
In vivo modulation of LPS-induced alterations in brain and peripheral cytokines and HPA axis...
11
In vivo modulation of LPS-induced alterations in brain and peripheral cytokines and HPA axis activity by cannabinoids Michelle Roche a , Michael Diamond b , John P. Kelly b , David P. Finn a, ⁎ a Department of Physiology, National University of Ireland, Galway, University Road, Galway, Ireland b Department of Pharmacology, National University of Ireland, Galway, University Road, Galway, Ireland Received 18 May 2006; received in revised form 2 August 2006; accepted 2 August 2006 Abstract This study investigated cannabinoid receptor-mediated regulation of brain and peripheral cytokines in vivo. The cannabinoid receptor agonist, HU210 attenuated lipopolysaccharide (LPS)-induced increases in IL-1β and TNFα in rat brain and IL-1β, TNFα, IL-6 and IFNγ in plasma. The CB 1 receptor antagonist, SR141716A, attenuated the immunosupressive effects of HU210 on IL-1β, but not TNFα. SR141716A or the CB 2 receptor antagonist, SR144528, alone attenuated LPS-induced cytokine increases. LPS and/or cannabinoids also reduced circulating lymphocyte numbers and increased corticosterone levels. These data provide evidence for modulation of pro-inflammatory cytokines in vivo by cannabinoid receptors and inform the development of cannabinoids for neuroinflammatory disorders. © 2006 Elsevier B.V. All rights reserved. Keywords: HU210; Corticosterone; SR141716A; SR144528; IL-1β; TNFα 1. Introduction Cannabinoids such as Δ 9 -tetrahydrocannabinol (Δ 9 - THC), the main psychoactive constituent of marijuana, and related analogues, modulate a range of physiological processes including appetite, memory, mood, cardiovascular system, pain and immune function (Di Marzo et al., 2004; Finn and Chapman, 2004; Grotenhermen, 2005; Klein, 2005). As a consequence, examination of the mechanisms underlying the action of cannabinoids is both of fundamental physiological importance and offers potential for the development of new therapeutic agents. Many plant-derived, endogenous or synthetic cannabinoids elicit their biological effects via two known cannabinoid receptor subtypes, CB 1 and CB 2 (Matsuda et al., 1990; Munro et al., 1993). A number of endogenous ligands (endocannabinoids) for these receptors have been discovered including the compounds anandamide and 2-arachidonoyl glycerol (Mechoulam et al., 1998). The CB 1 receptor, located extensively throughout the central nervous system (Herkenham et al., 1990), mediates many of the behavioural effects of cannabinoids. CB 1 receptors (Bouaboula et al., 1993; Galiegue et al., 1995) and CB 2 receptors (Bouaboula et al., 1993; Munro et al., 1993) are expressed on cells and tissues of the immune system and recent evidence indicates that both receptors are present on brain glial cells (Waksman et al., 1999; Walter et al., 2003). The endocannabinoid system may play a significant role in modulating the physiology of the immune system as well as representing an important therapeutic target for a number of peripheral and central inflammatory disorders (Di Marzo et al., 2004; Klein, 2005). Cytokines including tumor necrosis factor α (TNFα), interleukin (IL)-1β, IL-6, IL-10 and interferon-γ (IFNγ) play crucial roles in mediating inflammatory and immune responses and facilitate commu- nication between immune, nervous and endocrine systems (Watkins et al., 1995). Interactions between these systems are important for maintaining homeostasis. For example, in- creased release of pro-inflammatory cytokines in response to immune activation results in activation of the hypothalamic– pituitary–adrenal (HPA) axis and increased glucocorticoid Journal of Neuroimmunology 181 (2006) 57 – 67 www.elsevier.com/locate/jneuroim ⁎ Corresponding author. Tel.: +353 91 492141; fax: +353 91 494544. E-mail address: [email protected] (D.P. Finn). 0165-5728/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.jneuroim.2006.08.001
-
Upload
michelle-roche -
Category
Documents
-
view
212 -
download
0
Transcript of In vivo modulation of LPS-induced alterations in brain and peripheral cytokines and HPA axis...

gy 181 (2006) 57–67www.elsevier.com/locate/jneuroim
Journal of Neuroimmunolo
In vivo modulation of LPS-induced alterations in brain and peripheralcytokines and HPA axis activity by cannabinoids
Michelle Roche a, Michael Diamond b, John P. Kelly b, David P. Finn a,⁎
a Department of Physiology, National University of Ireland, Galway, University Road, Galway, Irelandb Department of Pharmacology, National University of Ireland, Galway, University Road, Galway, Ireland
Received 18 May 2006; received in revised form 2 August 2006; accepted 2 August 2006
Abstract
This study investigated cannabinoid receptor-mediated regulation of brain and peripheral cytokines in vivo. The cannabinoid receptoragonist, HU210 attenuated lipopolysaccharide (LPS)-induced increases in IL-1β and TNFα in rat brain and IL-1β, TNFα, IL-6 and IFNγ inplasma. The CB1 receptor antagonist, SR141716A, attenuated the immunosupressive effects of HU210 on IL-1β, but not TNFα. SR141716Aor the CB2 receptor antagonist, SR144528, alone attenuated LPS-induced cytokine increases. LPS and/or cannabinoids also reducedcirculating lymphocyte numbers and increased corticosterone levels. These data provide evidence for modulation of pro-inflammatorycytokines in vivo by cannabinoid receptors and inform the development of cannabinoids for neuroinflammatory disorders.© 2006 Elsevier B.V. All rights reserved.
Keywords: HU210; Corticosterone; SR141716A; SR144528; IL-1β; TNFα
1. Introduction
Cannabinoids such as Δ9-tetrahydrocannabinol (Δ9-THC), the main psychoactive constituent of marijuana, andrelated analogues, modulate a range of physiologicalprocesses including appetite, memory, mood, cardiovascularsystem, pain and immune function (Di Marzo et al., 2004;Finn and Chapman, 2004; Grotenhermen, 2005; Klein,2005). As a consequence, examination of the mechanismsunderlying the action of cannabinoids is both of fundamentalphysiological importance and offers potential for thedevelopment of new therapeutic agents. Many plant-derived,endogenous or synthetic cannabinoids elicit their biologicaleffects via two known cannabinoid receptor subtypes, CB1
and CB2 (Matsuda et al., 1990; Munro et al., 1993). Anumber of endogenous ligands (endocannabinoids) for thesereceptors have been discovered including the compoundsanandamide and 2-arachidonoyl glycerol (Mechoulam et al.,
⁎ Corresponding author. Tel.: +353 91 492141; fax: +353 91 494544.E-mail address: [email protected] (D.P. Finn).
0165-5728/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.jneuroim.2006.08.001
1998). The CB1 receptor, located extensively throughout thecentral nervous system (Herkenham et al., 1990), mediatesmany of the behavioural effects of cannabinoids. CB1
receptors (Bouaboula et al., 1993; Galiegue et al., 1995) andCB2 receptors (Bouaboula et al., 1993; Munro et al., 1993)are expressed on cells and tissues of the immune system andrecent evidence indicates that both receptors are present onbrain glial cells (Waksman et al., 1999; Walter et al., 2003).
The endocannabinoid system may play a significant rolein modulating the physiology of the immune system as wellas representing an important therapeutic target for a numberof peripheral and central inflammatory disorders (Di Marzoet al., 2004; Klein, 2005). Cytokines including tumornecrosis factor α (TNFα), interleukin (IL)-1β, IL-6, IL-10and interferon-γ (IFNγ) play crucial roles in mediatinginflammatory and immune responses and facilitate commu-nication between immune, nervous and endocrine systems(Watkins et al., 1995). Interactions between these systems areimportant for maintaining homeostasis. For example, in-creased release of pro-inflammatory cytokines in response toimmune activation results in activation of the hypothalamic–pituitary–adrenal (HPA) axis and increased glucocorticoid

58 M. Roche et al. / Journal of Neuroimmunology 181 (2006) 57–67
release, inhibiting further cytokine release and protectingagainst endotoxemic shock (Beishuizen and Thijs, 2003).There is substantial evidence from in vitro studies thatcannabinoid receptor agonists exert anti-inflammatory effectsby inhibiting pro-inflammatory cytokine release (Puffenbar-ger et al., 2000; Chang et al., 2001; Facchinetti et al., 2003;Klegeris et al., 2003; Kaplan et al., 2003), increasing anti-inflammatory cytokine release (Molina-Holgado et al.,2003), inhibiting mast cell and granulocyte activation(Mazzari et al., 1996) and lymphocyte proliferation (Schwarzet al., 1994). In vivo studies, both pre-clinical (Gallily et al.,1997; Smith et al., 2000; Panikashvili et al., 2001) andclinical (Roth et al., 2002), also demonstrate the immuno-suppressive effects of cannabinoid receptor activation. Theseimmunomodulatory effects of cannabinoid receptors supportthe large body of evidence implicating a role for the endo-cannabinoid system in neuroprotection (Mechoulam, 2002;Jackson et al., 2005) and its potential as a therapeutic target inneuroinflammatory disease such as multiple sclerosis(Mechoulam et al., 2002; Walter and Stella, 2004). To date,however, studies on the effects of cannabinoids on immunefunction have focused almost entirely on peripheral immuneresponses and there is a paucity of in vivo studies inves-tigating the effects of cannabinoids on brain cytokines.
The present study investigated the effects of the potent,synthetic cannabinoid receptor agonist and Δ9-tetrahydro-cannabinol (THC) analogue, HU210, on cytokine levels indiscrete rat brain regions and in plasma, following the in vivoadministration of the bacterial endotoxin and immunogen,lipopolysaccaride (LPS). LPS induces the release of cy-tokines, eicosanoids and glucocorticoids and is a widelyemployed tool for the investigation of neuro-immune-endocrine responses and interactions. In addition, LPS hasbeen shown to induce the activity of a number of enzymesincluding inducible nitric oxide synthase (iNOS) andcyclooxgenases (Chang et al., 2001) and down-regulate theexpression of the endocannabinoid metabolising enzyme,fatty acid amide hydroylase (FAAH) (Maccarrone et al.,2001). Effects of HU210 on LPS-induced alterations incirculating lymphocyte numbers and plasma corticosteronelevels were also assessed as further indices of its effects onthe immune and endocrine systems respectively. The se-lective CB1 and CB2 receptor antagonists, SR141716A andSR144528 respectively, were used to identify the receptorsubtype through which HU210 mediates its effects. More-over, administration of these receptor antagonists aloneenabled investigation of putative endocannabinoid tone onLPS-induced cytokine and glucocorticoid release and lym-phocyte number.
2. Materials and methods
2.1. Animals
Experiments were carried out on male Sprague Dawleyrats (weight 220–260 g; Harlan, UK), housed singly in
plastic bottomed cages (45×25×20 cm) containing woodshavings as bedding. The animals were maintained at aconstant temperature (20±2°C) under standard lightingconditions (12:12 h light–dark, lights on from 0800 to2000 h). All experiments were carried out during the lightphase between 0830 h and 1500 h. Food and water wereavailable ad libitum. Animals were habituated to handlingand received an intraperitoneal (i.p.) injection of sterilesaline (0.89% NaCl) for 4 days prior to experimentation inorder to minimise the influence of injection procedure onbiological endpoints. The experimental protocol was carriedout in accordance with the guidelines of the Animal WelfareCommittee, National University of Ireland, Galway underlicence from the Irish Department of Health and Childrenand in compliance with the European Communities Councildirective 86/609.
2.2. Experimental design
Rats received either HU210 [(−)-11-hydroxy- Δ8tetrahy-drocannabinol-dimethylheptyl] (100 μg/kg), SR141716A [N-(piperdin-1-yl)-5-(4-chloropheny)-1-(2,4-dichloropheny)-4-methyl-1H-pyrazole-3-carboxamide hydrochloride] (3 mg/kg), SR144528 [N-[(1s)-endo-1,3,3-timethylbicyclo[2.2.1]heptan-2-yl]5-(4-choro-3-methylpanyl)-1-(4-methlbenzyl)pyrazole-3-carboxamide] (3 mg/kg), co-administration ofHU210 (100 μg/kg)+SR141716A (3 mg/kg), co-administra-tion of HU210 (100 μg/kg)+SR144528 (3 mg/kg) or vehicle(ethanol: Tween 80: saline; 1:1:18) at an injection volume of2 ml/kg i.p. Rats were injected 30 min prior to a second i.p.injection of either lipopolysaccaride (LPS: 100 μg/kg) orsaline vehicle (sterile 0.89% NaCl) administered at a volumeof 1 ml/kg. Rats were randomly assigned to experimentalgroups and the sequence of testing was randomised in order tocontrol for any confounding factors. The dose and time of LPSadministration were chosen on the basis of a previous study inour laboratory which demonstrated increases in peripheral orcentral cytokine (IL-1β, IL-6, IL-10 and IFNγ) levels at 2 and4 h post LPS administration (Roche et al., 2005). The vehicle-saline treated control group was comprised of 4 rats at eachtime point (2 and 4 h) and data from both time points weresubsequently pooled following statistical analysis (Student'sunpaired t-test) demonstrating no significant differencebetween cytokine levels at these two time points. There were6–9 rats in each of the other experimental groups. The doseand time of HU210 administration were based on studiesdemonstrating its effectiveness in attenuating LPS-inducedincreases in serum cytokine levels (Smith et al., 2000).Antagonist doses were chosen based on previous studiesdemonstrating their ability to block the effects of HU210(Cheer et al., 2000; Finn et al., 2004). Blood samples weretaken 2 and 4 h post LPS or saline administration, via cardiacpuncture into a heparinized syringe under CO2 anaesthesia.Differential blood cell counts were determined by a Vet ABCanimal blood counter (ABX Diagnostics, France) andexpressed as count ×106 per ml of blood. Remaining blood
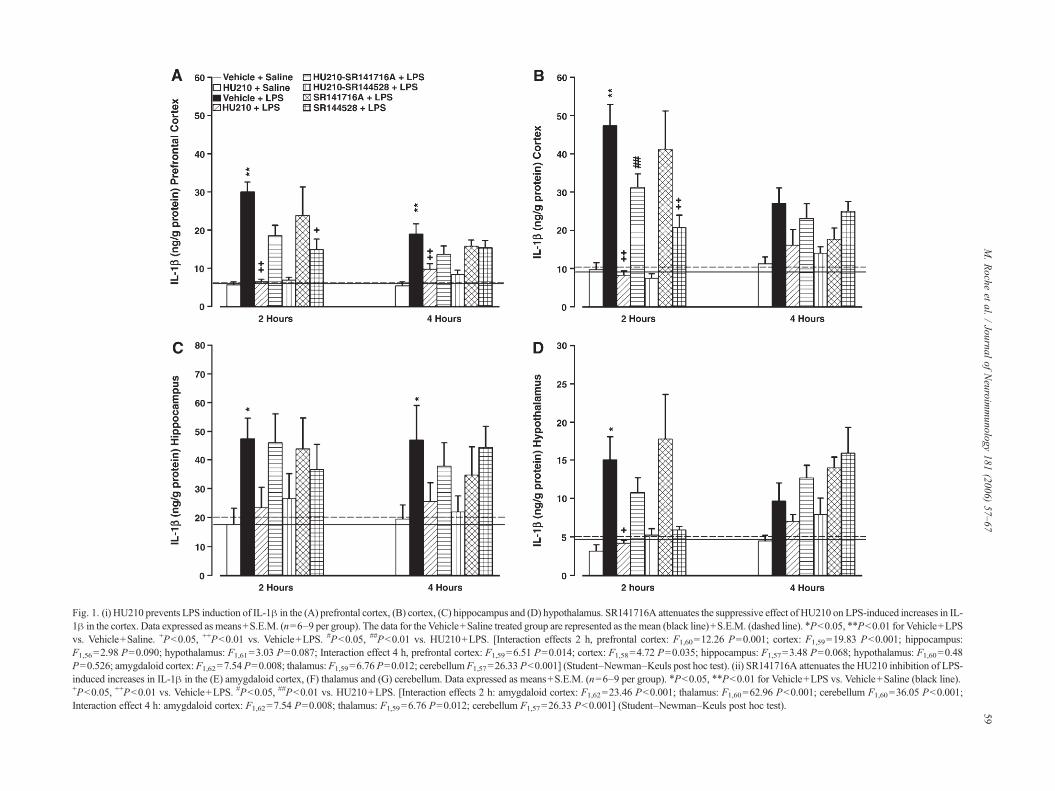
Fig. 1. (i) HU210 prevents LPS induction of IL-1β in the (A) prefrontal cortex, (B) cortex, (C) hippocampus and (D) hypothalamus. SR141716A attenuates the suppressive effect of HU210 on LPS-induced increases in IL-1β in the cortex. Data expressed as means+S.E.M. (n=6–9 per group). The data for the Vehicle+Saline treated group are represented as the mean (black line)+S.E.M. (dashed line). ⁎P<0.05, ⁎⁎P<0.01 for Vehicle+LPSvs. Vehicle+Saline. +P<0.05, ++P<0.01 vs. Vehicle+LPS. #P<0.05, ##P<0.01 vs. HU210+LPS. [Interaction effects 2 h, prefrontal cortex: F1,60=12.26 P=0.001; cortex: F1,59=19.83 P<0.001; hippocampus:F1,56=2.98 P=0.090; hypothalamus: F1,61=3.03 P=0.087; Interaction effect 4 h, prefrontal cortex: F1,59=6.51 P=0.014; cortex: F1,58=4.72 P=0.035; hippocampus: F1,57=3.48 P=0.068; hypothalamus: F1,60=0.48P=0.526; amygdaloid cortex: F1,62=7.54 P=0.008; thalamus: F1,59=6.76 P=0.012; cerebellumF1,57=26.33 P<0.001] (Student–Newman–Keuls post hoc test). (ii) SR141716A attenuates the HU210 inhibition of LPS-induced increases in IL-1β in the (E) amygdaloid cortex, (F) thalamus and (G) cerebellum. Data expressed as means+S.E.M. (n=6–9 per group). ⁎P<0.05, ⁎⁎P<0.01 for Vehicle+LPS vs. Vehicle+Saline (black line).+P<0.05, ++P<0.01 vs. Vehicle+LPS. #P<0.05, ##P<0.01 vs. HU210+LPS. [Interaction effects 2 h: amygdaloid cortex: F1,62=23.46 P<0.001; thalamus: F1,60=62.96 P<0.001; cerebellum F1,60=36.05 P<0.001;Interaction effect 4 h: amygdaloid cortex: F1,62=7.54 P=0.008; thalamus: F1,59=6.76 P=0.012; cerebellum F1,57=26.33 P<0.001] (Student–Newman–Keuls post hoc test). 59
M.Roche
etal.
/Journal
ofNeuroim
munology
181(2006)
57–67
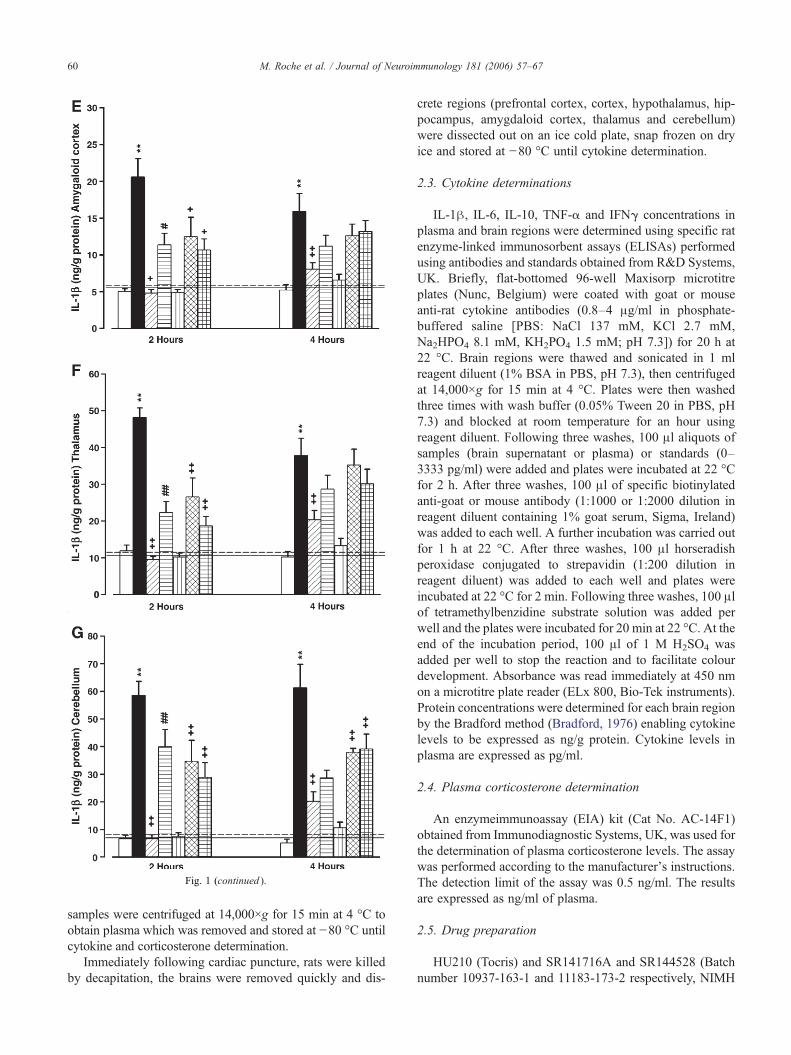
Fig. 1 (continued ).
60 M. Roche et al. / Journal of Neuroimmunology 181 (2006) 57–67
samples were centrifuged at 14,000×g for 15 min at 4 °C toobtain plasma which was removed and stored at −80 °C untilcytokine and corticosterone determination.
Immediately following cardiac puncture, rats were killedby decapitation, the brains were removed quickly and dis-
crete regions (prefrontal cortex, cortex, hypothalamus, hip-pocampus, amygdaloid cortex, thalamus and cerebellum)were dissected out on an ice cold plate, snap frozen on dryice and stored at −80 °C until cytokine determination.
2.3. Cytokine determinations
IL-1β, IL-6, IL-10, TNF-α and IFNγ concentrations inplasma and brain regions were determined using specific ratenzyme-linked immunosorbent assays (ELISAs) performedusing antibodies and standards obtained from R&D Systems,UK. Briefly, flat-bottomed 96-well Maxisorp microtitreplates (Nunc, Belgium) were coated with goat or mouseanti-rat cytokine antibodies (0.8–4 μg/ml in phosphate-buffered saline [PBS: NaCl 137 mM, KCl 2.7 mM,Na2HPO4 8.1 mM, KH2PO4 1.5 mM; pH 7.3]) for 20 h at22 °C. Brain regions were thawed and sonicated in 1 mlreagent diluent (1% BSA in PBS, pH 7.3), then centrifugedat 14,000×g for 15 min at 4 °C. Plates were then washedthree times with wash buffer (0.05% Tween 20 in PBS, pH7.3) and blocked at room temperature for an hour usingreagent diluent. Following three washes, 100 μl aliquots ofsamples (brain supernatant or plasma) or standards (0–3333 pg/ml) were added and plates were incubated at 22 °Cfor 2 h. After three washes, 100 μl of specific biotinylatedanti-goat or mouse antibody (1:1000 or 1:2000 dilution inreagent diluent containing 1% goat serum, Sigma, Ireland)was added to each well. A further incubation was carried outfor 1 h at 22 °C. After three washes, 100 μl horseradishperoxidase conjugated to strepavidin (1:200 dilution inreagent diluent) was added to each well and plates wereincubated at 22 °C for 2 min. Following three washes, 100 μlof tetramethylbenzidine substrate solution was added perwell and the plates were incubated for 20 min at 22 °C. At theend of the incubation period, 100 μl of 1 M H2SO4 wasadded per well to stop the reaction and to facilitate colourdevelopment. Absorbance was read immediately at 450 nmon a microtitre plate reader (ELx 800, Bio-Tek instruments).Protein concentrations were determined for each brain regionby the Bradford method (Bradford, 1976) enabling cytokinelevels to be expressed as ng/g protein. Cytokine levels inplasma are expressed as pg/ml.
2.4. Plasma corticosterone determination
An enzymeimmunoassay (EIA) kit (Cat No. AC-14F1)obtained from Immunodiagnostic Systems, UK, was used forthe determination of plasma corticosterone levels. The assaywas performed according to the manufacturer's instructions.The detection limit of the assay was 0.5 ng/ml. The resultsare expressed as ng/ml of plasma.
2.5. Drug preparation
HU210 (Tocris) and SR141716A and SR144528 (Batchnumber 10937-163-1 and 11183-173-2 respectively, NIMH
61M. Roche et al. / Journal of Neuroimmunology 181 (2006) 57–67
chemical synthesis program) were dissolved in 100%ethanol. Surfactant (Tween 80, Sigma Aldrich, Ireland) andsterile saline (0.89% NaCl) were then added to produce afinal concentration of either 50 μg/ml of HU210 or 1.5 mg/ml of SR141716A and SR144528 in vehicle (ethanol: tween80: saline at 1:1:18). An emulsion mixture of HU210+SR1411716A or HU210+SR144528 was obtained bydissolving the SR compounds in the HU210 ethanolicsolution followed by addition of Tween 80 and saline in theratio of 1:1:18. LPS (Escherichia coli serotype 0111:B4,Sigma Aldrich, Ireland) was dissolved in sterile saline toproduce a final volume of 100 μg/ml. Drugs or corre-sponding vehicle were administered i.p. in a volume of 1 or2 ml/kg.
2.6. Data analysis
All data are presented as means±S.E.M. Cytokine con-centrations were analysed by two-way ANOVA using SPSS(Version 12.0.1), with LPS and drug treatment as factors,followed by Student–Newman–Keuls post hoc test whereappropriate. Comparisons between vehicle-saline treatedcontrols at 2 and 4 h were made using Student's unpaired t-test. The level of significance was set at P<0.05.
3. Results
3.1. HU210 prevents LPS induction of IL-1β in discretebrain regions, attenuation by SR141716A
Since cytokine levels in the vehicle-saline group at the 2 htime point were not significantly different to levels in thisgroup at the 4 h time point, (Student's unpaired t-test) thesetwo groups were pooled and comparisons made against thisone set of vehicle-treated controls. The effects of HU210,SR141716A and SR144528, alone and in combination, oncentral IL-1β production following LPS administration areshown in Fig. 1 together with results of statistical analysis.LPS administration induced a significant increase in IL-1β inthe prefrontal cortex, cortex, hippocampus, hypothalamus,amygdaloid cortex, thalamus and cerebellum 2 h followingadministration compared with vehicle-treated controls. Al-though an increase was also seen in most brain regions at the4 h time point, the magnitude of the IL-1β increase was notas pronounced as that seen 2 h post-LPS injection. HU210alone did not alter cytokine levels in any of the brain regionsexamined when compared with vehicle-treated controls.However, HU210 fully attenuated the LPS-induced increasein IL-1β levels in all regions examined 2 h following admi-nistration, with the exception of the hippocampus where the
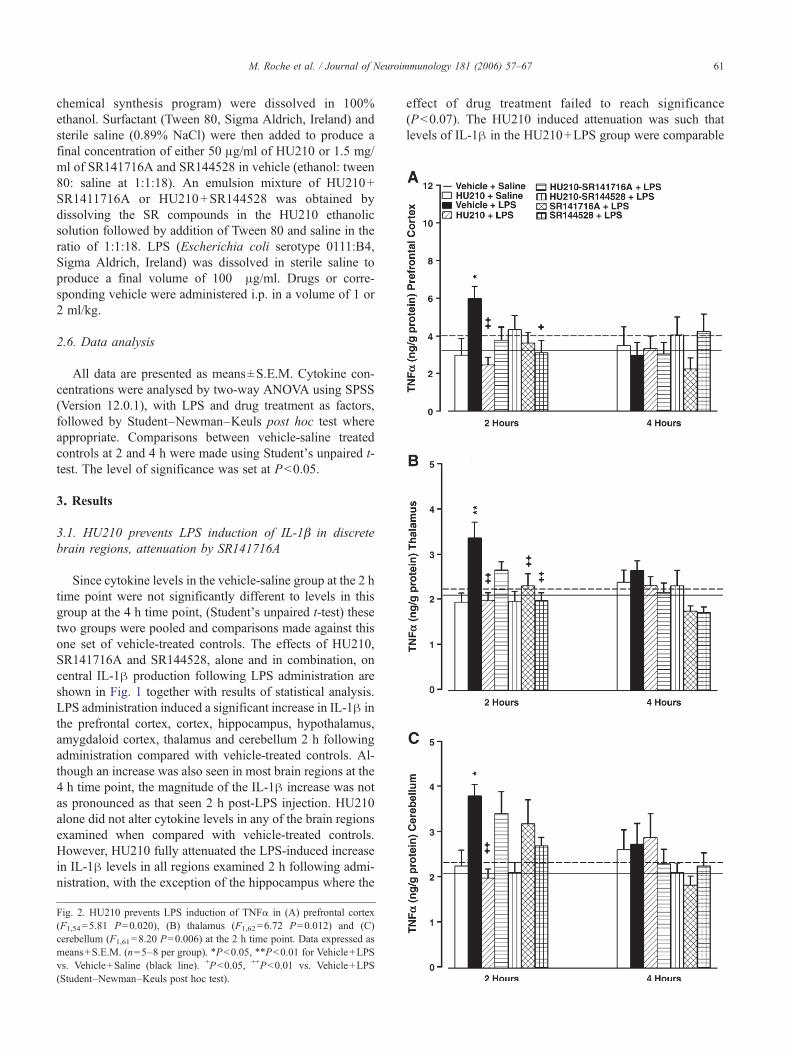
Fig. 2. HU210 prevents LPS induction of TNFα in (A) prefrontal cortex(F1,54=5.81 P=0.020), (B) thalamus (F1,62=6.72 P=0.012) and (C)cerebellum (F1,61=8.20 P=0.006) at the 2 h time point. Data expressed asmeans+S.E.M. (n=5–8 per group). ⁎P<0.05, ⁎⁎P<0.01 for Vehicle+LPSvs. Vehicle+Saline (black line). +P<0.05, ++P<0.01 vs. Vehicle+LPS(Student–Newman–Keuls post hoc test).
effect of drug treatment failed to reach significance(P<0.07). The HU210 induced attenuation was such thatlevels of IL-1β in the HU210+LPS group were comparable
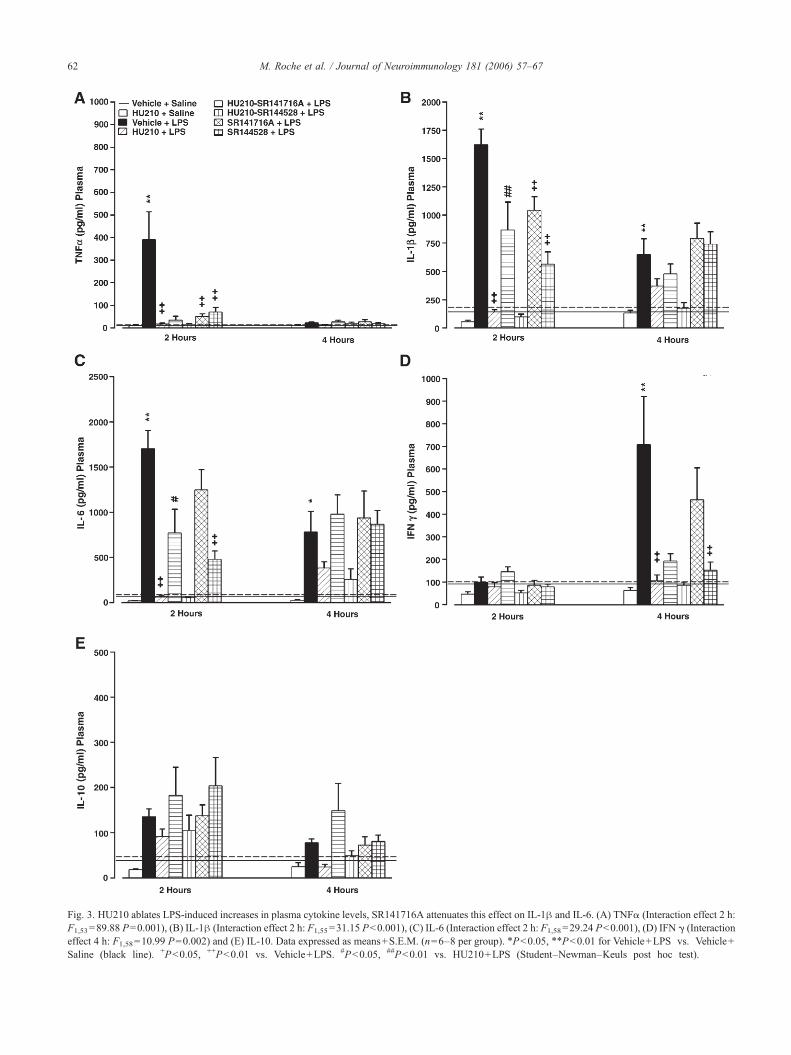
Fig. 3. HU210 ablates LPS-induced increases in plasma cytokine levels, SR141716A attenuates this effect on IL-1β and IL-6. (A) TNFα (Interaction effect 2 h:F1,53=89.88 P=0.001), (B) IL-1β (Interaction effect 2 h: F1,55=31.15 P<0.001), (C) IL-6 (Interaction effect 2 h: F1,58=29.24 P<0.001), (D) IFN γ (Interactioneffect 4 h: F1,58=10.99 P=0.002) and (E) IL-10. Data expressed as means+S.E.M. (n=6–8 per group). ⁎P<0.05, ⁎⁎P<0.01 for Vehicle+LPS vs. Vehicle+Saline (black line). +P<0.05, ++P<0.01 vs. Vehicle+LPS. #P<0.05, ##P<0.01 vs. HU210+LPS (Student–Newman–Keuls post hoc test).
62 M. Roche et al. / Journal of Neuroimmunology 181 (2006) 57–67

Fig. 5. Circulating corticosterone levels are increased 2 (Interaction effect:F1,61=7.10 P=0.002) and 4 (Interaction effect: F1,58=6.65 P=0.013) hoursfollowing acute administration of LPS and cannabinoids, alone or incombination. Data expressed as means+S.E.M. (n=6–8 per group).⁎⁎P<0.01 vs. Vehicle+Saline (black line). +P<0.05 vs. Vehicle+LPS(Student–Newman–Keuls post hoc test).
Fig. 4. Acute administration of cannabinoids and LPS administration, aloneand in combination, reduced plasma lymphocyte concentration at both the 2(Interaction effect: F1,62 =8.90 P=0.004) and 4 (Interaction effect:F1,62=6.39 P=0.014) hour time points. Data expressed as means+S.E.M.(n=6–8 per group). ⁎⁎P<0.01 vs. Vehicle+Saline (black line) (Student–Newman–Keuls post hoc test).
63M. Roche et al. / Journal of Neuroimmunology 181 (2006) 57–67
with levels in vehicle+ saline group. HU210 also signifi-cantly attenuated the IL-1β production in the prefrontalcortex, amygdaloid cortex, thalamus and cerebellum 4 h postLPS administration. Co-administration of the CB1 antagonistSR141716A with HU210 resulted in a significant, albeitpartial, attenuation of the HU210-induced inhibition of IL-1β production in the cortex, amygdaloid cortex, thalamusand cerebellum 2 h post LPS. In contrast, co-administrationof the CB2 antagonist SR144528 with HU210 did notprevent the inhibitory effect of HU210 on LPS-induced IL-1β increases. SR141716A alone significantly reduced LPS-induced IL-1β production in the amygdaloid cortex andthalamus at the 2 h time point and in the cerebellum at boththe 2 and 4 h time points. In addition, LPS-induced IL-1βproduction in the prefrontal cortex, cortex, amygdaloid cor-tex and thalamus was significantly reduced by SR144528alone at the 2 h time point. SR144528 also reduced IL-1βlevels in the cerebellum at both the 2 and 4 h post LPSadministration.
3.2. HU210 prevents LPS induction of TNFα in discretebrain regions
Increased TNFα levels in the prefrontal cortex, thalamusand cerebellum were observed 2, but not 4, hours followingLPS administration (Fig. 2), with levels remaining un-changed in other brain regions (data not shown). HU210alone had no significant effect on TNFα production in anyof the brain regions examined. However, HU210 fully atten-uated the LPS induction of TNFα in the prefrontal cortex,thalamus and cerebellum. Co-administration of HU210 witheither SR141716A or SR144528 did not alter the effect ofHU210 on LPS-induced production of TNFα. SR141716Aalone reduced the effect of LPS on TNFα production in the
thalamus while SR144528 reduced the LPS-induced increasein TNFα in both the prefrontal cortex and thalamus.
3.3. No effect of LPS or cannabinoid drugs on centralproduction of IL-6, IL-10 or IFNγ
There was no significant effect of LPS administration oncentral IL-6, IL-10 or IFNγ levels in any of the brain regionsat either the 2 or 4 h time period (data not shown). HU210,either alone or in combination with LPS, exhibited no effecton central production of the aforementioned cytokines in anyof the brain regions examined.
3.4. HU210 blocks LPS-induced increases in cytokine levelsin plasma, SR141716A attenuates this effect on IL-1β andIL-6
LPS induced a significant increase in plasma TNFα levels2 h, IL-1β and IL-6 levels 2 and 4 h and IFNγ levels 4 hpost injection when compared with vehicle-treated controls(Fig. 3). HU210 alone had no significant effect on plasmacytokine levels. However, HU210 fully attenuated the LPS-induced increase in the levels of IL-1β, TNFα and IL-6 at the2 h time point and IFNγ at 4 h. Co-administration of HU210with SR141716A resulted in a significant, albeit partial,attenuation of the HU210 reduction in LPS-induced IL-1βand IL-6 levels but not the TNFα levels, at the 2 h time point.Co-administration of the CB2 antagonist SR144528 withHU210 did not prevent the inhibitory effect of HU210 onLPS-induced plasma cytokine increases. SR141716A aloneattenuated the LPS-induced IL-1β and TNFα levels at the2 h time point. SR144528 alone attenuated the LPS inductionof plasma IL-1β, TNFα and IL-6 at the 2 h time point andIFN at the 4 h time.

64 M. Roche et al. / Journal of Neuroimmunology 181 (2006) 57–67
3.5. Lymphocyte concentrations are reduced following acuteLPS and HU210 administration
Acute administration of LPS alone reduced total circu-lating white blood cell counts at both 2 and 4 h post-administration, which was found to be due to a decrease inlymphocyte cell numbers (Fig. 4), an effect that was notmodulated by HU210 or the antagonists. Administration ofHU210 alone also reduced circulating lymphocyte numbersat both time points to an extent comparable with the effectsof LPS.
3.6. HU210 and LPS increases plasma corticosterone levels
Plasma corticosterone levels were significantly increasedat 2 and 4 h following the administration of LPS whencompared with vehicle-treated controls (Fig. 5). Adminis-tration of the cannabinoid receptor agonist HU210 alone alsoincreased plasma corticosterone levels to an extent compa-rable with the effects of LPS at both time points. The co-administration of HU210 and the antagonists did not affectthe LPS-induced increase in corticosterone levels. However,SR141716A alone significantly potentiated the LPS-inducedincrease in plasma corticosterone levels at the 2 h time point.
4. Discussion
LPS increased the release of the pro-inflammatorycytokines IL-1β and TNFα in discrete rat brain regions, aneffect that was attenuated by the cannabinoid receptoragonist HU210. Although previous studies have revealed theimmunosuppressive effect of cannabinoids, this is the first invivo study to examine cannabinoid effects on LPS-inducedbrain cytokine levels. The study also confirms previousfindings that HU210 attenuates LPS-induced increases inplasma cytokine levels. CB1 receptors contribute to theimmunosuppressive effects of HU210, both centrally andperipherally, since SR141716A attenuated, albeit partially,the decrease in LPS-induced cytokine release induced bythis cannabinoid receptor agonist. Interestingly, systemicadministration of the CB1 or CB2 receptor antagonists aloneblocked LPS-induced increases in cytokine levels in manybrain regions and in plasma. Changes in cytokine levels wereaccompanied by reduced circulating lymphocyte numbersand increased plasma corticosterone levels in response toacute administration of LPS and/or cannabinoid drugs.
Acute LPS administration has long been associated withincreases in plasma pro-inflammatory cytokines (Beishuizenand Thijs, 2003; Turrin et al., 2001). The majority of studiesinvestigating the effects of LPS on brain cytokines, however,have measured cytokine mRNA levels with few studiesdirectly measuring cytokine protein levels. The present studyprovides direct evidence generated in vivo for cannabinoidreceptor-mediated suppression of pro-inflammatory cytokineproduction in discrete brain regions following immune chal-lenge. The attenuation by HU210 of LPS-induced increases
in levels of TNFα and IL-1β in the brain supports studies invitro demonstrating that endogenous and synthetic cannabi-noids, including HU210, attenuate the release of TNFα(Facchinetti et al., 2003) and pro-inflammatory cytokinemRNA expression (Puffenbarger et al., 2000) in LPS-stimulated rat cortical microglial cells.
Our data indicate that the immunosuppressive effects ofHU210 on IL-1β levels in the cortex, amygdaloid cortex,thalamus and cerebellum are mediated, at least in part, by theCB1 receptor since co-administration of the CB1 receptorantagonist, SR141716A, but not the CB2 receptor antagonist,SR144528, attenuated the effects of HU210. However, inline with previous studies in vitro (Puffenbarger et al., 2000;Facchinetti et al., 2003), neither SR141716A nor SR144528attenuated the HU210-induced suppression of central TNFαproduction following LPS administration. Taken together,these findings suggest that CB1 receptors are differentiallyinvolved in modulating central levels of individual pro-inflammatory cytokines and that a novel, as yet undiscov-ered, cannabinoid receptor subtype(s) may mediate some ofthe immunosuppressive effects of cannabinoid receptoragonists including HU210.
Several studies have reported the immunosuppressiveeffect of cannabinoids on peripheral, circulating cytokinesincluding TNFα, IL-10 and IL-12 (Gallily et al., 1997; Smithet al., 2000; Croci et al., 2003). The present study confirmsthese findings and further expands this field by investigatingeffects on a broad range of cytokines examined concurrentlyat two different time points. SR141716A, but not SR144528,attenuated the immunosuppressive effect of HU210 on LPS-induced increases in IL-1β and IL-6 levels in plasma,reaffirming a role for the CB1 receptor in modulating peri-pheral cytokine levels (Smith et al., 2000; Croci et al., 2003).As was the case for central TNFα, neither of the antagonistsaltered the HU210 effect on plasma TNFα or IFNγ levels,suggesting the potential involvement of novel cannabinoidreceptor subtypes in regulating peripheral levels of thesecytokines. The failure of the CB2 receptor antagonist toattenuate the HU210-induced effects is interesting in light ofsubstantial evidence demonstrating the high expression ofCB2 receptors in immune tissue (Bouaboula et al., 1993;Munro et al., 1993) and an important role for CB2 receptorsin mediating the anti-inflammatory effects of cannabinoids(Mazzari et al., 1996; Sacerdote et al., 2000; Germain et al.,2002). It seems likely therefore, that the respective rolesof CB1 and CB2 receptors in the regulation of cytokinesare dependent on the type of immune or inflammatory chal-lenge and the particular cytokine or immune mediator underinvestigation.
Of further interest is the finding that administration ofSR141716A or SR144528 alone was sufficient to suppresscentral and peripheral cytokine responses, in a manner qual-itatively and quantitatively similar to the immunosuppres-sive effects of HU210. Indeed, immunosuppressive and anti-inflammatory effects of SR141716A have previously beenreported (Smith et al., 2000; Croci et al., 2003; Costa et al.,

65M. Roche et al. / Journal of Neuroimmunology 181 (2006) 57–67
2005). Thus, Croci et al. (2003) demonstrated thatSR141716A inhibited LPS-induced increases in TNFαlevels in wild-type mice but not in mice lacking the CB1
receptor. Smith et al. (2000) reported a similar attenuation ofLPS-induced increases in circulating TNFα levels bySR141716A in mice, prompting these authors to proposethat SR141716A may act as a partial agonist at the CB1
receptor when administered alone. Our data support thishypothesis and extend it to include apparent partial agonisteffects of SR141716A on both peripheral and centralcytokines in rats. Less is known about the potential partialagonist effects of SR144528 at the CB2 receptor, however,suppressive effects on LPS-induced increases in levels ofcirculating TNFα and IL-12 and on development of inflam-mation in mice have been reported following intra-cerebraladministration of this CB2 receptor antagonist (Smith et al.,2001a,b).
Alternatively, the immunosuppressive effects of theantagonists may be associated with their ability to blockthe effects of released endocannabinoids on CB1 and CB2
receptors. LPS itself induces the release of endocannabinoidsby macrophages (Di Marzo et al., 1999; Maccarrone et al.,2002) and platelets (Maccarrone et al., 2002) and inhibitsendocannabinoid degradation in lymphocytes (Maccarroneet al., 2001), effects that may also occur in microglial cellsand astrocytes in the brain. In the presence of high levels ofendocannabinoids, therefore, the effects of the antagonistsmay result from unmasking the action of endocannabinoidsat alternative targets such as the transient receptor potentialvanilloid type 1 (TRPV1) receptor (Zygmunt et al., 1999;Cabranes et al., 2005; Cristino et al., 2006) or peroxisomeproliferator activated receptor (PPAR)-gamma (Rockwellet al., 2006) following blockade of the CB1 or CB2 receptors.It is also possible that whilst potent synthetic agonists such asHU210 exert an immunosuppressive effect, endocannabi-noids such as anandamide and/or 2-arachidonoyl glycerolmay signal through CB1 and CB2 receptors to facilitate LPS-induced cytokine release in vivo. Such a mechanism ofaction would explain the immunosuppressive effects ob-served following pharmacological blockade of these recep-tors. It should be noted, however, that the weight of evidencefrom in vitro studies argues against this hypothesis. Thus,treatment of astrocytes and microglia with endocannabinoidsand inhibition of endocannabinoid degradation or re-uptakehas been shown to reduce production of cytokines includingIL-1β, TNFα and IL-6 in a manner similar to that whichoccurs following direct activation of cannabinoid receptorswith synthetic agonists (Facchinetti et al., 2003; Ortega-Gutierrez et al., 2005a,b; Mestre et al., 2005). Furthermore,Panikashvili et al. (2006) have recently shown in vivo thatthe endocannabinoid 2-AG attenuates increases in levels ofIL-1β, TNFα and IL-6 mRNA in the mouse brain followingclosed head injury. However, a study by Molina-Holgadoet al. (1998) demonstrated a stimulatory effect of ananda-mide on IL-6 release from astrocytes infected with Theiler'smurine encephalomyelitis virus, supporting the notion that
endocannabinoids may, under some circumstances, act tofacilitate pro-inflammatory cytokine release. Taken together,these studies highlight the complexity of endocannabinoidsystem involvement in neuro-immune regulation. Future invivo studies should build on the data generated in the presentstudy to investigate the effects of inhibiting endocannabinoiddegradation and uptake on LPS-induced alterations in braincytokines.
The LPS-induced increase in central and peripheralcytokine levels was accompanied by an increase in plasmacorticosterone levels in accordance with several previousstudies (Beishuizen and Thijs, 2003; Bik et al., 2004). LPS-induced increases in plasma IL-1β, TNFα and IL-6 mayact directly on the pituitary and adrenal glands in orderto increase corticosterone levels (Lenczowski et al., 1997;Mazzocchi et al., 1998; Bethin et al., 2000). Increased cyto-kine levels, for example IL-1β, in certain brain regions suchas the hypothalamus may be a further mechanism by whichLPS activates the HPA axis (Rivier et al., 1989). HU210,administered alone, also increased plasma corticosteronelevels but did not affect peripheral or central cytokine levelssuggesting that the effects of this cannabinoid receptoragonist on HPA axis activity are not mediated by alterationsin cytokine levels. Several studies have previously demon-strated that cannabinoids, including HU210, activate theHPA axis and increase circulating corticosterone levels(Weidenfeld et al., 1994; Martin-Calderon et al., 1998; Finnet al., 2004). Cannabinoid activation of central CB1 receptorsmay disinhibit hypothalamic neurons and elicit a numberof endocrine effects (Rodriguez de Fonseca et al., 1996;Martin-Calderon et al., 1998). Whilst HU210 attenuatedLPS-induced increases in peripheral and central cytokinelevels it did not attenuate the LPS-induced increase in plasmacorticosterone levels, further indicating the likely dissocia-tion between the effects of this cannabinoid receptor agoniston the immune and endocrine systems. Moreover, admin-istration of SR141716A alone, but not SR144528, potenti-ated the LPS-induced increase in corticosterone. Since thepotent agonist, HU210, failed to produce this effect, it isunlikely that the effects of SR141716A reflect partial agonistactivity but instead may be due to blockade of endocanna-binoids signalling through the CB1 receptor or unmaskingtheir effects at additional receptors as discussed above. Theincrease in plasma corticosterone in response to LPS and/orcannabinoid drugs was accompanied by a reduction in cir-culating lymphocyte numbers. It has been demonstrated thatboth glucocorticoids and increased central IL-1β cansuppress lymphocyte proliferation (Sundar et al., 1990;Wiegers et al., 1993) and either of these mechanisms mayaccount for the reduced lymphocyte number observed post-LPS administration. HU210 alone suppressed circulatinglymphocyte numbers but had no effect on the quantitativelycomparable LPS-induced suppression of lymphocyte numb-ers. Studies have shown that Δ9-THC and other cannabi-noids directly reduce proliferation and cytolytic responsesand induce apoptosis of lymphocytes, acting mainly via the

66 M. Roche et al. / Journal of Neuroimmunology 181 (2006) 57–67
CB2 receptor (Klein et al., 1991; Schwarz et al., 1994;McKallip et al., 2002). However, in the present study, thesuppressive effects of HU210 on circulating lymphocytenumbers were not blocked by either the CB2 or CB1
antagonist. Further studies employing adrenalectomy andcorticosterone replacement should test the hypothesis thatthe HU210-induced suppression of lymphocyte numbers ismediated by increased corticosterone levels.
In summary, the present study demonstrates the immuno-suppressive effect of HU210 on LPS-induced cytokineproduction both in the brain and periphery. This effect waspartially mediated by the CB1 receptor, however, the potentialinvolvement of additional receptor targets for cannabinoidscannot be excluded. In addition, both CB1 and CB2 receptorantagonists alone suppressed the LPS-induced increase incytokine levels, in the central nervous system and plasma.Increases in central pro-inflammatory cytokine productionhave been associatedwith a number of neuroimmune disordersincluding Alzheimer's disease, multiple sclerosis and chronicpain. Elucidation of the fundamental physiology of endocan-nabinoid–neuroimmune interactions and the immunosuppres-sive properties of cannabinoid agonists and antagonists willcontinue to inform the development of these compounds forthe treatment of neuroinflammatory disorders.
Acknowledgements
This work was supported by grants from the Irish HealthResearch Board and the National University of Ireland,Galway Millennium Fund. Technical assistance from DanielKerr, Ruth McNamara, Catherine Diskin and José ManuelEntrena is gratefully acknowledged.
References
Beishuizen, A., Thijs, L.G., 2003. Endotoxin and the hypothalamo–pituitary–adrenal (HPA) axis. J. Endotoxin Res. 9, 3–24.
Bethin, K.E., Vogt, S.K., Muglia, L.J., 2000. Interleukin-6 is an essential,corticotropin-releasing hormone-independent stimulator of the adrenalaxis during immune system activation. Proc. Natl. Acad. Sci. U. S. A.97, 9317–9322.
Bik, W., Wolinska-Witort, E., Chmielowska, M., Baranowska-Bik, A.,Rusiecka-Kuczalek, E., Baranowska, B., 2004. Vasoactive intestinalpeptide can modulate immune and endocrine responses duringlipopolysaccharide-induced acute inflammation. Neuroimmunomodula-tion 11, 358–364.
Bouaboula, M., Rinaldi, M., Carayon, P., Carillon, C., Delpech, B., Shire,D., Le Fur, G., Casellas, P., 1993. Cannabinoid-receptor expression inhuman leukocytes. Eur. J. Biochem. 214, 173–180.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal. Biochem. 72, 248–254.
Cabranes, A., Venderova, K., de Lago, E., Fezza, F., Sanchez, A., Mestre, L.,Valenti, M., Garcia-Merino, A., Ramos, J.A., Di Marzo, V., Fernandez-Ruiz, J., 2005. Decreased endocannabinoid levels in the brain andbeneficial effects of agents activating cannabinoid and/or vanilloidreceptors in a rat model of multiple sclerosis. Neurobiol. Dis. 20, 207–217.
Chang, Y.H., Lee, S.T., Lin, W.W., 2001. Effects of cannabinoids on LPS-stimulated inflammatory mediator release from macrophages: involve-ment of eicosanoids. J. Cell. Biochem. 81, 715–723.
Cheer, J.F., Kendall, D.A., Marsden, C.A., 2000. Cannabinoid receptors andreward in the rat: a conditioned place preference study. Psychopharma-cology 151, 25–30.
Costa, B., Trovato, A.E., Colleoni, M., Giagnoni, G., Zarini, E., Croci, T.,2005. Effect of the cannabinoid CB1 receptor antagonist, SR141716, onnociceptive response and nerve demyelination in rodents with chronicconstriction injury of the sciatic nerve. Pain 116, 52–61.
Cristino, L., de Petrocellis, L., Pryce, G., Baker, D., Guglielmotti, V., DiMarzo, V., 2006. Immunohistochemical localization of cannabinoid type1 and vanilloid transient receptor potential vanilloid type 1 receptors inthe mouse brain. Neuroscience 139, 1405–1415.
Croci, T., Landi, M., Galzin, A.M., Marini, P., 2003. Role of cannabinoidCB1 receptors and tumor necrosis factor-alpha in the gut and systemicanti-inflammatory activity of SR 141716 (rimonabant) in rodents. Br. J.Pharmacol. 140, 115–122.
Di Marzo, V., Bisogno, T., De Petrocellis, L., Melck, D., Orlando, P.,Wagner, J.A., Kunos, G., 1999. Biosynthesis and inactivation of theendocannabinoid 2-arachidonoylglycerol in circulating and tumoralmacrophages. Eur. J. Biochem. 264, 258–267.
Di Marzo, V., Bifulco, M., De Petrocellis, L., 2004. The endocannabinoidsystem and its therapeutic exploitation. Nat. Rev., Drug Discov. 3,771–784.
Facchinetti, F., Del Giudice, E., Furegato, S., Passarotto, M., Leon, A., 2003.Cannabinoids ablate release of TNFalpha in rat microglial cellsstimulated with lypopolysaccharide. Glia 41, 161–168.
Finn, D.P., Chapman, V., 2004. Cannabinoids as analgesic agents: evidencefrom in vivo studies. Curr. Neuropharmacol. 2, 75–89.
Finn, D.P., Beckett, S.R., Roe, C.H., Madjd, A., Fone, K.C., Kendall, D.A.,Marsden, C.A., Chapman, V., 2004. Effects of coadministration ofcannabinoids and morphine on nociceptive behaviour, brain mono-amines and HPA axis activity in a rat model of persistent pain. Eur. J.Neurosci. 19, 678–686.
Galiegue, S., Mary, S., Marchand, J., Dussossoy, D., Carriere, D., Carayon,P., Bouaboula, M., Shire, D., Le Fur, G., Casellas, P., 1995. Expressionof central and peripheral cannabinoid receptors in human immunetissues and leukocyte subpopulations. Eur. J. Biochem. 232, 54–61.
Gallily, R., Yamin, A., Waksmann, Y., Ovadia, H., Weidenfeld, J., Bar-Joseph, A., Biegon, A., Mechoulam, R., Shohami, E., 1997. Protectionagainst septic shock and suppression of tumor necrosis factor alpha andnitric oxide production by dexanabinol (HU-211), a nonpsychotropiccannabinoid. J. Pharmacol. Exp. Ther. 283, 918–924.
Germain, N., Boichot, E., Advenier, C., Berdyshev, E.V., Lagente, V., 2002.Effect of the cannabinoid receptor ligand, WIN 55,212-2, on superoxideanion and TNF-alpha production by human mononuclear cells. Int.Immunopharmacol. 2, 537–543.
Grotenhermen, F., 2005. Cannabinoids. Curr. Drug Targets CNS Neurol.Disord. 4, 507–530.
Herkenham, M., Lynn, A.B., Little, M.D., Johnson, M.R., Melvin, L.S., deCosta, B.R., Rice, K.C., 1990. Cannabinoid receptor localization inbrain. Proc. Natl. Acad. Sci. U. S. A. 87, 1932–1936.
Jackson, S.J., Diemel, L.T., Pryce, G., Baker, D., 2005. Cannabinoids andneuroprotection in CNS inflammatory disease. J. Neurol. Sci. 233,21–25.
Kaplan, B.L., Rockwell, C.E., Kaminski, N.E., 2003. Evidence forcannabinoid receptor-dependent and -independent mechanisms of actionin leukocytes. J. Pharmacol. Exp Ther. 306, 1077–1085.
Klegeris, A., Bissonnette, C.J., McGeer, P.L., 2003. Reduction of humanmonocytic cell neurotoxicity and cytokine secretion by ligands of thecannabinoid-type CB2 receptor. Br. J. Pharmacol. 139, 775–786.
Klein, T.W., 2005. Cannabinoid-based drugs as anti-inflammatory thera-peutics. Nat. Rev., Immunol. 5, 400–411.
Klein, T.W., Kawakami, Y., Newton, C., Friedman, H., 1991. Marijuanacomponents suppress induction and cytolytic function of murinecytotoxic T cells in vitro and in vivo. J. Toxicol. Environ. Health 32,465–477.
Lenczowski, M.J., Van Dam, A.M., Poole, S., Larrick, J.W., Tilders, F.J.,1997. Role of circulating endotoxin and interleukin-6 in the ACTH and

67M. Roche et al. / Journal of Neuroimmunology 181 (2006) 57–67
corticosterone response to intraperitoneal LPS. Am. J. Physiol. 273,R1870–R1877.
Maccarrone, M., De Petrocellis, L., Bari, M., Fezza, F., Salvati, S., DiMarzo, V., Finazzi-Agro, A., 2001. Lipopolysaccharide downregulatesfatty acid amide hydrolase expression and increases anandamide levelsin human peripheral lymphocytes. Arch. Biochem. Biophys. 393,321–328.
Maccarrone, M., Bari, M., Battista, N., Finazzi-Agro, A., 2002. Endocan-nabinoid degradation, endotoxic shock and inflammation. Curr. DrugTargets Inflamm. Allergy 1, 53–63.
Martin-Calderon, J.L., Munoz, R.M., Villanua, M.A., del Arco, I., Moreno,J.L., de Fonseca, F.R., Navarro, M., 1998. Characterization of the acuteendocrine actions of (−)-11-hydroxy-delta8-tetrahydrocannabinol-dimethylheptyl (HU-210), a potent synthetic cannabinoid in rats. Eur.J. Pharmacol. 344, 77–86.
Matsuda, L.A., Lolait, S.J., Brownstein, M.J., Young, A.C., Bonner, T.I.,1990. Structure of a cannabinoid receptor and functional expression ofthe cloned cDNA. Nature 346, 561–564.
Mazzari, S., Canella, R., Petrelli, L., Marcolongo, G., Leon, A., 1996. N-(2-hydroxyethyl)hexadecanamide is orally active in reducing edemaformation and inflammatory hyperalgesia by down-modulating mastcell activation. Eur. J. Pharmacol. 300, 227–236.
Mazzocchi, G., Gottardo, G., Nussdorfer, G.G., 1998. A local immuno-endocrine interaction may mediate rat adrenal glucocorticoid response tobacterial endotoxins. Life Sci. 62, 1783–1787.
McKallip, R.J., Lombard, C., Martin, B.R., Nagarkatti, M., Nagarkatti, P.S.,2002. Delta(9)-tetrahydrocannabinol-induced apoptosis in the thymusand spleen as a mechanism of immunosuppression in vitro and in vivo.J. Pharmacol. Exp. Ther. 302, 451–465.
Mechoulam, R., 2002. Discovery of endocannabinoids and some randomthoughts on their possible roles in neuroprotection and aggression.Prostaglandins Leukot. Essent. Fat. Acids 66, 93–99.
Mechoulam, R., Fride, E., Di Marzo, V., 1998. Endocannabinoids. Eur. J.Pharmacol. 359, 1–18.
Mechoulam, R., Panikashvili, D., Shohami, E., 2002. Cannabinoids andbrain injury: therapeutic implications. Trends Mol. Med. 8, 58–61.
Mestre, L., Correa, F., Arevalo-Martin, A., Molina-Holgado, E., Valenti, M.,Ortar, G., Di Marzo, V., Guaza, C., 2005. Pharmacological modulationof the endocannabinoid system in a viral model of multiple sclerosis.J. Neurochem. 92, 1327–1339.
Molina-Holgado, F., Molina-Holgado, E., Guaza, C., 1998. The endogenouscannabinoid anandamide potentiates interleukin-6 production by astro-cytes infected with Theiler's murine encephalomyelitis virus by areceptor-mediated pathway. FEBS Lett. 433, 139–142.
Molina-Holgado, F., Pinteaux, E., Moore, J.D., Molina-Holgado, E., Guaza,C., Gibson, R.M., Rothwell, N.J., 2003. Endogenous interleukin-1receptor antagonist mediates anti-inflammatory and neuroprotectiveactions of cannabinoids in neurons and glia. J. Neurosci. 23, 6470–6474.
Munro, S., Thomas, K.L., Abu-Shaar, M., 1993. Molecular characterizationof a peripheral receptor for cannabinoids. Nature 365, 61–65.
Ortega-Gutierrez, S., Molina-Holgado, E., Arevalo-Martin, A., Correa, F.,Viso, A., Lopez-Rodriguez, M.L., Di Marzo, V., Guaza, C., 2005a.Activation of the endocannabinoid system as therapeutic approach in amurine model of multiple sclerosis. Faseb J. 19, 1338–1340.
Ortega-Gutierrez, S., Molina-Holgado, E., Guaza, C., 2005b. Effect ofanandamide uptake inhibition in the production of nitric oxide and in therelease of cytokines in astrocyte cultures. Glia 52, 163–168.
Panikashvili, D., Simeonidou, C., Ben-Shabat, S., Hanus, L., Breuer, A.,Mechoulam, R., Shohami, E., 2001. An endogenous cannabinoid (2-AG) is neuroprotective after brain injury. Nature 413, 527–531.
Panikashvili, D., Shein, N.A, Mechoulam, R., Trembovler, V., Kohen, R.,Alexandrovich,A., Shohami, E., 2006. The endocannabinoid 2-AGprotectsthe blood–brain barrier after closed head injury and inhibits mRNAexpression of proinflammatory cytokines. Neurobiol. Dis. 22, 257–264.
Puffenbarger, R.A., Boothe, A.C., Cabral, G.A., 2000. Cannabinoids inhibitLPS-inducible cytokine mRNA expression in rat microglial cells. Glia29, 58–69.
Rivier, C., Chizzonite, R., Vale, W., 1989. In the mouse, the activation of thehypothalamic–pituitary–adrenal axis by a lipopolysaccharide (endotox-in) is mediated through interleukin-1. Endocrinology 125, 2800–2805.
Roche, M., Diamond, M., Lonergan, I., Keeney, G., Kelly, J.P., Finn, D.P.,2005. The effects of acute lipopolysaccaride injection on levels of pro-and anti-inflammatory cytokines in discrete rat brain regions. J. Psycho-pharmacol. 19, A37.
Rockwell, C.E., Snider, N.T., Thompson, J.T., Vanden Heuvel, J.P.,Kaminski, N.E., 2006. Interleukin-2 suppression by 2-arachidonylglycerol is mediated through peroxisome proliferator activated receptor{gamma} independently of cannabinoid receptors 1 and 2. Mol.Pharmacol. 70 (1), 101–111.
Rodriguez de Fonseca, F., Rubio, P., Menzaghi, F., Merlo-Pich, E., Rivier, J.,Koob, G.F., Navarro, M., 1996. Corticotropin-releasing factor (CRF)antagonist [D-Phe12,Nle21,38,C alpha MeLeu37]CRF attenuates the acuteactions of the highly potent cannabinoid receptor agonist HU-210 ondefensive–withdrawal behavior in rats. J. Pharmacol. Exp. Ther. 276, 56–64.
Roth, M.D., Baldwin, G.C., Tashkin, D.P., 2002. Effects of delta-9-tetrahydrocannabinol on human immune function and host defense.Chem. Phys. Lipids 121, 229–239.
Sacerdote, P., Massi, P., Panerai, A.E., Parolaro, D., 2000. In vivo and invitro treatment with the synthetic cannabinoid CP55, 940 decreases thein vitro migration of macrophages in the rat: involvement of both CB1and CB2 receptors. J. Neuroimmunol. 109, 155–163.
Schwarz, H., Blanco, F.J., Lotz, M., 1994. Anadamide, an endogenouscannabinoid receptor agonist inhibits lymphocyte proliferation andinduces apoptosis. J. Neuroimmunol. 55, 107–115.
Smith, S.R., Terminelli, C., Denhardt, G., 2000. Effects of cannabinoidreceptor agonist and antagonist ligands on production of inflammatorycytokines and anti-inflammatory interleukin-10 in endotoxemic mice.J. Pharmacol. Exp. Ther. 293, 136–150.
Smith, S.R., Denhardt, G., Terminelli, C., 2001a. The anti-inflammatoryactivities of cannabinoid receptor ligands in mouse peritonitis models.Eur. J. Pharmacol. 432, 107–119.
Smith, S.R., Terminelli, C., Denhardt, G., 2001b. Modulation of cytokineresponses in Corynebacterium parvum-primed endotoxemic mice bycentrally administered cannabinoid ligands. Eur. J. Pharmacol. 425, 73–83.
Sundar, S.K., Cierpial, M.A., Kilts, C., Ritchie, J.C., Weiss, J.M., 1990.Brain IL-1-induced immunosuppression occurs through activation ofboth pituitary–adrenal axis and sympathetic nervous system bycorticotropin-releasing factor. J. Neurosci. 10, 3701–3706.
Turrin, N.P., Gayle, D., Ilyin, S.E., Flynn, M.C., Langhans, W., Schwartz, G.J., Plata-Salaman, C.R., 2001. Pro-inflammatory and anti-inflammatorycytokine mRNA induction in the periphery and brain followingintraperitoneal administration of bacterial lipopolysaccharide. BrainRes. Bull. 54, 443–453.
Waksman, Y., Olson, J.M., Carlisle, S.J., Cabral, G.A., 1999. The centralcannabinoid receptor (CB1) mediates inhibition of nitric oxide productionby rat microglial cells. J. Pharmacol. Exp. Ther. 288, 1357–1366.
Walter, L., Stella, N., 2004. Cannabinoids and neuroinflammation. Br. J.Pharmacol. 141, 775–785.
Walter, L., Franklin, A., Witting, A., Wade, C., Xie, Y., Kunos, G., Mackie,K., Stella, N., 2003. Nonpsychotropic cannabinoid receptors regulatemicroglial cell migration. J. Neurosci. 23, 1398–1405.
Watkins, L.R., Maier, S.F., Goehler, L.E., 1995. Cytokine-to-braincommunication: a review and analysis of alternative mechanisms. LifeSci. 57, 1011–1026.
Weidenfeld, J., Feldman, S., Mechoulam, R., 1994. Effect of the brainconstituent anandamide, a cannabinoid receptor agonist, on the hypotha-lamo–pituitary–adrenal axis in the rat. Neuroendocrinology 59, 110–112.
Wiegers, G.J., Croiset, G., Reul, J.M., Holsboer, F., de Kloet, E.R., 1993.Differential effects of corticosteroids on rat peripheral blood T-lymphocytemitogenesis in vivo and in vitro. Am. J. Physiol. 265, E825–E830.
Zygmunt, P.M., Petersson, J., Andersson, D.A., Chuang, H., Sorgard, M., DiMarzo, V., Julius, D., Hogestatt, E.D., 1999. Vanilloid receptors onsensory nerves mediate the vasodilator action of anandamide. Nature400, 452–457.


















