
Inhibition of Paclitaxel-induced A-fiber...
31
JPET #103614 1 Inhibition of Paclitaxel-induced A-fiber-Hypersensitization by Gabapentin Misaki Matsumoto, Makoto Inoue, Andreas Hald, Weijiao Xie and Hiroshi Ueda Division of Molecular Pharmacology and Neuroscience, Nagasaki University Graduate School of Biomedical Sciences, Nagasaki 852-8521, Japan (M.M., M.I., A.H., W.X., H.U.) JPET Fast Forward. Published on May 10, 2006 as DOI:10.1124/jpet.106.103614 Copyright 2006 by the American Society for Pharmacology and Experimental Therapeutics. This article has not been copyedited and formatted. The final version may differ from this version. JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614 at ASPET Journals on June 18, 2018 jpet.aspetjournals.org Downloaded from
Transcript of Inhibition of Paclitaxel-induced A-fiber...

JPET #103614
1
Inhibition of Paclitaxel-induced A-fiber-Hypersensitization
by Gabapentin
Misaki Matsumoto, Makoto Inoue, Andreas Hald, Weijiao Xie
and Hiroshi Ueda
Division of Molecular Pharmacology and Neuroscience, Nagasaki University Graduate
School of Biomedical Sciences, Nagasaki 852-8521, Japan (M.M., M.I., A.H., W.X.,
H.U.)
JPET Fast Forward. Published on May 10, 2006 as DOI:10.1124/jpet.106.103614
Copyright 2006 by the American Society for Pharmacology and Experimental Therapeutics.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
2
a) Running title: Gabapentin inhibits paclitaxel-induced neuropathic pain.
b) Address for correspondence: Dr. Hiroshi Ueda,
Division of Molecular Pharmacology and Neuroscience, Nagasaki University Graduate
School of Biomedical Sciences, 1-14 Bunkyo-machi, Nagasaki 852-8521, Japan
Tel: +81-95-819-2421; Fax: +81-95-819-2420; Email: [email protected]
c) Number of text pages, figures and references:
Text pages: 27
Figures: 4
Tables: 0
References: 40
Number of words in
-Abstract: 224
-Introduction: 344
-Discussion: 752
d) Abbreviations: EPW, electrical-stimulus induced paw withdrawal; DRG, dorsal root
ganglion; GAPDH, Glyceraldehyde-3-phosphate dehydrogenase; Caα2δ-1, voltage-gated
calcium channel α2δ-1subunit.
e) Recommended section: Neuropharmacology
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
3
Abstract
Paclitaxel (Taxol®) is a widely used chemotherapeutic agent in the treatment of several
tumors. However, its use is often associated with the generation of peripheral neuropathic
pain expressed as mechanical allodynia and thermal hyperalgesia. The molecular
mechanism behind this debilitating side effect is obscure and efficient drugs for its
prevention are required. We sought to clarify the cellular changes in the involved
nociceptor types underlying paclitaxel-induced neuropathic pain, and to test for an
alleviating effect of gabapentin treatment in a murine model of paclitaxel-induced
neuropathic pain. We found that a single treatment with paclitaxel (4 mg/kg, i.p.) led to a
decrease in both the thermal and mechanical nociceptive thresholds, as well as reducing
the thresholds for 250 Hz (Aδ-fiber) and 2000 Hz (Aβ-fiber) but not 5 Hz (C-fiber)
sine-wave electrical stimuli-induced paw withdrawal (EPW). The paclitaxel-induced
neuropathic pain was completely abrogated by gabapentin (30 mg/kg, i.p.) treatment.
Furthermore, we found that mRNA and protein levels of the calcium channel subunit
α2δ-1 (Caα2δ-1), one of the putative targets for gabapentin, was upregulated in dorsal
root ganglions (DRG), as well as increased expression of Caα2δ-1 protein in
medium/large DRG neurons by immohistochemistry, following paclitaxel treatment.
This suggests, that paclitaxel induces A-fiber specific hypersensitization, which may
contribute to the functional mechanical allodynia and hyperalgesia, and that gabapentin
could be a potential therapeutic agent for paclitaxel-induced neuropathic pain.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
4
Introduction
Paclitaxel (Taxol®) is widely used in cancer chemotherapy for the treatment of solid
carcinomas (Kohler and Goldspiel, 1994; Socinski, 1999). Its mechanism is known to
involve an increase in the stability of tubulin polymers and inhibit cellular replication
(Schiff et al., 1979; Horwitz, 1992). However, it has serious side effects such as
peripheral neuropathic pain (Rowinsky et al., 1993; Chaudhry et al., 1994; Cavaletti et al.,
1995; Forsyth et al., 1997), which become more marked with continued therapy. In some
cases, the side effects may outweigh the beneficial effects on life quality, necessitating a
dose reduction (Bristol-Myers Squibb Company, 2003). However, the molecular
mechanism underlying paclitaxel-induced neuropathic pain is enigmatic and no
therapeutics for its prevention are known.
Recently, the anticonvulsant gabapentin has proven effective in the treatment of
neuropathic pain, and in neuropathic pain models such as the peripheral nerve ligation
and diabetic neuropathic pain model, gabapentin inhibits thermal hyperalgesia and tactile
allodynia (Hunter et al., 1997; Hwang and Yaksh, 1997; Cesena and Calcutt, 1999; Miki
et al., 2001; Luo et al., 2002). Although gabapentin was originally designed as a structural
analogue of the neurotransmitter GABA, it was revealed that gabapentin binds with
high-affinity to Caα2δ-1 (Gee et al., 1996), which most likely accounts for its analgesic
effects (Cheng and Chiou, 2006). Furthermore, gabapentin has been reported to inhibit
high-threshold Ca2+ channels and modulate synaptic neurotransmission (Patel et al.,
2000; Shimoyama et al., 2000; Sutton et al., 2002; Maneuf et al., 2004). In addition, Luo
et al. (2002) reported that gabapentin efficacy is correlated with the increase of Caα2δ-1
in DRG in peripheral nerve injury models.
In the present study, we demonstrated that gabapentin inhibits
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
5
paclitaxel-induced peripheral neuropathic pain in mice. Furthermore, Neurometer tests,
which are known to determine nociceptor sensitivity, as well as to distinguish between the
different types of sensory neurons in humans and rodents (Masson and Boulton, 1991;
Pitei et al., 1994; Kiso et al., 2001), showed an A-fiber specific hypersensitization
following paclitaxel treatment. Finally, we report an increased expression of Caα2δ-1 in
medium/large DRG neurons after paclitaxel treatment.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
6
Methods
Animals
Male ddY mice weighing 20 - 22 g were used. They were kept in a room maintained at 21
± 2ºC with free access to standard laboratory diet and tap water. Procedures were
approved by Nagasaki University Animal Care Committee and complied with the
recommendations of the International Association for the Study of Pain (Zimmermann,
1983).
Drugs
The following drugs were used: Paclitaxel (Taxol®) was generously provided by
Bristol–Myers Squibb Company, Paris, France. Paclitaxel was dissolved in saline just
before administration. Gabapentin [1-(Aminomethyl)-cyclohexaneacetic acid] was
purchased from Sigma Chemical (St Louis, MO, U.S.A.), dissolved in saline.
Drug injection and analysis
We used three types of paclitaxel (4 mg/kg) -injection schedule as follows; single i.p.
injection, multiple (four times on alternate days; days 0, 2, 4 and 6) i.p. injection and
single i.v. injection. In the “area under the curve” (AUC) analysis for paclitaxel effect on
pain thresholds, we calculated the area under the curve generated by the paw withdrawal
threshold (PWT)-time course from week 1 to 4 after paclitaxel treatment using a
trapezoidal method. Gabapentin (3 – 30 mg/kg) was injected (i.p.) 30 minutes before
behavioral tests.
Conventional nociceptive tests
In thermal paw withdrawal tests, nociception was measured as the latency to paw
withdrawal evoked by exposure to a thermal stimulus (Hargreaves et al., 1988; Inoue et
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
7
al., 2004). Unanesthetized animals were placed in plexiglas cages on top of a glass sheet
and an adaptation period of 1 hour was allowed. The thermal stimulator (IITC Inc.,
Woodland Hills, CA, USA) was positioned under the glass sheet and the focus of the
projection bulb was aimed exactly on the middle of the plantar surface of the animal. A
mirror attached to the stimulator, permitted visualization of the plantar surface. A cut-off
time of 20 seconds was set in order to prevent tissue damage. The paw pressure test was
performed as described previously (Rashid et al., 2003; Inoue et al., 2004). Briefly, mice
were placed into a plexiglas chamber on a 6×6 mm wire mesh grid floor and were
allowed to acclimatize for a period of 1 hour. The mechanical stimulus was then delivered
onto the middle of the plantar surface of the right hind-paw using a Transducer Indicator
(Model 1601, IITC Inc., Woodland Hills, USA). The pressure needed to induce a flexor
response was defined as the pain threshold. All behavioral experiments were carried out
by investigators blinded to the drug-treatment.
Electrical stimulation-induced paw withdrawal (EPW) test
Electrodes (Neurotron Inc., Blatimore, MD) were fastened to the right plantar surface and
instep of the mice. Transcutaneous nerve stimuli with each of the three sine-wave pulses
(5, 250 and 2000Hz) were applied using the Neurometer CPT/C (Neurotron Inc.). The
minimum intensity (µA) at which each mouse withdrew its paw was defined as the
current stimulus threshold. Stimuli were applied with 10 minutes intervals. All behavioral
experiments were carried out by investigators blinded to the drug-treatment.
Quantitative real-time PCR
On the 3rd, 7th and 14th day after paclitaxel treatment, the L4-6 DRGs were removed and
RNA was purified using TRIzol (Invitrogen, CA, USA). Total RNA (1 µg) was used for
cDNA synthesis with Superscript II reverse transcriptase and random hexamer primers
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
8
(Invitrogen, CA, USA). Real-time quantitative PCR (RT-PCR) was performed using an
ABI Prism 7000 Sequence Detection system (Applied Biosystems, Tokyo, Japan), using
qPCR MasterMix Plus for SYBR® Green I (Eurogentec, Seraing, Belgium) containing
dNTPs (+dUTP), Hot Goldstar DNA polymerase, and Urasil-N Glycosilase, according to
the manufactures instructions. The cycling conditions for all primers were the following:
10 minutes at 95°C to activate the Hot Goldstar DNA polymerase, followed by 50 cycles
consisting of two steps, 15 seconds at 95°C (denaturation) and 1 minute at 60°C
(annealing-extension). The Caα2δ-1mRNA levels were evaluated by comparison to
glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Primer sequences were as
follows; Caα2δ-1, (F)5′-CTC ATG CAA GCG GAA CAG ACT TCT-3′ and (R)5′-TCA
AAG CAG ACA TCA GGT CCT TT-3′ corresponding to the mouse Caα2δ-1 gene (NCBI
accession number; NM_009784) (Martin et al., 2002), GAPDH, (F)5′- TAT GAC TCC
ACT CAC GGC AAA T-3′ and (R)5′-GGG TCT CGC TCC TGG AAG AT-3’
corresponding to the mouse GAPDH (NCBI accession number; NM_001001303). In all
cases, the validity of amplification was confirmed by the presence of a single peak in the
melting temperature analysis and linear amplification against the PCR cycles.
Western blot analysis
On the 7th, 14th and 28th day after paclitaxel treatment, the L4-6 DRGs were removed
for Western blot analysis. Mixed L4-6 DRG protein (30 µg) were applied to an
SDS-polyacrylamide gel (8 %). The anti- Caα2δ-1 antibody (Sigma, USA) diluted 1:200
(Inoue et al., 2004) was used and anti-β−Tubulin antibody (Santa Cruz Biotechnology,
CA, USA) diluted 1:1000 was used as internal control. HRP-labeled rabbit antibody was
used as 2nd antibody at 1:2000. Visualization of immunoreactive bands was performed
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
9
by the Light Capture (AE-6960/C/FC, Atto, Tokyo, Japan) with an enhanced
chemiluminescent substrate for the detection of horseradish peroxidase, SuperSignal®
West Pico Chemiluminescent Substrate (PIERCE, Rockford, IL). The intensities of
immunoreactive bands were analyzed by NIH imaging for Macintosh.
Immunohistochemistry
The immunohistochemical analysis were performed as described previously (Inoue et al.,
2004). Briefly, mice were deeply anesthetized with sodium pentobarbital (50 mg/kg i.p.)
and perfused transcardially with 20 ml of potassium-free phosphate-buffered saline
(K+-free PBS, pH 7.4) followed by 50 ml of 4% paraformaldehyde solution. The L4-6
DRGs were removed, postfixed for 3 hours, and cryoprotected overnight in 25% sucrose
solution. The DRGs were fast-frozen in cryo-embedding compound on a mixture of
ethanol and dry ice and stored at -80°C until use. DRGs were cut at 10 µm thickness,
thaw-mounted on a silane-coated glass slides, and air-dried overnight at room
temperature (RT). Before immunolabeling, antigen unmasking was performed by
microwave treatment for 15 minutes in10 mM citrate buffer (pH 6.0). DRG sections were
incubated with excess blocking buffer containing 2% skim milk in PBST (0.1% Triton
X-100 in K+-free PBS) and subsequently reacted overnight at 4°C with anti-
Caα2δ-1antibodies (1:200; Sigma, USA) in 2% BSA/PBST. The sections were then
placed in fluorescein isothiocyanate (FITC)-conjugated anti-rabbit IgG (1:200; Santa
Cruz Biotechnology, Inc., Santa Cruz, CA) for 2 hours at RT. All sections were treated
with Permafluor (Thermo Shandon, Pittsburgh, PA) and coverslipped and evaluated by
microscopy (Keyence, Tokyo, Japan).
Measurements of DRG neuron diameters were done using the BZ Image
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
10
Measurement software (Keyence, Tokyo, Japan). Total cell counts was carried out in
bright field images as reported elsewhere (Josephson et al., 2001; Dai et al., 2004).
Therefore, the quantification of the data may represent a biased estimate of the actual
number of neurons. However, in order to determine the Caα2δ-1 positive neuron profile,
we have carried out the counts using only neurons with clearly visible nuclei. In this
experiment, we divided neurons into 2 groups, small and medium/large neurons. We used
24 µm in diameter as dividing line between small and medium/large neurons (Gold MS et
al., 1996; Hiruma et al., 2000). The total 1684 DRG neurons from vehicle-treated mice
(n=3) and 2724 DRG neurons from paclitaxel-treated mice (n=4) were measured.
Statistical analysis
The behavioral time course data were analyzed using a Student’s t-test to detect
differences between vehicle- and paclitaxel- treated groups. The behavioral data on
gabapentin and the RT-PCR data as well as the western blot data of Caα2δ-1 were
analyzed using a one-way ANOVA and Tukey multiple comparison post-hoc analysis.
The criterion of significance was set at p<0.05. All results were expressed as mean ±
S.E.M.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
11
Results
Paclitaxel-induced thermal hyperalgesia and mechanical allodynia.
We treated mice with paclitaxel (4 mg/kg) single (i.p. or i.v.) or multiple times (four i.p.
injections on alternate days; days 0, 2, 4 and 6), and evaluated the threshold in several
nociceptive tests from pre-injection (control) to 4 weeks after the first treatment. No
significant side effects such as ascites, loss of body weight or alopecia was observed in
this experimental schedule. A single i.p. injection of paclitaxel (4 mg/kg) led to a
decreased threshold in the paw pressure test, which was significant at 1 and 2 weeks after
treatment but increased to baseline level at week 4 post treatment (Fig. 1A). When
comparing the mechanical allodynia after a single i.p. injection, multiple i.p. injections or
single i.v. injection of paclitaxel, no significantly different outcomes were observed,
between the numbers of administrations or the route, as shown as the AUC values for the
paw pressure test from week 1 to 4 after treatment (Fig. 1B). The thermal nociceptive
threshold level decreased in a manner similar to the decrease in threshold for mechanical
allodynia, though the effect was less pronounced. However, the effect was significant 2
weeks after a single i.p. injection of paclitaxel (Fig. 1C). Based on these results, we
determined to adopt the single i.p. injection for the paclitaxel-induced neuropathic pain
model in mice and to evaluate the effect of gabapentin 2 weeks after treatment which was
the peak time point of neuropathic pain.
Blockade of paclitaxel-induced neuropathic pain by gabapentin.
Gabapentin, injected i.p. 30 minutes before behavioral tests, inhibited mechanical
allodynia 2 weeks after paclitaxel-treatment (4 mg/kg, single i.p. injection) in a
dose-dependent manner from 3 to 30 mg/kg, while gabapentin (30 mg/kg) did not cause
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
12
any significant effects in vehicle-treated mice (Fig. 1D). The duration of the analgesic
effect from treatment with 30 mg/kg gabapentin lasted for more than 24 hours but the
significant effect had vanished by 48 hours after treatment (Fig. 1E). Moreover,
gabapentin injection (30 mg/kg, i.p.) 2 weeks after paclitaxel-treatment reversed thermal
hyperalgesia within 30 minutes. However, gabapentin treatment of vehicle-treated mice
did not affect the thresholds of thermal paw withdrawal latency (Fig. 1F).
Paclitaxel-induced myelinated A-fiber specific hypersensitization
Transcutaneous nerve stimuli with each of the three sine-wave pulses of different
frequencies of 5, 250 and 2000 Hz to activate C-, Aδ- and Aβ-fibers, respectively
(Katims, 1997; Lengyel et al., 1998), were applied to the right hind paw of the mice, and
the intensity (µA) was gradually increased. Thresholds were determined, at which paw
withdrawal was observed, from pre-injection (control) to 4 weeks after paclitaxel
treatment (Fig. 2A). The reduction in threshold by the 250 Hz (Aδ) and 2000 Hz (Aβ)
stimuli at 1 and 2 weeks after paclitaxel-treatment (4 mg/kg, single i.p. injection), had
completely vanished 4 weeks after treatment. No significant change in the threshold by 5
Hz (C) stimuli was observed at any time. These results suggest that paclitaxel induces
myelinated Aδ and Aβ-fiber specific hypersensitization.
Blockade of paclitaxel-induced myelinated A-fiber specific hypersensitization by
gabapentin.
Gabapentin injection (30 mg/kg, i.p.) 2 weeks after paclitaxel-treatment reversed the
hyperalgesia within 30 minutes as determined by 250 Hz (Aδ) and 2000 Hz (Aβ)
stimulations. However, gabapentin treatment of vehicle-treated mice did not affect the
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
13
thresholds of neither 250 Hz (Aδ) and 2000 Hz (Aβ) stimuli induced responses (Fig. 2B).
Increased expression of Caα2δ-1 mRNA and protein in DRG after paclitaxel
treatment.
We removed L4-6 DRGs at the 3rd, 7th and 14th day after paclitaxel treatment and
Caα2δ-1 mRNA expression was quantified by RT-PCR analysis and compared to
GAPDH mRNA expression. The specificity of the PCR primers was confirmed by the
occurrence of a single peak in the melting curves for the PCR products, and the efficiency
of the reaction was found to be approximately 100 % shown by a threshold cycle (Ct) vs.
sample-dilution plot (Fig. 3A). Caα2δ-1 mRNA expression was slightly but not
significantly increased at day 3 but a larger and significant increase was observed at day 7
in paclitaxel-treated mice compared to controls (Fig. 3A). At day 14, the Caα2δ-1 mRNA
expression was reduced to control level.
Moreover, we removed L4-6 DRGs at 7th, 14th and 28th day after paclitaxel
treatment and Caα2δ-1 protein expression was quantified by western blot analysis. The
result indicated that Caα2δ-1 up-regulation was significant at day 7, maximum at day 14
and returned to baseline level 28 days post paclitaxel injection (Fig. 3B).
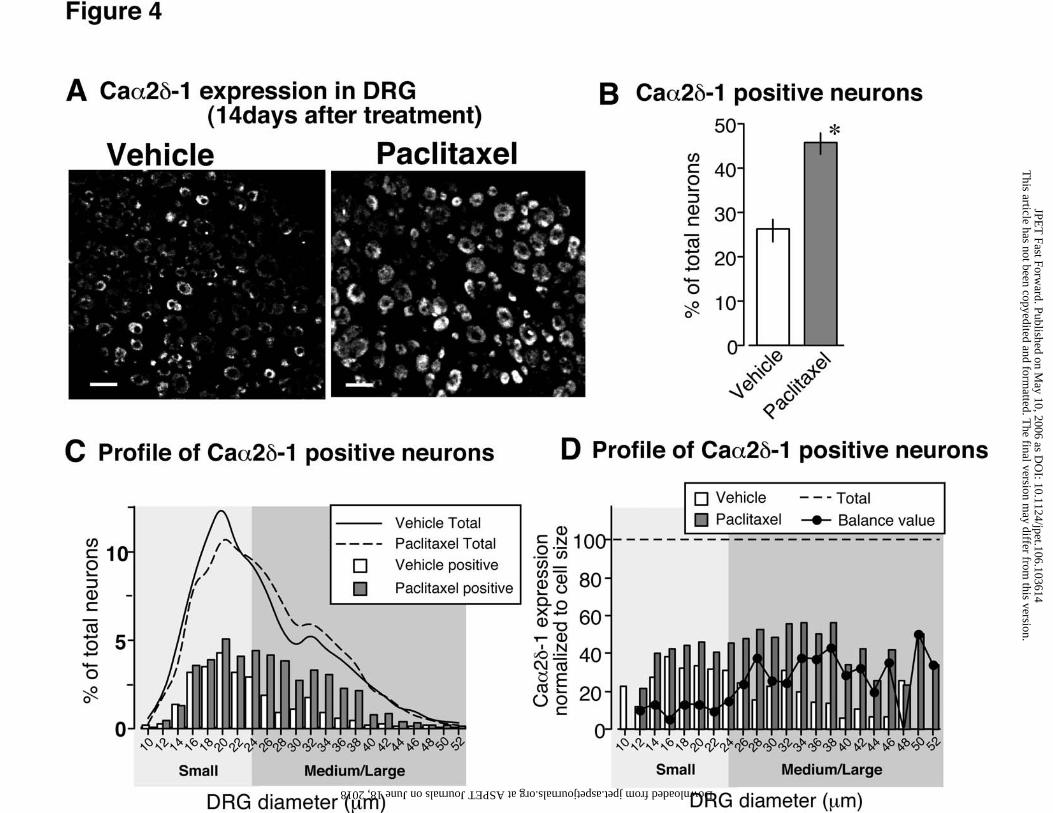
Increased expression of Caα2δ-1 in medium/large sized DRG neurons following
paclitaxel treatment.
We detected Caα2δ-1 protein expression in DRGs at day 14 after treatment using
immunohistochemistry. In vehicle-treated mice, Caα2δ-1 expression was mainly
localized to small diameter DRG neurons (Fig. 4A), while in paclitaxel-treated mice,
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
14
Caα2δ-1 protein expression was both observed in small diameter DRG neurons and in
medium/large diameter DRG neurons (Fig. 4A). The total amount of neurons in the
DRGs, which stained positive for Caα2δ-1 protein, was counted and their individual sizes
measured both in paclitaxel- and vehicle- treated mice. A significant increase in the
frequency of stained neurons was found in the paclitaxel-treated mice compared to
vehicle-treated controls (Fig. 4B). Furthermore the cell-size measurements showed that
the increased number of Caα2δ-1 stained neurons reflected an increased expression of
Caα2δ-1 in medium/large sized neurons, while the number of positively stained small
neurons remained almost same between vehicle- and paclitaxel- treated groups (Fig. 4C
and D). However, no significant difference in the general size distribution of DRG
neurons was found between the two groups, supporting the notion, that the increased
Caα2δ-1 protein expression indeed resulted from an up-regulation in medium/large sized
neurons (Fig. 4C).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
15
Discussion
Here we found that single administration of paclitaxel at a dose of 4 mg/kg i.p. caused
peripheral neuropathic pain. According to the paclitaxel injection prescribing information
sheet, the recommended dose is 135-175 mg/m2 (Bristol-Myers Squibb Company, 2003),
which is converted into 3.6-4.7 mg/kg approximately (Height 1.7 m, Weight 65 kg; Du
Bois formula). In addition, it states that paclitaxel should be given every 3 weeks in
clinical states. However, in previous experimental studies, the paclitaxel administration
has been performed multiple times on consecutive/alternate days or in a single but high
dose (Authier et al.,2000; Dina et al., 2001; Polomano et al., 2001; Flatters and Bennett,
2004; Smith et al., 2004). Therefore, the present finding seems to have some advantages.
We conclude that the protocol using single administration of paclitaxel (4 mg/kg) is
proper and easy approach to study paclitaxel-induced neuropathic pain.
In our experimental model, we observed mechanical allodynia, thermal
hyperalgesia and A-fiber specific hypersensitization using the Neuromater test. The
Neurometer test is widely used clinically in the evaluation of sensory function in humans
with complex regional pain syndrome or other types of peripheral neuropathic pain,
which can result from common diseases such as diabetes mellitus (Masson and Boulton,
1991; Katims, 1997; Lengyel et al., 1998). Our results are supported by previous data
from animal models of paclitaxel-induced hypersensitivity (Smith et al., 2004), as well as
neurometer analysis of paclitaxel-treated patients, which also show Aβ-fiber
hypersensitivity, as determined by stimulating the median nerve with 2000 Hz pulses
(Doi et al., 2003). Therefore, we conclude that our model is useful for investigating the
mechanism by which paclitaxel induces neuropathic pain and to test new drug candidates.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
16
Frequent reports of the alleviating effect of gabapentin treatment in various
models of neuropathic pain such as the peripheral nerve ligation model and diabetic
neuropathic pain models, are published (Hunter et al., 1997; Hwang and Yaksh, 1997;
Cesena and Calcutt, 1999; Miki et al., 2001; Luo et al., 2002). The present study is the
first to report an anti-nociceptive effect of gabapentin on paclitaxel-induced neuropathic
pain in mice. We evaluated the effect by EPW tests as well as conventional nociceptive
tests measuring mechanical nociception. Gabapentin revealed specific analgesic effects
against paclitaxel-induced mechanical allodynia, thermal hyperalgesia and A-fiber
hypersensitization without affecting the pain threshold of naïve mice. These results
suggest that gabapentin could be used to treat paclitaxel-induced neuropathic pain in the
clinic.
Referring to the mechanism of anti-nociceptive effects of gabapentin, we
measured the expression of Caα2δ-1, which is known to possess gabapentin-binding
properties (Gee et al., 1996). We first demonstrated that the expression of Caα2δ-1
mRNA and protein was increased in the DRGs of paclitaxel-treated mice. The difference
in the peak time point between mRNA and protein expression seems to be attributable to
the longer life-time of protein. It should be noted that the time course of Caα2δ-1 protein
expression was very close to that of paclitaxel-induced neuropathic pain (Fig. 1A, 1C and
Fig. 2A). Interestingly, immunohistochemistry revealed that Caα2δ-1 protein was
increased in medium/large diameter neurons following paclitaxel-treatment. As the
function of the calcium channel containing Caα2δ-1 is related to modulation of synaptic
neurotransmission (Patel et al., 2000; Shimoyama et al., 2000; Maneuf et al., 2004),
up-regulation of Caα2δ-1 expression in medium/large-sized neurons may contribute to
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
17
the paclitaxel-induced mechanical allodynia and A-fiber dependent hypersensitization.
Recently, we have reported that Caα2δ-1 is upregulated in medium/large-sized DRG
neurons following peripheral nerve injury or intrathecal injection of lysophosphatidic
acid, an initiator of neuropathic pain (Inoue et al., 2004). Luo et al. (2002) reported that
inhibitory effects of gabapentin on neuropathic pain correlates to Caα2δ-1 expression.
Gabapentin sensitivity and Caα2δ-1 up-regulation were observed in spinal nerve ligation,
spinal nerve transection, sciatic nerve chronic constriction injury and diabetic models, but
not in the vincristine-induced neuropathic pain model. In the present study, we confirmed
the gabapentin sensitivity and Caα2δ-1 up-regulation in the paclitaxel-induced
neuropathic pain model. It is interesting that the gabapentin sensitivity and Caα2δ-1
up-regulation differed between the vincristine and paclitaxel models, when taken into
account that both drugs are known to act through tubulin assembly. Although the
underlying mechanism remains to be determined, it might be attributed different manners
of action; disruption of tubulin assembly by vincristine and increasing stabilization by
paclitaxel (Kohler and Goldspiel, 1994).
In conclusion, we first report, that paclitaxel induces A-fiber hypersensitization
and novel expression of Caα2δ-1 in medium/large DRG neurons, which may functionally
contribute to mechanical allodynia and hyperalgesia. Furthermore, we identify
gabapentin as a potential therapeutic agent for paclitaxel-induced peripheral neuropathic
pain.
Acknowledgements
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
18
We thank N. Uminotaira and A. Yamaguchi for technical help and behavioral
studies. Paclitaxel (Taxol®) was generously provided by Bristol–Myers Squibb Company,
Paris, France.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
19
References
Authier N, Gillet JP, Fialip J, Eschalier A and Coudore F (2000) Description of a
short-term Taxol-induced nociceptive neuropahy in rats. Brain Res 887:239-249.
Bristol-Myers Squibb Company (2003) TAXOL (paclitaxel) Injection Prescribing
Information.
Cavaletti G, Bogliun G, Marzorati L, Zincone A, Marzola M, Colombo N and Tredici G
(1995) Peripheral neurotoxicity of taxol in patients previously treated with
cisplatin. Cancer 75:1141-1150.
Cesena RM and Calcutt NA (1999) Gabapentin prevents hyperalgesia during the formalin
test in diabetic rats. Neurosci Lett 262:101-104.
Chaudhry V, Rowinsky EK, Sartorius SE, Donehower RC and Cornblath DR (1994)
Peripheral neuropathy from taxol and cisplatin combination chemotherapy:
clinical and electrophysiological studies. Ann Neurol 35:304-311.
Cheng JK and Chiou LC (2006) Mechanisms of the Antinociceptive Action of
Gabapentin. J Pharmacol Sci. in press.
Dai Y, Fukuoka T, Wang H, Yamanaka H, Obata K, Tokunaga A and Noguchi K (2004)
Contribution of sensitized P2X receptors in inflamed tissue to the mechanical
hypersensitivity revealed by phosphorylated ERK in DRG neurons. Pain
108:258-266.
Dina OA, Chen X, Reichling D and Levine JD (2001) Role of protein kinase Cepsilon
and protein kinase A in a model of paclitaxel-induced painful peripheral
neuropathy in the rat. Neuroscience 108:507-515.
Doi D, Ota Y, Konishi H, Yoneyama K and Araki T (2003) Evaluation of the
neurotoxicity of paclitaxel and carboplatin by current perception threshold in
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
20
ovarian cancer patients. J Nippon Med Sch 70:129-134.
Flatters SJ and Bennett GJ (2004) Ethosuximide reverses paclitaxel- and
vincristine-induced painful peripheral neuropathy. Pain 109:150-161.
Forsyth PA, Balmaceda C, Peterson K, Seidman AD, Brasher P and DeAngelis LM
(1997) Prospective study of paclitaxel-induced peripheral neuropathy with
quantitative sensory testing. J Neurooncol 35:47-53.
Gee NS, Brown JP, Dissanayake VU, Offord J, Thurlow R and Woodruff GN (1996) The
novel anticonvulsant drug, gabapentin (Neurontin), binds to the alpha2delta
subunit of a calcium channel. J Biol Chem 271:5768-5776.
Gold MS, Dastmalchi S and Levine JD (1996) Co-expression of nociceptor properties in
dorsal root ganglion neurons from the adult rat in vitro. Neuroscience 71:265-275.
Hargreaves K, Dubner R, Brown F, Flores C and Joris J (1988) A new and sensitive
method for measuring thermal nociception in cutaneous hyperalgesia. Pain
32:77-88.
Hiruma H, Saito A, Ichikawa T, Kiriyama Y, Hoka S, Kusakabe T, Kobayashi H and
Kawakami T (2000) Effects of substance P and calcitonin gene-related peptide on
axonal transport in isolated and cultured adult mouse dorsal root ganglion neurons.
Brain Res 883:184-191.
Horwitz SB (1992) Mechanism of action of taxol. Trends Pharmacol Sci 13:134-136.
Hunter JC, Gogas KR, Hedley LR, Jacobson LO, Kassotakis L, Thompson J and Fontana
DJ (1997) The effect of novel anti-epileptic drugs in rat experimental models of
acute and chronic pain. Eur J Pharmacol 324:153-160.
Hwang JH and Yaksh TL (1997) Effect of subarachnoid gabapentin on tactile-evoked
allodynia in a surgically induced neuropathic pain model in the rat. Reg Anesth
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
21
22:249-256.
Inoue M, Rashid MH, Fujita R, Contos JJ, Chun J and Ueda H (2004) Initiation of
neuropathic pain requires lysophosphatidic acid receptor signaling. Nat Med
10:712-718.
Josephson A, Widenfalk J, Trifunovski A, Widmer HR, Olson L and Spenger C (2001)
GDNF and NGF family members and receptors in human fetal and adult spinal
cord and dorsal root ganglia. J Comp Neurol 440:204-217.
Katims JJ (1997) Neuroselective current perception threshold quantitative sensory test.
Muscle Nerve 20:1468-1469.
Kiso T, Nagakura Y, Toya T, Matsumoto N, Tamura S, Ito H, Okada M and Yamaguchi T
(2001) Neurometer measurement of current stimulus threshold in rats. J
Pharmacol Exp Ther 297:352-356.
Kohler DR and Goldspiel BR (1994) Paclitaxel (taxol). Pharmacotherapy 14:3-34.
Lengyel C, Torok T, Varkonyi T, Kempler P and Rudas L (1998) Baroreflex sensitivity
and heart-rate variability in insulin-dependent diabetics with polyneuropathy.
Lancet 351:1436-1437.
Luo ZD, Calcutt NA, Higuera ES, Valder CR, Song YH, Svensson CI and Myers RR
(2002) Injury type-specific calcium channel alpha 2 delta-1 subunit up-regulation
in rat neuropathic pain models correlates with antiallodynic effects of gabapentin.
J Pharmacol Exp Ther 303:1199-1205.
Maneuf YP, Blake R, Andrews NA and McKnight AT (2004) Reduction by gabapentin of
K+-evoked release of [3H]-glutamate from the caudal trigeminal nucleus of the
streptozotocin-treated rat. Br J Pharmacol 141:574-579.
Martin DJ, McClelland D, Herd MB, Sutton KG, Hall MD, Lee K, Pinnock RD and Scott
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
22
RH (2002) Gabapentin-mediated inhibition of voltage-activated Ca2+ channel
currents in cultured sensory neurones is dependent on culture conditions and
channel subunit expression. Neuropharmacology 42:353-366.
Masson EA and Boulton AJ (1991) The Neurometer: validation and comparison with
conventional tests for diabetic neuropathy. Diabet Med 8 Spec No:S63-66.
Miki S, Yoshinaga N, Iwamoto T, Yasuda T and Sato S (2001) Antinociceptive effect of
the novel compound OT-7100 in a diabetic neuropathy model. Eur J Pharmacol
430:229-234.
Patel MK, Gonzalez MI, Bramwell S, Pinnock RD and Lee K (2000) Gabapentin inhibits
excitatory synaptic transmission in the hyperalgesic spinal cord. Br J Pharmacol
130:1731-1734.
Pitei DL, Watkins PJ, Stevens MJ and Edmonds ME (1994) The value of the Neurometer
in assessing diabetic neuropathy by measurement of the current perception
threshold. Diabet Med 11:872-876.
Polomano RC, Mannes AJ, Clark US and Bennett GJ (2001) A painful peripheral
neuropathy in the rat produced by the chemotherapeutic drug, paclitaxel. Pain
94:293-304.
Rashid MH, Inoue M, Kondo S, Kawashima T, Bakoshi S and Ueda H (2003) Novel
expression of vanilloid receptor 1 on capsaicin-insensitive fibers accounts for the
analgesic effect of capsaicin cream in neuropathic pain. J Pharmacol Exp Ther
304:940-948.
Rowinsky EK, Eisenhauer EA, Chaudhry V, Arbuck SG and Donehower RC (1993)
Clinical toxicities encountered with paclitaxel (Taxol). Semin Oncol 20:1-15.
Schiff PB, Fant J and Horwitz SB (1979) Promotion of microtubule assembly in vitro by
taxol. Nature 277:665-667.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
23
Shimoyama M, Shimoyama N and Hori Y (2000) Gabapentin affects glutamatergic
excitatory neurotransmission in the rat dorsal horn. Pain 85:405-414.
Smith SB, Crager SE and Mogil JS (2004) Paclitaxel-induced neuropathic
hypersensitivity in mice: responses in 10 inbred mouse strains. Life Sci
74:2593-2604.
Socinski MA (1999) Single-agent paclitaxel in the treatment of advanced non-small cell
lung cancer. Oncologist 4:408-416.
Sutton KG, Martin DJ, Pinnock RD, Lee K and Scott RH (2002) Gabapentin inhibits
high-threshold calcium channel currents in cultured rat dorsal root ganglion
neurones. Br J Pharmacol 135:257-265.
Zimmermann M (1983) Ethical guidelines for investigations of experimental pain in
conscious animals. Pain 16:109-110.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
24
Footnotes
a) Financial assistance: Part of this study was supported by the Grant-in-Aid for the
Cancer Research (15-7) from the Ministry of Helth, Labour, and Welfare.
b) Reprint requests should be addressed to: Dr. Hiroshi Ueda,
Division of Molecular Pharmacology and Neuroscience, Nagasaki University Graduate
School of Biomedical Sciences, 1-14 Bunkyo-machi, Nagasaki 852-8521, Japan
Tel: +81-95-819-2421; Fax: +81-95-819-2420; Email: [email protected]
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
25
Legends for Figures
Fig. 1. Paclitaxel-induced neuropathic pain and antinociceptive effect of gabapentin.
(A) Time course of paclitaxel-induced mechanical allodynia from pre-injection (control)
to 4 weeks post-treatment. (B) Summation of paclitaxel-induced mechanical allodynia
determined as the AUC from week 1 to 4 in response to a single i.p. injection, single i.v.
injection or 4 i.p. injections on alternate days (multi). (C) Time course of
paclitaxel-induced thermal hyperalgesia from pre-injection to 4 weeks after treatment.
(D) Dose-dependent analgesic effects of gabapentin in mechanical paw pressure tests 2
weeks after paclitaxel treatment. (E) Time course of gabapentin (30 mg/kg)-induced
analgesia 2 weeks after paclitaxel treatment (4 mg/kg, i.p.). (F) Analgesic effects of
gabapentin in thermal paw withdrawal tests 2 weeks after paclitaxel treatment (4 mg/kg,
i.p.). Animals were analyzed 30 minutes after gabapentin (30 mg/kg) treatment: control
(C), saline (Sal) and gabapentin (GBP), *:p<0.05, **:p<0.01 vs. control (vehicle).
#:p<0.05, ##:p<0.01 vs. paclitaxel/saline. Data are presented as mean ± S.E.M. from
experiments using at least 6 mice.
Fig. 2. Paclitaxel-induced A-fiber hypersensitization and inhibitory effects of gabapentin.
(A) Time courses of paclitaxel (4 mg/kg, i.p.)-induced sensory fiber sensitizations
determined by EPW tests using Neurometer from pre-injection to 4 weeks after treatment.
(B) Alleviating effects of gabapentin on paclitaxel-induced A-fiber hypersensitivity
determined by EPW tests using 250 Hz and 2000 Hz stimulations 2 weeks after paclitaxel
treatment. All animals were analyzed 30 minutes after gabapentin treatment: Control (C),
saline (Sal) and gabapentin (GBP). *:p<0.05, **:p<0.01 vs. control or vehicle/saline.
##:p<0.01 vs. paclitaxel/saline. Data are presented as mean ± S.E.M. from experiments
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
26
using at least 6 mice.
Fig. 3. Expression of Caα2δ-1 mRNA and protein in L4-6 DRGs after
paclitaxel-treatment. (A. Left and Middle) Dissociation curves of PCR products from
reactions using primers specific for mRNA encoding Caα2δ-1 and GAPDH, and
amplification efficiency analysis. (A. Right) Quantitative RT-PCR analysis. Caα2δ-1 and
GAPDH mRNAs in L4-6 DRGs on 3rd, 7th and 14th day after paclitaxel treatment were
measured and Caα2δ-1 was normalized to GAPDH. The graph shows the relative amount
of Caα2δ-1 mRNA in paclitaxel-treated mice compared to the controls. (B) Western blot
analysis. Caα2δ-1 and β-Tubulin protein in L4-6 DRGs 7, 14 and 28 days after paclitaxel
treatment were measured and Caα2δ-1 was normalized to β-Tubulin. The graph shows
the relative amount of Caα2δ-1 protein in paclitaxel-treated mice compared to the
controls. Photograph shows representative data. *:p<0.05, **:p<0.01 vs. control (C).
Data are presented as mean ± S.E.M. from experiments using at least 8 mice.
Fig. 4. Increased expression of Caα2δ-1 protein in medium/large DRG neurons. (A) DRG
slices of 10 µm from day 14 after vehicle or paclitaxel treatment were stained with
antibodies against Caα2δ-1. (B) The percentage of Caα2δ-1 stained neurons in paclitaxel-
and vehicle-treated mice 2 weeks after treatment. The total 1684 DRG neurons from
vehicle-treated mice (n=3) and 2724 DRG neurons from paclitaxel treated mice (n=4)
were measured. (C) Size distribution of all neurons (total) and of Caα2δ-1 expressing
neurons in DRG in paclitaxel- and vehicle-treated mice, respectively. (D) Bar graph
represents the frequency distribution of Caα2δ-1 positive neurons normalized to cell size.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #103614
27
Line graph (solid line) represents the balance value of the vehicle- and paclitaxel- treated
group. Scale bar, 50 µm. *:p<0.05 vs. vehicle.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

This article has not been copyedited and form
atted. The final version m
ay differ from this version.
JPET
Fast Forward. Published on M
ay 10, 2006 as DO
I: 10.1124/jpet.106.103614 at ASPET Journals on June 18, 2018 jpet.aspetjournals.org Downloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from

This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 10, 2006 as DOI: 10.1124/jpet.106.103614
at ASPE
T Journals on June 18, 2018
jpet.aspetjournals.orgD
ownloaded from
This article has not been copyedited and form
atted. The final version m
ay differ from this version.
JPET
Fast Forward. Published on M
ay 10, 2006 as DO
I: 10.1124/jpet.106.103614 at ASPET Journals on June 18, 2018 jpet.aspetjournals.org Downloaded from


















