
In vitro phosphorylation of AlgR, a regulator of mucoidy ... · 2762 V. Deretic, J. H. J. Leveau,...
8
Molecular Microbiology (1992) 6(19), 2761-2767 In Vitro phosphorylation of AlgR, a regulator of mucoidy in Pseudomonas aeruginosa, by a histidine protein kinase and effects of small phospho-donor molecules V. Deretic,* J. H. J. Leveau, C. D. Mohr and N. S. Hibler Department of Microbiology, University of Texas Health Science Center at San Antonio, San Antonio, Texas 78284-7758. USA. Summary AlgR is a transcriptional regulator of mucoidy in Pseudomonas aeruginosa, a critical virulence factor expressed in cystic fibrosis. AlgR belongs to the superfamily of bacterial signal transduction systems, and has been shown to bind to the algD promoter, a critical point in the regulation of mucoidy. This protein, like other typical response regulators, contains highly conserved residues known to be critical for the phos- phorylation and signal transduction processes. However, a typical second component Interacting with AlgR has not been identified. Here we demonstrate that AlgR undergoes phosphorylation in vitro when interacting with the well-characterized histidine pro- tein kinase CheA. These results Indicate that AlgR is capable of undergoing phosphorylation typical of other two-component signal transduction systems. Moreover, the phosphotransfer reaction between CheA and AtgR was found to be affected by the presence of carbamoyl phosphate, acetyl phosphate, and salts of phosphoramidic acid, recently shown to act as small-motecular-weight phospho-donors in the process of phosphorylation of several response regulators. These findings suggest that AlgR may react with intermediary metabolites such as carba- moyl phosphate and acetyl phosphate, and that these processes may play a role in the control of mucoidy in P. aeruginosa. Introduction Respiratory infections with mucoid Pseudomonas aerugi- nosa strains are the major cause of high mortality and morbidity in cystic fibrosis (Govan, 1988), Mucoidy is the Received 17 April, 1992; revised and accepted 11 June, 1992, *For corresponder>ce. Tel. (512) 567 3962; Fax (512) 567 6612. result of the overproduction and excessive secretion of the exopolysaccharide alginate {Govan, 1988). Transcrip- tional activation of algD (Deretic et ai, 1987), a gene encoding GDPmannose dehydrogenase (Deretic et ai, 1987), is a sine qua non for the establishment of mucoidy (Deretic e^ al., 1991). AlgR is absolutely required for the transcription of algD and binds to three sites within the algD promoter (Mohr et ai, 1992), two of which are located in a region positioned unusually far upstream from the mRNA start site (Mohr et ai, 1990). AlgR is a member of the superfamily of bacterial signal transduction systems (Deretic et al., 1989; Stock et ai, 1989b), and contains a conserved W-terminal domain typical of response-regula- tors, as well as a unique C-terminal domain (Deretic e( ai, 1989). A sensory component interacting with AlgR, commonly present in other two-component systems, has not been identified. It has been suggested that the algQ gene (Deretic and Konyecsni, 1989; Konyecsni and Dere- tic, 1990), also known as AlgR2 (Katoef a/., 1989), located several kilobases downstream of atgR, plays a role in this process (Deretic and Konyecsni, 1989; Kato ef ai, 1989; Konyecsni and Deretic, 1990). However, this relatively small acidic polypeptide does not contain any recogniz- able histidine kinase motifs (Kato et al., 1989; Konyecsni and Deretic, 1990). The A/-terminal domain of AlgR has its primary structure closely resembling all members of the class, and it contains the critical residues (corresponding to Asp-12, Asp-13, Asp-57, and Lys-109 of CheY, the paradigm of these types of signal transducers (Stock etai, 19B9a,b)) found in all characterized response regulators that undergo phosphorylation (Stock et ai, 1989a,b; Bourret et ai, 1991). In this work we examined the possibitity that AlgR can interact with a typical histidine protein kinase using CheA, a protein involved in the regulation of chemotaxis in Salmonella typhimurium (Wylieefa/., 1988). In addition, since mucoidy appears to be modulated by a variety of different environmental signals, including growth rate (Terry et ai, 1991) and carbon source (DeVaulte/a/., 1991), we also investigated whether AlgR can interact with small-molecular-weight phospho-donors such as carbamoyl phosphate and acetyl phosphate, as has been recently described for CheY and PhoB (Lukat et ai, 1992; Wanner and Wilmer- Reisenberg, 1992).
Transcript of In vitro phosphorylation of AlgR, a regulator of mucoidy ... · 2762 V. Deretic, J. H. J. Leveau,...

Molecular Microbiology (1992) 6(19), 2761-2767
In Vitro phosphorylation of AlgR, a regulator of mucoidyin Pseudomonas aeruginosa, by a histidine proteinkinase and effects of small phospho-donor molecules
V. Deretic,* J. H. J. Leveau, C. D. Mohr and N. S.HiblerDepartment of Microbiology, University of Texas HealthScience Center at San Antonio, San Antonio, Texas78284-7758. USA.
Summary
AlgR is a transcriptional regulator of mucoidy inPseudomonas aeruginosa, a critical virulence factorexpressed in cystic fibrosis. AlgR belongs to thesuperfamily of bacterial signal transduction systems,and has been shown to bind to the algD promoter, acritical point in the regulation of mucoidy. This protein,like other typical response regulators, contains highlyconserved residues known to be critical for the phos-phorylation and signal transduction processes.However, a typical second component Interacting withAlgR has not been identified. Here we demonstratethat AlgR undergoes phosphorylation in vitro wheninteracting with the well-characterized histidine pro-tein kinase CheA. These results Indicate that AlgR iscapable of undergoing phosphorylation typical ofother two-component signal transduction systems.Moreover, the phosphotransfer reaction betweenCheA and AtgR was found to be affected by thepresence of carbamoyl phosphate, acetyl phosphate,and salts of phosphoramidic acid, recently shown toact as small-motecular-weight phospho-donors in theprocess of phosphorylation of several responseregulators. These findings suggest that AlgR mayreact with intermediary metabolites such as carba-moyl phosphate and acetyl phosphate, and that theseprocesses may play a role in the control of mucoidy inP. aeruginosa.
Introduction
Respiratory infections with mucoid Pseudomonas aerugi-nosa strains are the major cause of high mortality andmorbidity in cystic fibrosis (Govan, 1988), Mucoidy is the
Received 17 April, 1992; revised and accepted 11 June, 1992, *Forcorresponder>ce. Tel. (512) 567 3962; Fax (512) 567 6612.
result of the overproduction and excessive secretion of theexopolysaccharide alginate {Govan, 1988). Transcrip-tional activation of algD (Deretic et ai, 1987), a geneencoding GDPmannose dehydrogenase (Deretic et ai,1987), is a sine qua non for the establishment of mucoidy(Deretic e^ al., 1991). AlgR is absolutely required for thetranscription of algD and binds to three sites within thealgD promoter (Mohr et ai, 1992), two of which are locatedin a region positioned unusually far upstream from themRNA start site (Mohr et ai, 1990). AlgR is a member ofthe superfamily of bacterial signal transduction systems(Deretic et al., 1989; Stock et ai, 1989b), and contains aconserved W-terminal domain typical of response-regula-tors, as well as a unique C-terminal domain (Deretic e( ai,1989). A sensory component interacting with AlgR,commonly present in other two-component systems, hasnot been identified. It has been suggested that the algQgene (Deretic and Konyecsni, 1989; Konyecsni and Dere-tic, 1990), also known as AlgR2 (Katoef a/., 1989), locatedseveral kilobases downstream of atgR, plays a role in thisprocess (Deretic and Konyecsni, 1989; Kato ef ai, 1989;Konyecsni and Deretic, 1990). However, this relativelysmall acidic polypeptide does not contain any recogniz-able histidine kinase motifs (Kato et al., 1989; Konyecsniand Deretic, 1990). The A/-terminal domain of AlgR has itsprimary structure closely resembling all members of theclass, and it contains the critical residues (correspondingto Asp-12, Asp-13, Asp-57, and Lys-109 of CheY, theparadigm of these types of signal transducers (Stock etai,19B9a,b)) found in all characterized response regulatorsthat undergo phosphorylation (Stock et ai, 1989a,b;Bourret et ai, 1991). In this work we examined thepossibitity that AlgR can interact with a typical histidineprotein kinase using CheA, a protein involved in theregulation of chemotaxis in Salmonella typhimurium(Wylieefa/., 1988). In addition, since mucoidy appears tobe modulated by a variety of different environmentalsignals, including growth rate (Terry et ai, 1991) andcarbon source (DeVaulte/a/., 1991), we also investigatedwhether AlgR can interact with small-molecular-weightphospho-donors such as carbamoyl phosphate andacetyl phosphate, as has been recently described forCheY and PhoB (Lukat et ai, 1992; Wanner and Wilmer-Reisenberg, 1992).
2762 V. Deretic, J. H. J. Leveau, C. D. MohrandN. S. Hibler
kDa 0 I2 0 0 -
lO' 30' Ih
6 9 -
4 6 -
-aCheA
3 0 - lAlgR
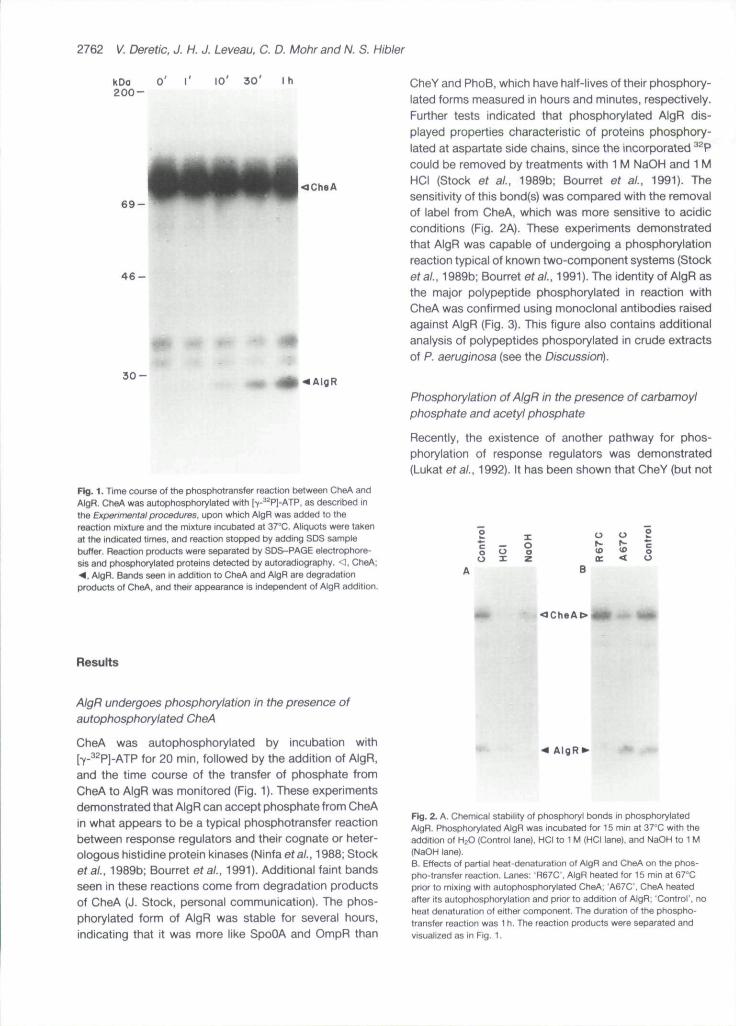
Fig. 1. Time course of the phosphotransfer reaction between CheA andAlgR. CheA was autophosphorylated with [y-^^PJ-ATP. as described inthe Experimental procedures, upon which AlgR was added to thereaction mixture and the mixture incubated at 37°C. Aliquots were takenat the indicated times, and reaction stopped by adding SDS samplebuffer. Reaction products were separated by SDS-PAGE electrophore-sis and phosphorylated proteins detected by autoradiography. <1. CheA;<. AlgR. Bands seen in addition to CheA and AlgR are degradationproducts of CheA. and their appearance is independent of AlgR addition.
CheY and PhoB, which have half-lives of their phosphory-lated forms measured in hours and minutes, respectively.Further tests indicated that phosphorylated AlgR dis-played properties characteristic cf proteins phosphory-lated at aspartate side chains, since the incorporated ̂ ^Pcould be removed by treatments with 1 M NaOH and 1 MHCI (Stock et ai, 1989b; Bourret ef ai, 1991). Thesensitivity of this bond(s) was compared with the removalof label from CheA, which was more sensitive to acidicconditions (Fig. 2A). These experiments demonstratedthat AlgR was capable of undergoing a phosphorylationreaction typical of known two-component systems (Stocketai, 1989b; Bourret eta/., 1991). The identity of AlgR asthe major polypeptide phosphorylated in reaction withCheA was confirmed using monoclonal antibodies raisedagainst AlgR (Fig. 3). This figure also contains additionalanalysis of polypeptides phosporylated in crude extractsof P. aeruginosa (see the Discussion).
Phosphoryiation of AlgR in the presence ofcarbamoyiphosphate and acetyi phosphate
Recently, the existence of another pathway for phos-phorylation of response regulators was demonstrated(Lukat et ai, 1992). It has been shown that CheY (but not
I 3 I(J I Z
Results
<lCheAl>
AlgR undergoes phosphorylation in the presence ofautophosphoryiated CheA
CheA was autophosphorylated by incubation with[-y-̂ ^P]-ATP for 20 min, followed by the addition of AlgR,and the time ccurse of the transfer of phosphate fromCheA to AlgR was monitored (Fig. 1). These experimentsdemonstrated that AlgR can accept phosphate from CheAin what appears to be a typical phosphotransfer reactionbetween response regulators and their cognate or heter-ologous histidine protein kinases (Ninfa efa/., 1988; Stocketai. 1989b; Bourret etai, 1991). Additional faint bandsseen in these reactions come from degradation productsof CheA (J. Stock, personal communication). The phos-phorylated form of AlgR was stable for several hours,indicating that it was more like SpoOA and OmpR than
•4 AlgRi
Fig. 2. A. Chemical stability of phosphoryl bonds in phosphorylatedAlgR. Phosphorylated AlgR was incubated for 15 min at 37°C with theaddition of H20 (Control lane). HCI to 1 M (HCt lane), and NaOH to 1 M(NaOH lane).B. Effects of partial heat-denatufation of AlgR and CheA on the phos-pho-transfer reaction. Lanes: 'R67C'. AigR heated for 15 min at 67°Cprior to mixing with autophosphorytated CheA: 'A67C', CheA heatedafter Its autophosphorylation and prior to addition of AlgR; 'Control', noheat denaturation of either component. The duration of the phospho-transfer reaction was 1 h. The reaction products were separated andvisualized as in Fig, 1,

Phosphorytation ofAtgR 2763
CheA M22.AlgR 0 6
I I
CheA
AlgR •
•f' kDa
—200
_IOO93
—69
-46
- 3 0
algR1 21 1
31
atgR41
algR
1 21 1
31
algR41
=9769
AlgR •
3] ^2p
1 2 3 4
CheA
AlgR •
kDa
- 4 6
- 3 0
C mAb(aAlgR)
2 3 4I I I kDa
_ 9 7- 6 9
- 4 6
- 3 0
— M - 1432 mAb(aAIgR)
Fig. 3. Identification of phosphofylation products by immunoblotting and autoradiographic analyses.A. Comparison of CheA-A)gR phosphotranster products (see the Discussion) with the phosphorylation of an uncharacterized polypeptide in crudeextracts of P. aeruginosa 8822 reported to represent AlgR by Rcychoudhury ef al. (1992a,b). Lanes: 'CheA-AlgR', transfer of ^^P phosphate from CheA toAlgR, as described in the Experimental procedures; •8822', 0.6M.g ml '.crude extracts (concentration of protein adjusted to 0,6 (ig ml ') preparedaccordinng to Roychoudhury etal. (1992a,b) incubated with [^-'^PJ-ATP for 5 min at room temperature: •8822', 160ng ml ', the same as in lane 88220.6, except that the concentration of proteins was 160tig mi '. Two polypeptides are seen (lane 8822 0.6t.Lg ml ') at positions as reported byRoychoudhury etal. (1992b), but the radiolabelled polypeptide above the 30kDa has a different mobility from that of phosphofylated AlgR.B and C. Autoradiogram and Western blot, respectively, of the same gel with crude protein preparations (Roychoudhury ef al., 1992b| from differentstrains of P. aeruginosa. Lanes: 1, PAO579 (mucoid {muc-23) derivative of PAO381); 2, PAO381 [non-mucoid strain); 3, PAO568 (mucoid {muc-2)derivative of PAO381, and parental to CDMl/1); 4, CDM1/1 {algR strain with insertionally inactivated algR in the PAO568 background} (Fyfe and Govan,1980; Mohr and Deretic. 1990; Mohr etal., 1990).
D and E. Autoradiogram and Western biot, respectively, of the same gel with 1, autophosphorylated CheA incubated with AlgR as described in theExperimental procedures. 2, 3, and 4, crude extracts of 8822 (160, 0.6 and 0.6ng ml ', respectively) prepared and incubated with [-y-̂ ^P]-ATP asdescribed by Roychoudhury ef al. (1992b). Lane 4 has a preparation of partially purified AlgR (50-60% pure AlgR) added to the reaction mixtureaccording to the procedure described by Roychoudhury er al. (1992a,b). An increase in the radiolabelling of the band migrating slightly slower than the30kDa standard is probably due to the presence of a contaminating protein in this partially purified AlgR preparation.
CheB) can be phosphorylated in the absence of histidineprotein kinases by acetyl phosphate and carbamoyiphosphate, cellular intermediates Important in carbonutilization and synthesis of both pyrimidine and arginine,respectively. This phenomenon entails that sources forphosphorylation other than typical histidine proteinkinases may be used by certain response regulators suchas AlgR. Since there is no commercially available radio-labelled carbamoyi phosphate or acetyl phosphate, v/eemployed a competition assay based on monitonng thetransfer of ^^P phosphate from CheA to AlgR in thepresence of these small-molecular-weight compounds.As shown in different panels of Fig. 4, the addition ofcarbamoyi phosphate and acetyl phosphate efficientlysuppressed the transfer of ^^P from CheA to AlgR. Thissuggested that these compounds interacted with AlgR
probably by competing with the transfer of the phosphorylgroup from CheA to AlgR.
Phosphorytation cfAigR in the presence ofphosphoramidates
It has recently been demonstrated that CheY as well asCheB, both homologues of AlgR, can undergo directphosphorylation in the presence of salts of phosphorami-dic acid (Lukat efa/., 1992). This suggests that phospho-transfer between histidine protein kinases and responseregulators may depend on a more active enzymatic role ofresponse regulators in their interaction with phosphohisti-dine residues located in kinases (Lukat et at., 1992). Toexplore this possibility in the case of CheA-AlgR interac-tion, we partially heat-denatured AlgR or phosphorylated

2764 y. Deretic, J. H. J. Leveau, C. D. Mohr and N. S. Hibler
^ z CP PRKCP C ^ ^ mM: I 10 I 10 C
ATP mM AcP0.2 1 10 0
<CheA <CheA • * < CheA
I AlgRAlgR
A B C
Fig. 4. Competition of carbamoyl phosphate, phosphoramidate, and acetyl phosphate with CheA for phosphotransfer to AlgR,A. Autophosphorylated CheA was incubated with AlgR in the presence of lOmM cartjamoyl phosphate (lane CP), no additions (control, lane C), lOmMpotassium hydrogen phosphoramidate (lane PRK), or lOmM ammonium hydrogen phosphoramidate (tane PRN),B. Autophosphorylated CheA was incubated with AlgR in the presence of different concentrations (1 and lOmM where indicated) ot carbamoylphosphate (CP), potassium hydrogen phosphoramidate (PRK), ATP, or no additions {lane C).C. Effects of different concentrations of acetyl phosphate (0,2, 1, 10mM)on the transfer ol phosphate from CheA to AlgR compared with no additions(lane labelled 0). In each case the incubation time was 1 h. The products of phosphotylation reactions were treated as described in the Experimentalprocedures, separated by SDS-PAGE, and visualized by autoradiography.
CheA prior to their mixing, and examined the effects ofsuch treatments on the efficiency of the phosphotransferreaction (Fig. 2B). These results indicated that heattreatment of AlgR had a more profound effect andsignificantly reduced the efficiency of the phosphotransferin contrast to the treatment of CheA, in support of thenotion that molecules such as AlgR may play an activeenzymatic role in the process of their phosphorylation. Toexamine further whether AlgR can interact with small-molecular-weight molecules with the chemistry of a phos-phoryl bond similar to the phosphoramidate bond in CheA,potassium and ammonium hydrogen salts of phosphora-midic acid were synthesized and used in the competitionassay with the CheA-AlgR phosphotransfer reaction (Fig.4A). Phosphoramidate proved to be as efficient a competi-tor of the transfer of ^^P from CheA to AlgR as werecarbamoyl phosphate and acetyl phosphate.
phosphoramidate, the anticipated mechanism of actionwas that the small-molecular-weight phospho-donorsmodified AlgR and reduced the number of availableacceptor sites or their affinity for the transfer of radio-labelled phosphate from CheA to AlgR. In support of directphosphorylation and competition for the similar residues,
1 2
AlgR I
Concentration-dependent effects of carbamoylphosphate, acetyl phosphate and phosphoramidate
In all three cases of the inhibition of the phosphotransferreaction by acetyl phosphate, carbamoyl phosphate, and
Fig. 5. Radiolabelling of AlgR with acetyl pP]-phosphate, AlgR wasincubated with acetyl PP]-phosphate (2mCi mmo! ') as described inthe Experimental procedures, and reaction products were analysed bySDS-PAGE and autoradiography. Lanes: 1, acetyl P^P]-phosphate; 2, aslane 1 with the addition of lOmM EDTA.

Phosphorylation of AlgR 2765
preliminary quantitative measurements with acetyi phos-phate indicate that this Is a competitive inhibition. Theconcentration-dependence of these processes is shownin Fig. 4, B and C. In addition, acetyi PP]-phosphate wassynthesized according to the method of Stadtman (1957).and it did radiolabel AlgR when incubated with this protein(Fig. 5); however, the specific activity of radiolabelledacetyi phosphate was very low (2 mCi mmol"^) and thedetectable incorporation of ^^P was far less than in theCheA-AlgR phosphotransfer reaction. In addition, whenEDTA was present in the reaction mixture this reduced theradiolabelling of AlgR with acetyi [^^P]-phosphate, but didnot completely eliminate it.
The results obtained in this study also suggest thatAlgR, like CheY (Lukat et ai, 1992), may play an active rolein its own phosphorylation. Moreover, the finding thatcarbamoyi phosphate and acetyi phosphate, two impor-tant intermediates in bacterial metabolism, can affect therate of AlgR phosphorylation in vitro, points to the possi-bility that the complex regulation of mucoidy in P. aerugi-nosa (Deretic efal., 1991) may be affected by intracellularconcentrations of such small-molecular-weight phospho-rylated intermediates.
Discussion
In this work we have examined whether AlgR is capable ofundergoing a phosphorylation modification observed withseveral other studied response regulators (Ninfa ef al..1988; Stock ef ai. 1989b; Bourret ef a/., 1991). The resultsshow that AlgR can be phosphorylated in a similar manner.This suggests that a typical histidine protein kinasecognate to AlgR may be present in P. aeruginosa, althoughsuch a protein has yet to be identified.
Taken together, the results presented here and those ofLukat ef ai (1992), as well as the recent genetic studies ofWanner and Wilmer-Riesenberg (1992), strongly suggestthat the phosphotransfer reactions involving responseregulators may be more complicated in vivo than originallyanticipated. It is possible that several types of macromol-ecules and small-molecular-weight phosphate donorscould interact with a given response regulator, thusbroadening the operational definition of the term 'cross-talk', a phenomenon formerly ascribed to interactionsamong non-cognate sensors and regulators (Ninfa etai,1988; Stock eta/., 1989b).
The role of carbamoyi phosphate and acetyi phosphatemay be of significance for the understanding of theenvironmental modulation of mucoidy and the emergenceof mucoid strains in cystic fibrosis (Deretic ef ai, 1991).Interpretations of phenomena related to environmentalmodulation of this virulence determinant (Deretic ef ai.1991) should now take into account the effects that grov r̂thconditions can have on intracellular levels of intermediates
such as carbamoyt phosphate and acetyi phosphate. Theformer is necessary for pyrimidine and arginine biosynthe-sis, and is also used by P. aeruginosa for ATP synthesisunder anaerobic conditions via the arginine deiminasepathway (Galimand ef ai. 1991); the latter is related to thesynthesis of acetyl-CoA, a key substrate for the tricarbox-ylic acid cycle, and is an important intermediate inutilization of pyruvate, glucose, and acetate, as well as inATP synthesis when oxygen is limiting in some organisms(Knappe, 1987).
It has been reported that slowing the growth rate of P.aeruginosa by limitation of different nutrients (Deretic efai, 1991; Terry etal.. 1991) can result in activation of algDand the emergence of mucoid strains. Exactly whatprocesses and messenger molecules are involved in thetransduction of such signals have never been understood.Based on the results reported here, we propose thatintermediates such as carbamoyi phosphate and acetyiphosphate, which may vary in concentration dependingon the growth rate and nutrition conditions (Lukat ef ai,1992; Wanner and Wilmer-Reisenberg, 1992), could affectthe signal transduction processes. In such a model,intracellular levels of these intermediates could affect AlgRphosphorylation or its capacity to be phosphorylated, andthereby contribute to the modulation of the alginatesystem in P. aeruginosa. In addition, the ability of simple,small-molecular-weight phospho-donors to interact withAlgR and affect its phosphorylation suggests that perhapsinhibitors of phosphotransfer reactions may be developedin an effort to provide alternative therapeutic strategies forcontrol of the currently intractable Pseudomonas infec-tions in cystic fibrosis.
In order to accomplish these goals it will be important toidentify protein kinases which interact with AlgR in vivo. Ofthe mutations affecting algD transcription, several havebeen suggested as potentially representing the putativesecond component (Deretic ef ai, 1991). The first geneproposed to play such a function was algO (Deretic andKonyecsni, 1989) (also termed AlgR2 (Kato etai, 1989)).However, AlgQ has no discernable similarity with theknown histidine kinases (Kato etai, 1989; Konyecsni andDeretic, 1990). While this manuscript was in preparation,Roychoudhury etai (1992a,b) presented evidence indi-cating that AlgQ (AlgR2) can undergo phosphorylation andtransfer the phosphate to a 30 kDa polypeptide in crudeextracts of P. aeruginosa. Furthermore, Roychoudhury efai (1992b) suggested that the 30kDa polypeptide wasAlgR, and reported that they were monitoring phosphory-lation of AlgR (Roychoudhury etai, 1992a,b). Since AlgRin our hands displayed the capacity to undergo phos-phorylation typical of the two-component signal transduc-tion systems, we repeated the experiments ofRoychoudhury ef ai (1992b). The results of these studiesare shown in Fig. 3. In these experiments, while we could

2766 V. Deretic, J. H. J. Leveau, C. D. Mohr and N. S. Hibier
clearly detect the phosphorylation of a 30 kDa polypeptide
(Fig. 3A) with electrophoretic mobility identical to that
reported by Royohoudhury et ai (1992b), monoclonal
antibodies against AlgR did not react with this polypeptide
(Fig. 3, C and E). Instead, monoclonal antibody 3H9
recognized AlgR in the same sample as a band located
below the 30 kDa polypeptide, at the position which was in
agreement with the previously determined M, of AlgR
(27600 Da); the same antibodies reacted with the CheA-
phosphorylated AlgR (Fig. 3E) at a position identical to that
of the band seen on autoradiograms (Fig. 3D). Moreover,
when an algR~ strain was tested, the 30 kDa phosphory-
lated protein was still present on autoradiograms (Fig. 3B)
while AlgR was completely absent in this strain, as judged
by Western blot analysis (Fig. 3C). Thus, the phosphory-
lated 30 kDa polypeptide reported to be AlgR by
Roychoudhury et ai (1992a,b) is not AlgR and its identity,
role, and any relevance to mucoidy are unclear.
We are currently in the process of identifying protein
kinase(s) interacting with AlgR in vivo, studying how
environmental conditions affect intracellular concentra-
tions of carbamoyi and acetyl phosphate, and determining
whether these processes modulate phosphorylation of
AlgR in vivo.
Experimental procedures
Proteins and reagents
AlgR was purified as previously described (Mohr etai., 1991). Thepurity was estimated to be greater than 95% (Mohr efa/., 1991).Purified CheA from S. typhimurium was a gift from Dr J. Stook.[-y-32pj_ATP, 6000Ci mmol ', was obtained from DuPont NEN.Potassium and ammonium salts of phosphoramidic acid(KHPO3NH2 and NH4HP03NH2) were synthesized according tothe method of Sheridan et ai (1971) and the synthesizedcompounds characterized by mass spectroscopy. AcetylP^P]-phosphate was synthesized using acetic anhydride andpotassium p^P]-orthophosphate (9000Ci mmol '; NEN DuPont)according to the method of Kornberg as described by Stadtman(1957), except that one tenth of the recommended volume of 4 NLiOH was used to precipitate the dilithium salt of acetyl phos-phate. Acetyl and carbamoyi phosphates were obtained fromSigma Chemical Company.
Phosphorylation of AlgR
CheA was autophosphorylated according to the published pro-cedure (Wylie et ai, 1988). After incubation of CheA with[.Y-32p]-ATP for 20 min at room temperature, purified AlgR wasadded so that the final concentrations of components werelOOmM Tris pH 8.0. 5mM MgCb, 50mM KCl. 2.5[xM CheA andIOM-M AlgR. When indicated, solutions of acetyl phosphate.carbamoyi phosphate, and K' or NH^ salts of phosphoramidicacid were added to the desired final concentrations. Phosphory-lation of AlgR was carried out for 1 h at 37°C, unless specifiedotherwise. The reaction was stopped by adding SDS sample
buffer and placing samples on dry ice. Radiolabelled productswere detected by autoradiography after their separation bySDS-polyacrylamide gel electrophoresis (SDS-PAGE). Whereindicated, the reaction products were treated by adding 50 \Lg ofcrude protein to the sample; this removed most of the labelassociated with proteolytic CheA products but did not affect theamount of ^^P incorporated in the AlgR band. In the case ofphosphorylation using acidified crude extracts of P. aeruginosathe protocols described by Roychoudhury ef ai (1992b) werefollowed. AlgR was radiolabelled with acetyl pP]-phosphateaccording to the protocol of Lukat etai (1992), using an estimated2.5-fold higher concentration of acetyl p^P]-phosphate (2mCimmol ').
Monoclonal antibodies against AigR and Western
biotting anaiysis
Monoclonal antibodies against AlgR were generated as pre-viously described (Deretic etai, 1992), using purified AlgR (Mohref ai, 1991). Hybridomas producing AlgR-reactive antibodieswere subcloned by the method of limiting cell dilution and werethen cryopreserved. In the case of monoclonal antibody 3H9 thesubclass was lgG1. Large-scale production of antibodies wasperformed from ascites fluid and antibodies purified by chroma-tography on a protein G sepharose 4 fast-flow column, using themonoclonal antibody Mab Trap G Kit (Pharmacia). Western blotswere performed as described before (Deretic etai., 1992), usinghorseradish peroxidase-conjugated goat anti-mouse IgG(HyClone). Bound antibodies were visualized by reaction withH2C2 and diaminobenzidine (Deretic ef ai, 1992).
Acknowledgements
We thank J. B. Stock for providing us with CheA, W. McCleary foradvice on acetyl phosphate synthesis, and A. Lazzell and theInstitutional Hybridoma Facility for the help with the generation ofmonoclonal antibodies. This work was supported by grantsAI31139 from the National Institutes of Health, G229 from theCystic Fibrosis Foundation, and in part by DMB-8807424 fromNational Science Foundation. C.D.M. was supported by traininggrant T32 AI0727-06 from National Institutes of Health and was arecipient of a Predoctoral Traineeship grant (5405) from CysticFibrosis Foundation.
References
Bourret, R.B., Borkowich, K.A.. and Simon, M.l. (1991) Signaltransduction pathways involving protein phosphorylation inprokaryotes. Annu Rev Biochem 60: 401 -441.
Deretic, V., Konyecsni, W.M. (1989) Control of mucoidy inPseudomonas aeruginosa: transcriptional regulation of algRand identification of the second regulatory gene, algQ. JBacterioi-17^: 3680-3688.
Deretic, V., Gill, J.F., and Chakrabarty. A.M. (1987) Gene algDcoding for GDPmannose dehydrogenase is transcriptionallyactivated in mucoid Pseudomonas aeruginosa. J Bacteriot AG9:351-358.
Deretic, V., Dikshit. R., Konyeosni, W.M.. Chakrabarty. A.M., andMisra, T.K. (1989) The algR gene, which regulates mucoidy inPseudomonas aeruginosa, belongs to a class of environ-mentally responsive genes. J Bacterioi 171: 1278-1283.

Phosphorytation of AtgR 2767
Deretic, V., Mohr, CD., and Martin, D.W. (1991) Mucoid Pseudo-monas aeruginosa in cystic fibrosis: signal transduction andhistone-IJke elements in the regulation of bacterial virulence.Mol Microbiot 5: 1557-1583,
Deretic, V., Hibler, N.S., and Holt, S.C. (1992) Immunocytochemi-cal analysis of AlgP (Hpi), a histone-like element participating incontrol of mucoidy in Pseudomonas aeruginosa. J Bacteriot174:824-831.
DeVault, J.D., Hendrickson, W., Kato, J., and Chakrabarty, A.M.(1991) Environmentally regulated aigD promoter is responsiveto the cAMP receptor protein in Escherichia coti. Moi Microbiol5: 2503-2509.
Fyfe, J.A.M., and Govan, J.R.W. (1980) Alginate synthesis inmucoid Pseudomonas aeruginosa: a chromosomal locusinvolved in control. J Gen Microbiot A^Q: 443-450.
Galimand,M., Gamper, M.,Zimmermann, A.,andHaas, D.(1991)Positive FNR-like control of anaerobic arginine degradationand nitrate respiration in Pseudomonas aeruginosa. J Bacteriol173: 1598-1606.
Govan, J.R.W. (1988) Alginate biosynthesis and other unusualcharacteristics associated with the pathogenesis of Pseudo-monas aeruginosa in cystic fibrosis. In Bacteriat infections ofRespiratory and Gastrointestinal Mucosae. Griffiths, E., Dona-chie. W., and Stephen, J. (eds). Oxford: IRL Press, pp. 67-96.
Kato, J., Chu, L., Kitano, K., DeVault, J.D., Kimbara, K., Chakra-barty, A.M., and Misra, T.K. (1989) Nucleotide sequence of aregulatory region controlling alginate synthesis in Pseudomo-nas aeruginosa: characterization of the algR2 gene. Gene 84:31-38.
Knappe, J. (1987) Anaerobic dissimilation of pyruvate. In Escher-ichia coli and Salmonella typhimurium: Cellular and MotecutarBiotogy. Neidhardt, F.C, Ingraham, J.L., Low, K.B., Magasanik,B., Schaechter, M., and Umbarger, H.E. (eds). Washington,D.C: American Society for Microbiology, pp. 151-155.
Konyecsni, W.M., and Deretic, V. (1990) DNA sequence andexpression analysis of algP and algQ, components of themultigene system transcriptionally regulating mucoidy inPseudomonas aeruginosa: atgP contains multiple directrepeats. J Bacteriot 172: 2511-2520.
Lukat, G.S.. McCleary, W.R., Stock, A.M., and Stock, J.B. (1992)Phosphorylation of bacterial response regulator proteins bylow molecular weight phospho-donors. Proc Natt Acad SciUSA8Q: 718-722.
Mohr, CD., and Deretic, V. (1990) Gene-scrambling mutagenesis:generation and analysis of insertional mutations in the alginateregulatory region of Pseudomonas aeruginosa. J Bacteriot A72:62552-6260.
Mohr, CD., Martin. D.W., Konyecsni. W.M., Govan, J.R.W., Lory,S., and Deretic, V. (1990) Role of the far-upstream sites of the
atgD promoter and the atgtl and rpoN genes in environmentalmodulation of mucoidy in Pseudomonas aeruginosa. JBacteriot 172: 6576-6580.
Mohr, CD., Hibler, N.S., and Deretic, V. (1991) AlgR, a responseregulator controlling mucoidy in Pseudomonas aeruginosa,binds to the FUS sites of the algD promoter located unusuallyfar upstream from the mRNA start site. J Bacteriol 173:5136-5143.
Mohr, CD.. Leveau, J.H.J., Krieg, D.P., Hibler, N.S., and Deretic,V. (1992) AlgR binding sites within the atgD promoter comprisea set of inverted repeats separated by a large interveningsegment of DNA. J Bacteriot, in press.
Ninfa, A.J., Ninfa, E.G., Lupas, A., Stock, A., Magasanik, B., andStock. J. (1988) Crosstalk between bacterial chemotaxis signaltransduction proteins and the regulators of transcription of theNtr regulon: evidence that nitrogen assimilation and chemo-taxis are controlled by a common phosphotransfer mechanism.Proc Natt Acad Sci USA 85: 5492-5496.
Roychoudhury. S., Sakai, K., and Chakrabarty, A.M. (1992a)AlgR2 is an ATP/GTP-dependent protein kinase involved inalginate synthesis by Pseudomonas aeruginosa. Proc NatIAcad Sci USA 89: 2659-2663.
Roychoudhury, S., Sakai. K., Schlictman, D., and Chakrabarty,A.M. (1992b) Signal transduction in exopolysaccharide alginatesynthesis: phosphorylation of the response regulator AlgRI inPseudomonas aeruginosa and Escherichia coii. Gene 112:45-51.
Sheridan. R.C, McCullough, J.F., and Wakefield, Z.T. (1971)Phosphoramidic acid and its salts, tnorg Synth 13: 23-26.
Stadtman, E.R. (1957) Preparation and assay of acetyl phos-phate. Meth Enzymot 3: 228-231.
Stock, A.M., Mottonen, J.M,, Stock, J.B., and Schutt. CE. (1989a)Three-dimensional structure of CheY, the response regulator ofbacterial chemotaxis. Nature 337: 745-749.
Stock, J.B., Ninfa, A.J., and Stock, A.M. (1989b) Protein phos-phorytation and regulation of adaptive responses in bacteria.Microbiot Rev 53: 450-^90.
Terry, J.M., Pina, S.E., and Mattingty. S.J. (1991) Environmentalconditions which influence mucoid conversion in Pseudomo-nas aeruginosa PA01. Infect tmmun 59: 471-477.
Wanner, B.L., and Wilmes-Reisenberg, M.R. (1992) Involvementof phosphotransacetylase, acetate kinase. and acetyl phos-phate synthesis in control of the phosphate regulon in Escher-ichia coti J Bacteriot 174: 2124-2130.
Wylie, D., Stock. A., Wong, C.-Y., and Stock, J. (1988) Sensorytransduction in bacterial chemotaxis involves phosphotransferbetween che proteins. Biochem Biophys Res Commun 151:891-896.



















![arXiv:1003.4416v2 [math-ph] 11 May 2010 · Then (AlgR,R) is a formal distribution Lie superalgebra. Note that Alg is a functor from the category of Lie conformal superalgebras to](https://static.fdocuments.in/doc/165x107/5f7a75c0b5aacc1e7630f9db/arxiv10034416v2-math-ph-11-may-2010-then-algrr-is-a-formal-distribution-lie.jpg)