
In Vitro Treatment of Carcinoma Cell Lines With Pancreatic (Pro)Enzymes Suppresses the EMT Programme...
14
ORIGINAL PAPER In vitro treatment of carcinoma cell lines with pancreatic (pro)enzymes suppresses the EMT programme and promotes cell differentiation Macarena Perán & Juan Antonio Marchal & Maria A. García & Julian Kenyon & David Tosh Accepted: 2 May 2013 / Published online: 16 May 2013 # International Society for Cellular Oncology 2013 Abstract Background Previous research has suggested a putative util- ity of pancreatic (pro)enzymes in cancer treatment. The aim of the present study was to investigate the in vitro effects of a mixture of two pancreatic pro-enzymes, i.e., Chymotryp- sinogen and Trypsinogen, and the enzyme Amylase on three human cancer cell lines, i.e., OE33 (derived from an oesophageal carcinoma), Panc1 (derived from a pancreatic carcinoma) and Caco-2 (derived from a colon carcinoma). Results After treatment of the three cancer cell lines with different doses of the (pro)enzymes for up to 7 days, we observed (i) growth inhibition in a dose-dependent manner, (ii) enhanced expression of β-catenin and E-cadherin and decreased expression of several epithelial-mesenchymal transition (EMT)-associated genes, such as Vimentin, Snail and Slug, (iii) differentiation of Caco-2 cells, including the appearance of cell-specific differentiated structures such as microvilli and tight junctions, the acquisition of a more regular polygonal morphology, and an increased expression of the intestinal differentiation markers alkaline phosphatase and cytokeratin 8, and (iv) differentiation of Panc1 cells, including the formation of cell aggregates, an increment on lamellar bodies and an increased expression of the pancre- atic differentiation markers glucagon and insulin. Conclusions Our results show that the treatment of three different human cancer cell lines with pancreatic (pro)enzymes results in an enhancement of cell adhesion, an attenuation of several EMT-associated markers, and an in- crease in the expression of several differentiation-associated markers, suggesting the acquisition of a less malignant phe- notype and a decrease in proliferative capacity due to lineage- specific cellular differentiation. Keywords Cell adhesion . Cell differentiation . Pancreatic (pro)enzymes . Epithelial-mesenchymal transition . Cancer treatment 1 Introduction Tumour metastases represent a major cause of mortality in cancer patients. Cell detachment, migration and invasion of the surrounding tissues are key events in this process [1]. M. Perán : D. Tosh Centre for Regenerative Medicine, Department of Biology & Biochemistry, University of Bath, Bath, UK M. Perán (*) Department of Health Sciences, University of Jaén, Paraje de las Lagunillas sn, B3-212, Jaén 23071, Spain e-mail: [email protected] J. A. Marchal Department of Human Anatomy and Embryology, University of Granada, Granada, Spain J. A. Marchal Biopathology and Regenerative Medicine Institute (IBIMER), University of Granada, Granada, Spain M. A. García Research Unit, Hospital Universitario Virgen de las Nieves, Granada, Spain J. Kenyon (*) The Dove Clinic for Integrated Medicine, The Old Brewery, High Street, Twyford SO21 1RG, UK e-mail: [email protected] Cell Oncol. (2013) 36:289–301 DOI 10.1007/s13402-013-0134-8
-
Upload
arunsomeshwar -
Category
Documents
-
view
48 -
download
0
description
Propanc Heath Group Corp.'s peer-reviewed paper
Transcript of In Vitro Treatment of Carcinoma Cell Lines With Pancreatic (Pro)Enzymes Suppresses the EMT Programme...

ORIGINAL PAPER
In vitro treatment of carcinoma cell lines with pancreatic(pro)enzymes suppresses the EMT programmeand promotes cell differentiation
Macarena Perán & Juan Antonio Marchal & Maria A. García &
Julian Kenyon & David Tosh
Accepted: 2 May 2013 /Published online: 16 May 2013# International Society for Cellular Oncology 2013
AbstractBackground Previous research has suggested a putative util-ity of pancreatic (pro)enzymes in cancer treatment. The aimof the present study was to investigate the in vitro effects ofa mixture of two pancreatic pro-enzymes, i.e., Chymotryp-sinogen and Trypsinogen, and the enzyme Amylase on threehuman cancer cell lines, i.e., OE33 (derived from anoesophageal carcinoma), Panc1 (derived from a pancreaticcarcinoma) and Caco-2 (derived from a colon carcinoma).
Results After treatment of the three cancer cell lines withdifferent doses of the (pro)enzymes for up to 7 days, weobserved (i) growth inhibition in a dose-dependent manner,(ii) enhanced expression of β-catenin and E-cadherin anddecreased expression of several epithelial-mesenchymaltransition (EMT)-associated genes, such as Vimentin, Snailand Slug, (iii) differentiation of Caco-2 cells, including theappearance of cell-specific differentiated structures such asmicrovilli and tight junctions, the acquisition of a moreregular polygonal morphology, and an increased expressionof the intestinal differentiation markers alkaline phosphataseand cytokeratin 8, and (iv) differentiation of Panc1 cells,including the formation of cell aggregates, an increment onlamellar bodies and an increased expression of the pancre-atic differentiation markers glucagon and insulin.Conclusions Our results show that the treatment of threedifferent human cancer cell lines with pancreatic(pro)enzymes results in an enhancement of cell adhesion, anattenuation of several EMT-associated markers, and an in-crease in the expression of several differentiation-associatedmarkers, suggesting the acquisition of a less malignant phe-notype and a decrease in proliferative capacity due to lineage-specific cellular differentiation.
Keywords Cell adhesion . Cell differentiation . Pancreatic(pro)enzymes . Epithelial-mesenchymal transition . Cancertreatment
1 Introduction
Tumour metastases represent a major cause of mortality incancer patients. Cell detachment, migration and invasion ofthe surrounding tissues are key events in this process [1].
M. Perán :D. ToshCentre for Regenerative Medicine,Department of Biology & Biochemistry, University of Bath,Bath, UK
M. Perán (*)Department of Health Sciences, University of Jaén,Paraje de las Lagunillas sn, B3-212,Jaén 23071, Spaine-mail: [email protected]
J. A. MarchalDepartment of Human Anatomy and Embryology,University of Granada, Granada, Spain
J. A. MarchalBiopathology and Regenerative Medicine Institute (IBIMER),University of Granada, Granada, Spain
M. A. GarcíaResearch Unit, Hospital Universitario Virgen de las Nieves,Granada, Spain
J. Kenyon (*)The Dove Clinic for Integrated Medicine,The Old Brewery, High Street,Twyford SO21 1RG, UKe-mail: [email protected]
Cell Oncol. (2013) 36:289–301DOI 10.1007/s13402-013-0134-8

Currently, the most common treatment modalities for pa-tients with cancers in a metastatic stage are chemotherapyand radiotherapy, but both are hampered by major sideeffects. Consequently, there is an urgent need to developnew approaches to overcome these side effects. The idea ofusing proteases for cancer treatment has already been putforward more than 100 years ago. In 1906, Beard proposedthe use of fresh pancreatic enzyme extracts as a possibletherapeutic approach [2] and, subsequently, found that in-jection of the Jensen’s mouse sarcoma model with trypsinresulted in tumour regression [3, 4]. Also, the treatment ofhuman cancer patients with crude enzyme extracts preparedfrom sheep pancreas appeared to reduce tumour sizes and toextend median survival rates in at least some of the patients(reviewed in [5]). Additional in vivo studies have shownthat the administration of a mixture of enzymes in a meta-static model of syngeneic melanoma B16 mice reduced theformation of metastases and improved the survival rates ofthe treated mice [6], and clinical trials using an oral enzymepreparation containing papain, trypsin and chymotrypsin(formulated to be unaffected by the gastric environment),administered in combination with standard chemotherapy,have shown efficacy in the treatment of patients sufferingfrom breast cancer [7], colorectal cancer [8] and multiplemyeloma [9]. The most relevant outcome of these latterstudies was an improvement in the quality of life throughreducing both the clinical symptoms and the adverse effectsassociated with adjuvant anti-cancer therapy. Additionally, amodest increase in patient survival was reported.
While these observations are encouraging, the cellularmechanisms whereby pancreatic enzymes elicit a beneficialeffect on tumour progression, including metastasis, are asyet poorly understood. One possible explanation could be achange in the expression of cell adhesion molecules [10].Many of the pathways involved in tumour progression ap-pear to converge to a down-regulation of E-cadherin [11]. E-cadherin is known to play an important role in establishingcontacts between epithelial cells and is known to be linkedto the cytoskeleton through associations with α-, β- and γ-catenin [12–14]. Since a general role of cell adhesion mol-ecules in human carcinogenesis is well established [15–17],several efforts have been made to develop strategies tomodulate the E-cadherin/β-catenin complex as a possibletherapeutic approach [18–20]. The clinical benefits of pan-creatic (pro)enzymes may also result from enhanced differ-entiation of tumour cells, which may have repercussions fortheir proliferative capacity [21] and concomitant loss ofmalignant potential [22]. These latter effects also constitutethe main goals of chemo-differentiation therapy regimensfor the treatment of cancer [23, 24].
In the present study, we evaluated the in vitro effects of acombination of pancreatic (pro)enzymes on the humancancer-derived cell lines Caco-2, Panc1 and OE33. We
found, for the first time, that this treatment induces (i) adose-dependent inhibition of cell growth, (ii) an enhancedexpression of epithelial markers and (iii) a decreased ex-pression of EMT markers in the three human carcinoma celllines tested, as well as (iv) differentiation of the Caco-2 andPanc1 cell lines. These observations may have implicationsfor the prevention of cancer metastases and underscore theuse of pancreatic (pro)enzymes for the treatment of cancer.
2 Materials and methods
2.1 Enzymes
The following (pro)enzymes were purchased from Sigma-Aldrich (St. Louis, MO): Trypsinogen (T) and α-Chymotripsinogen (C), both from bovine pancreas origin,and α-Amylase (A) from Bacillus sp. These (pro)enzymeswere dissolved in phosphate buffered saline (PBS, Sigma) toreach stock solutions of 19.5×106 ng/ml for T and C, and4.5×106 ng/ml for A. From these stock solutions a mixturewas made that contained T (6.5×106 ng/ml), C (6.5×106 ng/ml) and A (1.5×106 ng/ml). Experiments wereconducted with the (pro)enzyme mixture diluted in differentconcentrations in culture media (see below). The final con-centrations of the (pro)enzymes were chosen based on liter-ature data and previous clinical experiences [25].
2.2 Cell cultures
The cells lines used were Panc1 (ECACC 87092802), Caco-2 (ECACC 86010202) and OE33 (ECACC 96070808).Panc1 is a pancreatic carcinoma cell line, OE33 anoesophageal carcinoma cell line and Caco-2 a colon carci-noma cell line. All cell lines were routinely maintained inT75 culture flasks at 37 °C in a humidified atmospherecontaining 5 % CO2. The culture media used were:Dulbecco’s Modified Eagle Medium (DMEM) for Panc1,Eagle’s Minimum Essential Medium (EMEM) for Caco-2,and RPMI-1640 Medium for OE33 (Invitrogen, Merelbeke,Belgium). All media were supplemented with 10 % foetalbovine serum (FBS), 100 IU/ml penicillin (Invitrogen),100 μg/ml streptomycin (Invitrogen) and 2 mM L-Glutamine (Sigma).
2.3 Cell growth inhibition assay
After (pro)enzyme treatment, cell numbers were determinedusing the Sulforhodamine B (SRB, Sigma) protein-stainingassay. To this end, cells were seeded at low densities (1,000–3,000 cells per well depending on the cell type) in 24-wellculture plates (day 1). One 24-well culture plate of each cellline was used to test the effect of the (pro)enzymes. This
290 M. Perán et al.

effect was tested by varying the concentrations of the(pro)enzymes in quintuplicate. In each case, another 24-well culture plate was used as control. For this, the samenumbers of cells were seeded and the culture media werechanged as for the treated cells. During the assay the cellswere treated twice, on day 2 and on day 5, whereby theculture media were removed and fresh culture media withthe indicated (pro)enzyme concentrations were added to thecells. On day 7, the cells were fixed with 10 % cold (4 °C)trichloroacetic acid (TCA, Sigma). Following gentle aspira-tion of the culture medium, TCA was gently added to thewells. The culture plates were left for 30 min at 4 °C,washed three times with deionized water and left to dry atroom temperature for at least 24 h. Next, the fixed cells werestained with Sulforhodamine B (SRB). To this end, 1 ml of0.4 % SRB (w/v in 1 % acetic acid solution) was added toeach well and left at room temperature for 20 min. ThenSRB was removed and the plates were washed three timeswith 1 % acetic acid before air drying. Bound SRB wassolubilised with 500 μl of 10 mM Tris-base (Sigma) solu-tion and the plates were put on a plate shaker for at least10 min. Finally, 100 μl solution from each well was trans-ferred to a well of a 96-well plate, in triplicate for each wellof the 24-well plate, and the absorbance was measured at492 nm in a 96-well plate reader (Rainbow Thermo A-5002; TECAN Spectra, Switzerland). The cell number foreach well was calculated using the following equation: %cell number = (absorbance tested cells/absorbance controlcells) × 100.
2.4 Marker expression analysis by flow cytometry
Caco-2, Panc1 and OE33 cells were grown in 25 cm2 flasksand treated twice (on day 2 and day 4) with T (65×103 ng/ml), C (65×103 ng/ml), and A (15×103 ng/ml). Onday 5, the cells were harvested by trypsin treatment andincubated in 10 % FBS-PBS for 15 min at room temperatureto inactivate the trypsin. Following this incubation, the cellswere centrifuged and resuspended in 200 μl 10 % FBS-PBSmonoclonal anti-human-E-cadherin-Phycoerythrin clone180224 (R and D Systems) and incubated for another30 min at 4 °C. Then the cells were washed once in PBSand resuspended in 200 μl PBS for final flow cytometryanalysis. For labelling with the monoclonal anti-human-β-catenin-fluorescein clone 196624 (R and D Systems), thecells were permeabilized with 0.4 % Triton X-100 afterwhich the same procedure was followed as described above.
2.5 Marker expression analysis by immunocytochemistry
Panc1, OE33 and Caco-2 cells were grown on glass cover-slips and treated twice (on day 2 and day 4) with T (65×103 ng/ml), C (65×103 ng/ml) and A (15×103 ng/ml).
Untreated cells were used as controls. On day 5, the cellswere washed three times with PBS and fixed with 4 %paraformaldehyde in PBS for 30 min at room temperature.Next, the cells were washed three times with PBS and post-fixed with ice cold acetone/methanol (1:1) for 5 min at−20 °C. Following this, the cells were washed three timesin PBS and blocked in 2 % blocking buffer solution (Roche)for 1 h at room temperature. Then, the cells were incubatedovernight with primary antibodies (monoclonal mouse anti-h uman β -Ca t e n i n o r an t i - human E - c adhe r i n ,DakoCytomation; clone β-Catenin-1 and clone NCH-38,respectively) diluted 1:50 for anti-β-Catenin and 1:100 foranti-E-cadherin in blocking buffer solution at 4 °C. Next,the cells were washed three times in PBS and incubated for2 h with a secondary antibody (goat anti-mouse IgG (H&L)-FITC conjugated secondary antibody, DakoCytomation) inblocking buffer solution 1:100. Finally, the cells werewashed three times in PBS and coverslips were mountedonto the slides with 4′,6-diamidino-2-phenylindole (DAPI)-containing mounting solution (Ultra CruzTM Mounting Me-dium, Santa Cruz). Photographs were taken using a LeicaDM 5500B (Solms, Germany) fluorescent microscope,equipped with Meta Systems Isis and NIH ImageJ software(National Institutes of Health, MA, USA). The integratedfluorescence intensity for each cell line was determined bycomputing the average fluorescence intensity of the image,subtraction of the background intensity and datanormalization.
2.6 Alkaline phosphatase dye deposition assay
Caco-2 cells (5×104) were seeded in 2 ml complete mediumin 35 mm culture plates (day 1), and treated with T (65×103 ng/ml), C (65×103 ng/ml) and A (15×103 ng/ml) twice(on day 2 and day 4). To assess alkaline phosphatase (ALP)activity in vitro, the cells were rinsed with PBS on day 5 and1 ml Western BlueTM Stabilized Substrate for ALP(Promega) was added for 10 min at 37 °C. Subsequently,the cells were rinsed once with PBS and photomicrographswere taken to visualize the area of dye deposition. Images ofcontrol untreated and treated cells were collected using aNikon Ti-S microscope equipped with a camera (DS-Qi1Mc; Nikon), and figures were collected using the NisElements F 2.3 software package (Nikon, Japan).
2.7 Caco-2 differentiation detection by scanning electronmicroscopy (SEM)
Caco-2 cells were grown on glass coverslips that wereplaced into the wells of a 24-well plate and treated twice(on day 2 and day 4) with T (65×103 ng/ml), C (65×103 ng/ml) and A (15×103 ng/ml). On day 5 the cells werewashed three times with PBS and fixed with 2.5 %
Pancreatic (pro)enzymes promote cell differentiation 291

paraformaldehyde in 0.1 M cacodylate for 1 h at roomtemperature. The cells were post-fixed in 1 % osmiumtetroxide in 0.1 M phosphate buffer in the dark at 4 °C for1 h and then at room temperature for 1 h. Next, the cellswere washed three times with distilled water for 10 min anddehydrated using a gradient of 50 %, 70 %, 90 % and (3×)100 % ethanol. Then, the samples were dried with carbondioxide in a Polaron CPD 7501 critical point drier and,finally, carbon-covered. The samples were examined usinga Zeiss DSM 950 scanning electron microscope.
2.8 Morphological analysis of Caco-2 and Panc-1 cellsby transmission electron microscopy (TEM)
Caco-2 and Panc1 cells were treated twice (on day 2 and day4) with T (65×103 ng/ml), C (65×103 ng/ml) and A (15×103 ng/ml). The cells were fixed on day 5 with 2.5 % glutar-aldehyde in 0.1 M sodium cacodylate buffer (pH 7.4; Sigma)for 1 h at room temperature, post-fixed with 1 % osmiumtetroxide in 0.1 M cacodylate buffer (Sigma) for 1 h at roomtemperature, and dehydrated in ethanol. Next, the cells weredetached from the culture vessels with propyleneoxide andembedded in Epon 812. After polymerization, the plastic wasremoved and ultrathin parallel and perpendicular sectionswere cut. The sections were contrasted with uranyl acetateand lead citrate and examined using a Carl Zeiss EM 902transmission electron microscope (TEM).
2.9 Immunological detection of cytokeratin 8, insulinand glucagon
Caco-2 and Panc1 cells were grown on glass coverslips andtreated without or with T (65×103 ng/ml), C (65×103 ng/ml)and A (15×103 ng/ml) twice (on day 2 and day 4). TheCaco-2 cells were fixed on day 5 with ice coldacetone/methanol (1:1) for 5 min at −20 °C. The Panc1 cellswere fixed on day 7 with 4 % paraformaldehyde in PBS for20 min. Next, the cells were washed three times with PBSand incubated in Citrate Antigen Retrieval solution (Abcam)for 1 h at 37 °C. After this, the cells were washed three timeswith PBS and blocked in 2 % blocking buffer solution(Roche) for 1 h at room temperature. Next, the cells wereincubated overnight at 4 °C with primary antibodies diluted1:100 in blocking buffer. The primary antibodies used weremonoclonal Troma I anti-CK8 (DSHB, University of Iowa)for the Caco-2 cells and guinea-pig anti-insulin whole anti-serum (Sigma) and mouse monoclonal anti-glucagon cloneK79bB10 (Sigma) for the Panc1 cells. Next, the cells werewashed three times in PBS and incubated for another 2 hwith secondary (H&L) FITC or TRIC conjugated antibodies(DakoCytomation) diluted 1:100 in blocking buffer solu-tion. After this, the cells were washed three times in PBSand incubated for 15 min in a 1:1000 dilution of DAPI in
PBS at room temperature. Coverslips were mounted onslides with DAPI-containing mounting solution (UltraCruzTM Mounting Medium, Santa Cruz). Photographs weretaken using a Leica DM 5500B fluorescent microscopeequipped with MetaSystems software.
2.10 RNA isolation and real-time PCR analysis
Real-time PCR was performed to assess the effect of(pro)enzyme treatment on the EMT markers E-cadherin,Vimentin, Snail and Slug. OE33, Panc1 and Caco-2 cells (5×104) were seeded in 2 ml complete medium in 35 mm cultureplates (day 1), and treated with T (65×103 ng/ml), C (65×103 ng/ml) and A (15×103 ng/ml) trice (on day 2, day 4 andday 6). The cells were collected on day 7. Total cellular RNAwas isolated using QIAcube equipment (QIAGEN) in con-junction with the miRNeasy Mini Kit (QIAGEN). TheQuantiTect Reverse Transcription Kit (QIAGEN) was usedfor cDNA synthesis. Real-time PCR was performed using theKAPA SYBR Fast qPCR Kit (KAPABIOSYSTEMS) in con-junction with an Eco Illumina System (Illumina). Sequencesof the primers used were: 5 ′GGACTCATGACCACAGTCCATGCC3′ and 5′TCAGGGATGACCTTGCCCACAG3′ for GAPDH, 5′GGAACTATGAAAAGTGGGCTTG3′ and 5′AAATTGCCAGGCTCAATGAC3′ for E-cadherin, 5′GTTTCCCCTAAACCGCTAGG3′ and 5′AGCGAGAGTGGCAGAGGA3′ for Vimentin, 5′CTTCGGCTCCAGGAGAGTC3′ and 5′TTCCCACTGTCCTCATCTGAC3 ′ for Snail and 5 ′TGGTTGCTTCAAGGACACAT3′ and 5′GTTGCAGTGAGGGCAAGAA3′ forSlug. The expression levels were normalized to correspondingfor GAPDH values and are shown as fold change relative tothe value of the control sample. All the analyses were carriedout in triplicate.
2.11 Statistical analysis
All analyses were performed at least in triplicate in each ofat least three independent experiments, except for the SEMand TEM analyses that were performed twice. Statisticalanalyses were performed using a one-way-ANOVA test(two-sided) and Fisher post-hoc comparison; p-values lessthan 0.05 were considered statistically significant. TheStatistica 6.0 Program was used to analyse the results.
3 Results
3.1 Effects of (pro)enzymes on Caco-2, Panc1 and OE33cellular growth
Initial experiments were performed to determine the effectsof the (pro)enzymes Trypsinogen (T), α-Chymotripsinogen
292 M. Perán et al.
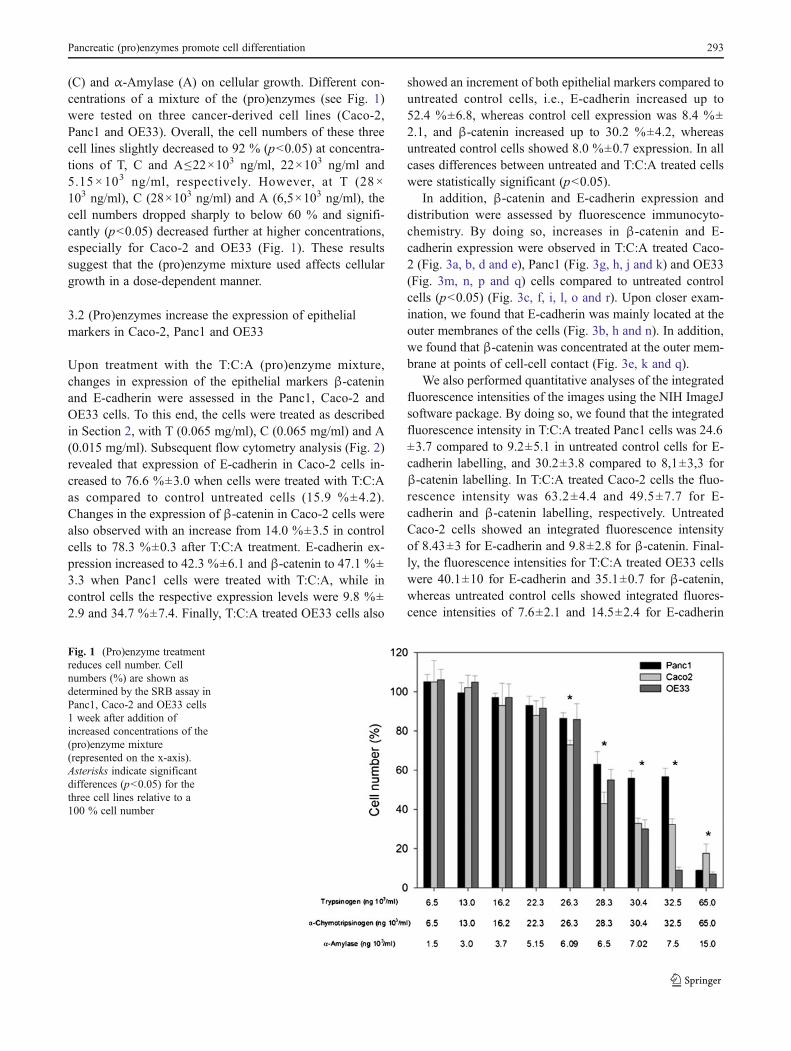
(C) and α-Amylase (A) on cellular growth. Different con-centrations of a mixture of the (pro)enzymes (see Fig. 1)were tested on three cancer-derived cell lines (Caco-2,Panc1 and OE33). Overall, the cell numbers of these threecell lines slightly decreased to 92 % (p<0.05) at concentra-tions of T, C and A≤22×103 ng/ml, 22×103 ng/ml and5.15×103 ng/ml, respectively. However, at T (28×103 ng/ml), C (28×103 ng/ml) and A (6,5×103 ng/ml), thecell numbers dropped sharply to below 60 % and signifi-cantly (p<0.05) decreased further at higher concentrations,especially for Caco-2 and OE33 (Fig. 1). These resultssuggest that the (pro)enzyme mixture used affects cellulargrowth in a dose-dependent manner.
3.2 (Pro)enzymes increase the expression of epithelialmarkers in Caco-2, Panc1 and OE33
Upon treatment with the T:C:A (pro)enzyme mixture,changes in expression of the epithelial markers β-cateninand E-cadherin were assessed in the Panc1, Caco-2 andOE33 cells. To this end, the cells were treated as describedin Section 2, with T (0.065 mg/ml), C (0.065 mg/ml) and A(0.015 mg/ml). Subsequent flow cytometry analysis (Fig. 2)revealed that expression of E-cadherin in Caco-2 cells in-creased to 76.6 %±3.0 when cells were treated with T:C:Aas compared to control untreated cells (15.9 %±4.2).Changes in the expression of β-catenin in Caco-2 cells werealso observed with an increase from 14.0 %±3.5 in controlcells to 78.3 %±0.3 after T:C:A treatment. E-cadherin ex-pression increased to 42.3 %±6.1 and β-catenin to 47.1 %±3.3 when Panc1 cells were treated with T:C:A, while incontrol cells the respective expression levels were 9.8 %±2.9 and 34.7 %±7.4. Finally, T:C:A treated OE33 cells also
showed an increment of both epithelial markers compared tountreated control cells, i.e., E-cadherin increased up to52.4 %±6.8, whereas control cell expression was 8.4 %±2.1, and β-catenin increased up to 30.2 %±4.2, whereasuntreated control cells showed 8.0 %±0.7 expression. In allcases differences between untreated and T:C:A treated cellswere statistically significant (p<0.05).
In addition, β-catenin and E-cadherin expression anddistribution were assessed by fluorescence immunocyto-chemistry. By doing so, increases in β-catenin and E-cadherin expression were observed in T:C:A treated Caco-2 (Fig. 3a, b, d and e), Panc1 (Fig. 3g, h, j and k) and OE33(Fig. 3m, n, p and q) cells compared to untreated controlcells (p<0.05) (Fig. 3c, f, i, l, o and r). Upon closer exam-ination, we found that E-cadherin was mainly located at theouter membranes of the cells (Fig. 3b, h and n). In addition,we found that β-catenin was concentrated at the outer mem-brane at points of cell-cell contact (Fig. 3e, k and q).
We also performed quantitative analyses of the integratedfluorescence intensities of the images using the NIH ImageJsoftware package. By doing so, we found that the integratedfluorescence intensity in T:C:A treated Panc1 cells was 24.6±3.7 compared to 9.2±5.1 in untreated control cells for E-cadherin labelling, and 30.2±3.8 compared to 8,1±3,3 forβ-catenin labelling. In T:C:A treated Caco-2 cells the fluo-rescence intensity was 63.2±4.4 and 49.5±7.7 for E-cadherin and β-catenin labelling, respectively. UntreatedCaco-2 cells showed an integrated fluorescence intensityof 8.43±3 for E-cadherin and 9.8±2.8 for β-catenin. Final-ly, the fluorescence intensities for T:C:A treated OE33 cellswere 40.1±10 for E-cadherin and 35.1±0.7 for β-catenin,whereas untreated control cells showed integrated fluores-cence intensities of 7.6±2.1 and 14.5±2.4 for E-cadherin
Fig. 1 (Pro)enzyme treatmentreduces cell number. Cellnumbers (%) are shown asdetermined by the SRB assay inPanc1, Caco-2 and OE33 cells1 week after addition ofincreased concentrations of the(pro)enzyme mixture(represented on the x-axis).Asterisks indicate significantdifferences (p<0.05) for thethree cell lines relative to a100 % cell number
Pancreatic (pro)enzymes promote cell differentiation 293

and β-catenin, respectively. Together, these results showand increment in the fluorescence intensities of T:C:A
treated cells, labelled with specific antibodies, as comparedto untreated control cells (p<0.05).
Fig. 2 Flow cytometry analysis of E-cadherin and β-catenin expres-sion in control untreated and (pro)enzyme treated Caco-2, Panc1 andOE33 cells. Numbers indicate the percentages of cells labelled withanti-E cadherin and anti- β-catenin antibodies. As a control, isotype
antibody labelling is shown. All data were obtained from three inde-pendent experiments and are expressed as mean ± SE. Significantdifferences (p <0.05) were observed between control and treated cells
Fig. 3 Epithelial marker expression in Caco-2, Panc2 and OE33 cellstreated with (pro)enzymes. Cell surface E-cadherin and β-cateninexpression was detected (in green) in fixed and permeabilized cells,respectively, and nuclei were counterstained with DAPI (blue). Themerged images show the membrane distribution of these markers,
which is significantly stronger as compared to untreated cells incubatedwith the same antibodies. Representative pictures from three indepen-dent experiments are shown. The original magnification was 40× forall the panels, except b, e, h, k, n and q (63×)
294 M. Perán et al.

3.3 (Pro)enzyme treatment induces differentiationin Caco-2 cells
We used Caco-2 cells as a model of cell differentiation andtested whether treatment with the (pro)enzymes could in-duce these cells to acquire an enterocyte-like phenotype.Confluent Caco-2 cell monolayers (Fig. 4a) were treatedwith the T (65×103 ng/ml), C (65×103 ng/ml) and A (15×103 ng/ml) combination. After 24 h of treatment the cellsstarted to form ring-like structures (Fig. 4b). Closer exami-nation of these cells revealed that the ring-like structureswhere surrounded by multicellular bands in which the cellswere elongated and had acquired a cord-like structure(Fig. 4c). Next, we investigated the activity of alkalinephosphatase (ALP) in these cells, which is a marker for
the differentiation of intestinal epithelial cells. ALP activitywas observed in Caco-2 cells after treatment with the(pro)enzyme mixture, and patches of ALP dye deposits werefound to be associated with the ring-like multi-cellularstructures (Fig. 4e). In contrast, untreated Caco-2 cells onlyshowed a slight blue labelling, which may be attributed tobackground staining (Fig. 4d). In addition, T:C:A treatedCaco-2 cells showed microvilli-like structures (Fig. 4f).
The morphological changes occurring in T:C:A treatedCaco-2 cells were analysed in further detail by scanningelectron microscopy (SEM). Untreated Caco-2 cellsscarcely showed microvilli and an irregular morphology(Fig. 4g). In contrast, Caco-2 cells treated with the(pro)enzyme mixture showed a more regular polygonalmorphology appearing as a homogenous carpet of microvilli
Fig. 4 Caco-2 cells treatedwith (pro)enzymes exhibitlineage-specific differentiation.Phase-contrast microscopyshowing highly confluentmonolayers of untreated (a andd) and treated (b, c, e and f)Caco-2 cells. In (b) and (c) thetreated cells exhibit ring-likestructures. In (d–f) the untreated(d) and treated (e and f) cellswere stained with the WesternBlueTM Stabilized Substratefor alkaline phosphatase (ALP).In (f) a detail is shown of (e) inwhich the arrows point atstructures with a microvillusappearance. Scanning electronmicrographs of untreated (g)and treated (h–j) cells showerected microvilli. In (j) anenlargement of (i) is shown.Representative pictures fromtwo independent experimentsare shown. Originalmagnifications: 20× (a, b ande), 40× (d), 63× (c) and 100× (f)
Pancreatic (pro)enzymes promote cell differentiation 295

on the cell surface (Figs. 4h and i). These features aretypical of end-stage differentiated cells. Upon closer obser-vation, these cells showed well-developed brush bordermicrovilli (Fig. 4j).
The ultrastructural analyses of the cells were completedby TEM, by which Caco-2 cells treated with the T:C:Acombination indeed showed a dramatic increment in thenumber of microvilli (Fig. 5b) as compared to untreatedcontrol cells (Fig. 5a). T:C:A treated cells also showed theappearance of desmosomes and tight junctions (Fig. 5c andd), structures that were not evident in untreated control cells.In addition, we observed an increase in glycogen accumu-lation in the T:C:A treated cells and an increased network of
intermediate filaments (Fig. 5c and d). Together, these ob-servations indicate that the pancreatic (pro)enzymes testedare able to induce lineage-specific Caco-2 differentiation.
To assess whether the observed morphological changesafter T:C:A treatment were accompanied by modifications inthe expression of the intermediate filament network, untreatedand (pro)enzyme-treated Caco-2 cells were immunostainedwith an anti-cytokeratin 8 (CK8) antibody. The T:C:A treatedcells showed an increment in fluorescence intensity, preciselyin the cells that formed the periphery of the ring-like structures(Fig. 6a, c, d, and f) as compared to untreated control cells(Fig. 6g and i). This result indicates that CK8 expressionincreases upon pancreatic (pro)enzymes treatment.
Fig. 5 Scanning electronmicrographs of untreated (a)and treated (b–d) Caco-2 cells.In (b) numerous microvilli canbe seen. Panels (c) and (d) showthe appearance of tightjunctions (arrowheads),desmosomes (arrows), theincrement in glycogendepositions (asterisk) andintermediate filaments (IF) of(pro)enzyme treated cells.Representative pictures fromtwo independent experimentsare shown
Fig. 6 Cytokeratin8 expression in treated anduntreated Caco-2 cells.Confluent monolayers of Caco-2 cells were treated with(pro)enzymes, fixed andimmunostained for cytokeratin8 (CK8) expression (green).Nuclei were counterstainedwith DAPI (blue). An increasedexpression of CK8 is observedin cells that form the ring-likestructures in treated cells (a–f)as compared to untreated cells(g–i). Representative picturesfrom three independentexperiments are shown. (c, fand i): merged images of (a, b),(d, e) and (g, h), respectively.Original magnifications: 20×(a–c) and 40× (d–i)
296 M. Perán et al.

3.4 (Pro)enzyme treatment induces aggregate formationand differentiation in Panc1 cells
Panc1 cells proliferate in an adherent monolayer composed ofmore or less homogeneous and flattened cells. After T:C:Atreatment of confluent Panc1 cells (Fig. 7a), they were foundto cluster and migrate to form rounded cells aggregates(Fig. 7b, c, e and f). In contrast, confluent untreated cells didnot form any cell aggregates (Fig. 7d). These observationsprompted us to analyse the morphological changes inuntreated and T:C:A treated Panc1 cells in further detail byelectron microscopy. By doing so, we observed an enhancedaccumulation of lamellar bodies, microvilli and tight junctionsin T:C:A treated cells (Fig. 8b–d) as compared to untreatedcontrol Panc1 cells (Fig. 8a). These results suggest the acqui-sition, by at least some of the cells, of an exocrine
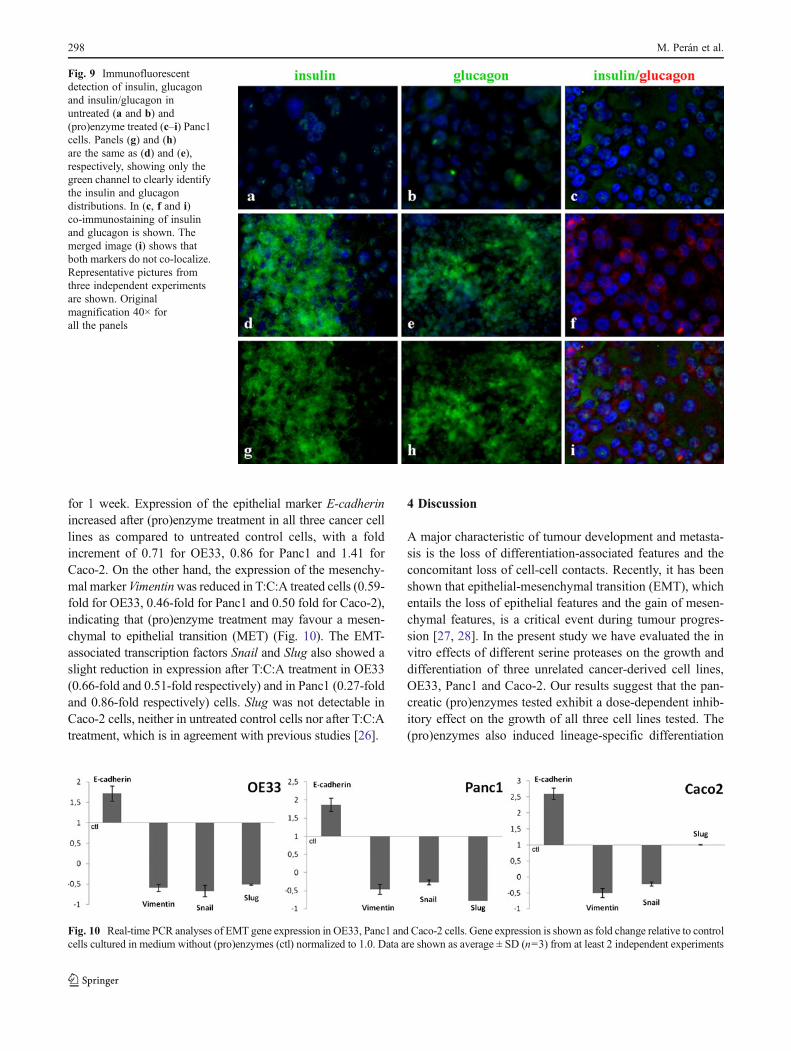
differentiated phenotype. In addition, we observed an incre-ment in the number of microvillus prolongations on the mem-branes of the cells exposed to the enzymatic mixture. Finally,we performed immunological analyses of the expression ofendocrine cell markers to further characterise the level ofdifferentiation of Panc1 cells after 7 days of T:C:A treatment(Fig. 9c–i). Our results show a significant increase in thenumber of insulin and glucagon expressing cells, as comparedto untreated control cells (Fig. 9a–b).
3.5 (Pro)enzyme treatment attenuates EMT gene expressionin OE33, Panc1 and Caco-2 cells
We further analyzed the expression of EMT-associatedmarkers in OE33, Panc1 and Caco-2 cells after treatment withT (65×103 ng/ml), C (65×103 ng/ml) and A (15×103 ng/ml)
Fig. 7 (Pro)enzyme treatmentinduces aggregation of Panc1cells. Untreated cells (a and d)are evenly distributed in amonolayer culture, whereastreated cells (b, c, e and f)cluster and form aggregates.Representative pictures fromthree independent experimentsare shown. Originalmagnifications: 20× (a–c), 40×(d and e) and 63× (f)
Fig. 8 Ultrastructure of(pro)enzyme treated Panc1 cellsas revealed by TEM. Untreatedcells are shown in (a). Thetreated cells exhibit anenhanced accumulation oflamellar bodies (arrows),microvilli (asterisk) and tightjunctions (arrowhead) (b–d). In(d) a magnification is shown ofthe lamellar bodies found in thecell shown in (b).Representative pictures fromtwo independent experimentsare shown
Pancreatic (pro)enzymes promote cell differentiation 297
for 1 week. Expression of the epithelial marker E-cadherinincreased after (pro)enzyme treatment in all three cancer celllines as compared to untreated control cells, with a foldincrement of 0.71 for OE33, 0.86 for Panc1 and 1.41 forCaco-2. On the other hand, the expression of the mesenchy-mal markerVimentinwas reduced in T:C:A treated cells (0.59-fold for OE33, 0.46-fold for Panc1 and 0.50 fold for Caco-2),indicating that (pro)enzyme treatment may favour a mesen-chymal to epithelial transition (MET) (Fig. 10). The EMT-associated transcription factors Snail and Slug also showed aslight reduction in expression after T:C:A treatment in OE33(0.66-fold and 0.51-fold respectively) and in Panc1 (0.27-foldand 0.86-fold respectively) cells. Slug was not detectable inCaco-2 cells, neither in untreated control cells nor after T:C:Atreatment, which is in agreement with previous studies [26].
4 Discussion
A major characteristic of tumour development and metasta-sis is the loss of differentiation-associated features and theconcomitant loss of cell-cell contacts. Recently, it has beenshown that epithelial-mesenchymal transition (EMT), whichentails the loss of epithelial features and the gain of mesen-chymal features, is a critical event during tumour progres-sion [27, 28]. In the present study we have evaluated the invitro effects of different serine proteases on the growth anddifferentiation of three unrelated cancer-derived cell lines,OE33, Panc1 and Caco-2. Our results suggest that the pan-creatic (pro)enzymes tested exhibit a dose-dependent inhib-itory effect on the growth of all three cell lines tested. The(pro)enzymes also induced lineage-specific differentiation
Fig. 9 Immunofluorescentdetection of insulin, glucagonand insulin/glucagon inuntreated (a and b) and(pro)enzyme treated (c–i) Panc1cells. Panels (g) and (h)are the same as (d) and (e),respectively, showing only thegreen channel to clearly identifythe insulin and glucagondistributions. In (c, f and i)co-immunostaining of insulinand glucagon is shown. Themerged image (i) shows thatboth markers do not co-localize.Representative pictures fromthree independent experimentsare shown. Originalmagnification 40× forall the panels
Fig. 10 Real-time PCR analyses of EMT gene expression in OE33, Panc1 and Caco-2 cells. Gene expression is shown as fold change relative to controlcells cultured in medium without (pro)enzymes (ctl) normalized to 1.0. Data are shown as average ± SD (n=3) from at least 2 independent experiments
298 M. Perán et al.

in the colon (Caco-2) and pancreas (Panc1) cancer-derivedcell lines.
Caco-2 cells, that may serve as a model of small intestinalcells [29], display an undifferentiated phenotype under nor-mal culture conditions, with an irregular morphology andscarce microvilli [21]. However, when cultured under post-confluent conditions [30] or following treatment with sodi-um butyrate [31], the cells display differentiated character-istics. Here we show that when Caco-2 cells are treated withthe (pro)enzyme mixture they acquire a more differentiated‘enterocyte-like’ phenotype, and form ring-like structures,characteristic of a multilayered disposition of cells [32].These morphological changes were found to be accompa-nied by an up-regulation of several differentiation markers,including alkaline phosphatase, cytokeratin 8, and E-cadherin, and the appearance of numerous tight junctionsat the EM level [33–35]. Tight junctions represent apicalconstituents of the intercellular junctional complexes inmammalian epithelial cell sheets where they form a semi-permeable barrier for the paracellular transport of ions andsolutes [36]. Since EMT is accompanied by the loss of cell-cell adhesion through the disintegration of tight, adherentsand gap junctions [11, 37], our results suggest that the(pro)enzyme treatment may reduce the conversion from anepithelial to a mesenchymal phenotype and, as such, mayreduce the metastatic potential of the tumour cells. Addi-tional work is required to test this possibility e.g. by usingan EMT inducer to assess if the (pro)enzymes induce areduction in EMT.
A differentiated phenotype of Panc1 cells was obtainedfollowing (pro)enzyme treatment. In agreement with a pre-vious study [38], we found that endocrine differentiation toislet-like clusters was accompanied by aggregation of Panc1cells to form spherical clusters. In addition, we observed amarked increment of lamellar bodies and microvilli forma-tion in some of the cells after (pro)enzyme treatment,suggesting the acquisition of an exocrine phenotype. Wedid, however, not detect any expression of digestive en-zymes. A phenotypic variability of Panc1 cells has beendescribed before [39]. Given this variability, it is possiblethat the (pro)enzyme treatment may induce a sub-populationof undifferentiated cells to undergo exocrine differentiation,while another sub-population may follow the endocrinepathway. Taken together, the morphological and structuralspecializations of Caco-2 and Panc1 cells exposed to pan-creatic (pro)enzyme treatment could represent a first mani-festation of a post-proliferative differentiated cell state. Inthe past, differentiation therapy has already been used as analternative to cytotoxic therapy and appears to offer prospectsas a useful therapeutic tool in cancer treatment [23, 24, 40].
E-cadherin and β-catenin complexes are widely regardedas suppressors of tumourigenesis and metastasis [15], andloss of their expression has been associated with a reduction
or loss of cell-cell adhesion and differentiated properties[41]. The gene encoding E-cadherin (CDH1, on chromo-some 16q22.1) [42] is considered to be a metastatic tumoursuppressor gene. Decreased E-cadherin expression is widelyencountered in cancers, including prostate cancer, lung can-cer, breast cancer, gastric cancer and colorectal cancer [16].Previously, it was reported that silencing of the CDH1 geneby aberrant promoter hypermethylation occurs in leukae-mias and undifferentiated solid tumours, such as gastriccarcinomas [43]. In addition, a correlation has been ob-served between reduced levels of E-cadherin expressionand pathological features such as poor differentiation,infiltrative growth, lymph node metastasis and decreasedpatient survival [44, 45]. Loss of E-cadherin results inprogression from a benign adenoma to an invasive tumour,and has been associated with a poor clinical prognosis [46,47]. β-catenin is also involved in cell adhesion throughbinding E-cadherin to the actin cytoskeleton, and is a criticalelement in the canonical Wnt signalling pathway [48]. Re-duced membranous β-catenin expression has been associat-ed with a poor prognosis in gastric carcinoma, whereaslonger postoperative survival rates have been noted forcarcinomas exhibiting high membranous β-catenin expres-sion levels [41]. Our data revealed increased expressionlevels of both E-cadherin and β-catenin, including a mem-branous β-catenin distribution, after (pro)enzyme treatment,suggesting that the main effects of these serine proteases are(i) differentiation induction and (ii) metastasis suppressionin cancer cells. The inverse relationship between prolifera-tion and differentiation implies an alternative approach tocancer therapy that does not primarily involve cell killingbut, instead, induction of malignant cells to differentiate tobenign forms with a reduced proliferative potential [49].Our data indicate that pancreatic (pro)enzymes may enforcethe re-entry of tumour cells into the normal pathway(s) ofdifferentiation and, as such, may comprise a new category ofexperimental cancer differentiation agents.
Finally, we found that the expression of the EMT-associated transcription factors Snail and Slug [50] wasreduced after pancreatic (pro)enzyme treatment. These datasuggest that the (pro)enzyme mixture tested may inducesuppression of the EMT program, which is accompaniedby an increased expression of E-cadherin (both at the proteinand mRNA level) and a modest, but significant, decrease inSnail and Slug expression. Interestingly, it has been reportedby others that over-expression of Snail and Slug cause EMT,which is associated with loss of E-cadherin expression [51,52]. Accordingly, over-expression of Slug in OE33 cellsrepresses E-cadherin and enhances vimentin and fibronectinexpression [53]. Similarly, it has been found that over-expression of Slug in Panc1 cells increases invasion andmetastasis and accelerates pancreatic cancer progressionin an in vivo mouse model [54]. The EMT program also
Pancreatic (pro)enzymes promote cell differentiation 299

involves the induction of mesenchymal protein expression,including vimentin [55]. Here, we observed an increase inE-cadherin expression with a concomitant decrease invimentin expression, which suggests that pancreatic(pro)enzymes may act by deregulating EMT progressionand suppressing tumorigenesis.
In conclusion, we show here that the pancreatic(pro)enzyme mixture tested induces enhanced cell-cell ad-hesion in different cancer-derived cell lines. This changemay promote the acquisition of a less malignant phenotypein addition to a decrease in proliferation due to lineage-specific cellular differentiation. Treatment of patients suffer-ing from malignant diseases with a mixture of (pro)enzymesmay represent a new therapeutic approach.
Acknowledgments Macarena Peran was supported by a grant fromJaen University, Spain (“Plan de Apoyo a la Investigación, DesarrolloTecnológico e Innovación . IV. Programa de Ayudas a losInvestigadores”). Work in the laboratory of Bath University wasfunded by Propanc Pty Ltd. Work in the laboratory of J.A. Marchalat Granada University was funded by the Instituto de Salud Carlos III(Fondo de Investigación Sanitaria, FEDER, grant number PI10/02295).
Conflict of interest Dr Julian Kenyon is the Founder and ScientificDirector of Propanc Pty Ltd and owns stock in the company. PropancPty Ltd and the University of Bath have filed a joint patent from thescientific research undertaken in this report
References
1. P.A. Andreasen, L. Kjoller, L. Christensen, M.J. Duffy, Theurokinase-type plasminogen activator system in cancer metastasis:a review. Int. J. Cancer 72, 1–22 (1997)
2. J. Beard, The action of trypsin upon the living cells of jensen’smouse-tumour: A preliminary note upon a research made (with aGrant from the Carnegie Trust). Br. Med. J. 1, 140–1 (1906)
3. R.W. Moss, An annotated bibliography of works by John Beard.Integr. Cancer Ther. 7, 317–21 (2008)
4. J. Beard, The cancer problem. Lancet 168, 281–283 (1905)5. J.F. Novak, F. Trnka, Proenzyme therapy of cancer. Anticancer.
Res. 25, 1157–77 (2005)6. M. Wald, T. Olejar, V. Sebkova, M. Zadinova, M. Boubelik, P.
Pouckova, Mixture of trypsin, chymotrypsin and papain reducesformation of metastases and extends survival time of C57Bl6 micewith syngeneic melanoma B16. Cancer Chemother. Pharmacol. 47,16–22 (2001)
7. J. Beuth, B. Ost, A. Pakdaman, E. Rethfeldt, P.R. Bock, J. Hanischet al., Impact of complementary oral enzyme application on thepostoperative treatment results of breast cancer patients–results ofan epidemiological multicentre retrolective cohort study. CancerChemother. Pharmacol. 47, 45–54 (2001)
8. T. Popiela, J. Kulig, J. Hanisch, P.R. Bock, Influence of a comple-mentary treatment with oral enzymes on patients with colorectalcancers–an epidemiological retrolective cohort study. CancerChemother. Pharmacol. 47, 55–63 (2001)
9. A. Sakalova, P.R. Bock, L. Dedik, J. Hanisch, W. Schiess, S.Gazova et al., Retrolective cohort study of an additive therapywith an oral enzyme preparation in patients with multiple myelo-ma. Cancer Chemother Pharmacol 47, 38–44 (2001)
10. W.J. Nelson, Regulation of cell-cell adhesion by the cadherin-catenin complex. Biochem. Soc. Trans. 36, 149–55 (2008)
11. H. Peinado, D. Olmeda, A. Cano, Snail, Zeb and bHLH factors intumour progression: an alliance against the epithelial phenotype?Nat. Rev. Cancer 7, 415–28 (2007)
12. R. Kemler, From cadherins to catenins: cytoplasmic protein in-teractions and regulation of cell adhesion. Trends. Genet. 93, 17–21 (1993)
13. M. Conacci-Sorrell, J. Zhurinsky, A. Ben-Ze’ev, The cadherin-catenin adhesion system in signaling and cancer. J. Clin. Invest.109, 987–91 (2002)
14. B.M. Gumbiner, P.D. McCrea, Catenins as mediators of the cyto-plasmic functions of cadherins. J. Cell Sci. Suppl. 17, 155–158(1993)
15. B. Zhai, H.X. Yan, S.Q. Liu, L. Chen, M.C. Wu, H.Y. Wang,Reduced expression of P120 catenin in cholangiocarcinoma corre-lated with tumor clinicopathologic parameters. World J.Gastroenterol. 14, 3739–44 (2008)
16. E. Margineanu, C.E. Cotrutz, C. Cotrutz, Correlation between E-cadherin abnormal expressions in different types of cancer and theprocess of metastasis. Rev. Med. Chir. Soc. Med. Nat. Iasi. 112,432–6 (2008)
17. I. Molina-Ortiz, R.A. Bartolome, P. Hernandez-Varas, G.P. Colo, J.Teixido, Overexpression of E-cadherin on melanoma cells inhibitschemokine-promoted invasion involving p190RhoGAP/p120ctn-dependent inactivation of RhoA. J. Biol. Chem. 284, 15147–57(2009)
18. R.W. Pang, R.T. Poon, From molecular biology to targeted thera-pies for hepatocellular carcinoma: the future is now. Oncology 72,30–44 (2007)
19. S. Tommasi, R. Pinto, B. Pilato, A. Paradiso, Molecular pathwaysand related target therapies in liver carcinoma. Curr. Pharm. Des.13, 3279–87 (2007)
20. S. Boyault, D.S. Rickman, A. de Reynies, C. Balabaud, S.Rebouissou, E. Jeannot et al., Transcriptome classification ofHCC is related to gene alterations and to new therapeutic targets.Hepatology 45, 42–52 (2007)
21. T. Parasassi, R. Brunelli, L. Bracci-Laudiero, G. Greco, A.C.Gustafsson, E.K. Krasnowska et al., Differentiation of normaland cancer cells induced by sulfhydryl reduction: biochemicaland molecular mechanisms. Cell Death Differ. 12, 1285–96 (2005)
22. J.C. Fleet, L. Wang, O. Vitek, B.A. Craig, H.J. Edenberg, Geneexpression profiling of Caco-2 BBe cells suggests a role for spe-cific signaling pathways during intestinal differentiation. Physiol.Genomics 13, 57–68 (2003)
23. S. Sell, Stem cell origin of cancer and differentiation therapy. Crit.Rev. Oncol. Hematol. 51, 1–28 (2004)
24. R.L. Eckert, J.F. Crish, T. Efimova, S. Balasubramanian, Antiox-idants regulate normal human keratinocyte differentiation.Biochem. Pharmacol. 68, 1125–31 (2004)
25. J. Leipner, R. Saller, Systemic enzyme therapy in oncology: Effectand mode of action. Drugs 59, 769–80 (2000)
26. E.R. Camp, V.J. Findlay, S.G. Vaena, J. Walsh, D.N. Lewin, D.P.Turner et al., Slug expression enhances tumor formation in anoninvasive rectal cancer model. J. Surg. Res. 170(1), 56–63(2011)
27. P.B. Gupta, T.T. Onder, G. Jiang, K. Tao, C. Kuperwasser, R.A.Weinberg et al., Identification of selective inhibitors of cancer stemcells by high-throughput screening. Cell 138, 645–59 (2009)
28. A. Voulgari, A. Pintzas, Epithelial-mesenchymal transition in can-cer metastasis: mechanisms, markers and strategies to overcomedrug resistance in the clinic. Biochim. Biophys. Acta 1796, 75–90(2009)
29. J. Fogh, J.M. Fogh, T. Orfeo, One hundred and twenty-sevencultured human tumor cell lines producing tumors in nude mice.J. Natl. Cancer Inst. 59, 221–6 (1977)
300 M. Perán et al.

30. M. Pinto, S. Robineleon, M.D. Appay, M. Kedinger, N. Triadou,E. Dussaulx et al., Enterocyte-like differentiation and polarizationof the human-colon carcinoma cell-line caco-2 in culture. Biol.Cell 47, 323–330 (1983)
31. M.D. Basson, G.A. Turowski, Z. Rashid, F. Hong, J.A. Madri,Regulation of human colonic cell line proliferation and phenotypeby sodium butyrate. Dig. Dis. Sci. 41, 1989–93 (1996)
32. S.R. Wood, Q. Zhao, L.H. Smith, C.K. Daniels, Altered morphol-ogy in cultured rat intestinal epithelial IEC-6 cells is associatedwith alkaline phosphatase expression. Tissue Cell 35, 47–58(2003)
33. S.K. Kurdistani, P. Arizti, C.L. Reimer, M.M. Sugrue, S.A.Aaronson, S.W. Lee, Inhibition of tumor cell growth by RTP/rit42 and its responsiveness to p53 and DNA damage. CancerRes. 58, 4439–44 (1998)
34. J.P. Brunet, N. Jourdan, J. Cotte-Laffitte, C. Linxe, M. Geniteau-Legendre, A. Servin et al., Rotavirus infection induces cytoskele-ton disorganization in human intestinal epithelial cells: implicationof an increase in intracellular calcium concentration. J. Virol. 74,10801–6 (2000)
35. R.J. Guang, J.L. Ford, Y.N. Fu, Y.Z. Li, L.M. Shaw, A.B. Pardee,Drg-1 as a differentiation-related, putative metastatic suppressorgene in human colon cancer. Cancer Res. 60, 749–755 (2000)
36. J.M. Anderson, Molecular structure of tight junctions and their rolein epithelial transport. News Physiol. Sci. 16, 126–30 (2001)
37. Y. Kang, J. Massague, Epithelial-mesenchymal transitions: Twistin development and metastasis. Cell 118, 277–9 (2004)
38. A.A. Hardikar, B. Marcus-Samuels, E. Geras-Raaka, B.M. Raaka,M.C. Gershengorn, Human pancreatic precursor cells secreteFGF2 to stimulate clustering into hormone-expressing islet-likecell aggregates. Proc. Natl. Acad. Sci. U. S. A. 100, 7117–22(2003)
39. Y. Wu, J. Li, S. Saleem, S.P. Yee, A.A. Hardikar, R. Wang, c-Kit andstem cell factor regulate PANC-1 cell differentiation into insulin- andglucagon-producing cells. Lab. Invest. 90(1373–84) (2010)
40. J.A. Marchal, J. Prados, J. Campos, F. González, C. Melguizo, C.Velez et al., Therapeutic potential of differentiation in cancer andnormal stem cells, in New Cell Differentiation Research Topics, ed.by H. Saitama (Nova Science Publisher, Inc, New York, 2008), pp.7–77
41. J. Czyzewska, K. Guzinska-Ustymowicz, M. Ustymowicz, A.Pryczynicz, A. Kemona, The expression of E-cadherin-catenincomplex in patients with advanced gastric cancer: role in formationof metastasis. Folia Histochem. Cytobiol. 48, 37–45 (2010)
42. I.J. Chalmers, M. Aubele, E. Hartmann, E. Braungart, M. Werner,H. Hofler, M.J. Atkinson, Mapping the chromosome 16 cadheringene cluster to a minimal deleted region in ductal breast cancer.Cancer Genet. Cytogenet. 126, 39–44 (2001)
43. W. Feng, R. Orlandi, N. Zhao, M.L. Carcangiu, E. Tagliabue, J. Xuet al., Tumor suppressor genes are frequently methylated in lymphnode metastases of breast cancers. B.M.C. Cancer 10(378–88)(2010)
44. M.T. Debies, D.R. Welch, Genetic basis of human breast cancermetastasis. J. Mammary Gland. Biol. Neoplasia 6, 441–51 (2001)
45. R.B. Hazan, R. Qiao, R. Keren, I. Badano, K. Suyama, Cadherinswitch in tumor progression. Ann. N. Y. Acad. Sci. 1014, 155–63(2004)
46. Y. Uchikado, H. Okumura, S. Ishigami, T. Setoyama, M.Matsumoto, T. Owaki et al., Increased Slug and decreased E-cadherin expression is related to poor prognosis in patients withgastric cancer. Gastric Cancer 14, 41–9 (2011)
47. Y. Li, C.Q. Chen, Y.L. He, S.R. Cai, D.J. Yang, W.L. He et al.,Abnormal expression of E-cadherin in tumor cells is associatedwith poor prognosis of gastric carcinoma. J. Surg. Oncol. 106(3),304–10 (2012)
48. X. Tian, Z. Liu, B. Niu, J. Zhang, T.K. Tan, S.R. Lee et al., E-cadherin/beta-catenin complex and the epithelial barrier. J.Biomed. Biotechnol. 2011, 1–6 (2011)
49. J.A. Marchal, M.C. Nunez, A. Aranega, M.A. Gallo, A. Espinosa,J.M. Campos, Acyclonucleosides, modified seco-nucleosides, andsalicyl- or catechol-derived acyclic 5-fluorouracil O, N-acetals:antiproliferative activities, cellular differentiation and apoptosis.Curr. Med. Chem. 16, 1166–83 (2009)
50. J.P. Thiery, J.P. Sleeman, Complex networks orchestrate epithelial-mesenchymal transitions. Nat. Rev. Mol. Cell Biol. 7, 131–42(2006)
51. V. Bolos, H. Peinado, M.A. Perez-Moreno, M.F. Fraga, M.Esteller, A. Cano, The transcription factor Slug represses E-cadherin expression and induces epithelial to mesenchymal transi-tions: a comparison with Snail and E47 repressors. J. Cell. Sci.116, 499–511 (2003)
52. C. Castro Alves, E. Rosivatz, C. Schott, R. Hollweck, I. Becker etal., Slug is overexpressed in gastric carcinomas and may actsynergistically with SIP1 and Snail in the down-regulation of E-cadherin. J. Pathol. 211, 507–15 (2007)
53. P. Jethwa, M. Naqvi, R.G. Hardy, N.A. Hotchin, S. Roberts, R.Spychal et al., Overexpression of Slug is associated with malignantprogression of esophageal adenocarcinoma. World J.Gastroenterol. 14, 1044–52 (2008)
54. K. Zhang, D. Chen, X. Jiao, S. Zhang, X. Liu, J. Cao et al., Slugenhances invasion ability of pancreatic cancer cells throughupregulation of matrix metalloproteinase-9 and actin cytoskeletonremodeling. Lab. Invest. 91, 426–38 (2011)
55. M.G. Mendez, S. Kojima, R.D. Goldman, Vimentin induceschanges in cell shape, motility, and adhesion during the epithelialto mesenchymal transition. FASEB J. 24, 1838–51 (2010)
Pancreatic (pro)enzymes promote cell differentiation 301

Copyright of Cellular Oncology (2211-3428) is the property of Springer Science & BusinessMedia B.V. and its content may not be copied or emailed to multiple sites or posted to alistserv without the copyright holder's express written permission. However, users may print,download, or email articles for individual use.













![MiR-26b suppresses cell proliferation and invasion by ... · Neurobiol. 2016. [Epub ahead of print] 6. Yao C, Li P, Song H, et al. CXCL12/CXCR4 Axis Upregulates Twist to Induce EMT](https://static.fdocuments.in/doc/165x107/5f7e1db4f0db084cb75cc44e/mir-26b-suppresses-cell-proliferation-and-invasion-by-neurobiol-2016-epub.jpg)




