
Impact of Tapes philippinarum farming on nutrient dynamics and benthic respiration in the Sacca di...
10
Hydrobiologia 455: 203–212, 2001. © 2001 Kluwer Academic Publishers. Printed in the Netherlands. 203 Impact of Tapes philippinarum farming on nutrient dynamics and benthic respiration in the Sacca di Goro Marco Bartoli 1,∗ , Daniele Nizzoli 1 , Pierluigi Viaroli 1 , Edoardo Turolla 2 , Giuseppe Castaldelli 2 , Elisa Anna Fano 2 & Remigio Rossi 2 1 Department of Environmental Sciences, Parma University, viale delle Scienze, 43100 Parma, Italy 2 Department of Evolutive Biology, Ferrara University, via Borsari, 44100 Ferrara, Italy Tel: 0521-905976, Fax: 0521-905402; E-mail: [email protected] Received 9 June 2000; in revised form 9 April 2001; accepted 20 April 2001 Key words: clam culture, surficial sediments, benthic fluxes, anoxia, nutrient budget Abstract The introduction of the short-necked clam Tapes philippinarum into the Sacca di Goro has over a short period made this coastal environment one of the top European clam production sites. In recent years, this activity has been seriously impacted due to the appearance in the lagoon of large macroalgal beds and the occurrence of dystrophic events causing anoxia and massive deaths of molluscs in the cultivated areas. Tapes cultivation sites now cover more than one third of the lagoon surface at densities sometimes attaining 2000–2500 adult individuals m −2 ; such densities and the harvesting methods, based on sediment dredging, prob- ably have a strong impact on the benthic system. Whilst a number of studies have reported water–sediment interface induced modifications due to oyster or mussel farming there have been few attempts to quantify how clam farming affects biogeochemical cycles of oxygen and nutrients, in particular in the Sacca di Goro. Two areas, a farmed and a control one, were compared for benthic fluxes and results were correlated with clam biomass. Oxygen, carbon dioxide, ammonium, reactive silica and phosphorus fluxes were stimulated several fold by the presence of Tapes due to the clams, respiration and excretion activities, but also to the reducing conditions in the surface sediments. On average, the whole lagoon dark sediment O 2 demand and CO 2 production were stimulated by a factor of, respectively, 1.8 and 3.3, whilst nutrient release was 6.5 times higher for NH 4 + and 4.6 times higher for PO 4 3− . Our results indicate that clam farmers should carefully consider sustainable densities of Tapes in order to prevent the risk of sediment and water anoxia. Rapid nutrient recycling (up to 4000 µmol NH 4 + m −2 h −1 and 150 µmol PO 4 3− m −2 h −1 ) stimulated by the high biodegradability of clam faeces and pseudofaeces could in turn favour macroalgal growth. Introduction In coastal lagoons, the water column is very shallow and varies from 0 (in the macrotidal environments where the sediments are periodically exposed to the atmosphere) to few meters depth; the chemical com- position of the water and the oxygen availability is strictly linked to surficial sediment features and meta- bolism. At the water–sediment interface, aerobic and anaerobic mineralization processes deplete oxygen, produce reduced compounds and regenerate nutrients (Jørgensen, 1983) . Important transformations such nitrification and denitrification, mediated by special- ised bacteria, or the formation of complexes and the precipitation of orthophosphate also occur at this level (Henriksen & Kemp, 1988; Golterman, 1995). The continuous sedimentation of particulate mat- ter, produced in the water column or transported by the rivers, makes the water–sediment interface a par- ticularly dynamic zone. In particular, in coastal areas, settling particulate matter represents an inexhaustible source of organic carbon for a diversified community of filter feeders and decomposers (Graf et al., 1982; Albertelli et al., 1999). The intensification of molluscs
-
Upload
marco-bartoli -
Category
Documents
-
view
219 -
download
0
Transcript of Impact of Tapes philippinarum farming on nutrient dynamics and benthic respiration in the Sacca di...

Hydrobiologia 455: 203–212, 2001.© 2001 Kluwer Academic Publishers. Printed in the Netherlands.
203
Impact of Tapes philippinarum farming on nutrient dynamicsand benthic respiration in the Sacca di Goro
Marco Bartoli1,∗, Daniele Nizzoli1, Pierluigi Viaroli1, Edoardo Turolla2,Giuseppe Castaldelli2, Elisa Anna Fano2 & Remigio Rossi21Department of Environmental Sciences, Parma University, viale delle Scienze, 43100 Parma, Italy2Department of Evolutive Biology, Ferrara University, via Borsari, 44100 Ferrara, ItalyTel: 0521-905976, Fax: 0521-905402; E-mail: [email protected]
Received 9 June 2000; in revised form 9 April 2001; accepted 20 April 2001
Key words: clam culture, surficial sediments, benthic fluxes, anoxia, nutrient budget
Abstract
The introduction of the short-necked clam Tapes philippinarum into the Sacca di Goro has over a short periodmade this coastal environment one of the top European clam production sites. In recent years, this activity has beenseriously impacted due to the appearance in the lagoon of large macroalgal beds and the occurrence of dystrophicevents causing anoxia and massive deaths of molluscs in the cultivated areas.
Tapes cultivation sites now cover more than one third of the lagoon surface at densities sometimes attaining2000–2500 adult individuals m−2; such densities and the harvesting methods, based on sediment dredging, prob-ably have a strong impact on the benthic system. Whilst a number of studies have reported water–sediment interfaceinduced modifications due to oyster or mussel farming there have been few attempts to quantify how clam farmingaffects biogeochemical cycles of oxygen and nutrients, in particular in the Sacca di Goro. Two areas, a farmed anda control one, were compared for benthic fluxes and results were correlated with clam biomass. Oxygen, carbondioxide, ammonium, reactive silica and phosphorus fluxes were stimulated several fold by the presence of Tapesdue to the clams, respiration and excretion activities, but also to the reducing conditions in the surface sediments.On average, the whole lagoon dark sediment O2 demand and CO2 production were stimulated by a factor of,respectively, 1.8 and 3.3, whilst nutrient release was 6.5 times higher for NH4
+ and 4.6 times higher for PO43−.
Our results indicate that clam farmers should carefully consider sustainable densities of Tapes in order to preventthe risk of sediment and water anoxia. Rapid nutrient recycling (up to 4000 µmol NH4
+ m−2 h−1 and 150 µmolPO4
3− m−2 h−1) stimulated by the high biodegradability of clam faeces and pseudofaeces could in turn favourmacroalgal growth.
Introduction
In coastal lagoons, the water column is very shallowand varies from 0 (in the macrotidal environmentswhere the sediments are periodically exposed to theatmosphere) to few meters depth; the chemical com-position of the water and the oxygen availability isstrictly linked to surficial sediment features and meta-bolism. At the water–sediment interface, aerobic andanaerobic mineralization processes deplete oxygen,produce reduced compounds and regenerate nutrients(Jørgensen, 1983) . Important transformations such
nitrification and denitrification, mediated by special-ised bacteria, or the formation of complexes and theprecipitation of orthophosphate also occur at this level(Henriksen & Kemp, 1988; Golterman, 1995).
The continuous sedimentation of particulate mat-ter, produced in the water column or transported bythe rivers, makes the water–sediment interface a par-ticularly dynamic zone. In particular, in coastal areas,settling particulate matter represents an inexhaustiblesource of organic carbon for a diversified communityof filter feeders and decomposers (Graf et al., 1982;Albertelli et al., 1999). The intensification of molluscs

204
sowing and collection practises is a disturbance factorfor the environment, and in particular for the benthicsystem where disequilibrium side-effects, due to theincreasing dominance of the cultivated species mainlyoccur. Another important side-effect is represented bythe higher input of organic matter (faecal pellets orpseudo faeces) at the sediment surface (Kaspar et al.,1985; Hatcher et al., 1994; Kaiser et al., 1998).
Molluscs such as oysters or mussels are generallyfarmed in boxes suspended in the water column orattached to supports. The study of their impact onthe environment due to increased organic matter de-position on sediments is relatively easy (Boucher &Boucher-Rodoni, 1988; Cheng et al., 1993; Hatcheret al., 1994; Kemp et al., 1997). In contrast, clamsare farmed directly within the sediments and theirimpact is for this reason more difficult to determine.High densities of clams affect the environment due totheir respiration activity, the excretion of nitrogenouscompounds and the enrichment of the sediment withorganic matter (Doering et al., 1987; Zhu et al., 1999).Clam culture enhances suspended matter sedimenta-tion and concentrates faeces and pseudofaeces in thefarmed areas. This results in an accumulation of or-ganic matter which stimulates sulphate reduction andleads to the accumulation of sulphide in the surfacesediments (Sorokin et al., 1999). Moreover, the dom-inance of clams within the benthic community resultsin the displacement of other organisms and the loss oftheir associated functions, for example sediment mix-ing or oxygen transport into the anoxic sediment ho-rizons. Finally, clam harvesting techniques, based onthe dredging of the surficial sediments, cause altera-tions in the natural sediment stratification and improvenutrient and reduced compounds mobility (Kaiser etal., 1998).
The introduction and the large-scale farming ofthe short-necked clam Tapes philippinarum (Adams& Reeve, 1850) represents the most relevant socio-economic event for Italian shellfish production. Theappearance of this bivalve in the Adriatic Sea is rel-atively recent and is a consequence of local farmingtrials initiated in Venice lagoon in 1983 (Breber, 1985;Cesari & Pellizzato, 1985). Further introductions andthe impressive adaptive capacity of this species haveresulted in a wide and rapid expansion into other envir-onments, firstly in the Po River Delta and then alongthe Adriatic coast (Paesanti, 1990; Rinaldi, 1991) andin different coastal Italian areas (Cottiglia & MasalaTagliasacchi, 1988; Zentilin, 1990). Italy is now thelargest producer of clams in the European Community
with an estimated crop comprising between 50 and60 000 ton yr−1.
The economic relevance of the species has drawnthe attention of the scientific community which isinterested in fish and acquaculture problems. Manystudies regarding reproduction strategies and techno-logies, as well as cultivation and harvesting techniquescan be found in the literature. In contrast, the ef-fect of the species on the farmed environment hasreceived little attention. The bibliography regardingmollusc farming describes mostly intuitive consider-ations about the relationships between mollusc cultureand eutrophication processes: the collection and re-moval from the system of organisms that feed onmicroalgae and particulate matter should result in anet reduction of nutrient loads (Cloern, 1982; Officeret al., 1982; Kaspar et al., 1985). This consideration isbased on the assumption that, contrary to fish farmingsystems in which food (and so N and P) are continu-ously supplied from outside, molluscs find their foodrequirements in situ.
The net removal of nutrients from the system asmollusc crop is easily quantifiable while this is not thecase for nutrient regeneration/mobilisation rates due tomollusc metabolic activity and sediment dredging.
Objectives
The aim of this work was to evaluate the impact ofTapes philippinarum farming on sediment features andbenthic metabolism in the Sacca di Goro. Oxygen,carbon dioxide and nutrient fluxes, sedimentary or-ganic matter and total N and P concentrations wereanalysed in a control and in a cultivated site. Relation-ships between fluxes and clam biomasses were alsoconsidered.
Surficial sediments were resuspended in order tosimulate sediment dredging during clams harvestingand oxygen consumption due to both chemical andbiological processes was estimated. Preliminary N andP mass balances were calculated in order to eval-uate the impact of clams on the whole ecosystemmetabolism, since the exploited area covers approx-imately 35–40% of the lagoon surface. The potentialeffects of clam farming with respect to eutrophicationand its ultimate manifestation, dystrophy, was alsoconsidered.

205
Figure 1. Map of the Sacca di Goro: the areas exploited for molluscs farming are shaded gray. C and F indicate, respectively, the control andthe farmed sites.
Study area
The Sacca di Goro (44◦ 82′ N, 12◦ 27′ E) is a shal-low coastal lagoon located in the southernmost part ofthe Po River Delta (Northern Italy); the total surfacearea is about 26 km2 and the average depth is 1.5 m(Fig. 1). The bottom sediments are a mosaic of sandy,silty and clayey zones; the distribution of these areasin the lagoon depends on water currents due in turnto tidal movements and the input of freshwater fromthe Po di Volano and the Po di Goro. Sandy areas arelocated close to sea mouths, whilst muddy sedimentsare common in the eastern corner were the waters arestagnant. At present more than one third of the lagoonsurface (about 10 km2) is cultivated with Tapes phil-ippinarum; local fishermen co-operatives manage thesowing and the harvesting of the clams in well definedlicensed areas called ‘concessioni’.
Young clams (5–10 mm in shell length) are con-tinuously collected within or in the sand banks justoutside the lagoon and are immediately sown in thelicensed areas until they reach commercial size; inthe Sacca di Goro, due to high food availability, thisoccurs within a few months.
Within the ‘concessioni’, according to the fish-ermen, Tapes densities are generally maintained ataround 1000 adult individuals m−2; the shell lengthof an adult is around 4 cm, its weight about 10 g.
Licensed areas for clam farming are mainly locatedin the southern area of the lagoon (Fig. 1) where thewater exchange rate is higher and the sediments aremostly sandy.
The first introduction of Tapes philippinarum intothe Sacca di Goro was in 1986 (Carrieri et al., 1992)and the first crop was harvested in the following year.In this lagoon, the cultivation of Tapes was so success-ful that in a few years the number of people involvedreached 1200 with a production of clams in 1991 ofmore than 9000 tons (Rossi & Paesanti, 1992) andin the mid 1990s around 15 000 tons (but 20–25 000tons are probably more realistic). In recent years,clam farming in the Sacca di Goro has suffered ser-ious set backs due to massive clam mortality in thesummer. The appearance in the lagoon of the mac-roalgae Ulva rigida and the occurrence of dystrophicevents triggered by macroalgae decomposition pro-cesses have been proposed as one of the main factorsdetermining the decline of clam culture. Dystrophicevents occur generally in the summer when huge matsof Ulva (up to 800 g dry weight m−2) suddenly col-lapse; decomposition processes turn the water columnanoxic and allow sulphide diffusion from the sedi-ments with catastrophic effects for the benthic fauna(Viaroli et al., 1996).

206
Materials and methods
Core sampling and maintenance
Sediments with and without clams were sampled on23 June 1999 within and just outside a licensed areaclose to the sand bank which separates the lagoon fromthe open sea (Fig. 1). According to the fishermen, thesediments in the chosen licensed area were not dis-turbed by recent harvesting activities and the clamswere close to commercial size; we sampled in summerto have high metabolic rates and because this is theperiod in which dystrophic events occur.
In each area, 5 sediment cores were hand collec-ted using transparent plexiglass tubes (height 40 cm;inner diameter 20 cm), the sediment depth within thecores was adjusted to 15 cm. The collected cores weretransported within 1 hour from the sampling site to theGoro hatchery (CRIM: Center for Molluscs Reproduc-tion) and submersed in a 300 l tank containing in situwater. The water inside each core was continuouslymixed by a small aquarium pump and the water in thetank continuously renewed with lagoon water at a rateof 100 l h−1 in order to maintain a constant supply ofmicroalgae and particulate matter for the clams and theother components of the benthic community. The tankwas placed outside under natural light and temperatureconditions; the cores were left to stabilise in the tankfor about 30 h before the beginning of the experiments.
Fluxes determination
Oxygen, CO2, DIN (dissolved inorganic nitrogen:NO3
−+NO2−+NH4
+), urea, reactive phosphorus andsilica and chlorophyll ‘a’ fluxes were determined.Only dark incubations were performed since our in-terest was mainly in the comparison of sedimentrespiration and nutrient regeneration rates. The in-cubation started when floating plexiglass lids werepositioned above the cores; initial samples for oxygen,CO2, nutrient and chlorophyll determinations werecollected in quadruplicate from the tank. After 1 h,water samples were collected by a syringe from eachcore through a small rubber septum in the lid. Thechlorophyll concentration at the end of the incubationwas determined filtering 1 l of water from each coreafter removal of the lid.
Fluxes across the sediment-water interface for O2,CO2, nutrients and Chl ‘a’ were calculated by con-centration difference before and after the incubation
according to equation 1:
f = (Cf − Ci) × V
t × A, (1)
where Cf and Ci represent the final and initial concen-tration, respectively, of the considered compound, Vthe volume of the water in the core, t the incubationtime and A the surface area.
After incubation, sediment subsamples were col-lected in triplicate from each microcosm using 2 cmi.d. Plexiglas cores and analysed for organic mat-ter and total N and P in the 0–2 and 2–5 cm depthhorizons.
Approximately 1 g of fresh material from each ofthe 2 layers was resuspended in a modified 50 mlbeaker containing oxygen saturated in situ water. Amagnetic bar allowed the complete resuspension of thesediment inside the beaker whilst a floating lid preven-ted oxygen exchanges with the atmosphere. Oxygendepletion was followed over a 1 h period using a Clarkstyle microelectrode (Diamond General, mod. 737-GC) inserted through the cap; chemical and biologicalO2 demand was then calculated.
The remaining sediment in each core was sievedthrough a 500 µm net in order to determine clamand other macrobenthic species densities. The collec-ted clams were measured for shell length, width andheight and for the determination of mollusc flesh dryweight. The shell and the dried flesh of the clamswere finally powdered and analysed for total P and Ncontent.
Methods for water, sediment and clam analysis
Dissolved oxygen was determined by the Winklermethod (APHA, 1975) and via polarographic sensors;total carbonates for CO2 fluxes by titration with 0.1 MHCl (Anderson et al., 1986). Ammonium was de-termined using the indophenol blue method accordingto Bower & Holm Hansen (1986). Nitrate was de-termined via diazotation after cadmium reduction tonitrite (APHA, 1975). Soluble reactive phosphorusand dissolved reactive silica were determined accord-ing to Valderrama (1977) and APHA (1975). Urea wasdetermined according to the method proposed by Mul-venna et al. (1992). Chlorophyll ‘a’ was determinedspectrophotometrically after extraction with 90% v/vacetone from GF/C Whatman filters (APHA, 1975).
Total phosphorus in the sediment and in the shelland flesh of clams was determined after incinerationand extraction of the ashes with concentrated HCl(Aspila et al., 1976).

207
Total nitrogen in the sediment and in the clamflesh was determined with the Kjeldhal method afterdigestion at 350 ◦C with H2SO4 and H2O2.
Sediment organic matter content was determinedafter 2 h muffle incineration at 550 ◦C.
Statistical analysis
The differences between sediment quality and fluxesin cultivated and control areas and the clam biomass-dependent changes of benthic exchanges were testedwith a one-way ANOVA.
Results
Macrofauna colonisation within and outside thelicensed area
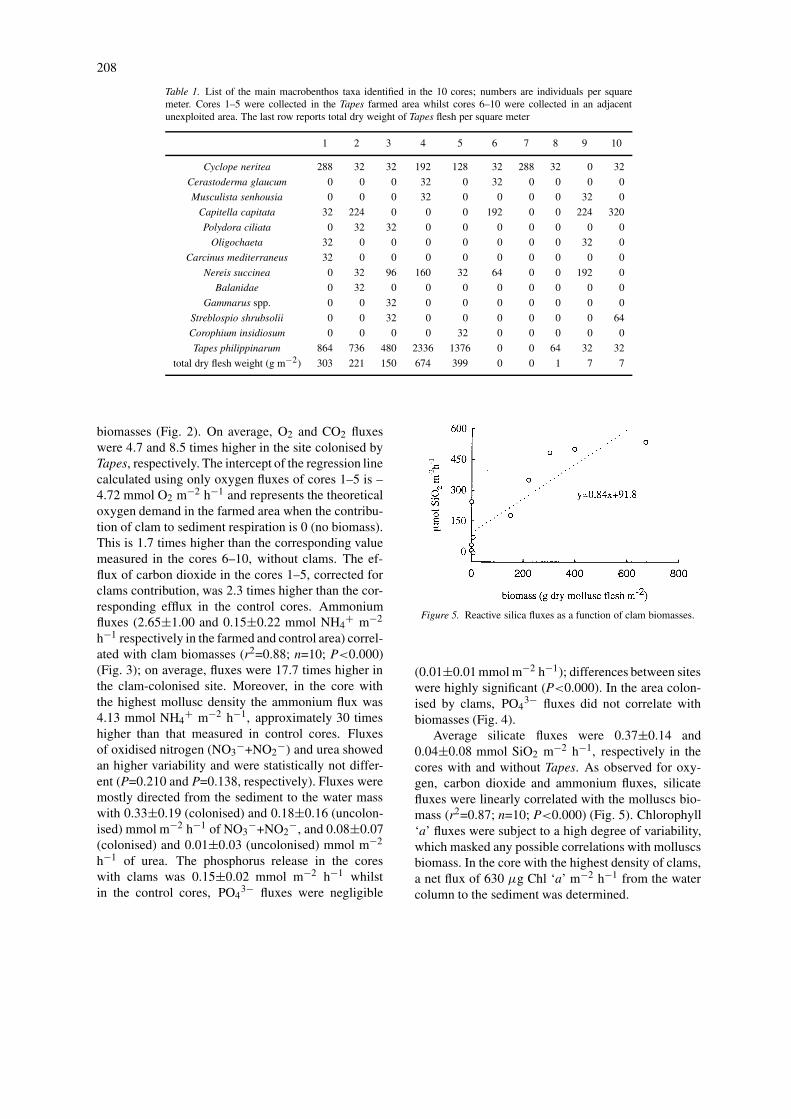
Densities of the main macrofauna taxa are reported atTable 1. Sediment collected outside the licensed area(cores 6–10) contained practically no clams, whilstextremely high densities occurred within the cultiv-ated areas, comprising from 480 to 2336 ind. m−2.Such variability probably depends on the sowing pro-cedure (scatter seeding) and on the clams tendency toassume contagious distributions. The highest densityof Tapes corresponded to about 674 g of flesh whichis equivalent to 15 kg of molluscs per square meter.Contrary to what would be expected the densities ofother macrobenthic organisms were generally low inboth sets of cores (Table 1). In the cores 1–5, theclams were close to the commercial size, with a shelllength of 3.0±0.7 cm and mollusc flesh dry weight of0.32±0.02 g ind−1. Within the licensed area the clamsize was relatively homogeneous suggesting that themolluscs were the progeny of a single seeding event.
Organic matter and total N and P in the surfacesediment
Along the surficial sediment horizons (0–2 and 2–5 cmdeep), organic matter, total P and N were constant anddifferences between colonised and uncolonised siteswere not statistically significant (Table 2).
O2, CO2 and nutrient fluxes
In the clam, colonised sediments the dark oxygenconsumption was –12.49±4.54 mmol O2 m−2 h−1,whereas in the uncolonised sediments the averageoxygen flux (–2.67±0.58 mmol O2 m−2 h−1) was
Figure 2. Benthic fluxes of oxygen and carbon dioxide as a func-tion of clam biomass (dry weight of flesh) per square meter. Linearregressions were calculated using fluxes determined in the 10 cores.
Figure 3. Sediment-water fluxes of ammonium as a function ofclam biomass (dry weight of flesh per square meter). Linearregressions were calculated using fluxes determined in 10 cores.
significantly lower (P=0.001). Average CO2 pro-duction rates, (10.42±5.61 and 1.22±0.81 mmolCO2 m−2 h−1, respectively in the cores with andwithout Tapes) were also different (P=0.003). Oxy-gen (r2=0.98; n=10; P<0.000) and carbon dioxide(r2=0.84; n=10; P<0.000) fluxes correlated with clam
Figure 4. Reactive phosphorus fluxes as a function of clam biomass.
208
Table 1. List of the main macrobenthos taxa identified in the 10 cores; numbers are individuals per squaremeter. Cores 1–5 were collected in the Tapes farmed area whilst cores 6–10 were collected in an adjacentunexploited area. The last row reports total dry weight of Tapes flesh per square meter
1 2 3 4 5 6 7 8 9 10
Cyclope neritea 288 32 32 192 128 32 288 32 0 32
Cerastoderma glaucum 0 0 0 32 0 32 0 0 0 0
Musculista senhousia 0 0 0 32 0 0 0 0 32 0
Capitella capitata 32 224 0 0 0 192 0 0 224 320
Polydora ciliata 0 32 32 0 0 0 0 0 0 0
Oligochaeta 32 0 0 0 0 0 0 0 32 0
Carcinus mediterraneus 32 0 0 0 0 0 0 0 0 0
Nereis succinea 0 32 96 160 32 64 0 0 192 0
Balanidae 0 32 0 0 0 0 0 0 0 0
Gammarus spp. 0 0 32 0 0 0 0 0 0 0
Streblospio shrubsolii 0 0 32 0 0 0 0 0 0 64
Corophium insidiosum 0 0 0 0 32 0 0 0 0 0
Tapes philippinarum 864 736 480 2336 1376 0 0 64 32 32
total dry flesh weight (g m−2) 303 221 150 674 399 0 0 1 7 7
biomasses (Fig. 2). On average, O2 and CO2 fluxeswere 4.7 and 8.5 times higher in the site colonised byTapes, respectively. The intercept of the regression linecalculated using only oxygen fluxes of cores 1–5 is –4.72 mmol O2 m−2 h−1 and represents the theoreticaloxygen demand in the farmed area when the contribu-tion of clam to sediment respiration is 0 (no biomass).This is 1.7 times higher than the corresponding valuemeasured in the cores 6–10, without clams. The ef-flux of carbon dioxide in the cores 1–5, corrected forclams contribution, was 2.3 times higher than the cor-responding efflux in the control cores. Ammoniumfluxes (2.65±1.00 and 0.15±0.22 mmol NH4
+ m−2
h−1 respectively in the farmed and control area) correl-ated with clam biomasses (r2=0.88; n=10; P<0.000)(Fig. 3); on average, fluxes were 17.7 times higher inthe clam-colonised site. Moreover, in the core withthe highest mollusc density the ammonium flux was4.13 mmol NH4
+ m−2 h−1, approximately 30 timeshigher than that measured in control cores. Fluxesof oxidised nitrogen (NO3
−+NO2−) and urea showed
an higher variability and were statistically not differ-ent (P=0.210 and P=0.138, respectively). Fluxes weremostly directed from the sediment to the water masswith 0.33±0.19 (colonised) and 0.18±0.16 (uncolon-ised) mmol m−2 h−1 of NO3
−+NO2−, and 0.08±0.07
(colonised) and 0.01±0.03 (uncolonised) mmol m−2
h−1 of urea. The phosphorus release in the coreswith clams was 0.15±0.02 mmol m−2 h−1 whilstin the control cores, PO4
3− fluxes were negligible
Figure 5. Reactive silica fluxes as a function of clam biomasses.
(0.01±0.01 mmol m−2 h−1); differences between siteswere highly significant (P<0.000). In the area colon-ised by clams, PO4
3− fluxes did not correlate withbiomasses (Fig. 4).
Average silicate fluxes were 0.37±0.14 and0.04±0.08 mmol SiO2 m−2 h−1, respectively in thecores with and without Tapes. As observed for oxy-gen, carbon dioxide and ammonium fluxes, silicatefluxes were linearly correlated with the molluscs bio-mass (r2=0.87; n=10; P<0.000) (Fig. 5). Chlorophyll‘a’ fluxes were subject to a high degree of variability,which masked any possible correlations with molluscsbiomass. In the core with the highest density of clams,a net flux of 630 µg Chl ‘a’ m−2 h−1 from the watercolumn to the sediment was determined.

209
Sediment resuspension
Resuspension of surficial sediment resulted in a rapidoxygen depletion in the overlying water. Results fromtwo sediment layers (0–2 and 2–5 cm depth) werepooled and gave potential oxygen consumption of56±12 and 27±9 mmol O2 m−2, respectively, in thefarmed and control areas. In both areas the resuspen-sion of the deeper layer determined a higher oxygendemand. Most of the oxygen was consumed withina few minutes, indicating a high chemical oxygendemand for reoxidation of reduced compounds.
Discussion
The farming of the clam Tapes philippinarum in theSacca di Goro strongly stimulated oxygen and car-bon dioxide fluxes and nutrient regeneration acrossthe water-sediment interface. In the farmed area, clamdensities were over 2000 adult individuals m−2, withvery low abundances of other macrobenthic species.Low biodiversity was found also in an adjacent un-exploited area. We explained these patterns as a con-sequence of both seasonal trends and farming impact.Macrobenthos density in the lagoon is generally lowin the late spring due to reducing conditions in thesurface sediment, as already reported by Fano et al.(1996). Lagoon areas close to the cultivated sites arestrongly impacted by the dense clam banks whichremove suspended particles (Danovaro & Fabiano,1997) and thus potential food availability is low forsediment-associated detritivores. Furthermore, clamsproduce large quantities of faeces and pseudofaeceswhich represent an ideal substrate for bacteria andshift the sediments towards a rapid consumption ofoxygen (Sorokin et al., 1999). In the areas close tothe farmed sites, increased densities of the infaunalCapitella capitata have been usually observed. Thisopportunistic polychaete is generally a good indicatorof organic matter enrichment of the sediment (Weston,1990).
Most of the sediment oxygen demand estimatedin the farming area of the Sacca di Goro was due tothe respiration of the clams. Measured values, of upto 20 mmol O2 m−2 h−1 are indicative of signific-ant amounts of oxygen driven from the water to thebenthic system. In eutrophic coastal lagoons, oxygenrespiration although usually high never exceeds 10–15 mmol m−2 h−1. Summer maximum are usuallyattained in systems which are dominated by macro-phytes (Murray & Wetzel, 1987; D’Avanzo et al.,
1996, Viaroli et al., 1996; Bartoli et al., 2001). If adrifting Ulva bed, common in the spring in the Saccadi Goro with biomasses (as dry weight) of 300/400g m−2, becomes stranded on the sediment of a farmedarea, dark benthic respiration up to 30–40 mmolO2 m−2 h−1 would be expected. Such oxygen demandcould turn a water column of 50 cm completely an-oxic in a few hours, under conditions of low waterexchange and light winds. Occasional Tapes mortal-ities in the Sacca di Goro could be simply explainedby the establishment of anoxia due to the metabolismof the clams and, probably, by a series of events suchas the saturation of iron pools in the sediment andthe consequent diffusion of toxic free sulphide to thewater-sediment interface. Samplings of Ulva withinthe licensed areas revealed that in summer the clamswere growing attached to the macroalgae thalli, out-side the sediment. Ulva mats are likely to change themicrocirculation of the water overlying the bottom andwhere Ulva develops and settles clams are probablyforced out of the sediments in order to respire and feed.
Carbon dioxide, ammonium, reactive phosphorusand silica fluxes were all strongly stimulated by thepresence of the clams. This is due to the reducingconditions that establish at the surface sediment (inparticular for phosphorus fluxes), to the clams excre-tion (Zhu et al., 1999) and to the rapid degradationof the organic matter (faecal pellets, faeces or pseudofaeces) produced by the clams and settling at thesediment-water interface. Average DIN fluxes in thecores with clams were 2.98±1.12 mmol m−2 h−1 andapproximately 10-fold higher than those measured inthe cores without clams (0.32±0.28 mmol m−2h−1).Ammonium was the dominant component of inorganicnitrogen released to the water column and represen-ted about 90% of the total in the cores in which themolluscs were present while in the controls it accoun-ted for only 30% of the DIN flux. Ammonium fluxesabove 2 mmol NH4
+ m−2 h−1 have never been re-ported in previous studies in the Sacca di Goro, evenin summer; rates of about 0.6–0.8 mmol NH4
+ m−2
h−1 were measured in July 1994 in the muddy Gorinoarea following the collapse of macroalgal mats (Bar-toli, 1996). Silica regeneration was on average 9.2times faster in the site farmed with clams and thisis probably due to clams directly feeding on diatomsand to the production of easily degradable faeces andpseudo faeces. In contrast, although higher in farmedareas, phosphate effluxes were not correlated with theclam biomasses and were apparently constant at bio-masses greater than 100 g m−2. Likewise, PO4
3−

210
effluxes were partially buffered by sedimentary pro-cesses. Nevertheless, a high amount of remineralisedPO4
3−- was released to the water column. Phosphorusfluxes were measured before in the Sacca di Goroby Giordani (1997) and Azzoni (1999) along a gridof uncultivated sites and never exceeded 20–30 µmolPO4
3−m−2 h−1.Filtration rates by Tapes philippinarum are mod-
ulated by the molluscs feeding and respiration re-quirements; in the Sacca di Goro, the high molluscsdensities in the farmed areas probably force the mol-luscs towards the filtration of large amounts of waterjust to satisfy metabolic oxygen demand. Many of thefiltered particles are wrapped with mucus by the clamand this happens whether the particle is ingested orexpelled. The need for large volumes of water for res-piration results in a the production of large quantitiesof pseudofaeces which represent an easily degradablematerial deposited on the sediment surface. This ex-plains the extreme reactivity of this sediment horizonin the farmed area and the rapid regeneration ratesfor C, N, P and Si coupled to high oxygen consump-tion. Similar results have been obtained for oyster beds(Boucher & Boucher-Rodoni, 1998; Chapelle et al.,2000), mussel (Kaspar et al., 1985; Baudinet et al.,1990; Hatcher et al., 1994) and clams (Doering et al.,1987).
The rapid circulation of nutrients can promote newphytoplankon blooms which is a positive feedbackfor clams but can also sustain macroalgal growth, inparticular in the warm season when external nutrientloads are low, whereas the consequences of dystrophicevents associated with macroalgal bloom collapses area negative feedback for clam culture (Naldi, 1994;Viaroli et al., 1996).
We tried to compare how oxygen, carbon dioxide,silica, ammonium and reactive phosphorus fluxes arestimulated by the clams biomass; results are shownin Fig. 6. Reported values are ratios between singlemeasurements in the cores with clams and the aver-age fluxes in the control cores. For example, in thecore with the highest biomass of Tapes ammoniumfluxes were 20 times higher than the correspondingrate measured in the controls, while the oxygen fluxwas 7 fold higher. The presence of high densities ofclams stimulated ammonium fluxes to a greater degreethan carbon dioxide, reactive silica, phosphorus andoxygen ones due probably to direct NH4
+ excretion(Zhu et al., 1999). The higher relative increase of CO2production compared to oxygen demand indicates ashift towards anaerobic mineralisation processes like,
Figure 6. Factor of increase of oxygen, carbon dioxide and nutri-ent fluxes in relation to mollusc biomass. Each number is the ratiobetween single fluxes measured in the cores 1–5 (with clams) andthe average flux measured in the control cores.
Table 2. Organic matter and total N and P content of theclam-colonised and control sediments; average values withstandard deviations are reported
Layer (cm) Farmed Control
Organic matter 0–2 2.8±0.3 2.5±0.2
(%) 23–5 2.5±0.3 2.3±0.2
Total P 0–2 19.5±1.1 17.4± 1.3
(µmol g−1dw) 2–5 19.1±1.5 17.4±2.1
Total N 0–2 67.5±9.5 68.5±2.3
(µmol g−1dw) 2–5 79.1±16.3 73.3±2.6
for example, iron and sulphate reduction, which couldalso explain the release of phosphorus (Hatcher et al.,1994; Giordani, 1996; Azzoni, 1999).
Simulated sediment dredging indicated that cultiv-ated sediments are probably more reduced comparedto control areas and strengthened the hypothesis re-garding the increasing importance of anaerobic pro-cesses for organic matter mineralisation. Furthermore,clam harvesting activities are disruptive for the sur-ficial sediment and result in sudden and localiseddeficits of oxygen in the water column which couldaffect the benthic fauna.
Table 3 reports some major characteristics of theshell and the flesh of 120 individuals of Tapes col-lected from the cores incubated in these experiments.Standard deviations, not reported, were less then 5%of the averages. The shells and the flesh of Tapeshave been analysed for total P and N content in or-der to quantify the amount of those nutrients that are

211
Table 3. Average composition of 120 in-dividuals of Tapes philippinarum collectedfrom the cores 1–5
Tapes philippinarum composition %
Shell, ashes 59.0
Shell, loss after ignition 1.6
Shell, water 0.6
Flesh, ashes 0.6
Flesh, organic matter 3.5
Flesh, water 34.7
removed from the lagoon during fishing activity; cal-culations were corrected for a clams crop of 15 000tons which is a peak of the Sacca di Goro productionand resulted in about 46 tons of N and 10 tons of Premoved.
It is interesting to compare these numbers withannual N and P loads to the lagoon from freshwater in-puts and then with the N and P amounts released fromthe sediment to the water column. N and P loads en-tering the lagoon have been estimated by in 1997 to be1300 and 40 tons, respectively (Viaroli, unpublisheddata); clams harvesting accounts for approximately 5and 25% of external inputs having a relatively highereffect on phosphorus loading. The presence of clams,however, stimulates inorganic N and P regenerationfrom the sediment to the water column. Fluxes determ-ined in the cores collected in the farmed area have beenintegrated for a lagoon surface of 10 km2, a period of 4months (mid May–mid September) in which the watertemperature is generally above 20 ◦C and the intensityof these fluxes can be realistic and a dark period of10 hours per day. Results indicate a regeneration fromthe sedimentary loads of 504 tons of ammonium andof 56 tons of reactive phosphorus. Thus, the amountof N and P potentially removed in the Tapes crop is asmall fraction compared to that regenerated from thesediment to the water column in cultivated areas.
Many authors reporting the effects of molluscscultivation on benthic fluxes have found that regenera-tion processes were stimulated several fold (Doeringet al., 1987; Baudinet et al., 1990; Hatcher et al.,1994; Chapelle et al., 2000) but the global effects werenegligible due to the relatively small farmed surfacecompared to the total lagoon area (Baudinet et al.,1990).
In the Sacca di Goro, the situation is different sinceclams are now farmed over the 35–40% of the total
lagoon surface and, at least in the summer, increasedbenthic mineralisation could affect the global nutrientcirculation in the lagoon. Taking into account the ratioof cultivated – not cultivated areas, we have estim-ated that, in the period of the present investigation,the clams farming potentially increased the sedimentoxygen demand by a factor of 1.8 and CO2 production3.3 times. Nutrient release was 6.5 times higher forammonium and 4.6 times higher for phosphorus.
In conclusion the hypothesis that clam farming actsas a control for eutrophication processes seems notrealistic for the Sacca di Goro; on the contrary, furtherinvestigations are needed to clarify possible feedbacksbetween Tapes farming, internal nutrient loading andmacroalgal growth.
References
Albertelli, G., A. Covazzi-Harrigue, R. Danovaro, M. Fabiano, S.Fraschetti & A. Pusceddu, 1999. Differential responses of bac-teria, meiofauna and macrofauna in a shelf area (Ligurian Sea,NW Mediterranean): role of food availability. J Sea Res. 42:11–26.
Anderson, L. G., P. O. J. Hall, A. Iverfeldt, M. M. R. Van Der Loeff,B. Sundby & S. F. G. Westerlund, 1986. Benthic respirationmeasured by total carbonate production. Limnol. Oceanogr. 31:319–329.
A.P.H.A., 1975. Standard Methods for the Examination of Waterand Wastewater. 14th edn. A.P.H.A., Washington, U.S.A.: 1193pp.
Aspila, K. J., H. Agemian & A. S. Y. Chau, 1976. A semiauto-mated method for the determination of inorganic, organic andtotal phosphate in sediments. Analyst 101: 187–197.
Azzoni, R., 1999. Sulphur, iron and phosphorus regulation in coastallagoon ecosystems with different primary producers. PhD thesis:113 pp.
Bartoli, M., 1996. Benthic fluxes of nitrogen and phosphorus incoastal lagoons with different primer producers. PhD thesis,Department of Environmental Sciences, Parma University: 136pp.
Bartoli, M., G. Castaldelli, D. Nizzoli, L. G. Gatti & P. Viaroli,2001. Benthic fluxes of oxygen, ammonium and nitrate andcoupled and uncoupled denitrification rates in three eutrophiccoastal lagoons with different primary producers. In Faranda, M.,L. Guglielmo & G. Spezie (eds), Structures and Processes in theMediterranean Ecosystems. Springer Verlag Italia, Milano 29:227–235.
Baudinet, D., E. Alliot, B. Berland, C. Grenz, M. Plante-Cuny,R. Plante & C. Salen-Picard, 1990. Incidence of mussel cul-ture on biogeochemical fluxes at the sediment-water interface.Hydrobiologia 207: 187–196.
Boucher, G. & R. Boucher-Rodoni, 1988. In situ measurement ofrespiratory metabolism and nitrogen fluxes at the interface ofoyster beds. Mar. Ecol. Prog. Ser. 44: 229–238.
Bower, C. E. & T. Holm-Hansen, 1980. A salicylate-hypochloritemethod for determining ammonia in seawater. Can. J. Fish aquat.Sci. 37: 794–798.

212
Breber, P., 1985. L’introduzione e l’allevamento in Italiadell’Arsella del Pacifico, Tapes semidecussatus Reeve (Bivalvia;Veneridae). Oebalia 9: 675–680.
Carrieri, A., F. Paesanti & R. Rossi, 1992. Risultatidell’introduzione di vongola filippina Tapes Philippinarum(Adams & Reeve, 1850) nella Sacca di Goro (Delta del Po).Oebalia 17: 97–104.
Castaldelli, G., 1997. Nitrification and ammonium pools in the sedi-ments of the Sacca di Goro. PhD thesis, Department of evolutiveBiology, Ferrara University: 102 pp.
Cesari, P. & M. Pellizzato, 1985. Molluschi pervenuti in laguna diVenezia per apporti volontari o casuali. Acclimazione di Sac-costrea commercialis (Iredale & Roghely, 1933 ) e di Tapesphilippinarum (Adams & Reeve, 1850). Boll. Malacologico 21:237–274.
Chapelle, A., A. Menesguen, J. Paoli, P. Souchu, N. Mazouni,A. Vaquer & B. Millet, 2000. Modelling nitrogen, primaryproduction and oxygen in a Mediterranean lagoon. Impact ofoysters farming and inputs from the watershed. Ecol. Model.127: 161–181.
Cheng, J. I., S. L. Jeffrey, M. McCartney, D. Martinez & M. J.Weissburg, 1993. A bioassay approach to sesonal variation inthe nutritional value of sediment.
Cloern, J. E., 1982. Does the benthos control phytoplankton biomassin south San Francisco Bay? Mar. Ecol. Prog. Ser. 9: 191–202.
Cottiglia, M. & M. L. Masala Tagliasacchi, 1988. Esperienze diallevamento di Tapes philippinarum in Sardegna. Quaderni Ist.Idrobiol. Acquacolt. Brunelli: 3–26.
Danovaro, R. & M. Fabiano, 1997. Seasonal changes in qualityand quantity of food available for benthic suspension-feeders inthe Golfo Marconi (north-western Mediterranean). Estuar. coastshelf Sci. 44: 723–736.
D’Avanzo, V., J. N. Kremer & S. C. Wainright, 1996. Ecosys-tem production and respiration in response to eutrophicationin shallow temperate estuaries. Mar. Ecol. Prog.s Ser. 141:263–274.
Doering, P. H., J. R Kelly, C. A. Oviatt & T. Sowers, 1987. Effectof the hard clam Mercenaria mercenaria on benthic fluxes ofinorganic nutrients and gases. Mar. Biol. 94: 377–383.
Fano, E. A., L. G. Gatti, C. Tita, F. Schiffino & V. Gaiani, 1996.Benthic communities in lagoon sediments and macroalgae. Biol.mar. Medit. 3: 459–463.
Giordani, G., 1997. Sulphide speciation and inorganic reactivephosphorus mobility in relation to iron availability in dystrophiclagoons sediments. PhD thesis: 131 pp.
Golterman, H. L., 1995. Theoretical aspects of the adsorption oforthophosphate onto iron-hydrpxide. Hydrobiologia 315: 59–68.
Graf, G., W. Bengtsson, U. Diesner, R. Schulz & H. Theede, 1982,Benthic response to sedimentation of a spring phytoplanktonbloom: process and budget. Mar. Biol. 67: 21–27.
Hatcher, A., J. Grant & B. Schofield, 1994. Effects of suspendedmussel culture (Mytilus spp.) on sedimentation, benthic respir-ation and sediment dynamics in a coastal bay. Mar. Ecol. Prog.Ser. 115: 219–235.
Henriksen, K. & W. Kemp, 1988. Nitrification in estuarine andcoastal marine sediments. In Blackburn, T. H. & J. Sorensen(eds), Nitrogen Cycling in Coastal Marine Environments. JohnWiley and Sons: 201–249.
Jørgensen, B. B., 1983. Processes at the sediment-water interface.In Bolin & Cook (eds), The Major Biogeochemical Cycles andtheir Interactions. SCOPE 33. John Wiley & Sons, New York.
Kaiser, M. J., I. Laing, S. D. Utting & G. M. Burnell, 1998. En-vironmental impacts of bivalve mariculture. J. Shel. Res. 17:59–66.
Kaspar, H. F., P. A. Gillespie, I. C. Boyer & A. L. Mac Kenzie,1985. Effect of mussel aquaculture on the nitrogen cycle andbenthic communities in Knepuru Sound, Marlborough Sounds,New Zealand. Mar. Biol. 85: 127–136.
Kemp, W. M., E. M. Smith, M. Marvin-DiPasquale & W. RBoynton, 1997. Organic carbon balance and net ecosystem meta-bolism in Chesapeake Bay. Mar. Ecol. Prog. Ser. 150: 229–248.
Mulvenna, P. F. and G. Savidge, 1992. A modified manualmethod for the determination of urea in seawater using diacetyl-monoxime reagent. Estuar. coast shellf Sci. 34: 429–438.
Murray, L. & R. L. Wetzel, 1987. Oxygen production and con-sumption associated with the major autotrophic components intwo temperate seagrass communities. Mar. Ecol. Prog. Ser. 38:231–239.
Naldi, M., 1994. Benthic macroalgae and nutrient cycling in a Poriver delta lagoon (Sacca di Goro) PhD Thesis: 124 pp.
Officer C. B., T. J. Smayda & R. Mann 1982. Benthic filter feeding:a natural eutrophication control. Mar. Ecol. Prog. Ser. 9: 203–210.
Paesanti F., 1990. Programmi di sviluppo della pesca edell’acquacoltura nella Sacca di Goro. Hydrores 8: 56–57.
Rinaldi, E., 1991. Le conchiglie della costa romagnola. Ed. Essegi -Ravenna.
Rossi, R. & F. Paesanti, 1992. Successful clam farming in Italy.Proc. XXIII Annual Shellfish Conference, 19 & 20 May 1992.Fishmongers’ Hall, London: 62–68.
Sorokin, I. I., O. Giovanardi, F. Pranovi & P. I. Sorokin, 1999. Needfor restricting bivalve culture in the southern basin of the Lagoonof Venice. Hydrobiologia 400: 141–148.
Valderrama, J. C., 1977. Methods used by the Hydrographic De-partment of National Board of Fisheries, Sweden. In Grasshof,K. (ed.), Report of the Baltic Intercalibration Workshop. Annex,Interim Commission for the Protection of the Environment of theBaltic Sea: 13–40.
Viaroli, P., M. Bartoli, C. Bondavalli, R. R. Christian, G. Giordani& M. Naldi, 1996. Macrophytes communities and their im-pact on benthic fluxes of oxygen, sulphides and nutrients inshallow eutrophic environments. In Caumette, P., J. Castel &R. Herbert (eds), Coastal Lagoon Eutrophication and Anaer-obic Processes: C.L.E.A.N. Developments in Hydrobiology 117.Kluwer Academic Publishers, Dordrecht: 105–119. Reprintedfrom Hydrobiologia 329.
Weston, D. P., 1990. Quantitative examination of macrobenthiccommunity changes along an organic enrichment gradient. Mar.Ecol. Prog. Ser. 61: 233–244.
Zentilin A., 1990. Venericoltura in Laguna di Marano. In Tapesphilippinarum: Biologia e sperimentazione. E.S.A.V.: 201–205.
Zhu, S., B. Saucier, J. Durfey, S. Chen & B. Dewey, 1999. Wasteexcretion characteristics of Manila clams (Tapes philippinarum)under different temperature conditions. Aquac. Engin. 20: 231–244.
















![Functional plasticity in oyster gut microbiomes along a … · 2021. 1. 6. · [35]. Varying pollution levels alter Manila clam, Rudi-tapes philippinarum, microbiome composition and](https://static.fdocuments.in/doc/165x107/60f84ac8de01d43d2b2e56d4/functional-plasticity-in-oyster-gut-microbiomes-along-a-2021-1-6-35-varying.jpg)
