
In situ recruitment of sporophytes of the giant kelp ... · Address for correspondence: Kelp...
23
J. Exp. Mar. Biol. Ecol., 1986, Vol. 103, pp. 41-63 Elsevier 41 JEM 00770 In situ recruitment of sporophytes of the giant kelp, Macrocystis pyrijkra (L.) C.A. Agardh: effects of physical factors Lawrence E. Deysher and Thomas A. Dean Marine Science Institute, Universityof Ca&omia, Santa Barbara, CA 93106, U.S.A. (Received 14 April 1986; revision received 21 July 1986; accepted 4 August 1986) Abstract: Quantitative relationships were established between the recruitment of sporophytes of the giant kelp, Mucrocystis pyrifera (L.) C. A. Agardh, and various environmental variables. Artificial substrata were inoculated with M. pyriferu zoospores in the laboratory and outplanted to a set of field stations in the San Mateo, San Onofre, and Barn kelp forests in southern California. Measurements oftemperature, irradiance, seston flux and nitrogen concentration were made concurrently with estimates of sporophyte recruitment during 39 outplants from September 1977 to August 1982. Temperature, irradiance, and seston flux were significantly correlated with the density of sporophytes recruited from the outplanted gametophytes. Recruitment of sporophytes at densities >50 sporophytes cm - 2 represents sporophyte production by approximately 0.1 y0 of the outplanted female gametophytes. This occurred only at temperatures below 16.3 “C and irradiation levels above 0.4 E. m ‘. day - r. We have defmed these conditions as a “recruitment window” within which sporophytes can be produced under natural conditions. Sporophyte recruitment was enhanced by fertilizing gametophytes with nitrogen and phosphorus. However, the relative level of recruitment on the fertilized substrata, which were outplanted in the late summer when sea temperatures were above 16”C, was low compared to recruitment on unfertilized substrata outplanted when temperatures were below 16 “C. The irradiance requirements for sporophyte recruitment as indicated by these field studies agree well with those derived in the laboratory. Temperature, on the other hand, was negatively correlated with recruitment in the field, but had little effect (at temperatures up to 20 “C) in the laboratory. These results coupled with the nutrient enrichment data indicate that a factor correlated with temperature, such as nutrient levels, may be the most important in determining recruitment success. Our results suggest that large scale temporal patterns of recruitment of giant kelp in southern California are determined, in part, by the infrequent occurrences of recruitment windows. However, proper irradiance and temperature conditions are not the sole requisites for recruitment as evidenced by a lack of recruitment on our artificial substrata during some apparent window periods. Key words: Gametogenesis; Gametophytes; Kelp forests; Macrocystk; Recruitment; Southern California Recruitment processes are receiving increasing attention as important factors con- trolling the distribution and abundance of marine populations. Studies in several Address for correspondence: Kelp Ecology Project, 531 Encinitas Blvd., Suite 118, Encinitas, CA 92024, U.S.A.
Transcript of In situ recruitment of sporophytes of the giant kelp ... · Address for correspondence: Kelp...

J. Exp. Mar. Biol. Ecol., 1986, Vol. 103, pp. 41-63 Elsevier
41
JEM 00770
In situ recruitment of sporophytes of the giant kelp, Macrocystis pyrijkra (L.) C.A. Agardh: effects of physical
factors
Lawrence E. Deysher and Thomas A. Dean Marine Science Institute, University of Ca&omia, Santa Barbara, CA 93106, U.S.A.
(Received 14 April 1986; revision received 21 July 1986; accepted 4 August 1986)
Abstract: Quantitative relationships were established between the recruitment of sporophytes of the giant kelp, Mucrocystis pyrifera (L.) C. A. Agardh, and various environmental variables. Artificial substrata were inoculated with M. pyriferu zoospores in the laboratory and outplanted to a set of field stations in the San Mateo, San Onofre, and Barn kelp forests in southern California. Measurements oftemperature, irradiance, seston flux and nitrogen concentration were made concurrently with estimates of sporophyte recruitment during 39 outplants from September 1977 to August 1982.
Temperature, irradiance, and seston flux were significantly correlated with the density of sporophytes recruited from the outplanted gametophytes. Recruitment of sporophytes at densities >50 sporophytes cm - 2 represents sporophyte production by approximately 0.1 y0 of the outplanted female gametophytes. This occurred only at temperatures below 16.3 “C and irradiation levels above 0.4 E. m ‘. day - r. We have defmed these conditions as a “recruitment window” within which sporophytes can be produced under natural conditions.
Sporophyte recruitment was enhanced by fertilizing gametophytes with nitrogen and phosphorus. However, the relative level of recruitment on the fertilized substrata, which were outplanted in the late summer when sea temperatures were above 16”C, was low compared to recruitment on unfertilized substrata outplanted when temperatures were below 16 “C.
The irradiance requirements for sporophyte recruitment as indicated by these field studies agree well with those derived in the laboratory. Temperature, on the other hand, was negatively correlated with recruitment in the field, but had little effect (at temperatures up to 20 “C) in the laboratory. These results coupled with the nutrient enrichment data indicate that a factor correlated with temperature, such as nutrient levels, may be the most important in determining recruitment success.
Our results suggest that large scale temporal patterns of recruitment of giant kelp in southern California are determined, in part, by the infrequent occurrences of recruitment windows. However, proper irradiance and temperature conditions are not the sole requisites for recruitment as evidenced by a lack of recruitment on our artificial substrata during some apparent window periods.
Key words: Gametogenesis; Gametophytes; Kelp forests; Macrocystk; Recruitment; Southern California
Recruitment processes are receiving increasing attention as important factors con- trolling the distribution and abundance of marine populations. Studies in several
Address for correspondence: Kelp Ecology Project, 531 Encinitas Blvd., Suite 118, Encinitas, CA 92024, U.S.A.

42 LAWRENCE E. DEYSHER AND THOMAS A. DEAN
intertidal and subtidal communities have suggested that the population structure of dominant species is controlled primarily by larval recruitment (Birkland, 1982; Yoshioka, 1982; Doherty, 1983; Ebert, 1983; Watanabe, 1984; Calfey, 1985; Connell, 1985 ; Gaines & Roughgarden, 1985 ; Roughgarden et al., 1985). Underwood & Denley (1984) have also suggested that variations in recruitment may be important in inter- preting the roles which competition and predation play in structuring intertidal com- munities.
Recruitment processes appear to be especially important in populations of the giant kelp, Macrocystis pyrijka (L.) C. A. Agardh in southern California. The abundance of adult sporophytes of this species undergoes large fluctuations which peak every 2 to 3 yr (Rosenthal et al., 1974; Dayton et al., 1984; Dean & Thies, unpubl. data). These population fluctuations are generally coincident in kelp forests between Palos Verdes and San Diego, a distance of 160 km (North, 1971). Although there is local variability in kelp density, primarily due to adult mortality caused by storms and urchins, these gross, large scale patterns appear to reflect the patterns of sporophyte recruitment. Peaks in adult abundance follow distinct episodes of recruitment which also have a periodicity of 2 to 3 yr. During these recruitment episodes, high densities of small sporophytes occur over large areas of kelp forests throughout southern California (Dayton & Tegner, 1984; Dayton et al., 1984).
The recruitment of the Macrocystis sporophyte involves several distinct processes that include the release of haploid zoospores from the adult sporophyte, the settlement of these zoospores onto suitable substratum to start the gametophyte generation, the production of eggs and sperm by the gametophyte generation, and the subsequent fertilization and production of the sporophyte. The year-round release of zoospores noted over 5 yr in this study and by Anderson & North (1967) argues against zoospore availability controlling the temporal patterns of recruitment that are observed. This process, however, needs further study to determine its relative importance in sporophyte recruitment. Very little is also known about the factors influencing the settlement of zoospores onto suitable substratum. Preliminary studies in our laboratory (L. Deysher & T.A. Dean, unpubl. data) indicate that gametophytes are widespread on substrata within the San Onofre kelp forest. It therefore appears that widespread occurrence of sporophytes over 10’s to 100’s of kilometers of coastline during recruitment episodes is due to large scale oceanographic conditions influencing the production of sporophytes from the gametophyte generation.
The small size of the gametophyte of laminarian algae makes it difficult to directly observe this life history stage in the field. Therefore, the response of gametophytes to various physicochemical regimes has been investigated almost entirely in the laboratory (see Kain, 1979; Luning, 1980 for review). Recent laboratory studies on Macrocystis pynifra have determined the relationships between n-radiance, temperature, and sedi- mentation (Liining & Dring, 1972, 1975; Devinny & Volse, 1978; Liming & Neushul, 1978; Deysher & Dean, 1984, 1986). The objective of the present study was to identify the physicochemical parameters which regulate the recruitment of sporophytes from

MACROCYSTISRECRUITMENT 33
populations of g~etoph~es in the field and to establish qu~titative relationships between remitment and these parameters. In addition, we compared relationships observed in situ with those established previously in the more controlled but less complex laboratory environment.
STUDY AREA,MATERIALS,AND METHODS
STUDY AREA
This investigation was conducted in three Macrocystis pyrifra forests located in northern San Diego County, California (Fig. 1). Three stations were established in the San Onofre Kelp forest (SOK) z 6 km south of San Clemente; two stations in the San Mateo Kelp forest (SMK), 4 km northwest of SOK; and one in Barn Kelp (BK 55) approximately 10 km southeast of SOK. The three stations in SOK were located in the upcoast (SOKU 49, downcoast (SOKD 45), and inshore (SOKD 35) portions of the forest. The San Onofre Kelp and San Mateo Kelp stations were at a depth of 13 to 14 m
. . . “.._. .
KELP
Kilometefs BARN
Fig. 1. Locations of kelp forests and stations used in the gametophyte outplant experiments; SONGS is the San Onofre Nuclear Generating Station.

44 LAWRENCE E. DEYSHER AND THOMAS A. DEAN
except the inshore SOK station (SOKD 35) which was at a depth of 11 m. The Barn Kelp station was at a depth of 16 m. The bottom subs~at~ at each station was composed of mixed cobble and sand which is the predominant bottom type in San Onofre area. The San Onofre kelp forest stations, however, had a higher percentage of sand than the other stations.
The six field stations were used for varying periods of time between the Fall 1977 and the Fall 1982. The station at SOKD 45 was used continuously since 1977, the station at SOKU 45 was added in June 1979, and the station at BK 55 was added in August 1980. The additional three stations at SOKD 35, SMK 45 and SMK Canopy were added in August 1981. All the stations were in areas cleared of kelp canopy except the SMK Canopy station (SMK CAN).
OUTPLANT SUBSTRATA AND INOCULATION PROCEDURES
The primary experimental tools in this study were artificial substrata which were inoculated with Mucrocystis pyt$rra zoospores in the laboratory and outplanted to the field stations. These inoculated substrata provided a defined population of g~etoph~es from which the recruitment of sporophytes could be quantified. The substrata consisted of Plexiglas plates (30.5 x 10.2 x 0.6 cm) onto which pieces of nylon line (10 cm long, 0.64 cm diameter) were fastened with plastic cable ties. This arrangement allowed for the simultaneous inoculation of a large number of sampling units which could be individually removed from the substratum for transport back to the laboratory. The nylon line was chosen as the artiIicial substratum because dense sporophyte recruitment onto this substratum has been noted on many occasions within the San Onofre kelp forest.
Inoculation of the substrata followed the methods of Foster et al. (1985). Sporophylls used to obtain the zoospores in the inoculations were taken from adult Mucrocystis pyri@a in the San Onofre Kelp forest. The zoospore concentration of the inoculation solution was 1 .O x lo5 spores/ml. Before transport to the field, one rope from each plate was removed to quantify the initial gametophyte density. Gametophyte densities were counted at a ma~i~cation of 400 x with a dipping cone objective on a microscope with an epi-ill~ination system. The mean initial g~etoph~e density for all experiments was 327.2 mm-’ rt 37.9 (95% CI).
The substrata were transported into the field 36 h after the start of inoculation. They were transported to the field in black Plexiglas boxes stored in insulated chests. The transport boxes were designed to avoid exposing the gametophytes to large temperature changes and direct sunlight. The effects of transporting the gametophytes into the field were monitored during the fust four outplants by taking inoculated substrata into the field and then returning them to the laboratory. The gametophyte survival on these transport substrata was compared to that on plates which remained in the culture room. Dense populations of sporophytes were produced on both sets of substrata during culturing indicating that the process of transporting the substrata to the field had little effect on the gametophyte populations.

MACROCYSTZS RECRUITMENT 45
FIELD STATIONS
We attached outplant substrata and instruments used to measure the physical environment on an array of racks at each station (Fig. 2). Four replicate inoculated plates were placed on the bottom and two at a height of 2 m above the bottom. In addition, two inoculated plates were placed at both 4 m and 6 m above the bottom at the SMK 45 station during 1981 and 1982. The plates were kept in position by PVC frames attached to a stainless steel cable which was anchored to the bottom by a 23 kg steel plate. The bottom substrata were used because under natural conditions Mucro- cystis would only find suitable sites for attachment on the bottom. However, recruitment is infrequent on the bottom because of highly turbid conditions. The sites above the bottom were selected to provide more frequent occurrences of successful recruitment and provide a wider range of physicochemical conditions.
PHYSICAL DATA
Field measurements of irradiance, temperature, and seston flux were made at each field station concurrent with the outplant experiments. Details of the methods for measuring these variables are given in Dean & Jacobsen (1984) and Dean (1985). Nitrogen measurements near the San Onofre kelp forest were made at approximately monthly intervals by Marine Ecological Consultants and Southern California Edison from 1977 through 1981. During 1982, weekly nutrient measurements were made by Ecosystems Management, Inc.
h-radiance was recorded continuously and reported as integrated values over either 2-h or daily intervals. Measurements were made with LI-COR Instruments (Lincoln, NE) LI-192s underwater quantum sensor in conjunction with LI-500 integrators. The data from these instruments were recorded on either a digital counter which was read at weekly intervals by divers in the field or were recorded on an EPROM from which the data were directly transferred to computer tiles (Zabloudil, 1984). The EPROM data were recorded over 2-h intervals for periods up to a month.
Temperature was recorded at each station/depth by either Ryan model-J thermo- graphs (Ryan Instruments, Inc., Kirkland, WA) or instruments using thermilinear thermistors (Zabloudil, 1984). Daily means were calculated from temperatures recorded at 2-h intervals.
Vertical seston flux was determined by the accumulation of sediment within sediment traps. The traps were 30 cm in length with an opening of 2.5 cm. Three replicate traps were placed at each station and depth. Traps on the bottom were attached to steel reinforcing rods driven into the bottom substratum while the traps at 2 m above the bottom were placed on the outplant racks. The mouth of the traps on the bottom was 30 cm above the bottom and the traps at 2 m above the bottom were positioned such that the opening of the trap was at approximately the same level as the outplant substratum. The traps were replaced at weekly intervals and the height of the accumu- lated sediment was measured in the laboratory.

46 LAWRENCE E. DEYSHER AND THOMAS A. DEAN
2m u#va eot?lrrm plates wfRop0 &bra-
PVC Rack
Fig, 2. Design of outpIant array showjng the racks hokkg autpiant substrata on the bottom and at 2 m above the bottom: the instrumentation for measuring physical variables is &a shown.

MACROCYS7”S RECRUITMENT 47
SAMPLING OF OUTPLANT EXPERIMENTS
The number of replicate inoculated plates used at each field station varied between outplants. During the initial outplants, three plates, each with 30 ropes, were placed at each depth. In later outplants, the number of ropes was reduced to 7 per plate and at 2, 4, and 6 m above the bottom the number of plates was reduced to two per depth. These changes in sample size were due to a reduction in the number of times each outplant was sampled after being placed in the field. Each inoculated plate was held on a separate rack to ensure that data were obtained even if a rack was lost.
As indicated above, the sampling schedules for the gametophyte outplant substrata varied between outplants. Substrata were sampled p~~ic~ly during outpl~ts in 1977 to 1981. Collection intervals during these outplants ranged from 4 to 120 days. During 1982, the experiments were censused for recruited sporophytes only once after an outplant period of 42 days. This 42-day period was selected because it represented the time when the maximum sporophyte density could be expected in these outplants. An interval of 30-35 days is adequate for gametophytes to produce sporophytes (Deysher & Dean, 1984) and have them grow to a size (even under sub-optimal conditions) which could be censused using a magnification of lo-20 x . Sporophyte densities, on the other hand, generally decreased at sampling intervals longer than 48 days. In 21 of the 26 cases in which sporophyte densities were greater than 1. cm - *, the m~imum sporophyte density occurred within a 48-day outplant period, The use of the 42-day outplant interval, therefore, maximized rec~itment input and minimized the contributions of sporophyte mortality to the final sporophyte density. Only the 42-day data is presented in this paper. All of the outplants had collections at this time period.
Collection of inoculated substrata during the outplants in 1977 and early 198 1, which were sampled periodically throughout the outplant period, involved removal of indi- vidual lines from the plates which remained in the field. The lines were placed in small plastic vials and returned to the laboratory for censusing. In the outplants from late 198 1 to 1982, in which the substrata were sampled only once at 42 days after outplant, the entire plate was collected from the field and lines were removed from the plates in the laboratory. In these cases the substrata were transported back to the laboratory in containers which were kept cool and out of direct sunlight. All lines were observed within a few days of collection and storage in the dark at 2 “C was adequate for preservation of the sporophytes. Sporophytes on the entire line were counted at a magnification of 12 to 20 x .
NUTRIENT ENRICHMENT STUDIES
The effect of nutrients on sporophyte recruitment was tested in a field experiment in which the levels of nitrogen and phosphorus were elevated in the vicinity of outplanted gametophytes. These experiments used the same rope substrata attached to Plexiglas plates as in the gametophyte outplants. Nitrogen levels on these plates were elevated above ambient with a ~ommerci~ly available slow-release inorganic fertilizer
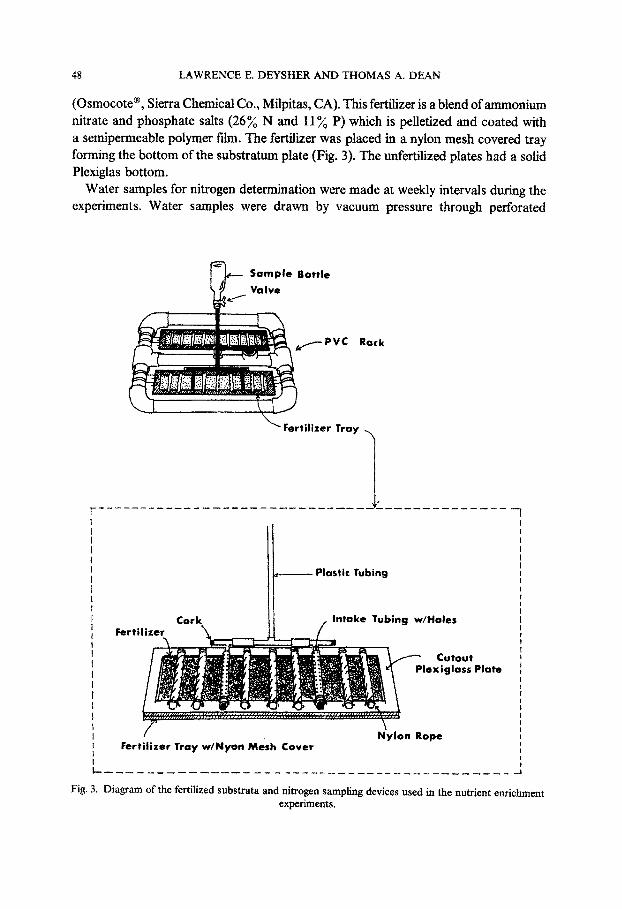
48 LAWRENCE E. DEYSHER AND THOMAS A. DEAN
(Osmocote@, Sierra Chemical Co., Milpitas, CA). This fertilizer is a blend of ammonium nitrate and phosphate salts (26% N and 11 yQ P) which is pelletized and coated with a semipermeable polymer f&n. The fertilizer was placed in a nylon mesh covered tray forming the bottom of the substratum plate (Fig. 3). The unfertilized plates had a solid Plexiglas bottom.
Water samples for nitrogen determination were made at weekly intervals during the experiments. Water samples were drawn by vacuum pressure through perforated
n-- Sample Bottle d id-- Volve
Fertilizer
vc Rack
Tray
r \1 __----------_____-__-~~~-~~~~~---------~~-~~- i I
I
Intake Tubing w/Holes I I I I
cutout lexigloss Plate i
I I I I
/ fertilizer Tray w/Nyon Mesh Cover
dylon Rape
L-____----_------ __-___- ---- -----------,---.I
Fig. 3. Diagram of the fertilized substrata and nitrogen sampling devices used in the nutrient enrichment experiments.

MACROCYSTZS RECRUITMENT 49
polyethylene tubes which alternated with the rope substrata on the outplant plate (Fig. 3). The samples were collected in acid washed glass bottles, immediately brought back to the boat, filtered, and frozen on dry ice. Analyses for NO; + NO; and NH,’ were made by Ecosystems Management, Inc. (Encinitas, CA).
The first nutrient enrichment experiment was put into the field on 6 August and the second on 10 September 1981. Both experiments were at the SMK 45 station. Each experiment consisted of four arrays, each having plates with attached lines at 0, 2, 4, and 6 m above the bottom. The plates in two of the arrays were fertilized, and the other two arrays served as unfertilized controls. The unfertilized arrays were located approxi- mately 10 m from the fertilized arrays. Preliminary experiments indicated that this was far enough from the controls to avoid any contamination by drift of the fertilizer (Dean & Jacobsen, 1986). Each plate with inoculated lines was paired with an uninoculated plate. The uninoculated plate served as a control to monitor the natural recruitment of sporophytes.
The fertilizer trays were replaced immediately after the weekly water samples were taken. The gametophyte substrata from the two experiments were collected after a 42-day outplant period on 16 September and 22 October 1981, for Experiments 1 and 2, respectively.
STATISTICAL ANALYSIS
The relationships between sporophyte recruitment and the various physicochemical factors were determined with a multiple regression analysis (SAS, 1982). The dependent variable used in the regression analysis was the 42-day sporophyte density adjusted for the initial gametophyte density:
adjusted density = log,@,, + 1)/(1,,/2))
where D,, is the average sporophyte density after 42 days of outplant and I, is the initial density of gametophytes. The initial density is divided by 2 so that only female gametophytes are taken into account for this calculation. We have assumed that the equal proportions of male and female gametophytes are produced. Casual observations of gametophyte cultures indicate that this is a fair assumption. Initial analyses performed to find the regression model which best fit the data indicated that a log, transformed model had a higher overall R2 than a linear model. Log, transformed variables, therefore, were used in all the regression equations for sporophyte recruitment.
The regression analyses were first performed using all four of the physicochemical variables for each of the depth classes. Separate analyses were performed for each depth and for all depths combined. The variables which were not significant at P < 0.10 were eliminated and the analysis was rerun using only the significant factors as independent variables. We also computed determinations of partial correlation, which provide the variance explained by the independent variable when the other variables are held constant (Belsley et al., 1980).
50 LAWRENCE E. DEYSHER AND THOMAS A. DEAN
Occasionally there were no irradiance or temperature data available from a particular location because of instrument fake. When possible, we estimated irradiance and temperature values for these cases using data from instruments at the other depths from the stations. The estimates were made using the long term relationships between values at the two depths. Estimates were required for < 10% of the 275 observations used in the analysis.
RESULTS
PATTERNS OF SPOROPHYTE RECRUITMENT
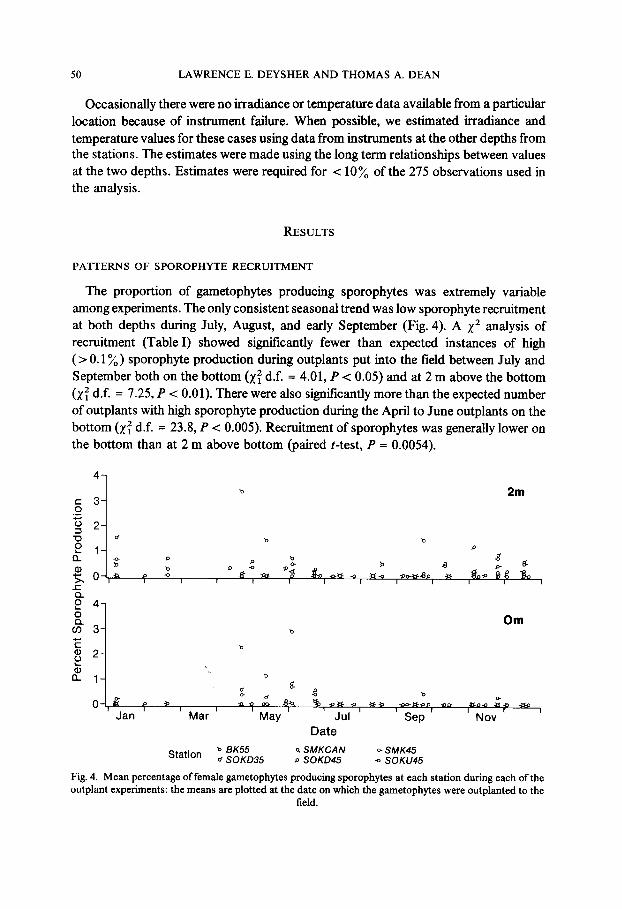
The proportion of gametophytes producing sporophytes was extremely variable among experiments. The only consistent seasonal trend was low sporophyte recruitment at both depths during July, August, and early September (Fig. 4). A x2 analysis of recruitment (Table I) showed significantly fewer than expected instances of high ( > 0.1%) sporophyte production during outplants put into the field between July and September both on the bottom (x: d.f. = 4.01, P < 0.05) and at 2 m above the bottom (x: d.f. = 7.25, P < 0.01). There were also significantly more than the expected number of outplants with high sporophyte production during the April to June outplants on the bottom (x: d.f. = 23.8, P < 0.005). Recruitment of sporophytes was generally lower on the bottom than at 2 m above bottom (paired t-test, P = 0.0054).
b 2m
Om b
b
Date
Station b 665 9 SMKCAN c- SMK45 @ SOKD35 P SOKD45 Q SOKlJ45
Fig. 4. Mean percentage of female gametophytes producing sporophytes at each station during each of the outplant experiments: the means are plotted at the date on which the gametophytes were outplanted to the
field.

MACROCYSTIS RECRUITMENT 51
TABLE I
x2 analysis of recruitment data from the gametophytes outplants to both 0 m and 2 m above the bottom: the three recruitment classes are based on the percentage of kmale gametophytes producing sporophytes.
Bottom recruitment classification
0 >O<O.l >O.l
Jan.-Mar. 6 6 1 Apr.-June 5 11 13 July-Sept. 12 18 1 Oct.-Dec. 11 24 1
Overall x2 = 30.6, P < 0.005
2 m above the bottom recruitment classification
0 >o < 0.1 >O.l
Jan.-Mar. 0 6 1
Apr.-June 0 16 15 July-Sept. 3 24 4 Oct.-Dec. 3 17 17
Overall x2 = 14.9, P i 0.05
The sporophyte densities on the uninoculated outplant control were always less than those on the inoculated substrata. Of the 275 observations of sporophyte recruitment on inoculated substrata, 160 of the controls had zero density and only 12 had sporophyte densities > 5 sporophytes . cm- 2. The natural settlement of zoospores on the substrata, therefore, appeared to be an insignificant factor in these experiments. The densities in the controls paralleled the inoculated substrata and appeared to result primarily from newly produced sporophytes being dislodged from gametophytes on the inoculated substrata and settling on the nearby control. This reattachment of microscopic sporophytes has been noted by North (1976), Moss et al. (198 l), and Chapman (1984).
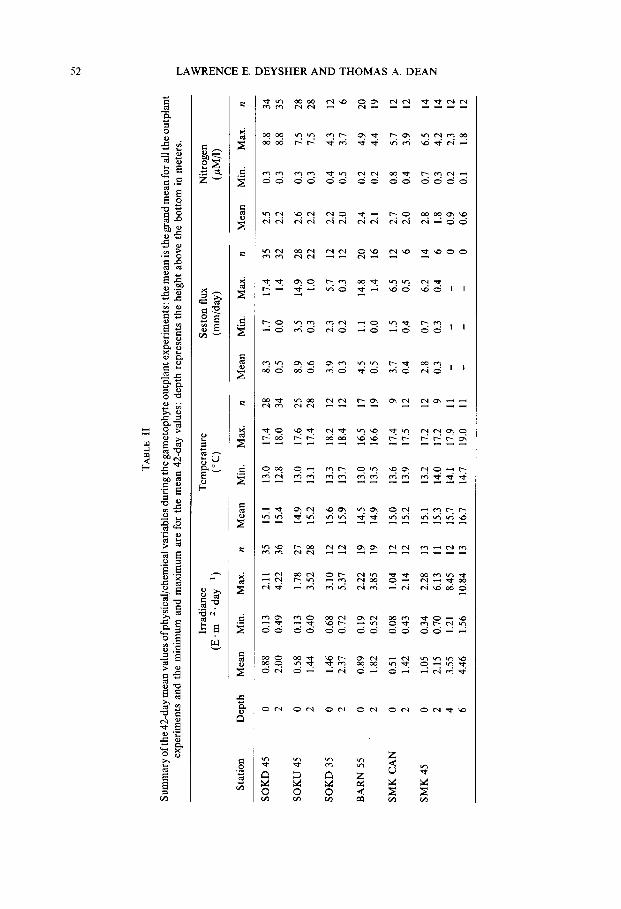
PHYSICOCHEMICAL MEASURES
It-radiances at the bottom were approximately one half of those at 2 m above the bottom (Table II). The lowest n-radiances were found under the Mucrocystis canopy in the San Mateo kelp bed (SMK CAN) and at the SOKU 45 station. The highest mean n-radiances were at the shallower inshore station (SOKD 35). Rates of seston flux were also greater on the bottom than at 2 m above the bottom. The mean rate on the bottom was 5.1 mm * day- ’ and the rate at 2 m above the bottom was ten times lower at 0.4 mm - day- ‘, suggesting that high seston flux was primarily due to resuspension of
TA
BLE
II
Sum
mar
y of
the
42-d
ay
mea
n va
lues
of
phys
icah
chem
ical
va
riab
les
duri
ng
the
gam
etop
hyte
ou
tpla
nt
expe
rim
ents
: th
e m
ean
is t
he g
rand
m
ean
for
all
the
outp
lant
ex
peri
men
ts
and
the
min
imum
an
d m
axim
um
are
for
the
mea
n 42
-day
va
lues
; de
pth
repr
esen
ts
the
heig
ht
abov
e th
e bo
ttom
in
met
ers.
Stat
ion
Irra
dian
ce
Tem
pera
ture
Se
ston
fl
ux
Nitr
ogen
(E
.m
-‘.d
ay-r
) (“
C)
(mm
/day
) (P
M/I
)
Dep
th
Mea
n M
in.
Max
. n
Mea
n M
in.
Max
. n
Mea
n M
in.
Max
. n
Mea
n M
in.
Max
. n
SOK
D
45
0 0.
88
0.13
2.
11
35
15.1
13
.0
17.4
28
8.
3 1.
7 17
.4
35
2.5
0.3
8.8
34
2 2.
00
0.49
4.
22
36
15.4
12
.8
18.0
34
0.
5 0.
0 1.
4 32
2.
2 0.
3 8.
8 35
SOK
U
45
0 0.
58
0.13
1.
78
27
14.9
13
.0
17.6
25
8.
9 3.
5 14
.9
28
2.6
0.3
7.5
28
2 1.
44
0.40
3.
52
28
15.2
13
.1
17.4
28
0.
6 0.
3 1.
0 22
2.
2 0.
3 7.
5 28
SOK
D
35
0 1.
46
0.68
3.
10
12
15.6
13
.3
18.2
12
3.
9 2.
3 5.
7 12
2.
2 0.
4 4.
3 12
2 2.
37
0.72
5.
37
12
15.9
13
.7
18.4
12
0.
3 0.
2 0.
3 12
2.
0 0.
5 3.
7 6
BA
RN
55
0
0.89
0.
19
2.22
19
14
.5
13.0
16
.5
17
4.5
1.1
14.8
20
2.
4 0.
2 4.
9 20
2 1.
82
0.52
3.
85
19
14.9
13
.5
16.6
19
0.
5 0.
0 1.
4 16
2.
1 0.
2 4.
4 19
SMK
C
AN
0
0.51
0.
08
1.04
12
15
.0
13.6
17
.4
9 3.
7 1.
5 6.
5 12
2.
7 0.
8 5.
7 12
2 1.
42
0.43
2.
14
12
15.2
13
.9
17.5
12
0.
4 0.
4 0.
5 6
2.0
0.4
3.9
12
SMK
45
0
1.05
0.
34
2.28
13
15
.1
13.2
17
.2
12
2.8
0.7
6.2
14
2.8
0.7
6.5
14
2 2.
15
0.70
6.
13
I1
15.3
14
.0
17.2
9
0.3
0.3
0.4
6 1.
8 0.
3 4.
2 14
4 3.
55
1.21
8.
45
12
15.7
14
.1
17.9
11
-
- -
0 0.
9 0.
2 2.
3 12
6 4.
46
1.56
10
.84
13
16.7
14
.7
19.0
11
-
- -
0 0.
6 0.
1 1.
8 12
MACROCYSTIS RECRUITMENT 53
sediments from the bottom. Temperatures were generally similar among stations and averaged 15.2 “C on the bottom and 15.4 “C at 2 m above the bottom from all periods when outplants were in the field. The lowest 42-day average was 12.7 “C and the highest was 18.4 “C. Mean nitrogen concentrations ranged from 0.2 to 8.8 PM al- ’ with an overall mean of 2.5 PM * l- ‘.
RELATIONSHIPS BETWEEN SPOROPHYTE RECRUITMENT AND PHYSICOCHEMICAL
FACTORS
Temperature was negatively correlated with sporophyte recruitment and explained more of the variability in recruitment than any of the other independent variables (Table III). Temperature was also the only variable which was significant in analyses for all depths. Irradiance showed a significant positive correlation with recruitment in all situations except for the substrata in the water column (> 2 m above the bottom) analyzed separately from substrata on the bottom.
TABLE III
Summary of regression analyses of sporophyte recruitment at various depth levels.
Depth R 2 of overall Significant
model variables Regression coefficient
Partial R2
0 0.25 Temp. - 13.1 0.24 Irr. 0.9 0.17
234, 6 0.27 Temp. - 11.6
0, 2 0.33 Temp. - 12.1 0.24 Irr. 0.8 0.10
Sed. - 0.2 0.03
0, 2, 4, 6 0.27 Temp. - 14.0 0.26 Irr. 0.9 0.16
Seston flux was not significantly correlated with sporophyte recruitment on the bottom substrata, but explained a small but significant proportion of the variance for substrates in the water column.
There were no significant effects of nitrogen noted in any of the analyses. The lack of a stronger correlation with nitrogen may have been because nitrogen was sampled too infrequently to provide a good estimate of the nitrogen levels that gametophytes actually experienced. In addition, the nitrogen levels in the microenvironments around the gametophytes may be different from nitrogen concentrations in the water column because of the regeneration of nitrogen by organisms living on the bottom. Because of a correlation between temperature and nutrients and the high frequency variability in nutrient levels, daily temperatures may actually provide a better estimator of changes in nutrients than bi-weekly nutrient samples (Zimmerman & Kremer, 1984).

54 LAWRENCE E. DEYSHER AND THOMAS A. DEAN
The recruitment of sporophytes from g~etoph~es was plotted against both temper- ature and irradiance to examine the relationships between these factors, especially any threshold responses or.interactive effects,(Fig. 5). The highest t~~rat~e at which recruitment occurred was 18.3 “C and the lowest irradiation level was 0.1 E. m- * + day- ‘. Sporophyte production by > 0.1 y0 of the female gametophytes, which represented approximately the top 15% of recruitment densities, occurred only at temperatures < 16.3 “C and it-radiances > 0.4 E - m - ’ . day- ‘. Greater than 1.0% of the gametoph~es produced sporoph~es only at temperat~es -K 14 “C and at irradiances > 0.4 E * m- 2 - day- ’ . These conditions, therefore, can be considered as being optimal for sporophyte recruitment. A possible interactive effect between tempera- ture and irradiance was also noted. Higher irradiance levels (approximately 1 E - m - ’ 1 day- ‘) appeared to be required for production of large numbers of sporophytes as temperatures approached 16 “C.
The ~omb~ations of temperature and irradiance that fall within the boundaries of 16.3 “C and 0.4 E * m- 2 * day- ’ can be considered “rec~tment windows”, i.e., times when conditions are favorable for gametogenesis and subsequent kelp recruitment.
Sporophyte Recruitment Response Surface
,
12 13 14 15 16 17 18 19
Mean Temperature (“C) ~ercenfage Q
Producing Sporophytes 0 0 . O-&Of l 0.07-0.1 e @i-1.0 0 > 7.0
Fig. 5. Percentage of female gametophytes producing sporophytes plotted against both mean temperature and irradiance values from the co~es~nding location and time: the outer solid line represents the boundary of temperature and irradiance conditions outside of which no sporophytes were produced; the inner solid line represents the boundary for sporophyte production by 0.01% of female gametophytes; the dashed line shows the boundary for >O.l % sporophyte production, and the dotted line shows the boundary for z 1.0%
production.
MACROCYSTIS RECRUITMENT 55
However, it is also evident that not all outplants that fell within the window produced a large number of sporophytes. There were many instances when conditions appeared favorable but < 0.1% of the females produced sporophytes.
NUTRIENT ENRICHMENT STUDIES
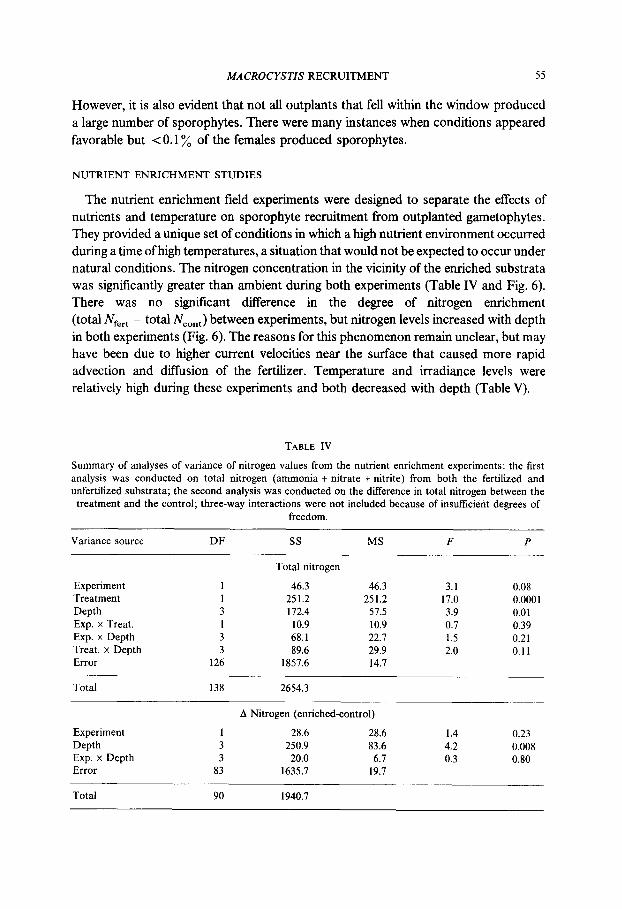
The nutrient enrichment field experiments were designed to separate the effects of nutrients and temperature on sporophyte recruitment from outplanted gametophytes. They provided a unique set of conditions in which a high nutrient environment occurred during a time of high temperatures, a situation that would not be expected to occur under natural conditions. The nitrogen concentration in the vicinity of the enriched substrata was significantly greater than ambient during both experiments (Table IV and Fig. 6). There was no significant difference in the degree of nitrogen enrichment (total N,,,, - total NC,“, ) between experiments, but nitrogen levels increased with depth in both experiments (Fig. 6). The reasons for this phenomenon remain unclear, but may have been due to higher current velocities near the surface that caused more rapid advection and diffusion of the fertilizer. Temperature and irradiance levels were relatively high during these experiments and both decreased with depth (Table V).
TABLE IV
Summary of analyses of variance of nitrogen values from the nutrient enrichment experiments: the first analysis was conducted on total nitrogen (ammonia + nitrate + nitrite) from both the fertilized and unfertilized substrata; the second analysis was conducted on the difference in total nitrogen between the
treatment and the control; three-way interactions were not included because of insufficient degrees of
freedom.
Variance source DF ss MS F P
Experiment 1 46.3 46.3 3.1 0.08 Treatment 1 251.2 251.2 17.0 0.0001 Depth 3 172.4 51.5 3.9 0.01 Exp. x Treat. 1 10.9 10.9 0.7 0.39 Exp. x Depth 3 68.1 22.7 1.5 0.21 Treat. x Depth 3 89.6 29.9 2.0 0.11 Error 126 1857.6 14.7
Total nitrogen
Total 138 2654.3
A Nitrogen (enriched-control)
Experiment 1 28.6 28.6 1.4 0.23
Depth 3 250.9 83.6 4.2 0.008 Exp. x Depth 3 20.0 6.7 0.3 0.80 Error 83 1635.7 19.7
Total 90 1940.7
56 LAWRENCE E. DEYSHER AND THOMAS A. DEAN
I_-o+ l-21 l-4-l t-6-i DEPTH
Fig. 6. Average total nitrogen levels (PM/I) on the fertilized (enriched) and unfertilized (control) substrata at 0,2,4 and 6 m above the bottom during the two nutrient enrichment experiments: these experiments were outplanted to SMK 45 on 6 August 1981 (A) and 10 September 1981 (B); the vertical bars indicate
one standard error of the mean.
TABLE V
Mean sporophyte densities (42 day) and associated physicochemical variables at various heights above the bottom for the nutrient addition experiments: the dates are the outplant date of the experiment.
Temperature Irradiation
(“Cl (E.m -**day-‘)
6 August 1981 Om 2m 16.6 6.1 4m 8.6 6m 18.4 11.1
10 September 1981 Om 16.9 2.3 tm 4m 6m 18.1 4.8
Mean sporophyte densities were signiftcantly higher on the enriched substrata than on the unfertilized controls in each of the experiments (P < 0.005, Wilcoxon matched pairs signed ranks test) (Fig. 7). In the first experiment, sporophyte recruitment on enriched substrata decreased with distance from the bottom in a manner similar to

MACROCYSTIS RECRUITMENT 5-l
6-
4-
2-
+0-l l-2-l l-4-l i-6-l DEPTH
Fig. 7. Mean sporophyte densities on fertilized (enriched) and unfertilized (control) gametophyte outplant substrata at 0, 2, 4 and 6 m above the bottom in the two nutrient enrichment experiments: sporophyte densities were determined at 42 days after outplanting; experiment 1 was outplanted on 6 August 1981 (A) and Experiment 2 on 10 September 1981 (B); the vertical bars indicate one standard error of the mean.
nitrogen concentrations. However, in the second experiment, sporophyte recruitment was highest at 6 m above the bottom. This may have been because of the relatively low light levels during the second experiment and possibly interactions between light and nutrient uptake. Although there were unexplained interactions among depths and experiments, it is clear that fertilizing substrata increased sporophyte production even at the high temperatures observed during these experiments.
DISCUSSION
Gametophytes of Macrocystis pynira, like those of most laminarian species, will undergo gametogenesis at relatively low light levels (Cosson, 1975; King & Schramm, 1976; Luning & Neushul, 1978). Recent laboratory studies on the relationship between gametogenesis and h-radiance in laminarian gametophytes (Llining, 1980; Deysher & Dean, 1984, 1986; Novaczek, 1984) indicate that a distinct irradiation threshold exists for gametophyte fertility. The relationships between fertility and daily quantum dose in these studies are steep exponential functions (almost step functions) in which fertility is quickly saturated at h-radiances above the lowest level which is sufficient to induce

58 LAWRENCE E. DEYSHER AND THOMAS A. DEAN
fertility. For M. pyriferu, fertility is induced at irradiation levels as low as 0.2 ET m - ’ * day - ’ and 100% of the gametophytes produce sporophytes at irradiances above 0.4 E. m - 2 - day- ’ (Deysher & Dean, 1984). In the field studies described here, the lowest irradiance at which sporophytes were produced was 0.1 E - m - * + day- ’ and > 0.1 y0 of the female gametophytes produced sporophytes only at irradiances above 0.4 E * m- 2 * d- i. The use of an irradiation level of 0.4 E * m - *. d - ’ in defining the “recruitment window” for M. pyrifera gametophytes is therefore consistent with levels required for high levels of sporophyte rec~itment in the laboratory (Deysher & Dean, 1984).
Ltining (1980) has emphasized the importance of the total quantum dose of blue light (400-512 nm) in stimulating gametogenesis in laminarian gametophytes. In the three species of Laminariu which he studied, the blue light quantum dose producing 50% fertility at 15 “C ranged from 2.1 to 5.6 E/m2. The blue light quantum dose required for 50% fertility in M. ~~~~~~ is 1.1 to 3.1 E ’ m- ’ * day- * (Deysher & Dean, 1984). In the present field study the blue quantum dose for sporophyte densities in the top 15 percentile was approximately 4.5 E/m’, slightly higher than observed for M. pynifera in the laboratory. The blue quantum dose in our field study was calculated by assuming that 40 y0 of the light received was in the range 400-5 10 nm and that average time from zoospore settlement to g~etogenesis was 28 days. This time period was calculated by taking the average length of sporophytes at 42 days (0.8 mm) and extrapolating back to the approximate time of gametogenesis using an average sporophyte growth rate of 65% . day- ’ (Deysher & Dean, unpubl. data). The value of 40% for the proportion of blue light is the average proportion observed in the vicinity of San Onofre (Zabloudil, 1984). The blue light fraction showed a strong log-linear correlation with total irradiance (r2 = 0.91). Therefore, variations in the blue light fraction would not be expected to cause failures of gametogenesis independent of the effects of total it-radiance.
The low partial coefficient for irradiance in the regression analyses is not surprising given that the relationship between irradiance and recruitment appears to be non-linear. Irradiances above the apparent threshold explain little of the observed variability in sporophyte recruitment. This can be seen in the regression analysis for substrata in the water column where it-radiance did not explain a signilicant portion of the variability in sporophyte recruitment. Irradiances at these locations were always above the threshold level for sporophyte production. On the bottom, however, irradiances ranged both above and below the threshold level and explained a significant portion of the v~iability in sporoph~e recrui~ent.
Abrasion and burial by sediments are known to be sources of mortality for kelp gametophytes (Devinny & Volse, 1978) but these factors explained little of the variability in sporophyte recruitment. The lack of a stronger effect may have been due to the generally strong negative correlation between seston flux and irradiance (Dean, 1985). Seston flux explained a significant portion of v~ab~ty in sporoph~e remitment at 2 m above the bottom, but not on bottom substrata. The lack of a significant correlation on the bottom may also have resulted from consistently high rates of seston flux and

MACROCYSTIS RECRUITMENT 59
consistently high rates of gametophyte mortality. At 2 m above the bottom, there may have been more variability in seston lhrx over the range critical to gametophytes.
Temperature explained the highest proportion of the variability in sporophyte recruit- ment. The highest densities of sporophytes occurred at temperatures below 14 “C and the upper temperature limit for sporophyte production by > 0.1 yQ of female gametophytes (a density of approximately 50 sporophytes - cm - 2, was 16.3 “C. Labo- ratory studies on MZC~OC_Y&S pyrifera gametophytes by Ltining & Neushul (1978), however, showed an optimal temperat~e range of 12 to 17 ‘C for g~etogenesis with no fertility at 7 and 20 o C. Deysher & Dean (1986) found 100% sporophyte recruitment over temperatures ranging from 11 to 19 “C in a laboratory study on gametophytes produced from sporophytes from southern California. Based on these laboratory studies, we expected that high densities of sporophytes would have been produced in the outplant experiments at temperat~es up to 17 “C. The low fertility values at temperatures at 17 “C and above suggest that some factor correlated with temperature, such as nutrient availability, was inhibiting recruitment at high temperatures. The results of the nutrient enrichment experiment support this conclusion. The enrichment of nitrogen and phosphorus in the vicinity of the gametophyte populations increased sporophyte recruitment, albeit at low densities, at temperatures up to 18.1 “C. It appears that the significant effect of temperature in the regression analyses is a reflection of the correlation between temperature and nitrogen which exists in the southern California nearshore environment (Jackson, 1977; Zentara & Kamykowski, 1977; Zimmerman & Kremer, 1984). Because of this correlation and because of high frequency temporal variability in nutrient levels, hourly temperature records may provide a better estimator of nitrogen availability than b&weekly nitrogen samples (Zimme~~ & Kremer, 1984). The case for temperature serving as proxy for nutrients is further supported by the similarity of the upper temperature boundary of the defined recruitment window, 16.3 “C, and the temperature level, 15.5 “C, above which nitrogen is uniformly low (Zimmerman & Kremer, 1984). Although temperature may not be the factor controlling rec~itment, it serves as a good index variable because it is more easily measured than the nutrient components of upwelled water such as nitrogen and various trace minerals which are essential to gametogenesis (Kuwabara & North, 1980).
The relationship between sporophyte recruitment, temperature, and irradiance sug- gests that there may be some interaction between ii-radiance and temperature (or nutrients) such that higher irradiance levels are required as temperature increases. However, laboratory studies indicate no such interaction between temperature and irradiance, at least over the range of temperatures observed in this field study (Deysher & Dean, 1986). There may be some interaction between nutrients and irradiauce, as suggested by ii-radiance dependent nitrate uptake in the sporophyte stage (Wheeler, 1978; Gerard, 1982). On the other hand, the suggested interaction may be an artifact due to the lack of low ii-radiance values at higher temperatures resulting from the positive correlation between field values of it-radiance and temperature (Fig. 5).
The observed patterns of Macrocystispyrifera abundance and distribution in southern

60 LAWRENCE E. DEYSHER AND THOMAS A. DEAN
California and elsewhere indicate that sporophyte recruitment is a critical component of the population dynamics of this species. Long term studies in three southern California kelp forests (Rosen~~ et al., 1974; Wilson et al., 1980; Dayton et al., 1984) show that M. pyriferu populations undergo pronounced fluctuations in adult density. During these fluctuations, densities of adult plants decline over a period of 2 to 3 yr primarily due to dislodgement caused by storms. Pulses of sporophyte recruitment often follow thinning of adult populations (Gerard, 1976; Pearse & I-Iines, 1979; Kimura & Foster, 1984; Reed & Foster, 1984; Santelices & Ojeda, 1984), suggesting that shading by adult sporophytes may reduce n-radiance and thereby limit recruitment.
Although pulses of recruitment may be stimulated by increased ii-radiance on the bottom due to decreased canopy of an adult population, increased n-radiance is not a sufficient condition in itself for recruitment. As shown in these outplant studies, rec~itment of new sporophytes will only take place when irradiance levels are above a threshold level and temperatures are below approximately 16 “C. Recruitment in southern California kelp forests, therefore, is, in part, regulated by changes in large scale oceanographic events which control patterns of light transmission and the input of cold, nutrient rich water into the shallow coastal zone. In central California recruitment occurs on a more annual basis. This is probably because water temperatures are lower (and associated nutrient levels higher) and the surface canopies of ~~c~~cy~~~ are regularly thinned by winter storms (Dayton et al., 1984). In a study on the gametophytes of Laminarkz saccharina in British Columbia, Hsaio & Druehl (1973) found that the production of sporophytes from gametophytes in the field was potentially limited by ii-radiance and not nutrients, again reflecting the higher nutrient levels in this region. Althou~ ii-radiance is often high enough and temperat~e low enough for remitment in southern California, the joint probability of high n-radiance and low temperature is small. We are currently analyzing the continuous records of temperature and irradiance to determine this probability and its relationship to natural recruitment events. These analyses are beyond the scope of this paper and will be presented in a future publication.
Although proper temperature and irradiance levels are necessary requisites for recruitment of sporophytes, not all apparent “recruitment windows” resulted in success- ful recruitment. This suggests that factors other than those measured in this study can also affect recruitment success. One factor that could be especially important is wave surge. High surge would increase the abrasion of seston particles, especially on the bottom, and thereby increase the mortality rate of gametophytes and recruited sporoph~es. Various biological factors, such as grazing and the settlement of other organisms, could also adversely affect the recruitment of sporophytes. On our outplant substrata the effects of these biological factors are minimized by the design of the outplant racks. Also, biological effects appeared to be very patchy both in space and time. Some of the outplant plates had ropes which suffered obvious grazing, but in none of these cases did all the ropes on the all plates suffer damage. Therefore, biological effects would be expected to increase the variance, i.e. noise, of the relationships that we have observed, but not significantly alter the relationships themselves.

MACROCYSTZS RECRUITMENT 61
Although sporophyte recruitment is controlled by a host of environmental factors, knowledge of the boundary conditions for important phys~coche~c~ variables such as irradiance, nutrients, temperature, and seston flux provide a usefti index in describing the potential ~~c~~y~?~ pyrifera habitat (sensu Whittaker etal., 1973). The total description of the physical habitat would allow an assessment of the importance of nonphysical environmental parameters which potentially control the distribution of this species. For example, the absence of&f. pyrifera from an area where physical parameters appear favorable for sporophyte recruitment would be an indication that biological factors such as predation or lack of dispersal were responsible for con~o~ng the distribution in this region. Additionally, an understanding of the role which physical factors play in the recruitment of M. pyriferu is important in regulating human-induced mo~cations to the marine en~o~ent which alter the physical characteristics of nearshore waters. The protection of environmentally important species, such as M. pynira, is enhanced when the boundary conditions within which these alterations must be contained are well defined.
ACKNOWLEDGEMENTS
We thank all the present and past members of the Kelp Ecology Project who made this study possible. Special recognition is due to L. Asakawa, F. Jacobsen, S. Lagos, K. Thies, C. Criger, S. Medler, K. Parker, and members of the Marine Review Com- mittee and their advisors, especially Allan Stewart-Oaten. P. Dayton and M. Foster reviewed a draft of the manuscript. The study was funded by the Marine Review Committee, Inc. (MRC), Encinitas, California. The MRC does not necessarily accept the results, findings, or conclusions stated herein.
REFERENCES
ANDERSON, E.K. & W.J. NORTH, 1967. Zoospore release rates in giant kelp, Macrocystis. Bull. Sough Calij Acad. Sci., Vol. 66, pp. 223-232.
BELSLEY, DA, E. KUH & R.E. WELSCH, 1980. Regression diagnostics. John Wiley & Sons, New York, 292 pp.
BIRKLAND, C., 1982. Terrestrial runoff as a cause of outbreaks of Acant~s~e~ planei (Echino- dermata: Asteroidea). Mar. Biol., Vol. 69, pp. 175-185.
CAFFEY, H.M., 1985. Spatial and temporal variation in settlement and recruitment of intertidal barnacles. Ecol. Monogr., Vol. 55, pp. 313-332.
CHAPMAN, A.R.O., 1984. Reproduction, recruitment, and mortality in two species of Larninariu in southwest Nova Scotia. J. Exp. Mar. Biol. Ecol., Vol. 78, pp. 99-109.
CONNELL, J. H., 1985. The consequences of variation in initial settlement vs. post-settlement mortality in rocky intertidal communities. .I. Exp. Mar. Biol, Eeol., Vol. 93, pp. 1 l-45.
COSSON, J., 1975. Action des conditions d’Clairement sur la croissance des gametophytes de Laminatiu digitutu (L.) Lamouroux (Pheophy&e, Laminariale). Bulf. Soe. Phycol. Fr., Vol. 20, pp_ 50-54.
DAYTON, P.K. & M.J. TEGNER, 1984. Catastrophic storms, El NiiIo, and patch stability in a southern California kelp community. Science, Vol. 224, pp. 283-285.

62 LAWRENCE E. DEYSHER AND THOMAS A. DEAN
DAYTON, P.K., V. CURRIE, T. GERRODETTE, B. KELLER, R. ROSENTHAL & D.V. TRESCA, 1984. Patch dynamics and stability of some California Kelp Communities. Ecol. Monogr., Vol. 54, pp. 253-289.
DEAN, T. A., 1985. The temporal and spatial distribution of underwater quantum irradiation in a southern California kelp forest. Esfuarine Coastal Shelf&i., Vol. 21, pp. 835-844.
DEAN, T. A. & F. R. JACOBSEN, 1984. Growth of juvenile Macrocystlr pyriira (Laminariales) in relation to environmental factors. Mar. Biol., Vol. 83, pp. 301-311.
DEAN, T. A. L F. R. JACOBSEN, 1986. Nutrient-limited growth of juvenile kelp, Macrocysris pynifra, during the 1982-1984 “El Nifio” in southern California. Mar. Biol., Vol. 90, pp. 597-601.
DEVINNY, J. S. & L. A. VOLSE, 1978. The effects of sediments on the development of Macrocystis pyn$.w gametoph~es. Mar. Biol., Vol. 48, pp. 343-348.
DEYSKER, L.E. & T.A. DEAN, 1984. Critical irradiance levels and the interactive effects of quantum irradiance and dose on gametogenesis in the giant kelp, Macrocystis pyn@ra. J. Phycol*, Vol. 20, pp. 520-524.
DEYSHER, L. E. & T.A. DEAN, 1986. Interactive effects of light and temperature on sporophyte production in the giant kelp, Macrocysrb pyrttra. Mar. Biol., (In press).
DOHERTY, P. J., 1983. Tropical territorial damselfishes: is density limited by aggression or recruitment? Ecology, Vol. 64, pp. 176-190.
EBERT, T.A., 1983. Recruitment in echinoderms. In, Echinodem Studies, Vol. 1, edited by M. Jangoux & J. M. Lawrence, A. A. Balkema, Rotterdam, pp. 169-203.
FOSTER, M. S., T.A. DEAN & L.E. DEYSHER, 1985. Subtidal techniques. In, Handbook of phycological methods: e~o~o~~a~~eId methods@ macroalgae, edited by M. M. Littler, Cambridge University Press, England, pp. 199-231.
GAINES, S. & J. ROUGHGARDEN, 1985. Larval settlement rate: a leading determinant of structure in an ecological community of the marine intertidal zone. Proc. Nuff. Acad. Sci., Vol. 82, pp. 3707-3711.
GERARD, V. A., 1976. Some aspects of the material dynamics and energy flow in a kelp forest in Monterey Bay, California. Ph.D. thesis, University of California at Santa Cruz, 173 pp.
GERARD, V.A., 1982. In situ rates of nitrate uptake by giant kelp Macrocystispyriifera (L.) C. Agardh: tissue differences, environmental effects, and predictions of nitrogen-limited growth. J. Exp. Mar. Biol. Ecal., Vol. 62, pp. 21 l-224.
HSAIO, S. I. & L. D. DRUEHL, 1973. Environmental control of gametogenesis in Laminaria saccharina. IV. In situ development of gametophytes and young sporophytes. J. Phycol., Vol. 9, pp. 160-164.
JACKSON, G. A., 1977. Nutrients and production of giant kelp, Macrocysfispyn@ra, off Southern California. Litnnol. Ocean., Vol. 22, pp. 979-995.
KAIN, J.M., 1979. A view of the genus Lam~a~a. Oceanogr. Mar. Bid. Ann. Rev., Vol. 17, pp. IOl-161. KIMURA, R. S. & M. S. FOSTER, 1984. The effects of harvesting Macrocysr~p~~ra on the algal assemblage
in a giant kelp forest. ~ydrobiolo~u, Vol. 116/l 17, pp. 425-428. KING, R. J. & W. SCHRAMM, 1976. Photosynthetic rates of benthic algae in relation to light intensity and
seasonal variations. Mar. Biol., Vol. 37, pp. 215-222. KUWABARA, J. S. & W.J. NORTH, 1980. Culturing microscopic stages ofMacrocystispyn@a (Phaeophyta)
in Aquil, a chemically defined medium. J. Phycol., Vol. 16, pp. 546-549. LONING, K., 1980. Critical levels of light and temperature regulating the gametogenesis of three Lam&aria
species (Phaeophyceae). J. Phycol., Vol. 16, pp. l-15. LUNING, K. & M. NEUSHUL, 1978. Light and temperature demands for growth and reproduction of
laminarian gametophytes in Southern and Central California. Mar. Biol., Vol. 45, pp. 297-310. Moss, B. L., D. TO~EY & P. COURT, 1981. Kelps as fouling organisms on North Sea platforms. Bof. Mar.,
Vol. 24, pp. 207-209. NORTH, W., 1971. Intr~uction and background. In, The Biolom of Giant Ke& Beds (~acrocysr~) in
Cul&mia, edited by W. North, Nova Hedr;igia, Suppl. to Vol. 32, pp. l-97. NORTH, W., 1976. Aquacultural techniques for creating and restoring beds of giant kelp, Macrocyst& spp.
J. Fish. Res. Board Can., Vol. 33, pp. 1015-1023. NOVACZEK, I., 1984. Response ofE&onia radiata (Laminariales) to light at 15 “C with reference to the field
light budget at Goat Island Bay. N.Z. Mar. Biol., Vol. 80, pp. 263-272. PEARSE, J. S. & A. H. HINES, 1979. Expansion ofa central California kelp forest following the mass mortality
of sea urchins. Mar. Biol., Vol. 51, pp. 83-91.

MA CROCYSTIS RECRUITMENT 63
REED, D. C. & M. S. FOSTER, 1984. The effect of canopy shading on algal rec~~ent and growth in a kelp forest. Ecology, Vol. 65, pp. 937-948.
ROSENTHAL, R. J., W. D. CLARKE & P. K. DAYTON, 1974. Ecology and natural history of a stand of giant kelp, Macrocysrir pyrifru, off Del Mar, California. Ftih. BuU. NOAA, Vol. 72, pp. 670-684.
ROUGHGARDEN, J., Y. IWASA & C. BAXTER, 1985. Theory of population processes for marine organisms. I. Demography of an open population with space-limited recruitment. Ecology, Vol. 66, pp. 54-67.
SAMELICES, B. & F.P. OJEDA, 1984. Effects of canopy removal on the understorey algal community structure of coastal forests of Macr~yst~ pynzru, from southern South America. Mar. Ecoi. Prog. Ser., Vol. 14, pp. 165-173.
SAS, 1982. SAS Users Guide: Statistics. SAS Institute Inc. Gary, North Carolina, pp. 3-110. UNDERWOOD, A. & L. DENLEY, 1984. Paradigms, explanations and generalizations in models for the
structure of intertidal communities on rocky shores. In, Ecological Communities: Conceptwd Issues and the Evidence, edited by D. R. Strong, D. Simberloff, L. G. Abele & A. B. Thistle, Princeton University Press, Princeton, New Jersey, pp. 15 l-1 80.
WATANABE, J.M., 1984. The influence of recruitment, competition, and benthic predation of spatial distributions of three species ofkelp forest gastropods (Trochidae : Tegula). Ecology, Vol. 65, pp. 920-936.
WHEELER, B.N., 1978. Ecophysiological studies on the giant kelp, Macrocystis. Ph.D. thesis, University of California at Santa Barbara, 179 pp.
WHITTAKER, R.H., S.A. LEVIN & R.B. ROOT, 1973. Niche, habitat, and ecotope. Am. Nat., Vol. 107, pp. 321-338.
WILSON, C.K., A. J. MEARNS & J.J. GRANT, 1980. Changes in kelp forests at Palos Verdes. In, Coastal Water Research Project, biennial report for the years 19794980. Southern California Coastal Water Research Project, Long Beach, California, pp. 77-92.
YOSHIOKA, P.M., 1982. Role of planktonic and benthic factors in the population dynamics of the bryozoan ~embran~ora membranacea. ~colo~, Vol. 63, pp. 457-468.
ZABLOIJDIL, K.A., 1984. A micro-powered underwater logger for recording photosynthetically active radiation and illumination. Ocean Optics VII, edited by M. A. Blizzard, Proceedings SPIE - International Society of Optical Engineering, Vol. 489, pp. 375-379.
ZENTARA, S. J. & D. KAMYKOWSKI, 1977. Latitudinal relationships among temperature and selected plant nutrients along the west coast of North and South America. J. Mar. Res., Vol. 85, pp. 321-336.
ZIMMERMAN, R.C. & KREMER, J.N., 1984. Episodic nutrient supply to a kelp forest ecosystem in southern California. J. Mur. Res., Vol. 42, pp. 591-604.


















