (1990) The relationship of spring and summer weather with density and breeding performance of the buzzard (Buteo buteo), goshawk
361 K. Steenhof, M.N. Kochert & T.L. McDonald
? 1997 British Ecological Society Journal of Animal Ecology, 66, 350-362

(Accipiter gentilis) and kestrel (Falco tinnunculus). Ibis, 132, 550-559.
Kostrzewa, R. & Kostrzewa, A. (1991) Winter weather, spring and summer density, and subsequent breeding suc- cess of Eurasian kestrels, common buzzards, and northern goshawks. Auk, 108, 342-347.
Marti, C.D. & Wagner, P.W. (1985) Winter mortality in common barn-owls and its effect on population density and reproduction. Condor, 87, 111-115.
Mosher, J.A. & White, C.M. (1976) Directional exposure of golden eagle nests. Canadian Field-Naturalist, 90, 356-359.
Newton, I. (1979) Population Ecology of Raptors. Buteo Books, Vermilion, South Dakota.
Newton, I. & Marquiss, M. (1982) Fidelity to breeding area and mate in sparrowhawks Accipiter nisus. Journal of Ani- mal Ecology, 54, 241-253.
Phillips, R.L., Wheeler, A.H., Lockhart, J.M., McEneaney, T.P. & Forrester, N.C. (1990) Nesting ecology of golden eagles and other raptors in southeastern Montana and north- ern Wyoming. US Fish and Wildlife Service Technical Report 26, Washington, DC.
Ridpath, M.G. & Brooker, M.G. (1986) The breeding of the wedge-tailed eagle (Aquila audax) in relation to its food supply in arid western Australia. Ibis, 128, 177-194.
Smith, D.G. & Murphy, J.R. (1979) Breeding responses of raptors to jackrabbit density in the eastern Great Basin desert of Utah. Raptor Research, 13, 1-14.
Smith, D.G., Murphy, J.R. & Woffinden, N.D. (1981) Relationships between jackrabbit abundance and fer- ruginous hawk reproduction. Condor, 83, 52-56.
Smith, G.W. (1987) Mortality and movement within a black- tailed jackrabbit population. PhD thesis, Utah State Uni- versity, Logan.
Smith, G.W. & Nydegger, N.C. (1985) A spotlight, line- transect method for surveying jack rabbits. Journal of Wildlife Management, 49, 699-702.
Southern, H.N. (1970) The natural control of a population of tawny owls (Strix aluco). Journal of Zoology, London, 162, 197-285.
Squires, J.R., Anderson, S.H. & Oakleaf, R. (1991) Prairie falcons quit nesting in response to spring snowstorm. Jour- nal of Field Ornithology, 62, 191-194.
Stalmaster, M.V. & Gessaman, J.A. (1984) Ecological ener- getics and foraging behavior of overwintering bald eagles. Ecological Monographs, 54, 407-428.
Statistical Sciences (1995) S-PLUS Guide to Statistical & Mathematical Analysis, Version 3.3, StatSci, a division of MathSoft, Inc., Seattle, WA.
Steenhof, K. (1987) Assessing raptor reproductive success and productivity. Raptor Management Techniques Manual (eds B. A. Giron Pendleton, B. A. Millsap, K. W. Cline &
D. M. Bird), pp. 157-170. National Wildlife Federation, Washington, DC.
Steenhof, K. & Kochert, M.N. (1982) An evaluation of methods used to estimate raptor nesting success. Journal of Wildlife Management, 46, 885-893.
Steenhof, K. & Kochert, M.N. (1988) Dietary responses of three raptor species to changing prey densities in a natural environment. Journal of Animal Ecology, 57, 37-48.
Steenhof, K., Kochert, M.N. & Moritsch, M.Q. (1984) Dis- persal and migration of southwestern Idaho raptors. Jour- nal of Field Ornithology, 55, 357-368.
Swenson, J.E., Alt, K.L. & Eng, R.L. (1986) Ecology of bald eagles in the greater Yellowstone ecosystem. Wildlife Monographs, 95, 46 pp.
Taylor, I. (1994) Barn owls: Predator-Prey Relationships and Conservation. Cambridge University Press, Cambridge.
Tjernberg, M. (1983) Prey abundance and reproductive suc- cess of the golden eagle Aquila chrysaetos in Sweden. Hol- arctic Ecology, 6, 17-23.
Tomback, D.F. & Murphy, J.R. (1981) Food deprivation and temperature regulation in nestling ferruginous hawks. Wilson Bulletin, 93, 92-97.
US Department of the Interior (1979) Snake River Birds of Prey Special Research Report. Bureau of Land Manage- ment, Boise, Idaho.
Village, A. (1981) The diet and breeding of long-eared owls in relation to vole numbers. Bird Study, 28, 215-224.
Village, A. (1986) Breeding performance of kestrels at Esk- dalemuir, south Scotland. Journal of Zoology, London (A), 208, 367-378.
Watson, A. (1957) The breeding success of golden eagles in the north-east highlands. Scottish Naturalist, 69, 153-169.
Watson, J. & Langslow, D.R. (1989) Can food supply explain variation in nesting density and breeding success amongst golden eagles Aquila chrysaetos? Raptors in the Modern World (eds B.-U. Meyburg & R. D. Chancellor), pp. 181- 186. World Working Group on Birds of Prey, Berlin, Lon- don & Paris.
Weisberg, S. (1985) Applied Linear Regression, 2nd edn. John Wiley & Sons, New York.
Wiens, J.A. (1977) On competition and variable environ- ments. American Scientist, 65, 590-597.
Wiklund, C.G. & Stigh, J. (1986) Breeding density of snowy owls Nyctea scandiaca in relation to food, nest sites, and weather. Ornis Scandinavica, 17, 268-274.
Wilkinson, L. (1989) Systat: the Systemfor Statistics. Systat, Inc. Evanston, IL.
Woffinden, N.D. & Murphy, J.R. (1977) Population dynam- ics of the ferruginous hawk during a prey decline. Great Basin Naturalist, 37, 411-425.
Received 11 December 1995; revision received 18 July 1996
362 Golden eagle reproduction
? 1997 British Ecological Society Journal of Animal Ecology, 66, 350-362


















