
Insertion of a Pax6 Consensus Binding Site into the A-Crystallin ...
10
Insertion of a Pax6 Consensus Binding Site into the A-Crystallin Promoter Acts as a Lens Epithelial Cell Enhancer in Transgenic Mice Haotian Zhao, 1,2 Ying Yang, 1 Christian M. Rizo, 1 Paul A. Overbeek, 3 and Michael L. Robinson 1,2,4 PURPOSE. Although the murine A-crystallin promoter is the most commonly used promoter for achieving transgene ex- pression in the developing lens, this promoter directs trans- gene expression efficiently only in lens fiber cells. The purpose of the present study was to generate promoters capable of directing transgene expression to the entire lens but not to the corneal epithelium. METHODS. Transgenic mice were generated with fragments of the murine A- and B-crystallin promoters, as well as with an A-crystallin promoter engineered with the insertion of a Pax6 consensus binding site driving either human growth hormone (hGH) or Cre recombinase genes. hGH expression was evalu- ated by in situ hybridization and immunohistochemistry. Cre expression was revealed by x-gal staining after crossing Cre transgenic mice with a Cre reporter strain. RESULTS. Within the lens, the 214/38 B-crystallin promoter fragment directed transgene expression in the lens epithelium, but not in fiber cells. The native 282/43 A-crystallin pro- moter drove transgene expression in the lens fiber cells of several independent lines of transgenic mice, but none of these mice demonstrated significant transgene expression in the lens epithelium. In contrast, the insertion of a 32-bp sequence containing a Pax6 consensus binding site into the 282/43 A-crystallin promoter reproducibly led to transgene expres- sion in the lens epithelium as well as the lens fiber cells. CONCLUSIONS. The inclusion of a Pax6 consensus binding site within the 282/43 A-crystallin promoter enhances the ability of this promoter to drive transgene expression in the lens epithelium. (Invest Ophthalmol Vis Sci. 2004;45: 1930 –1939) DOI:10.1167/iovs.03-0856 T he lens consists of a proliferative epithelium that lines the anterior surface of the structure, covering the apical sur- face of a core of postmitotic terminally differentiated fiber cells. These two types of cells represent a single lineage, in that lens epithelial cells migrate toward the equatorial region of the lens where they exit the cell cycle and differentiate into fiber cells. Genetic engineering has emerged as a powerful tool for the investigation of the molecular basis of lens fiber differen- tiation. The mouse lens was one of the first tissues in which tissue-specific transgene expression was demonstrated 1 and was the first tissue in which Cre-mediated recombination was used to catalyze tissue-specific genomic engineering. 2 Most of the promoters used for transgene expression in the lens are derivations of crystallin promoters, and with the exception of the B-crystallin promoter, 3–5 they are capable of driving high- level transgene expression only in the lens fiber cells. 6 –13 To investigate mechanisms of fiber differentiation, however, it is desirable to manipulate gene expression in the undifferenti- ated lens epithelium. Crystallins are major water-soluble cytoplasmic proteins in the lens. The -, -, and -crystallins, found in all mammalian lenses, are expressed in either a lens-specific or a lens-pre- ferred manner. The -crystallins consist of A- and B-crystal- lin. The lens is the major site of A-crystallin expression, where it is detected in both epithelial and fiber cells, but a dramatic increase in the expression of A-crystallin occurs as the lens epithelial cells differentiate into lens fibers. 14 Only trace amounts of A-crystallin have been detected in a small number of non–lens tissues. 15,16 Multiple cis-acting transcriptional reg- ulatory elements residing in the 5-flanking region of the mu- rine A-crystallin gene contribute to its lens-preferred expres- sion. 17–22 In contrast, murine B-crystallin is expressed in the lens, retinal pigment epithelium (RPE), heart, skeletal muscle, and brain. 3,23–28 A short fragment of the B-crystallin promoter (164/44), lacking upstream elements shown to be impor- tant for non–lens expression of B-crystallin, 25–27 was shown to confer lens specificity in transgenic mice. 29 However, a later report demonstrated that this promoter is also active in the postnatal corneal epithelium. 5 Two regulatory elements within this region bind Pax6 and mediate transcriptional activation of the B-crystallin gene in the lens epithelium. 4 Unlike A-crys- tallin, B-crystallin has more pronounced expression in the lens epithelium than in the fibers. 14 In the embryonic and adult lens, Pax6 mRNA is detected mainly in the epithelium. 30,31 Although Pax6 protein is detect- able in newly differentiated lens fibers, the protein is gradually lost as fiber differentiation proceeds. 30,31 Pax6 plays many critical roles in early vertebrate eye development, and lens formation does not occur in the absence of Pax6. 32,33 Pax6 plays a role in regulating the spatial and temporal expression of A-, B-, E-, and F-crystallins in the mouse; A-, B1- and 1-crystallins in chick, and -crystallin in the guinea pig. 4,17,31,34 –37 In addition, Pax6 has been shown to act up- stream of transcription factors that are essential in lens devel- opment including Sox2, Six3, L-Maf, and Prox1. 32,38,39 An element from the Pax6 P0 promoter, known as the ectoderm enhancer, has been shown to direct transgene ex- pression in the lens placode and placode-derived structures, From the 1 Center for Molecular and Human Genetics, Columbus Children’s Research Institute, Columbus, Ohio; the 2 Graduate Program in Molecular, Cellular, and Developmental Biology, College of Biolog- ical Sciences, The Ohio State University, Columbus, Ohio; the 3 Depart- ment of Cellular and Molecular Biology, Baylor College of Medicine, Houston, Texas; and the 4 Department of Pediatrics, College of Medi- cine, The Ohio State University, Columbus, Ohio. Supported by National Eye Institute Grants EY06511 (MLR), EY12995 (MLR), EY10448 (PAO), and EY10803 (PAO); and the Colum- bus Children’s Research Institute and NCI The Comprehensive Cancer Center at The Ohio State University. Submitted for publication August 7, 2003; revised December 26, 2003; accepted January 29, 2004. Disclosure: H. Zhao, None; Y. Yang, None; C.M. Rizo, None; P.A. Overbeek, None; M.L. Robinson, None The publication costs of this article were defrayed in part by page charge payment. This article must therefore be marked “advertise- ment” in accordance with 18 U.S.C. §1734 solely to indicate this fact. Corresponding author: Michael L. Robinson, Center for Molecular and Human Genetics, Columbus Children’s Research Institute, Colum- bus, OH 43205; [email protected]. Investigative Ophthalmology & Visual Science, June 2004, Vol. 45, No. 6 1930 Copyright © Association for Research in Vision and Ophthalmology
Transcript of Insertion of a Pax6 Consensus Binding Site into the A-Crystallin ...

Insertion of a Pax6 Consensus Binding Site into the�A-Crystallin Promoter Acts as a Lens Epithelial CellEnhancer in Transgenic Mice
Haotian Zhao,1,2 Ying Yang,1 Christian M. Rizo,1 Paul A. Overbeek,3 andMichael L. Robinson1,2,4
PURPOSE. Although the murine �A-crystallin promoter is themost commonly used promoter for achieving transgene ex-pression in the developing lens, this promoter directs trans-gene expression efficiently only in lens fiber cells. The purposeof the present study was to generate promoters capable ofdirecting transgene expression to the entire lens but not to thecorneal epithelium.
METHODS. Transgenic mice were generated with fragments ofthe murine �A- and �B-crystallin promoters, as well as with an�A-crystallin promoter engineered with the insertion of a Pax6consensus binding site driving either human growth hormone(hGH) or Cre recombinase genes. hGH expression was evalu-ated by in situ hybridization and immunohistochemistry. Creexpression was revealed by x-gal staining after crossing Cretransgenic mice with a Cre reporter strain.
RESULTS. Within the lens, the �214/�38 �B-crystallin promoterfragment directed transgene expression in the lens epithelium,but not in fiber cells. The native �282/�43 �A-crystallin pro-moter drove transgene expression in the lens fiber cells ofseveral independent lines of transgenic mice, but none of thesemice demonstrated significant transgene expression in the lensepithelium. In contrast, the insertion of a 32-bp sequencecontaining a Pax6 consensus binding site into the �282/�43�A-crystallin promoter reproducibly led to transgene expres-sion in the lens epithelium as well as the lens fiber cells.
CONCLUSIONS. The inclusion of a Pax6 consensus binding sitewithin the �282/�43 �A-crystallin promoter enhances theability of this promoter to drive transgene expression in thelens epithelium. (Invest Ophthalmol Vis Sci. 2004;45:1930–1939) DOI:10.1167/iovs.03-0856
The lens consists of a proliferative epithelium that lines theanterior surface of the structure, covering the apical sur-
face of a core of postmitotic terminally differentiated fiber
cells. These two types of cells represent a single lineage, in thatlens epithelial cells migrate toward the equatorial region of thelens where they exit the cell cycle and differentiate into fibercells. Genetic engineering has emerged as a powerful tool forthe investigation of the molecular basis of lens fiber differen-tiation. The mouse lens was one of the first tissues in whichtissue-specific transgene expression was demonstrated1 andwas the first tissue in which Cre-mediated recombination wasused to catalyze tissue-specific genomic engineering.2 Most ofthe promoters used for transgene expression in the lens arederivations of crystallin promoters, and with the exception ofthe �B-crystallin promoter,3–5 they are capable of driving high-level transgene expression only in the lens fiber cells.6–13 Toinvestigate mechanisms of fiber differentiation, however, it isdesirable to manipulate gene expression in the undifferenti-ated lens epithelium.
Crystallins are major water-soluble cytoplasmic proteins inthe lens. The �-, �-, and �-crystallins, found in all mammalianlenses, are expressed in either a lens-specific or a lens-pre-ferred manner. The �-crystallins consist of �A- and �B-crystal-lin. The lens is the major site of �A-crystallin expression, whereit is detected in both epithelial and fiber cells, but a dramaticincrease in the expression of �A-crystallin occurs as the lensepithelial cells differentiate into lens fibers.14 Only traceamounts of �A-crystallin have been detected in a small numberof non–lens tissues.15,16 Multiple cis-acting transcriptional reg-ulatory elements residing in the 5�-flanking region of the mu-rine �A-crystallin gene contribute to its lens-preferred expres-sion.17–22
In contrast, murine �B-crystallin is expressed in the lens,retinal pigment epithelium (RPE), heart, skeletal muscle, andbrain.3,23–28 A short fragment of the �B-crystallin promoter(�164/�44), lacking upstream elements shown to be impor-tant for non–lens expression of �B-crystallin,25–27 was shownto confer lens specificity in transgenic mice.29 However, a laterreport demonstrated that this promoter is also active in thepostnatal corneal epithelium.5 Two regulatory elements withinthis region bind Pax6 and mediate transcriptional activation ofthe �B-crystallin gene in the lens epithelium.4 Unlike �A-crys-tallin, �B-crystallin has more pronounced expression in thelens epithelium than in the fibers.14
In the embryonic and adult lens, Pax6 mRNA is detectedmainly in the epithelium.30,31 Although Pax6 protein is detect-able in newly differentiated lens fibers, the protein is graduallylost as fiber differentiation proceeds.30,31 Pax6 plays manycritical roles in early vertebrate eye development, and lensformation does not occur in the absence of Pax6.32,33 Pax6plays a role in regulating the spatial and temporal expression of�A-, �B-, �E-, and �F-crystallins in the mouse; �A-, �B1- and�1-crystallins in chick, and �-crystallin in the guineapig.4,17,31,34–37 In addition, Pax6 has been shown to act up-stream of transcription factors that are essential in lens devel-opment including Sox2, Six3, L-Maf, and Prox1.32,38,39
An element from the Pax6 P0 promoter, known as theectoderm enhancer, has been shown to direct transgene ex-pression in the lens placode and placode-derived structures,
From the 1Center for Molecular and Human Genetics, ColumbusChildren’s Research Institute, Columbus, Ohio; the 2Graduate Programin Molecular, Cellular, and Developmental Biology, College of Biolog-ical Sciences, The Ohio State University, Columbus, Ohio; the 3Depart-ment of Cellular and Molecular Biology, Baylor College of Medicine,Houston, Texas; and the 4Department of Pediatrics, College of Medi-cine, The Ohio State University, Columbus, Ohio.
Supported by National Eye Institute Grants EY06511 (MLR),EY12995 (MLR), EY10448 (PAO), and EY10803 (PAO); and the Colum-bus Children’s Research Institute and NCI The Comprehensive CancerCenter at The Ohio State University.
Submitted for publication August 7, 2003; revised December 26,2003; accepted January 29, 2004.
Disclosure: H. Zhao, None; Y. Yang, None; C.M. Rizo, None;P.A. Overbeek, None; M.L. Robinson, None
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be marked “advertise-ment” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Corresponding author: Michael L. Robinson, Center for Molecularand Human Genetics, Columbus Children’s Research Institute, Colum-bus, OH 43205; [email protected].
Investigative Ophthalmology & Visual Science, June 2004, Vol. 45, No. 61930 Copyright © Association for Research in Vision and Ophthalmology

such as the epithelium of the lens, cornea, conjunctiva, andlacrimal gland.40,41 Transgene expression with this enhancer isnot maintained in lens fiber cells. Our purpose was to find orengineer promoters capable of driving transgene expression inthe entire lens, including the lens epithelium, but without theinclusion of other ocular epithelia. We chose to use two dif-ferent genes to evaluate the promoter expression pattern tocontrol for possible influences of coding sequence on pro-moter activity. The first of these, human growth hormone(hGH), is not normally expressed in the eye, but can easily bedetected by in situ hybridization or immunohistochemistry.The second gene was Cre recombinase. Cre is a P1 bacterio-phage-derived DNA recombinase with a specific 34-bp recog-nition sequence called a loxP site. Cre recombinase was usedto test these promoters with the thought that the resultantmice could not only provide more in vivo evidence about theeffect of the Pax6 consensus binding site on the �A-crystallinpromoter activity, but also could serve as tools to create con-ditional inactivation of genes in a tissue-specific manner.
Within the eye, transgenic constructs containing the murine�214/�38 �B-crystallin promoter most often drove transgeneexpression in the lens epithelium and RPE, whereas constructscontaining the native murine �282/�43 �A-crystallin pro-moter most often drove transgene expression only in fiber cellsof the lens. Insertion of a DNA sequence that binds Pax634 intothe �A-crystallin promoter enhanced the expression of twodifferent reporter genes in the lens epithelium while maintain-ing fiber cell expression.
METHODS
Construction of Transgenic Vectors
All primer sequences and detailed methods are available in the onlineappendix at www.iovs.org/cgi/content/full/45/6/1930/DC1. A novelBglII site was inserted into the CPV2 vector42 at �86 relative to themurine �A-crystallin transcription start site by overlapping PCR, creat-ing the intermediate CPV7 vector. A consensus Pax6 binding site,34
created by annealing two partially complementary oligonucleotideswith 5� overhangs compatible with the BglII restriction site, wasinserted into the BglII site of CPV7 to generate CPV14 and CPV15vectors, differing only by the orientation of the single Pax6 bindingsite. A genomic fragment containing the hGH gene from pOGH (ob-tained from Francesco DeMayo, Baylor College of Medicine, Houston,TX43) was subcloned into CPV2, CPV14, and CPV15, creating CPV2/hGH, CPV14/hGH, and CPV15/hGH, respectively.
To create the �B1/hGH construct, genomic sequence from �214to �38 of the mouse �B-crystallin gene (relative to the major lenstranscription start site, GenBank accession no. M7374144; http://www.ncbi.nlm.nih.gov/Genbank; provided in the public domain by the Na-tional Center for Biotechnology Information, Bethesda, MD) was am-plified by PCR and inserted into CPV2, replacing the murine�A-crystallin promoter and creating the vector �B1. The promoter of�B1 was ligated into the vector backbone of CPV2/hGH, replacing the�A-crystallin promoter to create �B1/hGH.
The �A/�B4/hGH construct was made through several steps. First,the 3� untranslated region and polyadenylation signal of the murine�B-crystallin gene was amplified by PCR and ligated into the largefragment of the �B1 vector, replacing the SV40-derived intron andpolyadenylation sequence of �B1. This intermediate plasmid wascalled �B2. The first intron of the murine �B-crystallin gene wasamplified by PCR and inserted into the �B1 vector, creating theintermediate vector �B3. A three-way ligation with the �B-crystallinpromoter and first intron from �B3, the hGH gene from pOGH,43 andthe �B-crystallin 3� untranslated region–polyadenylation sequencesand plasmid backbone of �B2 created �B4/hGH. To generate thecomposite �A/�B promoter, CPV7 (described earlier) was digestedwith BglII and BamHI to remove the �86/�43 region of the �A-crystallin gene. The PCR-amplified murine �B-crystallin promoter (de-
scribed earlier) was digested with BglII and ligated into the BglII/BamHI-cut CPV7, creating the intermediate �A/�B compositepromoter vector. The �A/�B composite promoter was then used toreplace the �B-crystallin promoter of �B4/hGH, completing the �A/�B4/hGH construct.
The Cre coding sequence and intron–polyadenylation sequencescontained in the murine metallothionein gene were excised frompBS216 (obtained from Brian Sauer, Stowers Institute, Kansas City,MO2) and ligated into HindIII/SalI cut CPV14 and CPV2, creatingCPV14/Cre and CPV2/Cre, respectively.
Generation of Transgenic Mice
All animals were treated in compliance with the ARVO Statement forthe Use of Animals in Ophthalmic and Vision Research. Microinjectionfragments were isolated from CPV2/hGH, CPV14/hGH, CPV15/hGH,�B1/hGH, TYBS (a tyrosinase minigene),45 �A/�B4/hGH, CPV14/Cre,and CPV2/Cre following digestion with appropriate restriction en-zymes and gel purification with a gel extraction kit (QiaexII; Qiagen,Hilden, Germany). All microinjection constructs were injected intopronuclear stage FVB/N mouse embryos as described.46 All hGH trans-genic constructs were injected independently, but Cre constructs wereeither injected independently or coinjected with the TYBS cassette.45
Histology, Immunohistochemistry, andIn Situ Hybridization
Eyes or embryos were collected and fixed in 4% paraformaldehyde,processed, and embedded in paraffin. Embedded samples were sec-tioned at 5 �m. Immunohistochemical staining for hGH was performedas described previously.43 An hGH-specific riboprobe vector, CB4, wascreated by ligating the 3� untranslated region of the human growthhormone gene from the pSW247 vector (obtained from Lewis T. Wil-liams, University of California, San Francisco) into pBluescript II KS(�)(Stratagene, La Jolla, CA). Antisense [35S]-UTP–labeled probes weresynthesized using a HindIII-digested CB4 template and T7 RNA poly-merase (Promega, Madison, WI). In situ hybridization of tissue sectionswas performed as described previously.42
X-Gal Staining for �-Galactosidase Activity
To evaluate the expression pattern of the Cre transgene, Cre transgenicmouse lines were crossed with a homozygous ROSA26 reporter line(B6;129S-Gtrosa26tm1Sor).48 Mouse embryos, neonatal eyes and lensepithelium from mice at weaning age were collected and analyzed for�-galactosidase activity by x-gal staining, as described.42
RESULTS
Characterization of Engineered�-Crystallin Promoters
We produced several transgenic constructs in which an hGHreporter gene was placed under the regulatory control ofmodified versions of the murine �A-crystallin promoter or ashort fragment (�214/�38) of the murine �B-crystallin pro-moter (Table 1). The activity of these promoters was initiallytested by using a transient transgenic approach in which trans-genic founder embryos were collected at 15.5 days after coitus(embryonic day [E]15.5). At least two transgenic founder em-bryos were generated for each construct and evaluated forhGH expression by both in situ hybridization and immunohis-tochemistry.
CPV2/hGH consisted of the hGH reporter gene downstreamof the (�282/�43) murine �A-crystallin promoter (Fig. 1A,Table 1). The �B1/hGH construct was identical to CPV2/hGHexcept that the �A-crystallin promoter was replaced with ashort fragment (�214/�38) of the murine �B-crystallin pro-moter (Fig. 1A, Table 1). A composite promoter including the�282/�86 region of the murine �A-crystallin promoter fol-lowed by the �214/�38 promoter fragment and the firstintron of murine �B-crystallin gene was also created. The hGH
IOVS, June 2004, Vol. 45, No. 6 Lens Epithelial Transgene Expression 1931
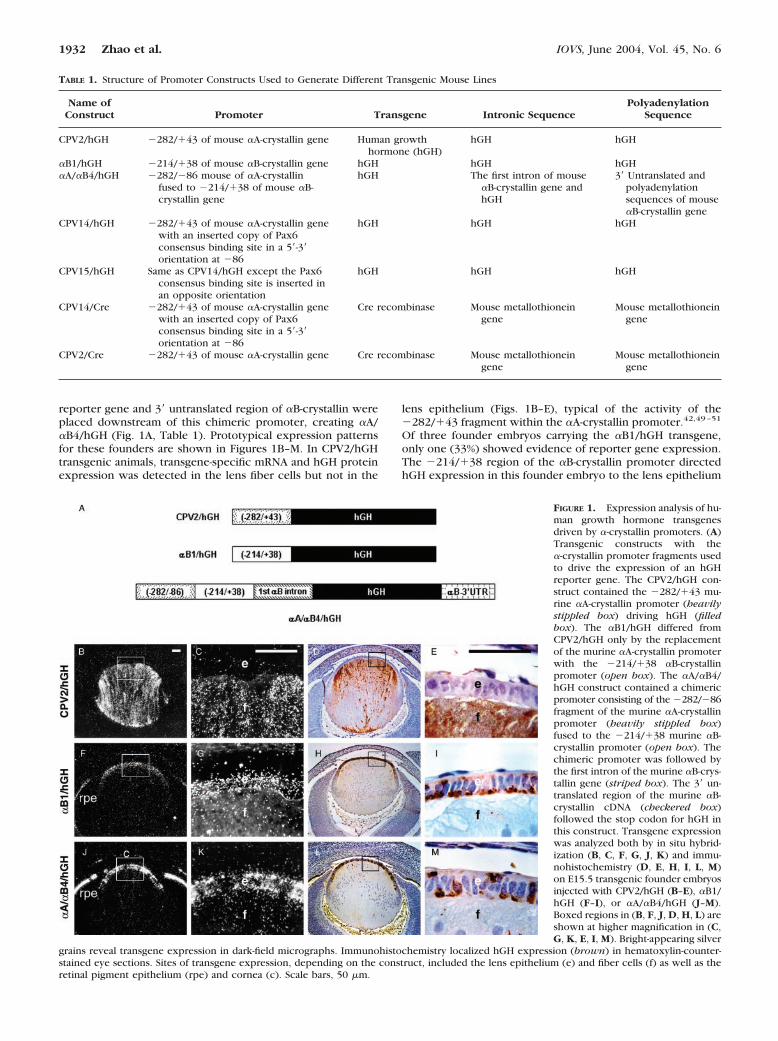
reporter gene and 3� untranslated region of �B-crystallin wereplaced downstream of this chimeric promoter, creating �A/�B4/hGH (Fig. 1A, Table 1). Prototypical expression patternsfor these founders are shown in Figures 1B–M. In CPV2/hGHtransgenic animals, transgene-specific mRNA and hGH proteinexpression was detected in the lens fiber cells but not in the
lens epithelium (Figs. 1B–E), typical of the activity of the�282/�43 fragment within the �A-crystallin promoter.42,49–51
Of three founder embryos carrying the �B1/hGH transgene,only one (33%) showed evidence of reporter gene expression.The �214/�38 region of the �B-crystallin promoter directedhGH expression in this founder embryo to the lens epithelium
TABLE 1. Structure of Promoter Constructs Used to Generate Different Transgenic Mouse Lines
Name ofConstruct Promoter Transgene Intronic Sequence
PolyadenylationSequence
CPV2/hGH �282/�43 of mouse �A-crystallin gene Human growthhormone (hGH)
hGH hGH
�B1/hGH �214/�38 of mouse �B-crystallin gene hGH hGH hGH�A/�B4/hGH �282/�86 mouse of �A-crystallin
fused to �214/�38 of mouse �B-crystallin gene
hGH The first intron of mouse�B-crystallin gene andhGH
3� Untranslated andpolyadenylationsequences of mouse�B-crystallin gene
CPV14/hGH �282/�43 of mouse �A-crystallin genewith an inserted copy of Pax6consensus binding site in a 5�-3�orientation at �86
hGH hGH hGH
CPV15/hGH Same as CPV14/hGH except the Pax6consensus binding site is inserted inan opposite orientation
hGH hGH hGH
CPV14/Cre �282/�43 of mouse �A-crystallin genewith an inserted copy of Pax6consensus binding site in a 5�-3�orientation at �86
Cre recombinase Mouse metallothioneingene
Mouse metallothioneingene
CPV2/Cre �282/�43 of mouse �A-crystallin gene Cre recombinase Mouse metallothioneingene
Mouse metallothioneingene
FIGURE 1. Expression analysis of hu-man growth hormone transgenesdriven by �-crystallin promoters. (A)Transgenic constructs with the�-crystallin promoter fragments usedto drive the expression of an hGHreporter gene. The CPV2/hGH con-struct contained the �282/�43 mu-rine �A-crystallin promoter (heavilystippled box) driving hGH (filledbox). The �B1/hGH differed fromCPV2/hGH only by the replacementof the murine �A-crystallin promoterwith the �214/�38 �B-crystallinpromoter (open box). The �A/�B4/hGH construct contained a chimericpromoter consisting of the �282/�86fragment of the murine �A-crystallinpromoter (heavily stippled box)fused to the �214/�38 murine �B-crystallin promoter (open box). Thechimeric promoter was followed bythe first intron of the murine �B-crys-tallin gene (striped box). The 3� un-translated region of the murine �B-crystallin cDNA (checkered box)followed the stop codon for hGH inthis construct. Transgene expressionwas analyzed both by in situ hybrid-ization (B, C, F, G, J, K) and immu-nohistochemistry (D, E, H, I, L, M)on E15.5 transgenic founder embryosinjected with CPV2/hGH (B–E), �B1/hGH (F–I), or �A/�B4/hGH (J–M).Boxed regions in (B, F, J, D, H, L) areshown at higher magnification in (C,G, K, E, I, M). Bright-appearing silver
grains reveal transgene expression in dark-field micrographs. Immunohistochemistry localized hGH expression (brown) in hematoxylin-counter-stained eye sections. Sites of transgene expression, depending on the construct, included the lens epithelium (e) and fiber cells (f) as well as theretinal pigment epithelium (rpe) and cornea (c). Scale bars, 50 �m.
1932 Zhao et al. IOVS, June 2004, Vol. 45, No. 6

and RPE with very little detectable expression in lens fiber cells(Figs. 1F–I). Three stable transgenic lines were generated withthe �B1/hGH construct (OVE1046, OVE1047, and OVE1048).Of these lines, two (OVE1046 and OVE1048) demonstratedtransgene expression in the lens epithelial cells, RPE, anddeveloping ciliary body in E15.5 embryos. One line (OVE1046)expressed the reporter gene weakly in the lens fiber cells (datanot shown). Overall, three (50%) of the six transgenic foundersor lines made with the �B1/hGH construct expressed thetransgene in the lens. In an attempt to increase the level of lenstransgene expression in the �B1/hGH founders, we created achimeric promoter consisting of the �282/�86 fragment ofthe mouse �A-crystallin promoter fused to the �214/�38mouse �B1-promoter, intron 1 of �B-crystallin, and the 3�untranslated sequence. Of the five �A/�B4/hGH transgenicfounder embryos, reporter gene expression was found in three(60%). In these founders, hGH was detected in the lens epi-thelium and RPE. In one (20%) founder, patchy expression inthe corneal epithelium was detected by in situ hybridization,but not by immunohistochemistry (Figs. 1J–M). None of the�A/�B4/hGH founder embryos expressed the transgene in thelens fiber cells. Thus, despite its activity in the lens epithelium,the �A/�B4 chimeric promoter failed to activate transgeneexpression in the lens fibers, indicating the �282/�86 �A-crystallin promoter fragment was insufficient to activate trans-gene expression in the lens fibers when coupled with the�214/�38 �B-crystallin promoter.
Effect of Insertion of the Pax6 Consensus BindingSite into the �A-crystallin Promoter on TransgeneExpression in the Lens Epithelium
The differential expression patterns of �A- and �B-crystallinswithin the lens and the observation that Pax6 directly regulatesexpression of �B-crystallin in the lens epithelium prompted usto hypothesize that we might be able to enhance the activity ofthe murine �A-crystallin promoter in the lens epithelium byincorporating additional Pax6-binding regulatory elements. To
test this hypothesis, we made a small modification in CPV2 byinserting a 32-bp sequence containing a Pax6 consensus bind-ing site in either orientation, creating CPV14 and CPV15, re-spectively. The insertion site is located at �86 (relative to thetranscription start site) of the �A-crystallin gene, a regiondevoid of elements known to be important for �A-crystallintranscriptional regulation. The hGH gene was then subclonedinto both of these vectors, creating CPV14/hGH and CPV15/hGH (Fig. 2A, Table 1).
Transgenic mice made with either CPV14/hGH or CPV15/hGH constructs expressed hGH mRNA and protein specificallyin both the lens epithelium and fibers (Figs. 2B–I). Becauseboth CPV14/hGH and CPV15/hGH supported transgene ex-pression in the lens epithelium, the inclusion of the Pax6consensus binding sequence in the �282/�43 region of �A-crystallin promoter enhanced lens epithelial transgene expres-sion in an orientation-independent manner. In these transgenicfounder embryos, no ocular expression of the hGH reportergene was detected outside the lens.
Generation of CPV2/Cre and CPV14/CreTransgenic Mice
Because hGH expression driven by both the CPV14 and CPV15promoters was specific to the lens and detected in both thelens epithelium and fiber cells in transgenic founder embryos,we decided to test the activity of the CPV14 promoter morethoroughly by using a different reporter gene. To this end, wecloned Cre recombinase into CPV2 and CPV14, creating CPV2/Cre and CPV14/Cre constructs, respectively. In these con-structs, the mouse metallothionein gene is downstream of theCre coding sequence, providing introns and a polyadenylationsignal (Table 1). In most cases, a tyrosinase minigene cassettewas coinjected with the Cre transgenes to facilitate identifica-tion of transgenic animals by coat color.45
Seven transgenic lines were established for each Cre trans-gene construct. Lines coinjected with tyrosinase minigenewere also transgenic for this marker (Table 2). The expression
FIGURE 2. The addition of a Pax6-binding site into the murine �A-crys-tallin promoter enhanced transgeneexpression in the lens epithelium inan orientation-independent manner.(A) Modified �A-crystallin promotertransgenic constructs. A single copyof Pax6 consensus binding site(striped box) was inserted at position�86 of the murine �A-crystallin pro-moter (heavily stippled box) to cre-ate the CPV14 promoter. The CPV15promoter differed from CPV14 onlyby the orientation of the Pax6 con-sensus binding site. The sequence ofthe inserted fragment was sur-rounded by the endogenous �A-crys-tallin promoter sequences (smallerbold font) and the Pax6 binding se-quence is shown within the filledbox. hGH (filled box) was placeddownstream of the modified �A-crys-tallin promoters. In situ hybridization(B, C, F, G) and immunohistochem-istry (D, E, H, I) were performedon E15.5 embryos transgenic forCPV14/hGH (B–E) or CPV15/hGH(F–I) to reveal transgene expression.Boxed regions in (B, F, D, H) areshown at higher magnification in (C,G, E, I). For in situ hybridization,transgene-specific signals were revealed by reflected light in dark-field pictures. For immunohistochemistry, brown staining signals indicated theexpression of transgene product. e, lens epithelium; f, lens fiber cells. Scale bars, 50 �m.
IOVS, June 2004, Vol. 45, No. 6 Lens Epithelial Transgene Expression 1933
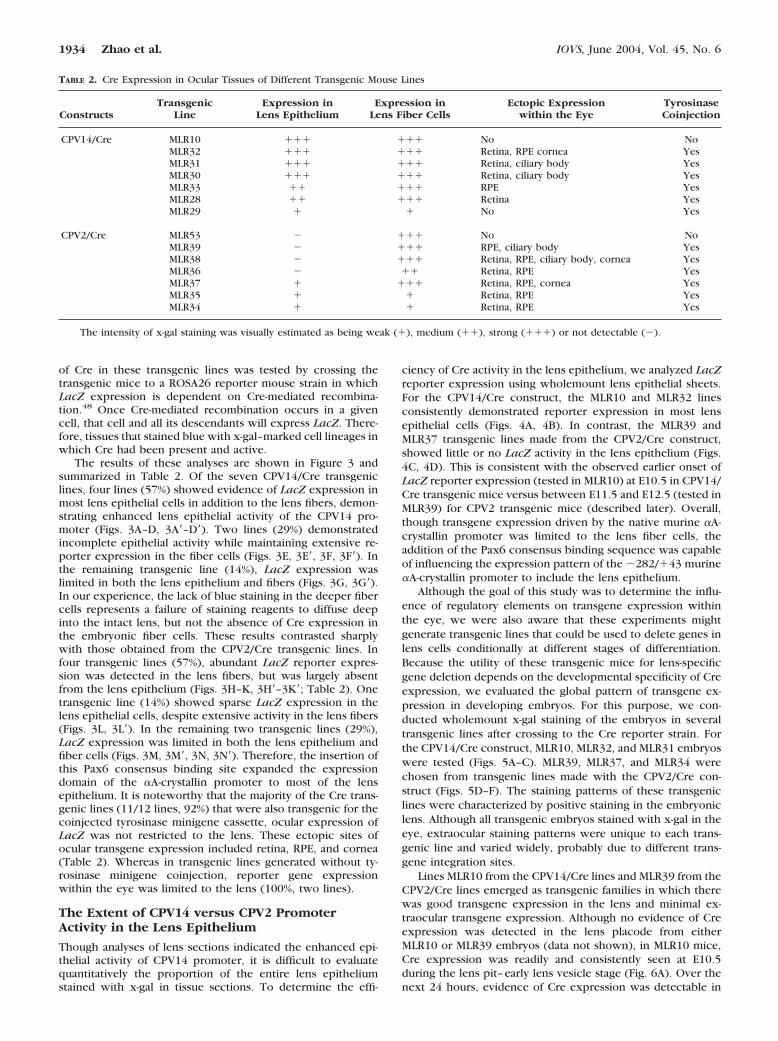
of Cre in these transgenic lines was tested by crossing thetransgenic mice to a ROSA26 reporter mouse strain in whichLacZ expression is dependent on Cre-mediated recombina-tion.48 Once Cre-mediated recombination occurs in a givencell, that cell and all its descendants will express LacZ. There-fore, tissues that stained blue with x-gal–marked cell lineages inwhich Cre had been present and active.
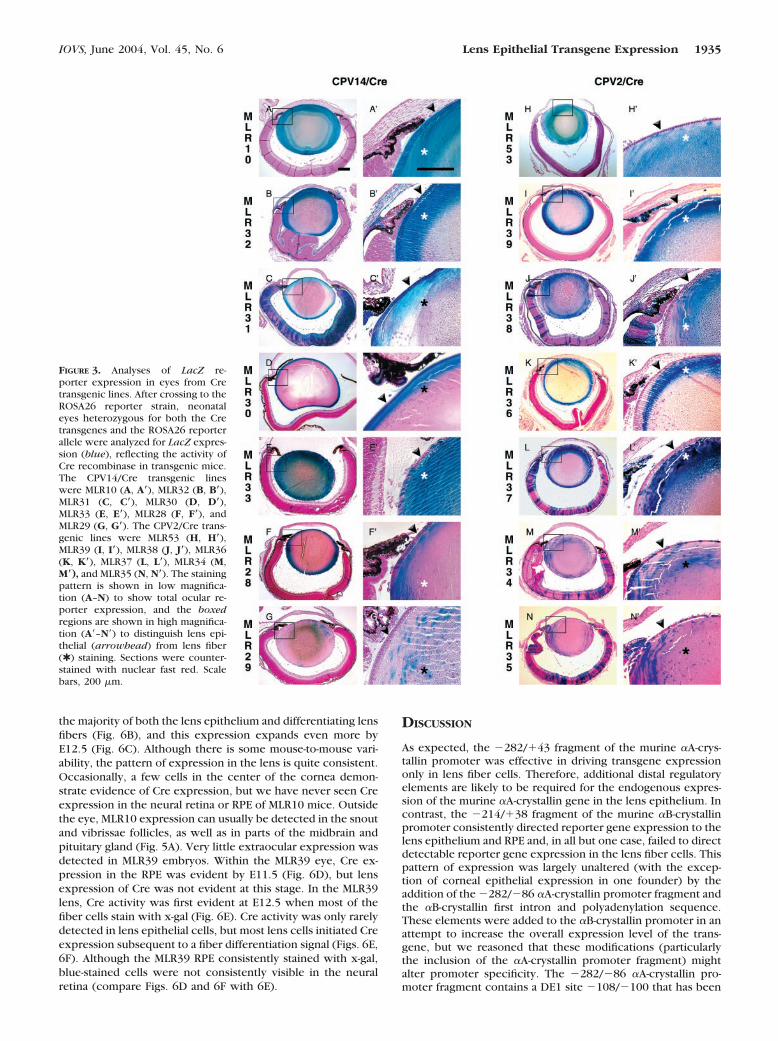
The results of these analyses are shown in Figure 3 andsummarized in Table 2. Of the seven CPV14/Cre transgeniclines, four lines (57%) showed evidence of LacZ expression inmost lens epithelial cells in addition to the lens fibers, demon-strating enhanced lens epithelial activity of the CPV14 pro-moter (Figs. 3A–D, 3A�–D�). Two lines (29%) demonstratedincomplete epithelial activity while maintaining extensive re-porter expression in the fiber cells (Figs. 3E, 3E�, 3F, 3F�). Inthe remaining transgenic line (14%), LacZ expression waslimited in both the lens epithelium and fibers (Figs. 3G, 3G�).In our experience, the lack of blue staining in the deeper fibercells represents a failure of staining reagents to diffuse deepinto the intact lens, but not the absence of Cre expression inthe embryonic fiber cells. These results contrasted sharplywith those obtained from the CPV2/Cre transgenic lines. Infour transgenic lines (57%), abundant LacZ reporter expres-sion was detected in the lens fibers, but was largely absentfrom the lens epithelium (Figs. 3H–K, 3H�–3K�; Table 2). Onetransgenic line (14%) showed sparse LacZ expression in thelens epithelial cells, despite extensive activity in the lens fibers(Figs. 3L, 3L�). In the remaining two transgenic lines (29%),LacZ expression was limited in both the lens epithelium andfiber cells (Figs. 3M, 3M�, 3N, 3N�). Therefore, the insertion ofthis Pax6 consensus binding site expanded the expressiondomain of the �A-crystallin promoter to most of the lensepithelium. It is noteworthy that the majority of the Cre trans-genic lines (11/12 lines, 92%) that were also transgenic for thecoinjected tyrosinase minigene cassette, ocular expression ofLacZ was not restricted to the lens. These ectopic sites ofocular transgene expression included retina, RPE, and cornea(Table 2). Whereas in transgenic lines generated without ty-rosinase minigene coinjection, reporter gene expressionwithin the eye was limited to the lens (100%, two lines).
The Extent of CPV14 versus CPV2 PromoterActivity in the Lens Epithelium
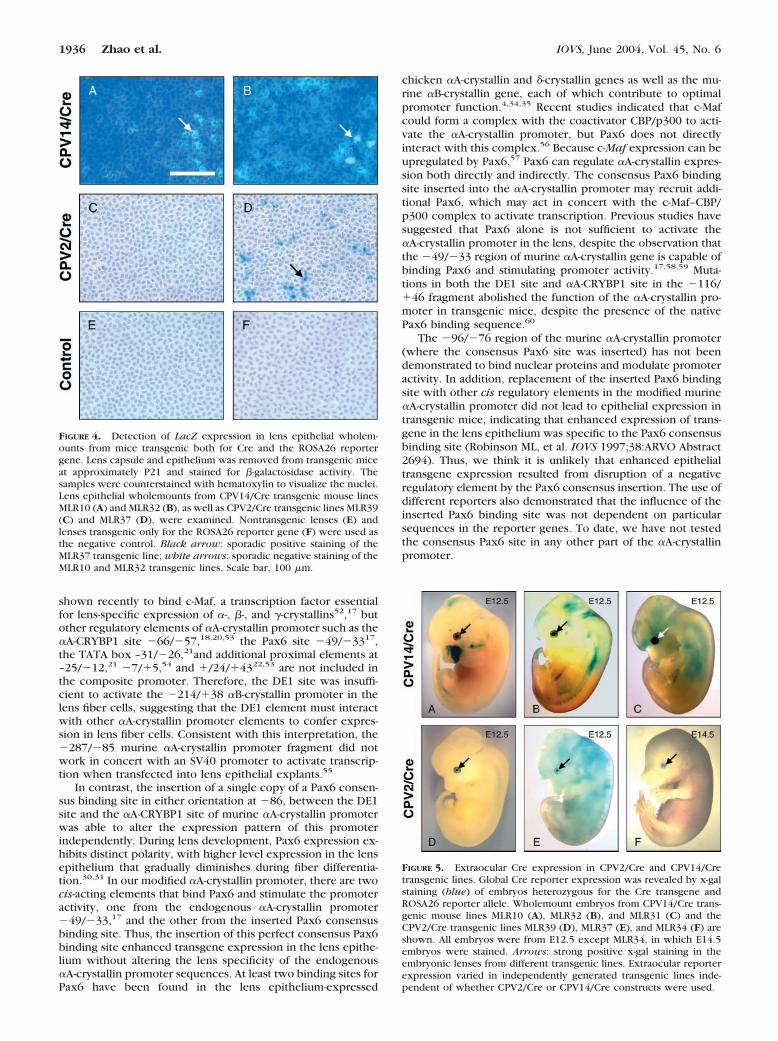
Though analyses of lens sections indicated the enhanced epi-thelial activity of CPV14 promoter, it is difficult to evaluatequantitatively the proportion of the entire lens epitheliumstained with x-gal in tissue sections. To determine the effi-
ciency of Cre activity in the lens epithelium, we analyzed LacZreporter expression using wholemount lens epithelial sheets.For the CPV14/Cre construct, the MLR10 and MLR32 linesconsistently demonstrated reporter expression in most lensepithelial cells (Figs. 4A, 4B). In contrast, the MLR39 andMLR37 transgenic lines made from the CPV2/Cre construct,showed little or no LacZ activity in the lens epithelium (Figs.4C, 4D). This is consistent with the observed earlier onset ofLacZ reporter expression (tested in MLR10) at E10.5 in CPV14/Cre transgenic mice versus between E11.5 and E12.5 (tested inMLR39) for CPV2 transgenic mice (described later). Overall,though transgene expression driven by the native murine �A-crystallin promoter was limited to the lens fiber cells, theaddition of the Pax6 consensus binding sequence was capableof influencing the expression pattern of the �282/�43 murine�A-crystallin promoter to include the lens epithelium.
Although the goal of this study was to determine the influ-ence of regulatory elements on transgene expression withinthe eye, we were also aware that these experiments mightgenerate transgenic lines that could be used to delete genes inlens cells conditionally at different stages of differentiation.Because the utility of these transgenic mice for lens-specificgene deletion depends on the developmental specificity of Creexpression, we evaluated the global pattern of transgene ex-pression in developing embryos. For this purpose, we con-ducted wholemount x-gal staining of the embryos in severaltransgenic lines after crossing to the Cre reporter strain. Forthe CPV14/Cre construct, MLR10, MLR32, and MLR31 embryoswere tested (Figs. 5A–C). MLR39, MLR37, and MLR34 werechosen from transgenic lines made with the CPV2/Cre con-struct (Figs. 5D–F). The staining patterns of these transgeniclines were characterized by positive staining in the embryoniclens. Although all transgenic embryos stained with x-gal in theeye, extraocular staining patterns were unique to each trans-genic line and varied widely, probably due to different trans-gene integration sites.
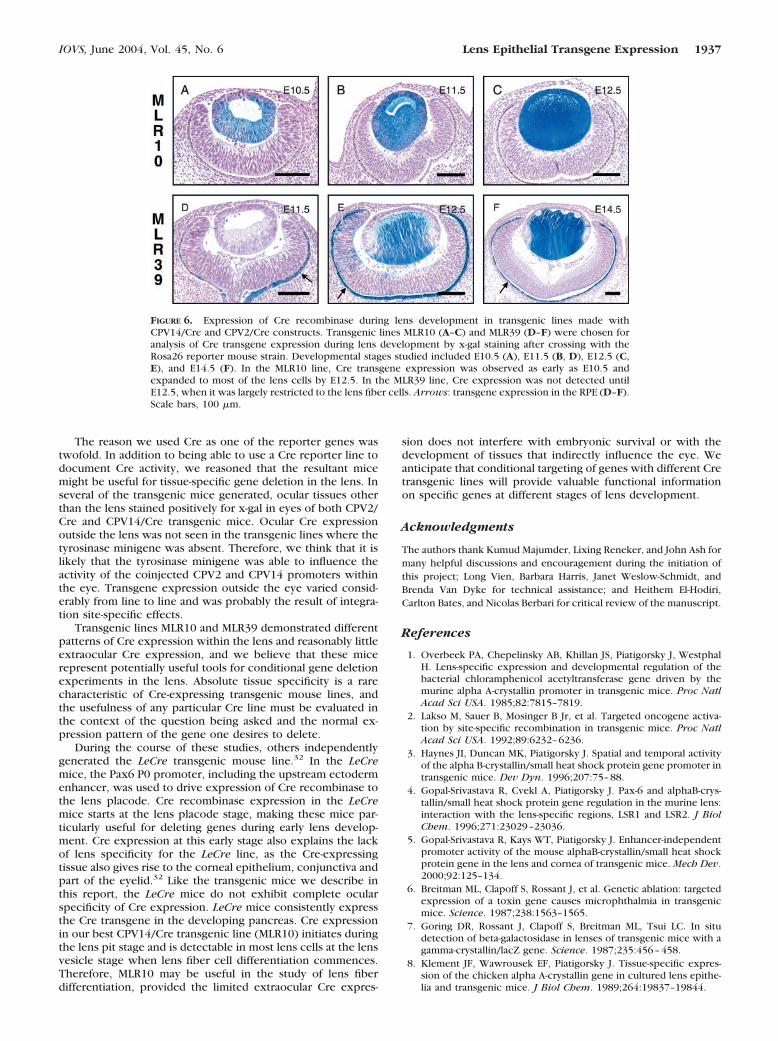
Lines MLR10 from the CPV14/Cre lines and MLR39 from theCPV2/Cre lines emerged as transgenic families in which therewas good transgene expression in the lens and minimal ex-traocular transgene expression. Although no evidence of Creexpression was detected in the lens placode from eitherMLR10 or MLR39 embryos (data not shown), in MLR10 mice,Cre expression was readily and consistently seen at E10.5during the lens pit–early lens vesicle stage (Fig. 6A). Over thenext 24 hours, evidence of Cre expression was detectable in
TABLE 2. Cre Expression in Ocular Tissues of Different Transgenic Mouse Lines
ConstructsTransgenic
LineExpression in
Lens EpitheliumExpression in
Lens Fiber CellsEctopic Expression
within the EyeTyrosinaseCoinjection
CPV14/Cre MLR10 ��� ��� No NoMLR32 ��� ��� Retina, RPE cornea YesMLR31 ��� ��� Retina, ciliary body YesMLR30 ��� ��� Retina, ciliary body YesMLR33 �� ��� RPE YesMLR28 �� ��� Retina YesMLR29 � � No Yes
CPV2/Cre MLR53 � ��� No NoMLR39 � ��� RPE, ciliary body YesMLR38 � ��� Retina, RPE, ciliary body, cornea YesMLR36 � �� Retina, RPE YesMLR37 � ��� Retina, RPE, cornea YesMLR35 � � Retina, RPE YesMLR34 � � Retina, RPE Yes
The intensity of x-gal staining was visually estimated as being weak (�), medium (��), strong (���) or not detectable (�).
1934 Zhao et al. IOVS, June 2004, Vol. 45, No. 6
the majority of both the lens epithelium and differentiating lensfibers (Fig. 6B), and this expression expands even more byE12.5 (Fig. 6C). Although there is some mouse-to-mouse vari-ability, the pattern of expression in the lens is quite consistent.Occasionally, a few cells in the center of the cornea demon-strate evidence of Cre expression, but we have never seen Creexpression in the neural retina or RPE of MLR10 mice. Outsidethe eye, MLR10 expression can usually be detected in the snoutand vibrissae follicles, as well as in parts of the midbrain andpituitary gland (Fig. 5A). Very little extraocular expression wasdetected in MLR39 embryos. Within the MLR39 eye, Cre ex-pression in the RPE was evident by E11.5 (Fig. 6D), but lensexpression of Cre was not evident at this stage. In the MLR39lens, Cre activity was first evident at E12.5 when most of thefiber cells stain with x-gal (Fig. 6E). Cre activity was only rarelydetected in lens epithelial cells, but most lens cells initiated Creexpression subsequent to a fiber differentiation signal (Figs. 6E,6F). Although the MLR39 RPE consistently stained with x-gal,blue-stained cells were not consistently visible in the neuralretina (compare Figs. 6D and 6F with 6E).
DISCUSSION
As expected, the �282/�43 fragment of the murine �A-crys-tallin promoter was effective in driving transgene expressiononly in lens fiber cells. Therefore, additional distal regulatoryelements are likely to be required for the endogenous expres-sion of the murine �A-crystallin gene in the lens epithelium. Incontrast, the �214/�38 fragment of the murine �B-crystallinpromoter consistently directed reporter gene expression to thelens epithelium and RPE and, in all but one case, failed to directdetectable reporter gene expression in the lens fiber cells. Thispattern of expression was largely unaltered (with the excep-tion of corneal epithelial expression in one founder) by theaddition of the �282/�86 �A-crystallin promoter fragment andthe �B-crystallin first intron and polyadenylation sequence.These elements were added to the �B-crystallin promoter in anattempt to increase the overall expression level of the trans-gene, but we reasoned that these modifications (particularlythe inclusion of the �A-crystallin promoter fragment) mightalter promoter specificity. The �282/�86 �A-crystallin pro-moter fragment contains a DE1 site �108/�100 that has been
FIGURE 3. Analyses of LacZ re-porter expression in eyes from Cretransgenic lines. After crossing to theROSA26 reporter strain, neonataleyes heterozygous for both the Cretransgenes and the ROSA26 reporterallele were analyzed for LacZ expres-sion (blue), reflecting the activity ofCre recombinase in transgenic mice.The CPV14/Cre transgenic lineswere MLR10 (A, A�), MLR32 (B, B�),MLR31 (C, C�), MLR30 (D, D�),MLR33 (E, E�), MLR28 (F, F�), andMLR29 (G, G�). The CPV2/Cre trans-genic lines were MLR53 (H, H�),MLR39 (I, I�), MLR38 (J, J�), MLR36(K, K�), MLR37 (L, L�), MLR34 (M,M�), and MLR35 (N, N�). The stainingpattern is shown in low magnifica-tion (A–N) to show total ocular re-porter expression, and the boxedregions are shown in high magnifica-tion (A�–N�) to distinguish lens epi-thelial (arrowhead) from lens fiber(✱ ) staining. Sections were counter-stained with nuclear fast red. Scalebars, 200 �m.
IOVS, June 2004, Vol. 45, No. 6 Lens Epithelial Transgene Expression 1935
shown recently to bind c-Maf, a transcription factor essentialfor lens-specific expression of �-, �-, and �-crystallins52,17 butother regulatory elements of �A-crystallin promoter such as the�A-CRYBP1 site �66/�57,18,20,53 the Pax6 site �49/�3317,the TATA box –31/�26,21and additional proximal elements at–25/�12,21 �7/�5,54 and �/24/�4322,53 are not included inthe composite promoter. Therefore, the DE1 site was insuffi-cient to activate the �214/�38 �B-crystallin promoter in thelens fiber cells, suggesting that the DE1 element must interactwith other �A-crystallin promoter elements to confer expres-sion in lens fiber cells. Consistent with this interpretation, the�287/�85 murine �A-crystallin promoter fragment did notwork in concert with an SV40 promoter to activate transcrip-tion when transfected into lens epithelial explants.55
In contrast, the insertion of a single copy of a Pax6 consen-sus binding site in either orientation at �86, between the DE1site and the �A-CRYBP1 site of murine �A-crystallin promoterwas able to alter the expression pattern of this promoterindependently. During lens development, Pax6 expression ex-hibits distinct polarity, with higher level expression in the lensepithelium that gradually diminishes during fiber differentia-tion.30,31 In our modified �A-crystallin promoter, there are twocis-acting elements that bind Pax6 and stimulate the promoteractivity, one from the endogenous �A-crystallin promoter�49/�33,17 and the other from the inserted Pax6 consensusbinding site. Thus, the insertion of this perfect consensus Pax6binding site enhanced transgene expression in the lens epithe-lium without altering the lens specificity of the endogenous�A-crystallin promoter sequences. At least two binding sites forPax6 have been found in the lens epithelium-expressed
chicken �A-crystallin and �-crystallin genes as well as the mu-rine �B-crystallin gene, each of which contribute to optimalpromoter function.4,34,35 Recent studies indicated that c-Mafcould form a complex with the coactivator CBP/p300 to acti-vate the �A-crystallin promoter, but Pax6 does not directlyinteract with this complex.56 Because c-Maf expression can beupregulated by Pax6,57 Pax6 can regulate �A-crystallin expres-sion both directly and indirectly. The consensus Pax6 bindingsite inserted into the �A-crystallin promoter may recruit addi-tional Pax6, which may act in concert with the c-Maf–CBP/p300 complex to activate transcription. Previous studies havesuggested that Pax6 alone is not sufficient to activate the�A-crystallin promoter in the lens, despite the observation thatthe �49/�33 region of murine �A-crystallin gene is capable ofbinding Pax6 and stimulating promoter activity.17,58,59 Muta-tions in both the DE1 site and �A-CRYBP1 site in the �116/�46 fragment abolished the function of the �A-crystallin pro-moter in transgenic mice, despite the presence of the nativePax6 binding sequence.60
The �96/�76 region of the murine �A-crystallin promoter(where the consensus Pax6 site was inserted) has not beendemonstrated to bind nuclear proteins and modulate promoteractivity. In addition, replacement of the inserted Pax6 bindingsite with other cis regulatory elements in the modified murine�A-crystallin promoter did not lead to epithelial expression intransgenic mice, indicating that enhanced expression of trans-gene in the lens epithelium was specific to the Pax6 consensusbinding site (Robinson ML, et al. IOVS 1997;38:ARVO Abstract2694). Thus, we think it is unlikely that enhanced epithelialtransgene expression resulted from disruption of a negativeregulatory element by the Pax6 consensus insertion. The use ofdifferent reporters also demonstrated that the influence of theinserted Pax6 binding site was not dependent on particularsequences in the reporter genes. To date, we have not testedthe consensus Pax6 site in any other part of the �A-crystallinpromoter.
FIGURE 5. Extraocular Cre expression in CPV2/Cre and CPV14/Cretransgenic lines. Global Cre reporter expression was revealed by x-galstaining (blue) of embryos heterozygous for the Cre transgene andROSA26 reporter allele. Wholemount embryos from CPV14/Cre trans-genic mouse lines MLR10 (A), MLR32 (B), and MLR31 (C) and theCPV2/Cre transgenic lines MLR39 (D), MLR37 (E), and MLR34 (F) areshown. All embryos were from E12.5 except MLR34, in which E14.5embryos were stained. Arrows: strong positive x-gal staining in theembryonic lenses from different transgenic lines. Extraocular reporterexpression varied in independently generated transgenic lines inde-pendent of whether CPV2/Cre or CPV14/Cre constructs were used.
FIGURE 4. Detection of LacZ expression in lens epithelial wholem-ounts from mice transgenic both for Cre and the ROSA26 reportergene. Lens capsule and epithelium was removed from transgenic miceat approximately P21 and stained for �-galactosidase activity. Thesamples were counterstained with hematoxylin to visualize the nuclei.Lens epithelial wholemounts from CPV14/Cre transgenic mouse linesMLR10 (A) and MLR32 (B), as well as CPV2/Cre transgenic lines MLR39(C) and MLR37 (D), were examined. Nontransgenic lenses (E) andlenses transgenic only for the ROSA26 reporter gene (F) were used asthe negative control. Black arrow: sporadic positive staining of theMLR37 transgenic line; white arrows: sporadic negative staining of theMLR10 and MLR32 transgenic lines. Scale bar, 100 �m.
1936 Zhao et al. IOVS, June 2004, Vol. 45, No. 6
The reason we used Cre as one of the reporter genes wastwofold. In addition to being able to use a Cre reporter line todocument Cre activity, we reasoned that the resultant micemight be useful for tissue-specific gene deletion in the lens. Inseveral of the transgenic mice generated, ocular tissues otherthan the lens stained positively for x-gal in eyes of both CPV2/Cre and CPV14/Cre transgenic mice. Ocular Cre expressionoutside the lens was not seen in the transgenic lines where thetyrosinase minigene was absent. Therefore, we think that it islikely that the tyrosinase minigene was able to influence theactivity of the coinjected CPV2 and CPV14 promoters withinthe eye. Transgene expression outside the eye varied consid-erably from line to line and was probably the result of integra-tion site-specific effects.
Transgenic lines MLR10 and MLR39 demonstrated differentpatterns of Cre expression within the lens and reasonably littleextraocular Cre expression, and we believe that these micerepresent potentially useful tools for conditional gene deletionexperiments in the lens. Absolute tissue specificity is a rarecharacteristic of Cre-expressing transgenic mouse lines, andthe usefulness of any particular Cre line must be evaluated inthe context of the question being asked and the normal ex-pression pattern of the gene one desires to delete.
During the course of these studies, others independentlygenerated the LeCre transgenic mouse line.32 In the LeCremice, the Pax6 P0 promoter, including the upstream ectodermenhancer, was used to drive expression of Cre recombinase tothe lens placode. Cre recombinase expression in the LeCremice starts at the lens placode stage, making these mice par-ticularly useful for deleting genes during early lens develop-ment. Cre expression at this early stage also explains the lackof lens specificity for the LeCre line, as the Cre-expressingtissue also gives rise to the corneal epithelium, conjunctiva andpart of the eyelid.32 Like the transgenic mice we describe inthis report, the LeCre mice do not exhibit complete ocularspecificity of Cre expression. LeCre mice consistently expressthe Cre transgene in the developing pancreas. Cre expressionin our best CPV14/Cre transgenic line (MLR10) initiates duringthe lens pit stage and is detectable in most lens cells at the lensvesicle stage when lens fiber cell differentiation commences.Therefore, MLR10 may be useful in the study of lens fiberdifferentiation, provided the limited extraocular Cre expres-
sion does not interfere with embryonic survival or with thedevelopment of tissues that indirectly influence the eye. Weanticipate that conditional targeting of genes with different Cretransgenic lines will provide valuable functional informationon specific genes at different stages of lens development.
Acknowledgments
The authors thank Kumud Majumder, Lixing Reneker, and John Ash formany helpful discussions and encouragement during the initiation ofthis project; Long Vien, Barbara Harris, Janet Weslow-Schmidt, andBrenda Van Dyke for technical assistance; and Heithem El-Hodiri,Carlton Bates, and Nicolas Berbari for critical review of the manuscript.
References
1. Overbeek PA, Chepelinsky AB, Khillan JS, Piatigorsky J, WestphalH. Lens-specific expression and developmental regulation of thebacterial chloramphenicol acetyltransferase gene driven by themurine alpha A-crystallin promoter in transgenic mice. Proc NatlAcad Sci USA. 1985;82:7815–7819.
2. Lakso M, Sauer B, Mosinger B Jr, et al. Targeted oncogene activa-tion by site-specific recombination in transgenic mice. Proc NatlAcad Sci USA. 1992;89:6232–6236.
3. Haynes JI, Duncan MK, Piatigorsky J. Spatial and temporal activityof the alpha B-crystallin/small heat shock protein gene promoter intransgenic mice. Dev Dyn. 1996;207:75–88.
4. Gopal-Srivastava R, Cvekl A, Piatigorsky J. Pax-6 and alphaB-crys-tallin/small heat shock protein gene regulation in the murine lens:interaction with the lens-specific regions, LSR1 and LSR2. J BiolChem. 1996;271:23029–23036.
5. Gopal-Srivastava R, Kays WT, Piatigorsky J. Enhancer-independentpromoter activity of the mouse alphaB-crystallin/small heat shockprotein gene in the lens and cornea of transgenic mice. Mech Dev.2000;92:125–134.
6. Breitman ML, Clapoff S, Rossant J, et al. Genetic ablation: targetedexpression of a toxin gene causes microphthalmia in transgenicmice. Science. 1987;238:1563–1565.
7. Goring DR, Rossant J, Clapoff S, Breitman ML, Tsui LC. In situdetection of beta-galactosidase in lenses of transgenic mice with agamma-crystallin/lacZ gene. Science. 1987;235:456–458.
8. Klement JF, Wawrousek EF, Piatigorsky J. Tissue-specific expres-sion of the chicken alpha A-crystallin gene in cultured lens epithe-lia and transgenic mice. J Biol Chem. 1989;264:19837–19844.
FIGURE 6. Expression of Cre recombinase during lens development in transgenic lines made withCPV14/Cre and CPV2/Cre constructs. Transgenic lines MLR10 (A–C) and MLR39 (D–F) were chosen foranalysis of Cre transgene expression during lens development by x-gal staining after crossing with theRosa26 reporter mouse strain. Developmental stages studied included E10.5 (A), E11.5 (B, D), E12.5 (C,E), and E14.5 (F). In the MLR10 line, Cre transgene expression was observed as early as E10.5 andexpanded to most of the lens cells by E12.5. In the MLR39 line, Cre expression was not detected untilE12.5, when it was largely restricted to the lens fiber cells. Arrows: transgene expression in the RPE (D–F).Scale bars, 100 �m.
IOVS, June 2004, Vol. 45, No. 6 Lens Epithelial Transgene Expression 1937

9. Yu CC, Tsui LC, Breitman ML. Homologous and heterologousenhancers modulate spatial expression but not cell-type specificityof the murine gamma F-crystallin promoter. Development. 1990;110:131–139.
10. Duncan MK, Roth HJ, Thompson M, Kantorow M, PiatigorskyJ. Chicken beta B1 crystallin: gene sequence and evidence forfunctional conservation of promoter activity between chicken andmouse. Biochim Biophys Acta. 1995;1261:68–76.
11. Duncan MK, Li X, Ogino H, Yasuda K, Piatigorsky J. Developmen-tal regulation of the chicken beta B1-crystallin promoter in trans-genic mice. Mech Dev. 1996;57:79–89.
12. McDermott JB, Cvekl A, Piatigorsky J. Lens-specific expression of achicken beta A3/A1-crystallin promoter fragment in transgenicmice. Biochem Biophys Res Commun. 1996;221:559–564.
13. Li X, Cvekl A, Bassnett S, Piatigorsky J. Lens-preferred activity ofchicken delta 1- and delta 2-crystallin enhancers in transgenic miceand evidence for retinoic acid-responsive regulation of the delta1-crystallin gene. Dev Genet. 1997;20:258–266.
14. Robinson ML, Overbeek PA. Differential expression of alpha A-andalpha B-crystallin during murine ocular development. Invest Oph-thalmol Vis Sci. 1996;37:2276–2284.
15. Kato K, Shinohara H, Kurobe N, Goto S, Inaguma Y, Ohshima K.Immunoreactive alpha A crystallin in rat non-lenticular tissuesdetected with a sensitive immunoassay method. Biochim BiophysActa. 1991;1080:173–180.
16. Srinivasan AN, Nagineni CN, Bhat SP. alpha A-crystallin is ex-pressed in non-ocular tissues. J Biol Chem. 1992;267:23337–23341.
17. Cvekl A, Kashanchi F, Sax CM, Brady JN, Piatigorsky J. Transcrip-tional regulation of the mouse alpha A-crystallin gene: activationdependent on a cyclic AMP-responsive element (DE1/CRE) and aPax-6-binding site. Mol Cell Biol. 1995;15:653–660.
18. Nakamura T, Donovan DM, Hamada K, et al. Regulation of themouse alpha A-crystallin gene: isolation of a cDNA encoding aprotein that binds to a cis sequence motif shared with the majorhistocompatibility complex class I gene and other genes. Mol CellBiol. 1990;10:3700–3708.
19. Kantorow M, Becker K, Sax CM, Ozato K, Piatigorsky J. Binding oftissue-specific forms of alpha A-CRYBP1 to their regulatory se-quence in the mouse alpha A-crystallin-encoding gene: double-label immunoblotting of UV-crosslinked complexes. Gene. 1993;131:159–165.
20. Brady JP, Kantorow M, Sax CM, Donovan DM, Piatigorsky J. Mu-rine transcription factor alpha A-crystallin binding protein. I. Com-plete sequence, gene structure, expression, and functional inhibi-tion via antisense RNA. J Biol Chem. 1995;270:1221–1229.
21. Sax CM, Cvekl A, Kantorow M, et al. Lens-specific activity of themouse alpha A-crystallin promoter in the absence of a TATA box:functional and protein binding analysis of the mouse alpha A-crys-tallin PE1 region. Nucleic Acids Res. 1995;23:442–451.
22. Ilagan JG, Cvekl A, Kantorow M, Piatigorsky J, Sax CM. Regulationof alphaA-crystallin gene expression: lens specificity achievedthrough the differential placement of similar transcriptional con-trol elements in mouse and chicken. J Biol Chem. 1999;274:19973–19978.
23. Dubin RA, Wawrousek EF, Piatigorsky J. Expression of the murinealpha B-crystallin gene is not restricted to the lens. Mol Cell Biol.1989;9:1083–1091.
24. Iwaki T, Kume-Iwaki A, Liem RK, Goldman JE. Alpha B-crystallin isexpressed in non-lenticular tissues and accumulates in Alexander’sdisease brain. Cell. 1989;57:71–78.
25. Gopal-Srivastava R, Piatigorsky J. The murine alpha B-crystallin/small heat shock protein enhancer: identification of alpha BE-1,alpha BE-2, alpha BE-3, and MRF control elements. Mol Cell Biol.1993;13:7144–7152.
26. Dubin RA, Gopal-Srivastava R, Wawrousek EF, Piatigorsky J. Ex-pression of the murine alpha B-crystallin gene in lens and skeletalmuscle: identification of a muscle-preferred enhancer. Mol CellBiol. 1991;11:4340–4349.
27. Gopal-Srivastava R, Haynes JI II, Piatigorsky J. Regulation of themurine alpha B-crystallin/small heat shock protein gene in cardiacmuscle. Mol Cell Biol. 1995;15:7081–7090.
28. Haynes JI II, Gopal-Srivastava R, Frederikse PH, Piatigorsky J. Dif-ferential use of the regulatory elements of the alpha B-crystallin
enhancer in cultured murine lung (MLg), lens (alpha TN4-1) andmuscle (C2C12) cells. Gene. 1995;155:151–158.
29. Gopal-Srivastava R, Piatigorsky J. Identification of a lens-specificregulatory region (LSR) of the murine alpha B-crystallin gene.Nucleic Acids Res. 1994;22:1281–1286.
30. Koroma BM, Yang JM, Sundin OH. The Pax-6 homeobox gene isexpressed throughout the corneal and conjunctival epithelia. In-vest Ophthalmol Vis Sci. 1997;38:108–120.
31. Richardson J, Cvekl A, Wistow G. Pax-6 is essential for lens-specificexpression of zeta-crystallin. Proc Natl Acad Sci USA. 1995;92:4676–4680.
32. Ashery-Padan R, Marquardt T, Zhou X, Gruss P. Pax6 activity in thelens primordium is required for lens formation and for correctplacement of a single retina in the eye. Genes Dev. 2000;14:2701–2711.
33. Marquardt T, Ashery-Padan R, Andrejewski N, Scardigli R, Guil-lemot F, Gruss P. Pax6 is required for the multipotent state ofretinal progenitor cells. Cell. 2001;105:43–55.
34. Cvekl A, Sax CM, Bresnick EH, Piatigorsky J. A complex array ofpositive and negative elements regulates the chicken alpha A-crys-tallin gene: involvement of Pax-6, USF, CREB and/or CREM, andAP-1 proteins. Mol Cell Biol. 1994;14:7363–7376.
35. Cvekl A, Sax CM, Li X, McDermott JB, Piatigorsky J. Pax-6 andlens-specific transcription of the chickens d1-crystallin gene. ProcNatl Acad Sci. 1995;92:4681–4685.
36. Duncan MK, Haynes JI II, Cvekl A, Piatigorsky J. Dual roles forPax-6: a transcriptional repressor of lens fiber cell-specific beta-crystallin genes. Mol Cell Biol. 1998;18:5579–5586.
37. Kralova J, Czerny T, Spanielova H, Ratajova V, Kozmik Z. Complexregulatory element within the gammaE- and gammaF-crystallinenhancers mediates Pax6 regulation and is required for inductionby retinoic acid. Gene. 2002;286:271–282.
38. Reza HM, Ogino H, Yasuda K. L-Maf, a downstream target of Pax6,is essential for chick lens development. Mech Dev. 2002;116:61–73.
39. Goudreau G, Petrou P, Reneker LW, Graw J, Loster J, Gruss P.Mutually regulated expression of Pax6 and Six3 and its implica-tions for the Pax6 haploinsufficient lens phenotype. Proc NatlAcad Sci USA. 2002;99:8719–8724.
40. Williams SC, Altmann CR, Chow RL, Hemmati-Brivanlou A, LangRA. A highly conserved lens transcriptional control element fromthe Pax-6 gene. Mech Dev. 1998;73:225–229.
41. Dimanlig PV, Faber SC, Auerbach W, Makarenkova HP, Lang RA.The upstream ectoderm enhancer in Pax6 has an important role inlens induction. Development. 2001;128:4415–4424.
42. Robinson ML, Overbeek PA, Verran DJ, et al. Extracellular FGF-1acts as a lens differentiation factor in transgenic mice. Develop-ment. 1995;121:505–514.
43. Ray MK, Magdaleno SW, Finegold MJ, DeMayo FJ. cis-Acting ele-ments involved in the regulation of mouse Clara cell-specific 10-kDa protein gene: in vitro and in vivo analysis. J Biol Chem.1995;270:2689–2694.
44. Frederikse PH, Dubin RA, Haynes JI II, Piatigorsky J. Structure andalternate tissue-preferred transcription initiation of the mouse al-pha B-crystallin/small heat shock protein gene. Nucleic Acids Res.1994;22:5686–5694.
45. Overbeek PA, Aguilar-Cordova E, Hanten G, et al. Coinjectionstrategy for visual identification of transgenic mice. TransgenicRes. 1991;1:31–37.
46. Taketo M, Schroeder AC, Mobraaten LE, et al. FVB/N: an inbredmouse strain preferable for transgenic analyses. Proc Natl Acad SciUSA. 1991;88:2065–2069.
47. Werner S, Weinberg W, Liao X, et al. Targeted expression of adominant-negative FGF receptor mutant in the epidermis of trans-genic mice reveals a role of FGF in keratinocyte organization anddifferentiation. EMBO J. 1993;12:2635–2643.
48. Soriano P, Generalized lacZ expression with the ROSA26 Crereporter strain. Nat Genet. 1999;21:70–71.
49. Robinson ML, MacMillan-Crow LA, Thompson JA, Overbeek PA.Expression of a truncated FGF receptor results in defective lensdevelopment in transgenic mice. Development. 1995;121:3959–3967.
1938 Zhao et al. IOVS, June 2004, Vol. 45, No. 6

50. Reneker LW, Overbeek PA. Lens-specific expression of PDGF-A intransgenic mice results in retinal astrocytic hamartomas. InvestOphthalmol Vis Sci. 1996;37:2455–2466.
51. Cammarata PR, Zhou C, Chen G, et al. A transgenic animal modelof osmotic cataract. Part 1: over-expression of bovine Na�/myo-inositol cotransporter in lens fibers. Invest Ophthalmol Vis Sci.1999;40:1727–1737.
52. Ring BZ, Cordes SP, Overbeek PA, Barsh GS. Regulation of mouselens fiber cell development and differentiation by the Maf gene.Development. 2000;127:307–317.
53. Kantorow M, Cvekl A, Sax CM, Piatigorsky J. Protein-DNA interac-tions of the mouse alpha A-crystallin control regions: differencesbetween expressing and non-expressing cells. J Mol Biol. 1993;230:425–435.
54. Sax CM, Cvekl A, Piatigorsky J. Transcriptional regulation of themouse alpha A-crystallin gene: binding of USF to the �7/�5region. Gene. 1997;185:209–216.
55. Chepelinsky AB, Sommer B, Wawrousek EF, Piatigorsky J. In:Piatigorsky J, Shinohara T, Zelenka PS, eds. Molecular Biology of
the Eye: Genes, Vision and Ocular Disease. New York: Alan R.Liss; 1988:215–227.
56. Chen Q, Dowhan DH, Liang D, Moore DD, Overbeek PA. CREB-binding protein/p300 co-activation of crystallin gene expression.J Biol Chem. 2002;277:24081–24089.
57. Sakai M, Serria MS, Ikeda H, Yoshida K, Imaki J, Nishi S. Regulationof c-maf gene expression by Pax6 in cultured cells. Nucleic AcidsRes. 2001;29:1228–1237.
58. Chepelinsky A, Sommer B, Piatigorsky J. Interaction between twodifferent regulatory elements activates the murine �A-crystallingene promoter in explanted lens epithelia. Mol Cell Biol. 1987;7:1807–1814.
59. Wawrousek EF, Chepelinsky AB, McDermott JB, Piatigorsky J.Regulation of the murine �A-crystallin promoter in transgenicmice. Dev Biol. 1990;137:68–76.
60. Sax CM, Ilagan JG, Piatigorsky J. Functional redundancy of theDE-1 and alpha A-CRYBP1 regulatory sites of the mouse alphaA-crystallin promoter. Nucleic Acids Res. 1993;21:2633–2640.
IOVS, June 2004, Vol. 45, No. 6 Lens Epithelial Transgene Expression 1939


















