
Impact of Chestnut and Quebracho Tannins on Rumen...
12
Research Article Impact of Chestnut and Quebracho Tannins on Rumen Microbiota of Bovines Juan María Díaz Carrasco, 1,2 Claudio Cabral, 3 Leandro Martín Redondo, 1,2 Natalia Daniela Pin Viso, 2,4 Darío Colombatto, 2,5 Marisa Diana Farber, 2,4 and Mariano Enrique Fernández Miyakawa 1,2 1 Instituto de Patobiolog´ ıa, Centro Nacional de Investigaciones Agropecuarias, Instituto Nacional de Tecnolog´ ıa Agropecuaria, Calle Las Caba˜ nas y Los Reseros s/n, Casilla de Correo 25, Castelar, 1712 Buenos Aires, Argentina 2 Consejo Nacional de Investigaciones Cient´ ıficas y T´ ecnicas, Godoy Cruz 2290, 1425 Buenos Aires, Argentina 3 Animal Nutrition, Silvateam, Indunor, Cerrito 1136, 1010 Buenos Aires, Argentina 4 Instituto de Biotecnolog´ ıa, Centro Nacional de Investigaciones Agropecuarias, Instituto Nacional de Tecnolog´ ıa Agropecuaria, Calle Las Caba˜ nas y Los Reseros s/n, Casilla de Correo 25, Castelar, 1712 Buenos Aires, Argentina 5 Departamento de Producci´ on Animal, Facultad de Agronom´ ıa, Universidad de Buenos Aires, Av. San Mart´ ın 4453, 1417 Buenos Aires, Argentina Correspondence should be addressed to Mariano Enrique Fern´ andez Miyakawa; [email protected] Received 28 July 2017; Accepted 3 December 2017; Published 28 December 2017 Academic Editor: Yiannis Kourkoutas Copyright © 2017 Juan Mar´ ıa D´ ıaz Carrasco et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. e use of phytogenic dietary additives is being evaluated as a means to improve animal productivity. e effect of tannins seems to be the influence not only directly on the digestive process through binding of dietary proteins but also indirectly over their effects on gastrointestinal microbiota. High-throughput sequencing of 16S rRNA gene was used to analyze the impact of dietary supplementation with a blend of chestnut and quebracho tannins on the rumen microbiota of Holstein steers. Bacterial richness was lower in tannins treated animals, while the overall population structure of rumen microbiota was not significantly disturbed by tannins. e ratio of the phyla Firmicutes and Bacteroidetes, a parameter associated with energy harvesting function, was increased in tannins supplemented animals, essentially due to the selective growth of Ruminococcaceae over members of genus Prevotella. Fibrolytic, amylolytic, and ureolytic bacterial communities in the rumen were altered by tannins, while methanogenic archaea were reduced. Furthermore, ruminal pH was significantly higher in animals supplemented with tannins than in the control group, while urease activity exhibited the opposite pattern. Further work is necessary to assess the relation between tannins impact on rumen microbiota and alteration of rumen fermentation parameters associated with bovine performance. 1. Introduction e use of bioactive phytochemicals as natural feed additives has been extensively studied as a strategy to manipulate rumen fermentation in pursuit of an improvement in cattle productivity by reducing methanogenesis and increasing the efficiency of nitrogen utilization [1–3]. However, the lack of sufficient understanding about the rumen microbiome is considered one of the major knowledge gaps that hinder effective enhancement of the rumen function. Tannins are a complex group of polyphenolic compounds found in many plants species which are commonly included in ruminant diets such as forage and sorghum [4]. Tannins are classified as hydrolysable or condensed based on their chemical structure. e molecular structure of hydrolysable tannins includes a core of glucose esterified with gallic and hexahydroxydiphenic acids. Condensed tannins are usually referred to as proanthocyanidins, polymers of flavon-3-ols or flavon-3, 4-diols such as catechin and epicatechin [5]. Tannins can complex with proteins, starch, vitamins, and minerals at moderate pH, as in ruminal conditions, and dissociate at lower pH, including abomasum and the initial portion of the duodenum [6]. e increased digestibility and efficiency of feed utilization induced by addition of tannins Hindawi BioMed Research International Volume 2017, Article ID 9610810, 11 pages https://doi.org/10.1155/2017/9610810
Transcript of Impact of Chestnut and Quebracho Tannins on Rumen...

Research ArticleImpact of Chestnut and Quebracho Tannins on RumenMicrobiota of Bovines
JuanMariacutea Diacuteaz Carrasco12 Claudio Cabral3
Leandro Martiacuten Redondo12 Natalia Daniela Pin Viso24 Dariacuteo Colombatto25
Marisa Diana Farber24 andMariano Enrique Fernaacutendez Miyakawa12
1 Instituto de Patobiologıa Centro Nacional de Investigaciones Agropecuarias Instituto Nacional de Tecnologıa AgropecuariaCalle Las Cabanas y Los Reseros sn Casilla de Correo 25 Castelar 1712 Buenos Aires Argentina2Consejo Nacional de Investigaciones Cientıficas y Tecnicas Godoy Cruz 2290 1425 Buenos Aires Argentina3Animal Nutrition Silvateam Indunor Cerrito 1136 1010 Buenos Aires Argentina4Instituto de Biotecnologıa Centro Nacional de Investigaciones Agropecuarias Instituto Nacional de Tecnologıa AgropecuariaCalle Las Cabanas y Los Reseros sn Casilla de Correo 25 Castelar 1712 Buenos Aires Argentina5Departamento de Produccion Animal Facultad de Agronomıa Universidad de Buenos Aires Av San Martın 44531417 Buenos Aires Argentina
Correspondence should be addressed to Mariano Enrique Fernandez Miyakawa fernandezmiyakawamintagobar
Received 28 July 2017 Accepted 3 December 2017 Published 28 December 2017
Academic Editor Yiannis Kourkoutas
Copyright copy 2017 JuanMarıaDıazCarrasco et alThis is an open access article distributed under theCreativeCommonsAttributionLicense which permits unrestricted use distribution and reproduction in anymedium provided the originalwork is properly cited
The use of phytogenic dietary additives is being evaluated as a means to improve animal productivity The effect of tannins seemsto be the influence not only directly on the digestive process through binding of dietary proteins but also indirectly over theireffects on gastrointestinal microbiota High-throughput sequencing of 16S rRNA gene was used to analyze the impact of dietarysupplementation with a blend of chestnut and quebracho tannins on the rumen microbiota of Holstein steers Bacterial richnesswas lower in tannins treated animals while the overall population structure of rumenmicrobiota was not significantly disturbed bytanninsThe ratio of the phyla Firmicutes and Bacteroidetes a parameter associated with energy harvesting function was increasedin tannins supplemented animals essentially due to the selective growth of Ruminococcaceae over members of genus PrevotellaFibrolytic amylolytic and ureolytic bacterial communities in the rumen were altered by tannins while methanogenic archaea werereduced Furthermore ruminal pH was significantly higher in animals supplemented with tannins than in the control group whileurease activity exhibited the opposite pattern Further work is necessary to assess the relation between tannins impact on rumenmicrobiota and alteration of rumen fermentation parameters associated with bovine performance
1 Introduction
The use of bioactive phytochemicals as natural feed additiveshas been extensively studied as a strategy to manipulaterumen fermentation in pursuit of an improvement in cattleproductivity by reducing methanogenesis and increasing theefficiency of nitrogen utilization [1ndash3] However the lackof sufficient understanding about the rumen microbiome isconsidered one of the major knowledge gaps that hindereffective enhancement of the rumen function
Tannins are a complex group of polyphenolic compoundsfound in many plants species which are commonly included
in ruminant diets such as forage and sorghum [4] Tanninsare classified as hydrolysable or condensed based on theirchemical structure The molecular structure of hydrolysabletannins includes a core of glucose esterified with gallic andhexahydroxydiphenic acids Condensed tannins are usuallyreferred to as proanthocyanidins polymers of flavon-3-olsor flavon-3 4-diols such as catechin and epicatechin [5]Tannins can complex with proteins starch vitamins andminerals at moderate pH as in ruminal conditions anddissociate at lower pH including abomasum and the initialportion of the duodenum [6] The increased digestibility andefficiency of feed utilization induced by addition of tannins
HindawiBioMed Research InternationalVolume 2017 Article ID 9610810 11 pageshttpsdoiorg10115520179610810
2 BioMed Research International
to ruminant diets have been attributed to their ability toprecipitate proteins allowing bypass of ruminal digestion andenhancing protein availability at small intestinal level How-ever ruminal bypass cannot entirely explain the performanceimprovement associated with tannins addition in feed
For a long time tannins were thought to reduce weightgain and efficiency of nutrient utilization However it is nowknown that their effectmay be either beneficial or detrimentaldepending on tannins origin molecular structure dosageand animal species [4 7ndash9] High doses of tannins reducevoluntary feed intake andnutrient digestibility whereasmod-erate concentrations can improve feed utilization [4] A blendof tannins extracted from quebracho and chestnut tree hasbeen used as additives to improve performance of ruminantsand to reduce urinary nitrogen excretion [10] A recent reportshowed that addition of moderate concentrations of chestnutand quebracho tannins to the diets of dairy cows did notaffect animal performance but increased milk protein yieldand decreased urinary nitrogen excretion [11] Other authorsobserved that inclusion of chestnut and quebracho tanninsincreased drymatter intake average daily gain and final bodyweight of steers during the finishing feedlot phase [12] Thesetwo types of tannins differ in their nutritional role and toxiceffects in livestock nutrition
Tannins modify the digestive processes of ruminants notonly by binding dietary protein but also through modulationof rumen microbiota [7 13] The bovine rumen houses acomplex and highly dense microbiota that is responsible forcattle ability to convert indigestible plant mass into energy Inrecent years a link between gut microbiota composition andenergy harvesting function has been observed in humans andmice [14 15] Rumen microbiota composition was found tobe strongly correlated with daily milk-fat yield in bovines [1617] The microbial populations of the rumen and the varia-tions associatedwith diet have been previously described [18]However although much research has been done regardingthe effects of tannins on ruminants physiology and theirmetabolic fate [19] the impact of chestnut and quebrachotannins on rumen microbiota of bovines has not been fullydescribed The hypothesis under study in the present workis that tannins alter the bacterial populations of the rumenand therefore can be used as a dietary strategy to modulaterumen function The aim of this study was to analyze thein vivo effects of a tannins blend derived from chestnut andquebracho on rumen bacterial populations of Holstein cattleby means of massive 16S rRNA gene sequencing exploringthe relationship between rumen microbiome compositionand physiological parameters
2 Materials and Methods
21 Animal Handling Dietary Treatments and Determinationof Rumen Parameters The study was carried out using rumi-nally fistulated Holstein cows of the HolandominusArgentinobreed (119899 = 6) with an average body weight of 584 plusmn 12 kg(mean plusmn SD) In order to emulate productive conditions dietwas gradually changed from low starch (60 alfalfa bale40 concentrate composed of 80 ground corn grain and20 soybean meal) to high starch (20 alfalfa bale 80
Table 1 Formulation and composition of diet as percentage of drymatter
Ingredients of DMAlfalfa bale 190Ground corn grain 640Soybean meal 160Trace mineral and vitamins 08Tannins blend 02Total 1000Composition analysis of DMCP 150RDP 93Total calcium 09Total phosphorus 04Energy analysis McalkgME 297NEm 201NEg 135DM dry matter CP crude protein RDP rumen degradable protein MEmetabolizable energy NEm net energy for maintenance NEg net energyfor gain
concentrate) over an adaptation period of 14 days (from d1 to d 14) Animals were kept on high starch diet for 21days (from d 15 to d 35) before the beginning of dietarytreatments in order to ensure complete adaptation of rumenmicrobiota On day 36 animals were randomly divided intotwo groups of 3 steers and each group was assigned to oneof the two dietary treatments (1) control group withoutadditives or (2) tannins-supplemented group in which ablend of chestnut and quebracho tannins was added to dietat a concentration of 2 g per kg of feed (Table 1) Controland tannins-supplemented diets were administered to theanimals until the end of the treatment period (from d 36to d 48) The twelve-day treatment period was repeatedonce after a ldquowashoutrdquo period of 21 days during which allanimals received the control diet Ruminal samples weretaken from each animal at the end of each treatment periodfor microbiota analysis Diet was offered ad libitum duringthe whole experiment as a total mixed ration once daily at0800 h The average dry matter intake was 128 plusmn 06 kg peranimal per day (mean plusmn SD)
The tannins blend was obtained from Silvateam (IndunorSA Argentina) and contained one-third chestnut wood(Castanea sativa) tannins extract and two-thirds quebracho(Schinopsis lorentzii) tannins extract by weight Quebrachoextract ismainly composed of flavan-3-ols condensed tannins(gt84) while chestnut extract contains mainly digalloylglucose hydrolysable tannins (gt78) A detailed descriptionof quebracho and chestnut extracts chemical composition hasbeen described elsewhere [11]
Ruminal contents were collected via a dorsal fistula fromthe ventral sac of the rumen taking handfuls of materialfrom the interface between the solid material and liquidlayer Samples were collected in sterile containers (200mLincluding solid and liquidmaterial) frozen in liquid nitrogen
BioMed Research International 3
and stored at minus80∘C until further processing Part of thesample was used for determination of ruminal physiologicalparameters The ruminal liquor pH was measured usinga standard pH meter Urease activity (UA) was measuredaccording to the CaskeyminusKnappmethodmodified by AACCNonprotein nitrogen (NPN) was determined through Kjel-dahl method (VELP Scientifica Italy) Animals were caredfor by trained personnel only and the experimental protocoland procedures used were conducted according to protocol272011 of the Institutional Committee for the Care and Useof Experimental Animals (CICVyA-INTA)
22 DNA Extraction Twenty milliliters of evenly homoge-nized ruminal liquor was lyophilized before DNA extractionin order tomaximizemicrobial density (1 g of drymaterial persample) DNA extraction was conducted using the QIAampDNA stool kit (Qiagen Hilden Germany) following theinstructions of the manufacturer with slight modificationsBriefly 100mg of lyophilized rumen was weighed and lysedby incubation for 5min at 95∘C DNA elution was donewith 100 120583l of Buffer AE after incubation for 10min atroom temperature DNA quality was assessed by agarosegel electrophoresis and DNA concentration was determinedusing a NanoDrop ND-1000 spectrophotometer (NanoDropTechnologies Wilmington DE USA) DNA was kept atminus20∘C until further processing
23 High-Throughput Sequencing of 16S rRNA Gene The16S rRNA gene V3-V4 regions were amplified using Illu-mina primers (forward 51015840 CCTACGGGNGGCWGCAG 31015840reverse 51015840 GGACTACHVGGGTATCTAATCC 31015840) with stan-dard adapter sequences attached for barcoding and multi-plexing 16S gene libraries construction and high-throughputsequencing were performed at Macrogen Inc (Seoul SouthKorea) in the Illumina MiSeq platform following manu-facturerrsquos instructions [20] In order to reduce unbalancedand biased base compositions 15 of PhiX control librarywas spiked into the amplicon pool Due to an issue withthe length of reverse reads which were not long enough toachievemerging of paired-end sequences only forward readscovering V3 and its flanking regions were used for furtherbioinformatics analysis The datasets generated in this studyare available under request
24 Bioinformatics Analysis FASTQ files were trimmedusing Trimmomatic v033 [21] which removed all primerand adapter sequences and also removed leading and trailingbases if quality value was below 9 and 3 respectivelySliding window trimming was also performed as well ascutting if the average quality within a 4-base window fallsbelow Q15 Demultiplexing and quality filtering were doneusing the script split libraries fastqpy which is part of theQIIME v191 software package [22] A threshold of Phredquality score (119876 gt 20) of the base was chosen forstringent quality control processing Chimeric sequenceswere filtered out in QIIME using the USERCH algorithmOpen-reference operational taxonomic units (OTUs) pickingwas performed using UCLUST and USEARCH algorithmsin QIIME Each sequence was assigned taxonomy against
Greengenes reference OTU build version 138 using a 97sequence similarity threshold OTUs with abundance below0005 were removed from final OTU table in order toavoid microbial diversity overestimation [23] Normalizationof OTU counts was done by performingmultiple rarefactionsfrom 10000 to 210000 sequences with steps of 10000 andwith 10 repetitions at each rarefaction depth The resultingmultiple rarefied OTU tables were used for all furtheranalysis Principal coordinate analysis (PCoA) plots weregenerated in QIIME with default options using a distancematrix calculated by unweighted UniFrac metric The signif-icance of grouping in the PCoA plot was tested by analysis ofsimilarity (ANOSIM) in QIIME with 999 permutationsThiswork used computational resources from the BioinformaticsUnit Instituto de Biotecnologıa (CICVyA-INTA)
25 Statistical Analysis Relative abundances of bacterialpopulations were statistically analyzed using STAMP v213[24] The relative abundances of bacterial taxa in controland tannins treated groups were compared at each levelof classification (phylum class order family and genus)usingWhitersquos nonparametric two-tailed 119905-test with 1000 per-mutations Comparisons in physiological data and diversityestimators were calculated using nonparametric two-tailedMannminusWhitney test (GraphPad Software San Diego CAUSA) and were considered statistically significant if 119901 lt 005
3 Results
After high-throughput sequencing 3812179 reads wereobtained with an average of 346562 plusmn 42326 reads persample Stringent filtration of the sequences based on lengthand quality was performed before taxonomy assignationresulting in 2951356 reads with an average length of 267 plusmn22 base pairs
The total number of OTUs detected after filtration was2263 but this number exhibited a high interindividualvariation (Figure 1) We found that the number of OTUstended to be lower in tannins-supplemented animals thanin the control group (119901 = 005) (Figure 2(a)) Shannonrsquosdiversity index which estimates the internal sample com-plexity was not significantly affected by tannins (119901 = 014)(Figure 2(b)) PCoA based on unweighted UniFrac metricwas performed in order to explore dissimilarities inmicrobialcomposition of the rumen among treated groups (Figure 3)ANOSIM detected no significant differences in bacterialdiversity between control and tannins dietary treatments (119901 =086)
Firmicutes andBacteroideteswere the dominant bacterialphyla in the bovine ruminal fluid accounting for nearly 90of total microbiota However large interindividual variancewas observed in the relative abundance of Bacteroidetes andFirmicutes among different animals In the control groupbacterial populations belonging to phylum Bacteroideteswere the most abundant in all animals (521 on average)while Firmicutes accounted for 376 of total microbiotaHowever this predominance was inverted in the tanninstreated animals with a significantly higher percentage ofFirmicutes (462 119901 = 002) and a reduction to 446 in
4 BioMed Research International
0
500
1000
1500
2000
2500
OTU
s
100000 150000 20000050000Sequences per sample
Figure 1 OTUs rarefaction curves of rumen microbiota based on16S rRNA gene sequences OTUs were picked using the UCLUSTmethod with 3 dissimilarity in QIIME Each curve corresponds toa single ruminal sample
Bacteroidetes (119901 = 018) Accordingly steers supplementedwith tannins presented a trend to higher Firmicutes toBacteroidetes ratio in comparison with the control group(108 versus 073 119901 = 009) (Figure 4)
Significant differences in the abundance of certain taxawere detected between control and tannins treated animals(Figure 5) Among Bacteroidetes the most abundant genuswas Prevotella accounting for more than 40 of this phylumThe average abundance of Prevotella was lower in tannins-supplemented animals than in the control group althoughit exhibited a high degree of variance among animals (165versus 219 119901 = 015) Clostridia was the predominantclass which accounted for more than 90 of total Firmicutesand it was significantly enhanced in tannins treated animals(415 versus 346 119901 lt 0001) Among Clostridia Ruminococ-caceae was the most abundant family and showed a sig-nificantly higher abundance in the tannins-supplementedanimals (178 versus 107 119901 = 0009) In the controlgroup most sequences corresponding to family Ruminococ-caceae belonged to unclassified members (79) and genusRuminococcus (26) Both taxa were enhanced in tanninstreated steers reaching abundances of 126 (119901 = 001) and49 (119901 = 007) respectively Other nonclostridial bacteriawithin the phylum Firmicutes were significantly altered bytannins including members of class Erysipelotrichi SomeErysipelotrichi were enhanced (genus L7A-E11 119901 = 002and genus p-75-A5 119901 = 006) while others were lowered(genus RFN20 119901 = 0001) in tannins-supplemented animalsMembers of class Bacilli (genera Streptococcus and Lacto-bacillus) showed onlymoderate increases in their abundanceMeanwhile genus Fibrobacter was significantly affected bytannins accounting for 010 of total microbiota in thecontrol group and only 0005 in the tannins treated animals(119901 = 001) Other minor fibrolytic bacteria were significantlymore abundant in tannins treated steers including genusBlautia (008 versus 002 119901 = 001) and member of familyEubacteriaceae genus Anaerofustis (006 versus 002 119901 =003)
Among sugar fermenting bacteria the most abundanttaxonwas genus Prevotella whose abundance was reduced by54 in tannins treated animals as mentioned above GenusTreponema was also reduced in tannins treated steers (041versus 121 119901 = 004) Among Veillonellaceae membersgenus Succiniclasticum which specializes in fermenting suc-cinate to propionate doubled their levels in tannins treatedanimals (399 versus 199 119901 = 008) Lipolytic genusAnaerovibrio was significantly enhanced by tannins (011 ver-sus 005 119901 = 001) Genus Selenomonas was also increasedin tannins supplemented animals (011 versus 005 119901 =007) Among ureolytic bacteria genus Butyrivibrio wasthe most abundant one and it was negatively affected bytannins treatment (180 versus 236) as well as Treponemaand Succinivibrio (0009 versus 002) On the other handmethanogens belonging to phylum Euryarchaeota were lessabundant in tannins supplemented steers (137 versus 203)and their levels were inversely correlated with rumen pH (119903 =minus080) Genus Methanosphaera was significantly reduced bytannins (006 versus 016 119901 = 001)
Determination of pH urease activity and NPN wasperformed in all rumen samples along with microbiotacomposition analysis Tannins treated steers had significantlyhigher ruminal pH than the control group (630 versus 588119901 = 002) (Figure 6(a)) Urease activity exhibited the oppositepattern showing a significant decline in the tannins treatedsteers (Figure 6(b)) Moreover a strong negative correlationbetween pH and urease activity was detected (119903 = minus095)NPNwas not significantly altered by treatments (Figure 6(c))
4 Discussion
Rumen microbiome diversity is a key feature of ruminantsthat confers cattle the ability to adapt to a wide range ofdietary conditions [25] In recent years the concept of hostmicrobiome individuality in ruminants is gaining supportsince numerous studies found a large number of taxa whosepresence or abundance in the rumen varies markedly amongindividuals [26ndash28] Dietary tannins diminished ruminalrichness but did not significantly affect the bacterial com-munitiesrsquo complexity (ie balance between the relative abun-dances of taxa) A recent report found an increase in rumenrichness but no change in Shannonrsquos diversity index aftersupplementation with a blend of polyphenols essential oil indairy heifers under a high grain diet supporting the idea thatpolyphenols can affect bacterial richness without disruptingthe overall rumen microbiota population structure [29] Inline with this 120573-diversity analysis detected no significantdifferences in rumen bacterial diversity between control andtannins treated steers Low microbial richness in the rumenhas been recently found to be tightly linked to a higher feedefficiency in dairy cows [30] The authors suggest that lowerrichness in the rumen of efficient animals results in a simplermetabolic network which leads to higher concentrationsof specific components that are used to support the hostrsquosenergy requirements Together diversity analyses suggest thatbacterial richness was decreased while the overall bacterialcomplexity of the rumen was not significantly affected bychestnut and quebracho tannins supplementation
BioMed Research International 5
Control Tannins1000
1100
1200
1300
1400
1500
1600
1700
1800
1900
2000
2100
2200N
umbe
r of O
TUs
p = 005
(a)Control Tannins
6
7
8
9
10
Shan
nonrsquos
inde
x
p = 014
(b)
Figure 2 Effect of tannins treatment on (a) bacterial richness (number of OTUs) and (b) Shannonrsquos diversity index of rumen microbiomeLine = median Box = 25ndash75 percentiles Bar = 5ndash95 percentiles
minus010
minus005
000
005
010
015
PC2
(11
1)
00 01 02 03 04 05minus01PC1 (556)
Figure 3 PCoA plot based on unweighted UniFrac metric Itemsshaped with triangles and circles correspond to samples fromcontrol and tannins treated animals respectively Axes (PC1 = 556and PC2 = 111) account for 667 of the total variation detected
The dominance of phyla Firmicutes and Bacteroidetesin the bovine ruminal fluid is a common feature in thegastrointestinal microbiome of monogastric organisms andruminants [15 17 28 31] Henderson et al also found thatan increase in the ruminal abundance of total Firmicutescorrelated with a decrease in the abundance of Bacteroidetesboth in cows (119903 = minus080) and in sheep (119903 = minus097) [32] Otherauthors found that Bacteroidetes were the most abundant
Control Tannins
p = 009
04
05
06
07
08
09
10
11
12
13
14
15
16
17
Firm
icut
esB
acte
roid
etes
Figure 4 Effect of tannins on the ratio of phyla Firmicutes andBacteroidetes in rumen microbiota Line = median Box = 25ndash75percentiles Bar = 5ndash95 percentiles
phylum in ruminal samples obtained from dairy cows butsome animals exhibited a higher percentage of Firmicutescompensating for a lower abundance of Bacteroidetes [17]Our results agree with these observations since a stronginverse correlation between the abundances of Firmicutesand Bacteroidetes was detected (119903 = minus099) These resultssuggest that members of Firmicutes and Bacteroidetes com-pete for available resources in the rumen and tannins wouldtip the balance in favor of Firmicutes
6 BioMed Research International
lowast
lowastlowast
lowast
lowastlowast
lowast
lowast
lowastlowastlowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
lowast
lowastlowast
lowast
lowast
lowast
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
minus4 minus2 minus1minus3minus5 0 2 3 4 51
Phylum FirmicutesGenus Butyrivibrio
Unc Family RuminococcaceaeGenus Ruminococcus
Genus SucciniclasticumGenus Streptococcus
Genus p-75-a5Genus L7A-E11
Genus SelenomonasGenus Anaerovibrio
Genus BlautiaGenus Anaerofustis
Genus RFN20Genus Lachnospira
Phylum BacteroidetesFamily Prevotellaceae
Family BacteroidaceaeGenus Prevotella
Genus BacteroidesGenus YRC22
Unc Family RF16
Unc Family CoriobacteriaceaePhylum Actinobacteria
Phylum SpirochaetesGenus Treponema
Genus Sphaerochaeta
Phylum EuryarchaeotaGenus Methanobrevibacter
Genus Methanosphaera
Phylum ProteobacteriaUnclass Order GMD14H09
Genus RuminobacterGenus Oxalobacter
Genus Succinivibrio
Phylum TenericutesUnc Family Anaeroplasmata
Genus Anaeroplasma
Phylum ChloroflexiGenus SHD-231
Phylum FibrobacteresGenus Fibrobacter
log 2 fold change
Figure 5 Relative fold changes (log 2 tanninscontrol) in the abundance of rumen bacterial taxa between control and tannins treated steerslowastlowast119901 lt 005 lowast119901 lt 010 Bar = SEM
The ratio of Firmicutes to Bacteroidetes has been shownto affect energy harvesting and body fat accumulation inhumans and mice [14 15] Along with increased fatty acidabsorption more energy was found to be efficiently obtainedfrom diet in obese mice illustrating the connection betweenFirmicutes and improved efficiency in energy harvesting [14]In cows the Firmicutes to Bacteroidetes ratio was found tobe strongly correlated with daily milk-fat yield [17] A recentstudy found that the abundance of Firmicutes in the rumen
positively correlates with the average daily body weight gainin steers suggesting that these bacteria play a significant rolein feed efficiency of bovines [33] Therefore it is possiblethat the increase of Firmicutes to Bacteroidetes ratio inducedby tannins can improve bovine performance as previouslysuggested by other authors [19]
Fiber degradation is a complex process carried out bya group of microorganisms that are able to digest plantpolysaccharides mainly through production of cellulolytic
BioMed Research International 7
Control Tannins50
52
54
56
58
60
62
64
66
68
70pH
p = 002
(a)Control Tannins
000
005
010
015
020
Ure
ase a
ctiv
ity (U
A)
p = 0007
(b)
Control Tannins
000
005
010
015
Non
prot
ein
nitro
gen
(NPN
)
p = 0849
(c)
Figure 6 Effect of tannins on pH urease activity and NPN in the rumen liquor Line = median Box = 25ndash75 percentiles Bar = 5ndash95percentiles Urease activity values are given in pH units proportional to urease activity NPN levels are expressed as a percentage of solublenitrogen in the rumen
hemicellulolytic and pectinolytic enzymes [34] Cellulolyticactivity in the rumen involves a diverse bacterial commu-nity whose main members belong to genera Ruminococcusand Fibrobacter while the main hemicellulolytic bacteriabelong to genera Ruminococcus Prevotella and Butyrivib-rio [34ndash37] The blend of tannins administrated increasedRuminococcaceae and other members of phylum Firmicuteswhile inhibited genera Prevotella and Fibrobacter Prevotellaspecies have a documented role in metabolism of starchhemicellulose pectin and protein catabolism [25] The highabundance of this genus might be the result of a largemetabolic niche that is occupied by different species with
similar metabolic capabilities or it might be associated witha high genetic variability that enables members of genusPrevotella to occupy different ecological niches within therumen [38ndash40] For instance Prevotella abundance has beenshown to be higher in the rumen of animals of beef cattlewith low feed efficiency [41] Previous studies reported a greatdiversity in the sensitivity ofPrevotella species to tannins [42]which could explain the lower abundance detected for thisgenus in tannins-supplemented steers Since genus Prevotellais characterized by a high genetic diversity and wide array ofoutputmetabolites [38ndash40] their replacement byRuminococ-cus may lead to a simpler product profile specialized to
8 BioMed Research International
support the hostrsquos energy requirements [30] Our resultsagree with previous studies that found a significant reduc-tion in Fibrobacter and Prevotella rumen populations aftersupplementation with condensed and hydrolysable tanninsincluding chestnut and quebracho tannins [43 44]
Rumen fibrolytic function is carried out by a largelyredundant microbial community with overlapping distribu-tion of metabolic capabilities and this fibrolytic communityhas the ability to restore its structure and function afterperturbation a phenomenon known as resilience [27] In thiscontext the observed changes in diversity of fibrolytic bacte-ria in tannins treated steers may result from a combinationof physicochemical and biological mechanisms described inthe literature including direct interaction of tannins withfiber [45] which could alter the available surface area formicrobial attack the inhibition of certain fibrolytic taxa bymeans of tannins antimicrobial activity [46] andmodulationof fibrolytic bacterial species driven by changes in rumen pHsince certain fibrolytic taxa of the rumen are inhibited at lowpH under high-grain diets [36 47]
Rumen amylolytic and saccharolytic bacteria were alsoaffected by dietary treatment with tannins mainly throughthe decrease of genera Prevotella and Treponema Otheramylolytic genera were moderately increased in tannins-fedsteers including Streptococcus Bifidobacterium and Lacto-bacillus Amylolytic activity is normally enhanced in rumi-nants consuming high-grain diets [18 48 49] The rate ofgrain degradation by these microbial communities plays akey role in maintaining rumen homeostasis since rapidstarch fermentation produces large amounts of organic acidstherefore producing a drop in ruminal pH that may leadto metabolic acidosis [50] A previous study showed thattannic acid and quebracho tannins lower the rate ofmicrobialhydrolysis of starch-rich grains in the rumen by physicalmodification of the endosperm protein matrix [45] Thisphysical modulation of starch degradation could explain thelower abundance of sugar fermenting taxa detected in tanninstreated steers as well as the higher ruminal pH observed inthis group
Fermentation products of microbial activity in rumenmainly short-chain volatile fatty acids serve as amajor sourceof energy for ruminants and have a direct impact on thephysiological parameters of the animal and feed utilizationefficiency [16] Some members of family Veillonellaceaewhich produce propionate as a major fermentation productand have been associated with lower methane emissions [51]were enhanced in tannins treated animals Interestingly someSelenomonas species can break tannin-protein complexes anduse tannins as energy source [52 53] Therefore the higherabundance of Selenomonas in tannins treated steers may bepartly due to availability of tannins as direct energy sourceMeanwhile members of class Erysipelotrichi which havebeen linked to beef cattle feed efficiency [33 51 54] were alsomodulated by tannins treatment
Methane production during fermentation of feeds in therumen represents a loss of 2ndash12 of gross energy [1] andit is performed by a group of archaea known collectivelyas methanogens which belong to phylum EuryarchaeotaThe microorganisms produce methane the second largest
anthropogenic greenhouse gas which has a global warmingpotential 25 times that of carbon dioxide [55] In the presentstudy a reduction of methanogenic archaea in tannins sup-plemented steers was detected as well as an inverse cor-relation between the abundances of phylum Euryarchaeotaand rumen pH (119903 = minus095) Tannins are thought todirectly inhibit methanogens as well as indirectly limitmethanogenesis through reduction of hydrogen availability[1 55] Saminathan et al found a significant decrease in genusMethanobrevibacter after treatment of rumen samples withcondensed tannins in vitro [56] Other authors describeda linear decrease of genus Methanobrevibacter after dietarysupplementation with condensed tannins from pine bark ingoats [31]
Ruminal pH was significantly higher in tannins treatedsteers than in the control group while urease activity exhib-ited the opposite pattern These results agree with previousreports which found an increase in ruminal pH after sup-plementation with chestnut and quebracho tannins [57 58]Other authors observed that feeding chestnut and quebrachotannins decreased urease activity in the faeces of cows[10 11]
Feed-grade urea is an effective source of nitrogen com-monly used in beef cattle diets Ureolytic bacteria in therumen produce urease to hydrolyse urea to ammonia whichis subsequently used for the synthesis of amino acids andmicrobial protein Normally the rate of urea hydrolysisexceeds the rate of ammonia utilization which leads to poorefficiency of urea utilization and increases toxic ammoniaconcentrations in blood [59] Ureolytic bacteria in the rumencomprise a highly diverse group whose main species belongto genera Succinivibrio Treponema Bacteroides ButyrivibrioStreptococcus andBifidobacterium [60 61]We found that themost abundant ureolytic generaButyrivibrio andTreponemawere negatively affected by tannins treatment Thus theobserved decline in ruminal urease activity may be relatedwith the decrease of these urease-producing taxa A previousstudy also observed a drop in ruminal urease activity afteraddition of tannins to diet but a direct interaction betweentannins and urease enzyme was postulated as responsible forthis inhibition [62]
5 Conclusions
The current study showed that chestnut and quebrachotannins added to the diet of Holstein steers modifiedrumen microbiota composition particularly fiber and starchdegrading bacteria mainly by reducing the abundanceof Prevotella and Fibrobacter while favoring Ruminococ-caceae and other members of phylum Firmicutes Tanninstreatment significantly increased pH and decreased ureaseactivity in the ruminal liquor Further work is necessaryto assess the possible relation between tannins ability tomodify rumen microbiota composition and the alterationof rumen fermentation parameters associated with energyand feed efficiency of beef cattle such as the profile ofshort-chain fatty acids and the emissions of ammonia andmethane
BioMed Research International 9
Conflicts of Interest
The authors declare that there are no known conflicts ofinterest associated with this publication and there has beenno financial support for this work that could have influencedits outcome
Acknowledgments
The authors thank Alfredo Ivan Martınez Caceres for histechnical assistance during installation and startup of thebioinformatics tools used in this study This work wassupported by grants from INTA (PNSA 1115052 and PNBIO1131043) Additional support was provided by CONICET
References
[1] A K Patra and J Saxena ldquoDietary phytochemicals as rumenmodifiers a review of the effects on microbial populationsrdquoInternational Journal of General and Molecular Microbiologyvol 96 no 4 pp 363ndash375 2009 Antonie Van Leeuwenhoek
[2] R Bodas N Prieto R Garcıa-Gonzalez S Andres F JGiraldez and S Lopez ldquoManipulation of rumen fermentationand methane production with plant secondary metabolitesrdquoAnimal Feed Science and Technology vol 176 no 1-4 pp 78ndash932012
[3] E Oskoueian N Abdullah and A Oskoueian ldquoEffects offlavonoids on rumen fermentation activity methane produc-tion andmicrobial populationrdquo BioMed Research Internationalvol 2013 Article ID 349129 8 pages 2013
[4] P Frutos GHervas F J Giraldez andA RMantecon ldquoReviewtannins and ruminant nutritionrdquo Spanish Journal of AgriculturalResearch vol 2 no 2 pp 191ndash202 2004
[5] K Khanbabaee and T van Ree ldquoTannins classification anddefinitionrdquo Natural Product Reports vol 18 no 6 pp 641ndash6492001
[6] L J Porter ldquoStructure and chemical properties of the con-densed tanninsrdquo Plant Polyphenols pp 245ndash258 1992 Springer Boston MA USA
[7] B R Min C Wright P Ho J-S Eun N Gurung and RShange ldquoThe effect of phytochemical tannins-containing dieton rumen fermentation characteristics and microbial diversitydynamics in goats using 16S rDNA amplicon pyrosequencingrdquoAgriculture Food and Analytical Bacteriology vol 4 no 3 pp195ndash211 2014
[8] B S Gemeda and A Hassen ldquoEffect of tannin and speciesvariation on in vitro digestibility gas and methane productionof tropical browse plantsrdquoAsian-Australasian Journal of AnimalSciences vol 28 no 2 pp 188ndash199 2015
[9] Z Li A-D G Wright H Liu et al ldquoResponse of the rumenmicrobiota of sika deer (cervus nippon) fed different concen-trations of tannin rich plantsrdquo PLoS ONE vol 10 no 5 ArticleID e0123481 2015
[10] JM PowelM J Aguerre andMAWattiaux ldquoTannin extractsabate ammonia emissions from simulated dairy barn floorsrdquoJournal of Environmental Quality vol 40 no 3 pp 907ndash9142011
[11] M J Aguerre M C Capozzolo P Lencioni C Cabral andM A Wattiaux ldquoEffect of quebracho-chestnut tannin extractsat 2 dietary crude protein levels on performance rumenfermentation and nitrogen partitioning in dairy cowsrdquo Journal
of Dairy Science vol 99 no 6 Article ID 74473 pp 4476ndash44862016
[12] C Rivera-Mendez A Plascencia N Torrentera andR A ZinnldquoEffect of level and source of supplemental tannin on growthperformance of steers during the late finishing phaserdquo Journalof Applied Animal Research vol 45 pp 199ndash203 2017
[13] B R Min T N Barry G T Attwood and W C McNabbldquoThe effect of condensed tannins on the nutrition and health ofruminants fed fresh temperate forages a reviewrdquo Animal FeedScience and Technology vol 106 no 1-4 pp 3ndash19 2003
[14] P J Turnbaugh R E Ley M A Mahowald V MagriniE R Mardis and J I Gordon ldquoAn obesity-associated gutmicrobiomewith increased capacity for energy harvestrdquoNaturevol 444 no 7122 pp 1027ndash1031 2006
[15] R E Ley R Knight and J I Gordon ldquoThe human micro-biome Eliminating the biomedicalenvironmental dichotomyin microbial ecologyrdquo Environmental Microbiology vol 9 no 1pp 3-4 2007
[16] I Mizrahi ldquoThe role of the rumen microbiota in determiningthe feed efficiency of dairy cowsrdquo Beneficial Microorganismsin Multicellular Life Forms pp 203ndash210 2012 Springer BerlinHeidelberg
[17] E Jami B AWhite and IMizrahi ldquoPotential role of the bovinerumen microbiome in modulating milk composition and feedefficiencyrdquo PLoS ONE vol 9 no 1 Article ID e85423 2014
[18] S C Fernando H T Purvis F Z Najar et al ldquoRumenmicrobialpopulation dynamics during adaptation to a high-grain dietrdquoApplied and Environmental Microbiology vol 76 no 22 pp7482ndash7490 2010
[19] H P S Makkar ldquoEffects and fate of tannins in ruminantanimals adaptation to tannins and strategies to overcomedetrimental effects of feeding tannin-rich feedsrdquo Small Rumi-nant Research vol 49 no 3 pp 241ndash256 2003
[20] Illumina Protocol 16S Metagenomic Sequencing Library 2013[21] AM BolgerM Lohse and B Usadel ldquoTrimmomatic a flexible
trimmer for Illumina sequence datardquoBioinformatics vol 30 no15 pp 2114ndash2120 2014
[22] J Kuczynski J Stombaugh W A Walters A Gonzalez J GCaporaso and R Knight ldquoUsing QIIME to analyze 16S rRNAgene sequences frommicrobial communitiesrdquoCurrent Protocolsin Microbiology vol 1 1E5 pages 2012
[23] N A Bokulich S Subramanian J J Faith et al ldquoQuality-filtering vastly improves diversity estimates from Illuminaamplicon sequencingrdquo Nature Methods vol 10 no 1 pp 57ndash592013
[24] D Parks and R Beiko ldquoSTAMP statistical analysis of metage-nomic profilesrdquo Encyclopedia of Metagenomics pp 1ndash6 2013Springer New York NY USA
[25] J C McCann T A Wickersham and J J Loor ldquoHigh-throughput methods redefine the rumen microbiome and itsrelationshipwith nutrition andmetabolismrdquoBioinformatics andBiology Insights vol 8 pp 109ndash125 2014
[26] G Henderson F Cox S Ganesh A Jonker W Young and PH Janssen ldquoRumen microbial community composition varieswith diet and host but a coremicrobiome is found across a widegeographical rangerdquo Scientific Reports vol 5 no 14567 pp 1ndash132015
[27] P J Weimer ldquoRedundancy resilience and host specificity ofthe ruminal microbiota Implications for engineering improvedruminal fermentationsrdquo Frontiers in Microbiology vol 6 articleno 296 2015
10 BioMed Research International
[28] E Jami and I Mizrahi ldquoComposition and similarity of bovinerumenmicrobiota across individual animalsrdquo PLoS ONE vol 7no 3 Article ID e33306 2012
[29] R De Nardi G Marchesini S Li et al ldquoMetagenomic analysisof rumen microbial population in dairy heifers fed a high graindiet supplemented with dicarboxylic acids or polyphenolsrdquoBMC Veterinary Research vol 12 no 1 article no 29 2016
[30] S Kruger Ben Shabat G Sasson A Doron-Faigenboim etal ldquoSpecific microbiome-dependent mechanisms underlie theenergy harvest efficiency of ruminantsrdquoThe ISME Journal vol10 no 12 pp 2958ndash2972 2016
[31] B R Min S Solaiman R Shange and J-S Eun ldquoGastroin-testinal bacterial andmethanogenic archaea diversity dynamicsassociated with condensed tannin-containing pine bark diet ingoats using 16S rDNA amplicon pyrosequencingrdquo InternationalJournal of Microbiology vol 2014 Article ID 141909 2014
[32] G Henderson F Cox S Kittelmann et al ldquoEffect of DNAextraction methods and sampling techniques on the apparentstructure of cow and sheep rumen microbial communitiesrdquoPLoS ONE vol 8 no 9 Article ID e74787 2013
[33] P R Myer T P L Smith J E Wells L A Kuehn and HC Freetly ldquoRumen microbiome from steers differing in feedefficiencyrdquo PLoS ONE vol 10 no 6 Article ID e0129174 2015
[34] Y Wang and T A McAllister ldquoRumen microbes enzymes andfeed digestion-a reviewrdquo Asian-Australasian Journal of AnimalSciences vol 15 no 11 pp 1659ndash1676 2002
[35] J B Russell R EMuck and P JWeimer ldquoQuantitative analysisof cellulose degradation and growth of cellulolytic bacteria inthe rumenrdquo FEMS Microbiology Ecology vol 67 no 2 pp 183ndash197 2009
[36] DOKrause S EDenmanR IMackie et al ldquoOpportunities toimprove fiber degradation in the rumen microbiology ecologyand genomicsrdquo FEMS Microbiology Reviews vol 27 no 5 pp663ndash693 2003
[37] M Zorec M Vodovnik and R Marinsek-Logar ldquoPotentialof selected rumen bacteria for cellulose and hemicellulosedegradationrdquo Food Technology and Biotechnology vol 52 no 2pp 210ndash221 2014
[38] A Z Bekele S Koike and Y Kobayashi ldquoGenetic diversityand diet specificity of ruminal prevotella revealed by 16S rRNAgene-based analysisrdquo FEMSMicrobiology Letters vol 305 no 1pp 49ndash57 2010
[39] A RamsakM Peterka K Tajima et al ldquoUnravelling the geneticdiversity of ruminal bacteria belonging to the CFB phylumrdquoFEMS Microbiology Ecology vol 33 no 1 pp 69ndash79 2000
[40] J Purushe D E Fouts M Morrison et al ldquoComparativegenome analysis of prevotella ruminicola and prevotella bryan-tii insights into their environmental nicherdquo Microbial Ecologyvol 60 no 4 pp 721ndash729 2010
[41] C A Carberry D A Kenny S Han M S McCabe and S MWaters ldquoEffect of phenotypic residual feed intake and dietaryforage content on the rumen microbial community of beefcattlerdquo Applied and Environmental Microbiology vol 78 no 14pp 4949ndash4958 2012
[42] A L Molan G T Attwood B R Min and W C McNabbldquoThe effect of condensed tannins from Lotus pedunculatus andLotus corniculatus on the growth of proteolytic rumen bacteriain vitro and their possible mode of actionrdquo Canadian Journal ofMicrobiology vol 47 no 7 pp 626ndash633 2001
[43] A Jayanegara G Goel H P S Makkar and K Becker ldquoDiver-gence between purified hydrolysable and condensed tannin
effects on methane emission rumen fermentation and micro-bial population in vitrordquo Animal Feed Science and Technologyvol 209 pp 60ndash68 2015
[44] BMinW Pinchak K Hernandez et al ldquoEffects of plant tanninsupplementation on animal responses and in vivo ruminalbacterial populations associated with bloat in heifers grazingwheat forage1rdquo The Professional Animal Scientist vol 28 no 4pp 464ndash472 2012
[45] T F Martınez T A McAllister Y Wang and T Reuter ldquoEffectsof tannic acid and quebracho tannins on in vitro ruminalfermentation of wheat and corn grainrdquo Journal of the Scienceof Food and Agriculture vol 86 no 8 pp 1244ndash1256 2006
[46] L Marın E M Miguelez C J Villar and F Lombo ldquoBioavail-ability of dietary polyphenols and gut microbiota metabolismantimicrobial propertiesrdquo BioMed Research International vol2015 Article ID 905215 18 pages 2015
[47] J B Russell and D B Wilson ldquoWhy are ruminal cellulolyticbacteria unable to digest cellulose at low pHrdquo Journal of DairyScience vol 79 no 8 pp 1503ndash1509 1996
[48] K Tajima S Arai K Ogata et al ldquoRumen bacterial communitytransition during adaptation to high-grain dietrdquo Anaerobe vol6 no 5 pp 273ndash284 2000
[49] D W Goad C L Goad and T G Nagaraja ldquoRuminal micro-bial and fermentative changes associated with experimentallyinduced subacute acidosis in steersrdquo Journal of Animal Sciencevol 76 no 1 pp 234ndash241 1998
[50] J Hernandez J L Benedito A Abuelo and C CastilloldquoRuminal acidosis in feedlot from aetiology to preventionrdquoTheScientific World Journal vol 2014 Article ID 702572 2014
[51] A B De Menezes E Lewis M OrsquoDonovan B F OrsquoNeill NClipson and E M Doyle ldquoMicrobiome analysis of dairy cowsfed pasture or total mixed ration dietsrdquo FEMS MicrobiologyEcology vol 78 no 2 pp 256ndash265 2011
[52] I K Skene and J D Brooker ldquoCharacterization of tanninacylhydrolase activity in the ruminal bacterium selenomonasruminantiumrdquo Anaerobe vol 1 no 6 pp 321ndash327 1995
[53] G Goel A K Puniya and K Singh ldquoTannic acid resistance inruminal streptococcal isolatesrdquo Journal of Basic Microbiologyvol 45 no 3 pp 243ndash245 2005
[54] P R Myer J E Wells T P L Smith L A Kuehn and H CFreetly ldquoCecum microbial communities from steers differingin feed efficiencyrdquo Journal of Animal Science vol 93 no 11 pp5327ndash5340 2015
[55] S E Hook A-D GWright and BWMcBride ldquoMethanogensmethane producers of the rumen and mitigation strategiesrdquoArchaea vol 2010 Article ID 945785 11 pages 2010
[56] M Saminathan C C Sieo H M Gan N Abdullah C M VL Wong and Y W Ho ldquoEffects of condensed tannin fractionsof different molecular weights on population and diversity ofbovine rumen methanogenic archaea in vitro as determinedby high-throughput sequencingrdquo Animal Feed Science andTechnology vol 216 pp 146ndash160 2016
[57] F Hassanat and C Benchaar ldquoAssessment of the effect ofcondensed (acacia and quebracho) and hydrolysable (chestnutand valonea) tannins on rumen fermentation andmethane pro-duction in vitrordquo Journal of the Science of Food and Agriculturevol 93 no 2 pp 332ndash339 2013
[58] J Balcells A Aris A Serrano A R Seradj J Crespo and MDevant ldquoEffects of an extract of plant flavonoids (bioflavex)on rumen fermentation and performance in heifers fed high-concentrate dietsrdquo Journal of Animal Science vol 90 no 13 pp4975ndash4984 2012
BioMed Research International 11
[59] A K Patra ldquoUreaammonia metabolism in the rumen andtoxicity in ruminantsrdquo Rumen Microbiology From Evolution toRevolution pp 329ndash341 2015
[60] MAWoznyM P Bryant L VHoldeman andW E CMooreldquoUrease assay and urease producing species of anaerobes in thebovine rumen and human fecesrdquo Applied and EnvironmentalMicrobiology vol 33 no 5 pp 1097ndash1104 1977
[61] D Jin S Zhao P Wang et al ldquoInsights into abundant rumenureolytic bacterial community using rumen simulation systemrdquoFrontiers in Microbiology vol 7 article no 1006 2016
[62] H P SMakkar B Singh andRKDawra ldquoEffect of tannin-richleaves of oak (Quercus incana) on various microbial enzymeactivities of the bovine rumenrdquo British Journal of Nutrition vol60 no 2 pp 287ndash296 1988
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 BioMed Research International
to ruminant diets have been attributed to their ability toprecipitate proteins allowing bypass of ruminal digestion andenhancing protein availability at small intestinal level How-ever ruminal bypass cannot entirely explain the performanceimprovement associated with tannins addition in feed
For a long time tannins were thought to reduce weightgain and efficiency of nutrient utilization However it is nowknown that their effectmay be either beneficial or detrimentaldepending on tannins origin molecular structure dosageand animal species [4 7ndash9] High doses of tannins reducevoluntary feed intake andnutrient digestibility whereasmod-erate concentrations can improve feed utilization [4] A blendof tannins extracted from quebracho and chestnut tree hasbeen used as additives to improve performance of ruminantsand to reduce urinary nitrogen excretion [10] A recent reportshowed that addition of moderate concentrations of chestnutand quebracho tannins to the diets of dairy cows did notaffect animal performance but increased milk protein yieldand decreased urinary nitrogen excretion [11] Other authorsobserved that inclusion of chestnut and quebracho tanninsincreased drymatter intake average daily gain and final bodyweight of steers during the finishing feedlot phase [12] Thesetwo types of tannins differ in their nutritional role and toxiceffects in livestock nutrition
Tannins modify the digestive processes of ruminants notonly by binding dietary protein but also through modulationof rumen microbiota [7 13] The bovine rumen houses acomplex and highly dense microbiota that is responsible forcattle ability to convert indigestible plant mass into energy Inrecent years a link between gut microbiota composition andenergy harvesting function has been observed in humans andmice [14 15] Rumen microbiota composition was found tobe strongly correlated with daily milk-fat yield in bovines [1617] The microbial populations of the rumen and the varia-tions associatedwith diet have been previously described [18]However although much research has been done regardingthe effects of tannins on ruminants physiology and theirmetabolic fate [19] the impact of chestnut and quebrachotannins on rumen microbiota of bovines has not been fullydescribed The hypothesis under study in the present workis that tannins alter the bacterial populations of the rumenand therefore can be used as a dietary strategy to modulaterumen function The aim of this study was to analyze thein vivo effects of a tannins blend derived from chestnut andquebracho on rumen bacterial populations of Holstein cattleby means of massive 16S rRNA gene sequencing exploringthe relationship between rumen microbiome compositionand physiological parameters
2 Materials and Methods
21 Animal Handling Dietary Treatments and Determinationof Rumen Parameters The study was carried out using rumi-nally fistulated Holstein cows of the HolandominusArgentinobreed (119899 = 6) with an average body weight of 584 plusmn 12 kg(mean plusmn SD) In order to emulate productive conditions dietwas gradually changed from low starch (60 alfalfa bale40 concentrate composed of 80 ground corn grain and20 soybean meal) to high starch (20 alfalfa bale 80
Table 1 Formulation and composition of diet as percentage of drymatter
Ingredients of DMAlfalfa bale 190Ground corn grain 640Soybean meal 160Trace mineral and vitamins 08Tannins blend 02Total 1000Composition analysis of DMCP 150RDP 93Total calcium 09Total phosphorus 04Energy analysis McalkgME 297NEm 201NEg 135DM dry matter CP crude protein RDP rumen degradable protein MEmetabolizable energy NEm net energy for maintenance NEg net energyfor gain
concentrate) over an adaptation period of 14 days (from d1 to d 14) Animals were kept on high starch diet for 21days (from d 15 to d 35) before the beginning of dietarytreatments in order to ensure complete adaptation of rumenmicrobiota On day 36 animals were randomly divided intotwo groups of 3 steers and each group was assigned to oneof the two dietary treatments (1) control group withoutadditives or (2) tannins-supplemented group in which ablend of chestnut and quebracho tannins was added to dietat a concentration of 2 g per kg of feed (Table 1) Controland tannins-supplemented diets were administered to theanimals until the end of the treatment period (from d 36to d 48) The twelve-day treatment period was repeatedonce after a ldquowashoutrdquo period of 21 days during which allanimals received the control diet Ruminal samples weretaken from each animal at the end of each treatment periodfor microbiota analysis Diet was offered ad libitum duringthe whole experiment as a total mixed ration once daily at0800 h The average dry matter intake was 128 plusmn 06 kg peranimal per day (mean plusmn SD)
The tannins blend was obtained from Silvateam (IndunorSA Argentina) and contained one-third chestnut wood(Castanea sativa) tannins extract and two-thirds quebracho(Schinopsis lorentzii) tannins extract by weight Quebrachoextract ismainly composed of flavan-3-ols condensed tannins(gt84) while chestnut extract contains mainly digalloylglucose hydrolysable tannins (gt78) A detailed descriptionof quebracho and chestnut extracts chemical composition hasbeen described elsewhere [11]
Ruminal contents were collected via a dorsal fistula fromthe ventral sac of the rumen taking handfuls of materialfrom the interface between the solid material and liquidlayer Samples were collected in sterile containers (200mLincluding solid and liquidmaterial) frozen in liquid nitrogen
BioMed Research International 3
and stored at minus80∘C until further processing Part of thesample was used for determination of ruminal physiologicalparameters The ruminal liquor pH was measured usinga standard pH meter Urease activity (UA) was measuredaccording to the CaskeyminusKnappmethodmodified by AACCNonprotein nitrogen (NPN) was determined through Kjel-dahl method (VELP Scientifica Italy) Animals were caredfor by trained personnel only and the experimental protocoland procedures used were conducted according to protocol272011 of the Institutional Committee for the Care and Useof Experimental Animals (CICVyA-INTA)
22 DNA Extraction Twenty milliliters of evenly homoge-nized ruminal liquor was lyophilized before DNA extractionin order tomaximizemicrobial density (1 g of drymaterial persample) DNA extraction was conducted using the QIAampDNA stool kit (Qiagen Hilden Germany) following theinstructions of the manufacturer with slight modificationsBriefly 100mg of lyophilized rumen was weighed and lysedby incubation for 5min at 95∘C DNA elution was donewith 100 120583l of Buffer AE after incubation for 10min atroom temperature DNA quality was assessed by agarosegel electrophoresis and DNA concentration was determinedusing a NanoDrop ND-1000 spectrophotometer (NanoDropTechnologies Wilmington DE USA) DNA was kept atminus20∘C until further processing
23 High-Throughput Sequencing of 16S rRNA Gene The16S rRNA gene V3-V4 regions were amplified using Illu-mina primers (forward 51015840 CCTACGGGNGGCWGCAG 31015840reverse 51015840 GGACTACHVGGGTATCTAATCC 31015840) with stan-dard adapter sequences attached for barcoding and multi-plexing 16S gene libraries construction and high-throughputsequencing were performed at Macrogen Inc (Seoul SouthKorea) in the Illumina MiSeq platform following manu-facturerrsquos instructions [20] In order to reduce unbalancedand biased base compositions 15 of PhiX control librarywas spiked into the amplicon pool Due to an issue withthe length of reverse reads which were not long enough toachievemerging of paired-end sequences only forward readscovering V3 and its flanking regions were used for furtherbioinformatics analysis The datasets generated in this studyare available under request
24 Bioinformatics Analysis FASTQ files were trimmedusing Trimmomatic v033 [21] which removed all primerand adapter sequences and also removed leading and trailingbases if quality value was below 9 and 3 respectivelySliding window trimming was also performed as well ascutting if the average quality within a 4-base window fallsbelow Q15 Demultiplexing and quality filtering were doneusing the script split libraries fastqpy which is part of theQIIME v191 software package [22] A threshold of Phredquality score (119876 gt 20) of the base was chosen forstringent quality control processing Chimeric sequenceswere filtered out in QIIME using the USERCH algorithmOpen-reference operational taxonomic units (OTUs) pickingwas performed using UCLUST and USEARCH algorithmsin QIIME Each sequence was assigned taxonomy against
Greengenes reference OTU build version 138 using a 97sequence similarity threshold OTUs with abundance below0005 were removed from final OTU table in order toavoid microbial diversity overestimation [23] Normalizationof OTU counts was done by performingmultiple rarefactionsfrom 10000 to 210000 sequences with steps of 10000 andwith 10 repetitions at each rarefaction depth The resultingmultiple rarefied OTU tables were used for all furtheranalysis Principal coordinate analysis (PCoA) plots weregenerated in QIIME with default options using a distancematrix calculated by unweighted UniFrac metric The signif-icance of grouping in the PCoA plot was tested by analysis ofsimilarity (ANOSIM) in QIIME with 999 permutationsThiswork used computational resources from the BioinformaticsUnit Instituto de Biotecnologıa (CICVyA-INTA)
25 Statistical Analysis Relative abundances of bacterialpopulations were statistically analyzed using STAMP v213[24] The relative abundances of bacterial taxa in controland tannins treated groups were compared at each levelof classification (phylum class order family and genus)usingWhitersquos nonparametric two-tailed 119905-test with 1000 per-mutations Comparisons in physiological data and diversityestimators were calculated using nonparametric two-tailedMannminusWhitney test (GraphPad Software San Diego CAUSA) and were considered statistically significant if 119901 lt 005
3 Results
After high-throughput sequencing 3812179 reads wereobtained with an average of 346562 plusmn 42326 reads persample Stringent filtration of the sequences based on lengthand quality was performed before taxonomy assignationresulting in 2951356 reads with an average length of 267 plusmn22 base pairs
The total number of OTUs detected after filtration was2263 but this number exhibited a high interindividualvariation (Figure 1) We found that the number of OTUstended to be lower in tannins-supplemented animals thanin the control group (119901 = 005) (Figure 2(a)) Shannonrsquosdiversity index which estimates the internal sample com-plexity was not significantly affected by tannins (119901 = 014)(Figure 2(b)) PCoA based on unweighted UniFrac metricwas performed in order to explore dissimilarities inmicrobialcomposition of the rumen among treated groups (Figure 3)ANOSIM detected no significant differences in bacterialdiversity between control and tannins dietary treatments (119901 =086)
Firmicutes andBacteroideteswere the dominant bacterialphyla in the bovine ruminal fluid accounting for nearly 90of total microbiota However large interindividual variancewas observed in the relative abundance of Bacteroidetes andFirmicutes among different animals In the control groupbacterial populations belonging to phylum Bacteroideteswere the most abundant in all animals (521 on average)while Firmicutes accounted for 376 of total microbiotaHowever this predominance was inverted in the tanninstreated animals with a significantly higher percentage ofFirmicutes (462 119901 = 002) and a reduction to 446 in
4 BioMed Research International
0
500
1000
1500
2000
2500
OTU
s
100000 150000 20000050000Sequences per sample
Figure 1 OTUs rarefaction curves of rumen microbiota based on16S rRNA gene sequences OTUs were picked using the UCLUSTmethod with 3 dissimilarity in QIIME Each curve corresponds toa single ruminal sample
Bacteroidetes (119901 = 018) Accordingly steers supplementedwith tannins presented a trend to higher Firmicutes toBacteroidetes ratio in comparison with the control group(108 versus 073 119901 = 009) (Figure 4)
Significant differences in the abundance of certain taxawere detected between control and tannins treated animals(Figure 5) Among Bacteroidetes the most abundant genuswas Prevotella accounting for more than 40 of this phylumThe average abundance of Prevotella was lower in tannins-supplemented animals than in the control group althoughit exhibited a high degree of variance among animals (165versus 219 119901 = 015) Clostridia was the predominantclass which accounted for more than 90 of total Firmicutesand it was significantly enhanced in tannins treated animals(415 versus 346 119901 lt 0001) Among Clostridia Ruminococ-caceae was the most abundant family and showed a sig-nificantly higher abundance in the tannins-supplementedanimals (178 versus 107 119901 = 0009) In the controlgroup most sequences corresponding to family Ruminococ-caceae belonged to unclassified members (79) and genusRuminococcus (26) Both taxa were enhanced in tanninstreated steers reaching abundances of 126 (119901 = 001) and49 (119901 = 007) respectively Other nonclostridial bacteriawithin the phylum Firmicutes were significantly altered bytannins including members of class Erysipelotrichi SomeErysipelotrichi were enhanced (genus L7A-E11 119901 = 002and genus p-75-A5 119901 = 006) while others were lowered(genus RFN20 119901 = 0001) in tannins-supplemented animalsMembers of class Bacilli (genera Streptococcus and Lacto-bacillus) showed onlymoderate increases in their abundanceMeanwhile genus Fibrobacter was significantly affected bytannins accounting for 010 of total microbiota in thecontrol group and only 0005 in the tannins treated animals(119901 = 001) Other minor fibrolytic bacteria were significantlymore abundant in tannins treated steers including genusBlautia (008 versus 002 119901 = 001) and member of familyEubacteriaceae genus Anaerofustis (006 versus 002 119901 =003)
Among sugar fermenting bacteria the most abundanttaxonwas genus Prevotella whose abundance was reduced by54 in tannins treated animals as mentioned above GenusTreponema was also reduced in tannins treated steers (041versus 121 119901 = 004) Among Veillonellaceae membersgenus Succiniclasticum which specializes in fermenting suc-cinate to propionate doubled their levels in tannins treatedanimals (399 versus 199 119901 = 008) Lipolytic genusAnaerovibrio was significantly enhanced by tannins (011 ver-sus 005 119901 = 001) Genus Selenomonas was also increasedin tannins supplemented animals (011 versus 005 119901 =007) Among ureolytic bacteria genus Butyrivibrio wasthe most abundant one and it was negatively affected bytannins treatment (180 versus 236) as well as Treponemaand Succinivibrio (0009 versus 002) On the other handmethanogens belonging to phylum Euryarchaeota were lessabundant in tannins supplemented steers (137 versus 203)and their levels were inversely correlated with rumen pH (119903 =minus080) Genus Methanosphaera was significantly reduced bytannins (006 versus 016 119901 = 001)
Determination of pH urease activity and NPN wasperformed in all rumen samples along with microbiotacomposition analysis Tannins treated steers had significantlyhigher ruminal pH than the control group (630 versus 588119901 = 002) (Figure 6(a)) Urease activity exhibited the oppositepattern showing a significant decline in the tannins treatedsteers (Figure 6(b)) Moreover a strong negative correlationbetween pH and urease activity was detected (119903 = minus095)NPNwas not significantly altered by treatments (Figure 6(c))
4 Discussion
Rumen microbiome diversity is a key feature of ruminantsthat confers cattle the ability to adapt to a wide range ofdietary conditions [25] In recent years the concept of hostmicrobiome individuality in ruminants is gaining supportsince numerous studies found a large number of taxa whosepresence or abundance in the rumen varies markedly amongindividuals [26ndash28] Dietary tannins diminished ruminalrichness but did not significantly affect the bacterial com-munitiesrsquo complexity (ie balance between the relative abun-dances of taxa) A recent report found an increase in rumenrichness but no change in Shannonrsquos diversity index aftersupplementation with a blend of polyphenols essential oil indairy heifers under a high grain diet supporting the idea thatpolyphenols can affect bacterial richness without disruptingthe overall rumen microbiota population structure [29] Inline with this 120573-diversity analysis detected no significantdifferences in rumen bacterial diversity between control andtannins treated steers Low microbial richness in the rumenhas been recently found to be tightly linked to a higher feedefficiency in dairy cows [30] The authors suggest that lowerrichness in the rumen of efficient animals results in a simplermetabolic network which leads to higher concentrationsof specific components that are used to support the hostrsquosenergy requirements Together diversity analyses suggest thatbacterial richness was decreased while the overall bacterialcomplexity of the rumen was not significantly affected bychestnut and quebracho tannins supplementation
BioMed Research International 5
Control Tannins1000
1100
1200
1300
1400
1500
1600
1700
1800
1900
2000
2100
2200N
umbe
r of O
TUs
p = 005
(a)Control Tannins
6
7
8
9
10
Shan
nonrsquos
inde
x
p = 014
(b)
Figure 2 Effect of tannins treatment on (a) bacterial richness (number of OTUs) and (b) Shannonrsquos diversity index of rumen microbiomeLine = median Box = 25ndash75 percentiles Bar = 5ndash95 percentiles
minus010
minus005
000
005
010
015
PC2
(11
1)
00 01 02 03 04 05minus01PC1 (556)
Figure 3 PCoA plot based on unweighted UniFrac metric Itemsshaped with triangles and circles correspond to samples fromcontrol and tannins treated animals respectively Axes (PC1 = 556and PC2 = 111) account for 667 of the total variation detected
The dominance of phyla Firmicutes and Bacteroidetesin the bovine ruminal fluid is a common feature in thegastrointestinal microbiome of monogastric organisms andruminants [15 17 28 31] Henderson et al also found thatan increase in the ruminal abundance of total Firmicutescorrelated with a decrease in the abundance of Bacteroidetesboth in cows (119903 = minus080) and in sheep (119903 = minus097) [32] Otherauthors found that Bacteroidetes were the most abundant
Control Tannins
p = 009
04
05
06
07
08
09
10
11
12
13
14
15
16
17
Firm
icut
esB
acte
roid
etes
Figure 4 Effect of tannins on the ratio of phyla Firmicutes andBacteroidetes in rumen microbiota Line = median Box = 25ndash75percentiles Bar = 5ndash95 percentiles
phylum in ruminal samples obtained from dairy cows butsome animals exhibited a higher percentage of Firmicutescompensating for a lower abundance of Bacteroidetes [17]Our results agree with these observations since a stronginverse correlation between the abundances of Firmicutesand Bacteroidetes was detected (119903 = minus099) These resultssuggest that members of Firmicutes and Bacteroidetes com-pete for available resources in the rumen and tannins wouldtip the balance in favor of Firmicutes
6 BioMed Research International
lowast
lowastlowast
lowast
lowastlowast
lowast
lowast
lowastlowastlowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
lowast
lowastlowast
lowast
lowast
lowast
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
minus4 minus2 minus1minus3minus5 0 2 3 4 51
Phylum FirmicutesGenus Butyrivibrio
Unc Family RuminococcaceaeGenus Ruminococcus
Genus SucciniclasticumGenus Streptococcus
Genus p-75-a5Genus L7A-E11
Genus SelenomonasGenus Anaerovibrio
Genus BlautiaGenus Anaerofustis
Genus RFN20Genus Lachnospira
Phylum BacteroidetesFamily Prevotellaceae
Family BacteroidaceaeGenus Prevotella
Genus BacteroidesGenus YRC22
Unc Family RF16
Unc Family CoriobacteriaceaePhylum Actinobacteria
Phylum SpirochaetesGenus Treponema
Genus Sphaerochaeta
Phylum EuryarchaeotaGenus Methanobrevibacter
Genus Methanosphaera
Phylum ProteobacteriaUnclass Order GMD14H09
Genus RuminobacterGenus Oxalobacter
Genus Succinivibrio
Phylum TenericutesUnc Family Anaeroplasmata
Genus Anaeroplasma
Phylum ChloroflexiGenus SHD-231
Phylum FibrobacteresGenus Fibrobacter
log 2 fold change
Figure 5 Relative fold changes (log 2 tanninscontrol) in the abundance of rumen bacterial taxa between control and tannins treated steerslowastlowast119901 lt 005 lowast119901 lt 010 Bar = SEM
The ratio of Firmicutes to Bacteroidetes has been shownto affect energy harvesting and body fat accumulation inhumans and mice [14 15] Along with increased fatty acidabsorption more energy was found to be efficiently obtainedfrom diet in obese mice illustrating the connection betweenFirmicutes and improved efficiency in energy harvesting [14]In cows the Firmicutes to Bacteroidetes ratio was found tobe strongly correlated with daily milk-fat yield [17] A recentstudy found that the abundance of Firmicutes in the rumen
positively correlates with the average daily body weight gainin steers suggesting that these bacteria play a significant rolein feed efficiency of bovines [33] Therefore it is possiblethat the increase of Firmicutes to Bacteroidetes ratio inducedby tannins can improve bovine performance as previouslysuggested by other authors [19]
Fiber degradation is a complex process carried out bya group of microorganisms that are able to digest plantpolysaccharides mainly through production of cellulolytic
BioMed Research International 7
Control Tannins50
52
54
56
58
60
62
64
66
68
70pH
p = 002
(a)Control Tannins
000
005
010
015
020
Ure
ase a
ctiv
ity (U
A)
p = 0007
(b)
Control Tannins
000
005
010
015
Non
prot
ein
nitro
gen
(NPN
)
p = 0849
(c)
Figure 6 Effect of tannins on pH urease activity and NPN in the rumen liquor Line = median Box = 25ndash75 percentiles Bar = 5ndash95percentiles Urease activity values are given in pH units proportional to urease activity NPN levels are expressed as a percentage of solublenitrogen in the rumen
hemicellulolytic and pectinolytic enzymes [34] Cellulolyticactivity in the rumen involves a diverse bacterial commu-nity whose main members belong to genera Ruminococcusand Fibrobacter while the main hemicellulolytic bacteriabelong to genera Ruminococcus Prevotella and Butyrivib-rio [34ndash37] The blend of tannins administrated increasedRuminococcaceae and other members of phylum Firmicuteswhile inhibited genera Prevotella and Fibrobacter Prevotellaspecies have a documented role in metabolism of starchhemicellulose pectin and protein catabolism [25] The highabundance of this genus might be the result of a largemetabolic niche that is occupied by different species with
similar metabolic capabilities or it might be associated witha high genetic variability that enables members of genusPrevotella to occupy different ecological niches within therumen [38ndash40] For instance Prevotella abundance has beenshown to be higher in the rumen of animals of beef cattlewith low feed efficiency [41] Previous studies reported a greatdiversity in the sensitivity ofPrevotella species to tannins [42]which could explain the lower abundance detected for thisgenus in tannins-supplemented steers Since genus Prevotellais characterized by a high genetic diversity and wide array ofoutputmetabolites [38ndash40] their replacement byRuminococ-cus may lead to a simpler product profile specialized to
8 BioMed Research International
support the hostrsquos energy requirements [30] Our resultsagree with previous studies that found a significant reduc-tion in Fibrobacter and Prevotella rumen populations aftersupplementation with condensed and hydrolysable tanninsincluding chestnut and quebracho tannins [43 44]
Rumen fibrolytic function is carried out by a largelyredundant microbial community with overlapping distribu-tion of metabolic capabilities and this fibrolytic communityhas the ability to restore its structure and function afterperturbation a phenomenon known as resilience [27] In thiscontext the observed changes in diversity of fibrolytic bacte-ria in tannins treated steers may result from a combinationof physicochemical and biological mechanisms described inthe literature including direct interaction of tannins withfiber [45] which could alter the available surface area formicrobial attack the inhibition of certain fibrolytic taxa bymeans of tannins antimicrobial activity [46] andmodulationof fibrolytic bacterial species driven by changes in rumen pHsince certain fibrolytic taxa of the rumen are inhibited at lowpH under high-grain diets [36 47]
Rumen amylolytic and saccharolytic bacteria were alsoaffected by dietary treatment with tannins mainly throughthe decrease of genera Prevotella and Treponema Otheramylolytic genera were moderately increased in tannins-fedsteers including Streptococcus Bifidobacterium and Lacto-bacillus Amylolytic activity is normally enhanced in rumi-nants consuming high-grain diets [18 48 49] The rate ofgrain degradation by these microbial communities plays akey role in maintaining rumen homeostasis since rapidstarch fermentation produces large amounts of organic acidstherefore producing a drop in ruminal pH that may leadto metabolic acidosis [50] A previous study showed thattannic acid and quebracho tannins lower the rate ofmicrobialhydrolysis of starch-rich grains in the rumen by physicalmodification of the endosperm protein matrix [45] Thisphysical modulation of starch degradation could explain thelower abundance of sugar fermenting taxa detected in tanninstreated steers as well as the higher ruminal pH observed inthis group
Fermentation products of microbial activity in rumenmainly short-chain volatile fatty acids serve as amajor sourceof energy for ruminants and have a direct impact on thephysiological parameters of the animal and feed utilizationefficiency [16] Some members of family Veillonellaceaewhich produce propionate as a major fermentation productand have been associated with lower methane emissions [51]were enhanced in tannins treated animals Interestingly someSelenomonas species can break tannin-protein complexes anduse tannins as energy source [52 53] Therefore the higherabundance of Selenomonas in tannins treated steers may bepartly due to availability of tannins as direct energy sourceMeanwhile members of class Erysipelotrichi which havebeen linked to beef cattle feed efficiency [33 51 54] were alsomodulated by tannins treatment
Methane production during fermentation of feeds in therumen represents a loss of 2ndash12 of gross energy [1] andit is performed by a group of archaea known collectivelyas methanogens which belong to phylum EuryarchaeotaThe microorganisms produce methane the second largest
anthropogenic greenhouse gas which has a global warmingpotential 25 times that of carbon dioxide [55] In the presentstudy a reduction of methanogenic archaea in tannins sup-plemented steers was detected as well as an inverse cor-relation between the abundances of phylum Euryarchaeotaand rumen pH (119903 = minus095) Tannins are thought todirectly inhibit methanogens as well as indirectly limitmethanogenesis through reduction of hydrogen availability[1 55] Saminathan et al found a significant decrease in genusMethanobrevibacter after treatment of rumen samples withcondensed tannins in vitro [56] Other authors describeda linear decrease of genus Methanobrevibacter after dietarysupplementation with condensed tannins from pine bark ingoats [31]
Ruminal pH was significantly higher in tannins treatedsteers than in the control group while urease activity exhib-ited the opposite pattern These results agree with previousreports which found an increase in ruminal pH after sup-plementation with chestnut and quebracho tannins [57 58]Other authors observed that feeding chestnut and quebrachotannins decreased urease activity in the faeces of cows[10 11]
Feed-grade urea is an effective source of nitrogen com-monly used in beef cattle diets Ureolytic bacteria in therumen produce urease to hydrolyse urea to ammonia whichis subsequently used for the synthesis of amino acids andmicrobial protein Normally the rate of urea hydrolysisexceeds the rate of ammonia utilization which leads to poorefficiency of urea utilization and increases toxic ammoniaconcentrations in blood [59] Ureolytic bacteria in the rumencomprise a highly diverse group whose main species belongto genera Succinivibrio Treponema Bacteroides ButyrivibrioStreptococcus andBifidobacterium [60 61]We found that themost abundant ureolytic generaButyrivibrio andTreponemawere negatively affected by tannins treatment Thus theobserved decline in ruminal urease activity may be relatedwith the decrease of these urease-producing taxa A previousstudy also observed a drop in ruminal urease activity afteraddition of tannins to diet but a direct interaction betweentannins and urease enzyme was postulated as responsible forthis inhibition [62]
5 Conclusions
The current study showed that chestnut and quebrachotannins added to the diet of Holstein steers modifiedrumen microbiota composition particularly fiber and starchdegrading bacteria mainly by reducing the abundanceof Prevotella and Fibrobacter while favoring Ruminococ-caceae and other members of phylum Firmicutes Tanninstreatment significantly increased pH and decreased ureaseactivity in the ruminal liquor Further work is necessaryto assess the possible relation between tannins ability tomodify rumen microbiota composition and the alterationof rumen fermentation parameters associated with energyand feed efficiency of beef cattle such as the profile ofshort-chain fatty acids and the emissions of ammonia andmethane
BioMed Research International 9
Conflicts of Interest
The authors declare that there are no known conflicts ofinterest associated with this publication and there has beenno financial support for this work that could have influencedits outcome
Acknowledgments
The authors thank Alfredo Ivan Martınez Caceres for histechnical assistance during installation and startup of thebioinformatics tools used in this study This work wassupported by grants from INTA (PNSA 1115052 and PNBIO1131043) Additional support was provided by CONICET
References
[1] A K Patra and J Saxena ldquoDietary phytochemicals as rumenmodifiers a review of the effects on microbial populationsrdquoInternational Journal of General and Molecular Microbiologyvol 96 no 4 pp 363ndash375 2009 Antonie Van Leeuwenhoek
[2] R Bodas N Prieto R Garcıa-Gonzalez S Andres F JGiraldez and S Lopez ldquoManipulation of rumen fermentationand methane production with plant secondary metabolitesrdquoAnimal Feed Science and Technology vol 176 no 1-4 pp 78ndash932012
[3] E Oskoueian N Abdullah and A Oskoueian ldquoEffects offlavonoids on rumen fermentation activity methane produc-tion andmicrobial populationrdquo BioMed Research Internationalvol 2013 Article ID 349129 8 pages 2013
[4] P Frutos GHervas F J Giraldez andA RMantecon ldquoReviewtannins and ruminant nutritionrdquo Spanish Journal of AgriculturalResearch vol 2 no 2 pp 191ndash202 2004
[5] K Khanbabaee and T van Ree ldquoTannins classification anddefinitionrdquo Natural Product Reports vol 18 no 6 pp 641ndash6492001
[6] L J Porter ldquoStructure and chemical properties of the con-densed tanninsrdquo Plant Polyphenols pp 245ndash258 1992 Springer Boston MA USA
[7] B R Min C Wright P Ho J-S Eun N Gurung and RShange ldquoThe effect of phytochemical tannins-containing dieton rumen fermentation characteristics and microbial diversitydynamics in goats using 16S rDNA amplicon pyrosequencingrdquoAgriculture Food and Analytical Bacteriology vol 4 no 3 pp195ndash211 2014
[8] B S Gemeda and A Hassen ldquoEffect of tannin and speciesvariation on in vitro digestibility gas and methane productionof tropical browse plantsrdquoAsian-Australasian Journal of AnimalSciences vol 28 no 2 pp 188ndash199 2015
[9] Z Li A-D G Wright H Liu et al ldquoResponse of the rumenmicrobiota of sika deer (cervus nippon) fed different concen-trations of tannin rich plantsrdquo PLoS ONE vol 10 no 5 ArticleID e0123481 2015
[10] JM PowelM J Aguerre andMAWattiaux ldquoTannin extractsabate ammonia emissions from simulated dairy barn floorsrdquoJournal of Environmental Quality vol 40 no 3 pp 907ndash9142011
[11] M J Aguerre M C Capozzolo P Lencioni C Cabral andM A Wattiaux ldquoEffect of quebracho-chestnut tannin extractsat 2 dietary crude protein levels on performance rumenfermentation and nitrogen partitioning in dairy cowsrdquo Journal
of Dairy Science vol 99 no 6 Article ID 74473 pp 4476ndash44862016
[12] C Rivera-Mendez A Plascencia N Torrentera andR A ZinnldquoEffect of level and source of supplemental tannin on growthperformance of steers during the late finishing phaserdquo Journalof Applied Animal Research vol 45 pp 199ndash203 2017
[13] B R Min T N Barry G T Attwood and W C McNabbldquoThe effect of condensed tannins on the nutrition and health ofruminants fed fresh temperate forages a reviewrdquo Animal FeedScience and Technology vol 106 no 1-4 pp 3ndash19 2003
[14] P J Turnbaugh R E Ley M A Mahowald V MagriniE R Mardis and J I Gordon ldquoAn obesity-associated gutmicrobiomewith increased capacity for energy harvestrdquoNaturevol 444 no 7122 pp 1027ndash1031 2006
[15] R E Ley R Knight and J I Gordon ldquoThe human micro-biome Eliminating the biomedicalenvironmental dichotomyin microbial ecologyrdquo Environmental Microbiology vol 9 no 1pp 3-4 2007
[16] I Mizrahi ldquoThe role of the rumen microbiota in determiningthe feed efficiency of dairy cowsrdquo Beneficial Microorganismsin Multicellular Life Forms pp 203ndash210 2012 Springer BerlinHeidelberg
[17] E Jami B AWhite and IMizrahi ldquoPotential role of the bovinerumen microbiome in modulating milk composition and feedefficiencyrdquo PLoS ONE vol 9 no 1 Article ID e85423 2014
[18] S C Fernando H T Purvis F Z Najar et al ldquoRumenmicrobialpopulation dynamics during adaptation to a high-grain dietrdquoApplied and Environmental Microbiology vol 76 no 22 pp7482ndash7490 2010
[19] H P S Makkar ldquoEffects and fate of tannins in ruminantanimals adaptation to tannins and strategies to overcomedetrimental effects of feeding tannin-rich feedsrdquo Small Rumi-nant Research vol 49 no 3 pp 241ndash256 2003
[20] Illumina Protocol 16S Metagenomic Sequencing Library 2013[21] AM BolgerM Lohse and B Usadel ldquoTrimmomatic a flexible
trimmer for Illumina sequence datardquoBioinformatics vol 30 no15 pp 2114ndash2120 2014
[22] J Kuczynski J Stombaugh W A Walters A Gonzalez J GCaporaso and R Knight ldquoUsing QIIME to analyze 16S rRNAgene sequences frommicrobial communitiesrdquoCurrent Protocolsin Microbiology vol 1 1E5 pages 2012
[23] N A Bokulich S Subramanian J J Faith et al ldquoQuality-filtering vastly improves diversity estimates from Illuminaamplicon sequencingrdquo Nature Methods vol 10 no 1 pp 57ndash592013
[24] D Parks and R Beiko ldquoSTAMP statistical analysis of metage-nomic profilesrdquo Encyclopedia of Metagenomics pp 1ndash6 2013Springer New York NY USA
[25] J C McCann T A Wickersham and J J Loor ldquoHigh-throughput methods redefine the rumen microbiome and itsrelationshipwith nutrition andmetabolismrdquoBioinformatics andBiology Insights vol 8 pp 109ndash125 2014
[26] G Henderson F Cox S Ganesh A Jonker W Young and PH Janssen ldquoRumen microbial community composition varieswith diet and host but a coremicrobiome is found across a widegeographical rangerdquo Scientific Reports vol 5 no 14567 pp 1ndash132015
[27] P J Weimer ldquoRedundancy resilience and host specificity ofthe ruminal microbiota Implications for engineering improvedruminal fermentationsrdquo Frontiers in Microbiology vol 6 articleno 296 2015
10 BioMed Research International
[28] E Jami and I Mizrahi ldquoComposition and similarity of bovinerumenmicrobiota across individual animalsrdquo PLoS ONE vol 7no 3 Article ID e33306 2012
[29] R De Nardi G Marchesini S Li et al ldquoMetagenomic analysisof rumen microbial population in dairy heifers fed a high graindiet supplemented with dicarboxylic acids or polyphenolsrdquoBMC Veterinary Research vol 12 no 1 article no 29 2016
[30] S Kruger Ben Shabat G Sasson A Doron-Faigenboim etal ldquoSpecific microbiome-dependent mechanisms underlie theenergy harvest efficiency of ruminantsrdquoThe ISME Journal vol10 no 12 pp 2958ndash2972 2016
[31] B R Min S Solaiman R Shange and J-S Eun ldquoGastroin-testinal bacterial andmethanogenic archaea diversity dynamicsassociated with condensed tannin-containing pine bark diet ingoats using 16S rDNA amplicon pyrosequencingrdquo InternationalJournal of Microbiology vol 2014 Article ID 141909 2014
[32] G Henderson F Cox S Kittelmann et al ldquoEffect of DNAextraction methods and sampling techniques on the apparentstructure of cow and sheep rumen microbial communitiesrdquoPLoS ONE vol 8 no 9 Article ID e74787 2013
[33] P R Myer T P L Smith J E Wells L A Kuehn and HC Freetly ldquoRumen microbiome from steers differing in feedefficiencyrdquo PLoS ONE vol 10 no 6 Article ID e0129174 2015
[34] Y Wang and T A McAllister ldquoRumen microbes enzymes andfeed digestion-a reviewrdquo Asian-Australasian Journal of AnimalSciences vol 15 no 11 pp 1659ndash1676 2002
[35] J B Russell R EMuck and P JWeimer ldquoQuantitative analysisof cellulose degradation and growth of cellulolytic bacteria inthe rumenrdquo FEMS Microbiology Ecology vol 67 no 2 pp 183ndash197 2009
[36] DOKrause S EDenmanR IMackie et al ldquoOpportunities toimprove fiber degradation in the rumen microbiology ecologyand genomicsrdquo FEMS Microbiology Reviews vol 27 no 5 pp663ndash693 2003
[37] M Zorec M Vodovnik and R Marinsek-Logar ldquoPotentialof selected rumen bacteria for cellulose and hemicellulosedegradationrdquo Food Technology and Biotechnology vol 52 no 2pp 210ndash221 2014
[38] A Z Bekele S Koike and Y Kobayashi ldquoGenetic diversityand diet specificity of ruminal prevotella revealed by 16S rRNAgene-based analysisrdquo FEMSMicrobiology Letters vol 305 no 1pp 49ndash57 2010
[39] A RamsakM Peterka K Tajima et al ldquoUnravelling the geneticdiversity of ruminal bacteria belonging to the CFB phylumrdquoFEMS Microbiology Ecology vol 33 no 1 pp 69ndash79 2000
[40] J Purushe D E Fouts M Morrison et al ldquoComparativegenome analysis of prevotella ruminicola and prevotella bryan-tii insights into their environmental nicherdquo Microbial Ecologyvol 60 no 4 pp 721ndash729 2010
[41] C A Carberry D A Kenny S Han M S McCabe and S MWaters ldquoEffect of phenotypic residual feed intake and dietaryforage content on the rumen microbial community of beefcattlerdquo Applied and Environmental Microbiology vol 78 no 14pp 4949ndash4958 2012
[42] A L Molan G T Attwood B R Min and W C McNabbldquoThe effect of condensed tannins from Lotus pedunculatus andLotus corniculatus on the growth of proteolytic rumen bacteriain vitro and their possible mode of actionrdquo Canadian Journal ofMicrobiology vol 47 no 7 pp 626ndash633 2001
[43] A Jayanegara G Goel H P S Makkar and K Becker ldquoDiver-gence between purified hydrolysable and condensed tannin
effects on methane emission rumen fermentation and micro-bial population in vitrordquo Animal Feed Science and Technologyvol 209 pp 60ndash68 2015
[44] BMinW Pinchak K Hernandez et al ldquoEffects of plant tanninsupplementation on animal responses and in vivo ruminalbacterial populations associated with bloat in heifers grazingwheat forage1rdquo The Professional Animal Scientist vol 28 no 4pp 464ndash472 2012
[45] T F Martınez T A McAllister Y Wang and T Reuter ldquoEffectsof tannic acid and quebracho tannins on in vitro ruminalfermentation of wheat and corn grainrdquo Journal of the Scienceof Food and Agriculture vol 86 no 8 pp 1244ndash1256 2006
[46] L Marın E M Miguelez C J Villar and F Lombo ldquoBioavail-ability of dietary polyphenols and gut microbiota metabolismantimicrobial propertiesrdquo BioMed Research International vol2015 Article ID 905215 18 pages 2015
[47] J B Russell and D B Wilson ldquoWhy are ruminal cellulolyticbacteria unable to digest cellulose at low pHrdquo Journal of DairyScience vol 79 no 8 pp 1503ndash1509 1996
[48] K Tajima S Arai K Ogata et al ldquoRumen bacterial communitytransition during adaptation to high-grain dietrdquo Anaerobe vol6 no 5 pp 273ndash284 2000
[49] D W Goad C L Goad and T G Nagaraja ldquoRuminal micro-bial and fermentative changes associated with experimentallyinduced subacute acidosis in steersrdquo Journal of Animal Sciencevol 76 no 1 pp 234ndash241 1998
[50] J Hernandez J L Benedito A Abuelo and C CastilloldquoRuminal acidosis in feedlot from aetiology to preventionrdquoTheScientific World Journal vol 2014 Article ID 702572 2014
[51] A B De Menezes E Lewis M OrsquoDonovan B F OrsquoNeill NClipson and E M Doyle ldquoMicrobiome analysis of dairy cowsfed pasture or total mixed ration dietsrdquo FEMS MicrobiologyEcology vol 78 no 2 pp 256ndash265 2011
[52] I K Skene and J D Brooker ldquoCharacterization of tanninacylhydrolase activity in the ruminal bacterium selenomonasruminantiumrdquo Anaerobe vol 1 no 6 pp 321ndash327 1995
[53] G Goel A K Puniya and K Singh ldquoTannic acid resistance inruminal streptococcal isolatesrdquo Journal of Basic Microbiologyvol 45 no 3 pp 243ndash245 2005
[54] P R Myer J E Wells T P L Smith L A Kuehn and H CFreetly ldquoCecum microbial communities from steers differingin feed efficiencyrdquo Journal of Animal Science vol 93 no 11 pp5327ndash5340 2015
[55] S E Hook A-D GWright and BWMcBride ldquoMethanogensmethane producers of the rumen and mitigation strategiesrdquoArchaea vol 2010 Article ID 945785 11 pages 2010
[56] M Saminathan C C Sieo H M Gan N Abdullah C M VL Wong and Y W Ho ldquoEffects of condensed tannin fractionsof different molecular weights on population and diversity ofbovine rumen methanogenic archaea in vitro as determinedby high-throughput sequencingrdquo Animal Feed Science andTechnology vol 216 pp 146ndash160 2016
[57] F Hassanat and C Benchaar ldquoAssessment of the effect ofcondensed (acacia and quebracho) and hydrolysable (chestnutand valonea) tannins on rumen fermentation andmethane pro-duction in vitrordquo Journal of the Science of Food and Agriculturevol 93 no 2 pp 332ndash339 2013
[58] J Balcells A Aris A Serrano A R Seradj J Crespo and MDevant ldquoEffects of an extract of plant flavonoids (bioflavex)on rumen fermentation and performance in heifers fed high-concentrate dietsrdquo Journal of Animal Science vol 90 no 13 pp4975ndash4984 2012
BioMed Research International 11
[59] A K Patra ldquoUreaammonia metabolism in the rumen andtoxicity in ruminantsrdquo Rumen Microbiology From Evolution toRevolution pp 329ndash341 2015
[60] MAWoznyM P Bryant L VHoldeman andW E CMooreldquoUrease assay and urease producing species of anaerobes in thebovine rumen and human fecesrdquo Applied and EnvironmentalMicrobiology vol 33 no 5 pp 1097ndash1104 1977
[61] D Jin S Zhao P Wang et al ldquoInsights into abundant rumenureolytic bacterial community using rumen simulation systemrdquoFrontiers in Microbiology vol 7 article no 1006 2016
[62] H P SMakkar B Singh andRKDawra ldquoEffect of tannin-richleaves of oak (Quercus incana) on various microbial enzymeactivities of the bovine rumenrdquo British Journal of Nutrition vol60 no 2 pp 287ndash296 1988
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 3
and stored at minus80∘C until further processing Part of thesample was used for determination of ruminal physiologicalparameters The ruminal liquor pH was measured usinga standard pH meter Urease activity (UA) was measuredaccording to the CaskeyminusKnappmethodmodified by AACCNonprotein nitrogen (NPN) was determined through Kjel-dahl method (VELP Scientifica Italy) Animals were caredfor by trained personnel only and the experimental protocoland procedures used were conducted according to protocol272011 of the Institutional Committee for the Care and Useof Experimental Animals (CICVyA-INTA)
22 DNA Extraction Twenty milliliters of evenly homoge-nized ruminal liquor was lyophilized before DNA extractionin order tomaximizemicrobial density (1 g of drymaterial persample) DNA extraction was conducted using the QIAampDNA stool kit (Qiagen Hilden Germany) following theinstructions of the manufacturer with slight modificationsBriefly 100mg of lyophilized rumen was weighed and lysedby incubation for 5min at 95∘C DNA elution was donewith 100 120583l of Buffer AE after incubation for 10min atroom temperature DNA quality was assessed by agarosegel electrophoresis and DNA concentration was determinedusing a NanoDrop ND-1000 spectrophotometer (NanoDropTechnologies Wilmington DE USA) DNA was kept atminus20∘C until further processing
23 High-Throughput Sequencing of 16S rRNA Gene The16S rRNA gene V3-V4 regions were amplified using Illu-mina primers (forward 51015840 CCTACGGGNGGCWGCAG 31015840reverse 51015840 GGACTACHVGGGTATCTAATCC 31015840) with stan-dard adapter sequences attached for barcoding and multi-plexing 16S gene libraries construction and high-throughputsequencing were performed at Macrogen Inc (Seoul SouthKorea) in the Illumina MiSeq platform following manu-facturerrsquos instructions [20] In order to reduce unbalancedand biased base compositions 15 of PhiX control librarywas spiked into the amplicon pool Due to an issue withthe length of reverse reads which were not long enough toachievemerging of paired-end sequences only forward readscovering V3 and its flanking regions were used for furtherbioinformatics analysis The datasets generated in this studyare available under request
24 Bioinformatics Analysis FASTQ files were trimmedusing Trimmomatic v033 [21] which removed all primerand adapter sequences and also removed leading and trailingbases if quality value was below 9 and 3 respectivelySliding window trimming was also performed as well ascutting if the average quality within a 4-base window fallsbelow Q15 Demultiplexing and quality filtering were doneusing the script split libraries fastqpy which is part of theQIIME v191 software package [22] A threshold of Phredquality score (119876 gt 20) of the base was chosen forstringent quality control processing Chimeric sequenceswere filtered out in QIIME using the USERCH algorithmOpen-reference operational taxonomic units (OTUs) pickingwas performed using UCLUST and USEARCH algorithmsin QIIME Each sequence was assigned taxonomy against
Greengenes reference OTU build version 138 using a 97sequence similarity threshold OTUs with abundance below0005 were removed from final OTU table in order toavoid microbial diversity overestimation [23] Normalizationof OTU counts was done by performingmultiple rarefactionsfrom 10000 to 210000 sequences with steps of 10000 andwith 10 repetitions at each rarefaction depth The resultingmultiple rarefied OTU tables were used for all furtheranalysis Principal coordinate analysis (PCoA) plots weregenerated in QIIME with default options using a distancematrix calculated by unweighted UniFrac metric The signif-icance of grouping in the PCoA plot was tested by analysis ofsimilarity (ANOSIM) in QIIME with 999 permutationsThiswork used computational resources from the BioinformaticsUnit Instituto de Biotecnologıa (CICVyA-INTA)
25 Statistical Analysis Relative abundances of bacterialpopulations were statistically analyzed using STAMP v213[24] The relative abundances of bacterial taxa in controland tannins treated groups were compared at each levelof classification (phylum class order family and genus)usingWhitersquos nonparametric two-tailed 119905-test with 1000 per-mutations Comparisons in physiological data and diversityestimators were calculated using nonparametric two-tailedMannminusWhitney test (GraphPad Software San Diego CAUSA) and were considered statistically significant if 119901 lt 005
3 Results
After high-throughput sequencing 3812179 reads wereobtained with an average of 346562 plusmn 42326 reads persample Stringent filtration of the sequences based on lengthand quality was performed before taxonomy assignationresulting in 2951356 reads with an average length of 267 plusmn22 base pairs
The total number of OTUs detected after filtration was2263 but this number exhibited a high interindividualvariation (Figure 1) We found that the number of OTUstended to be lower in tannins-supplemented animals thanin the control group (119901 = 005) (Figure 2(a)) Shannonrsquosdiversity index which estimates the internal sample com-plexity was not significantly affected by tannins (119901 = 014)(Figure 2(b)) PCoA based on unweighted UniFrac metricwas performed in order to explore dissimilarities inmicrobialcomposition of the rumen among treated groups (Figure 3)ANOSIM detected no significant differences in bacterialdiversity between control and tannins dietary treatments (119901 =086)
Firmicutes andBacteroideteswere the dominant bacterialphyla in the bovine ruminal fluid accounting for nearly 90of total microbiota However large interindividual variancewas observed in the relative abundance of Bacteroidetes andFirmicutes among different animals In the control groupbacterial populations belonging to phylum Bacteroideteswere the most abundant in all animals (521 on average)while Firmicutes accounted for 376 of total microbiotaHowever this predominance was inverted in the tanninstreated animals with a significantly higher percentage ofFirmicutes (462 119901 = 002) and a reduction to 446 in
4 BioMed Research International
0
500
1000
1500
2000
2500
OTU
s
100000 150000 20000050000Sequences per sample
Figure 1 OTUs rarefaction curves of rumen microbiota based on16S rRNA gene sequences OTUs were picked using the UCLUSTmethod with 3 dissimilarity in QIIME Each curve corresponds toa single ruminal sample
Bacteroidetes (119901 = 018) Accordingly steers supplementedwith tannins presented a trend to higher Firmicutes toBacteroidetes ratio in comparison with the control group(108 versus 073 119901 = 009) (Figure 4)
Significant differences in the abundance of certain taxawere detected between control and tannins treated animals(Figure 5) Among Bacteroidetes the most abundant genuswas Prevotella accounting for more than 40 of this phylumThe average abundance of Prevotella was lower in tannins-supplemented animals than in the control group althoughit exhibited a high degree of variance among animals (165versus 219 119901 = 015) Clostridia was the predominantclass which accounted for more than 90 of total Firmicutesand it was significantly enhanced in tannins treated animals(415 versus 346 119901 lt 0001) Among Clostridia Ruminococ-caceae was the most abundant family and showed a sig-nificantly higher abundance in the tannins-supplementedanimals (178 versus 107 119901 = 0009) In the controlgroup most sequences corresponding to family Ruminococ-caceae belonged to unclassified members (79) and genusRuminococcus (26) Both taxa were enhanced in tanninstreated steers reaching abundances of 126 (119901 = 001) and49 (119901 = 007) respectively Other nonclostridial bacteriawithin the phylum Firmicutes were significantly altered bytannins including members of class Erysipelotrichi SomeErysipelotrichi were enhanced (genus L7A-E11 119901 = 002and genus p-75-A5 119901 = 006) while others were lowered(genus RFN20 119901 = 0001) in tannins-supplemented animalsMembers of class Bacilli (genera Streptococcus and Lacto-bacillus) showed onlymoderate increases in their abundanceMeanwhile genus Fibrobacter was significantly affected bytannins accounting for 010 of total microbiota in thecontrol group and only 0005 in the tannins treated animals(119901 = 001) Other minor fibrolytic bacteria were significantlymore abundant in tannins treated steers including genusBlautia (008 versus 002 119901 = 001) and member of familyEubacteriaceae genus Anaerofustis (006 versus 002 119901 =003)
Among sugar fermenting bacteria the most abundanttaxonwas genus Prevotella whose abundance was reduced by54 in tannins treated animals as mentioned above GenusTreponema was also reduced in tannins treated steers (041versus 121 119901 = 004) Among Veillonellaceae membersgenus Succiniclasticum which specializes in fermenting suc-cinate to propionate doubled their levels in tannins treatedanimals (399 versus 199 119901 = 008) Lipolytic genusAnaerovibrio was significantly enhanced by tannins (011 ver-sus 005 119901 = 001) Genus Selenomonas was also increasedin tannins supplemented animals (011 versus 005 119901 =007) Among ureolytic bacteria genus Butyrivibrio wasthe most abundant one and it was negatively affected bytannins treatment (180 versus 236) as well as Treponemaand Succinivibrio (0009 versus 002) On the other handmethanogens belonging to phylum Euryarchaeota were lessabundant in tannins supplemented steers (137 versus 203)and their levels were inversely correlated with rumen pH (119903 =minus080) Genus Methanosphaera was significantly reduced bytannins (006 versus 016 119901 = 001)
Determination of pH urease activity and NPN wasperformed in all rumen samples along with microbiotacomposition analysis Tannins treated steers had significantlyhigher ruminal pH than the control group (630 versus 588119901 = 002) (Figure 6(a)) Urease activity exhibited the oppositepattern showing a significant decline in the tannins treatedsteers (Figure 6(b)) Moreover a strong negative correlationbetween pH and urease activity was detected (119903 = minus095)NPNwas not significantly altered by treatments (Figure 6(c))
4 Discussion
Rumen microbiome diversity is a key feature of ruminantsthat confers cattle the ability to adapt to a wide range ofdietary conditions [25] In recent years the concept of hostmicrobiome individuality in ruminants is gaining supportsince numerous studies found a large number of taxa whosepresence or abundance in the rumen varies markedly amongindividuals [26ndash28] Dietary tannins diminished ruminalrichness but did not significantly affect the bacterial com-munitiesrsquo complexity (ie balance between the relative abun-dances of taxa) A recent report found an increase in rumenrichness but no change in Shannonrsquos diversity index aftersupplementation with a blend of polyphenols essential oil indairy heifers under a high grain diet supporting the idea thatpolyphenols can affect bacterial richness without disruptingthe overall rumen microbiota population structure [29] Inline with this 120573-diversity analysis detected no significantdifferences in rumen bacterial diversity between control andtannins treated steers Low microbial richness in the rumenhas been recently found to be tightly linked to a higher feedefficiency in dairy cows [30] The authors suggest that lowerrichness in the rumen of efficient animals results in a simplermetabolic network which leads to higher concentrationsof specific components that are used to support the hostrsquosenergy requirements Together diversity analyses suggest thatbacterial richness was decreased while the overall bacterialcomplexity of the rumen was not significantly affected bychestnut and quebracho tannins supplementation
BioMed Research International 5
Control Tannins1000
1100
1200
1300
1400
1500
1600
1700
1800
1900
2000
2100
2200N
umbe
r of O
TUs
p = 005
(a)Control Tannins
6
7
8
9
10
Shan
nonrsquos
inde
x
p = 014
(b)
Figure 2 Effect of tannins treatment on (a) bacterial richness (number of OTUs) and (b) Shannonrsquos diversity index of rumen microbiomeLine = median Box = 25ndash75 percentiles Bar = 5ndash95 percentiles
minus010
minus005
000
005
010
015
PC2
(11
1)
00 01 02 03 04 05minus01PC1 (556)
Figure 3 PCoA plot based on unweighted UniFrac metric Itemsshaped with triangles and circles correspond to samples fromcontrol and tannins treated animals respectively Axes (PC1 = 556and PC2 = 111) account for 667 of the total variation detected
The dominance of phyla Firmicutes and Bacteroidetesin the bovine ruminal fluid is a common feature in thegastrointestinal microbiome of monogastric organisms andruminants [15 17 28 31] Henderson et al also found thatan increase in the ruminal abundance of total Firmicutescorrelated with a decrease in the abundance of Bacteroidetesboth in cows (119903 = minus080) and in sheep (119903 = minus097) [32] Otherauthors found that Bacteroidetes were the most abundant
Control Tannins
p = 009
04
05
06
07
08
09
10
11
12
13
14
15
16
17
Firm
icut
esB
acte
roid
etes
Figure 4 Effect of tannins on the ratio of phyla Firmicutes andBacteroidetes in rumen microbiota Line = median Box = 25ndash75percentiles Bar = 5ndash95 percentiles
phylum in ruminal samples obtained from dairy cows butsome animals exhibited a higher percentage of Firmicutescompensating for a lower abundance of Bacteroidetes [17]Our results agree with these observations since a stronginverse correlation between the abundances of Firmicutesand Bacteroidetes was detected (119903 = minus099) These resultssuggest that members of Firmicutes and Bacteroidetes com-pete for available resources in the rumen and tannins wouldtip the balance in favor of Firmicutes
6 BioMed Research International
lowast
lowastlowast
lowast
lowastlowast
lowast
lowast
lowastlowastlowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
lowast
lowastlowast
lowast
lowast
lowast
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
minus4 minus2 minus1minus3minus5 0 2 3 4 51
Phylum FirmicutesGenus Butyrivibrio
Unc Family RuminococcaceaeGenus Ruminococcus
Genus SucciniclasticumGenus Streptococcus
Genus p-75-a5Genus L7A-E11
Genus SelenomonasGenus Anaerovibrio
Genus BlautiaGenus Anaerofustis
Genus RFN20Genus Lachnospira
Phylum BacteroidetesFamily Prevotellaceae
Family BacteroidaceaeGenus Prevotella
Genus BacteroidesGenus YRC22
Unc Family RF16
Unc Family CoriobacteriaceaePhylum Actinobacteria
Phylum SpirochaetesGenus Treponema
Genus Sphaerochaeta
Phylum EuryarchaeotaGenus Methanobrevibacter
Genus Methanosphaera
Phylum ProteobacteriaUnclass Order GMD14H09
Genus RuminobacterGenus Oxalobacter
Genus Succinivibrio
Phylum TenericutesUnc Family Anaeroplasmata
Genus Anaeroplasma
Phylum ChloroflexiGenus SHD-231
Phylum FibrobacteresGenus Fibrobacter
log 2 fold change
Figure 5 Relative fold changes (log 2 tanninscontrol) in the abundance of rumen bacterial taxa between control and tannins treated steerslowastlowast119901 lt 005 lowast119901 lt 010 Bar = SEM
The ratio of Firmicutes to Bacteroidetes has been shownto affect energy harvesting and body fat accumulation inhumans and mice [14 15] Along with increased fatty acidabsorption more energy was found to be efficiently obtainedfrom diet in obese mice illustrating the connection betweenFirmicutes and improved efficiency in energy harvesting [14]In cows the Firmicutes to Bacteroidetes ratio was found tobe strongly correlated with daily milk-fat yield [17] A recentstudy found that the abundance of Firmicutes in the rumen
positively correlates with the average daily body weight gainin steers suggesting that these bacteria play a significant rolein feed efficiency of bovines [33] Therefore it is possiblethat the increase of Firmicutes to Bacteroidetes ratio inducedby tannins can improve bovine performance as previouslysuggested by other authors [19]
Fiber degradation is a complex process carried out bya group of microorganisms that are able to digest plantpolysaccharides mainly through production of cellulolytic
BioMed Research International 7
Control Tannins50
52
54
56
58
60
62
64
66
68
70pH
p = 002
(a)Control Tannins
000
005
010
015
020
Ure
ase a
ctiv
ity (U
A)
p = 0007
(b)
Control Tannins
000
005
010
015
Non
prot
ein
nitro
gen
(NPN
)
p = 0849
(c)
Figure 6 Effect of tannins on pH urease activity and NPN in the rumen liquor Line = median Box = 25ndash75 percentiles Bar = 5ndash95percentiles Urease activity values are given in pH units proportional to urease activity NPN levels are expressed as a percentage of solublenitrogen in the rumen
hemicellulolytic and pectinolytic enzymes [34] Cellulolyticactivity in the rumen involves a diverse bacterial commu-nity whose main members belong to genera Ruminococcusand Fibrobacter while the main hemicellulolytic bacteriabelong to genera Ruminococcus Prevotella and Butyrivib-rio [34ndash37] The blend of tannins administrated increasedRuminococcaceae and other members of phylum Firmicuteswhile inhibited genera Prevotella and Fibrobacter Prevotellaspecies have a documented role in metabolism of starchhemicellulose pectin and protein catabolism [25] The highabundance of this genus might be the result of a largemetabolic niche that is occupied by different species with
similar metabolic capabilities or it might be associated witha high genetic variability that enables members of genusPrevotella to occupy different ecological niches within therumen [38ndash40] For instance Prevotella abundance has beenshown to be higher in the rumen of animals of beef cattlewith low feed efficiency [41] Previous studies reported a greatdiversity in the sensitivity ofPrevotella species to tannins [42]which could explain the lower abundance detected for thisgenus in tannins-supplemented steers Since genus Prevotellais characterized by a high genetic diversity and wide array ofoutputmetabolites [38ndash40] their replacement byRuminococ-cus may lead to a simpler product profile specialized to
8 BioMed Research International
support the hostrsquos energy requirements [30] Our resultsagree with previous studies that found a significant reduc-tion in Fibrobacter and Prevotella rumen populations aftersupplementation with condensed and hydrolysable tanninsincluding chestnut and quebracho tannins [43 44]
Rumen fibrolytic function is carried out by a largelyredundant microbial community with overlapping distribu-tion of metabolic capabilities and this fibrolytic communityhas the ability to restore its structure and function afterperturbation a phenomenon known as resilience [27] In thiscontext the observed changes in diversity of fibrolytic bacte-ria in tannins treated steers may result from a combinationof physicochemical and biological mechanisms described inthe literature including direct interaction of tannins withfiber [45] which could alter the available surface area formicrobial attack the inhibition of certain fibrolytic taxa bymeans of tannins antimicrobial activity [46] andmodulationof fibrolytic bacterial species driven by changes in rumen pHsince certain fibrolytic taxa of the rumen are inhibited at lowpH under high-grain diets [36 47]
Rumen amylolytic and saccharolytic bacteria were alsoaffected by dietary treatment with tannins mainly throughthe decrease of genera Prevotella and Treponema Otheramylolytic genera were moderately increased in tannins-fedsteers including Streptococcus Bifidobacterium and Lacto-bacillus Amylolytic activity is normally enhanced in rumi-nants consuming high-grain diets [18 48 49] The rate ofgrain degradation by these microbial communities plays akey role in maintaining rumen homeostasis since rapidstarch fermentation produces large amounts of organic acidstherefore producing a drop in ruminal pH that may leadto metabolic acidosis [50] A previous study showed thattannic acid and quebracho tannins lower the rate ofmicrobialhydrolysis of starch-rich grains in the rumen by physicalmodification of the endosperm protein matrix [45] Thisphysical modulation of starch degradation could explain thelower abundance of sugar fermenting taxa detected in tanninstreated steers as well as the higher ruminal pH observed inthis group
Fermentation products of microbial activity in rumenmainly short-chain volatile fatty acids serve as amajor sourceof energy for ruminants and have a direct impact on thephysiological parameters of the animal and feed utilizationefficiency [16] Some members of family Veillonellaceaewhich produce propionate as a major fermentation productand have been associated with lower methane emissions [51]were enhanced in tannins treated animals Interestingly someSelenomonas species can break tannin-protein complexes anduse tannins as energy source [52 53] Therefore the higherabundance of Selenomonas in tannins treated steers may bepartly due to availability of tannins as direct energy sourceMeanwhile members of class Erysipelotrichi which havebeen linked to beef cattle feed efficiency [33 51 54] were alsomodulated by tannins treatment
Methane production during fermentation of feeds in therumen represents a loss of 2ndash12 of gross energy [1] andit is performed by a group of archaea known collectivelyas methanogens which belong to phylum EuryarchaeotaThe microorganisms produce methane the second largest
anthropogenic greenhouse gas which has a global warmingpotential 25 times that of carbon dioxide [55] In the presentstudy a reduction of methanogenic archaea in tannins sup-plemented steers was detected as well as an inverse cor-relation between the abundances of phylum Euryarchaeotaand rumen pH (119903 = minus095) Tannins are thought todirectly inhibit methanogens as well as indirectly limitmethanogenesis through reduction of hydrogen availability[1 55] Saminathan et al found a significant decrease in genusMethanobrevibacter after treatment of rumen samples withcondensed tannins in vitro [56] Other authors describeda linear decrease of genus Methanobrevibacter after dietarysupplementation with condensed tannins from pine bark ingoats [31]
Ruminal pH was significantly higher in tannins treatedsteers than in the control group while urease activity exhib-ited the opposite pattern These results agree with previousreports which found an increase in ruminal pH after sup-plementation with chestnut and quebracho tannins [57 58]Other authors observed that feeding chestnut and quebrachotannins decreased urease activity in the faeces of cows[10 11]
Feed-grade urea is an effective source of nitrogen com-monly used in beef cattle diets Ureolytic bacteria in therumen produce urease to hydrolyse urea to ammonia whichis subsequently used for the synthesis of amino acids andmicrobial protein Normally the rate of urea hydrolysisexceeds the rate of ammonia utilization which leads to poorefficiency of urea utilization and increases toxic ammoniaconcentrations in blood [59] Ureolytic bacteria in the rumencomprise a highly diverse group whose main species belongto genera Succinivibrio Treponema Bacteroides ButyrivibrioStreptococcus andBifidobacterium [60 61]We found that themost abundant ureolytic generaButyrivibrio andTreponemawere negatively affected by tannins treatment Thus theobserved decline in ruminal urease activity may be relatedwith the decrease of these urease-producing taxa A previousstudy also observed a drop in ruminal urease activity afteraddition of tannins to diet but a direct interaction betweentannins and urease enzyme was postulated as responsible forthis inhibition [62]
5 Conclusions
The current study showed that chestnut and quebrachotannins added to the diet of Holstein steers modifiedrumen microbiota composition particularly fiber and starchdegrading bacteria mainly by reducing the abundanceof Prevotella and Fibrobacter while favoring Ruminococ-caceae and other members of phylum Firmicutes Tanninstreatment significantly increased pH and decreased ureaseactivity in the ruminal liquor Further work is necessaryto assess the possible relation between tannins ability tomodify rumen microbiota composition and the alterationof rumen fermentation parameters associated with energyand feed efficiency of beef cattle such as the profile ofshort-chain fatty acids and the emissions of ammonia andmethane
BioMed Research International 9
Conflicts of Interest
The authors declare that there are no known conflicts ofinterest associated with this publication and there has beenno financial support for this work that could have influencedits outcome
Acknowledgments
The authors thank Alfredo Ivan Martınez Caceres for histechnical assistance during installation and startup of thebioinformatics tools used in this study This work wassupported by grants from INTA (PNSA 1115052 and PNBIO1131043) Additional support was provided by CONICET
References
[1] A K Patra and J Saxena ldquoDietary phytochemicals as rumenmodifiers a review of the effects on microbial populationsrdquoInternational Journal of General and Molecular Microbiologyvol 96 no 4 pp 363ndash375 2009 Antonie Van Leeuwenhoek
[2] R Bodas N Prieto R Garcıa-Gonzalez S Andres F JGiraldez and S Lopez ldquoManipulation of rumen fermentationand methane production with plant secondary metabolitesrdquoAnimal Feed Science and Technology vol 176 no 1-4 pp 78ndash932012
[3] E Oskoueian N Abdullah and A Oskoueian ldquoEffects offlavonoids on rumen fermentation activity methane produc-tion andmicrobial populationrdquo BioMed Research Internationalvol 2013 Article ID 349129 8 pages 2013
[4] P Frutos GHervas F J Giraldez andA RMantecon ldquoReviewtannins and ruminant nutritionrdquo Spanish Journal of AgriculturalResearch vol 2 no 2 pp 191ndash202 2004
[5] K Khanbabaee and T van Ree ldquoTannins classification anddefinitionrdquo Natural Product Reports vol 18 no 6 pp 641ndash6492001
[6] L J Porter ldquoStructure and chemical properties of the con-densed tanninsrdquo Plant Polyphenols pp 245ndash258 1992 Springer Boston MA USA
[7] B R Min C Wright P Ho J-S Eun N Gurung and RShange ldquoThe effect of phytochemical tannins-containing dieton rumen fermentation characteristics and microbial diversitydynamics in goats using 16S rDNA amplicon pyrosequencingrdquoAgriculture Food and Analytical Bacteriology vol 4 no 3 pp195ndash211 2014
[8] B S Gemeda and A Hassen ldquoEffect of tannin and speciesvariation on in vitro digestibility gas and methane productionof tropical browse plantsrdquoAsian-Australasian Journal of AnimalSciences vol 28 no 2 pp 188ndash199 2015
[9] Z Li A-D G Wright H Liu et al ldquoResponse of the rumenmicrobiota of sika deer (cervus nippon) fed different concen-trations of tannin rich plantsrdquo PLoS ONE vol 10 no 5 ArticleID e0123481 2015
[10] JM PowelM J Aguerre andMAWattiaux ldquoTannin extractsabate ammonia emissions from simulated dairy barn floorsrdquoJournal of Environmental Quality vol 40 no 3 pp 907ndash9142011
[11] M J Aguerre M C Capozzolo P Lencioni C Cabral andM A Wattiaux ldquoEffect of quebracho-chestnut tannin extractsat 2 dietary crude protein levels on performance rumenfermentation and nitrogen partitioning in dairy cowsrdquo Journal
of Dairy Science vol 99 no 6 Article ID 74473 pp 4476ndash44862016
[12] C Rivera-Mendez A Plascencia N Torrentera andR A ZinnldquoEffect of level and source of supplemental tannin on growthperformance of steers during the late finishing phaserdquo Journalof Applied Animal Research vol 45 pp 199ndash203 2017
[13] B R Min T N Barry G T Attwood and W C McNabbldquoThe effect of condensed tannins on the nutrition and health ofruminants fed fresh temperate forages a reviewrdquo Animal FeedScience and Technology vol 106 no 1-4 pp 3ndash19 2003
[14] P J Turnbaugh R E Ley M A Mahowald V MagriniE R Mardis and J I Gordon ldquoAn obesity-associated gutmicrobiomewith increased capacity for energy harvestrdquoNaturevol 444 no 7122 pp 1027ndash1031 2006
[15] R E Ley R Knight and J I Gordon ldquoThe human micro-biome Eliminating the biomedicalenvironmental dichotomyin microbial ecologyrdquo Environmental Microbiology vol 9 no 1pp 3-4 2007
[16] I Mizrahi ldquoThe role of the rumen microbiota in determiningthe feed efficiency of dairy cowsrdquo Beneficial Microorganismsin Multicellular Life Forms pp 203ndash210 2012 Springer BerlinHeidelberg
[17] E Jami B AWhite and IMizrahi ldquoPotential role of the bovinerumen microbiome in modulating milk composition and feedefficiencyrdquo PLoS ONE vol 9 no 1 Article ID e85423 2014
[18] S C Fernando H T Purvis F Z Najar et al ldquoRumenmicrobialpopulation dynamics during adaptation to a high-grain dietrdquoApplied and Environmental Microbiology vol 76 no 22 pp7482ndash7490 2010
[19] H P S Makkar ldquoEffects and fate of tannins in ruminantanimals adaptation to tannins and strategies to overcomedetrimental effects of feeding tannin-rich feedsrdquo Small Rumi-nant Research vol 49 no 3 pp 241ndash256 2003
[20] Illumina Protocol 16S Metagenomic Sequencing Library 2013[21] AM BolgerM Lohse and B Usadel ldquoTrimmomatic a flexible
trimmer for Illumina sequence datardquoBioinformatics vol 30 no15 pp 2114ndash2120 2014
[22] J Kuczynski J Stombaugh W A Walters A Gonzalez J GCaporaso and R Knight ldquoUsing QIIME to analyze 16S rRNAgene sequences frommicrobial communitiesrdquoCurrent Protocolsin Microbiology vol 1 1E5 pages 2012
[23] N A Bokulich S Subramanian J J Faith et al ldquoQuality-filtering vastly improves diversity estimates from Illuminaamplicon sequencingrdquo Nature Methods vol 10 no 1 pp 57ndash592013
[24] D Parks and R Beiko ldquoSTAMP statistical analysis of metage-nomic profilesrdquo Encyclopedia of Metagenomics pp 1ndash6 2013Springer New York NY USA
[25] J C McCann T A Wickersham and J J Loor ldquoHigh-throughput methods redefine the rumen microbiome and itsrelationshipwith nutrition andmetabolismrdquoBioinformatics andBiology Insights vol 8 pp 109ndash125 2014
[26] G Henderson F Cox S Ganesh A Jonker W Young and PH Janssen ldquoRumen microbial community composition varieswith diet and host but a coremicrobiome is found across a widegeographical rangerdquo Scientific Reports vol 5 no 14567 pp 1ndash132015
[27] P J Weimer ldquoRedundancy resilience and host specificity ofthe ruminal microbiota Implications for engineering improvedruminal fermentationsrdquo Frontiers in Microbiology vol 6 articleno 296 2015
10 BioMed Research International
[28] E Jami and I Mizrahi ldquoComposition and similarity of bovinerumenmicrobiota across individual animalsrdquo PLoS ONE vol 7no 3 Article ID e33306 2012
[29] R De Nardi G Marchesini S Li et al ldquoMetagenomic analysisof rumen microbial population in dairy heifers fed a high graindiet supplemented with dicarboxylic acids or polyphenolsrdquoBMC Veterinary Research vol 12 no 1 article no 29 2016
[30] S Kruger Ben Shabat G Sasson A Doron-Faigenboim etal ldquoSpecific microbiome-dependent mechanisms underlie theenergy harvest efficiency of ruminantsrdquoThe ISME Journal vol10 no 12 pp 2958ndash2972 2016
[31] B R Min S Solaiman R Shange and J-S Eun ldquoGastroin-testinal bacterial andmethanogenic archaea diversity dynamicsassociated with condensed tannin-containing pine bark diet ingoats using 16S rDNA amplicon pyrosequencingrdquo InternationalJournal of Microbiology vol 2014 Article ID 141909 2014
[32] G Henderson F Cox S Kittelmann et al ldquoEffect of DNAextraction methods and sampling techniques on the apparentstructure of cow and sheep rumen microbial communitiesrdquoPLoS ONE vol 8 no 9 Article ID e74787 2013
[33] P R Myer T P L Smith J E Wells L A Kuehn and HC Freetly ldquoRumen microbiome from steers differing in feedefficiencyrdquo PLoS ONE vol 10 no 6 Article ID e0129174 2015
[34] Y Wang and T A McAllister ldquoRumen microbes enzymes andfeed digestion-a reviewrdquo Asian-Australasian Journal of AnimalSciences vol 15 no 11 pp 1659ndash1676 2002
[35] J B Russell R EMuck and P JWeimer ldquoQuantitative analysisof cellulose degradation and growth of cellulolytic bacteria inthe rumenrdquo FEMS Microbiology Ecology vol 67 no 2 pp 183ndash197 2009
[36] DOKrause S EDenmanR IMackie et al ldquoOpportunities toimprove fiber degradation in the rumen microbiology ecologyand genomicsrdquo FEMS Microbiology Reviews vol 27 no 5 pp663ndash693 2003
[37] M Zorec M Vodovnik and R Marinsek-Logar ldquoPotentialof selected rumen bacteria for cellulose and hemicellulosedegradationrdquo Food Technology and Biotechnology vol 52 no 2pp 210ndash221 2014
[38] A Z Bekele S Koike and Y Kobayashi ldquoGenetic diversityand diet specificity of ruminal prevotella revealed by 16S rRNAgene-based analysisrdquo FEMSMicrobiology Letters vol 305 no 1pp 49ndash57 2010
[39] A RamsakM Peterka K Tajima et al ldquoUnravelling the geneticdiversity of ruminal bacteria belonging to the CFB phylumrdquoFEMS Microbiology Ecology vol 33 no 1 pp 69ndash79 2000
[40] J Purushe D E Fouts M Morrison et al ldquoComparativegenome analysis of prevotella ruminicola and prevotella bryan-tii insights into their environmental nicherdquo Microbial Ecologyvol 60 no 4 pp 721ndash729 2010
[41] C A Carberry D A Kenny S Han M S McCabe and S MWaters ldquoEffect of phenotypic residual feed intake and dietaryforage content on the rumen microbial community of beefcattlerdquo Applied and Environmental Microbiology vol 78 no 14pp 4949ndash4958 2012
[42] A L Molan G T Attwood B R Min and W C McNabbldquoThe effect of condensed tannins from Lotus pedunculatus andLotus corniculatus on the growth of proteolytic rumen bacteriain vitro and their possible mode of actionrdquo Canadian Journal ofMicrobiology vol 47 no 7 pp 626ndash633 2001
[43] A Jayanegara G Goel H P S Makkar and K Becker ldquoDiver-gence between purified hydrolysable and condensed tannin
effects on methane emission rumen fermentation and micro-bial population in vitrordquo Animal Feed Science and Technologyvol 209 pp 60ndash68 2015
[44] BMinW Pinchak K Hernandez et al ldquoEffects of plant tanninsupplementation on animal responses and in vivo ruminalbacterial populations associated with bloat in heifers grazingwheat forage1rdquo The Professional Animal Scientist vol 28 no 4pp 464ndash472 2012
[45] T F Martınez T A McAllister Y Wang and T Reuter ldquoEffectsof tannic acid and quebracho tannins on in vitro ruminalfermentation of wheat and corn grainrdquo Journal of the Scienceof Food and Agriculture vol 86 no 8 pp 1244ndash1256 2006
[46] L Marın E M Miguelez C J Villar and F Lombo ldquoBioavail-ability of dietary polyphenols and gut microbiota metabolismantimicrobial propertiesrdquo BioMed Research International vol2015 Article ID 905215 18 pages 2015
[47] J B Russell and D B Wilson ldquoWhy are ruminal cellulolyticbacteria unable to digest cellulose at low pHrdquo Journal of DairyScience vol 79 no 8 pp 1503ndash1509 1996
[48] K Tajima S Arai K Ogata et al ldquoRumen bacterial communitytransition during adaptation to high-grain dietrdquo Anaerobe vol6 no 5 pp 273ndash284 2000
[49] D W Goad C L Goad and T G Nagaraja ldquoRuminal micro-bial and fermentative changes associated with experimentallyinduced subacute acidosis in steersrdquo Journal of Animal Sciencevol 76 no 1 pp 234ndash241 1998
[50] J Hernandez J L Benedito A Abuelo and C CastilloldquoRuminal acidosis in feedlot from aetiology to preventionrdquoTheScientific World Journal vol 2014 Article ID 702572 2014
[51] A B De Menezes E Lewis M OrsquoDonovan B F OrsquoNeill NClipson and E M Doyle ldquoMicrobiome analysis of dairy cowsfed pasture or total mixed ration dietsrdquo FEMS MicrobiologyEcology vol 78 no 2 pp 256ndash265 2011
[52] I K Skene and J D Brooker ldquoCharacterization of tanninacylhydrolase activity in the ruminal bacterium selenomonasruminantiumrdquo Anaerobe vol 1 no 6 pp 321ndash327 1995
[53] G Goel A K Puniya and K Singh ldquoTannic acid resistance inruminal streptococcal isolatesrdquo Journal of Basic Microbiologyvol 45 no 3 pp 243ndash245 2005
[54] P R Myer J E Wells T P L Smith L A Kuehn and H CFreetly ldquoCecum microbial communities from steers differingin feed efficiencyrdquo Journal of Animal Science vol 93 no 11 pp5327ndash5340 2015
[55] S E Hook A-D GWright and BWMcBride ldquoMethanogensmethane producers of the rumen and mitigation strategiesrdquoArchaea vol 2010 Article ID 945785 11 pages 2010
[56] M Saminathan C C Sieo H M Gan N Abdullah C M VL Wong and Y W Ho ldquoEffects of condensed tannin fractionsof different molecular weights on population and diversity ofbovine rumen methanogenic archaea in vitro as determinedby high-throughput sequencingrdquo Animal Feed Science andTechnology vol 216 pp 146ndash160 2016
[57] F Hassanat and C Benchaar ldquoAssessment of the effect ofcondensed (acacia and quebracho) and hydrolysable (chestnutand valonea) tannins on rumen fermentation andmethane pro-duction in vitrordquo Journal of the Science of Food and Agriculturevol 93 no 2 pp 332ndash339 2013
[58] J Balcells A Aris A Serrano A R Seradj J Crespo and MDevant ldquoEffects of an extract of plant flavonoids (bioflavex)on rumen fermentation and performance in heifers fed high-concentrate dietsrdquo Journal of Animal Science vol 90 no 13 pp4975ndash4984 2012
BioMed Research International 11
[59] A K Patra ldquoUreaammonia metabolism in the rumen andtoxicity in ruminantsrdquo Rumen Microbiology From Evolution toRevolution pp 329ndash341 2015
[60] MAWoznyM P Bryant L VHoldeman andW E CMooreldquoUrease assay and urease producing species of anaerobes in thebovine rumen and human fecesrdquo Applied and EnvironmentalMicrobiology vol 33 no 5 pp 1097ndash1104 1977
[61] D Jin S Zhao P Wang et al ldquoInsights into abundant rumenureolytic bacterial community using rumen simulation systemrdquoFrontiers in Microbiology vol 7 article no 1006 2016
[62] H P SMakkar B Singh andRKDawra ldquoEffect of tannin-richleaves of oak (Quercus incana) on various microbial enzymeactivities of the bovine rumenrdquo British Journal of Nutrition vol60 no 2 pp 287ndash296 1988
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
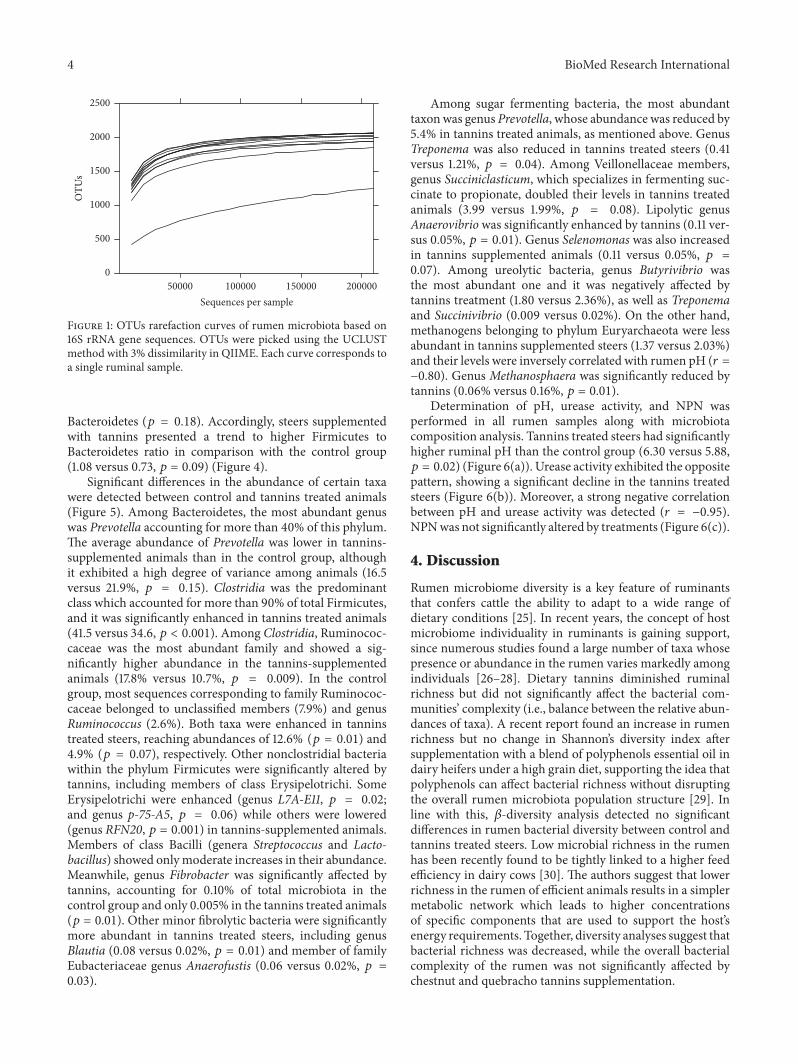
4 BioMed Research International
0
500
1000
1500
2000
2500
OTU
s
100000 150000 20000050000Sequences per sample
Figure 1 OTUs rarefaction curves of rumen microbiota based on16S rRNA gene sequences OTUs were picked using the UCLUSTmethod with 3 dissimilarity in QIIME Each curve corresponds toa single ruminal sample
Bacteroidetes (119901 = 018) Accordingly steers supplementedwith tannins presented a trend to higher Firmicutes toBacteroidetes ratio in comparison with the control group(108 versus 073 119901 = 009) (Figure 4)
Significant differences in the abundance of certain taxawere detected between control and tannins treated animals(Figure 5) Among Bacteroidetes the most abundant genuswas Prevotella accounting for more than 40 of this phylumThe average abundance of Prevotella was lower in tannins-supplemented animals than in the control group althoughit exhibited a high degree of variance among animals (165versus 219 119901 = 015) Clostridia was the predominantclass which accounted for more than 90 of total Firmicutesand it was significantly enhanced in tannins treated animals(415 versus 346 119901 lt 0001) Among Clostridia Ruminococ-caceae was the most abundant family and showed a sig-nificantly higher abundance in the tannins-supplementedanimals (178 versus 107 119901 = 0009) In the controlgroup most sequences corresponding to family Ruminococ-caceae belonged to unclassified members (79) and genusRuminococcus (26) Both taxa were enhanced in tanninstreated steers reaching abundances of 126 (119901 = 001) and49 (119901 = 007) respectively Other nonclostridial bacteriawithin the phylum Firmicutes were significantly altered bytannins including members of class Erysipelotrichi SomeErysipelotrichi were enhanced (genus L7A-E11 119901 = 002and genus p-75-A5 119901 = 006) while others were lowered(genus RFN20 119901 = 0001) in tannins-supplemented animalsMembers of class Bacilli (genera Streptococcus and Lacto-bacillus) showed onlymoderate increases in their abundanceMeanwhile genus Fibrobacter was significantly affected bytannins accounting for 010 of total microbiota in thecontrol group and only 0005 in the tannins treated animals(119901 = 001) Other minor fibrolytic bacteria were significantlymore abundant in tannins treated steers including genusBlautia (008 versus 002 119901 = 001) and member of familyEubacteriaceae genus Anaerofustis (006 versus 002 119901 =003)
Among sugar fermenting bacteria the most abundanttaxonwas genus Prevotella whose abundance was reduced by54 in tannins treated animals as mentioned above GenusTreponema was also reduced in tannins treated steers (041versus 121 119901 = 004) Among Veillonellaceae membersgenus Succiniclasticum which specializes in fermenting suc-cinate to propionate doubled their levels in tannins treatedanimals (399 versus 199 119901 = 008) Lipolytic genusAnaerovibrio was significantly enhanced by tannins (011 ver-sus 005 119901 = 001) Genus Selenomonas was also increasedin tannins supplemented animals (011 versus 005 119901 =007) Among ureolytic bacteria genus Butyrivibrio wasthe most abundant one and it was negatively affected bytannins treatment (180 versus 236) as well as Treponemaand Succinivibrio (0009 versus 002) On the other handmethanogens belonging to phylum Euryarchaeota were lessabundant in tannins supplemented steers (137 versus 203)and their levels were inversely correlated with rumen pH (119903 =minus080) Genus Methanosphaera was significantly reduced bytannins (006 versus 016 119901 = 001)
Determination of pH urease activity and NPN wasperformed in all rumen samples along with microbiotacomposition analysis Tannins treated steers had significantlyhigher ruminal pH than the control group (630 versus 588119901 = 002) (Figure 6(a)) Urease activity exhibited the oppositepattern showing a significant decline in the tannins treatedsteers (Figure 6(b)) Moreover a strong negative correlationbetween pH and urease activity was detected (119903 = minus095)NPNwas not significantly altered by treatments (Figure 6(c))
4 Discussion
Rumen microbiome diversity is a key feature of ruminantsthat confers cattle the ability to adapt to a wide range ofdietary conditions [25] In recent years the concept of hostmicrobiome individuality in ruminants is gaining supportsince numerous studies found a large number of taxa whosepresence or abundance in the rumen varies markedly amongindividuals [26ndash28] Dietary tannins diminished ruminalrichness but did not significantly affect the bacterial com-munitiesrsquo complexity (ie balance between the relative abun-dances of taxa) A recent report found an increase in rumenrichness but no change in Shannonrsquos diversity index aftersupplementation with a blend of polyphenols essential oil indairy heifers under a high grain diet supporting the idea thatpolyphenols can affect bacterial richness without disruptingthe overall rumen microbiota population structure [29] Inline with this 120573-diversity analysis detected no significantdifferences in rumen bacterial diversity between control andtannins treated steers Low microbial richness in the rumenhas been recently found to be tightly linked to a higher feedefficiency in dairy cows [30] The authors suggest that lowerrichness in the rumen of efficient animals results in a simplermetabolic network which leads to higher concentrationsof specific components that are used to support the hostrsquosenergy requirements Together diversity analyses suggest thatbacterial richness was decreased while the overall bacterialcomplexity of the rumen was not significantly affected bychestnut and quebracho tannins supplementation
BioMed Research International 5
Control Tannins1000
1100
1200
1300
1400
1500
1600
1700
1800
1900
2000
2100
2200N
umbe
r of O
TUs
p = 005
(a)Control Tannins
6
7
8
9
10
Shan
nonrsquos
inde
x
p = 014
(b)
Figure 2 Effect of tannins treatment on (a) bacterial richness (number of OTUs) and (b) Shannonrsquos diversity index of rumen microbiomeLine = median Box = 25ndash75 percentiles Bar = 5ndash95 percentiles
minus010
minus005
000
005
010
015
PC2
(11
1)
00 01 02 03 04 05minus01PC1 (556)
Figure 3 PCoA plot based on unweighted UniFrac metric Itemsshaped with triangles and circles correspond to samples fromcontrol and tannins treated animals respectively Axes (PC1 = 556and PC2 = 111) account for 667 of the total variation detected
The dominance of phyla Firmicutes and Bacteroidetesin the bovine ruminal fluid is a common feature in thegastrointestinal microbiome of monogastric organisms andruminants [15 17 28 31] Henderson et al also found thatan increase in the ruminal abundance of total Firmicutescorrelated with a decrease in the abundance of Bacteroidetesboth in cows (119903 = minus080) and in sheep (119903 = minus097) [32] Otherauthors found that Bacteroidetes were the most abundant
Control Tannins
p = 009
04
05
06
07
08
09
10
11
12
13
14
15
16
17
Firm
icut
esB
acte
roid
etes
Figure 4 Effect of tannins on the ratio of phyla Firmicutes andBacteroidetes in rumen microbiota Line = median Box = 25ndash75percentiles Bar = 5ndash95 percentiles
phylum in ruminal samples obtained from dairy cows butsome animals exhibited a higher percentage of Firmicutescompensating for a lower abundance of Bacteroidetes [17]Our results agree with these observations since a stronginverse correlation between the abundances of Firmicutesand Bacteroidetes was detected (119903 = minus099) These resultssuggest that members of Firmicutes and Bacteroidetes com-pete for available resources in the rumen and tannins wouldtip the balance in favor of Firmicutes
6 BioMed Research International
lowast
lowastlowast
lowast
lowastlowast
lowast
lowast
lowastlowastlowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
lowast
lowastlowast
lowast
lowast
lowast
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
minus4 minus2 minus1minus3minus5 0 2 3 4 51
Phylum FirmicutesGenus Butyrivibrio
Unc Family RuminococcaceaeGenus Ruminococcus
Genus SucciniclasticumGenus Streptococcus
Genus p-75-a5Genus L7A-E11
Genus SelenomonasGenus Anaerovibrio
Genus BlautiaGenus Anaerofustis
Genus RFN20Genus Lachnospira
Phylum BacteroidetesFamily Prevotellaceae
Family BacteroidaceaeGenus Prevotella
Genus BacteroidesGenus YRC22
Unc Family RF16
Unc Family CoriobacteriaceaePhylum Actinobacteria
Phylum SpirochaetesGenus Treponema
Genus Sphaerochaeta
Phylum EuryarchaeotaGenus Methanobrevibacter
Genus Methanosphaera
Phylum ProteobacteriaUnclass Order GMD14H09
Genus RuminobacterGenus Oxalobacter
Genus Succinivibrio
Phylum TenericutesUnc Family Anaeroplasmata
Genus Anaeroplasma
Phylum ChloroflexiGenus SHD-231
Phylum FibrobacteresGenus Fibrobacter
log 2 fold change
Figure 5 Relative fold changes (log 2 tanninscontrol) in the abundance of rumen bacterial taxa between control and tannins treated steerslowastlowast119901 lt 005 lowast119901 lt 010 Bar = SEM
The ratio of Firmicutes to Bacteroidetes has been shownto affect energy harvesting and body fat accumulation inhumans and mice [14 15] Along with increased fatty acidabsorption more energy was found to be efficiently obtainedfrom diet in obese mice illustrating the connection betweenFirmicutes and improved efficiency in energy harvesting [14]In cows the Firmicutes to Bacteroidetes ratio was found tobe strongly correlated with daily milk-fat yield [17] A recentstudy found that the abundance of Firmicutes in the rumen
positively correlates with the average daily body weight gainin steers suggesting that these bacteria play a significant rolein feed efficiency of bovines [33] Therefore it is possiblethat the increase of Firmicutes to Bacteroidetes ratio inducedby tannins can improve bovine performance as previouslysuggested by other authors [19]
Fiber degradation is a complex process carried out bya group of microorganisms that are able to digest plantpolysaccharides mainly through production of cellulolytic
BioMed Research International 7
Control Tannins50
52
54
56
58
60
62
64
66
68
70pH
p = 002
(a)Control Tannins
000
005
010
015
020
Ure
ase a
ctiv
ity (U
A)
p = 0007
(b)
Control Tannins
000
005
010
015
Non
prot
ein
nitro
gen
(NPN
)
p = 0849
(c)
Figure 6 Effect of tannins on pH urease activity and NPN in the rumen liquor Line = median Box = 25ndash75 percentiles Bar = 5ndash95percentiles Urease activity values are given in pH units proportional to urease activity NPN levels are expressed as a percentage of solublenitrogen in the rumen
hemicellulolytic and pectinolytic enzymes [34] Cellulolyticactivity in the rumen involves a diverse bacterial commu-nity whose main members belong to genera Ruminococcusand Fibrobacter while the main hemicellulolytic bacteriabelong to genera Ruminococcus Prevotella and Butyrivib-rio [34ndash37] The blend of tannins administrated increasedRuminococcaceae and other members of phylum Firmicuteswhile inhibited genera Prevotella and Fibrobacter Prevotellaspecies have a documented role in metabolism of starchhemicellulose pectin and protein catabolism [25] The highabundance of this genus might be the result of a largemetabolic niche that is occupied by different species with
similar metabolic capabilities or it might be associated witha high genetic variability that enables members of genusPrevotella to occupy different ecological niches within therumen [38ndash40] For instance Prevotella abundance has beenshown to be higher in the rumen of animals of beef cattlewith low feed efficiency [41] Previous studies reported a greatdiversity in the sensitivity ofPrevotella species to tannins [42]which could explain the lower abundance detected for thisgenus in tannins-supplemented steers Since genus Prevotellais characterized by a high genetic diversity and wide array ofoutputmetabolites [38ndash40] their replacement byRuminococ-cus may lead to a simpler product profile specialized to
8 BioMed Research International
support the hostrsquos energy requirements [30] Our resultsagree with previous studies that found a significant reduc-tion in Fibrobacter and Prevotella rumen populations aftersupplementation with condensed and hydrolysable tanninsincluding chestnut and quebracho tannins [43 44]
Rumen fibrolytic function is carried out by a largelyredundant microbial community with overlapping distribu-tion of metabolic capabilities and this fibrolytic communityhas the ability to restore its structure and function afterperturbation a phenomenon known as resilience [27] In thiscontext the observed changes in diversity of fibrolytic bacte-ria in tannins treated steers may result from a combinationof physicochemical and biological mechanisms described inthe literature including direct interaction of tannins withfiber [45] which could alter the available surface area formicrobial attack the inhibition of certain fibrolytic taxa bymeans of tannins antimicrobial activity [46] andmodulationof fibrolytic bacterial species driven by changes in rumen pHsince certain fibrolytic taxa of the rumen are inhibited at lowpH under high-grain diets [36 47]
Rumen amylolytic and saccharolytic bacteria were alsoaffected by dietary treatment with tannins mainly throughthe decrease of genera Prevotella and Treponema Otheramylolytic genera were moderately increased in tannins-fedsteers including Streptococcus Bifidobacterium and Lacto-bacillus Amylolytic activity is normally enhanced in rumi-nants consuming high-grain diets [18 48 49] The rate ofgrain degradation by these microbial communities plays akey role in maintaining rumen homeostasis since rapidstarch fermentation produces large amounts of organic acidstherefore producing a drop in ruminal pH that may leadto metabolic acidosis [50] A previous study showed thattannic acid and quebracho tannins lower the rate ofmicrobialhydrolysis of starch-rich grains in the rumen by physicalmodification of the endosperm protein matrix [45] Thisphysical modulation of starch degradation could explain thelower abundance of sugar fermenting taxa detected in tanninstreated steers as well as the higher ruminal pH observed inthis group
Fermentation products of microbial activity in rumenmainly short-chain volatile fatty acids serve as amajor sourceof energy for ruminants and have a direct impact on thephysiological parameters of the animal and feed utilizationefficiency [16] Some members of family Veillonellaceaewhich produce propionate as a major fermentation productand have been associated with lower methane emissions [51]were enhanced in tannins treated animals Interestingly someSelenomonas species can break tannin-protein complexes anduse tannins as energy source [52 53] Therefore the higherabundance of Selenomonas in tannins treated steers may bepartly due to availability of tannins as direct energy sourceMeanwhile members of class Erysipelotrichi which havebeen linked to beef cattle feed efficiency [33 51 54] were alsomodulated by tannins treatment
Methane production during fermentation of feeds in therumen represents a loss of 2ndash12 of gross energy [1] andit is performed by a group of archaea known collectivelyas methanogens which belong to phylum EuryarchaeotaThe microorganisms produce methane the second largest
anthropogenic greenhouse gas which has a global warmingpotential 25 times that of carbon dioxide [55] In the presentstudy a reduction of methanogenic archaea in tannins sup-plemented steers was detected as well as an inverse cor-relation between the abundances of phylum Euryarchaeotaand rumen pH (119903 = minus095) Tannins are thought todirectly inhibit methanogens as well as indirectly limitmethanogenesis through reduction of hydrogen availability[1 55] Saminathan et al found a significant decrease in genusMethanobrevibacter after treatment of rumen samples withcondensed tannins in vitro [56] Other authors describeda linear decrease of genus Methanobrevibacter after dietarysupplementation with condensed tannins from pine bark ingoats [31]
Ruminal pH was significantly higher in tannins treatedsteers than in the control group while urease activity exhib-ited the opposite pattern These results agree with previousreports which found an increase in ruminal pH after sup-plementation with chestnut and quebracho tannins [57 58]Other authors observed that feeding chestnut and quebrachotannins decreased urease activity in the faeces of cows[10 11]
Feed-grade urea is an effective source of nitrogen com-monly used in beef cattle diets Ureolytic bacteria in therumen produce urease to hydrolyse urea to ammonia whichis subsequently used for the synthesis of amino acids andmicrobial protein Normally the rate of urea hydrolysisexceeds the rate of ammonia utilization which leads to poorefficiency of urea utilization and increases toxic ammoniaconcentrations in blood [59] Ureolytic bacteria in the rumencomprise a highly diverse group whose main species belongto genera Succinivibrio Treponema Bacteroides ButyrivibrioStreptococcus andBifidobacterium [60 61]We found that themost abundant ureolytic generaButyrivibrio andTreponemawere negatively affected by tannins treatment Thus theobserved decline in ruminal urease activity may be relatedwith the decrease of these urease-producing taxa A previousstudy also observed a drop in ruminal urease activity afteraddition of tannins to diet but a direct interaction betweentannins and urease enzyme was postulated as responsible forthis inhibition [62]
5 Conclusions
The current study showed that chestnut and quebrachotannins added to the diet of Holstein steers modifiedrumen microbiota composition particularly fiber and starchdegrading bacteria mainly by reducing the abundanceof Prevotella and Fibrobacter while favoring Ruminococ-caceae and other members of phylum Firmicutes Tanninstreatment significantly increased pH and decreased ureaseactivity in the ruminal liquor Further work is necessaryto assess the possible relation between tannins ability tomodify rumen microbiota composition and the alterationof rumen fermentation parameters associated with energyand feed efficiency of beef cattle such as the profile ofshort-chain fatty acids and the emissions of ammonia andmethane
BioMed Research International 9
Conflicts of Interest
The authors declare that there are no known conflicts ofinterest associated with this publication and there has beenno financial support for this work that could have influencedits outcome
Acknowledgments
The authors thank Alfredo Ivan Martınez Caceres for histechnical assistance during installation and startup of thebioinformatics tools used in this study This work wassupported by grants from INTA (PNSA 1115052 and PNBIO1131043) Additional support was provided by CONICET
References
[1] A K Patra and J Saxena ldquoDietary phytochemicals as rumenmodifiers a review of the effects on microbial populationsrdquoInternational Journal of General and Molecular Microbiologyvol 96 no 4 pp 363ndash375 2009 Antonie Van Leeuwenhoek
[2] R Bodas N Prieto R Garcıa-Gonzalez S Andres F JGiraldez and S Lopez ldquoManipulation of rumen fermentationand methane production with plant secondary metabolitesrdquoAnimal Feed Science and Technology vol 176 no 1-4 pp 78ndash932012
[3] E Oskoueian N Abdullah and A Oskoueian ldquoEffects offlavonoids on rumen fermentation activity methane produc-tion andmicrobial populationrdquo BioMed Research Internationalvol 2013 Article ID 349129 8 pages 2013
[4] P Frutos GHervas F J Giraldez andA RMantecon ldquoReviewtannins and ruminant nutritionrdquo Spanish Journal of AgriculturalResearch vol 2 no 2 pp 191ndash202 2004
[5] K Khanbabaee and T van Ree ldquoTannins classification anddefinitionrdquo Natural Product Reports vol 18 no 6 pp 641ndash6492001
[6] L J Porter ldquoStructure and chemical properties of the con-densed tanninsrdquo Plant Polyphenols pp 245ndash258 1992 Springer Boston MA USA
[7] B R Min C Wright P Ho J-S Eun N Gurung and RShange ldquoThe effect of phytochemical tannins-containing dieton rumen fermentation characteristics and microbial diversitydynamics in goats using 16S rDNA amplicon pyrosequencingrdquoAgriculture Food and Analytical Bacteriology vol 4 no 3 pp195ndash211 2014
[8] B S Gemeda and A Hassen ldquoEffect of tannin and speciesvariation on in vitro digestibility gas and methane productionof tropical browse plantsrdquoAsian-Australasian Journal of AnimalSciences vol 28 no 2 pp 188ndash199 2015
[9] Z Li A-D G Wright H Liu et al ldquoResponse of the rumenmicrobiota of sika deer (cervus nippon) fed different concen-trations of tannin rich plantsrdquo PLoS ONE vol 10 no 5 ArticleID e0123481 2015
[10] JM PowelM J Aguerre andMAWattiaux ldquoTannin extractsabate ammonia emissions from simulated dairy barn floorsrdquoJournal of Environmental Quality vol 40 no 3 pp 907ndash9142011
[11] M J Aguerre M C Capozzolo P Lencioni C Cabral andM A Wattiaux ldquoEffect of quebracho-chestnut tannin extractsat 2 dietary crude protein levels on performance rumenfermentation and nitrogen partitioning in dairy cowsrdquo Journal
of Dairy Science vol 99 no 6 Article ID 74473 pp 4476ndash44862016
[12] C Rivera-Mendez A Plascencia N Torrentera andR A ZinnldquoEffect of level and source of supplemental tannin on growthperformance of steers during the late finishing phaserdquo Journalof Applied Animal Research vol 45 pp 199ndash203 2017
[13] B R Min T N Barry G T Attwood and W C McNabbldquoThe effect of condensed tannins on the nutrition and health ofruminants fed fresh temperate forages a reviewrdquo Animal FeedScience and Technology vol 106 no 1-4 pp 3ndash19 2003
[14] P J Turnbaugh R E Ley M A Mahowald V MagriniE R Mardis and J I Gordon ldquoAn obesity-associated gutmicrobiomewith increased capacity for energy harvestrdquoNaturevol 444 no 7122 pp 1027ndash1031 2006
[15] R E Ley R Knight and J I Gordon ldquoThe human micro-biome Eliminating the biomedicalenvironmental dichotomyin microbial ecologyrdquo Environmental Microbiology vol 9 no 1pp 3-4 2007
[16] I Mizrahi ldquoThe role of the rumen microbiota in determiningthe feed efficiency of dairy cowsrdquo Beneficial Microorganismsin Multicellular Life Forms pp 203ndash210 2012 Springer BerlinHeidelberg
[17] E Jami B AWhite and IMizrahi ldquoPotential role of the bovinerumen microbiome in modulating milk composition and feedefficiencyrdquo PLoS ONE vol 9 no 1 Article ID e85423 2014
[18] S C Fernando H T Purvis F Z Najar et al ldquoRumenmicrobialpopulation dynamics during adaptation to a high-grain dietrdquoApplied and Environmental Microbiology vol 76 no 22 pp7482ndash7490 2010
[19] H P S Makkar ldquoEffects and fate of tannins in ruminantanimals adaptation to tannins and strategies to overcomedetrimental effects of feeding tannin-rich feedsrdquo Small Rumi-nant Research vol 49 no 3 pp 241ndash256 2003
[20] Illumina Protocol 16S Metagenomic Sequencing Library 2013[21] AM BolgerM Lohse and B Usadel ldquoTrimmomatic a flexible
trimmer for Illumina sequence datardquoBioinformatics vol 30 no15 pp 2114ndash2120 2014
[22] J Kuczynski J Stombaugh W A Walters A Gonzalez J GCaporaso and R Knight ldquoUsing QIIME to analyze 16S rRNAgene sequences frommicrobial communitiesrdquoCurrent Protocolsin Microbiology vol 1 1E5 pages 2012
[23] N A Bokulich S Subramanian J J Faith et al ldquoQuality-filtering vastly improves diversity estimates from Illuminaamplicon sequencingrdquo Nature Methods vol 10 no 1 pp 57ndash592013
[24] D Parks and R Beiko ldquoSTAMP statistical analysis of metage-nomic profilesrdquo Encyclopedia of Metagenomics pp 1ndash6 2013Springer New York NY USA
[25] J C McCann T A Wickersham and J J Loor ldquoHigh-throughput methods redefine the rumen microbiome and itsrelationshipwith nutrition andmetabolismrdquoBioinformatics andBiology Insights vol 8 pp 109ndash125 2014
[26] G Henderson F Cox S Ganesh A Jonker W Young and PH Janssen ldquoRumen microbial community composition varieswith diet and host but a coremicrobiome is found across a widegeographical rangerdquo Scientific Reports vol 5 no 14567 pp 1ndash132015
[27] P J Weimer ldquoRedundancy resilience and host specificity ofthe ruminal microbiota Implications for engineering improvedruminal fermentationsrdquo Frontiers in Microbiology vol 6 articleno 296 2015
10 BioMed Research International
[28] E Jami and I Mizrahi ldquoComposition and similarity of bovinerumenmicrobiota across individual animalsrdquo PLoS ONE vol 7no 3 Article ID e33306 2012
[29] R De Nardi G Marchesini S Li et al ldquoMetagenomic analysisof rumen microbial population in dairy heifers fed a high graindiet supplemented with dicarboxylic acids or polyphenolsrdquoBMC Veterinary Research vol 12 no 1 article no 29 2016
[30] S Kruger Ben Shabat G Sasson A Doron-Faigenboim etal ldquoSpecific microbiome-dependent mechanisms underlie theenergy harvest efficiency of ruminantsrdquoThe ISME Journal vol10 no 12 pp 2958ndash2972 2016
[31] B R Min S Solaiman R Shange and J-S Eun ldquoGastroin-testinal bacterial andmethanogenic archaea diversity dynamicsassociated with condensed tannin-containing pine bark diet ingoats using 16S rDNA amplicon pyrosequencingrdquo InternationalJournal of Microbiology vol 2014 Article ID 141909 2014
[32] G Henderson F Cox S Kittelmann et al ldquoEffect of DNAextraction methods and sampling techniques on the apparentstructure of cow and sheep rumen microbial communitiesrdquoPLoS ONE vol 8 no 9 Article ID e74787 2013
[33] P R Myer T P L Smith J E Wells L A Kuehn and HC Freetly ldquoRumen microbiome from steers differing in feedefficiencyrdquo PLoS ONE vol 10 no 6 Article ID e0129174 2015
[34] Y Wang and T A McAllister ldquoRumen microbes enzymes andfeed digestion-a reviewrdquo Asian-Australasian Journal of AnimalSciences vol 15 no 11 pp 1659ndash1676 2002
[35] J B Russell R EMuck and P JWeimer ldquoQuantitative analysisof cellulose degradation and growth of cellulolytic bacteria inthe rumenrdquo FEMS Microbiology Ecology vol 67 no 2 pp 183ndash197 2009
[36] DOKrause S EDenmanR IMackie et al ldquoOpportunities toimprove fiber degradation in the rumen microbiology ecologyand genomicsrdquo FEMS Microbiology Reviews vol 27 no 5 pp663ndash693 2003
[37] M Zorec M Vodovnik and R Marinsek-Logar ldquoPotentialof selected rumen bacteria for cellulose and hemicellulosedegradationrdquo Food Technology and Biotechnology vol 52 no 2pp 210ndash221 2014
[38] A Z Bekele S Koike and Y Kobayashi ldquoGenetic diversityand diet specificity of ruminal prevotella revealed by 16S rRNAgene-based analysisrdquo FEMSMicrobiology Letters vol 305 no 1pp 49ndash57 2010
[39] A RamsakM Peterka K Tajima et al ldquoUnravelling the geneticdiversity of ruminal bacteria belonging to the CFB phylumrdquoFEMS Microbiology Ecology vol 33 no 1 pp 69ndash79 2000
[40] J Purushe D E Fouts M Morrison et al ldquoComparativegenome analysis of prevotella ruminicola and prevotella bryan-tii insights into their environmental nicherdquo Microbial Ecologyvol 60 no 4 pp 721ndash729 2010
[41] C A Carberry D A Kenny S Han M S McCabe and S MWaters ldquoEffect of phenotypic residual feed intake and dietaryforage content on the rumen microbial community of beefcattlerdquo Applied and Environmental Microbiology vol 78 no 14pp 4949ndash4958 2012
[42] A L Molan G T Attwood B R Min and W C McNabbldquoThe effect of condensed tannins from Lotus pedunculatus andLotus corniculatus on the growth of proteolytic rumen bacteriain vitro and their possible mode of actionrdquo Canadian Journal ofMicrobiology vol 47 no 7 pp 626ndash633 2001
[43] A Jayanegara G Goel H P S Makkar and K Becker ldquoDiver-gence between purified hydrolysable and condensed tannin
effects on methane emission rumen fermentation and micro-bial population in vitrordquo Animal Feed Science and Technologyvol 209 pp 60ndash68 2015
[44] BMinW Pinchak K Hernandez et al ldquoEffects of plant tanninsupplementation on animal responses and in vivo ruminalbacterial populations associated with bloat in heifers grazingwheat forage1rdquo The Professional Animal Scientist vol 28 no 4pp 464ndash472 2012
[45] T F Martınez T A McAllister Y Wang and T Reuter ldquoEffectsof tannic acid and quebracho tannins on in vitro ruminalfermentation of wheat and corn grainrdquo Journal of the Scienceof Food and Agriculture vol 86 no 8 pp 1244ndash1256 2006
[46] L Marın E M Miguelez C J Villar and F Lombo ldquoBioavail-ability of dietary polyphenols and gut microbiota metabolismantimicrobial propertiesrdquo BioMed Research International vol2015 Article ID 905215 18 pages 2015
[47] J B Russell and D B Wilson ldquoWhy are ruminal cellulolyticbacteria unable to digest cellulose at low pHrdquo Journal of DairyScience vol 79 no 8 pp 1503ndash1509 1996
[48] K Tajima S Arai K Ogata et al ldquoRumen bacterial communitytransition during adaptation to high-grain dietrdquo Anaerobe vol6 no 5 pp 273ndash284 2000
[49] D W Goad C L Goad and T G Nagaraja ldquoRuminal micro-bial and fermentative changes associated with experimentallyinduced subacute acidosis in steersrdquo Journal of Animal Sciencevol 76 no 1 pp 234ndash241 1998
[50] J Hernandez J L Benedito A Abuelo and C CastilloldquoRuminal acidosis in feedlot from aetiology to preventionrdquoTheScientific World Journal vol 2014 Article ID 702572 2014
[51] A B De Menezes E Lewis M OrsquoDonovan B F OrsquoNeill NClipson and E M Doyle ldquoMicrobiome analysis of dairy cowsfed pasture or total mixed ration dietsrdquo FEMS MicrobiologyEcology vol 78 no 2 pp 256ndash265 2011
[52] I K Skene and J D Brooker ldquoCharacterization of tanninacylhydrolase activity in the ruminal bacterium selenomonasruminantiumrdquo Anaerobe vol 1 no 6 pp 321ndash327 1995
[53] G Goel A K Puniya and K Singh ldquoTannic acid resistance inruminal streptococcal isolatesrdquo Journal of Basic Microbiologyvol 45 no 3 pp 243ndash245 2005
[54] P R Myer J E Wells T P L Smith L A Kuehn and H CFreetly ldquoCecum microbial communities from steers differingin feed efficiencyrdquo Journal of Animal Science vol 93 no 11 pp5327ndash5340 2015
[55] S E Hook A-D GWright and BWMcBride ldquoMethanogensmethane producers of the rumen and mitigation strategiesrdquoArchaea vol 2010 Article ID 945785 11 pages 2010
[56] M Saminathan C C Sieo H M Gan N Abdullah C M VL Wong and Y W Ho ldquoEffects of condensed tannin fractionsof different molecular weights on population and diversity ofbovine rumen methanogenic archaea in vitro as determinedby high-throughput sequencingrdquo Animal Feed Science andTechnology vol 216 pp 146ndash160 2016
[57] F Hassanat and C Benchaar ldquoAssessment of the effect ofcondensed (acacia and quebracho) and hydrolysable (chestnutand valonea) tannins on rumen fermentation andmethane pro-duction in vitrordquo Journal of the Science of Food and Agriculturevol 93 no 2 pp 332ndash339 2013
[58] J Balcells A Aris A Serrano A R Seradj J Crespo and MDevant ldquoEffects of an extract of plant flavonoids (bioflavex)on rumen fermentation and performance in heifers fed high-concentrate dietsrdquo Journal of Animal Science vol 90 no 13 pp4975ndash4984 2012
BioMed Research International 11
[59] A K Patra ldquoUreaammonia metabolism in the rumen andtoxicity in ruminantsrdquo Rumen Microbiology From Evolution toRevolution pp 329ndash341 2015
[60] MAWoznyM P Bryant L VHoldeman andW E CMooreldquoUrease assay and urease producing species of anaerobes in thebovine rumen and human fecesrdquo Applied and EnvironmentalMicrobiology vol 33 no 5 pp 1097ndash1104 1977
[61] D Jin S Zhao P Wang et al ldquoInsights into abundant rumenureolytic bacterial community using rumen simulation systemrdquoFrontiers in Microbiology vol 7 article no 1006 2016
[62] H P SMakkar B Singh andRKDawra ldquoEffect of tannin-richleaves of oak (Quercus incana) on various microbial enzymeactivities of the bovine rumenrdquo British Journal of Nutrition vol60 no 2 pp 287ndash296 1988
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
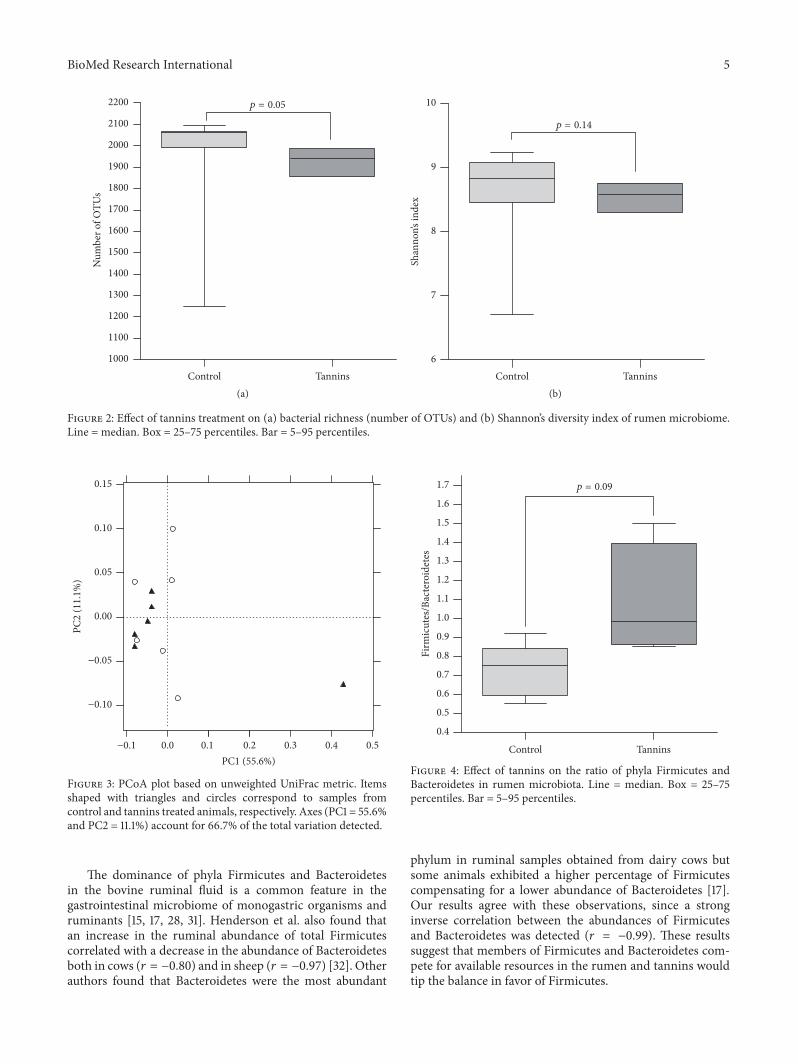
BioMed Research International 5
Control Tannins1000
1100
1200
1300
1400
1500
1600
1700
1800
1900
2000
2100
2200N
umbe
r of O
TUs
p = 005
(a)Control Tannins
6
7
8
9
10
Shan
nonrsquos
inde
x
p = 014
(b)
Figure 2 Effect of tannins treatment on (a) bacterial richness (number of OTUs) and (b) Shannonrsquos diversity index of rumen microbiomeLine = median Box = 25ndash75 percentiles Bar = 5ndash95 percentiles
minus010
minus005
000
005
010
015
PC2
(11
1)
00 01 02 03 04 05minus01PC1 (556)
Figure 3 PCoA plot based on unweighted UniFrac metric Itemsshaped with triangles and circles correspond to samples fromcontrol and tannins treated animals respectively Axes (PC1 = 556and PC2 = 111) account for 667 of the total variation detected
The dominance of phyla Firmicutes and Bacteroidetesin the bovine ruminal fluid is a common feature in thegastrointestinal microbiome of monogastric organisms andruminants [15 17 28 31] Henderson et al also found thatan increase in the ruminal abundance of total Firmicutescorrelated with a decrease in the abundance of Bacteroidetesboth in cows (119903 = minus080) and in sheep (119903 = minus097) [32] Otherauthors found that Bacteroidetes were the most abundant
Control Tannins
p = 009
04
05
06
07
08
09
10
11
12
13
14
15
16
17
Firm
icut
esB
acte
roid
etes
Figure 4 Effect of tannins on the ratio of phyla Firmicutes andBacteroidetes in rumen microbiota Line = median Box = 25ndash75percentiles Bar = 5ndash95 percentiles
phylum in ruminal samples obtained from dairy cows butsome animals exhibited a higher percentage of Firmicutescompensating for a lower abundance of Bacteroidetes [17]Our results agree with these observations since a stronginverse correlation between the abundances of Firmicutesand Bacteroidetes was detected (119903 = minus099) These resultssuggest that members of Firmicutes and Bacteroidetes com-pete for available resources in the rumen and tannins wouldtip the balance in favor of Firmicutes
6 BioMed Research International
lowast
lowastlowast
lowast
lowastlowast
lowast
lowast
lowastlowastlowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
lowast
lowastlowast
lowast
lowast
lowast
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
minus4 minus2 minus1minus3minus5 0 2 3 4 51
Phylum FirmicutesGenus Butyrivibrio
Unc Family RuminococcaceaeGenus Ruminococcus
Genus SucciniclasticumGenus Streptococcus
Genus p-75-a5Genus L7A-E11
Genus SelenomonasGenus Anaerovibrio
Genus BlautiaGenus Anaerofustis
Genus RFN20Genus Lachnospira
Phylum BacteroidetesFamily Prevotellaceae
Family BacteroidaceaeGenus Prevotella
Genus BacteroidesGenus YRC22
Unc Family RF16
Unc Family CoriobacteriaceaePhylum Actinobacteria
Phylum SpirochaetesGenus Treponema
Genus Sphaerochaeta
Phylum EuryarchaeotaGenus Methanobrevibacter
Genus Methanosphaera
Phylum ProteobacteriaUnclass Order GMD14H09
Genus RuminobacterGenus Oxalobacter
Genus Succinivibrio
Phylum TenericutesUnc Family Anaeroplasmata
Genus Anaeroplasma
Phylum ChloroflexiGenus SHD-231
Phylum FibrobacteresGenus Fibrobacter
log 2 fold change
Figure 5 Relative fold changes (log 2 tanninscontrol) in the abundance of rumen bacterial taxa between control and tannins treated steerslowastlowast119901 lt 005 lowast119901 lt 010 Bar = SEM
The ratio of Firmicutes to Bacteroidetes has been shownto affect energy harvesting and body fat accumulation inhumans and mice [14 15] Along with increased fatty acidabsorption more energy was found to be efficiently obtainedfrom diet in obese mice illustrating the connection betweenFirmicutes and improved efficiency in energy harvesting [14]In cows the Firmicutes to Bacteroidetes ratio was found tobe strongly correlated with daily milk-fat yield [17] A recentstudy found that the abundance of Firmicutes in the rumen
positively correlates with the average daily body weight gainin steers suggesting that these bacteria play a significant rolein feed efficiency of bovines [33] Therefore it is possiblethat the increase of Firmicutes to Bacteroidetes ratio inducedby tannins can improve bovine performance as previouslysuggested by other authors [19]
Fiber degradation is a complex process carried out bya group of microorganisms that are able to digest plantpolysaccharides mainly through production of cellulolytic
BioMed Research International 7
Control Tannins50
52
54
56
58
60
62
64
66
68
70pH
p = 002
(a)Control Tannins
000
005
010
015
020
Ure
ase a
ctiv
ity (U
A)
p = 0007
(b)
Control Tannins
000
005
010
015
Non
prot
ein
nitro
gen
(NPN
)
p = 0849
(c)
Figure 6 Effect of tannins on pH urease activity and NPN in the rumen liquor Line = median Box = 25ndash75 percentiles Bar = 5ndash95percentiles Urease activity values are given in pH units proportional to urease activity NPN levels are expressed as a percentage of solublenitrogen in the rumen
hemicellulolytic and pectinolytic enzymes [34] Cellulolyticactivity in the rumen involves a diverse bacterial commu-nity whose main members belong to genera Ruminococcusand Fibrobacter while the main hemicellulolytic bacteriabelong to genera Ruminococcus Prevotella and Butyrivib-rio [34ndash37] The blend of tannins administrated increasedRuminococcaceae and other members of phylum Firmicuteswhile inhibited genera Prevotella and Fibrobacter Prevotellaspecies have a documented role in metabolism of starchhemicellulose pectin and protein catabolism [25] The highabundance of this genus might be the result of a largemetabolic niche that is occupied by different species with
similar metabolic capabilities or it might be associated witha high genetic variability that enables members of genusPrevotella to occupy different ecological niches within therumen [38ndash40] For instance Prevotella abundance has beenshown to be higher in the rumen of animals of beef cattlewith low feed efficiency [41] Previous studies reported a greatdiversity in the sensitivity ofPrevotella species to tannins [42]which could explain the lower abundance detected for thisgenus in tannins-supplemented steers Since genus Prevotellais characterized by a high genetic diversity and wide array ofoutputmetabolites [38ndash40] their replacement byRuminococ-cus may lead to a simpler product profile specialized to
8 BioMed Research International
support the hostrsquos energy requirements [30] Our resultsagree with previous studies that found a significant reduc-tion in Fibrobacter and Prevotella rumen populations aftersupplementation with condensed and hydrolysable tanninsincluding chestnut and quebracho tannins [43 44]
Rumen fibrolytic function is carried out by a largelyredundant microbial community with overlapping distribu-tion of metabolic capabilities and this fibrolytic communityhas the ability to restore its structure and function afterperturbation a phenomenon known as resilience [27] In thiscontext the observed changes in diversity of fibrolytic bacte-ria in tannins treated steers may result from a combinationof physicochemical and biological mechanisms described inthe literature including direct interaction of tannins withfiber [45] which could alter the available surface area formicrobial attack the inhibition of certain fibrolytic taxa bymeans of tannins antimicrobial activity [46] andmodulationof fibrolytic bacterial species driven by changes in rumen pHsince certain fibrolytic taxa of the rumen are inhibited at lowpH under high-grain diets [36 47]
Rumen amylolytic and saccharolytic bacteria were alsoaffected by dietary treatment with tannins mainly throughthe decrease of genera Prevotella and Treponema Otheramylolytic genera were moderately increased in tannins-fedsteers including Streptococcus Bifidobacterium and Lacto-bacillus Amylolytic activity is normally enhanced in rumi-nants consuming high-grain diets [18 48 49] The rate ofgrain degradation by these microbial communities plays akey role in maintaining rumen homeostasis since rapidstarch fermentation produces large amounts of organic acidstherefore producing a drop in ruminal pH that may leadto metabolic acidosis [50] A previous study showed thattannic acid and quebracho tannins lower the rate ofmicrobialhydrolysis of starch-rich grains in the rumen by physicalmodification of the endosperm protein matrix [45] Thisphysical modulation of starch degradation could explain thelower abundance of sugar fermenting taxa detected in tanninstreated steers as well as the higher ruminal pH observed inthis group
Fermentation products of microbial activity in rumenmainly short-chain volatile fatty acids serve as amajor sourceof energy for ruminants and have a direct impact on thephysiological parameters of the animal and feed utilizationefficiency [16] Some members of family Veillonellaceaewhich produce propionate as a major fermentation productand have been associated with lower methane emissions [51]were enhanced in tannins treated animals Interestingly someSelenomonas species can break tannin-protein complexes anduse tannins as energy source [52 53] Therefore the higherabundance of Selenomonas in tannins treated steers may bepartly due to availability of tannins as direct energy sourceMeanwhile members of class Erysipelotrichi which havebeen linked to beef cattle feed efficiency [33 51 54] were alsomodulated by tannins treatment
Methane production during fermentation of feeds in therumen represents a loss of 2ndash12 of gross energy [1] andit is performed by a group of archaea known collectivelyas methanogens which belong to phylum EuryarchaeotaThe microorganisms produce methane the second largest
anthropogenic greenhouse gas which has a global warmingpotential 25 times that of carbon dioxide [55] In the presentstudy a reduction of methanogenic archaea in tannins sup-plemented steers was detected as well as an inverse cor-relation between the abundances of phylum Euryarchaeotaand rumen pH (119903 = minus095) Tannins are thought todirectly inhibit methanogens as well as indirectly limitmethanogenesis through reduction of hydrogen availability[1 55] Saminathan et al found a significant decrease in genusMethanobrevibacter after treatment of rumen samples withcondensed tannins in vitro [56] Other authors describeda linear decrease of genus Methanobrevibacter after dietarysupplementation with condensed tannins from pine bark ingoats [31]
Ruminal pH was significantly higher in tannins treatedsteers than in the control group while urease activity exhib-ited the opposite pattern These results agree with previousreports which found an increase in ruminal pH after sup-plementation with chestnut and quebracho tannins [57 58]Other authors observed that feeding chestnut and quebrachotannins decreased urease activity in the faeces of cows[10 11]
Feed-grade urea is an effective source of nitrogen com-monly used in beef cattle diets Ureolytic bacteria in therumen produce urease to hydrolyse urea to ammonia whichis subsequently used for the synthesis of amino acids andmicrobial protein Normally the rate of urea hydrolysisexceeds the rate of ammonia utilization which leads to poorefficiency of urea utilization and increases toxic ammoniaconcentrations in blood [59] Ureolytic bacteria in the rumencomprise a highly diverse group whose main species belongto genera Succinivibrio Treponema Bacteroides ButyrivibrioStreptococcus andBifidobacterium [60 61]We found that themost abundant ureolytic generaButyrivibrio andTreponemawere negatively affected by tannins treatment Thus theobserved decline in ruminal urease activity may be relatedwith the decrease of these urease-producing taxa A previousstudy also observed a drop in ruminal urease activity afteraddition of tannins to diet but a direct interaction betweentannins and urease enzyme was postulated as responsible forthis inhibition [62]
5 Conclusions
The current study showed that chestnut and quebrachotannins added to the diet of Holstein steers modifiedrumen microbiota composition particularly fiber and starchdegrading bacteria mainly by reducing the abundanceof Prevotella and Fibrobacter while favoring Ruminococ-caceae and other members of phylum Firmicutes Tanninstreatment significantly increased pH and decreased ureaseactivity in the ruminal liquor Further work is necessaryto assess the possible relation between tannins ability tomodify rumen microbiota composition and the alterationof rumen fermentation parameters associated with energyand feed efficiency of beef cattle such as the profile ofshort-chain fatty acids and the emissions of ammonia andmethane
BioMed Research International 9
Conflicts of Interest
The authors declare that there are no known conflicts ofinterest associated with this publication and there has beenno financial support for this work that could have influencedits outcome
Acknowledgments
The authors thank Alfredo Ivan Martınez Caceres for histechnical assistance during installation and startup of thebioinformatics tools used in this study This work wassupported by grants from INTA (PNSA 1115052 and PNBIO1131043) Additional support was provided by CONICET
References
[1] A K Patra and J Saxena ldquoDietary phytochemicals as rumenmodifiers a review of the effects on microbial populationsrdquoInternational Journal of General and Molecular Microbiologyvol 96 no 4 pp 363ndash375 2009 Antonie Van Leeuwenhoek
[2] R Bodas N Prieto R Garcıa-Gonzalez S Andres F JGiraldez and S Lopez ldquoManipulation of rumen fermentationand methane production with plant secondary metabolitesrdquoAnimal Feed Science and Technology vol 176 no 1-4 pp 78ndash932012
[3] E Oskoueian N Abdullah and A Oskoueian ldquoEffects offlavonoids on rumen fermentation activity methane produc-tion andmicrobial populationrdquo BioMed Research Internationalvol 2013 Article ID 349129 8 pages 2013
[4] P Frutos GHervas F J Giraldez andA RMantecon ldquoReviewtannins and ruminant nutritionrdquo Spanish Journal of AgriculturalResearch vol 2 no 2 pp 191ndash202 2004
[5] K Khanbabaee and T van Ree ldquoTannins classification anddefinitionrdquo Natural Product Reports vol 18 no 6 pp 641ndash6492001
[6] L J Porter ldquoStructure and chemical properties of the con-densed tanninsrdquo Plant Polyphenols pp 245ndash258 1992 Springer Boston MA USA
[7] B R Min C Wright P Ho J-S Eun N Gurung and RShange ldquoThe effect of phytochemical tannins-containing dieton rumen fermentation characteristics and microbial diversitydynamics in goats using 16S rDNA amplicon pyrosequencingrdquoAgriculture Food and Analytical Bacteriology vol 4 no 3 pp195ndash211 2014
[8] B S Gemeda and A Hassen ldquoEffect of tannin and speciesvariation on in vitro digestibility gas and methane productionof tropical browse plantsrdquoAsian-Australasian Journal of AnimalSciences vol 28 no 2 pp 188ndash199 2015
[9] Z Li A-D G Wright H Liu et al ldquoResponse of the rumenmicrobiota of sika deer (cervus nippon) fed different concen-trations of tannin rich plantsrdquo PLoS ONE vol 10 no 5 ArticleID e0123481 2015
[10] JM PowelM J Aguerre andMAWattiaux ldquoTannin extractsabate ammonia emissions from simulated dairy barn floorsrdquoJournal of Environmental Quality vol 40 no 3 pp 907ndash9142011
[11] M J Aguerre M C Capozzolo P Lencioni C Cabral andM A Wattiaux ldquoEffect of quebracho-chestnut tannin extractsat 2 dietary crude protein levels on performance rumenfermentation and nitrogen partitioning in dairy cowsrdquo Journal
of Dairy Science vol 99 no 6 Article ID 74473 pp 4476ndash44862016
[12] C Rivera-Mendez A Plascencia N Torrentera andR A ZinnldquoEffect of level and source of supplemental tannin on growthperformance of steers during the late finishing phaserdquo Journalof Applied Animal Research vol 45 pp 199ndash203 2017
[13] B R Min T N Barry G T Attwood and W C McNabbldquoThe effect of condensed tannins on the nutrition and health ofruminants fed fresh temperate forages a reviewrdquo Animal FeedScience and Technology vol 106 no 1-4 pp 3ndash19 2003
[14] P J Turnbaugh R E Ley M A Mahowald V MagriniE R Mardis and J I Gordon ldquoAn obesity-associated gutmicrobiomewith increased capacity for energy harvestrdquoNaturevol 444 no 7122 pp 1027ndash1031 2006
[15] R E Ley R Knight and J I Gordon ldquoThe human micro-biome Eliminating the biomedicalenvironmental dichotomyin microbial ecologyrdquo Environmental Microbiology vol 9 no 1pp 3-4 2007
[16] I Mizrahi ldquoThe role of the rumen microbiota in determiningthe feed efficiency of dairy cowsrdquo Beneficial Microorganismsin Multicellular Life Forms pp 203ndash210 2012 Springer BerlinHeidelberg
[17] E Jami B AWhite and IMizrahi ldquoPotential role of the bovinerumen microbiome in modulating milk composition and feedefficiencyrdquo PLoS ONE vol 9 no 1 Article ID e85423 2014
[18] S C Fernando H T Purvis F Z Najar et al ldquoRumenmicrobialpopulation dynamics during adaptation to a high-grain dietrdquoApplied and Environmental Microbiology vol 76 no 22 pp7482ndash7490 2010
[19] H P S Makkar ldquoEffects and fate of tannins in ruminantanimals adaptation to tannins and strategies to overcomedetrimental effects of feeding tannin-rich feedsrdquo Small Rumi-nant Research vol 49 no 3 pp 241ndash256 2003
[20] Illumina Protocol 16S Metagenomic Sequencing Library 2013[21] AM BolgerM Lohse and B Usadel ldquoTrimmomatic a flexible
trimmer for Illumina sequence datardquoBioinformatics vol 30 no15 pp 2114ndash2120 2014
[22] J Kuczynski J Stombaugh W A Walters A Gonzalez J GCaporaso and R Knight ldquoUsing QIIME to analyze 16S rRNAgene sequences frommicrobial communitiesrdquoCurrent Protocolsin Microbiology vol 1 1E5 pages 2012
[23] N A Bokulich S Subramanian J J Faith et al ldquoQuality-filtering vastly improves diversity estimates from Illuminaamplicon sequencingrdquo Nature Methods vol 10 no 1 pp 57ndash592013
[24] D Parks and R Beiko ldquoSTAMP statistical analysis of metage-nomic profilesrdquo Encyclopedia of Metagenomics pp 1ndash6 2013Springer New York NY USA
[25] J C McCann T A Wickersham and J J Loor ldquoHigh-throughput methods redefine the rumen microbiome and itsrelationshipwith nutrition andmetabolismrdquoBioinformatics andBiology Insights vol 8 pp 109ndash125 2014
[26] G Henderson F Cox S Ganesh A Jonker W Young and PH Janssen ldquoRumen microbial community composition varieswith diet and host but a coremicrobiome is found across a widegeographical rangerdquo Scientific Reports vol 5 no 14567 pp 1ndash132015
[27] P J Weimer ldquoRedundancy resilience and host specificity ofthe ruminal microbiota Implications for engineering improvedruminal fermentationsrdquo Frontiers in Microbiology vol 6 articleno 296 2015
10 BioMed Research International
[28] E Jami and I Mizrahi ldquoComposition and similarity of bovinerumenmicrobiota across individual animalsrdquo PLoS ONE vol 7no 3 Article ID e33306 2012
[29] R De Nardi G Marchesini S Li et al ldquoMetagenomic analysisof rumen microbial population in dairy heifers fed a high graindiet supplemented with dicarboxylic acids or polyphenolsrdquoBMC Veterinary Research vol 12 no 1 article no 29 2016
[30] S Kruger Ben Shabat G Sasson A Doron-Faigenboim etal ldquoSpecific microbiome-dependent mechanisms underlie theenergy harvest efficiency of ruminantsrdquoThe ISME Journal vol10 no 12 pp 2958ndash2972 2016
[31] B R Min S Solaiman R Shange and J-S Eun ldquoGastroin-testinal bacterial andmethanogenic archaea diversity dynamicsassociated with condensed tannin-containing pine bark diet ingoats using 16S rDNA amplicon pyrosequencingrdquo InternationalJournal of Microbiology vol 2014 Article ID 141909 2014
[32] G Henderson F Cox S Kittelmann et al ldquoEffect of DNAextraction methods and sampling techniques on the apparentstructure of cow and sheep rumen microbial communitiesrdquoPLoS ONE vol 8 no 9 Article ID e74787 2013
[33] P R Myer T P L Smith J E Wells L A Kuehn and HC Freetly ldquoRumen microbiome from steers differing in feedefficiencyrdquo PLoS ONE vol 10 no 6 Article ID e0129174 2015
[34] Y Wang and T A McAllister ldquoRumen microbes enzymes andfeed digestion-a reviewrdquo Asian-Australasian Journal of AnimalSciences vol 15 no 11 pp 1659ndash1676 2002
[35] J B Russell R EMuck and P JWeimer ldquoQuantitative analysisof cellulose degradation and growth of cellulolytic bacteria inthe rumenrdquo FEMS Microbiology Ecology vol 67 no 2 pp 183ndash197 2009
[36] DOKrause S EDenmanR IMackie et al ldquoOpportunities toimprove fiber degradation in the rumen microbiology ecologyand genomicsrdquo FEMS Microbiology Reviews vol 27 no 5 pp663ndash693 2003
[37] M Zorec M Vodovnik and R Marinsek-Logar ldquoPotentialof selected rumen bacteria for cellulose and hemicellulosedegradationrdquo Food Technology and Biotechnology vol 52 no 2pp 210ndash221 2014
[38] A Z Bekele S Koike and Y Kobayashi ldquoGenetic diversityand diet specificity of ruminal prevotella revealed by 16S rRNAgene-based analysisrdquo FEMSMicrobiology Letters vol 305 no 1pp 49ndash57 2010
[39] A RamsakM Peterka K Tajima et al ldquoUnravelling the geneticdiversity of ruminal bacteria belonging to the CFB phylumrdquoFEMS Microbiology Ecology vol 33 no 1 pp 69ndash79 2000
[40] J Purushe D E Fouts M Morrison et al ldquoComparativegenome analysis of prevotella ruminicola and prevotella bryan-tii insights into their environmental nicherdquo Microbial Ecologyvol 60 no 4 pp 721ndash729 2010
[41] C A Carberry D A Kenny S Han M S McCabe and S MWaters ldquoEffect of phenotypic residual feed intake and dietaryforage content on the rumen microbial community of beefcattlerdquo Applied and Environmental Microbiology vol 78 no 14pp 4949ndash4958 2012
[42] A L Molan G T Attwood B R Min and W C McNabbldquoThe effect of condensed tannins from Lotus pedunculatus andLotus corniculatus on the growth of proteolytic rumen bacteriain vitro and their possible mode of actionrdquo Canadian Journal ofMicrobiology vol 47 no 7 pp 626ndash633 2001
[43] A Jayanegara G Goel H P S Makkar and K Becker ldquoDiver-gence between purified hydrolysable and condensed tannin
effects on methane emission rumen fermentation and micro-bial population in vitrordquo Animal Feed Science and Technologyvol 209 pp 60ndash68 2015
[44] BMinW Pinchak K Hernandez et al ldquoEffects of plant tanninsupplementation on animal responses and in vivo ruminalbacterial populations associated with bloat in heifers grazingwheat forage1rdquo The Professional Animal Scientist vol 28 no 4pp 464ndash472 2012
[45] T F Martınez T A McAllister Y Wang and T Reuter ldquoEffectsof tannic acid and quebracho tannins on in vitro ruminalfermentation of wheat and corn grainrdquo Journal of the Scienceof Food and Agriculture vol 86 no 8 pp 1244ndash1256 2006
[46] L Marın E M Miguelez C J Villar and F Lombo ldquoBioavail-ability of dietary polyphenols and gut microbiota metabolismantimicrobial propertiesrdquo BioMed Research International vol2015 Article ID 905215 18 pages 2015
[47] J B Russell and D B Wilson ldquoWhy are ruminal cellulolyticbacteria unable to digest cellulose at low pHrdquo Journal of DairyScience vol 79 no 8 pp 1503ndash1509 1996
[48] K Tajima S Arai K Ogata et al ldquoRumen bacterial communitytransition during adaptation to high-grain dietrdquo Anaerobe vol6 no 5 pp 273ndash284 2000
[49] D W Goad C L Goad and T G Nagaraja ldquoRuminal micro-bial and fermentative changes associated with experimentallyinduced subacute acidosis in steersrdquo Journal of Animal Sciencevol 76 no 1 pp 234ndash241 1998
[50] J Hernandez J L Benedito A Abuelo and C CastilloldquoRuminal acidosis in feedlot from aetiology to preventionrdquoTheScientific World Journal vol 2014 Article ID 702572 2014
[51] A B De Menezes E Lewis M OrsquoDonovan B F OrsquoNeill NClipson and E M Doyle ldquoMicrobiome analysis of dairy cowsfed pasture or total mixed ration dietsrdquo FEMS MicrobiologyEcology vol 78 no 2 pp 256ndash265 2011
[52] I K Skene and J D Brooker ldquoCharacterization of tanninacylhydrolase activity in the ruminal bacterium selenomonasruminantiumrdquo Anaerobe vol 1 no 6 pp 321ndash327 1995
[53] G Goel A K Puniya and K Singh ldquoTannic acid resistance inruminal streptococcal isolatesrdquo Journal of Basic Microbiologyvol 45 no 3 pp 243ndash245 2005
[54] P R Myer J E Wells T P L Smith L A Kuehn and H CFreetly ldquoCecum microbial communities from steers differingin feed efficiencyrdquo Journal of Animal Science vol 93 no 11 pp5327ndash5340 2015
[55] S E Hook A-D GWright and BWMcBride ldquoMethanogensmethane producers of the rumen and mitigation strategiesrdquoArchaea vol 2010 Article ID 945785 11 pages 2010
[56] M Saminathan C C Sieo H M Gan N Abdullah C M VL Wong and Y W Ho ldquoEffects of condensed tannin fractionsof different molecular weights on population and diversity ofbovine rumen methanogenic archaea in vitro as determinedby high-throughput sequencingrdquo Animal Feed Science andTechnology vol 216 pp 146ndash160 2016
[57] F Hassanat and C Benchaar ldquoAssessment of the effect ofcondensed (acacia and quebracho) and hydrolysable (chestnutand valonea) tannins on rumen fermentation andmethane pro-duction in vitrordquo Journal of the Science of Food and Agriculturevol 93 no 2 pp 332ndash339 2013
[58] J Balcells A Aris A Serrano A R Seradj J Crespo and MDevant ldquoEffects of an extract of plant flavonoids (bioflavex)on rumen fermentation and performance in heifers fed high-concentrate dietsrdquo Journal of Animal Science vol 90 no 13 pp4975ndash4984 2012
BioMed Research International 11
[59] A K Patra ldquoUreaammonia metabolism in the rumen andtoxicity in ruminantsrdquo Rumen Microbiology From Evolution toRevolution pp 329ndash341 2015
[60] MAWoznyM P Bryant L VHoldeman andW E CMooreldquoUrease assay and urease producing species of anaerobes in thebovine rumen and human fecesrdquo Applied and EnvironmentalMicrobiology vol 33 no 5 pp 1097ndash1104 1977
[61] D Jin S Zhao P Wang et al ldquoInsights into abundant rumenureolytic bacterial community using rumen simulation systemrdquoFrontiers in Microbiology vol 7 article no 1006 2016
[62] H P SMakkar B Singh andRKDawra ldquoEffect of tannin-richleaves of oak (Quercus incana) on various microbial enzymeactivities of the bovine rumenrdquo British Journal of Nutrition vol60 no 2 pp 287ndash296 1988
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 BioMed Research International
lowast
lowastlowast
lowast
lowastlowast
lowast
lowast
lowastlowastlowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
lowast
lowastlowast
lowast
lowast
lowast
lowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
minus4 minus2 minus1minus3minus5 0 2 3 4 51
Phylum FirmicutesGenus Butyrivibrio
Unc Family RuminococcaceaeGenus Ruminococcus
Genus SucciniclasticumGenus Streptococcus
Genus p-75-a5Genus L7A-E11
Genus SelenomonasGenus Anaerovibrio
Genus BlautiaGenus Anaerofustis
Genus RFN20Genus Lachnospira
Phylum BacteroidetesFamily Prevotellaceae
Family BacteroidaceaeGenus Prevotella
Genus BacteroidesGenus YRC22
Unc Family RF16
Unc Family CoriobacteriaceaePhylum Actinobacteria
Phylum SpirochaetesGenus Treponema
Genus Sphaerochaeta
Phylum EuryarchaeotaGenus Methanobrevibacter
Genus Methanosphaera
Phylum ProteobacteriaUnclass Order GMD14H09
Genus RuminobacterGenus Oxalobacter
Genus Succinivibrio
Phylum TenericutesUnc Family Anaeroplasmata
Genus Anaeroplasma
Phylum ChloroflexiGenus SHD-231
Phylum FibrobacteresGenus Fibrobacter
log 2 fold change
Figure 5 Relative fold changes (log 2 tanninscontrol) in the abundance of rumen bacterial taxa between control and tannins treated steerslowastlowast119901 lt 005 lowast119901 lt 010 Bar = SEM
The ratio of Firmicutes to Bacteroidetes has been shownto affect energy harvesting and body fat accumulation inhumans and mice [14 15] Along with increased fatty acidabsorption more energy was found to be efficiently obtainedfrom diet in obese mice illustrating the connection betweenFirmicutes and improved efficiency in energy harvesting [14]In cows the Firmicutes to Bacteroidetes ratio was found tobe strongly correlated with daily milk-fat yield [17] A recentstudy found that the abundance of Firmicutes in the rumen
positively correlates with the average daily body weight gainin steers suggesting that these bacteria play a significant rolein feed efficiency of bovines [33] Therefore it is possiblethat the increase of Firmicutes to Bacteroidetes ratio inducedby tannins can improve bovine performance as previouslysuggested by other authors [19]
Fiber degradation is a complex process carried out bya group of microorganisms that are able to digest plantpolysaccharides mainly through production of cellulolytic
BioMed Research International 7
Control Tannins50
52
54
56
58
60
62
64
66
68
70pH
p = 002
(a)Control Tannins
000
005
010
015
020
Ure
ase a
ctiv
ity (U
A)
p = 0007
(b)
Control Tannins
000
005
010
015
Non
prot
ein
nitro
gen
(NPN
)
p = 0849
(c)
Figure 6 Effect of tannins on pH urease activity and NPN in the rumen liquor Line = median Box = 25ndash75 percentiles Bar = 5ndash95percentiles Urease activity values are given in pH units proportional to urease activity NPN levels are expressed as a percentage of solublenitrogen in the rumen
hemicellulolytic and pectinolytic enzymes [34] Cellulolyticactivity in the rumen involves a diverse bacterial commu-nity whose main members belong to genera Ruminococcusand Fibrobacter while the main hemicellulolytic bacteriabelong to genera Ruminococcus Prevotella and Butyrivib-rio [34ndash37] The blend of tannins administrated increasedRuminococcaceae and other members of phylum Firmicuteswhile inhibited genera Prevotella and Fibrobacter Prevotellaspecies have a documented role in metabolism of starchhemicellulose pectin and protein catabolism [25] The highabundance of this genus might be the result of a largemetabolic niche that is occupied by different species with
similar metabolic capabilities or it might be associated witha high genetic variability that enables members of genusPrevotella to occupy different ecological niches within therumen [38ndash40] For instance Prevotella abundance has beenshown to be higher in the rumen of animals of beef cattlewith low feed efficiency [41] Previous studies reported a greatdiversity in the sensitivity ofPrevotella species to tannins [42]which could explain the lower abundance detected for thisgenus in tannins-supplemented steers Since genus Prevotellais characterized by a high genetic diversity and wide array ofoutputmetabolites [38ndash40] their replacement byRuminococ-cus may lead to a simpler product profile specialized to
8 BioMed Research International
support the hostrsquos energy requirements [30] Our resultsagree with previous studies that found a significant reduc-tion in Fibrobacter and Prevotella rumen populations aftersupplementation with condensed and hydrolysable tanninsincluding chestnut and quebracho tannins [43 44]
Rumen fibrolytic function is carried out by a largelyredundant microbial community with overlapping distribu-tion of metabolic capabilities and this fibrolytic communityhas the ability to restore its structure and function afterperturbation a phenomenon known as resilience [27] In thiscontext the observed changes in diversity of fibrolytic bacte-ria in tannins treated steers may result from a combinationof physicochemical and biological mechanisms described inthe literature including direct interaction of tannins withfiber [45] which could alter the available surface area formicrobial attack the inhibition of certain fibrolytic taxa bymeans of tannins antimicrobial activity [46] andmodulationof fibrolytic bacterial species driven by changes in rumen pHsince certain fibrolytic taxa of the rumen are inhibited at lowpH under high-grain diets [36 47]
Rumen amylolytic and saccharolytic bacteria were alsoaffected by dietary treatment with tannins mainly throughthe decrease of genera Prevotella and Treponema Otheramylolytic genera were moderately increased in tannins-fedsteers including Streptococcus Bifidobacterium and Lacto-bacillus Amylolytic activity is normally enhanced in rumi-nants consuming high-grain diets [18 48 49] The rate ofgrain degradation by these microbial communities plays akey role in maintaining rumen homeostasis since rapidstarch fermentation produces large amounts of organic acidstherefore producing a drop in ruminal pH that may leadto metabolic acidosis [50] A previous study showed thattannic acid and quebracho tannins lower the rate ofmicrobialhydrolysis of starch-rich grains in the rumen by physicalmodification of the endosperm protein matrix [45] Thisphysical modulation of starch degradation could explain thelower abundance of sugar fermenting taxa detected in tanninstreated steers as well as the higher ruminal pH observed inthis group
Fermentation products of microbial activity in rumenmainly short-chain volatile fatty acids serve as amajor sourceof energy for ruminants and have a direct impact on thephysiological parameters of the animal and feed utilizationefficiency [16] Some members of family Veillonellaceaewhich produce propionate as a major fermentation productand have been associated with lower methane emissions [51]were enhanced in tannins treated animals Interestingly someSelenomonas species can break tannin-protein complexes anduse tannins as energy source [52 53] Therefore the higherabundance of Selenomonas in tannins treated steers may bepartly due to availability of tannins as direct energy sourceMeanwhile members of class Erysipelotrichi which havebeen linked to beef cattle feed efficiency [33 51 54] were alsomodulated by tannins treatment
Methane production during fermentation of feeds in therumen represents a loss of 2ndash12 of gross energy [1] andit is performed by a group of archaea known collectivelyas methanogens which belong to phylum EuryarchaeotaThe microorganisms produce methane the second largest
anthropogenic greenhouse gas which has a global warmingpotential 25 times that of carbon dioxide [55] In the presentstudy a reduction of methanogenic archaea in tannins sup-plemented steers was detected as well as an inverse cor-relation between the abundances of phylum Euryarchaeotaand rumen pH (119903 = minus095) Tannins are thought todirectly inhibit methanogens as well as indirectly limitmethanogenesis through reduction of hydrogen availability[1 55] Saminathan et al found a significant decrease in genusMethanobrevibacter after treatment of rumen samples withcondensed tannins in vitro [56] Other authors describeda linear decrease of genus Methanobrevibacter after dietarysupplementation with condensed tannins from pine bark ingoats [31]
Ruminal pH was significantly higher in tannins treatedsteers than in the control group while urease activity exhib-ited the opposite pattern These results agree with previousreports which found an increase in ruminal pH after sup-plementation with chestnut and quebracho tannins [57 58]Other authors observed that feeding chestnut and quebrachotannins decreased urease activity in the faeces of cows[10 11]
Feed-grade urea is an effective source of nitrogen com-monly used in beef cattle diets Ureolytic bacteria in therumen produce urease to hydrolyse urea to ammonia whichis subsequently used for the synthesis of amino acids andmicrobial protein Normally the rate of urea hydrolysisexceeds the rate of ammonia utilization which leads to poorefficiency of urea utilization and increases toxic ammoniaconcentrations in blood [59] Ureolytic bacteria in the rumencomprise a highly diverse group whose main species belongto genera Succinivibrio Treponema Bacteroides ButyrivibrioStreptococcus andBifidobacterium [60 61]We found that themost abundant ureolytic generaButyrivibrio andTreponemawere negatively affected by tannins treatment Thus theobserved decline in ruminal urease activity may be relatedwith the decrease of these urease-producing taxa A previousstudy also observed a drop in ruminal urease activity afteraddition of tannins to diet but a direct interaction betweentannins and urease enzyme was postulated as responsible forthis inhibition [62]
5 Conclusions
The current study showed that chestnut and quebrachotannins added to the diet of Holstein steers modifiedrumen microbiota composition particularly fiber and starchdegrading bacteria mainly by reducing the abundanceof Prevotella and Fibrobacter while favoring Ruminococ-caceae and other members of phylum Firmicutes Tanninstreatment significantly increased pH and decreased ureaseactivity in the ruminal liquor Further work is necessaryto assess the possible relation between tannins ability tomodify rumen microbiota composition and the alterationof rumen fermentation parameters associated with energyand feed efficiency of beef cattle such as the profile ofshort-chain fatty acids and the emissions of ammonia andmethane
BioMed Research International 9
Conflicts of Interest
The authors declare that there are no known conflicts ofinterest associated with this publication and there has beenno financial support for this work that could have influencedits outcome
Acknowledgments
The authors thank Alfredo Ivan Martınez Caceres for histechnical assistance during installation and startup of thebioinformatics tools used in this study This work wassupported by grants from INTA (PNSA 1115052 and PNBIO1131043) Additional support was provided by CONICET
References
[1] A K Patra and J Saxena ldquoDietary phytochemicals as rumenmodifiers a review of the effects on microbial populationsrdquoInternational Journal of General and Molecular Microbiologyvol 96 no 4 pp 363ndash375 2009 Antonie Van Leeuwenhoek
[2] R Bodas N Prieto R Garcıa-Gonzalez S Andres F JGiraldez and S Lopez ldquoManipulation of rumen fermentationand methane production with plant secondary metabolitesrdquoAnimal Feed Science and Technology vol 176 no 1-4 pp 78ndash932012
[3] E Oskoueian N Abdullah and A Oskoueian ldquoEffects offlavonoids on rumen fermentation activity methane produc-tion andmicrobial populationrdquo BioMed Research Internationalvol 2013 Article ID 349129 8 pages 2013
[4] P Frutos GHervas F J Giraldez andA RMantecon ldquoReviewtannins and ruminant nutritionrdquo Spanish Journal of AgriculturalResearch vol 2 no 2 pp 191ndash202 2004
[5] K Khanbabaee and T van Ree ldquoTannins classification anddefinitionrdquo Natural Product Reports vol 18 no 6 pp 641ndash6492001
[6] L J Porter ldquoStructure and chemical properties of the con-densed tanninsrdquo Plant Polyphenols pp 245ndash258 1992 Springer Boston MA USA
[7] B R Min C Wright P Ho J-S Eun N Gurung and RShange ldquoThe effect of phytochemical tannins-containing dieton rumen fermentation characteristics and microbial diversitydynamics in goats using 16S rDNA amplicon pyrosequencingrdquoAgriculture Food and Analytical Bacteriology vol 4 no 3 pp195ndash211 2014
[8] B S Gemeda and A Hassen ldquoEffect of tannin and speciesvariation on in vitro digestibility gas and methane productionof tropical browse plantsrdquoAsian-Australasian Journal of AnimalSciences vol 28 no 2 pp 188ndash199 2015
[9] Z Li A-D G Wright H Liu et al ldquoResponse of the rumenmicrobiota of sika deer (cervus nippon) fed different concen-trations of tannin rich plantsrdquo PLoS ONE vol 10 no 5 ArticleID e0123481 2015
[10] JM PowelM J Aguerre andMAWattiaux ldquoTannin extractsabate ammonia emissions from simulated dairy barn floorsrdquoJournal of Environmental Quality vol 40 no 3 pp 907ndash9142011
[11] M J Aguerre M C Capozzolo P Lencioni C Cabral andM A Wattiaux ldquoEffect of quebracho-chestnut tannin extractsat 2 dietary crude protein levels on performance rumenfermentation and nitrogen partitioning in dairy cowsrdquo Journal
of Dairy Science vol 99 no 6 Article ID 74473 pp 4476ndash44862016
[12] C Rivera-Mendez A Plascencia N Torrentera andR A ZinnldquoEffect of level and source of supplemental tannin on growthperformance of steers during the late finishing phaserdquo Journalof Applied Animal Research vol 45 pp 199ndash203 2017
[13] B R Min T N Barry G T Attwood and W C McNabbldquoThe effect of condensed tannins on the nutrition and health ofruminants fed fresh temperate forages a reviewrdquo Animal FeedScience and Technology vol 106 no 1-4 pp 3ndash19 2003
[14] P J Turnbaugh R E Ley M A Mahowald V MagriniE R Mardis and J I Gordon ldquoAn obesity-associated gutmicrobiomewith increased capacity for energy harvestrdquoNaturevol 444 no 7122 pp 1027ndash1031 2006
[15] R E Ley R Knight and J I Gordon ldquoThe human micro-biome Eliminating the biomedicalenvironmental dichotomyin microbial ecologyrdquo Environmental Microbiology vol 9 no 1pp 3-4 2007
[16] I Mizrahi ldquoThe role of the rumen microbiota in determiningthe feed efficiency of dairy cowsrdquo Beneficial Microorganismsin Multicellular Life Forms pp 203ndash210 2012 Springer BerlinHeidelberg
[17] E Jami B AWhite and IMizrahi ldquoPotential role of the bovinerumen microbiome in modulating milk composition and feedefficiencyrdquo PLoS ONE vol 9 no 1 Article ID e85423 2014
[18] S C Fernando H T Purvis F Z Najar et al ldquoRumenmicrobialpopulation dynamics during adaptation to a high-grain dietrdquoApplied and Environmental Microbiology vol 76 no 22 pp7482ndash7490 2010
[19] H P S Makkar ldquoEffects and fate of tannins in ruminantanimals adaptation to tannins and strategies to overcomedetrimental effects of feeding tannin-rich feedsrdquo Small Rumi-nant Research vol 49 no 3 pp 241ndash256 2003
[20] Illumina Protocol 16S Metagenomic Sequencing Library 2013[21] AM BolgerM Lohse and B Usadel ldquoTrimmomatic a flexible
trimmer for Illumina sequence datardquoBioinformatics vol 30 no15 pp 2114ndash2120 2014
[22] J Kuczynski J Stombaugh W A Walters A Gonzalez J GCaporaso and R Knight ldquoUsing QIIME to analyze 16S rRNAgene sequences frommicrobial communitiesrdquoCurrent Protocolsin Microbiology vol 1 1E5 pages 2012
[23] N A Bokulich S Subramanian J J Faith et al ldquoQuality-filtering vastly improves diversity estimates from Illuminaamplicon sequencingrdquo Nature Methods vol 10 no 1 pp 57ndash592013
[24] D Parks and R Beiko ldquoSTAMP statistical analysis of metage-nomic profilesrdquo Encyclopedia of Metagenomics pp 1ndash6 2013Springer New York NY USA
[25] J C McCann T A Wickersham and J J Loor ldquoHigh-throughput methods redefine the rumen microbiome and itsrelationshipwith nutrition andmetabolismrdquoBioinformatics andBiology Insights vol 8 pp 109ndash125 2014
[26] G Henderson F Cox S Ganesh A Jonker W Young and PH Janssen ldquoRumen microbial community composition varieswith diet and host but a coremicrobiome is found across a widegeographical rangerdquo Scientific Reports vol 5 no 14567 pp 1ndash132015
[27] P J Weimer ldquoRedundancy resilience and host specificity ofthe ruminal microbiota Implications for engineering improvedruminal fermentationsrdquo Frontiers in Microbiology vol 6 articleno 296 2015
10 BioMed Research International
[28] E Jami and I Mizrahi ldquoComposition and similarity of bovinerumenmicrobiota across individual animalsrdquo PLoS ONE vol 7no 3 Article ID e33306 2012
[29] R De Nardi G Marchesini S Li et al ldquoMetagenomic analysisof rumen microbial population in dairy heifers fed a high graindiet supplemented with dicarboxylic acids or polyphenolsrdquoBMC Veterinary Research vol 12 no 1 article no 29 2016
[30] S Kruger Ben Shabat G Sasson A Doron-Faigenboim etal ldquoSpecific microbiome-dependent mechanisms underlie theenergy harvest efficiency of ruminantsrdquoThe ISME Journal vol10 no 12 pp 2958ndash2972 2016
[31] B R Min S Solaiman R Shange and J-S Eun ldquoGastroin-testinal bacterial andmethanogenic archaea diversity dynamicsassociated with condensed tannin-containing pine bark diet ingoats using 16S rDNA amplicon pyrosequencingrdquo InternationalJournal of Microbiology vol 2014 Article ID 141909 2014
[32] G Henderson F Cox S Kittelmann et al ldquoEffect of DNAextraction methods and sampling techniques on the apparentstructure of cow and sheep rumen microbial communitiesrdquoPLoS ONE vol 8 no 9 Article ID e74787 2013
[33] P R Myer T P L Smith J E Wells L A Kuehn and HC Freetly ldquoRumen microbiome from steers differing in feedefficiencyrdquo PLoS ONE vol 10 no 6 Article ID e0129174 2015
[34] Y Wang and T A McAllister ldquoRumen microbes enzymes andfeed digestion-a reviewrdquo Asian-Australasian Journal of AnimalSciences vol 15 no 11 pp 1659ndash1676 2002
[35] J B Russell R EMuck and P JWeimer ldquoQuantitative analysisof cellulose degradation and growth of cellulolytic bacteria inthe rumenrdquo FEMS Microbiology Ecology vol 67 no 2 pp 183ndash197 2009
[36] DOKrause S EDenmanR IMackie et al ldquoOpportunities toimprove fiber degradation in the rumen microbiology ecologyand genomicsrdquo FEMS Microbiology Reviews vol 27 no 5 pp663ndash693 2003
[37] M Zorec M Vodovnik and R Marinsek-Logar ldquoPotentialof selected rumen bacteria for cellulose and hemicellulosedegradationrdquo Food Technology and Biotechnology vol 52 no 2pp 210ndash221 2014
[38] A Z Bekele S Koike and Y Kobayashi ldquoGenetic diversityand diet specificity of ruminal prevotella revealed by 16S rRNAgene-based analysisrdquo FEMSMicrobiology Letters vol 305 no 1pp 49ndash57 2010
[39] A RamsakM Peterka K Tajima et al ldquoUnravelling the geneticdiversity of ruminal bacteria belonging to the CFB phylumrdquoFEMS Microbiology Ecology vol 33 no 1 pp 69ndash79 2000
[40] J Purushe D E Fouts M Morrison et al ldquoComparativegenome analysis of prevotella ruminicola and prevotella bryan-tii insights into their environmental nicherdquo Microbial Ecologyvol 60 no 4 pp 721ndash729 2010
[41] C A Carberry D A Kenny S Han M S McCabe and S MWaters ldquoEffect of phenotypic residual feed intake and dietaryforage content on the rumen microbial community of beefcattlerdquo Applied and Environmental Microbiology vol 78 no 14pp 4949ndash4958 2012
[42] A L Molan G T Attwood B R Min and W C McNabbldquoThe effect of condensed tannins from Lotus pedunculatus andLotus corniculatus on the growth of proteolytic rumen bacteriain vitro and their possible mode of actionrdquo Canadian Journal ofMicrobiology vol 47 no 7 pp 626ndash633 2001
[43] A Jayanegara G Goel H P S Makkar and K Becker ldquoDiver-gence between purified hydrolysable and condensed tannin
effects on methane emission rumen fermentation and micro-bial population in vitrordquo Animal Feed Science and Technologyvol 209 pp 60ndash68 2015
[44] BMinW Pinchak K Hernandez et al ldquoEffects of plant tanninsupplementation on animal responses and in vivo ruminalbacterial populations associated with bloat in heifers grazingwheat forage1rdquo The Professional Animal Scientist vol 28 no 4pp 464ndash472 2012
[45] T F Martınez T A McAllister Y Wang and T Reuter ldquoEffectsof tannic acid and quebracho tannins on in vitro ruminalfermentation of wheat and corn grainrdquo Journal of the Scienceof Food and Agriculture vol 86 no 8 pp 1244ndash1256 2006
[46] L Marın E M Miguelez C J Villar and F Lombo ldquoBioavail-ability of dietary polyphenols and gut microbiota metabolismantimicrobial propertiesrdquo BioMed Research International vol2015 Article ID 905215 18 pages 2015
[47] J B Russell and D B Wilson ldquoWhy are ruminal cellulolyticbacteria unable to digest cellulose at low pHrdquo Journal of DairyScience vol 79 no 8 pp 1503ndash1509 1996
[48] K Tajima S Arai K Ogata et al ldquoRumen bacterial communitytransition during adaptation to high-grain dietrdquo Anaerobe vol6 no 5 pp 273ndash284 2000
[49] D W Goad C L Goad and T G Nagaraja ldquoRuminal micro-bial and fermentative changes associated with experimentallyinduced subacute acidosis in steersrdquo Journal of Animal Sciencevol 76 no 1 pp 234ndash241 1998
[50] J Hernandez J L Benedito A Abuelo and C CastilloldquoRuminal acidosis in feedlot from aetiology to preventionrdquoTheScientific World Journal vol 2014 Article ID 702572 2014
[51] A B De Menezes E Lewis M OrsquoDonovan B F OrsquoNeill NClipson and E M Doyle ldquoMicrobiome analysis of dairy cowsfed pasture or total mixed ration dietsrdquo FEMS MicrobiologyEcology vol 78 no 2 pp 256ndash265 2011
[52] I K Skene and J D Brooker ldquoCharacterization of tanninacylhydrolase activity in the ruminal bacterium selenomonasruminantiumrdquo Anaerobe vol 1 no 6 pp 321ndash327 1995
[53] G Goel A K Puniya and K Singh ldquoTannic acid resistance inruminal streptococcal isolatesrdquo Journal of Basic Microbiologyvol 45 no 3 pp 243ndash245 2005
[54] P R Myer J E Wells T P L Smith L A Kuehn and H CFreetly ldquoCecum microbial communities from steers differingin feed efficiencyrdquo Journal of Animal Science vol 93 no 11 pp5327ndash5340 2015
[55] S E Hook A-D GWright and BWMcBride ldquoMethanogensmethane producers of the rumen and mitigation strategiesrdquoArchaea vol 2010 Article ID 945785 11 pages 2010
[56] M Saminathan C C Sieo H M Gan N Abdullah C M VL Wong and Y W Ho ldquoEffects of condensed tannin fractionsof different molecular weights on population and diversity ofbovine rumen methanogenic archaea in vitro as determinedby high-throughput sequencingrdquo Animal Feed Science andTechnology vol 216 pp 146ndash160 2016
[57] F Hassanat and C Benchaar ldquoAssessment of the effect ofcondensed (acacia and quebracho) and hydrolysable (chestnutand valonea) tannins on rumen fermentation andmethane pro-duction in vitrordquo Journal of the Science of Food and Agriculturevol 93 no 2 pp 332ndash339 2013
[58] J Balcells A Aris A Serrano A R Seradj J Crespo and MDevant ldquoEffects of an extract of plant flavonoids (bioflavex)on rumen fermentation and performance in heifers fed high-concentrate dietsrdquo Journal of Animal Science vol 90 no 13 pp4975ndash4984 2012
BioMed Research International 11
[59] A K Patra ldquoUreaammonia metabolism in the rumen andtoxicity in ruminantsrdquo Rumen Microbiology From Evolution toRevolution pp 329ndash341 2015
[60] MAWoznyM P Bryant L VHoldeman andW E CMooreldquoUrease assay and urease producing species of anaerobes in thebovine rumen and human fecesrdquo Applied and EnvironmentalMicrobiology vol 33 no 5 pp 1097ndash1104 1977
[61] D Jin S Zhao P Wang et al ldquoInsights into abundant rumenureolytic bacterial community using rumen simulation systemrdquoFrontiers in Microbiology vol 7 article no 1006 2016
[62] H P SMakkar B Singh andRKDawra ldquoEffect of tannin-richleaves of oak (Quercus incana) on various microbial enzymeactivities of the bovine rumenrdquo British Journal of Nutrition vol60 no 2 pp 287ndash296 1988
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
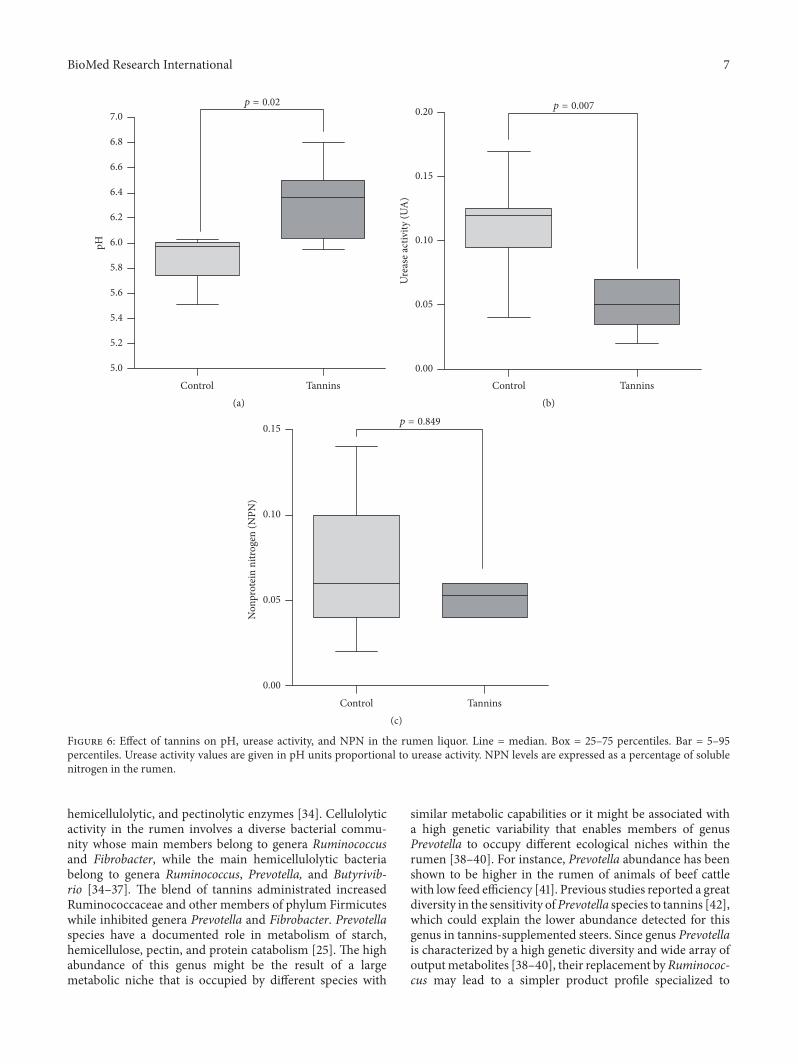
BioMed Research International 7
Control Tannins50
52
54
56
58
60
62
64
66
68
70pH
p = 002
(a)Control Tannins
000
005
010
015
020
Ure
ase a
ctiv
ity (U
A)
p = 0007
(b)
Control Tannins
000
005
010
015
Non
prot
ein
nitro
gen
(NPN
)
p = 0849
(c)
Figure 6 Effect of tannins on pH urease activity and NPN in the rumen liquor Line = median Box = 25ndash75 percentiles Bar = 5ndash95percentiles Urease activity values are given in pH units proportional to urease activity NPN levels are expressed as a percentage of solublenitrogen in the rumen
hemicellulolytic and pectinolytic enzymes [34] Cellulolyticactivity in the rumen involves a diverse bacterial commu-nity whose main members belong to genera Ruminococcusand Fibrobacter while the main hemicellulolytic bacteriabelong to genera Ruminococcus Prevotella and Butyrivib-rio [34ndash37] The blend of tannins administrated increasedRuminococcaceae and other members of phylum Firmicuteswhile inhibited genera Prevotella and Fibrobacter Prevotellaspecies have a documented role in metabolism of starchhemicellulose pectin and protein catabolism [25] The highabundance of this genus might be the result of a largemetabolic niche that is occupied by different species with
similar metabolic capabilities or it might be associated witha high genetic variability that enables members of genusPrevotella to occupy different ecological niches within therumen [38ndash40] For instance Prevotella abundance has beenshown to be higher in the rumen of animals of beef cattlewith low feed efficiency [41] Previous studies reported a greatdiversity in the sensitivity ofPrevotella species to tannins [42]which could explain the lower abundance detected for thisgenus in tannins-supplemented steers Since genus Prevotellais characterized by a high genetic diversity and wide array ofoutputmetabolites [38ndash40] their replacement byRuminococ-cus may lead to a simpler product profile specialized to
8 BioMed Research International
support the hostrsquos energy requirements [30] Our resultsagree with previous studies that found a significant reduc-tion in Fibrobacter and Prevotella rumen populations aftersupplementation with condensed and hydrolysable tanninsincluding chestnut and quebracho tannins [43 44]
Rumen fibrolytic function is carried out by a largelyredundant microbial community with overlapping distribu-tion of metabolic capabilities and this fibrolytic communityhas the ability to restore its structure and function afterperturbation a phenomenon known as resilience [27] In thiscontext the observed changes in diversity of fibrolytic bacte-ria in tannins treated steers may result from a combinationof physicochemical and biological mechanisms described inthe literature including direct interaction of tannins withfiber [45] which could alter the available surface area formicrobial attack the inhibition of certain fibrolytic taxa bymeans of tannins antimicrobial activity [46] andmodulationof fibrolytic bacterial species driven by changes in rumen pHsince certain fibrolytic taxa of the rumen are inhibited at lowpH under high-grain diets [36 47]
Rumen amylolytic and saccharolytic bacteria were alsoaffected by dietary treatment with tannins mainly throughthe decrease of genera Prevotella and Treponema Otheramylolytic genera were moderately increased in tannins-fedsteers including Streptococcus Bifidobacterium and Lacto-bacillus Amylolytic activity is normally enhanced in rumi-nants consuming high-grain diets [18 48 49] The rate ofgrain degradation by these microbial communities plays akey role in maintaining rumen homeostasis since rapidstarch fermentation produces large amounts of organic acidstherefore producing a drop in ruminal pH that may leadto metabolic acidosis [50] A previous study showed thattannic acid and quebracho tannins lower the rate ofmicrobialhydrolysis of starch-rich grains in the rumen by physicalmodification of the endosperm protein matrix [45] Thisphysical modulation of starch degradation could explain thelower abundance of sugar fermenting taxa detected in tanninstreated steers as well as the higher ruminal pH observed inthis group
Fermentation products of microbial activity in rumenmainly short-chain volatile fatty acids serve as amajor sourceof energy for ruminants and have a direct impact on thephysiological parameters of the animal and feed utilizationefficiency [16] Some members of family Veillonellaceaewhich produce propionate as a major fermentation productand have been associated with lower methane emissions [51]were enhanced in tannins treated animals Interestingly someSelenomonas species can break tannin-protein complexes anduse tannins as energy source [52 53] Therefore the higherabundance of Selenomonas in tannins treated steers may bepartly due to availability of tannins as direct energy sourceMeanwhile members of class Erysipelotrichi which havebeen linked to beef cattle feed efficiency [33 51 54] were alsomodulated by tannins treatment
Methane production during fermentation of feeds in therumen represents a loss of 2ndash12 of gross energy [1] andit is performed by a group of archaea known collectivelyas methanogens which belong to phylum EuryarchaeotaThe microorganisms produce methane the second largest
anthropogenic greenhouse gas which has a global warmingpotential 25 times that of carbon dioxide [55] In the presentstudy a reduction of methanogenic archaea in tannins sup-plemented steers was detected as well as an inverse cor-relation between the abundances of phylum Euryarchaeotaand rumen pH (119903 = minus095) Tannins are thought todirectly inhibit methanogens as well as indirectly limitmethanogenesis through reduction of hydrogen availability[1 55] Saminathan et al found a significant decrease in genusMethanobrevibacter after treatment of rumen samples withcondensed tannins in vitro [56] Other authors describeda linear decrease of genus Methanobrevibacter after dietarysupplementation with condensed tannins from pine bark ingoats [31]
Ruminal pH was significantly higher in tannins treatedsteers than in the control group while urease activity exhib-ited the opposite pattern These results agree with previousreports which found an increase in ruminal pH after sup-plementation with chestnut and quebracho tannins [57 58]Other authors observed that feeding chestnut and quebrachotannins decreased urease activity in the faeces of cows[10 11]
Feed-grade urea is an effective source of nitrogen com-monly used in beef cattle diets Ureolytic bacteria in therumen produce urease to hydrolyse urea to ammonia whichis subsequently used for the synthesis of amino acids andmicrobial protein Normally the rate of urea hydrolysisexceeds the rate of ammonia utilization which leads to poorefficiency of urea utilization and increases toxic ammoniaconcentrations in blood [59] Ureolytic bacteria in the rumencomprise a highly diverse group whose main species belongto genera Succinivibrio Treponema Bacteroides ButyrivibrioStreptococcus andBifidobacterium [60 61]We found that themost abundant ureolytic generaButyrivibrio andTreponemawere negatively affected by tannins treatment Thus theobserved decline in ruminal urease activity may be relatedwith the decrease of these urease-producing taxa A previousstudy also observed a drop in ruminal urease activity afteraddition of tannins to diet but a direct interaction betweentannins and urease enzyme was postulated as responsible forthis inhibition [62]
5 Conclusions
The current study showed that chestnut and quebrachotannins added to the diet of Holstein steers modifiedrumen microbiota composition particularly fiber and starchdegrading bacteria mainly by reducing the abundanceof Prevotella and Fibrobacter while favoring Ruminococ-caceae and other members of phylum Firmicutes Tanninstreatment significantly increased pH and decreased ureaseactivity in the ruminal liquor Further work is necessaryto assess the possible relation between tannins ability tomodify rumen microbiota composition and the alterationof rumen fermentation parameters associated with energyand feed efficiency of beef cattle such as the profile ofshort-chain fatty acids and the emissions of ammonia andmethane
BioMed Research International 9
Conflicts of Interest
The authors declare that there are no known conflicts ofinterest associated with this publication and there has beenno financial support for this work that could have influencedits outcome
Acknowledgments
The authors thank Alfredo Ivan Martınez Caceres for histechnical assistance during installation and startup of thebioinformatics tools used in this study This work wassupported by grants from INTA (PNSA 1115052 and PNBIO1131043) Additional support was provided by CONICET
References
[1] A K Patra and J Saxena ldquoDietary phytochemicals as rumenmodifiers a review of the effects on microbial populationsrdquoInternational Journal of General and Molecular Microbiologyvol 96 no 4 pp 363ndash375 2009 Antonie Van Leeuwenhoek
[2] R Bodas N Prieto R Garcıa-Gonzalez S Andres F JGiraldez and S Lopez ldquoManipulation of rumen fermentationand methane production with plant secondary metabolitesrdquoAnimal Feed Science and Technology vol 176 no 1-4 pp 78ndash932012
[3] E Oskoueian N Abdullah and A Oskoueian ldquoEffects offlavonoids on rumen fermentation activity methane produc-tion andmicrobial populationrdquo BioMed Research Internationalvol 2013 Article ID 349129 8 pages 2013
[4] P Frutos GHervas F J Giraldez andA RMantecon ldquoReviewtannins and ruminant nutritionrdquo Spanish Journal of AgriculturalResearch vol 2 no 2 pp 191ndash202 2004
[5] K Khanbabaee and T van Ree ldquoTannins classification anddefinitionrdquo Natural Product Reports vol 18 no 6 pp 641ndash6492001
[6] L J Porter ldquoStructure and chemical properties of the con-densed tanninsrdquo Plant Polyphenols pp 245ndash258 1992 Springer Boston MA USA
[7] B R Min C Wright P Ho J-S Eun N Gurung and RShange ldquoThe effect of phytochemical tannins-containing dieton rumen fermentation characteristics and microbial diversitydynamics in goats using 16S rDNA amplicon pyrosequencingrdquoAgriculture Food and Analytical Bacteriology vol 4 no 3 pp195ndash211 2014
[8] B S Gemeda and A Hassen ldquoEffect of tannin and speciesvariation on in vitro digestibility gas and methane productionof tropical browse plantsrdquoAsian-Australasian Journal of AnimalSciences vol 28 no 2 pp 188ndash199 2015
[9] Z Li A-D G Wright H Liu et al ldquoResponse of the rumenmicrobiota of sika deer (cervus nippon) fed different concen-trations of tannin rich plantsrdquo PLoS ONE vol 10 no 5 ArticleID e0123481 2015
[10] JM PowelM J Aguerre andMAWattiaux ldquoTannin extractsabate ammonia emissions from simulated dairy barn floorsrdquoJournal of Environmental Quality vol 40 no 3 pp 907ndash9142011
[11] M J Aguerre M C Capozzolo P Lencioni C Cabral andM A Wattiaux ldquoEffect of quebracho-chestnut tannin extractsat 2 dietary crude protein levels on performance rumenfermentation and nitrogen partitioning in dairy cowsrdquo Journal
of Dairy Science vol 99 no 6 Article ID 74473 pp 4476ndash44862016
[12] C Rivera-Mendez A Plascencia N Torrentera andR A ZinnldquoEffect of level and source of supplemental tannin on growthperformance of steers during the late finishing phaserdquo Journalof Applied Animal Research vol 45 pp 199ndash203 2017
[13] B R Min T N Barry G T Attwood and W C McNabbldquoThe effect of condensed tannins on the nutrition and health ofruminants fed fresh temperate forages a reviewrdquo Animal FeedScience and Technology vol 106 no 1-4 pp 3ndash19 2003
[14] P J Turnbaugh R E Ley M A Mahowald V MagriniE R Mardis and J I Gordon ldquoAn obesity-associated gutmicrobiomewith increased capacity for energy harvestrdquoNaturevol 444 no 7122 pp 1027ndash1031 2006
[15] R E Ley R Knight and J I Gordon ldquoThe human micro-biome Eliminating the biomedicalenvironmental dichotomyin microbial ecologyrdquo Environmental Microbiology vol 9 no 1pp 3-4 2007
[16] I Mizrahi ldquoThe role of the rumen microbiota in determiningthe feed efficiency of dairy cowsrdquo Beneficial Microorganismsin Multicellular Life Forms pp 203ndash210 2012 Springer BerlinHeidelberg
[17] E Jami B AWhite and IMizrahi ldquoPotential role of the bovinerumen microbiome in modulating milk composition and feedefficiencyrdquo PLoS ONE vol 9 no 1 Article ID e85423 2014
[18] S C Fernando H T Purvis F Z Najar et al ldquoRumenmicrobialpopulation dynamics during adaptation to a high-grain dietrdquoApplied and Environmental Microbiology vol 76 no 22 pp7482ndash7490 2010
[19] H P S Makkar ldquoEffects and fate of tannins in ruminantanimals adaptation to tannins and strategies to overcomedetrimental effects of feeding tannin-rich feedsrdquo Small Rumi-nant Research vol 49 no 3 pp 241ndash256 2003
[20] Illumina Protocol 16S Metagenomic Sequencing Library 2013[21] AM BolgerM Lohse and B Usadel ldquoTrimmomatic a flexible
trimmer for Illumina sequence datardquoBioinformatics vol 30 no15 pp 2114ndash2120 2014
[22] J Kuczynski J Stombaugh W A Walters A Gonzalez J GCaporaso and R Knight ldquoUsing QIIME to analyze 16S rRNAgene sequences frommicrobial communitiesrdquoCurrent Protocolsin Microbiology vol 1 1E5 pages 2012
[23] N A Bokulich S Subramanian J J Faith et al ldquoQuality-filtering vastly improves diversity estimates from Illuminaamplicon sequencingrdquo Nature Methods vol 10 no 1 pp 57ndash592013
[24] D Parks and R Beiko ldquoSTAMP statistical analysis of metage-nomic profilesrdquo Encyclopedia of Metagenomics pp 1ndash6 2013Springer New York NY USA
[25] J C McCann T A Wickersham and J J Loor ldquoHigh-throughput methods redefine the rumen microbiome and itsrelationshipwith nutrition andmetabolismrdquoBioinformatics andBiology Insights vol 8 pp 109ndash125 2014
[26] G Henderson F Cox S Ganesh A Jonker W Young and PH Janssen ldquoRumen microbial community composition varieswith diet and host but a coremicrobiome is found across a widegeographical rangerdquo Scientific Reports vol 5 no 14567 pp 1ndash132015
[27] P J Weimer ldquoRedundancy resilience and host specificity ofthe ruminal microbiota Implications for engineering improvedruminal fermentationsrdquo Frontiers in Microbiology vol 6 articleno 296 2015
10 BioMed Research International
[28] E Jami and I Mizrahi ldquoComposition and similarity of bovinerumenmicrobiota across individual animalsrdquo PLoS ONE vol 7no 3 Article ID e33306 2012
[29] R De Nardi G Marchesini S Li et al ldquoMetagenomic analysisof rumen microbial population in dairy heifers fed a high graindiet supplemented with dicarboxylic acids or polyphenolsrdquoBMC Veterinary Research vol 12 no 1 article no 29 2016
[30] S Kruger Ben Shabat G Sasson A Doron-Faigenboim etal ldquoSpecific microbiome-dependent mechanisms underlie theenergy harvest efficiency of ruminantsrdquoThe ISME Journal vol10 no 12 pp 2958ndash2972 2016
[31] B R Min S Solaiman R Shange and J-S Eun ldquoGastroin-testinal bacterial andmethanogenic archaea diversity dynamicsassociated with condensed tannin-containing pine bark diet ingoats using 16S rDNA amplicon pyrosequencingrdquo InternationalJournal of Microbiology vol 2014 Article ID 141909 2014
[32] G Henderson F Cox S Kittelmann et al ldquoEffect of DNAextraction methods and sampling techniques on the apparentstructure of cow and sheep rumen microbial communitiesrdquoPLoS ONE vol 8 no 9 Article ID e74787 2013
[33] P R Myer T P L Smith J E Wells L A Kuehn and HC Freetly ldquoRumen microbiome from steers differing in feedefficiencyrdquo PLoS ONE vol 10 no 6 Article ID e0129174 2015
[34] Y Wang and T A McAllister ldquoRumen microbes enzymes andfeed digestion-a reviewrdquo Asian-Australasian Journal of AnimalSciences vol 15 no 11 pp 1659ndash1676 2002
[35] J B Russell R EMuck and P JWeimer ldquoQuantitative analysisof cellulose degradation and growth of cellulolytic bacteria inthe rumenrdquo FEMS Microbiology Ecology vol 67 no 2 pp 183ndash197 2009
[36] DOKrause S EDenmanR IMackie et al ldquoOpportunities toimprove fiber degradation in the rumen microbiology ecologyand genomicsrdquo FEMS Microbiology Reviews vol 27 no 5 pp663ndash693 2003
[37] M Zorec M Vodovnik and R Marinsek-Logar ldquoPotentialof selected rumen bacteria for cellulose and hemicellulosedegradationrdquo Food Technology and Biotechnology vol 52 no 2pp 210ndash221 2014
[38] A Z Bekele S Koike and Y Kobayashi ldquoGenetic diversityand diet specificity of ruminal prevotella revealed by 16S rRNAgene-based analysisrdquo FEMSMicrobiology Letters vol 305 no 1pp 49ndash57 2010
[39] A RamsakM Peterka K Tajima et al ldquoUnravelling the geneticdiversity of ruminal bacteria belonging to the CFB phylumrdquoFEMS Microbiology Ecology vol 33 no 1 pp 69ndash79 2000
[40] J Purushe D E Fouts M Morrison et al ldquoComparativegenome analysis of prevotella ruminicola and prevotella bryan-tii insights into their environmental nicherdquo Microbial Ecologyvol 60 no 4 pp 721ndash729 2010
[41] C A Carberry D A Kenny S Han M S McCabe and S MWaters ldquoEffect of phenotypic residual feed intake and dietaryforage content on the rumen microbial community of beefcattlerdquo Applied and Environmental Microbiology vol 78 no 14pp 4949ndash4958 2012
[42] A L Molan G T Attwood B R Min and W C McNabbldquoThe effect of condensed tannins from Lotus pedunculatus andLotus corniculatus on the growth of proteolytic rumen bacteriain vitro and their possible mode of actionrdquo Canadian Journal ofMicrobiology vol 47 no 7 pp 626ndash633 2001
[43] A Jayanegara G Goel H P S Makkar and K Becker ldquoDiver-gence between purified hydrolysable and condensed tannin
effects on methane emission rumen fermentation and micro-bial population in vitrordquo Animal Feed Science and Technologyvol 209 pp 60ndash68 2015
[44] BMinW Pinchak K Hernandez et al ldquoEffects of plant tanninsupplementation on animal responses and in vivo ruminalbacterial populations associated with bloat in heifers grazingwheat forage1rdquo The Professional Animal Scientist vol 28 no 4pp 464ndash472 2012
[45] T F Martınez T A McAllister Y Wang and T Reuter ldquoEffectsof tannic acid and quebracho tannins on in vitro ruminalfermentation of wheat and corn grainrdquo Journal of the Scienceof Food and Agriculture vol 86 no 8 pp 1244ndash1256 2006
[46] L Marın E M Miguelez C J Villar and F Lombo ldquoBioavail-ability of dietary polyphenols and gut microbiota metabolismantimicrobial propertiesrdquo BioMed Research International vol2015 Article ID 905215 18 pages 2015
[47] J B Russell and D B Wilson ldquoWhy are ruminal cellulolyticbacteria unable to digest cellulose at low pHrdquo Journal of DairyScience vol 79 no 8 pp 1503ndash1509 1996
[48] K Tajima S Arai K Ogata et al ldquoRumen bacterial communitytransition during adaptation to high-grain dietrdquo Anaerobe vol6 no 5 pp 273ndash284 2000
[49] D W Goad C L Goad and T G Nagaraja ldquoRuminal micro-bial and fermentative changes associated with experimentallyinduced subacute acidosis in steersrdquo Journal of Animal Sciencevol 76 no 1 pp 234ndash241 1998
[50] J Hernandez J L Benedito A Abuelo and C CastilloldquoRuminal acidosis in feedlot from aetiology to preventionrdquoTheScientific World Journal vol 2014 Article ID 702572 2014
[51] A B De Menezes E Lewis M OrsquoDonovan B F OrsquoNeill NClipson and E M Doyle ldquoMicrobiome analysis of dairy cowsfed pasture or total mixed ration dietsrdquo FEMS MicrobiologyEcology vol 78 no 2 pp 256ndash265 2011
[52] I K Skene and J D Brooker ldquoCharacterization of tanninacylhydrolase activity in the ruminal bacterium selenomonasruminantiumrdquo Anaerobe vol 1 no 6 pp 321ndash327 1995
[53] G Goel A K Puniya and K Singh ldquoTannic acid resistance inruminal streptococcal isolatesrdquo Journal of Basic Microbiologyvol 45 no 3 pp 243ndash245 2005
[54] P R Myer J E Wells T P L Smith L A Kuehn and H CFreetly ldquoCecum microbial communities from steers differingin feed efficiencyrdquo Journal of Animal Science vol 93 no 11 pp5327ndash5340 2015
[55] S E Hook A-D GWright and BWMcBride ldquoMethanogensmethane producers of the rumen and mitigation strategiesrdquoArchaea vol 2010 Article ID 945785 11 pages 2010
[56] M Saminathan C C Sieo H M Gan N Abdullah C M VL Wong and Y W Ho ldquoEffects of condensed tannin fractionsof different molecular weights on population and diversity ofbovine rumen methanogenic archaea in vitro as determinedby high-throughput sequencingrdquo Animal Feed Science andTechnology vol 216 pp 146ndash160 2016
[57] F Hassanat and C Benchaar ldquoAssessment of the effect ofcondensed (acacia and quebracho) and hydrolysable (chestnutand valonea) tannins on rumen fermentation andmethane pro-duction in vitrordquo Journal of the Science of Food and Agriculturevol 93 no 2 pp 332ndash339 2013
[58] J Balcells A Aris A Serrano A R Seradj J Crespo and MDevant ldquoEffects of an extract of plant flavonoids (bioflavex)on rumen fermentation and performance in heifers fed high-concentrate dietsrdquo Journal of Animal Science vol 90 no 13 pp4975ndash4984 2012
BioMed Research International 11
[59] A K Patra ldquoUreaammonia metabolism in the rumen andtoxicity in ruminantsrdquo Rumen Microbiology From Evolution toRevolution pp 329ndash341 2015
[60] MAWoznyM P Bryant L VHoldeman andW E CMooreldquoUrease assay and urease producing species of anaerobes in thebovine rumen and human fecesrdquo Applied and EnvironmentalMicrobiology vol 33 no 5 pp 1097ndash1104 1977
[61] D Jin S Zhao P Wang et al ldquoInsights into abundant rumenureolytic bacterial community using rumen simulation systemrdquoFrontiers in Microbiology vol 7 article no 1006 2016
[62] H P SMakkar B Singh andRKDawra ldquoEffect of tannin-richleaves of oak (Quercus incana) on various microbial enzymeactivities of the bovine rumenrdquo British Journal of Nutrition vol60 no 2 pp 287ndash296 1988
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 BioMed Research International
support the hostrsquos energy requirements [30] Our resultsagree with previous studies that found a significant reduc-tion in Fibrobacter and Prevotella rumen populations aftersupplementation with condensed and hydrolysable tanninsincluding chestnut and quebracho tannins [43 44]
Rumen fibrolytic function is carried out by a largelyredundant microbial community with overlapping distribu-tion of metabolic capabilities and this fibrolytic communityhas the ability to restore its structure and function afterperturbation a phenomenon known as resilience [27] In thiscontext the observed changes in diversity of fibrolytic bacte-ria in tannins treated steers may result from a combinationof physicochemical and biological mechanisms described inthe literature including direct interaction of tannins withfiber [45] which could alter the available surface area formicrobial attack the inhibition of certain fibrolytic taxa bymeans of tannins antimicrobial activity [46] andmodulationof fibrolytic bacterial species driven by changes in rumen pHsince certain fibrolytic taxa of the rumen are inhibited at lowpH under high-grain diets [36 47]
Rumen amylolytic and saccharolytic bacteria were alsoaffected by dietary treatment with tannins mainly throughthe decrease of genera Prevotella and Treponema Otheramylolytic genera were moderately increased in tannins-fedsteers including Streptococcus Bifidobacterium and Lacto-bacillus Amylolytic activity is normally enhanced in rumi-nants consuming high-grain diets [18 48 49] The rate ofgrain degradation by these microbial communities plays akey role in maintaining rumen homeostasis since rapidstarch fermentation produces large amounts of organic acidstherefore producing a drop in ruminal pH that may leadto metabolic acidosis [50] A previous study showed thattannic acid and quebracho tannins lower the rate ofmicrobialhydrolysis of starch-rich grains in the rumen by physicalmodification of the endosperm protein matrix [45] Thisphysical modulation of starch degradation could explain thelower abundance of sugar fermenting taxa detected in tanninstreated steers as well as the higher ruminal pH observed inthis group
Fermentation products of microbial activity in rumenmainly short-chain volatile fatty acids serve as amajor sourceof energy for ruminants and have a direct impact on thephysiological parameters of the animal and feed utilizationefficiency [16] Some members of family Veillonellaceaewhich produce propionate as a major fermentation productand have been associated with lower methane emissions [51]were enhanced in tannins treated animals Interestingly someSelenomonas species can break tannin-protein complexes anduse tannins as energy source [52 53] Therefore the higherabundance of Selenomonas in tannins treated steers may bepartly due to availability of tannins as direct energy sourceMeanwhile members of class Erysipelotrichi which havebeen linked to beef cattle feed efficiency [33 51 54] were alsomodulated by tannins treatment
Methane production during fermentation of feeds in therumen represents a loss of 2ndash12 of gross energy [1] andit is performed by a group of archaea known collectivelyas methanogens which belong to phylum EuryarchaeotaThe microorganisms produce methane the second largest
anthropogenic greenhouse gas which has a global warmingpotential 25 times that of carbon dioxide [55] In the presentstudy a reduction of methanogenic archaea in tannins sup-plemented steers was detected as well as an inverse cor-relation between the abundances of phylum Euryarchaeotaand rumen pH (119903 = minus095) Tannins are thought todirectly inhibit methanogens as well as indirectly limitmethanogenesis through reduction of hydrogen availability[1 55] Saminathan et al found a significant decrease in genusMethanobrevibacter after treatment of rumen samples withcondensed tannins in vitro [56] Other authors describeda linear decrease of genus Methanobrevibacter after dietarysupplementation with condensed tannins from pine bark ingoats [31]
Ruminal pH was significantly higher in tannins treatedsteers than in the control group while urease activity exhib-ited the opposite pattern These results agree with previousreports which found an increase in ruminal pH after sup-plementation with chestnut and quebracho tannins [57 58]Other authors observed that feeding chestnut and quebrachotannins decreased urease activity in the faeces of cows[10 11]
Feed-grade urea is an effective source of nitrogen com-monly used in beef cattle diets Ureolytic bacteria in therumen produce urease to hydrolyse urea to ammonia whichis subsequently used for the synthesis of amino acids andmicrobial protein Normally the rate of urea hydrolysisexceeds the rate of ammonia utilization which leads to poorefficiency of urea utilization and increases toxic ammoniaconcentrations in blood [59] Ureolytic bacteria in the rumencomprise a highly diverse group whose main species belongto genera Succinivibrio Treponema Bacteroides ButyrivibrioStreptococcus andBifidobacterium [60 61]We found that themost abundant ureolytic generaButyrivibrio andTreponemawere negatively affected by tannins treatment Thus theobserved decline in ruminal urease activity may be relatedwith the decrease of these urease-producing taxa A previousstudy also observed a drop in ruminal urease activity afteraddition of tannins to diet but a direct interaction betweentannins and urease enzyme was postulated as responsible forthis inhibition [62]
5 Conclusions
The current study showed that chestnut and quebrachotannins added to the diet of Holstein steers modifiedrumen microbiota composition particularly fiber and starchdegrading bacteria mainly by reducing the abundanceof Prevotella and Fibrobacter while favoring Ruminococ-caceae and other members of phylum Firmicutes Tanninstreatment significantly increased pH and decreased ureaseactivity in the ruminal liquor Further work is necessaryto assess the possible relation between tannins ability tomodify rumen microbiota composition and the alterationof rumen fermentation parameters associated with energyand feed efficiency of beef cattle such as the profile ofshort-chain fatty acids and the emissions of ammonia andmethane
BioMed Research International 9
Conflicts of Interest
The authors declare that there are no known conflicts ofinterest associated with this publication and there has beenno financial support for this work that could have influencedits outcome
Acknowledgments
The authors thank Alfredo Ivan Martınez Caceres for histechnical assistance during installation and startup of thebioinformatics tools used in this study This work wassupported by grants from INTA (PNSA 1115052 and PNBIO1131043) Additional support was provided by CONICET
References
[1] A K Patra and J Saxena ldquoDietary phytochemicals as rumenmodifiers a review of the effects on microbial populationsrdquoInternational Journal of General and Molecular Microbiologyvol 96 no 4 pp 363ndash375 2009 Antonie Van Leeuwenhoek
[2] R Bodas N Prieto R Garcıa-Gonzalez S Andres F JGiraldez and S Lopez ldquoManipulation of rumen fermentationand methane production with plant secondary metabolitesrdquoAnimal Feed Science and Technology vol 176 no 1-4 pp 78ndash932012
[3] E Oskoueian N Abdullah and A Oskoueian ldquoEffects offlavonoids on rumen fermentation activity methane produc-tion andmicrobial populationrdquo BioMed Research Internationalvol 2013 Article ID 349129 8 pages 2013
[4] P Frutos GHervas F J Giraldez andA RMantecon ldquoReviewtannins and ruminant nutritionrdquo Spanish Journal of AgriculturalResearch vol 2 no 2 pp 191ndash202 2004
[5] K Khanbabaee and T van Ree ldquoTannins classification anddefinitionrdquo Natural Product Reports vol 18 no 6 pp 641ndash6492001
[6] L J Porter ldquoStructure and chemical properties of the con-densed tanninsrdquo Plant Polyphenols pp 245ndash258 1992 Springer Boston MA USA
[7] B R Min C Wright P Ho J-S Eun N Gurung and RShange ldquoThe effect of phytochemical tannins-containing dieton rumen fermentation characteristics and microbial diversitydynamics in goats using 16S rDNA amplicon pyrosequencingrdquoAgriculture Food and Analytical Bacteriology vol 4 no 3 pp195ndash211 2014
[8] B S Gemeda and A Hassen ldquoEffect of tannin and speciesvariation on in vitro digestibility gas and methane productionof tropical browse plantsrdquoAsian-Australasian Journal of AnimalSciences vol 28 no 2 pp 188ndash199 2015
[9] Z Li A-D G Wright H Liu et al ldquoResponse of the rumenmicrobiota of sika deer (cervus nippon) fed different concen-trations of tannin rich plantsrdquo PLoS ONE vol 10 no 5 ArticleID e0123481 2015
[10] JM PowelM J Aguerre andMAWattiaux ldquoTannin extractsabate ammonia emissions from simulated dairy barn floorsrdquoJournal of Environmental Quality vol 40 no 3 pp 907ndash9142011
[11] M J Aguerre M C Capozzolo P Lencioni C Cabral andM A Wattiaux ldquoEffect of quebracho-chestnut tannin extractsat 2 dietary crude protein levels on performance rumenfermentation and nitrogen partitioning in dairy cowsrdquo Journal
of Dairy Science vol 99 no 6 Article ID 74473 pp 4476ndash44862016
[12] C Rivera-Mendez A Plascencia N Torrentera andR A ZinnldquoEffect of level and source of supplemental tannin on growthperformance of steers during the late finishing phaserdquo Journalof Applied Animal Research vol 45 pp 199ndash203 2017
[13] B R Min T N Barry G T Attwood and W C McNabbldquoThe effect of condensed tannins on the nutrition and health ofruminants fed fresh temperate forages a reviewrdquo Animal FeedScience and Technology vol 106 no 1-4 pp 3ndash19 2003
[14] P J Turnbaugh R E Ley M A Mahowald V MagriniE R Mardis and J I Gordon ldquoAn obesity-associated gutmicrobiomewith increased capacity for energy harvestrdquoNaturevol 444 no 7122 pp 1027ndash1031 2006
[15] R E Ley R Knight and J I Gordon ldquoThe human micro-biome Eliminating the biomedicalenvironmental dichotomyin microbial ecologyrdquo Environmental Microbiology vol 9 no 1pp 3-4 2007
[16] I Mizrahi ldquoThe role of the rumen microbiota in determiningthe feed efficiency of dairy cowsrdquo Beneficial Microorganismsin Multicellular Life Forms pp 203ndash210 2012 Springer BerlinHeidelberg
[17] E Jami B AWhite and IMizrahi ldquoPotential role of the bovinerumen microbiome in modulating milk composition and feedefficiencyrdquo PLoS ONE vol 9 no 1 Article ID e85423 2014
[18] S C Fernando H T Purvis F Z Najar et al ldquoRumenmicrobialpopulation dynamics during adaptation to a high-grain dietrdquoApplied and Environmental Microbiology vol 76 no 22 pp7482ndash7490 2010
[19] H P S Makkar ldquoEffects and fate of tannins in ruminantanimals adaptation to tannins and strategies to overcomedetrimental effects of feeding tannin-rich feedsrdquo Small Rumi-nant Research vol 49 no 3 pp 241ndash256 2003
[20] Illumina Protocol 16S Metagenomic Sequencing Library 2013[21] AM BolgerM Lohse and B Usadel ldquoTrimmomatic a flexible
trimmer for Illumina sequence datardquoBioinformatics vol 30 no15 pp 2114ndash2120 2014
[22] J Kuczynski J Stombaugh W A Walters A Gonzalez J GCaporaso and R Knight ldquoUsing QIIME to analyze 16S rRNAgene sequences frommicrobial communitiesrdquoCurrent Protocolsin Microbiology vol 1 1E5 pages 2012
[23] N A Bokulich S Subramanian J J Faith et al ldquoQuality-filtering vastly improves diversity estimates from Illuminaamplicon sequencingrdquo Nature Methods vol 10 no 1 pp 57ndash592013
[24] D Parks and R Beiko ldquoSTAMP statistical analysis of metage-nomic profilesrdquo Encyclopedia of Metagenomics pp 1ndash6 2013Springer New York NY USA
[25] J C McCann T A Wickersham and J J Loor ldquoHigh-throughput methods redefine the rumen microbiome and itsrelationshipwith nutrition andmetabolismrdquoBioinformatics andBiology Insights vol 8 pp 109ndash125 2014
[26] G Henderson F Cox S Ganesh A Jonker W Young and PH Janssen ldquoRumen microbial community composition varieswith diet and host but a coremicrobiome is found across a widegeographical rangerdquo Scientific Reports vol 5 no 14567 pp 1ndash132015
[27] P J Weimer ldquoRedundancy resilience and host specificity ofthe ruminal microbiota Implications for engineering improvedruminal fermentationsrdquo Frontiers in Microbiology vol 6 articleno 296 2015
10 BioMed Research International
[28] E Jami and I Mizrahi ldquoComposition and similarity of bovinerumenmicrobiota across individual animalsrdquo PLoS ONE vol 7no 3 Article ID e33306 2012
[29] R De Nardi G Marchesini S Li et al ldquoMetagenomic analysisof rumen microbial population in dairy heifers fed a high graindiet supplemented with dicarboxylic acids or polyphenolsrdquoBMC Veterinary Research vol 12 no 1 article no 29 2016
[30] S Kruger Ben Shabat G Sasson A Doron-Faigenboim etal ldquoSpecific microbiome-dependent mechanisms underlie theenergy harvest efficiency of ruminantsrdquoThe ISME Journal vol10 no 12 pp 2958ndash2972 2016
[31] B R Min S Solaiman R Shange and J-S Eun ldquoGastroin-testinal bacterial andmethanogenic archaea diversity dynamicsassociated with condensed tannin-containing pine bark diet ingoats using 16S rDNA amplicon pyrosequencingrdquo InternationalJournal of Microbiology vol 2014 Article ID 141909 2014
[32] G Henderson F Cox S Kittelmann et al ldquoEffect of DNAextraction methods and sampling techniques on the apparentstructure of cow and sheep rumen microbial communitiesrdquoPLoS ONE vol 8 no 9 Article ID e74787 2013
[33] P R Myer T P L Smith J E Wells L A Kuehn and HC Freetly ldquoRumen microbiome from steers differing in feedefficiencyrdquo PLoS ONE vol 10 no 6 Article ID e0129174 2015
[34] Y Wang and T A McAllister ldquoRumen microbes enzymes andfeed digestion-a reviewrdquo Asian-Australasian Journal of AnimalSciences vol 15 no 11 pp 1659ndash1676 2002
[35] J B Russell R EMuck and P JWeimer ldquoQuantitative analysisof cellulose degradation and growth of cellulolytic bacteria inthe rumenrdquo FEMS Microbiology Ecology vol 67 no 2 pp 183ndash197 2009
[36] DOKrause S EDenmanR IMackie et al ldquoOpportunities toimprove fiber degradation in the rumen microbiology ecologyand genomicsrdquo FEMS Microbiology Reviews vol 27 no 5 pp663ndash693 2003
[37] M Zorec M Vodovnik and R Marinsek-Logar ldquoPotentialof selected rumen bacteria for cellulose and hemicellulosedegradationrdquo Food Technology and Biotechnology vol 52 no 2pp 210ndash221 2014
[38] A Z Bekele S Koike and Y Kobayashi ldquoGenetic diversityand diet specificity of ruminal prevotella revealed by 16S rRNAgene-based analysisrdquo FEMSMicrobiology Letters vol 305 no 1pp 49ndash57 2010
[39] A RamsakM Peterka K Tajima et al ldquoUnravelling the geneticdiversity of ruminal bacteria belonging to the CFB phylumrdquoFEMS Microbiology Ecology vol 33 no 1 pp 69ndash79 2000
[40] J Purushe D E Fouts M Morrison et al ldquoComparativegenome analysis of prevotella ruminicola and prevotella bryan-tii insights into their environmental nicherdquo Microbial Ecologyvol 60 no 4 pp 721ndash729 2010
[41] C A Carberry D A Kenny S Han M S McCabe and S MWaters ldquoEffect of phenotypic residual feed intake and dietaryforage content on the rumen microbial community of beefcattlerdquo Applied and Environmental Microbiology vol 78 no 14pp 4949ndash4958 2012
[42] A L Molan G T Attwood B R Min and W C McNabbldquoThe effect of condensed tannins from Lotus pedunculatus andLotus corniculatus on the growth of proteolytic rumen bacteriain vitro and their possible mode of actionrdquo Canadian Journal ofMicrobiology vol 47 no 7 pp 626ndash633 2001
[43] A Jayanegara G Goel H P S Makkar and K Becker ldquoDiver-gence between purified hydrolysable and condensed tannin
effects on methane emission rumen fermentation and micro-bial population in vitrordquo Animal Feed Science and Technologyvol 209 pp 60ndash68 2015
[44] BMinW Pinchak K Hernandez et al ldquoEffects of plant tanninsupplementation on animal responses and in vivo ruminalbacterial populations associated with bloat in heifers grazingwheat forage1rdquo The Professional Animal Scientist vol 28 no 4pp 464ndash472 2012
[45] T F Martınez T A McAllister Y Wang and T Reuter ldquoEffectsof tannic acid and quebracho tannins on in vitro ruminalfermentation of wheat and corn grainrdquo Journal of the Scienceof Food and Agriculture vol 86 no 8 pp 1244ndash1256 2006
[46] L Marın E M Miguelez C J Villar and F Lombo ldquoBioavail-ability of dietary polyphenols and gut microbiota metabolismantimicrobial propertiesrdquo BioMed Research International vol2015 Article ID 905215 18 pages 2015
[47] J B Russell and D B Wilson ldquoWhy are ruminal cellulolyticbacteria unable to digest cellulose at low pHrdquo Journal of DairyScience vol 79 no 8 pp 1503ndash1509 1996
[48] K Tajima S Arai K Ogata et al ldquoRumen bacterial communitytransition during adaptation to high-grain dietrdquo Anaerobe vol6 no 5 pp 273ndash284 2000
[49] D W Goad C L Goad and T G Nagaraja ldquoRuminal micro-bial and fermentative changes associated with experimentallyinduced subacute acidosis in steersrdquo Journal of Animal Sciencevol 76 no 1 pp 234ndash241 1998
[50] J Hernandez J L Benedito A Abuelo and C CastilloldquoRuminal acidosis in feedlot from aetiology to preventionrdquoTheScientific World Journal vol 2014 Article ID 702572 2014
[51] A B De Menezes E Lewis M OrsquoDonovan B F OrsquoNeill NClipson and E M Doyle ldquoMicrobiome analysis of dairy cowsfed pasture or total mixed ration dietsrdquo FEMS MicrobiologyEcology vol 78 no 2 pp 256ndash265 2011
[52] I K Skene and J D Brooker ldquoCharacterization of tanninacylhydrolase activity in the ruminal bacterium selenomonasruminantiumrdquo Anaerobe vol 1 no 6 pp 321ndash327 1995
[53] G Goel A K Puniya and K Singh ldquoTannic acid resistance inruminal streptococcal isolatesrdquo Journal of Basic Microbiologyvol 45 no 3 pp 243ndash245 2005
[54] P R Myer J E Wells T P L Smith L A Kuehn and H CFreetly ldquoCecum microbial communities from steers differingin feed efficiencyrdquo Journal of Animal Science vol 93 no 11 pp5327ndash5340 2015
[55] S E Hook A-D GWright and BWMcBride ldquoMethanogensmethane producers of the rumen and mitigation strategiesrdquoArchaea vol 2010 Article ID 945785 11 pages 2010
[56] M Saminathan C C Sieo H M Gan N Abdullah C M VL Wong and Y W Ho ldquoEffects of condensed tannin fractionsof different molecular weights on population and diversity ofbovine rumen methanogenic archaea in vitro as determinedby high-throughput sequencingrdquo Animal Feed Science andTechnology vol 216 pp 146ndash160 2016
[57] F Hassanat and C Benchaar ldquoAssessment of the effect ofcondensed (acacia and quebracho) and hydrolysable (chestnutand valonea) tannins on rumen fermentation andmethane pro-duction in vitrordquo Journal of the Science of Food and Agriculturevol 93 no 2 pp 332ndash339 2013
[58] J Balcells A Aris A Serrano A R Seradj J Crespo and MDevant ldquoEffects of an extract of plant flavonoids (bioflavex)on rumen fermentation and performance in heifers fed high-concentrate dietsrdquo Journal of Animal Science vol 90 no 13 pp4975ndash4984 2012
BioMed Research International 11
[59] A K Patra ldquoUreaammonia metabolism in the rumen andtoxicity in ruminantsrdquo Rumen Microbiology From Evolution toRevolution pp 329ndash341 2015
[60] MAWoznyM P Bryant L VHoldeman andW E CMooreldquoUrease assay and urease producing species of anaerobes in thebovine rumen and human fecesrdquo Applied and EnvironmentalMicrobiology vol 33 no 5 pp 1097ndash1104 1977
[61] D Jin S Zhao P Wang et al ldquoInsights into abundant rumenureolytic bacterial community using rumen simulation systemrdquoFrontiers in Microbiology vol 7 article no 1006 2016
[62] H P SMakkar B Singh andRKDawra ldquoEffect of tannin-richleaves of oak (Quercus incana) on various microbial enzymeactivities of the bovine rumenrdquo British Journal of Nutrition vol60 no 2 pp 287ndash296 1988
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 9
Conflicts of Interest
The authors declare that there are no known conflicts ofinterest associated with this publication and there has beenno financial support for this work that could have influencedits outcome
Acknowledgments
The authors thank Alfredo Ivan Martınez Caceres for histechnical assistance during installation and startup of thebioinformatics tools used in this study This work wassupported by grants from INTA (PNSA 1115052 and PNBIO1131043) Additional support was provided by CONICET
References
[1] A K Patra and J Saxena ldquoDietary phytochemicals as rumenmodifiers a review of the effects on microbial populationsrdquoInternational Journal of General and Molecular Microbiologyvol 96 no 4 pp 363ndash375 2009 Antonie Van Leeuwenhoek
[2] R Bodas N Prieto R Garcıa-Gonzalez S Andres F JGiraldez and S Lopez ldquoManipulation of rumen fermentationand methane production with plant secondary metabolitesrdquoAnimal Feed Science and Technology vol 176 no 1-4 pp 78ndash932012
[3] E Oskoueian N Abdullah and A Oskoueian ldquoEffects offlavonoids on rumen fermentation activity methane produc-tion andmicrobial populationrdquo BioMed Research Internationalvol 2013 Article ID 349129 8 pages 2013
[4] P Frutos GHervas F J Giraldez andA RMantecon ldquoReviewtannins and ruminant nutritionrdquo Spanish Journal of AgriculturalResearch vol 2 no 2 pp 191ndash202 2004
[5] K Khanbabaee and T van Ree ldquoTannins classification anddefinitionrdquo Natural Product Reports vol 18 no 6 pp 641ndash6492001
[6] L J Porter ldquoStructure and chemical properties of the con-densed tanninsrdquo Plant Polyphenols pp 245ndash258 1992 Springer Boston MA USA
[7] B R Min C Wright P Ho J-S Eun N Gurung and RShange ldquoThe effect of phytochemical tannins-containing dieton rumen fermentation characteristics and microbial diversitydynamics in goats using 16S rDNA amplicon pyrosequencingrdquoAgriculture Food and Analytical Bacteriology vol 4 no 3 pp195ndash211 2014
[8] B S Gemeda and A Hassen ldquoEffect of tannin and speciesvariation on in vitro digestibility gas and methane productionof tropical browse plantsrdquoAsian-Australasian Journal of AnimalSciences vol 28 no 2 pp 188ndash199 2015
[9] Z Li A-D G Wright H Liu et al ldquoResponse of the rumenmicrobiota of sika deer (cervus nippon) fed different concen-trations of tannin rich plantsrdquo PLoS ONE vol 10 no 5 ArticleID e0123481 2015
[10] JM PowelM J Aguerre andMAWattiaux ldquoTannin extractsabate ammonia emissions from simulated dairy barn floorsrdquoJournal of Environmental Quality vol 40 no 3 pp 907ndash9142011
[11] M J Aguerre M C Capozzolo P Lencioni C Cabral andM A Wattiaux ldquoEffect of quebracho-chestnut tannin extractsat 2 dietary crude protein levels on performance rumenfermentation and nitrogen partitioning in dairy cowsrdquo Journal
of Dairy Science vol 99 no 6 Article ID 74473 pp 4476ndash44862016
[12] C Rivera-Mendez A Plascencia N Torrentera andR A ZinnldquoEffect of level and source of supplemental tannin on growthperformance of steers during the late finishing phaserdquo Journalof Applied Animal Research vol 45 pp 199ndash203 2017
[13] B R Min T N Barry G T Attwood and W C McNabbldquoThe effect of condensed tannins on the nutrition and health ofruminants fed fresh temperate forages a reviewrdquo Animal FeedScience and Technology vol 106 no 1-4 pp 3ndash19 2003
[14] P J Turnbaugh R E Ley M A Mahowald V MagriniE R Mardis and J I Gordon ldquoAn obesity-associated gutmicrobiomewith increased capacity for energy harvestrdquoNaturevol 444 no 7122 pp 1027ndash1031 2006
[15] R E Ley R Knight and J I Gordon ldquoThe human micro-biome Eliminating the biomedicalenvironmental dichotomyin microbial ecologyrdquo Environmental Microbiology vol 9 no 1pp 3-4 2007
[16] I Mizrahi ldquoThe role of the rumen microbiota in determiningthe feed efficiency of dairy cowsrdquo Beneficial Microorganismsin Multicellular Life Forms pp 203ndash210 2012 Springer BerlinHeidelberg
[17] E Jami B AWhite and IMizrahi ldquoPotential role of the bovinerumen microbiome in modulating milk composition and feedefficiencyrdquo PLoS ONE vol 9 no 1 Article ID e85423 2014
[18] S C Fernando H T Purvis F Z Najar et al ldquoRumenmicrobialpopulation dynamics during adaptation to a high-grain dietrdquoApplied and Environmental Microbiology vol 76 no 22 pp7482ndash7490 2010
[19] H P S Makkar ldquoEffects and fate of tannins in ruminantanimals adaptation to tannins and strategies to overcomedetrimental effects of feeding tannin-rich feedsrdquo Small Rumi-nant Research vol 49 no 3 pp 241ndash256 2003
[20] Illumina Protocol 16S Metagenomic Sequencing Library 2013[21] AM BolgerM Lohse and B Usadel ldquoTrimmomatic a flexible
trimmer for Illumina sequence datardquoBioinformatics vol 30 no15 pp 2114ndash2120 2014
[22] J Kuczynski J Stombaugh W A Walters A Gonzalez J GCaporaso and R Knight ldquoUsing QIIME to analyze 16S rRNAgene sequences frommicrobial communitiesrdquoCurrent Protocolsin Microbiology vol 1 1E5 pages 2012
[23] N A Bokulich S Subramanian J J Faith et al ldquoQuality-filtering vastly improves diversity estimates from Illuminaamplicon sequencingrdquo Nature Methods vol 10 no 1 pp 57ndash592013
[24] D Parks and R Beiko ldquoSTAMP statistical analysis of metage-nomic profilesrdquo Encyclopedia of Metagenomics pp 1ndash6 2013Springer New York NY USA
[25] J C McCann T A Wickersham and J J Loor ldquoHigh-throughput methods redefine the rumen microbiome and itsrelationshipwith nutrition andmetabolismrdquoBioinformatics andBiology Insights vol 8 pp 109ndash125 2014
[26] G Henderson F Cox S Ganesh A Jonker W Young and PH Janssen ldquoRumen microbial community composition varieswith diet and host but a coremicrobiome is found across a widegeographical rangerdquo Scientific Reports vol 5 no 14567 pp 1ndash132015
[27] P J Weimer ldquoRedundancy resilience and host specificity ofthe ruminal microbiota Implications for engineering improvedruminal fermentationsrdquo Frontiers in Microbiology vol 6 articleno 296 2015
10 BioMed Research International
[28] E Jami and I Mizrahi ldquoComposition and similarity of bovinerumenmicrobiota across individual animalsrdquo PLoS ONE vol 7no 3 Article ID e33306 2012
[29] R De Nardi G Marchesini S Li et al ldquoMetagenomic analysisof rumen microbial population in dairy heifers fed a high graindiet supplemented with dicarboxylic acids or polyphenolsrdquoBMC Veterinary Research vol 12 no 1 article no 29 2016
[30] S Kruger Ben Shabat G Sasson A Doron-Faigenboim etal ldquoSpecific microbiome-dependent mechanisms underlie theenergy harvest efficiency of ruminantsrdquoThe ISME Journal vol10 no 12 pp 2958ndash2972 2016
[31] B R Min S Solaiman R Shange and J-S Eun ldquoGastroin-testinal bacterial andmethanogenic archaea diversity dynamicsassociated with condensed tannin-containing pine bark diet ingoats using 16S rDNA amplicon pyrosequencingrdquo InternationalJournal of Microbiology vol 2014 Article ID 141909 2014
[32] G Henderson F Cox S Kittelmann et al ldquoEffect of DNAextraction methods and sampling techniques on the apparentstructure of cow and sheep rumen microbial communitiesrdquoPLoS ONE vol 8 no 9 Article ID e74787 2013
[33] P R Myer T P L Smith J E Wells L A Kuehn and HC Freetly ldquoRumen microbiome from steers differing in feedefficiencyrdquo PLoS ONE vol 10 no 6 Article ID e0129174 2015
[34] Y Wang and T A McAllister ldquoRumen microbes enzymes andfeed digestion-a reviewrdquo Asian-Australasian Journal of AnimalSciences vol 15 no 11 pp 1659ndash1676 2002
[35] J B Russell R EMuck and P JWeimer ldquoQuantitative analysisof cellulose degradation and growth of cellulolytic bacteria inthe rumenrdquo FEMS Microbiology Ecology vol 67 no 2 pp 183ndash197 2009
[36] DOKrause S EDenmanR IMackie et al ldquoOpportunities toimprove fiber degradation in the rumen microbiology ecologyand genomicsrdquo FEMS Microbiology Reviews vol 27 no 5 pp663ndash693 2003
[37] M Zorec M Vodovnik and R Marinsek-Logar ldquoPotentialof selected rumen bacteria for cellulose and hemicellulosedegradationrdquo Food Technology and Biotechnology vol 52 no 2pp 210ndash221 2014
[38] A Z Bekele S Koike and Y Kobayashi ldquoGenetic diversityand diet specificity of ruminal prevotella revealed by 16S rRNAgene-based analysisrdquo FEMSMicrobiology Letters vol 305 no 1pp 49ndash57 2010
[39] A RamsakM Peterka K Tajima et al ldquoUnravelling the geneticdiversity of ruminal bacteria belonging to the CFB phylumrdquoFEMS Microbiology Ecology vol 33 no 1 pp 69ndash79 2000
[40] J Purushe D E Fouts M Morrison et al ldquoComparativegenome analysis of prevotella ruminicola and prevotella bryan-tii insights into their environmental nicherdquo Microbial Ecologyvol 60 no 4 pp 721ndash729 2010
[41] C A Carberry D A Kenny S Han M S McCabe and S MWaters ldquoEffect of phenotypic residual feed intake and dietaryforage content on the rumen microbial community of beefcattlerdquo Applied and Environmental Microbiology vol 78 no 14pp 4949ndash4958 2012
[42] A L Molan G T Attwood B R Min and W C McNabbldquoThe effect of condensed tannins from Lotus pedunculatus andLotus corniculatus on the growth of proteolytic rumen bacteriain vitro and their possible mode of actionrdquo Canadian Journal ofMicrobiology vol 47 no 7 pp 626ndash633 2001
[43] A Jayanegara G Goel H P S Makkar and K Becker ldquoDiver-gence between purified hydrolysable and condensed tannin
effects on methane emission rumen fermentation and micro-bial population in vitrordquo Animal Feed Science and Technologyvol 209 pp 60ndash68 2015
[44] BMinW Pinchak K Hernandez et al ldquoEffects of plant tanninsupplementation on animal responses and in vivo ruminalbacterial populations associated with bloat in heifers grazingwheat forage1rdquo The Professional Animal Scientist vol 28 no 4pp 464ndash472 2012
[45] T F Martınez T A McAllister Y Wang and T Reuter ldquoEffectsof tannic acid and quebracho tannins on in vitro ruminalfermentation of wheat and corn grainrdquo Journal of the Scienceof Food and Agriculture vol 86 no 8 pp 1244ndash1256 2006
[46] L Marın E M Miguelez C J Villar and F Lombo ldquoBioavail-ability of dietary polyphenols and gut microbiota metabolismantimicrobial propertiesrdquo BioMed Research International vol2015 Article ID 905215 18 pages 2015
[47] J B Russell and D B Wilson ldquoWhy are ruminal cellulolyticbacteria unable to digest cellulose at low pHrdquo Journal of DairyScience vol 79 no 8 pp 1503ndash1509 1996
[48] K Tajima S Arai K Ogata et al ldquoRumen bacterial communitytransition during adaptation to high-grain dietrdquo Anaerobe vol6 no 5 pp 273ndash284 2000
[49] D W Goad C L Goad and T G Nagaraja ldquoRuminal micro-bial and fermentative changes associated with experimentallyinduced subacute acidosis in steersrdquo Journal of Animal Sciencevol 76 no 1 pp 234ndash241 1998
[50] J Hernandez J L Benedito A Abuelo and C CastilloldquoRuminal acidosis in feedlot from aetiology to preventionrdquoTheScientific World Journal vol 2014 Article ID 702572 2014
[51] A B De Menezes E Lewis M OrsquoDonovan B F OrsquoNeill NClipson and E M Doyle ldquoMicrobiome analysis of dairy cowsfed pasture or total mixed ration dietsrdquo FEMS MicrobiologyEcology vol 78 no 2 pp 256ndash265 2011
[52] I K Skene and J D Brooker ldquoCharacterization of tanninacylhydrolase activity in the ruminal bacterium selenomonasruminantiumrdquo Anaerobe vol 1 no 6 pp 321ndash327 1995
[53] G Goel A K Puniya and K Singh ldquoTannic acid resistance inruminal streptococcal isolatesrdquo Journal of Basic Microbiologyvol 45 no 3 pp 243ndash245 2005
[54] P R Myer J E Wells T P L Smith L A Kuehn and H CFreetly ldquoCecum microbial communities from steers differingin feed efficiencyrdquo Journal of Animal Science vol 93 no 11 pp5327ndash5340 2015
[55] S E Hook A-D GWright and BWMcBride ldquoMethanogensmethane producers of the rumen and mitigation strategiesrdquoArchaea vol 2010 Article ID 945785 11 pages 2010
[56] M Saminathan C C Sieo H M Gan N Abdullah C M VL Wong and Y W Ho ldquoEffects of condensed tannin fractionsof different molecular weights on population and diversity ofbovine rumen methanogenic archaea in vitro as determinedby high-throughput sequencingrdquo Animal Feed Science andTechnology vol 216 pp 146ndash160 2016
[57] F Hassanat and C Benchaar ldquoAssessment of the effect ofcondensed (acacia and quebracho) and hydrolysable (chestnutand valonea) tannins on rumen fermentation andmethane pro-duction in vitrordquo Journal of the Science of Food and Agriculturevol 93 no 2 pp 332ndash339 2013
[58] J Balcells A Aris A Serrano A R Seradj J Crespo and MDevant ldquoEffects of an extract of plant flavonoids (bioflavex)on rumen fermentation and performance in heifers fed high-concentrate dietsrdquo Journal of Animal Science vol 90 no 13 pp4975ndash4984 2012
BioMed Research International 11
[59] A K Patra ldquoUreaammonia metabolism in the rumen andtoxicity in ruminantsrdquo Rumen Microbiology From Evolution toRevolution pp 329ndash341 2015
[60] MAWoznyM P Bryant L VHoldeman andW E CMooreldquoUrease assay and urease producing species of anaerobes in thebovine rumen and human fecesrdquo Applied and EnvironmentalMicrobiology vol 33 no 5 pp 1097ndash1104 1977
[61] D Jin S Zhao P Wang et al ldquoInsights into abundant rumenureolytic bacterial community using rumen simulation systemrdquoFrontiers in Microbiology vol 7 article no 1006 2016
[62] H P SMakkar B Singh andRKDawra ldquoEffect of tannin-richleaves of oak (Quercus incana) on various microbial enzymeactivities of the bovine rumenrdquo British Journal of Nutrition vol60 no 2 pp 287ndash296 1988
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 BioMed Research International
[28] E Jami and I Mizrahi ldquoComposition and similarity of bovinerumenmicrobiota across individual animalsrdquo PLoS ONE vol 7no 3 Article ID e33306 2012
[29] R De Nardi G Marchesini S Li et al ldquoMetagenomic analysisof rumen microbial population in dairy heifers fed a high graindiet supplemented with dicarboxylic acids or polyphenolsrdquoBMC Veterinary Research vol 12 no 1 article no 29 2016
[30] S Kruger Ben Shabat G Sasson A Doron-Faigenboim etal ldquoSpecific microbiome-dependent mechanisms underlie theenergy harvest efficiency of ruminantsrdquoThe ISME Journal vol10 no 12 pp 2958ndash2972 2016
[31] B R Min S Solaiman R Shange and J-S Eun ldquoGastroin-testinal bacterial andmethanogenic archaea diversity dynamicsassociated with condensed tannin-containing pine bark diet ingoats using 16S rDNA amplicon pyrosequencingrdquo InternationalJournal of Microbiology vol 2014 Article ID 141909 2014
[32] G Henderson F Cox S Kittelmann et al ldquoEffect of DNAextraction methods and sampling techniques on the apparentstructure of cow and sheep rumen microbial communitiesrdquoPLoS ONE vol 8 no 9 Article ID e74787 2013
[33] P R Myer T P L Smith J E Wells L A Kuehn and HC Freetly ldquoRumen microbiome from steers differing in feedefficiencyrdquo PLoS ONE vol 10 no 6 Article ID e0129174 2015
[34] Y Wang and T A McAllister ldquoRumen microbes enzymes andfeed digestion-a reviewrdquo Asian-Australasian Journal of AnimalSciences vol 15 no 11 pp 1659ndash1676 2002
[35] J B Russell R EMuck and P JWeimer ldquoQuantitative analysisof cellulose degradation and growth of cellulolytic bacteria inthe rumenrdquo FEMS Microbiology Ecology vol 67 no 2 pp 183ndash197 2009
[36] DOKrause S EDenmanR IMackie et al ldquoOpportunities toimprove fiber degradation in the rumen microbiology ecologyand genomicsrdquo FEMS Microbiology Reviews vol 27 no 5 pp663ndash693 2003
[37] M Zorec M Vodovnik and R Marinsek-Logar ldquoPotentialof selected rumen bacteria for cellulose and hemicellulosedegradationrdquo Food Technology and Biotechnology vol 52 no 2pp 210ndash221 2014
[38] A Z Bekele S Koike and Y Kobayashi ldquoGenetic diversityand diet specificity of ruminal prevotella revealed by 16S rRNAgene-based analysisrdquo FEMSMicrobiology Letters vol 305 no 1pp 49ndash57 2010
[39] A RamsakM Peterka K Tajima et al ldquoUnravelling the geneticdiversity of ruminal bacteria belonging to the CFB phylumrdquoFEMS Microbiology Ecology vol 33 no 1 pp 69ndash79 2000
[40] J Purushe D E Fouts M Morrison et al ldquoComparativegenome analysis of prevotella ruminicola and prevotella bryan-tii insights into their environmental nicherdquo Microbial Ecologyvol 60 no 4 pp 721ndash729 2010
[41] C A Carberry D A Kenny S Han M S McCabe and S MWaters ldquoEffect of phenotypic residual feed intake and dietaryforage content on the rumen microbial community of beefcattlerdquo Applied and Environmental Microbiology vol 78 no 14pp 4949ndash4958 2012
[42] A L Molan G T Attwood B R Min and W C McNabbldquoThe effect of condensed tannins from Lotus pedunculatus andLotus corniculatus on the growth of proteolytic rumen bacteriain vitro and their possible mode of actionrdquo Canadian Journal ofMicrobiology vol 47 no 7 pp 626ndash633 2001
[43] A Jayanegara G Goel H P S Makkar and K Becker ldquoDiver-gence between purified hydrolysable and condensed tannin
effects on methane emission rumen fermentation and micro-bial population in vitrordquo Animal Feed Science and Technologyvol 209 pp 60ndash68 2015
[44] BMinW Pinchak K Hernandez et al ldquoEffects of plant tanninsupplementation on animal responses and in vivo ruminalbacterial populations associated with bloat in heifers grazingwheat forage1rdquo The Professional Animal Scientist vol 28 no 4pp 464ndash472 2012
[45] T F Martınez T A McAllister Y Wang and T Reuter ldquoEffectsof tannic acid and quebracho tannins on in vitro ruminalfermentation of wheat and corn grainrdquo Journal of the Scienceof Food and Agriculture vol 86 no 8 pp 1244ndash1256 2006
[46] L Marın E M Miguelez C J Villar and F Lombo ldquoBioavail-ability of dietary polyphenols and gut microbiota metabolismantimicrobial propertiesrdquo BioMed Research International vol2015 Article ID 905215 18 pages 2015
[47] J B Russell and D B Wilson ldquoWhy are ruminal cellulolyticbacteria unable to digest cellulose at low pHrdquo Journal of DairyScience vol 79 no 8 pp 1503ndash1509 1996
[48] K Tajima S Arai K Ogata et al ldquoRumen bacterial communitytransition during adaptation to high-grain dietrdquo Anaerobe vol6 no 5 pp 273ndash284 2000
[49] D W Goad C L Goad and T G Nagaraja ldquoRuminal micro-bial and fermentative changes associated with experimentallyinduced subacute acidosis in steersrdquo Journal of Animal Sciencevol 76 no 1 pp 234ndash241 1998
[50] J Hernandez J L Benedito A Abuelo and C CastilloldquoRuminal acidosis in feedlot from aetiology to preventionrdquoTheScientific World Journal vol 2014 Article ID 702572 2014
[51] A B De Menezes E Lewis M OrsquoDonovan B F OrsquoNeill NClipson and E M Doyle ldquoMicrobiome analysis of dairy cowsfed pasture or total mixed ration dietsrdquo FEMS MicrobiologyEcology vol 78 no 2 pp 256ndash265 2011
[52] I K Skene and J D Brooker ldquoCharacterization of tanninacylhydrolase activity in the ruminal bacterium selenomonasruminantiumrdquo Anaerobe vol 1 no 6 pp 321ndash327 1995
[53] G Goel A K Puniya and K Singh ldquoTannic acid resistance inruminal streptococcal isolatesrdquo Journal of Basic Microbiologyvol 45 no 3 pp 243ndash245 2005
[54] P R Myer J E Wells T P L Smith L A Kuehn and H CFreetly ldquoCecum microbial communities from steers differingin feed efficiencyrdquo Journal of Animal Science vol 93 no 11 pp5327ndash5340 2015
[55] S E Hook A-D GWright and BWMcBride ldquoMethanogensmethane producers of the rumen and mitigation strategiesrdquoArchaea vol 2010 Article ID 945785 11 pages 2010
[56] M Saminathan C C Sieo H M Gan N Abdullah C M VL Wong and Y W Ho ldquoEffects of condensed tannin fractionsof different molecular weights on population and diversity ofbovine rumen methanogenic archaea in vitro as determinedby high-throughput sequencingrdquo Animal Feed Science andTechnology vol 216 pp 146ndash160 2016
[57] F Hassanat and C Benchaar ldquoAssessment of the effect ofcondensed (acacia and quebracho) and hydrolysable (chestnutand valonea) tannins on rumen fermentation andmethane pro-duction in vitrordquo Journal of the Science of Food and Agriculturevol 93 no 2 pp 332ndash339 2013
[58] J Balcells A Aris A Serrano A R Seradj J Crespo and MDevant ldquoEffects of an extract of plant flavonoids (bioflavex)on rumen fermentation and performance in heifers fed high-concentrate dietsrdquo Journal of Animal Science vol 90 no 13 pp4975ndash4984 2012
BioMed Research International 11
[59] A K Patra ldquoUreaammonia metabolism in the rumen andtoxicity in ruminantsrdquo Rumen Microbiology From Evolution toRevolution pp 329ndash341 2015
[60] MAWoznyM P Bryant L VHoldeman andW E CMooreldquoUrease assay and urease producing species of anaerobes in thebovine rumen and human fecesrdquo Applied and EnvironmentalMicrobiology vol 33 no 5 pp 1097ndash1104 1977
[61] D Jin S Zhao P Wang et al ldquoInsights into abundant rumenureolytic bacterial community using rumen simulation systemrdquoFrontiers in Microbiology vol 7 article no 1006 2016
[62] H P SMakkar B Singh andRKDawra ldquoEffect of tannin-richleaves of oak (Quercus incana) on various microbial enzymeactivities of the bovine rumenrdquo British Journal of Nutrition vol60 no 2 pp 287ndash296 1988
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 11
[59] A K Patra ldquoUreaammonia metabolism in the rumen andtoxicity in ruminantsrdquo Rumen Microbiology From Evolution toRevolution pp 329ndash341 2015
[60] MAWoznyM P Bryant L VHoldeman andW E CMooreldquoUrease assay and urease producing species of anaerobes in thebovine rumen and human fecesrdquo Applied and EnvironmentalMicrobiology vol 33 no 5 pp 1097ndash1104 1977
[61] D Jin S Zhao P Wang et al ldquoInsights into abundant rumenureolytic bacterial community using rumen simulation systemrdquoFrontiers in Microbiology vol 7 article no 1006 2016
[62] H P SMakkar B Singh andRKDawra ldquoEffect of tannin-richleaves of oak (Quercus incana) on various microbial enzymeactivities of the bovine rumenrdquo British Journal of Nutrition vol60 no 2 pp 287ndash296 1988
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology


















