
Interaction of conserved signaling pathways during ...
109
Interaction of conserved signaling pathways during cellular development in Sordaria macrospora Dissertation to obtain the degree Doctor Rerum Naturalium (Dr. rer. nat.) at the Faculty of Biology and Biotechnology Ruhr-University Bochum International Graduate School of Biosciences Ruhr-University Bochum Department of General and Molecular Botany submitted by Sarah Schmidt from Bochum, Germany Bochum October 2019 1 st supervisor: Prof. Dr. Ulrich Kück 2 nd supervisor: Prof. Dr. Franz Narberhaus
Transcript of Interaction of conserved signaling pathways during ...

Interaction of conserved signaling pathways during cellular
development in Sordaria macrospora
Dissertation to obtain the degree
Doctor Rerum Naturalium (Dr. rer. nat.)
at the Faculty of Biology and Biotechnology
Ruhr-University Bochum
International Graduate School of Biosciences
Ruhr-University Bochum
Department of General and Molecular Botany
submitted by
Sarah Schmidt
from
Bochum, Germany
Bochum
October 2019
1st supervisor: Prof. Dr. Ulrich Kück
2nd supervisor: Prof. Dr. Franz Narberhaus

Interaktion konservierter Signaltransduktionswege
während der Zellentwicklung von Sordaria macrospora
Dissertation zur Erlangung des Grades
eines Doktors der Naturwissenschaften
der Fakultät für Biologie und Biotechnologie
der Ruhr-Universität Bochum
Internationale Graduiertenschule Biowissenschaften
Ruhr-Universität Bochum
Lehrstuhl für Allgemeine und Molekulare Botanik
vorgelegt von
Sarah Schmidt
aus
Bochum
Bochum
Oktober 2019
Referent: Prof. Dr. Ulrich Kück
Korreferent: Prof. Dr. Franz Narberhaus

Danksagung
Ich danke meinem Doktorvater Herrn Prof. Dr. Ulrich Kück für die attraktive und aktuelle
Themenstellung, die geistige und materielle Unterstützung, die lehrreichen Diskussionen und
sein immerwährendes Interesse an der Fortführung der Arbeit.
Prof. Dr. Franz Narberhaus möchte ich für die freundliche Übernahme des Korreferates
danken.
Mein besonderer Dank gilt PD Dr. Ines Teichert für die großartige Hilfe während meiner
Promotionszeit und das intensive Korrekturlesen dieser Arbeit.
Bei den aktuellen Mitgliedern des Arbeitskreises „Allgemeine und molekulare Botanik“ als auch
allen ehemaligen Mitgliedern des Lehrstuhls bedanke ich mich für das nette Arbeitsklima und
die gute Zusammenarbeit. Bei allen Mitgliedern der Sordaria-Gruppe möchte ich mich
bedanken für viele hilfreiche Diskussionen und die gegenseitige Unterstützung. PD Dr. Minou
Nowrousian danke ich für wertvolle Ratschläge. Ich danke Stephanie Lorenz für das
Autoklavieren von unzähligen Puffern und Medien und Susanne Schlewinski für die technische
Unterstützung bei manchen Experimenten. Für Hilfe bei IT Problemen danke ich Dr. Tim
Dahlmann und Dr. Dominik Terfehr. Tim Dahlmann danke ich zudem für das schnelle
Korrekturlesen dieser Arbeit.
Meinen Doktorschwestern danke ich für die tolle Zeit im und außerhalb des Labors. Ich danke
Ramona Märker, Ines Teichert und Yasaman Mahmoudjanlou für viele schöne und lustige
gemeinsame Pausen. Ramona Märker gilt mein besonderer Dank für eine unvergesslich
schöne und lustige Zeit im Labor und Büro.
Schließlich möchte ich allen meinen Freunden und meiner Familie für die immerwährende
Unterstützung danken.

Index of Contents
Interaction of conserved signaling pathways during cellular
development in Sordaria macrospora
Abbreviations ______________________________________________________ I
List of Tables _____________________________________________________ III
List of Figures _____________________________________________________ III
I. Introduction ______________________________________________________ 1
1. Signal transduction through MAPK cascades in mammals .............................................. 1
1.1 Scaffolds improve signaling performance .................................................................................. 3
1.2 KSR1 – Regulation of scaffold activity ....................................................................................... 5
1.3 MP1 and MORG1 – Specific targeting of MAPK cascade to organelles ................................... 6
2. MAPK cascades in yeast ................................................................................................ 8
2.1 Ste5 – Composition of the signaling cascade is determined by scaffolds ...............................10
3. Summary ...................................................................................................................... 11
II. Scope of the thesis _______________________________________________ 12
III. Materials and methods ___________________________________________ 15
1. Materials ....................................................................................................................... 15
1.1 Strains.......................................................................................................................................15
1.2 Oligonucleotides .......................................................................................................................17
1.3 Plasmids ...................................................................................................................................19
1.4 Chemicals .................................................................................................................................20
1.5 Buffers and solutions ................................................................................................................20
1.6 Media ........................................................................................................................................21
1.7 Kits ............................................................................................................................................21
1.8 Fluorescent dyes ......................................................................................................................22
1.9 Antibodies and enzymes ..........................................................................................................22
1.10 Software and websites ...........................................................................................................22

Index of Contents
2. Methods ........................................................................................................................ 22
2.1 Culture conditions .....................................................................................................................22
2.2 Transformations ........................................................................................................................23
2.3 Polymerase chain reaction (PCR) ............................................................................................23
2.4 DNA gel electrophoresis ...........................................................................................................23
2.5 Isolation of nucleic acids ..........................................................................................................23
2.6 Plasmid construction ................................................................................................................24
2.7 Sequencing of DNA ..................................................................................................................24
2.8 Crosses of S. macrospora ........................................................................................................24
2.9 Growth and stress test..............................................................................................................24
2.10 Quantification of fruiting body formation .................................................................................24
2.11 Ascospore germination assay ................................................................................................24
2.12 Microscopic investigation .......................................................................................................25
2.13 Safety precautions ..................................................................................................................25
IV. Results ________________________________________________________ 26
1. Imaging of organelle and cytoskeletal markers ............................................................. 26
1.1 Organelle markers ....................................................................................................................26
1.2 Cytoskeletal markers ................................................................................................................30
2. Characterization of the PR pathway .............................................................................. 32
2.1 Deletion of genes coding for pheromone response kinases leads to defects in sexual
development, hyphal fusion, and vegetative growth ................................................................33
2.2 Subcellular localization of PR components during vegetative growth and sexual
development .............................................................................................................................37
3. Interaction of the PR pathway with the NADPH oxidase complex ................................. 43
3.1 HAM5 and NOR1 co-localize in older hyphae ..........................................................................43
3.2 Initiation of fruiting body formation is regulated by the PR and NOX complexes.....................46
3.3 The PR kinases and NOX2 complex regulate ascospore germination ....................................48
V. Discussion _____________________________________________________ 52
1. HAM5-dependent interaction of PR cascade and NOX1 complex mediates chemotropic
interaction during hyphal fusion .................................................................................... 53
2. PR regulation of ascospore germination by activation of NOX2 via STE12 is
independent
of HAM5 ....................................................................................................................... 57
3. How is sexual development influenced by PR signaling? .............................................. 60
4. Microtubule organizing centers – signaling hubs for the integration of PR signaling with
other signaling pathways? ............................................................................................ 64

Index of Contents
VI. Summary ______________________________________________________ 70
VII. Zusammenfassung______________________________________________ 71
VIII. References ____________________________________________________ 72
IX. Supplements ___________________________________________________ 90
X. Curriculum Vitae ________________________________________________ 97
XI. Erklärung ______________________________________________________ 99

Abbreviations I
Abbreviations
A.dest aqua destillata
aa amino acids
ATP adenosine tri-phosphate
bp base pairs
ChIP-Seq chromatin immunoprecipitation DNA-sequencing
CWI cell wall integrity
DAPI 4′,6-diamidin-2-phenylindol
DPSS diode pumped solid-state
EGFP enhanced green fluorescent protein
ER endoplasmic reticulum
ERK extracellular signal-regulated kinase
GCK germinal center kinase
GDP guanosine diphosphate
GEF guanine nucleotide exchange factor
GF growth factor
GPCR G protein-coupled receptors
GRB2 growth factor receptor-bound protein 2
GTP guanosine triphosphate
H2A histone 2A
H2O2 hydrogen peroxide
HAM hyphal anastomosis
hph hygromycin B resistance gene
IDP intrinsically disordered protein
IMP impedes mitogenic signal propagation
JNK1/2 Jun N-terminal kinase 1-3
KSR1 kinase suppressor of Ras 1
MAPK mitogen-activated protein kinase
MAPKK MAPK kinase
MAPKKK MAPK kinase kinase
MOR morphogenesis Orb6
MORG1 mitogen-activated protein kinase organizer 1
MP1 MEK Partner 1
mRFP monomeric red fluorescent protein
MTOC microtubule-organizing centers
nat nourseothricin resistance gene

Abbreviations II
NBT nitroblue tetrazolium
NDR nuclear Dbf2p-related
NOR1 NOX regulator 1
NOX NADPH oxidase
O2- superoxide
p38α-δ p38MAPK α, β, γ and δ
PAK p21-activated kinase
PCR polymerase chain reaction
PMA phorbol 12-myristate-13-acetate
PP2A protein phosphatase-2A
PPP protoplast buffer
PR pheromone response
PRE pheromone response elements
RAS rat sarcoma
RPE retinal pigmented epithelial
RTK receptor tyrosine
SHC Src homology 2 containing
SIN septation initiation network
SO SOFT
SOS son of sevenless
SPA septal pore-associated
SPB spindle pole body
STRIPAK striatin-interacting phosphatases and kinases
TF transcription factor
wt wild type
Y2H yeast two-hybrid
Δ deletion of a gene

List of Tables & Figures III
List of Tables
Table 1: S. macrospora strains used in this study. ________________________________ 15
Table 2: Oligonucleotides used in this study. ____________________________________ 17
Table 3: Plasmids used in this study. __________________________________________ 19
Table 4: Software and websites used in this work. _______________________________ 22
Table 5: Frequency of hygromycin B resistance or sterile phenotype (in %) in black- and
brown-spored progeny from indicated crosses. ___________________________ 49
Table S1: Absolute values for ascogonia and protoperithecia formation per 0.5 cm². _____ 95
List of Figures
Figure 1: Activation of the ERK MAPK pathway. __________________________________ 2
Figure 2: Regulation of KSR1 localization and activity. _____________________________ 6
Figure 3: MP1- and MORG1-dependent ERK cascade localization to endosomes. _______ 8
Figure 4 : Pheromone response and filamentous growth pathways in yeast. ___________ 10
Figure 5: Life cycle of S. macrospora. _________________________________________ 13
Figure 6: Localization of nuclei, nuclear pores and spindle pole bodies. _______________ 27
Figure 7: Confocal microscopy of a fluorescent marker for the endoplasmic reticulum. ___ 29
Figure 8: Confocal microscopy of a fluorescent marker for peroxisomes. ______________ 30
Figure 9: Confocal microscopy of a fluorescent marker for actin filaments. _____________ 31
Figure 10: Confocal microscopy of a fluorescent marker for microtubules. _____________ 32
Figure 11: Microscopic investigation of sexual development of Δmak2, Δmek2, Δmik2 and
Δham5. ________________________________________________________ 34
Figure 12: Complementation of PR kinase deletion strains in sexual development and hyphal
fusion. _________________________________________________________ 35
Figure 13: Vegetative growth and response to oxidative stress of PR kinase deletion and
complementation strains. __________________________________________ 36
Figure 14 Localization of PR components in hyphae. _____________________________ 38
Figure 15: MIK2 and MEK2 localize to spindle pole bodies. ________________________ 39
Figure 16: GFP-HAM5 associates with nuclei. ___________________________________ 40
Figure 17: MAK2 accumulates in nuclei of older hyphae. __________________________ 41
Figure 18: PR kinases localize to vacuolar structures in ascogonia. __________________ 42
Figure 19: Spot-like localization of NOR1-GFP is similar to GFP-HAM5. ______________ 44
Figure 20: NOR1 is targeted to hyphal tips and forms spherical aggregates located around
septal pores. ____________________________________________________ 45

List of Tables & Figures IV
Figure 21: NOR1 and HAM5 co-localize in older hyphae. __________________________ 46
Figure 22: Generation of fruiting bodies in PR and NOX deletion strains. ______________ 47
Figure 23: MAK2 accumulation in nuclei of ascospores is not dependent on the NOX2
complex. _______________________________________________________ 50
Figure 24: Germling fusion is regulated by crosstalk of several signaling modules. ______ 54
Figure 25: PR kinases, STE12, and the NOX2 complex act in the same pathway controlling
ascospore germination. ___________________________________________ 59
Figure 26: Model for the initiation of fruiting body formation. ________________________ 61
Figure 27: Steps of fruiting body formation and involved signaling complexes. __________ 62
Figure 28: SPBs and septal pores are MTOCs that may function as signaling hubs. _____ 67
Figure 29: Septal pore-associated proteins recruit several signaling modules to septal
pores. _________________________________________________________ 68
Figure S1: Generation and verification of Δmak2 deletion strains. ___________________ 91
Figure S2: Generation and verification of Δmek2 deletion strains. ___________________ 92
Figure S3: Generation and verification of Δmik2 deletion strains. ____________________ 93
Figure S4: Ascogonial septation in Δmak2, Δmek2, Δmik2 and Δham5. _______________ 94
Figure S5: The deletion strain Δste12 is not impaired in hyphal fusion. ________________ 95

I. Introduction 1
I. Introduction
The ability to react and adapt to the surrounding environment is a necessity for cell survival
in all organisms. Information is perceived by sensing of external stimuli, which can lead to
activation of many different signaling pathways within the cell. These pathways, in turn,
trigger a variety of cellular reactions, including DNA damage response, cell cycle arrest and
progression, differentiation, stress adaptation, and apoptosis (Chang et al. 2003; Rezatabar
et al. 2019). A breakthrough in the investigation of signal transduction was achieved in
Saccharomyces cerevisiae, where Zhou et al. (1993) first identified a mitogen-activated
protein kinase (MAPK) cascade as a signal transduction system. MAPK cascades are, like
many other signaling modules, conserved through all eukaryotes. They are comprised of a
MAPK kinase kinase (MAPKKK), a MAPK kinase (MAPKK) and a MAPK. Activation of these
three-tiered kinase cascades are triggered by specific extracellular cues, leading to
consecutive activation of the kinases by phosphorylation of specific tyrosine/threonine
residues (Payne et al. 1991). The last MAPK will phosphorylate and activate transcription
factors that determine the cellular reaction by induction or repression of gene expression
(Martínez-Soto & Ruiz-Herrera 2017). Through this kinase system, it is possible to multiply
the effect of the initial signal, since every MAPKKK phosphorylates several MAPKKs and
those each activate several MAPK molecules. Intriguingly, a single linear MAPK cascade is
able to regulate many different and sometimes antagonistic processes. This multi-
functionality is enabled by spatio-temporal regulation and fine-tuning of MAPK activity
through a variety of mechanisms, like scaffolding proteins, feedback-loops, and interaction
with other signaling modules. The following paragraphs will provide insight into the impact
of different scaffolding proteins on spatio-temporal regulation of MAPK pathways in
mammals and yeast.
1. Signal transduction through MAPK cascades in mammals
Since the discovery of MAPK cascades in 1993, extensive research has broadened our
understanding of MAPK cascades, including their impact on cellular processes and their
role in diseases. In mammals, MAPK cascades can be distinguished into four distinct
groups, which are named after their MAPKs: extracellular signal-regulated kinase 1 and 2
(ERK1/2), c-Jun N-terminal kinase 1-3 (JNK1-3), p38MAPK α, β, γ and δ (p38α-δ), and
ERK5 (Wortzel & Seger 2011). As mentioned above, all of these cascades are based on a
three-tiered MAPK phosphorylation system and often consist of more than one component
for each level of the cascade. The best-characterized MAPK cascade, which will also be
the focus of this introduction, is the ERK1/2 MAPK cascade. Diverse MAPKKKs can be part

I. Introduction 2
of this pathway, including RAF family members (RAF-1, B-RAF, and A-RAF) as well as
MOS and TLP2. MAPKKs and MAPKs for this pathway are named MEK1/2 and ERK1/2,
respectively (Wortzel & Seger 2011; Morrison 2012). ERK1/2 will further be referred to as
ERK, except indicated otherwise.
The ERK MAPK cascade can be activated by growth factors (GFs), which bind to receptor
tyrosine kinases (RTKs) (Figure 1, left side). Although GFs are also known to activate the
other MAPK cascades, they are the main activators for the ERK pathway, while other
cascades are only partially activated by GFs (Katz et al. 2007). RTKs consist of an
extracellular ligand-binding domain, a single transmembrane domain, and a cytoplasmic
domain. The cytoplasmic domain contains a tyrosine kinase domain as well as several
regulatory tyrosines. Upon binding of GFs, RTKs dimerize and are activated through auto-
or trans-phosphorylation of their regulatory kinases (Meister et al. 2013). The adaptor
protein GRB2 (growth factor receptor-bound protein 2) binds to the activated RTKs by
interaction with the phosphorylated tyrosines, or it is recruited by another adaptor protein
named SHC (Src homology 2 containing). GRB2 associates with the guanine nucleotide
exchange factor (GEF) SOS (son of sevenless), targeting it to the plasma membrane.
There, SOS catalyzes the exchange of GDP for GTP of the small G protein RAS (Rat
sarcoma) (Katz et al. 2007). This exchange triggers conformational changes in the RAS
Figure 1: Activation of the ERK MAPK pathway. The ERK pathway can be activated through ligand binding to receptor tyrosine kinases (RTKs) and G protein-coupled receptors (GPCRs). Left side: RTKs dimerize upon activation by growth factors and phosphorylate tyrosine residues in their cytoplasmic tail. This recruits SHC and GRB2 proteins as well as SOS, which catalyzes the exchange of RAS-bound GDP to GTP, resulting in its activation. Active RAS phosphorylates RAF, which subsequently leads to phosphorylation of MEK and ERK and thus to ERK activation. Right side: The ER cascade can further be activated by GPCRs, which is mediated by activation and dissociation of heterotrimeric G proteins. See text for more information about the indirect activation by GPCRs (indicated with dashed lines). Modified from Dhanasekaran et al. (2007) and Schöneborn et al. (2018).

I. Introduction 3
protein, leading to its activation (Shima et al. 2010). Activated RAS will bind to RAF and
induce conformational changes, leading to phosphorylation and activation of the
serine/threonine kinase activity of RAF and thus to activation of the MAPK pathway (Hibino
et al. 2009).
While activation of ERK through RTKs is typical for this cascade, it can also be activated by
G protein-coupled receptors (GPCRs) (Watson et al. 2018). A huge variety of ligands is able
to initiate signal transduction through GPCRs, i.e. hormones, neurotransmitters,
chemoattractants or calcium ions. The cytoplasmic side of GPCRs is associated with
heterotrimeric G proteins, consisting of Gα, Gβ, and Gγ subunits. Receptor activation
induces the exchange of GDP for GTP on the Gα subunit, promoting its dissociation from
the Gβγ dimer. Both, Gα and Gβγ, can subsequently associate with downstream targets of
the cascade (Figure 1, right side) (Vilardaga et al. 2010). In some cases, activated G
proteins lead to trans-activation of a specific RTK subset, which induced RAS-mediated
ERK activation as described above (Daub et al. 1997). In other cases, RAF is activated by
GPCR signaling via PKC and PI3K activity mediated by the Gα subunit (Van Biesen et al.
1996; Antonelli et al. 2000).
An activated cascade can induce a variety of distinctly different cellular responses,
dependent on cell or tissue type or the context of activation, i.e. proliferation, survival or
motility. As mentioned above, regulation of the signaling outcome is, among other
mechanisms, achieved by spatio-temporal regulation of MAPK activity though scaffold
proteins.
1.1 Scaffolds improve signaling performance
Through recent advances in the study of MAPK signaling, it became clear that the MAPK
cascades do not transmit signals by diffusing freely through the cytoplasm, but rather
increase the interaction of the three kinases by assembly into complexes. This assembly is
mediated by scaffold proteins, which bind, per definition, at least two members of a signaling
cascade (Zeke et al. 2009). In general, scaffolding proteins act by binding of signaling
partners, leading to an increased local concentration and phosphorylation rate of target
proteins. This leads to positive regulation of signal transduction and is necessary for
sufficient signal transmission of otherwise weak signals (Takahashi & Pryciak 2008). The
ability of a scaffold protein to intensify signaling is dependent on its synthesis level. An
increase of scaffold concentration up to the optimal level facilitates signaling by assembling
all kinases at one location. At higher concentrations, however, signal transduction is
attenuated, since an overload of scaffold proteins would lead to the incomplete assembly
of the kinase cascade. This effect of scaffold overproduction is referred to as ‘combinational

I. Introduction 4
inhibition’ (Levchenko et al. 2000). Besides the enhancement of signal initiation, scaffolds
also play a role in extending signal transduction by protection of the kinases against
dephosphorylation. The interaction of phosphatases with active MAP kinases can either be
sterically hindered by the scaffold, or dephosphorylation is minimized since phosphatases
diffuse freely in the cytoplasm, leading to much lower local concentrations than the MAP
kinases (Witzel et al. 2012). Another important aspect is the reduction of crosstalk with other
pathways.
As mentioned above, a given MAPK cascade acts as signal transducer for many different
cellular processes. MAPK cascades gain this ability to transduce different signals through
the insulation of the kinases by different, signal-specific scaffold proteins (Dhanasekaran et
al. 2007). Scaffolds modify signal transduction by regulation of the spatial distribution of
MAPK cascades. There is a huge variety of scaffold proteins, which all operate in a
sublocalization-specific manner. These scaffolds selectively target ERK signaling to i.e. the
plasma membrane (KSR1), endosomes (MP1), or focal adhesions (Paxillin) (Teis et al.
2002; Ishibe et al. 2003). Which pathway is activated depends on the origin of RAS
activation (Casar et al. 2009a). Changes in subcellular localization of RAS will be explained
together with detailed explanations of KSR1 and MP1 in upcoming sections. Interestingly,
one difference between nuclear and scaffold-dependent activity of ERK is the preference
for dimerization. ERK monomers are preferred for interaction with nuclear targets, while
scaffolds utilize ERK dimers. It is suggested that one part of the dimer is associated with
the binding domain of the scaffold, whereas the other part associates with substrates in the
vicinity (Casar et al. 2008). Different studies have shown that cytoplasmic substrates of ERK
bind exclusively to its dimerized form and that this interaction is abolished in dimerization-
deficient ERK mutants (Casar et al. 2008; Casar et al. 2009b).
A very interesting observation is the detrimental effect of changes in production levels of a
single scaffold on overall ERK signaling. Up- or downregulation of synthesis levels of one
scaffold affects ERK signaling on a much higher scale than would be suggested (Casar &
Crespo 2016). Some cases reported a decrease in ERK signal intensity by up to 80% after
decreased protection levels of a single scaffold gene (Nguyen et al. 2002; Lozano et al.
2003; Teis et al. 2006). There are several hypotheses trying to explain this phenomenon.
First, higher concentrations of one scaffold protein might influence the activity of overall
ERK signaling by competition with other scaffolds, since they all regulate the same set of
enzymes and compete for their binding. This would produce fewer scaffolds that are fully
occupied and thus decrease the number of complete and functional signaling complexes.
Second, depletion of a scaffold might produce MAP kinase concentrations that are too high
for the remaining scaffolds leading to combinational inhibition and thus also to incomplete
complexes. Third, different scaffold proteins sometimes interact to form ‘macro’ signaling

I. Introduction 5
platforms and thus mediate crosstalk between different pathways (Pan et al. 2012).
Because of this connection, alternation of expression of one scaffold might also influence
signaling through other pathways.
1.2 KSR1 – Regulation of scaffold activity
As mentioned above, scaffold ‘Kinase suppressor of RAS 1’ (KSR1) is one of the best
characterized MAPK scaffolds. It was initially identified as a modifier of RAS and a positive
regulator of MAPK signaling in Caenorhabditis elegans and Drosophila melanogaster. While
D. melanogaster only has a single ksr gene, which is essential for viability, two KSR genes,
ksr-1 and ksr-2, can be found in C. elegans as well as higher organisms (Zhang et al. 2013;
Frodyma et al. 2017). KSR1 has the ability to bind all three kinases of the ERK cascade
(Figure 2). It continuously associates with MEK, while interaction with RAF and ERK is only
triggered after GF stimulation (Müller et al. 2001). In quiescent cells, phosphorylation of
serine residues 297 and 392 of KSR1 by different kinases (C-TAK1 or Nm23) enables
binding of 14-3-3 proteins, which leads to retention of the KSR1-MEK complex in the
cytoplasm (Kolch 2005). Similarly, 14-3-3 proteins also bind to RAF to keep it in the
cytoplasm (Figure 2, left side). In this inactive state, the N-terminal domain of RAF acts as
an auto-inhibitor for its kinase domain, which is located at the C-terminus. Phosphorylated
sites on both N- and C-terminus promote the binding to 14-3-3 proteins and thus stabilize
this autoinhibitory effect (McKay & Morrison 2007).
The regulation of MAPK cascades by 14-3-3 proteins is not restricted to mammalian cells. In the fission yeast Schizosaccharomyces pombe, the 14-3-3 protein homologs Rad24 and Rad25 interact with N- and C-terminal domains of the RAF homolog Byr2. Similar to mammalian cells, this association results in retention of Byr2 in the cytoplasm (Ozoe et al. 2002).
Stimulation by mitogens leads to dephosphorylation of serine residues on KSR1 and RAF
by protein phosphatase-2A (PP2A), resulting in dissociation from 14-3-3 proteins. RAF is
then recruited to the plasma membrane by active GTP-bound RAS. Release of KSR1 from
14-3-3 proteins exposes its docking site for ERK as well as the atypical C1 domain, which
promotes localization to the plasma membrane by interaction with phospholipids (McKay &
Morrison 2007; Koveal et al. 2012). At the plasma membrane, the KSR1-MEK-ERK complex
comes into contact with activated RAF-RAS, initiating ERK signaling (Figure 2, right side).
Besides regulation by 14-3-3 proteins, the inactive KSR1-MEK complex is also sequestered
in a triton-insoluble compartment by interaction with a protein named ‘impedes mitogenic
signal propagation’ (IMP). This isolation of KSR1 insulates it from other effectors until
activated RAS binds to IMP and promotes its autoubiquitination, leading to release of the
KSR1-MEK complex (Matheny et al. 2004; Ory & Morrison 2004).

I. Introduction 6
When the ERK complex is active, KSR1 and RAF can act as substrates of activated ERK.
Each contains four serine/threonine sites that can be phosphorylated by ERK, resulting in
interruption of the interaction of KSR1 and RAF and subsequent dissociation of KSR from
the plasma membrane. Thus, activated ERK regulates a negative feedback loop for KSR1
mediated activation (McKay et al. 2009).
1.3 MP1 and MORG1 – Specific targeting of MAPK cascade to organelles
Scaffolds do not only regulate the activity of the cascade but can also target the signaling
complex to different subcellular localizations by mediating the interaction with specific
cellular organelles. Targeting of the ERK cascade to endosomes by the scaffold MP1 will
be discussed as an example for such a regulation.
Initially, cell signaling was thought to be regulated exclusively from the plasma membrane
(McKay & Morrison 2007). However, it was shown that endocytosis regulates receptor
signaling for both, RTKs and GPCRs. The receptors are internalized by endocytosis upon
ligand binding (Sorkin & von Zastrow 2009).
Endocytosis of receptors is mediated by lysine 63-linked polyubiquitylation. This ubiquitinylation pattern does not result in association with the proteasome, which would lead to degradation, but instead facilitates the interaction of the receptors with
Figure 2: Regulation of KSR1 localization and activity. Left side: In quiescent cells, KSR only binds MEK. KSR1 and RAF are kept in the cytoplasm by interaction of 14-3-3 proteins with phosphorylated serine residues. Right side: Stimulation by mitogens activates RTK signaling via RAS-dependent activation of RAF. Mitogen stimulation also triggers the phosphatase PP2A, which dephosphorylates serine residues on KSR1 and RAF. Binding of both by 14-3-3 is disrupted upon phosphorylation. Unbound RAF then associates with activated RAS at the plasma membrane. Simultaneously, KSR1 is also recruited to the plasma membrane, where it binds RAF and ERK and facilitates ERK signaling. Modified from Kolch (2005) and McKay & Morrison (2007).

I. Introduction 7
the protein machinery involved in endocytic trafficking. This method of marking receptors for endocytosis is conserved from yeast to mammals (Sorkin & von Zastrow 2009).
Several RTKs remain ligand-bound after endocytosis and stay activated until late stages of
endocytic trafficking (Sorkin & von Zastrow 2009). RAS proteins, which are also bound to
the plasma membrane, are endocytosed together with the receptors. Thus, the active
receptor can rapidly induce the activation of the RAS protein by recruitment of the adaptor
proteins GRB2 and SOS from the cytoplasm (McKay & Morrison 2007; Taguchi & Misaki
2011). GTP bound RAS then recruits RAF to the endosomal membrane and induces its
phosphorylation. The interaction of RAF with the other two MAP kinases, MEK and ERK, is
not mediated by KSR1 since this scaffold exclusively localizes to the cytoplasm and plasma
membrane. A different scaffold named MEK Partner 1 (MP1) is specialized in ERK cascade
signal transduction at endosomes. MP1 preferably binds to MEK1 and ERK1 and targets
these kinases to endosomes by associating with the adaptor protein p14, which is located
at endosomal membranes (Teis et al. 2006). This interaction brings the MP1-MEK-ERK
complex into close proximity of the already activated RAS-RAF complex, leading to initiation
of signal transduction of the ERK cascade (Figure 3, left side) (Witzel et al. 2012).
The targeting of the ERK cascade to endosomes by MP1 is not restricted to RTK signaling
as it also occurs after the activation of GPCRs. When activated by GPCRs, the MAP kinase
cascade is organized by the ‘superscaffold’ ‘mitogen-activated protein kinase organizer 1’
(MORG1), which has the ability to bind to all of the kinases as well as the scaffold MP1.
Localization to endosomes is then mediated by MP1 interaction with p14 at the endosomal
membrane (Figure 3, right side). The mechanism of MORG1 activation by GPCRs is not
yet know; however, it is evident that MORG1 is dispensable for RTK activation of the ERK
cascade. It was shown that GPCRs can also be internalized via endocytosis; however, this
process has only been linked to desensitization of to GPCR signaling (Ferguson 2001). The
activity of MORG1 can be induced by several specific stimuli like lysophatidic acid, phorbol
12-myristate-13-acetate (PMA) or serum (Meister et al. 2013).
Knockout studies in mice and Drosophila addressed the functions of the MP1 scaffold
in vivo. Conditional p14 knockout mice showed impaired proliferation and differentiation of
the epidermis during embryonal development (Teis et al. 2006). An impact on differentiation
can also be observed in Drosophila, where a tightly regulated expression of
MP1 is essential for the correct differentiation and placement of vein cells during wing
development (Mouchel-Vielh et al. 2008). Thus, important processes like cellular
differentiation are dependent on correct spatial regulation of ERK signaling.

I. Introduction 8
2. MAPK cascades in yeast
This section will focus on S. cerevisiae, although this study utilizes a filamentous fungi as
model organism since MAPK regulations by scaffolds are well-investigated in this unicellular
fungus. Studies in this fungus led to the discovery of not only MAP kinases as signal
transduction components, but were also first to describe the function of a scaffold protein in
the regulation of MAPK signaling (Choi et al. 1994; Zeke et al. 2009). Fungi provide
excellent model organisms for the investigation of major signaling pathways as these often
show high conservation between species of the eukaryotic kingdom. Moreover, regulation
of signaling pathways is not as complex as in mammalian cells, enabling straightforward
exploration of connections within or between signaling pathways.
Five different MAPK cascades have been described in yeast. MAPKs of the pheromone
response and filamentous growth pathways show strong homology to MAPKs of the ERK1/2
pathway in mammals (Widmann et al. 1999). MAPKs of the high osmolarity and cell integrity
pathways are homologous to MAPKs of p38 and ERK5 pathways, respectively (Westfall et
al. 2008; Kim et al. 2010). The spore wall assembly pathway is only conserved in some
Figure 3: MP1- and MORG1-dependent ERK cascade localization to endosomes. Left side: MAPK pathway targeting to endosomes is initiated by endocytosis of the activated RTK together with the membrane-associated RAS protein. Active RTK signaling triggers GTP exchange and thus activation of RAS, which subsequently targets RAF to the endosomal membrane. MEK and ERK1 are bound by the scaffold protein MP1. Association of MP1 with the adaptor protein p14 brings the kinases into close proximity and enables signaling. Right side: Endosome-specific signaling of ERK1 cascades can also be initiated by GPCRs, which activate the superscaffold MORG1. MORG1 is able to bind all three kinases as well as MP1, which is already located at the endosomal membrane by interaction with p14. Modified from Kolch (2005) and McKay & Morrison (2007).

I. Introduction 9
other ascomycete fungi (Hamel et al. 2012). This section will further focus on the pheromone
response (PR) and the filamentous growth (FG) pathway.
The PR pathway is, as the name suggests, responsible for pheromone response and mating
(Figure 4, left side). Haploid cells of S. cerevisiae carry either genes for mating type a or α,
which induce production of pheromone peptides, named a- and α-factor, respectively.
These pheromone peptides are able to bind GPCRs at the yeast cell wall of cells carrying
the opposite mating type. During this process, the a-factor acts as a ligand for a receptor
named Ste3 and the α-factor binds the receptor Ste2, which both activate the same
heterotrimeric G protein. Upon activation by pheromone binding, the Gα (Gpa1) subunit
exchanges the bound GDP for GTP and dissociates from the Gβ (Ste4) and Gγ (Ste18)
subunits. When released to the cytoplasm, the Gβ subunit directly interacts with
downstream components of the mating signaling pathway.
It has been hypothesized that the Rac protein Cdc42 and its GEF Cdc14 are involved in transduction of the pheromone signal to the MAPK cascade; however, it remains controversial if they function in the direct relay or in modification of the signal (Heinrich et al. 2007).
One of these components is the activated p21-activated kinase (PAK)-like kinase Ste20,
which acts as a MAPKKK kinase. The Gβ-Ste20 complex is brought into contact with the
MAPK cascade at the plasma membrane by the scaffold protein Ste5, which can bind to all
three kinases of the cascade as well as the Gβ subunit (Whiteway et al. 1995). Ste20
phosphorylates the MAPKKK Ste11, which then relays the phosphorylation to the MAPKK
Ste7 and subsequently to the MAPK Fus3. Phosphorylated Fus3 dissociates from Ste5 and
translocates to the nucleus, where it phosphorylates the inhibitors of the transcription factor
(TF) Ste12. Subsequently, Ste12 binds to ‘pheromone response elements’ (PRE) on the
DNA and induces the transcription of mating-specific genes (Chou et al. 2006).
The FG pathway, which also consists of the MAPKKK Ste11 and the MAPKK Ste7, utilizes
Kss1 as MAPK. The Kss1 pathway is scaffold-independent and is activated by limited
nutrient concentration in the environment and by membrane receptors different from
ste2/ste3 (Figure 4, right side). The aspartyl protease Yps1 is expressed upon starvation,
which locates at the plasma membrane and processes the extracellular domain of the
signaling mucin Msb2. The processed part of Msb2 can activate the receptor Sho1, which
leads to binding and activation of Cdc24. Subsequently, Cdc24 is brought into close
proximity of Msb2-associated Cdc42, leading to exchange of GDP for GTP. GTP-bound
Cdc42 then activates Ste20, which initiates the FG response by phosphorylating Ste11,
similar to the PR pathway. Phosphorylation of either Fus3 or Kss1 by Ste7 is determined by
activity of the scaffold Ste5. The following section gives insight into the regulation of this
process.

I. Introduction 10
2.1 Ste5 – Composition of the signaling cascade is determined by scaffolds
It is remarkable that different signaling outcomes can be achieved mainly through the
involvement of the scaffold protein Ste5, which is specific for the pheromone response
pathway. Ste5 is able to enhance PR signaling by enhancing the local concentration of the
Ste11-Ste7-Fus3 cascade at the plasma membrane. Interestingly, it has been shown that
Ste5 not only brings the kinases into close proximity, but is also indispensable for Ste7-
mediated phosphorylation of Fus3. In vitro experiments have shown that Fus3 is an
intrinsically poor substrate of Ste7, in contrast to Kss1, which is a good substrate. However,
mutation of the Fus3-binding site of Ste5 did not abolish mating signaling, indicating that
Fus3 phosphorylation is regulated by a different mechanism than simple complex formation.
Ste5 also contains an independently folding domain termed Ste5-minimal scaffold (Ste5-
ms). This domain binds strongly to Ste7 and is able to catalytically unlock Fus3 for
phosphorylation. While this modification increases the Ste7-mediated Fus3 phosphorylation
Figure 4 : Pheromone response and filamentous growth pathways in yeast. Left side: Pheromone response (PR) pathway signaling is initiated by binding of pheromones to Ste2 or Ste3 receptors at the plasma membrane. These GPCRs trigger activation of a heterotrimeric G protein and dissociation of the α-subunit Gpa1 from G protein subunits Ste4 and Ste18. Ste4 promotes activity of the MAPKKKK Ste20, leading to subsequent phosphorylation of the MAPKKK Ste11, MAPKK Ste7 and MAPK Fus3. The MAPK cascade is insulated by the scaffold Ste5 and targeted to the plasma membrane by Ste5-Ste4 interaction. Right side: The filamentous growth pathway comprises the same set of signaling molecules as the PR pathway but utilizes the MAPK Kss1 instead of Fus3 and lacks the scaffold Ste5. It is activated by limited nutrient concentrations, and signal transmission to Ste20 is mediated via Cdc24 and Cdc42. See text for further information.

I. Introduction 11
by ~5000 fold, it has no effect on the phosphorylation activity of Ste7 to Kss1. Thus,
selective activation of the pheromone response pathway is achieved by Ste5-mediated
regulation of the catalytic activity of Ste7 (Good et al. 2009).
The mechanism of Ste5 is highly similar to mammalian KSR1, despite having no amino acid
sequence similarity (Witzel et al. 2012). Both scaffolds are able to bind all three kinases and
enhance their local concentration at the plasma membrane. Like ERK-mediated negative
feedback on KSR1, Ste5 is also negatively regulated by the MAPK Fus3 during mating
response (Bhattacharyya et al. 2006). Furthermore, a similar mechanism as the effect of
Ste5 in the Fus3-directed catalytic activity of Ste7 was found in mammalian cells, where
KSR2 has the ability to phosphorylate MEK in vitro, leading to a conformational change that
promotes MEK activation by RAF (Brennan et al. 2011). This similarity between yeast and
mammalian scaffold mechanisms confirms fungi as excellent model organism. Further
investigation of Ste5 and other scaffolds in yeast might provide important hints for scaffold
regulations in mammals.
3. Summary
MAPK signaling cascades belong to the major signal transduction systems that are
conserved through all eukaryotes. The transduction of signals through this pathway is tightly
regulated by scaffold proteins. These scaffolds can modify the activity of MAP kinases in a
space-, time- and dose-dependent manner. This regulation, along with the huge variety of
different scaffold proteins, is crucial to generate the multipurpose signaling mediated by
MAPK cascades. Therefore, investigation of scaffolds is very important to understand the
overall regulation of MAPK cascade signaling. For this purpose, fungi have been proven to
function as good model organisms since scaffold regulation in fungi shows high similarity
mammalians, even without sequence homology. New insights into signaling mechanisms
of MAPK cascades and their scaffold proteins might provide new therapeutic targets for a
large variety of diseases associated with dysregulation of MAPK signaling.

II. Scope of the thesis 12
II. Scope of the thesis
The use of fungi as model organisms for the investigation of highly conserved cell signaling
mechanisms does not only include unicellular yeast, like Saccharomyces cerevisiae or
Schizosaccharomyces pombe, but also involves filamentous fungi. While research on
cellular mechanisms is easier in yeast, filamentous fungi provide means to study the
differentiation of more complex structures like fruiting bodies or conidiophores (Kück et al.
2009). Several filamentous fungi have already been established as model organisms, i.e.,
Aspergillus nidulans, Neurospora crassa, Podospora anserina and Sordaria macrospora
(Golduran & Morris 1995; Davis & Perkins 2002; Scheckhuber & Osiewacz 2008; Kück et
al. 2009; Bennett & Turgeon 2016). Each of these model organisms has its own advantages
and disadvantages for experimental research.
One of the major advantages of the fungus that is used in this study, S. macrospora, is the
absence of an asexual reproduction program, which facilitates the investigation of fruiting
body development during sexual reproduction. In contrast to heterothallic fungi like
N. crassa, which need a mating partner for reproduction, the homothallic S. macrospora is
able to reproduce via selfing. While a mating partner is not required for completion of the
life cycle, crossing with other strains is still possible. Recombinant perithecia can easily be
identified by crossing of spore color mutants against wild type strains, resulting in two spore
colors within one ascus (Teichert et al. 2014a). Thus, recombinant progeny of two strains
can be obtained by simple crossing experiments. Another advantage is the establishment
of tools for microscopic investigation. A broad range of fluorescent proteins was
successfully used for localization experiments, including EGFP, mRFP1, and mCherry.
Furthermore, several markers for the labeling of different organelles have already been
established (Engh et al. 2010).
The life cycle of S. macrospora is completed within seven days under laboratory conditions,
beginning with the germination of an ascospore (Figure 5). The outgrowing hyphae form an
expanding mycelium, which starts to generate small hyphal coils named ascogonia after
three days. These ascogonia represent the initial stage of fruiting body development. They
become surrounded by vegetative hyphae, producing small spherical structures named
protoperithecia. Differentiation into pear-shaped mature fruiting bodies, also named
perithecia, takes place after melanization of the outer peridial layer. After seven days, newly
formed ascospores are discharged through the ostiole, an opening at the tip of the
perithecium (Kück et al. 2009; Engh et al. 2010; Lord & Read 2011).
To investigate fruiting body differentiation, mutants with developmental defects were
generated by UV and chemical mutagenesis. Mutants were categorized as asc, pro and per
and stopp differentiation at the stage of ascogonia, protoperithecia, and immature

II. Scope of the thesis 13
perithecia, respectively (Teichert et al. 2014a). Several signaling complexes have been
found to be involved in the generation of fruiting bodies. Interestingly, mutants of some
highly conserved signaling complexes all arrest development at the protoperithecial stage,
indicating that further development is mediated by crosstalk of these signaling modules.
Among them are the NADPH oxidase 1 (NOX1) complex, the striatin-interacting
phosphatases and kinases (STRIPAK) complex as well as a MAPK cascade named the cell
wall integrity (CWI) complex (Bloemendal et al. 2010; Bloemendal et al. 2012; Dirschnabel
et al. 2014; Teichert et al. 2014b; Nordzieke et al. 2015; Kück et al. 2016).
In A. nidulans, N. crassa and P. anserina, a second MAPK cascade is involved in fruiting
body development, which is homologous to the pheromone response pathway in
S. cerevisiae (Li et al. 2005; Bayram et al. 2012; Lalucque et al. 2012). This pathway was
found to be regulated by a scaffold protein named HAM-5 in N. crassa, which has, however,
no sequence homology to the scaffold Ste5 from S. cerevisiae (Jonkers et al. 2014).
Intriguingly, yeast two-hybrid (Y2H) experiments conducted by our group have confirmed
HAM5 association with the PR kinases in S. macrospora and indicated an interaction
between HAM5 and NOR1, which is the regulator of NADPH oxidase complexes 1 and 2
(Beier, personal communication).
Crosstalk between the PR and NOX complex has already been suggested based on studies
conducted in other filamentous fungi. In P. anserina, mutants of the NOX2 complex as well
as the PR kinase pathway are impaired in the germination of melanized ascospores
(Lalucque et al. 2012). A similar germination defect was already observed in S. macrospora
for strains lacking components of the NOX2 complex. A connection of the PR cascade to
the NOX1 complex was found in Claviceps purpurea, where deletion of a PR MAPK
homolog named mk1 reduced expression levels of the nox1 gene (Rolke & Tudzynski
2008). Functional cooperation between RAS and NOX was also suggested for mammalian
Figure 5: Life cycle of S. macrospora. The life cycle starts with germination of an ascospore, leading to formation of a mycelium. Ascogonial coils are formed as initial stage of fruiting body formation after 2-3 days, which are enveloped by sterile hyphae to produce spherical protoperithecia. After melanization of the outer layer, protoperithecia differentiate into mature fruiting bodies (perithecia) containing several asci with new ascospores after 7 days. Spores are ejected through an opening at the tip of the peer-shaped perithecium. Figure from Teichert et al (2014a).

II. Scope of the thesis 14
cells, since both show a similar subcellular distribution (membrane rafts, endosomes, and
endomembranes) (Wu & Terada 2009). Several studies suggested that mammalian NOX1
might act downstream of RAS during RAS-dependent tumor formation (Suh et al. 1999;
Mitsushita et al. 2004; Lim et al. 2005; Laurent et al. 2008).
This study pursues three major objectives. The first objective was the establishment of a
collection of strains expressing organelle markers and construction of a new marker for
spindle pole bodies (SPBs). Distribution of organelles was assessed in different parts of the
mycelium and during different developmental stages using conventional and confocal
microscopy. The second objective was the characterization of deletion strains of
components of the PR pathway, as well as the investigation of their subcellular localization
by means of fluorescence microscopy. The third objective was the analysis of crosstalk
between the PR cascade and both NOX complexes in S. macrospora. For this, the
localization of NOR1, as well as NOR1-HAM5 co-localization, was examined to verify the
interaction between these two proteins. The generation of fruiting bodies was quantified in
all PR and NOX deletion strains to investigate a connection of these pathways during the
initiation of sexual development. Furthermore, deletion strains of PR kinases and HAM5
were assayed for a defect in ascospore germination. The results will contribute to the
general understanding of signal transduction networks during cellular differentiation in fungi
and provide cues for potential connections in mammalian cells.

III. Materials and methods 15
III. Materials and methods
1. Materials
1.1 Strains
Escherichia coli E. coli strain XL1 Blue MRF’ was used for plasmid cloning. XL1 Blue MRF’ ∆(mcrA)183∆(mcrCB-hsdSMR-mrr)173, recA1, endA1, gyrA96, thi-
1, hsdR17, supE44, relA1, lac (Jerpseth et al. 1992) Sordaria macrospora All S. macrospora strains used in this work are listed in Table 1.
Table 1: S. macrospora strains used in this study.
Strain Relevant genotype and phenotype Source / Reference
R19027 Wild type, fertile Culture collection
S70823 Spore color mutant, fertile
fus1-1
Culture collection
DD194 ASI, fertile
Δnox2::hph, fus1-1
Dirschnalbel et al (2014)
DD574 ASI, sterile
Δnor1::hph, fus1-1
Dirschnabel et al (2014)
RM1941 ASI, ectopic integration of pGG-N-EGFP-HAM5 into RM329, fertile
Δham5::hph, gpd(p)::egfp::ham5::trpC(t), trpC(p)::nat
Märker (2019)
S143932 ASI, sterile
Δmek2::hph, fus1-1
This work
S147281 ASI, sterile
Δham5::hph, fus1-1
Märker (2019)
S156605 ASI, sterile
Δham5::hph, Δnor1::hph, fus1-1
Märker (2019)
S68567 ASI, fertile
Δste12::hph, fus1-1
Nolting & Pöggeler (2006)
S96888 Strain for homologous recombination, fertile
Δku70::nat
Culture collection
SaS1090 ASI, sterile
Δmik2::hph, fus1-1
This work
SaS1097 ASI, ectopic integration of pH2A-mRFP into R19027, fertile
gpd(p)::h2a::mrfp1::trpC(t), trpC(p)::nat
This work
SaS1298 ASI, sterile
Δmak2, fus1-1
This work
III. Materials and methods 16
Strain Relevant genotype and phenotype Source / Reference
SaS1339 ASI, ectopic integration of pgfp-mik2 into SaS1090, fertile
∆mik2::hph, gpd(p)::egfp::mik2::trpC(t), trpC(p)::nat, fus1-1
This work
SaS1669 ASI, ectopic integration of pgfp-mak2 into SaS1298, fertile
∆mak2, gpd(p)::egfp::mak2::trpC(t), trpC(p)::nat, fus1-1
This work
SaS1787 ASI, ectopic integration of pgfp-mek2 into S143932, fertile
∆mek2::hph, gpd(p)::egfp::mek2::trpC(t), trpC(p)::nat, fus1-1
This work
SaS1821 ASI, ectopic integration of pnor1-gfp into DD574, fertile
∆nor1::hph, gpd(p)::nor1::egfp::trpC(t), trpC(p)::nat, fus1-1
This work
SaS1938 ASI from crossing of SaS1821 and SaS1097, fertile
Δnor1::hph, gpd(p)::nor1::egfp::trpC(t), trpC(p)::nat, gpd(p)::h2a::mrfp::trpC(t), trpC(p)::nat, fus1-1
This work
SaS2308 ASI from crossing of RM1941 and SaS1097, fertile
Δham5::hph, gpd(p)::egfp::ham5::trpC(t), trpC(p)::nat, gpd(p)::h2a::mrfp::trpC(t), trpC(p)::nat
This work
SaS2372 ASI, ectopic integration of pGG-mRFP-Grc1 and pTub4-mCherry into R19027, fertile
gpd(p)::mrfp::grc-1::trpC(t), trpC(p)::nat, gpd(p)::tub4::mcherry::trpC(t), trpC(p)::hph
This work
SaS2451 ASI from crossing of SaS2372 and S70823, fertile
gpd(p)::mrfp::grc-1::trpC(t), trpC(p)::nat, gpd(p)::tub4::mcherry::trpC(t), trpC(p)::hph, fus1-1
This work
SaS2485 ASI, ectopic integration of pDsRed-SKL into R19027, fertile
gpd(p)::dsredskl::trpC(t), trpC(p)::nat
This work
SaS2538 ASI from crossing of SaS1821 and SaS2451, fertile
Δnor1::hph, gpd(p)::nor1::egfp::trpC(t), trpC(p)::nat, gpd(p)::mrfp::grc-1::trpC(t), trpC(p)::nat, gpd(p)::tub4::mcherry::trpC(t), trpC(p)::hph, fus1-1
This work
SaS2558 ASI, ectopic integration of pLA-GFP into R19027, fertile
gpd(p)::lifeact::gfp::trpC(t), trpC(p)::nat
This work
SaS886 ASI, ectopic integration of pmRFP-KDEL into R19027, fertile
gpd(p)::Sppg1::mrfp::kdel::trpC(t), trpC(p)::nat
This work
SaS926 ASI, ectopic integration of pmRFP-SON1 into R19027, fertile
gpd(p)::mrfp::son1::trpC(t), trpC(p)::nat
This work

III. Materials and methods 17
Strain Relevant genotype and phenotype Source / Reference
TSA132D1 Primary transformant, ectopic integration of pMF309 and pABnat into R19027, fertile
ccg1(p)::sgfp::bml, gpd(p)::trpC(t), trpC(p)::nat
This work
TSA133D2 Primary transformant, ectopic integration of pgfp-mak2 and pH2A-mRFP into R19027, fertile
gpd(p)::h2a::mrfp1::trpC(t), trpC(p)::nat
gpd(p)::egfp::mak2::trpC(t), trpC(p)::nat
This work
TSA149A3 Primary transformant, ectopic integration of pGG-N-EGFP-HAM5 and pNor1-mCherry into S156605, fertile
Δham5::hph, Δnor1::hph, gpd(p)::nor1::mcherry::trpC(t), trpC(p)::nat, gpd(p)::egfp::ham5::trpC(t), trpC(p)::nat, fus1-1
This work
TSA152D2 Primary transformant, ectopic integration of pgfp-mak2 into DD194, fertile
∆nox2::hph, gpd(p)::egfp::mak2::trpC(t), trpC(p)::nat
This work
TSA153D3 Primary transformant, ectopic integration of ptub4-mcherry and pGG-N-EGFP-HAM5 into R19027, fertile
gpd(p)::tub4::mcherry::trpC(t), trpC(p)::hph, gpd(p)::egfp::ham5::trpC(t), trpC(p)::nat
This work
ASI: ascospore isolate, hph: hygromycin B resistance gene from E. coli, Sppg1: signal sequence of ppg1 gene for co-translational insertion into the endoplasmic reticulum, nat: nourseothricin resistance gene from Streptomyces noursei, gpd(p): constitutive gpd promoter from Aspergillus nidulans , trpC(t): trpC terminator from A. nidulans.
1.2 Oligonucleotides
Oligonucleotides used for PCR and sequencing in this study are listed in Table 2. Table 2: Oligonucleotides used in this study.
Oligonucleotide Sequence (5’-3’) Specificity
mik2_5_nhe agagctagcggggttttccatggtcgcttgcgac mik2 5‘ flank forward
mik2_5_hin aagcttggttgaagcgacggtcagct mik2 5‘ flank reverse
mik2_3_bam tctggatccgatcgttcaatatcatgagg mik2 3‘ flank forward
mik2_3_sna cgttacgtacgggagataagtcccatgtgt mik2 3‘ flank reverse
KO_5356_1 gcaggagctcctcctccagttttc upstream of mik2 5‘ flank forward
KO_5356_2 ggaaagcgccgatatttaaagcaatc downstream of mik2 3‘ flank reverse
KO_5356_3 acgactgggaacccaacgagg mik2 reverse
mek2_5_fw atcggatccagaattcaacttttgagaagtgacagaa mek2 5‘ flank forward
mek2_5_rv ccgggaaccagttgaggtgtaggtttaagcgtaat mek2 5‘ flank reverse
mek2_3_fw ctacgactggctctcagc mek2 3‘ flank forward
mek2_3_rv gcttgtcgacgaattcggtaagggggtgacgcgg mek2 3‘ flank reverse

III. Materials and methods 18
Oligonucleotide Sequence (5’-3’) Specificity
MEK2KO1 ctggttgtctccgttcccaaaatc upstream of mek2 5‘ flank forward
MEK2KO2 ctggtggatcatgaacatacg downstream of mek2 3‘ flank reverse
hph1MN cgatggctgtgtagaagtactcgc hph reverse
hph2MN atccgcctggacgactaaaccaa hph forward
MAK2-KO1-KpnI ggtacccacaatattgccctcgaaacg mak2 5‘ flank forward
MAK2-KO2-SnaBI tacgtatttggcgtgtccctgaggg mak2 5‘ flank reverse
MAK2-KO3-HindIII
aagcttacgtctacttgcatacaagctgtggg mak2 3‘ flank forward
MAK2-KO4-BglII agatctgtcttgccttgccttgctgg mak2 3‘ flank reverse
Hph_split_3’_fw ttggcgacctcgtattgggaatc hph forward
Hph_split_5’_rv cgttgcaagacctgcctgaaacc hph split reverse
MAK2KO1 ctcctgtttattcctccatcagct upstream of mak2 5‘ flank forward
MAK2KO2 cctcctcgagagcgaacacatcat downstream of mak2 5‘ flank reverse
mRFP1-FW atccatggcctcctccgagg mrfp forward
mRFP-KDEL ggatccttagagctcgtccttggcgccggtggagtggcg mrfp reverse with KDEL sequence
GG_01693_for1 tcactcggtctcgtggtatgggatcatcagcaacc grc1 forward
GG_01693_rev1 tcactcggtctcgtacgcgtcatgaccccccacca grc1 reverse, bsaI mutation
GG_01693_for2 tcactcggtctcggttcctcgtaggggttgta grc1 forward, bsaI mutation
GG_01693_rev2 tcactcggtctcggaacgcgaccgcttgtcaggg grc1 reverse
SaS_01693_for ttgtaccggggaacatgacg forward sequencing grc1
SaS_01693_rev aaaactcggaatgtgggtcg reverse sequencing grc1
GG-N-mRFP-fw gctagcatggcctcctccgaggacg mRFP forward
NotI-tub4-for gctcagcggccgcatgcccaggtacgcgtcc tub4 forward
EcoRI-tub4-rev cctggaattcacaagccatccgtttgtccgtg tub4 reverse
1751 gccatattttcctgctctcc gpd(p) forward
1757 agctgacatcgacaccaacg trpC(t) reverse
GG-nor1-rev
tctctcggtctccatcccgatatctcctggacccag
nor1 reverse
GG-nor1-for
tctctcggtctcctggtatgtcgctaaaacaggtca
nor1 forward
02124 comp 5'rv ctatcatccttgacttccagtttcc nor1 reverse
02124 comp 3'fw gtgctgaaatcaaaaaatgttagtcttg
nor1 forward
EcoRV-mCherry-for
gggtccaggagatatcgggatctggctctggtatgg
mcherry forward
III. Materials and methods 19
1.3 Plasmids
All plasmids used for S. macrospora transformations are listed in Table 3. Table 3: Plasmids used in this study.
Plasmid Relevant features Reference
pDS23 Cloning vector gpd(p)::egfp::trpC(t) in pRSnat, URA3, bla, nat
Schindler & Nowrousian (2014)
pH2A-mRFP gpd(p)::h2a::mRFP::trpc(t), URA3, bla, nat
Teichert
(pers. communication)
pmRFP-KDEL gpd(p)::Sppg1::mrfp::kdel::trpC(t), cloned NcoI-BamHI in pEGFP-KDEL, bla, nat
This work
pGG-N-mRFP Plasmid for GoldenGate cloning,
gpd(p)::mRFP::trpC(t), bla, nat
Teichert
(pers. communication)
pGG-mRFP-Grc1
gpd(p)::grc1::mrfp::trpC(t) in pGG-N-mRFP1, bla, nat
This work
pCherry gpd(p)::mcherry::trpC(t), bla, hph Engh et al. (2010)
ptub4-mCherry gpd(p)::tub4::mcherry::trpC(t) in pCherry, bla, hph
This work
pMF309 ccg1(p)::sgfp::bml, bla Freitag et al. (2004)
pABnat gpd(p)::trpC(t), bla, nat Beier
(pers. communication)
pLA-GFP gpd(p)::lifeact::gfp::trpC(t), trpC(p)::nat Lutomski (2015)
pGG-mrfp-son1 gpd(p)::mrfp::son1::trpC(t), bla, nat Beier
(pers. communication)
pDsRedSKL gpd(p)::dsredskl::trpC(t)::nat Elleuche & Pöggeler (2008)
pEGFP-mik2 gpd(p)::egfp::mik2::trpC(t) in pGG-N-EGFP, bla, nat
Ebbing (2017)
pEGFP-mek2 gpd(p)::egfp::mek2::trpC(t) in pDS23, URA3, bla, nat
Beier
(pers. communication)
pEGFP-mak2 gpd(p)::egfp::mak2::trpC(t) in pDS23, URA3, bla, nat
Beier
(pers. communication)
pGG-N-EGFP-HAM5
gpd(p)::egfp::ham5::trpC(t) in pGG-N-EGFP, bla, nat
Märker (2019)
pGG-C-EGFP Plasmid for GoldenGate cloning,
gpd(p)::egfp::trpC(t), bla, nat
Teichert
(pers. communication)
pGG-Nor1-gfp gpd(p)::nor1::egfp::trpC(t) in pGG-C-EGFP, bla, nat
This work
pNor1-mCherry gpd(p)::nor1::mcherry::trpC(t) in pGG-Nor1-gfp, bla, nat
This work
pFlip5-MAK2 Deletion plasmid for SMAC_03492
smxyl(p)::Pcflp, trpC(p)::hph
Teichert
(pers. communication)
pFlip3-MAK2 Deletion plasmid for SMAC_03492
smxyl(p)::Pcflp, trpC(p)::hph
Teichert
(pers. communication)

III. Materials and methods 20
Plasmid Relevant features Reference
pKO-mek2 Deletion plasmid for SMAC_06526
5’ flank mek2, trpC(p)::hph,
3’ flank mek2, bla, kanR
Beier
(pers. communication)
pKO-mik2 Deletion plasmid for SMAC_05356
5’ flank mik2, trpC(p)::hph,
3’ flank mik2, ura3, bla
Beier
(pers. communication)
Sppg1: signal sequence of ppg1 gene for co-translational insertion into the ER, bla: ampicillin resistance gene, hph: hygromycin B resistance gene from E. coli, kanR: kanamycin resistance gene from Streptomyces kanamyceticus, nat: nourseothricin resistance gene from Streptomyces noursei, gpd(p): constitutive gpd promoter from Aspergillus nidulans, trpC(t): trpC terminator from A. nidulans
1.4 Chemicals
Agar-agar (AppliChem, Serva), LE agarose (Biozym), ampicillin (Applichem), ammonium chloride (Baker), ammonium iron(II) sulfate hexahydrate (Riedel-de Haën), biotin (AppliChem), boric acid (Roth), blue dextran (Sigma-Aldrich), boric acid (AppliChem), calcium chloride (Riedel-de Haën), CFW = Calcofluor White (Sigma-Aldrich), chloroform (Normapur), citric acid monohydrate (Roth), cornmeal (Husener Mühle), copper sulfate pentahydrate (Baker), dipotassium hydrogen phosphate trihydrate (Merck), disodium hydrogen phosphate (Merck), dNTPs = 2´-Desoxyribonucleosid-5´-triphosphate (Roth), EDTA = ethylene diamine tetraacetic acid (Merck), ethanol (Sigma-Aldrich), ethidium bromide (Boehringer), glucose (Applichem), hydrochloric acid (Baker), hydrogen peroxide (Sigma Aldrich), hygromycin B (CalBiochem), IPTG = isopropyl β-D-1-thiogalactopyranoside (Applichem), iron(II) chloride (Riedel-de Haën), isoamylalcohol (Riedel-de Haën), isopropyl alcohol (Normapur), L-arginine (AppliChem), magnesium chloride (Baker), magnesium sulfate (Riedel-de Haën), manganese(II) chloride (Applichem), manganese(II) sulfate (Merck), malt (CSM Deutschland GmbH), nourseothricin (Werner BioAgents), PEG 6000 = polyethylene glycol 6000 (AppliChem), phenol (AppliChem), potassium chloride (Baker), potassium hydroxide (Roth), potassium dihydrogen phosphate (Normapur), potassium nitrate (Roth), sodium acetate (Baker), sodium chloride (Baker), sodium dihydrogen phosphate (Baker), sodium hydrogen phosphate (Baker), sodium hydroxide (Sigma Aldrich), SDS = sodium dodecyl sulfate (AppliChem), Sodium molybdate (Applichem), soluble starch (Difco), sorbitol (AcrosOrganics), sucrose (AppliChem), Tris = Tri-(hydroxymethyl)-aminomethane (Sigma-Aldrich), tryptone (Roth, Difco), urea (AppliChem), X -Gal = 5-Chlor-4-brom-3-indolyl-ß-D-galactopyranoside (Applichem), yeast extract (Difco), zinc sulfate (Riedel-de Haën), zinc sulfate heptahydrate (Riedel-de Haën). Further chemicals used in this study were purchased in analytical grade from Amersham Biosciences, AppliChem, Baker, BioRad, Difco, GE Healthcare, Life Technologies, Merck Millipore, Molecular Probes, Perkin-Elmer, Promega, Qiagen, Riedel-de Haën, Roche, Roth, Serva, Sigma-Aldrich, Thermo Scientific, or VWR International.
1.5 Buffers and solutions
Protoplast buffer (PPP) 13 mM Na2HPO4 x 2 H2O, 45 mM KH2PO4, 600 mM KCl, pH 6.0
Trace elements solution 1 1 % (w/v) ZnSO4, 1 % (w/v) Fe(II)Cl2, 1 % (w/v) MnCl2
Trace elements solution 2 5 % (w/v) citric acid x H2O, 5 % (w/v) ZnSO4 x 7 H2O, 1 % (w/v) Fe(NH4)2(SO4) x 6 H2O, 0.25 % (w/v) CuSO4

III. Materials and methods 21
x 5 H2O, 0.05 % (w/v) MnSO4, 0.05 % (w/v) H3BO3, 0.05 % (w/v) Na2MoO4
TBE 100 mM Tris, 100 mM boric acid, 2 mM EDTA, pH 8.3
TE(H) 50 mM Tris, 20 mM EDTA, pH 8.0
Neutralization buffer 3M NaAc pH 5.5, 5 M NaCl,
Lysis buffer (E. coli) 0.4 M NaOH, 2% SDS
Lysis buffer (S. macrospora) 10 mM Tris, 2.5 mM EDTA, 50 mM NaCl, 0.2% (w/v) SDS, pH 8.0
Westergaard’s solution 0.5 % (w/v) KNO3, 0.5 % (w/v) KH2PO4, 0.25 % (w/v) MgSO4 x 7 H2O, 0.05 % (w/v) NaCl, 0.05 % (w/v) CaCl2, 0.05 % (v/v) trace elements solution 2
1.6 Media
After preparation, media were sterilized by autoclaving at 121 °C for 30 min.
E. coli
LB 1 % (w/v) tryptone (Roth), 0.5 % (w/v) yeast extract, 0.5 % (w/v) NaCl, pH 7.2; solid medium contained 1.5 % (w/v) agar-agar (AppliChem); 100 μg/ml ampicillin was added for antibiotic selection, 0.004% X-Gal and 0.2 mM IPTG were used for blue-white screening
SOC 2 % (w/v) tryptone (Difco), 0.5 % (w/v) yeast extract (Difco), 10 mM NaCl, 2.5 mM KCl, 10 mM MgSO4, 10 mM MgCl2, 20 mM glucose, pH 7.5
1.7 Kits
E.Z.N.A.® Plasmid Mini Kit I (Omega), NucleoBond® AX500 Kit (Macherey & Nagel), NucleoSpin® Gel and PCR Clean-up kit (Macherey & Nagel), SureClean Plus (Bioline), CloneJet PCR Cloning Kit (ThermoFisher).
S. macrospora
BMM 0.8 % (w/v) malt in filtered cornmeal extract, pH 6.4-6.6, solid medium contained 1.5 % (w/v) agar-agar (AppliChem); 80 U/ml hygromycin B and/or 50 μg/ml nourseothricin were added for selection
BMM-NaAc BMM with 0.5 % (w/v) NaAc, pH 6.4-6.6
CM 1 % (w/v) glucose, 0.2 % (w/v) yeast extract, 0.2 % (w/v) tryptone (Roth), 0.05 % (w/v) KCl, 0.15 % (w/v) KH2PO4, 0.05 % (w/v) MgSO4, 0.37 % (w/v) NH4Cl, 0.1 % (v/v) trace elements solution 1, pH 6.4-6.6
CMS CM with 0.8 M sucrose, solid medium contained 2 % (w/v) agar-agar (AppliChem)
Cornmeal extract
2.5 % (w/v) cornmeal in tap water incubated over night at 60 °C
MMS 1.8 mM KH2PO4, 1.7 mM K2HPO4 x 3 H2O, 8.3 mM urea, 1 mM MgSO4 x 7 H2O, 0.01 % (v/v) trace elements solution 2, 0.1 % soluble starch, 5 µM biotin, pH 6.6-6.8; solid medium contained 1.5 % (w/v) agar-agar (Rech et al. 2007)
Prep agar 6 % (w/v) agar-agar (Serva) in A. dest.
SWG 1x Westergaard‘s, 0.1 % (w/v) L-arginine, 2 % (w/v) glucose, 4 % (w/v) soluble starch, 415 nM biotin, pH 6.5, solid medium contained 1.5 % agar-agar
Top agar 0.8 M NaCl, 0.8 % (w/v) agar-agar (Serva); 400 U/ml hygromycin B and/or 216 μg/ml nourseothricin were added to sterile medium for selection

III. Materials and methods 22
1.8 Fluorescent dyes
MitoTracker® Orange CMTMRos (Invitrogen), DAPI = 4′,6-Diamidin-2-phenylindol (Serva), CFW = Calcofluor White (Sigma-Aldrich)
1.9 Antibodies and enzymes
Chitinase (ASA Spezialenzyme), GoTaq® polymerase (Promega), Phusion® polymerase (Thermo Scientific), T4 DNA ligase (Roche), restriction endonucleases (Thermo Scientific, Roche, New England Biolabs), rShrimp Alkaline Phosphatase (Affymetrix), RNase A (Thermo Scientific), VinoTaste Pro (Novozymes).
1.10 Software and websites
Software and website that were applied in this study are listed in Table 4.
Table 4: Software and websites used in this work.
Software Purpose Source / Reference
Adobe Illustrator CS6 Figure design Adobe Creative Suite 6
Adobe Photoshop CS6 Image processing Adobe Creative Suite 6
Artemis Release 13.1 Genome browser Rutherford et al. (2000)
Biostep ArgusX1 Documentation of agarose gels
Biostep
Blast Local alignments Altschul et al. (1990)
ClustalW Execution of multiple sequence alignments
Larkin et al. (2007)
ImageJ Image processing Collins (2007)
LasAF Confocal microscopy Leica
LasX Processing of confocal images
Leica
Mendeley Reference manager Lo Russo et al. (2013)
MetaMorph 7.7 Fluorescence and light microscopy
Universal imaging
Microsoft Office Data and text processing Microsoft
OligoCalc 3.27 Oligonucleotide design Kibbe (2007)
SnapGene 3.3.4 In silico cloning, sequence analysis
GSL Biotech
2. Methods
All standard molecular genetic methods not described in this section were performed according to Sambrook and Russell (2001).
2.1 Culture conditions
E. coli E. coli strains were cultured in solid or liquid LB medium overnight at 37°C. Liquid cultures were placed on a shaking device at 250 rpm.
S. macrospora S. macrospora strains were grown at 27°C on solid BMM containing the antibiotics nourseothricin or hygromycin for selection. Isolated ascospores were incubated at 27°C on solid BMM-NaAc medium overnight for germination.

III. Materials and methods 23
2.2 Transformations
E. coli Electrocompetent E. coli strains were transformed with plasmid DNA by electroporation with a Multiporator system (Eppendorf) according to the method described by Dower et al. (1988). Cells were plated on solid ampicillin-containing LB medium overnight at 37°C.
S. macrospora Protoplasts were generated from 3-day old CM liquid cultures by incubation with 1.3 g VinoTaste Pro (Novozymes) and 27 U chitinase (ASA Spezialenzyme) resulting in enzymatic breakdown of the fungal cell wall (Walz & Kück 1995; Engh et al. 2007). Afterward, protoplasts were transformed with 10–20 μg plasmid DNA using PEG and subsequently incubated on solid CMS plates for one day. Plates were covered with Top-Agar containing either nourseothricin (216 μg/ml) or hygromycin B (400 U/ml) for selection, and positive transformants were isolated after two to three days.
2.3 Polymerase chain reaction (PCR)
Polymerase chain reaction (PCR) was performed to obtain selectively amplified DNA fragments as described by Mullis et al. (1996). PCRs were conducted in “GeneAmp PCR system 9700” (Applied Biosystems) or “MyCyclerTM Thermal Cycler” (BioRad). The reaction mixture contained either 1-5 ng plasmid DNA or 100-200 ng genomic DNA as template. For verification of gene integration, PCR mixtures contained 1 U GoTaq® DNA polymerase (Promega), 1 x GoTaq® buffer, 0.2 mM of each dNTP and 0.5 μM oligonucleotides in 25 μl total volume. For amplification of fragments used for cloning, Phusion® polymerase (Thermo Scientific) was used in a total volume of 50 μl containing 1 U polymerase, 1 x Phusion HF or GC buffer, 0.2 mM of each dNTP and 0.5 μM oligonucleotides. PCR programs were modified for oligonucleotide melting temperatures and amplicon length and conducted according to manufacturers’ protocols.
2.4 DNA gel electrophoresis
Horizontal gel electrophoresis was applied to separate DNA using a gel containing 1 % agarose dissolved in 0.5x TBE buffer and run in a Mupid-One gel chamber (Nippon Genetics). DNA fragments were purified using the Sure Clean Plus Kit (Bioline). For sizing and quantification of DNA on gels, GeneRuler™ DNA Ladder Mix (Thermo Scientific) was used as a standard. NucleoSpin® Gel and PCR Clean-up Kit (Macherey & Nagel) was used when the cloning procedure required the elution of DNA fragments.
2.5 Isolation of nucleic acids
E. coli For isolation of plasmids, single colonies of the transformed E. coli strains were inoculated in 5 ml liquid LB medium. DNA was isolated using alkaline lysis following the protocol of Birnboim and Dolly (1979). Sequencing was performed on high-quality plasmid DNA obtained with E.Z.N.A.® Plasmid Mini Kit 1 (Omega). Isolation of larger quantities of plasmid DNA was achieved by extraction from 200 ml cultures using the Nucleobond PC500 Kit (Machery-Nagel).
S. macrospora Nucleic acids were isolated from S. macrospora strains that were grown for 3 days in liquid BMM medium in three petri dishes per strain (Nowrousian et al. 1999). After addition of lysis buffer, mycelium was subjected to three cycles of 1 min vortexing, 30 sec freezing in liquid nitrogen and 1 min thawing in a 70°C water bath to disrupt the cell walls. Genomic DNA was isolated by phenol-chloroform extraction. After precipitation, DNA was solubilized in A. dest.

III. Materials and methods 24
2.6 Plasmid construction
Plasmids were cloned by restriction and ligation of PCR or restriction fragments into vector backbones using T4 DNA ligase. Alternatively, plasmids were constructed using BsaI-mediated Golden Gate cloning as described by Marillonnet & Werner (2015).
2.7 Sequencing of DNA
Sequencing of plasmid DNA was performed by Eurofins Genomics (Ebersberg, Germany) or the Department for Biochemistry (Ruhr-University Bochum). Sequencing results were analyzed with SnapGene.
2.8 Crosses of S. macrospora
For crossing of two S. macrospora strains, a strain with wild type spore color was crossed against a strain carrying the spore color mutation fus1-1. After cultivation on BMM for 7-9 days, recombinant perithecia containing spores with two different colors in a single ascus were isolated and ascospores from these perithecia were inoculated on BMM-NaAc for germination. Afterward, germinated spores were tested for the required resistance or phenotype to ensure successful recombination.
2.9 Growth and stress test
Growth and stress tests were conducted three times for each observed strain using three technical replicates in each experiment. After pre-culturing the strains on BMM for 4 days, a standardized piece of the pre-culture was used to inoculate SWG plates containing 30 ml SWG. Mycelial growth was measured after 24 and 48 hours. SWG medium contained 0.001% (v/v) H2O2 for oxidative stress assays.
2.10 Quantification of fruiting body formation
The initiation of fruiting body development was quantified after three days of incubation on BMM-covered microscope slides. Ascogonia and protoperithecia were counted separately within an area of 0.5 cm2 located 1 cm behind the growth front. For assessment of fruiting body formation, pictures of strains were taken after seven days of incubation in petri dishes on solid BMM with a Zeiss Stemi 2000-C binocular, using an AxioCam ERc5s with the software Zen 2 core (version 2.5, Zeiss).
2.11 Ascospore germination assay
Since several deletion strains analyzed in this thesis could only be recovered in a fus1-1 mutant background, an ascospore germination assay was performed to quantitatively assess the germination capacity of black ascospores carrying distinct gene deletions. This assay was conducted by crossing deletion strains carrying the fus1-1 mutation against wild type strain. After 11 days, black and brown ascospores were isolated from recombinant perithecia and recovered on BMM-NaAc. The experiment was repeated until at least 100 ascospores of each spore color germinated. For strains with a generally decreased germination rate, only 50 germinated ascospores of each color were collected per strain. Germinated ascospores were tested for hygromycin resistance and ascospores from crosses of marker-free strains for sterility or fertility.

III. Materials and methods 25
2.12 Microscopic investigation
Microscopic investigation of strains was conducted using either conventional or confocal light microscopy.
Conventional light and fluorescence microscopy The Axioimager.M1 microscope (Zeiss) with Metamorph 7.7 software was used for conventional microscopy. For fluorescent images, a SPECTRA X 6 LCR SA LED lamp (Lumencor) was used for excitation of fluorophores. For detection of DAPI and CFW stainings, Chroma filter set 31000v2 (excitation filter D350/50, emission filter D460/50, beam splitter 400dclp) was used. EGFP was detected using the filter set 49002 (excitation filter HQ470/40, emission filter HQ525/50, beamsplitter T495LPXR), and mRFP, mCherry, DsRed as well as MitoTracker were detected with the filter set 49008 (excitation filter HQ560/40, emission filter ET630/75m, beamsplitter T585lp). Images were taken with a Photometrix Cool SnapHQ camera (Roper Scientific). Images were converted with Metamorph software and processed using Adobe Photoshop. For microscopic investigation of sexual development and fluorescent imaging, strains were inoculated on BMM-covered slides and mounted with a coverslip using 0.7% NaCl solution. For assessment of sexual development, strains were grown for three days (ascogonia, unpigmented protoperithecia), five days (pigmented protoperithecia) and seven days (perithecia). Fluorescent imaging of hyphae was conducted after two days of incubation. Nuclei were stained using DAPI (Serva) in a concentration of 5 mg/ml diluted in 0.7% NaCl solution. MitoTracker (0.5 μM in 0.7% NaCl solution; Invitrogen) was used for visualization of mitochondria. Ascogonial septation was assessed by staining of the cell wall using CFW (Sigma Aldrich).The 1 μg/ml CFW stock solution was diluted 1:400 in a 0.7% NaCl solution. Hyphal fusion events were investigated by observation of strains grown on a layer of cellophane on top of solid MMS medium for two days. For localization studies in ascospores, strains were inoculated 7-10 days on solid BMM. Mature perithecia were collected and opened on a glass slide. Obtained spores were suspended in 0.7% NaCl solution. Micro dishes (Cat.No:80136, Ibidi) were used for a minimum of stress exposure. The bottom of the micro dishes was covered with a thin layer of BMM and inoculated strains were grown for 1 day. Confocal microscopy Confocal microscopy was carried out using a Leica TCS SP5 II microscope (Leica DMI6000 B) with Leica Application Suite Advanced Fluorescence software (LAS AF 2.6). All images were taken after growth of strains for two days on BMM-covered slides as described above. Fluorescence was detected via fluorophore excitation using a 488 nm (EGFP) argon laser or 561 nm (mRFP, mCherry, DsRed, MitoTracker) diode pumped solid-state (DPSS) laser. Images of DAPI-stained hyphae were taken at 405 nm excitation using a 405 diode laser. Images were converted with Metamorph 7.7 (AxioImager) or LasX (Leica) and further processed with Photoshop CS6 and ImageJ.
2.13 Safety precautions
All experiments regarding genetic engineering of security level 1 (S1/B1) were conform to the ‘Richtlinen des Genetikgesetzes’ (GenTG) from 07/01/1990 including the last changes of 1 G from 07/22/2017.

IV. Results 26
IV. Results
Fruiting body development during sexual reproduction in filamentous fungi can serve as an
excellent model for investigation of cell differentiation. Previous studies indicated that the
PR pathway is involved in sexual reproduction in N. crassa and P. anserina (Li et al. 2005;
Lalucque et al. 2012) but proof in S. macrospora is still lacking. The aim of this work was
the characterization and localization of the PR pathway as well as the scaffold protein
HAM5, which was first described in N. crassa. Previous Y2H experiments already confirmed
the interaction of HAM5 with the PR kinases MAK2 and MEK2 in
S. macrospora and indicated a connection between HAM5 and NOR1, a component of the
NOX complexes (Ebbing 2017). Therefore, part of this study focuses on the investigation of
genetic and physical interactions between the PR pathway and the NOX1 and NOX2
complexes. Examination of the PR pathway and its association with the NOX complexes
was primarily performed by fluorescence imaging. For this reason, the first part of the results
section will cover the imaging analysis of several fluorescent organelle markers during
different developmental stages.
1. Imaging of organelle and cytoskeletal markers
Subcellular localizations of proteins can be determined via co-localization with different
organelle markers. There are several methods for visualization of organelles. In filamentous
fungi, organelle markers are often generated by genetically fusing fluorescent tags to the
proteins of interest. Proteins used for organelle markers are usually core components of
specific cellular structures or localize exclusively to specific organelles. A different approach
is the utilization of short signal sequences that mediate the insertion and/or retention of the
fluorophore into specific cellular compartment. Organelles can also be visualized by
application of different stains (Hickey & Read 2009). This section describes the construction
of fluorescent organelle marker strains and their investigation during different
developmental stages of S. macrospora using conventional and confocal microscopy. Some
of the constructs for fluorescent organelle markers were generated in this study, whereas
others had already been established.
1.1 Organelle markers
Specific labeling of nuclei was achieved by tagging the protein sequence of the histone H2A
with the fluorophore mRFP (Teichert per. communication). Microscopic investigation of
strain SaS1097 indicated that nuclei were evenly distributed throughout hyphae and sexual
structures (Figure 6A). An exception to this distribution was the absence of nuclei

IV. Results 27
near hyphal tips as already described by Teichert (2017). For co-localization of nuclei with
other red-fluorescent markers, nuclei were stained with DAPI (Figure 6B+C). DAPI exhibits
a blue fluorescence after binding to AT-rich regions of double-stranded DNA (Kapuscinski
1995).
Figure 6: Localization of nuclei, nuclear pores and spindle pole bodies. (A) Strain SaS1097 contains nuclei labeled with the marker H2A-mRFP. Nuclei show an even distribution in hyphae, ascogonia and protoperithecia, but are absent from the very hyphal tip. (B) Counterstaining with DAPI shows the punctate localization of the nuclear pore marker mRFP-SON1 around the nuclei (SaS926). (C) Images of strain SaS2372 carrying the SPB marker consisting of mRFP-GRC1 and TUB4-mCherry. Nuclear association of SPBs is assessed by counterstaining with DAPI. Images were obtained by confocal (A-B) and epifluorescence (C) microscopy. All scale bars indicate 10 μm. BF: Bright-field; DIC: Differential interference contrast

IV. Results 28
For visualization of nuclear pores, the nucleoporin SON1, which is a component of the
nuclear pore complex, was N-terminally tagged with mRFP (Beier, pers. communication).
Nucleoporins were already successfully used for visualization of nuclear pores in mammals
and fungi (Kubitscheck et al. 1996; Steinberg et al. 2012). Images of ascospore isolate
SaS926, which carries the mRFP-SON1 fusion protein, were taken using confocal
microscopy (Figure 6B). Counterstaining of nuclei with DAPI confirmed localization of the
nuclear pore marker around nuclei. However, although nuclear pores are visible as rough
spots around nuclei, higher resolution would be necessary for co-localization experiments,
since individual pores are not distinguishable.
A new marker for visualization of spindle pole bodies (SPBs) in S. macrospora was
constructed in this study. This marker is based on ZtGrc1 from Zymospetoria tritici (Schuster
et al. 2015), a protein localized at the γ-tubulin ring complex at the minus-end of
microtubules. For generation of a marker for S. macrospora, the ZtGrc1 homolog GRC1
was tagged N-terminally with mRFP. Primary transformants containing this construct
exhibited weak fluorescent signals at SPBs, insufficient for co-localization experiments.
Commonly, ascospore isolates of positive transformants were used for microscopic
investigation in this study since these showed a stronger fluorescent signal, which is less
prone to photobleaching. However, ascospore isolates of mRFP-GRC1-expressing primary
transformants did not contain the desired construct. A possible explanation for this can be
found from data concerning the homolog of grc1 in S.cerevisiae, named spc97, which leads
to lethality when overexpressed. As this lethality can be avoided by co-overexpression of
tub4 (γ-tubulin) (Knop et al. 1997), a second marker was generated, consisting of
S. macrospora TUB4 tagged C-terminally with mCherry. Ascospores were successfully
isolated from primary transformants overexpressing both, GRC1-mRFP and TUB4-
mCherry, resulting in strains with a stable red fluorescent signal (SaS2372). SPBs were
detected as spots in the cytoplasm, and counterstaining with DAPI confirmed the
association with nuclei (Figure 6C). SPBs belong to the microtubule-organizing centers,
which regulate microtubule assembly. Both, GRC1 and TUB4, are core components of
these complexes (Wu & Akhmanova 2017). Since it is debated whether fungi contain
microtubule-organizing centers other than SPBs (Zhang et al. 2017), hyphae were
investigated for other subcellular localizations of GRC1 and TUB4. However, other locations
could not be identified.
A plasmid for endoplasmic reticulum (ER)-localized DsRed has already been described by
Nowrousian et al. (2007). This construct contains an N-terminal signal sequence for co-
translational insertion into the ER-derived from PRO41 and the ER retention amino acid
sequence KDEL at the C-terminus (Nowrousian et al. 2007). Since the monomers of the
tetrameric DsRed protein can exhibit green fluorescence (Baird et al. 2000), it may possibly

IV. Results 29
interfere with co-localization experiments featuring GFP-tagged proteins. To prevent this,
DsRed was replaced with the monomeric mRFP protein in this construct. Fluorescence of
the ER marker was examined in the ascospore isolate SaS886, containing the mRFP-KDEL
construct. Confocal images show a net-like distribution of the endoplasmic reticulum in
hyphae and sexual structures (Figure 7). Besides enhanced localization to small filamental
structures in older hyphae, no changes in ER distribution were observed during different
developmental stages.
Peroxisomes were labeled using DsRed-SKL, which was described in Elleuche & Pöggeler
(2008). In this construct, DsRed is targeted to peroxisomes by a C-terminal SKL tripeptide,
which is a peroxisomal targeting sequence 1 signal. Expression of DsRed-SKL in strain
SaS2485 produces bright fluorescence of small spots in the cytoplasm, which are less
abundant near the hyphal tip and otherwise evenly distributed in hyphae and sexual
structures (Figure 8).
Figure 7: Confocal microscopy of a fluorescent marker for the endoplasmic reticulum. Microscopic investigation of strain SaS886, carrying the KDEL-mRFP construct, shows the distribution of the endoplasmic reticulum at hyphal tips and hyphae as well as ascogonia and protoperithecia. All scale bars indicate 10 μm. BF: Bright-field.

IV. Results 30
The constructs H2A-mRFP, mRFP-SON1, GRC1-mRFP/TUB4-mCherry, mRFP-KDEL
and DsRed-SKL were suitable organelle markers for labeling of the nucleus, nuclear
pore, spindle pole body, ER and peroxisomes, respectively, and enabled
investigations by confocal microscopy. Distribution of these organelles did not
change significantly in different developmental stages.
1.2 Cytoskeletal markers
In addition to the organelle marker strains described in 1.1, strains carrying cytoskeletal
markers for actin filaments and microtubules were generated. These strains could serve for
investigation of directed protein transport since PR pathway components were shown to
undergo a directed transport in A. nidulans (Bayram et al. 2012).
Actin was visualized with a fusion construct of GFP and LifeAct constructed by Lutomski
(2015). LifeAct is a 17-amino-acid peptide from S. cerevisiae, which has the ability to stain
filamentous actin and was generated for actin labeling in mammalian cells (Riedl et al.
2008). In S. macrospora, LifeAct exhibits a bright fluorescence at distinct locations (strain
SaS2558). Figure 9 shows LifeAct-labeling of Actin filaments and patches during different
developmental stages. Images are derived from Z-stacks of the indicated compartments.
Figure 8: Confocal microscopy of a fluorescent marker for peroxisomes. The strain SaS2485 carrying the peroxisome marker DsRed-SKL shows a dot-like fluorescence throughout the whole cytoplasm, which is less abundant near hyphal tips. Distribution of peroxisomes does not change in hyphae, ascogonia or protoperithecia. All scale bars indicate 10 μm. BF: Bright-field.

IV. Results 31
The upper part of the figure shows localization of actin in the middle focal plane of the Z-
stack, while the maximum projection shows the most intense signal of all planes and gives
an indication about the 3D structure of actin filaments. At hyphal tips, LifeAct is visible in a
distinct spot, representing the Spitzenkörper (Riquelme et al. 2018). Enhanced fluorescence
in subapical regions indicates sites with endocytic activity. Association of actin with the
plasma membrane can also be observed farther from the hyphal tip, with strong
fluorescence at the membrane on both sides of the hypha. The maximum projection shows
that this signal expands in a ring-shaped structure around the hyphal membrane, a
phenomenon that can be observed during septum formation. In ascogonia, actin forms
small spots (actin patches) that associate with the plasma membrane. This association is
more prominent at the tip of ascogonia. Localization of actin at newly forming septa is also
visible in maximum projections of ascogonia and protoperithecia.
Strain TSA132D1, carrying β-tubulin labeled with GFP, was generated to assess the
distribution of microtubules (Figure 10). The plasmid used for transformation was described
in Freitag et al. (2004) and contains the bml gene from N. crassa expressed from the ccg1
promoter. Microtubules labeled with β-tubulin-GFP showed pronounced
Figure 9: Confocal microscopy of a fluorescent marker for actin filaments. (A) LifeAct-GFP shows actin localization at the Spitzenkörper and in association with the plasma membrane at subapical regions in the strain SaS2558. Ring-like localization at new forming septa can be observed in hyphae as well as sexual structures. Actin patches are visible in ascogonia and are most prominent at the tip. Images are derived from Z-stack. The figure shows the middle plane and maximum projection of the stack. Scale bars indicate 10 μm. BF: Bright-field.

IV. Results 32
longitudinal orientation along the plasma membrane in subapical regions. In more
peripheral compartments of hyphae and in sexual structures, microtubules appeared in non-
linear organization.
Labeling of Actin with LifeAct-GFP showed actin accumulation at the Spitzenkörper,
near the plasma membrane in subapical regions, and at forming septa. Actin patches
in ascogonia were most prominent at the ascogonial tip. Microtubules were
successfully labeled using chimeric β-tubulin-GFP from
N. crassa. Microtubules show a non-linear distribution with the exception of the
subapical region where long microtubules are located along the plasma membrane.
2. Characterization of the PR pathway
This section focusses on the characterization and the localization of the PR pathway during
fungal development. PR kinases are encoded by SMAC_03492 (mak2), SMAC_06526
(mek2) and SMAC_05356 (mik2). All share a high sequence identity to respective homologs
in N. crassa, with rates of 94.07% (mak2), 91.18% (mek2) and 91.84% (mik2). For functional
characterization, deletion strains of mak2, mek2, and mik2 had been generated prior to this
Figure 10: Confocal microscopy of a fluorescent marker for microtubules. The strain TSA132D1 contains microtubules labeled with β-tubulin-GFP, which are organized in longitudinal filaments in subapical regions and show non-linear organization in more peripheral hyphae and sexual structures. Scale bars indicates 10 μm. BF: Bright-field.

IV. Results 33
study (Beier, pers. communication). PCR fragements amplified from plasmids pFlip5-MAK2
and pFlip3-MAK2 containing the 5’ and 3’ flanking region of mak2, respectively, were co-
transformed into Δku70 (Figure S1A+B). In contrast to other deletion strains used in this
study, the deletion strain of mak2 is marker-free since it was generated using the FLP/FRT
recombination system (Bloemendal et al. 2014; Teichert et al. 2017). For generation of
Δmek2 (Figure S2A) and Δmik2 (Figure S3A), linearized plasmids pKO-mek2 and pKO-
mik2 were transformed into Δku70, respectively. Primary transformants of all strains were
tested for hygromycin resistance and analyzed by PCR (data not shown). Transformants
carrying the knockout construct were crossed against fus to obtain ascospore isolates
without Δku70 background (crossing of Δmek2/Δku70 was performed by Anna Beier (pers.
communication), while crossing of Δmak2/Δku70 and Δmik2/Δku70 was conducted during
this study). All ascospore isolates from kinase deletion strains carry the fus1-1 mutation
since ascospores with black wild type spore color carrying a kinase deletion never
germinated (see section 3.3). PCR was performed for verification of correct recombination
of deletion cassettes in Δmak2 (Figure S1B), Δmek2 (Figure S2B) and Δmik2 (Figure S3B).
Strains SaS1298 (Δmak2), S143932 (Δmek2) and SaS1090 (Δmik2) were chosen for
further analysis. The deletion strain of ham5 was generated and verified by Ramona Märker
(Märker 2019).
2.1 Deletion of genes coding for pheromone response kinases leads to defects in
sexual development, hyphal fusion, and vegetative growth
As mentioned above, several studies have indicated an involvement of PR kinases in
developmental processes during sexual reproduction of filamentous fungi (Li et al. 2005;
Lalucque et al. 2012). The role of mak2, mek2, and mik2 as well as ham5 during sexual
development of S. macrospora was investigated over time. As shown in Figure 11, wild type
and fus control strains show normal fruiting body development and generate ascogonia after
three days. Ascogonia develop into unpigmented and pigmented protoperithecia after five
days and ultimately produce mature perithecia after seven days. Deletion strains of PR
kinases show severe defects in fruiting body formation, only producing very small
unpigmented protoperithecia that are unable to differentiate further. In contrast to the PR
kinase deletion strains, Δham5 produces larger protoperithecia, which show pigmentation.
However, Δham5 fails to produce mature perithecia like the kinase deletion strains
(characterization of Δham5 was already conducted by Märker (2019)). Ectopic integration
of genes encoding for N-terminal GFP-fusions of MAK2, MEK2, MIK2 and HAM5 into the
corresponding deletion strain restored fertility, resulting in mature perithecia (Figure 12A)
and ascospore generation after seven days.

IV. Results 34
The PR pathway plays a major role during cell communication in N. crassa, where deletion
of homologous genes results in the lack of hyphal fusion (Pandey et al. 2004). Microscopic
investigation of strains lacking components of the PR pathway revealed the same hyphal
fusion deficiency in S. macrospora (Figure 12B, upper lane). Integration of the GFP-fusions
constructs restored hyphal fusion in Δmak2, Δmek2, Δmik2; however, expression of gfp-
ham5 in Δham5 failed to rescue this defect (Figure 12B, lower lane).
Another conserved signaling complex controlling sexual development is the STRIPAK
complex. For some STRIPAK deletion strains, a defect in the septation of ascogonial coils
was described (Bloemendal et al. 2010; Beier et al. 2016). Since fruiting body maturation of
PR and STRIPAK deletion strains arrests at the same developmental stage, it was
investigated if Δmak2, Δmek2, Δmik2, and Δham5 strains also exhibit a defect in ascogonial
septation. However, staining of ascogonial coils with the cell wall marker calcofluor white
revealed that PR deletion strains are not impaired in ascogonial septation (Figure S4).
Figure 11: Microscopic investigation of sexual development of Δmak2, Δmek2, Δmik2 and Δham5. Sexual development of deletion strains of PR kinase and ham5 was observed after three (ascogonia), five (unpigmented and pigmented protoperithecia) and seven days (perithecia). The fus and wild type strains were used as control for PR kinase deletion strains and Δham5, respectively. All kinase deletion strains show an arrest of fruiting body formation at the stage of unpigmented protoperithecia, while larger and pigmented protoperithecia are formed in Δham5. Scale bars indicate 20 μm (black) or 100 μm (white).

IV. Results 35
A reduced growth rate was observed for the PR kinase deletion strains. To quantify the
vegetative growth of PR kinase deletion and complemented strains, growth rates were
measured after growth for 24 and 48h at 27°C on solid SWG. In comparison to the control
strain fus, which had vegetative growth rates of 2.62 ± 0.36 cm/day, PR kinase deletion
strains displayed significantly reduced growth with rates of 0.82 ± 0.04 cm/day for Δmak2,
0.81 ± 0.05 cm/day for Δmek2 and 0.78 ± 0.09 cm/day for Δmik2 (Figure 13A). Growth was
restored in all strains carrying the GFP-fusion constructs, indicating that the gene deletions
were the underlying cause for decreased growth.
Figure 12: Complementation of PR kinase deletion strains in sexual development and hyphal fusion. (A) Strains were grown on solid BMM for seven days. PR kinase deletion strains and Δham5 did not generate mature perithecia. Expression of N-terminally gfp-tagged fusion constructs of PR kinase genes and the scaffold gene ham5 in corresponding deletion strains leads to restoration of fertility. (B) For investigation of hyphal fusions, strains from (A) were grown on a layer of cellophane on MMS for two days. Hyphal fusion is impaired in PR kinase deletion strains as well as Δham5. Hyphal fusion bridges (arrowheads) observed in the control strains (fus and wild type) could not be observed in the deletion strains (asterisks) but were present in complemented strains. The hyphal fusion defect of Δham5 could not be rescued by the GFP-HAM5 construct. Scale bars indicate 20 μm (black) and 2 mm (white).

IV. Results 36
It has been reported that deletion strains of PR kinases are hypersensitive to oxidative
stress in N. crassa (März et al. 2008). The response to oxidative stress was tested by
addition of 0.01% H2O2 to the SWG medium and was otherwise performed as described for
the vegetative growth assay. The control strain fus showed 50.9 ± 5.2 % growth on oxidative
stress medium compared to growth on SWG (Figure 13B). Kinase deletion strains were
significantly more sensitive than fus, showing rates of 27.1 ± 2.9 % (Δmak2), 25.9 ± 1.1 %
(Δmek2) and 26.9 ±1.0 % (Δmik2). All of the complemented strains showed more sensitivity
to oxidative stress than fus; however, only Δmak2/fus::gfp-mak2 showed a significant
difference. Thus, the impaired adaptation to oxidative stress that could be observed in
kinase deletion strains was only partially restored by expression of GFP-fusion constructs.
Vegetative growth and oxidative stress response of Δham5 were already investigated by
Ramona Märker (Märker 2019). While PR kinase deletion strains showed growth rates
reduced by 70% as compared to the control strain fus in this thesis, only a 30% reduction
was decribed for the Δham5/fus strain. The strain Δham5/fus also showed a slightly higher
sensitivity to oxidative stress than the fus strain.
Figure 13: Vegetative growth and response to oxidative stress of PR kinase deletion and complementation strains. (A) Vegetative growth rates were measured for fus (control), PR kinase deletion and complementation strains. Measurements were taken after 24 and 48h of incubation on solid SWG medium at 27°C. Growth rates are indicated in cm per day. (B) Growth response to oxidative stress was examined like vegetative growth with 0.01% H2O2 added to the SWG medium. Stress response is depicted in % growth on SWG + H2O2 compared to vegetative growth rates of each strain on SWG. Graphs in A and B represent means and standard deviations of three biological replicats per strain. Asterisks indicate significant difference of p≤0.01 compared to control the strain fus according to student’s t-test.

IV. Results 37
Deletion strains of PR kinase genes and ham5 are defective in sexual development
and vegetative growth and are hypersensitive to oxidative stress, whereby PR kinase
gene deletion results in a slightly more severe phenotype than deletion of ham5.
Fruiting body development and vegetative growth were restored by ectopic
integration of GFP-fusion constructs, confirming the gene deletions as the cause for
these phenotypes. All deletion strains are hyphal fusion deficient and
complementation of this defect could be achieved for PR kinase deletions but not for
Δham5.
2.2 Subcellular localization of PR components during vegetative growth and sexual
development
After characterization of PR deletion strains, subcellular localization of all PR components
was examined using strains Δmak2/fus::gfp-mak2 (SaS1669), Δmek2/fus::gfp-mek2
(SaS1787), Δmik2/fus::gfp-mik2 (SaS1339) and Δham5::gfp-ham5 (RM1941). All strains
express the gfp-fusions from the gpd promoter from A. nidulans, resulting in overexpression
of the genes. However, the functionality of these fusion proteins can be assumed since
phenotypes of all deletion strains were complemented using these construct (as described
in section 2.1). As shown in Figure 14, all PR components localize around septal pores
(indicated by white arrowhead). The septal pores are small openings in septa, which allow
the distribution of cytoplasm throughout the mycelium. Localization to these pores can be
seen as thin line (at the side of the septal pore) or as two dots, if the focal plane goes though
the opening of the pore. Additionally, MAK2 accumulates in round structures in older
hyphae, which presumably are nuclei (Figure 14, red arrowheads). PR kinases MEK2 and
MIK2 as well as HAM5 localize to small spots in the cytoplasm (Figure 14: C+D, yellow
arrowhead; Figure 15E, white arrowhead). MIK2 only exhibits spot-like localization in older
hyphae. In contrast, HAM5 spots are observable throughout the whole hyphae, including
the vicinity of the hyphal tip.
For further insight into the subcellular localization of the observed distinct fluorescent spots,
the strain expressing GFP-MIK2 was used in a co-localization experiment together with the
nuclear marker strain carrying H2A-mRFP. Both strains were grown on a single BMM-
covered slide as shown in Figure 15A. The exchange of transformed nuclei through hyphal
fusion bridges allows expression of both signals in one hypha and enables co-localization
approaches. Punctate fluorescence of GFP-MIK2 was observed either in the cytoplasm
(Figure 15B, blue arrowhead) or in association with nuclear membranes (Figure 15B, white
arrowhead). Interestingly, MIK2 spots were present at both sides of

IV. Results 38
Figure 14 Localization of PR components in hyphae. (A-D) All PR components are recruited to septal pores (white arrowheads). (A) GFP-MAK2 (SaS1669) localizes to the cytoplasm and shows enhanced fluorescence in round structures resembling nuclei (red arrowhead). (B) Apart from localization to septal pores, GFP-MEK2 (SaS1787) shows faint localization the cytoplam. (C) GFP-MIK2 (SaS1339) localizes to small spots in the cytoplasm (yellow arrowhead), which can be observed in older hyphae but are lacking near the hyphal tip. (D) In contrast to MIK2, spot-like localization of GFP-HAM5 (RM1941) is more abundant and can also be found near hyphal tips (yellow arrowhead). All scale bars indicate 10 μm. BF: Bright-field.
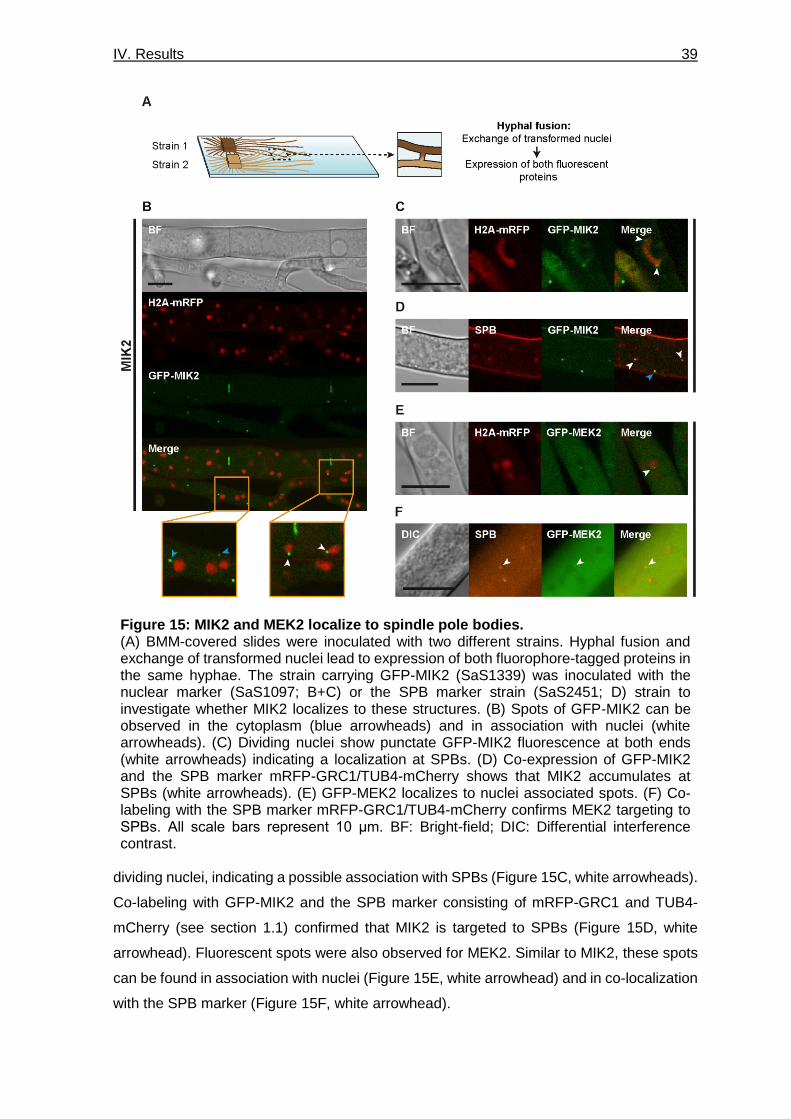
IV. Results 39
dividing nuclei, indicating a possible association with SPBs (Figure 15C, white arrowheads).
Co-labeling with GFP-MIK2 and the SPB marker consisting of mRFP-GRC1 and TUB4-
mCherry (see section 1.1) confirmed that MIK2 is targeted to SPBs (Figure 15D, white
arrowhead). Fluorescent spots were also observed for MEK2. Similar to MIK2, these spots
can be found in association with nuclei (Figure 15E, white arrowhead) and in co-localization
with the SPB marker (Figure 15F, white arrowhead).
Figure 15: MIK2 and MEK2 localize to spindle pole bodies. (A) BMM-covered slides were inoculated with two different strains. Hyphal fusion and exchange of transformed nuclei lead to expression of both fluorophore-tagged proteins in the same hyphae. The strain carrying GFP-MIK2 (SaS1339) was inoculated with the nuclear marker (SaS1097; B+C) or the SPB marker strain (SaS2451; D) strain to investigate whether MIK2 localizes to these structures. (B) Spots of GFP-MIK2 can be observed in the cytoplasm (blue arrowheads) and in association with nuclei (white arrowheads). (C) Dividing nuclei show punctate GFP-MIK2 fluorescence at both ends (white arrowheads) indicating a localization at SPBs. (D) Co-expression of GFP-MIK2 and the SPB marker mRFP-GRC1/TUB4-mCherry shows that MIK2 accumulates at SPBs (white arrowheads). (E) GFP-MEK2 localizes to nuclei associated spots. (F) Co-labeling with the SPB marker mRFP-GRC1/TUB4-mCherry confirms MEK2 targeting to SPBs. All scale bars represent 10 μm. BF: Bright-field; DIC: Differential interference contrast.
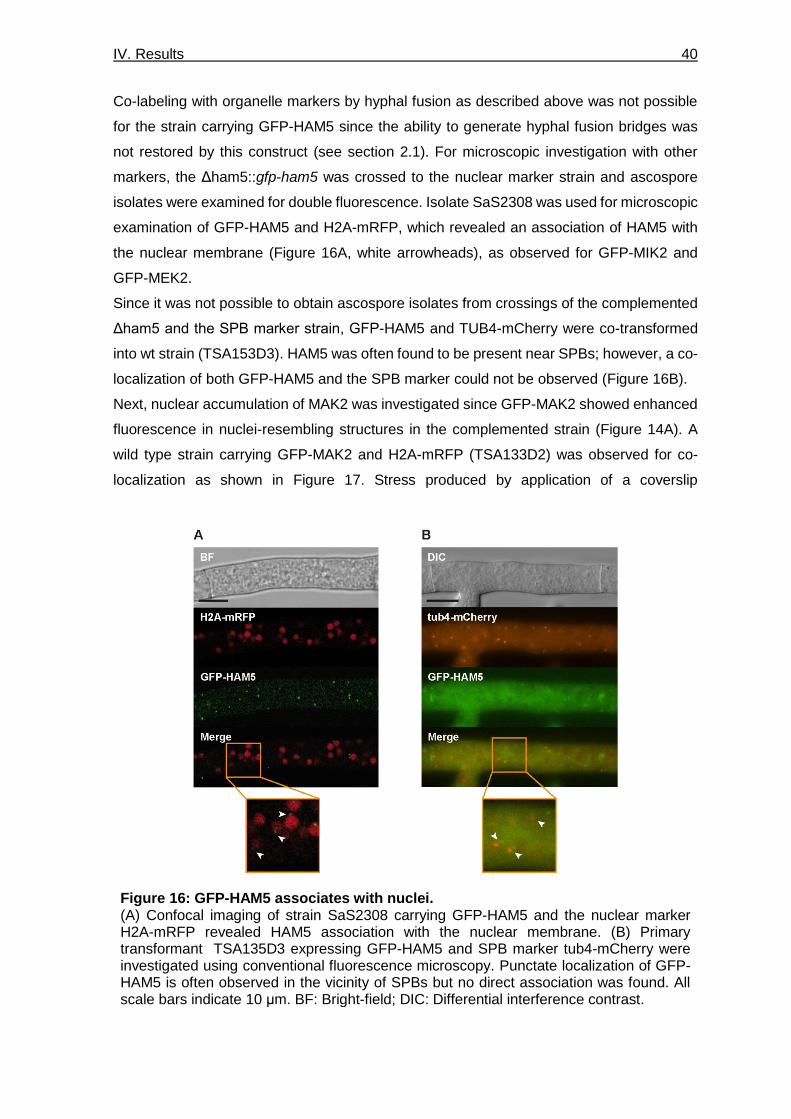
IV. Results 40
Co-labeling with organelle markers by hyphal fusion as described above was not possible
for the strain carrying GFP-HAM5 since the ability to generate hyphal fusion bridges was
not restored by this construct (see section 2.1). For microscopic investigation with other
markers, the Δham5::gfp-ham5 was crossed to the nuclear marker strain and ascospore
isolates were examined for double fluorescence. Isolate SaS2308 was used for microscopic
examination of GFP-HAM5 and H2A-mRFP, which revealed an association of HAM5 with
the nuclear membrane (Figure 16A, white arrowheads), as observed for GFP-MIK2 and
GFP-MEK2.
Since it was not possible to obtain ascospore isolates from crossings of the complemented
Δham5 and the SPB marker strain, GFP-HAM5 and TUB4-mCherry were co-transformed
into wt strain (TSA153D3). HAM5 was often found to be present near SPBs; however, a co-
localization of both GFP-HAM5 and the SPB marker could not be observed (Figure 16B).
Next, nuclear accumulation of MAK2 was investigated since GFP-MAK2 showed enhanced
fluorescence in nuclei-resembling structures in the complemented strain (Figure 14A). A
wild type strain carrying GFP-MAK2 and H2A-mRFP (TSA133D2) was observed for co-
localization as shown in Figure 17. Stress produced by application of a coverslip
Figure 16: GFP-HAM5 associates with nuclei. (A) Confocal imaging of strain SaS2308 carrying GFP-HAM5 and the nuclear marker H2A-mRFP revealed HAM5 association with the nuclear membrane. (B) Primary transformant TSA135D3 expressing GFP-HAM5 and SPB marker tub4-mCherry were investigated using conventional fluorescence microscopy. Punctate localization of GFP-HAM5 is often observed in the vicinity of SPBs but no direct association was found. All scale bars indicate 10 μm. BF: Bright-field; DIC: Differential interference contrast.

IV. Results 41
was prevented by culturing of the strain in microdishes (Figure 17A). The bottom of these
micro dishes was covered with BMM and strains were grown for 1 day. Microscopic
investigation revealed that GFP-MAK2 showed mainly cytoplasmic distribution and nearly
no accumulation in nuclei around hyphal tips (Figure 17B, white arrowhead indicates the
location of a nucleus that correlates with reduced fluorescence of GFP-MAK2). In contrast
to its localization near hyphal tips, GFP-MAK2 exhibits enhanced fluorescence in nuclei in
older hyphae (Figure 17C, white arrowhead indicates localization of a nucleus with
enhanced GFP-MAK2 fluorescence).
As described above, investigation of sexual development revealed a defect of all PR
deletion strains in the maturation of protoperithecia (Figure 11). For further insight into the
role of the PR pathway during the early stages of fruiting body development, the localization
of the PR components in ascogonia was examined using strains SaS1669 (GFP-MAK2),
SaS1787(GFP-MEK2), SaS1339 (GFP-MIK2) and RM1941 (GFP-HAM5).
While localization of GFP-HAM5 remained the same as observed in hyphae (Figure 18A,
yellow arrowheads), localization of PR kinases MAK2, MEK2, and MIK2 changed to an
Figure 17: MAK2 accumulates in nuclei of older hyphae. (A) Utilization of BMM-covered micro dishes allows microscopic observation of samples with a minimum of stress exposure. (B) Co-expression with the nuclear marker H2A-mRFP revealed that GFP-MAK2 is absent form nuclei near hyphal tips (white arrowhead). (C) MAK2 accumulates in nuclei in older hyphae (white arrowhead). Strain TSA133D2 was used for imaging. All scale bars indicate 10 μm. DIC: Differential interference contrast.

IV. Results 42
accumulation in vesicular structures (Figure 18A, white arrowheads). To better distinguish
the cellular compartments, strain SaS1669 carrying GFP-MAK2 was used for co-localization
experiments with different organelle markers. Co-labeling with markers for nuclei (H2A-
mRFP), ER (mRFP-KDEL) and peroxisomes (DsRed-SKL) as well as counterstaining with
MitoTracker was conducted to identify the localization to or association with any of these
organelles. However, a co-localization of these markers with GFP-MAK2 was not observed
(Figure 18B). Vesicular localization of PR components might be located in vacuoles.
Staining with the dye FM4-64 was used for examination of PR kinase targeting to vacuoles;
however, vacuoles of ascogonia could not be visualized using this dye (data not shown).
Microscopic investigation of PR components revealed different subcellular
localizations. MAK2 is distributed in the cytoplasm and accumulates in the nuclei of
older hyphae. MEK2, MIK2, and HAM5 are located in spots that associate with nuclei,
and MEK2 and MIK2 spots co-localize with SPBs. In ascogonia, PR kinases are
targeted to vesicular structures, while HAM5 retains the spot-like distribution.
Figure 18: PR kinases localize to vacuolar structures in ascogonia. (A) In ascogonial coils, PR kinases all show accumulation to vesicular structures. HAM5 retains the spot-like localization that was also observed for hyphae. Images were taken using strains SaS1669 (MAK2), SaS1787 (MEK2), SaS1339 (MIK2) and RM1941 (HAM5). (B) For assessment of MAK2 localization at specific subcellular organelles, strains were inoculated on BMM slides as described in Figure 15A. GFP-MAK2 (SaS1669) does not co-localize with markers for nuclei (SaS1097), endoplasmic reticulum (ER, SaS886), mitochondria (MitoTracker) or peroxisomes (SaS2485). All scale bars indicate 10 μm. BF: Bright-field.

IV. Results 43
3. Interaction of the PR pathway with the NADPH oxidase complex
Prior to this study, experimental results hinted to a genetic and physical interaction between
the PR pathway and the NOX complexes. NOX complexes function by transferring NADPH-
derived electrons across biological membranes, which catalyzes the extracellular
transformation of O2 to superoxide (O2-) (Lambeth 2004). Two complexes have been
described and characterized in S. macrospora, and both are regulated by the same protein
named NADPH oxidase regulator 1 (NOR1). It was shown that these complexes regulate
different cellular processes. While the NOX1 complex plays a role in hyphal fusion and
sexual development, the NOX2 complex is necessary for ascospore germination
(Dirschnabel et al. 2014). In this section, crosstalk between the PR pathway and the NOX
complexes was investigated during different developmental stages.
3.1 HAM5 and NOR1 co-localize in older hyphae
Previous Y2H experiments indicated a physical interaction between HAM5 of the PR
pathway and NOR1 of both NOX complexes. Before co-localization of these two proteins
by fluorescence microscopy was assessed, cellular distribution of NOR1 was investigated.
For this purpose, strain SaS1821, carrying a C-terminally GFP-tagged version of NOR1 in
a Δnor1 background, was generated. A C-terminal tag was chosen since it was already
successfully applied in studies conducted in Epichloë festucae (Takemoto et al. 2011).
Strain Δnor1/fus is defective in sexual development and hyphal fusion (Dirschnabel et al.
2014). Introducing the NOR1-GFP construct in this strain rescued both defects (Figure
19A+B), indicating that the GFP-tag did not impair NOR1 activity. Fluorescence microscopy
revealed that NOR1 had a distribution very similar to HAM5 (see section 2.2). A spot-like
localization could be observed for NOR1, which was not only present in older hyphae but
also near hyphal tips (Figure 19 C). Strain SaS1938 was used for co-labeling of the nuclear
marker H2A-mRFP and NOR1-GFP, revealing that NOR1 also associates with nuclear
membranes (Figure 19C, white arrowheads). To investigate a potential targeting to SPBs,
the complementation strain was crossed to the strain carrying the SPB marker, resulting in
strain SaS2538. NOR1 localization at SPB was neither observed with conventional nor
confocal fluorescence microscopy (Figure 19D).
Figure 20 reveals two additional localization sites of NOR1. One of these sites is the hyphal
tip, where it locates in a crescent shape (Figure 20A+B). Like all PR components
(section 2.2.), NOR1 also accumulates around septal pores. In a primary transformant with
a very strong fluorescent signal at septal pores, it was possible to observe that NOR1 forms
spherical aggregations, which then locate around the septal pore. Figure 20C shows the
distribution of these spherical aggregates at two different focal planes of the same septum.
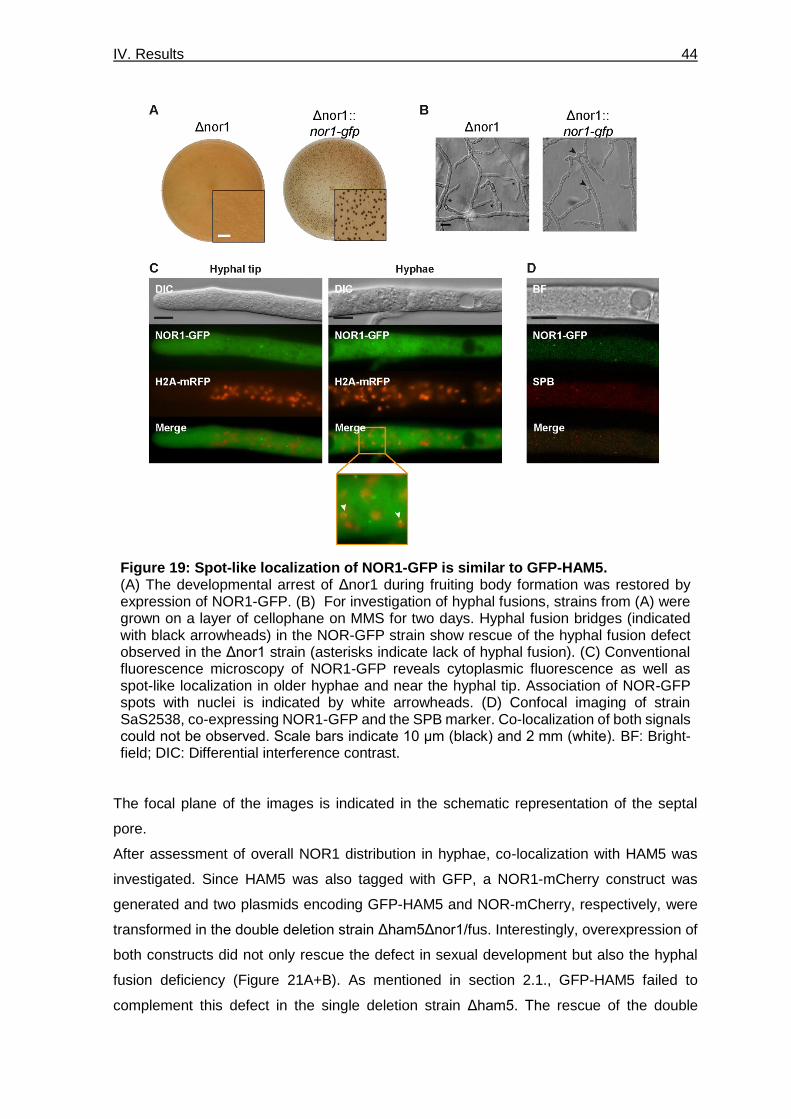
IV. Results 44
The focal plane of the images is indicated in the schematic representation of the septal
pore.
After assessment of overall NOR1 distribution in hyphae, co-localization with HAM5 was
investigated. Since HAM5 was also tagged with GFP, a NOR1-mCherry construct was
generated and two plasmids encoding GFP-HAM5 and NOR-mCherry, respectively, were
transformed in the double deletion strain Δham5Δnor1/fus. Interestingly, overexpression of
both constructs did not only rescue the defect in sexual development but also the hyphal
fusion deficiency (Figure 21A+B). As mentioned in section 2.1., GFP-HAM5 failed to
complement this defect in the single deletion strain Δham5. The rescue of the double
Figure 19: Spot-like localization of NOR1-GFP is similar to GFP-HAM5. (A) The developmental arrest of Δnor1 during fruiting body formation was restored by expression of NOR1-GFP. (B) For investigation of hyphal fusions, strains from (A) were grown on a layer of cellophane on MMS for two days. Hyphal fusion bridges (indicated with black arrowheads) in the NOR-GFP strain show rescue of the hyphal fusion defect observed in the Δnor1 strain (asterisks indicate lack of hyphal fusion). (C) Conventional fluorescence microscopy of NOR1-GFP reveals cytoplasmic fluorescence as well as spot-like localization in older hyphae and near the hyphal tip. Association of NOR-GFP spots with nuclei is indicated by white arrowheads. (D) Confocal imaging of strain SaS2538, co-expressing NOR1-GFP and the SPB marker. Co-localization of both signals could not be observed. Scale bars indicate 10 μm (black) and 2 mm (white). BF: Bright-field; DIC: Differential interference contrast.

IV. Results 45
deletion strain with the same construct might imply that overexpression of ham5 is only able
to complement this defect with simultaneous overexpression of nor1. Microscopic
investigation revealed the co-localization of GFP-HAM5 and NOR1-mCherry to spots in the
cytoplasm in older hyphae (Figure 21C).
NOR1 shows a spot-like distribution in hyphae similar to the localization observed
for HAM5. Association of NOR1 spots with nuclei was observed but co-localization
with the SPB marker was not detected. Co-expression of gfp-ham5 and nor1-mCherry
in the double deletion strain Δham5Δnor1/fus rescued the hyphal fusion defect,
which was not achieved by the integration of gfp-ham5 into the Δham5 single deletion
strain. NOR1 and HAM5 co-localize in cytoplasmic spots observed in older hyphae.
Figure 20: NOR1 is targeted to hyphal tips and forms spherical aggregates located around septal pores. (A) Schematic representation of NOR1 localization throughout the hyphae. (B) NOR1-GFP accumulates at the plasma membrane at hyphal tips of side branches. (C) Spherical aggregates of NOR1 can be observed around septal pores. Images were taken in two different focal planes of the same hyphae, showing one cross-section through the septal pore and one next to the pore. The schematic representation of NOR1 localization around septal pores indicates the focal plane of depicted fluorescent images. All scale bars represent 10 µm. DIC: Differential interference contrast.

IV. Results 46
3.2 Initiation of fruiting body formation is regulated by the PR and NOX complexes
During microscopic investigation of the Δham5 and Δham5Δnor1 strains it was observed
that both strains generate a high amount of strongly melanized protoperithecia, while such
an amount of protoperithecia could not be seen in single deletion strains of NOX
components or PR kinases. Macroscopic images of all strains for assessment of
ascogonium and protoperithecium formation are shown in Figure 22A. The control strain
fus generates mature fruiting bodies after seven days of incubation on BMM. PR kinase
deletion strains show the most severe phenotype with no macroscopically detectable
generation of fruiting bodies. As mentioned above, Δham5/fus exhibits a strikingly different
phenotype with production of macroscopically visible protoperithecia that group together in
small clusters (Figure 22A, white arrowheads). Interestingly, the double deletion strain
Δham5Δnor1/fus (provided by Ramona Märker, phenotype described in Märker (2019)) has
the same phenotype as the ham5 single deletion strain including clustered protoperithecia
(Figure 22A, white arrowheads), while Δnor1/fus and Δnox1/fus show smaller and less
Figure 21: NOR1 and HAM5 co-localize in older hyphae. (A) Strains S156605 (Δham5Δnor1/fus) and TSA149A3 (double deletion strain expressing gfp-ham5 and nor1-mCherry) were grown on solid BMM for seven days. Overexpression of GFP-HAM5 and NOR1-mCherry in the double deletion strain Δham5Δnor1/fus restores sexual development. (B) For investigation of hyphal fusions, strains from (A) were grown on a layer of cellophane on MMS for two days. Even though GFP-HAM5 failed to rescue the hyphal fusion defect of Δham5, many hyphal fusion events could be observed after expression of GFP-HAM5 and NOR1-mCherry in the double deletion strain (hyphal fusions are indicated by arrowheads, hyphal contacts without fusion by asterisks). (C) Fluorescence microscopy of TSA149A3 reveals cytoplasmic spots of NOR1-mCherry and GFP-HAM5 that co-localize in older hyphae. Scale bars indicate 10 µm (black) and 2 mm (white). DIC: Differential interference contrast.

IV. Results 47
Figure 22: Generation of fruiting bodies in PR and NOX deletion strains. (A) Fruiting body generation is shown after incubation for seven days on solid BMM. Macroscopic images of deletion strains of PR components and NOX complexes as well as the double deletion strain Δham5Δnor1/fus. Images were all taken from a the same defined part of the plate. Fus is used as a control since all examined strains carried the fus1-1 mutation. Clusters of protoperithecia are indicated by white arrowheads. (B) The quantification of ascogonia and protoperithecia of all strains shown in (A). Graphs represent means and standard deviations of three biological replicats per strain. For each strain, ascogonia and protoperithecia were counted in an area of 0.5 cm2 located 1 cm behind the growth front. Statistical analysis was performed using students t-test. Significant differences (p≤0.05) to fus (a), Δham5/fus (b1), Δham5/fus and Δham5Δnor1/fus (b2) or Δnox2/fus (c) are indicated on top of each bar. The scale bar represents 200 µm.

IV. Results 48
abundant protoperithecia. Deletion of nox2 resulted in the production of larger
protoperithecia (Figure 22A) and also mature perithecia (not shown, described in
Dirschnabel et al. (2014)).
For further analysis of these observed differences, initial stages of fruiting body generation
were quantified. For all strains, ascogonia and protoperithecia were counted in an area of
0.5 cm² located 1 cm behind the growth front. Figure 22B shows graphic representation of
counted ascogonia (dark grey bars) and protoperithecia (light grey bars). Absolute values
of means and standard deviations for all strains are listed in Table S1. Strains Δham5/fus
and Δham5Δnor1/fus produced slightly fewer ascogonia and protoperithecia than the
control, but these results were not significant. In contrast, numbers of ascogonia and
protoperithecia of PR kinase deletion strains as well as Δnor1/fus and Δnox1/fus were
significantly reduced in comparison to fus, indicating a defect during the initiation of fruiting
body generation. Interestingly, PR kinase deletion strains, as well as Δnor1/fus and
Δnox1/fus, were also significantly reduced compared to Δham5/fus in the number of
ascogonia. The number of protoperithecia of these strains was significantly reduced
compared to both, Δham5/fus and Δham5Δnor1/fus. Thus, the more severe phenotype of
Δnor1/fus is masked in the double deletion strain Δham5Δnor1/fus. Deletion of nox2
resulted in the generation of a high number of ascogonia and protoperithecia, which was
significantly higher compared to Δnor1/fus and Δnox1/fus but not fus. Thus, initiation of
fruiting body generation is regulated by the NOX1 complex comprising NOR1 and NOX1,
while the NOX2 complex has no influence.
Deletion strains of PR kinases and components of the NOX1 complex show a defect
in the initiation of fruiting body formation. This phenotype was not observed in
Δham5/fus, indicating that PR kinases operate independent of HAM5 during fruiting
body generation. Double deletion of ham5 and nor1 masks the effect of the nor1
single deletion and results in a phenotype similar to ham5 single deletion.
3.3 The PR kinases and NOX2 complex regulate ascospore germination
As described in section 2.1, deletion strains of mak2, mek2 and mik2 were generated
through homologous recombination at the 5’ and 3’ regions flanking the target genes in a
ku70 deletion strain, resulting in a replacement of the genes with a hygromycin B resistance
cassette. The Δku70 strain lacks the KU70 subunit of the KU heterodimer, which binds to
DNA double-strand breaks and initiates the non-homologous end joining (NHEJ) pathway
of DNA repair. This strain is required for generation of deletion strains by homologous
recombination. KU70 is recovered from the deletion strains by crossing to the color mutant
fus. This prevents a higher mutation rate caused by the disrupted repair systems and results
in the removal of the nourseothricin resistance cassette from the Δku70 strain. Strikingly,

IV. Results 49
ascospore isolates containing the PR kinase deletion cassette were only obtained in
combination with a fus1-1 mutation. A similar phenomenon had previously been observed
for mutants of the NOX2 complex including Δnor1 (Dirschnabel et al. 2014). To assess,
whether the PR components show the same defect in germination as the NOX2 complex,
black and brown ascospores were isolated from crosses of the deletion strains (carrying a
fus1-1 mutation) against a wild type strain. The experiment was repeated until at least 100
(Δham5) or 50 (Δmak2, Δmek2, Δmik2) germinated ascospores were obtained for black as
well as brown spores. Fewer germinated spores were obtained for the kinases because
isolated spores had lower germination rates. After germination, spores were tested for
hygromycin B resistance, which corresponds to gene deletion of ham5, mek2, and mik2
(Table 5). Resistance to hygromycin B could not be used as a selection marker for
assessment of mak2 deletion since the knockout construct for this gene contained an
FLP/FRT recombination system, which resulted in excision of the hph gene (Figure S1).
Therefore, mak2 deletion strains were distinguished from wild type strains by their inability
to generate fruiting bodies after one week of incubation on solid BMM at 27°C (Table 5,
indicated with fertile/sterile).
Since none of the isolated black spores showed resistance to hygromycin B, it can be
assumed that PR kinase deletion strains Δmak2, Δmek2, and Δmik2 all have a defect in the
germination of black ascospores similar to nox2 and nor1 deletion strains. Germinated
brown spores showed an even distribution of fus and Δmak2/fus phenotypes, which was
Table 5: Frequency of hygromycin B resistance or sterile phenotype (in %) in black- and brown-spored progeny from indicated crosses. Ascospores were isolated from crosses of the wild type strain against mutant strains carrying the fus1-1 deletion. 100 (wt x Δham5) or 50 (wt x Δmak2 / Δmek2 / Δmik2) germinated spores were collected for both spore colors (black and brown). Strains from germinated ascospores were tested for hygromycin resistance or fruiting body formation. Source of data produced in other studies is indicated in the Table. 1 Data from Dirschnabel et al. (2014).
Cross Black ascospores Brown ascospores Reference
HygS Hyg
R Hyg
S Hyg
R
Δnor1/fus x wt 100 0 48 52 1
Δnox1/fus x wt 65 35 49 51 1
Δnox2/fus x wt 100 0 43 57 1
Δste12/fus x wt 100 0 47 53 1
Δham5/fus x wt 53 47 53 47 This work
Δmik2/fus x wt 100 0 78 22 This work
Δmek2/fus x wt 100 0 66 34 This work
Fertile Sterile Fertile Sterile
Δmak2/fus x wt 100 0 56 44 This work

IV. Results 50
comparable to results obtained for Δnox2 and Δnor1 (Table 5, data from Dirschnabel et al.
(2014)). Interestingly, germinated brown spores had no equal distribution of fus1-1
background compared to Δmek2/fus and Δmik2/fus, showing ratios of approximately 2:1 for
the mek2 and 3:1 for the mik2 deletion. In contrast to the PR kinases, deletion of ham5 did
not impair the germination of black ascospores, as both black and brown spores showed
an even distribution of hygromycin sensitivity and resistance after germination. The same
result was observed for Δnox1/fus by Dirschnabel et al. (2014). Thus, germination of
melanized ascospores is mediated by the NOX2 complex including NOR1 as well as the
HAM5-independent PR pathway.
The germination defect of melanized ascospores was also observed for strains lacking the
transcription factor ste12 (Table 5). Beside a defect in ascospore germination, deletion of
the ste12 gene results in impaired ascospore generation. Unlike deletions of PR and NOX
components, neither vegetative growth nor sexual development was affected in Δste12
(Nolting & Pöggeler 2006). The ability to generate hyphal fusions was investigated in this
thesis since it has not yet been described in this strain. Fewer hyphae were detected in
older parts of the mycelium in Δste12/fus, but hyphal fusions could still be found (Figure
S5). Therefore, impaired germination of melanized ascospores seems to be the only
phenotype shared by deletion strains of PR kinases, NOX2 complex and Δste12.
To investigate whether the NOX2 complex acts upstream of the PR kinases, localization of
MAK2 was assessed in wild type spores and spores of Δnox2 (Figure 23). For the
localization of MAK2 in wt and Δnox2 spores, ascospores were collected from perithecia
Figure 23: MAK2 accumulation in nuclei of ascospores is not dependent on the NOX2 complex. Microscopic investigation of strain TSA133D2 shows accumulation of GFP-MAK2 in nuclei of ascospores by co-labeling with the nuclear marker H2A-mRFP. Nuclear localization of GFP-MAK2 is not altered in Δnox2/fus (TSA152D2). All scale bars represent 10 μm. DIC: Differential interference contrast; wt: wild type.

IV. Results 51
generated by primary transformants TSA133D2 (wt strain carrying GFP-MAK2 and H2A-
mRFP) and TSA152D2 (Δnox2 strain carrying GFP-MAK2) after seven days of incubation
on solid BMM. In wt spores, GFP-MAK2 showed cytoplasmic fluorescence and enhanced
fluorescence in two round structures, which were confirmed to be nuclei using the marker
H2A-mRFP. This accumulation of MAK2 in nuclei was not affected in nox2 deletion strains,
indicating that the NOX2 complex acts downstream of the PR kinases or that nuclear
accumulation of MAK2 is not necessary for germination of ascospores.
Strains lacking PR kinases, NOX2 complex components and the transcription factor
STE12 share a defect in the germination of melanized ascospores. MAK2 shows an
accumulation in nuclei of ascospores, which is not altered by deletion of nox2. These
results indicate that the PR pathway might operate upstream of the NOX2 complex.

V. Discussion 52
V. Discussion
Signal transduction systems are necessary for the regulation of processes like
differentiation and growth in all eukaryotes. Some of these systems are only specific for a
small subgroup of organisms while others are conserved from yeast to mammals.
Filamentous ascomycetes are well-established systems for investigation of signal
transduction during diverse cellular processes. For example, the development of complex
multicellular fruiting bodies for sexual reproduction serves as an excellent model for the
analysis of cell differentiation. Furthermore, the mechanism of cell-cell communication can
be investigated by analysis of hyphal and germling fusion events as well as analysis of the
interactions of plant pathogens with their host plant. These processes are regulated by the
interaction of several highly conserved signaling pathways. Thus, crosstalk of different
signal transduction pathways can be investigated easily during a variety of different cellular
processes in fungi (Hammond-Kosack & Parker 2003; Teichert et al. 2014a).
The filamentous ascomycete S. macrospora, which was used as a model organism in this
study, is well-investigated for the generation of fruiting bodies during sexual development
(Kück et al. 2009; Engh et al. 2010). Through genetic approaches, several signaling
pathways and complexes that play a role in fruiting body development have been identified.
Among them are the STRIPAK complex, the CWI pathway, and the NOX complex
(Bloemendal et al. 2010; Bloemendal et al. 2012; Dirschnabel et al. 2014; Teichert et al.
2014b; Nordzieke et al. 2015; Beier et al. 2016). Studies from other filamentous fungi
indicate that the PR pathway, a MAPK cascade, is also involved in the development of
sexual structures (Li et al. 2005; Bayram et al. 2012; Lalucque et al. 2012).
In this study, the PR pathway was characterized in S. macrospora and localized in different
developmental stages using fluorescence microscopy. In addition, crosstalk of the PR
pathway with the NOX complexes was investigated during different developmental
processes. Since strains lacking genes for PR and NOX components share defects in the
same cellular and developmental processes, their connection during these processes will
be discussed in the following sections. Furthermore, fluorescent imaging of PR components
and NOR1, the regulator of NOX complexes 1 and 2, revealed that both are recruited
similarly to distinct subcellular localizations. Thus, the last section will focus on the possible
role of these distinct locations as signaling hubs for PR and NOX signaling as well as other
signaling pathways.

V. Discussion 53
1. HAM5-dependent interaction of PR cascade and NOX1 complex
mediates chemotropic interaction during hyphal fusion
Characterization of PR deletion strains revealed a hyphal fusion defect for deletion strains
of mak2, mek2, and mik2 as well as for the PR scaffold ham5. Similar findings have already
been reported in N. crassa (Aldabbous et al. 2010; Fu et al. 2011), which is a well-
established model for investigation of cell-communication during fusion events in conidial
germlings. During germling fusion in N. crassa, it is proposed that MAK-2 and SOFT (SO)
are recruited to the growing cell tip in an oscillatory manner, which occurs in exact antiphase
to each other. MAK-2 is the homolog of MAK2 in S. macrospora (MEK2, MIK2, and HAM5
homologs are named MEK-2, NRC-1 and HAM-5, respectively). SO is the homolog of
PRO40, which is a scaffold for the CWI pathway. The CWI pathway comprises kinases
MAK-1, MEK-1, and MIK-1 in N. crassa (MAK1, MEK1, and MIK1 in S. macrospora), which
are all essential for hyphal fusion (März et al. 2008; Teichert et al. 2014b). During
chemotropic interaction, all PR kinases and the scaffold protein HAM-5 localize to the
hyphal tip of one germling, while only SO, but not the CWI kinases, localizes to the tip of
the communication partner (Fleißner et al. 2009; Dettmann et al. 2012). After contact of the
two germlings, SO, HAM-5 and MAK-2 accumulate at the contact site until fusion is
completed (Fleißner & Herzog 2016; Weichert et al. 2016). To date, the exact mechanisms
of how the oscillation of MAK-2 and SO is regulated are not fully understood; however,
several signaling pathways and other proteins that influence this process have been
identified (Figure 24). One of these signaling pathways is the NOX-1 complex in N. crassa,
which was also investigated during this study in S. macrospora. A link between PR and
NOX signaling during hyphal fusion is currently strongly debated (Serrano et al. 2018).
The results of this thesis provide new information about this potential link. The interaction
of NOR1 and HAM5, which had been shown prior to this study using the Y2H technique,
was further confirmed by fluorescence microscopy (Figure 19). NOR1 and HAM5 show a
similar localization pattern in hyphae and were shown to co-localize in older hyphae.
Furthermore, overexpression of both gfp-ham5 and nor1-mcherry in the Δham5Δnor1 strain
rescued the hyphal fusion defect of this double deletion strain, while overexpression of gfp-
ham5 alone failed to do the same in Δham5 (Figure 9). A possible explanation for this is
that the HAM5-NOR1-mediated interaction of PR and NOX1 complex signaling is potentially
necessary for regulation of hyphal fusion. This signaling can be disrupted by overproduction
of HAM5 since scaffold concentrations that are higher than the optimum will lead to an
incomplete assembly of the cascade (see chapter I, 1.1) (Levchenko et al. 2000). NOR1
expression under its native promoter would result in drastically lower concentrations of
NOR1 than HAM5, which is expressed from a gpd promoter, meaning that NOR1 is only

V. Discussion 54
able to interact with a fraction of HAM5-PR modules. Thus, the chance to interact with a
fully occupied HAM5-PR module is significantly lower in Δham5::gfp-ham5 than under wild-
type conditions and thus might not be sufficient to induce signal transduction above a certain
threshold level that is necessary for hyphal fusion. However, increased concentrations of
both, HAM5 and NOR1, leads to a higher possibility of interaction of NOR1 with a complete
cascade. This model could explain why the hyphal fusion defect of the double deletion strain
could be complemented since both nor1 and ham5 are overexpressed while the same was
not possible for the ham5 single deletion strain, which only had overexpression of ham5.
Expression of ham5 and nor1 under native promoters could be a possible way to verify this
hypothesis. While the expression of ham5 and nor1 under their native promoters in
Δham5Δnor1 should rescue the hyphal fusion defect, hyphal fusion should still be impaired
Figure 24: Germling fusion is regulated by crosstalk of several signaling modules. Cell-cell communication during germling fusion is regulated by the oscillation of the pheromone response (PR) module (including HAM5) and the scaffold protein SOFT (SO) to the germling tip (oscillating proteins are framed by thick black lines). SO is a scaffold for the cell wall integrity (CWI) kinases, which do not take part in the oscillation process. It was described that MAK-1 translocation to the nucleus is dependent on PR and STRIPAK (striatin-associated phosphatases and kinases) signaling. In addition, the NOX1 (NADPH oxidase 1) complex plays a role in nuclear accumulation of MAK-1 as well as oscillation of MAK-2. Lines indicate subcellular translocation. Dashed lines indicate an influence on a translocation process.

V. Discussion 55
in a double deletion strain with native expression of nor1 and simultaneous overexpression
of ham5.
Since hyphal fusion is evidently regulated by the PR as well as the NOX1 pathway, the
question arises how these pathways regulate each other. For the comprehension of this
regulation, it is important to understand how the NOX1 complex is organized. In
E. festucae, it has been shown that the NOR1 homolog NoxR forms a complex with the
scaffold BemA, the GTPase RacA and the GEF Cdc24 at hyphal tips (Takemoto et al. 2011).
These components work together with the NOX1 homolog in maintaining a mutualistic
relationship between E. festucae and its host plant (Tanaka et al. 2006; Takemoto et al.
2011). The localization of these components to germling tips has also been shown for the
respective homologs in N. crassa, named NOR-1, BEM1, RAC-1, and CDC-24. In contrast
to SO and PR components, these proteins are localized permanently to the hyphal tip
(Schürg et al. 2012; Lichius et al. 2014). The localization of these proteins to the hyphal tip
is important for regulation of polarity, i.e. the regulation of hyphal growth direction (Araujo-
Palomares et al. 2011; Takemoto et al. 2011).
For BEM1, it has been reported that localization to the hyphal tip is permanent, indicating
that the NOX1 complex (or at least BEM1) does not take part in the oscillation during cell
communication (Schürg et al. 2012). In Botrytis cinerea, NOR1 localization was also
observed at the hyphal tip as well as in granules that were transported to the hyphal tip
(Siegmund et al. 2013). A spot-like accumulation of NOR-1 in germlings was observed in
N. crassa, where it partially co-localized with NOX-1 signals (Cano-Domínguez et al. 2019).
Similarly, homologs of NOR1 and NOX1 in P. anserina were shown to co-localize in ER-
derived vesicles, while accumulation at the hyphal tip during fusion events of vegetative
hyphae was only detected for the NOR1 homolog (Lacaze et al. 2015). Homologs of NOR1
and RAC1 in E. festucae were also reported to co-localize at fusion sites of vegetative
hyphae (Kayano et al. 2018). Apart from these studies, NOX1 and also NOX2 homologs
were observed solely in association with the ER in other fungi (Schürmann et al. 2013;
Siegmund et al. 2013). Data on NOX-PR interaction during hyphal fusion indicates that the
PR pathway is epistatic to the NOX complex during this process. For example, the
oscillation of MAK-2 during germling fusion in N. crassa was abolished after inhibition of
RAC-1 (Lichius et al. 2014). The same was observed for strains lacking the NOX1 complex
component BEM1 (Schürg et al. 2012).
The mammalian homolog of NOR1, p67phox, can be phosphorylated by ERK2 in vitro as well
as in stimulated neutrophils (Dang et al. 2003), suggesting that MAPK signaling operates
upstream of NOX signaling. In N. crassa, phosphorylation targets of MAK-2 were
investigated by combining a global phosphoproteomic approach with specific inhibition of a
MAK2 allele by the ATP analog 1NM-PP1. However, NOR-1 or other proteins related to the

V. Discussion 56
NOX1 complex were not identified as MAK-2 phosphorylation targets (Jonkers et al. 2014),
which supports the hypothesis that the PR pathway is epistatic to NOX1 signaling.
It is still debated which external stimuli are required for cell-cell communication. A very
interesting model has recently been described for the communication between Fusarium
oxysporum and its host plant. It was shown that ROS production of a homolog of NOX2 is
necessary for interaction with the host plant (Nordzieke et al. 2019). As mentioned above,
NOX-mediated transfer of NADPH-derived electrons across biological membranes results
in the production of superoxide (Lambeth 2004). It was proposed that this superoxide will
be transformed into H2O2 by fungal extracellular superoxide dismutase and will
subsequently be reduced by plant peroxidases, which have been released by the host plant.
This will subsequently lead to the activation of the MAK1 homolog via a yet unknown
mechanism. However, it was discovered that Ste2, which is a homolog of the pheromone
receptor Ste2 in S. cerevisiae, is needed for this activation (Nordzieke et al. 2019). Since
Ste2 leads to the activation of the PR cascade in yeast, PR signaling might also play a role
in the communication of E. festucae and its host plant.
Receptors that play a role in signal transduction during hyphal fusion have also been
identified for N. crassa. One example is the GPI-anchored cell wall protein HAM-7, which
regulates the activity of MAK-1 and, to a lesser extent, also MAK-2 in germlings as well as
vegetative hyphae (Maddi et al. 2012). It was proposed that HAM-7 forms a complex at the
plasma membrane with HAM-6 and HAM-8, which show a similar regulation of MAPK
activity in germlings, but the mechanism behind this regulation MAPK signaling is still
unknown (Fu et al. 2014). The homolog of HAM-6 in P. anserina was identified as a
component of the NOX1 complex and has been described to co-localize with NOX1 at the
ER (Lacaze et al. 2015). Thus, signaling via HAM-6/HAM-7/HAM-8 might be mediated via
activation of the NOX1 complex.
Besides the regulation of MAK-1 activity by plasma membrane-associated complexes, it
has been described that other signaling pathways influence MAK-1 localization.
Accumulation of MAK-1 in the nucleus is dependent on the crosstalk between the STRIPAK
complex and the PR pathway. The STRIPAK complex in S. macrospora consists of PRO22,
SmMOB3, the protein phosphatase 2A (PP2A) subunits PP2AA, PP2Ac1, and PRO11, as
well as the kinases SmKIN3 and SmKIN24 (Kück et al. 2016). It orchestrates several cellular
processes by regulating the phosphorylation status of a variety of proteins (Märker 2019).
Mass spectrometry has identified an interaction between the PR kinases and homologs of
PRO11, PP2AA, and SmMOB3 in N. crassa (Dettmann et al. 2014). The interaction of MAK-
2 and HAM-3 (PRO11) was verified by co-immunoprecipitation (Dettmann et al. 2013).
STRIPAK deletion strains show the same hyphal fusion defect as observed for PR, NOX
and CWI deletion strains. It was shown that the nuclear accumulation of MAK-1 is

V. Discussion 57
dependent on the phosphorylation of the STRIPAK subunit MOB-3 (SmMOB3 homolog) by
MAK-2. Furthermore, it was described that homologs of NOX1 also regulate the nuclear
import of the MAK1 homolog in P. anserina, indicating that the NOX1 complex might
function together with the STRIPAK complex and the PR pathway in the regulation of MAK1
translocation to the nucleus (Kicka et al. 2006). This mechanism of nuclear import of MAK-
1 is included in present theories about the hyphal fusion process; however, since it was only
investigated near the tips of vegetative hyphae and not during germling communication, it
remains unclear whether the PR pathway and the STRIPAK complex have the same effect
on MAK-1 localization during this stage. If the nuclear import of MAK1 would be regulated
similarly in S. macrospora, it could also be hypothesized that this accumulation of MAK1 in
nuclei near the tips of vegetative hyphae, together with the absence of MAK2 in nuclei in
this region (Figure 15) could be important for the regulation of vegetative growth.
While it cannot be concluded that PR-STRIPAK-NOX-dependent accumulation of MAK-1 is
needed for germling communication, it is evident that the PR pathway is essential for
activation of MAK-1 in germlings and thus likely acts upstream of the CWI pathway during
germling fusion (Fu et al. 2014). Since the PR signaling is most likely epistatic to NOX1
signaling, it could be concluded that the NOX1 complex also acts upstream of the CWI
pathway.
In summary, hyphal fusion is regulated by crosstalk of the PR pathway with the NOX1
complex, which is potentially mediated by an interaction of HAM5 and NOR1. Hyphal fusion
is similar to fusion of conidial germlings, a process that is well-investigated in
N. crassa. Cell communication during germling fusion is dependent on the oscillation of SO
and MAK-2-HAM-5. The NOX-1 and STRIPAK complexes, as well as the CWI pathway,
play a role in the regulation of these highly complex cellular dynamics; however, further
research needs to be conducted to unravel the exact process. Furthermore, it also needs
to be investigated if cell communication during fusion of vegetative hyphae is regulated in
the same way as during germling fusion.
2. PR regulation of ascospore germination by activation of NOX2 via
STE12 is independent of HAM5
While the NOX1 complex primarily functions in hyphal fusion and sexual development, the
NOX2 complex, including NOR1, has been associated with regulation of ascospore
germination in S. macrospora, N. crassa, and P. anserina (Malagnac et al. 2004; Cano-
Domínguez et al. 2008; Dirschnabel et al. 2014). The germination defect observed in
S. macrospora was dependent on the presence of melanin in the cell wall of the ascospore
since deletion of nox1 or nor1 in the melanin deficient mutant fus rescued this defect in
S. macrospora (Dirschnabel et al. 2014). A similar observation was made for nox2 deletion

V. Discussion 58
strains in P. anserina (Lambou et al. 2008). The composition of the NOX2 complex is not
yet known but it is assumed that it contains similar components as the NOX1 complex
(Teichert et al. 2014a). An exception is the transmembrane protein PRO41 (homolog of
HAM-6 of N. crassa), which is excluded as a component of the NOX2 complex since it does
not share the ascospore germination defect. Instead, a different membrane protein named
PLS1 is associated with the NOX2 complex (Lambou et al. 2008; Siegmund et al. 2013;
Green et al. 2019). If the homologs of CDC-24, RAC-1, and BEM1 are part of the NOX2
complex has not yet been investigated; however, it was shown that the bem1 deletion strain
had an ascospore lethal phenotype in N. crassa (Schürg et al. 2012).
In this study, investigation of PR deletion strains revealed a defect in the germination of
melanized ascospores in strains lacking PR kinases. Melanin-dependent germination
defects were also reported for strains carrying deletions of mak2, mek2 and mik2 homologs
in P. anserina (Lalucque et al. 2012). In contrast to deletions of PR kinase genes, deletion
of ham5 did not result in a defect in ascospore germination in this study (Table 5). This is
consistent with reports for IDC1, which is a homolog of HAM5 in
P. anserina (Jamet-Vierny et al. 2007). Based on these results, it can be concluded that the
PR kinases interact with the NOX2 complex in the regulation of germination of melanized
ascospores and that this interaction is independent of the scaffolding function of HAM5
(Figure 25). It has been suggested that ascospore germination is triggered via ROS
production by the NOX2 complex. In the absence of NOX2 or the regulator NOR1, it is not
possible to produce sufficient amounts of ROS to induce germination. The residual ROS
produced by other sources than NOX2 would be enough to induce the germination process
in fus but not melanized spores since melanin is able to function as a ROS scavenger
(Dirschnabel et al. 2014).
It is still unclear how the NOX2 activity is regulated during ascospore germination.
Dirschnabel et al. (2014) proposed that the NOX2 complex acts in the same genetic
pathway as the transcription factor STE12 since the deletion of ste12 results in the same
melanin-dependent germination defect. As mentioned before, a hyphal fusion defect in the
ste12 deletion strain could not be confirmed in this study. Since Δste12 has also no defect
in sexual development, it seems that ascospore germination is the only process linking
STE12 to PR and NOX2 signaling in S. macrospora. Homologs of the transcription factor
STE12 are regulated via phosphorylation by the MAK2 homolog in S. cerevisiae (Elion et
al. 1993). It is generally assumed that the PR pathway acts primarily by activation and
phosphorylation of STE12 in other filamentous fungi. RNA sequencing conducted in
N. crassa revealed that nox-2 may be downregulated in strains lacking the gene encoding
the STE12 homolog PP-1 (Fischer et al. 2018). This reduction was not significant, possibly

V. Discussion 59
because the experiment was conducted during conditions that promote germling fusion.
Transcriptional regulation by STE12/PP-1 is likely different during ascospore germination.
The transcription profile of PP-1 also showed a slight downregulation of pls1. As mentioned
before, it is assumed that the tetraspanin PLS1 is a component of the NOX2 complex since
deletion of pls1 results in the same defect as observed in nox2 and PR deletion strains in
P. anserina (Lambou et al. 2008). Tetraspanins have been described to regulate signal
transduction by acting as a scaffold for signaling pathways (Termini & Gillette 2017), so
PLS1 could be a potential scaffold protein during ascospore germination. It can be assumed
that PLS1 interacts with NOX2 since similar subcellular localizations have been reported
for both proteins. Both are described to localize to the ER in vegetative hyphae in Botrytis
cinerea and Claviceps purpurea. However, while NOX2 and PLS1 homologs were
necessary for penetration of the host plant by B. cinerea, NOX2 homologs were dispensable
for infection of the host by C. purpurea (Schürmann et al. 2013; Siegmund et al. 2013). The
localization of NOX2 complex components including PLS1 in ascospores has not been
investigated so far.
A different germination defect was observed for strains lacking sac1 and gsa3, which
showed a general reduction in ascospore germination that was not dependent on melanin.
It is likely that GSA3, which is a heterotrimeric G protein α-subunit, and the adenylate
cyclase SAC1 act in the same genetic pathway in the control of the germination of
ascospores. Adenylyl cyclases produce cAMP, which has been shown to selectively inhibit
Figure 25: PR kinases, STE12, and the NOX2 complex act in the same pathway controlling ascospore germination. Hypothetical model showing the regulation of ascospore germination via a PR-STE12-NOX2 pathway. Activation of MAPKs leads to translocation of MAK2 to the nucleus, where it phosphorylates and activates the transcription factor STE12. Activation of STE12 induces either direct upregulation of nox2 and pls1 or it induces expression of a gene that mediates the activation of the NOX2 complex. ROS produced by NOX2 activity will initiate ascospore germination. Bold letters indicate that proteins play a role in the germination of melanized ascospores in S. macrospora. Proteins are colored in grey if an influence on ascospore germination has not yet been described. PR: pheromone response, NOX: NADPH oxidase, ROS: reactive oxygen species.

V. Discussion 60
the germination of melanized ascospores. This creates a potential link between GSA3-
SAC1 and PR-STE12-NOX2 signaling. It has been suggested that cAMP might interfere
with NOX2 signaling by inhibition of RAC1, which has been associated with NOX signaling;
however, RAC1 interaction has so far only been described for the NOX1 complex
(Dirschnabel et al. 2014). Furthermore, a defect in ascospore germination has not been
described for strains carrying deletions of rac1 or other components of NOX complexes (like
CDC-24) (Lichius et al. 2014; Kayano et al. 2018).
In summary, the germination of black ascospores is regulated by HAM5-independent
signaling of the PR pathway as well as the activation of the transcription factor STE12 and
the NOX1 complex, including NOR1. To gain further insight into the regulation of this
process, analysis of the expression levels of genes coding for NOX2 complex components
in Δste12 or PR kinase deletion strains could be used to provide a closer insight into the
organization of the complexes.
3. How is sexual development influenced by PR signaling?
Several studies have shown that the PR pathway is indispensable for the formation of
fruiting bodies in different ascomycetes (Li et al. 2005; Bayram et al. 2012; Lalucque et al.
2012). Interestingly, the characterization of strains with deletions of PR kinase genes
revealed slightly different defects in sexual development compared to deletion strains of
ham5. Protoperithecia of PR kinase deletion strains were smaller compared to those
observed for Δham5. In addition, PR kinase deletion strains showed strongly reduced
numbers of ascogonia and protoperithecia, which was not observed in this extent for
Δham5. Macroscopic investigation after seven days of growth might even indicate that
Δham5 produces ultimately more protoperithecia than the wild type strain. This implies that
the PR kinase cascade influences the initiation of fruiting body formation as well as early
protoperithecia development independent of HAM5.
A strong reduction in the fruiting body initiation was also observed in nox1 and nor1 deletion
strains. This phenotype was suppressed in the Δham5Δnor1 double mutant, which had
similar numbers of ascogonia and protoperithecia as Δham5 and also showed abundant
protoperithecia after one week of incubation. A hypothesis for explaining this phenomenon
could be that the presence of the NOX1 complex influences the binding of the kinases to
the scaffold HAM5. It is evident that HAM5 plays no role in the initiation of sexual
development; however, it remains unclear whether the kinases are regulated by a different
scaffold (further referred to as scaffold A) during this process. It could be hypothesized that
such a scaffold A exists in a low concentration and/or with a weak affinity for the kinases
(Figure 26). During vegetative growth, the PR kinase activity would be mainly regulated by

V. Discussion 61
HAM5 and only low concentrations of PR kinases would be unbound and thus available for
other scaffolds. As already mentioned in section 1.1, the PR cascade mediates hyphal
fusion dependent on HAM5. The occurrence of hyphal fusion events could induce a
negative feedback mechanism, leading to disassembly of the PR kinases from HAM5.
Unbound PR kinases could then interact with scaffold A, which would result in induction of
sexual development. In case of the deletion of the PR kinases, initiation of sexual
development would be reduced since signaling through these kinases is necessary for this
process. Deletion of either nox1 or nor1 would abolish hyphal fusion and thus also reduce
dissociation of PR kinases from HAM5. The deletion of ham5 would lead to an increased
number of ascogonia and protoperithecia since PR kinases are present in an unbound state
and can interact with scaffold A. This would also explain the masking of the Δnor1
phenotype in Δham5Δnor1. However, this theory is highly speculative and further research
will be needed to confirm these assumptions. Affinity purification of PR kinases with
subsequent mass spectrometry analysis during conditions of sexual development could be
applied for identification of an unkown scaffold, which is specific for this developmental
process. It is also possible that the PR kinases act independent of a scaffolding protein
during the initiation of fruiting body formation. This could be examined by mutation of HAM5
binding sites of all PR kinases. This approach should lead to PR kinases activation, which
is excusively independent of HAM5. According to the hypothesis, strains carrying these
constructs should have normal numbers of ascogonia and protoperithecia, but still have a
hyphal fusion defect.
Figure 26: Model for the initiation of fruiting body formation. MAPKs are bound to and regulated by HAM5 under conditions of hyphal fusion. After the occurrence of hyphal fusion in older hyphae, negative feedback regulates the disassembly of the pheromone response kinases from HAM5. Unbound kinases can subsequently interact with scaffold A (hypothetical scaffold), which will facilitate the initiation of fruiting body formation.

V. Discussion 62
Besides PR and NOX1 pathways, several components of different signaling complexes
have been reported to play a role in the formation of ascogonia and protoperithecia (Figure
27). These include STRIPAK components PRO11, PRO22, PP2Ac1, and SmKIN3 as well
as CWI component MAK1 (Pöggeler & Kück 2004; Bloemendal et al. 2010; Teichert et al.
2014b; Beier et al. 2016; Radchenko et al. 2018). However, comparison of this data is
difficult since there was no standardized method used for the assessment of the numbers
of ascogonia and protoperithecia.
A rather distinct defect that occurs after the formation of ascogonia has been described for
several STRIPAK deletion strains, which are impaired in the septation of ascogonial coils.
Ascogonial coils are not septated in pro22, pro11, pp2ac1, and smkin3 deletion strains
(Bloemendal et al. 2010; Bloemendal et al. 2012; Beier et al. 2016; Radchenko et al. 2018).
So far, this phenotype was only observed for STRIPAK deletion strains and it is still
unknown how this process is regulated. Ascogonial septation was also examined in PR
deletion strains in this study, but no defect was observed (Figure S4). It has been
hypothesized that the compartmentalization of ascogonia via septum formation is required
for the accumulation of specific signaling components, which are necessary for further
development. Since the accumulation of these signaling components fails in STRIPAK
deletion strains, they are arrested at an early protoperithecial stage (Bloemendal et al.
2010). The STRIPAK complex may also have additional functions in sexual development
after septum formation in ascogonia. A potential link between PR and STRIPAK signaling
was detected in a phosphoproteomic analysis of STRIPAK deletion strains, which revealed
HAM5 as a dephosphorylation target of the STRIPAK complex (Märker 2019). However, it
is still unknown if this dephosphorylation of HAM5 has a function in sexual development.
Figure 27: Steps of fruiting body formation and involved signaling complexes. Initiation of sexual development, ascogonial septation, and maturation of protoperithecia are regulated by several signaling complexes. Signaling pathways that regulate a specific process are indicated under the respective image. PR: pheromone response pathway, CWI: cell wall integrity pathway, NOX1: NADPH oxidase 1, STRIPAK: striatin-associated phosphatases and kinases complex.

V. Discussion 63
Lack of the NOX1 complex resulted in a similar developmental arrest as lack of PR and
CWI components. It has been suggested that an oxidative burst is necessary for the
maturation of protoperithecia. This hypothesis is supported by the detection of ROS during
differentiation of protoperithecia in N. crassa (Cano-Domínguez et al. 2008). Surprisingly, a
similar experiment revealed that the amount of ROS in protoperithecia was not reduced in
nox1 and nor1 deletion strains in S. macrospora (Dirschnabel et al. 2014). Both experiments
were conducted using a nitroblue tetrazolium (NBT) stain for visualization of superoxide,
which is the type of ROS that is produced by NOX complexes (Lambeth 2004). However,
NBT staining is not very specific for reduction by superoxide since NBT can also be reduced
by several cellular reductases and it was described that NBT staining itself can create
superoxide by autoxidation (Aitken 2018). Thus, it is not possible to draw a conclusion about
superoxide production by NOX1 during sexual development. Interestingly, BEM1 seems to
be excluded from the NOX1 complex during sexual development since bem1 deletion
strains in N. crassa were not impaired in fruiting body formation (Schürg et al. 2012). Up to
date, there is no other data about the interaction of the NOX1 complex and the PR cascade
during sexual development.
In S. cerevisiae, the PR pathway is activated by binding of pheromones to the pheromone
receptors Ste2 and Ste3 (see chapter I, section 2). Homologs in S. macrospora are named
PRE2 and PRE1, respectively. Deletion of either pre1 or pre2 has no effect on fruiting body
formation, while double deletion of these genes results in a strain with a sterile phenotype
(Mayrhofer et al. 2006). Signal transduction downstream of pheromone receptors is
normally mediated via trimeric G proteins. Three G proteins α-subunits that play a role in
sexual development in S. macrospora are named GSA1, GSA2, and GSA3 (GSA3 has
already been mentioned in section 2); however, it has not yet been investigated if these G
protein subunits function downstream of PRE1 and/or PRE2. Similar to the pheromone
receptors, the deletion of the genes coding for either of these proteins did not result in
sterility; however, double deletion of either gsa1/gsa2 or gsa1/gsa3 abolishes perithecium
formation (Kamerewerd et al. 2008). This indicates that PRE1 and PRE2 signaling is most
likely transferred via GSA1. GSA2 and GSA3 are able to compensate for the lack of GSA1
in Δgsa1 strains. Two pheromone precursor genes have been described in S. macrospora,
named ppg1 and ppg2. These genes are predicted to encode an α-factor- and an a-factor-
like pheromone, respectively. However, deletion of these genes and even a double deletion
of both genes did not result in sterility, indicating that they are dispensable for fruiting body
formation (Mayrhofer et al. 2006). However, it was shown that ppg1 is only expressed in
ascogonia and in some hyphae surrounding young protoperithecia (Teichert et al. 2012).
This specific expression pattern indicates a function during sexual development. It is
possible that other genes compensate for the deletion of ppg1 and ppg2 during fruiting body

V. Discussion 64
formation. Hyphal fusion was not investigated in any of these strains lacking pheromone
receptor, G protein α-subunit, or pheromone precursor coding genes. Moreover, activation
of the PR kinases downstream of PRE1-PRE2 has not been confirmed for S. macrospora.
Further investigation of PRE1 and PRE2 functions could provide data about upstream
regulation of the PR pathway during sexual development.
Masloff et al. (1999) identified the transcription factor PRO1 as an additional factor
regulating sexual development. Interestingly, chromatin immunoprecipitation followed by
DNA-sequencing (ChIP-Seq) revealed binding sites for PRO1 in the promotor regions of
genes coding for components of PR, CWI, NOX, and STRIPAK complexes, suggesting that
PRO1 might be a key regulator of sexual development (Steffens et al. 2016). These results
are supported by data from a combined approach of laser microdissection and RNA-seq
analysis, which allows the assessment of gene expression in protoperithecia. It was shown
that the expression of genes coding for components of these complexes was significantly
downregulated in Δpro1 protoperithecia (Teichert et al. 2012). It is still not known how PRO1
is activated specifically during maturation of protoperithecia.
In conclusion, sexual development is regulated by complex crosstalk between several
signaling pathways. The PR pathway plays a role in the initiation of sexual development as
well as the maturation of protoperithecia. Both processes are likely regulated HAM5-
independent since the initiation of sexual development is not affected and protoperithecia
development arrests at a later stage in Δham5 than in the PR kinase deletion strains.
4. Microtubule organizing centers – signaling hubs for the integration
of PR signaling with other signaling pathways?
As described above, PR components and NOR1 localize to distinct subcellular locations.
Accumulation of different signal transduction modules at specific locations might indicate
that these locations function as signaling hubs and thus provide a platform for crosstalk of
several signal transduction pathways. Targeting of this crosstalk to distinct subcellular
locations provides spatio-temporal regulation of signaling during different developmental
processes. The resulting higher local concentration of signaling modules facilitates the
attainment of the threshold value needed for signal transduction. This section will therefore
discuss if SPBs and septal pores, the two distinct localization spots for PR components and
NOR1 observed in this study, act as signaling hubs and thus mediate the crosstalk of the
PR pathway and NOX complexes as well as other signaling pathways in filamentous fungi.
The first part will focus on the localization of the PR pathway and the NOX complexes to
spindle poles. The second part will discuss the targeting of those signaling modules to septal
pores as well as the role of septal pores as potential signaling hubs. The last part will review

V. Discussion 65
additional signaling modules that localize to these places. SPBs are embedded into the
nuclear envelope and consist of three layers, which connect the cytoplasmic and nuclear
microtubules (Kilmartin 2014). They belong to the microtubule-organizing centers (MTOCs)
and are thus involved in the remodeling of the cytoskeleton. SPBs and centrosomes, which
are the functional equivalents of SPBs in higher eukaryotes, have already been suggested
to function as signaling hubs in the regulation of diverse cellular processes (Arquint et al.
2014).
In this study, PR kinases MEK2 and MIK2 were confirmed to locate at SPBs (Figure 13).
Spots observed for the PR scaffold HAM5 did not co-localize with the SPB marker but were
often visible close to SPBs (Figure 14). The investigation of HAM5 localization at SPBs was
conducted using conventional fluorescence microscopy. The microscope uses different
filter sets for detection of different fluorophores and thus needs to switch these filters
between the productions of two images. This switch produces a time-lag between the
images. Since cells were not fixed for microscopic investigation, proteins are able to
relocate during this time lag due to movement of the cytoplasm, resulting in a shift of protein
signals in the two images. The close localization of HAM5 spots to SPBs might be caused
by such a shift. Thus, although HAM5 spots did not co-localize with the SPB marker, it might
still be possible that HAM5 is recruited to these structures. Furthermore, localization of
HAM-5 and MEK-2 to nuclei-associated spots in vegetative hyphae has been observed in
N. crassa. Interestingly, spot-like localization of MEK-2 at SPBs was abolished in a ham5
deletion background (Jonkers et al. 2014), indicating that the scaffolding function of
HAM-5 is needed for this localization. In A. nidulans, homologs of all PR components have
been found in association with nuclei for a short time period. PR components localized in
spots that traveled between from plasma membrane to the nucleus (Bayram et al. 2012).
However, experiments were performed with conidial germlings and cellular localization of
PR components is most likely different in vegetative hyphae.
An association between ERK signaling and centrosomes has also been described in
mammals. In mouse fibroblasts (3T3 cells), localization of MEK and ERK at centrosomes
has been linked to the reorganization of spindle microtubules during mitosis (Kalive & Capco
2013). Similarly, in chinese hamster ovary (CHO-K1) cells, microtubule nucleation was
regulated via integrin-induced ERK signaling (Colello et al. 2012). It was further described
that inhibition of ERK signaling induced separation of the centrosome in human retinal
pigmented epithelial (RPE) cells, a phenomenon that has been linked to inhibition of cell
migration (Ou et al. 2017).
Similar to the localization of PR components, NOR1 spots were associated with nuclei.
However, since it was not possible to detect NOR1 at SPBs, no conclusion about the
function of this distinct localization can be drawn. So far, localization of NOR1 or other

V. Discussion 66
components of NOX complexes has not been described at SPBs in fungi or at centrosomes
in higher eukaryotes. Besides the PR pathway, other complexes in fungi have been
described to localize to SPBs (Figure 28). These complexes will be described at the end of
this section.
MTOCs also locate in other places than SPBs or centrosomes, i.e. in the cytoplasm or
associated with organelles like Golgi or ER (Zhang et al. 2017; Tillery et al. 2018). Septal
pores were also previously described as MTOCs in A. nidulans. It was shown that γ-tubulin,
which builds the basis for microtubule nucleation, locates at septal pores (Zekert et al.
2010). This, and the observation that microtubules bundle together at septal pores (Freitag
et al. 2004), support the hypothesis that these structures operate as MTOCs in filamentous
fungi. Interestingly, several intrinsically disordered proteins (IDPs) have been found to
associate with the septal pore and were accordingly named septal pore-associated (SPA)
proteins (Lai et al. 2011). IDPs have been identified at the center of signaling hubs and can
fold differently depending on their interaction partner. They are thus able to interact with a
variety of proteins (Wright & Dyson 2009). Therefore, it has been suggested that septal
pores might act as an additional signaling hub in filamentous fungi (Seiler & Justa-Schuch
2010; Riquelme et al. 2011; Dettmann et al. 2012).
As mentioned above, PR kinases MAK2, MEK2, and MIK2, as well as the PR scaffold
HAM5, were all shown to localize around the septal pore in this study (Figure 12). A similar
finding was reported in A. nidulans, where all PR components localized to septa.
Additionally, HamE-SteC (HAM5-MIK2), as well as SteC-MkkB (MIK2-MEK2) interactions
at septa were confirmed using bimolecular fluorescence complementation (BiFC) (Bayram
et al. 2012; Frawley et al. 2018). In N. crassa, all PR components are found in association
with septal pores and HAM-5 is recruited to septa during hyphal fusion (Jonkers et al. 2014).
Septal pore association of MEK-2 was also assessed in a ham5 deletion background, but
no changes could be detected. Furthermore, mass-spectrometry experiments conducted by
Dettmann et al. (2014) indicated that all PR kinases interact with proteins NCU00627,
NCU02972, and NCU08957, which are SPA1, SPA2, and SPA15, respectively. SPA1 was
also included in the list of phosphorylation targets of MAK-2 by Jonkers et al. (2014). Thus,
the interaction of PR kinases and HAM5 takes place at septal pores. However, data suggest
that the recruitment of PR kinases to septa is independent of HAM5 and instead mediated
via association with SPA proteins. The function of this recruitment is still unclear; however,
an influence on germling fusion can be ruled out since strains carrying deletions of N. crassa
homologs of either spa1, spa2, or spa15 were not impaired in cell communication during
this process (Dettmann et al. 2014).
Similar to components of the PR pathway, NOR1 was shown to localize to septal pores in

V. Discussion 67
this study (Figure 18). In E. festucae, the NOR1 homolog NoxR also localizes at septal
pores, where it likely forms a complex with BemA, RacA, and Cdc24 (Takemoto et al. 2011).
Targeting to septal pores was also confirmed for homologs of BemA and RacA in N. crassa
(Araujo-Palomares et al. 2011; Schürg et al. 2012). While homologs of NOR1 have not been
associated with MTOCs in higher eukaryotes, it was shown that NOX signaling has major
functions in the reorganization of the cytoskeleton, including microtubules as well as actin
and intermediate filaments (Valdivia et al. 2015).
As mentioned above, other signaling complexes besides the PR pathway and the NOX1
complex were described to localize to SPBs (Figure 28) and/or to septal pores (Figure 29).
One of these signaling pathways is the CWI pathway, which consists of the three kinases
MAK1, MEK1, and MIK1. The CWI pathway functions in hyphal fusion, sexual development,
vegetative growth, and response to cell wall stress (Teichert et al. 2014b). All CWI kinases
were described to localize to septal pores in S. macrospora (Teichert, pers.
communication). The CWI scaffold PRO40 also associates with septa; however, this
association was linked to septal pore occlusion by Woronin bodies in response to hyphal
damage in S. macrospora and the same could be detected for the N. crassa homolog SO
Figure 28: SPBs and septal pores are MTOCs that may function as signaling hubs. (A) Components of the PR, SIN, and STRIPAK modules are recruited to the spindle pole bodies (SPBs). The colors indicate that the targeting of proteins to SPBs was found in filamentous fungi (S. macrospora: dark blue; other fungi: light blue) and yeast (green). Red indicates that proteins have not yet been detected at these locations. PR: pheromone response, SIN: septation initiation network, STRIPAK: striatin-interacting phosphatases and kinases.

V. Discussion 68
(Engh et al. 2007; Fleißner & Glass 2007). Interestingly, the interaction of PRO40 and a
homolog of SPA3 from N. crassa indicates that the CWI pathway is recruited to the SPA
complex at septal pores in S. macrospora (Teichert et al. 2014b).
Another signal transduction pathway, which is linked to SPB and septal pore localization, is
the septation initiation network (SIN). It comprises germinal center kinases (GCKs) as well
as nuclear Dbf2p-related (NDR) kinases, which act together in a three-tiered kinase
cascade to regulate cell cycle progression and initiation of cytokinesis (Heilig et al. 2014;
Simanis 2015). In N. crassa, localization to SPBs and septal pores was observed for many
SIN components, including the GCKs SID-1, MST-1 and CDC-14, the STE kinase CDC-7,
and the NDR kinase DBF-2 (Heilig et al. 2013; Heilig et al. 2014). Interestingly, MST-1 was
identified as an interaction partner of MEK-2 and MIK-2 in this organism (Dettmann et al.
2014).
In S. macrospora, SID-1 and MST-1 homologs are named SmKIN3 and SmKIN24,
respectively. Both have also been described as components of the STRIPAK complex (see
section 1). Strains carrying deletions of STRIPAK components were impaired in sexual
development, hyphal fusion and vegetative growth (Kück et al. 2016). Since SmKIN3
combines developmental defects observed for the deletion of genes related to SIN and
Figure 29: Septal pore-associated proteins recruit several signaling modules to septal pores. A complex of various SPA (septal pore-associated) proteins forms the basis of the signaling hub located at septal pores. Components of PR, CWI, NOX, SIN, and MOR modules have been shown to localize to pores. The colors indicate that the targeting of proteins to septal pores was found in S. macrospora (dark blue) or other filamentous fungi (light blue). Red indicates that proteins have not yet been detected at these locations. PR: pheromone response, SIN: septation initiation network, CWI: cell wall integrity, MOR: morphogenesis orb6.

V. Discussion 69
STRIPAK signaling, it has been hypothesized that it acts as a functional link between these
signaling modules (Radchenko et al. 2018). However, no other components of the STRIPAK
complex could be detected at septal pores or spindle pole bodies in filamentous fungi,
excluding these structures as potential interaction sites. One exception is the STRIPAK
component PRO45, which was located at SPBs in deletion strains of other STRIPAK
components (Nordzieke et al. 2015). In addition to PRO45, homologs of STRIPAK
components PRO11, PRO22, and PP2A were shown to localize to the SPB in S. pombe,
where they counteract SIN signaling (Singh et al. 2011).
Another septal-pore associated complex is the morphogenesis Orb6 (MOR) network. In
N. crassa, the MOR pathway comprises the NDR kinase COT-1 with the regulator MOB-2
as well as the upstream GCK POD-6 and the scaffold HYM-1 (März et al. 2009; Heilig et al.
2014). The pathway has been associated with the regulation of polarized growth, cell
elongation, hyphal branching, and cell wall composition. While COT-1 and POD-6
accumulate exclusively in vesicles, which are actively transported to the cell tip, POD-6 and
HYM-1 additionally localize to septal pores (Seiler et al. 2006; Dettmann et al. 2012). As
mentioned above, septal pores were described as MTOCs in A. nidulans, based on the
localization of γ-tubulin at these places (Zekert et al. 2010). The γ-tubulin homolog in
S. macrospora, TUB4, was used as a marker for spindle pole bodies in this study. Since
this marker showed no accumulation at septal pores, it was not possible to confirm septal
pores as MTOCs in S. macrospora. However, targeting of TUB4 to septal pores might also
be disturbed by C-terminal fusion to mCherry.
In summary, the recruitment of several signaling modules to SPBs and septal pores
indicates that these structures function as signaling hubs in S. macrospora. SPBs might
facilitate the crosstalk of PR, SIN, and STRIPAK, while septal pores might enhance the
interaction of PR, CWI, NOX, SIN, and MOR. Further research is needed to unravel the
site-specific crosstalk of these signaling pathways and the distinct role of the PR pathway.

VI. Summary 70
VI. Summary
Crosstalk of diverse signal transduction pathways is necessary for the regulation of
developmental processes in all eukaryotes. The filamentous fungus Sordaria macrospora
is a well-investigated model organism for the generation of highly complex three-
dimensional fruiting bodies during sexual development. Investigation of deletion mutants
indicated that several conserved signaling complexes orchestrate this developmental
process, among them the pheromone response (PR) pathway, a homolog of the
mammalian ERK1/2 pathway.
The focus of this thesis was the characterization and localization of the PR pathway
including the PR scaffold HAM5. Furthermore, the physical and genetic interaction of the
PR pathway with both NADPH oxidase (NOX) complexes was investigated.
Characterization of PR deletion strains revealed defects in sexual development, vegetative
growth, hyphal fusion and the response to oxidative stress. Fluorescence imaging showed
that all PR components localize around septal pores. In addition, HAM5 and the two PR
kinases MEK2 and MIK2 localize to spots that are found in the cytoplasm and in association
with spindle pole bodies, the fungal equivalent of centrosomes. The downstream kinase
MAK2 accumulates in nuclei of older hyphae but not in nuclei close to hyphal tips. In
ascogonia, the initial stage of fruiting body formation, the localization of PR kinases changes
to vacuolar structures. The interaction of the PR pathway and the NOX complexes 1 and 2
was investigated since previous experiments showed an interaction of HAM5 with the NOX
regulator NOR1, which is essential for both NOX complexes. In this thesis, this interaction
was supported by microscopic investigation, showing co-localization of HAM5 and NOR1 in
older hyphae. Since both the PR pathway and the NOX1 complex are needed for hyphal
fusion, it is hypothesized that these signaling modules interact during the regulation of this
process. In addition to hyphal fusion, PR and NOX1 signaling is needed for the initiation of
fruiting body formation as well as the maturation of early fruiting body stages. Interestingly,
the PR kinases seem to act independently of the scaffold HAM5 in the initiation of fruiting
body formation. The same was observed for the regulation of ascospore germination.
During this process, the PR kinases likely interact with the transcription factor STE12 and
the NOX2 complex, including NOR1 to induce germination of ascospores.
In summary, the PR pathway interacts differently with the NOX complexes depending on
the developmental stage. During these interactions, the PR scaffold HAM5 is only
necessary for crosstalk between the PR pathway and the NOX1 complex during hyphal
fusion and potentially also during maturation of fruiting bodies, while crosstalk with the
NOX2 complex is independent of HAM5.

VII. Zusammenfassung 71
VII. Zusammenfassung
Die meisten eukaryotische Entwicklungsprozesse werden durch die Interaktion von
verschiedenen Signaltransduktionswegen reguliert. Der Hyphenpilz Sordaria macrospora
ist ein etablierter Modellorganismus zur Erforschung der Zelldifferenzierung während der
sexuellen Fruchtkörperentwicklung. Die Untersuchung von Entwicklungsmutanten ergab,
dass mehrere konservierte Signalkomplexe an diesem Entwicklungsprozess beteiligt sind,
u.A. der Pheromon (PR)-Signalweg, der dem ERK1/2-Signalweg in Säugetieren entspricht.
Der Fokus dieser Arbeit war die Charakterisierung und Lokalisierung des PR-Signalwegs,
einschließlich des Gerüstproteins HAM5. Des Weiteren wurde die physikalische und
genetische Interaktion des PR-Signalwegs mit den beiden NADPH-Oxidase (NOX)-
Komplexen untersucht. Deletionsstämme des PR-Signalwegs wiesen Defekte in der
sexuellen Entwicklung, dem vegetativen Wachstum, der Hyphenfusion und der Anpassung
an oxidativen Stress auf. Fluoreszenzmikroskopische Aufnahmen zeigten, dass alle PR-
Komponenten an Septenporen lokalisieren. Zudem sind HAM5 und die PR-Kinasen MEK2
und MIK2 in Punkten aufzufinden, welche sowohl im Zytoplasma als auch an
Spindelpolköpern (dem pilzlichen Äquivalent zu Zentrosomen) lokalisieren. Die Kinase
MAK2 ist kernlokalisiert, jedoch nicht in der Nähe von Hyphenspitzen. Die PR Kinasen
akkumulieren in den Initialstadien der sexuellen Entwicklung (Ascogonen) in vakuolären
Strukturen. Vorherige Experimente wiesen auf eine Interaktion zwischen HAM5 und NOR1,
dem gemeinsamen Regulator beider NOX-Komplexe, hin. Diese Interaktion wurde im Zuge
dieser Arbeit mit fluoreszenzmikroskopischen Aufnahmen bestätigt, da die Kolokalisation
beider Proteine in älteren Hyphen nachgewiesen werden konnte. Da sowohl der PR- als
auch der NOX1-Signalkomplex für die Hyphenfusion erforderlich sind, ist es wahrscheinlich,
dass beide Komplexe gemeinsam diesen Prozess regulieren. Die Signalweiterleitung durch
PR- und NOX1-Komplex wird zudem für die Initiierung und Fortschreitung der
Fruchtkörperentwicklung benötigt. Dabei ist der Einfluss der PR-Kinasen auf die Initiierung
der Fruchtkörperentwicklung unabhängig vom Gerüstprotein HAM5. Dies konnte auch in
der Regulation der Sporenkeimung festgestellt werden, welche die PR-Kinasen
wahrscheinlich in einem Zusammenspiel mit dem Transkriptionsfaktor STE12 und dem
NOX2-Komplex, einschließlich NOR1, regulieren.
Insgesamt zeigt diese Arbeit, dass der PR-Signalweg, je nach Entwicklungsstadium, auf
verschiedene Weise mit den NOX-Komplexen interagieren kann. Das Gerüstprotein HAM5
spielt dabei lediglich eine Rolle für die Interaktionen des PR-Signalwegs mit dem NOX1-
Komplex während der Hyphenfusion und der Fruchtkörperentwicklung. Für Interaktionen
des PR-Signalwegs mit dem NOX2-Komplex während der Sporenkeimung ist HAM5 jedoch
nicht notwendig.

VIII. References 72
VIII. References
Aitken R (2018) Nitroblue tetrazolium (Nbt). Reprod Biomed Online 36:90–91. doi:
10.1016/j.rbmo.2017.09.005
Aldabbous MS, Roca MG, Stout A, Huang IC, Read ND, Free SJ (2010) The ham-5,
rcm-1 and rco-1 genes regulate hyphal fusion in Neurospora crassa. Microbiology
156:2621–2629. doi: 10.1099/mic.0.040147-0
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment
search tool. J Mol Biol 215:403–410. doi: 10.1016/S0022-2836(05)80360-2
Antonelli V, Bernasconi F, Wong YH, Vallar L (2000) Activation of B-Raf and regulation
of the mitogen-activated protein kinase pathway by the G(o) α chain. Mol Biol Cell
11:1129–1142. doi: 10.1091/mbc.11.4.1129
Araujo-Palomares CL, Richthammer C, Seiler S, Castro-Longoria E (2011) Functional
characterization and cellular dynamics of the CDC-42 - RAC - CDC-24 module in
Neurospora crassa. PLoS One 6:e27148. doi: 10.1371/journal.pone.0027148
Arquint C, Gabryjonczyk AM, Nigg EA (2014) Centrosomes as signalling centres.
Philos Trans R Soc Lond B Biol Sci 369:20130464. doi: 10.1098/rstb.2013.0464
Baird GS, Zacharias DA, Tsien RY (2000) Biochemistry, mutagenesis, and
oligomerization of DsRed, a red fluorescent protein from coral. Proc Natl Acad Sci U
S A 97:11984–11989. doi: 10.1073/pnas.97.22.11984
Bayram Ö, Bayram ÖS, Ahmed YL, Maruyama J, Valerius O, Rizzoli SO, Ficner R,
Irniger S, Braus GH (2012) The Aspergillus nidulans MAPK module AnSte11-Ste50-
Ste7-Fus3 controls development and secondary metabolism. PLoS Genet
8:e1002816. doi: 10.1371/journal.pgen.1002816
Beier A, Teichert I, Krisp C, Wolters DA, Kück U (2016) Catalytic subunit 1 of protein
phosphatase 2A is a subunit of the STRIPAK complex and governs fungal sexual
development. mBio 7:e00870-16. doi: 10.1128/mBio.00870-16
Bennett RJ, Turgeon BG (2016) Fungal sex: The ascomycota. Microbiol Spectr 4: doi:
10.1128/microbiolspec.FUNK-0005-2016
Bhattacharyya RP, Reményi A, Good MC, Bashor CJ, Falick AM, Lim WA (2006) The
Ste5 scaffold allosterically modulates signaling output of the yeast mating pathway.
Science 311:822–826. doi: 10.1126/science.1120941

VIII. References 73
Birnboim HC, Dolly J (1979) A rapid alkaline extraction procedure for screening
recombinant plasmid DNA. Nucleic Acids Res 7:1513–1523. doi:
10.1093/nar/7.6.1513
Bloemendal S, Bernhards Y, Bartho K, Dettmann A, Voigt O, Teichert I, Seiler S,
Wolters DA, Pöggeler S, Kück U (2012) A homologue of the human STRIPAK
complex controls sexual development in fungi. Mol Microbiol 84:310–323. doi:
10.1111/j.1365-2958.2012.08024.x
Bloemendal S, Löper D, Terfehr D, Kopke K, Kluge J, Teichert I, Kück U (2014) Tools
for advanced and targeted genetic manipulation of the β-lactam antibiotic producer
Acremonium chrysogenum. J Biotechnol 169:51–62. doi:
10.1016/j.jbiotec.2013.10.036
Bloemendal S, Lord KM, Rech C, Hoff B, Engh I, Read ND, Kück U (2010) A mutant
defective in sexual development produces aseptate ascogonia. Eukaryot Cell
9:1856–1866. doi: 10.1128/EC.00186-10
Brennan DF, Dar AC, Hertz NT, Chao WCH, Burlingame AL, Shokat KM, Barford D
(2011) A Raf-induced allosteric transition of KSR stimulates phosphorylation of MEK.
Nature 472:366–369. doi: 10.1038/nature09860
Cano-Domínguez N, Álvarez-Delfín K, Hansberg W, Aguirre J (2008) NADPH
oxidases NOX-1 and NOX-2 require the regulatory subunit NOR-1 to control cell
differentiation and growth in Neurospora crassa. Eukaryot Cell 7:1352–1361. doi:
10.1128/EC.00137-08
Cano-Domínguez N, Bowman B, Peraza-Reyes L, Aguirre J (2019) Neurospora crassa
NADPH oxidase NOX-1 Is localized in the vacuolar system and the plasma
membrane. Front Microbiol 10:1825. doi: 10.3389/fmicb.2019.01825
Casar B, Arozarena I, Sanz-Moreno V, Pinto A, Agudo-Ibanez L, Marais R, Lewis RE,
Berciano MT, Crespo P (2009a) Ras subcellular localization defines extracellular
signal-regulated kinase 1 and 2 substrate specificity through distinct utilization of
scaffold proteins. Mol Cell Biol 29:1338–1353. doi: 10.1128/MCB.01359-08
Casar B, Crespo P (2016) ERK signals: Scaffolding scaffolds? Front Cell Dev Biol 4:49.
doi: 10.3389/fcell.2016.00049
Casar B, Pinto A, Crespo P (2008) Essential role of ERK dimers in the activation of
cytoplasmic but not nuclear substrates by ERK-scaffold complexes. Mol Cell 31:708–
721. doi: 10.1016/j.molcel.2008.07.024

VIII. References 74
Casar B, Pinto A, Crespo P (2009b) ERK dimers and scaffold proteins: Unexpected
partners for a forgotten (cytoplasmic) task. Cell Cycle 8:1007–1013. doi:
10.4161/cc.8.7.8078
Chang F, Steelman LS, Shelton JG, Lee JT, Navolanic PM, Blalock WL, Franklin R,
McCubrey JA (2003) Regulation of cell cycle progression and apoptosis by the
Ras/Raf/MEK/ERK pathway. Int J Oncol 22:469–480
Choi KY, Satterberg B, Lyons DM, Elion EA (1994) Ste5 tethers multiple protein
kinases in the MAP kinase cascade required for mating in S. cerevisiae. Cell 78:499–
512. doi: 10.1016/0092-8674(94)90427-8
Chou S, Lane S, Liu H (2006) Regulation of mating and filamentation genes by two
distinct Ste12 complexes in Saccharomyces cerevisiae. Mol Cell Biol 26:4794–4805.
doi: 10.1128/MCB.02053-05
Colello D, Mathew S, Ward R, Pumiglia K, LaFlamme SE (2012) Integrins regulate
microtubule nucleating activity of centrosome through mitogen-activated protein
kinase/extracellular signal-regulated kinase kinase/extracellular signal-regulated
kinase (MEK/ERK) signaling. J Biol Chem 287:2520–2530. doi:
10.1074/jbc.M111.254128
Collins TJ (2007) ImageJ for microscopy. Biotechniques 43(1 Suppl):25–30. doi:
10.2144/000112517
Dang PM, Morel F, Gougerot-Pocidalo MA, El Benna J (2003) Phosphorylation of the
NADPH oxidase component p67PHOX by ERK2 and P38MAPK: Selectivity of
phosphorylated sites and existence of an intramolecular regulatory domain in the
tetratricopeptide-rich region. Biochemistry 42:4520–4526. doi: 10.1021/bi0205754
Daub H, Wallasch C, Lankenau A, Herrlich A, Ullrich A (1997) Signal characteristics of
G protein-transactivated EGF receptor. EMBO J 16:7032–7044. doi:
10.1093/emboj/16.23.7032
Davis RH, Perkins DD (2002) Neurospora: A model of model microbes. Nat Rev Genet
3:397–403. doi: 10.1038/nrg797
Dettmann A, Heilig Y, Ludwig S, Schmitt K, Illgen J, Fleißner A, Valerius O, Seiler S
(2013) HAM-2 and HAM-3 are central for the assembly of the Neurospora STRIPAK
complex at the nuclear envelope and regulate nuclear accumulation of the MAP
kinase MAK-1 in a MAK-2-dependent manner. Mol Microbiol 90:796–812. doi:
10.1111/mmi.12399

VIII. References 75
Dettmann A, Heilig Y, Valerius O, Ludwig S, Seiler S (2014) Fungal communication
requires the MAK-2 pathway elements STE-20 and RAS-2, the NRC-1 adapter STE-
50 and the MAP kinase scaffold HAM-5. PLoS Genet 10:e1004762. doi:
10.1371/journal.pgen.1004762
Dettmann A, Illgen J, März S, Schürg T, Fleissner A, Seiler S (2012) The NDR kinase
scaffold HYM1/MO25 is essential for MAK2 MAP kinase signaling in Neurospora
crassa. PLoS Genet 8:e1002950. doi: 10.1371/journal.pgen.1002950
Dhanasekaran DN, Kashef K, Lee CM, Xu H, Reddy EP (2007) Scaffold proteins of
MAP-kinase modules. Oncogene 26:3185–3202. doi: 10.1038/sj.onc.1210411
Dirschnabel DE, Nowrousian M, Cano-Domínguez N, Aguirre J, Teichert I, Kück U
(2014) New insights into the roles of NADPH oxidases in sexual development and
ascospore germination in Sordaria macrospora. Genetics 196:729–744. doi:
10.1534/genetics.113.159368
Dower WJ, Miller JF, Ragsdale CW (1988) High effiency transformation of E. coli by high
volate electroporation. Nucleic Acids Res 16:6127–6145. doi:
10.1093/nar/16.13.6127
Ebbing T (2017) Signaling during sexual developement in filamentous fungi: Functional
analysis of the MAPKKK MIK2. Bachelor Thesis, Ruhr-University Bochum
Elion EA, Satterberg B, Kranz JE (1993) FUS3 phosphorylates multiple components of
the mating signal transduction cascade: Evidence for STE12 and FAR1. Mol Biol Cell
4:495–510. doi: 10.1091/mbc.4.5.495
Elleuche S, Pöggeler S (2008) Visualization of peroxisomes via SKL-tagged DsRed
protein in Sordaria macrospora. Fungal Genet Rep 55: doi: 10.4148/1941-4765.1083
Engh I, Nowrousian M, Kück U (2010) Sordaria macrospora, a model organism to study
fungal cellular development. Eur J Cell Biol 89:864–872. doi:
10.1016/j.ejcb.2010.07.002
Engh I, Würtz C, Witzel-Schlömp K, Zhang HY, Hoff B, Nowrousian M, Rottensteiner
H, Kück U (2007) The WW domain protein PRO40 is required for fungal fertility and
associates with woronin bodies. Eukaryot Cell 6:831–843. doi: 10.1128/EC.00269-06
Ferguson SS (2001) Evolving concepts in G protein-coupled receptor endocytosis: The
role in receptor desensitization and signaling. Pharmacol Rev 53:1–24

VIII. References 76
Fischer MS, Wu VW, Lee JE, O’Malley RC, Glass NL (2018) Regulation of cell-to-cell
communication and cell wall integrity by a network of map kinase pathways and
transcription factors in Neurospora crassa. Genetics 209:489–506. doi:
10.1534/genetics.118.300904
Fleißner A, Glass NL (2007) SO, a protein involved in hyphal fusion in Neurospora
crassa, localizes to septal plugs. Eukaryot Cell 6:84–94. doi: 10.1128/EC.00268-06
Fleißner A, Herzog S (2016) Signal exchange and integration during self-fusion in
filamentous fungi. Semin Cell Dev Biol 57:76–83. doi: 10.1016/j.semcdb.2016.03.016
Fleißner A, Leeder AC, Roca MG, Read ND, Glass NL (2009) Oscillatory recruitment of
signaling proteins to cell tips promotes coordinated behavior during cell fusion. Proc
Natl Acad Sci U S A 106:19387–19392. doi: 10.1073/pnas.0907039106
Frawley D, Karahoda B, Bayram ÖS, Bayram Ö (2018) The HamE scaffold positively
regulates MpkB phosphorylation to promote development and secondary metabolism
in Aspergillus nidulans. Sci Rep 8:16588. doi: 10.1038/s41598-018-34895-6
Freitag M, Hickey PC, Raju NB, Selker EU, Read ND (2004) GFP as a tool to analyze
the organization, dynamics and function of nuclei and microtubules in Neurospora
crassa. Fungal Genet Biol 41:897–910. doi: 10.1016/j.fgb.2004.06.008
Frodyma D, Neilsen B, Costanzo-Garvey D, Fisher K, Lewis R (2017) Coordinating
ERK signaling via the molecular scaffold Kinase Suppressor of Ras. F1000Research
6:1621. doi: 10.12688/f1000research.11895.1
Fu C, Ao J, Dettmann A, Seiler S, Free SJ (2014) Characterization of the Neurospora
crassa cell fusion proteins, HAM-6, HAM-7, HAM-8, HAM-9, HAM-10, AMPH-1 and
WHI-2. PLoS One 9:e107773. doi: 10.1371/journal.pone.0107773
Fu C, Iyer P, Herkal A, Abdullah J, Stout A, Free SJ (2011) Identification and
characterization of genes required for cell-to-cell fusion in Neurospora crassa.
Eukaryot Cell 10:1100–1109. doi: 10.1128/EC.05003-11
Golduran GH, Morris NR (1995) Aspergillus nidulans as a model system for cell and
molecular biology studies. Methods Mol Genet 6:48–65. doi: 10.1016/S1067-
2389(06)80006-3
Good M, Tang G, Singleton J, Reményi A, Lim WA (2009) The Ste5 scaffold directs
mating signaling by catalytically unlocking the Fus3 MAP kinase for activation. Cell
136:1085–1097. doi: 10.1016/j.cell.2009.01.049

VIII. References 77
Green KA, Eaton CJ, Savoian MS, Scott B (2019) A homologue of the fungal
tetraspanin Pls1 is required for Epichloë festucae expressorium formation and
establishment of a mutualistic interaction with Lolium perenne. Mol Plant Pathol
20:961–975. doi: 10.1111/mpp.12805
Hamel LP, Nicole MC, Duplessis S, Ellis BE (2012) Mitogen-activated protein kinase
signaling in plant-interacting fungi: Distinct messages from conserved messengers.
Plant Cell 24:1327–1351. doi: 10.1105/tpc.112.096156
Hammond-Kosack KE, Parker JE (2003) Deciphering plant-pathogen communication:
Fresh perspectives for molecular resistance breeding. Curr Opin Biotechnol 14:177–
193. doi: 10.1016/s0958-1669(03)00035-1
Heilig Y, Dettmann A, Mouriño-Pérez RR, Schmitt K, Valerius O, Seiler S (2014)
Proper actin ring formation and septum constriction requires coordinated regulation
of SIN and MOR pathways through the germinal centre kinase MST-1. PLoS Genet
10:e1004306. doi: 10.1371/journal.pgen.1004306
Heilig Y, Schmitt K, Seiler S (2013) Phospho-regulation of the Neurospora crassa
septation initiation network. PLoS One 8:e79464. doi: 10.1371/journal.pone.0079464
Heinrich M, Köhler T, Mösch HU (2007) Role of Cdc42-Cla4 interaction in the
pheromone response of Saccharomyces cerevisiae. Eukaryot Cell 6:317–327. doi:
10.1128/EC.00102-06
Hibino K, Shibata T, Yanagida T, Sako Y (2009) A RasGTP-induced conformational
change in C-RAF is essential for accurate molecular recognition. Biophys J 97:1277–
1287. doi: 10.1016/j.bpj.2009.05.048
Hickey PC, Read ND (2009) Imaging living cells of Aspergillus in vitro. Med Mycol 47
Suppl 1:S110–S119. doi: 10.1080/13693780802546541
Ishibe S, Joly D, Zhu X, Cantley LG (2003) Phosphorylation-dependent paxillin-ERK
association mediates hepatocyte growth factor-stimulated epithelial morphogenesis.
Mol Cell 12:1275–1285. doi: 10.1016/s1097-2765(03)00406-4
Jamet-Vierny C, Debuchy R, Prigent M, Silar P (2007) IDC1, a Pezizomycotina-specific
gene that belongs to the PaMpk1 MAP kinase transduction cascade of the
filamentous fungus Podospora anserina. Fungal Genet Biol 44:1219–1230. doi:
10.1016/j.fgb.2007.04.005

VIII. References 78
Jerpseth B, Greener A, Short J, Viola J, Kretz PL (1992) XL1-Blue MRF’ E. coli cells:
mcrA-, mcrCB-, mcrF-, mmr-, hsdR- derivative of XL1-Blue cells. Strategies 5:81–83
Jonkers W, Leeder AC, Ansong C, Wang Y, Yang F, Starr TL, Camp DG 2nd, Smith
RD, Glass NL (2014) HAM-5 functions as a MAP kinase scaffold during cell fusion in
Neurospora crassa. PLoS Genet 10:e1004783. doi: 10.1371/journal.pgen.1004783
Kalive M, Capco DG (2013) Involvement of MEK, ERK, PKC and GSK3B in maintaining
the mitotic spindle. Int J Biol 5: doi: 10.5539/ijb.v5n2p1
Kamerewerd J, Jansson M, Nowrousian M, Pöggeler S, Kück U (2008) Three α-
subunits of heterotrimeric G proteins and an adenylyl cyclase have distinct roles in
fruiting body development in the homothallic fungus Sordaria macrospora. Genetics
180:191–206. doi: 10.1534/genetics.108.091603
Kapuscinski J (1995) DAPI: a DNA-specific fluorescent probe. Biotech Histochem
70:220–233. doi: 10.3109/10520299509108199
Katz M, Amit I, Yarden Y (2007) Regulation of MAPKs by growth factors and receptor
tyrosine kinases. Biochim Biophys Acta 1773:1161–1176. doi:
10.1016/j.bbamcr.2007.01.002
Kayano Y, Tanaka A, Takemoto D (2018) Two closely related Rho GTPases, Cdc42 and
RacA, of the endophytic fungus Epichloë festucae have contrasting roles for ROS
production and symbiotic infection synchronized with the host plant. PLoS Pathog
14:e1006840. doi: 10.1371/journal.ppat.1006840
Kibbe WA (2007) OligoCalc: An online oligonucleotide properties calculator. Nucleic
Acids Res 35(Web Server Issue):W43–W46. doi: 10.1093/nar/gkm234
Kicka S, Bonnet C, Sobering AK, Ganesan LP, Silar P (2006) A mitotically inheritable
unit containing a MAP kinase module. Proc Natl Acad Sci U S A 103:13445–13450.
doi: 10.1073/pnas.0603693103
Kilmartin JV (2014) Lessons from yeast : The spindle pole body and the centrosome.
Philos Trans R Soc Lond B Biol Sci 369:20130456. doi: 10.1098/rstb.2013.0456
Kim KY, Truman AW, Caesar S, Schlenstedt G, Levin DE (2010) Yeast Mpk1 cell wall
integrity mitogen-activated protein kinase regulates nucleocytoplasmic shuttling of
the Swi6 transcriptional regulator. Mol Biol Cell 21:1609–1619. doi:
10.1091/mbc.e09-11-0923

VIII. References 79
Knop M, Pereira G, Geissler S, Grein K, Schiebel E (1997) The spindle pole body
component Spc97p interacts with the γ-tubulin of Saccharomyces cerevisiae and
functions in microtubule organization and spindle pole body duplication. EMBO J
16:1550–1564. doi: 10.1093/emboj/16.7.1550
Kolch W (2005) Coordinating ERK/MAPK signalling through scaffolds and inhibitors. Nat
Rev Mol Cell Biol 6:827–837. doi: 10.1038/nrm1743
Koveal D, Schuh-Nuhfer N, Ritt D, Page R, Morrison DK, Peti W (2012) A CC-SAM, for
coiled coil-sterile α motif, domain targets the scaffold KSR-1 to specific sites in the
plasma membrane. Sci Signal 5:ra94. doi: 10.1126/scisignal.2003289
Kubitscheck U, Wedekind P, Zeidler O, Grote M, Peters R (1996) Single nuclear pores
visualized by confocal microscopy and image processing. Biophys J 70:2067–2077.
doi: 10.1016/S0006-3495(96)79811-9
Kück U, Beier AM, Teichert I (2016) The composition and function of the striatin-
interacting phosphatases and kinases (STRIPAK) complex in fungi. Fungal Genet
Biol 90:31–38. doi: 10.1016/j.fgb.2015.10.001
Kück U, Pöggeler S, Nowrousian M, Nolting N, Engh I (2009) Sordaria macrospora, a
model system for fungal development. In: Anke T (ed) THE MYCOTA XV, Springer
Verlag, Heidelberg, New York, Tokyo. pp 17–39
Lacaze I, Lalucque H, Siegmund U, Silar P, Brun S (2015) Identification of NoxD/Pro41
as the homologue of the p22phox NADPH oxidase subunit in fungi. Mol Microbiol
95:1006–1024. doi: 10.1111/mmi.12876
Lai F, Wu R, Wang J, Li C, Zou L, Lu Y, Liang C (2011) Far3p domains involved in the
interactions of Far proteins and pheromone-induced cell cycle arrest in budding
yeast. FEMS Yeast Res 11:72–79. doi: 10.1111/j.1567-1364.2010.00691.x
Lalucque H, Malagnac F, Brun S, Kicka S, Silar P (2012) A non-mendelian MAPK-
generated hereditary unit controlled by a second MAPK pathway in Podospora
anserina. Genetics 191:419–433. doi: 10.1534/genetics.112.139469
Lambeth JD (2004) NOX enzymes and the biology of reactive oxygen. Nat Rev Immunol
4:181–189. doi: 10.1038/nri1312

VIII. References 80
Lambou K, Malagnac F, Barbisan C, Tharreau D, Lebrun MH, Silar P (2008) The
crucial role of the Pls1 tetraspanin during ascospore germination in Podospora
anserina provides an example of the convergent evolution of morphogenetic
processes in fungal plant pathogens and saprobes. Eukaryot Cell 7:1809–1818. doi:
10.1128/EC.00149-08
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H,
Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG
(2007) Clustal W and Clustal X version 2.0. Bioinformatics 23:2947–2948. doi:
10.1093/bioinformatics/btm404
Laurent E, McCoy JW 3rd, Macina RA, Liu W, Cheng G, Robine S, Papkoff J,
Lambeth JD (2008) Nox1 is over-expressed in human colon cancers and correlates
with activating mutations in K-Ras. Int J Cancer 123:100–107. doi: 10.1002/ijc.23423
Levchenko A, Bruck J, Sternberg PW (2000) Scaffold proteins may biphasically affect
the levels of mitogen-activated protein kinase signaling and reduce its threshold
properties. Proc Natl Acad Sci U S A 97:5818–5823. doi: 10.1073/pnas.97.11.5818
Li D, Bobrowicz P, Wilkinson HH, Ebbole DJ (2005) A mitogen-activated protein kinase
pathway essential for mating and contributing to vegetative growth in Neurospora
crassa. Genetics 170:1091–1104. doi: 10.1534/genetics.104.036772
Lichius A, Goryachev AB, Fricker MD, Obara B, Castro-Longoria E, Read ND (2014)
CDC-42 and RAC-1 regulate opposite chemotropisms in Neurospora crassa. J Cell
Sci 127:1953–1965. doi: 10.1242/jcs.141630
Lim SD, Sun C, Lambeth JD, Marshall F, Amin M, Chung L, Petros JA, Arnold RS
(2005) Increased Nox1 and hydrogen peroxide in prostate cancer. Prostate 62:200–
207. doi: 10.1002/pros.20137
Lo Russo G, Spolveri F, Ciancio F, Mori A (2013) Mendeley: An easy way to manage,
share, and synchronize papers and citations. Plast Reconstr Surg 131:946e–947e.
doi: 10.1097/PRS.0b013e31828bd400
Lord KM, Read ND (2011) Perithecium morphogenesis in Sordaria macrospora. Fungal
Genet Biol 48:388–399. doi: 10.1016/j.fgb.2010.11.009
Lozano J, Xing R, Cai Z, Jensen HL, Trempus C, Mark W, Cannon R, Kolesnick R
(2003) Deficiency of kinase suppressor of Ras1 prevents oncogenic Ras signaling in
mice. Cancer Res 63:4232–4238

VIII. References 81
Lutomski M (2015) Complementation and functional characterization of the Sordaria
macrospora sterile mutant spd. Master thesis, Ruhr-University Bochum
Maddi A, Dettmann A, Fu C, Seiler S, Free SJ (2012) WSC-1 and HAM-7 are MAK-1
MAP kinase pathway sensors required for cell wall integrity and hyphal fusion in
Neurospora crassa. PLoS One 7:e42374. doi: 10.1371/journal.pone.0042374
Malagnac F, Lalucque H, Lepere G, Silar P (2004) Two NADPH oxidase isoforms are
required for sexual reproduction and ascospore germination in the filamentous
fungus Podospora anserina. Fungal Genet Biol 41:982–997. doi:
10.1016/j.fgb.2004.07.008
Marillonnet S, Werner S (2015) Assembly of multigene constructs using golden gate
cloning. Glyco-Engineering Methods Protoc 1321:269–284. doi: 10.1007/978-1-4939-
2760-9_19
Märker R (2019) Phospho-regulated signal transduction pathways in sexual development
of the filamentous fungus Sordaria macrospora. PhD thesis, Ruhr-University Bochum
Martínez-Soto D, Ruiz-Herrera J (2017) Functional analysis of the MAPK pathways in
fungi. Rev Iberoam Micol 34:192–202. doi: 10.1016/j.riam.2017.02.006
März S, Dettmann A, Ziv C, Liu Y, Valerius O, Yarden O, Seiler S (2009) Two NDR
kinase-MOB complexes function as distinct modules during septum formation and tip
extension in Neurospora crassa. Mol Microbiol 74:707–723. doi: 10.1111/j.1365-
2958.2009.06896.x
März S, Ziv C, Vogt N, Helmstaedt K, Cohen N, Gorovits R, Yarden O, Seiler S (2008)
The nuclear Dbf2-related kinase COT1 and the mitogen-activated protein kinases
MAK1 and MAK2 genetically interact to regulate filamentous growth, hyphal fusion
and sexual development in Neurospora crassa. Genetics 179:1313–1325. doi:
10.1534/genetics.108.089425
Masloff S, Pöggeler S, Kück U (1999) The pro1+ gene from Sordaria macrospora
encodes a C6 zinc finger transcription factor required for fruiting body development.
Genetics 152:191–199
Matheny SA, Chen C, Kortum RL, Razidlo GL, Lewis RE, White MA (2004) Ras
regulates assembly of mitogenic signalling complexes through the effector protein
IMP. Nature 427:256–260. doi: 10.1038/nature02237

VIII. References 82
Mayrhofer S, Weber JM, Pöggeler S (2006) Pheromones and pheromone receptors are
required for proper sexual development in the homothallic ascomycete Sordaria
macrospora. Genetics 172:1521–1533. doi: 10.1534/genetics.105.047381
McKay MM, Morrison DK (2007) Integrating signals from RTKs to ERK/MAPK.
Oncogene 26:3113–3121. doi: 10.1038/sj.onc.1210394
McKay MM, Ritt DA, Morrison DK (2009) Signaling dynamics of the KSR1 scaffold
complex. Proc Natl Acad Sci U S A 106:11022–11027. doi:
10.1073/pnas.0901590106
Meister M, Tomasovic A, Banning A, Tikkanen R (2013) Mitogen-activated protein
(MAP) kinase scaffolding proteins: A recount. Int J Mol Sci 14:4854–4884. doi:
10.3390/ijms14034854
Mitsushita J, Lambeth JD, Kamata T (2004) The superoxide-generating oxidase Nox1 is
functionally required for Ras oncogene transformation. Cancer Res 64:3580–3585.
doi: 10.1158/0008-5472.CAN-03-3909
Morrison DK (2012) MAP kinase pathways. Cold Spring Harb Perspect Biol 4:a011254.
doi: 10.1101/cshperspect.a011254
Mouchel-Vielh E, Bloyer S, Salvaing J, Randsholt NB, Peronnet F (2008) Involvement
of the MP1 scaffold protein in ERK signaling regulation during Drosophila wing
development. Genes Cells 13:1099–1111. doi: 10.1111/j.1365-2443.2008.01231.x
Müller J, Ory S, Copeland T, Piwnica-worms H, Morrison DK (2001) C-TAK1 regulates
ras signaling by phosphorylating the MAPK scaffold, KSR1. Mol Cell 8:983–993. doi:
10.1016/s1097-2765(01)00383-5
Mullis K, Faloona F, Scharf S, Saiki R, Horn G, Erlich H (1986) Specific enzymatic
amplification of DNA in vitro: The polymerase chain reaction. Cold Spring Harb Symp
Quant Biol 51 Pt 1:263–273. doi: 10.1101/sqb.1986.051.01.032
Nguyen A, Burack WR, Stock JL, Kortum R, Chaika OV, Afkarian M, Muller WJ,
Murphy KM, Morrison DK, Lewis RE, McNeish J, Shaw AS (2002) Kinase
suppressor of Ras (KSR) is a scaffold which facilitates mitogen-activated protein
kinase activation in vivo. Mol Cell Biol 22:3035–3045. doi: 10.1128/mcb.22.9.3035-
3045.2002

VIII. References 83
Nolting N, Pöggeler S (2006) A STE12 homologue of the homothallic ascomycete
Sordaria macrospora interacts with the MADS box protein MCM1 and is required for
ascosporogenesis. Mol Microbiol 62:853–868. doi: 10.1111/j.1365-
2958.2006.05415.x
Nordzieke DE, Fernandes TR, El Ghalid M, Turrà D, Di Pietro A (2019) NADPH
oxidase regulates chemotropic growth of the fungal pathogen Fusarium oxysporum
towards the host plant. New Phytol 224:1600-1612. doi: 10.1111/nph.16085
Nordzieke S, Zobel T, Fränzel B, Wolters DA, Kück U, Teichert I (2015) A fungal
sarcolemmal membrane-associated protein (SLMAP) homolog plays a fundamental
role in development and localizes to the nuclear envelope, endoplasmic reticulum,
and mitochondria. Eukaryot Cell 14:345–358. doi: 10.1128/EC.00241-14
Nowrousian M, Frank S, Koers S, Strauch P, Weitner T, Ringelberg C, Dunlap JC,
Loros JJ, Kück U (2007) The novel ER membrane protein PRO41 is essential for
sexual development in the filamentous fungus Sordaria macrospora. Mol Microbiol
64:923–937. doi: 10.1111/j.1365-2958.2007.05694.x
Nowrousian M, Masloff S, Pöggeler S, Kück U (1999) Cell differentiation during sexual
development of the fungus Sordaria macrospora requires ATP citrate lyase activity.
Mol Cell Biol 19:450–460. doi: 10.1128/mcb.19.1.450
Ory S, Morrison DK (2004) Signal transduction: IMPlications for Ras-dependent ERK
signaling. Curr Biol 14:277–278. doi: 10.1016/j.cub.2004.03.023
Ou Y, Xuan Pan C, Zuo J, Van Der Hoorn FA (2017) Ouabain affects cell migration via
Na,K-ATPase-p130cas and via nucleus-centrosome association. PLoS One
12:e0183343. doi: 10.1371/journal.pone.0183343
Ozoe F, Kurokawa R, Kobayashi Y, Jeong HT, Tanaka K, Sen K, Nakagawa T,
Matsuda H, Kawamukai M (2002) The 14-3-3 proteins Rad24 and Rad25 negatively
regulate Byr2 by affecting its localization in Schizosaccharomyces pombe. Mol Cell
Biol 22:7105–7119. doi: 10.1128/mcb.22.20.7105-7119.2002
Pan CQ, Sudol M, Sheetz M, Low BC (2012) Modularity and functional plasticity of
scaffold proteins as p(l)acemakers in cell signaling. Cell Signal 24:2143–2165. doi:
10.1016/j.cellsig.2012.06.002
Pandey A, Roca MG, Read ND, Glass NL (2004) Role of a mitogen-activated protein
kinase pathway during conidial germination and hyphal fusion in Neurospora crassa.
Eukaryot Cell 3:348–358. doi: 10.1128/ec.3.2.348-358.2004

VIII. References 84
Payne DM, Rossomando AJ, Martino P, Erickson AK, Her JH, Shabanowitz J, Hunt
DF, Weber MJ, Sturgill TW (1991) Identification of the regulatory phosphorylation
sites in pp42/mitogen-activated protein kinase (MAP kinase). EMBO J 10:885–892.
Pöggeler S, Kück U (2004) A WD40 repeat protein regulates fungal cell differentiation
and can be replaced functionally by the mammalian homologue striatin. Eukaryot Cell
3:232–240. doi: 10.1128/ec.3.1.232-240.2004
Radchenko D, Teichert I, Pöggeler S, Kück U (2018) A Hippo pathway-related GCK
controls both sexual and vegetative developmental processes in the fungus Sordaria
macrospora. Genetics 210:137–153. doi: 10.1534/genetics.118.301261
Rezatabar S, Karimian A, Rameshknia V, Parsian H, Majidinia M, Kopi TA, Bishayee
A, Sadeghinia A, Yousefi M, Monirialamdari M, Yousefi B (2019) RAS/MAPK
signaling functions in oxidative stress, DNA damage response and cancer
progression. J Cell Physiol 234:14951–14965. doi: 10.1002/jcp.28334
Riedl J, Crevenna AH, Kessenbrock K, Yu JH, Neukirchen D, Bista M, Bradke F,
Jenne D, Holak TA, Werb Z, Sixt M, Wedlich-Söldner R (2008) Lifeact: A versatile
marker to visualize F-actin. Nat Methods 5:605–607. doi: 10.1038/nmeth.1220
Riquelme M, Aguirre J, Bartnicki-García S, Braus GH, Feldbrügge M, Fleig U,
Hansberg W, Herrera-Estrella A, Kämper J, Kück U, Mouriño-Pérez RR,
Takeshita N, Fischer R (2018) Fungal morphogenesis, from the polarized growth of
hyphae to complex reproduction and infection structures. Microbiol Mol Biol Rev
82:e00068-17. doi: 10.1128/MMBR.00068-17
Riquelme M, Yarden O, Bartnicki-Garcia S, Bowman B, Castro-Longoria E, Free SJ,
Fleißner A, Freitag M, Lew RR, Mouriño-Pérez R, Plamann M, Rasmussen C,
Richthammer C, Roberson RW, Sanchez-Leon E, Seiler S, Watters MK (2011)
Architecture and development of the Neurospora crassa hypha - a model cell for
polarized growth. Fungal Biol 115:446–474. doi: 10.1016/j.funbio.2011.02.008
Rolke Y, Tudzynski P (2008) The small GTPase Rac and the p21-activated kinase Cla4
in Claviceps purpurea: Interaction and impact on polarity, development and
pathogenicity. Mol Microbiol 68:405–423. doi: 10.1111/j.1365-2958.2008.06159.x
Rutherford K, Parkhill J, Crook J, Horsnell T, Rice P, Rajandream MA, Barrell B
(2000) Artemis: Sequence visualization and annotation. Bioinformatics 16:944–945.
doi: 10.1093/bioinformatics/16.10.944

VIII. References 85
Sambrook J, Russell D (2001) Molecular cloning: A laboratory manual. Cold Spring
Harbor Laboratory Press, Cold Spring Harbor NY
Scheckhuber CQ, Osiewacz HD (2008) Podospora anserina: A model organism to study
mechanisms of healthy ageing. Mol Genet Genomics 280:365–374. doi:
10.1007/s00438-008-0378-6
Schindler D, Nowrousian M (2014) The polyketide synthase gene pks4 is essential for
sexual development and regulates fruiting body morphology in Sordaria macrospora.
Fungal Genet Biol 68:48–59. doi: 10.1016/j.fgb.2014.04.008
Schöneborn H, Raudzus F, Coppey M, Neumann S, Heumann R (2018) Perspectives
of RAS and RHEB GTPase signaling pathways in regenerating brain neurons. Int J
Mol Sci 19:4052. doi: 10.3390/ijms19124052
Schürg T, Brandt U, Adis C, Fleißner A (2012) The Saccharomyces cerevisiae BEM1
homologue in Neurospora crassa promotes co-ordinated cell behaviour resulting in
cell fusion. Mol Microbiol 86:349–366. doi: 10.1111/j.1365-2958.2012.08197.x
Schürmann J, Buttermann D, Herrmann A, Giesbert S, Tudzynski P (2013) Molecular
characterization of the NADPH oxidase complex in the ergot fungus Claviceps
purpurea: CpNox2 and CpPls1 are important for a balanced host-pathogen
interaction. Mol Plant-Microbe Interact 26:1151–1164. doi: 10.1094/MPMI-03-13-
0064-R
Schuster M, Kilaru S, Latz M, Steinberg G (2015) Fluorescent markers of the
microtubule cytoskeleton in Zymoseptoria tritici. Fungal Genet Biol 79:141–149. doi:
10.1016/j.fgb.2015.03.005
Seiler S, Justa-Schuch D (2010) Conserved components, but distinct mechanisms for
the placement and assembly of the cell division machinery in unicellular and
filamentous ascomycetes. Mol Microbiol 78:1058–1076. doi: 10.1111/j.1365-
2958.2010.07392.x
Seiler S, Vogt N, Ziv C, Gorovits R, Yarden O (2006) The STE20/germinal center
kinase POD6 interacts with the NDR kinase COT1 and is involved in polar tip
extension in Neurospora crassa. Mol Biol Cell 17:4080–4092. doi: 10.1091/mbc.e06-
01-0072
Serrano A, Illgen J, Brandt U, Thieme N, Letz A, Lichius A, Read ND, Fleißner A
(2018) Spatio-temporal MAPK dynamics mediate cell behavior coordination during
fungal somatic cell fusion. J Cell Sci 131:jcs213462. doi: 10.1242/jcs.213462

VIII. References 86
Shima F, Ijiri Y, Muraoka S, Liao J, Ye M, Araki M, Matsumoto K, Yamamoto N,
Sugimoto T, Yoshikawa Y, Kumasaka T, Yamamoto M, Tamura A, Kataoka T
(2010) Structural basis for conformational dynamics of GTP-bound ras protein. J Biol
Chem 285:22696–22705. doi: 10.1074/jbc.M110.125161
Siegmund U, Heller J, van Kan JAL, Tudzynski P (2013) The NADPH oxidase
complexes in Botrytis cinerea: Evidence for a close association with the ER and the
tetraspanin Pls1. PLoS One 8:e55879. doi: 10.1371/journal.pone.0055879
Simanis V (2015) Pombe’s thirteen - control of fission yeast cell division by the septation
initiation network. J Cell Sci 128:1465-1474. doi: 10.1242/jcs.094821
Singh NS, Shao N, McLean JR, Sevugan M, Ren L, Chew TG, Bimbo A, Sharma R,
Tang X, Gould KL, Balasubramanian MK (2011) SIN-inhibitory phosphatase
complex promotes Cdc11p dephosphorylation and propagates SIN asymmetry in
fission yeast. Curr Biol 21:1968–1978. doi: 10.1016/j.cub.2011.10.051
Sorkin A, von Zastrow M (2009) Endocytosis and signalling: intertwining molecular
networks. Nat Rev Mol Cell Biol 10:609–622. doi: 10.1038/nrm2748
Steffens EK, Becker K, Krevet S, Teichert I, Kück U (2016) Transcription factor PRO1
targets genes encoding conserved components of fungal developmental signaling
pathways. Mol Microbiol 102:792–809. doi: 10.1111/mmi.13491
Steinberg G, Schuster M, Theisen U, Kilaru S, Forge A, Martin-Urdiroz M (2012)
Motor-driven motility of fungal nuclear pores organizes chromosomes and fosters
nucleocytoplasmic transport. J Cell Biol 198:343–355. doi: 10.1083/jcb.201201087
Suh Y, Arnold RS, Lassegue B, Shi J, Xu X, Sorescu D, Chung AB, Griendling KK,
Lambeth JD (1999) Cell transformation by the superoxide-generating oxidase Mox1.
Nature 401:79–82. doi: 10.1038/43459
Taguchi T, Misaki R (2011) Palmitoylation pilots Ras to recycling endosomes. Small
GTPases 2:82–84. doi: 10.4161/sgtp.2.2.15245
Takahashi S, Pryciak PM (2008) Membrane localization of scaffold proteins promotes
graded signaling in the yeast MAP kinase cascade. Curr Biol 18:1184–1191. doi:
10.1016/j.cub.2008.07.050

VIII. References 87
Takemoto D, Kamakura S, Saikia S, Becker Y, Wrenn R, Tanaka A, Sumimoto H,
Scott B (2011) Polarity proteins Bem1 and Cdc24 are components of the filamentous
fungal NADPH oxidase complex. Proc Natl Acad Sci 108:2861–2866. doi:
10.1073/pnas.1017309108
Tanaka A, Christensen MJ, Takemoto D, Park P, Scott B (2006) Reactive oxygen
species play a role in regulating a fungus-perennial ryegrass mutualistic interaction.
Plant Cell 18:1052–1066. doi: 10.1105/tpc.105.039263
Teichert I (2017) Nuclear dynamics during ascospore germination in Sordaria
macrospora. Fungal Genet Biol 98:20–22. doi: 10.1016/j.fgb.2016.11.004
Teichert I, Lutomski M, Märker R, Nowrousian M, Kück U (2017) New insights from an
old mutant: SPADIX4 governs fruiting body development but not hyphal fusion in
Sordaria macrospora. Mol Genet Genomics 292:93–104. doi: 10.1007/s00438-016-
1258-0
Teichert I, Nowrousian M, Pöggeler S, Kück U (2014a) The filamentous fungus
Sordaria macrospora as a genetic model to study fruiting body development. Adv
Genet 87:199–244. doi: 10.1016/B978-0-12-800149-3.00004-4
Teichert I, Steffens EK, Schnaß N, Fränzel B, Krisp C, Wolters DA, Kück U (2014b)
PRO40 is a scaffold protein of the cell wall integrity pathway, linking the MAP kinase
module to the upstream activator protein kinase C. PLoS Genet 10:e1004582. doi:
10.1371/journal.pgen.1004582
Teichert I, Wolff G, Kück U, Nowrousian M (2012) Combining laser microdissection and
RNA-seq to chart the transcriptional landscape of fungal development. BMC
Genomics 13:511. doi: 10.1186/1471-2164-13-511
Teis D, Taub N, Kurzbauer R, Hilber D, De Araujo ME, Erlacher M, Offterdinger M,
Villunger A, Geley S, Bohn G, Klein C, Hess MW, Huber LA (2006) p14-MP1-
MEK1 signaling regulates endosomal traffic and cellular proliferation during tissue
homeostasis. J Cell Biol 175:861–868. doi: 10.1083/jcb.200607025
Teis D, Wunderlich W, Huber LA (2002) Localization of the MP1-MAPK scaffold
complex to endosomes is mediated by p14 and required for signal transduction. Dev
Cell 3:803–814. doi: 10.1016/s1534-5807(02)00364-7
Termini CM, Gillette JM (2017) Tetraspanins function as regulators of cellular signaling.
Front Cell Dev Biol 5:34. doi: 10.3389/fcell.2017.00034

VIII. References 88
Tillery M, Blake-Hedges C, Zheng Y, Buchwalter R, Megraw T (2018) Centrosomal and
non-centrosomal microtubule-organizing centers (MTOCs) in Drosophila
melanogaster. Cells 7:121. doi: 10.3390/cells7090121
Valdivia A, Duran C, San Martin A (2015) The role of Nox-mediated oxidation in the
regulation of cytoskeletal dynamics. Curr Pharm Des 21:6009–6022. doi:
10.2174/1381612821666151029112624
Van Biesen T, Hawes BE, Raymond JR, Luttrell LM, Koch WJ, Lefkowitz RJ (1996)
Go-protein α-subunits activate mitogen-activated protein kinase via a novel protein
kinase C-dependent mechanism. J Biol Chem 271:1266–1269. doi:
10.1074/jbc.271.3.1266
Vilardaga JP, Agnati LF, Fuxe K, Ciruela F (2010) G-protein-coupled receptor
heteromer dynamics. J Cell Sci 123:4215–4220. doi: 10.1242/jcs.063354
Walz M, Kück U (1995) Transformation of Sordaria macrospora to hygromycin B
resistance: Characterization of transformants by electrophoretic karyotyping and
tetrad analysis. Curr Genet 29:88–95. doi: 10.1007/BF00313198
Watson U, Jain R, Asthana S, Saini DK (2018) Spatiotemporal modulation of ERK
activation by GPCRs. Int Rev Cell Mol Biol 338:111–140. doi:
10.1016/bs.ircmb.2018.02.004
Weichert M, Lichius A, Priegnitz BE, Brandt U, Gottschalk J, Nawrath T,
Groenhagen U, Read ND, Schulz S, Fleißner A (2016) Accumulation of specific
sterol precursors targets a MAP kinase cascade mediating cell–cell recognition and
fusion. Proc Natl Acad Sci U S A 113:11877–11882. doi: 10.1073/pnas.1610527113
Westfall PJ, Patterson JC, Chen RE, Thorner J (2008) Stress resistance and signal
fidelity independent of nuclear MAPK function. Proc Natl Acad Sci U S A 105:12212–
12217. doi: 10.1073/pnas.0805797105
Whiteway MS, Wu C, Leeuw T, Clark K, Fourest-Lieuvin A, Thomas DY, Leberer E
(1995) Association of the yeast pheromone response G protein β;γ subunits with the
MAP kinase scaffold Ste5p. Science 269:1572–1575. doi: 10.1126/science.7667635
Widmann C, Gibson S, Jarpe MB, Johnson GL (1999) Mitogen-activated protein
kinase: Conservation of a three-kinase module from yeast to human. Physiol Rev
79:143–180. doi: 10.1152/physrev.1999.79.1.143

VIII. References 89
Witzel F, Maddison L, Blüthgen N (2012) How scaffolds shape MAPK signaling: What
we know and opportunities for systems approaches. Front Physiol 3:475. doi:
10.3389/fphys.2012.00475
Wortzel I, Seger R (2011) The ERK cascade: Distinct functions within various subcellular
organelles. Genes Cancer 2:195–209. doi: 10.1177/1947601911407328
Wright PE, Dyson HJ (2009) Linking folding and binding. Curr Opin Struct Biol 19:31–38.
doi: 10.1016/j.sbi.2008.12.003
Wu J, Akhmanova A (2017) Microtubule-organizing centers. Annu Rev Cell Dev Biol
33:51–75. doi: 10.1146/annurev-cellbio-100616-060615
Wu RF, Terada LS (2009) Ras and Nox: linked signaling networks? Free Radic Biol Med
47:1276–1281. doi: 10.1016/j.freeradbiomed.2009.05.037
Zeke A, Lukacs M, Lim WA, Remenyi A (2009) Scaffolds: interaction platforms for the
engineering of signalling circuits. Trends Cell Biol 19:364–374. doi:
10.1016/j.tcb.2009.05.007
Zekert N, Veith D, Fischer R (2010) Interaction of the Aspergillus nidulans microtubule-
organizing center (MTOC) component ApsB with gamma-tubulin and evidence for a
role of a subclass of peroxisomes in the formation of septal MTOCs. Eukaryot Cell
9:795–805. doi: 10.1128/EC.00058-10
Zhang H, Koo CY, Stebbing J, Giamas G (2013) The dual function of KSR1: A
pseudokinase and beyond. Biochem Soc Trans 41:1078–1082. doi:
10.1042/BST20130042
Zhang Y, Gao X, Manck R, Schmid M, Osmani AH, Osmani SA, Takeshita N, Fischer
R (2017) Microtubule-organizing centers of Aspergillus nidulans are anchored at
septa by a disordered protein. Mol Microbiol 106:285–303. doi: 10.1111/mmi.13763
Zhou Z, Gartner A, Cade R, Ammerer G, Errede B (1993) Pheromone-induced signal
transduction in Saccharomyces cerevisiae requires the sequential function of three
protein kinases. Mol Cell Biol 13:2069–2080. doi: 10.1128/mcb.13.4.2069

IX. Supplements 90
IX. Supplements
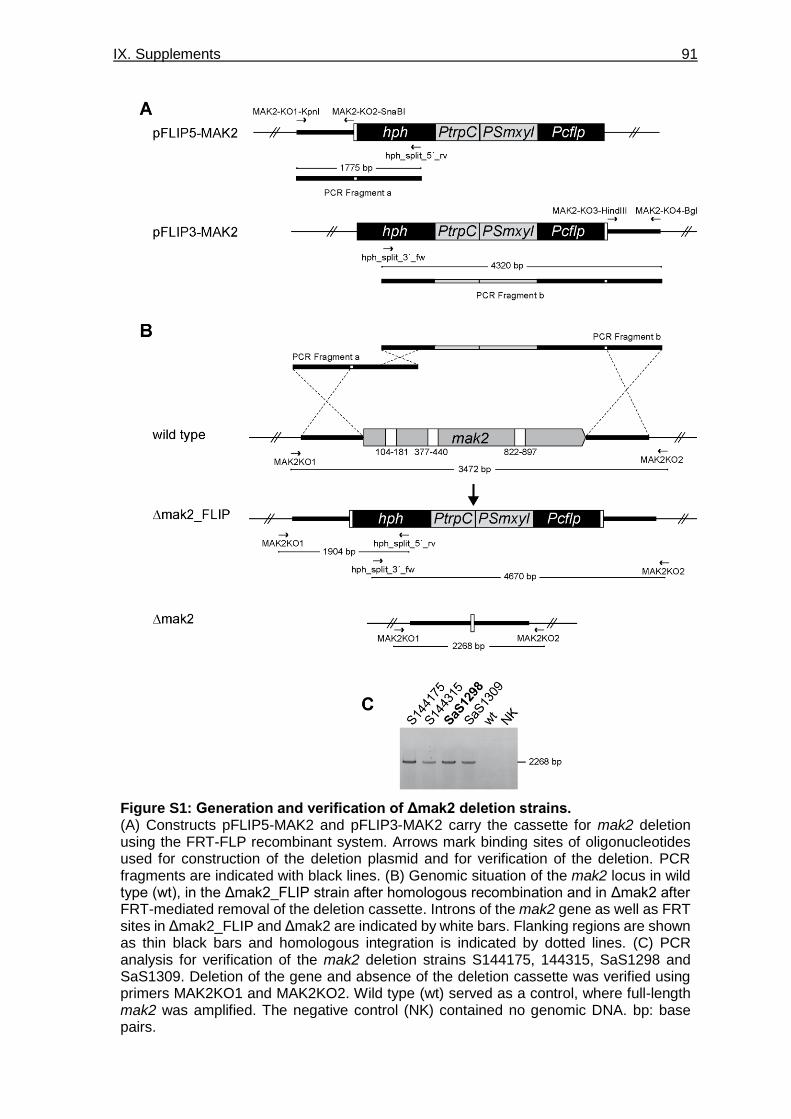
IX. Supplements 91
Figure S1: Generation and verification of Δmak2 deletion strains. (A) Constructs pFLIP5-MAK2 and pFLIP3-MAK2 carry the cassette for mak2 deletion using the FRT-FLP recombinant system. Arrows mark binding sites of oligonucleotides used for construction of the deletion plasmid and for verification of the deletion. PCR fragments are indicated with black lines. (B) Genomic situation of the mak2 locus in wild type (wt), in the Δmak2_FLIP strain after homologous recombination and in Δmak2 after FRT-mediated removal of the deletion cassette. Introns of the mak2 gene as well as FRT sites in Δmak2_FLIP and Δmak2 are indicated by white bars. Flanking regions are shown as thin black bars and homologous integration is indicated by dotted lines. (C) PCR analysis for verification of the mak2 deletion strains S144175, 144315, SaS1298 and SaS1309. Deletion of the gene and absence of the deletion cassette was verified using primers MAK2KO1 and MAK2KO2. Wild type (wt) served as a control, where full-length mak2 was amplified. The negative control (NK) contained no genomic DNA. bp: base pairs.

IX. Supplements 92
Figure S2: Generation and verification of Δmek2 deletion strains. (A) Genomic situation of the mek2 locus in wild type (wt) and in the Δmek2 strain after homologous recombination. Introns are indicated by white bars. Arrows mark binding sites of oligonucleotides used for construction of the deletion plasmid and for verification of the deletion. Flanking regions are shown as thin black bars and homologous integration is indicated by dotted lines. PCR fragments are indicated with black lines. (B) PCR analysis for verification of the mek2 deletion strains S143932 and S1143995. Gene deletion was verified using primer pairs MEK2KO1/hph1MN, hph2MN/MEK2KO2 and gfp-mek2-fw/gfp-mek2-rv for amplification of the 5’ flank, 3’ flank and mek2 gene, respectively. Wild type (wt) served as a control, where full length mek2 was amplified. The negative control (NK) contained no genomic DNA. bp: base pairs.

IX. Supplements 93
Figure S3: Generation and verification of Δmik2 deletion strains. (A) Genomic situation of the mik2 locus in wild type (wt) and in the Δmik2 strain after homologous recombination. Introns are indicated by white bars. Arrows mark binding sites of oligonucleotides used for construction of the deletion plasmid and for verification of the deletion. PCR fragments are indicated with black lines. Flanking regions are shown as thin black bars and homologous integration is indicated by dotted lines. (B) PCR analysis for verification of the mik2 deletion strains SaS1090, SaS1091, SaS1092 and SaS1093. Gene deletion was verified using primer pairs KO-1/hph1MN, hph2MN/KO-2 and KO-1/KO-3 for amplification of the 5’ flank, 3’ flank and mik2 gene, respectively. Wild type (wt) served as a control, where full length mek2 was amplified. The negative control (NK) contained no genomic DNA. bp: base pairs.

IX. Supplements 94
Figure S4: Ascogonial septation in Δmak2, Δmek2, Δmik2 and Δham5. Ascogonial coils were observed for septation using CFW staining for visualization of the cell wall. Strains were grown on BMM-covered slides for 2 days and 2.5 ng/ml CFW in 0.9% NaCl solution was applied before microscopic investigation. Fus strain was used as control for Δmak2/fus, Δmek2/fus and Δmik2/fus, while the wild type (wt) strain served as a control for Δham5. Scale bars indicate 10 μm.

IX. Supplements 95
Table S1: Absolute values for ascogonia and protoperithecia formation per 0.5 cm².
Ascogonia and protoperithecia were counted on microscopic slides that were incubated for three days. Numbers were measured in an area of 0.5 cm2, located 1 cm behind the growth front. Examination included the strains fus, Δmak2/fus, Δmek2/fus, Δmik2/fus, Δham5/fus, Δham5Δnor1/fus, Δnor1/fus, Δnox1/fus and Δnox2/fus. Means and standard deviation for ascogonia as well as protoperithecia formation is shown for each strain.
Strain Ascogonia Protoperithecia
fus 82.3 ± 40.0 113.3 ± 58.1
Δmak2/fus 8.0 ± 7.9 10.7 ± 14.2
Δmek2/fus 6.0 ± 3.5 9.0 ± 6.2
Δmik2/fus 3.7 ± 1.5 4.7 ± 4.7
Δham5/fus 41.0 ± 13.5 58.3 ± 8.4
Δham5Δnor1/fus 47.0 ± 32.9 71.7 ± 18.7
Δnor1/fus 10.7 ± 6.7 23.3 ± 12.7
Δnox1/fus 24.7 ± 15.8 16.7 ± 6.4
Δnox2/fus 165.0 ± 63.6 170.3 ± 93.1

IX. Supplements 96
Figure S 5: The deletion strain Δste12/fus is not impaired in hyphal fusion. For investigation of hyphal fusions, the strain Δste12/fus was grown on a layer of cellophane on MMS for two days. Hyphal fusion events can be observed and are indicated with arrowheads. The scale bar indicates 10 μm.

X. Curriculum Vitae 97
X. Curriculum Vitae
Personal data
Name Sarah Schmidt
Date of birth 17.11.1990
Place of birth Bochum
Citizenship German
Academic education
Since 10/2016 Graduation at Ruhr-University Bochum,
Department of General and Molecular Botany: “Interaction of
conserved signaling pathways during cellular development in
S. macrospora”
Supervisor: Prof. Dr. U. Kück,
09/2014 – 08/2016 Master of Science in Biomedical Sciences at Maastricht
University, Maastricht, the Netherlands
Department of Surgery: “The role of digestive enzymes in
human intestinal ischemia-reperfusion”
Department of Toxicology: “Influence of oxidized quercetin on
apoptosis induction in cells”
09/2013 – 08/2014 Pre-master in Medicine at UMC St. Radboud, Nijmegen, the
Netherlands
09/2010 – 08/2013 Bachelor of Science in Biology at Radboud University,
Nijmegen, the Netherlands
Department of Pathology: “Obstructed intestinal motility due to
amyloidosis”
School education
09/2001 – 08/2010 Graf-Engelbert Gymnasium, Bochum
Graduation qualifying for university admission (Abitur)

X. Curriculum Vitae 98
Congress contributions
Schmidt S, Märker R, Beier A, Teichert I, Kück U (2019) HAM5 mediates the interaction
of the pheromone response pathway and NOX1 complex during fungal development. 1st
joint meeting with the GeneAG “Fungal Genetics” of the German Genetics Society,
Göttingen, Germany, Poster
Schmidt S, Teichert I, Kück U (2018) Time and development dependent localization of the
STRIPAK complex. Annual Conference of the Association for General and Applied
Microbiology (VAAM), Wolfsburg, Germany, Poster
Schmidt S, Beier A, Teichert I, Kück U (2017) Cellular localization of components of the
STRIPAK signaling complex by fluorescence microscopy. Annual Conference of the
German Genetics Society (GfG), Bochum, Germany, Poster
Schmidt S, Beier A, Teichert I, Kück U (2017) Cellular localization of components of the
STRIPAK signaling complex by fluorescence microscopy. Novel tools to investigate cellular
physiology at the nanoscale, Bochum, Germany, Poster

XI. Erklärung 99
XI. Erklärung
Ich versichere an Eides statt, dass ich die eingereichte Dissertation selbstständig und ohne
unzulässige fremde Hilfe verfasst, andere als die in ihr angegebene Literatur nicht benutzt
und dass ich alle ganz oder annähernd übernommenen Textstellen sowie verwendete
Grafiken und Tabellen kenntlich gemacht habe. Weiterhin erkläre ich, dass digitale
Abbildungen nur die originalen Daten enthalten oder eine eindeutige Dokumentation von
Art und Umfang der inhaltsverändernden Bildbearbeitung vorliegt. Außerdem versichere
ich, dass es sich bei der von mir vorgelegten Dissertation (elektronische und gedruckte
Version) um völlig übereinstimmende Exemplare handelt und die Dissertation in dieser oder
ähnlicher Form noch nicht anderweitig als Promotionsleistung vorgelegt und bewertet
wurde.
Es wurden keine anderen als die angegebenen Hilfsmittel verwendet. Die Dissertation
wurde gemäß der Promotionsordnung und der Betreuungsvereinbarung angefertigt.
Bochum, den 01.07.2020
_____________________________________
(Unterschrift)


















