
Identification of Nanoviruses in banana from Pakistan and ...
166
i Identification of Nanoviruses in banana from Pakistan and possible control through RNAi By Rohina Bashir M.Phil (UAF) A thesis submitted in partial fulfillment of requirements for the degree of DOCTOR OF PHILOSPPHY IN BOTANY Department of Botany Faculty of Sciences University of Agriculture, Faisalabad 2012
Transcript of Identification of Nanoviruses in banana from Pakistan and ...

i
Identification of Nanoviruses in banana from Pakistan and possible control through RNAi
By
Rohina Bashir
M.Phil (UAF)
A thesis submitted in partial fulfillment of requirements
for the degree of
DOCTOR OF PHILOSPPHY
IN
BOTANY
Department of Botany
Faculty of Sciences
University of Agriculture, Faisalabad
2012

i
The Controller of Examinations, University of Agriculture, Faisalabad. We, the supervisory committee, certify that the contents and form of this thesis submitted by Rohina Bashir D/o Bashir Ahmad Shad, Reg No: 2001-ag-1629 have been found satisfactory and recommend that it be processed for evaluation by the external examiner(s) for the award of the degree. I declare that this work is the result of my own investigation and has not already been accepted in substance for any degree, nor it is currently being submitted for any other degree All authors’ works referred to in this thesis have been fully acknowledged. SUPERVISORY COMMITTEE We certify that above statement is correct. 1) Chairman: _______________________________ (Dr. Farrukh Javed) 2) Co-supervisor: _______________________________
(Dr Shahid Mansoor) 3) Member: _______________________________
(Prof Dr. Abdul Wahid) 4) Member: _______________________________
(Dr. Rashid Ahmad)

ii
DECLARATION
I hereby certify that contents of the thesis “Identification of Nanoviruses in banana
from Pakistan and possible control through RNAi” research and no part has been
copied from any published source (except the references, some standard mathematical
or genetic models/equations/protocols etc.). I further declare that this work has not
been submitted for award of any other diploma/degree. The University may take
action if the above statement is found inaccurate at any stage.
________________________
Name: Rohina Bashir
Regd No. 2001-ag-1629

iii
TABLE OF CONTENTS
Chapter No.
Chapter Pages
Detailed contents Iv
Acknowledgements X
List of abbreviationss xi
List of figures xiii
List of tables xiv
1 Introduction 1
2 Review of literature 11
3 Materials and Methods 43
4 Results 57
5 Discussion 105
Conclusions 113
Future Prospects 114
Summary 115
Literature Cited 117

iv
DETAILED CONTENTS
ABSTRACT .............................................................................................................................. vii
ACKNOWLEDGMENTS ............................................................................................................. x
Chapter 1 ................................................................................................................................. 1
INTRODUCTION ....................................................................................................................... 1
1.1 General ............................................................................................................ 1
1.2 Control of Plant Viruses .................................................................................. 6
1.2.1 Cultural Practices ............................................................................................ 7
1.2.2 Conventional Resistance to Plants .................................................................. 8
1.2.3 Genetic Engineering ........................................................................................ 8
Chapter 2 ............................................................................................................................... 11
REVIEW OF LITERATURE ........................................................................................................ 11
2.1 Nanoviruses .................................................................................................. 11
2.1.1 Faba bean necrotic yellow virus (FBNYV) .............................................. 11
2.1.2 Milk vetch dwarf virus (MDV) .................................................................. 13
2.1.3 Coconut foliar decay virus (CFDV) .......................................................... 13
2.1.4 Banana bunchy top virus (BBTV) ............................................................. 13
2.2 Banana: The General Introduction ................................................................ 16
2.2.1 Soil and Climate for Banana Production ................................................... 16
2.2.2 Composition of Banana ............................................................................. 16
2.2.3 Medicinal Properties of Banana ............................................................... 17
2.2.4 World Banana Production ........................................................................ 17
2.2.5 Banana Cultivation in Pakistan................................................................. 17
2.2.6 Major Threats to Banana Cultivation in Pakistan ...................................... 17
2.2.7 Disease Management ................................................................................. 18
2.2.8 Application of Biotechnological Techniques in Banana ........................... 18
2.3 Genome Organization of BBTV ................................................................... 19
2.4 BBTV Satellite Molecules ............................................................................ 21
2.5 Replication of Circular ssDNA Viruses ........................................................ 22
2.6 Proteins Encoded By ssDNA ........................................................................ 23
2.6.1 Replication-associated protein (Rep) ........................................................ 23
2.6.2 Coat Protein .............................................................................................. 25
2.6.3 Nuclear Shuttle Protein ............................................................................. 26

v
2.6.4 Movement Protein ..................................................................................... 27
2.6.5 Cell Cycle Link Protein ............................................................................ 28
2.7 Genetic Approaches for Virus Resistance .................................................... 29
2.7.1 Non Pathogen Derived Resistance ........................................................... 29
2.7.2 Pathogen Derived Resistance ................................................................... 30
2.7.3 Protein Based Resistance .......................................................................... 31
2.8 RNA Interference (RNAi) ............................................................................ 32
2.8.1 History of RNAi ........................................................................................ 32
2.8.2 Discovery of RNAi ................................................................................... 33
2.8.3 Mechanism of RNAi ................................................................................. 34
2.9 Gene Expression System .............................................................................. 34
2.10 Suppressors Encoded By Plant Viruses ....................................................... 36
2.11 Promoters of ssDNA ..................................................................................... 37
2.12 Promoters Used in Transgenic Banana ........................................................ 40
Chapter 3 ............................................................................................................................... 43
MATERIALS AND METHODS ................................................................................................... 43
3.1 DNA Manipulation Techniques .................................................................... 43
3.1.1 CTAB method........................................................................................... 43
3.1.2 Polymerase Chain Reaction (PCR) ........................................................... 43
3.1.3 Rolling Circle Amplification (RCA) ........................................................ 43
3.1.4 Cloning of PCR Products ......................................................................... 44
3.1.5 Cloning of RCA Product .......................................................................... 44
3.1.6 Restriction Analysis .................................................................................. 44
3.1.7 Phenol Chloroform ................................................................................... 44
3.1.8 Plasmid isolation (Miniprep) .................................................................... 45
3.2 DNA Analysis Techniques ........................................................................... 48
3.2.1 Quantification of DNA ............................................................................. 48
3.2.2 Agarose Gel Electrophoresis .................................................................... 48
3.2.3 Southern Hybridization............................................................................. 48
3.2.3.1 Preparation of Radio‐Labeled Probe, Pre‐Hybridization, Hybridization and Signal
Detection ....................................................................................................................... 48
3.2.3.2 Biotin‐Labeled Probe Preparation, Pre‐Hybridiztion, Hybridization and Detection
of Probe ......................................................................................................................... 49
3.3 Microbiology Techniques ............................................................................ 51

vi
3.3.1 Preparation of Competent Cells of E. Coli ............................................... 51
3.3.2 Transformation of E. coli.......................................................................... 51
3.3.3 Preparation of Electro-Competent Cells of Agrobacterium ..................... 51
3.3.4 Transformation of Clones in Agrobacterium tumefaciens by Electroporation ......................................................................................................... 52
3.3.5 Agrobacterium-Mediated Infiltration of Host Plants ................................. 52
Chapter 4 ............................................................................................................................... 57
RESULTS ................................................................................................................................. 57
4.1 RFLP Analysis of Banana Infected Samples ................................................ 57
4.1.1 Phylogenetic Relationship of BBTV components from Pakistan Cloned from RCA Product ................................................................................................... 57
4.1.2 RFLP Analysis of Banana Infected Samples ............................................. 63
4.2 Transient Expression of BBTV Genes from PVX Vector ........................... 70
4.2.1 MRep-PVX ............................................................................................... 70
4.2.2 MP-PVX ................................................................................................... 71
4.2.3 NSP-PVX ................................................................................................. 71
4.2.4 Clink-PVX ................................................................................................ 72
4.2.5 CP-PVX .................................................................................................... 72
4.3 RNA Silencing Suppressors by BBTV ......................................................... 78
4.4 BBTV Promoter Study .................................................................................. 85
4.4.1 Cloning of Intergenic Region of BBTV Genes ........................................ 85
4.4.2. Transient Activity of Promoters in N. benthamiana.................................. 85
4.4.3 Cloning of Deleted Components of BBTV Genes ................................... 93
4.4.4 Transient Activity of Deleted Components in N. benthamiana ................ 93
4.5 BBTV Promoter Study in Monocots ............................................................. 97
4.6 Development of RNAi Construct against BBTV .......................................... 99
Chapter 5 ............................................................................................................................. 107
DISCUSSION……………………………………………………………………………….105
CONCLUSIONS…………………………………………………………………………….113
FUTURE PROSPECTS……………………………………………………………………..114
SUMMARY…………………………………………………………………………………115
LITERATURE CITED……………………………………………………………………...117

vii
ABSTRACT
Banana Bunchy Top Virus (BBTV) is a member of genus Babuvirus of the
family Nanoviridae, ssDNA virus transmitted by Pentalonia nigronervosa. Family
Nanoviridae is divided into two genera: Nanovirus and Babuvirus. Nanovirus
includes FBNYV, MDV, SCSV, while the genus Babuvirus include BBTV. In
Pakistan, banana production is under severe loss due to BBTV. In the absence of
natural resistance, the use of genetically engineered resistance is an attractive option.
The main objective of this study was to develop resistance in banana against banana
bunchy top virus through RNAi and the identification of unknown components of
BBTV by a new technique called Rolling Circle Amplification (RCA). Rolling circle
amplification (RCA) is a novel technique for the amplification of circular DNAs. This
technique has been widely used for the amplification of geminiviruses but its use for
the characterization of nanoviruses has not been reported. The identification of
unknown component is also necessary to find out whether any additional component
is associated with infectious unit or not. An analysis of the genetic diversity of BBTV
was made by this valuable technique across Tando Jam, Sindh, Pakistan, to
characterize components of banana bunchy top virus. The RCA product was digested
with several restriction enzymes and was resolved in agarose gel. The resulting RFLP
pattern resembled those expected for BBTV. In order to confirm the RFLP analysis,
the DNA was probed with cloned components of BBTV. The probes for components
DNA-S, DNA-N and DNA-M correctly hybridized to their respective fragment. We
further cloned two components of BBTV to verify results. The cloned components
were highly homologous to South Pacific group of BBTV as reported from Pakistan.
The results of present studies confirmed that RCA technology can be used for
characterization of nanoviruses. The technique is of great value to nanovirus research
since the components that make up this group are still being discovered. This diversity
(low) is also helpful in generating resistance against viruses. So, RNAi construct was
made against MRep of BBTV to engineer resistance against BBTV. This construct
was transiently checked in banana male flower bud. The buds agro-infiltrated with
EHA105 gave better expression as compared to GV3101.
Expression of BBTV genes from PVX and under 35S promoter was also
observed. Expression of MRep and MP under PVX resulted in necrosis and cell death
at the site of inoculation and severe leaf curling and necrosis in newly emerging

viii
leaves in MP. Clink, NSP and CP produced mild symptoms of leaf curling and
mosaic, while CP produced necrotic response in inoculated leaves. When all these
genes were expressed under 35S promoter in N. benthamiana 16c line, MP and Clink
stabilized GFP specific mRNA and reduced GFP specific siRNA. MRep, NSP and CP
did not show accumulation of GFP specific mRNA. These results identified that MP
and Clink are supressors of silencing. The ability of MP to induce severe necrosis in
inoculated and systemic leaves and RNA silencing suppressors indicates that MP is a
major pathogenecity determinant in BBTV genome.
Promoter regions of BBTV components may have application for
heterologous transgene expression. Promoter regions of BBTV components were
cloned in expression vector and checked it in N. benthamiana plants. Out of five
components of BBTV, DNA-S, DNA-C and DNA-R did not show any GUS
expression in N. benthamiana, while DNA-N showed some level of expression. The
deletion of 200bp from 5’ end of DNA-N increased the promoter activity but was still
low as compared to CaMV, 35S.

ix
This humble effort is dedicated
to
my Family

x
ACKNOWLEDGMENTS
All thanks are for “ALLAH” whose blessings enabled me to seek knowledge
and invigorate me for this task. Words can’t express my feelings of thankfulness for
“ALLAH” almighty. I offer my salutations to The Holy Prophet Muhammad
(PBUH) for the source of guidance for humanity as a whole forever.
At first, I would like to extend my heartiest gratitude to my supervisor, Dr
Farrukh Javed who made a great contribution for the successful completion of this
work. His skilful advices, sincere cooperation and learned guidance enabled me to
complete this work. I would like to thank my Co-supervisor Dr Shahid Mansoor
whose presence was always a source of confidence for me. I want to thank him from
the core of my heart for his personal interest, ample support, valuable guidance,
suggestions and help during this research work and writing of this manuscript. I shall
always be thankful to him for his affectionate behaviour towards me.
I am indebted to Mrs Javaria Qazi for her all time available cooperation,
encouragement and useful suggestions in accomplishment of this work. I am also
thankful to Dr Abdul Wahid and Dr Rashid Ahmad for their help and valuable
suggestions. I want to thank all my lab fellows at nibge for their help and sincere
cooperation. I want to acknowledge the most important people, the lab supporting
staff, Yasmeen and Ghulam Mustafa for their cooperation in lab work. I want to
thank my friends Sumera Anwer, Sumaira Ikram, Nusrat Parveen, Huma and
Shaista for their memorable company during the whole study.
At last, but not the least, I am indebted to all of my family members. No words
can pay my thanks to my parents for their prayers, encouragement, sacrifices and
moral support throughout my life and during this study. I am also very grateful to my
Brothers and Sisters for their moral support and prayers on a number of tough times
during my Ph.D studies. I am also very gratified to my in laws as during the last years
of my studies they helped and encouraged me a lot for the completion of this work. I
want to show my gratitude and respect for my life partner and friend
Muhammad Awais for his all time support and encouragement. I also
want to express gratitude to my pretty daughter Mashal as during the
last year of this work she was with me all the time and was a source
of inspiration and encouragement for compilation of this work.

xi
Rohina Bashir
List of abbreviations
μL micorlitre asRNA anti-sense RNA AZPs artificial zinc finger proteins BC Before Christ BSA bovine serum albumin CaCl2 calcium chloride cccDNA covalently closed circular DNA CIAP calf intestine alkaline phosphatase CLCuD cotton leaf curl disease CP coat protein CR common region CTAB cetyl trimethyl ammonium bromide DEAE diethylaminoethyl cellulose DNA deoxyribonucleic acid RNAi RNA interference dNTP deoxyribonucleotide triphosphate dsDNA double-stranded DNA dsRNA double-stranded RNA DTT dithiothreitol EDTA ethylene diamine tetraacetic acid FeSO4.7H2O ferrous sulphate hepta hydrate GFP green fluorescence protein GUS beta-glucuronidase hpRNA hairpin RNA HR hypersensitive response ICTV International Committee on Taxonomy of Viruses IPTG isopropyl-beta-D-1-thiogalactopyranoside IR intergenic region IRD iteron related domain K2HPO4 dipotassium phosphate KCl potassium chloride kDa kilo Dalton Kv kilo Volt LB Lauria broth LIR large intergenic region BBTV Banana bunchy top virus MCS multiple cloning site mg milligram MgSO4 magnesium sulphate MgSO4.7H2O magnesium sulphate heptahydrate miRNA microRNA mM millimolar MP movement protein mRNAs messenger RNA NaCl sodium chloride NaH2PO4 sodium phosphate

xii
NaOH sodium hydroxid ng nanogram NH4Cl ammonium chloride NSP nuclear shuttle protein nt. nucleotide ORF open reading frame PCR polymerase chain reaction PDR pathogen derived resistance pH paviour of hydrogen pre-miRNA precursor miRNA PVP polyvinyl pyrrolidone RCA rolling circle amplification RCR rolling circle replication RDR recombination-dependent replication RdRP RNA dependent RNA polymerase REn replication enhancer protein Rep replication associated protein RISC RNA-induced silencing complex RNA ribonucleic acid rpm revolutions per minute SCR satellite-conserved region SDS sodium dodecyl sulphate SIR small intergenic region siRNA small interfering RNA SSC standard sodium citrate ssDNA single-stranded DNA TAE tris-acetate EDTA Taq Thermus aquaticus ta-siRNAs trans-acting siRNAs TGS transcriptional gene silencing TrAP transcriptional activator protein T-Rep truncated Rep UV ultra violet VIGS virus induced gene silencing X-Gal 5-bromo-4-chloro-3-indolyl-b-D-galactopyranoside BAP Benzyl amino purine CaMV Cauliflower mosaic virus Hr Hour/hours GFP Green fluorescent protein µM micromolar µg microgram 0C degree centigrade

xiii
LIST OF FIGURES
Fig 2.1 Symptoms produced by Banana bunchy top virus (BBTV) 15
Fig 4.1 (a) Rolling Circle Amplification of BBTV samples. 58
(b) Cloning of RCA product into pTZ57R. 58
Fig 4.2 Phylogenetic dendrogram based upon a nucleotide sequence 59 alignment of selected BBTV DNA-N from NCBI database.
Fig 4.3 Phylogenetic dendrogram based upon a nucleotide sequence 60 alignment of selected BBTV DNA-M from NCBI database
Fig 4.4 RFLP analyzed restriction fragments of Phi product of BBTV. 62
Fig. 4.5 Southern hybridization of blots probed with DNA-S. 63
Fig. 4.6 (a) Southern blot analysis with probe of DNA-N. 64
(b) Southern hybridization of blots probed with DNA-M. 64
Fig 4.7 Genome organization of BBTV DNA-S. 65
Fig 4.8 Genome organization of BBTV DNA-N. 66
Fig 4.9 Genome organization of BBTV DNA-M. 67
Fig 4.10 Phenotypic behaviour of the expression of N. benthamiana plants 71 inoculated with (a) healthy control plant (b) PVX vector (c) PVX vector expressing GFP viewed under UV illumination (d) MRep-PVX
Fig 4.11 Phenotypic effects of the expression of BBTV genes from a PVX 72 vector in N. benthamiana (f,g) shows the plant infected with MP-PVX, (h,i) plants infected with NSP-PVX
Fig 4.12 Phenotypic effects of the expression of BBTV genes from a PVX 73 vector in N. benthamiana (j,k) shows the plant infected with Clink-PVX, (l) plants infected with CP-PVX Fig 4.13 Identification of BBTV gene products with PTGS suppressor 79
activity (a) shows the leaves from non-transgenic N. benthamiana plant, (b) a line 16c plant, (c) a leaf of a line 16c plant inoculated with RNAi-GFP and (d) and a leaf of a line 16c plant co-inoculated with RNAi-GFP and pGreen0029.
Fig 4.14 Plants co-inoculated with RNAi-GFP and constructs for the 80 expression of HC-Pro (e), TCV-CP (f), BBTV-NSP (g), BBTV-CP (h)
Fig 4.15 Plants co-inoculated with RNAi-GFP and constructs for the 81 expression of BBTV-MRep (i), BBTV-MP (j), BBTV-Clink (k)
Fig 4.16 Northern blot analysis of total RNAs extracted from inoculated N. 82 benthamiana plants with BBTV suppressors

xiv
Fig. 4.17 PCR amplification of MRep, Clink and NSP intergenic regions. 84
Fig 4.18 Cloning of intergenic regions in pTZ57R/T. 85
Fig 4.19 Cloning of intergenic region in vector designated as pGreen-GUS. 86
Fig 4.20 Cloning of Clink in vector pGreen-GUS. 87
Fig 4.21 Cloning of pJit166 into pGreen. 88
Fig 4.22 GUS assay for BBTV-NSP promoter. 89
Fig 4.23 GUS assay for BBTV-MRep promoter. 90
Fig. 4.24 A: PCR amplification of deleted MRep intergenic regions. 92
B: PCR amplification of deleted intergenic region of NSP. 92
Fig. 4.25 A: Cloning of deleted MRep intergenic region in pTZ57R/T. 93 B: Cloning of deleted NSP in pTZ57R/T. 93
Fig 4.26 GUS assay for N. benthamiana leaves for BBTV-NSP and
BBTV-MRep promoter. 94
Fig 4.27 GUS assay for Wheat callus. 96
Fig 4.28 PCR amplification of MRep-RNAi construct. 99
Fig 4.29 Confirmation of clones in pFGC5941. 100
Fig 4.30 Transient assay of banana male flower buds with RNAi construct. 101
Fig 4.31 In vitro multiplication of banana. 104

xv
LIST OF TABLES
Table 2.1 Proteins encoded by the integral genome components of BBTV. 20
Table 3.1 Composition of reagents used in DNA manipulation techniques. 46
Table 3.2 Composition of reagents used in DNA analysis techniques. 50
Table 3.3 Primers used during this study. 53
Table 3.4 Details of constructs produced during this study. 55
Table 3.5 Vectors used during this study. 56
Table 4.1 Summary of results of expression of BBTV genes from PVX 74
vector in N. benthamiana.
Table 4.2 Study of plants inoculated with BBTV genes in expression vector. 75
Table 4.3 Effect of different concentrations of BAP on multiplication 103
of banana explants.

1
Chapter 1 INTRODUCTION
1.1 General
The word, “virus” is a Latin word means “Poison”. Viruses are the obligate
intracellular parasites and they used host molecular machinery for their replication.
They can be transmitted horizontally between cells. The most reasonable definition of
virus is “a virus is a set of one or more nucleic acid template molecules, normally
encased in a protective coat or coats of protein or lipoprotein, which is able to
organize its own replication only within suitable host cells” Martinus Beijerinck
discovered the first virus in 1898 which was Tobacco mosaic virus (TMV)
(Beijerinck, 1898).
Most of the plant viruses (approximately 90%) have RNA (single-stranded or
double-stranded) as genetic material whereas some of the viruses have DNA genome
that can be single stranded (ssDNA) or double stranded (dsDNA). They require some
type of vectors for transmission from one host to other. Vectors include: fungi,
nematodes, insects (aphids, whiteflies, thrips, mites, leafhopper, and treehoppers).
Other means of transmission include pollen grains, seeds and soil. However, there are
several viruses that are transmitted by mechanical means and are relatively stable and
occur in higher concentration in infected leaves. Plant viruses have been characterized
on the basis of their host range, source of transmission, comparison of their nucleic
acid sequences and serological relationship. International Committee on Taxonomy of
viruses (ICTV) in its 8th report of classification and nomenclature of viruses has
approved 5 orders, 82 families, 11 subfamilies, 307 genera and 2083 species of
viruses.
There are many plant viruses that cause a loss of yield and vectors are the
major cause of their spread. So, efforts are made to control vectors or weeds that serve
as alternate hosts. Plant virus infection causes a huge loss in the yield and production
of important crops, thereby, affecting the economy of many countries in the whole
world. Plants have developed defense mechanisms to combat these viruses. These
defense mechanisms include R gene resistance mechanism and RNAi. R gene
resistance mechanism causes the cell death at the site of inoculation and thus inhibits
the spread of infection (Dinesh-Kumar et al., 2000). When viruses infect plant they

2
also cause the production of natural disinfectants like salicylic acid (SA), nitric oxide
(NO) and reactive oxygen species (ROS) (Soosaar et al., 2005).
Virus replication is dependent on the host cell machinery for their replication.
Changes in the nucleic acid of virus molecule give rise to variants of virus.
Caulimoviruses (F- Caulimoviridae) are dsDNA viruses, whereas Geminiviruses
(Geminividae) and Nanoviruses (Nanoviridae) are ssDNA viruses. All these viruses
cause a huge loss to economically important crops.
Geminiviruses is a large group of viruses that infect both monocots and dicots.
Therefore, viruses in this genus cause significant losses to economically important
crops. Geminiviruses have genomes of one or two circular ssDNA genome that is
packaged into icosahederal capsid (18-20 nm in diameter) and can be seen as twinned
particles under electron microscope. These viruses replicate through dsDNA
intermediates in the nucleus of host plants and are dependent on the host proteins for
their replication as they encode only a few proteins, which are not enough to carry out
these processes. These viruses are transmitted by insect vectors in a persistent,
circulative and non-propagated manner (Hanley-Bowdein et al., 1999; Gutierrez,
2000). Geminiviruses are often phloem limited and restricted to vascular tissues while
some of them can escape to mesophyll cells (Morra and Petty, 2000). Geminiviruses
consists of four genera, namely: Begomovirus, Mastrevirus, Topocovirus and
Curtovirus (Fauquet et al., 2008). All these genera differ in their host range, vector
and genome organization.
Geminiviruses comprise a large group of plant viruses that cause huge
economic losses and thus, they are a subject of vast distress worldwide. These viruses
use host replication machinery for their replication and encode only a few proteins for
their replication (Hanley-Bowdoin et al., 1999). Geminivirus group was established in
1979 (Matthews, 1979) and was upgraded to the family Geminiviridae in 1995
(Murphy et al., 1995). Geminiviruses include 199 species, of which 181 species
belong to Begomovirus. The complete nucleotide sequences of this group have been
submitted to databases (Fauquet et al., 2008), which indicates their vast diversity,
distribution and host adaptation.
Nanoviridae is considered as a separate family of plant viruses with a multiple
circular ssDNA genome components encapsidated in small icosahederal particles (18-
20 nm). Nanoviridae is separated into two genera: Nanovirus and Babuvirus. The

3
name Nanovirus is derived from Greek word “Nanos” means small refers to the fact
that these viruses are smallest among all known viruses. The genus nanovirus has
narrow host range and infects leguminous species whereas genus babuvirus infect
only banana plants and have no additional hosts. A first complete description of
nanovirus was given by Chu et al. (1995). Symptoms of nanoviruses include leaf
chlorosis, stunting and malformed fruit production. The vector for nanoviruses is
aphids that are transmitted in a circulative manner. These viruses are not mechanically
transmitted (Franz et al., 1998). The minimum feeding time for aphids to pick up
virus is from 15 to 30 min for FBNYV and 4 hours for BBTV. The inoculation time
for both viruses is 5-15 min and vector can transmit virus throughout its life span,
which may be the cause of its multicomponent genome (Hull, 2002; Hu et al., 1996).
Nanovirus components have a major ORF and they transcribed
unidirectionally. The coding region of each component contains a TATA box which is
followed by a polyadenylation signal. One of the components of nanovirus encodes
replication initiator protein (32.4-33.6 kDa) and other components encode non-Rep
proteins (Chu et al., 1995; Katul et al., 1997; Burns et al., 1995). All components of
nanoviruses have a stem loop region in their non-coding region. This feature of
nanovirus is similar for all nanoviruses and is also similar to geminiviruses. Banana is
a monocotyledonous, large herbaceous perennial crop in tropical and sub tropical
regions of the world. Banana is one of the world’s main crops with production of 70.7
m tonnes in 2006 (Anon, 2006). Latin America and Asia are the major banana
production areas and South America is the major banana exporter country. Most of
bananas are cultivated for local consumption and are very good source of
carbohydrates, fiber and the elements potassium, magnesium, phosphorous, calcium
and iron (Marriott and Lancaster, 1983). Bananas are staple food in rising countries
like Ethiopia and are also a source of income and employment to rural populations.
Banana belongs to Musa genus and Family Musaceae (Robinson, 1996).
Today’s edible bananas are hybrid of two diploid species Musa acuminata and Musa
balbisiana. The resultant hybrids are mostly triploid and sterile (Novak, 1992). The
most common triploid is Cavendish cultivar that is used today. The Cavendish
cultivar forms the basis of the world export banana trade, but the major threat to this
cultivar is the susceptibility to diseases. This cultivar is also cultivated in Pakistan,
where annual production is 209,820 metric tones (PARC, 2005).

4
Bananas are most susceptible to diseases caused by fungi, bacteria and viruses
(Jones, 2000). Bacterial diseases include Moko and blood caused by the bacteria
Ralstonia solanacearum have caused local crop loss (Taghavi et al., 1996; Ploetz et
al., 2003; Wicker et al., 2007). Viral diseases are more dangerous than bacterial and
fungal diseases in two respects. First is that the viral infection can not be eliminated
by chemotherapy, which means that control of viral diseases must be based on
preventing infection rather than on curing disease. The second is that viral damage to
plant is longer term than bacterial and fungal damage and has direct effect on growth
and yield. There are numerous viral diseases of banana, which include Banana bract
mosaic disease caused by Banana bract mosaic virus, Banana mild mosaic disease
caused by Banana mild mosaic virus, Banana mosaic disease caused by Cucumber
mosaic virus, banana streak disease caused by Banana streak virus and banana
bunchy top disease (BBTD) caused by Banana bunchy top virus (Dale, 1987;
Srivastava et al., 1995; Diekmann and Putter, 1996; Adams et al., 2004; Geering et
al., 2005; Teycheney et al., 2005). BBTD is one of the most important disease
infecting banana worldwide (Dale, 1987).
Recently, a new disease, Banana Bacterial Wilt disease has been reported from
Uganda. This disease has devastated over 50 districts in the country and destroyed 90
percent of banana farms. This disease is also mechanically transmitted. This disease
kills the plant and makes their fruit inedible. No banana variety is known to be
resistant to the disease and there is no chemical control effective against it. So this
disease can spread like wildfire (FAO, 2007).
BBTD has been reported from many areas including Africa, Asia and Pacific
region (Dale, 1987; Diekmann and Putter, 1996; Magnaye and Valmayor, 1995) while
Central and South America and the Caribbean are free from BBTD (Dale, 1987).
BBTD was first reported in 1889 in Fiji and caused huge loss to their industry. In
Pakistan, the area under banana cultivation was reduced from 23,000 hac to 11,300
hac and production from 209,800 tonnes to 44,200 tonnes due to BBTV epidemics
(Khalid and Soomro, 1993). As a result of which, farmers started growing bananas in
newer areas. This result in the spread of disease to new areas and this disease spread
in all areas of Sindh. The cause of this disease is Banana bunchy top virus (BBTV).
There is no cure for this disease and Cavendish cultivar is more readily infected with
this virus but there is no resistant variety.

5
RNA interference (RNAi) is concerned with the inhibition of viruses and
silencing of transposable elements in plants, insects, fungi and nematodes by small
interfering (si)RNAs (21-nt dsRNA) that are processed from dsRNA viral replication
intermediates (Waterhouse et al., 2001; Voinnet, 2001; Wang et al., 2005; Segers et
al., 2007). These siRNAs are loaded into RISC and target the fully complementary
viral RNAs for damage or translational repression (Haasnoot et al., 2003). RNaseIII-
type enzymes, termed Drosha and Dicer (DCR) in animals or DCR-LIKE (DCL) in
plants, catalyze processing of siRNA precursors to 21- to 24-nt duplexes (Baulcombe,
2004). Distinct pathways are concerned in the synthesis of siRNAs. Heterochromatin-
associated siRNAs (predominantly 24-nt) form through the activities of RDR2, RNA
polymerase IV, and DCL3 and require AGO4 for activity to direct or reinforce
cytosine methylation of DNA and histone H3 methylation at Lys-9 (Herr et al., 2005;
Onodera et al., 2005; Hamilton et al., 2002; Zilberman et al., 2003). Formation of
post-transcriptionally active siRNAs from exogenous (viral and transgenic) sources
may involve RDR1 or RDR6 and, for some viruses, DCL2 (Baulcombe, 2004).
Endogenous, trans-acting small interfering (ta-si) RNAs arise from PolII genes and
guide cleavage of target messenger RNAs (Allen et al., 2005; Peragine et al., 2004;
Vazquez et al., 2004). ta-siRNAs require RDR6 and suppressor of gene silencing 3
(SGS3) for precursor formation (Peragine et al., 2004; Vazquez et al., 2004). ta-
siRNA formation also requires DCL1, although the specific role of DCL1 may be
indirect (Allen et al.,2005; Peragine et al.,2004; Vazquez et al.,2004). All known
classes of endogenous small RNAs in Arabidopsis require HEN1, an RNA
methyltransferase that modifies the 3′ end (Yu et al., 2005). A. thaliana has three
known families of ta-siRNAencoding genes, designated TAS1, TAS2, and TAS3
(Allen et al., 2005; Peragine et al., 2004; Vazquez et al., 2004). The TAS1 family is
composed of three genes that encode a closely related set of ta-siRNAs (for example,
siR255 and siR480) that target four messenger RNAs encoding proteins of unknown
function (Allen et al., 2005; Peragine et al., 2004; Vazquez et al., 2004). TAS2-
derived ta-siRNAs (for example, siR1511) targets a set of messenger RNAs encoding
pentatricopeptide repeat proteins (Allen et al., 2005; Peragine et al., 2004). The TAS3
locus specifies two ta-siRNAs that target a set of messenger RNAs for several Auxin
response factors (ARFs), including ARF3 (ETTIN) and ARF4 (Allen et al., 2005;
Williams et al., 2005). Arabidopsis mutants with defects in RDR6 and SGS3 lack ta-
siRNAs and exhibit accelerated transition from juvenile to adult phase during

6
vegetative development (Peragine et al., 2004; Vazquez et al., 2004), suggesting that
ta-siRNAs regulate developmental timing, presumably through regulation of ta-
siRNA target genes.
Several lines of research indicate that RNA silencing is a general antiviral
defense mechanism in plants. The first indication came from the studies of pathogen
derived resistance (PDR) in plants. In PDR, resistance to a particular virus is
engineered by stably transforming plants with a transgene derived from the virus.
Eventually it became clear, that one class of PDR was the result of RNA silencing of
the viral transgene. Once transgenic RNA silencing had been established, all RNAs
with homology to the transgene were degraded, including those derived from an
infecting virus (Lindbo et al., 1993). Thus plant viruses could be the target of RNA
silencing induced by a transgene. The same work demonstrated that plant viruses
could also induce RNA silencing. VIGS can be targeted to either transgenes or
endogenous genes (Ruiz et al., 1998) and the technique has been used to screen for
gene function using libraries of endogenous sequences cloned into a viral vector
(Vance and Vaucheret, 2001). Transient expression of reporter genes encoding either
green fluorescence protein (GFP) or red fluorescent protein from Discosoma was
specifically reduced by 58% and 47%, respectively, at 24 h after codelivery of
cognate siRNAs in BY2 protoplasts. In contrast to mammalian systems, the
siRNAinduced silencing of GFP expression was transitive as indicated by the
presence of siRNAs representing parts of the target RNA outside the region
homologous to the triggering siRNA. Codelivery of a siRNA designed to target the
messenger RNA encoding the Rep of the geminivirus ACMV from Cameroon
blocked Rep messenger RNA accumulation by 91% and inhibited accumulation of the
ACMV genomic D by 66% at 36 and 48 h after transfection. As with siRNA-induced
reporter gene silencing, the siRNA targeting ACMV Rep was specific and did not
affect the replication of EACMCV.
1.2 Control of Plant Viruses
Viruses cause considerable losses to crops and there is a number of ways to
control this problem. But the correct identification of the virus infecting a particular
crop is necessary for the control of virus. Most virus diseases are mainly caused
directly or indirectly by human activities (Thresh, 2006). Some of points that cause
epidemics include:

7
Introduction of viruses into new area by transporting infected planting material
or seeds
Repeated use of same area for the same crop
Introduction of virus vectors into new areas
Introduction of new variety into field which is susceptible to disease
Use of monocultures instead of polycultures
These are practices to control viruses.
1.2.1 Cultural Practices
Seed is an important source of virus infection. If the contaminated seed is
grown then it will introduce the virus into plant at an early stage, allowing the
infection to be transmitted to nearby plants at an early stage. When the seed is only
source of virus infection, the use of virus free seeds provide an effective means for the
control of disease e.g infected tomato seeds carries the virus on the surface of seed
coat. As the seed germinates the virus is transmitted to cotyledons and ultimately into
plant.
The use of virus free propagating material is also helpful in control of virus.
As in case banana, the major cause of the disease is the propagating material. The use
of infected planting material may cause the spread of disease to other areas. Tissue
culture is a most useful method for the production of virus free plants. Cultures from
apical meristem may lead to the production of virus free plant from infected plants.
BBTV infected plants of Cavendish dwarf were exposed to 40oC for 16 hours for 12
weeks, but this failed to eliminate the virus. However, culturing plants for 3 months at
35oC gave some healthy plants (Wu and Su, 1992).
A change in the sowing date of planting is also effective in the control of
virus. The best time for the sowing will depend on the time of vector migration. If
vectors migrate early, then late sowing will be preferred. The early sowing of crop
will help plants to grow in a healthy environment until the vector is introduced into
the field when plants will be less susceptible to virus. Closely spaced plants are less
susceptible to virus infection than are widely spaced ones because aphid densities are
higher in widely spaced plants. The grain yield in rice was much higher significant in
closely spaced fields in which the incidence of rice tungro disease was reduced
compared to conventionally planted fields (Shukla and Anjaneyulu, 1981).

8
It is clear that there will be no virus problem in the crop if the field is free of
virus sources. Additional hosts of viruses may include weeds, ornamental plants etc.
Weeds are the major source of virus infection. Plant remains of previously infected
plants in the soil are also major source of infection in plants. Ploughing and burning
of these materials is best way to control the spread of virus. Sometimes, it is better to
remove the infected plants from field. If the virus spread is slow and within the crop
then rouging is beneficial. Sometimes, the virus also spread from infected to healthy
plants in a single field, so the eradication of the infected material is helpful.
1.2.2 Conventional Resistance to Plants
Conventional breeding is the introduction of resistant genes into crop plants is
one of the best solutions to virus problems. There are many difficulties in the
conventional breeding programmes to develop resistance. Sometimes source of
resistance gene is not available among crops and if it is available in wild species it is
sometime difficult to introduce into cultivated crops due to incompatibility at the
nuclear or cytoplasmic level.
In banana, conventional resistance has not proved successful in improvement
of the crop, due to triploidy, low fertility, long generation time and large area
requirement for field testing. Furthermore, banana has a narrow genetic base which is
impoverished in respect to genes for disease resistance. Some disease resistant genes
may be found among wild species of banana cultivars but these species are not
suitable for commercial banana production. Musa acuminata is a wild, diploid or
triploid banana which produces an extremely low quantity of inedible fruit and only
some accessions are resistant to pathogens, including Foc and Mycosphaerella
fijiensis & musicola (Gowen, 1995).
1.2.3 Genetic Engineering
Recombinant DNA and tissue culture technologies offer alternative means of
generating resistance in plants. Banana transformation procedure will help us to
overcome many problems associated with conventional breeding programme and will
ultimately lead us to introduction of resistant genes into susceptible banana cultivars
(May et al., 1995; Sagi et al., 1995a).
Tissue culture is also a very useful technique for germplasm conservation,
multiplication and generating resistance in plants against viruses (Azam et al., 2010).

9
Banana production is increasingly suffering losses due to viral diseases in many
banana growing countries and the major cause of virus spread is the propagation
material. The successful solution to this problem is production of virus free banana
through tissue culture and then the introduction of plants were introduced into field
(Resmi and Nair, 2011). This is a cheaper and easier method for multiplication of
bananas and also for production of virus free banana. There are also chances of virus
transmission in tissue culture technique if the source plant is virus infected.
Micropropagation in banana is mostly done by the shoot tip culture method.
Cytokinin type and its concentration effect rate of shoot proliferation and elongation
(Resmi and Nair, 2011). However, micropropagation of the male flower can help to
increase the efficiency compared to micropropagation of soil born suckers and the
chances of virus entry into plants is also decreased (Darvari et al., 2010). Application
of this tissue culture technique for banana plantlet production benefited our growers
and helped us to produce disease free banana and also rapid multiplication of banana
(Hwang and Su, 1998).
Before 1990, banana viruses were detected visually by symptom appearance
and electron microscope. But there are chances of error in these techniques. In last
decade, the advances in biotechnological techniques led to the development of new
sensitive methods for virus detection like PCR. Recently, five components of BBTV
in Pakistan have been cloned by PCR (Amin et al., 2008). In addition to
micropropagation techniques, gene silencing techniques like RNA interference
(RNAi) will prove successful in the development of resistance against BBTV, as
RNAi has been successfully used in virus resistance in many crops (Watanabe et al.,
2006; Fusaro et al., 2006). The development of RNAi construct requires the complete
sequencing of BBTV components. This information will be helpful in the
development of resistance strategies against BBTV.
Banana bunchy top disease continues to cause heavy yield losses to banana
production in Pakistan. In the absence of sources of natural resistance, the use of
genenetically-engineered resistance is an attractive option. Several options are
available for genetically-engineered resistance and among them RNA silencing based
technologies are likely to result in viable resistance. However, before embarking on
such program, it is important to know the diversity of the pathogen, viral genes that
are important for pathogenicity and suppressors of RNA silencing encoded by BBTV
complex prevalent in Pakistan. So, this thesis was aimed at understanding these

10
aspects of BBTV so that prospects of genetically-engineered resistance can be
explored.

11
Chapter 2 REVIEW OF LITERATURE
2.1 Nanoviruses
Nanoviruses were previously known as non-geminated ssDNA viruses (Chu et
al., 1995). But recently they were established as a separate genus (Randles et al.,
2000). Nanoviruses include four species: Suberranean clover stunt virus (SCSV)
(Chu and Helms, 1988), Faba bean necrotic yellow virus (FBNYV) (Katul et al.,
1997, 1998), Milk vetch dwarf virus (MDV) (Sano et al., 1998) and Banana bunchy
top virus (BBTV) (Harding et al., 1991). Coconut foliar decay virus (CFDV) may
also be considered as a tentative member of nanovirus (Rohde et al., 1990). The first
complete description of these viruses was given by Chu et al. (1995). All these viruses
are single stranded with a multipartite genome. All ssDNAs of nanoviruses are
individually encapsidated within small isometric virions of 18-20 nm in size (Franz et
al., 1998) and are transmitted by aphids. Functions of a few nanovirus proteins have
been studied. Of five proteins, functions of master Rep (MRep) (Timchenko et al.,
2000) and the cell cycle link protein (Clink) (Aronson et al., 2000) are well studied.
Other nanovirus proteins include a movement protein (MP) and a nuclear shuttle
protein (NSP), which are very much similar to the geminiviruses (Lazarowitz and
Beachy, 1999; Wanitchakorn et al., 2000). Functions of all other proteins have not
been identified yet. FBNYV requires a helper protein for aphid transmission,
however, the nature of that protein is also not identified yet (Franz et al., 1999).
2.1.1 Faba bean necrotic yellow virus (FBNYV)
Faba bean is the fourth most important pulse crop in the world, is a source of
protein in the human diet, and its dry seeds are used as animal feed. The total area
under faba bean cultivation in China is 1,100,000 ha with the annual production of
2,450,000 metric tones (FAO, 2007) Many viruses infect faba bean crops world wide
(Makkouk et al., 2003; Kumari and Makkouk, 2007). Out of these, FBNYV is the
economically most important virus infecting several legume crops in African and
Asian countries (Makkouk and Comeau, 1994). This virus has a wide host range and
so far more than 50 plant species belonging to Fabaceae have been identified as hosts
for this virus (Katul et al., 1993; Franz et al., 1997). Symptoms of the disease include

12
stunting and leaf yellowing. This virus is transmitted by aphids: Aphis craccivora and
Acyrthosiphon pisum (Franz et al., 1998).
In the Egyptian isolate, ten genome components of FBNYV have been
reported and designated as C1- C10, four of which encode Rep proteins and six
encode non-rep proteins. The Syrian isolate also contains ten ssDNA components
(Katul et al., 1995, 1997). A single Rep protein i.e MRep initiates the replication of
all other components and none of other Rep protein was able to initiate the replication
of other components. These Rep like components were identified from different
isolates of FBNYV. The identification of four different Rep components from two
different geographical isolates raised the question as to what extent these components
are associated with other isolates of FBNYV. Therefore, 55 different samples from
different geographical areas were analyzed. In all these samples only rep 2 like
components was detected in each sample, which indicates that rep 2 encodes the
master replication associated protein of FBNYV. Both Syrian and Egyptian isolates
were similar in their nucleotide sequence (94%) while there was striking difference in
their amino acid sequence in coding (61%) (Katul et al., 1998). The C10 component
of FBNYV contained amino acid motif LXCXE which is also present in Rep A
protein of Mastreviruses and is required for efficient virus replication (Katul et al.,
1998).
Recently, another nanovirus, Faba bean necrotic stunt virus (FBNSV) was
reported from Ethiopia. This virus is amplified by using rolling circle amplification
(RCA). This virus produced different symptoms from those of FBNYV, which
include leaf rolling, severe plant stunting and apical necrosis. This virus is
serologically different from FBNYV Syrian and Egyptian isolates and also from Milk
vetch dwarf virus (MDV) (Grigoras et al., 2009). Like other nanoviruses, this virus
also contains eight components of 1 kb. Each genomic DNA contains a common
region stem loop structure (CR-SL) and a single ORF which is preceded by TATA
box and followed by a polyadenylation signal (Burns et al., 1995; Katul et al.,1998;
Sharman et al., 2008; Timchenko et al., 2000). DNA components of FBNSV are more
closely related to FBNYV and MDV than to SCSV. MRep is the most conserved
protein among all these nanoviruses (83% amino acid sequence identity with SCSV
and 54-57% with babuvirus). FBNSV, FBNYV and MDV share about 80-84% amino
acid sequence similarity in CP and less than 75% in other proteins of unknown
functions. In contrast to FBNYV, the host range of FBNSV is limited (Grigoras et al.,

13
2009). The FBNYV was the first virus for which infectivity was achieved by cloned
DNAs (Timchenko et al., 2006) and now FBNSV infectivity has also been achieved.
This infectivity assay showed that all eight components of FBNSV are required for
disease symptoms (Grigoras et al., 2009). In FBNYV, an additional helper component
for aphid transmission has been identified but the nature of that component remains to
be identified (Franz et al., 1999).
2.1.2 Milk vetch dwarf virus (MDV)
MDV is also composed of ten ssDNA. Components C1, C2, C3, and C10
encoded putative Rep associated proteins. These rep proteins showed similarity with
other rep components associated with FBNYV, SCSV which indicate that all these
viruses may have a common origin. FBNYV and MDV are more closely related
species than SCSV. Other proteins encoded by MDV contain the retinoblastoma
binding motif LXCXE suggesting that they may be involved in controlling host cell
cycle. MRep protein helps in the replication of non-rep DNAs, which have been
tested in various combinations (Sano et al., 1998).
2.1.3 Coconut foliar decay virus (CFDV)
Coconut foliar decay virus (CFDV) is a tentative member of the genus
Nanovirus with an ssDNA genome (Randles et al., 2000). The only known genome
component is of 1291 nucleotides and contain six open reading frames (Rohde et al.,
1990). The largest frame encodes the replication initiator protein, which is different
from other nanoviruses in its slightly larger size. The sequenced CFDV DNA is
similar to Rep encoding components of other nanoviruses. This virus is also phloem-
specific like other nanoviruses (Merits et al., 2000).
2.1.4 Banana bunchy top virus (BBTV)
The initial symptoms of BBTD include “morse code” pattern that is formed by
dark green streaks in the lower portion of midrib, leaf stem and floral bracts. These
streaks are irregular and resemble a series of dots and dashes. Disease progression
results in the leaves at the apex of the plant becoming short and narrow with chlorotic,
brittle leaf margins that tend to curl upward. Plants that are infected late in the season
may produce fruit that is small and distorted, while plants infected early give the
plants a bunchy appearance.

14
Infection caused by BBTD may be primary or secondary. Primary infection is
caused by infected plant material and it produces more severe symptoms that result in
the production of dwarf plants of about one metre that may not produce any fruit.
Secondary infection is caused by virus transmitted by vectors to plants that are
initially healthy. Secondary infection causes symptoms that are mild compared to
those of primary infection (Magnaye and Valmayor, 1995; Dale et al., 1986).
However, symptoms produced by the BBTD are distinctive across all Musa sp, but
cases of mild and symptomless banana have been reported from Taiwan (Diekmann
and Putter, 1996). However, no natural resistance in any banana cultivar has been
found (Magnaye and Valmayor, 1995).
BBTV infects Musa species like Musa acuminate, M. balbisiana, M. coccinea,
M. ornate, M. textiles, M. velutina (Espino et al., 1993; Thomas and Caruana, 2000;
Furuya et al., 2003). Other hosts of BBTV include Alpinia purpurata (red ginger) and
Colocasia esculenta, which have been reported to be symptomless hosts in India
(Ram and Summanwar, 1984). Canna indica and Hedychium coronarium have been
reported as hosts of BBTV in Taiwan, but in Australia and Hawaii (Su et al., 1993;
Hu et al., 1996).
BBTD is not mechanically transmitted but is transmitted locally by the aphid,
Pentalonia nigronervosa. The long distance spread of this disease is caused by the
transport of infected planting material. Aphids mainly colonize on banana but
alternate hosts include Heliconia, Strelitzia and Ravenala (Magnaye and Valmayor,
1995). Aphids also colonize for a short period on plants in the genera Zingiber and
Canna (Magnaye and Valmayor, 1995). Banana aphids require a feeding period of at
least 12 hours to become infective (Hafner et al., 1993; Hu et al., 1996). The virus
remains in the aphid after molting but it is not transferred to progeny (Magnaye and
Valmayor, 1995).

15
A
B
Fig 2.1 Symptoms produced by Banana bunchy top virus (BBTV) (A) Showing the morse code symptoms as short dark, parallel streaks.
(http://www.ctahr.hawaii.edu/banana/morse_code.asp)
(B) Showing banana aphid Pentalonia nigronervosa
(http://www.agnet.org/images/library/nc122b4.jpg)

16
2.2 Banana: The General Introduction
Banana is used as a clip food for 400 million public in many mounting
countries like Uganda. This crop is under constant threat of large numbers of biotic
and abiotic stresses (Msogoya and Grout, 2008). Among biotic stresses, BBTV
represents a major constrain, limiting banana production worldwide. Ecuador is the
major banana export country and export about 80% of banana exports, the United
States is the major banana importing country (FAO, 2004). India is the major banana
producer country growing about 11,000,000 metric tones, while banana production in
Pakistan is 95,000 metric tones (FAO, 2004).
2.2.1 Soil and Climate for Banana Production
Sindh is a main banana producing country in Pakistan. The best soil and
climate requisite for banana production is present in Sindh. Banana can grow up in all
types of soils with adequate amount of moisture (Robinson, 1996). Well drained and
friable loamy soil, but not fine sand, which holds water, is favorable for its growth
(Stover and Simmonds, 1987).
Banana can grow from sea level to 1200m in elevation and from 15oC to 40oC.
It is basically a plant of the humid tropics and can grow from the humid tropics to dry
climates. Colder climates are limiting factors for banana growth because the crop
needs more time for maturation. Warm winds blowing in the summer break the
foliage of banana plants. On the average, 10mm rainfall is enough for the
development of banana plants (Ikisan, 2000).
2.2.2 Composition of Banana
Banana is the cheapest, abundant and most nourishing of all fruits. It is a rich
source of carbohydrates and provides energy similar to vegetables. They are suitable
for children because of their digestibility and high mineral content. Bananas are very
tender fruits providing more calories. Because of its high potassium content, they help
to maintain water in animal bodies by counteracting sodium. Bananas are suitable for
those who want to reduce their weight. It is also helpful for the students to increase
their memory. Bananas are free from substances that provide uric acid and thus best
for patients with gout or arthritis.

17
2.2.3 Medicinal Properties of Banana
Banana is rich in iron and thus helps in the production of hemoglobin. The
unique feature of hemoglobin is that it is high is sodium contents and low in
potassium contents. A research report by the University of Michigan Medical Centre
in 2010 found that the lectin present in banana is useful in inhibiting the HIV
infection by blocking the virus entry into the body. According to a newsletter
published in 2003 by INIBAP, a natural extract from pseudostem of banana is useful
in cure of cancer. In Africa, the peels of banana are used to make poultice for wounds.
2.2.4 World Banana Production
98% of world banana production is in developing countries. In 2007, 130
countries produced bananas. The 10 major banana producing countries accounted for
more than 75% of the total banana production. Furthermore, India, China, the
Philippines, Brazil and Ecuador alone produced more than 60% of the total world
banana production. Asia began to increase banana production and finally reach to
58% of total production in 2007 (FAO, 2007). The banana cultivation increased up to
95.60 million tones in 2009 from 66.84 m tonnes in 2001. Total banana production is
app. 100 m tonnes (FAO, 2007).
2.2.5 Banana Cultivation in Pakistan
Banana is one of the most significant fruit crops of Pakistan. It is mainly
cultivated in the Sindh province. It is grown on an area of 23.500 ha with an annual
production of 209,820 tonnes (PARC, 2005). About 90% of area under banana
cultivation in Pakistan is planted with a single cultivar, Cavendish dwarf, locally
known as Basrai. Banana is propagated in Pakistan by suckers from parent plants.
This unmonitored practice is a major cause of disease. So, it is necessary to check for
the presence of off-types of diseases.
2.2.6 Major Threats to Banana Cultivation in Pakistan
In December 1988, a farmer observed unusual symptoms on banana plants in
the Thatta district of Pakistan. These plants were brought to Sindh Agricultural
extension staff in May, 1989. Later on, this disease was diagnosed as Banana bunchy
top disease (BBTD) caused by virus known as Banana bunchy top virus (BBTV)
(Khalid et al., 1993). Due to this disease, the banana cultivation area was reduced
from 23,500 ha to 11,300 ha and production from 209,800 tonnes to 44,200 tonnes by

18
the end of 1992 (Khalid and Soomro, 1993) due to this virus. Farmers propagated the
infected suckers for further cultivation which is major cause of spread of disease. This
resulted in the spread of disease to newer areas where it was not found previously.
The farmers have no choice and cultivate the infected suckers as disease free banana
was not found. A number of factors are playing roles in the spread of disease
including the un-availability of virus free banana, the presence of vectors and a lack
of awareness of farmers regarding the clean cultivation and management of fields for
virus control (PARC, 2005).
2.2.7 Disease Management
When BBTV is prevalent in banana growing areas, it causes a huge loss in
production of banana. A comprehensive management, strict inspection and
eradication are essential. If the detection and eradication program continues then
spread of disease can be controlled to some extent. The major cause of spread of
disease is aphids. Thus, the control of aphids requires the spraying of plants with
kerosene emulsion. Vandenveken, 1977 suggested that kerosene oil modifies the
charge of stylets, which inhibits the adsorption and elusion of virus particles. Banana
virus can be eliminated by means of heat and chemotherapy from infected plants. If
the diseased mother plant is kept at 36 oC for 4 months, the virus can be eliminated in
up to 53.33% of diseased plants. As far as chemotherapy is concerned, virazol will
eradicate BBTV better than 2-thiouracil (Abdel-Aziz et al., 1998). But in
micropropagation these managements are not required because micropropagation
results in the isolation of BBTV free plant tissue for culture (Wu and Su, 1992).
Wellings et al. (1994) proposed that the introduction of Aphidius colemani to banana
plants result in the control of the banana aphid, Pentalonia nigronervosa.
2.2.8 Application of Biotechnological Techniques in Banana
A large number of techniques are being used worldwide to improve the banana
germplasm. Tissue culture is being widely used for the multiplication of banana and
germplasm exchange (Crouch et al., 1998). Anther culture method is useful in
conventional breeding programmes. Assani et al. (2003) reported successful
regeneration of haploid plants of Musa balbisiana (BB), which is known to carry
resistant genes against economically important banana diseases. Protoplast fusion
techniques were put forwarded by Matsomoto et al. (2002), to be useful in
introducing disease resistance from wild species to cultivated varieties. Mutation

19
breeding could also be used in genetic improvement of banana. So far, there has been
no practical implementation of mutation breeding for banana. Bhagwat and Duncan
(1998) used gamma radiations on explants of in vitro grown cultures of banana to
produce variant resistant to the fungus Fusarium oxysporum. Somatic embryogenesis
techniques are also aimed at developoing new, high performance micropropagation
techniques and cell regeneration systems useful for genetic transformation and
cultivar improvement (Kosky et al., 2002).
Molecular techniques have been applied to detect somaclonal variations in
banana. Ramage et al. (2004) used random amplified polymorphic molecular marker
(RAPD) to detect dwarf off types in micropropagated Cavendish bananas (AAA
group). Molecular markers were also used to study the diversity by genotyping and
mapping of Musa species (Kaemmer et al., 1997). In addition to RAPD, SSRs can
also be used to study the diversity. Creste et al. (2004) used microsatellite markers to
study the genetic diversity of banana. AFLP and VNTR can also be used to study the
polymorphism in banana.
2.3 Genome Organization of BBTV
BBTV has 18-20 nm isometric virions and a multicomponent genome of
circular ssDNA (Harding et al., 1991, 1993; Burns et al., 1994, 1995). The genome of
BBTV consists of at least six components, all approximately 1 kb in size (Harding et
al., 1993; Burns et al., 1995). Each DNA component contains the stem loop common
region (CR-SL) and the major common region (CR-M) (Burns et al., 1995). The stem
loop common region consists of 69 nucleotides which is 62% identical between all
components of BBTV and incorporates a putative stem loop structure. The loop
sequence has the highly conserved nanonucleotide sequence similar to all
geminiviruses (Lazarowitz, 1992). The major common region incorporates 66-92
nucleotides that is 76% identical between components (Burns et al., 1995). Each of
the six components also contains one open reading frame (ORF) in the virion sense
(Burns et al., 1995) with a potential TATA box 3’ of CR-SL and with associated
polyadenylation signals.
The large ORF (856nt) of DNA 1 encodes a replication protein (Rep) of 33.6
kDa (Harding et al., 1993). This DNA 1 has been isolated from all BBTV infected
plants from 11 countries (Karan et al., 1994). In addition to this, additional DNA
components have been isolated from Taiwan isolates that encode putative Reps and

20
these Reps are different from the Rep of DNA 1 (Yeh et al., 1994; Wu et al., 1994).
Further, these components are different in their organization from BBTV DNA 1-6
(Burns et al., 1995)
BBTV isolates are divided into two groups based on sequence comparisons of
DNA 1, 3 and 6. The South Pacific group comprises isolates from Australia, Burundi,
Egypt, Fiji, India, Pakistan, Tonga and Western Samoa while the Asian group
comprises the isolates from Philippines, Taiwan and Vietnam (Karan et al., 1994;
1997; Wanitchakorn et al., 2000). Interestingly, all BBTV components have been
identified in all isolates tested from both the South Pacific and Asian groups,
indicating that all six components of BBTV are the integral part of the genome (Karan
et al., 1997). BBTV genome encodes all proteins in a virion sense. BBTV DNA R
contains two ORFs, one internal to the other; the major ORF encodes a Rep protein
(MRep) (Hafner et al., 1997). DNA R of BBTV has been identified in all isolates. The
encoded MRep protein helps in the replication of all other components of BBTV
(Horser et al., 2001). Satellite DNA also encodes a Rep protein but it does not help in
the replication of integral components of BBTV. DNA S encodes viral coat protein
(Beetham et al., 1999) and DNA M encodes movement protein (MP) with a
hydrophobic N-terminus (Wanitchakorn et al., 1997). DNA C encodes Clink proteins,
which have a LXCXE binding domain, which binds to plant retinoblastoma binding
protein (pRb). DNA N encodes the nuclear shuttle protein (NSP) (Wanitchakorn et
al., 2000).
Table 2.1 Proteins encoded by the integral genome components of BBTV.
Component Size (bp)
Functions of encoded proteins
Predicted coding capacity of encoded proteins (amino acids)
DNA R 1111 MRep 286 DNA S 1075 CP 175 DNA M 1042 MP 117 DNA C 1018 Clink 161 DNA N 1096 NSP 156

21
2.4 BBTV Satellite Molecules
The first satellite DNA molecule which was 262 nucleotide ssDNA was found
in a ToLCV isolate from Northern Australia (Dry et al., 1997). The replication of the
satellite molecule is dependent upon host replication associated proteins and is
encapsidated by the helper virus coat protein. Satellite molecules have no ORF. Other
begomoviruses also have several non-viral ssDNA like African cassava mosaic virus
(ACMV), Tomato yellow leaf curl virus (TYLCV) and Beet curly top virus (BCTV).
Several monopartite begomoviruses such as Cotton leaf curl Multan virus (CLCuMV)
and Ageratum yellow vein virus (AYVV) are associated with nanovirus-like
components similar to nanovirus a component that encodes Rep proteins (Mansoor et
al., 1999; Saunders and Stanley, 1999). AYVV also contains another ssDNA called
betasatellite which is approximately half the size (1347 nucleotides) of DNA A
(Saunders et al., 2000). This satellite is dependent on DNA A for its replication and it
is encapsidated by coat protein encoded by DNA A. Both of these characteristics are
similar to satellite molecules. The symptoms in Ageratum conyzoides are dependent
on co-inoculation of DNA A and betasatellite, thus nanovirus like satellite has no
effect on symptom development (Saunders et al., 2000).
In BBTV, in addition to six components, other ssDNA, DNA S1, S2, S3, Y,
W1 and W2, are found in some isolates so they are named as satellite molecules of
BBTV. DNA S1, S2, W1, W2 and Y were originally found in Taiwanese isolates of
BBTV (Horser et al., 2001; Yeh et al., 1994; Wu et al., 1994) while S3 was originally
found in Vietnam isolates of BBTV (Bell et al., 2002). These satellite molecules are
not present in all BBTV infected areas, despite that this disease is highly prevalent in
Asian countries. These satellite molecules were detected only from South Pacific
group from Tonga and Samoa (Horser et al., 2001). DNA sequence of S2 and W2
indicated that they are homologous so they are considered to be same component.
Sequences of DNA Y1 and W1 are also the same (Horser et al., 2001).
Six integral components of BBTV and satellite molecules have some similar features.
1. All components are of 1 kb in size.
2. TATA box is conserved.
3. Transcription termination elements are conserved.
4. Stem loop region is conserved.

22
5. Nanonucleotide (TAnTATTAC) is present in stem loop region (Horser et al.,
2001; Bell et al., 2002).
While there are also some differences:
1. BBTV satellites do not have internal ORF as present in DNA R.
2. Major common region is absent.
3. TATA box is present immediately upstream of stem loop structure in satellite
DNA molecules while in integral genome components it is present down
stream of stem loop region.
4. Rest of CR-SL is not conserved (Horser et al., 2001; Bell et al., 2002).
To determine whether BBTV S1 and S2 are present in other BBTV isolates or not,
samples were collected from both the South Pacific and Asian groups. These samples
were hybridized with probes generated BBTV S1, DNA 1 and DNA 3. With probes of
BBTV S1/S2, hybridization was only observed from isolates of Tonga (1/1). Samoa
(1/2), Philippines (2/3), Taiwan (1/1) and Vietnam (13/13). This probe did not
hybridize with isolates of Australia, Egypt, Fiji and India (Horser et al., 2001).
2.5 Replication of Circular ssDNA Viruses
Nanoviruses replicate their ssDNA by Rolling circle replication (RCR)
mechanism as in geminiviruses (Gronenborn, 2003; Hafner, 1998). Upon entry into
the host plant nucleus, virus uncoats and their ssDNA is converted into dsDNA by
host DNA polymerase. In nanoviruses, 80 nucleotide long DNA primer is found
associated with BBTV viral genomic DNA. This primer anneals to CR-M within
intergenic region of cssDNA to initiate the synthesis of complementary strand of
DNA. This dsDNA then expresses the specific viral proteins including Rep.
In geminiviruses, like Maize streak virus (MSV), Digiteria streak virus (DSV)
(Donson et al.,1987) and Chloris striate mosaic virus (CSMV) (Anderson et al.,
1988) these DNA primers have also been found, which are capable of forming
complementary viral DNA strands. In both geminiviruses and nanoviruses, this DNA
primer is of 80 nucleotide is present in the intergenic region of viral genome. In
BBTV, there are variable 5’ initiation sites of these DNA primers while in
geminiviruses there is a conserved 5’ initiation sites where ribonucleotides are
attached.

23
All components of BBTV were capable of self primed extension. The
component 5 shows the highest concentration of extension while component 1,2,4,6
show similar extension and there is no extension in component 3. The intergenic
region of BBTV is almost similar in all components of BBTV (Hafner et al., 1997).
Rep protein binds to the specific nanonucleotide sequence in the stem loop region.
Rep nicks the positive strand of the stem loop region to produce a 3’OH end that
initiates the synthesis of positive sense ssDNA to result in the formation of full length
viral ssDNA. Rep protein still binds to the 5’ end and religates the viral DNA for
further replication and transcription.
2.6 Proteins Encoded By ssDNA
2.6.1 Replication-associated protein (Rep)
This protein of about 41 kDa, is encoded by ORF C1 (also called AC1) in all
whitefly-transmitted geminiviruses and due to its similarity with rolling circle DNA
replication initiator proteins of some prokaryotic plasmids (Koonin and Ilyina, 1992),
it has been called Rep protein (Laufs et al., 1995). Rep is known to possess modular
functions (Campos-Olivas et al., 2002; Orozco et al., 1997). The N-terminal part of
Rep contains DNA-binding, nicking ligation and oligomerization domains, while C-
terminal half contains ATP-binding and ATPase activity domains (Raghavan et al.,
2004; Orozco et al., 1997; Fontes et al., 1992; 1994). During rolling circle replication
(RCR), Rep binds to repeat elements near the stem loop structure, makes a site-
specific nick at TAATATT↓AC in the loop region of the hairpin structure of the plus
strand to initiate replication and binds to the 5′ end of the nicked DNA via a tyrosine
residue. The 3′-OH end of the DNA is probably then used as a primer for synthesis of
the viral DNA (Heyraud-Nitschke et al., 1995). Rep also acts as a replicative helicase
by forming a large oligomeric complex and the helicase activity is dependent on the
oligomeric conformation (~24 mer) (Choudhury et al., 2006). Mastreviruses encode
two replication-associated proteins, RepA and Rep, which are translated from
messenger RNA splicing variants. The Rep protein of mastreviruses is the functional
homologue of Rep of begomoviruses (Heyraud-Nitschke et al., 1995).
The three dimensional solution NMR structure has been elucidated for the
catalytic domain of the Rep protein of the TYLCV (Campos-Olivas et al., 2002).
Structural analysis points towards a conserved architecture of the above domain in

24
prokaryotic and eukaryotic Rep proteins and in the number of functionally diverse
proteins. Rep is also known to induce host replication machinery presumably to
enable the virus to replicate in differentiated cells (Egelkrout et al., 2002; Kong et al.,
2000). The protein binds with retinoblastoma related proteins (pRBR) involved in the
cell cycle, which prevents cell entry into the S phase by sequestering transcription
factors (Collin et al., 1996). In nanoviruses, Clink protein binds to pRB and inhibits
the cell cycle. It means that cell death in nanoviruses and geminiviruses are initiated
by different mechanisms. The Rep protein of TGMV binds to pRB through an 80-
amino acid region that contains two predicted α-helices designated 3 and 4 (Arguello-
Astorga et al., 2004). It activates host transcription in mature leaves by relieving
pRBR/E2F repression. The pRBR binding activity of Rep and the ability of TGMV
infection to overcome E2F-mediated repression of the proliferating cell nuclear
antigen (PCNA; the processivity factor for DNA polymerase δ (Castillo et al., 2003)
promoter support a model whereby geminivirus replication proteins modulate host
gene expression through the pRBR/E2F pathway. According to this model, in mature
plant cells, E2F binds to the PCNA promoter and recruits pRBR, which in turn
recruits chromatin remodelling activities, such as histone deacetylases and SWI/SNF
like enzyme, to create a repressor complex (Zhang and Dean, 2001). In this case host
gene expression is activated, leading to the production of the requisite host DNA
replication machinery. Geminivirus replication proteins also interact with the
replication factor C (RFC) complex, the clamp loader that transfers PCNA to the
replication fork (Castillo et al., 2003; Luque et al., 2002). These interactions are likely
to represent early steps in the assembly of a DNA replication complex on the
geminivirus origin. ACMV Rep protein induces re-replication in fission yeast. Upon
expression of Rep, cells exhibited morphological changes. They were elongated
threefold on average and possessed a single, but enlarged and less compact nucleus in
comparison to non-induced cells. Rep expressing cells exhibited DNA contents
beyond 2C indicating ongoing replication without intervening mitosis (Kittelmann et
al., 2009). TGMV Rep also binds histone H3, a mitotic kinesin, a novel protein kinase
(GRIK) (Kong and Hanley-Bowdoin, 2002), and Ubc9, a component of the
sumoylation pathway (Castillo et al., 2004).
Master Rep (MRep) of BBTV not only initiates the replication of itself but
also replication of all other components (Horser et al., 2001). BBTV satellite
molecules also encode a Rep, but it initiates only replication of itself (Horser et al.,

25
2001). There are several functional domains in Rep of BBTV which include RCR
domain, an ATPase domain and an oligomerization domain. The RCR domain is
located on N- terminus and possesses three motifs and this site is responsible for the
interaction between viral DNA and the Rep protein (Jupin et al., 1994; Orozco et al.,
1997). These RCR domains are conserved between nanoviruses and geminiviruses.
MRep of all other nanoviruses like SCSV, FBNYV, MDV also contain similar
iterons. Therefore, MRep can bind to similar iterons in the integral components of all
three viruses to initiate replication (Timchenko et al., 2000). BBTV MRep is unlikely
to initiate the replication of other nanoviruses because the sequence of first 30 amino
acids of BBTV is different from other nanoviruses. However, the iterons of BBTV
and begomoviruses are similar (Horser, 2000) suggesting that BBTV MRep may
initiate the replication of begomoviruses and vice versa. Similarly, BBTV and
Mastrevirus satellite DNA have similar iterons and can initiate the replication of each
other (Horser, 2000). CFDV which is a tentative member of the genus Nanovirus also
contain similar motifs that are present in geminiviruses (Boevink et al., 1995).
An oligomereization domain is present between RCR and ATPase domain of
BBTV. Rep of geminiviruses has an oligomerization domain that interacts with plant
Rb-related proteins, viral coat proteins and may bind with other Rep proteins to form
multimers (Orozco et al., 2000; Arguello-Astorga et al., 2004; Shepherd et al., 2005;
Malik et al., 2005). But Rep of BBTV did not interact with Rb-related protein because
Rep of BBTV does not encode Rb-related protein, this protein is encoded by DNA C
of BBTV which contain LxCxE motif (Gronenborn, 2003). The oligomarization
domain in Rep of geminiviruses is also involved in non specific ssDNA binding and
appropriate assembly of Rep at the replication origin (Gomez-Llorente et al., 2005;
Hickman and Dyda, 2005). ATPase domain of BBTV also contains three motifs and
is located near C-terminus F (Koonin, 1993). Truncated Rep without ATPase did not
have ATPase activity but still have RCR and oligomerization activities (Hong and
Stanley, 1995; Brunetti et al., 2001).
2.6.2 Coat Protein
The coat protein (CP) of geminiviruses is a multifunctional protein necessary
for a range of functions associated with encapsidation, accumulation of viral ssDNA,
insect transmission and both intracellular and intercellular movement. The most
important function of CP is to produce the protein coat in which viral ssDNA is

26
encapsidated. A study based on MSV revealed that geminate particles are assembled
from 110 protein subunits, organized as 22 pentameric capsomers (Zhang et al.,
2001). For geminiviruses, CP also helps in the systemic spread of the virus. So,
mutation in this gene decreased the accumulation of viral DNA in the infected plants
but the level of viral DNA is not affected in protoplast, suggesting impairment of
movement function (Boulton et al., 1991). The localization of the product of this ORF
in secondary plasmodesmata with the start of viral lesions is consistent with its role in
the movement of monopartite geminiviruses (Dickinson et al., 1996).
CP of TYLCV is restricted to the nucleus and nucleolus and help in the
transport of viral DNA out of nucleus (Rojas et al., 2001). The CP of mungbean
yellow mosaic virus (MYMV) is thought to be involved in transport of the viral
genome from nucleus to cytoplasm. In vitro analysis showed the interaction of
MYMV CP with the nuclear import factor importin α suggesting that CP is imported
into the nucleus via an importin α- dependent pathway (Guerra-Peraza et al., 2005).
The localization of geiminivirus CPs has been described in TYLCV (Kunik et al.,
1998; Rojas et al., 2001), MSV (Liu et al., 1999) and ACMV (Unseld et al., 2001).
Interruption in the CP of ToLCV disrupted spread of the virus (Rigden et al., 1993).
CP of BCTV is essential for its infectivity (Briddon et al., 1989). MSV requires CP to
produce symptomatic systemic infection and mutants containing insertions or
deletions in CP gene were able to replicate to low levels, producing dsDNA although
virion ssDNA was not detected and symptoms were not observed (Boulton et al.,
1989). Insect vector specificity of geminiviruses is associated with the CP.
Replacement of CP of ACMV with that of BCTV changed to insect vector from
whitefly to leafhopper (Briddon et al., 1990).
2.6.3 Nuclear Shuttle Protein
Since geminiviruses replicate in the nucleus, they require movement protein
(MP) for movement from cell to cell and nuclea shuttle protein (NSP) for the
movement from cytoplasm into the nucleus. Both these proteins are encoded by
DNA-B of bipartite geminiviruses. These gene products have been implicated in local
and systemic spread, symptom development, host-range determination and virus
transmission (putting the virus where the insect feeds) by whiteflies. The requirement
of DNA-B products for above functions varies for different viruses and specific host-
virus combinations. Mutation in either of these genes eliminated virus infectivity but

27
did not affect virus replication or encapsidation (Brough et al., 1988; Etessami et al.,
1988; Pascal et al., 1994) provided direct evidence that NSP binds strongly to ssDNA
with high affinity and localizes to the cell nucleus. NSP is also shown to have
sequence independent affinity for dsDNA (Hehnle et al., 2004). NSP is a very basic
protein and contains two NLSs. A mutation in either of the potential NLSs severely
impaired or eliminated viral infectivity. The C-terminus of NSP is required for
interaction with MP (Sanderfoot et al., 1996). A number of host factors have been
identified that interact with NSP, including an Arabidopsis acetyltransferase and a
receptor-like protein kinase from tomato and soybean (McGarry et al., 2003; Mariano
et al., 2004). Kinase helps in the phosphorylation of NSP and regulates the movement
of viral DNA, or mask host resistance response against viruses (Mariano et al., 2004).
Thus, NSP has shown avirulence factor in common bean (Garrido-Ramirez et al.,
2000). Based on differential interaction with NSP and CP, it has been proposed that
acetylation may be involved in the segregation of ssDNA for movement and
encapsidation, respectively (McGarry et al., 2003). The synthesis of NSP is regulated
at the transcriptional level by TrAP transactivation (Sunter and Bisaro, 1992) and
unlike other begomoviruses NSP of ToLCNDV is a pathogenicity determinant
(Hussain et al., 2005).
In nanoviruses, DNA-N encodes nuclear shuttle protein of 17.4 kDa for the
movement of virus out of nucleus. This protein binds to the viral ssDNA and help in
the movement of virus out of nucleus (Wanitchakorn et al., 2000). Function of NSP is
equivalent to geminiviruses (Sudarshana et al., 1998).
2.6.4 Movement Protein
The movement protein (MP) of geminiviruses is involved in the movement of
virus from cell to cell. MP helps to increase the size of plasmodesmata and facilitates
the movement of dsDNA (Sudarshana et al., 1998; Noueiry et al., 1994). There are
two hypotheses i.e. relay race model and couple-skating model about the functions of
MP. According to “relay race model” NSP helps in the transfer of dsDNA freom
nucleus to cytoplasm and then transferred to MP for its movement through
plasmodesmata. According to couple skating model viral ssDNA binds to NSP
containing complex and transfer from nucleus to cytoplasm and interact with MP for
cell to cell movement. It means that both MP and NSP are required for cell to cell
movement of viral DNA. It has been demonstrated for BDMV, where mutations in

28
MP and NSP limited the movement of viral DNA from cell to cell (Sudarshana et al.,
1998). The movement protein of BDMV increases the size of plasmodesmata and
helps in the movement of DNA from cell to cell (Noueiry et al., 1994; Rojas et al.,
1998). Fission yeast model have revealed that MP redirects the NSP of ABMV to
plama memebrane (Frischmuth et al., 2007). It has been demonstrated that 117-160
amino acids of MP is necessary for movement of viral DNA to the periphery (Zhang
et al., 2002). Sequence analysis of this domain revealed a amphiphilic helix which
could provide attachment of the protein in one leaflet of the membrane, whereas no
transmembrane helices were recognizable (Zhang et al., 2002). The combined
properties of the geminivirus-encoded MP and plasmodesmata were discovered that
enforce a severe restraint on the size of the viral genome at the level of cell-to-cell
movement (Gilbertson et al., 2003).
The MP also plays an important role in the development of symtoms. It has
been demonstrated by the expression of MP of SqLCV in model plant N. benthamiana
(Pascal et al., 1993) and the expression of Tomato mottle virus (ToMoV) N. tabacum
(Duan et al., 1997), in which they produced phenotypes suggestive of the symptoms
of disease. In a related study, Pascal et al. (1993) expressed both MP and NSP of
SqLCV in transgenic plants. Their results showed that the expression of MP alone is
sufficient to cause mosaic and leaf curl symptoms, typical of SqLCV infection. These
results suggest a role of MP in symptom development in bipartite geminiviruses.
Movement protein of nanoviruses helps in the movement of virus from cytoplasm to
cell periphery. Thus, NSP and MP work together for the movement of virus in the
plant. This function of MP and NSP is similar to geminiviruses (Sanderfoot and
Lazarowitz, 1995).
2.6.5 Cell Cycle Link Protein
In geminiviruses, Rep proteins bind to retinoblastoma protein (pRB) and thus
trigger S- phase of cell cycle by the alleviation and sequestration of transcription
factors of E2F-family (Hanley-Bowdoin et al., 2000; Gutierrez, 2000). In
nanoviruses, a separate protein of 19kDa is produced called Clink. This protein binds
to retinoblastoma protein and thus helps in the activation of S-phase of cell cycle
(Aronson et al., 2000). Clink protein also binds to SKP1 member of the SCF complex.
This SCF complex is a part of the ubiquitin mediated protein pathway (Ciechanover et
al., 2000) but the function of that protein is unknown.

29
2.7 Genetic Approaches for Virus Resistance
Genetic engineering is another important tool for virus resistance, as today it is
possible to introduce any gene into plant through transformation. Therefore, we can
introduce any viral gene into plant and obtain the expression of that gene. The genes
can also be introduced into plant that interferes with virus replication cycle. Two
approaches are being widely used in plants to develop resistance against viruses.
Pathogen derived resistance
Non-pathogen derived resistance
2.7.1 Non Pathogen Derived Resistance
It involves the introduction of gene from non-pathogenic sources into plant.
The first symptom of R gene mediated resistance is the hypersensitive response (HR)
at the site of inoculation, so the virus can not spread to other parts of plant. They
recognize pathogens and defensive pathways are activated in plant. Numerous, R
genes have been identified, cloned and sequenced (Hammond-Kosack and Jones,
1997; Richter and Ronald, 2000). Flor (1971) proposed a “gene-for-gene” model that
states that, for resistance to occur the interaction between avr genes of pathogen and
R genes of plant is required. A loss of interaction between R gene and avr gene may
lead to the disease. R genes in the plant act as a receptor for pathogen avr genes,
which then prompts a signal transduction cascades. The local signaling cascade
triggers the response of host at the site of infection commonly known as
hypersensitive response (HR). This causes cell death at the site of infection and
inhibits the systemic spread of virus. The systemic cascade primes the defense system
in other parts of the plant. Thus, R genes have two functions: first to act as a receptor
for avr gene and second to produce signaling cascades.
R genes are now placed into five groups based on their structural similarities,
Leucine rich region (LRR) which is a pathogen recognition domain, a serine-
threonine kinase domain, a leucine zipper (LZ), Nucleotide binding site (NBS) and
immune response gene TIR from mammals (Parker et al., 2000; Baker et al., 1997).
The Rx1 gene present in potato gives extreme resistance to PVX. This gene is
transformed into Nicotiana benthamiana and N. tabacum (Bendahmane et al., 1999)
where it provides resistance against viruses. N. glutinosa has N gene related to R
gene, which provide resistance in plants against Tobacco mosaic virus (TMV). The N
gene has two domains, recognition site and signaling domain, LRR domain acts as a

30
recognition site and TIR domain acts as a signaling domain. This N- gene of
Nicotiana glutinosa has been successfully transformed to tomato to develop resistance
against Tobacco mosaic virus (Whitham, 1996). Current hypothesis is that N gene
interacts directly or indirectly with the TMV protein and lead to the cell death. N gene
identifies the replicase region as avr factor and causes cell death at the site of
infection. The N gene has been isolated by transposon tagging using maize activator
transposon (Whitham et al., 1994; Dinesh-Kumar et al., 1995). Rx gene in potato
provides extreme resistance to PVX, which is expressed in protoplast or in whole
plant (Kohm et al., 1993)
R genes are located in plant genome and can provide resistance in plants
against diverse group of pathogens. The most common example is Rx gene which
provides resistance in plants against PVX (Rouppe van der Voort et al., 1997). Thus
proper understanding of the molecular interactions between R genes products and
their elicitors will be helpful in providing resistance against a broad spectrum of
pathogens. So, this R gene mediated resistance could help in developing resistance for
disease control. As viruses causes a great loss to our economy. When R gene is
inserted into plant timely, this can stop the pathogen spread in plant with minimum
damage to plant. But there is a drawback of this R gene mediated resistance that it can
recognize limited number of pathogens, thus can not provide broad spectrum
resistance against viruses. These R genes are also destructed by the pathogens. There
is no need from the farmer to control the disease and it has no adverse environmental
effects (Pink, 2002). R genes and RNA silencing are antiviral defence mechanisms
against viruses. In plants these two pathways are linked with eachother to stop the
viral infections.
2.7.2 Pathogen Derived Resistance
Resistance against a particular virus or a strain of virus is conferred by the
transformation of plant with portion of viral genome. The first demonstration of this
resistance was given by Powell-Abel et al. (1986), they expressed Tobacco mosaic
virus coat protein (CP) in tobacco plants. These transgenic plants were protected
against TMV infection. After this demonstration an extensive research was started on
protecting crop species against viruses and on the mechanisms involved. Gonsalves et
al. (1998) expressed coat protein of Papaya ringspot virus (PRV) in papaya plants.
These viral capsid proteins provided resistance to Papaya plants against PRV. These

31
transgenic papayas i.e. Rainbow and SunUp were now successfully used in Hawaii
against PRV. Presently PDR is thought to be operated by two basic molecular
mechanisms protein-based resistance and nucleic acid-based resistance.
2.7.3 Protein Based Resistance
Resistance achieved by the expression of a viral gene product in transgenic
plant is termed as protein-based resistance. Protein-based protection attained by the
expression of coat protein of TMV was the first demonstration involving the PDR
(Powell-Abel et al., 1986). Transgenic plants either escaped infection following
inoculation or developed systemic disease symptoms significantly later than plants not
expressing the gene. Coat protein of TMV also provided resistance against other
tabamoviruses but it is not effective against viruses which were not closely related to
TMV. The mechanism of resistance is by the inhibition of spread of virions from
infected cells to other parts of plant (Bendahmane and Gronenborn, 1997). The
expression of this protein blocks the normal uncoating and replication of invading
viruses. The evidence support this was provided by the use of TMV pseudoviruses, in
which the GUS reporter gene attached to the viral origin of assembly, was packaged
into intact viral particles. When these particles were applied to protoplast, the level of
GUS activity was significantly reduced from that when control plants were
inoculated. It showed that pseudovirion was unable to disassemble in the transgenic
plants (Dickinson et al., 1996).
Transgenic tobacco expressing the PVX coat protein gene was significantly
protected against PVX infection. These plants showed delayed symptom development
numbers of lesions were also reduced. Inoculated and systemic leaves both showed
reduction in virus accumulation (Hemenway et al., 1988). The resistance is affected
by various environmental conditions and inoculums concentration. Coat protein-based
resistance is the most widely used for conferring protection in plants. Numerous crop
plants transformed to express viral CP have been reported and released for
commercial cultivation (Dasgupta et al., 2003). It includes tomato plants resistant to
TMV, Tomato mosaic virus (ToMV) and Cucumber mosaic virus (CMV); squash
resistant to Zucchinia yellow mosaic virus (ZYMV) and cucumber resistant to CMV.
This type of resistance is not only true for RNA viruses rather can also use equally in
developing protection against DNA viruses. This could be used for bipartite
begomoviruses that can compensate for CP in systemic infection.

32
Expression of other viral gene products also gives protection to a greater or
lesser extent against the target viruses. Replicase (Rep) protein could be used for
replicase-mediated resistance against viruses. Rep-mediated resistance was first
demonstrated against TMV, where the 54 kilo Dalton (kDa) putative gene was
expressed in transgenic tobacco under 35S promoter. Blocking the movement proteins
(MPs) could be more effective means of preventing spread of both related and
unrelated viruses, to result in a broad-spectrum resistance (Heinlein, 2002).
Transgenic tobacco plants expressing the Brome mosaic virus (BMV) MP were found
resistant against the unrelated TMV, and plants transgenic for a defective mutant of
the TMV MP were resistant to a wide range of unrelated viruses (von Arnim and
Stanley, 1992).
2.8 RNA Interference (RNAi)
It is the degradation of cellular RNA through a complex of enzymes and also
called as post-transcriptional gene silencing in plants. RNAi plays an important role in
the establishment of defense mechanism in plants against viruses by transcriptional
gene silencing (TGS) and post-trancriptional gene silencing (PTGS) (Akhtar et al.,
2010). The major role in RNAi is played by small interfering RNA (siRNA), which
acts as an intermediate in the silencing pathway (Voinnet, 2001). dsRNA is cleaved
into siRNAs of 21-26nt (Hamilton and Baulcombe, 1999) by enzymes called Dicers
(Bernstein et al., 2001). These siRNA serve as a template for degradation of mRNA
in sequence specific manner by being integrated into RNA induced silencing complex
(Hammond et al., 2000). So, this mechanism can be used to develop the resistance in
plants against viruses (Tenllado et al., 2004). Viral gene in sense and antisense
orientation within one transcript can show resistance in plant upon infection of that
particular virus through RNAi. A number of vectors like pFGC5941 have been
developed for the expression of dsRNA in plants for RNAi (Wesley et al., 2001; Miki
and Shimamoto, 2004). First studies on RNAi in plants were done in tobacco to
compare the ability of sense, antisense and or dsRNA that generate resistance via
PTGS (Waterhouse et al., 1998).
2.8.1 History of RNAi
For many years, it was believed that mRNA is an unremitting nucleotide
sequence in DNA. In 1977, Phillip and Richard Roberts showed that the mRNA

33
sequence is scattered intermittently in the genome. Later on, the discovery by Sidney
and Thomas in 1989 showed that RNA act as a catalyst in enzyme reaction gave a
new role of RNA. In the early 1980s it was revealed that in E. coli small sequence of
about 100bp can bind to the complementary sequence of mRNA and stop translation
(Mizuno et al., 1984; Nordstrom and Wagner, 1994). It was first demonstrated in
1993 that translation by antisense RNA can also be regulated in eukaryotes when
genes governing the development of the worm Caenorhabditis elegans was studied
(Lee et al., 1993; Wightman et al., 1993). This mechanism of posttranscriptional
regulation was regarded as an oddity for many years. However, the situation changed
drastically when small regulatory RNA called microRNAs was found (Lee and
Ambros, 2001).
Before the discovery of RNAi, gene silencing mechanism was first observed
in petunia plants in 1990, when Rich Jorgensen tried to engineer plants with the aim
to get deeper purple color of petals. Unexpectedly, the transgenic plants showed
variegated pigmentation due to the silencing of newly introduced genes. This
phenomenon was named as “co-suppression of gene expression”. This mechanism
was also recognized in fungus Neurospora crassa (Romano and Macino, 1992). But
the molecular mechanism at the back of gene silencing remained unidentified until the
discovery of RNAi which gave the amazing details about this phenomenon.
2.8.2 Discovery of RNAi
First of all, Guo and Kempheus (1995) showed that the introduction of sense
or antisense RNA for a particular gene was effective in sequence specific gene
silencing in Caenorhabditis elegans. Later it was shown by Fire et al., (1998) that
dsRNA was tenfolds more effective as compared to sense or antisense RNAs alone.
Fire and Mello was awarded Nobel Prize in Physiology or Medicine in 2006 for their
work published in 1998 for the discovery of RNAi. Within one year after its
discovery, it was recognized in hydra, fungi, Drosophila, zebra fish (Tuschl, 1999)
and this phenomenon was proven very rapidly among eukaryotes. After RNAi
discovery, it was shown that post transcriptional gene silencing in plants was
associated with the prescence of small RNA sequences in both sense and antisense
orientation (Hamilton and Baulcombe, 1999). In mammalian cultured cells, it was not
possible to get a strong and particular RNAi response initially. However, when short

34
21 nucleotide long dsRNA was introduced into the cell, silencing was achieved in
these cells (Elbashir et al., 2001).
2.8.3 Mechanism of RNAi
It can be divided into two steps (Bass, 2000). In the first step dsRNA was
cleaved into small interfering RNAs (siRNA) (Elbashir et al., 2001) by enzymes
called Dicer (Hamilton and Baulcombe, 1999; Bernstein et al., 2001; Zamore et al.,
2000). There are two dicer proteins in Drosophila and Neurospora, four in
Arabidopsis and one in mammals and C. elegans. Dicer has four domains:
1. An N-terminal helicase
2. Dual RNase III motifs
3. C-terminal dsRNA binding domain
4. PAZ domain (Piwi/Argonaute/Zwille)
All these domains have specific functions (Agarwal et al., 2003). These siRNA target
mRNA for its degradtaion.
In the second step, the siRNA are integrated into RNA-induced silencing
complex (RISC), which destroy mRNAs that are homologous to siRNA (Bernstein et
al., 2001; Hammond et al., 2000). The target mRNA is cleaved in the region which is
complementary to siRNA and result a decrease in protein expression (Elbashir et al.,
2001). RISC complex contains both protein and RNA. The protein component
contains PAZ domain and also has ribonuclease activity with the ability to cut RNA.
In certain plants, fungi and worms an RNA dependent RNA polymerase (RdRP) plays
a key role in producing siRNA (Cogoni and Macino, 2000). RdRP synthesizes dsRNA
from a siRNA template to initiate RNAi process.
2.9 Gene Expression System
PVX is a member of Potexvirus of RNA plant viruses. It is a rod shaped
filamentous virus. RNA of PVX is polyadenylated and capped (Huisman et al., 1988).
It infects plant species of Solanceae and 15 other plant species. This virus is
transmitted by mechanical means and sap transmissible. In potato, this virus causes
mild mosaics and in tomato it causes mottling or necrotic spotting. In other hosts, the
severity of symptoms increases when co- inoculated with other viruses. The virus can
be detected by ELISA test. There are five open reading frames (ORFs) in PVX, out of
which only ORF1 encodes the replicase which is concerned in synthesis of RNA.

35
PVX depend on host machinery for the synthesis of minus and genomic and
subgenomic plus strand RNAs but this protein is synthesized directly from viral RNA
upon infection. These subgenomic RNAs are then used for protein translation of triple
gene block (TGB), movement protein which is required for virus movement (Angell
et al., 1996; Lough et al., 2000) and coat protein (CP) which is necessary for viral
encpsidation and movement (Chapman et al., 1992a; Baulcombe et al., 1995).
PVX can be used as a model system gene function study of viruses. So, the
PVX based vector was designed for the gene expression study and was designed to
express the forign genes (Chapman et al., 1992b). The foreign genes are inserted in
the vector under the control of duplicated coat protein promoter. The foreign gene
expressed along with other proteins of PVX during infection. PVX has also been used
as a VIGS vector (Angell and Baulcombe, 1999). A number of virus groups, including
potexviruses, hordeiviruses, benyviruses and pecluviruses contain three overlapping
open reading frames, called as triple gene block (TGB) which encodes three proteins
involved in the movement of virus from one cell to other (Beck et al., 1991; Gilmer et
al., 1992; Petty and Jackson, 1990). These three proteins were reffered to as TGBp1,
p2 and p3 (Solovyev et al., 1996). RNA helicase increases the size of plasmodesmata
and help in the movement from cell to cell (Kalinina et al., 2002), while all proteins
are required for replication. Different experimental approaches have shown that a
defect in viral MP can be compensated by the viral protein of a second, unrelated
helper virus. MPs of different viruses are functionally interchangeable which involved
double infection with movement defective isolates and helper viruses (Atabekov and
Taliansky, 1990). In another experiment, it was shown that MP produced in
transgenic plants could complment a transport deficient virus (Deom et al., 1987;
Ziegler-Graff et al., 1991). In third experiment, wild type MP gene was replaced by
the MP gene from heterologous virus (De Jong and Ahlquist, 1992; Solovyev et al.,
1996). The inserted MP facilitates the movement of the hybrid virus from cell to cell
which further confirmed the compatibility of MPs of different viruses.
PVX vector has been used to express foreign genes in many host plants to
study their functions (Toth et al., 2001). This vector has diverse applications such as
gene silencing, virus movement, protein expression and RNA replication. In addition
to this, Tobacco mosaic virus (TMV) (Selth et al., 2004) has also proven very useful
tools for this purpose. The PVX vector (pGR107) itself gives very mild symptoms in
Solanaceous plants. However, when a gene is inserted in the vector under the control

36
of the duplicated coat protein promoter, it is expressed along with the other PVX
proteins during infection and may induce a phenotype above and beyond what the
virus vector produces. For example, PVX-mediated expression of the CP gene of
Turnip crinkle virus has revealed the suppression of RNA silencing. Similarly the
chimeric PVX containing the p22 gene of Tomato bushy stunt virus (TBSV) trans-
complement a TBSV cell-to-cell movement mutant and the vector containing p19 of
TBSV induced systemic necrosis in N. benthamiana (Scholthof, 2004).
Apart from the study of gene expression and their functions, gene silencing
techniques can also be used since the disruption of gene expression often shows a
more critical phenotype. The geminiviruses which have been used as silencing vectors
has been discussed earlier. The PVX vector is also used as VIGS vector (Faivre-
Rampant et al., 2004; Lacomme and Chapman, 2008) in which plant endogenous
genes are silenced using viruses. So, the PVX vector was used to study gene functions
of BBTV.
2.10 Suppressors Encoded By Plant Viruses
Viruses have a number of suppressors and some even encode various
suppressors that hinder at different steps of the silencing pathway. The first known
suppressor was the HC-Pro of Tobacco etch virus (TEV). This suppressor has the
ability to block local silencing and to reverse the already established silencing in
plants (Anandalakshmi et al., 1998; Kasschau and Carrington, 1998; Llave et al.,
2000). HC-Pro interacts with theThe mechanism of silencing suppressor is that this
protein interacts with the cellular calmodulin-related protein (rgsCaM) that is itself a
silencing suppressor, suggesting that HC-Pro stimulates an endogenous mechanism
that negatively regulates RNA silencing (Anandalakshmi et al.,2000). HC-Pro has
also been shown to interfere with dsRNA processing by Dicer and inhibit the
unwinding of duplex siRNA, thereby preventing the incorporation of targeting
information into RISC complex (Dunoyer et al., 2004; Chapman et al., 2004). In
contrast, the p19 protein of Cymbidium ringspot virus (CymRSV) cannot suppress
established silencing, although it can suppress local silencing and block production of
the systemic silencing signal. It has the ability to bind and sequester siRNAs,
preventing their incorporation into RISC complex (Lakatos et al., 2004; Silhavy et al.,
2002; Vargason et al., 2003). The ability of suppressors such as p19 and HC-Pro to
interfere with both siRNA and miRNA metabolism shows the overlapping nature of

37
these pathways (Chapman et al., 2004; Dunoyer et al., 2004). Coat protein of Turnip
crinkle virus (TCV-CP) does not interfere the miRNA pathway. The suppressor
activity of this protein is attributed to its ability to interfere with the activity of DCL-2
that does not play major role in processing miRNA precursors. The 2b protein of
Cucumber mosaic virus (CMV) cannot inhibit the initiation of silencing but
effectively prevents its systemic spread to neighbouring tissues (Brigneti et al., 1998;
Guo and Ding, 2002). Thus RNA viruses have adopted many different counter-
defense strategies aimed at different aspects of RNA silencing. From this perspective,
the molecular basis for synergistic diseases that can result from mixed infections with
viruses carrying unrelated suppressors become clear (Pruss et al., 1997). Information
of viral suppressors can provide a powerful tool for the analysis of the mechanism of
RNA silencing and the relationship between the different silencing pathways (Bisaro,
2006).
2.11 Promoters of ssDNA
Gene expression in eukaryotes require a series of multiple events including
histone modifications, binding of activators and coactivators to enhancer and
promoters of eukaryotic gene and recruitment of the basal mechanism to the central
part of promoter for transcription initiation (Smale and Kadonaga, 2003). Chromatin
remodeling controls the availability of DNA sequences known as promoters which
have to be in contact with the transcription machinery in order to start transcription.
DNA is packaged into chromatin and therefore the chromatin structure plays a crucial
role in gene expression and regulation (Mellor, 2006). Once the gene is accessible, the
promoter is key factor in controlling the expression of the corresponding gene.
Promoter is the basic structure needed for gene expression. Promoter is a DNA
region which is present upstream to the transcription start site and directs transcription
initiation of a gene (Praz et al., 2002). Gene regulatory sequences placed in the DNA
region upstream of the transcription start site are sites for transcriptional activators
and repressors (Wu et al., 2001) and they are also called transcription factor (TF)
binding sites (Venter and Botha, 2004). Core promoter is a DNA region present in
contact with the basal transcription machinery and thus help in initiation of basal
transcription (Lee and Young, 2000). RNA polymerase II binds to transcription
initiation apparatus of gene which is involved in protein synthesis (Lee and Young,
2000). RNA polymerase II is involved in assembly of TFs such as TFIID and binds to

38
the TATA box of the core promoter, and in combination with other TFs the RNA
polymeraseII is then recruited to the core promoter to form a preinitiation complex
(PIC) (Venter and Botha, 2004). The presence of TFs on a promoter allows the
interaction of other TFs with other cis-acting elements to activate transcription (Berk,
1999; Struhl, 2001). Specific activation of the common bean β-phaseolin promoter
during embryogenesis of transgenic tobacco lines is driven by the combinatorial effort
of specific cis-acting elements needed for phas expression in certain tissues of the
embryo such as the hypocotyl which are present in the 295nt proximal region
proximal to the TSS (Li and Hall, 1999).
In a ‘typical’ eukaryotic core promoter TATA box is located about 30
nucleotide upstream TSS (Breathnach and Chambon, 1981), an initiator region
located at the TSS (Smale, 1997) and a downstream promoter element (DPE) is
located app 30 nucleotide downstream of the TSS (Kutach and Kadonaga, 2000). 30-
50% of known plant promoters contain a TATA box upstream of the TSS
(Shahmuradov et al., 2005). Although most of plant promoters contain a TATA box,
but some photosynthesis related genes and house keeping genes lack a TATA box
(Dynan, 1986; Nakamura et al., 2002). Promoters without TATA boxes are
denominated TATA-less promoters and their TSS might be controlled by the
sequence surrounding the TSS (Smale, 1997) or the DPE (Shahmuradov et al., 2005).
Recently, a pyrimidine patch (composed of C and T) was identified around the TSS of
plant promoters (Yamamoto et al., 2007a, 2007b, 2007c). The ‘YR Rule’, which
consist of a pyrimidine (C/T) and a purine (A/G) at positions -1 and +1 at the TSS,
respectively, is found in 77% (10,806) of the Arabidopsis promoters analyzed and is
also applicable to rice promoters (Yamamoto et al., 2007a). The ‘YR Rule’ can be
considered as a less stringent initiator region and therefore might be recognized by
TFIID (Yamamoto et al., 2007a). In addition, the ‘YR Rule’ is in agreement with the
consensus around the TSS found in 217 dicot promoters (Shahmuradov et al., 2003).
Moreover, DNA elements further downstream of the TSS and located in gene regions,
such as the 5’ untranslated region (UTR), can (co-)regulate a specific level or pattern
of expression. For instance, among the versions tested the maize Ubilquitin promoter
including 5’ UTR and first intron has the highest activity (Christensen et al., 1992).
The presence of the Adh1 (maize) intron between the promoter of the phosphate
transporter gene from barley and the reporter gene also increased the level of
expression (Schunmann et al., 2004).

39
Cauliflower mosaic virus (CaMV) was reported as a first plant virus which
contains DNA as a genetic material (Zaitlin and Palukaitis, 2000). CaMV promoter
gives high level of expression in most plant tissues. So it can be used as a promoter
for the foreign genes expression in transgenic studies. Although CaMV is a dicot
infecting virus but it can also used in monocots in transgenic studies (Terada and
Shimamoto, 1990). Caulimoviruses are icosahederal in shape and 50 nm in diameter.
They produced coat protein and replicase as a distinct protein. Commelina yellow
mottle virus (CoYMV) is a monocot virus and it is a type member of Badnavirus
(Lockhart, 1990). It infects only monocot species like Commelina diffusa.
Badnaviruses are bacilliform in shape with 30 to 130 nm in diameter. These viruses
are transmitted by different vectors.
Nanoviruses like geminiviruses are also ssDNA plant viruses. Promoter
activity of nanoviruses like SCSV, CFDV and MDV has been characterized. In
SCSV, promoter activity varies between seven components and is phloem specific
(Surin et al., 1996). In CFDV, like BBTV there is one component encoding a Rep. A
promoter associated with this component has weak phloem specific activity in
tobacco which indicates the tissue limited accumulation of the virus within its natural
monocot host (Rohde et al., 1995). In MDV, among 11 promoters, the promoter of
component 8 (MP) confers a strong and constitutive promoter activity than all other
promoters of components. This promoter is a characteristics promoter different from
CaMV 35S promoter in both monocot and dicot species.
In BBTV, each of six components encodes at least one gene (Beetham et al.,
1997). Three regions in the intergenic region of BBTV components i.e. major
common region (CR-M), stem loop common region (CR-SL) and consensus
nanonucleotide sequence are similar in all components. The major common region
comprises of 66-92 nucleotides region (Burns et al., 1995) and is the site for binding
of endogenous ssDNA primer which helps in the synthesis of complementary strand
of full length (Hafner et al., 1997). CR-SL comprises of 69nt region and is similar to
stem loop region of geminiviruses (Lazarowitz et al., 1992). The third region is a
potential TATA box, with a nanonucleotide sequence CTATATATATAA and is one
of a number of transcription elements identified within the intergenic region,
suggesting the potential promoter function of these BBTV sequences. In order to
better understand gene functions of BBTV, it is better to study the promoter
sequences in BBTV components.

40
2.12 Promoters Used in Transgenic Banana
Transformation of monocot plants with Agrobacterium tumefacians is very
difficult task. So, genetic transformation of banana, a monocot plant is also not an
easy task. However, research on the suitability of Agrobacterium mediated genetic
transformation for banana resulted in the successful generation of transgenic plants
(Arinaitwe, 2008; Ganapathi et al., 2001) and banana has been successfully
transformed with Agrobacterium in Uganda. Banana can also be transformed by other
methods like particle bombardment (Sagi et al., 1995a, 1995b, 2995c; Becker et al.,
2000) and electroporation of banana protoplasts (Sagi et al., 1992, 1995b, 1995c).
Various varieties of banana can be used for transformation. The first successful
transformation in banana was done in the cultivar Bluggoe (genomic group, AAB)
(Dugdale et al., 1998; Remy et al., 1998a; Sagi et al., 1992, 1994). Later on, other
cultivars like Three Hand Planty (AAB, Perez Hernandez et al., 1998, 2006a, 2006b;
Remy et al., 2005; Arinaitwe et al., 2004; Santos et al., 2007; Arinaitwe, 2008),
Grand naine (AAA, Remy et al., 1998a; Schenk et al., 1999; Becker et al., 2000;
Yang et al., 2003; Arinaitwe et al., 2004; Atkinson et al., 2004; Khanna et al., 2004;
Acereto-Escoffie et al., 2005; Arinaitwe, 2008) and Williams (AAA, Bosque-Perez et
al., 2000; Sagi et alc., 1995a). Multiple promoters have been used in transient and
stable transformation of bananas (Swennen et al., 2003). CaMV have been widely
used as a constitutive promoter in transgenic studies. CaMV is a promoter which is
derived from CaMV virus, which is RNA virus. Several other viral promoters have
been used in banana transformation like nopaline synthase promoter from
Agrobacterium tumefacians especially to take the expression of the selectable marker
genes. Few plant promoters have been used in banana genetic transformation so far.
Uptill now only actin gene and genes concerned with fruit formation have been used
in banana genetic transformation.
Genetic transformation in banana can also be achieved by particle
bombardment. Particle bombardment in banana ECS revealed the highest level of
expression with the maize Ubil promoter, as measured by histochemical and
flourometric GUS assays. Promoters derived from intergenic regions of banana
components revealed a very weak transient GUS expression as compared to the
activity of CaMV and Ubil promoters after particle bombardment (Dugdale et al.,
1998). Howver, GFP expression was observed in promoter BT6.1 in banana embryos

41
and roots especially in root meristems and in leaf phloem and stomata (Dugdale et al.,
1998). However, further analysis revealed that BBTV components 4 and 5 showed the
maximum expression (Dugdale et al., 2000).
However, BBTV promoters showed the GUS expression in vascular tissues in
transgenic banana and intron-mediated expression was observed when maize Ubil
intron was used as a promoter (Dugdale et al., 2000). Similarly, the promoters derived
from satellite associated molecules S1 also direct the vascular specific expression
(Hermann et al., 2001b). Intergenic regions of Taro bacilliform virus (TaBV) in
banana showed the strongest activity in vascular tissue (Yang et al., 2003). Promoter
activity has also been tested in greenhouse mature banana plants. uidA reporter gene
driven by Gelvin or Super promoter was observed in leaf segments of greenhouse
plants and in ripe banana fruits (Ganapathi et al., 2001). Banana streak badnavirus
(BSV) promoters were analyzed in banana by stable transformation (Schenk et al.,
2001). Transgenic banana plants containing BSV promoters isolated from the
cultivars ‘Mysore’ (My) and Williams (Cv) showed greater GUS activity in the corm
and vascular tissues (Schenk et al., 2001). My promoter was more active in transgenic
banana than the Ubil promoter of maize, while Cv promoter showed the comparable
level of expression with Ubil promoter. Although the activity of nopaline synthase
(nos) was not compared with other promoters, its activity is confirmed by the
production of transgenic banana plants.
Recombinant promoters have been synthesized in few reports to achieve high
level of gene expression. Emu promoter is a recombinant containing a maize alcohol
dehydrogenase (Adh1) promoter, anaerobic responsive element from Adh1 promoter
and octopine synthase (Last et al., 1991). Transient assay in banana showed the lower
Emu promoter activity than enhanced 35S promoter and Ubil promoter of maize (Sagi
et al., 1995a). Similarly, Emu also showed the lower level of GUS expression in
banana plants as compared to the improved 35S promoter and Ubil promoter of maize
(Remy et al., 1998b). Maize Ubil promoter exhibited similar or highest level of
promoter activity when compared with other promoters (Dugdale et al., 1998;
Hermann et al., 2001a, 2001b) apart from BSV My promoter (Schenk et al., 2001)
and TaBV promoter with 5’ deletion (Yang et al., 2003). In case of BBTV, BT5.1
and BT4.1 intergenic regions showed the higher level of expression than Ubil
promoter of maize, when the reporter gene was gfp (Dugdale et al., 1998). In case of
GUS expression, the Ubil promoter of maize showed the higher level of expression as

42
compared to six BBTV intergenic regions in transient assay. TaBV promoters (T1200,
T600 and T500) were stably transformed in banana fused with uidA reporter gene.
Leaf tissues of banana plants showed similar activities for Ubil and T1200 and T500.
But T600 showed the four fold higher GUS activity than Ubil promoter (Yang et al.,
2003). Banana deleted actin 1 promoter exhibited two fold higher transient expression
in bombarded banana ECS than CaMV 35S promoter but maize Ubil promoter
showed four folds higher activity than banana actin 1 promoter. Similar results were
obtained in transgenic leaf extracts of banana (Hermann et al., 2001a).

43
Chapter 3 MATERIALS AND METHODS
3.1 DNA Manipulation Techniques
3.1.1 CTAB method
Total nucleic acids were extracted from the frozen leaves by CTAB method
(Doyle and Doyle, 1990). Leaves were placed in liquid nitrogen in autoclave pestle
and mortar and ground into a fine powder. The fine powder was taken into falcon tube
and 2% Cetyltrimethyl ammonium bromide (CTAB) buffer was added to the tube.
The tubes were placed at 65°C for half an hour. Then equal volume of chloroform:
isoamylalcohol (24:1) was added and mixed lightly. This mixture was centrifuged at
4000rpm for 15min. After centrifugation the top aqueous phase was taken into
separate tubes. This phase contains total nucleic acids while the remaining phases
containing proteins and debris were discarded. 2/3 volume of isopropanol was added
to the tube and centrifuged for 15min at 4000rpm to pellet down the total nucleic
acids. The pellet was washed with 70% ethanol and dried. The pellet was then
dissolved in sterile double distilled water (ddH2O) and DNA was quantified.
3.1.2 Polymerase Chain Reaction (PCR)
Polymerase chain reaction (PCR) was performed for DNA amplification. 50µl
of reaction mixture containing template DNA, 10X Taq polymerase buffer
(Fermentas), 2mM dNTPs, 1.5mM MgCl2, forward and reverse primers and Taq DNA
polymerase (Fermentas) was prepared in PCR tube. PCR mixture was denatured at
95°C for 5min followed by 35-40 cycles and final extension at 72°C for 10min. 5µl of
PCR product was then run on 1% agarose gel. Agarose gel is prepared in 0.5X TAE
buffer and 2µl of ethidium bromide (10mg/ml) was added. The sample was mixed
with loading dye prior to loading on the gel. The sizes of fragments were compared
with 1kb DNA ladder. For diagnostic PCR the volume was reduced to 25 µl per tube
by reducing the ingredients accordingly.
3.1.3 Rolling Circle Amplification (RCA)
Rolling circle amplification was used to amplify the circular DNA molecule
from nucleic acid extracts (Liu et al., 1996; Lizardi et al., 1998). Amplification
reaction (20µl) comprised of 100 to 200ng of template DNA, 50µl of random

44
hexamer primers, 1mM dNTPs, 2µl of 10X Φ29 DNA polymerase reaction buffer and
enzyme mixture comprised of Φ 29 polymerase and pyrophosphatase in 1X reaction
buffer and incubated for 3min at 94oC to denature ds DNA. The mixture was then
cool down to room temperature and mixed with 5-7units of Φ 29 DNA polymerase
and 0.02 units of pyrophosphatase and placed at 30°C for 18-20hrs. After this, the
reaction was heated in Thermal cycler for 10min at 65°C to discontinue the activity of
Φ 29 DNA polymerase enzymes. 2µl of RCA product was resolved on agarose gel as
mentioned above for PCR product.
3.1.4 Cloning of PCR Products
Amplified PCR product was ligated into InsTA clone PCR cloning kit
(Fermentas). Ligation mixture (20µl) comprised of 3µl of plasmid vector pTZ57R/T,
18 to 540ng purified PCR product, 5X ligation buffer, 5units T4 DNA ligase and
sterile dH2O. This reaction was placed in a ligation bath at 16°C overnight.
3.1.5 Cloning of RCA Product
RCA product was restricted to identify a unique restriction site in the viral
molecule. Then RCA product and cloning vector pTZ57R was restricted with the
same enzyme and ligation reaction was performed as mentioned above for PCR
product.
3.1.6 Restriction Analysis
Restriction was performed in a microfuge tube containing 20µl of reaction
mixture with a composition of 1X or 2X reaction buffer, 5U of reaction enzyme. The
reaction was incubated for 1hr at 37°C.
3.1.7 Phenol Chloroform
Phenol was added to the restricted product in equal volume and centrifuged for
10min. The supernatant was collected in another microfuge tube and 3M sodium
acetate and chilled 100% ethanol was added and placed at -20oC for two hour. The
microfuge tubes were centrifuged at 13,200rpm for 15min. The supernatant was
useless and pellet was washed with 70% ethanol. Then pellet was air dried and finally
dissolved in sterile dH2O.

45
3.1.8 Plasmid isolation (Miniprep)
A single colony from plate was inoculated into a test tube containing 2ml of
LB medium with appropriate antibiotic. These were incubated at 37°C with vigorous
shaking and left it overnight. After about 16hrs 1ml culture was poured into a
microfuge tube and centrifuged at 13,200rpm for 1min. Upper aqueous phase was
removed and 100µl of resuspension solution was added to pellet for resuspension by
vigorous mixing with vortex. Resuspended cells were lysed by adding 150µl of
alkaline lysis solution and mixed it gently by inverting the tubes four to five times.
Then 200µl of neutralization solution was added to neutralize the lysate and mixing
the contents by inverting the tubes four to five times. The tubes were centrifuged at
13,200rpm for 15min. Upper aqueous phase was collected in a fresh tube and 100%
chilled absolute ethanol was added to double the volume and centrifuged for 20min.
The supernatant was removed and pellet was washed with 100µl of 70% ethanol and
air dried. Finally the pellet was dissolved in sterilized water and stored in freezer.
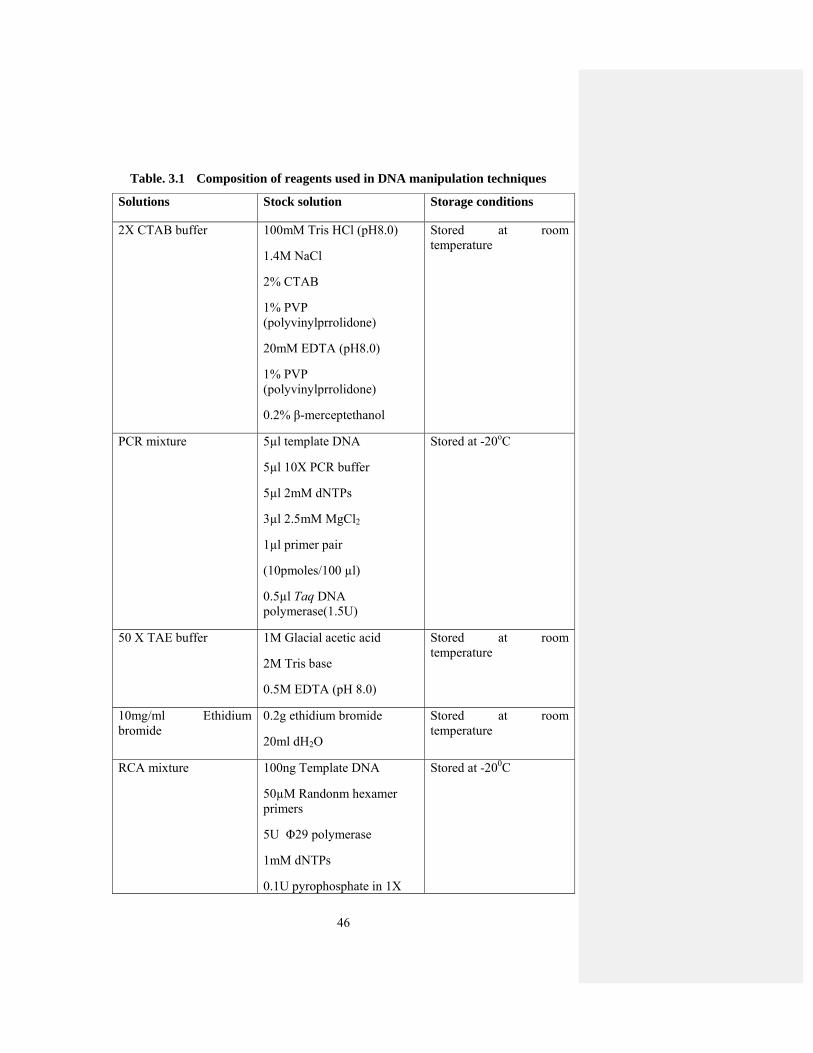
46
Table. 3.1 Composition of reagents used in DNA manipulation techniques
Solutions Stock solution Storage conditions
2X CTAB buffer 100mM Tris HCl (pH8.0)
1.4M NaCl
2% CTAB
1% PVP (polyvinylprrolidone)
20mM EDTA (pH8.0)
1% PVP (polyvinylprrolidone)
0.2% β-merceptethanol
Stored at room temperature
PCR mixture 5µl template DNA
5µl 10X PCR buffer
5µl 2mM dNTPs
3µl 2.5mM MgCl2
1µl primer pair
(10pmoles/100 µl)
0.5µl Taq DNA polymerase(1.5U)
Stored at -20oC
50 X TAE buffer 1M Glacial acetic acid
2M Tris base
0.5M EDTA (pH 8.0)
Stored at room temperature
10mg/ml Ethidium bromide
0.2g ethidium bromide
20ml dH2O
Stored at room temperature
RCA mixture 100ng Template DNA
50µM Randonm hexamer primers
5U Φ29 polymerase
1mM dNTPs
0.1U pyrophosphate in 1X
Stored at -200C

47
reaction buffer
Ligation mixture 0.165µg plasmid vector
3µg purified PCR product
2µl ligation buffer
5U T4 DNA ligase
Stores at -20oC
LB (Luria-Bertini) medium
1.0% Tryptone
0.5% Yeast extract
0.5% NaCl
1.5% Agar pH 6.2 - 7.2 and autoclave
Stored at room temperature
Resuspension Solution (Sol I)
25mM Tris HCl
10mM EDTA pH 8
50mM Glucose
Stored at room temperature
Denaturation Solution (Sol II)
0.5M NaOH
1M NaCl
Stored at room temperature
Neutralization Solution (Sol III)
11.5ml Glacial acetic acid
5M Potassium acetate
Stored at room temperature

48
3.2 DNA Analysis Techniques
3.2.1 Quantification of DNA
DNA concentration in total nucleic acid extract was determined by
spectrophotometer (SmartSpec Plus, BIORAD). A 20 times dilution was made for
each sample and DNA concentration was measured at 260nm with a conversion factor
of l = 50µg/ml for absorbance (A).
3.2.2 Agarose Gel Electrophoresis
1g of agarose was added in 100ml of TAE buffer (0.5X) in a 200ml flask to
prepare 1% agarose gel. The contents were boiled in a microwave oven for 2min to
get a transparent and clear solution. After cooling the contents, 2µl of ethidium
bromide (10mg/ml) was added to the solution and poured into a gel tray and left to
cool down. DNA samples were loaded on the gel with 6X loading dye. The gel was
run at 100V until the DNA was separated into different bands. Different sizes of DNA
were compared with 1kb DNA ladder. The gel was visualized and photographed
under UV light.
3.2.3 Southern Hybridization
10µg of DNA was resolved on 1.5% agarose gel. The agarose gel was put in a
glass dish and agitating continuously in depurination solution for 20min. After
removing this solution gel was rinsed with dH2O and agitated in denaturation solution
for 20min and treated with neutralization solution for 40min. After rinsing the gel
with distilled water, DNA was blotted to Hybond membrane. For blotting, gel was
placed on a wedge of filter paper pre-soaked in 4X SSC. The edges of filter paper and
piece of membrane were also dipped in 4X SSC. Some filter papers were placed over
the membrane and kept under weight overnight. Next day the membrane was UV
cross linked and air dried.
3.2.3.1 Preparation of Radio-Labeled Probe, Pre-Hybridization, Hybridization and Signal Detection
The membrane was transferred to a hybridization bottle and incubated with
pre- hybridization solution in a hybridization oven at 65oC. The radioactively labeled
probe was prepared by HexaLabelTM DNA labeling Kit (Fermentas) and the probe
was denatured before use. Depending on the choice of labeled dNTP (dATP or dCTP)
3µl mix dNTPs were added. After it 6µl of labeled dNTP was added and 5units of

49
Klenow fragment was added. The contents in microfuge were spin down for 5sec and
incubated for 30min at 37oC. The denatured probe was mixed in the pre-hybridization
solution and blot was incubated again at 65oC overnight for hybridization. Next day
three successive washing were given to blot with different concentrations of SSC i.e.
2X, 1X and 0.5X SSC, while keeping the SDS concentration constant (0.1%) at 65oC.
Then the blot was dried on a filter paper and wrapped in cling film. Fuji RX film was
exposed to radio-labeled blot overnight at -70oC.
3.2.3.2 Biotin-Labeled Probe Preparation, Pre-Hybridiztion, Hybridization and Detection of Probe
The membrane was transferred to a hybridization bottle and incubated with
pre-hybridization solution at the rate of 0.2ml/cm2 of membrane. Incubation was done
at 42oC for 2hrs with shaking in a Hybrid (Midi dual-14) hybridization oven. Biotin-
labeled probe was prepared by Biotin DecaLabelTM DNA Ladeling Kit (Fermentas).
The probe was denatured and 5µl of Biotin-Labeling Mix (provided by the
manufacturers) and 1µl of Klenow fragment were added in the tube and incubated at
37oC for 2hrs. Then 0.5mM EDTA was added to stop the reaction. Probe was
denatured by incubating at 100oC for 5min and chilled on ice. Denatured probe was
added to hybridization solution and this hybridization solution was added to the
membrane. Membrane was incubated overnight in hybridization solution at 42oC with
continuous shaking. Next day the membrane was washed twice with 2X SSC and
0.1% SDS. Then two more washings were performed with 0.1X SSC and 0.1% SDS.
Each washing was given for 20min at 65oC. After completing all washings excess
liquid was removed by briefly placing the membrane on filter paper. This labeled
DNA was then detected by following the detection procedure described by the
manufacturer.

50
Table. 3.2 Composition of reagents used in DNA analysis techniques
Solutions
Stock Solutions Storage Conditions
Depurination Solution 0.25M HCl
Stored at room temperature
Denaturation Solution 1.5M NaCl
0.5M NaOH
Stored at room temperature
Neutralization Solution 1M Tris HCl
1.5M NaCl
pH 7.5
Stored at room temperature
20X SSC 0.3MSodium citrate
3M NaCl
pH 7.5
Stored at room temperature
100X Denhardt Solution 2% w/v PVP
2% w/v ficoll
2% BSA
Stored at -20oC
Pre-hybridization Solution
(Radio-labelled)
8X SSC
0.08% (w/v) PVP
0.08% ficoll
0.08% BSA
1% SDS
4µg/ml denaturedcalf thymus DNA
Stored at room temperature
Radioactive Probe 10ng Template DNA
10µl hexanucleotide
5X reaction buffer
3µl dNTPs
5U klenow fragment
nuclease free water
Stored at -20oC

51
Biotin labeled probe 200ng template DNA
10µl decanucleotides
5µl Biotin-labeling mix
1µl Klenow fragment
0.5mM EDTA
Stored at -20oC
3.3 Microbiology Techniques
3.3.1 Preparation of Competent Cells of E. Coli
E. coli cells were streaked on Luria-Bertani (LB) plates. A single colony from
freshly streaked plate was inoculated to 50ml autoclaved LB medium in 250ml flask.
This was incubated at 37oC overnight with vigorous shaking. Next day, 2ml of this
overnight culture was put in 250ml LB in 1 litre flask and incubated at 37oC with
vigorous shaking until an optical density of 0.5-1.0 (1010cells/ml) was achieved. After
getting desired OD value, culture was cooled on ice for 30min. The culture was
transferred to sterile 50ml propylene tubes aseptically. The cells were pelleted down
by centrifugation at 4oC at 4000rpm for 5min and re-suspended into 5ml of 0.1M
MgCl2. The cells were again centrifuged under same conditions for 5min and re-
suspended in 0.1M CaCl2 and kept on ice for 30min. The cells were again centrifuged
and finally suspended in 0.1M CaCl2 (1-1.5ml) and 100% sterile glycerol. Finally the
cells were stored at -70oC
3.3.2 Transformation of E. coli
5µl ligation mixture was mixed in a vial of E. coli cells (100µl) and incubated
on ice for 30min. The cells were then heat shocked at 42oC in a dry bath for 2min.
The cells were immediately transferred on ice and 1ml LB medium was added. The
cells were then placed at 37oC for half an hour and then centrifuged at 13,200rpm for
1min and resuspended in 200µl of LB media. The cells were then spread on LB plates
containing required antibiotics. Plates were placed at 37oC overnight.
3.3.3 Preparation of Electro-Competent Cells of Agrobacterium
A single colony of Agrobacterium tumefaciens was inoculated into 20ml of
LB media containing appropriate antibiotic from a freshly streaked plate by a sterilized
wire-loop and incubated at 28oC for two days with vigorous shaking. After two days

52
5ml of the culture was inoculated into 300ml LB media and placed at 28oC until
OD600 of the cells is 0.5-1.0 (1010cells/ml). The culture was then transferred to 50ml
ice cold propylene tubes aseptically and left on ice for half an hour. These tubes were
centrifuged at 4000rpm for 10min in a tabletop refrigerated centrifuge at 4oC. The
supernatant was discarded and the cells were re-suspended in 50ml of sterilized cold
water. The cells were again centrifuged at 4000rpm at 4oC for 10min and again re-
suspended in 25ml of sterilized cold water and this washing was repeated thrice.
Finally the cells were dissolved in 5ml of sterilized 10% glycerol and 50μl aliquots of
electro-competent cells were made and placed at -70oC.
3.3.4 Transformation of Clones in Agrobacterium tumefaciens by
Electroporation
1μl of recombinant plasmid was put in one aliquot (50μl) of electro-competent
cells. These cells were then transferred to chilled electroporation cuvette (1mm gap).
The electroporation cuvette was placed in the electroporator and electric shock was
given at 1.44kV. After shock 1ml of liquid LB medium was added immediately and
then transferred to a microfuge tube and placed at 28C for 2hr with vigorous shaking.
The microfuge tubes were centrifuged for 1min and pellet was dissolved in 200μl of
LB media. This LB media was then spread on LB plates containing appropriate
antibiotic. The plates were placed at 28oC for two days. Colonies of Agrobacterium
were used to inoculate 2ml LB medium containing rifampicin (100µg/ml) and
kanamycin (50µg/ml) and incubated at 28oC for 48hrs with vigorous shaking.
Recombinant plasmid was confirmed by PCR.
3.3.5 Agrobacterium-Mediated Infiltration of Host Plants
Nicotiana tabacum and Nicotiana benthamiana were grown under controlled
conditions at 25°C with a photoperiod of 16hrs. The infectivity constructs were
electroporated into Agrobacterium strain GV3101. The bacterial cells were pelleted
down at 4000rpm for 15min and re-suspended in 10mM MgCl2. 150μg/ml
acetosyringone was added to the bacterial suspension for the activation of bacteria and
left for 3hrs and then infiltrated into the fully expanded leaves of N. tabacum and N.
benthamiana plants. Infection on inoculated plants was confirmed by visual
observation of symptom development and by biotechnological techniques i.e PCR by
using specific primers and Southern hybridization.

53
Table. 3.3 Primers used during this study
Primer Gene Sequence
NCMRF MRep AAGCTTACAAGTAATGACTTTACAGC
NCMRR MRep ATCGATGTTCTGTCGTCGATGATGATCT
NCCPF CP AAGCTTTATGTTTATGTAAACATAAA
NCCPR CP ATCGATCTAACTCGACACTGGTATTT
NCNSPF NSP GATATCAAGTTGTGCTGTAATGTT
NCNSPR NSP AAGCTTCGCTTCTGCTTTGCTTTC
NCClinkF Clink AAGCTTGAGTAATATATAGGAATGTT
NCClinkR Clink ATCGATTTCTCTTCCGACGAGTGATT
NCdelMRF MRep AAGCTTCGACAAAAGTACACTATGAC
NCdelMRR MRep ATCGATGTTCTGTCGTCGATGATGATCT
NCdelNSPF NSP GATATCAACATAAACATAAACGAAGT
NCdelNSPR NSP AAGCTTCGCTTCTGCTTTGCTTTC
MR107F MRep CGAATCGATGGCGCGATATGTGGTATG
MR107R MRep ATTGTCGACTCAGCAAGAAACCAATTTATTC
CP107F CP GAGATCGATGTTCAAACAACAAATGGC
CP107R CP CATAGTCGACTCAAACATGATATGTAATTC
MP107F MP CCCATCGATATGGCATTAACAACAGAGCG
MP107R MP TACGTCGACTTAAAACATAGGTCCGACG
NSP107F NSP GCACCCGGGATGGATTGGGCGGAATC
NSP107R NSP GCAGTCGACTTATTCCTTGATTCTTAACG

54
Clink107F Clink GGAATCGATATGGAGTTCTGGGAATCG
Clink107R Clink TATGTCGACTACATCTTCTTCCTCTGTC
MR35SF MRep GACAAGCTTATGGCGCGATATGTGGTATG
MR35SR MRep ATTGTCGACTCAGCAAGAAACCAACTTTATTC
CP35SF CP TCGAAGCTTATGTTCAAACAACAAATGGC
CP35SR CP CATAGTCGACTCAAACATGATATGTAATTC
MP35SF MP CCCAAGCTTATGGCATTAACAACAGACG
MP35SR MP TACGTCGACTTAAAACATAGGTCCGACG
NSP35SF NSP GCACCCGGGATGGATTGGGCGGAATC
NSP35SR NSP GCAGAATTCTTATTCCTTGATTCTTAACG
Clink35SF Clink GGAAAGCTTATGGAGTTCTGGGAATC
Clink35SR Clink TATGTCGACTACATCTTCTTCCTCTGTC
GFPSenseF GFP CTCGAGATGAGTAAAGGAGAAGAACT
GFPSenseR GFP CCATGGCCATAGGTGAAGGTAGTGAC
GFPantiF GFP GGATCCCCATAGGTGAAGGTAGTGAC
GFPantiR GFP TCTAGAATGAGTAAAGGAGAAGAAT

55
Table. 3.4 Details of constructs produced during this study
Clone name
Insert detail Sites Primers Details of cloning
MR-PVX 850bp of MRep gene of BBTV
ClaI/SalI MR107F
MR107R
The gene was amplified by PCR. PCR product was cloned into pTZ57R/T. From TA clone the gene was transferred to pGR107
CP-PVX 414bp of CP gene of BBTV
ClaI/SalI CP107F
CP107R
As above
NSP-PVX 475bp of NSP gene of BBTV
ClaI/SalI NSP107F
NSP107R
As above
Clink-PVX 386bp of Clink gene of BBTV
ClaI/SalI Clink107F
Clink107R
As above
MP-PVX 526bp of MP gene of BBTV
ClaI/SalI MP107F
MP107R
As above
MRNC 250bp non-coding region of MRep of BBTV
ClaI/HindIIII NCMRF
NCMRR
The gene was amplified by PCR. PCR product was cloned into pTZ57R/T. From TA clone the gene was cloned into expression vector. The expression cassette is made by restriction of pJit166 with SalI and XhoI and ligated into pGreen restricted with same enzymes.
NSPNC 625bp fragment of NSP of BBTV
HindIII/EcoRV NCNSPF
NCNSPR
As above

56
ClinkNC 714bp fragment of Clink of BBTV
ClaI/HindIII NCClinkF
NCClinkR
As above
CPNC 686bp fragment of CP of BBTV
ClaI/HindIII NCCPF
NCCPR
As above
MRdelNC 230bp fragment of MRep deleted of BBTV
ClaI/HindIII NCdelMRF
NCdelMRR
As above
NSPdelNC 432bp fragment of NSP of BBTV
HindIII/EcoRV NCdelNSPF
NCdelNSPR
As above
Table. 3.5 Vectors used during this study
Vector Size Type
Antibiotic selection
pTZ57R/T
2886bp
Cloning vector
Ampicillin
pGreen0029
4632bp
Transformation vector
Kanamycin
pJIT166
3727bp
Expression vector
Ampicillin
pGR107
10359bp
Expression vector
Kanamycin
pFGC5941
11405bp
Expression vector
Kanamycin

57
Chapter 4 RESULTS
4.1 RFLP Analysis of Banana Infected Samples
4.1.1 Phylogenetic Relationship of BBTV components from Pakistan Cloned
from RCA Product
BBTV infected samples were collected from Tando Jam Sindh. These samples
were amplified by Rolling circle amplification (RCA) method. Out of seven samples,
only three samples gave amplification (Fig. 4.1A). These phi products were then
restricted with hexacutters like HindIII and BglII. Out of these three samples, only
one sample gave band of 1.1kb with restriction enzyme HindIII, while other samples
remain unrestricted. So, this sample was then again restricted randomly with
hexacutters like EcoRI, BamHI, XhoI, NotI. But this sample remains unrestricted with
all these restriction enzymes. The restricted product with HindIII was then cloned into
circular vector pTZ57R. On the basis of restriction analysis 3 clones of 1.1kb sizes
were selected for sequencing (Fig. 4.1B). The sequences obtained were compared
with already available sequences in the NCBI database using BLAST. Comparisons
of sequence data showed similarity with sequences of South Pacific group of BBTV.
One sequence showed 98% homology to DNA-N component of South pacific group.
These results suggest that there was very little sequence heterogeneity among
Pakistani isolates of BBTV. It was further confirmed by phylogenetic analysis which
showed that they belong to the South Pacific group of BBTV. They showed the
closest relationship with isolates from India and Australia (Fig. 4.2). South pacific
group and Asian group showed greatest similarity, they showed only 5% sequence
diversity at nucleotide level between groups and 3% sequence diversity within groups.
Two clones isolated from Pakistani isolate were also sequenced. These clones
also showed 99% homology to South Pacific group of BBTV. They showed 91%
nucleotide sequence identity of already cloned component of DNA-M. They also
showed closest similarity to Australian and Indian isolates. Phylogenetic analysis also
showed that DNA-M of Pakistani isolate is closely related to South Pacific group
(Fig. 4.3).

58
Fig 4.1. (A) Rolling Circle Amplification of BBTV samples. Lane 4, 5, 6 represents amplification by RCA.
(B) Cloning of RCA product into pTZ57R. Lane 1, 3, 5 represents fragment of 1.1kb released after restriction with appropriate enzymes.

59

60
Fig 4.2. Phylogenetic dendrogram based upon a nucleotide sequence alignment of reported DNA-N of BBTV, showing their relationship to the newly characterized component of BBTV DNA-N from Pakistan. For each component the database accession number and its geographic origin using country name is indicated.

61

62
Fig 4.3. Phylogenetic dendrogram based upon a nucleotide sequence alignment of reported sequence of BBTV DNA-M, showing their relationship to the newly characterized component of BBTV DNA-M from Pakistan. For each component the database accession number and its geographic origin using country name is indicated.

63
4.1.2 RFLP Analysis of Banana Infected Samples
Five to six components of BBTV have been characterized from both Asian
group and South pacific group. South pacific group includes isolates of Egypt, India,
Fiji, Australia, Tonga and Pakistan, while Asian group includes isolates from Taiwan,
Vietnam, and Philippines. DNA-R has also been investigated from both groups.
While, additional Rep encoding components have been isolated and characterized
only in Vietnam isolates, belongs to Asian group of BBTV isolates. These
components were designated as DNA Y, W1 and W2 of approximately 1kb in size.
So, in order to find out these additional Rep encoding components from South pacific
group, we performed RFLP analysis of infected banana samples. For this purpose, the
infected samples were amplified by RCA method. This RCA product was then
restricted with tetracutters like HpaII, MseI, MboI, HaeIII and their restriction pattern
was checked. These enzymes gave the bands of different sizes (Fig. 4.4). These
results were further confirmed by southern hybridization with probes of already
cloned BBTV components of DNA-S, DNA-N, DNA-M present in our lab. For this
purpose, phi product was digested with hexacutters like KpnI, ClaI, EcoRV, BglII,
they gave the bands of 1.1kb and also with tetracutters. The restricted phi product was
run on the gel and transferred to nylon membrane and these membranes were
hybridized with probes of DNA-S, DNA-N and DNA-M. All bands were hybridized
with probe. It means that there is not new component in this sample. Rep-encoding
satellite DNAs have been isolated from Asian isolate. On the basis of sequence
analysis the present BBTV isolate in Pakistan belong to South pacific group. The
restriction patterns and Southern hybridization reflected that a single homogenous
population of virus is prevalent in Pakistan and no satellite molecules are associated
with this isolate. So, there is very low level of genetic diversity in South pacific group
than Asian group.

64
Fig 4.4. RFLP analyzed restriction fragments of Phi product of BBTV. Phi product restricted with ClaI (1), KpnI (2s), EcoRV (3), BglII (4), HpaII (5),
MseI (6), MboI (7) and HaeIII (8)
Formatted: Indent: Left: 0", Hanging: 0.5",Space After: 0 pt, Tab stops: Not at 0.5"

65
Fig. 4.5. (A) Southern hybridization of blots probed with DNA-S. Phi product was digested with enzymes ClaI (1), KpnI (2), EcoRV (3), BglII (4), HpaII (5), MseI (6), MboI (7) and HaeIII (8)

66
Fig. 4.6 (A) Southern blot analysis with DNA-N probe. Phi product was digested with enzymes ClaI (1), KpnI (2), EcoRV (3), BglII (4), HpaII (5), MseI (6), MboI (7) and HaeIII (8)
(B) Southern hybridization of blots probed with DNA-M. Phi product was digested with enzymes ClaI (1), KpnI (2), EcoRV (3), BglII (4), HpaII (5), MseI (6), MboI (7) and HaeIII (8)

67
Component Enzyme Expected size Size on gel Southern
hybridization
DNA-S EcoRV 1075bp 1075bp 1075bp
HpaII 167,908bp 167,908bp 167,908bp
MseI 60,371,644bp 60,371,644bp 60,371,644bp
MboI 520,555bp 520,555bp 520,555bp
HaeIII 176,801bp 167,801bp 167,801bp
Fig 4.7. Genome organization of BBTV DNA-S. Show the position of tetracutters in DNA-S. Possible sizes of bands with tetarcutters are shown in the table below.

68
Component Enzyme Expected size Size on gel Southern
hybridization
DNA-M KpnI 1042bp 1042bp 1042bp
HpaII 349,693bp 349,693bp 349,693bp
MseI 64,118,162,239,459bp 64,118,162,239bp 64,118,162,239bp
MboI 294,748bp 294,748bp 294,748bp
HaeIII 460,582bp 460,582bp 460,582bp
Fig 4.8. Genome organization of BBTV DNA-M. Show the position of tetracutters in DNA-M. Possible sizes of bands with tetarcutters are shown in the table below.

69
Component Enzyme Expected size Size on gel Southern hybridization
DNA-N ClaI 1096bp 1096bp 1096bp
HpaII 25,1071bp 1071bp 1071bp
MseI 111,114,226,317,328bp 111,114,226,317,328bp
111,114,226,317,328bp
MboI 79,202,245,570bp 202,245,570bp 202,245,570bp
HaeIII 137,164,795bp 795bp 795bp
Fig 4.9. Genome organization of BBTV DNA-N. Show the position of tetracutters in DNA-N. Possible sizes of bands with tetarcutters are shown in the table below.

70
4.2 Transient Expression of BBTV proteins Proteins from PVX
Vector
All BBTV genes were cloned into PVX vector generating constructs MRep-
PVX, MP-PVX, NSP-PVX, Clink-PVX and CP-PVX. These constructs along with
PVX vector alone was expressed in model plants N. benthamiana and N. tabacum for
symptom expression. PVX vector without insert was agro-inoculated as a control. The
presence of PVX in inoculated and control plants were confirmed through ELISA.
Plants inoculated with vector alone showed typical symptoms of PVX, consisting of a
very mild yellowing of the veins initially followed by faint mosaics. As PVX showed
very mild symptoms in these model plants which make it a good model system for the
study of foreign gene expression (Lacomme and Chapman, 2008). Symptoms
produced by constructs are discussed in detail.
4.2.1 MRep-PVX
N. benthamiana and N. tabacum was used as a host for this study. MRep-PVX
was agro-inoculated to 10 healthy plants of N. benthamiana by syringe method. As a
control to this experiment PVX was also agro-inoculated to same number of healthy
plants. 5 healthy control plants were also kept under same conditions along with
inoculated plants. Healthy control plants remained symptomless (Fig 4.10. a). Plants
inoculated with PVX alone showed mild vein yellowing initially and a faint mosaic
appeared 10dpi but these plants recovered from symptoms 20dpi (Fig 4.10. b). Plants
inoculated with MRep-PVX showed cell death at the site of inoculation within 7dpi
(Fig 4.10. d). The newly emerging leaves showed mild symptoms of necrosis. Any
leaf or shoot that emerged after one month were normal and did not show any
symptoms. The plants were kept for flower formation and seed set. They produced
flowers and seeds normally. All results were also confirmed through ELISA. This
experiment was repeated in three batches.
MRep-PVX construct was also agro-inoculated to 10 healthy plants of N.
tabacum to check differences in symptom development. PVX was agro-inoculated to
same number of healthy plants. 5 healthy control plants were also kept under same
conditions along with inoculated plants and these plants remained symptomless.
Symptom of PVX in this host was much milder as compared to N. tabacum that was
difficult to photograph. Plants inoculated with MRep-PVX did not show any

71
symptoms as these plants were left for one month. All experiments were repeated
thrice.
4.2.2 MP-PVX
MP-PVX was agro-inoculated to 10 N. benthamiana and N. tabacum plants.
PVX vector alone was also inoculated to the same number of plants. All these plants
were kept under same conditions along with 8 healthy control plants. N. benthamiana
plants showed severe necrosis at 7dpi (Fig. 4.11. f&g). When young plants at 3-4 leaf
stage were inoculated with this construct, the plants died after few days of
inoculation. When slightly older plants were inoculated, the plants were not killed by
the infection but ceased to grow. The newly emerging leaves showed severe leaf
necrosis and leaf curling adjacent to the veins. Symptoms showed by MP-PVX were
severe as compared to MRep-PVX. The newly developing leaves frequently was dark
colored. Plants inoculated with PVX vector alone showed mild symptoms of vein
yellowing initially and later mosaics were appeared. Healthy control plants remained
symptomless. Theses results were also confirmed through ELISA. The experiment
was repeated in three batches.
10 N. tabacum plants were also agro-inoculated with same construct but they
did not show any symptoms. These plants were left for one month but they did not
show any symptoms. PVX vector alone was used as a control. In N. tabacum PVX
showed very mild symptoms which can not be seen visually.
4.2.3 NSP-PVX
10 plants of N. benthamiana were agro-inoculated with NSP-PVX. This
construct showed relatively less symptoms as compared to MRep-PVX and MP-PVX.
These plants showed symptoms of necrotic lesions at the site of inoculation at 14dpi.
The newly emerging leaves showed faint mosaics that were slightly more severe than
that induced by PVX vector (Fig 4.11. h&i). They also showed slight upward curling
of leaf margins. PVX showed very slight symptoms of necrosis. These results were
confirmed by ELISA.
Similarly in N. tabacum they did not show any obvious symptoms. These
plants were left for one month but they did not show any symptoms. PVX vector
alone also showed very little symptoms of PVX which can not be photographed.

72
4.2.4 Clink-PVX
Expression of Clink gene from PVX vector in N. benthamiana and N. tabacum
showed mild symptoms of foliar mosaics. 10 plants of N. benthamiana were agro-
inoculated with this construct by syringe method. PVX was also agro-inoculated to
the same number of plants and same numbers of healthy plants were also placed under
same conditions. N. benthamiana plants showed very mild symptoms of foliar
mosaics and leaf deformation at 10dpi (Fig 4.12. j&k). There was no necrosis in
inoculated and systemic leaves. PVX showed very mild symptoms of leaf mosaics.
When N. tabacum plants were agro-inoculated with this construct. There were
no visible symptoms on this plant like other constructs. These plants were left for one
month but there are no visible symptoms on this plant. These plants only show the
typical symptoms of PVX.
4.2.5 CP-PVX
N. benthamiana and N. tabacum was used as a host to study the functions of
CP gene. CP-PVX was agro-inoculated to 10 healthy plants of N. benthamiana by
syringe method. As a control to this experiment PVX was also agro-inoculated to
same number of healthy plants. 5 healthy control plants were also kept under same
conditions along with inoculated plants. Healthy control plants remained
symptomless. Plants inoculated with PVX alone showed mild vein yellowing initially
and a faint mosaic appeared 10dpi (Fig 4.12. l) but these plants recovered from
symptoms 20dpi. Plants inoculated with CP-PVX showed very mild symptoms as in
Clink. They showed typical symptoms of PVX but these symptoms are slightly severe
than that produced by PVX in systemic leaves. There is also some leaf deformation at
1odpi. The newly emerging leaves did not show any symptoms. The plants were kept
for flower formation and seed set. They produced flowers and seeds normally. All
results were also confirmed through ELISA. This experiment was repeated thrice to
check reproducibility of experiment.
CP-PVX construct was also agro-inoculated to 10 healthy plants of N.
tabacum to check differences in symptom development. PVX was agro-inoculated to
same number of healthy plants. 5 healthy control plants were also kept under same
conditions along with inoculated plants and these plants remained symptomless.
Symptom of PVX in this host was much milder as compared to N. tabacum that was
difficult to photograph. Plants inoculated with CP-PVX did not show any symptoms

73
as these plants were left for one month. All experiments were repeated thrice in order
to check the reproducibility of experiment.
Fig 4.10. Phenotypic effects of the expression of BBTV genes from a PVX vector in N. benthamiana (a) shows the healthy control plant of N. benthamiana (b) N. benthamina plant infected with PVX vector (c) PVX vector expressing GFP viewed under UV illumination (d) shows the plant infected with MRep-PVX

74
Fig 4.11. Phenotypic effects of the expression of BBTV genes from a PVX vector in N. benthamiana (f,g) shows the plant infected with MP-PVX, (h,i) plants infected with NSP-PVX

75
Fig 4.12. Phenotypic effects of the expression of BBTV genes from a PVX vector in N. benthamiana (j,k) shows the plant infected with Clink-PVX, (l) plants infected with CP-PVX
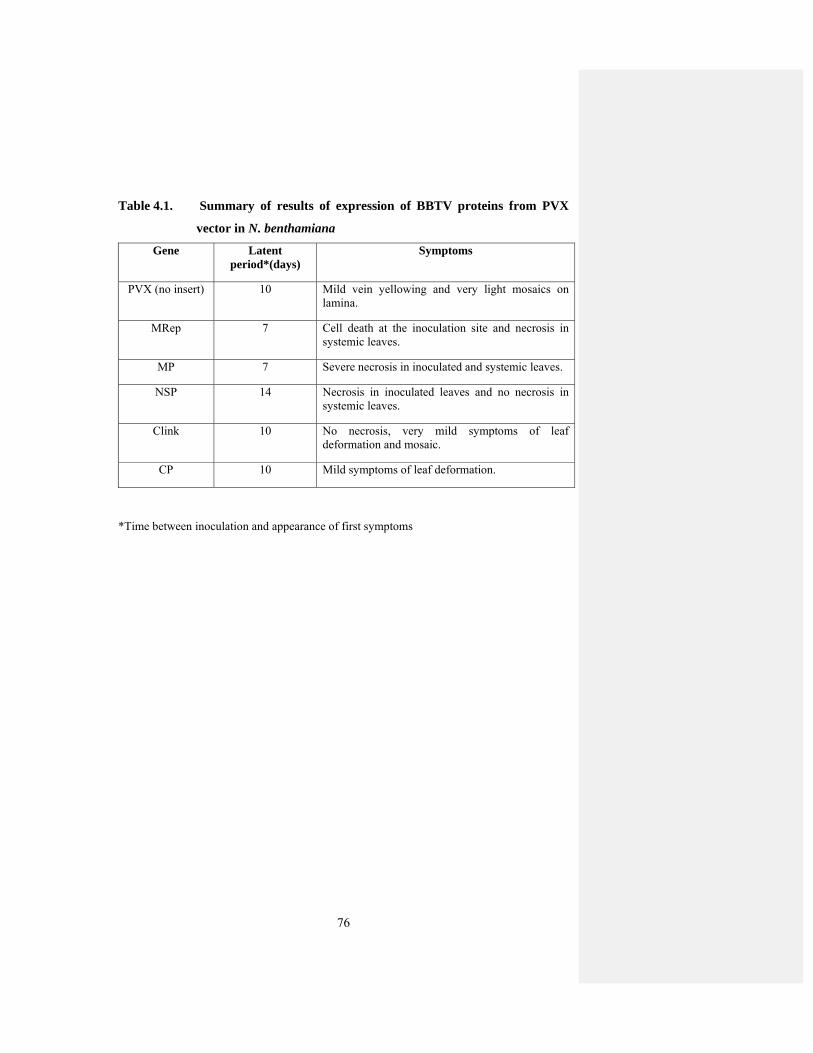
76
Table 4.1. Summary of results of expression of BBTV proteins from PVX
vector in N. benthamiana
Gene Latent period*(days)
Symptoms
PVX (no insert) 10 Mild vein yellowing and very light mosaics on lamina.
MRep 7 Cell death at the inoculation site and necrosis in systemic leaves.
MP 7 Severe necrosis in inoculated and systemic leaves.
NSP 14 Necrosis in inoculated leaves and no necrosis in systemic leaves.
Clink 10 No necrosis, very mild symptoms of leaf deformation and mosaic.
CP 10 Mild symptoms of leaf deformation.
*Time between inoculation and appearance of first symptoms

77
Table 4.2. Study of plants inoculated with BBTV proteins.
Inoculum Plant sp Infectivity* Symptoms
MRep-PVX N.B 6/10 7/10 7/10 Showed symptoms
N.T 0/10 0/10 0/10 No symptoms
MP-PVX N.B 7/10 8/10 7/10 Showed symptoms
N.T 0/10 0/10 0/10 No symptoms
Clink-PVX N.B 7/10 6/10 5/10 Showed symptoms
N.T 0/10 0/10 0/10 No symptoms
NSP-PVX N.B 8/10 6/10 7/10 Showed symptoms
N.T 0/10 0/10 0/10 No symptoms
CP-PVX N.B 7/10 4/10 7/10 Showed symptoms
N.T 0/10 0/10 0/10 No symptoms
* Represents infected plants/total plants

78
4.3 RNA Silencing Suppressors by BBTV
RNA silencing is a general defense mechanism in plants against viruses. Once,
RNA silencing transgene has been established, all RNAs are degraded which show
the homology to viral DNA (Lindbo et al., 1993). For successful infection viruses
have developed mechanisms to suppress RNA silencing in the host (Voinnet et al.,
1999; Vance and Vaucheret, 2001). Some suppressors of plant DNA viruses have
been identified but in nanoviruses it has not been reported. In order to study whether
BBTV genes have the capacity to suppress RNAi, the five available genes were
transiently expressed under the control of CaMV 35S promoter together with RNAi-
GFP construct in N. benthamiana line 16c plants. These N. benthamiana 16c plants
were used that carry a constitutive GFP gene. These plants fluoresce green under UV
light due to constitutive expression of GFP transgene (Fig 4.13 b), while N.
benthamiana plants fluoresce red under UV light (Fig 4.13 a). In order to study
whether BBTV genes are silencing suppressors or not, five BBTV genes were
transiently expressed under the control of 35S promoter in N. benthamiana line 16c
plants. RNAi-GFP construct was designed to express an inverted repeat of GFP gene
sequence which leads to the silencing and these leaves appear red under UV light. Co-
inoculation of RNAi-GFP (silencing inducer) along with BBTV genes onto 16c plants
will thus help to identify protein which can block silencing. 16c plants were also co-
inoculated with RNAi-GFP and two well known suppressors, HC-Pro of TEV and CP
of TCV which maintained GFP fluoresce in inoculated leaves. The level of GFP
expression in HC-Pro of TEV (Fig 4.14 e) and CP of TCV was then used to compare
the intensity of suppression in other BBTV genes.
`Northern blot analysis of inoculated leaves was also performed to check the
level of GFP specific mRNA and siRNA at 3dpi. Neither control 16c nor N.
benthamiana leaves showed the accumulation of GFP-mRNA, while 16c plants
showed the accumulation of GFP specific mRNA (Fig 4.16). However, when 16c
plants inoculated with the RNAi-GFP construct, the leaves produced a large quantity
of siRNA and reduction in GFP-mRNA, which indicate the silencing of GFP
transgene. In contrast, co-inoculation of 16c plants with RNAi-GFP constructs and CP
of TCV accumulated only minute amount of GFP specific siRNA, indicative of
suppressor activity. However, when plants co-inoculated with GFP-RNAi and HC-Pro
of TEV resulted in accumulation of significant amount of GFP-specific siRNA.

79
When 16c plants were inoculated with silencing inducer, it led to the silencing
and loss of GFP fluoresce under UV light. When MRep under 35S was co-inoculated
into 16c plants with GFP-RNAi construct led to silencing of the transgene and loss of
GFP fluorescence under UV light (Fig 4.15 i). These plants were observed visually by
comparison with two well known suppressors. These plants fluoresce green under UV
light indicative of silencing. Thus this protein showed lack of suppressor activity.
Northern blot analysis also showed the production of large quantity of siRNA and
small amount of mRNA, which further confirmed the results. It showed the reduction
in GFP-mRNA but produced large quantities of siRNA as observed in case of GFP-
RNAi alone and vector as a control. Thus, this protein does not help in suppression of
silencing activity.
Similarly, 16c plants were co-inoculated with NSP along with GFP-RNAi.
After 3dpi these plants were observed under UV light (Fig 4.14 g). These plants
appear green under UV light. Thus this protein also did not help in suppression of
silencing activity and appear green under UV light. These results were further
confirmed by Northern blot analysis (Fig 4.16). These plants showed reduction in
GFP-mRNA but production of large quantities of siRNA as in case of GFP-RNAi
construct alone, which also indicate the lack of suppressor activity.
When 16c plants were co-inoculated with GFP-RNAi and BBTV-CP under
35S promoter. The same number of plants was also co-inoculated with GFP-RNAi
and HC-Pro of TEV and CP of TCV. These plants were left for three days. After 3
days, these plants were examined under UV light and led to silencing of the transgene
and GFP fluorescence in inoculated leaves of 16c N. benthamiana (Fig 4.14 h). These
results were further confirmed by Northern blot analysis. This indicated that NSP, CP
and MRep of BBTV do not have suppressor activity, at least under the conditions of
the assay used here.
However, inoculation of 16c plants with MP and Clink under 35S along with
silencing inducer maintained GFP fluorescence in inoculated tissues (Fig 4.15 j&k).
The level of GFP expression was estimated visually by the intensity of GFP
fluorescence which was lower as compared to HC-Pro of TEV and CP of TCV which
were used as control in these assays. Northern blot analysis of inoculated leaves at
72dpi showed the accumulation of GFP-mRNA but no accumulation of GFP specific
siRNA as in case of non transgenic N. benthamiana plants (Fig 4.16). However, for

80
BBTV Clink and MP co-expression in 16c plants with GFP-RNAi construct yields
only minute amount of GFP specific siRNA and maintains GFP mRNA levels. This
indicates that the clink and MP proteins of BBTV have suppressor of gene silencing
activity, confirming the results of GFP fluorescence assays. Thus, in case of BBTV
genes MRep, NSP and CP does not have suppression of gene silencing activity, while
MP and clink have suppressor of gene silencing activity.
Neither non-trangenic nor 16c N. benthamiana showed accumulation of GFP-
specific siRNA, although the line 16c plants contained GFP-specific mRNA.
However, when inoculated with the RNAi-GFP construct, line 16c plants showed a
reduction in GFP mRNA and the production of large quantities of GFP specific
siRNAs, indicative of silencing of the GFP transgene. In contrast, coinoculation of
16c plants with the RNAi-GFP construct and the two well characterized suppressors
did not lead to a significant reduction in GFP mRNA levels and for TCV-CP only
minute amounts of GFP-specific siRNA accumulated; indicative of suppressor
activity. However, for plants co-inoculated with HC-Pro significant amounts of GFP-
specific siRNA were detected (Fig 4.16). This apparently anomalous behavior of HC-
Pro is due to its unusual suppressor activity. HC-Pro has been shown to mediate
suppression by stabilizing ds siRNA, thereby preventing its incorporation into RISC
and preventing any further mRNA degradation. Therefore, unlike many other
suppressors, HC-Pro does not prevent the accumulation of siRNA.

81
Fig 4.13. Identification of BBTV gene products with PTGS suppressor activity (a) shows the leaves from non-transgenic N. benthamiana plant, (b) a line 16c plant, (c) a leaf of a line 16c plant inoculated with RNAi-GFP and (d) and a leaf of a line 16c plant co-inoculated with RNAi-GFP and pGreen0029.

82
Fig 4.14. Identification of BBTV gene products with PTGS suppressor activity. The leaves in panels e–h from plants co-inoculated with RNAi-GFP and constructs for the expression of HC-Pro (e), TCV-CP (f), BBTV-NSP (g), BBTV-CP (h)

83
Fig 4.15. Identification of BBTV gene products with PTGS suppressor activity. The leaves in panels i–k from plants co-inoculated with RNAi-GFP and constructs for the expression of BBTV-MRep (i), BBTV-MP (j), BBTV-Clink (k)
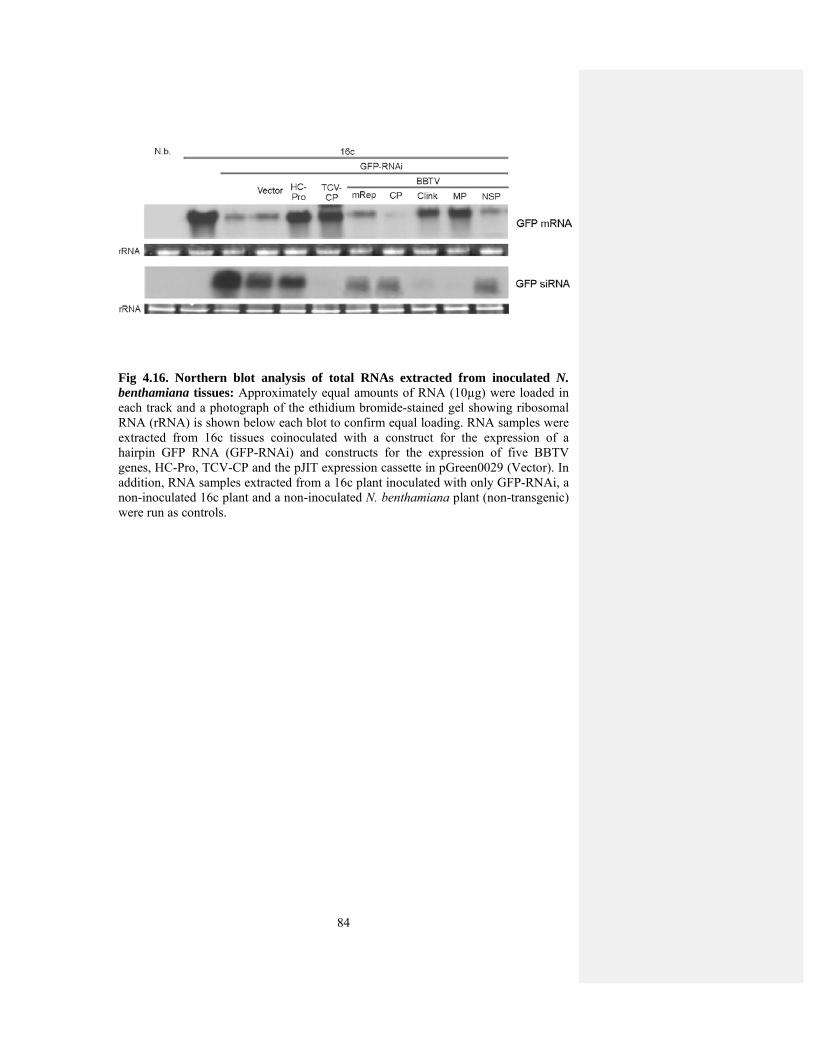
84
Fig 4.16. Northern blot analysis of total RNAs extracted from inoculated N. benthamiana tissues: Approximately equal amounts of RNA (10µg) were loaded in each track and a photograph of the ethidium bromide-stained gel showing ribosomal RNA (rRNA) is shown below each blot to confirm equal loading. RNA samples were extracted from 16c tissues coinoculated with a construct for the expression of a hairpin GFP RNA (GFP-RNAi) and constructs for the expression of five BBTV genes, HC-Pro, TCV-CP and the pJIT expression cassette in pGreen0029 (Vector). In addition, RNA samples extracted from a 16c plant inoculated with only GFP-RNAi, a non-inoculated 16c plant and a non-inoculated N. benthamiana plant (non-transgenic) were run as controls.

85
4.4 BBTV Promoter Study
4.4.1 Cloning of Intergenic Region of BBTV Genes
To determine the promoter activity of BBTV genes, intergenic regions of
BBTV components were cloned into expression vector by specific primers as
described in Table 3.3. The desired fragments were amplified by using the specific
primers of all components of BBTV (Fig 4.17 a). Then this product was ligated into
pTZ57R/T vector (Fig 4.17 b & Fig 4.18 a&b). This product was then further ligated
into expression vector (Fig 4.19 a&b, Fig 4.20a). For the construction of expression
vector, pJIT166 was restricted with SalI and XhoI and ligated into pGreen vector
which is also restricted with the same enzymes (Fig 4.21). So, BBTV promoter region
is ligated into this expression vector to check the specificity of promoter. This
specificity could be checked by the expression pattern of GUS gene. After cloning of
all constructs these constructs were transiently checked in model plant N.
benthamiana.
4.4.2. Transient Activity of Promoters in N. benthamiana
N. benthamiana was used as a model plant in this study. Five to six leaves of
N. benthamiana was agro-inoculated with MRep construct. As a control to this
experiment, CaMV under 35S was also agro-inoculated to the same number of
healthy plants. Healthy control plants of the same age were also kept under the same
conditions along with the inoculated plants. These plants were left for three days and
then stained for GUS expression by Jefferson (1987). Inoculated leaves were detached
from plants and placed in GUS substrate at 37oC and left it overnight. Next day leaves
were washed with 100% ethanol to destain the leaf and checked for GUS activity. The
leaves showed the blue color in the areas where gene is transformed. MRep did not
show any visual GUS expression (Fig 4.23 c&d).
When NSP was agro-inoculated into N. benthamiana leaves. It showed
moderate level of GUS activity than MRep. But this was also significantly lower than
GUS activity showed by CaMV, 35S promoter (Fig 4.22 c&d). There is no visible
GUS expression in leaves inoculated with CP and Clink. Thus, out of four genes, NSP
showed some level of GUS activity and CP, MRep and Clink did not show any GUS
activity. This experiment was repeated thrice to check the reproducibility of
experiment.

86
Fig. 4.17. (A) PCR amplification of MRep, Clink and NSP intergenic regions. Lane 1, 5, 7: negative control. Lane 2: 250bp fragment of MRep, Lane 6: 714bp fragment of Clink, Lane 8: 625bp fragment of NSP.
(B) Cloning of intergenic regions in pTZ57R/T. Lane 1-8 MRep fragment released from pTZ57R/T after restriction with EcoRI and HindIII, Lane 9-11 NSP fragment released from pTZ57R/T after restriction with EcoRI and HindIII.

87
Fig 4.18. Cloning of intergenic regions in pTZ57R/T. (A) Lane 1-5 NSP fragment released from pTZ57R/T after restriction with EcoRI and HindIII.
(B) Lane 1-6 represents Clink fragment released from pTZ57R/T after restriction with EcoRI and HindIII.

88
Fig 4.19. Cloning of intergenic region in vector designated as pGreen-GUS.
(A) Lane 3 & 4 250bp of MRep released from recombinant plasmid with ClaI and HindIII.
(B) Lane 1-4 625bp fragment of NSP released from recombinanat plasmid with EcoRV and HindIII restriction enzymes

89
Fig 4.20. (A) Cloning of Clink in vector pGreen-GUS. Lane 4 represents the 714bp fragment released from vector after restriction with HindIII and ClaI.
(B) lane 1 & 2 restriction of pGreen with SalI and XhoI. Lane 3 & 4 restriction of pJit166 with SalI and XhoI

90
Fig 4.21. Cloning of pJit166 into pGreen. Lane 3 represents confirmed clone after restriction with SalI and XhoI.

91
Fig 4.22. Leaves of N. benthamiana stained for GUS expression after transient analysis of BBTV promoter genes (A) leaves infiltrated with the GUS gene under the control of Cauliflower mosaic virus 35S promoter, (B) negative control, (C) & (D) leaves infiltrated with promoter region of NSP and GUS
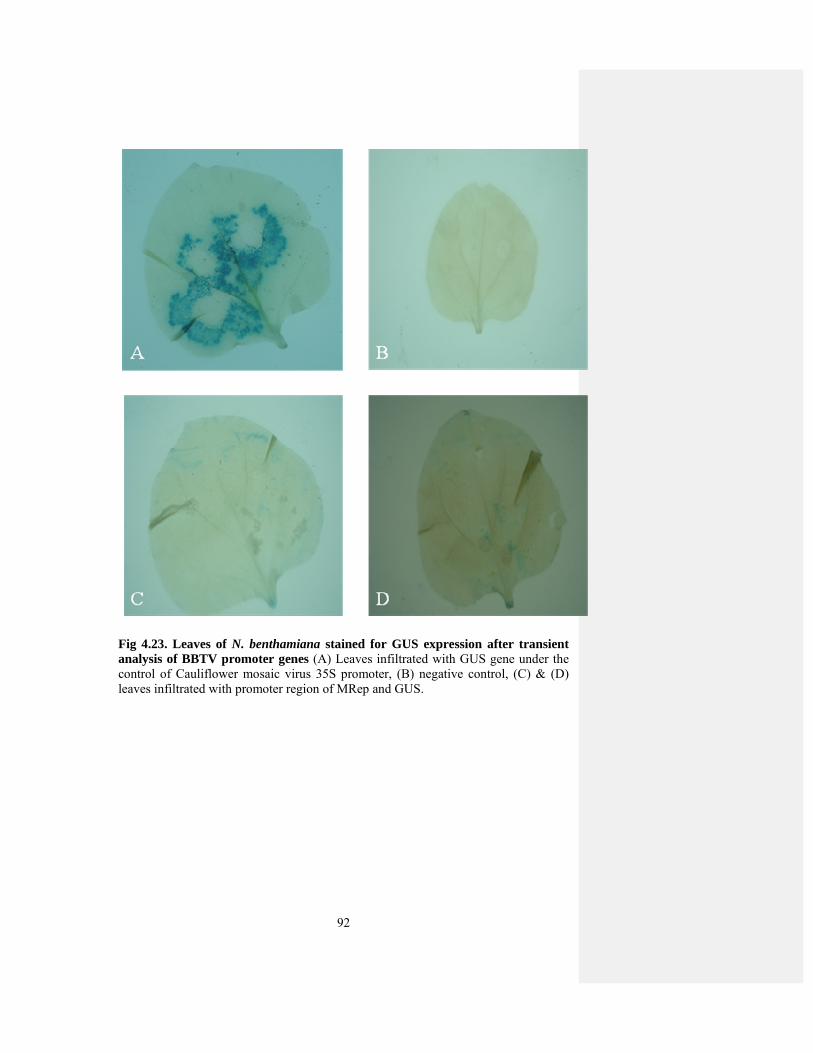
92
Fig 4.23. Leaves of N. benthamiana stained for GUS expression after transient analysis of BBTV promoter genes (A) Leaves infiltrated with GUS gene under the control of Cauliflower mosaic virus 35S promoter, (B) negative control, (C) & (D) leaves infiltrated with promoter region of MRep and GUS.

93
4.4.3 Cloning of Deleted Components of BBTV Genes
As in the previous experiment, GUS activity was observed in only one BBTV
gene i.e. NSP but MRep and Clink did not show any visible GUS expression. So,
deletions were made in MRep and NSP to check whether GUS activity is increased or
decreased. 5’ deletion of MRep and NSP was generated using specific primers (Table
3.3) which were generated by deleting 20bp fragment from MRep and 200bp from
NSP. The deleted intergenic regions of NSP and MRep were amplified using specific
primers (Fig 4.24 a&b). PCR products were cloned into pTZ57R/T vector (Fig 4.25
a&b). This product was further ligated into construction vector. This vector contained
a promoterless GUS gene downstream the MCS, so that the specificity of the
promoter cloned in MCS could be revealed by the expression pattern of GUS gene.
After cloning of constructs, these constructs were transiently checked in N.
benthamiana plants.
4.4.4 Transient Activity of Deleted Components in N. benthamiana
These promoters were also checked in model plant N. benthamiana. Five to
six leaves of three N. benthemiana plants were agro-inoculated with deleted
components of NSP and MRep. CaMV, 35S was also inoculated to the same number
of healthy plants as a control. Healthy control plants of same age were also placed
under same conditions. These plants were left for three days and then stained for GUS
activity. Inoculated leaves were detached from plants and placed in GUS substrate
and incubate at 37oC overnight. Next day leaves were washed with 100% ethanol to
destain the leaf. It was observed that promoter activity of both NSP and MRep
increased. But NSP promoter gave the higher level of GUS expression than MRep
(Fig 4.26 c&d). GUS activity was higher in deleted NSP as compared to full length
NSP promoter. Similarly, MRep also gave the higher level of GUS activity than full
length MRep promoter. But the expression is still lower than CaMV, 35S. CaMV, 35S
gave the higher level of GUS expression than MRep and NSP. There is no GUS
expression in leaves which were used as negative control. This experiment was
repeated in three batches to confirm the results.

94
Fig. 4.24. PCR amplification of deleted MRep intergenic regions. (A) Lane 1: negative control. Lane 2: 230bp fragment of MRep
(B) PCR amplification of deleted intergenic region of NSP. Lane 1: negative control, Lane 2: 432 bp fragment of NSP

95
Fig. 4.25. Cloning of deleted MRep intergenic region in pTZ57R/T. (A) Lane 1, 2, 4, 6, 7 represents MRep fragment released from vector after restriction wih EcoRI and HindIII.
(B) Cloning of deleted NSP in pTZ57R/T. Lane 1, 2, 3 represents NSP fragment released after restriction with EcoRI and HindIII.

96
Fig 4.26. GUS assay for deleted BBTV promoters. (A) Leaves of N. benthamiana plants infiltrated with the GUS gene under the control of Cauliflower mosaic virus 35S promoter, (B) negative control, (C) leaves stained for deleted MRep (D) leaves infiltrated with deleted NSP.

97
4.5 BBTV Promoter Study in Monocots
As promoter activity is checked N. benthamiana plant, which is dicot. But
banana is a monocot plant so promoter activity was then checked in monocot plant
wheat. Promoter activity was checked in wheat callus tissues. The induction of callus
in wheat genotypes can be controlled by the concentrtation of 2,4-D (Sears and
Deckard, 1982). Different concentrations of 2,4-D (0.5 to 8.0 mg/l) have been
reported for callus induction (Ozias-Akins and Vasil, 1982; Altpeter et al., 1996;
Dobrzanska et al., 1997). Most of these studies indicate that 2,4-D concentration of
1.0-2.0 mg/l are optimum for callus induction from scutellum tissue of immature
embryos of wheat. Therefore, we used 2mg/l of 2,4-D concentration for callus
induction. Callus was produced from scutellum of wheat after 3-4 days. During the
period of callus formation all tissues were continuously observed and any
unorganized or loose callus mass was removed to ensure the induction of true callus
from scutellum cells. Plumule and radicle was also removed to ensure true callus
induction.
Then, BBTV NSP promoter was transiently checked in wheat callus tissues
via micro-particle bombardment. The wheat callus was bombarded with NSP via
biolistic gun. Wheat calli which were bombarded at the distance of 9cm showed GUS
expression than calli which was bombarded at the distance of 6cm. As BBTV NSP
promoter showed the higher GUS activity than other components. So, this promoter
was checked in wheat to check its efficiency in monocots as banana is also a monocot.
Wheat callus tissues were bombarded with BBTV NSP promoter and CaMV, 35S
which is used as a positive control. The callus was also bombarded with the
microparticles containing gold particles without plasmid. Some calli was also placed
under same conditions without bombardment. All plates were placed in growth
chamber under controlled conditions of temperature and light. After 4 days, the calli
was checked for GUS expression. Calli was removed from plates and placed in GUS
substrate at 37oC overnight. Then GUS substrate was replaced by 100% ethanol. A
moderate level of GUS expression was observed in wheat calli bombarded with NSP.
The intensity of blue color in NSP promoter was low as compared to CaMV, 35S.
There was no GUS expression in calli witout plasmid (Fig 4.32). This experiment was
repeated thrice to check the reproducibilty of the experiment.
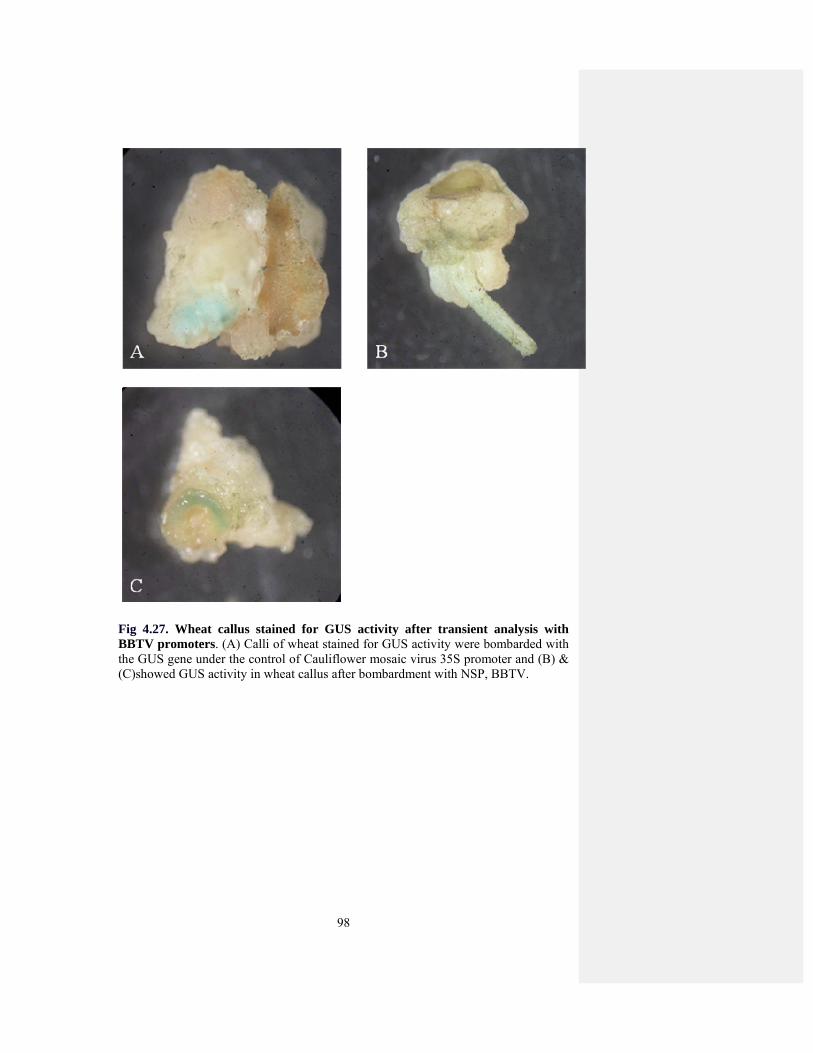
98
Fig 4.27. Wheat callus stained for GUS activity after transient analysis with BBTV promoters. (A) Calli of wheat stained for GUS activity were bombarded with the GUS gene under the control of Cauliflower mosaic virus 35S promoter and (B) & (C)showed GUS activity in wheat callus after bombardment with NSP, BBTV.

99
4.6 Development of RNAi Construct against BBTV
RNAi is believed to be very useful technique in developing resistance against
viruses in plants (Fusaro et al., 2006). This technique can be used for the study of
gene silencing and viral genes which are necessary for its replication and infection
(Baulcombe, 2004). As BBTD continues to cause heavy losses to banana production
in Pakistan. In the absence of natural sources of resistance in banana against BBTV,
the use of genetically engineered resistance is an alternate approach. In this study,
RNAi construct was made MRep, which is the key component essential for the
replication of all other components. The genetic variability is very important for
development of RNAi constructs against viruses. As genetic variability in Pakistani
isolate of BBTV is very low and we have confirmed through RFLP analysis. So this
technique can be effectively used for developing resistance against BBTV. As BBTV
genome consists of six components of 1kb in size. Out of these components DNA-R
helps in the replication of all other components of BBTV and this component is
present in all other isolates of BBTV. So, in this study RNAi construct was made
against DNA-R of BBTV. For this purpose, 600bp from N-terminal region of DNA-R
was amplified by specific primers (Fig 4.28) and cloned into T/A cloning vector. The
clone is confirmed by restriction analysis. These fragments were cloned in pFGC5941
vector in both sense and antisense orientation to direct the silencing. These clones
were confirmed by restriction with NcoI, XhoI and XbaI, BamHI restrirction enzymes
(Fig 4.29 a&b). The choice of Agrobacterium strain also has major effect on
transfortmation efficiency. So, this construct was transformed into two Agrobacterium
strains GV3101 and EHA105. This construct was then checked in bud of banana male
flower transiently. It can be checked by stable transformation in banana plants, but
establishment of banana regeneration system is very difficult task because of
excretion of phenolic compounds. Different varieties of banana have been
successfully transformed but transformation of Dwarf Cavendish (Basrai) has not
been reported. Cavendish dwarf is the most commonly used cultivar in Pakistan.
For the optimization of transformation protocols, transient expression is most
commonly used. It is used to test the expression efficiency and transfer of gene in host
plant. Most commonly used reporter genes are gusA gene and gfp gene. In this study
we used gusA gene for the optimization of transformation protocols. For this purpose
bud of male flower of banana was used as explant and placed on MS media for two

100
days. These plates were placed in growth room at 25±2oC with cool white fluorescent
light. MRep-RNAi construct cloned in Agrobacterium strains GV3101 and EHA105.
Single colony was picked from the plate and put in LB media containing appropriate
antibiotic. Acetosyringone was added in the culture before use. Male flower were
placed in the culture media for 30min and were blotted dry with a sterile filter paper
and placed on the MS media.
Co-cultivation period also plays a key role in transformation efficiency, and
co-cultivation for two days was considered to be suitable for Agrobacterium mediated
transformation. If buds were placed for more than 2 days, the buds turned black due to
the release of phenolic compounds. After 2 days, these buds were placed in GUS
substrate and incubated at 37oC overnight. Agrobacterium strain EHA105 showed
better GUS expression than GV3101. When the buds were directly placed in GUS
substrate without co-cultivation, there was no GUS expression in these buds. So,
transformation efficiency was better after 2 days co-cultivation, after this expression
efficiency decreased. Buds gave better GUS expression with EHA105 as compared to
GV3101 (Fig 4.30 a&b). After successful transient analysis, these constructs were
used for stable transformation in banana. For this purpose male flowers were used as
explants. There is callus induction after four weeks, but callus turned black due to the
release of phenolic compounds. So, an alternate host is needed to check the
effectiveness of this construct in other hosts like tobacco. Tobacco can be used as an
alternate host for BBTV, as we have studied pathogenecity and gene suppression
analysis in this host.

101
Fig 4.28. PCR amplification of MRep-RNAi construct. Lane 1: negative control. Lane 2: 600bp fragment of MRep-RNAi. .

102
Fig 4.29. Confirmation of clones in pFGC5941. (A) Lane 1,3,4 represents the 600bp fragment released after restriction with NcoI and XhoI.
(B) Lane 1,3 represents the 600bp fragment released after restriction with XbaI and BamHI.

103
Fig 4.30. Transient assay of banana male flower buds with RNAi construct. (A) shows the buds treated with MRep-RNAi cloned in GV3101 (B) shows the bud treated with MRep-RNAi cloned in EHA105.

104
4.7 Micropropagation of Banana
Micropropagation of banana is an alternate method for the production of virus
free banana. For this purpose, banana plants were collected from field and used for the
micropropagation. The suckers were trimmed to the size of 3mm long and placed in
jars containing culture media. Both liquid and solid media was used for
micropropagation. In solid media, agar is used as a gelling agent; while in liquid
media agar is not used. The cultures were placed in growth room under controlled
conditions. The media was optimized by using various concentrations of growth
regulators like BAP along with kinetin (1mg/l) in order to determine the media in
which maximum number of shoots was produced. After one week of culturing, the
creamy white meristem of explants turned green and blackening was observed at the
base of the explant due to the release of phenolic compounds. This black portion was
trimmed with the help of sterile scalpel and transferred to new jars. The size of
explant is increased and the shoots were produced on the sides of bud. Different
concentrations of BAP (1mg/l, 2mg/l, 3mg/l and 4mg/l) were used to get the
maximum shoots. Shoot multiplication increased upto certain level of BAP and then
decreased. Maximum shoots were observed at 3 mg/l of BAP along with kinetin,
followed by 4mg/l, 2mg/l and 1mg/l of BAP (Fig 4.31). All explants did not behave
alike in terms of multiplication; it varied from cultivar to cultivar. Liquid media
produced large number of shoots as compared to solid media. But the production of
shoot decreased after certain level of BAP. Subculturing of explants also played an
important role on the multiplication rate. These shoots were subcultured after two
week. When the shoots attained a sufficient length, they were transferred to rooting
media under aseptic conditions. Different concentrations of NAA were used for root
induction. Maximum number of roots was obtained at concentration of 2mg/l. Further
increase in concentration resulted in a decrease of root production. These plants were
then transferred to field conditions.

105
Table 4.3. Effect of different concentrations of BAP on multiplication of banana explants
BAP (mg/l) Number of shoots produced from explants
Results
1 0 Poor micropropagation
2 3-4 Good micropropagation
3 10-12 Highest micropropagation
4 5-7 Good micropropagation

106
Fig 4.31. In vitro multiplication of banana (A) shows meristem turned green after one week, (B) shows the multiplication of explant at 2 mg/l, (C) shows multiplication rate at 3 mg/l and (D) shows multiplication rate at 4 mg/l.

107
Chapter 5 DISCUSSION
Banana bunchy top disease continues to cause heavy yield losses to banana
production in Pakistan. In the absence of sources of natural resistance, the use of
genenetically-engineered resistance is an attractive option. Several options are
available for genetically-engineered resistance and among them RNA silencing based
technologies are likely to result in viable resistance. However, before embarking on
such program, it is important to know the diversity of pathogen, viral genes that are
important for pathogenicity and study of RNA silencing suppressors of BBTV
complex prevalent in Pakistan. This thesis was aimed at understanding these aspects
of BBTV so that prospects of genetically-engineered resistance can be explored.
Nanoviruses caused several diseases in many important crops and caused
severe losses to the economy of country. As FBNYV causes diseases in faba beans
and BBTV infect banana plants. In Pakistan banana production is severely affected by
BBTV since 1989. Banana is a monocot plant and is cultivated in southern coastal
areas of Pakistan. However, due to increasing pressure of BBTV, the banana
cultivation has moved upward in Sindh and adjoining areas of Balochistan to avoid
BBTV. Previous molecular analysis of BBTV from Pakistan has shown that it is
composed of five components (Amin et al., 2008) but another component (DNA U3)
has also been cloned from Pakistan and this is the most diverse BBTV component in
all components of BBTV (Hyder et al., 2011). Six components of BBTV have also
been characterized from geographically different isolates of banana (Karan et al.,
1994). These components have been cloned by PCR method. In this study, we used a
new technique, RCA for the amplification of unknown components of BBTV. Since
RCA is a very useful technique for the molecular characterization of unknown
components with circular genome. It can amplify the viral circular DNA of unknown
identity. In BBTV, there is a need to find out whether any additional component is
associated with the infectious unit or not. As BBTV is divided into two groups South
Pacific group and Asian group. These additional components have been characterized
from Asian group of BBTV. These were referred as DNA Y, W1, W2, S1, S2 (Horser
et al., 2001). The isolate prevalent in Pakistan belongs to South Pacific group, so there
is a need to identify these additional components from this group. RFLP technique
was used to find out these additional components. The results showed that there is no

108
additional component associated with South Pacific group of BBTV. It is possible that
these components are more widespread in Asian group as compared to South Pacific
group or it is not detected by probes used in this study. Similarly another family of
ssDNA Geminiviridae encodes only one Rep. In addition to this, additional Rep
encoding components have been identified for other members of family Nanoviridae.
In FBNYV four additional Rep encoding components have been identified (Katul et
al., 1998). In SCSV and MDV two and three additional Rep encoding components
have been identified (Boevinck et al., 1995; Sano et al., 1998). Coconut foliar decay
(CFDV) which is a possible member of Nanoviridae also showed one additional Rep
encoding components (Rohde et al., 1990). These additional Rep encoding
components are associated with nanoviruses and play an important role in infectivity
of nanoviruses.
As far as genetic diversity is concerned, phylogenetic analysis of BBTV
showed that there is very less diversity in BBTV components. Previous study has also
shown that genetic variability in South Pacific group of BBTV is less variable. These
components showed 98% homology to South Pacific group. These isolates were
closely related to Australian and Indian isolates of BBTV, which further confirmed
the results that this was disease introduced in Pakistan from Australia or India to
Pakistan. So there is single homogenous population of BBTV prevalent in Pakistan.
This low level of diversity in Pakistan can be used for obtaining resistance to the virus
by RNA silencing based technologies.
An important aspect of the research on BBTV components is the identification
of all components that make up infectious unit of BBTV. The use of RCA technology
suggests that six components identified so far make infectious unit of South Pacific
isolate of BBTV. It will there exciting to see if these six components can produce
symptoms of BBTV when inoculated on healthy banana plants. Such components can
be delivered into banana by biolistic method.
In order to study the gene functions of BBTV, there is not an established
system in banana. So, N. benthamiana was used as a host plant to study the gene
functions. But banana is a monocot plant, so there is a need to find out whether the
expression of proteins from monocots in a dicot plant has any effect on results. When
these genes were expressed in N. benthamiana they produced phenotype. Out of five
genes of BBTV, MRep and MP-PVX induced necrosis upon expression in N.
benthamiana. But necrosis is not a usual symptom of BBTV infection (Hooks et al.,

109
2009), which may indicate that this is a host specific response. In begomoviruses, the
AC2 gene prevents necrotic response induced by NSP (for the bipartite
begomovoiruses) and the V2 (for the monopartite begomoviruses) (Hussain et al.,
2005; Mubin et al., 2010). So it is possible that like geminiviruses, nanoviruses may
encode a protein that prevents necrosis in host plants, which is a host response but this
requires further analysis.
Out of five genes of BBTV, MP-PVX showed severe symptoms including
hypersensitive response and severe leaf curling of newly emerging leaves. In
monopartite begomoviruses, V1 and C4 proteins performed similar functions as MP
of BBTV (Rojas et al., 2001) and in some cases the V1 causes a necrotic response in
plants (Mubin et al., 2010). The sequence of MP of BBTV is different from MP of
begomoviruses but they performed similar functions. Thus, like begomoviruses,
proteins of BBTV are involved in movement and DNA replication produces a necrotic
response in plants (Selth et al., 2004; Wezel et al., 2002; Qazi et al., 2007). As in
bipartite begomoviruses, MP protein has the ability to redirect the NSP from nucleus
to cell periphery in GFP localization study (Wanitchakorn et al., 2000).
In geminiviruses, Rep protein binds to retinoblastoma binding protein (pRB)
of the cell cycle and induced necrosis (Selth et al., 2004). In case of nanoviruses,
Clink binds to pRB and interferes with the cell cycle (Aronson et al., 2000; Lageix et
al., 2007). This shows that cell death in geminiviruse and nanoviruses are initiated by
different mechanisms. In tobacco, silencing of pRB induced cell death and some other
abnormalities that resemble those induced by virus infection (Jordan et al., 2007).
In case of PVX expression system, MP showed severe leaf necrosis in
inoculated and infected leaves. While in case of suppressor study, MP and Clink have
the ability to suppress gene silencing activity, which is judged by the levels of GFP
fluorescence. These results showed that BBTV like other viruses has evolved a
general counter-defense mechanism. In BBTV, only two suppressors were found but
there is a possibility that there may be more than two suppressors. This conclusion is
supported by the demonstration that expression of MP and Clink-PVX increases the
viral RNA, suggesting that the proteins overcome some RNAi mediated host
resistance that limits either viral replication or spread. But there is a need to check all
these components in banana.
As MP showed severe symptoms when expressed from PVX and also have
suppressor of silencing activity suggests that this protein might be a major

110
pathogenecity determinant in BBTV. But this conclusion needs confirmation by
expression in banana. Future efforts will be aimed at examining the functions of other
unknown components from Pakistan and investigating the mechanisms by which MP
and Clink interfere with the gene silencing pathway.
Our findings are opposite to Niu et al. (2009). They reported that DNA-S and
DNA-M can suppress the silencing activity and we have shown that MP and Clink
showed the suppressor activity. One possibility is that the prevalent isolate in Pakistan
belongs to South Pacific group (Amin et al., 2008) while Hainan isolate belongs to
Asian group. These two groups are distinct from one another and during the course of
evolution; these groups may have evolved distinct suppressors. However, Niu et al.
(2009) used a field collected isolate for the amplification of the genes for expression.
They provided no sequence data to confirm the identity of the genes in their
expression constructs, leaving some doubt about the identity of the clones. In contrast,
sequence confirmed clones were used here for the isolation of genes for expression.
Clearly further analyses of the genes expressed for the “Asian” isolate will be
required to ascertain whether there is in fact a difference in the use of gene products
for suppression between “Asian” and “South Pacific” isolates of BBTV. Nevertheless,
it is clear that, in common with all other viruses so far examined, nanoviruses encode
proteins that are involved in overcoming host-defense based on RNAi. Niu et al.
(2009) also examined the phenotype induced upon PVX-mediated expression of the
same 5 BBTV genes in N. benthamiana. Generally, the symptoms induced by the
gene products of their Asian isolate were similar to those we have found for our South
Pacific isolate with the exception of the MRep protein (referred to by them as the B1
protein) which here induced necrosis but only mild yellow flecks for the Asian isolate.
RNAi has been found to be the robust technology for silencing of genes at
transcriptional and post transcriptional level. It has been shown that PTGS in plants
can be triggered at high efficiency by the presence of an inverted repeat in the
transcribed region of a transgene (Chuang and Meyerowitz, 2000). Globally, strong
evidence supports a key role of dsDNA as an inducer of PTGS. This technology has
been used to develop resistance against RNA viruses and DNA viruses. In DNA
viruses, this technology has been used against geminiviruses and there is no evidence
of its use in nanoviruses. Tenllado and Ruiz (2001) used pepper mild mottle virus

111
(PMMoV), tobacco etch virus (TEV) and alfalfa mosaic virus (AMV) for the study of
dsRNA mediated interference in plant virus infection.
The advantages of RNAi against viruses have been reported by many
researchers. In geminiviruses, RNAi has targeted a group of DNA viruses with a
circular ssDNA. The idea behind this approach is that DNA viruses transcribe mRNA
and silencing of these mRNA would result in developing resistance against viruses.
Similarly, the silencing of promoters of DNA would result in transcriptional gene
silencing. It has been shown that wide range of plant viruses is able to suppress PTGS
upon viral infection and these genes have been identified. These genes which are
called suppressors have been identified from both monocot and dicot viruses. Their
utility for overcoming PTGS has been demonstrated in transgenic N. benthamiana
where silencing of GFP was suppressed by these viral proteins (Voinnet et al., 1999).
The spatial pattern and degree of suppression varied extensively between viruses.
As RNAi technique has not been used against nanoviruses. In this study we
used this technique for developing resistance in banana against BBTV. The
assessment of genetic variability is important for developing any RNAi strategy
against viruses. Our assessment of genetic variability indicates low level of genetic
variability in the Pakistani population of BBTV. It therefore, offers great advantage of
using RNAi against this devastating virus. MRep is important gene which is
responsible for the replication of all other components of BBTV. So, RNAi construct
was made against MRep. The construct made in this study needs to be verified for its
effectiveness against BBTV in banana. This requires an effective and reproducible
banana transformation system which unfortunately is not available currently because
in banana callus induction and regeneration are very difficult due to its polyploidy and
excretion of phenolic compounds during the culturing procedure. Very few examples
have been reported for successful banana transformation for different varieties but
transformation of Musa acuminata Colla “Dwarf Cavendish” (Basrai), the major
cultivar in Pakistan, has rarely been reported. Different cultivars of banana like
Bluggoe, Grand Naine, Three Hand Planty and Williams have been successfully
transformed (Dugdale et al., 1998; Arinaitwe, 2008; Sagi et al., 1995a) Therefore, it is
needed to develop an alternative to check the effectiveness of this and similar
constructs against BBTV in some other plants which can easily be transformed like
tobacco. As we found that BBTV genes showed symptoms in tobacco plants. So,
tobacco can be used as a host plant to check RNAi constructs.

112
RNAi construct was transiently checked in bud of banana male flower.
Transient analysis helped us in the optimization of banana transformation protocol.
Co-cultivation of buds for GUS expression plays an important role in the
transformation. It has been observed that co-cultivation for 3 days is suitable for
transformation. Similar results have been reported in broccoli (Metz et al., 1995) and
Brassica napus (Takasaki et al., 1997). Co-cultivation for more than 3 days led to a
decrease in transformation frequency and 50% of explants loss due to the overgrowth
of bacteria in experiments to produce transgenic Datura (Ducrocq et al., 1994).
Similarly, in wheat, the longer co-cultivation reduced the capacity of the immature
embryos to form embryogenic callus and regeneration (Wu et al., 2003). Although
prolonged co-cultivation period for more than 3 days have been successfully used for
certain plants (Suzuki et al., 2001; Yu et al., 2001). But 2-3 days co-cultivation has
been routinely used in most reported transformation protocols. Mondal et al. (2001)
reported that the differential requirement of co-cultivation period largely depends
upon Agrobacterium strain used or the medium for bacterial culture.
The choice of Agrobactertium strain can have the major effect on
transformation efficiencies. Agrobacterium strain EHA105 was more effective as
compared to GV3101. Virulence of Agrobacterium strains varies widely among host
plants (Davis et al., 1991) and is particularly important for the transformation of
recalcitrant species. Several studies have shown that EHA105 to be more effective for
transformation than other strains of Agrobacterium tumefaciens (Tsukazaki et al.,
2002). Our findings are different from this, in our study GV3101 gave GUS
expression more than EHA105. The size of explant of 3mm is suitable for
transformation. This could be due to the small and delicate explants, perhaps received
more DNA. Increase in size was found to have lower GUS expression. Therefore,
3mm size of single bud was chosen as a suitable target size for transformation study.
In this case, buds showed the GUS expression in buds treated with EHA105 as
compared to GV3101. These buds produced callus but after few days, the callus
turned brown due to the release of phenolic compounds. One possibility is that it may
be due to cultivar used in Pakistan. As banana transformation is greatly effected by
cultivar and there is no report of successful banana transformation from Pakistan.
Micropropagation of banana is also a good method to produce virus free
banana by tissue culture. In banana micropropagation, different concentrations of
cytokinin play a key role. Moderate level of cytokinin increased the shoot

113
proliferation rate, but very high concentration decreased multiplication and depressed
shoot elongation. Shoot multiplication is increased upto the level of 3mg/l.
Furthermore, the joint effect of BAP and kinetin (1mg/l) increased shoot length more
as compared to BAP alone. The optimum BAP concentration does not vary
significantly among researchers. Arinaitwe et al. (2000) stated that shoot proliferation
was cultivar dependent. They also reported that increased concentration of BAP did
not significantly increase the shoot proliferation. Our results showed that 3mg/l
concentration of BAP is optimum for maximum shoot multiplication and beyond this
concentration the shoot proliferation rate is decreased. Rooting can be stimulated
when individual shoots are transferred to basal medium alone (Jarret et al., 1985).
However, auxins may induce further root initiation and NAA was more effective than
IAA (Vuylsteke, 1989). Our results also showed that NAA gave the maximum
number of shoots at the concentration of 2mg/l. Cronaeur and Krikorian (1984)
reported that there is no effect on rooting in NAA, IAA in the presence of 0.025%
charcoal. Our results also showed that NAA is necessary for root proliferation as
compared to IAA. When RNAi construct was agro-inoculated into banana leaves,
they did not show any GUS expression due to blackening of leaves.
We have demonstrated by transient assays that the non-coding regions of
BBTV components have promoter activity in N. benthamiana. The activities of
different non-coding regions vary considerably. The relative promoter activities from
different components are, however, probably relevant to BBTV replication. Promoter
activity in N. benthamiana was checked using GUS as a reporter gene. BBTV CP and
Clink did not show any GUS activity, while NSP and MRep showed some level of
GUS activity in N. benthamiana. But the expression of GUS in both genes is less as
compared to CaMV, 35S. However, it is likely that BBTV gene expression and
replication is a complex mechanism. In geminviruses, promoter activity is influenced
via the interaction of virus encoded gene products (Haley et al., 1992; Sunter and
Bisaro, 1992; Zhan et al., 1993) and gene expression in certain cell types may be
dependent on complex transcriptional transactivation mechanisms (Sunter and Bisaro,
1997). Therefore, the expression patterns associated with each BBTV promoter alone
may differ from the expression patterns from each promoter in a natural infection.
Apart from their involvement in the regulation of BBTV gene expression, the non-
coding regions of BBTV may also have applications for heterologous transgene
expression. Therefore, we found that whether any of cis-elements identified in BBTV

114
DNA-N non-coding region were involved in promoter activity. Removal of 200bp
from 5’ end of DNA-N promoter increased the promoter activity but is still less as
compared to CaMV, 35S. Similarly the removal of 20bp from 5’ end of MRep
showed a small increase in GUS expression but the expression is less as compared to
NSP. Our results similar to Dugdale et al. (1998) they also showed that GUS
expression is strong in BBTV-NSP as compared to other components in tobacco and
banana plants. Promoters derived from BBTV DNA 4 and 5 generated the highest
level of transient activity, which were greater than that of maize ubi- 1 promoter. In
transgenic banana plants the promoter activity is restricted to phloem of roots, leaves
and roots (Dugdale et al., 1998). We found that in full length promoter sequence GUS
activity is low, while in deleted NSP GUS activity was higher and it is phloem
specific. We used N. benthamiana as a host plant, which is a dicot and banana is a
monocot plant. So we tried to establish transformation protocol in alternate host,
wheat, which is monocot. For this purpose, we bombarded wheat callus with BBTV-
NSP by particle bombardment. The wheat callus did not show GUS expression in
NSP bombarded calli as compared to CaMV, 35S. The possible reason is that wheat is
not the host of BBTV and phloem specificity can not be checked in callus as it is
undifferentiated mass of cells. There is a need to check the promoter activity in
banana. So, promoter analysis should be performed under greenhouse and field
conditions to assess their possible use for transgenic banana production.

115
CONCLUSIONS
On the basis of results of present study, it can be concluded that
1. Single homogenous population of BBTV is prevalent in Pakistan.
2. There is no diversity of BBTV as MP and NSP showed 98% homolgy to
already known sequences from Pakistan.
3. BBTV isolate of Pakistan belongs to South pacific group.
4. All BBTV genes induced a phenotype when inoculated in plants.
5. In case of PVX expressed genes, MP showed severe symptoms of leaf
necrosis and curling, while other genes produced mild symptoms in N.
benthamiana.
6. In case of suppressor study, MP and Clink have the ability to suppress
gene silencing activity, which is judged by the levels of GFP fluorescence.
7. MP and Clink showed the minute amount of GFP specific siRNAand
maintains GFP mRNA levels.
8. MRep, NSP and CP did not show silencing activity in study of gene
suppression analysis.
9. Out of five genes of BBTV, MP is considered as major pathogenecity
determinant.
10. NSP promoter of BBTV showed some level of gene expression, while all
other genes did not show any promoter activity.
11. Full length intergenic region showed lower level of expression as
compared to deleted region.
12. Utility of BBTV promoters in monocot transformation.

116
FUTURE PROSPECTS
Banana bunchy top disease continues to cause heavy yield losses to banana
production in Pakistan. BBTV is a ssDNA virus belong to family Nanoviridae. This
family is divided into two genera: Nanoviruses and Babuviruses. Babuvirus include
Banana bunchy top virus (BBTV) and this virus has been characterized from
Pakistan. Nanoviruses include SCSV, MDV and FBNYV these viruses have not been
characterized from Pakistan. So, there is a need to characterize these viruses from
Pakistan. As it is evident from the present study that a single homogenous population
of BBTV is prevalent in Pakistan. But there is a need to collect the samples from
different areas of Sindh and evaluated for their genetic variability. There is still a need
to characterize and study the functions of other unknown components from Pakistan.
As these additional components have been characterized from Asian group of BBTV.
In the abscence of sources of natural resistance, the use of genenetically-
engineered resistance is an attractive option. RNA silencing techniques like RNAi is
very useful for developing resistance. RNAi construct was made against MRep of
BBTV. This construct was transiently checked in the bud of banana male flower.
Future efforts will be aimed at stable transformation of banana against MRep-RNAi.
Gene function study of BBTV genes were transiently checked in N.
benthamiana plants. Future efforts will be aimed at examining the functions of these
BBTV genes in banana and investigating the mechanism by which MP and Clink
interfere with the gene silencing pathway.
Micropropagation of banana is lacking in Pakistan and thus can be further
explored for the production of somatic embryos and production of banana on large
scale. As it is a very suitable method for the production of virus free banana. Promoter
study should be performed under greenhouse and field conditions to assess their
possible use for transgenic banana production.

117
SUMMARY
Banana bunchy top disease continues to cause heavy losses to banana
production in Pakistan. Several options are available for genetically engineered
resistance and among them RNA silencing based technologies are likely to result in
viable resistance. In this study, RNAi technology was used to develop resistance in
banana against BBTV and the identification of unknown components of BBTV by a
new technique called RCA. BBTV is the member of genus Babuvirus of Family
Nanoviridae. Nanoviruses are ssDNA viruses transmitted by different vectors. BBTV
is transmitted by Pentalonia nigronervosa. There are four genera of nanoviruses
which include FBNYV, MDV, SCSV and BBTV. An analysis of the genetic diversity
of BBTV across Tando Jam, Sindh, Pakistan was conducted. Sequence analysis
showed that there is low diversity in BBTV. RCA technique to identify the unknown
components of BBTV, as RCA can amplify the unknown components of circular
genome. The identification of unknown component is also necessary to find out
whether any additional component is associated with infectious unit or not. We did
not find any additional component with BBTV isolate. These components have been
characterized from Asian group and Pakistani BBTV isolates belong to Asian group.
This low diversity was also helpful in generating resistance against viruses. So, RNAi
construct was made against MRep of BBTV to engineer resistance against BBTV.
This construct was transiently checked in banana male flower bud. The buds agro-
infiltrated with EHA105 gave better expression as compared to GV3101.
Expression of BBTV genes from PVX and under 35S promoter was also
observed. Expression of MRep and MP under PVX resulted in necrosis and cell death
at the site of inoculation and severe leaf curling and necrosis in newly emerging
leaves in MP. Clink, NSP and CP produced mild symptoms of leaf curling and
mosaic, while CP produced necrotic response in inoculated leaves. When all these
genes were expressed under 35S promoter in N. benthamiana 16c line, MP and Clink
stabilized GFP specific mRNA and reduced GFP specific siRNA. MRep, NSP and CP
did not show accumulation of GFP mRNA. These results identified that MP and Clink
are supressors of silencing. The ability of MP to induce severe necrosis in inoculated
and systemic leaves and RNA silencing suppressors indicates that MP is a major
pathogenecity determinant in BBTV genome.

118
To study the expression of desired genes via genetic transformation promoters
are required. However, only a limited number of promoters from banana have been
isolated, characterized and used in transgenic bananas. In the present study, promoters
region of BBTV genes were cloned into pJIT166. In all BBTV genes, NSP is involved
in promoter activity. The deleted NSP of 200bp from 5’ end increases promoter
activity but was still less as compared to CaMV, 35S. As banana is a monocot plant,
we tried to establish transformation protocol in monocot plant because transformation
in banana is very difficult task. Wheat callus is bombarded with NSP-BBTV by
particle bombardment. They did not show any remarkable GUS expression in NSP
bombarded calli as compared to CaMV, 35S. So there is a need to check promoter
activity in banana. So, promoter analysis should be performed under greenhouse and
field conditions to assess their possible use for transgenic banana production.

119
LITERATURE CITED
Abdel-Aziz, N.A., A.M. Abdel-Salam, H. N. Soliman and S. M. El-Saghir. 1998.
Heat and chemotherapeutic agents as tools for elimination of two banana
viruses. Egyp. J. Phytopathol., 26:13-28.
Acereto-Escoffie, P. O. M., B. H .C. Mazanero, S. E. Echeverria, R. Grijalva, A. J.
Kay, T. G. Estarada, E. Castano, L. C. R. Zapata. 2005. Agrobacterium
mediated transformation of Musa acuminate cv. Grand Nain scalps by vacuum
infiltration. Sci. Hortic., 105: 359-371.
Adams, M. J., J. F. Antoniw, M. Bar-Joseph, A. A. Brunt, T. Candresse, G. D. Foster,
G. P. Martelli, R.G. Milne, S. K. Zavriev and C. M. Fauquet. 2004. The new
plant virus family Flexiviridae and assessment of molecular criteria for species
demarcation. Arch. Virol., 149: 1045-1060.
Agarwal, N., P. V. N. Dasaradhi, A. Mohammed, P. Malhotra, R. K. Bhatnager and S.
K. Mukherjee. 2003. RNA interference: Biology, Mechanism and
Applications. Microbiol. Mol. Biol. Rev., 67:657-685.
Akhtar, S., S. A. Wani, M.A. Bhat, N. A. Zeerak, S. Chalkoo, H. Ashraf, K. A. Bhat,
N. Wani, A. Anwar, S. A. Wani and M. R. Mir. 2010. RNA interference:
present status and future prospects in ameliorating the crop yield. Int. J. Curr.
Res., 4: 127-134.
Allen, E., Z. Xie, A. M. Gustafson and J. C. Carrington. 2005. microRNA-directed
phasing during trans-acting siRNA biogenesis in plants. Cell, 121:207–211.
Altpeter, F., V. Vasil, V. Srivastava and E. Stoger. 1996. Accelerated production of
transgenic wheat (Triticum aestivum L.) plants. Plant Cell Rep., 16:12-16.
Amin, I., J. Qazi, S. Mansoor, M. Ilyas and R. W. Briddon. 2008. Molecular
characterization of Banana bunchy top virus from Pakistan. Virus Genes,
36:191-198.
Anandalakshmi, R., G. J. Pruss, X. Ge, R. Marathe, A. C. Mallory, T. H. Smith and V.
B. Vance. 1998. A viral suppressor of gene silencing in plants. Proc. Nat.
Acad. Sci. USA, 95: 13079–13084.
Anandalakshmi, R., R. Marathe, X. Ge, J.M. Herr, C. Mau, A. Mallory, G. Pruss, L.
Bowman and V.B. Vance. 2000. A calmodulin-related protein that suppresses
posttranscriptional gene silencing in plants. Science, 290:142-144.

120
Andersen, M. T., K. A. Richardson, S. A. Harbison and B. A. M. Morris. 1988.
Nucleotide sequence of the geminivirus Chloris striate mosaic virus.
Virology, 164:443-449.
Angell, S.M. and D.C. Baulcombe. 1999. Potato virus X amplicon mediated silencing
of nuclear genes. Plant J., 20:357-362
Angell, S. M., C. Davies and D. C. Baulcombe. 1996. Cell-to-cell movement of
Potato virus X is associated with a change in the size exclusion limit of
plasmodesmata in trichome cells of Nicotiana clevelandii. Virology, 215: 197-
201.
Anon. 2006. FAOSTAT Database Food and Agricultural Organisation of the United
Nations Rome. (available from: http://faostat.fao.org)
Arguello-Astorga, G., L. Lopez-Ochoa, L. J. Kong, B. M. Orozco, S. B. Settlage and
L. Hanley-Bowdoin. 2004. A novel motif in geminivirus replication proteins
interacts with the plant retinoblastoma-related protein. J. Virol., 78: 4817–
4826.
Arinaitwe, G. 2008. An improved Agrobacterium-mediated transformation method
for banana and plantain (Musa spp.). Dissertationes de agricultura. Ph.D.
thesis 781. Katholieke Universiteit Leuven, Belgium, p. 156
Arinaitwe, G., P. R. Rubaihayo and M. J. S. Magambo. 2000. Proliferation rate effects
of cytokinins on banana (Musa spp.) cultivars. Sci. Hortic., 86: 13–21.
Arinaitwe, G., S. Remy, H. Strosse, R. Swennen and L. Sagi. 2004. Agrobacterium-
and particle bombardment-mediated transformation of a wide range of banana
cultivars. In: Banana Improvement: Cellular, Molecular Biology and Induced
Mutations. S. M. Jain and R. Swennen, editors. Science Publishers Inc.,
Enfield, NH, USA. pp. 351-357.
Aronson, M. N., A. D. Meyer, J. Gyorgyey, L. Katul, H. J. Vetten, B. Gronenborn and
T. Timchenko. 2000. Clink, a nanovirus-encoded protein, binds both pRB and
SKP1. J. Virol., 74: 2967–2972.
Assani, A., F. Bakry, F. Kerbellec, R. Haicour, G. Wenzel and B. Foroughi-Wher. 2003.
Production of haploids from anther culture of banana (Musa balbisiana). Plant
Cell Rep., 21: 511–516.
Atabekov, J. G. and M. E. Taliansky. 1990. Expression of a plant virus coded
transport function by different viral genomes. Adv. Virus Res., 38: 201-248.

121
Atkinson, H. J., S. Grimwood, K. Johnston and J. Green. 2004. Prototype
demonstration of transgenic resistance to the nematode Radopholus similis
conferred on banana by a cystatin. Trans. Res., 13:135-142.
Azam, F. M. S., S. Islam, M. Rahmatullah and A. Zaman. 2010. Clonal Propagation
of Banana (Musa spp.) cultivar ‘BARI-1’ (AAA Genome, Sapientum
Subgroup). Acta Hortic., 879: 537-544.
Baker, B., P. Zambryski, B. Staskawicz and S. P. Dinesh-Kumar. 1997. Signaling in
plant-microbe interactions. Science, 276: 726–733.
Bass, B. 2000. Double-stranded RNA as a template for gene silencing. Cell, 101:
235–238.
Baulcombe, D. 2004. RNA silencing in Plants. Nature, 431: 356–363.
Baulcombe, D. C., S. Chapman and S. Santa Cruz. 1995. Jellyfish green fluorescent
protein as a reporter for virus infections. Plant J., 7: 1045-1053.
Beck, D. L., P. J. Guilford, D. M. Voot, M. T. Andersen and R. L. S. Forster. 1991.
Triple gene block proteins of white clover mosaic potexvirus are required for
transport. Virol., 183: 695-702.
Becker, D., K. B. Dugdale, M. K. Smith, R. M. Harding and J. L. Dale. 2000. Genetic
transformation of Cavendish banana (Musa spp. AAA group) cv ‘Grand Nain’
via microprojectile bombardment. Plant Cell Rep., 19: 229-234.
Beetham, P. R., G. J. Hafner, R. M. Harding and J. L. Dale. 1997. Two mRNAs are
transcribed from Banana bunchy top virus DNA-1. J. Gen. Virol., 78: 229-
236.
Beetham, P. R., R. M. Harding and J. L. Dale. 1999. Banana bunchy top virus DNA-2
to 6 are monocistronic. Arch. Virol., 144: 89-105.
Beijerinck, M.W. 1898. Over een contagium vivum fluidum als oorzak van de
vlekziekte der tabaksbladen. Verslag van de Gewone Vergadering van de
Afdeling Wis-en Natuurkunde, Koninklijke Akademie van Wetenschappen,
Amsterdam, 7: 229-235.
Bell, K., J. L. Dale, C. Ha, M. Vu and P. Revill. 2002. Characterisation of Rep-
encoding components associated with Banana bunchy top virus in Vietnam.
Arch. Virol., 147: 695-707.
Bendahmane, A., K. Kanyuka and D. C. Baulcombe. 1999. The Rx gene from potato
controls separate virus resistance and cell death responses. Plant Cell, 11:
781–791.

122
Bendahmane, M. and B. Gronenborn. 1997. Engineering resistance against Tomato
yellow leaf curl virus (TYLCV) using antisense RNA. Plant Mol. Biol., 33:
351–357.
Berk, A.J. 1999. Activation of RNA polymerase II transcription. Curr. Opin. Cell
Biol., 11: 330-335.
Bernstein, E., A. A. Caudy, S. A. Hammond and G. J. Hannon. 2001. Role for a
bidentate ribonuclease in the initiation step of RNA
interference. Nature, 409:363-366.
Bhagwat, B. and E. J. Duncan. 1998. Mutation breeding of Highgate (Musa
acuminata, AAA) for tolerance to Fusarium oxysporum F. sp. Cubense using
gamma irradiation. Euphytica, 101: 143–150.
Bisaro, D. M. 2006. Silencing suppression by geminivirus proteins. Virology, 344:
158-168.
Boevinck, P., P. W. G. Chu and P. Keese. 1995. Sequence of subterranean clover
stunt virus DNA: affinities with geminiviruses. Virology, 207: 354-361.
Bosque-Perez, N. A., G. D. May and C. J. Arntzen. 2000. Applicability of an
Agrobacterium-based system for the transformation of Musa species with
diverse genomic constitution and polidy level. Acta Hortic., 540: 193-201.
Boulton, M. I., D. I. King, J. Donson and J. W. Davies. 1991. Point substitutions in a
promotor-like region and the V1 gene affect the host range and symptoms of
maize streak virus. Virology, 183: 114-121.
Boulton, M. I., H. Steinkellner, J. Donson, P. G. Markham, D. I. King and J. W.
Davies. 1989. Mutational analysis of the virion-sense genes of Maize streak
virus. J. Gen. Virol., 70: 2309–2323.
Breathnach, R. and P. Chambon. 1981. Organization and expression of eukaryotic
split genes-coding for proteins. Annu. Rev. Biochem., 50: 349-383.
Briddon, R. W., J. Watts, P.G. Markham and J. Stanley. 1989. The coat protein of
beet curly top virus is essential for infectivity. Virology, 172: 628–633.
Briddon, R. W., M. S. Pinner, J. Stanley and P. G. Markham. 1990. Geminivirus coat
protein gene replacement alters insect specificity. Virology, 177: 85–94.
Brigneti, B., O. Voinnet, W. X. Li, L.H. Ji, S. W. Ding and D. C. Baulcombe. 1998.
Viral pathogenicity determinants are suppressors of transgene silencing in
Nicotiana benthamiana. EMBO J., 17:6739-6746.

123
Brough, C. L., R. J. Hayes, A. J. Morgan, R. H. A. Coutts and K. W. Buck. 1988.
Effects of mutagenesis in vitro on the ability of cloned Tomato golden mosaic
virus DNA to infect Nicotiana benthamiana plants. J. Gen. Virol., 69: 503–
514.
Brunetti, A., R. Tavazza, E. Noris, A. Lucioli, G. P. Accotto and M. Tavazza. 2001.
Transgenic expressed T-Rep of Tomato yellow leaf curl Sardinia virus acts as
a trans- dominant-negative mutant, inhibiting viral transcription and
replication. J. Gen. Virol., 75: 10573-10581.
Burns, T. M., R. M. Harding and J. L. Dale. 1994. Evidence that Banana bunchy top
virus has a multiple component genome. Arch Virol., 137: 371-380.
Burns, T. M., R. M. Harding and J. L. Dale. 1995. The genome organization of
Banana bunchy top virus: analysis of six ssDNA components. J. Gen. Virol.,
76: 1471-1482.
Campos-Olivas, R., J. M. Louis, D. Clerot, B. Gronenborn and A. M. Gronenborn.
2002. The structure of a replication initiation protein unites diverse aspects of
nucleic acid metabolism. Proc. Nat. Acad. Sci. USA, 99: 10310–10315.
Castillo, A. G., D. Collinet, S. Deret, A. Kashoggi and E. R. Bejarano. 2003. Dual
interaction of plant PCNA with geminivirus replication accessory protein
(REn) and viral replication protein (Rep). Virology, 312: 381–394.
Castillo, A. G., L. J. Kong, L. Hanley-Bowdoin and E. R. Bejarano. 2004. Interaction
between a geminivirus replication protein and the plant sumoylation system. J.
Virol., 78: 2758-2769.
Chapman, E. J., A.I. Prokhnevsky, K. Gopinath, V. V. Dolja and J. C. Carrington.
2004. Viral RNA silencing suppressors inhibit the microRNA pathway at an
intermediate step. Genes Dev., 18: 1179-1186.
Chapman, S., G. Hills, J. Watts and D. C. Baulcombe. 1992a. Mutational analysis of
the coat protein gene of potato virus X: Effects on virion morphology and viral
pathogenicity. Virology, 191: 223-230.
Chapman, S., T. Kavanagh and D. C. Baulcombe. 1992b. Potato virus X as vector for
gene expression in plants. Plant J., 2: 549-557
Choudhury, N. R., P. S. Malik, D. K. Singh, M. N. Islam, K. Kaliappan and S. K.
Mukherjee. 2006. The oligomeric Rep protein of Mungbean yellow mosaic
India virus (MYMIV) is a likely replicative helicase. Nucl. Acids Res., 34:
6362–6377.

124
Christensen, A. H., R. A. Sharrock and P. H. Quail. 1992. Maize polyubiquitin genes:
structure, thermal perturbation of expression and transcript splicing, and
promoter activity following transfer to protoplasts by electroporation. Plant
Mol. Biol., 18: 675-689.
Chu, P., P. Boevink, B. Surin, P. Larkin, P. Keese and P. Waterhouse. 1995. Non-
geminated single-stranded DNA plant viruses. In: Singh, R.P., Singh, U.S.,
Kohmoto, K. (Eds.), Pathogenesis and Host Specificity in Plant Diseases, vol.
III, Viruses and Viroids. Pergamon Press, Tarrytown, NY, pp. 311–341.
Chu, P.W. and K. Helms. 1988. Novel virus-like particles containing circular single-
stranded DNAs associated with subterranean clover stunt disease. Virology,
167:38–49.
Chuang, C. F. and E. M. Meyerowitz. 2000. Specific and heritable genetic
interference by double-stranded RNA in Arabidopsis thaliana. Proc. Nat.
Acad. Sci., 97: 4985–4990.
Ciechanover, A., A. Orian and A. L. Schwartz. 2000. Ubiquitin mediated proteolysis:
biological regulation via destruction. Bioassays, 22:442–451.
Cogoni, C. and G. Macino. 2000. Post-transcriptional gene silencing across
kingdoms. Genes Dev., 10: 638-643.
Collin, S., M. Fernandezlobato, P. S. Gooding, P. M. Mullineaux and C. Fenoll. 1996.
The two non-structural proteins from Wheat dwarf virus involved in viral gene
expression and replication are retinoblastoma-binding proteins. Virology, 219:
324–329.
Creste, S., A. T. Neto, R. Vencovsky, S. Oliveira Silva and A. Figueira. 2004. Genetic
diversity of Musa diploid and triploid accessions from the Brazilian banana
program estimated by microsatellite markers. Gen. Res. Crop Evol., 51:723–
733.
Cronauer, S. S. and A. D. Krikorian. 1984. Rapid multiplication of banana and
plantains by in vitro. Hortic. Sci., 19: 234–235.
Crouch, J.H., D. Vuylsteke and R. Ortiz. 1998. Perspectives and applications of
biotechnology to assist the genetic enhancement of plantain and banana (Musa
spp.). Electro. J. Biotech., 1:11–22.
Dale, J. L. 1987. Banana bunchy top: an economically important tropical plant virus
disease. Adv. Virus Res., 33:301-325.

125
Dale, J. L., D. A. Phillips and J. N. Parry. 1986. Double stranded RNA in banana
plants with bunchy top disesase. J. Gen. Virol., 67: 371-375.
Darvari, F. M., M. Sariah, M. P. Puad and M. Maziah. 2010. Micropropagation of
some Malaysian banana and plantain (Musa sp.) cultivars using male flowers.
African J. Biotechnol., 45: 2360-2366.
Dasgupta, I., V. G. Malathi and S. K. Mukherjee. 2003. Genetic engineering for virus
resistance. Curr. Sci., 84: 341-354.
Davis, M. E., A. R. Miller and R. D. Lineberger. 1991. Temporal competence for
transformation of Lycopersicon esculentum (L.) Mill) cotyledons by
Agrobacterium tumefaciens. Relation to woundhealing and soluble plant
factors. J. Exp. Bot., 42: 359-364.
De Jong, W. and P. Ahlquist. 1992. A hybrid plant RNA virus made by transferring
the noncapsid movement protein from a rod-shaped to an icosahedral virus is
competent for systemic infection. Proc. Nat. Acad. Sci. USA, 89: 6808-6812.
Deom, C. M., M. J. Oliver and R. M. Beachy. 1987. The 30 kilodalton gene product
of tobacco mosaic virus potentiates virus movement. Science, 237: 389-394.
Dickinson, V. J., J. Halder and C. J. Woolston. 1996. The product of Maize streak
virus ORF V1 is associated with secondary plasmodesmata and is first
detected with the onset of viral lesions. Virology, 220:51–59.
Diekmann, M. and C. A. J. Putter. 1996. FAO/IPGRI Technical Guidelines for the
Safe Movement of Germplasm; No.15. Musa. 2nd edition. Rome Food and
Agriculture Organization of the United Nations, Rome/International Plant
Genetic Resources Institute. Rome.
Dinesh-Kumar, S. P., S. Whitham, D. Choi, R. Hehl, C. Corr and B. Baker. 1995.
Transposon tagging of tobacco mosaic virus resistance gene N: Its possible
role in the TMV-N-mediated signal transduction pathway. Proc. Nat. Acad.
Sci., 92: 4175-4180.
Dinesh-Kumar, S. P., W. H. Tham and B. Baker. 2000. The structure–function
analysis of the tobacco mosaic resistance gene N. Proc. Nat. Acad. Sci., 97:
14789–14794.
Dobrzanska, M., C. Krysiak and E. Kraszewska. 1997. Transient and stable
transformation of wheat with DNA preparations delivered by a biolistic
method. Acta Physiol. Plant., 19: 277-284.

126
Donson, J., G. P. Accotto, M. I. Boulton, P. M. Mullineaux and J. W. Davies. 1987.
The nucleotide sequence of a geminivirus from Digitaria sanguinalis.
Virology, 161: 160-169.
Doyle, J. J. and J. L. Doyle. 1990. Isolation of plant DNA from fresh tissue. Focus,
12: 13–15.
Dry, I., J. Krake, J. Rigden and A. Rezaian. 1997. A novel subviral agent associated
with geminivirus: the first report of a DNA satellite. Proc. Nat. Acad. Sci.
USA, 94: 7088-7093.
Duan, Y. P., C. A. Powell, D. E. Purcifull, P. Broglio and E. Hiebert. 1997.
Phenotypic variation in transgenic tobacco expressing mutated geminivirus
movement/pathogenicity (BC1) proteins. Mol. Plant-Microbe Interactions, 10:
1065–1074.
Ducrocq, C., S. R. Sangwan and S. B. Sangwan-Norrel. 1994. Production of
Agrobacterium-mediated transgenic fertile plants by direct somatic
embryogenesis from immature zygotic embryos of Datura innoxia. Plant Mol.
Biol., 25:995-1009.
Dugdale B., D. K. Becker, P. R. Beetham, R. M. Harding and J. L. Dale. 2000.
Promoters derived from Banana bunchy top virus DNA-1 to -5 direct
vascular-associated expressions in transgenic banana (Musa spp.). Plant Cell
Rep., 19: 810-814.
Dugdale B., P. R. Beetham, D K. Becker, R. M. Harding and J. L. Dale. 1998.
Promoter activity associated with the intergenic regions of Banana bunchy top
virus DNA-1 to-6 in transgenic tobacco and banana cells. J. Gen. Virol., 79:
2301-2311.
Dunoyer, P., C. H. Lecellier, E. A. Parizotto, C. Himber and O. Voinnet. 2004.
Probing the microRNA and small interfering RNA pathways with virus-
encoded suppressors of RNA silencing. Plant Cell, 16: 1235-1250.
Dynan, W. S. 1986. Promoters for housekeeping genes. Trends Genet., 2: 196-197.
Egelkrout, E. M., L. Mariconti, S. B. Settlage, R. Cella, D. Robertson and S. Hanley-
Bowdoin. 2002. Two E2F elements regulate the proliferating cell nuclear
antigen promoter differently during leaf development. Plant Cell, 14: 3225–
3236.
Elbashir, S. M., W. Lendeckel and T. Tuschl. 2001. RNA interference is mediated by
21- and 22-nucleotide RNAs. Gene Dev., 15: 188-200.

127
Espino, R. C., L. V. Magnaye, A. P. John and C. Juanillo. 1993. Evaluation of
Phillipine banana bunchy top and fusarium wilt. Pages 89-103 in Proc. Int.
Symop. Recent Developments in banana cultivation technology, edited by
Valmayor, R.V, S.C. Hwang, R. Ploetz, S.C. Lee, N.V. Rao, Taiwan Banana
Research Institute, 14-18 December 1992, Chiuju, Pingtung, Taiwan,
INIBAP/ASPNET, Los Banos, the Phillipines.
Etessami, P., R. Callis, S. Ellwood and J. Stanley. 1988. Delimitation of essential
genes of Cassava latent virus DNA 2. Nucl. Acids Res., 16: 4811–4829.
Faivre-Rampant, O., E.M. Gilroy, K. Hrubikova, I. Hein, S. Millam, G.J. Loake, P.
Birch, M. Taylor and C. Lacomme. 2004. Potato virus X-induced gene
silencing in leaves and tubers of potato. Plant Physiol., 134: 1308-1316.
Food and Agriculture Organization (FAO). 2004. Production Year Book. FAO,
Rome, Italy
Food and Agriculture Organization (FAO). 2007. The International Food and
Agricultural Statistics. Database of the Food and Agriculture Organization
(FAO) of the United Nations. Available at: http://faostat.fao.org.
Fauquet, C., R. Briddon, J. Brown, E. Moriones, J. Stanley, M. Zerbini and X. Zhou.
2008. Geminivirus strain demarcation and nomenclature. Arch. Virol.,
153:783–821.
Fire, A., S. Xu, M. K. Montgomery, S. A. Kostas, S. E. Driver and C. C. Mello. 1998.
Potent and specific genetic interference by double-stranded RNA
in Caenorhabditis elegans. Nature, 391: 806-811.
Flor, H. H. 1971. Current status of the gene-for-gene concept. Annu. Rev.
Phytopathol., 9: 275–296.
Fontes, E. P. B., P. A. Eagle, P. S. Sipe, V. A. Luckow and L. Hanley Bowdoin. 1994.
Interaction between a geminivirus replication protein and origin DNA is
essential for viral replication. J. Biol. Chem., 269: 8459–8463.
Fontes, E. P. B., V.A. Luckow and L. Hanley-Bowdoin. 1992. A geminivirus
replication protein is a sequence-specific DNA binding protein. Plant Cell, 4:
597–608.
Franz, A., K. M. Makkouk and H. J. Vetten. 1997. Host range of faba bean necrotic
yellows virus and potential yield loss in infected faba bean. Phytopathol.
Mediterr., 36: 94-103.

128
Franz, A., K. M. Makkouk and H. J. Vetten. 1998. Acquisition, retention and
transmission of faba bean necrotic yellows virus by two of its aphid vectors,
Aphis craccivora (Koch) and Acyrthosiphon pisum (Harris). J. Phytopathol.,
146: 347–355.
Franz, A.W., F. van der Wilk, M. Verbeek, A. M. Dullemans and J. F. van den
Heuvel. 1999. Faba bean necrotic yellows virus (genus Nanovirus) requires a
helper factor for its aphid transmission. Virology, 262: 210-219.
Frischmuth, S., C. Wege, D. Hülser and H. Jeske. 2007. The movement protein BC1
promoters redirection of the nuclear shuttle protein BV1 of Abutilon mosaic
geminivirus to the plasma membrane in fission yeast. Protoplasma, 230: 117–
123.
Furuya, N., N. B. Bahet, S. Kawano and K. Natsuki. 2003. Detection of viruses and
genetic analysis of Banana bunchy top virus (BBTV) associated with abaca.
Ann. Phytopathol. Soc. Jap., 69: 26-327.
Fusaro, A.F., L. Matthew, N. A. Smith, S. J. Curtin, J. Dedic-Hagan, G.A.
Ellacott, J.M. Watson, M. B. Wang, C. Brosnan, B. J. Carroll and P. M.
Waterhouse. 2006. RNA interference-inducing hairpin RNAs in plants act
through the viral defense pathway. EMBO Rep., 7: 1168-1175.
Ganapathi, T. R., N. S. Higgs, P. J. Balint-Kurti, C. J. Arntzen, G. D. May and J. M.
Van Eck. 2001. Agrobacterium-mediated transformation of embryogenic cell
suspensions of the banana cultivar Rasthali (AAB). Plant Cell Rep., 20: 157-
162.
Garrido-Ramirez, E. R., M. R. Sudarshana, W. J. Lucas and R. L. Gilbertson. 2000.
Bean dwarf mosaic virus BV1 protein is a determinant of the hypersensitive
response and avirulence in Phaseolus vulgaris. Mol. Plant Microbe
Interactions, 13: 1184–1194.
Geering, A. D., N. E. Olszewski, G. Harper, B. E. Lockhart, R. Hull and J. E.
Thomas. 2005.
Gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell,
75: 855-862.
Gilbertson, R. L., M. Sudarshana, H. Jiang, M.R. Rojas and W. J. Lucas. 2003.
Limitations on geminivirus genome size imposed by plasmodesmata and
virus-encoded movement protein: Insights into DNA Trafficking. Plant Cell,
15: 2578–2591.

129
Gilmer, D., S. Bouzoubaa, A. Hehn, H. Guilley, K. Richards and G. Jonard. 1992.
Efficient cell-to-cell movement of beet necrotic yellow vein virus requires 3′
proximal genes located on RNA 2. Virology, 189: 40-47.
Gormez-Llorente, Y., R. J. Fletcher, X. S. Chen, J. M. Carazo and C. S. Martin. 2005.
Polymorphism and random hexamer structure in the archaeal
minichoromosome maitenance (MCM) helicase from Methanobacterium
thermoautotrophicum. J. Biol. Chem., 280: 40909-40915.
Gowen, S. R. 1995. Bananas and Plantains. Chapman & Hall, London, p.612.
Grigoras, I., T. Timchenko, L. Katul, A. Grande-Perez, H. J. Vetten and B.
Gronenborn, 2009. Reconstitution of authentic nanovirus from multiple cloned
DNAs. J. Virol., 83: 10778-10787.
Gronenborn, B. 2003. Nanovirus genome organization and protein function. Vet.
Microbiol., 98: 103-109.
Guerra-Peraza, O., D. Kirk, V. Seltzer, K. Veluthambi, A. C. Schmit, T. Hohn and E.
Herzog. 2005. Coat proteins of Rice tungro bacilliform virus and Mungbean
yellow mosaic virus contain multiple nuclear-localization signals and interact
with importin alpha. J. Gen. Virology, 86:1815–1826.
Guo, H. S. and S. W. Ding. 2002. A viral protein inhibits the long range signaling
activity of the gene silencing signal. EMBO J., 21: 398-407.
Guo, S. and K. J. Kempheus. 1995. Par-1, a gene required for establishing polarity
in C. elegans embryos, encodes a putative Ser/Thr kinase that is
asymmetrically distributed. Cell, 81: 611-620.
Gutierrez, C., 2000. DNA replication and cell cycle in plants: learning from
geminiviruses. EMBO J., 19: 792–799.
Haasnoot, P. C. J., D. Cupac and B. Berkhout. 2003. Inhibition of virus replication by
RNA interference. J. Biomed. Sci., 10: 607–616.
Hafner, G., M. Stafford, L. Wolter, R. M. Harding and J. L. Dale. 1997. Nicking and
joining activity of Banana bunchy top virus replication protein in vitro. J. Gen.
Virol., 78: 795-799.
Hafner, G., R. Dietzgen and J.L. Dale. 1993. Nucleotide sequence of one components
of the BBTV genome contains a putative replicase gene. J. Gen. Virol.,
74:323-328.

130
Hafner, G. J. 1998. Replication of Banana bunchy top virus. Ph.D Thesis, Queensland
University of Technology, Brisbane, Australia.
Haley, A., X. Zhan, K. Rivhardson, K. Head and B. Morris. 1992. Regulation of the
activities of African cassava mosaic virus promoters by the AC1, AC2 and
AC3 gene products. Virology, 188: 905-909.
Hamilton, A., O. Voinnet, L. Chappell and D. Baulcombe. 2002. Two classes of short
interfering RNA in RNA silencing. EMBO J., 21: 4671–4679.
Hamilton, A. J. and D. C. Baulcombe. 1999. A species of small antisense RNA in
posttranscriptional gene silencing in plants. Science, 286: 950-952.
Hammond, S., E. Bernstein, D. Beach and G. Hannon. 2000. An RNA-directed
nuclease mediates post-transcriptional gene silencing
in Drosophila cells. Nature, 404:293-298.
Hammond-Kosack, K. E. and Jones J. D. G. 1997. Plant disease resistance genes.
Annu. Rev. Plant Physiol., 48: 575–607.
Hanley-Bowdoin, L., S.B. Settlage, B.M. Orozco, S. Nagar and D. Robertson. 1999.
Geminiviruses: models for plant DNA replication, transcription, and cell cycle
regulation. Cri. Rev. Plant Sci., 18: 71–106.
Hanley-Bowdoin, L., S. B. Settlage, B. M. Orozco, S. Nagar and D. Robertson. 2000.
Geminiviruses: models for plant DNA replication, transcription, and cell cycle
regulation. Crit. Rev. Biochem. Mol. Biol., 35:105–140.
Harding, R. M., T. M. Burns and J. L. Dale. 1991. Virus-like particles associated with
banana bunchy top disease contain small single-stranded DNA. J. Gen. Virol.,
72: 225–230.
Harding, R. M., T. M. Burns, G. Hafner, R. Dietzgen and J. L. Dale. 1993. Nucleotide
sequence of one component of BBTV genome contains a putative replicase
gene. J. Gen. Virol., 74: 323-328.
Hehnle, S., C. Wege and H. Jeske. 2004. Interaction of DNA with the movement
proteins of geminiviruses revisited. J. Virol., 78: 7698–7706.
Heinlein, M. 2002. Plasmodesmata: dynamic regulation and role in macromolecular
cell-to-cell signalling. Curr. Opin. Plant Biol., 5: 543–552.
Hemenway, C., R. X. Fang, W. K. Kaniewski, N. H. Chua and N. E. Tumer. 1988.
Analysis of the mechanism of protection in transgenic plants expressing the
potato virus X coat protein or its antisense RNA. EMBO J., 7: 1273–1280.

131
Hermann, S. R., D. K. Becker, R. M. Harding and J. L. Dale. 2001b. Promoters
derived from Banana bunchy top virus-associated components S1 and S2 drive
transgene expression in both tobacco and banana. Plant Cell Rep., 20:642-646.
Hermann, S. R., R. M. Harding and J. L. Dale. 2001a. The banana actin 1 promoter
drives near-constitutive transgene expression in vegetative tissues of banana
(Musa spp.). Plant Cell Rep., 20: 525-530.
Herr, A. J., M. B. Jensen, T. Dalmay and D. C. Baulcombe. 2005. RNA polymerase
IV directs silencing of endogenous DNA. Science, 308: 118–120.
Heyraud-Nitschke, F., S. Shumacher, J. Laufs, S. Schaefer, J. Schell and B.
Gronenborn. 1995. Determination of the origin cleavage and joining domain
of geminivirus Rep proteins. Nucl. Acids Res., 23: 910–916.
Hickman, A. B. and F. Dyda. 2005. Binding and unwimding SF3 viral helicases. Curr.
Opin. Struc. Biol., 15: 77-85.
Hong, Y. and J. Stanley. 1995. Regulation of African cassava mosaic virus
complementary-sense gene expression by N-terminal sequences of the
replication associated protein AC1. J. Gen. Virol., 76: 2415-2422.
Hooks, C. R. R., R. Manandhar, E.P. Perez, K.H. Wang and R.P.P. Almeida. 2009.
Comaprative susceptibility of two banana cultivars to Banana bunchy top
virus under laboratory and field environment. J. Econ. Entomol., 102: 897-
904.
Horser, C. L., 2000. Charaterisation of the putative satellite DNAs associated with
Banana bunchy top virus. Ph.D Thesis. Queensland University of Technology,
Brisbane, Australia, pp.168.
Horser, C. L., M. Karan, R. M. Harding and J. L. Dale. 2001. Additional rep-encoding
DNAs associated with Banana bunchy top virus. Arch. Virol., 146:71–86.
Horser, C. L., R. M. Harding and J. L. Dale. 2001. Banana bunchy top nanovirus
DNA-1 encodes the ‘master’ replication initiation protein. J. Gen. Virol., 82:
459-464.
Hu, J. S., M. Wang, D. Sether, W. Xie and K. W. Leondhart. 1996. Use of
Polymerase chain reaction (PCR) to study transmission of Banana bunchy top
virus by banana aphid (Pentalonia nigronervosa). Ann. Appl. Biol., 128: 55-
64
Huisman, M. J., H. J. M. Linthorst, J. F. Bol and B. J. C. Cornelissen. 1988. The
complete nucleotide sequence of potato virus X and its homologies at the

132
amino acid level with various plus-stranded RNA viruses. J. Gen. Virol., 69:
1789-1798.
Hull, R. 2002. Matthew's Plant Virology, 4 ed, San Diego: Academic Press, p. 1001.
Hussain, M., S. Mansoor, S. Iram, A. N. Fatima and Y. Zafar. 2005. The nuclear
shuttle protein of Tomato leaf curl New Delhi virus is a pathogenicity
determinant. J. Virol., 79: 4434–4439.
Hwang, S. C. and H. J. Su. 1998. Production of Virus-free Banana Plantlets in
Taiwan. Taiwan Banana Research Institute, Chiuju, Pingtung.
Hyder, M .Z., S. H. Shah, S. Hameed and S. M. Naqvi. 2011. Evidence of
recombination in the BBTV genome. Infec. Gen. Evol. (In Press).
IKISAN. 2000. www.Ikisan.com
Jarret, R. L., W. Rodriguez and R. Fernandez. 1985. Evaluation, tissue culture
propagation and dissemination of ‘Saba’ and ‘Pelipita’ plantain in Costa Rica.
Hortic. Sci., 25:137–147.
Jefferson, R.A. 1987. Assaying chimeric genes in plants: The GUS gene fusion
system. Plant Mol. Biol. Rep., 5: 387-405.
Jones, D.R. 2000. Diseases caused by viruses. Diseases of banana, abaca and enset.
CABI Publishing, Wallingford, UK/New York, USA, pp.241-293.
Jordan, C. V, W. Shen, L. Hanley-Bowdoin and D. Robertson. 2007. Geminivirus-
induced gene silencing of the tobacco retinoblastoma-related gene results in
cell death and altered development. Plant Mol. Biol., 65: 163-175.
Jupin, I., F. D. Kouchkovsky, F. Jouanneau and B. Gronenborn. 1994. Movement of
Tomato yellow leaf curl geminivirus (TYLCV): Involvement of the protein
encoded by ORF C4. Virology, 204: 82–90.
Kaemmer, D., D. Fischer and R.L. Jarret. 1997. Molecular breeding in the genus
Musa: a strong case for STMS marker technology. Euphytica, 96: 49–63.
Kalinina, N. O., D.V. Rakitina, A. G. Solovyev, J. Schiemann and S. Y. Morozov.
2002. RNA helicase activity of the plant virus movement proteins encoded by
the first gene of the triple gene block. Virology, 296: 321-329.
Karan, M., R. M. Harding and J. L. Dale. 1994. Evidence for two groups of Banana
bunchy top virus isolates. J. Gen. Virol., 75: 3541-3546.

133
Karan, M., R. M. Harding and J. L. Dale. 1997. Association of Banana bunchy top
virus DNA components 2 to 6 with bunchy top disease. Mol. Plant. Pathol.,
On line http://www.bspp.org.uk/mppol/1997/0624 Karan.
Kasschau, K. D. and J. C. Carrington. 1998. Suppression of posttranscriptional gene
silencing. Cell, 95: 461-470.
Katul, L., E. Maiss and H. Vetten. 1995. Sequence analysis of faba bean necrotic
yellows virus DNA component containing a putative replicase gene. J. Gen.
Virol., 76:475-479.
Katul, L., E. Maiss, S.Y. Morozov and H. J. Vetten. 1997. Analysis of six DNA
components of the faba bean necrotic yellows virus genome and their
structural affinity to related plant virus genomes. Virology, 233: 247–259.
Katul, L., H. J. Vetten, E. Maiss, K. M. Makkouk, D. E. Lesemann and R. Casper.
1993. Characterization and serology of virus-like particles associated with
faba bean necrotic yellows virus. Ann. Appl. Biol., 123:629-647.
Katul, L., T. Timchenko, B. Gronenborn and H. J. Vetten. 1998. Ten distinct circular
ssDNA components, four of which encode putative replication-associated
proteins, are associated with the faba bean necrotic yellows virus genome. J.
Gen. Virol., 79: 3101–3109.
Khalid, S. and M. H. Soomro. 1993. Banana bunchy top disease in Pakistan. Plant
Pathol., 42: 923-926.
Khalid, S., M. H. Soomro and R. H. Stover. 1993. First report of banana bunchy top
virus in Pakistan. Plant Dis., 77:101.
Khanna, H., D. Becker, J. Kleidon and J. L. Dale. 2004. Centrifugation Assisted
Agrobacterium tumefaciens-mediated Transformation (CAAT) of
embryogenic cell suspensions of banana (Musa spp. Cavendish AAA and
Lady finger AAB). Mol. Breeding, 14: 239-252.
Kittelmann, K., P. Rau, B. Gronenborn and H. Jeske. 2009. Plant geminivirus Rep
protein induces re-replication in fission yeast. J. Virol., 83: 6769–6778.
Kohm, B. A., M. G. Goulden, J. E. Gilbert, T. A. Kavanagh and D. C. Baulcombe.
1993. A potato virus X resistance gene mediates an induced, nonspecific
resistance in protoplasts. Plant Cell, 5: 913-920.
Kong, L. and L. Hanley-Bowdoin. 2002. A geminivirus replication protein interacts
with a protein kinase and a motor protein that display different expression
patterns during plant development and infection. Plant Cell, 14: 1817–1832.

134
Kong, L .J., B. M. Orozco, J. L. Roe, S. Nagar, S. Ou, H. S. Feiler, T. Durfee, A. B.
Miller, W. Gruissem, D. Robertson and L. Hanley-Bowdoin. 2000. A
geminivirus replication protein interacts with the retinoblastoma protein
through a novel domain to determine symptoms and tissue specificity of
infection in plants. EMBO J., 19: 3485–3495.
Koonin, E.V. 1993. A common set of conserved motifs in a vast variety of putative
nucleic acid dependent ATPases including MCM proteins involved in the
initiation of eukaryotic DNA replication. Nucl. Acids Res., 21: 2541-2547.
Koonin, E. V. and T. V. Ilyina. 1992. Geminivirus replication proteins are related to
prokaryotic plasmid rolling circle DNA-replication initiator proteins. J. Gen.
Virol., 73: 2763–2766.
Kosky, R. G., M. Silva and L. P. Perez. 2002. Somatic embryogenesis of the banana
hybrid cultivar FHIA-18 (AAAB) in liquid medium and scaled-up in a
bioreactor. Plant Cell Tiss. Org. Cult., 68: 21–26.
Kumari, S. G and K. M. Makkouk. 2007. Virus diseases of faba bean (Vicia faba L.)
in Asia and Africa. Plant Viruses, 1: 93–105.
Kunik, T., K. Palanichelvam, H. Czosnek, V. Citovsky and Y. Gafni. 1998. Nuclear
import of the capsid protein of Tomato yellow leaf curl virus (TYLCV) in
plant and insect cells. Plant J., 13: 393–399.
Kutach, A. K. and J. T. Kadonaga. 2000. The downstream promoter element DPE
appears to be as widely used as the TATA box in Drosophila core promoters.
Mol. Cell. Biol., 20: 4754-4764.
Lacomme, C. and S. Chapman. 2008. Use of Potato virus X (PVX) based vector for
gene expression and virus-induced gene silencing (VIGS). Curr. Prot.
Microbiol., 8: 11-13.
Lageix, S., O. Catrice, J. M. Deragon, B. Gronenborn, T. Pelissier and B.C. Ramirez.
2007. The Nanovirus-encoded clink protein affects plant cell cycle regulation
through interaction with the retinoblastoma-related protein. J. Virol., 81: 4177-
4185.
Lakatos, L., G. Szittya, D. Silhavy and J. Burgyan. 2004. Molecular mechanism of
RNA silencing suppression mediated by p19 protein of tombus viruses.
EMBO J., 23:876-884.

135
Last, D. I., R. I. S. Brettell, D.A. Chamberlain, A. M. Chaudhury, P. J. Larkin, E. L.
Marsh, W. J. Peacock and E. S. Dennis. 1991. pEmu: an improved promoter
for gene expression in cereal cells. Theor. Appl. Genet., 81: 581-588.
Laufs, J., W. Traut, F. Heyraud, V. Matzeit, S.G. Rogers, J. Schell and B.
Gronenborn. 1995. In vitro cleavage and joining at the viral origin of
replication by the replication initiator protein of Tomato yellow leaf curl virus.
Proc. Nat. Acad. Sci. USA, 92: 3879–3883.
Lazarowitz, S. G. 1992. Geminiviruses: genome structure and gene function. Crit.
Rev. Plant Sci., 11: 327-349.
Lazarowitz, S. G. and R. N. Beachy. 1999. Viral movement proteins as probes for
intracellular and intercellular trafficking in plants. Plant Cell, 11: 535–548.
Lazarowitz, S. G., L. C. Wu, S. G. Rogers and J. S. Elmer. 1992. Sequence-specific
interaction with the viral AL1 protein identifies a geminivirus DNA
replication origin. Plant Cell, 4: 799–809.
Lee, R. C., R. L. Feinbaum and V. Ambros. 1993. The C. elegans heterochronic gene
lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell, 75:
843-854.
Lee, R. C. and V. Ambros. 2001. An extensive class of small RNAs in
Caenorhabditis elegans. Science, 294: 862-864.
Lee, T. I. and R. A. Young. 2000. Transcription of eukaryotic protein-coding genes.
Annu. Rev. Gen., 34: 77-137.
Li, G. F. and T.C. Hall. 1999. Footprinting in vivo reveals changing profiles of
multiple factor interactions with the β-phaseolin promoter during
embryogenesis. Plant J., 18: 633-641.
Lindbo, J. A., L. Silva-Rosales, W.M. Proebsting and W. G. Dougherty. 1993.
lnduction of a highly specific antiviral state in transgenic plants: implications
for regulation of gene expression and virus resistance. Plant Cell, 5: 1749-
1759.
Liu, D., S. L. Daubendiek, M. A. Zillman, K. Ryan and E.T. Kool. 1996. Rolling
circle DNA synthesis: Small circular oligonucleotides as efficient templates
for DNA polymerases. J. Amer. Chem. Soc. 118: 1587–1594.
Liu, H., M. I. Boulton, C.L. Thomas, D. A. Prior, K. J. Oparka and J. W. Davies.
1999. Maize streak virus coat protein is karyophyllic and facilitates nuclear
transport of viral DNA. Mol. Plant Microbe Interactions, 12: 894–900.

136
Lizardi, P.M., X. Huang, Z. Zhu, P. Bray-Ward, D .C. Thomas and D.C. Ward. 1998.
Mutation detection and single-molecule counting using isothermal rolling-
circle amplification. Nat. Genet., 19: 225–232.
Llave, C., K. Kasschau and J.C. Carrington. 2000. Virus-encoded suppressor of
posttranscriptional gene silencing targets a maintenance step in the silencing
pathway. Proc. Nat. Acad. Sci. USA, 97: 13401-13406.
Lockhart, B .E. L. 1990. Evidence for a double-stranded circular genome in a second
group of plant viruses. Phytopathology, 80:127-131.
Lough, T. J., N. E. Netzler, S. J. Emerson, P. Sutherland, F. Carr, D. L. Beck, W. J.
Lucas and R. L S. Forster. 2000. Cell-to-cell movement of potexviruses:
Evidence for a ribonucleoprotein complex involving the coat protein and first
triple gene block protein. Mol. Plant Microbe Interactions, 13: 962-974.
Luque, A., A. P. Sanz-Burgos, E. Ramirez-Parra, M. M. Castellano and C. Gutiérrez.
2002. Interaction of geminivirus Rep protein with replication factor C and its
potential role during geminivirus DNA replication. Virology, 302: 83–94.
Magnaye, L.V. and R.V. Valmayor. 1995. BBTV, CMV and other viruses affecting
banana in Asia and the Pacific. INIBAP Asia and the Pacific Network.
Makkouk, K. M. and A. Comeau. 1994. Evaluation of various methods for the
detection of barley yellow dwarf luteovirus by the tissue blot immunoassay
and its use for BYDV detection in cereals inoculated at different growth
stages. Eur. J. Plant Pathol., 100: 71–80.
Makkouk, K. M., S. G. Kumari, J. A. Hughes, V. Muniyappa and N. K. Kulkarni.
2003. Other legumes: Faba bean, chickpea, lentil, pigeon pea, mungbean,
black gram, lima bean, horse gram, bambara groundnut and winged bean. In:
Loebenstein G, Thottappilly G (eds), Virus and Virus-like Diseases of Major
Crops in Developing Countries, Dordrecht, the Netherlands, Kluwer Acad.
Pub., pp. 447–476.
Malik, R. S., V. Jumar, B. Bagewadi and S. K. Mukherjee. 2005. Interaction between
coat protien and replication initiation protein of Mung bean yellow mosaic
India virus might lead to control of viral DNA replication. Virology, 337: 273-
283.
Mansoor, S., S. H. Khan, A. Bashir, M. Saeed, Y. Zafar, K. A. Malik, R. Briddon, J.
Stanley and P. G. Markham. 1999. Identification of a novel circular single-

137
stranded DNA associated with cotton leaf curl disease in Pakistan. Virology,
259: 190–199.
Mariano, A. C., M. O. Andrade, A. A. Santos, S. M B. Carolino, M. L. Oliveira, M. C.
Baracat-Pereira, S. H. Brommonshenkel and E. P. B. Fontes. 2004.
Identification of a novel receptor-like protein kinase that interacts with a
geminivirus nuclear shuttle protein. Virology, 318: 24–31.
Marriott, H. and P. A. Lancaster. 1983. Banana and Plantains. In Hand book of
Tropical Foods. Edited by H. T. Chan. Marce Dekker, New York. pp. 85-143.
Matsomoto, K., A. D. Vilarinhos and S. Oka. 2002. Somatic hybridization by
electrofusion of banana protoplasts. Euphytica, 125: 317–324.
Matthew, R. E. F. 1991. Plant viruses. San Diego: Academic Press.
Matthews, R. E. F. 1979. Classification and nomenclature of viruses. Third Report of
the International Committee on Taxonomy of Viruses. Intervirology, 12: 132–
296.
May, G. D., R. Afza, H. S. Mason, A. Wiecko, F.J. Novak and C.J. Arntzen. 1995.
Generation of transgenic banana (Musa acuminata) plants via Agrobacterium-
mediated transformation. Biotechnology, 13: 486-492.
McGarry, R. C., Y. D. Barron, M. F. Carvalho, J. E. Hill, D. Gold, E. Cheung, W.L.
Kraus and S. G. Lazarowitz. 2003. A novel Arabidopsis acetyltransferase
interacts with the geminivirus movement protein NSP. Plant Cell, 15: 1605–
1618.
Mellor, J. 2006. Dynamic nucleosomes and gene transcription. Trends Genet., 22:
320-329.
Merits, A., O. N. Fedorkin, D. Guo, N. O. Kalinina and S. Y. Morozov. 2000.
Activities associated with putative replication initiation protein of Coconut
foliar decay virus; a tentative member of genus nanovirus. J. Gen. Virol., 82:3
099-3106.
Metz, T. D., R. Dixit and E. D. Earle. 1995. Agrobacterium tumefaciens-mediated
treansformation of broccoli (Brasicca oleracea var. italica) and cabbage
(Brassica oleracea var. capitata). Plant Cell Rep., 15: 287-292.
Miki, D. and K. Shimamoto. 2004. Simple RNAi vectors for stable and transient
suppression of gene function in Rice. Plant Cell Physiol., 45: 490-495.

138
Mizuno, T., M. Y. Chou and M. Inouye. 1984. A unique mechanism of regulating
gene expression: Translational inhibition by a complementary RNA transcript
(microRNA). Proc. Nat. Acad. Sci., 81: 1966-1970.
Mondal, T. K., A. Bhattacharya, P. S. Ahuja and P. K. Chand. 2001. Transgenic tea
[Camellia sinensis (L) O. Kuntze cv. Kangra Jat] plants obtained by
Agrobacterium-mediated transformation of somatic embryos. Plant Cell Rep.,
20: 712-720.
Morra, M. R. and I. T. D. Petty. 2000. Tissue specificity of geminivirus infections is
genetically determined. Plant Cell, 12: 2259–2270.
Msogoya, T. J. and B. Grout. 2008. Altered response to biotic and abiotic stress in
tissue culture-induced off-type plants of east African highland banana (Musa
AAA East Africa). J. Appl. Sci., 8:2703 – 2710.
Mubin, M., I. Amin, L. Amrao, R. W. Briddon and S. Mansoor. 2010. The
hypersensitive response induced by the V2 protein of a monopartite
begomovirus is countered by the C2 protein. Mol. Plant Pathol., 11: 245–254.
Murphy, F.A., C. M. Fauquet, D.H. L. Bishop, S. A. Ghabrial, A.W. Jarvis, G. P.
Martelli, A. Mayo and M .D. Summers. 1995. Virus Taxonomy: Sixth Report
of the International Committee on Taxonomy of Viruses.Wien, New York:
Springer-Verlag, pp. 585.
Nakamura, M., T. Tsunoda and J. Obokata. 2002. Photosynthesis nuclear genes
generally lack TATA-boxes: a tobacco photosystem I gene responds to light
through an initiator. Plant J., 29: 1-10.
Niu, S., B. Wang, X. Guo, J. Yu, X. Wang, K. Xu, Y. Zhai, J. Wang and Z. Liu. 2009.
Identification of two RNA silencing suppressors from Banana bunchy top
virus. Arch. Virol., 154: 1775-1783.
Nordstrom, K. and E. G. Wagner. 1994. Kinetic aspects of control of plasmid
replication by antisense RNA, Trends Biochem. Sci., 19: 294-300.
Noueiry, A. O., W. J. Lucas and R. L. Gilbertson. 1994. Two proteins of a plant DNA
virus coordinate nuclear and plasmodesmal transport. Cell, 76: 925–932.
Novak, F. J. 1992. Musa (Bananas and Plantains) In Biotechnology of Perennial Fruit
Crops. Edited by F.A. Hammerschlag and R.E. Litz. C.A.B. International:
Wallingford, Oxon, pp. 449-488.

139
Onodera, Y., J. R. Haag, T. Ream, P.C. Nunes, O. Pontes and C. S. Pikaard. 2005.
Plant nuclear RNA polymerase IV mediates siRNA and DNA-methylation
dependent heterochromtin formation. Cell, 120: 613–622.
Orozco, B. M., A. B. Miller, S. B. Settlage and L. Hanley-Bowdoin 1997. Functional
domains of geminivirus replication protein; J. Biol. Chem., 272: 9840–9846.
Orozco, B. M., L. J. Kong, L. A. Batts, S. Elledge and L. Hanley-Bowdoin. 2000. The
multifunctional character of a geminivirus replication protein is reflected by its
complex oligomerization properties. J. Biol. Chem., 275: 6114–6122.
Ozias-Akins, P. and I. K. Vasil. 1982. Plant regeneration from immature embryos and
inflorescences of Triticum aestivum (wheat): evidence for somatic
embryogenesis. Protoplasma, 110: 95-105.
Pakistan Agriculture Research Council (PARC). 2005. Current crop production
Statistic, Year Book, Pakistan Agriculture Research Council, Government of
Pakistan.
Parker, A. J., K. C. Parker and H. Wiggins-Brown. 2000. Disturbance and scale
effects on southern old-growth forests (USA): The sand pine example, Nat.
Areas J., 20: 273-279.
Pascal, E., A. A. Sanderfoot, B. M. Ward, R. Medville, R. Turgeon and S.G.
Lazarowitz. 1994. The geminivirus BR1 movement protein binds single-
stranded DNA and localizes to the cell nucleus. Plant Cell, 6:995–1006.
Pascal, E., P. E. Goodlove, L. C. Wu and S. G. Lazarowitz. 1993. Transgenic tobacco
plants expressing the geminivirus BL1 protein exhibit symptoms of viral
disease. Plant Cell, 5: 795–807.
Peragine, A., M. Yoshikawa, G. Wu, H. L. Albrecht and R. S. Poethig. 2004. SGS3
and SGS2/SDE1/RDR6 are required for juvenile development and the
production of trans-acting siRNAs in Arabidopsis. Genes Dev., 18: 2368–
2379.
Perez Hernandez, J. B., S. Remy, R. Swennen and L. Sagi. 2006b. Banana (Musa sp.).
In: Methods in Molecular Biology, 344: Agrobacterium Protocols, volume 2.
K. Wang, editor. Humana Press Inc., Totowa, NJ, USA, pp. 167-175
Perez Hernandez, J. B., S. Remy, V. Galan Sauco, R. Swennen and L. Sagi. 1998.
Chemotaxis, attachment and transgene expression in the Agrobacterium-
mediated banana transformation system. Mededelingen Faculteit
Landbouwwetenschappen Universiteit Gent., 63: 1603-1606.

140
Perez-Hernandez, J. B., R. Swennen and L. Sagi. 2006a. Number and accuracy of T-
DNA insertions in transgenic banana (Musa spp.) plants characterized by an
improved anchored PCR technique. Trans. Res., 15:139-150.
Petty, I.. T. and A. O. Jackson. 1990. Mutational analysis of Barley stripe mosaic
virus RNA. Virology, 179:712-718.
Pink, D. A. 2002. Strategies using genes for nondurable disease resistance. Euphytica,
124:227–236.
Ploetz, R. C., J. E. Thomas and W.R. Slabaugh. 2003. Diseases of banana and
Plantain. In Diseases of Tropical fruit crops. Edited by R.C. Ploetz, C.A.B.
International Wallingford, Oxon, pp. 73-134
Powell-Abel, P., R. Nelson, B. De, N. Hoffman, S. Rogers, R. Fraley and R. Beachy.
1986. Delay in disease development in transgenic plants that express the
tobacco mosaic virus coat protein gene. Science, 232: 738-743.
Praz, V., R. Perier, C. Bonnard and P. Bucher. 2002. The Eukaryotic Promoter
Database, EPD: new entry types and links to gene expression data. Nucl.
Acids Res., 30: 322-324.
Pruss, P., X. Ge, X. M. Shi, J. C. Carrington and V. B. Vance. 1997. Plant viral
synergism: the potyviral genome encodes a broad-range pathogenicity
enhancer that transactivates replication of heterologous viruses. Plant Cell, 9:
859-868.
Qazi, J., M. Ilyas, S. Mansoor and R. W. Briddon. 2007. Legume yellow mosaic
viruses: genetically isolated begomoviruses. Mol. Plant Pathol., 8: 343-348.
Raghavan, V., P. S. Malik, N. R. Choudhury and S. K. Mukherjee. 2004. The DNA A
component of a plant geminivirus (Indian mung bean yellow mosaic virus)
replicates in budding yeast cells. J. Virol., 78: 2405–2413.
Ram, R. D. and A. S. Summanwar. 1984. Colocasia esculenta (L) Schott. A reservoir
of banana bunchy top disease of banana. Curr. Sci., 53: 145-146.
Ramage, C. M., A. M. Borda, S. D. Hamill and M. K. Smith. 2004. A simplified PCR
test for early detection of dwarf off-types in micropropagated Cavendish
bananas (Musa spp. AAA). Sci. Hortic., 103: 145–151.
Randles, J. W., P. W. G. Chu, J. L. Dale, R. M. Harding, J. Hu, M. Kojima, K. M.
Makkouk, Y. Sano, J.E. Thomas and H.J. Vetten. 2000. Genus Nanovirus. In:
Van Regenmortel, M. H. V., C.M. Fauquet and D. H. L. Bishop, Virus

141
Taxonomy, Seventh Report of the International Committee on Taxonomy of
Viruses. Academic Press, San Diego, London, pp. 303–309.
Remy, S., A. Buyens, B. P. A. Cammue, R. Swennen and L. Sagi. 1998b. Production
of transgenic banana plants expressing antifungal proteins. Acta Hortic., 490:
433-436.
Remy, S., E. Thiry, B. Coemans, S. Windelinckx, R. Swennen and L. Sagi. 2005.
Improved T-DNA vector for tagging plant promoters via high-throughput
luciferase screening. Biotechnology, 38: 763-770.
Remy, S., I. François, B. P. A. Cammue, R. Swennen and L. Sagi. 1998a. Co-
transformation as a potential tool to create multiple and durable resistance in
banana (Musa spp.). Acta Hortic., 461: 361-365.
Resmi, L. and A. S. Nair. 2011. Differential effects of cytokinins in the
micropropagation of diploid and triploid Musa cultivars. Int. J. Integr. Biol.,
11: 35-38.
Richter, T. E. and P. C. Ronald. 2000. The evolution of disease resistance genes. Plant
Mol. Biol., 42: 195–204.
Rigden, J .E., I. B. Dry, P. M. Mullineaux and M. A. Rezaian. 1993. Mutagenesis of
the virion-sense open reading frames of Tomato leaf curl virus. Virology, 193:
1001–1005.
Robinson, J. C. 1996. Banana and Plantains, Crop production in Horticulture, CAB
International, Cambridge Printing Press.
Rohde, W., D. Becker and J. W. Randles. 1995. The promoter of Coconut foliar decay
virus associated circular single-stranded DNA directs phloem-specific reporter
gene expression in transgenic tobacco. Plant Mol. Biol., 27: 623-628.
Rohde, W., J. W. Randles, P. Langridge and D. Hanold. 1990. Nucleotide sequence of
a circular single-stranded DNA associated with coconut foliar decay virus.
Virology, 176: 648–651.
Rojas, M. R., A. O. Noueiry, W. J. Lucas and R. L. Gilbertson. 1998. Bean dwarf
mosaic geminivirus movement proteins recognize DNA in a form and size
specific manner. Cell, 95: 105–113.
Rojas, M. R., H. Jiang, R. Salati, B. Xoconostle-Cazares, M. R. Sudarshana, W. J.
Lucas and R. L. Gilbertson. 2001. Functional analysis of proteins involved in
movement of the monopartite begomovirus, Tomato yellow leaf curl virus.
Virology, 291: 110–125.

142
Romano, N. and G. Macino. 1992. Quelling: Transient inactivation of gene expression
in Neurospora crassa by transformation with homologous sequences. Mol.
Microbiol., 6: 3343-3353.
Rouppe van der Voort J., P. Wolters, R. Folkertsma, R. Hutten, P. van Zandvoort, H.
Vinke, K. Kanyuka and A. Bendahmane. 1997. Mapping of the cyst nematode
resistance locus Gpa2 in potato using a strategy based on comigrating AFLP
markers. Theor. Appl. Genet., 95: 874–880.
Ruiz, T. M., O. Voinnet and D. C. Baulcombe. 1998. Initiation and maintenance of
virus-induced gene silencing. Plant Cell, 10: 937-946.
Sagi L., B. Panis, S. Remy, H. Schoofs, K. Desmet, R. Swennen and B.P.A. Cammue.
1995a. Genetic transformation of banana and plantain (Musa spp.) via particle
bombardment. Biotechnology, 13: 481-485.
Sagi L., S. Remy, B. Panis, G. Volckaert and R. Swennen. 1992. Transient gene
expression in banana protoplasts. Banana Newslet., 5: 42-47.
Sagi, L., B. Panis, S. Remy, B. P. A. Cammue and R. Swennen. 1995c. Stable and
transient genetic transformation of banana (Musa spp. cv. ‘Bluggoe’, ABB-
group) using cells and protoplasts. In: Proceedings of the 11th ACORBAT
Meeting. V. Morales Soto, editor. San José, Costa Rica, 13-18 February 1994,
pp. 85-102.
Sagi, L., S. Remy, B. Panis, R. Swennen and G. Volckaert. 1994. Transient gene
expression in electroporated banana (Musa spp., cv. Bluggoe, ABB group)
protoplasts isolated from regenerable embryogenetic cell suspensions. Plant
Cell Rep., 13: 262-266.
Sagi, L., S. Remy, B. Verelst, B. Panis, B. P. A. Cammue, G. Volckaert and R.
Swennen. 1995b. Transient gene expression in transformed banana (Musa cv.
‘Bluggoe’) protoplasts and embryogenic cell suspensions. Euphytica, 85: 89-
95.
Sanderfoot, A. A. and S. G. Lazarowitz. 1995. Cooperation in viral movement: the
geminivirus BL1 movement protein interacts with BR1 and redirects it from
the nucleus to the periphery. Plant Cell, 7: 1185–1194.
Sanderfoot, A. A., D. J. Ingham and S. C. Lazarowitz. 1996. A viral movement protein
as a nuclear shuttle. Plant Physiol., 11: 22–33.

143
Sano, Y., M. Wada, Y. Hashimoto, T. Matsumoto and M. Kojima. 1998. Sequences of
ten circular ssDNA components associated with the milk vetch dwarf virus
genome. J. Gen. Virol., 79: 3111-3118.
Santos, E., S. Remy, E. Thiry, S. Windelinckx, R. Swennen and L. Sagi. 2007.
Tagging novel promoters in banana using the luciferase rerporter gene. Acta
Hortic., 763: 99-107.
Sauders, K. and J. Stanley. 1999. A nanovirus like DNA component associated with
yellow vein disease of Ageratum conyzoides: evidence for interfamilial
recombination between plant DNA viruses. Virology, 264:142-152.
Saunders, K., I. D. Bedford, R.W. Briddon, P. G. Markham, S. M. Wong and J.
Stanley. 2000. A unique virus complex causes Ageratum yellow vein disease.
Proc. Nat. Acad. Sci. USA, 97: 6890–6895.
Schenk, P. M., L. Sagi, T. Remans, R. G. Dietzgen, M. J. Bernard, M. W. Graham and
J. M. Manners. 1999. A promoter from sugarcane bacilliform badnavirus
drives transgene expression in banana and other monocot and dicot plants.
Plant Mol. Biol., 39: 1221-1230.
Schenk, P. M., T. Remans, L. Sagi, A. R. Elliott, R. G. Dietzgen, R. Swennen, P.R.
Ebert, C. P. L. Grof and J. M. Manners. 2001. Promoters for pregenomic RNA
of banana streak badnavirus are active for transgene expression in monocot
and dicot plants. Plant Mol. Biol., 47:3 99-412.
Scholthof, K. B. G. 2004. Tobacco Mosaic Virus: A Model System for Plant Biology.
Annu. Rev. Phytopathol., 42: 13-34.
Schunmann, P. H. D., A. E. Richardson, C. E. Vickers and E. Delhaize. 2004.
Promoter analysis of the barley Pht1 phosphate transporter gene identifies
regions controlling root expression and responsiveness to phosphate
deprivation. Plant Physiol., 136: 4205-4214.
Sears, R. G. and E. L. Deckard. 1982. Tissue culture variability in wheat: callus
induction and plant regeneration. Crop Sci., 22: 546-550.
Segers, G. C., X. Zhang, F. Deng, Q. Sun and D.L. Nuss. 2007. Evidence that RNA
silencing functions as an antiviral defense mechanism in fungi. Proc. Nat.
Acad. Sci. USA, 104: 12902–12906.
Selth, L. A., J. W. Randles and M. A. Rezaian. 2004. Host responses to transient
expression of individual genes encoded by Tomato leaf curl virus. Mol. Plant
Microbe Interactions, 17:2 7-33.

144
Shahmuradov, I. A., A. J. Gammerman, J. M. Hancock, P. M. Bramley and V.V.
Solovyev. 2003. Plant Prom: a database of plant promoter sequences. Nucl.
Acids Res., 31: 114-117.
Shahmuradov, I. A., V. V. Solovyev and A. J. Gammerman. 2005. Plant promoter
prediction with confidence estimation. Nucl. Acids Res., 33: 1069-1076.
Sharman, M., J. E. Thomas, S. Skabo and T. A. Holton. 2008. Abacá bunchy top
virus, a new member of the genus Babuvirus (family Nanoviridae). Arch.
Virol., 153: 135-147.
Shepherd, D. N., D. P. Martin, D. R. McGivern, M.I. Boulton, J.A. Thomas and E.P.
Rybicki. 2005. A three nucleotiude mutation altering the Maize streak virus
Rep pBRB interaction motif reduces symptom severity in maize and partially
reverts at high frequency without restoring pRBR-Rep binding. J. Gen. Virol.,
86:803-813.
Shukla, V. D. and A. Anjaneyulu. 1981. Plant spacing to reduce rice tungro incidence.
Plant Dis., 65: 584-586.
Silhavy, D., A. Molnar, A. Lucioli, G. Szittya, C. Hornyik, M. Tavazza and J.
Burgyan. 2002. A viral protein suppresses RNA silencing and binds silencing-
generated, 21- to 25-nucleotide double-stranded RNAs. EMBO J., 21: 3070-
3080.
Smale, S. T. 1997. Transcription initiation from TATA-less promoters within
eukaryotic protein-coding genes. Biochimica et Biophysica Acta-Gene
Structure and Expression, 1351: 73-88.
Smale, S. T. and J. T. Kadonaga. 2003. The RNA polymerase II core promoter. Annu.
Rev. Biochem., 72: 449-479.
Solovyev, A. G., E. I. Savenkov, A. A. Agranovsky and S. Y. Morozov. 1996.
Comparisons of the genomic cis-elements and coding regions in RNA β
components of the shordeiviruses: barley stripe mosaic virus, lychnis ringspot
virus and poa semilatent virus. Virology, 219: 9-18.
Soosaar, J. L., T. M. Burch-Smith and S. P. Dinesh-Kumar. 2005. Mechanisms of
plant resistance to viruses. Nat. Rev. Microbiol., 3: 789-798.
Srivastava, A., S. K. Raj, Q. M. Haq, K. M. Srivastava, B. P. Singh and P. V. Sane.
1995. Association of Cucumber mosaic virus strain with mosaic disease of
banana Musa paradisiaca-an evidence using immuno/nucleic acid probe. Ind.
J. Exp. Biol., 33: 986-988.

145
Stover, R. H. and A. Simmonds. 1987. Bananas 3rd edition, Longman Scientific and
Technical, Harlow, UK.
Struhl, K. 2001. Gene regulation - A paradigm for precision. Science, 293: 1054-
1055.
Su, H. ., R. Y. Wu and L.Y. Tsao. 1993. Ecology of Banana bunchy top virus disease,
in Proceedings of International Symopsium on Recent Developments in
banana cultivation technology, edited by Valmayor, R.V, S.C. Hwang, R.
Ploetz, S.C. Lee, N.V. Rao, Taiwan Banana Research Institute, 14-18
December 1992, Chiuju, Pingtung, Taiwan, INIBAP/ASPNET, Los Banos,
the Phillipines, pp. 308-312.
Sudarshana, M. R., H. L. Wang, W. J. Lucas and R. L. Gilbertson. 1998. Dynamics of
bean dwarf mosaic geminivirus cell-to-cell and long-distance movement in
Phaseolus vulgaris revealed, using the green fluorescent protein. Mol. Plant
Microbe Interactions, 11: 277–291.
Sunter, G. and D. M. Bisaro. 1992. Transactivation of geminivirus AR1 and BR1
gene expression by the viral AL2 gene product occurs at the level of
transcription. Plant Cell, 4: 1321–1331.
Sunter, G. and D. M. Bisaro. 1997. Regulation of a geminivirus coat protein promoter
by AL2 protein (TrAP): Evidence for activation and derepression mechanisms.
Virology, 232: 269–280.
Surin, B., P. Boevink, P. Keese, P. Chu, P. Larkin and P. Waterhouse. 1996.
Expression patterns of novel plant-active promoters isolated from the genome
of subterranean clover stunt virus. Aust. Society of Biochemistry and
Molecular Biology and Aust. Society of Plant Physiology Combined
Conference, SYM-21-03, Canberra, Australia.
Suzuki, S., K. Supaibulwatana, M. Mii and M. Mii and M. Nakano. 2001. Production
of transgenic plants of the Liliaceous ornamental plant Agapanthus praecox
ssp. Orientalis (Leighton) Leighton via Agrobacterium-mediated
transformation of embryogenic calli. Plant Sci., 161: 89-97.
Swennen, R., G. Arinaitwe, B.P.A. Cammue, I. François, B. Panis. S. Remy, L. Sagi,
E. Santos, H. Strosse and I. Van den houwe. 2003. Transgenic approaches for
resistance to Mycosphaerella leaf spot diseases in Musa spp. L. Jacome, P.
Lepoivre, D. Marin, R. Ortiz, R. Romero and J.V. Escalant, editors.
Mycosphaerella leaf spot diseases of bananas: present status and outlook.

146
Proceedings of the 2nd International workshop on Mycosphaerella leaf spot
diseases of bananas. San José, Costa Rica, 20-23 May 2002. INIBAP,
Montpellier, France, pp.209-238.
Taghavi, M., C. Hayward, L. I. Sly and M. Fegan. 1996. Analysis of the phylogenetic
relationships of strains of Burkholderia solanacearum, Pseudomonas syzygii
and the blood disease bacterium of banana based on 16s rRNA gene
sequences. Int. J. Syst. Bacteriol., 46: 10-15.
Takasaki, T., K. Hatakeyama, K. Ojima, M. Watanabe, K. Toriyama and K. Hinata.
1997. Factors influencing Agrobacterium-mediated transformation of Brassica
rapa L. Breed Sci., 47: 127-134.
Tenllado, F. and J. R. D. Ruiz. 2001. Double-stranded RNA-mediated interference
with plant virus infection. J. Virol., 75: 12288-12297.
Tenllado, F., C. Llave, J. Ramon and D. Ruiz. 2004. RNA interference as a new
biotechnological tool for the control of virus diseases in plants. Virus Res.,
102: 85–96
Terada, R. and K. Shimamoto. 1990. Expression of CaMV35S-GUS gene in
transgenic rice plants. Mol. Gen., 220: 389–392.
Teycheney. D. Y., A. Marais, L. Svanell-Dumas, M. J. Dulucq and T. Candresse.
2005. Molecular characterization of Banana strain X (BVX), a novel member
of the Flexiviridae family. Arch. Virol., 150: 1715-1722.
Thomas J. E., and I. Caruana. 2000. Bunchy Top. In DR Jones, ed. Diseases of
banana, abaca and enset CABI Publishing, Oxon, UK, pp. 241-253
Thresh, J. M. 2006. Crop viruses and virus diseases: A global perspective. Virus
diseases and Crop, pp.9-32.
Timchenko, T., L. Katul, M. Aronson, J. C. Vega-Arreguin, B.C. Ramirez, H. J.
Vetten, and B. Gronenborn. 2006. Infectivity of nanovirus DNAs: induction of
disease by cloned genome components of Faba bean necrotic yellows virus. J.
Gen. Virol., 87: 1735-1743.
Timchenko, T., L. Katul, Y. Sano, F.D. Kouchovsky, H. Vetten and B. Gornenborn.
2000. The master Rep concept in nanovirus replication: identification of
missing genome components and potential for natural genetic reassortment.
Virology, 274: 189-195.

147
Toth, R. L., S. Chapman, F. Carr and S. Santa Cruz. 2001. A novel strategy for the
expression of foreign genes from plant virus vectors. FEBS Letters, 489: 215-
219.
Tsukazki, H., Y. Kuginuki, R. Aida and T. Suzuki. 2002. Agrobacterium-mediated
transformation of a double haploid line of cabbage. Plant Cell Rep., 21: 257-
262.
Tuschl, T., P. D. Zamore, R. Lehmann, D. P. Bartel and P.A. Sharp. 1999. Targeted
mRNA degradation by double-stranded RNA in vitro. Genes Dev., 13: 3191-
1397.
Unseld, S., M. Hohnle, M. Ringel and T. Frischmuth. 2001. Subcellular targeting of
the coat protein of African cassava mosaic geminivirus. Virology, 286: 373–
383.
Vance, V. B. and H. Vaucheret. 2001. RNA silencing in plants: defense and counter
defense. Science, 292: 2277–2280.
Vargason, J. M., G. Szittya, J. Burgyan and T. M. Hall. 2003. Size selective
recognition of siRNA by an RNA silencing suppressor. Cell, 115: 799-811.
Vazquez, F., H. Vaucheret, R. Rajagopalan, C. Lepers, V. Gasciolli, A. C. Mallory, J.
L. Hilbert, D.P. Bartel and P. Crete. 2004. Endogenous trans-acting siRNAs
regulate the accumulation of Arabidopsis mRNAs. Mol. Cell, 16: 69–79.
Venter, M. and F.C. Botha. 2004. Promoter analysis and transcription profiling:
Integration of genetic data enhances understanding of gene expression.
Physiol. Plant., 120: 74-83.
Voinnet, O. 2001. RNA silencing as a plant immune system against viruses. Trends
Genet., 17: 449–459.
Voinnet, O., Y. M. Pinto and D. C. Baulcombe. 1999. Suppression of gene silencing:
a general strategy used by diverse DNA and RNA viruses. Proc. Nat. Acad.
Sci., 96:1 4147–14152.
von Arnim, A. and J. Stanley. 1992. Determinants of Tomato golden mosaic
virus symptom development located on DNA B. Virology, 186: 286–293.
Vuylsteke, D. 1989. Shoot-tip culture for the propagation, conservation and exchange
of Musa germplasm. practical manuals for handling crop germplasm in vitro.
International Board for Plant Genetic Resources, Rome.

148
Wang, H., K. J. Buckley, X. Yang, R.C. Buchmann and D. M. Bisaro. 2005.
Adenosine kinase inhibition and suppression of RNA silencing by geminivirus
AL2 and L2 proteins. J. Virol., 79: 7410–7418.
Wanitchakorn, R., G. J. Hafner, R. M. Harding and J. L. Dale. 2000. Functional
analysis of proteins encoded by Banana bunchy top virus DNA4 to 6. J. Gen.
Virol., 81: 299-306.
Wanitchakorn, R., R. M. Harding and J. L. Dale. 1997. Banana bunchy top virus
DNA-3 encodes the viral coat protein. Arch. Virol., 142: 1673-1680.
Watanabe, T., M. Sudoh, M. Miyagishi, H. Akashi, M. Arai, K. Inuoe, K. Taira, M.
Yoshiba and M. Kohara. 2006. Intracellular-diced dsRNA has enhanced
efficacy for silencing HCV RNA and overcomes variation in the viral
genotype. Genet. Theor., 13: 883-892.
Waterhouse, P. M., M. B. Wang and T. Lough. 2001. Gene silencing as an adaptive
defense against viruses. Nature, 411: 834–842.
Waterhouse, P. M., M. W. Graham and M.B. Wang. 1998. Virus resistance and gene
silencing in plants can be induced by simultaneous expression of sense and
antisense RNA. Proc. Nat. Acad. Sci., 951: 3959-13964.
Wellings, P. W., P. H. Hart, V. Kami and D. C. Morneau. 1994. The introduction and
establishment of Aphidius colemani Viereck (Hym., Aphidiinae) in Tonga. J.
Appl. Ent., 118: 419-428.
Wesley, S. V., C. Helliwell, N. A. Smith, M. B. Wang, D. Rouse, Q. Liu, P. Gooding,
S. Singh, D. Abbott, P. Stoutjesdijk, S. Robinson, A. Gleave, A. Green and
P.M. Waterhouse. 2001. Constructs for Efficient, Effective and High
Throughput Gene Silencing in Plants. Plant J., 27: 581-590.
Wezel, R. V., X. Dong, P. Blake, J. Stanley and Y. Hong. 2002. Differential roles of
geminivirus Rep and AC4 in the induction of necrosis in N. benthamiana.
Mol. Plant Pathol., 3: 461-471.
Whitham, S., S. McCormick and B. Baker. 1996. The N gene of tobacco confers
resistance to tobacco mosaic virus in transgenic tomato. Proc. Nat Acad. Sci.,
93: 8776–878.
Whitham, S., S. P. Dinesh-Kumar, D. Choi, R. Hehl, C. Corr and B. Baker. 1994. The
product of the tobacco mosaic virus resistance gene N: Similarity to Toll and
the interleukin-1 receptor. Cell, 78: 1101-1115.

149
Wicker, E., L. Grassart, R. Coranson-Beaudu, I. D. Mian, C. Guilbard, M. Fegan and
P. Prior. 2007. Ralstonia solanoecearum strains from Martinique (French
West Indies) exhibiting a new pathogenic potential. Appl. Environ. Microbiol.,
73: 6790-6801.
Wightman, B., I. Ha and G. Ruvkun. 1993. Posttranscriptional regulation of the
heterochronic gene lin-14 by lin- 4 mediates temporal pattern formation in C.
elegans. Cell, 75: 855-862.
Williams, L., C. C. Carles, K. S. Osmont and J. C. Fletcher. 2005. A novel approach
identifies an endogenous trans-acting siRNA that targets the Arabidopsis
ARF2, ARF3/ETT and ARF4 genes. Proc. Nat. Acad. Sci. USA, 102: 9703–
9708.
Wu, C. H., L. Madabusi, H. Nishioka, P. Emanuel, M. Sypes, I. Arkhipova, D. S.
Gilmour. 2001. Analysis of core promoter sequences located downstream
from the TATA element in the hsp70 promoter from Drosophila
melanogaster. Mol. Cell. Biol., 21: 1593-1602.
Wu, H., C. Sparks, B. Amoah and H. D. Jones. 2003. Factors influencing successful
Agrobacterium-mediated transformation of wheat. Plant Cell Rep., 21: 659-
668.
Wu, R. Y. and H. J. Su. 1992. Detection of Banana bunchy top virus in diseased and
symptomless banana plants with monoclonal antibody. Trop. Agric.
(Trinidad), 69: 397-399.
Wu, R.Y., L. R. You and T. S. Soong. 1994. Nucleotide sequences of two circular
single-stranded DNAs associated with Banana bunchy top virus.
Phytopathology, 84: 952-956.
Yamamoto Y. Y. and J. Obokata. 2007c. ppdb: a plant promoter database. Nucl.
Acids Res., 36: 977-981.
Yamamoto Y. Y., H. Ichida, M. Matsui, J. Obokata, T. Sakurai, M. Satou, M. Seki, K.
Shinozaki and T. Abe. 2007a. Identification of plant promoter constituents by
analysis of local distribution of short sequences. BMC Genomics, 8: 7-72.
Yamamoto, Y. Y., H. Ichida, T. Abe, Y. Suzuki, S. Sugano and J. Obokata. 2007b
Differentiation of core promoter architecture between plants and mammals
revealed by LDSS analysis. Nucl. Acids Res., 35: 6219-6226.
Yang, I. C, J. P. Iommarini, D. K. Becker, G. J. Hafner, J. L. Dale and R. M. Harding.
2003. A promoter derived from taro bacilliform badnavirus drives strong

150
expression in transgenic banana and tobacco plants. Plant Cell Rep., 21: 1199-
1206.
Yeh, H. H., H. J. Su and Y. C. Chao. 1994. Genome characterization and
identification of viral associated dsDNA component of Banana bunchy top
virus. Virology, 198: 645-652.
Yu, B., Z. Yang, J. Li, S. Minakhina, M. Yang, R.W. Padgett, R. Steward and X.
Chen. 2005. Methylation as a crucial step in plant microRNA biogenesis.
Science, 307: 932–935.
Yu, H., S. H. Yang and C. J. Goh. 2001. Agrobacterium-mediated transformation of a
Dendrobium orchid with the class 1 knox gene DOH1. Plant Cell Rep., 20:
301-305.
Zaitlin, M. and P. Palukaitis. 2000. Advances in understanding plant viruses and virus
diseases. Ann. Rev. Phytopathol., 38: 117-143.
Zamore, P. D., T. Tuschl, P. A. Sharp and D. P. Bartel. 2000. RNAi: Double-stranded
RNA directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide
intervals. Cell, 101: 25-33.
Zhan, X., K. A. Richardson, A. Haley and B. A. M. Morris. 1993. The activity of the
coat protein promoter of chloris striate mosaic virus is enhanced by its own
and C1-C2 gene products. Virology, 193:498-502.
Zhang, H. S. and D. C. Dean. 2001. Rb-mediated chromatin structure regulation and
transcriptional repression. Oncogene, 20: 3134–3138.
Zhang, S. C., R. Ghosh and H. Jeske. 2002. Subcellular targeting domains of Abutilon
mosaic geminivirus movement protein BC1. Arch. Virol., 147: 2349–2363.
Zhang, W., N. H. Olson, T.S. Baker, L. Faulkner, M. Agbandje-McKenna, M.I.
Boulton, J. W. Davies and R.M. Kenna. 2001. Structure of the Maize streak
virus geminate particle. Virology, 279: 471– 477.
Ziegler-Graff, V., P. J. Guilford and D. C. Baulcombe. 1991. Tobacco rattle virus
RNA-1 29K gene product potentiates viral movement and also affects
symptom induction in tobacco. Virology, 182:145-155.
Zilberman, D., X. Cao and S. E. Jacobsen. 2003. Argonuate for control of locus
specific siRNA accumulation and DNA and histone methylation. Science,
299: 716–719.


















