
Induction of Antimicrobial Pathways during Early-Phase ... · typhoid, which remains a disease of...
9
INFECTION AND IMMUNITY, Aug. 2003, p. 4733–4741 Vol. 71, No. 8 0019-9567/03/$08.000 DOI: 10.1128/IAI.71.8.4733–4741.2003 Copyright © 2003, American Society for Microbiology. All Rights Reserved. Induction of Antimicrobial Pathways during Early-Phase Immune Response to Salmonella spp. in Murine Macrophages: Gamma Interferon (IFN-) and Upregulation of IFN- Receptor Alpha Expression Are Required for NADPH Phagocytic Oxidase gp91-Stimulated Oxidative Burst and Control of Virulent Salmonella spp. N. Foster, S. D. Hulme, and P. A. Barrow* Division of Environmental Microbiology, Institute for Animal Health, Compton Laboratory, Newbury, Berkshire RG20 7NN, United Kingdom Received 4 December 2002/Returned for modification 11 February 2003/Accepted 1 May 2003 The effect of gamma interferon (IFN-) on elevation of reactive oxygen species and the viability of virulent wild-type and avirulent mutants of Salmonella enterica serovar Typhimurium and S. enterica serovar Infantis was studied in a murine macrophage cell line (J774.2 cells). S. enterica serovar Typhimurium 14028 phoP and a rough lipopolysaccharide mutant of S. enterica serovar Infantis 1326/28 ( r ) (avirulent mutants) induced NADPH phagocytic oxidase gp91 (gp91 phox ) activity and a significant (P < 0.05) elevation of reactive oxygen species within 12 h without coculture with IFN-. This coincided with reduced survival of S. enterica serovar Typhimurium14028 phoP or stasis of S. enterica serovar Infantis r . Fluorometric studies indicated that expression of IFN- on infected J774.2 cells was not significantly (P > 0.05) elevated. However, studies with the virulent S. enterica serovar Typhimurium strains showed that a comparable level of control of bacterial numbers could only be achieved by coculture with IFN-. This coincided with significant upregulation of IFN- receptor alpha expression on the surface of J774.2 cells and was completely abolished by N-acetyl-L-cysteine captopril (an inhibitor of reactive oxygen species). Delay in reactive oxygen species induction due to a requirement for IFN- and upregulation of IFN- receptor alpha in macrophages infected with virulent salmonellae may result in greater dissemination of virulent salmonellae in host tissue. Over a number of years, Salmonella enterica serovar Typhi- murium infection of mice has been used as a model of human typhoid, which remains a disease of global importance. Mu- tants of S. enterica serovar Typhimurium that are unable to survive in murine macrophages are avirulent (10), and this has stimulated a great deal of interest in the nature of the killing pathways employed by murine macrophages, including the generation of reactive nitrogen species via the inducible nitric oxide synthase pathway and reactive oxygen species (ROS) via the NADPH phagocytic oxidase system. Previous studies have suggested that the oxidative burst is an important killing pathway employed by murine macrophages infected with salmonellae. Mutations in the S. enterica serovar Typhimurium recombination genes induce avirulence due to an inability of the salmonellae to repair DNA following expo- sure to the oxidative burst in J774 cells (2). Other studies have suggested that S. enterica serovar Typhimurium utilizes super- oxide dismutase to protect periplasmic membranes against the effect of peroxynitrite formation and that sodC mutants are susceptible to peroxynitrite killing and are attenuated (5). Re- cent reports have indicated that reactive oxygen species (ROS) precede inducible nitric oxide synthase when knockout mice are challenged with S. enterica serovar Typhimurium, and it has been suggested that the timing of these responses is augmented by gamma interferon (IFN-) (24, 32). The generation of ROS occurs via a membrane-bound fla- vocytochrome b 558 , consisting of two phagocytic oxidase com- ponents (gp91 phox and p22 phox ) and four cytosolic components, p40 phox , 47 phox , p67 phox , and a GTP-binding Rac protein. Dur- ing activation, the cytosolic components translocate to the site of gp91 phox /p22 phox on the phagosomal membrane, forming a functional enzyme complex which generates ROS by catalyzing electron transfer from NADPH to molecular oxygen (21, 31). NADPH phagocytic oxidase (NADPH phox ) is active in murine macrophages infected with S. enterica serovar Typhimurium (5), and greater numbers of S. enterica serovar Typhimurium are recovered from the livers and spleens of gp91 phox / C57BL/6 mice than from wild-type mice (24). Furthermore, a more recent study suggested that Salmonella pathogenicity is- land 2 genes are required for evasion of the murine macro- phage oxidative burst by preventing oxidase trafficking (32). Therefore, wild-type salmonellae may survive longer in mac- rophages due to their ability to evade ROS via Salmonella pathogenicity island 2 genes, but the NADPH phox system ulti- mately controls the early phase of infection and possibly pre- vents it from overwhelming the host immune response. In the animal studies mentioned above, murine macro- phages would have been stimulated by cytokines produced by * Corresponding author. Mailing address: Division of Environmen- tal Microbiology, Institute for Animal Health, Compton Laboratory, Compton, Newbury, Berkshire RG20 7NN, United Kingdom. Phone: 44 1635 578411. Fax: 44 1635 577263. E-mail: [email protected] .uk. 4733 on August 19, 2019 by guest http://iai.asm.org/ Downloaded from
Transcript of Induction of Antimicrobial Pathways during Early-Phase ... · typhoid, which remains a disease of...

INFECTION AND IMMUNITY, Aug. 2003, p. 4733–4741 Vol. 71, No. 80019-9567/03/$08.00�0 DOI: 10.1128/IAI.71.8.4733–4741.2003Copyright © 2003, American Society for Microbiology. All Rights Reserved.
Induction of Antimicrobial Pathways during Early-Phase ImmuneResponse to Salmonella spp. in Murine Macrophages: Gamma
Interferon (IFN-�) and Upregulation of IFN-� ReceptorAlpha Expression Are Required for NADPH Phagocytic
Oxidase gp91-Stimulated Oxidative Burst andControl of Virulent Salmonella spp.
N. Foster, S. D. Hulme, and P. A. Barrow*Division of Environmental Microbiology, Institute for Animal Health, Compton Laboratory,
Newbury, Berkshire RG20 7NN, United Kingdom
Received 4 December 2002/Returned for modification 11 February 2003/Accepted 1 May 2003
The effect of gamma interferon (IFN-�) on elevation of reactive oxygen species and the viability of virulentwild-type and avirulent mutants of Salmonella enterica serovar Typhimurium and S. enterica serovar Infantiswas studied in a murine macrophage cell line (J774.2 cells). S. enterica serovar Typhimurium 14028 phoP anda rough lipopolysaccharide mutant of S. enterica serovar Infantis 1326/28 (�r) (avirulent mutants) inducedNADPH phagocytic oxidase gp91 (gp91phox) activity and a significant (P < 0.05) elevation of reactive oxygenspecies within 12 h without coculture with IFN-�. This coincided with reduced survival of S. enterica serovarTyphimurium14028 phoP or stasis of S. enterica serovar Infantis �r. Fluorometric studies indicated thatexpression of IFN-� on infected J774.2 cells was not significantly (P > 0.05) elevated. However, studies with thevirulent S. enterica serovar Typhimurium strains showed that a comparable level of control of bacterialnumbers could only be achieved by coculture with IFN-�. This coincided with significant upregulation of IFN-�receptor alpha expression on the surface of J774.2 cells and was completely abolished by N-acetyl-L-cysteinecaptopril (an inhibitor of reactive oxygen species). Delay in reactive oxygen species induction due to arequirement for IFN-� and upregulation of IFN-� receptor alpha in macrophages infected with virulentsalmonellae may result in greater dissemination of virulent salmonellae in host tissue.
Over a number of years, Salmonella enterica serovar Typhi-murium infection of mice has been used as a model of humantyphoid, which remains a disease of global importance. Mu-tants of S. enterica serovar Typhimurium that are unable tosurvive in murine macrophages are avirulent (10), and this hasstimulated a great deal of interest in the nature of the killingpathways employed by murine macrophages, including thegeneration of reactive nitrogen species via the inducible nitricoxide synthase pathway and reactive oxygen species (ROS) viathe NADPH phagocytic oxidase system.
Previous studies have suggested that the oxidative burst is animportant killing pathway employed by murine macrophagesinfected with salmonellae. Mutations in the S. enterica serovarTyphimurium recombination genes induce avirulence due toan inability of the salmonellae to repair DNA following expo-sure to the oxidative burst in J774 cells (2). Other studies havesuggested that S. enterica serovar Typhimurium utilizes super-oxide dismutase to protect periplasmic membranes against theeffect of peroxynitrite formation and that sodC mutants aresusceptible to peroxynitrite killing and are attenuated (5). Re-cent reports have indicated that reactive oxygen species (ROS)
precede inducible nitric oxide synthase when knockout miceare challenged with S. enterica serovar Typhimurium, and it hasbeen suggested that the timing of these responses is augmentedby gamma interferon (IFN-�) (24, 32).
The generation of ROS occurs via a membrane-bound fla-vocytochrome b558, consisting of two phagocytic oxidase com-ponents (gp91phox and p22phox) and four cytosolic components,p40phox, 47phox, p67phox, and a GTP-binding Rac protein. Dur-ing activation, the cytosolic components translocate to the siteof gp91phox/p22phox on the phagosomal membrane, forming afunctional enzyme complex which generates ROS by catalyzingelectron transfer from NADPH to molecular oxygen (21, 31).NADPH phagocytic oxidase (NADPHphox) is active in murinemacrophages infected with S. enterica serovar Typhimurium(5), and greater numbers of S. enterica serovar Typhimuriumare recovered from the livers and spleens of gp91phox�/�
C57BL/6 mice than from wild-type mice (24). Furthermore, amore recent study suggested that Salmonella pathogenicity is-land 2 genes are required for evasion of the murine macro-phage oxidative burst by preventing oxidase trafficking (32).Therefore, wild-type salmonellae may survive longer in mac-rophages due to their ability to evade ROS via Salmonellapathogenicity island 2 genes, but the NADPHphox system ulti-mately controls the early phase of infection and possibly pre-vents it from overwhelming the host immune response.
In the animal studies mentioned above, murine macro-phages would have been stimulated by cytokines produced by
* Corresponding author. Mailing address: Division of Environmen-tal Microbiology, Institute for Animal Health, Compton Laboratory,Compton, Newbury, Berkshire RG20 7NN, United Kingdom. Phone:44 1635 578411. Fax: 44 1635 577263. E-mail: [email protected].
4733
on August 19, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

infected cells at the site of invasion while ex vivo macrophageswere stimulated with 20 U of IFN-� per ml (24). Lower levelsof IFN-� are produced by natural killer cells early in infection,while the most productive source of IFN-� comes from Tlymphocytes active later in infection (19). Lipopolysaccharideand IFN-� synergistically stimulate the signal-transducing ac-tivator of transcription 1 (STAT1) signaling pathway in murinemacrophage-like RAW 264.7 cells (14). In vivo studies havealso shown the importance of IFN-� in mediating responses tolipopolysaccharide in mice, especially in the lethal shock reac-tion (Schwartzman reaction) (15). However, none of thesestudies elucidated the full effect of IFN-� in terms of thegeneration of ROS during the early phase of infection withvirulent or avirulent salmonellae.
The aim of the work described here was to investigate theoxidative burst in Salmonella-infected J774.2 cells and to de-termine the effect of IFN-� on this killing pathway in theabsence of an effective inducible nitric oxide synthase pathwayduring infection of J774.2 cells with virulent and avirulent S.enterica serovar Typhimurium. Furthermore, the study was de-signed to ascertain why ROS are important in Salmonella con-trol early in infection compared to later control by induciblenitric oxide synthase.
MATERIALS AND METHODS
Bacterial culture and strains. Bacteria were grown in Luria-Bertani (LB)broth (Life Technologies Ltd., Paisley, United Kingdom) for 18 h at 37°C underagitation. The bacteria were then subcultured in fresh LB broth for 4 h to late logphase (established by conventional counts of CFU). Prior to incubation withJ774.2 cells, bacteria were adjusted to a multiplicity of infection (MOI) of 10.During this study, the following strains and mutants were used: S. enterica serovarTyphimurium 14028 (American Type Culture Collection strain); S. enterica se-rovar Typhimurium CS022 (phoP mutant of 14028, a gift from S. I. Miller,Harvard University), which does not survive in macrophages (25); S. entericaserovar Typhimurium F98 (virulent in mice); and a rough mutant of S. entericaserovar Infantis, 1326/28 �r, which is avirulent in mice (1).
In a separate study, the effect of other murine typhoid-inducing (S. entericaserovar Typhimurium 4/74, S. enterica serovar Enteritidis KMS1977, S. entericaserovar Dublin 2229, and S. enterica serovar Choleraesuis A50) and non-typhoid-inducing (S. enterica serovar Gallinarum 9, S. enterica serovar Kedougou GP, andS. enterica serovar Montevideo KMS) strains were analyzed. Growth curves foreach serovar were obtained as previously stated.
Cell culture. J774.2 cells were grown to confluence in 96-well plates (Nunc,Naperville, Ill.) containing RPMI 1640 medium at 37°C in CO2 (5%, vol/vol).The cells were then washed three times in phosphate-buffered saline (PBS) toremove medium and nonadherent cells and incubated in PBS at 22°C for 15 minprior to infection. Cells at passages of between 4 and 16 were used throughoutthis study.
Invasion and survival of salmonellae in J774.2 cells. Survival of salmonellae inmedium not supplemented with IFN-� or supplemented with 10, 100, or 1,000 Uof IFN-� per ml was assessed by the method described previously (10). Inseparate experiments, Salmonella survival was also investigated in J774.2 cellsincubated in medium supplemented with 1,000 U of IFN-� per ml and N-acetyl-L-cysteine captopril (ACC) (10 mM) (Sigma, Poole, United Kingdom), which isan inhibitor of the oxidative burst.
Measurement of oxidative burst. A nitroblue tetrazolium reduction assay (30)was used to detect oxygen metabolite activity in J774.2 cells. Briefly, cells wereincubated with salmonellae (MOI, 10) in 96-well plates (Nunc, Naperville, Ill.)for 60 min at 37°C in CO2 (5%, vol/vol). The wells were washed with PBS andthen incubated with 100 �g of gentamicin per ml for a further 60 min at 37°C. Atvarious time points between 2 and 24 h postinfection, cells were incubated at37°C with 50 �l of nitroblue tetrazolium (Sigma) (1 mg/ml in PBS) for 45 min.To stop the reaction, 100 �l of 1 N hydrochloric acid was added, and the platewas shaken gently at room temperature for 10 min. The wells were then washedwith PBS and shaken again for 10 min at ambient temperature prior to resus-pending the cells in 150 �l of dimethyl sulfoxide and adding 10 �l of 1 N sodiumhydroxide to develop the color. The optical density of the reaction was read in a
plate reader (Anthos Labtech Instruments, Hamburg, Germany) at 620 nm. Testsamples were compared with induction of oxidative burst in J774 monolayers byzymosan (15) and cells that were not incubated with zymosan or bacteria (neg-ative controls). In further experiments, oxidative burst was assessed in infectedcells which had been cocultured in IFN-�.
Construction of IFN-�R�-coated fluorescent microspheres. Early experimentsindicated that IFN-� did not increase ROS activity in J774 cells infected withvirulent salmonellae to the same degree as when cells were infected with aviru-lent mutants over a 24-h period. We therefore wanted to ascertain the effect ofIFN-� on anti-IFN-� receptor alpha (IFN-�R�) expression on the surface ofJ774.2 cells following IFN-� stimulation or following S. enterica serovar Typhi-murium invasion prior to IFN-� stimulation. A quantitative fluorescent micro-sphere assay for IFN-�R� (3) was used in which rabbit anti-mouse IFN-�R�(Autogenbioclear, Calne, Wiltshire, United Kingdom) was attached to Fluores-brite carboxylated microspheres (0.5-�m diameter) (Polyscience, Warrington,Pa.) by a previously reported method (11). Briefly, 0.25 ml of microspheresuspension was centrifuged in a series of buffers (phosphate, carbonate, andborate) for 10 min at 13,000 � g in a bench top microcentrifuge. The micro-spheres were then resuspended in PBS containing carbodiimide (2%, wt/vol)(Sigma) and mixed at room temperature on an end-to-end shaker for 4 h. After4 h, the microspheres were washed three times in PBS and resuspended in 200�g of rabbit anti-mouse IFN-�R� in 1 ml of PBS. The microspheres wereincubated overnight at 4°C on an end-to-end shaker, washed three times in PBS,and resuspended in bovine serum albumin (10 mg/ml; Sigma) for 60 min to blockany remaining free binding sites. After 60 min, the microspheres were washedthree times in PBS and stored in 1 ml of glycerol-sodium azide (Sigma) orresuspended in PBS and used within 48 h. To control for nonspecific microspherebinding via bovine serum albumin, microspheres which were coated only withbovine serum albumin were also constructed. To test whether proteins hadsuccessfully coated the microspheres, the microspheres were reacted with ap-propriate antibodies to analyze agglutination. Autoagglutination was also tested(11).
Quantitative determination of IFN-�R� expression on J774.2 cells. J774.2cells were grown to confluency in 96-well plates containing RPMI 1640 medium.We ascertained that the peak of IFN-�-induced oxidative burst over the 24-hpostinfection period occurred at 12 h. We therefore measured IFN-�R� expres-sion from 2 to 12 h postinfection. Salmonellae were grown and J774.2 cells wereinfected in the presence or absence of IFN-� (1,000 U/ml). After 2, 7, and 12 h,cells were washed in PBS and fixed for 45 min in 3% para-formaldehyde at roomtemperature. After being washed in PBS, monolayers were incubated for 60 minat room temperature in 100 �l of PBS containing 5 �l of anti-IFN-�R�-coatedmicrospheres (equivalent to 1.4 � 104 microspheres per cell) (Polyscience Inc.Biotechnology Product Handbook 1992–1993). After 60 min, the monolayerswere washed three times in PBS, and fluorescent emission was measured at 485and 535 nm in a Victor 1420 multichannel counter (PerkinElmer, Zaventem,Belgium). Fixed cells incubated with bovine serum albumin-coated microsphereswere used as a control for bovine serum albumin used to block uncoupledbinding sites on the surface of IFN-�R�-positive microspheres. Other controlsincluded incubating IFN-�R�-positive microspheres with fixed cells which hadnot been exposed to salmonellae or IFN-�. Autofluorescence was also measuredin uninfected J774.2 cells which had not been exposed to IFN-� or fluorescentmicrospheres. Each test was repeated three times on three occasions, and sta-tistical comparisons were made between the mean fluorescent counts per secondat each time point.
Immunocytochemical analysis of gp91phox activity. The activity of gp91phox ininfected and uninfected J774.2 cells was determined by standard immunocyto-chemical methods. Salmonella-infected cells which had or had not been incu-bated with IFN-� (1,000 U/ml) were washed free of medium and permeabilizedfor 10 min at ambient temperature in Triton X-100 (0.05%) at 2, 7, 12, and 24 hpostinfection. The cells were then fixed for 15 min in paraformaldehyde (5%) atroom temperature and washed in PBS. Fixed cells were then incubated on anend-to-end shaker for 60 min at 4°C in the dark with goat anti-mouse gp91phox
immunoglobulin G (IgG) (Autogenbioclear, London, United Kingdom) diluted1:100 in PBS-Tween. The cells were then washed three times in PBS and incu-bated in identical conditions but with rabbit anti-goat IgG conjugated to fluo-rescein isothiocyanate (Sigma). Propidium iodide was used to stain bacteria.Samples were viewed on a TCS-NT confocal laser scanning microscope equippedwith an argon laser (Leica, Hamburg, Germany). Controls included J774.2 cellswhich had not been infected or incubated with IFN-� and also uninfected cellswhich had been incubated with IFN-� (1,000 U/ml).
Effect of hydrogen peroxide on survival of salmonellae in LB broth. To inves-tigate whether there was a differential reaction to ROS by virulent wild-type andavirulent mutant salmonellae, Salmonella cultures were added to fresh LB con-
4734 FOSTER ET AL. INFECT. IMMUN.
on August 19, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

taining H2O2 at 0.02, 0.04, and 0.06%. Salmonella numbers were estimated asstated earlier at time zero and every hour for 4 h. All treatments were comparedwith an H2O2-free control.
Statistical analysis. Mann-Whitney analysis (Minitab) was used to measuresignificant difference at the 95% confidence limit between different test groupsand between the same test groups at different time points.
RESULTS
Survival of salmonellae in J774.2 monolayers. The numbersof both wild-type S. enterica serovar Typhimurium strains in-creased in J774.2 monolayers over the 24-h period measured,compared with a steady decline or stasis in the number ofviable S. enterica serovar Typhimurium phoP and S. entericaserovar Infantis 1326/28 �r, respectively (Fig. 1A).
When 10 U of IFN-� per ml was added to the culturemedium, the numbers of both virulent S. enterica serovar Ty-phimurium 14028 and F98 exhibited stasis, while a bactericidaleffect was observed for both mutants (Fig. 1C). Further in-creasing the IFN-� concentration (100 or 1,000 U/ml) did notincrease the bactericidal effect on the Salmonella mutants butdid so in both virulent strains (Fig. 1E and 1G). Thus, theoverall effect of increasing the IFN-� concentration on virulentSalmonella populations in J774.2 cells 24 h postinfection was tostimulate survival trends similar to those observed for avirulentmutants, and this was most apparent at high IFN-� concentra-tions.
Induction of ROS in Salmonella-infected J774.2 cells. In-fected J774.2 monolayers were analyzed for oxidative burstwith a nitroblue tetrazolium reduction assay. Significantlyhigher (P � 0.05) levels of ROS activity were measured inJ774.2 cells infected with S. enterica serovar Typhimurium14028 phoP or S. enterica serovar Infantis 1328/28 �r from 12 hpostinfection compared with uninfected control monolayers(Fig. 1B). Small but nonsignificant increases in ROS activitywere detected after infection with virulent S. enterica serovarTyphimurium 14028 or F98 throughout the 24-h postinfectionperiod studied.
When IFN-� (10 U/ml) was added to the culture medium,the mutant strains significantly (P � 0.05) elevated oxidativeburst from 7 h postinfection, and wild-type strains significantly(P � 0.05) increased oxidative burst from 12 h postinfection(Fig. 1D). This trend continued when the infected cells werecocultured with 100 U of IFN-� per ml (Fig. 1F) and 1,000 Uof IFN-� per ml (Fig. 1H), but at these higher IFN-� concen-trations, a significant level of oxidative burst was shifted to anearlier time point (Fig. 2F and H). Further experimentsshowed that significantly higher levels of oxidative burst couldbe achieved at earlier time points by increasing the MOI to100, without IFN-�, and when both an MOI of 100 and anIFN-� concentration of 1,000 U/ml were investigated, signifi-cant induction of oxidative burst could be achieved at 2 hpostinfection (data not shown).
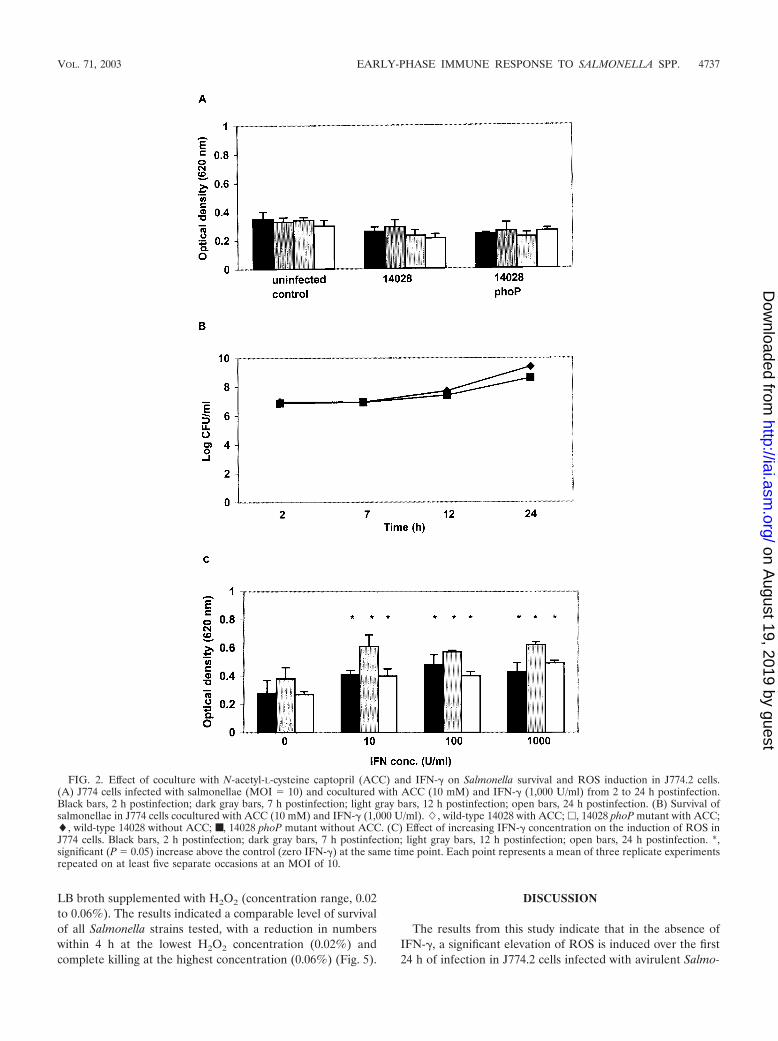
Effect of ACC on ROS induction and Salmonella survival inJ774.2 cells. Throughout the infection period, addition of N-acetyl-L-cysteine captopril (ACC) to the culture medium in-hibited ROS activity to levels recorded in uninfected controls,even in the presence of IFN-� (1,000 U/ml) (Fig. 2A). Inhibi-tion of ROS in these cells coincided with recovery of signifi-cantly (P � 0.05) higher numbers of S. enterica serovar Typhi-murium 14028 and its phoP mutant (Fig. 2B).
Effect of IFN-� on oxidative burst in uninfected J774.2 cells.When uninfected J774.2 cells were cultured with IFN-�, ROSactivity increased significantly (P � 0.05) in comparison toROS activity measured in uninfected control cells which hadnot been incubated with IFN-� (Fig. 2C). However, the effectof IFN-� on ROS was not dose dependent, since concentra-tions of IFN-� ranging from 10 to 1,000 U/ml did not have anincreased effect (Fig. 2C). The length of time that cells wereexposed to IFN-�, however, did have an effect, with 12 h beingthe optimal time of exposure for greatest ROS activity (Fig. 2C).
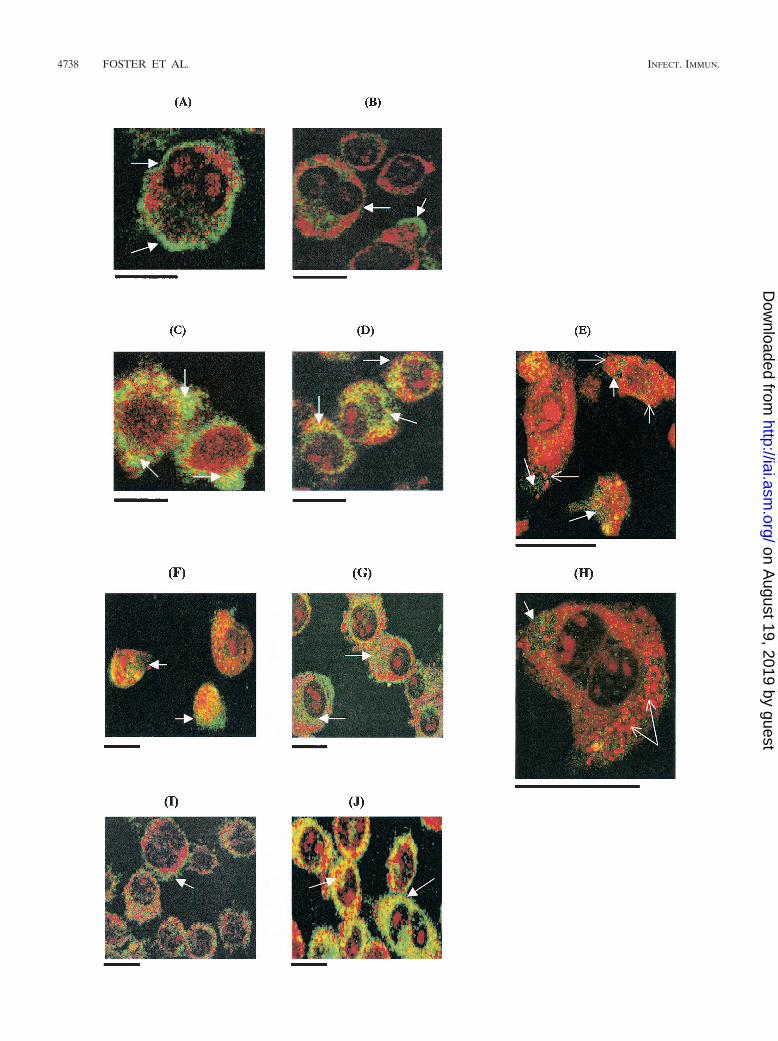
Immunocytochemical detection of gp91phox in Salmonella-infected J774.2 cells. Confocal laser scanning microscopy wasused to compare gp91phox activity in J774.2 cells infected withvirulent S. enterica serovar Typhimurium 14028 or avirulent S.enterica serovar Typhimurium phoP. Cells were infected withsalmonellae without the addition of IFN-� to the culture me-dium or in the presence of IFN-� (1,000 U/ml). Uninfectedcells exhibited cell membrane localization of gp91phox after 2and 12 h in culture, although slight cytoplasmic activity wasnoted after 12 h (Fig. 3A and B). Cells infected with S. entericaserovar Typhimurium 14028 exhibited little cytoplasmic local-ization of gp91phox even after 12 h of infection (Fig. 3C), butgp91phox cytoplasmic activity during infection was greatly en-hanced by the addition of IFN-� to the culture medium for12 h (Fig. 3D). Cells infected with S. enterica serovar Typhi-murium 14028 phoP, however, began to show strong cytoplas-mic gp91phox activity at 12 h postinfection without addition ofIFN-� to the culture medium (Fig. 3F), while the addition ofIFN-� to the medium increased the intensity of the responseeven further (Fig. 3G). Further analysis, below the cell surfaceand at higher power, indicated that gp91phox activity waspresent in cytoplasmic compartments but was not strictly lo-calized to areas which were infected by the salmonellae (Fig.3E and H). The effect of IFN-� (1,000 U/ml) alone on gp91phox
activity in J774.2 cells was also investigated and showed thatcytoplasmic localization was present after 2 h in culture withIFN-� (Fig. 3I), and this was further enhanced by 12 h (Fig.3J).
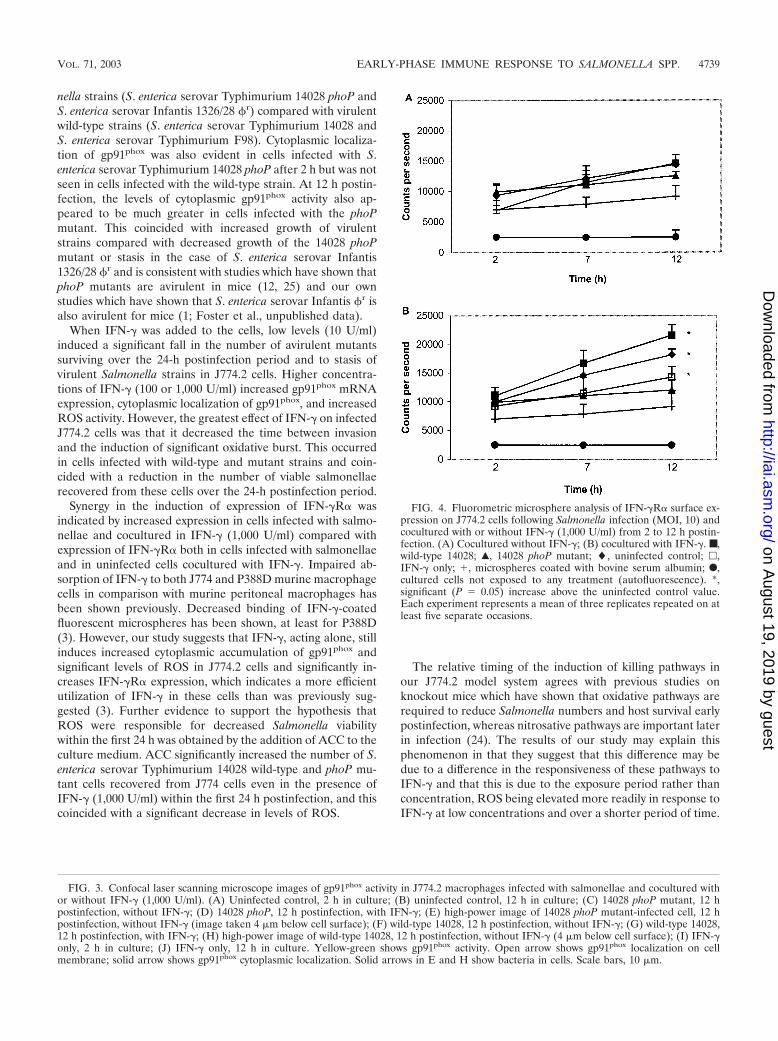
IFN-�R� expression on Salmonella-infected J774.2 cells.Fluorometry was used to measure changes in surface expres-sion of IFN-�R� following infection of J774.2 cells with S.enterica serovar Typhimurium 14028 or its phoP mutant, withor without the addition of IFN-� (1,000 U/ml) to the culturemedium. In cells infected with the wild type or phoP mutant,IFN-�R� expression was not significantly (P 0.05) raisedabove levels detected on the surface of uninfected cells, al-though expression of IFN-�R� on the surface of uninfectedcells was actually raised in culture during the 12-h periodinvestigated (Fig. 4A). The addition of IFN-� (1,000 U/ml) tothe culture medium significantly (P � 0.05) elevated IFN-�R�expression on the surface of cells infected with either wild-type14028 or the phoP mutant after 7 h, with the greatest increasemeasured on the surface of cells infected with wild-type 14028(Fig. 4B).
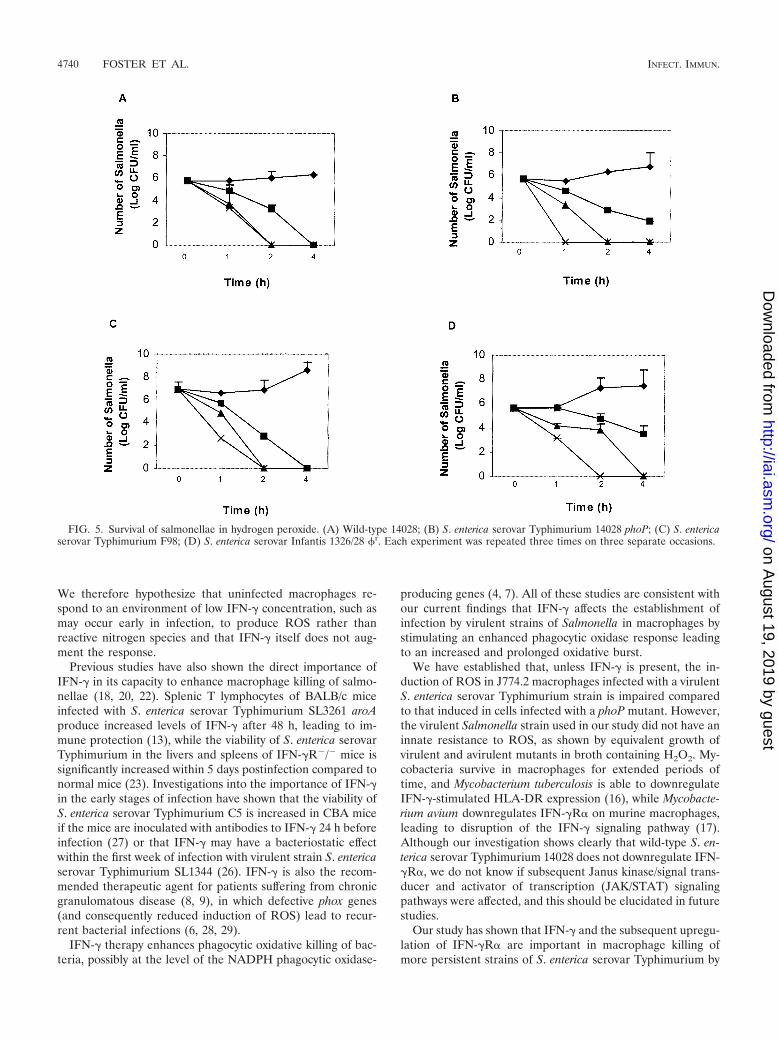
Effect of hydrogen peroxide on Salmonella survival in LBbroth. To examine whether or not some of the Salmonellastrains used in this study have a natural resistance or suscep-tibility to ROS, we grew virulent S. enterica serovar Typhi-murium 14028 and F98 and avirulent S. enterica serovar Ty-phimurium 14028 phoP and S. enterica serovar Infantis �r in
VOL. 71, 2003 EARLY-PHASE IMMUNE RESPONSE TO SALMONELLA SPP. 4735
on August 19, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
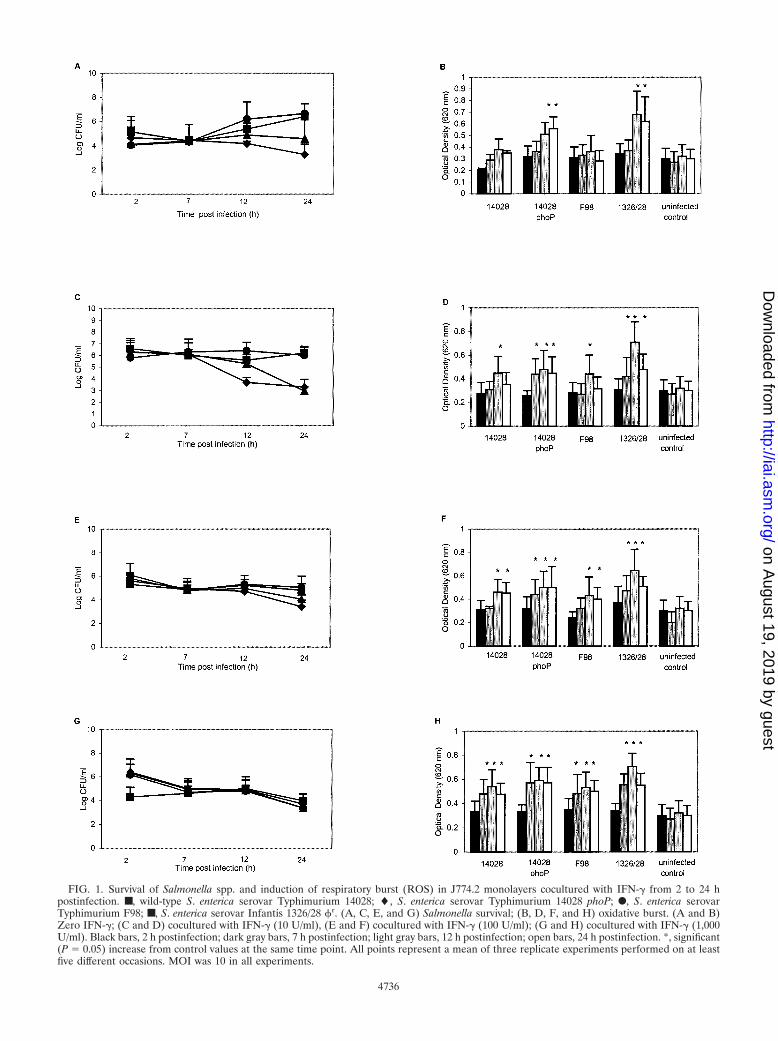
FIG. 1. Survival of Salmonella spp. and induction of respiratory burst (ROS) in J774.2 monolayers cocultured with IFN-� from 2 to 24 hpostinfection. ■, wild-type S. enterica serovar Typhimurium 14028; �, S. enterica serovar Typhimurium 14028 phoP; F, S. enterica serovarTyphimurium F98; ■, S. enterica serovar Infantis 1326/28 �r. (A, C, E, and G) Salmonella survival; (B, D, F, and H) oxidative burst. (A and B)Zero IFN-�; (C and D) cocultured with IFN-� (10 U/ml), (E and F) cocultured with IFN-� (100 U/ml); (G and H) cocultured with IFN-� (1,000U/ml). Black bars, 2 h postinfection; dark gray bars, 7 h postinfection; light gray bars, 12 h postinfection; open bars, 24 h postinfection. *, significant(P 0.05) increase from control values at the same time point. All points represent a mean of three replicate experiments performed on at leastfive different occasions. MOI was 10 in all experiments.
4736
on August 19, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
LB broth supplemented with H2O2 (concentration range, 0.02to 0.06%). The results indicated a comparable level of survivalof all Salmonella strains tested, with a reduction in numberswithin 4 h at the lowest H2O2 concentration (0.02%) andcomplete killing at the highest concentration (0.06%) (Fig. 5).
DISCUSSION
The results from this study indicate that in the absence ofIFN-�, a significant elevation of ROS is induced over the first24 h of infection in J774.2 cells infected with avirulent Salmo-
FIG. 2. Effect of coculture with N-acetyl-L-cysteine captopril (ACC) and IFN-� on Salmonella survival and ROS induction in J774.2 cells.(A) J774 cells infected with salmonellae (MOI 10) and cocultured with ACC (10 mM) and IFN-� (1,000 U/ml) from 2 to 24 h postinfection.Black bars, 2 h postinfection; dark gray bars, 7 h postinfection; light gray bars, 12 h postinfection; open bars, 24 h postinfection. (B) Survival ofsalmonellae in J774 cells cocultured with ACC (10 mM) and IFN-� (1,000 U/ml). �, wild-type 14028 with ACC; �, 14028 phoP mutant with ACC;�, wild-type 14028 without ACC; ■, 14028 phoP mutant without ACC. (C) Effect of increasing IFN-� concentration on the induction of ROS inJ774 cells. Black bars, 2 h postinfection; dark gray bars, 7 h postinfection; light gray bars, 12 h postinfection; open bars, 24 h postinfection. *,significant (P 0.05) increase above the control (zero IFN-�) at the same time point. Each point represents a mean of three replicate experimentsrepeated on at least five separate occasions at an MOI of 10.
VOL. 71, 2003 EARLY-PHASE IMMUNE RESPONSE TO SALMONELLA SPP. 4737
on August 19, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
4738 FOSTER ET AL. INFECT. IMMUN.
on August 19, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
nella strains (S. enterica serovar Typhimurium 14028 phoP andS. enterica serovar Infantis 1326/28 �r) compared with virulentwild-type strains (S. enterica serovar Typhimurium 14028 andS. enterica serovar Typhimurium F98). Cytoplasmic localiza-tion of gp91phox was also evident in cells infected with S.enterica serovar Typhimurium 14028 phoP after 2 h but was notseen in cells infected with the wild-type strain. At 12 h postin-fection, the levels of cytoplasmic gp91phox activity also ap-peared to be much greater in cells infected with the phoPmutant. This coincided with increased growth of virulentstrains compared with decreased growth of the 14028 phoPmutant or stasis in the case of S. enterica serovar Infantis1326/28 �r and is consistent with studies which have shown thatphoP mutants are avirulent in mice (12, 25) and our ownstudies which have shown that S. enterica serovar Infantis �r isalso avirulent for mice (1; Foster et al., unpublished data).
When IFN-� was added to the cells, low levels (10 U/ml)induced a significant fall in the number of avirulent mutantssurviving over the 24-h postinfection period and to stasis ofvirulent Salmonella strains in J774.2 cells. Higher concentra-tions of IFN-� (100 or 1,000 U/ml) increased gp91phox mRNAexpression, cytoplasmic localization of gp91phox, and increasedROS activity. However, the greatest effect of IFN-� on infectedJ774.2 cells was that it decreased the time between invasionand the induction of significant oxidative burst. This occurredin cells infected with wild-type and mutant strains and coin-cided with a reduction in the number of viable salmonellaerecovered from these cells over the 24-h postinfection period.
Synergy in the induction of expression of IFN-�R� wasindicated by increased expression in cells infected with salmo-nellae and cocultured in IFN-� (1,000 U/ml) compared withexpression of IFN-�R� both in cells infected with salmonellaeand in uninfected cells cocultured with IFN-�. Impaired ab-sorption of IFN-� to both J774 and P388D murine macrophagecells in comparison with murine peritoneal macrophages hasbeen shown previously. Decreased binding of IFN-�-coatedfluorescent microspheres has been shown, at least for P388D(3). However, our study suggests that IFN-�, acting alone, stillinduces increased cytoplasmic accumulation of gp91phox andsignificant levels of ROS in J774.2 cells and significantly in-creases IFN-�R� expression, which indicates a more efficientutilization of IFN-� in these cells than was previously sug-gested (3). Further evidence to support the hypothesis thatROS were responsible for decreased Salmonella viabilitywithin the first 24 h was obtained by the addition of ACC to theculture medium. ACC significantly increased the number of S.enterica serovar Typhimurium 14028 wild-type and phoP mu-tant cells recovered from J774 cells even in the presence ofIFN-� (1,000 U/ml) within the first 24 h postinfection, and thiscoincided with a significant decrease in levels of ROS.
The relative timing of the induction of killing pathways inour J774.2 model system agrees with previous studies onknockout mice which have shown that oxidative pathways arerequired to reduce Salmonella numbers and host survival earlypostinfection, whereas nitrosative pathways are important laterin infection (24). The results of our study may explain thisphenomenon in that they suggest that this difference may bedue to a difference in the responsiveness of these pathways toIFN-� and that this is due to the exposure period rather thanconcentration, ROS being elevated more readily in response toIFN-� at low concentrations and over a shorter period of time.
FIG. 4. Fluorometric microsphere analysis of IFN-�R� surface ex-pression on J774.2 cells following Salmonella infection (MOI, 10) andcocultured with or without IFN-� (1,000 U/ml) from 2 to 12 h postin-fection. (A) Cocultured without IFN-�; (B) cocultured with IFN-�. ■,wild-type 14028; Œ, 14028 phoP mutant; �, uninfected control; �,IFN-� only; �, microspheres coated with bovine serum albumin; F,cultured cells not exposed to any treatment (autofluorescence). *,significant (P 0.05) increase above the uninfected control value.Each experiment represents a mean of three replicates repeated on atleast five separate occasions.
FIG. 3. Confocal laser scanning microscope images of gp91phox activity in J774.2 macrophages infected with salmonellae and cocultured withor without IFN-� (1,000 U/ml). (A) Uninfected control, 2 h in culture; (B) uninfected control, 12 h in culture; (C) 14028 phoP mutant, 12 hpostinfection, without IFN-�; (D) 14028 phoP, 12 h postinfection, with IFN-�; (E) high-power image of 14028 phoP mutant-infected cell, 12 hpostinfection, without IFN-� (image taken 4 �m below cell surface); (F) wild-type 14028, 12 h postinfection, without IFN-�; (G) wild-type 14028,12 h postinfection, with IFN-�; (H) high-power image of wild-type 14028, 12 h postinfection, without IFN-� (4 �m below cell surface); (I) IFN-�only, 2 h in culture; (J) IFN-� only, 12 h in culture. Yellow-green shows gp91phox activity. Open arrow shows gp91phox localization on cellmembrane; solid arrow shows gp91phox cytoplasmic localization. Solid arrows in E and H show bacteria in cells. Scale bars, 10 �m.
VOL. 71, 2003 EARLY-PHASE IMMUNE RESPONSE TO SALMONELLA SPP. 4739
on August 19, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
We therefore hypothesize that uninfected macrophages re-spond to an environment of low IFN-� concentration, such asmay occur early in infection, to produce ROS rather thanreactive nitrogen species and that IFN-� itself does not aug-ment the response.
Previous studies have also shown the direct importance ofIFN-� in its capacity to enhance macrophage killing of salmo-nellae (18, 20, 22). Splenic T lymphocytes of BALB/c miceinfected with S. enterica serovar Typhimurium SL3261 aroAproduce increased levels of IFN-� after 48 h, leading to im-mune protection (13), while the viability of S. enterica serovarTyphimurium in the livers and spleens of IFN-�R�/� mice issignificantly increased within 5 days postinfection compared tonormal mice (23). Investigations into the importance of IFN-�in the early stages of infection have shown that the viability ofS. enterica serovar Typhimurium C5 is increased in CBA miceif the mice are inoculated with antibodies to IFN-� 24 h beforeinfection (27) or that IFN-� may have a bacteriostatic effectwithin the first week of infection with virulent strain S. entericaserovar Typhimurium SL1344 (26). IFN-� is also the recom-mended therapeutic agent for patients suffering from chronicgranulomatous disease (8, 9), in which defective phox genes(and consequently reduced induction of ROS) lead to recur-rent bacterial infections (6, 28, 29).
IFN-� therapy enhances phagocytic oxidative killing of bac-teria, possibly at the level of the NADPH phagocytic oxidase-
producing genes (4, 7). All of these studies are consistent withour current findings that IFN-� affects the establishment ofinfection by virulent strains of Salmonella in macrophages bystimulating an enhanced phagocytic oxidase response leadingto an increased and prolonged oxidative burst.
We have established that, unless IFN-� is present, the in-duction of ROS in J774.2 macrophages infected with a virulentS. enterica serovar Typhimurium strain is impaired comparedto that induced in cells infected with a phoP mutant. However,the virulent Salmonella strain used in our study did not have aninnate resistance to ROS, as shown by equivalent growth ofvirulent and avirulent mutants in broth containing H2O2. My-cobacteria survive in macrophages for extended periods oftime, and Mycobacterium tuberculosis is able to downregulateIFN-�-stimulated HLA-DR expression (16), while Mycobacte-rium avium downregulates IFN-�R� on murine macrophages,leading to disruption of the IFN-� signaling pathway (17).Although our investigation shows clearly that wild-type S. en-terica serovar Typhimurium 14028 does not downregulate IFN-�R�, we do not know if subsequent Janus kinase/signal trans-ducer and activator of transcription (JAK/STAT) signalingpathways were affected, and this should be elucidated in futurestudies.
Our study has shown that IFN-� and the subsequent upregu-lation of IFN-�R� are important in macrophage killing ofmore persistent strains of S. enterica serovar Typhimurium by
FIG. 5. Survival of salmonellae in hydrogen peroxide. (A) Wild-type 14028; (B) S. enterica serovar Typhimurium 14028 phoP; (C) S. entericaserovar Typhimurium F98; (D) S. enterica serovar Infantis 1326/28 �r. Each experiment was repeated three times on three separate occasions.
4740 FOSTER ET AL. INFECT. IMMUN.
on August 19, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

stimulating an earlier and more prolonged increase in ROS.This occurs via its cytoplasmic localization, but regulation atthe level of transcription may also be involved. However, thisrequirement for IFN-� may allow dissemination of the infec-tion in mice until adequate stimulation of macrophages byIFN-�-producing cells can be achieved. In comparison, aviru-lent strains, which induce high levels of ROS via gp91phox inJ774.2 cells without the associated IFN-�R� upregulation orthe need for IFN-�, may be killed in macrophages before theyare allowed to disseminate.
ACKNOWLEDGMENTS
We thank Elaine Bennett for assistance with cell culture.This work was supported by a European Union grant (Fair 98-4006)
and the Department of Food, the Environment and Rural Affairs,United Kingdom.
REFERENCES
1. Barrow, P. A., M. B. Huggins, and M. A. Lovell. 1994. Host specificity ofSalmonella infection in chickens and mice is expressed in vivo primarily atthe level of the reticuloendothelial system. Infect. Immun. 62:4602–4610.
2. Buchmeier, N. A., C. J. Lipps, Y. Magdalene, Y. So, and F. Heffron. 1993.Recombinant deficient mutants of Salmonella are avirulent and sensitive tooxidative burst. Mol. Microbiol. 7:933–936.
3. Cellada, A., P. W. Gray, E. Rinderknecht, and R. D. Shreiber. 1984. Evi-dence for a gamma-receptor that regulates macrophage tumoricidal activity.J. Exp. Med. 160:55–74.
4. Condino-Neto, A., and P. E. Newburger. 2000. Interferon-gamma improvessplicing efficiency of CYBB gene transcripts in an interferon responsivevariant of chronic granulomatous disease due to splice site consensus regionmutation. Blood 95:3548–3554.
5. De Groote, M. A., U. A. Ochsner, M. U. Shiloh, C. Nathan, J. M. McCord,M. C. Dinauer, S. J. Libby, A. Vasquez-Torres, Y. Xu, and F. C. Fang. 1997.Periplasmic superoxide dismutase protects Salmonella from products ofphagocyte NADPH-oxidase and nitric oxide synthase. Proc. Natl. Acad. Sci.USA 94:13997–14001.
6. Dinauer, M. C., L. L. Ling, H. Bjorgvinsdottir, C. Ding, and N. Pech. 1999.Long term correction of phagocyte NADPH oxidase activity by retroviral-mediated gene transfer in murine X-linked chronic granulomatous disease.Blood 94:914–922.
7. Eklund, E. L., W. Luo, and D. G. Skalnik. 1996. Characterization of threepromoter elements and cognate DNA binding protein(s) necessary forIFN-� induction of gp91-phox transcription. J. Immunol. 157:2418–2429.
8. Ezekowitz, R. A., M. C. Dinauer, H. S. Jaffe, H. Orkin, and P. E. Newburger.1988. Partial correction of the phagocyte defect in patients with chronicgranulomatous disease by subcutaneous interferon gamma. N. Engl. J. Med.319:146–151.
9. Ezekowitz, R. A., C. A. Sieff, M. C. Dinauer, C. Nathan, S. H. Orkin, andP. E. Newburger. 1990. Restoration of phagocyte function by interferon-gamma in X-linked chronic granulomatous disease occurs at the level of theprogenitor cell. Blood 76:2443–2448.
10. Fields, P. I., R. V. Swanson, C. G. Haidaris, and F. Heffron. 1986. Mutantsof Salmonella typhimurium that cannot survive within the macrophage areavirulent. Proc. Natl. Acad. Sci. USA 83:5189–5193.
11. Foster, N., M. A. Clark, M. A. Jepson, and B. H. Hirst. 1998. Ulex europaeus1 lectin targets microspheres to mouse Peyer’s patch M cells in vivo. Vaccine16:536–541.
12. Groisman, E. A., and M. H. Saier. 1990. Salmonella virulence: new clues tointramacrophage survival. Trends Biochem. Sci. 15:30–33.
13. Harrison, J. A., B. Villarreal-Ramos, P. Mastroeni, R. Demarco De Hor-maeche, and C. E. Hormaeche. 1997. Correlates of protection induced by live
Aro� Salmonella typhimurium vaccines in the murine typhoid model. Immu-nology 90:618–625.
14. Held, T. K., W. Xiao W. Yuan, D. V. Kavakolanu, and A. S. Cross. 1999.Gamma interferon augments macrophage activation by lipopolysaccharideby two distinct mechanisms, at the signal transduction level via an autocrinemechanism involving tumor necrosis factor alpha and interleukin-1. Infect.Immun. 67:206–212.
15. Heremans, H., J. Van Damme, C. Dillon, R. Dijkmans, and A. Billau. 1990.Interferon gamma, a mediator of lethal Schwartzman-like shock reaction inmice. J. Exp. Med. 171:1853–1869.
16. Hmama, Z., R. Gabathuler, W. A. Jefferies, G. De Jong, and N. E. Reiner.1998. Attenuation of HLA-DR expression by mononuclear phagocytes in-fected with Mycobacterium tuberculosis is related to intracellular sequestra-tion of immiature class II heterodimers. J. Immunol. 161:4882–4893.
17. Hussain, S., B. S. Zwilling, and W. P. Lafuse. 1999. Mycobacterium aviuminfection of mouse macrophages inhibits IFN-� Janus kinase-STAT signal-ling and gene induction by downregulation of the IFN-� receptor. J. Immu-nol. 163:2041–2048.
18. Kagaya, K., K. Watnabe, and J. Fukazawa. 1989. Capacity of recombinantgamma interferon to activate macrophages for Salmonella-killing activity.Infect. Immun. 57:609–615.
19. Kaufman, S. H. E. 1993. Immunity to intracellular bacteria. Annu. Rev.Immunol. 11:129–163.
20. Kincy-Cain, T., J. D. Clements, and L. Bost. 1996. Endogenous and exoge-nous interleukin-12 augment protective immune response in mice orallychallenged with Salmonella dublin. Infect. Immun. 64:1437–1440.
21. Leusen, J. H. W., M. deBoer, B. G. J. M. Bolscher, P. M. Hilaries, R. S.Weening, H. J. Ochs, D. Roos, and A. J. Verhoeven. 1994. A point mutationin gp91phox of cytochrome b558 of human NADPH oxidase leading to defec-tive translocation of the cytosolic proteins p47-phox and p67-phox. J. Clin.Investig. 93:2120–2126.
22. Mastroeni, P., J. A. Harrison, J. A. Chabalgoity, and C. E. Hormaeche. 1996.Effect of interleukin-12 neutralization on host resistance and gamma inter-feron production in mouse typhoid. Infect. Immun. 64:189–196.
23. Mastroeni, P., S. Clare, S. Khan, J. E. Harrison, C. E. Hormaeche, H.Okamura, M. Kurimoto, and G. Dougan. 1999. Interleukin-18 contributes tohost resistance and gamma interferon production in mice infected withvirulent Salmonella typhimurium. Infect. Immun. 67:478–483.
24. Mastroeni, P., A. Vasquez-Torres, F. C. Fang, Y. Yisheng, S. Khan, C. E.Hormaeche, and G. Dougan. 2000. Antimicrobial actions of NADPH phago-cyte oxidase and inducible nitric oxide synthase in experimental salmonello-sis. II. Effects on microbial proliferation and host survival. J. Exp. Med.192:237–247.
25. Miller, S. I., and J. J. Mekalanos. 1990. Constitutive expression of the phoPregulon attenuates Salmonella virulence and survival within macrophages. J.Bacteriol. 172:2485–2490.
26. Muotiala, A., and P. H. Makela. 1993. Role of gamma interferon in latestages of murine salmonellosis. Infect. Immunol. 61:4248–4252.
27. Nauciel, C., and F. Espinasse-Maes. 1992. Role of gamma interferon andtumor necrosis factor alpha in resistance to Salmonella typhimurium infec-tion. Infect. Immun. 60:450–454.
28. Rae, J., P. E. Newburger, M. C. Dinauern, D. Noack, P. J. Hopkins, R.Kuruto, and J. T. Curnutte. 1998. X-linked chronic granulomatous disease:mutations in the CYBB gene encoding the gp91-phox component of therespiratory burst oxidase. Am. J. Hum. Genet. 62:1320–1331.
29. Roos, D., M. de Boer, F. Kuribayashi, C. Meischel, R. Weening, A. Segal, A.Ahlin, K. Nemet, J. Hossle, E. Bernatowska-Matuszkiewicz, and H. Middle-ton-Price. 1996. Mutations in the x-linked and autosomal recessive forms ofchronic granulomatous disease. Blood 87:1663.
30. Segal, A. W. 1974. Nitroblue-tetrazolium tests. Lancet ii:1248–1252.31. Segal, A. W., and K. P. Shatwell. 1997. The NADPH oxidase of phagocytic
leucocytes. Ann. N.Y. Acad. Sci. 832:215–222.32. Vasquez-Torres, A., J. Jones-Carson, P. Mastroeni, H. Ischiropoulous, and
F. C. Fang. 2000. Antimicrobial actions of the NADPH phagocyte oxidaseand inducible nitric oxide synthase in experimental salmonellosis. I. Effectson microbial killing by activated peritoneal macrophages in vitro. J. Exp.Med. 192:227–236.
Editor: B. B. Finlay
VOL. 71, 2003 EARLY-PHASE IMMUNE RESPONSE TO SALMONELLA SPP. 4741
on August 19, 2019 by guest
http://iai.asm.org/
Dow
nloaded from


















