
Integrated Effects of Matrix Mechanics and Vascular Endothelial Growth Factor (VEGF) on Capillary...
13
Integrated Effects of Matrix Mechanics and Vascular Endothelial Growth Factor (VEGF) on Capillary Sprouting YANG WU, 1 MOHAMMAD ALI AL-AMEEN, 2 and GARGI GHOSH 2 1 Environmental Science, University of Michigan, Dearborn, 4901 Evergreen Road, Dearborn, MI 48128, USA; and 2 Bioengineering Program, Department of Mechanical Engineering, University of Michigan, Dearborn, 4901 Evergreen Road, Dearborn, MI 48128, USA (Received 10 October 2013; accepted 6 February 2014) Associate Editor K. A. Athanasiou oversaw the review of this article. Abstract—Angiogenesis is the growth of new capillaries from existing vasculature. Vascular network formation is known to be regulated by the biophysical and biochemical signals emanating from the microenvironment. However, it is not clear how endothelial cells integrate these signals to drive the capillary morphogenesis. In this study, human umbilical endothelial cells (HUVECs) were seeded on scaffolds with varying stiffness and capillary formation was quantified. Our study revealed that cells formed well defined networks on compliant gels. Increase in stiffness resulted in a decrease in sprouting. VEGF was encapsulated within the scaffolds at concentrations ranging from 0 to 100 ng/mL to investigate the interplay between VEGF concentration profiles and matrix stiffness in guiding network formation. Quantitative analysis revealed that while VEGF disrupted sprout forma- tion on compliant substrates, it elicited a sprouting response in cells seeded on scaffolds of intermediate rigidity. Addi- tionally, we also observed that a minimum cell density is required for the formation of well-connected vascular net- works. Keywords—Matrix rigidity/compliance, VEGF, Angiogenesis, Gelatin methacrylate. INTRODUCTION Angiogenesis, i.e., the growth of new capillaries from existing vasculature is necessary for physiological con- ditions including embryonic development, wound healing, and reproduction. 13 Dysregulated angiogenesis is a hallmark in many diseases including cancer, psori- asis, arthritis, osteoporosis, and cardiovascular dis- eases. 12 A key feature common to all these diseases is the alteration of tissue microenvironment which in turn may affect the neovascularization by up regulating or down regulating vascular morphogenesis. 7 Understanding the dependence of angiogenesis on microenvironmental cues may play a pivotal role in identification of thera- peutic interventions for controlling these diseases. In addition, in the field of tissue engineering, the con- struction of thick and complex tissues such as heart, liver, kidney, and lung has still remained a distant milestone due to the ischemic cell death resulting from the paucity of oxygen and nutrients required for cell metabolism. 3 In vivo, most cells are found within 100– 200 lm from the nearest capillary, which correlates with passive oxygen diffusion limit. 16 The challenge related to the construction of these complex tissues can only be addressed by promoting vascular networks within the bioengineered constructs. Without such a network, cell proliferation is restricted to the periphery of the engi- neered tissue construct. 29 Mounting evidence has revealed that the capillary sprouting is orchestrated via integrated interactions between the endothelial cells, growth factors including vascular endothelial growth factor (VEGF), and extracellular matrix (ECM). In vitro investigations have found cell–matrix interactions and ECM degra- dation and deposition serve a critical role in capillary formation. 21 Cellular sensory machineries sense, ana- lyze, and integrate the microenvironmental cues, par- ticularly, ECM compliance, topography, and adhesive ligand density to influence the endothelial cell behav- iors as manifested in altered cell shape, increased proliferation, focal adhesions, and cell generated con- tractile forces. 20 This suggests that understanding and harnessing the bidirectional crosstalk between endo- thelial cells and ECM is pivotal to stimulate/inhibit the formation of capillary networks. Address correspondence to Gargi Ghosh, Bioengineering Pro- gram, Department of Mechanical Engineering, University of Mich- igan, Dearborn, 4901 Evergreen Road, Dearborn, MI 48128, USA. Electronic mail: [email protected] Annals of Biomedical Engineering (ȑ 2014) DOI: 10.1007/s10439-014-0987-7 ȑ 2014 Biomedical Engineering Society
Transcript of Integrated Effects of Matrix Mechanics and Vascular Endothelial Growth Factor (VEGF) on Capillary...

Integrated Effects of Matrix Mechanics and Vascular Endothelial Growth
Factor (VEGF) on Capillary Sprouting
YANG WU,1 MOHAMMAD ALI AL-AMEEN,2 and GARGI GHOSH2
1Environmental Science, University of Michigan, Dearborn, 4901 Evergreen Road, Dearborn, MI 48128, USA; and2Bioengineering Program, Department of Mechanical Engineering, University of Michigan, Dearborn, 4901 Evergreen Road,
Dearborn, MI 48128, USA
(Received 10 October 2013; accepted 6 February 2014)
Associate Editor K. A. Athanasiou oversaw the review of this article.
Abstract—Angiogenesis is the growth of new capillaries fromexisting vasculature. Vascular network formation is knownto be regulated by the biophysical and biochemical signalsemanating from the microenvironment. However, it is notclear how endothelial cells integrate these signals to drive thecapillary morphogenesis. In this study, human umbilicalendothelial cells (HUVECs) were seeded on scaffolds withvarying stiffness and capillary formation was quantified. Ourstudy revealed that cells formed well defined networks oncompliant gels. Increase in stiffness resulted in a decrease insprouting. VEGF was encapsulated within the scaffolds atconcentrations ranging from 0 to 100 ng/mL to investigatethe interplay between VEGF concentration profiles andmatrix stiffness in guiding network formation. Quantitativeanalysis revealed that while VEGF disrupted sprout forma-tion on compliant substrates, it elicited a sprouting responsein cells seeded on scaffolds of intermediate rigidity. Addi-tionally, we also observed that a minimum cell density isrequired for the formation of well-connected vascular net-works.
Keywords—Matrix rigidity/compliance, VEGF,
Angiogenesis, Gelatin methacrylate.
INTRODUCTION
Angiogenesis, i.e., the growth of new capillaries fromexisting vasculature is necessary for physiological con-ditions including embryonic development, woundhealing, and reproduction.13 Dysregulated angiogenesisis a hallmark in many diseases including cancer, psori-asis, arthritis, osteoporosis, and cardiovascular dis-eases.12 A key feature common to all these diseases is the
alteration of tissuemicroenvironmentwhich in turnmayaffect the neovascularization by up regulating or downregulating vascular morphogenesis.7 Understanding thedependence of angiogenesis on microenvironmentalcues may play a pivotal role in identification of thera-peutic interventions for controlling these diseases. Inaddition, in the field of tissue engineering, the con-struction of thick and complex tissues such as heart,liver, kidney, and lung has still remained a distantmilestone due to the ischemic cell death resulting fromthe paucity of oxygen and nutrients required for cellmetabolism.3 In vivo, most cells are found within 100–200 lm from the nearest capillary, which correlates withpassive oxygendiffusion limit.16 The challenge related tothe construction of these complex tissues can only beaddressed by promoting vascular networks within thebioengineered constructs. Without such a network, cellproliferation is restricted to the periphery of the engi-neered tissue construct.29
Mounting evidence has revealed that the capillarysprouting is orchestrated via integrated interactionsbetween the endothelial cells, growth factors includingvascular endothelial growth factor (VEGF), andextracellular matrix (ECM). In vitro investigationshave found cell–matrix interactions and ECM degra-dation and deposition serve a critical role in capillaryformation.21 Cellular sensory machineries sense, ana-lyze, and integrate the microenvironmental cues, par-ticularly, ECM compliance, topography, and adhesiveligand density to influence the endothelial cell behav-iors as manifested in altered cell shape, increasedproliferation, focal adhesions, and cell generated con-tractile forces.20 This suggests that understanding andharnessing the bidirectional crosstalk between endo-thelial cells and ECM is pivotal to stimulate/inhibit theformation of capillary networks.
Address correspondence to Gargi Ghosh, Bioengineering Pro-
gram, Department of Mechanical Engineering, University of Mich-
igan, Dearborn, 4901 Evergreen Road, Dearborn, MI 48128, USA.
Electronic mail: [email protected]
Annals of Biomedical Engineering (� 2014)
DOI: 10.1007/s10439-014-0987-7
� 2014 Biomedical Engineering Society

Attempts undertaken to elucidate the role of matrixmechanics in angiogenesis have demonstrated thatenhancing the matrix stiffness restricts the formation ofvascular network.5,17Although these systems have helpedto advance our understanding of mechano-responsivecellular behavior, they suffer from several limitations.Mainly due to the fact that manipulation of rigidity ofthese matrices (naturally derived materials such as colla-gen, fibrin, andmatrigel8,24,38 or synthetic ECManaloguesuch as polyacrylamide gels11) by varying the proteinconcentration or the cross-linker density has been asso-ciated with alteration in other ECM properties such aspore size and porosity.32 As a result, increasedmechanicalproperty of the matrix is associated with restricted diffu-sional transport of putative angiogenic factors which inturn can play an important role in the regulation of cap-illary morphogenesis.17 This underscores the importanceof decoupling different intertwined ECM properties e.g.,the pore characteristics from matrix compliance to parsethe specific contributions of these micro-environmentalcues in stimulating angiogenesis.
In this study, we investigated the integrated effects ofVEGF and matrix mechanics on endothelial cell mor-phology and capillary formation. Specifically, humanumbilical vein endothelial cells (HUVECs) were seeded onpoly (ethylene glycol) diacrylate (PEGDA) and gelatinmethacrylate (GelMA) composite scaffolds. Rigidity ofthe matrices was manipulated independent of the scaffoldporosity and consequently, of the macromolecule release.GelMA, obtained via conjugation of methacrylate groupsto gelatin, provides the cell binding motifs e.g., RGD aswell as matrix metalloproteinase (MMP) sensitive degra-dation groups.33 VEGF was encapsulated within thescaffolds at different concentrations to evaluate howendothelial cells integrate the biophysical and biochemicalsignals to direct the capillary morphogenesis. Our resultsindicate that when the substrate compliance was increasedfrom 11 to 36 kPa, no significant difference in capillarynetwork formation was observed. Further increase inhydrogel rigidity to 78 kPa reduced network assemblysignificantly.To investigate the integrated effects ofVEGFand matrix compliances, cells were seeded on scaffoldswith compliances varying from soft (11 kPa), intermediate(36 kPa), and stiff (78 kPa). Our study revealed thatVEGF disrupted sprout formation on soft gels. However,on stiffer gels, VEGF elicited a stimulatory response.
MATERIALS AND METHODS
Materials
Poly (ethylene) glycol diacrylate 575 (PEG575DA),poly (ethylene) glycol diacrylate 6000 (PEG6kDA),phosphate buffer saline (PBS), gelatin fromporcine skin,
fluorescein isothiocyanate—dextran (FITC-Dextran70 kDa), photo-initiator (2-Hydroxy-4¢-(2-hydroxy-ethoxy)-2-methylpropionphenone), ethylene glycol, andmethacrylic anhydride were procured from SigmaAldrich (St. Louis, MO). Poly (ethylene) glycol diacry-late 10,000 (PEG10kDA)was obtained fromLaysanBioInc. (Alabama). Vascular endothelial growth factor(VEGF) was purchased from R&D systems (Minneap-olis, MN) and collagenase from Worthington (Lake-wood, NJ).
Synthesis of Methacrylated Gelatin
Gelatin methacrylate was synthesized as describedelsewhere.31 Briefly, gelatin type A (gelatin from por-cine skin) was added into Dulbecco’s phosphate buf-fered saline (DPBS) and stirred continuously at 50 �Cuntil fully dissolved to prepare 10% (w/v) solution.Methacrylation was achieved by adding 10% (w/v) ofmethacrylic anhydride at a rate 0.5 mL/min andreacting at 60 �C for 1 h. Following a 59 dilution withwarm DPBS (50 �C), the mixture was dialyzed usingSlide-A-lyzer against distilled water at 50 �C for15 days to remove salts and acid. The sample was thenfreeze dried and GelMA was generated as porous foamand stored at 280 �C until further use.
Fabrication and Characterization of Scaffolds
The scaffolds were fabricated by adding 125 lL ofthe pre-polymer solution consisting of PEGDA(PEG575DA, PEG6kDA or PEG10kDA), 5% GelMAand 1% photoinitiator to each well of 48 well platesand then exposing the plates to UV (CL-1000 UVCrosslinker (UVP), 365 nm) for 5 min. The concen-tration of PEGDA was varied from 5 to 15%. Tofabricate VEGF impregnated scaffolds, 25 lL ofVEGF of varying concentrations was added to the pre-polymer solution.
Mechanical Properties
To characterize the mechanical properties of thehydrogels, cylindrical samples of diameter 10 mm werefabricated and compressed along the thickness tomeasure stiffness. The polymer precursor solutionswere placed in 10 mm diameters cylindrical mold(~3 mm in thickness,) and UV polymerized. The cyl-inders were then incubated in PBS for 72 h to facilitateremoval of the unreacted monomers. Prior to analysis,excess fluids on the surface of the hydrated sampleswere removed by dabbing with Kimwipe. The com-pressive tests were carried out by using uniaxial testingmachine (TestResources, USA) at a loading rate of1.2 mm/min with a precision load up to 9 N. At least 3
WU et al.

samples per group were used for each test. Maximumstrain and stress at the moment of fracture wasrecorded and compression modulus was calculatedfrom the initial 10% compression.
Swelling Ratio
For measuring the mass swelling ratio (Qm), thehydrogels were incubated in PBS for 72 h in a 37 �Cshaker (50 rpm). Following incubation, excess fluidswere removed from the swollen hydrogels andweights ofthe samples (Wwet) were measured. The samples werethen dried at 60 �C for 24 h and the dry weights weremeasured (Wdry). Qm of the hydrogel was calculatedfrom the equation:Qm = Wwet/Wdry.
19 The average andstandard deviation of three independent experimentseach carried out in triplicate were calculated.
Degradation Ratio
To assess the degradation of the hydrogels, theweight of the hydrogel disks were measured followingfabrication (day 0). The samples were then incubatedwith 2.5 unit/mL of collagenase solution in a 37 �Cshaker (50 rpm) and the weights were measured everyday. Degradation of the hydrogels was then estimatedfrom the ratio of weights at different time points to theweights on the day of fabrication.23
Porosity
To measure the porosity of hydrogels, liquiddisplacement method was used.30 Briefly, the scaffoldswere dried over night at 60 �C and weighted(Wscaffolds). Following which the scaffolds wereimmersed in graduated cylinders containing knownvolume (V1) of distilled water for 2 h. Followingwhich, the total volume of water and water impreg-nated scaffolds were recorded as V2. The impregnatedscaffolds were removed and the weight (Wscaffolds+water)of the hydrated scaffolds were measured. Volume ofthe scaffolds (Vscaffolds) was calculated from thevolume difference (V2 2 V1). Volume of water whichcorresponds to the void volume within the scaffoldwas calculated from the difference between theweight of the hydrated scaffold and the initial weight.The porosity of the scaffold was obtained from theratio of volume of water (Vwater) to the volume ofthe scaffold (Vscaffolds). Porosity = Vwater/Vscaffolds =
((Wscaffolds+water 2 Wscaffolds)/qwater)/Vscaffolds.
Scanning Electron Microscopy
The hydrogel disks were made in 48 well plates.Following which the hydrogels were incubated in waterand allowed to swell overnight. The samples were thentreated in liquid nitrogen for 5 min and freeze dried,37
coated with gold using a sputter coater and thenimaged using a scanning electron microscope (HitachiS-2600 N). The pore size was measured manually fromthe SEM images using the Quartz PCI Image man-agement system software (Quartz Imaging Corpora-tion, Vancouver, Canada).
Diffusion
To estimate the release kinetics of proteins from thehydrogels, the scaffolds were fabricated by incorpo-rating FITC-dextran 70 kDa within the precursorsolution. The concentration of dextran within eachsample was maintained at 50 lg/mL. After photo-polymerization, the hydrogels were washed with PBS.To facilitate the release of dextran from the scaffolds,the hydrogels were incubated in PBS at 37 �C. Over aperiod of 6 days, half of PBS was collected every 24 hand replaced with fresh buffer. The withdrawn bufferwas stored at 4 �C. All the collected samples wereanalyzed using PerkinElmer LS55 fluorescence spec-trometer to assess release of FITC.
Cell Culture and Angiogenesis Assay
HUVECs were purchased from American Type Cul-tureCollection (ATCC,Manassas,VA) and expanded invascular basal medium with endothelial growth supple-ment (Ascorbic, fetal bovine serum (FBS), rh EGF,heparin sulfate, L-glutamine, hydrocortisone, bovinebrain extract) and 1% (v/v) penicillin streptomycin. Cellsup to passages 6 were used in this study.42
Morphology
HUVECs were dissociated from flask using 0.25%Trypsin–EDTA and suspended in cell growth media.Cells at a 1.0 9 104 cells/cm2 density were seeded ondifferent scaffolds and the cell morphology was mea-sured 72 h after seeding. For this purpose, images ofthe cells were captured to measure morphology usingZeiss Axio Observer A1 with integrated CCD camera.At least five random images were captured for eachscaffold. Area and aspect ratio (width/length of cells)of the cells in each image were calculated.9 Experi-ments were performed in three replicates. To investi-gate the effect of VEGF concentration on cellmorphology, HUVECs were seeded on 36 kPa scaf-folds containing 0, 25, 50, 75, 100 and 150 (ng/mL) ofVEGF and images captured after 72 h seeding.
Angiogenesis Assay
To determinate the effect of cell seeding density onsprouting, initially HUVECs were plated on 11 and36 kPa scaffold at seeding density ranging from 2.5 9 104
Integrated Effects of Matrix Mechanics and VEGF

to 7.5 9 104 cells/cm2. To monitor and quantifysprouting, images were taken after 72 h. At least 5 imageswere captured for each scaffold. Experiments were per-formed in three replicates. Capillary formation wasquantified by manually counting the number and lengthsof sprouts branching out from a branch point/node.
In order to investigate the effect of matrix mechanicson capillary formation, 5 9 104 cells/cm2 were seededon scaffolds of varying compliances. At least 5 imageswere captured for each scaffold 3 days after seeding.Experiments were performed in three replicates. Toevaluate the integrated effects of VEGF and matrixmechanics on sprouting, cells were seeded at a densityof 5 9 104 cells/cm2 on compliant (11 kPa), interme-diate (36 kPa), and rigid (78 kPa) scaffolds encapsu-lating 0, 25, 50, and 100 ng/mL of VEGF. Sproutingwas quantified after 72 h. Experiments were performedin three replicates.
VEGF Release
To measure the concentration of VEGF released inthe media, 36 kPa scaffolds encapsulating 0, 25, 50,and 100 ng/mL were incubated in 500 lL of media at37 �C. Media was collected every 24 h and replacedwith equal volume of fresh media. The cumulativerelease of VEGF in media was measured using VEGFELISA kit (R&D systems, Minneapolis, MN).
Statistical Analysis
For all experiments, data reported as mean ± SEMof three independent experiments. Statistical analyseswere carried out with one way ANOVA. Differencesbetween two sets of data were considered significant atp value< 0.05.
RESULTS
Mechanical microenvironment regulates the signaltransduction in endothelial cells and thereby controlsthe capillary morphogenesis. Since different ECMproperties are intertwined, when ECM stiffens, otherbiophysical properties including pore size and porosityare altered. This in turn directly impacts the interactionbetween the migrating cells and the matrix by affectingthe availability of growth factors. Therefore, the abilityto disentangle the porosity from hydrogel modulus iscrucial for parsing the contribution of matrixmechanics on capillary sprouting.
Mechanically Tunable Substrate for HUVEC Sprouting
The structural and mechanical properties of thehydrogels can be controlled by varying the concentration
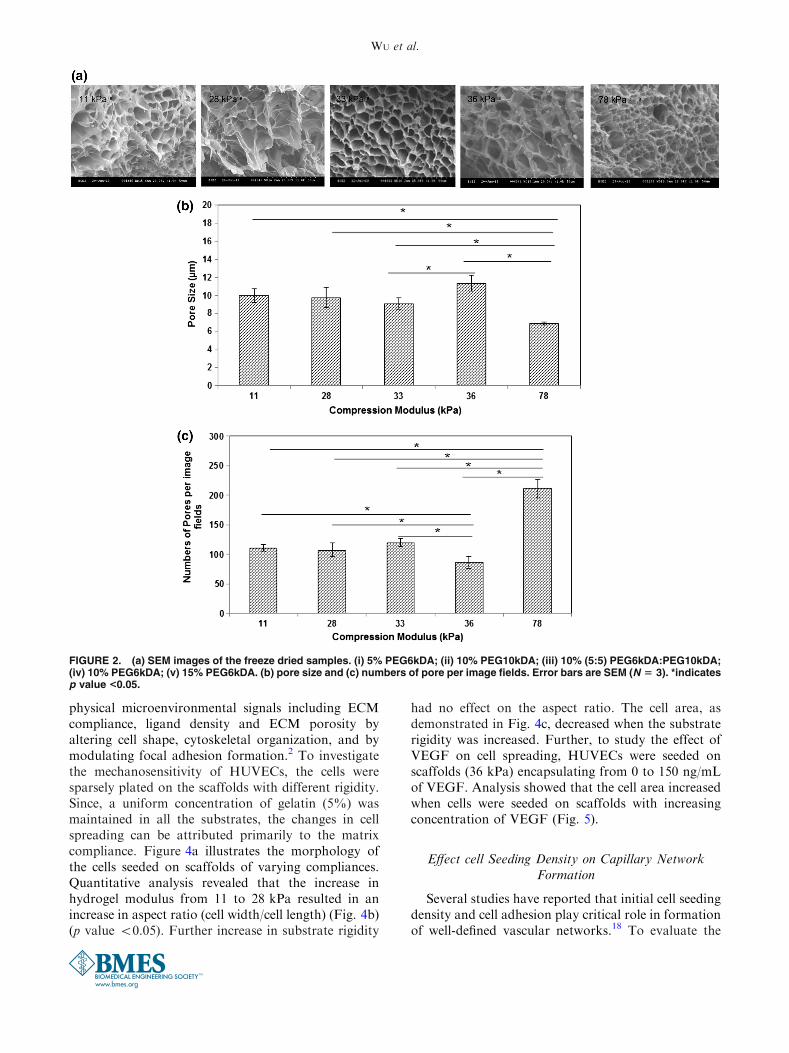
and molecular weight of PEGDA during polymeriza-tion. In this study, to tune the matrix stiffness inde-pendent of pore characteristics, scaffolds werefabricated by varying the concentrations of PEGDAfrom 5 to 15%, by altering the molecular weight of thePEGDA precursors from 575 to 10 k, as well as byblending PEGDA of different molecular weight whilemaintaining uniform presentation of cell adhesionmolecules (i.e., constant gelatin concentration)(Table 1). As demonstrated in Fig. 1a, alterations inthe potential crosslinking groups per volume signifi-cantly increased the hydrogel modulus from 7.4 ± 1.33to 287.2 ± 1.3 kPa. Several hydrogel formulationswere then identified in which the modulation of cross-linking density had minimal effect on the porosity ofthe hydrogel. The scatter plot of compressive modulusvs. porosity (Fig. 1b) illustrates the ability to decouplethe hydrogel modulus from the porosity of the scaf-folds. We next investigated the pore characteristics ofthese scaffolds. Figure 2a displays the SEM images ofthe freeze dried scaffolds. The pore size and the numberof pores were measured manually (Figs. 2b, 2c). Nosignificant differences were observed in pore diameterof the scaffolds (p value <0.05) when the compressivemodulus of the hydrogels were increased from 11 kPa(10.0 ± 0.73 lm) to 33 kPa (9.1 ± 0.63 lm). However,for the hydrogel with a modulus of 36 kPa, pores withbigger diameter (11.4 ± 0.86 lm) were observed. Thiswas accompanied with reduced number of pores. Onthe other hand, in case of hydrogel with modulus78 kPa, smaller pore diameter (6.9 ± 0.17 lm) andenhanced number of pores were observed. In addition,to functionally determine the macromolecular releasefrom these scaffolds, hydrogels were fabricated withentrapped FITC-conjugated dextran. Figure 3a illus-trates the cumulative release of dextran from thesescaffolds over a time span of 6 days. As anticipated, nosignificant difference was observed in the release profileof dextran from all of these scaffolds (p value >0.05).
To determine the stability of scaffolds, hydrogelmatrices were incubated with PBS at 37 �C and deg-radation over time was measured. As hydrogelsdegrade over time, bonds are broken which in turnfacilitates diffusion of more aqueous solution inside thegel. Thus, increased swollen weight of the hydrogelindicates degradation of the gel. As demonstrated inFig. 3b, swollen weight increased until day 15. Beyondthat, for all the different scaffolds swollen weightdecreased indicating the breakdown of polymeric net-work during which loss of polymers exceeds diffusionof aqueous solution inside the scaffold. However, allthe matrices retained their physical integrity duringthis time period. Taken together the studies demon-strate that scaffolds with varying compressive moduliindependent of pore size, porosity, and release of
WU et al.

macromolecules were fabricated. Since, the concen-tration of GelMA was maintained constant (5%), thechanges observed in cellular behavior can be correlatedto the matrix mechanics only.
Effect of Matrix Compliance on Cell Spreading
Numerous studies have demonstrated that the cellsrespond to the biochemical cues as well as the bio-
TABLE 1. Composition and compression moduli of 12 samples at constant GelMa concentration of 5%.
Composition Polymer concentration (%) PEG575DA (%) PEG6kDA (%) PEG10kDA (%) Compression modulus (kPa)
1 5 5 – – 21.8
2 5 – 5 – 11.4
3 5 – – 5 7.6
4 10 10 – – 116.1
5 10 – 10 – 35.8
6 10 – – 10 27.9
7 10 5 5 – 84.1
8 10 5 – 5 47.6
9 10 – 5 5 32.5
10 15 15 – – 287.2
11 15 – 15 – 78.1
12 15 – – 15 40.6
0
50
100
150
200
250
300
350
1 2 3 4 5 6 7 8 9 10 11 12
Co
mp
ress
ion
Mo
du
lus
(kP
a)
Polymer compositions
0.5
0.6
0.7
0.8
0.9
1
0 20 40 60 80 100
Po
rosi
ty
Compression Modulus (kPa)
(a)
(b)
FIGURE 1. (a) Effect of PEGDA concentration and molecular weight on compression modulus and (b) correlation between matrixcompliance and porosity of PEGDA scaffolds. Error bars are SEM (N 5 3). The compression moduli of all the samples werestatistically different.
Integrated Effects of Matrix Mechanics and VEGF
physical microenvironmental signals including ECMcompliance, ligand density and ECM porosity byaltering cell shape, cytoskeletal organization, and bymodulating focal adhesion formation.2 To investigatethe mechanosensitivity of HUVECs, the cells weresparsely plated on the scaffolds with different rigidity.Since, a uniform concentration of gelatin (5%) wasmaintained in all the substrates, the changes in cellspreading can be attributed primarily to the matrixcompliance. Figure 4a illustrates the morphology ofthe cells seeded on scaffolds of varying compliances.Quantitative analysis revealed that the increase inhydrogel modulus from 11 to 28 kPa resulted in anincrease in aspect ratio (cell width/cell length) (Fig. 4b)(p value <0.05). Further increase in substrate rigidity
had no effect on the aspect ratio. The cell area, asdemonstrated in Fig. 4c, decreased when the substraterigidity was increased. Further, to study the effect ofVEGF on cell spreading, HUVECs were seeded onscaffolds (36 kPa) encapsulating from 0 to 150 ng/mLof VEGF. Analysis showed that the cell area increasedwhen cells were seeded on scaffolds with increasingconcentration of VEGF (Fig. 5).
Effect cell Seeding Density on Capillary NetworkFormation
Several studies have reported that initial cell seedingdensity and cell adhesion play critical role in formationof well-defined vascular networks.18 To evaluate the
FIGURE 2. (a) SEM images of the freeze dried samples. (i) 5% PEG6kDA; (ii) 10% PEG10kDA; (iii) 10% (5:5) PEG6kDA:PEG10kDA;(iv) 10% PEG6kDA; (v) 15% PEG6kDA. (b) pore size and (c) numbers of pore per image fields. Error bars are SEM (N 5 3). *indicatesp value <0.05.
WU et al.

effect of initial cell seeding density on capillarysprouting, cells with densities varying from 2.5 9 104
to 7.5 9 104 cells/cm2 were seeded on the hydrogel(11 kPa and 36 kPa) and incubated at 37 �C and 5%CO2 in a humidified chamber. As demonstrated inFig. 6a, at a seeding density of 2.5 9 104 cells/cm2,most of the cells condense into islands of cells tomaximize the cell–cell contact. As the initial seedingdensity was increased to 5 9 104 cells/cm2 (Fig. 6b),formation of capillary networks was observed. How-ever, with further increase in seeding density to7.5 9 104 cells/cm2, HUVECs formed cell clusterrather than capillary structures over hydrogel surface(Fig. 6c). At a density lower than 2.5 9 104 cells/cm2,no capillary like structures were observed. The cellseeding density of 5 9 104 cells/cm2 also resulted in theformation of well-connected capillary networks on36 kPa scaffolds (Fig. 6d). Since, with initial cell den-
sity of 5 9 104 cells/cm2 maximum sprouting wasobserved, henceforth all the experiments were carriedout with this cell density.
Effects of Matrix Rigidity and VEGF on CapillaryNetwork Formation
In order to investigate the interplay between matrixrigidity and vascular sprouting, HUVECs at a densityof 5 9 104 cells/cm2 were seeded on different scaffoldswith compression modulus ranging from 11 to 78 kPa.72 h after plating, images were captured and thenumber of sprouts was manually calculated. Figure 7ademonstrates the SEM images of capillary networks.Capillaries or tubes emanating from a branch pointleading to the formation of polygonal structures wereconsidered as sprouts. To examine the effect of matrixrigidity on sprout formation, number of sprouts
FIGURE 3. (a) Comparison of diffusion of FITC-Dextran 70 kDa from the scaffolds over a span of 6 days. (b) Degradation ofPEGDA scaffolds monitored over 15 days. Error bars are SEM (N 5 3).
Integrated Effects of Matrix Mechanics and VEGF

formed on the scaffolds of varying compliances wasmeasured. As demonstrated in Fig. 7b, no significantdifference in number of sprouts were observed whenHUVECs were seeded on matrices with rigidity rang-ing from 11 to 36 kPa (p value >0.05). On the otherhand, network assembly of HUVECs was attenuatedwhen the matrix modulus was increased to 78 kPa (pvalue <0.05).
VEGF is known to stimulate angiogenic responsesby facilitating cell proliferation, migration, and capil-
lary network formation.25 However, it is not knownwhether VEGF and matrix mechanics act indepen-dently or in concert to induce capillary morphogenesis.To characterize the capillary network formation as afunction of VEGF release and matrix stiffness, HU-VECs were seeded on soft (11 kPa), intermediate(36 kPa) and stiff (78 kPa) scaffolds. Increasing theconcentrations of encapsulated VEGF from 0 to100 ng/mL displayed a sustained release of 0, 230, 350,and 915 pg/mL of VEGF in media from 0, 25, 50 and
FIGURE 4. HUVECs were seeded on scaffolds of varying stiffness. (a) Images of HUVECs cultured on scaffolds of varyingcompliances. Comparison of the effect matrix mechanics on HUVECs cells morphology as measured by (b) aspect ratio (c) cellarea. At least five random images for each scaffold were captured. The experiments were performed in three replicates. Cell areaand aspect ratio of the cells in each image were calculated. Error bars are SEM (N 5 3). *indicates p value < 0.05.
WU et al.

100 ng/mL scaffolds respectively. Figure 7c demon-strates how stimulation of HUVECs by VEGF affectsnetwork formation. Presentation of VEGF had no
effect on capillary morphogenesis when cells wereseeded on soft gels. However, when cells were platedon scaffolds of intermediate stiffness, VEGF elicited a
0
200
400
600
800
1000
1200
1400
0 25 50 75 100 150
Cel
l are
a p
er im
age
fiel
d (
µm
²)
VEGF concentration (ng/ml)
* **
**
*
FIGURE 5. HUVECs were seeded on scaffolds encapsulating VEGF at concentrations ranging from 0 to 150 ng/mL and cellmorphology was evaluated. Comparison of the effect of VEGF on (a) aspect ratio (b) cell area. At least five random images for eachscaffold were captured. The experiments were performed in three replicates. Cell area and aspect ratio of the cells in each imagewere calculated. Error bars are SEM (N 5 3). *Indicates p value <0.05.
FIGURE 6. Effect of seeding HUVECs at a density of (a) 2.5 3 104 cells/cm2; (b) 5 3 104 cells/cm2 and (c) 7.5 3 104 cells/cm2 on11 kPa scaffolds was compared. (d) Sprouting when HUVECs seeded at 5 3 104 cells/cm2 on 36 kPa scaffolds. Arrows indicate thesprouts or capillaries branching out from a node point.
Integrated Effects of Matrix Mechanics and VEGF

sprouting response and the network formationincreased in response to increased concentration ofVEGF released in the media. In case of stiff gels, eventhough network assembly increased in the presence ofVEGF, the response was less dramatic.
DISCUSSION
As a major component of tissue microenvironment,ECM plays a complex and multivariate role in regu-
lating capillary morphogenesis. Mounting evidencecorroborates that in addition to the biochemical cues,the mechanical signals emanating from the ECMdirectly impact tubulogenesis.4,5,24 Manipulation ofmatrix rigidity with naturally derived materials isassociated with the alteration in the availability ofligands facilitating cell adhesion, the protease suscep-tibility, and matrix architecture including altered porecharacteristics.36 This in turn can lead to a change inthe availability of growth factors which guide and
FIGURE 7. (a) SEM images of capillary network formation when HUVECs were cultured on scaffolds of varying stiffness. (b)Comparison of the effect of compliance on sprout number. (c) Integrated effects of VEGF and matrix mechanics on sprout number.At least five random images for each scaffold were captured. The experiments were performed in three replicates. Sprout numberin each image was calculated. Error bars are SEM (N 5 3). Arrows indicate the sprouts or capillaries branching out from a nodepoint. *indicates p value < 0.05.
WU et al.

regulate the process of capillary network formation.Therefore, to deconstruct the multivariate instructiverole of ECM in regulating tubulogenesis, syntheticpolymers have been utilized.4,24,36 These studiesestablished that cells sense and accordingly respond tothe mechanical resistivity of the substrate by alteringthe cell-substrate interactions and cytoskeletal orga-nizations. On the cellular level, the modulated cell-substrate interactions have a decisive impact on vari-ous cellular functions including cell spreading, migra-tion, and consequently capillary network formation. Abody of evidence has demonstrated that endothelialcells adopt an elongated spindle-shaped morphologyon compliant surfaces. However, on stiffer substratesthese cells exhibit a more isotropic spreading.27
Characteristics such as cell size and elongation reflectthe propensity of endothelial cells to organize intovascular networks. Reinhart King and co-workersdemonstrated that bovine aortic endothelial cellsorchestrate capillary formation in the presence ofadhesive and mechanical cues of the matrices.5 Inaddition, prior studies have also demonstrated thatnetwork formation by HUVEC is also dependent onsubstrate compliance and cell density.6 These studies,therefore, have established that the mechanical tensionbetween the cells and the matrices is sufficient totrigger intercellular signaling cascade leading tomigration of cells and eventually the vasculature for-mation. In this study we have fabricated scaffolds withrigidity varying from 11 to 78 kPa. This matrix rigiditycovers a broad range of stiffness reported for differenthuman tissue including adipose tissue, breast tissue,and stromal tissue as well as cancer tissue,14,15,39
thereby highlighting the physiological relevance ofthese matrices. When cultured on compliant scaffold,HUVECs exhibited elongated spindle shaped mor-phology. The compliant gels facilitated the cell–celladhesion via stimulating the growth of cells with longprocesses. This also explains why more capillary net-works were formed on compliant gels. On the otherhand, on intermediate or rigid substrates, the cellsdisplayed a more rounded morphology. The weakercell–cell contacts reduced the ability to form sproutson stiffer substrates. Prior studies have reported en-hanced endothelial cell spread area with increasedmatrix stiffness.5,10 In contrast, we observed a decreasein cell spread area with increasing matrix stiffness.These observations are however, in agreement with thefindings reported by Wood and co-workers.35 Thisdiscrepancy in our observations from others may bedue to utilization of different endothelial cells. As amatter of fact, Wood and co-workers demonstratedthat the substrate compliance has different impact oncell spreading when endothelial cells from differentanatomical sites were used.
Formation of adhesions due to clustering of inte-grins to ECM ligands leads to the recruitment ofgrowth factor receptors within the focal adhe-sions.28,41 VEGF signal synergize with integrins toactivate Rho-ERK pathway which in turn facilitatesin modulating angiogenesis.34 A recent analysis ofcellular mechanotransduction revealed that matrixrigidity can regulate the balance of activities of twoantagonistic transcription factors TFII-I and GATA2which in turn govern the expression of VEGFR2.22
Studies investigating the responsiveness of endothelialcells to VEGF mediated spouting suggested markeddependence on cell density and vessel density.26
Recently, Saunders and co-workers36 demonstrated thatpreincubation of HUVECs with 1 nM growth factorsfor 20 min prior to plating on gels disrupts networkformation. The study demonstrated that the networkinstability stemmed from increased cell migration in-duced by the growth factors. We further extended thework to demonstrate the impact of sustained releaseof VEGF on capillary morphogenesis. Towards this,we plated HUVECs on scaffolds encapsulating vary-ing concentration of VEGF to investigate how ECsintegrate signals from matrix mechanics and VEGF todirect vascular network formation. For this purpose,we utilized scaffolds whose compliances can be tunedindependent of porosity. To characterize the func-tional porosity of these scaffolds, the release ofencapsulated FITC-dextran 70 KDa from the hydro-gel matrices were evaluated. The release profile of themacromolecule did not vary significantly between thedifferent matrices, thereby suggesting that the diffu-sional transport of the angiogenic factors will be notbe affected by the increased mechanics of the scaf-folds. One caveat to utilizing dextran macromoleculesto model the transport of VEGF from the hydrogelmatrices is that the dextran molecules have differentstructure, charge, and susceptibility to matrix bindingas compared to VEGF. This in turn can likely affectthe diffusion of VEGF from the scaffolds. Because ofthe tendency of dextran macromolecules to get hy-drated easily, protein molecule of comparable mass asdextran will possess a smaller hydrodynamic radius,RH.
40 Since, the diffusion of a solute is inverselyproportional to RH, the potentially smaller hydrody-namic radius will counter the effect of charge andbinding interactions of VEGF with matrices. Whileassessing the quantitative relation between VEGFpresentation, matrix mechanics, and sprout forma-tion, our study revealed that in case of compliantscaffolds, VEGF did not have any effect on sprouting.However, in case of scaffolds with intermediate stiff-ness, VEGF enhanced the sprout formation and thesprouting response correlated with increased VEGFconcentration. When the cells were seeded on stiff
Integrated Effects of Matrix Mechanics and VEGF

scaffolds, VEGF facilitated capillary morphogenesisalbeit with inferior outcomes when compared toscaffolds of intermediate stiffness. Our results are inagreement with the findings of Mammoto andco-workers.1 Their study demonstrated that anappropriate level of ECM compliance is required foroptimal expression of VEGFR2. Our study suggeststhat matrix mechanics mediate VEGF inducedsprouting response of endothelial cells; highlighting acomplex relationship between the physical and bio-chemical signals in angiogenesis.
ACKNOWLEDGMENTS
The authors would like to thank Dr. Wonsuk Kimand Jacob Mack for their help in performing themechanical testing of the samples and Mao Ye for hishelp in SEM imaging. We would also like to thankUniversity of Michigan-Dearborn and University ofMichigan-Ann Arbor: Office of the Vice President forResearch for the financial support.
REFERENCES
1Arrio-Dupont, M., S. Cribier, G. Foucault, P. F. Devaux,and A. d’Albis. Diffusion of fluorescently labeled macro-molecules in cultured muscle cells. Biophys. J. 70(5):2327–2332, 1996.2Berrier, A. L., and K. M. Yamada. Cell–matrix adhesion.J. Cell. Physiol. 213(3):565–573, 2007.3Buzzai, M., R. G. Jones, R. K. Amaravadi, J. J. Lum, R. J.DeBerardinis, F. Zhao, and C. B. Thompson. Systemictreatment with the antidiabetic drug metformin selectivelyimpairs p53-deficient tumor cell growth. Cancer Res.67(14):6745–6752, 2007.4Califano, J. P., and C. A. Reinhart-King. The effects ofsubstrate elasticity on endothelial cell network formationand traction force generation. Conf. Proc. IEEE Eng. Med.Biol. Soc., Vol. 1, 2009, pp. 3343–3345.5Califano, J. P., and C. A. Reinhart-King. A balance ofsubstrate mechanics and matrix chemistry regulates endo-thelial cell network assembly. Cell. Mol. Bioeng. 1(2–3):122–132, 2008.6Califano, J. P., and C. A. Reinhart-King. Substrate stiff-ness and cell area predict cellular traction stresses in singlecells and cells in contact. Cell. Mol. Bioeng. 3(1):68–75,2010.7Carlevaro, M. F., S. Cermelli, R. Cancedda, and F. D.Cancedda. Vascular endothelial growth factor (VEGF) incartilage neovascularization and chondrocyte differentia-tion: auto-paracrine role during endochondral bone for-mation. J. Cell. Sci. 113(1):59–69, 2000.8Critser, P. J., S. T. Kreger, S. L. Voytik-Harbin, and M. C.Yoder. Collagen matrix physical properties modulateendothelial colony forming cell-derived vessels in vivo.Microvasc. Res. 80(1):23–30, 2000.
9Daugman, J. G. Uncertainty relation for resolution inspace, spatial frequency, and orientation optimized by two-dimensional visual cortical filters. Opt. Soc. Am. A2(7):1160–1169, 1985.
10Discher, D. E., P. Janmey, and Y. L. Wang. Tissue cells feeland respond to the stiffness of their substrate. Science310(5751):1139–1143, 2005.
11Doillon, C. J., A. J. Wasserman, R. A. Berg, and F. H.Silver. Behaviour of fibroblasts and epidermal cells culti-vated on analogues of extracellular matrix. Biomaterials9(1):91–96, 1988.
12Duong, H. T., S. C. Erzurum, and K. Asosingh. Pro-angiogenic hematopoietic progenitor cells and endothelialcolony-forming cells in pathological angiogenesis of bron-chial and pulmonary circulation. Angiogenesis 14(4):411–422, 2011.
13Ekstrand, A. J., R. Cao, M. Bjorndahl, S. Nystrom, A. C.Jonsson-Rylander, H. Hassani, and Y. Cao. Deletion ofneuropeptide Y (NPY) 2 receptor in mice results inblockage of NPY-induced angiogenesis and delayed woundhealing. Proc. Natl Acad. Sci. USA. 100(10):6033–6038,2003.
14Engler, A. J., M. A. Griffin, S. Sen, C. G. Bonnemann, H.L. Sweeney, and D. E. Discher. Myotubes differentiateoptimally on substrates with tissue-like stiffness patholog-ical implications for soft or stiff microenvironments. J.Cell. Biol. 166(6):877–887, 2004.
15Engler, A. J., L. Richert, J. Y. Wong, C. Picart, and D. E.Discher. Surface probe measurements of the elasticity ofsectioned tissue, thin gels and polyelectrolyte multilayerfilms: correlations between substrate stiffness and celladhesion. Surf. Sci. 570(1):142–154, 2004.
16Federspiel, W. J., and A. S. Popel. A theoretical analysis ofthe effect of the particulate nature of blood on oxygen re-lease in capillaries. Microvasc. Res. 32(2):164–189, 1986.
17Ghajar, C. M., X. Chen, J. W. Harris, V. Suresh, C. C.Hughes, N. L. Jeon, and S. C. George. The effect of matrixdensity on the regulation of 3-D capillary morphogenesis.Biophys. J. 94(5):1930–1941, 2008.
18Gospodarowicz, D., S. Massoglia, J. Cheng, and D. K.Fujii. Effect of fibroblast growth factor and lipoproteins onthe proliferation of endothelial cells derived from bovineadrenal cortex, brain cortex, and corpus luteum capillaries.J. Cell. Physiol. 127(1):121–136, 1986.
19Haraguchi, K., T. Takehisa, and S. Fan. Effects of claycontent on the properties of nanocomposite hydrogelscomposed of poly (N-isopropylacrylamide) and clay.Macromolecules 35(27):10162–10171, 2002.
20Hersel, U., C. Dahmen, and H. Kessler. RGD modifiedpolymers: biomaterials for stimulated cell adhesion andbeyond. Biomaterials 24(24):4385–4415, 2003.
21Hoeben, A., B. Landuyt, M. S. Highley, H. Wildiers, A. T.Van Oosterom, and E. A. De Bruijn. Vascular endothelialgrowth factor and angiogenesis. Pharmacol. Rev.56(4):549–580, 2004.
22Hood, J. D., R. Frausto, W. B. Kiosses, M. A. Schwartz,and D. A. Cheresh. Differential av integrin-mediated Ras-ERK signaling during two pathways of angiogenesis. J.Cell. Biol. 162(5):933–943, 2003.
23Kang, H. W., Y. Tabata, and Y. Ikada. Effect of porousstructure on the degradation of freeze-dried gelatinhydrogels. J. Bioactive Compat. Polym. 14(4):331–343,1999.
24Kniazeva, E., and A. J. Putnam. Endothelial cell tractionand ECM density influence both capillary morphogenesis
WU et al.

and maintenance in 3-D. Am. J. Physiol. Cell Physiol.297(1):C179–C187, 2009.
25Lammert, E., G. Gu, M. McLaughlin, D. Brown, R.Brekken, L. C. Murtaugh, and D. A. Melton. Role ofVEGF-A in vascularization of pancreatic islets. Curr. Biol.13(12):1070–1074, 2003.
26Mammoto, A., K. M. Connor, T. Mammoto, C. W. Yung,D. Huh, C. M. Aderman, and D. E. Ingber. A mechano-sensitive transcriptional mechanism that controls angio-genesis. Nature 457(7233):1103–1108, 2009.
27Miller, J. S., C. J. Shen, W. R. Legant, J. D. Baranski, B. L.Blakely, and C. S. Chen. Bioactive hydrogels made fromstep-growth derived PEGepeptide macromers. Biomaterials31:3736–3743, 2010.
28Miyamoto, S., H. Teramoto, O. A. Coso, J. S. Gutkind, P.D. Burbelo, S. K. Akiyama, and K. M. Yamada. Integrinfunction: molecular hierarchies of cytoskeletal and signal-ing molecules. J. Cell. Biol. 131(3):791–805, 1995.
29Murphy, C. M., M. G. Haugh, and F. J. O’Brien. Theeffect of mean pore size on cell attachment, proliferationand migration in collagen–glycosaminoglycan scaffolds forbone tissue engineering. Biomaterials 31(3):461–466, 2010.
30Nazarov, R., H. J. Jin, and D. L. Kaplan. Porous 3-Dscaffolds from regenerated silk fibroin. Biomacromolecules5(3):718–726, 2004.
31Nichol, J. W., S. T. Koshy, H. Bae, C. M. Hwang, S.Yamanlar, and A. Khademhosseini. Cell-laden microengi-neered gelatin methacrylate hydrogels. Biomaterials31(21):5536–5544, 2010.
32Park, S. N., J. C. Park, H. O. Kim, M. J. Song, and H. Suh.Characterization of porous collagen/hyaluronic acid scaf-fold modified by 1-ethyl-3-(3-dimethylaminopropyl) car-bodiimide cross-linking. Biomaterials 23(4):1205–1212,2002.
33Pedron, S., and B. A. C. Harley. Impact of the biophysicalfeatures of a 3D gelatin microenvironment on glioblastoma
malignancy. J. Biomed. Mater. Res A. 101A:3404–3415,2013.
34Plopper, G. E., H. P. McNamee, L. E. Dike, K. Boja-nowski, and D. E. Ingber. Convergence of integrin andgrowth factor receptor signaling pathways within the focaladhesion complex. Mol. Biol. Cell. 6(10):1349, 1995.
35Reinhart-King, C. A., M. Dembo, and D. A. Hammer. Thedynamics and mechanics of endothelial cell spreading.Biophys. J. 89(1):676–689, 2005.
36Saunders, R. L., and D. A. Hammer. Assembly of humanumbilical vein endothelial cells on compliant hydrogels.Cell. Mol. Bioeng. 3(1):60–67, 2010.
37Shen, F., Y. L. Cui, L. F. Yang, K. D. Yao, X. H. Dong,W. Y. Jia, and H. D. Shi. A study on the fabrication ofporous chitosan/gelatin network scaffold for tissue engi-neering. Polym. Int. 49(12):1596–1599, 2000.
38Sieminski, A. L., R. P. Hebbel, and K. J. Gooch. The rel-ative magnitudes of endothelial force generation and ma-trix stiffness modulate capillary morphogenesis in vitro.Exp. Cell. Res. 297(2):574–584, 2004.
39Sieminski, A. L., A. S. Was, G. Kim, H. Gong, and R. D.Kamm. The stiffness of three-dimensional ionic self-assembling peptide gels affects the extent of capillary-likenetwork formation. Cell. Biochem. Biophys. 49(2):73–83,2007.
40Verbridge, S. S., A. Chakrabarti, P. DelNero, B. Kwee, J.D. Varner, A. D. Stroock, and C. Fischbach. Physico-chemical regulation of endothelial sprouting in a 3Dmicrofluidic angiogenesis model. J. Biomed. Mater. Res. A101(10):2948–2956, 2013.
41Wood, J. A., N. M. Shah, C. T. McKee, M. L. Hughbanks,S. J. Liliensiek, P. Russell, and C. J. Murphy. The role ofsubstratum compliance of hydrogels on vascular endothe-lial cell behavior. Biomaterials 32(22):5056–5064, 2011.
42ATCC� STem Cell CulTure Guide: tips and techniques forculturing stem cells.
Integrated Effects of Matrix Mechanics and VEGF


















