
Induction of haploid plants in iris (Iris pseudacorus) by...
5
596 http://journals.tubitak.gov.tr/agriculture/ Turkish Journal of Agriculture and Forestry Turk J Agric For (2015) 39: 596-600 © TÜBİTAK doi:10.3906/tar-1407-18 Induction of haploid plants in iris (Iris pseudacorus) by pollen irradiation Mohammad SADAT HOSSEINI GROUH 1,2, *, Naeimeh SOUSARAEI 3 , Mohammad AKBARI 4 , Vahid RAHIMI 5 , Hamid Reza BAYAT 3 1 Department of Horticulture, Faculty of Agriculture, University of Jiroſt, Jiroſt, Iran 2 Department of Horticultural Crops, Faculty of Agriculture, University of Jiroſt, Jiroſt, Iran 3 Department of Horticulture, College of Abouraihan, University of Tehran, Tehran, Iran 4 Department of Horticultural Sciences, Faculty of Agriculture, University of Tabriz, Tabriz, Iran 5 Department of Biotechnology and Molecular Genetics, Faculty of Agriculture, University of Mohagheghe Ardabili, Ardabil, Iran * Correspondence: [email protected] 1. Introduction Production of haploid plants that inherit chromosomes from just one parent can effectively accelerate plant breeding (Ravi and Chan, 2010). About 80 different genera and 1500 different species of the iris, which belong to the family Iridaceae, are found worldwide (Park et al., 2006). Because of the beautiful shape of the flowers and the elegance of the elongated leaves, these plants are commonly found in gardens and widely used in floral arrangements. eir popularity has encouraged the development and breeding of new varieties by the horticultural industry. Wendelbo (1977) reported that there are apparently twenty different species of the wild iris growing in Iran. Doubled haploidy is and will continue to be a very efficient tool for the production of completely homozygous lines from heterozygous donor plants in a single step. Haploids and doubled haploids have been extensively utilized in genetic studies such as gene mapping, marker/ trait association studies, location of QTLs, and genomics and as targets for transformations. Pollination with irradiated pollen is one method that is used to induce the formation of maternal haploids via intraspecific pollination. Embryo development is stimulated by pollen germination on the stigma, furthered by the growth of the pollen tube within the style. Nevertheless, irradiated pollen is unable to fertilize the egg cell under this condition. In vitro embryo rescue is necessary to recover the haploid embryos for most plant species. Spontaneous parthenogenesis is rather exceptional in plant species, and yet it is possible to be induced via several techniques. Haploid induction via in situ parthenogenesis has been reported in some plants. Examples are carnations (Dianthus caryophyllus L.) (Sato et al., 2000), apples (Malus domestica) (Zhang and Lespinasse, 1991; Höfer and Grafe, 2003), pears (Pyrus communis) (Bouvier et al., 1993), kiwi fruits (Actidinia deliciosa) (Chalak and Legave, 1997; Pandey et al., 1990), tangerines (Citrus reticulata) (Ollitrault et al., 1996), sweet cherries (Prunus avium) (Höfer and Grafe, 2003), and mandarins (Froelicher et al., 2007). In the Persian walnut, haploid induction was successfully performed via in situ parthenogenesis followed by immature embryo culture (Grouh et al., 2011). In this regard, X-rays have proven Abstract: is is the first report of successful regeneration of haploid lines in iris (Iris pseudacorus) developed by in situ parthenogenesis followed by embryo rescue. Native genotypes of I. pseudacorus (2n = 38) were prepared as female parents and I. spuria was set as the males. Anthers collected from the pollen donor were irradiated by X-ray with doses of 0, 100, 200, 300, and 400 Gy in 2012. A second round of experiments was conducted in 2013 with 300 and 400 Gy. e ovaries from each pollinated flower were collected and cultured and the produced plantlets were then acclimatized. Chromosome counting and flow cytometry were used to determine the ploidy levels. e viability of irradiated pollens was also evaluated. Pollination with irradiated pollen at doses ranging from 100 to 400 Gy can significantly reduce fruit set, particularly at higher doses (300 and 400 Gy). e embryo production was reduced in I. pseudacorus as doses increased; the highest doses led to reduction in the number of regenerants with the formation of just 6 and 4 embryos at 300 and 400 Gy, respectively. Haploid plantlets were produced when the pollen grains were exposed to 300 and 400 Gy of X-ray. Irradiation at 100 and 200 Gy was found to be insufficient to cause pollen sterility. e best dose for haploid production in I. pseudacorus is hereby found to be 300 or 400 Gy of X-ray. Key words: Line, flow cytometry, chromosome counting, X-ray irradiation, embryo rescue Received: 05.07.2014 Accepted: 07.06.2015 Published Online: 08.07.2015 Printed: 30.07.2015 Research Article
-
Upload
dangkhuong -
Category
Documents
-
view
214 -
download
0
Transcript of Induction of haploid plants in iris (Iris pseudacorus) by...

596
http://journals.tubitak.gov.tr/agriculture/
Turkish Journal of Agriculture and Forestry Turk J Agric For(2015) 39: 596-600© TÜBİTAKdoi:10.3906/tar-1407-18
Induction of haploid plants in iris (Iris pseudacorus) by pollen irradiation
Mohammad SADAT HOSSEINI GROUH1,2,*, Naeimeh SOUSARAEI3, Mohammad AKBARI4,Vahid RAHIMI5, Hamid Reza BAYAT3
1Department of Horticulture, Faculty of Agriculture, University of Jiroft, Jiroft, Iran2Department of Horticultural Crops, Faculty of Agriculture, University of Jiroft, Jiroft, Iran
3Department of Horticulture, College of Abouraihan, University of Tehran, Tehran, Iran4Department of Horticultural Sciences, Faculty of Agriculture, University of Tabriz, Tabriz, Iran
5Department of Biotechnology and Molecular Genetics, Faculty of Agriculture, University of Mohagheghe Ardabili, Ardabil, Iran
* Correspondence: [email protected]
1. IntroductionProduction of haploid plants that inherit chromosomes from just one parent can effectively accelerate plant breeding (Ravi and Chan, 2010). About 80 different genera and 1500 different species of the iris, which belong to the family Iridaceae, are found worldwide (Park et al., 2006). Because of the beautiful shape of the flowers and the elegance of the elongated leaves, these plants are commonly found in gardens and widely used in floral arrangements. Their popularity has encouraged the development and breeding of new varieties by the horticultural industry. Wendelbo (1977) reported that there are apparently twenty different species of the wild iris growing in Iran.
Doubled haploidy is and will continue to be a very efficient tool for the production of completely homozygous lines from heterozygous donor plants in a single step. Haploids and doubled haploids have been extensively utilized in genetic studies such as gene mapping, marker/trait association studies, location of QTLs, and genomics and as targets for transformations. Pollination with irradiated pollen is one method that is used to induce the
formation of maternal haploids via intraspecific pollination. Embryo development is stimulated by pollen germination on the stigma, furthered by the growth of the pollen tube within the style. Nevertheless, irradiated pollen is unable to fertilize the egg cell under this condition. In vitro embryo rescue is necessary to recover the haploid embryos for most plant species. Spontaneous parthenogenesis is rather exceptional in plant species, and yet it is possible to be induced via several techniques. Haploid induction via in situ parthenogenesis has been reported in some plants. Examples are carnations (Dianthus caryophyllus L.) (Sato et al., 2000), apples (Malus domestica) (Zhang and Lespinasse, 1991; Höfer and Grafe, 2003), pears (Pyrus communis) (Bouvier et al., 1993), kiwi fruits (Actidinia deliciosa) (Chalak and Legave, 1997; Pandey et al., 1990), tangerines (Citrus reticulata) (Ollitrault et al., 1996), sweet cherries (Prunus avium) (Höfer and Grafe, 2003), and mandarins (Froelicher et al., 2007). In the Persian walnut, haploid induction was successfully performed via in situ parthenogenesis followed by immature embryo culture (Grouh et al., 2011). In this regard, X-rays have proven
Abstract: This is the first report of successful regeneration of haploid lines in iris (Iris pseudacorus) developed by in situ parthenogenesis followed by embryo rescue. Native genotypes of I. pseudacorus (2n = 38) were prepared as female parents and I. spuria was set as the males. Anthers collected from the pollen donor were irradiated by X-ray with doses of 0, 100, 200, 300, and 400 Gy in 2012. A second round of experiments was conducted in 2013 with 300 and 400 Gy. The ovaries from each pollinated flower were collected and cultured and the produced plantlets were then acclimatized. Chromosome counting and flow cytometry were used to determine the ploidy levels. The viability of irradiated pollens was also evaluated. Pollination with irradiated pollen at doses ranging from 100 to 400 Gy can significantly reduce fruit set, particularly at higher doses (300 and 400 Gy). The embryo production was reduced in I. pseudacorus as doses increased; the highest doses led to reduction in the number of regenerants with the formation of just 6 and 4 embryos at 300 and 400 Gy, respectively. Haploid plantlets were produced when the pollen grains were exposed to 300 and 400 Gy of X-ray. Irradiation at 100 and 200 Gy was found to be insufficient to cause pollen sterility. The best dose for haploid production in I. pseudacorus is hereby found to be 300 or 400 Gy of X-ray.
Key words: Line, flow cytometry, chromosome counting, X-ray irradiation, embryo rescue
Received: 05.07.2014 Accepted: 07.06.2015 Published Online: 08.07.2015 Printed: 30.07.2015
Research Article

597
SADAT HOSSEINI GROUH et al. / Turk J Agric For
to be efficient for melons (Katoh et al., 1993), carnations (Dianthus caryophyllus) (Sato et al., 2000), pomelos (Citrus maxima) (Yahata et al., 2010), and pumpkins (Košmrlj et al., 2013). As mentioned above, the iris is an important ornamental plant that has not been reported to have been bred through haploid lines as of yet. Therefore, the present investigation was set to evaluate the function of pollen grains, irradiated by X-ray, with the aim of producing haploid plants in the iris (Iris pseudacorus). The viability of irradiated pollens and the development of parthenogenetic embryos are also investigated.
2. Materials and methods2.1. Plant material and culture conditions The experiments were carried out in 2012 and 2013 in the laboratory and greenhouse complex of the Plant Science Department of the University of Jiroft, Jiroft, Kerman, Iran. Female parents were native genotypes of I. pseudacorus (2n = 38) collected from a rice farm area of Mazandaran, Iran (altitude: 100 m), on 15.05.2006. The male parents were specimens of I. spuria collected from Farash village, Jiroft, Kerman (altitude: 600 m), on 14.02.2007. Plants were grown under air temperature of 12 to 17 °C with 16 h of light and 8 h of darkness in the greenhouse. The soil temperature was maintained at 15 °C.2.2. PseudofertilizationPollen grains were collected from undehisced anthers of flower buds that were placed on petri dishes until the occurrence of dehiscence. This step was performed under sterilized conditions. The collected anthers from the pollen donor were irradiated with X-rays (TR50G, Triup Co., Nanjing, China) of 0, 100, 200, 300, and 400 Gy in 2012 and with 300 and 400 Gy in 2013. Immature flower buds of the ovule donors were emasculated 5 days prior to anthesis and were bagged to avoid unwanted outcrossing. Irradiated pollens were used for pollinating the flower buds of the ovule donor. Pollinated flowers were isolated to avoid undesirable pollen contamination. Paper bags were removed the second or third day after pollination. 2.3. In vitro germination of irradiation pollen Germination rates of the irradiated pollens were evaluated on a medium containing 10% sucrose, 1.0 mM CaCl2, and 0.16 mM boric acid solidified with 0.65% Difco Bacto-Agar (Merck, Darmstadt, Germany) at pH 5.7 in petri dishes (55 mm) at 28 °C and 100% relative humidity (Grouh et al., 2011). Germination was recorded after 24 h. Three replications of 100 pollen grains in each plate were observed.2.4. Ovule cultureAfter 2–4 weeks following pollination with irradiated pollen grains, the ovaries from each pollinated flower were collected. The ovaries were then sterilized with 70% ethanol, inundated in hypochlorite solution (containing
1% chlorine) for 15 min, and washed with distilled sterilized water five times. Isolated ovules from the ovaries (along with the placenta) were placed on MS medium (Murashige and Skoog, 1962) containing 2 mM NAA, 2 mM BAP, and 6% sucrose as indicated by Sato et al. (2000). The ovules were maintained at 24 °C with 12 h of light and 12 h of darkness for 5 weeks. The regenerated shoots were transferred to MS medium including 3% sucrose and 0.8% agar without any plant growth regulators. The produced plantlets were moved to mist conditions for acclimatization and were transferred to the greenhouse thereafter.2.5. Chromosome number determinationChromosome number was counted in the cells of the root tip obtained from in vitro-grown haploid and diploid plantlets using the standard Feulgen technique (Lillie, 1951). The root samples were pretreated with 0.002 M hydroxyquinoline for 8 h (4 h at room temperature and then 4 h at 4 °C). The samples were fixed for 24 h in 3:1 ethanol acetic acid and were then stored in 70% ethanol at 4 °C until the screening of chromosomes. Fixed root tips were placed in aceto-iron hematoxylin dye in order to stain chromosomes (Lu and Raju, 1970). The fixed root tips were hydrolyzed in 1 N HCl for 12 min at 60 °C and were then squashed in a drop of 45% (v/v) acetic acid.2.6. Flow cytometry analysisFlow cytometry was used to determine the ploidy level of samples prepared from leaves of the haploid and diploid (control) plants. Nuclei were released from 0.5 cm2 of leaf tissue by chopping with a razor blade for 25 s in 500 mL of modified Galbraith’s nuclei isolation buffer (Galbraith et al., 1983) containing 200 mM Tris, 4 mM MgCl2.6H2O, pH 7.5, and 0.5% Triton X-100 (Partec, Munster, Germany). Subsequently, 500 mL of the staining solution 4’,6-diamidino-2-phenylindole was added for DNA staining. After 2 min of incubation, nuclei were passed through a 30-mm nylon filter to eliminate cell debris. The samples were analyzed using a flow cytometer (PA-I; Partec).
3. Results and discussion 3.1. In vitro viability of irradiated pollenIn vitro germination of irradiated pollen was determined in both 2012 and 2013. The germination of nonirradiated pollen was about 84% compared to 65% for 100 Gy and 53% for 200 Gy in 2012. Germination capacity for pollens irradiated at doses of 400 Gy was significantly reduced to about 27% (Table 1). This also implies that I. spuria is more sensitive to ionizing radiation than other crops such as mandarin, whose gamma ray-irradiated pollens demonstrated no significant differences in germination rates between all applied doses (150, 300, 600, and 900 Gy) of radiation (Froelicher et al., 2007). In a study conducted by Höfer and Grafe (2003), sweet cherry pollens γ-irradiated

598
SADAT HOSSEINI GROUH et al. / Turk J Agric For
at doses ranging from 250 to 1200 Gy had no significant in vitro germination differences from the control. It has also been indicated that irradiation of pollen barely affected the in vitro germination in pomelo, such that X-ray irradiation at 1000 Gy had no significant effect on pollen germination (Yahata et al., 2010). On the other hand, similar to our observations, De Lange and Vincent (1988) found that in vitro pollen germination of trifoliate orange, pomelo, and sweet lime was reduced at irradiation doses over 500 Gy γ-rays. In European plum (Prunus domestica cv. Rainha Claudia Verde) it has also been reported that all applied irradiation levels had a significant effect on pollen viability (Peixe et al., 2000). It can be inferred, therefore, that the response of pollen to radiation probably relies on the plant genotype and the type of radiation used.3.2. Fruit set and haploid productionIn this study, the I. pseudacorus species responded positively to X-rays for the induction of haploid embryogenesis. For the first time haploid plants of Iris were obtained. Data for
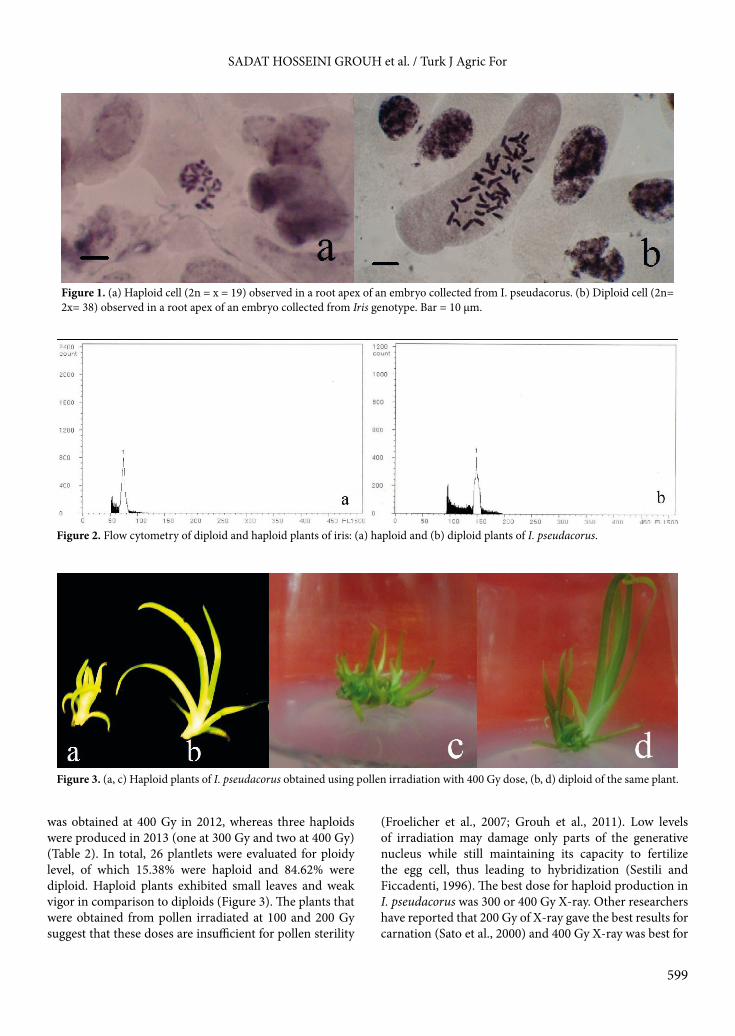
fruit set and haploid induction were recorded in 2012 and 2013. Results indicated that pollination with irradiated pollen at doses ranging from 100 to 400 Gy significantly reduced the fruit set, particularly at higher doses (300 and 400 Gy) (Table 2). As the irradiation doses increased, the embryo production reduced in I. pseudacorus. The number of regenerants derived from embryos reduced to 6 and 4 at 300 and 400 Gy, respectively (Table 2). These results are somewhat parallel to our observation in walnut pollen, where irradiation at doses ranging from 50 to 900 Gy considerably decreased the fruit set. According to those observations, we formed the idea that the degree to which irradiated pollen affected fruit set was dependent on genotype and irradiation dose (Grouh et al., 2011). As the literature indicates, the fruit set, mean embryo number, and haploid embryo induction are probably affected by the plant genotype, type of applied radiation, and amount of radiation dose (Froelicher et al., 2007; Gałązka and Niemirowicz-Szczytt, 2013). Increasing the X-ray radiation doses from 0 to 350 Gy in hull-less seed pumpkin (Cucurbita pepo subsp. pepo var. styriaca) demonstrated a gradual reduction in fruit set and mean number of embryos from the control (55.6% and 73.4) to 350 Gy (25.0% and 18.8), respectively. Fruit set reduction with increasing radiation doses was also reported in melon (Lotfi et al., 2003), European plum (Peixe et al., 2000), and cacao (Falque et al., 1992). Here in iris it seems that the fruit set and haploid production is, after all, hardly dependent on the type of applied radiation and the amount of radiation dose.3.3. Determination of ploidy levelPloidy level was determined by chromosome counting and flow cytometry (Figures 1 and 2). Nineteen chromosomes were visible in haploids. All the plantlets that were obtained at from 0 to 200 Gy were diploid. Haploid plantlets were produced when 300 or 400 Gy was applied. One haploid
Table 1. Mean in vitro germination percentage of X-irradiated pollen of native genotypes I. spuria in 2012 and 2013.
Irradiation dose (Gy)Germination rate (%)
2012 2013
0 84.23 a -
100 65.2 b -
200 53.33 c -
300 52 c 53.60 a
400 27 d 26.06 b
Percentages followed by the same letters are not significantly different at the 5% level by Duncan’s multiple range test.
Table 2. Effect of irradiation dose on fruit set, regenerant formation, and haploid induction using I. spuria as a pollen donor and I. pseudacorus as a female parent in 2012 and 2013.
Year Irradiation dose (Gy)
Pollinated flower number Fruit set (%) Regenerant
numberPloidy leveln 2n
2012 0 35 47 10 0 10
100 30 25 8 0 7
200 29 24 8 0 7
300 32 19 5 0 3
400 31 18 5 1 2
2013 300 30 21 6 1 2
400 28 19 4 2 1
599
SADAT HOSSEINI GROUH et al. / Turk J Agric For
was obtained at 400 Gy in 2012, whereas three haploids were produced in 2013 (one at 300 Gy and two at 400 Gy) (Table 2). In total, 26 plantlets were evaluated for ploidy level, of which 15.38% were haploid and 84.62% were diploid. Haploid plants exhibited small leaves and weak vigor in comparison to diploids (Figure 3). The plants that were obtained from pollen irradiated at 100 and 200 Gy suggest that these doses are insufficient for pollen sterility
(Froelicher et al., 2007; Grouh et al., 2011). Low levels of irradiation may damage only parts of the generative nucleus while still maintaining its capacity to fertilize the egg cell, thus leading to hybridization (Sestili and Ficcadenti, 1996). The best dose for haploid production in I. pseudacorus was 300 or 400 Gy X-ray. Other researchers have reported that 200 Gy of X-ray gave the best results for carnation (Sato et al., 2000) and 400 Gy X-ray was best for
Figure 1. (a) Haploid cell (2n = x = 19) observed in a root apex of an embryo collected from I. pseudacorus. (b) Diploid cell (2n= 2x= 38) observed in a root apex of an embryo collected from Iris genotype. Bar = 10 µm.
Figure 2. Flow cytometry of diploid and haploid plants of iris: (a) haploid and (b) diploid plants of I. pseudacorus.
Figure 3. (a, c) Haploid plants of I. pseudacorus obtained using pollen irradiation with 400 Gy dose, (b, d) diploid of the same plant.

600
SADAT HOSSEINI GROUH et al. / Turk J Agric For
pomelo (Yahata et al., 2010). Future studies could test 350, 500, and 600 Gy to find the capacity of these treatments to produce more haploids with fewer diploids and also to increase seed production. Thus far, the produced plantlets have been moved to the misters to be acclimatized and have also been transferred to the greenhouse. Doubling of the chromosome number will be carried out for the purpose of breeding programs. In conclusion, this work
shows efficient inductions of in situ parthenogenesis in I. pseudacorus. However, it remains to be recognized as the origin of haploid and diploid plants.
Acknowledgments We thank the University of Jiroft for financial support. Mr Mohsen Hamedpour Darabi is also acknowledged for editing the manuscript.
References
Bouvier L, Zhang YX, Lespinasse Y (1993). Two methods of haploidization in pear, Pyrus communis L.: greenhouse seedling selection and in situ parthenogenesis induced by irradiated pollen. Theor Appl Genet 87: 229–232.
Chalak L, Legave J (1997). Effects of pollination by irradiated pollen in Hayward kiwifruit and spontaneous doubling of induced parthenogenetic trihaploids. Sci Hort 68: 83–93.
De Lange J H, Vincent A P (1988). Studies on Citrus pollination using gamma-irradiated pollen. S Afr J Bot 54: 257–264.
Falque M, Kodia A, Sounigo O, Eskes A, Charrier A (1992). Gamma-irradiation of cacao (Theobroma cacao L.) pollen: effect on pollen grain viability, germination and mitosis and on fruit set. Euphytica 64: 167–172.
Froelicher Y, Bassene JB, Jedidi-Neji E, Dambier D, Morillon R, Bernardini G, Costantino G, Ollitrault P (2007). Induced parthenogenesis in mandarin for haploid production: induction procedures and genetic analysis of plantlets. Plant Cell Rep 26: 937–944.
Gałązka J, Niemirowicz-Szczytt K (2013). Review of research on haploid production in cucumber and other cucurbits. Fol Hort 25: 67–78.
Galbraith DW, Harkins KR, Maddox JM, Ayres NM, Sharma DP, Firoozabady E (1983). Rapid flow cytometric analysis of the cell cycle in intact plant tissues. Science 220: 1049–1051.
Grouh MSH, Vahdati K, Lotfi M, Hassani D, Biranvand NP (2011). Production of haploids in Persian walnut through parthenogenesis induced by gamma-irradiated pollen. J Amer Soc Hort Sci 136: 198–204.
Höfer M, Grafe C (2003). Induction of doubled haploids in sweet cherry (Prunus avium L.). Euphytica 130: 191–197.
Katoh N, Hagimori M, Iwai S (1993). Production of haploid plants of melon by pseudofertilized ovule culture. Plant Tissue Cult Lett 10: 60–66.
Košmrlj K, Murovec J, Bohanec B (2013). Haploid induction in hull-less seed pumpkin through parthenogenesis induced by X-ray-irradiated pollen. J Amer Soc Hort Sci 138: 310–316.
Lillie R (1951). Simplification of the manufacture of Schiff reagent for use in histochemical procedures. Biotec His 26: 163–165.
Lotfi M, Alan A, Henning M, Jahn M, Earle E (2003). Production of haploid and doubled haploid plants of melon (Cucumis melo L.) for use in breeding for multiple virus resistance. Plant Cell Rep 21: 1121–1128.
Lu BC, Raju NB (1970). Meiosis in Coprinus. II. Chromosome pairing and the lamphrush diploid stage of meiotic prophase. Chromosoma 29: 305–316.
Murashige T, Skoog F (1962). A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plant 15: 473–497.
Ollitrault P, Allent V, Luro F (1996). Production of haploid plants and embryogenic calli of clementine (Citrus reticulata Blanco) after in situ parthenogenesis induced by irradiated pollen. Proc Int Soc Cit S Afr 2: 913–917.
Pandey K, Przywara L, Sanders P (1990). Induced parthenogenesis in kiwifruit (Actinidia deliciosa) through the use of lethally irradiated pollen. Euphytica 51: 1–9.
Park YW, Kim DM, Hwang YJ, Lim KB, Kim HH (2006). Karyotype analysis of three Korean native Iris species. Hort Environ Biotech 47: 51–54.
Peixe A, Campos M, Cavaleiro C, Barroso J, Pais M (2000). Gamma-irradiated pollen induces the formation of 2n endosperm and abnormal embryo development in European plum (Prunus domestica L., cv.“Rainha Cláudia Verde”). Sci Hort 86: 267–278.
Ravi M, Chan SW (2010). Haploid plants produced by centromere-mediated genome elimination. Nature 464: 615–618.
Sato S, Katoh N, Yoshida H, Iwai S, Hagimori M (2000). Production of doubled haploid plants of carnation (Dianthus caryophyllus L.) by pseudofertilized ovule culture. Sci Hort 83: 301–310.
Sestili S, Ficcadenti N (1996). Irradiated pollen for haploid production. In: Jain SM, Sopory SK, Veilleux RE, editors. In Vitro Haploid Production in Higher Plants. 3rd Ed. Dordrecht, the Netherlands: Kluwer, pp. 263–247.
Wendelbo P (1977). Tulips and Irises of Iran and Their Relatives. Tehran, Iran: Ariamehr Botanical Garden.
Yahata M, Yasuda K, Nagasawa K, Harusaki S, Komatsu H, Kunitake H (2010). Production of haploid plant of ‘Banpeiyu’ pummelo [Citrus maxima (Burm.) Merr.] by pollination with soft X-ray-irradiated pollen. J Japan Soc Hort Sci 79: 239–245.
Zhang Y, Lespinasse Y (1991). Pollination with gamma-irradiated pollen and development of fruits, seeds and parthenogenetic plants in apple. Euphytica 54: 101–109.


















