
Inna Iņaškina. Cloning and biological activity of nuclear ...€¦ · Web viewInna Iņaškina....
73
UNIVERSITY OF LATVIA BIOMEDICAL RESEARCH AND STUDY CENTRE Inna Iņaškina CLONING AND BIOLOGICAL ACTIVITY OF NUCLEAR FACTOR OF ACTIVATED T CELL (NFAT) Abstract Author: Inna Iņaškina Supervisors: Professor Edgar Serfling Dr. biol. Ēriks Jankevics
Transcript of Inna Iņaškina. Cloning and biological activity of nuclear ...€¦ · Web viewInna Iņaškina....

UNIVERSITY OF LATVIA
BIOMEDICAL RESEARCH AND STUDY CENTRE
Inna Iņaškina
CLONING AND BIOLOGICAL ACTIVITY OF NUCLEAR FACTOR OF ACTIVATED T CELL (NFAT)
Abstract
Author: Inna Iņaškina
Supervisors: Professor Edgar Serfling
Dr. biol. Ēriks Jankevics
Riga 2005

SUMMARY
During the last decade when nuclear factor of activated T cells (NFAT) have been intensively studied, is became clear that calcium-regulated NFAT proteins play a major role in regulating inducible gene expression in the immune system, and that they also have important biological functions in other cell types, both during development and in the adult mammal.
In the present study we have cloned and investigated multiply NFATc1 and NFATc3 isoforms in lymphoid cells. The transcription factor NFATc1 is synthesised in three prominent isoforms. These differ in the length of their C terminal peptide and mode of synthesis. NFATc1/A, the shortest of three isoforms, is only one inducibly expressed in T cells. In contrast, the two longer isoforms NFATc1/B and NFATc1/C span extra C-terminal peptides of 128 and 246 aa, respectively, and are constitutively synthesised in many T cells. We also tested cloned cDNA of corresponding isoforms in transient transfection assays for promoter induction. NFATc1/A and NFATc1/C showed a very similar, if not identical, strong stimulatory effect on the IL-2 promoter, while NFATc1/B appeared to be the weakest transactivator. We also isolated three novel types of murine NFATc3 cDNA, designated as NFATc3-37, NFATc3-20, and NFATc3-l, and determined as isoforms alternatively spliced at the C-part. Using a number of specific antisera in Western Blotting assay, we found that NFATc3-37 is an exclusive form of NFATc3, constitutively expressed in the lymphoid tissues. An endogenous NFATc3-37 from thymocytes or EL4 cells, but not from Jurkat cells, bound to the distal NFAT site from the murine IL-2 promoter. This binding did not require the involvement of AP-1 related proteins. None of the isoforms overexpressed in 293 or Jurkat cells were able to activate the transiently transfected reporter gene driven by NFAT site from the IL-2 promoter. However, both NFATc3-37 and NFATc3-20 activated transcription via the NFAT I site from the Lck proximal promoter even without induction. Thus, albeit NFATc3-37 is widely expressed in lymphoid tissues, it might be involved in the regulation of the lck gene expression and, possibly, in the regulation of other genes during thymocyte development.
The second part of the work is devoted to the investigation of localisation and conditions of nuclear translocation of NFATc1, NFATc2, and NFATc3 in various T and B cell lines. The intracellular translocation of NFAT transcription factors is mediated by the activity of calcium-dependent serine/threonine phosphatase calcineurin and therefore is considered to be specifically inhibited by cyclosporin A (CsA). Our results suggest distinct behaviour of NFAT factors in response to treatment with CsA. Whereas the nuclear residence of NFATc2 and NFATc3 is always sensitive to treatment with CsA, i.e. these proteins are exported back to the cytoplasm, NFATc1 is considerably less affected by CsA and does not leave the cell nucleus. NFATc1 in high concentration forms granular structures within the cell nuclei. The treatment of cells with CsA leads to a decrease in the mobility of NFAT proteins and DNA-protein complex during electrophoresis due to hyperphosphorylation. These observations are validated by western blots, EMSA, immunostaining, and confocal microscopy of chimeric NFAT -green fluorescent protein. In addition, functional studies show that CsA inhibits transcription activity of endogenous NFATc1, but it has no effect on the overexpressed protein.
The transcriptional activation of NFAT factors is generally regulated by dynamic interplay between calcineurin and Ras/Raf/Erk protein kinase cascade on one hand and active serine/threonine protein kinases counteracting activity of calcineurin,

on the other hand. In the last part of the work we investigated the effect of glycogen syntase kinase-3 (GSK-3) and protein kinase A (PKA) on the transcriptional activity of NFAT family proteins - NFATc1/A, NFATc1/C, NFATc2 and NFATc3 using full-length expression vectors, NFATc1/Gal4 chimeras and different reporters. We observed that overexpression of GSK-3 dramatically inhibited transcriptional activity of all coexpressed proteins, while overexpression of PKA significantly increased the level of NFATc-mediated reporter gene production. We suggest that positive effect of PKA is a sum of several distinct effects caused by different mechanisms. The functional assay using NFATc1/Gal4 chimera with serine 269 mutated to alanine suppose that direct effect of PKA on the transcription activity of NFATc1 is accomplishing through phosphorylation of Ser269 and it is negative.
The work was carried out at the Biomedical Research and Study Centre, University ofLatvia and the Institute of Pathology, University of Würzburg, Germany.The study was supported by grants 96.0733, 01.0246, and 04.1145 of the LatvianCouncil of Science, the Graduirtenkollegs Regulation des Zellwachstums" (WürzburgDFG-2.22-10/96-II 13), DFG grant (DFG Wuerzburg; SE 469/12-1) and the NATOCollaborative Linkage Grant (HITECH.LG973289).Results of this work are reflected in four papers and one manuscript and have beenpresented in three international conferences and meetings.

CONTENTS
ORIGINAL PAPERS 5ABBREVIATIONS 6INTRODUCTION 7
Structure and evolution of NFAT proteins 7Activation of NFAT proteins 8NFAT-binding partners 9Biological function 10
RESULTS AND DISCUSSION 11The identification and characterisation of novel isoforms oftranscription factors NFATc1 and NFATc3 in lymphoid cells 11Constitutive and inducible synthesis of NFATc1 isoforms in lymphocytes 11The NFATc1 isoforms differ in their C and N terminal peptides 11NFATc1 isoforms differ in their transcriptional capacity 12Molecular cloning of murine NFATc3 cDNA 13The distribution of NFATc3 isoforms in lymphoid cell lines 15The binding activity of endogenous nuclear NFATc3 depends from cell type 17The transcription activity of NFATc3 isoforms 18Concluding remarks 19The different effect of immunosupressor cyclosporin A on the intracellulartrafficking and functional activity of NFAT transcription factors 20Nuclear export of NFAT proteins is differentially affected by CsA 20Cyclosporin A has no inhibitory effect on the translocation of NFATc1/A-GFP 23The transcription activity of the overexpressed NFATc1 is notCsA-sensitive in Jurkat cells 23The immuno fluorescence staining of endogenous NFATc1 in theJurkat and EL4 cells 25Concluding remarks 26The complex effect of Protein Kinase A on the transcriptionalactivity of NFAT family proteins 28Concluding remarks 30CONCLUSIONS 33REFERENCES 34

ORIGINAL PAPERS
This thesis is based on the following papers, which are referred by their Roman numerals in the text:
I. Iņaškina I., Serfling E. and Jankevics Ē. The Positive Effect of Protein KinaseA on the Transcriptional Activity of NFAT Family Proteins Occurs ViaDifferent Signal Transduction Mechanism, (in manuscript)
II. Iņaškina I., Serfling E. and Jankevics Ē. Intracellular Trafficking andFunctional Activity of Nuclear Factors of Activated T Cells Are DifferentlyAffected by Cyclosporine A. Proc Latvian Acad Sci, 2004, 58 (l):l-8.
III. Iņaškina I., Jackevica L., Zajakins P., Serfling E. and Jankevics Ē.Transcription Factor NFATc3 Isoforms in Lymphoid Cells. Proc Latvian AcadSci, 2003, 57 (1/2):11-16.
IV. Chuvpilo S., Avots A., Berberich-Siebelt F., Glockner J., Fischer C, Kerstan A.,Escher C, Inashkina I., Hlubek F., Jankevics E., Brabletz T., Serfling E.Multiple NFATc isoforms with individual transcriptional properties aresynthesized in T lymphocytes. J Immunol 1999,162 (12):7294-301.
V. Chuvpilo S., Zimmer M., Kerstan A., Glockner J., Avots A., Escher C, FischerC, Inashkina I., Jankevics E., Berberich-Siebelt F., Schmitt E., Serfling E.Alternative polyadenylation events contribute to the induction of NFATc ineffector T cells. Immunity, 1999,10 (2):261-9.
Results of this work have been presents in the following international conferences and scientific meetings:
I. Inashkina I., Jankevics E. and Serfling E. Nuclear translocation of NFATfactors. NATO/FEBS International Summer School on Molecular Mechanism ofsignal Transduetion. Island of Spetsai, Greece, August 16-28, 1999, Abstracts,p29.
II. Chuvpilo S., Zimmer M., Kerstan A., Glockner J., Avots A., Escher C. f FischerC, Inashkina I., Jankevics E., Berberich-Siebelt F., Schmitt E., Serfling E.Alternative splice/polyadenylation events contribute to the inducible synthesis ofNFATc in T effector cells. 15. Fruhjahrstagung Deutsche Gesellschaft furImmunologie, Stuttgart, March 03-06,1999, Abstracts, p106.
III. Chuvpilo S., Kerstan A., Glockner J., Avots A., Escher C, Fischer C,Inashkina I., Jankevics E., Siebelt F., and Serfling E. Coordinated synthesis andnuclear translocation of NFATc isoforms duting T cell activation. 28 th AnnualMeeting of the Deutsche Gesellschaft fur Immunologie, Würzburg, September25-27,1997. Immunobiol 1997,197:133-427, E 6.

ABBREVIATIONS
aa amino acidAb antibodiesAICD activation-induced cell deathAP-1 activator protein 1bp base pairCDK cycline-dependent kinaseCKI, II casein kinase I, IICn calcineurinCsA cyclosporin AEMSA electrophoretic mobility shift assayErk extracellular signal-regulated kinaseFasL Fas ligandGFP green fluorescent proteinGM-CSF granulocyte/macrophage colony-stimulating factorGSK-3 glycogen syntase kinase-3GTP guanosine triphosphateHDAC histone deacetylaseIL-2, 3, 4... interleukin-2, 3, 4...INF- interferon JNK c-Jun N-terminal kinaseMAP/SAP mitogen-activated protein/ stress activated proteinMEF2 myocyte enhancer factor-2NFAT nuclear factor of activated T cellsNFKB nuclear factor KBPAAG polyacrylamide gelPBL peripheral blood leukocytesPKA protein kinase APKC protein kinase CPTKs protein tyrosine kinasesSAP shrimp alkaline phosphataseSD standard deviationTCR T cell receptorTNF- tumour necrosis factor TonEBP tonicity enhancer binding proteinTPA 12-O-tetradecanoylphorbol 13-acetateWB western blot

INTRODUCTION
The NFAT (nuclear factor of activated T cells) family of transcription factors encompasses five proteins functionally and evolutionary related to the Rel/NFkB family of transcriptional activators [1, 2]. These include NFATc1 (NFATc/NFAT2), NFATc2 (NFATp/NFATl), NFATc3 (NFAT4/NFATx), NFATc4 (NFAT3), and NFAT5 (TonEBP: tonicity element binding protein or OREBP: osmotic response element binding protein) (reviewed in [3]). The NFATc1-c4 proteins are regulated by calcium/calcineurin dependent signalling - a rise in intracellular calcium activates the serine/threonine phosphatase calcineurin [4], which dephosphorylates NFATc1-c4 proteins [5]. NFATs are expressed in numerous cell type but they nevertheless play a particularly important role of activation, proliferation and activation-induced cell death (AICD) of lymphoid cells [4, 6-8].
Structure and evolution of NFAT proteinsThe structures of NFAT proteins are schematically compiled in Figure 1A. All NFAT proteins are characterised by the presence of a highly conserved DNA binding domain, often designated Rel similarity domain (RSD) due to sequence similarities with the DNA binding (Rel) domain of Rel/NFicB factors, which harbours the sequences motifs for DNA binding, interaction with AP-1 and, as shown for NFATc1, one nuclear localisation signal (NLS) [9], Moreover, they contain a regulatory domain in front of the RSD and at least one transactivation domain (TAD) near the N-terminus. The regulatory domain harbours numerous phosphorylation sites which are organised in the conserved serine-rich region (SRR) and three so-called SP motifs. These sites are substrates for several serine/threonine protein kinases and of the protein phosphatase calcineurine, which binds to this region (see Activation of NFAT proteins). In addition one NLS and one nuclear export signal (NES) have been identified within this domain of NFATc1 [9, 10].
All NFAT factors appear to be synthesised in several isoforms (Figure 1A) which can differ in both their N- and C-terminal peptides [11-16], The C-terminal sequences of the longest isoform NFATc1/C and NFATc2 harbour a second, albeit weak TAD [17, 18]. One isoform of NFATc3 (early designated NFATxl) revealed a strong TAD within 15 aa near its C-terminus which are highly conserved among all NFAT factors. Deletion of this C-terminal peptide led to a strong decrease in NFATxl activity after transfection into 293 and T cells. [14, 19].
Among NFAT proteins, NFAT5 is unique and unlike the calcium-regulated NFAT proteins. NFAT5 is constitutively dimeric, and dimerisation is essential for DNA binding and transcriptional activity [20-22]. The RSD of NFAT5 (Figure 1A) shares only 41-45% sequence similarity with the RSD of other NFATs [23]. Also NFAT5 is not regulated by calcium/calcineurin but rather works as a tonicity-responsive transcription factor required for certain aspects of T cell function as well as for kidney homeostasis and function [20-22]. Unlike genuine NFATs, NFAT5 does not interact with AP-1 factors, although it binds to the NFAT core binding motif TGGAAA. Moreover, NFAT5 lacks clear sequence homology to NFAT peptides outside the RSD, including the "regulatory region" [2].
NFAT5 seems to represent an evolutionary link between NFKB and genuine NFAT factors [2, 22]. In Drosophila, several NFkB-like proteins, such as Dorsal, Dif and Relish, have been detected which act downstream of signalling cascades similar to the NFicB-inducing cascades in mammalian cells. In analogy to mammalian RelA/p50 and RelB/pl05 NFKB complexes, Dif and Relish play important roles in the control of

Figure 1. Scheme of structure (A) and activation in T cells (B) of NFAT factors (from Serfling et al., 2004).A. DNA binding domains (RSD) are shown in yellow, transactivating domains (TAD), in red,and the regulatory domains of NFATs in blue. NLS, nuclear localisation signals; NES, nuclearexport signal; SP, serine-proline rich motifs; SRR, serine-rich motifs. CBP, CN indicates thebinding of transcriptional co-factor CBP/p300 and of phosphatase calcineurin (CN) to the N-termini of NFATs. NFATc1 was first cloned by Northrop et al. [24], NFATc2 by McCaffrey etal. [25], NFATc3 by Masuda et al. [26] and Hoey et al. [27], NFATc4 by Hoey et al. [27],respectively.B. NFAT activation: the central position of the calcium/calmodulin-dependent phosphatasecalcineurin in control of nuclear NFAT translocation and activation is indicated. It is thoughtthat calcineurin-mediated dephosphorylation of regulatory region (RR) unmasks NLSs andleads to their nuclear translocation. The immunosuppressants CsA and FK506 which bind tocalcineurin in concert with immunophilins are efficient inhibitors of calcineurin activity. Severalkinases, in particular GSK3, have been described to phophorylate the SRR and SP motifs inthe regulatory region of NFATs. The classicalRas/Raf/Erkcascade (but also other signalingmolecules, e.g. PKC members) was described to contribute to transacting capacity of NFATc1,but also in control of AP-1 activity.
innate immune responses by activating anti-microbial genes (reviewed in [28]). There is one NFAT-like gene of unknown function in the Drosophila genome, whose protein product has approximately 56% sequence identity with the RSD of NFAT5 and 42-43% identity with the RSDs of genuine NFATs [29]. Similarly, in the genome of the ancestral chordate Ciona intestinalis there is only one NFAT-like gene which harbours a RSD displaying 56% sequence identity at the protein level with the RSD of human NFAT5 and 38-40% identity with those of other human NFATs [30]. For teleost fish, NFAT-like transcription complexes controlled by a calcineurin-like activity have been described, and there are five NFAT genes in the genome of the teleost fish Fugu rubripes [31] corresponding to the five mammalian NFATs. This suggests that genuine NFATs appeared with the emergence of the adaptive immune system and lymphocytes approximately 500 million years ago, before the diversification of jawed fish.
Activation of NFAT proteinsThe efficient transcriptional activation of NFAT factors in T cells needs at least two signals, which are provided by activation of the TCR. These TCR-mediated signals lead

to (1) a rise in intracellular free calcium and calcineurin activation, and (2) the stimulation of several PTKs, e.g. p56lck, and p21ras and other small GTP bounding proteins, which activate a number of serine/threonine protein kinase cascades. While the activation of calcineurin mediates the nuclear translocation of NFAT factors [32], [33], activation of classical Ras/Raf/Erk and further protein kinase cascades controls the transcriptional activation of NFATs [34] and the induction of AP-1 [35].
In resting cells, NFAT proteins are phosphorylated, reside in the cytoplasm, and show low affinity for DNA in vitro. NFAT activation is initiated by dephosphorylation of the NFAT regulatory domain, a conserved ~300-aa region (Figure 1A). This domain is encoded in a single exon in all four NFAT proteins from all vertebrate species for which sequences are available [2]. Serine/threonine phosphatase calcineurin directly dephosphorylates serines of SRR and SP repeats located within regulatory domain, thus exposing NLS (Figure 1 A, B) and therefore leading to NFAT rapid nuclear import and increase in the affinity of protein for its target sites in DNA [36-42]. Inhibition of the phosphatase activity of calcineurin by FK506 or CsA results in the relocalisaion of NFAT to the cytosol and loss of its DNA-binding ability [43].
Although several serine/threonine protein kinases have been identified to phosphorylate NFAT proteins, thereby counteracting the activity of calcineurin, an integrated picture of NFAT phosphorylation has not yet emerged. CKI and GSK-3 are constitutive NFAT kinases that promote NFAT nuclear export [44, 45]; phosphorylation by GSK-3 requires prior phosphorylation by a priming kinase such as PKA [46, 47]. Consistent with its known sequence preference [48], GSK-3 phosphorylates the SPxx motifs of NFATc1 [44]. The MAP kinases p38 and JNK are inducible kinases that promote NFAT nuclear export, by selectively phosphorylating NFAT proteins at the SP sequences at the beginning of their SRR-1 regions: JNK1 phosphorylates NFATc1 and NFATc3 [49], whereas p38 selectively targets NFATc2 and NFATc4 [50, 51]. For JNK1, a proposed mechanism is that phospholylation of the calcineurin-docking site of NFATc1 blocks the interaction of NFATc1 with calcineurin [52].
NFAT-binding partnersSynergistic cooperation between NFAT and unrelated transcription factors AP-1 (Fos-Jun proteins) on composite DNA elements which contain adjacent NFAT (T/AGGAAAA/T) and AP-1 (TGAGTCA) binding sites regulates the expression of diverse inducible genes (reviewed in [53]). The outcome of cooperative interaction between NFAT and AP-1 is essential for a productive immune response, by stimulating the expression of a growing number of genes encoding cytokines, chemokines, and cell surface receptors (reviewed in [4]). The paradigm of NFAT-AP-1 cooperation has been recognised in a number of different cytokine promoters/enhancer regions (reviewed in [4, 6]). These include IL-2, IL-4, IL-5, GM-CSF, CD40L, IFN-y, IL-13, CTLA4, human TNF-a, human IL-8, and E-selectin. The strong cooperative binding of NFAT-AP-1 on specific DNA composite sites forms significantly more stable and higher affinity complexes than binding of the individual proteins alone [4].
In addition to its interaction with AP-1, NFAT engages in direct protein-protein interactions and influences transcription synergistically with several families of transcription factors: proteins such as Maf ICER that belongs to the same basic region-leucine zipper family as AP-1 [54, 55]; the zinc finger proteins GAT A (see below), and EGR [56]; the helix-turn-helix domain proteins Oct, HNF3, and IRE-4 [57-60]; the MADS-box protein MEF2 [8, 61, 62]; and the nuclear receptor PPAR-y [63]. Cooperation of NFAT with GATA family members has been observed in many systems such as functional assays using reporter constructs [64-66], yeast two-hybrid assay [64],

and co-immunoprecipitation experiments [67]. However, sequence inspection of regulatory regions has not led to unambiguous identification of a composite NFAT-GATA element with specific spacing and orientation, and cooperative NFAT-GATA binding on DN A has not been reported.
Biological functionNFATc1, NFATc2 and NFATc3 are expressed in cells of the immune system where they play a key role in regulating a large number of inducible genes during the immune response (reviewed in [4]). These include IL-2, IL-S, GM-CSF, IL-4, IL-5, IL-10 [68], IL-13, IFN-y, TNF-a, CD40L, FasL, CD5, IgK, CD25, IL-8, MIP4a and Cox 2. The transcriptional activity of NFATs may be either activating or deactivating depending on their binding partners. When NFATs cooperatively interact with activation partners such as AP-1, MEF2, and GATA, they activate a set of NFAT-regulated genes that mediate lymphocyte activation. On the other hand, interaction of NFAT with silencing complexes (e.g., HDACs) on specific gene loci (e.g., CDK4 promoter) turns off gene expression [69]. In addition, NFAT can induce expression of certain genes without the need for cooperative recruitment of Fos and Jun [70]. In this situation NFAT may turn on a completely new set of "anergy-associated genes" that mediates T cell anergy [71].
Mice that are deficient for both NFATc2 and NFATc3 show a striking allergic phenotype, suggesting a role for these proteins in suppressing production of the specific cytokines that control the development of allergy [72]. In contrast, T cells deficient in both NFATc1 and NFATc2 producing almost no cytokines upon stimulation, indicating that NFAT is essential for activating transcription of most T cell cytokine genes. At the same time, cells from these mice show an unprecedented phenotype of hyperproliferation, suggesting a role for NFAT in suppressing B cell responses [73].
All five NFAT proteins are also expressed in various non-lymphoid tissues, where they are involved in the regulation of diverse cellular functions in organs other than the immune system. The functions of NFAT proteins in extra-immune tissues have been largely inferred from the phenotypes of gene-disrupted mice lacking individual or multiple family members, and the identity of only a few target genes is known. Thus targeted disruption of the NFATc1 gene results in intrauterine death of the NFATc1-deflcient embryos due to a defect in cardiac valve formation [74, 75]. In muscle tissue NFATc1 is thought to cooperate with GATA2 to induce myocyte hypertrophy [67]. NFATc4 is expressed in hippocampal neurones where it induces transcription of NFAT dependent genes in response to depolarisation [76]. A calcineurin-dependent pathway that induces cardiac hypertrophy and involves a possible NFATc4-GATA4 cooperation has also been described [64]. Finally, NFAT proteins have been implicated in the regulation of chondrogenesis and adipogenesis [77, 78]. Whether these extra-immune NFAT-regulated processes require NFAT-AP-1 cooperation, or whether new transcriptional partners such as GATA proteins are required for NFAT activity outside the immune system, remains to be elucidated.
The aim of this work is investigation of biological functions of transcription factors NFATc1 and NFATc3 multiply isoforms.
In the present study we have focused on the three aspects of the investigation of NFAT family proteins: cloning and analysis of multiply NFATc1 and NFATc3 isoforms in lymphoid cells; localisation and conditions of nuclear translocation of NFATc1, NFATc2, and NFATc3 particularly in response to treatment with CsA in various cells; the effect of kinases GSK-3 and PKA on the NFAT proteins-mediated transcriptional activity.

RESULTS AND DISCUSSION
THE IDENTIFICATION AND CHARACTERISATION OF NOVEL ISOFORMS OF TRANSCRIPTION FACTORS NFATC1 AND NFATC3 IN
LYMPHOID CELLS
Although several common structural features among the NFATc proteins are known, but inactivation of the different NFATc genes in mice resulting in contrasting effects on cell differentiation and activation. One mechanism contributing to the pleiotropic effect of NFAT factors could be the mode of their expression. All NFAT factors appear to be synthesised in several isoforms that can differ in both their N- and C-terminal peptides [13, 14, 17]. In this chapter we report isolation and identification of human NFATc1 and murine NFATc3 isoforms. The properties of these proteins in expression manner and transcriptional activity were also investigated in different cells.
Constitutive and inducible synthesis of NFATc1 isoforms in lymphocytesThe nuclei of untreated Jurkat cells contain relatively low amounts of NFATc1 proteins larger than 100 kDa. After treatment with TPA plus ionomycin (TPA/Iono), the concentrations of these nuclear proteins increase steadily over a period of 3-4 hr (Figure 2A, lanes 1-5). However, 3-4 hr after treatment, a shorter prominent NFATc1 protein of about 90 kDa appears (Figure 2A, lane 5) that corresponds in size to NFATc cloned by Northrop et al. [24]. Similar patterns of nuclear NFATc1 proteins appear to be typical for many T and B cell lines, e.g., A3.01 T and Daudi B cells (Figure 2B and C, lanes 1-2). Other lymphoid cells express the 90 kDa NFATc1 protein in very different amounts. Thus, Raji B lymphoma cells synthesise it in low amounts (Figure 2C, lanes 3-4), EL-4 (Figure 9 A, lines 7-8) and L-5178Y T lymphoma cells in considerable, but H9 T cells not at all (Figure 2B, lanes 3-4).
To elucidate whether this heterogeneity in nuclear NFATc1 proteins is a peculiarity of lymphoid tumour cells, we investigated nuclear NFATc1 proteins from naive murine CD4+ T cells and Th1 and Th2 effector cells after differentiation in vitro. As shown in Figure 2D (line 1), naive T cells do not contain any detectable amount of nuclear NFATc1. TPA/Iono treatment for 6-12 hr leads to the predominant expression of two NFATc1 proteins larger than 100 kDa (Figure 2D, lane 2). TCR stimulation of these cells and differentiation for 8-10 days to Th1 or Th2 cells results in a nuclear accumulation of NFATc1, in particular of proteins larger than 100 kDa (Figure 2D, lines 2 and 5). However, stimulation of effector Th1 and Th2 cells with TPA/Iono gives rise to a predominant accumulation of the 90 kDa NFATc1 protein (Figure 2D, lines 4 and 6). The similar results were obtained in WB assay using nuclear proteins from PBL T cells. Figure 2E (line 1) shows that untreated PBL T cells did not contain any detectable nuclear NFATc1. Stimulation of PBL T cells with TPA/Iono resulted in the detection of NFATc1 isoforms B and C after 2 h (Figure 2E, line 2). After 6 h, NFATc1 isoform A of about 90 kDa became detectable (Figure 2E, line 3).
The NFATc1 isoforms differ in their C and N terminal peptidesThe occurrence of multiple nuclear NFATc1 proteins prompted us to clone NFATc1 cDNAs from a human Namalwa B cell cDNA library. Six out often positive NFATc1 cDNA clones isolated correspond to NFATc1 described previously [24], which we will designate as NFATc1/A. The residual four cDNAs were considerably longer. Two, designated as NFATc 1/B, lack the last C-terminal 19 aa residues of NFATc1/A and,

Figure 2. Three nuclear NFATc1 isoforms are expressed in lymphoid cells. A, B, C. 10 ng nuclear proteins from uninduced and induced for different time Jurkat T cells (A), uninduced and induced for 4 hr A3.01 and H9 T cells (B) or Raji and Daudi B cells (C) were fractionated and immunodetected using the 7A6 Ab. Proteins from 293 cells transfected with NFATc1/A, B, or C cDNA expression vectors were fractionated in the same gels (three right lines of each panel).D. WB of nuclear proteins isolated from the differentiated murine Th1 and Th2 effector cells.Naive CD4+CD62L+ T cells were isolated from the spleens of BALB/c mice, stimulated in vitrowith anti-TCRp Ab and anti-CD28 Ab, incubated in the presence of either IL-12 for Th1 or IL-4for Th2 differentiation, and stimulated with TPA and Iono for 5 hr as described previously [79].E. PBL-Ts were either left uninduced or were induced for 2-12 h with TPA+Iono and obtainednuclear proteins were electrophoretically separated and immunoblotted. The NFATc1 isoformsA, B, and C are indicated.
instead, contain a C-terminal stretch of 128 aa. A third cDNA, designated as NFATc1/C, encodes in addition to the 128 aa stretch an extra C terminal peptide of 118 aa and an N terminal peptide of 29 aa, instead of the 42 aa N terminal stretch of NFATc1/A. Both NFATc1/B and C cDNAs carry a 3’ untranslated mRNA sequence of 1844 or 1796 nucleotides, respectively, instead of the 360 nucleotides 3' tail found in NFATc1/A (Figure 3A).
The extra C terminal peptides in NFATc1/B and C show sequence homologies of 30.6% to those in NFATc2 (Figure 3B) [11]. The more proximal region (aa 685 to 813 in NFATc1/C) exhibits 36.7% homology while the proline-rich NFATc1/C-specific peptide shows a poor homology to NFATc2, except an identical decapeptide spanning the aa 911-920 inNFATc1/C (Figure 3B).
When the three NFATc1 cDNAs were expressed in human 293 cells, the proteins synthesised were identical in size to the NFATc1 proteins in Jurkat cells and other lymphoid tumour cells (Figures 3 A-C).
NFATc1 isoforms differ in their transcriptional capacityTo test the transcriptional activity of individual NFATc1 isoforms we cotransfected expression vectors for NFATc1/A, -B, or -C together with a variety of promoter/reporter gene constructs into 293 cells, which express only minor amounts of endogenous NFATc1 (data not shown). Figure 4A shows that all three NFATc1 isoforms were properly expressed and translocated into the nuclei as proteins of approximately 90, 110, and 140 kDa, respectively. Dephosphorylation of nuclear

Figure 3. Organisation of human NFATc1 isoforms.A. Scheme of NFATc1 cDNAs isolated from a human Namalwa B cell cDNA library. Thetransactivation domains of NFATc1 isoforms, TAD-A and TAD-B, and the Rel similaritydomain (RSD) are indicated. The 3* untranslated mRNA segments of 360 bp (in NFATc1/A),1844 bp (NFATc1/B), and 1796 bp (NFATc1/C) are shown as solid brown lines. No. 1 indicatesthe translational start codon and the numbers after the protein coding parts indicate the numbersof aa residues for each NFATc protein. The 5' untranslated mRNA segments are not shown.B. Sequence comparison between the extra C terminal peptides in NFATc 1/B and /C with thosein NFATc2 [11]. Identical aa residues between the C terminal peptides are indicated by longvertical dashes, similar residues by short dashes. Gaps are indicated by horizontal dashes. Notethe identical decapeptide near the C termini of NFATc1/C and NFATc2.
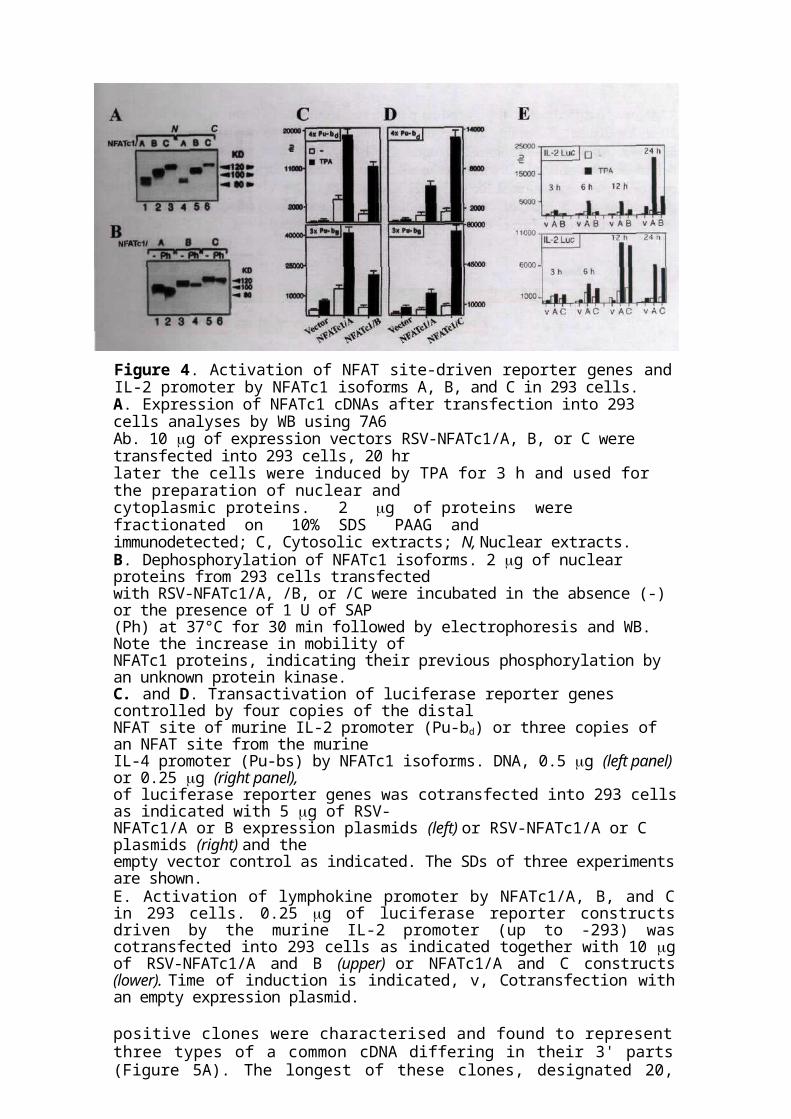
proteins led to an increase in their electrophoretic mobility, indicating phosphorylation of all three isoforms in these cells (Figure 4B). All three NFATc1 proteins stimulated the inducible activation of luciferase reporter genes driven by four copies of the distal NFAT site (Pu-bd) from the murine IL-2 promoter [80] or three copies of Pu-be from the murine IL-4 promoter (Figure 4C and D) [81]. In all transfection assays, NFATc 1/B was the weakest trans-activator (Figure 4C), whereas NFATc1/C appeared to be the strongest NFATc1, being 2-fold stronger than NFATc1/A in the activation of Pu-ba and 3-fold stronger hi PU-DB activation (Figure 4D).
We also tested the activities of individual NFATc1 isoforms on the TPA-mediated induction of the IL-2 promoters after transfection into 293 cells. Luciferase reporter gene construct governed by full-length murine IL-2 promoter was cotransfected with NFATc1 expression vectors. All three isoforms increased the induction of IL-2 promoters (Figure 4E), in particular after stimulation with TPA. In this assay, NFATc 1/B again appeared to be the weakest transactivator, leading to a 2- to 5-fold weaker activation of the IL-2 promoters than NFATc1/A. Surprisingly, NFATc1/A exerted the same or even a stronger effect on the induction of both promoters as the longest isoform NFATc1/C carrying two TADs (Figure 4E).
Molecular cloning of murine NFATc3 cDNAFull-length cDNA of human NFATc3 was used to screen a murine thymus cDNA library. High stringency hybridisation resulted in 41 positive plaques. The obtained

Figure 4. Activation of NFAT site-driven reporter genes and IL-2 promoter by NFATc1 isoforms A, B, and C in 293 cells.A. Expression of NFATc1 cDNAs after transfection into 293 cells analyses by WB using 7A6Ab. 10 g of expression vectors RSV-NFATc1/A, B, or C were transfected into 293 cells, 20 hrlater the cells were induced by TPA for 3 h and used for the preparation of nuclear andcytoplasmic proteins. 2 g of proteins were fractionated on 10% SDS PAAG andimmunodetected; C, Cytosolic extracts; N, Nuclear extracts.B. Dephosphorylation of NFATc1 isoforms. 2 g of nuclear proteins from 293 cells transfectedwith RSV-NFATc1/A, /B, or /C were incubated in the absence (-) or the presence of 1 U of SAP(Ph) at 37°C for 30 min followed by electrophoresis and WB. Note the increase in mobility ofNFATc1 proteins, indicating their previous phosphorylation by an unknown protein kinase.C. and D. Transactivation of luciferase reporter genes controlled by four copies of the distalNFAT site of murine IL-2 promoter (Pu-bd) or three copies of an NFAT site from the murineIL-4 promoter (Pu-bs) by NFATc1 isoforms. DNA, 0.5 g (left panel) or 0.25 g (right panel),of luciferase reporter genes was cotransfected into 293 cells as indicated with 5 g of RSV-NFATc1/A or B expression plasmids (left) or RSV-NFATc1/A or C plasmids (right) and theempty vector control as indicated. The SDs of three experiments are shown.E. Activation of lymphokine promoter by NFATc1/A, B, and C in 293 cells. 0.25 g of luciferase reporter constructs driven by the murine IL-2 promoter (up to -293) was cotransfected into 293 cells as indicated together with 10 g of RSV-NFATc1/A and B (upper) or NFATc1/A and C constructs (lower). Time of induction is indicated, v, Cotransfection with an empty expression plasmid.
positive clones were characterised and found to represent three types of a common cDNA differing in their 3' parts (Figure 5A). The longest of these clones, designated 20, had 3357 nucleotides and a predicted open reading frame of 903 amino acids with deduced molecular weight of 98 kDa.
There were a 92, 98, and 70% ammo acid identity with hNFATc3 hi the N-terminal, Rel homology domain, and C-terminal portions, respectively. Within the Rel homology domain, isolated molecule displayed a 67-70% homology to sequences of other members of the NFATc family that is characteristic feature of the family. The N-terminal part demonstrated another region containing a conserved motif characterised by a serine/proline repeat consensus sequence SPxxSPxxSPrxsxt[D/E][D/E]swl, which is itself repeated three times (Fig. 4B).
Two other isolated clones, designated 37 and 1, lacked the in-frame initiation codon and part of the 5’ coding region; therefore they represented a partial cDNA of alternatively spliced isoforms. The clone 1 had a 111-amino acid deletion of the C-part of the Rel homology domain replaced with a 13-amino acid unrelated sequence. Clone

37 showed a difference in the 3 ‘-coding region: replacement of 30-amino acids of clone 20 with a distinct 203-amino acid sequence. 3'untranslated parts of all clones also were different. Although N-terminus heterogeneity of NFATc factors has been reported, we determined the isolated cDNA clones as alternatively spliced forms differing in their C-part only. We reconstituted the full cDNA sequences of NFATc3 clones 37 and 1 by assembling the 5' end of clone 20 containing the initiation codon and the cDNA sequence of the other two clones respectively. The reconstituted clone 37 (NFATc3-37)
with a predicted length of 1080 amino acids was similar to human NFAT4 [27] and NFATX [26]. Therefore, NFATc3-37 represented the most abundant form, but the other two were novel isolated clones of NFATc3 that possibly exist in the cells.
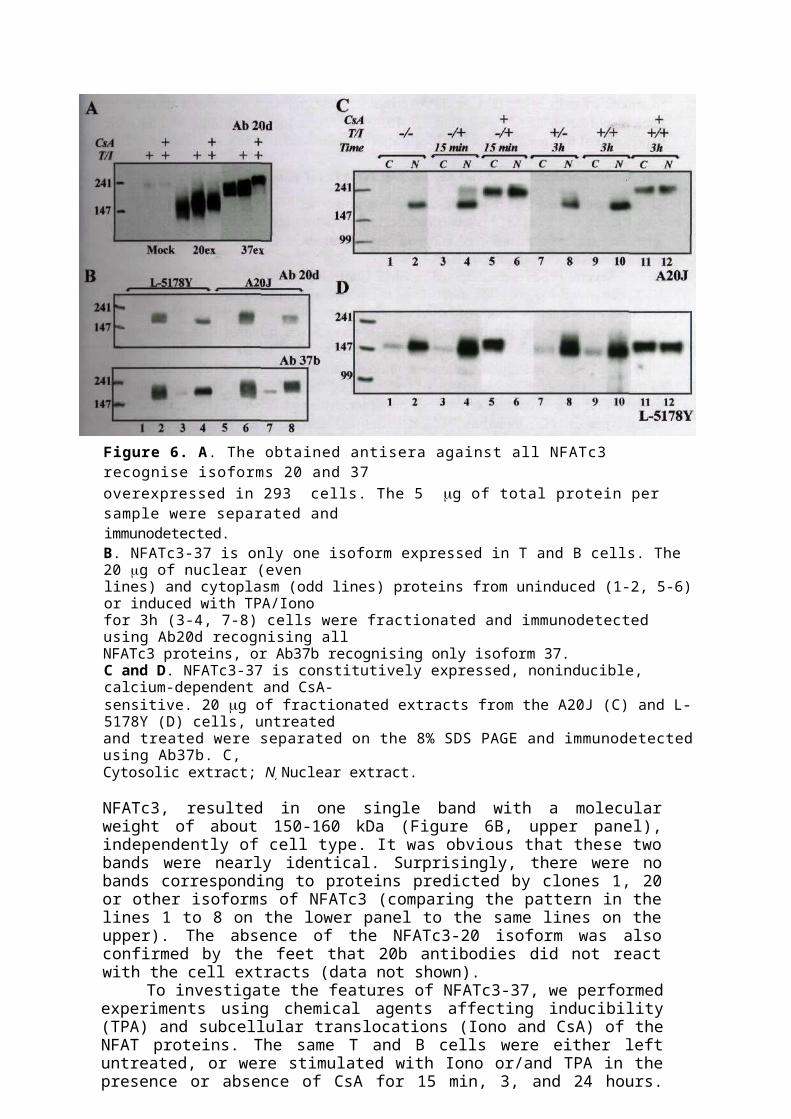
The distribution ofNFATc3 isoforms in lymphoid cell linesTo investigate the distribution of NFATc3 in different cell types and to distinguish the various isoforms, we raised antisera directed against unique isoforms (Figure 5A). In addition, we created an antiserum against a conserved motif in the N-terminus of NFATc3, recognising all of the NFATc3 isoforms. To test the ability of each serum to recognise its target protein in a specific manner, we prepared immunoblot assay using nuclear extracts from human 293 cells that were transiently transfected with pNFATc3-20ex and pNFATc3-37ex. As expected, the antiserum 20d recognised both isoforms, 20 and 37 (Figure 6A). Antisera 20b and 37b specifically reacted with the corresponding isoforms (data not shown).
To analyse the distribution of identified isoforms of NFATc3 and to investigate whether any of them had a predominant expression, we tested several lymphoid cell lines representing different types and developmental stages - EL4, L-5178Y, Jurkat TAg, A20J, and 70Z. Cells were treated (or not) with TPA and Iono for different periods of time, cytoplasmic and nuclear extracts were prepared, and WB experiments were performed. The same cell extracts were tested in parallel with 20b, 20d, and 37b antibodies. The 37b antibodies reacted with one single protein of about 155 kDa (Figure 6B, lower panel), the size of which corresponds to that predicted by the reconstituted clone 37 cDNA. The WB assay with 20d antibodies, recognising all isoforms of

Figure 6. A. The obtained antisera against all NFATc3 recognise isoforms 20 and 37overexpressed in 293 cells. The 5 g of total protein per sample were separated andimmunodetected.B. NFATc3-37 is only one isoform expressed in T and B cells. The 20 g of nuclear (evenlines) and cytoplasm (odd lines) proteins from uninduced (1-2, 5-6) or induced with TPA/Ionofor 3h (3-4, 7-8) cells were fractionated and immunodetected using Ab20d recognising allNFATc3 proteins, or Ab37b recognising only isoform 37.C and D. NFATc3-37 is constitutively expressed, noninducible, calcium-dependent and CsA-sensitive. 20 g of fractionated extracts from the A20J (C) and L-5178Y (D) cells, untreatedand treated were separated on the 8% SDS PAGE and immunodetected using Ab37b. C,Cytosolic extract; N, Nuclear extract.
NFATc3, resulted in one single band with a molecular weight of about 150-160 kDa (Figure 6B, upper panel), independently of cell type. It was obvious that these two bands were nearly identical. Surprisingly, there were no bands corresponding to proteins predicted by clones 1, 20 or other isoforms of NFATc3 (comparing the pattern in the lines 1 to 8 on the lower panel to the same lines on the upper). The absence of the NFATc3-20 isoform was also confirmed by the feet that 20b antibodies did not react with the cell extracts (data not shown).
To investigate the features of NFATc3-37, we performed experiments using chemical agents affecting inducibility (TPA) and subcellular translocations (Iono and CsA) of the NFAT proteins. The same T and B cells were either left untreated, or were stimulated with Iono or/and TPA in the presence or absence of CsA for 15 min, 3, and 24 hours. Figure 6C and D shows an example of expression pattern and localisation of protein in two types of cells - A20J (C) and L-5178Y (D). NFATc3-37 is constitutively expressed in lymphoid lineage cells and its relative amount varies from weak expression (in Jurkat cells) to high (in EL4 and 70Z, data not shown), which may reflect different roles of this transcription factor in different types of cells. The ratio between nuclear and cytoplasmatic protein is different among cell lines (Figure 6C and D, lanes 1-2), which can be explained by individual features of the cells. Treatment of the cells with Iono for 15 min leads to rapid translocation of protein into the nucleus, which can be inhibited partially or completely by CsA (Figure 6C and D, lines 3-6). Prolonged

treatment of cells with TPA or TPA/Iono does not lead to a significant increase of nuclear nor total NFATc3-37 (lines 7-10), confirming the constitutive noninducible expression of this protein. Nuclear localisation remains sensitive to CsA (lines 11-12).
The binding activity of endogenous nuclear NFATc3 depends from cell typeSince the published data regarding the binding properties of overexpressed NFATc3 are very discrepant [82],[83] and since transcription factors are often overexpressed to a level that may permit non-physiological binding in such studies, we investigated the binding activity of endogenous NFATc3.
Several bands were detected in EMSA with the distal NFAT site of the murine IL-2 promoter using nuclear extracts from stimulated and unstimulated Jurkat cells (data not shown), but the presence of NFATc3 within the complexes could not be proven by super shift. While the sensitivity of EMSA method is much higher than sensitivity of WB analysis, and since a weak band corresponding to NFATc3-37 was obtained in WB with Jurkat cells, we suggest that only a minimal part of nuclear NFATc3 is able to bind to this site. The one possible reason of that could be the inability of NFATc3-37 to bind to the NFAT site in context of the IL-2 promoter. Although NFATc3-37 is expressed in many types of cells, including PBL, its maximal expression has been detected in the thymus and its main function must lie within this area. From this point of view, the IL-2 promoter specific for PBL (Jurkat cells generally belong to this type) can not be the appropriate partner for NFATc3 in DNA-protein interactions. Secondly, efficient binding of NFAT family proteins to this DNA site may obligatory require the presence of AP-1-related proteins in the binding complex, as it has been observed for some of them (reviewed in [4]; [70]). In this context, the inability of NFATc3 to form a complex with AP-1 proteins leads to the inability to bind efficiently to the IL-2 NFAT site.
To elucidate these suggestions, nuclear extracts from induced or noninduced EL4 cells and thymocytes were examined in EMSA using the same NFAT site. Two inducible DNA-protein complexes were observed in the nuclear extracts from EL4 cells (Figure 7 A, lines 1,3,5). The upper complex gave a clear shift with Ab37b (Figure 7 A, lines 2,4,6) and therefore represented the NFATc3 complex with a distal NFAT site

from murine IL-2 promoter. TPA/Iono induced thymocytes found several binding complexes (Figure 7B, line 1). One band, corresponding to the heaviest complex, was shifted conspicuously by antibodies against NFATc3 (line 2). Additional super shift experiments using Ab against AP-1 proteins (cFos, FosB, JunB, and JunD) did not detect a contribution of these proteins in the upper complex nor in the others (lines 3-6), Additional proof of this is data suggesting that the thymocyte-specific NFAT binding site of the Lck proximal promoter is a composite element of NFAT and Myb sites, but not the AP-1 site (A. Avots, C. Stibbe, personal communications).
Taken together, the source of NFATc3 (the type of lymphoid cell) determines the binding activity of this protein with respect to different binding sites, while the origin of the promoters plays only a secondary role. This can be explained by specific conditions under which NFATc3 binds to the NFAT site with maximum efficiency. Possibly the involvement of a cofactor that interacts with NFATc3 is required for the sufficient binding of the complex to the DNA, or perhaps NFATc3 needs to become activated through specific modification (e.g. phosphorylation) that is not required in the case of NFATc1 or NFATc2. In any case, these specific conditions occur in the thymocytes and other lymphoid cells, but not in PBL or related cells (Jurkat).
The transcription activity ofNFATc3 isoforms
To determine whether NFATc3, in addition to binding to DNA in vitro, could also activate transcription from NFAT site in vivo, the NFATc3 expression constructs were overexpressed into 293 cells. Neither pNFATc3-20ex, nor pNFATc3-37ex, was able to induce expression of reporter gene driven by three copies of the distal NFAT binding site from murine IL-2 promoter into 293 cells, over those in cells transfected with mock plasmid DNA as a control, independently on stimulation (data not shown). WB and EMSA experiments using extracts from transiently transfected 293 cells confirmed a very high level of expression, but detected only weak binding of NFATc1 and did not
Figure 8. Transcription activity of the NFATc3 isoforms 37 and 20.A. Neither NFATc3-37 nor NFATc3-20 expression construct is able to transactivate the distalNFAT site from IL-2 promoter. 3 g of expression vectors pRSV-NFATc1/A, pNFATc2,pNFATc3-37ex, pNFATc3-20ex or mock plasmid were cotransfected into Jurkat cells with 1 gof a luciferase reporter gene construct driven by three copies of the distal NFAT site from theIL-2 promoter.B. Both NFATc3-20 and NFATc3-37 are able to activate transcription via an NFAT binding sitewithin the Lck proximal promoter. 3 g of expression plasmids were cotransfected into Jurkatcells with 1 g of luciferase reporter construct driven by four copies of NFAT I site from theLck proximal promoter.

detect any notable binding of overexpressed NFATc3 (data not shown). Similar results were observed in the transfections of Jurkat cells (Figure 8A). Transfection of the NFATc1 expression construct resulted in an increase of the inducible level of reporter gene expression. Despite this, there was no effect observed on the level of reporter gene production in cells transfected with the pNFATc3-37ex or pNFATc3-20ex plasmids.
Next, the NFATc3, NFATc2 and NFATc1 cDNA expression constructs were cotransfected into Jurkat cells with the reporter gene construct containing four copies of the NFATI binding site from the Lck proximal promoter. The Lck proximal promoter is active in the thymus only at an early developmental stage of T lymphopoesis, however it is essentially silent in peripheral T cells ([84]). Transfection of each of the NFATc family members resulted in an increase in the basal level of reporter gene expression in unstimulated Jurkat cells (Figure 8B), as compared to that in the cells transfected with mock plasmid as a control. NFATc3-37 appeared the strongest trans-activator within the family in the context of Lck promoter, activating reporter gene production by tenfold under the nonstimulating conditions. Upon stimulation, cells transfected with pNFATc3-20ex and pNFATc3-37ex did not show a significant increase in the level of reporter gene product in comparison to unstimulated cells. The transcription activity of pNFATc3-20ex and pNFATc3-37ex, in contrast to that of NFATc1 and NFATc2 was completely abolished by CsA.
Concluding remarksAll members of NFATc transcription factor family are expressed in multiple isoforms. This has been shown in detail for NFATc2 and NFATc3, which are expressed in several isoforms in T lymphocytes and other cells [11, 14]. The NFATc1 isoforms A, B and C described here are the most prominent NFATc1 proteins in lymphoid cells. In addition to alternative splicing at the 3'end, the detection of a different 5'peptide in NFATc1/C cDNA (Figure 3A) and published data [12, 13] suggest the existence of 5'heterogeneity in NFATc1 proteins. All three major NFATc1 proteins contain a strong transactivation domain, TAD-A, near the N terminus (Figure 3A, [34]).
Contrary to NFATc2 that is constitutively synthesised in numerous lymphoid cells, NFATc1 was reported to be inducibly expressed in T lymphocytes [13, 24]. However, this only is true for NFATc1/A, the shortest of three isoforms. In contrast, the two longer isoforms NFATc1/B and C are constitutively synthesised in many T cells, similar to NFATc2. All three NFATc2 isoforms are similar in length or even longer than NFATc1/C, the longest NFATc1 isoform. A short isoform lacking an extra C-terminal peptide, as does NFATc1/A, has also been described for NFATc3 [27], but it never have been shown whether it is inducibly synthesised like NFATc1/A. We also isolated three types of murine NFATc3 cDNA, one of them was short and lacked C-terminal part, therefore reminded NFATc1/A. However, further experiments identified long isoform NFATc3-37 as a prominent NFATc3 protein constitutively expressed in the lymphoid cells and did not found any protein corresponded to others cDNAs. NFATc3-37 protein is similar NFATc3 proteins, identified by other groups - human NFAT4 [27] and NFATX [26].
The C-terminal peptide of NFATc1/C shows >30% homology to a C-terminal QP-rich stretch of approximately 220 aa shared by NFATc2 isoforms and is able to act as a TAD [18]. Such a second TAD has also been identified within the C-terminal portion of the longest isoform of NFATc3, (early designated as NFATxl [14]). Thus, the existence of a C-terminal TAD is not a peculiarity of the long NFATc1 isoform C but is a typical component of numerous NFAT proteins.

The identification of TAD-B in NFATc1/C raises the question of which roles these isoforms play in gene control in T lymphocytes and other cells where they are expressed at different relative levels. When tested in transient transfections assays for promoter induction, NFATc 1/A and C showed a very similar, if not identical, strong stimulatory effect on the IL-2 (Figure 4E) and IL-4 [17] promoters while NFATc 1/B reached about 50% of this effect. Although NFATc factors alone are poor activators of the IL-5 promoter, in cooperation with other transcription factors, such us GATA-3 and Ets-1, they strongly stimulate the protein kinase A-mediated IL-5 promoter induction [66]. No marked differences among the three NFATc1 isoforms in respect to IL-5 promoter-driven reporter gene expression were detected when they were cotransfected with GATA-3 or/and Ets-1 [17]. The conspicuous differences between the NFATc1 isoforms in activation of the IL-2 and IL-4 promoters, on the one hand, and of the IL-5 promoter, on the other, suggest important functional roles of individual isoforms in promoter control. Several lines of evidence indicate that threshold levels of NFATc play a crucial role in the induction of promoters in T cells. In addition, due to the different transcriptional capacities of NFATc isoforms, changes in isoform composition will result in marked differences in specific transcriptional activity of nuclear NFATc1.
Unlike other NFATs, NFATc3 did not show efficient binding to the distal NFAT binding site from murine IL-2 promoter in the Jurkat cells (Figura 7A), and both NFATc3-20 and NFATc3-37, were not able to activate significantly transcription from the distal IL-2 NFAT DNA binding site. At the same time both could activate transcription via the NFAT binding site within the Lck proximal promoter (Figure 8). These results, together with the data on Lck promoter footprinting (A. Avots, personal communications), our EMSA results, and the expression of NFATc3 and NFATc2 in the thymocytes suggest that the main biological role of the NFATc3-37 lies in the regulation of the genes during the development of the thymocytes, especially lck gene.
THE DIFFERENT EFFECT OF IMMUNOSUPRESSOR CYCLOSPORIN A ONTHE INTRACELLULAR TRAFFICKING AND FUNCTIONAL ACTIVITY OF
NFAT TRANSCRIPTION FACTORS
How it has been described in details in Literature Review, all NFAT femily members have been reported as calcium/calcineurin-pathway-dependent [4, 33] and sensitive to the immunosupressive drugs CsA and FK506 [43]; reviewed in [85]. After induction of cells, NFAT factors rapidly translocate from cytoplasm to nucleus. The treatment of cells with immunosuppressant drugs blocks this translocation. However, our previous studies indicated that CsA might not affect nuclear localisation of NFATc1. In the presented work we investigated the cytoplasmic/nuclear localisation and conditions of nuclear translocation of NFATc1 in comparison with NFATc2 and NFATc3 in various human and murine T- and B- cell lines at distinct differentiation stages.
Nuclear export of NFAT proteins is differentially affected by CsA
To investigate the cytoplasmic/nuclear localisation of NFATc proteins, several lymphoid cell lines representing different types and developmental stages - EL4, L-5178Y, Jurkat TAg, A20J, and 70Z were treated with TPA and Iono in presence or absence of CsA for 15 min and 3 hours, cytoplasmic and nuclear extracts were prepared and immunoblot assays were performed. The same cell extracts were run in parallel on three gels and tested with antibodies against NFATc1, NFATc2, and NFATc3. Figure 9 shows the localisation of endogenous NFATc1 (A), NFATc2 (C), and NFATc3 (B) in

Figure 9. A, B, C. Cytoplasmic/nuclear localisation and translocation of NFATc proteins in EL4 cells. 20 g of fractionated extracts from cells differentially treated in the presence or absence of CsA were separated on the SDS PAAG and immunodetected using antibodies against NFATc1 (A), NFATc2 (C) and NFATc3 (B).D. Hyperphosphorylation of NFATc1 and NFATc3 in L-5178Y cells after treatment with CsA.Green arrows indicate the dephosphorylated form; red arrows indicate the hyperphosphorylatedform of NFATs. C, cytosolic extract; N, nuclear extract; the absence of contamination ofextracts was proved using antibodies against cytoplasm-specific protein lactate dehydrogenaseand nuclear protein upstream stimulatory factor (data not shown).E. Dephosphorylation of NFATc3 in A20J cells. 20 g of fractionated extracts from A20J cellstreated as indicated were incubated in the absence (-) or the presence of 1 U of SAP (Ph) at37°C for 30 min followed by electrophoresis and WB. Note the increase in mobility of NFATc3proteins from the cells treated with CsA after dephosphorylation by SAP, indicating then-previous phosphorylation.
differentially treated EL4 cells. The cytoplasmic fraction of untreated cells indicated very low amounts of NFAT proteins. This could be due to higher dilution of proteins during preparation of extracts. The three major isoforms of endogenous NFATc1, described in the first part of Results, similarly to NFATc2 and NFATc3, were localised in the nuclear fraction of untreated EL4 cells (Figure 9 A, B, and C, lines 2). The treatment of cells with Iono for 15 min led to additional accumulation of protein in the nucleus (compare lines 4 to lines 2), as it could be expected. Prolonged treatment of cells with TPA and Iono led to a considerable increase of nuclear protein (lines 8) due to the synthesis de novo. Treatment of cells with CsA before stimulation inhibited nuclear import of NFATc2 and NFATc3 and caused their partial accumulation in the cytoplasm. Under the same conditions, the nuclear residence of NFATc1 was not affected (Figure 9A, lines 5-6, 9-10). Similar results were obtained when cells were treated with CsA alone (Figure 9D).
The WB assays using extracts from the other cell lines showed similar results (data not shown) - while NFATc2 and NFATc3 always remained sensitive to CsA and were exported back to cytoplasm, NFATc1 accumulated in the nucleus and was not affected by CsA. An individual threshold level of calcineurin for each cell line may

Figure 10. The complex-formation between the distal NFAT site of the murine IL-2 promoter and NFATc1 proteins in the lymphoid cells is not disturbed by CsA treatment. EMSA was carried out with 2 or 2.5 g of nuclear proteins from EL4 (A) or Jurkat (B) cells treated as indicated. 1 l of undiluted Ab 7A6 was used for super shift (ss). The specific complexes detected are indicated by black arrows. The complexes shifted by Ab are indicated by green arrows. Note the shift of DNA-protein complexes from the cells treated with CsA.
explain the minor differences in expression pattern within different cell lines. For example, the NFATc1 pattern in resting EL4 cells is explained by the mutation in calcineurin gene that leads to high constitutive activity of endogenous calcineurin [86]. Two additional observations should be noted here. First, the relative concentrations of NFATc2 and NFATc3 in the cytoplasmic fraction from cells treated with CsA seemed to be considerably higher in comparison to that in the untreated cells (Figure 9B and C, compare lines 5-6 to 1-2). Such an increase in concentration could not be explained only by disruption of equilibrium between import and export of NFAT proteins by CsA. We therefore propose that additional NFAT was released from the subcellular compartment where it was not accessible during regular preparation of cell extracts. The intracellular membranes may be an example of such a compartment and CsA treatment may serve as a signal for the release of attached NFAT. Second, the treatment of cells with CsA led to hyperphosphorylation of all NFAT factors regardless to their localisation, which resulted in a decrease in mobility in the PAAG and a shift in WB (Figure 9A, B, and C, lines 5-6 and 9-10). Dephosphorylation of nuclear proteins by treatment with SAP led to an increase in their electrophoretic mobility (Figure 9E). The treatment of resting cells with CsA alone led to the same results (Figure 9D, lines 3-4 and 7-8), supporting the notion that such hyperphosphorylation occurs independent of the calcium/calcineurin signal.
The nuclear localisation of NFAT proteins itself does not guarantee its binding activity. In addition, results obtained in EMSA experiments using the distal NFAT site of the murine IL-2 promoter confirmed complex-formation between the NFAT binding site and NFATc1 proteins (Figure 10). Several DNA-protein complexes, apparently shifted by antibodies against NFATc1, were observed with extracts from EL4 (Figure 10A), Jurkat (Figure 10B), and A20J cells (data not shown). The treatment of cells with CsA did not disturb the complex-formation between NFAT binding site and NFATc1 protein, independently of the cell type. Moreover, comparison between EMSA pattern

of EL4 cells, nonstimulated and treated with CsA alone (Figure 10A, compare lines 1-2 with 5-6) or differently stimulated in the presence or absence of CsA as indicated (compare lines 3-4 with 7-8, 9-10 with 11-12 in Figure 10A, and also lines 9-10 with 11-12 in Figure 10B) revealed a slight shift of the bands, similar to the shift observed in WB.
Cyclosporin A has no inhibitory effect on the transhcation of NFATc1/A-GFPTo consolidate the WB results, we constructed chimeric proteins consisting of NFATc1 or NFATc3 and GFP. The chimeric constructs pF143-NFATc1 and pF143-NFATc3 were overexpressed in 293 cells and the subcellular localisation and CsA-dependent nuclear import/export of overexpressed proteins were examined by confocal microscopy.
The overexpressed NFATc3-GFP was localised in the cytoplasm of unstimulated 293 cells (Figure 11, image 4). The stimulation of cells with Iono resulted in the nuclear import of NFATc3-GFP, while the same treatment in the presence of CsA prevented it and allowed protein to remain in the cytoplasm (Figure 11, 5-6).
The other protein, NFATc1-GFP was expressed at a very high level and in the resting 293 cells was present in both compartments, but mostly in the nucleus (Figure 11, image 1). Similarly to NFATc3, stimulation of cells with Iono caused the translocation of NFATc1-GFP to the nucleus (Figure 11, 2). However, the presence of CsA during induction with Iono did not inhibit nuclear import of NFATc1-GFP (Figure 11, 3). Several cells with NFATc1 localised completely in the cytoplasm were found in uninduced and CsA/Iono-treated samples (Figure 11, images 7-8).
The transcription activity of the overexpressed NFATc1 is not CsA-sensitive in Jurkat cellsIn order to test whether CsA inhibits transcription activity of NFATc1 factors, the NFATc1/A and NFATc3 expression vectors were cotransfected into Jurkat cells with a reporter gene construct driven by four copies of NFAT I site from the Lck proximal promoter. Figure 12A shows that transfection of both pRSV-NFATc1/A and pNFATc3-37ex plasmids resulted in an increase in the inducible level of reporter gene expression.
Either inducible or basic transcription activity of overexpressed NFATc3 was completely abolished in the presence of CsA. There was no effect on the level of reporter gene production after the same treatment in the cells transfected with pRSV-NFATc1/A plasmid. However, reporter gene expression was clearly inhibited in the presence of CsA in the cells transfected with mock plasmid DNA as a control. Thus, the endogenous NFAT factors (NFATc1, NFATc2, and NFATc3) in toto are sensitive to the CsA.
Summarising data on functional studies we suppose that the difference of NFATc1 versus NFATc2 and NFATc3 could be quantitative rather qualitative, which would mean that translocation of NFATc1 is generally less sensitive to CsA than in case of the other NFAT factors. Confirmation experiments were performed by inducing Jurkat cells transfected with pRSV-NFATc1/A and pNFATc3-37ex plasmids with TPA/Iono in the presence of increasing concentration of CsA. The use of a two-fold higher concentration of CsA led to approximately a 30% decrease in the level of NFATc1/A activation (Figure 12B) in comparison to the cells treated with the conventional concentration of CsA. A further increase of the concentration of CsA did not result in significant inhibition of inducible transcription activity of NFATc1.
Figure 12. Cyclosporin A has no inhibitory effect on reporter gene transactivation by overexpressed NFATc1 in Jurkat cells. Standard conditions of stimulation (A) and increasing concentrations of CsA (B) were used.3 g of expression plasmids were cotransfected into Jurkat cells with 1 g of luciferase reporter construct driven by four copies of the NFAT I site from the Lck proximal promoter. The chart legends show the treatment of the cells: 2xCsA, two-times standard concentration of the CsA; 3xCsA, three-times standard concentration, et cetera. DMSO+TPA/Iono, control for the effect of solvent (CsA is dissolved in DMSO).
* not detected for mock transfection.
In the next experiment Jurkat cells were transiently transfected with different amount of pRSV-NFATc1/A expression plasmid and induced with TPA/Iono in the presence of CsA in conventional concentration. The results of this experiment (not shown) revealed that very low amounts of expression vector (up to 200 ng) did not increase the reporter gene activity over those in mock sample and did not affect it's CsA sensitivity. The further increase in amount (from 400 ng up to 1.5 g) resulted in decrease of sensitivity to CsA. At the same time the level of reporter gene expression

was comparable to that in mock transfection or only slightly higher. Under the concentration conveniently used in the functional assay (3-5 g/sample), transcription activity of overexpressed NFATc1/A was maximal but there was no inhibitory effect of CsA observed. Taken together, the transcription activity of overexpessed NFATc1/A was not inhibited by CsA, while the pool of endogenous NFATc factors was sensitive to CsA and transcription activity was abolished. In the similar experiment using Jurkat cells transfected with different amount of pNFATc3-37ex plasmid, reporter gene expression was completely inhibited in the presence of CsA independently on expression plasmid concentration.
The immunofluorescence staining of endogenous NFATc1 in the Jurkat and EL4 cellsProteins of the NFAT family, overexpressed in the 293 cells, are not always regulated according to physiological mechanisms in the lymphoid cells (see previous part). The transfection efficiency in Jurkat cells was too low to use the NFATc1-GFP and NFATc3-GFP constructs for fluorescent microscopy. Instead, immunofluorescence staining using antibodies against endogenous NFATc1 was performed with Jurkat and
Therefore, CsA decreased nuclear import of endogenous NFATc1 in the EL4 cells, but did not affect the NFATc1 overexpressed in the 293 cells. Similar data were obtained in an experiment using Jurkat cells (data not shown).

Concluding remarksWe investigated the cytoplasmic/nuclear localisation and conditions of nuclear translocation of NFATc1, NFATc2 and NFATc3 in various T- and B- cell lines. The WB studies showed a marked difference in the translocation manner between NFATc1 and other NFAT factors. Whereas the nuclear residence of NFATc2 and NFATc3 was always sensitive to treatment with CsA and these factors demonstrated cytoplasmic distribution (Figure 9B and C), NFATc1 was not affected by CsA and accumulated in the nucleus (Figure 9A). The similar results were obtained in the EMSA experiments using the distal NFAT site of the murine IL-2 promoter. Independently of the cell type, the treatment of cells with CsA did not disturb the complex-formation between NFAT binding site and NFATc1 protein, but resulted in a slight shift of the bands (Figure 10), similar to the shift observed in WB.
The examination of 293 cells, overexpressing chimeric proteins NFATc1/A-GFP and NFATc3-GFP, by confocal microscopy showed that nuclear import of NFATc3-GFP in stimulated cells was completely inhibited by CsA, while NFATc1/A-GFP accumulated in the nucleus and was not affected by CsA (Figure 11). Furthermore, functional studies using a reporter gene construct driven by four copies of the NFAT I site from the Lck proximal promoter demonstrated a high level of transcription activity of NFATc1 under conditions when transcription activity of NFATc3 was abolished by CsA (Figure 12A).
Thus, the observation of different responses of NFAT proteins to the treatment with inhibitor CsA was validated by different methods, WB and EMSA in the case of endogenous NFATc1 and confocal microscopy and transient transfection assay for the overexpressed NFATc1. We suggest that translocation of NFATc1 is generally less sensitive to CsA than in case of the other NFAT factors that is quantitative difference of NFATc1 versus NFATc2 and NFATc3. The transient transfection experiments performed with increasing concentration of CsA did not provide obvious supportive evidence for that hypothesis. A five-fold increase of CsA did not significantly affect NFATc1/A transcription activity (Figure 12B). Next, the titration of NFATc1/A expression vector showed that level of reporter gene production, considerably exceeding those in cells transfected with mock plasmid as a control, is not inhibited by CsA (data not shown), while transcription activity comparable to that of endogenous NFATc in control sample is abolished by CsA.
The results of immunofluorescence staining using antibodies against endogenous NFATc1 performed with Jurkat and EL4 cells showed that CsA was able to inhibit nuclear translocation of endogenous NFATc1 (Figure 13). Nevertheless, slight differences in the subcellular localisation in untreated and CsA treated cells could be observed. In untreated cells, NFATc1 was evenly distributed throughout the cytoplasm (Figure 13, image 3). After CsA treatment NFATc1 was mainly localised close to nuclear surface (Figure 13, image 9). We suggest that localisation of NFATc1 near the nuclear surface and its possible interaction with the nuclear membrane could be a cause of the consequent presence of protein in the nuclear fraction of cell extracts. Evidence to support this notion was a high concentration of NFAT proteins (data not shown) in WB experiments using nuclear membrane fractions.
Inhibition of the reporter gene production by CsA in the mock sample (Figure 12A), where transcription activity of transfected NFATc1/A did not decrease, can be explained in two ways. First, the pool of endogenous NFAT proteins in the Jurkat cells consists mainly not of NFATc1, but of NFATc2 and NFATc3, which are inhibited by CsA. However, the WB experiment showed that NFATc2 and NFATc1 are the major forms expressed in these cells and that NFATc3 was much less expressed. Second, the

Figure 14. The scheme of nuclear/cytoplasmic shuttling of transcriptions factors NFATc2/ NFATc3 (a-c) and NFATc1 (d-f).(a; d) The NFAT transcription factors are continuously subjects of transport, (b; e) The calcium-activated phosphatase calcineurin dephosphorylates NFAT's, resulting in their nuclear import. Dephosphorylated NFATc1 can form granular structures, (c) The immunosuppressant CsA inhibits calcineurin and therefore prevents nuclear import of NFATc2 and NFATc3, and enhances their return to the cytoplasm, (f) After exposure to the CsA, the majority of NFATc1 remains in the nucleus and localises near the nuclear membrane. The arrows indicate the direction of change in the balance between import and export of NFATc factors.
regulatory mechanism of the Jurkat cells, similarly to 293 cells, was not able to control properly the transcription activity of NFATc1, if it was overexpressed at very high level. NFATc 1-GFP when expressed in excess in 293 cells also formed granular structures within the nuclei (Figure 11, images 1-3). We suggest that these structures were formed by overexpressed protein itself or in interaction with other nuclear elements, and once being formed, further disturbed nuclear export. Such granular structures, therefore, could retain protein in the nucleus independently of the presence of CsA.
Our results suggest a differential effect of CsA on the endogenous and overexpressed NFATc1 and distinct behaviour of NFAT factors in response to treatment with CsA. While CsA inhibited transcription activity of endogenous NFATc1, it had no effect on the overexpressed protein.
Our observations are summarised in Figure 14. In the uninduced state NFAT's are localised in both nucleus and cytoplasm (Figure 14, a; d). After induction via the calcium pathway, the majority of the NFAT's rapidly translocates to the nucleus and only a minor fraction of the transcription factor remains in the cytoplasm (Figure 14, b; e). NFATc1 in a high concentration forms granular structures in the cell nuclei (Figure 14, e; f). The treatment of cells with CsA blocks nuclear translocation of NFAT's and enhances nuclear export of NFATc2 or NFATc3 (Figure 14, c), but exerts considerably less effect on NFATc1 (Figure 14, f). Despite the effect of CsA, the majority of NFATc1 does not leave the cell nucleus.
THE COMPLEX EFFECT OF PROTEIN KINASE A ON THE TRANSCRIPTIONAL ACTIVITY OF NFAT FAMILY PROTEINS
The relations between two kinases - GSK-3 and PKA and their involvement in the regulation of NFATc1 lie in the focus of this chapter. G. Crabtree and collegues described that GSK-3 phosphorylates NFATc1 only if it was first phosphorylated by PKA. PKA phosphorylates the NFATc1 directly at serines in positions 245, 269 and 294. The phosphorylation of PKA sites at Ser245 and Ser294 creates a series of overlaping GSK-3 substrate sites and therefore, PKA should be determinated as priming kinase for GSK-3 [44, 46, 47]. Recent data suggesting that interaction between PKA, GSK-3 and NFAT is not described by simple scheme: priming kinase - NFAT kinase - NFAT as a substrate. Here we investigate the molecular effects of PKA and GSK-3 on the function of NFATc1 in cells.
Figure 15. GSK-3 has an inhibitory, but PKA a stimulatory effect on the level of transactivation of the distal NFAT site from IL-2 promoter by overexpressed NFATc1/A (A), NFATc1/C (B), NFATc3 (C) and NFATc2 (D).0.4 g (A and B) or 1 g (C and D) of expression vectors pRSV-NFATc1/A, pRSV-NFATc1/C, pNFATc2, pNFATc3ex or corresponding amount of mock plasmid were cotransfected into Jurkat cells with 1 g of a luciferase reporter gene construct driven by three copies of the distal NFAT site from the IL-2 promoter alone or together with GSK-3 or PKA expression vector, in following amount - 0, 0.4, 0.8, 1.2, and 1.6 g of each or as combination of 1.2 g of both. Results of one representative experiment are shown here. Red columns indicate nonstimulating, but green columns stimulating conditions. m, mock plasmid.

In order to determine the effect of GSK-3 and PKA on the NFAT transcriptional activity, NFATc1/A was coexpressed in Jurkat cells either with increasing amount of GSK-3, or PKA, or both kinases together, and activities of reporter genes driven by three copies of the distal NFAT binding site from murine IL-2 promoter were measured. Figure 15A demonstrates that overexpression of GSK-3 led to a strong inhibition of NFAT-dependent reporter gene production, while overexpression of PKA resulted in significant increase in the level of NFATc1/A transcriptional activity. Parallel transfections, performed with pRSV-NFATc1/C, pNFATc2 and pNFATc3-37ex constructs, did not show as strong transactivation as NFATc1/A, but, in general, demonstrated similar pattern (Figure 15B - D).
In order to clarify is this up-regulation of reporter gene production a consequence of activation of NFAT or of AP-1 component, we repeated the same experiment using different reporter gene driven by four copies of NFAT I site from the Lck proximal promoter (data not shown). This site is a composite element of NFAT and Myb sites (A. Avots, C. Stibbe, personal communications) and does not contain AP-1 component. Like in the first experiment, overexpression of GSK-3 resulted in diminishing of transcriptional activity of all coexpressed proteins - NFATc1/A, NFATc 1/C, NFATc2, and NFATc3 In contrast, the presence of PKA did not show any significant effect on the level of reporter gene production in the cells expressing NFATc proteins (data not shown). These results suggested that up-regulating effect of the PKA on the reporter gene driven by three copies of the distal NFAT binding site from murine IL-2 promoter is explained by acting prevalently at the level of AP-1 component, and not NFATc component. Another explanation may be the presence of Myb binding site within the structure of NFAT I site. The influence of PKA on the reporter activity via Myb proteins is still unclear although both direct interaction between PKA and Myb and involving of other proteins in such interaction have been studied [87-89].
The strong inhibition of NFAT transcriptional activity by GSK-3 in the absence of PKA controverts with previously published data suggesting that GSK-3 completely opposes ionomycin-induced nuclear accumulation of NFATc1 only in synergism with PKA [46]. A possible explanation is that the limited endogenous PKA activity is sufficient to prime overexpressed GSK-3 or it may efficiently act with the aid of other endogenous kinases.
To create the conditions when any possibility of PKA to affect the level of reporter gene production via activating AP-1 proteins is completely excluded we constructed a Ga14/NFATc1 chimeric protein. This protein consists of the 2-400 aa of NFATc1/A harbouring predicted binding sites for the PKA and GSK-3 kinases, fused to the 1-147 aa of the yeast transcription factor Gal4 containing the DNA binding and dimerization domain. The use of such chimeric protein in cotransfection with luciferase reporter gene construct driven by five copies of Gal4 binding site avoids activation of reporter gene by endogenous NFATc proteins and influence of other transcription factors, particularly AP-1. The chimeric constructs pGal4/2-400NFATc1 and pGal4/2-400NFATc1m269, where serine 269 has mutated to alanine, were coexpressed in Jurkat cells with the PKA in different concentrations and activity of reporter gene was detected. Figure 16 shows that PKA enhanced NFATc-dependent production of reporter gene in cells transfected with both constructs - normal and mutated. This is similar to our results obtained in experiments using reporter driven by three copies of the distal NFAT binding site from murine IL-2 promoter, therefore proves that PKA affects the level of reporter gene production by enhancing of transcriptional activity of NFATc proteins, but without involving AP-1 proteins. Next, the reporter gene production in the

Figure 16. PKA enhances the transcription activity of both Gal4/NFATc1 chimeric proteins -wild type and those where serine 269 has mutated to alanine. 1 g of pGal4/2-400NFATc1, pGal4/2-400NFATc1m269 or mock plasmid were cotransfected into Jurkat cells with 1 g of luciferase reporter construct driven by five copies of Gal4 binding site and PKA expression vector in amount 0, 0.4, 0.8, and 1.6 fig. Results of one representative experiment are shown here.
cells transfected with mutated construct pGal4/2-400NFATc1m269 was significantly higher in comparison to cells transfected with wild type pGal4/2-400NFATc1. This means that inactivation of one of PKA phosphorylation sites results in up-regulation of NFATc1-mediated transcriptional activity.
Concluding remarksTaking together, the results of our experiments using full-length expression vectors as well Gal4/NFATc chimeras demonstrated that overexpression of GSK-3 dramatically inhibited transcriptional activity of all coexpressed proteins - NFATc1/A, NFATc1/C, NFATc2 and NFATc3 (Figure 15), while overexpression of PKA significantly increased the level of NFATc-mediated reporter gene production (Figure 15, 16).
Generally, the effect of PKA on the NFAT mediated reporter gene production does not have a single meaning, but is a result of several different mechanisms (Figure 17).
(1) The PKA works as the priming kinase for GSK-3. It has been shown thatGSK-3 phosphorylates NFATc1 only if it is first phosphorylated by PKA at Ser245 andSer294 that creates a series of overlaping GSK-3 substrate sites [44, 46, 47]. Then GSK-3phosphorylates NFATc1 (Figure 17, pathway 1) and, similar to several other kinases,exports it to the cytoplasm and therefore decreases transcriptional activity of NFATc1.
(2) PKA phosphorylates GSK-3 itself and inactivates it [90] that eventuallyinhibit the export of NFAT and could result in increase of transcriptional activity(Figure 17, pathway 2).
(3) PKA phosphorylates NFATc1 directly at serines in positions 245, 269 and294. The Ser245 and Ser294 are necessary for further phosphorylation by GSK-3 [46], butSer269 might be responsible for the direct effect of PKA on NFATc1 (Figure 17,pathway 3).

Figure 17. The scheme explaining the effect of PKA on the NFAT mediated reporter gene production as a sum result of several different mechanisms: 1) PKA works as the priming kinase for GSK-3; 2) PKA inactivates GSK-3 by phosphorylation; 3) direct effect of PKA on the NFAT transcriptional activity; 4) PKA activates transcription via AP-1. See Concluding remarks for the details.
(4) At last, the distal NFAT site from murine IL-2 promoter, often used as a part of a reporter construct in the transcriptional activity assays, can bind AP-1 proteins [36, 91]. At the same time, PKA can increase AP-1 activity [92, 93], therefore it is possible that up-regulation of reporter gene production occurs via activation of AP-1 component (Figure 17, pathway 4) and this effect may mask the real effect of PKA on the transcriptional activity of NFATc1. The use of Gal4/NFATc1 chimeric proteins instead of full-length NFATc1 avoids the involvement of AP-1 proteins and prevents the distortion of the results.
The first two mechanisms (Figure 17, pathways 1 and 2), GSK-3 priming by PKA and GSK-3 inactivation by PKA, start at the same point of pathway, carry on simultaneously and lead to the opposite results. Because of this it is very difficult to distinguish these two mechanisms and their effects in the frames of our experiment. The outcome of the third mechanism may be distinguished from the previous two. We assume that direct effect of PKA on the NFATc1 is accomplishing through its phosphorylation at Ser269 (Figure 17, pathway 3) and replacement of serine to alanine in this position will disrupt this effect. Mutation of Ser26 led to increase in the level of NFAT-dependent reporter gene production (Figure 16) and therefore suggested that phosphorylation of this site negatively affects NFATc1 transcriptional activity.
Altogether, we propose that in the different experiments under the diverse conditions, various mechanisms play a prevalent role. The NFATc proteins, overexpressed alone in the cells (Figure 15) are regulated by limited amounts of
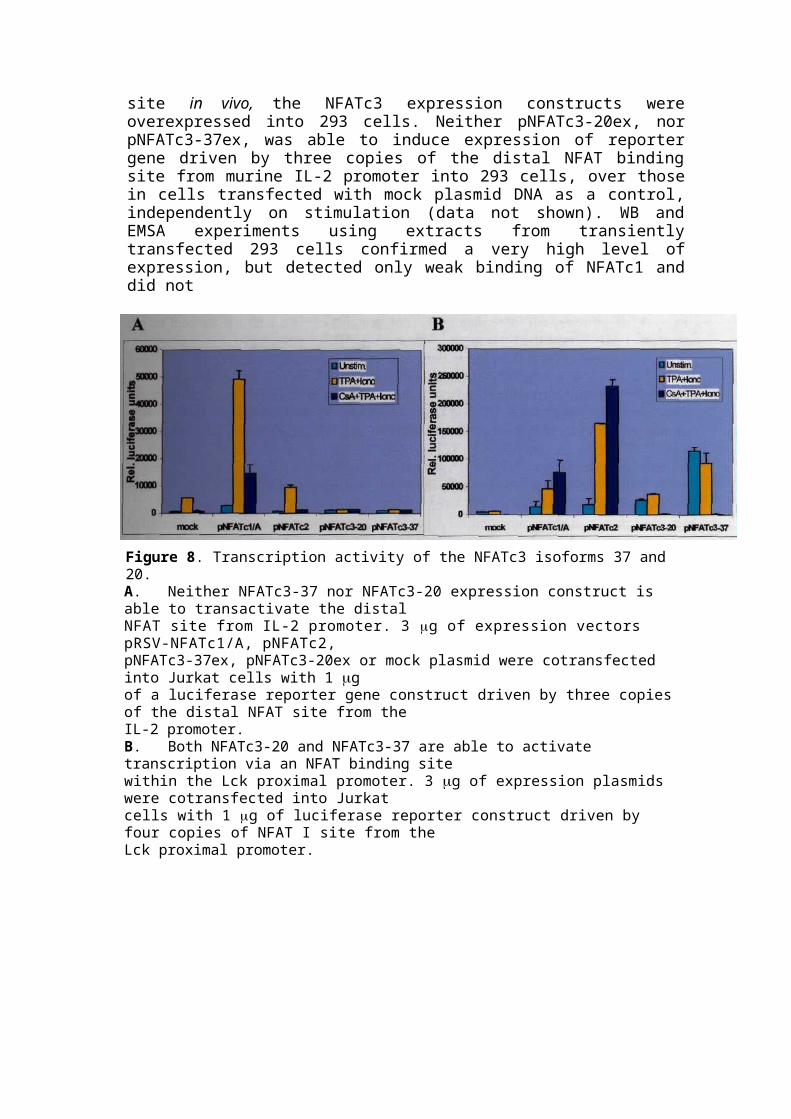
endogenous GSK-3 and PKA that resulted in moderate level of transcriptional activity. The coexpression of GSK-3 (Figure 15, data not shown) causes running of first mechanism and abolishing of NFATc-mediated transcriptional activity, while results of others two are masked, because the endogenous PKA is not sufficient to inactivate overexpressed GSK-3. The coexpression of PKA with NFATc1 (Figure 15, 16) causes inhibition of GSK-3 over usual level, deficiency of its active form and, as a result, arrest of the first mechanism. Therefore, in that case, NFATc1 transcriptional activity is a cumulative effect of the second and the third pathways.
The cells transfected with wild type or mutated Gal4/NFATc1 chimeric constructs are equally processing first two mechanisms while differ in respect to third pathway, which is inactivated in the cells expressing mutated fusion protein. This allow us to conclude that direct effect of PKA on the transcriptional activity of NFATc1 really exists, otherwise, there would not be difference between cells expressing wild type and mutant fusion proteins, and identify it as negative.

CONCLUSIONS
> Three isoforms of human transcription factor NFATc1, designated NFATc1/A,NFATc1/B and NFATc1/C, were cloned and characterised. The shortest isoformNFATc1/A corresponded to NFATc1 described previously, the two longer isoformsNFATc1/B and NFATc1/C spanned extra C-terminal peptides. The shortest isoformNFATc1/A is only one inducibly expressed in many T and B cell lines, and inTh1/Th2 cells as well.
> Three novel types of murine NFATc3 cDNA, designated an NFATc3-37, NFATc3-20 and NFATc3-l were isolated and determinated as isoforms alternatively splicedat the C-part. Using a number of raised specific antisera, NFATc3-37 isoform wasidentified as a prominent form of NFATc3, constitutively expressed in the lymphoidcells.
> An endogenous NFATc3-37 from thymocytes or EL4 cells, but not from Jurkatcells, was able to bind to the distal NFAT site from the murine IL-2 promoter. Thisbinding did not require the involvement of AP-1 related proteins. None of theisoforms were able to activate the transiently transfected reporter gene driven byNFAT site from the IL-2 promoter. However, both NFATc3-37 and NFATc3-20activated transcription via the NFAT I site from the Lck proximal promoter evenwithout induction.
> Distinct behaviour of NFATc factors in response to treatment withimmunosuppressant cyclosporin A (CsA) was observed. Whereas the nuclearresidence of NFATc2 and NFATc3 was always sensitive to treatment with CsA, i.e.these proteins were exported back to the cytoplasm, NFATc1 was considerably lessaffected by CsA and did not leave the cell nucleus. NFATc1 in high concentrationforms granular structures within the cell nuclei. The treatment of cells with CsA ledto a decrease in the mobility of NFAT proteins and DNA-protein complex duringelectrophoresis due to hyperphosphorylation. These observations were validated bywestern blots, EMSA, immunostaining, and confocal microscopy of chimericNFAT-GFP. In addition, functional studies showed that CsA inhibits transcriptionactivity of endogenous NFATc1, but it has no effect on the overexpressed protein.
> Overexpression of GSK-3 had dramatic inhibitory effect on the transcriptionalactivity of all coexpressed proteins - NFATc1/A, NFATc1/C, NFATc2 andNFATc3, while overexpression of PKA significantly increased the level of NFATc-mediated reporter gene production.
> The functional assay using Gal4/NFATc1 chimera suggested that effect of PKA isnot mediated via activation of AP-1 proteins. Replacement of serine to alanine inposition 269 of Gal4/NFATc1 construct led to increase in NFATc1-dependentreporter gene production. This may suggest that direct effect of PKA on thetranscription activity of NFATc1 is accomplishing through phosphorylation ofSer269 and it is negative.

REFERENCES1. Chytil M. and Verdine G.L. (1996) The Rel family of eukaryotic transcription factors.
Curr.Opin.Stuct.Biol. 6,91-100.
2. Graef I.A., Gastier J.M., Francke U. and Crabtree G.R. (2001) Evolutionary relationships amongRel domains indicate functional diversification by recombination. Proc.Natl.Acad.Sci.U.S.A 98,5740-5745.
3. Feske S., Okamura R, Hogan P.G. and Rao A. (2003) Ca2+/calcineurin signalling in cells of theimmune system. Biochem.Biophys.Res.Commun, 311,1117-1132.
4. Rao A., Luo C. and Hogan P.G. (1997) Transcription factors of the NFAT family: regulation andfunction. Annu.Rev.Immunol. 15, 707-747.
5. Clipstone N.A and Crabtree G.R. (1992) Identification of calcineurin as a key signalling enzymein T- lymphocyte activation. Nature 357, 695-697.
6. Hogan P.G., Chen L., Nardone J. and Rao A. (2003) Transcriptional regulation by calcium,calcineurin, and NFAT. Genes Dev. 17,2205-2232.
7. Serfling R, Berberich-Siebelt F., Chuvpilo S., Jankevics E., Klein-Hessling S., Twardzik T. andAvots A. (2000) The role of NF-AT transcription factors in T cell activation anddifferentiation(l) [In Process Citation], Biochim.Biophys.Acta 1498, 1-18.
8. Crabtree G.R. and Olson E.N. (2002) NFAT signaling: choreographing the social lives of cells.Cell 109 Suppl, S67-S79.
9. Beals C.R., Clipstone N.A, Ho S.N. and Crabiree G.R. (1997) Nuclear localization of NF-ATcby a calcineurin-dependent, cyclosporin- sensitive intramolecular interaction. Genes Dev. 11,824-834.
10. Klemm J.D., Beals C.R. and Crabtree G.R. (1997) Rapid targeting of nuclear proteins to thecytoplasm. Curr.Biol. 7, 638-644.
11. Luo C, Burgeon E., Carew J.A., McCaffrey P.G., Badalian T.M., Lane W.S., Hogan P.G. andRao A. (1996) Recombinant NFAT1 (NFATp) is regulated by calcineurin in T cells andmediates transcription of several cytokine genes. Mol.Cell Biol. 16, 3955-3966.
12. Park J., Takeuchi A. and Sharma S. (1996) Characterization of a new isoform of the NFAT(nuclear factor of activated T cells) gene family member NFATc [published erratum appears in JBiol Chem 1996 Dec 27; 271(52):33705]. J.Biol.Chem. 271, 20914-20921.
13. Lyakh L., Ghosh P. and Rice N.R. (1997) Expression of NFAT-family proteins in normal humanT cells. Mol.Cell Biol. 17, 2475-2484.
14. Imamura R., Masuda E.S., Naito Y., Imai S., Fujino T., Takano T., Arai K. and Arai N. (1998)Carboxyl-terminal 15-amino acid sequence of NFATxl is possibly created by tissue-specificsplicing and is essential for transactivation activity in T cells. J.Immunol. 161,3455-3463.
15. Chuvpilo S., Zimmer M., Kerstan A, Glockner J., Avots A., Escher C, Fischer C, Inashkina I.,Jankevics E., Berberich-Siebelt F., Schmitt E. and Serfling E. (1999) Alternativepolyadenylation events contribute to the induction of NF- ATc in effector T cells. Immunity. 10,261-269.
16. Sherman M.A., Powell D.R., Weiss DX. and Brown M.A. (1999) NF-ATc isoforms aredifferentially expressed and regulated in murine T and mast cells. J.Immunol. 162, 2820-2828.

10. Chuvpilo S., Avots A., Berberich-Siebelt F., Glockner J., Fischer C, Kerstan A., Escher C,Inashkina I., Hlubek F., Jankevics E., Brabletz T. and Serfling E. (1999) Multiple NF-ATcisoforms with individual transcriptional properties are synthesized in T lymphocytes. J.Immunol.162,7294-7301.
11. Luo C, Burgeon E. and Rao A. (1996) Mechanisms of transactivation by nuclear factor ofactivated T cells-1. J.Exp.Med 184,141-147.
12. Masuda E.S., Liu J., Imamura R., Imai S.I., Arai K.I. and Arai N. (1997) Control of NFATxlnuclear translocation by a calcineurin-regulated inhibitory domain. Mol.Cell Biol. 17, 2066-2075.
13. Lopez-Rodriguez C, Aramburu J., Rakeman A.S. and Rao A. (1999) NFAT5, a constitutivelynuclear NFAT protein that does not cooperate with Fos and Jun. Proc.Natl.Acad.Sci.U.S.A 96,7214-7219.
14. Lopez-Rodriguez C, Antos C.L., Shelton J.M., Richardson J.A., Lin F., Novobrantseva T.I.,Bronson R.T., Igarashi P., Rao A. and Olson E.N. (2004) Loss of NFAT5 results in renal atrophyand lack of tonicity-responsive gene expression. Proc.Natl.Acad.Sci.U.S.A 101,2392-2397.
15. Lopez-Rodriguez C, Aramburu J., Jin L., Rakeman A.S., Michino M. and Rao A. (2001)Bridging the NFAT and NF-kappaB families: NFAT5 dimerization regulates cytokine genetranscription in response to osmotic stress. Immunity. 15,47-58.
16. Miyakawa H, Woo S.K., Dahl S.C., Handler J.S. and Kwon H.M. (1999) Tonicity-responsiveenhancer binding protein, a rel-like protein that stimulates transcription in response tohypertonicity. Proc.Natl.Acad.Sci.U.S.A 96, 2538-2542.
17. Northrop J.P., Ho S.N., Chen L., Thomas D.J., Timmerman L.A., Nolan G.P., Admon A. andCrabtree G.R. (1994) NF-AT components define a family of transcription factors targeted in T-cell activation [see comments]. Nature 369,497-502.
18. McCaffrey P.G., Luo C, Kerppola T.K., Jain J., Badalian T.M., Ho A.M., Burgeon E., LaneW.S., Lambert J.N. and Curran T. (1993) Isolation of the cyclosporin-sensitive T celltranscription factor NFATp. Science 262, 750-754.
19. Masuda E.S., Naito Y., Tokumitsu H., Campbell D., Saito F., Hannum G, Arai K. and Arai N.(1995) NFATx, a novel member of the nuclear factor of activated T cells family that is expressedpredominantly in the thymus. Mol.Cell Biol. 15,2 697-2706.
20. Hoey T., Sun Y.L., Williamson K. and Xu X. (1995) Isolation of two new members of the NFAT gene family and functional characterization of the NF-AT proteins. Immunity. 2, 461-472.
21. Serfling E., Berberich-Siebelt F., Avots A., Chuvpilo S., Klein-Hessling S., Jha M.K., Kondo E.,Pagel P., Schulze-Luehrmann J. and Palmetshofer A. (2004) NFAT and NF-kappaB factors-thedistant relatives. IntJ.Biochem.Cell Biol. 36, 1166-1170.
22. Adams M.D. and others (2000) The genome sequence of Drosophila melanogaster. Science 287,2185-2195.
23. Dehal P. and others (2002) The draft genome of Ciona intestinalis: insights into chordate andvertebrate origins. Science 298,2157-2167.
24. Aparicio S. and others (2002) Whole-genome shotgun assembly and analysis of the genome ofFugu rubripes. Science 297, 1301-1310.
25. Liu J. (1993) FK506 and ciclosporin: molecular probes for studying intracellular signaltransduction. Trends Pharmacol. Sci. 14,182-188.

26. Crabtree G.R. (1999) Generic signals and specific outcomes: signaling through Ca2+,calcineurin, and NF-AT. Celt 96, 611-614.
27. Avots A., Buttmann M, Chuvpilo S., Escher C, Smola U., Bannister A.J., Rapp U.R.,Kouzarides T. and Serfling E. (1999) CBP/p300 integrates Rat7Rac-signaling pathways in thetranscriptional induction of NF-ATc during T cell activation. Immunity. 10, 515-524.
28. Treisman R. (1996) Regulation of transcription by MAP kinase cascades. Curr.Opin..Cell Biol. 8,205-215.
29. Jain J., McCaffrey P.G., Miner Z., Kerppola T.K., Lambert J.N., Verdine G.L., Curran T. andRao A. (1993) The T-cell transcription factor NFATp is a substrate for calcineurin and interactswith Fos and Jun. Nature 365, 352-355.
30. Luo C, Shaw K.T., Raghavan A., Aramburu J., Garcia-Cozar F., Perrino B.A., Hogan P.G. andRao A. (1996) Interaction of calcineurin with a domain of the transcription factor NFAT1 that controls nuclear import. Proc.Natl.AcadSci.U.S.A 93, 8907-8912.
38. Ruff V.A. and Leach K.L. (1995) Direct demonstration of NFATp dephosphorylation andnuclear localization in activated HT-2 cells using a specific NFATp polyclonal antibody.J.Biol.Chem. 270,22602-22607.
39. Park J., Yaseen N.R., Hogan P.G., Rao A. and Sharma S. (1995) Phosphorylation of thetranscription factor NFATp inhibits its DNA binding activity in cyclosporin A-treated human Band T cells. J.Biol.Chem. 270,20653-20659.
40. Okamura H., Aramburu J., Garcia-Rodriguez C, Viola J.P., Raghavan A., Tahiliani M., ZhangX., Qin J., Hogan P.G. and Rao A. (2000) Concerted dephosphorylation of the transcriptionfactor NFAT1 induces a con format ional switch that regulates transcriptional activity [In ProcessCitation]. Mol.Cell 6, 539-550.
41. Porter CM., Havens M.A. and Clipstone N.A. (2000) Identification of amino acid residues andprotein kinases involved in the regulation of NFATc subcellular localization. J.BiolChem. 275,3543-3551.
42. Neal J.W. and Clipstone N.A. (2000) Glycogen synthase kinase-3 inhibits the DNA bindingactivity of NFATc. J.Biol.Chem.
43. Liu J., Farmer J.D., Jr., Lane W.S., Friedman J., Weissman I. and Schreiber S.L. (1991)Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP- FK506 complexes. Cell66, 807-815.
44. Beals C.R., Sheridan C.M, Turck C.W., Gardner P. and Crabtree G.R. (1997) Nuclear export ofNF-ATc enhanced by glycogen synthase kinase-3. Science 275,1930-1934.
45. Zhu J., Shibasaki F-, Price R., Guillemot J.C, Yano T., Dotsch V., Wagner G., Ferrara P. andMcKeon F. (1998) Intramolecular masking of nuclear import signal on NF-AT4 by casein kinaseI and MEKK1. Cell 93, 851-861.
46. Neilson J., Stankunas K. and Crabtree G.R. (2001) Monitoring the duration of antigen-receptoroccupancy by calcineurin/glycogen-synthase-kinase-3 control of NF-AT nuclear shuttling.Curr.Opin.Immunol. 13,346-350.
47. Sheridan CM., Heist E.K., Beals C.R., Crabtree G.R. and Gardner P. (2002) Protein kinase Anegatively modulates the nuclear accumulation of NF-ATc1 by priming for subsequentphosphorylation by glycogen synthase kinase-3. J.Biol.Chem. 277, 48664-48676.
48. Harwood A.J. (2001) Regulation of GSK-3: a cellular multiprocessor. Cell 105, 821-824.
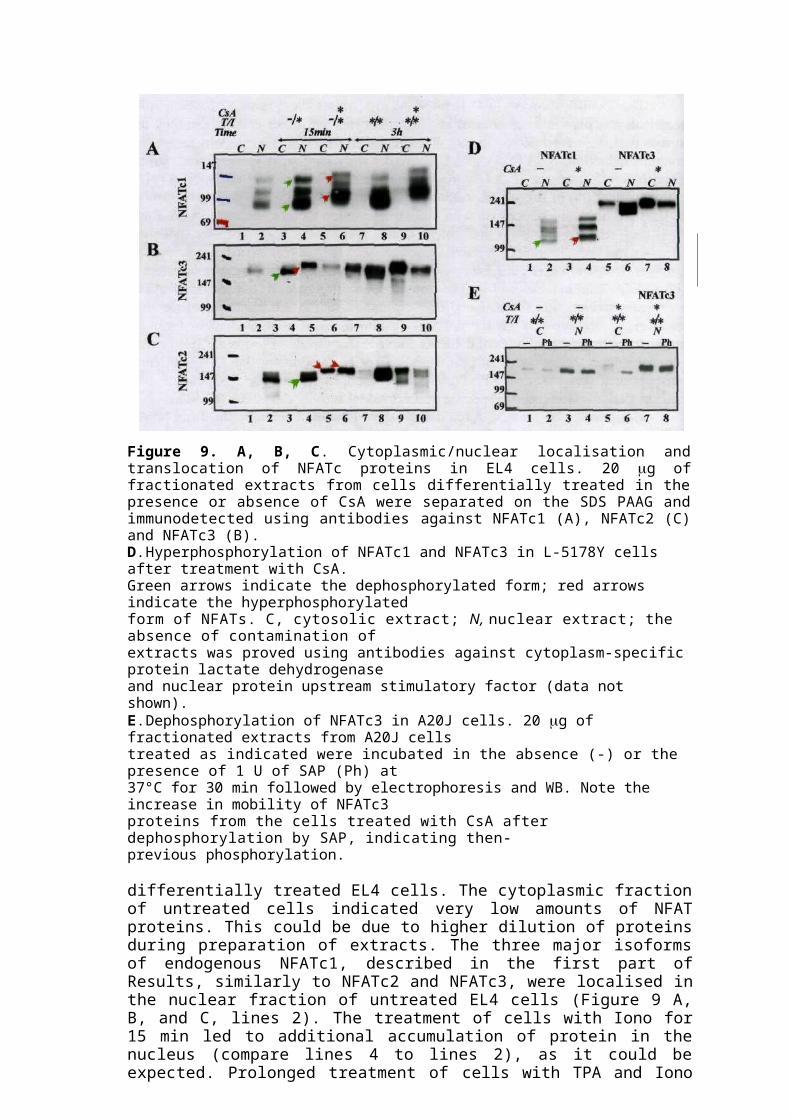
38. Chow C.W., Rincon M., Cavanagh X, Dickens M. and Davis R.J. (1997) Nuclear accumulationof NFAT4 opposed by the JNK signal transduction pathway. Science 278,1638-1641.
39. Gomez d.A., Martinez-Martinez S., Maldonado J.L., Ortega-Perez I. and Redondo J.M. (2000) Arole for the p38 MAP kinase pathway in the nuclear shuttling of NFATp. J.Biol.Chem. 275,13872-13878.
40. Yang T.T., Xiong Q., Enslen H., Davis R.J. and Chow C.W. (2002) Phosphorylation of NFATc4by p38 mitogen-activated protein kinases. Mol.Cell Biol. 22,3892-3904.
41. Chow C.W. and Davis R.J. (2000) Integration of calcium and cyclic AMP signaling pathways by14-3-3. MolCellBiol 20, 702-712.
42. Macian F., Lopez-Rodriguez C. and Rao A. (2001) Partners in transcription: NFAT and AP-1.Oncogene 20, 2476-2489.
43. Ho I.C., Hodge M.R., Rooney J.W. and Glimcher L.H. (1996) The proto-oncogene c-maf isresponsible for tissue-specific expression of interleukin-4. Cell 85,973-983.
44. Bodor J., Bodorova J. and Gress R.E. (2000) Suppression of T cell function: a potential role fortranscriptional repressor ICER. J.Leukoc.Biol 67,.774-779.
45. Decker E.L., Skerka C. and Zipfel P.F. (1998) The early growth response protein (EGR-1)regulates interleukin-2 transcription by synergistic interaction with the nuclear factor of activatedT cells. J.Biol.Chem. 273, 26923-26930.
46. Furstenau U., Schwaninger M., Blume R., Jendrusch E.M. and Knepel W. (1999)Characterization of a novel calcium response element in the glucagon gene. J.BioLChem. 274,5851-5860.
47. Bert A.G., Burrows J., Hawwari A., Vadas M.A. and Cockerill P.N. (2000) Reconstitution of Tcell-specific transcription directed by composite NFAT/Oct elements [In Process Citation].J.Immunol. 165, 5646-5655.
48. Rengarajan J., Mowen K.A., McBride K.D., Smith E.D., Singh H. and Glimcher L.H. (2002)Interferon regulatory factor 4 (IRF4) interacts with NFATc2 to modulate interleukin 4 geneexpression. J.Exp.Med. 195,1003-1012.
49. Hu CM., Jang S.Y., Fanzo J.C. and Pernis A.B. (2002) Modulation of T cell cytokineproduction by interferon regulatory factor-4. J.BioLChem. 277, 9238-49246.
50. McKinsey T.A., Zhang C.L. and Olson E.N. (2002) MEF2: a calcium-dependent regulator ofcell division, differentiation and death. Trends BiochemSci. 27, 40-47.
51. Olson E.N. and Williams R.S. (2000) Remodeling muscles with calcineurin. Bioessays 22, 510-519.
63. Yang X.Y., Wang L.H., Chen T., Hodge D.R., Resau J.H., DaSilva L. and Farrar W.L. (2000) Activation of human T lymphocytes is inhibited by peroxisome proliferator-activated receptor gamma (PPARgamma) agonists. PPARgamma co-association with transcription factor NFAT. J.Biol.Chem. 275, 4541-4544.
64. Molkentin J.D., Lu J.R., Antos C.L., Markham B., Richardson J., Robbins J., Grant S.R. andOlson E.N. (1998) A calcineurin-dependent transcriptional pathway for cardiac hypertrophy.Cell 93, 215-228.
65. Avni O., Lee D., Macian F., Szabo S.J., Glimcher L.H. and Rao A. (2002) T(H) celldifferentiation is accompanied by dynamic changes in histone acetylation of cytokine genes.Nat.Immunol. 3, 643-651.

66. Klein-Hessling S., Jha M.K., Santner-Nanan B., Berberich-Siebelt F., Baumruker T., Schimpl A.and Serfling E. (2003) Protein kinase A regulates GATA-3-dependent activation of IL-5 geneexpression in Th2 cells. J.Immunol. 170,2956-2961.
67. Musaro A., McCullagh K.J., Naya F.J., Olson E.N. and Rosenthal N. (1999) IGF-1 inducesskeletal myocyte hypertrophy through calcineurin in association with GATA-2 and NF-ATc1.Nature 400,581-585.
68. Im S.H., Hueber A., Monticelli S., Kang K.R and Rao A. (2004) Chromatin-level regulation ofthe IL10 gene in T cells. J.Biol.Chem. 279, 46818-46825.
69. Baksh S., Widlund H.R., Frazer-Abel A.A., Du J., Fosmire S., Fisher D.E., DeCaprio J.A.,Modiano J.F. and Burakoff S.J. (2002) NFATc2-mediated repression of cyclin-dependent kinase4 expression. Mol.Cell 10,1071-1081.
70. Marian F., Garcia-Rodriguez C. and Rao A. (2000) Gene expression elicited by NFAT in thepresence or absence of cooperative recruitment of fos and Jun [In Process Citation]. EMBO J.19,4783-4795.
71. Marian F., Garcia-Cozar F., 1m S.H., Horton H.F., Byrne M.C. and Rao A. (2002)Transcriptional mechanisms underlying lymphocyte tolerance. Cell 109, 719-731.
72. Ranger AM., Oukka M., Rengarajan J. and Glimcher L.R (1998) Inhibitory function of twoNFAT family members in lymphoid homeostasis and Th2 development. Immunity. 9, 627-635.
73. Peng S.L., Gerth A.J., Ranger A.M. and Glimcher L.R (2001) NFATc1 and NFATc2 togethercontrol both T and B cell activation and differentiation. Immunity. 14,13-20.
74. Ranger A.M., Grusby M.J., Hodge M.R., Gravallese E.M., de la Brousse F.C., Hoey T.,Mickanin C, Baldwin H.S. and Glimcher L.H. (1998) The transcription factor NF-ATc isessential for cardiac valve formation [see comments]. Nature 392, 186-190.
75. de la Pompa J.L., Timmerman L.A., Takimoto H., Yoshida R, Elia A.J., Samper E., Potter J.,Wakeham A., Marengere L., Langille B.L., Crabtree G.R. and Mak T.W. (1998) Role of the NF-ATc transcription factor in morphogenesis of cardiac valves and septum [see comments]. Nature392, 182-186.
76. Graef I.A., Mermelstein P.G., Stankunas K., Neilson J.R., Deisseroth K., Tsien R.W. andCrabtree G.R. (1999) L-type calcium channels and GSK-3 regulate the activity of NF-ATc4 inhippocampal neurons. Nature 401, 703-708.
77. Ranger A.M., Gerstenfeld L.C., Wang J., Kon T., Bae R, Gravallese E.M., Glimcher M.J. andGlimcher L.H. (2000) The nuclear factor of activated T cells (NFAT) transcription factorNFATp (NFATc2) is arepressor of chondrogenesis [see comments]. J.Exp.Med 191,9-22.
78. Ho I.C., Kim J.H., Rooney J.W., Spiegelman B.M. and Glimcher L.R (1998) A potential rolefor the nuclear factor of activated T cells family of transcriptional regulatory proteins inadipogenesis. Proc.Natl.Acad.SciU.S.A 95, 15537-15541.
79. Schmitt E., Hoehn P., Germann T. and Rude E. (1994) Differential effects of interleukin-12 onthe development of naive mouse CD4+ T cells. Eur.J.Immunol. 24, 343-347.
80. Randak C, Brabletz T., Hergenrother M., Sobotta I. and Serfling E. (1990) Cyclosporin Asuppresses the expression of the interleukin 2 gene by inhibiting the binding of lymphocyte-specific factors to the IL-2 enhancer. EMBOJ. 9, 2529-2536.
81. Chuvpilo S., Schomberg C, Gerwig R., Heinfling A., Reeves R., Grummt F. and Serfling E.(1993) Multiple closely-linked NFAT/octamer and HMG I(Y) binding sites are part of theinterleukin-4 promoter. Nucleic Acids Res. 21,5694-5704.

82. Liu J., Koyano-Nakagawa N., Amasaki Y., Saito-Ohara F., Ikeuchi T., Imai S., Takano T., AraiN., Yokota T. and Arai K. (1997) Calcineurin-dependent nuclear translocation of a murinetranscription factor NFATx: molecular cloning and functional characterization. Mol.Biol.Cell 8,157-170.
83. Ho S.N., Thomas D.J., Timmerman L.A., Li X., Francke U. and Crabtree G.R. (1995) NFATc3,a lymphoid-specific NFATc family member that is calcium- regulated and exhibits distinct DNAbinding specificity. J.Biol.Chem. 270, 19898-19907.
84. Reynolds P.J., Lesley J., Trotter J., Schulte R., Hyman R. and Sefton B.M. (1990) Changes inthe relative abundance of type I and type II lck mRNA transcripts suggest differential promoterusage during T-cell development. MolCell Biol. 10, 4266-4270.
85. Hemenway C.S. and Heitman J. (1999) Calcineurin. Structure, function, and inhibition. CellBiochem.Biophys. 30,115-151.
86. Fruman D.A., Pai S.Y., Burakoff S.J. and Bierer B.E. (1995) Characterization of a mutantcalcineurin A alpha gene expressed by EL4 lymphoma cells. MolCell Biol. 15, 3857-3863.
87. Dai P., Akimaru H., Tanaka Y., Hou D.X., Yasukawa T., Kanei-Ishii C, Takahashi T. and IshiiS. (1996) CBP as a transcriptional coactivator of c-Myb. Genes Dev. 10,528-540.
88. Bergholtz S., Andersen T.O., Andersson K.B., Borrebaek J., Luscher B. and Gabrielsen O.S.(2001) The highly conserved DNA-binding domains of A-, B- and c-Myb differ with respect toDNA-binding, phosphorylation and redox properties. Nucleic Acids Res. 29, 3546-3556.
89. Xu H., Inouye M., Hines E.R., Collins J.F. and Ghishan F.K. (2003) Transcriptional regulationof the human NaPi-IIb cotransporter by EGF in Caco-2 cells involves c-myb. AmJ.Physiol CellPhysiol 284, C1262-C1271.
90. Fang X., Yu S.X., Lu Y., Bast R.C., Jr., Woodgett J.R. and Mills G.B. (2000) Phosphorylationand inactivation of glycogen synthase kinase 3 by protein kinase A. Proc.Natl.Acad.Sci.U.S.A97,11960-11965.
91. Jain J., McCaffrey P.G., Valge-Archer V.E. and Rao A. (1992) Nuclear factor of activated Tcells contains Fos and Jun. Nature 356, 801-804.
92. Barton K., Muthusamy N., Chanyangam M., Fischer C, Clendenin C. and Leiden J.M. (1996)Defective thymocyte proliferation and IL-2 production in transgenic mice expressing adominant-negative form of CREB. Nature 379, 81-85.
93. de Groot R.P. and Sassone-Corsi P. (1992) Activation of Jun/AP-1 by protein kinase A,Oncogene 7, 2281-2286.


















