
Identification of Immunodominant Epitopes of -Gliadin in
10
of April 3, 2019. This information is current as following Oral Immunization -Gliadin in HLA-DQ8 Transgenic Mice α of Identification of Immunodominant Epitopes Rossi Troncone, Salvatore Auricchio, Rosa A. Siciliano and Mauro Marcello Gaita, Olga Fierro, Chella S. David, Riccardo Stefania Senger, Francesco Maurano, Maria F. Mazzeo, http://www.jimmunol.org/content/175/12/8087 doi: 10.4049/jimmunol.175.12.8087 2005; 175:8087-8095; ; J Immunol References http://www.jimmunol.org/content/175/12/8087.full#ref-list-1 , 20 of which you can access for free at: cites 55 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2005 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on April 3, 2019 http://www.jimmunol.org/ Downloaded from by guest on April 3, 2019 http://www.jimmunol.org/ Downloaded from
Transcript of Identification of Immunodominant Epitopes of -Gliadin in

of April 3, 2019.This information is current as
following Oral Immunization-Gliadin in HLA-DQ8 Transgenic Miceαof
Identification of Immunodominant Epitopes
RossiTroncone, Salvatore Auricchio, Rosa A. Siciliano and MauroMarcello Gaita, Olga Fierro, Chella S. David, Riccardo Stefania Senger, Francesco Maurano, Maria F. Mazzeo,
http://www.jimmunol.org/content/175/12/8087doi: 10.4049/jimmunol.175.12.8087
2005; 175:8087-8095; ;J Immunol
Referenceshttp://www.jimmunol.org/content/175/12/8087.full#ref-list-1
, 20 of which you can access for free at: cites 55 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2005 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

Identification of Immunodominant Epitopes of �-Gliadin inHLA-DQ8 Transgenic Mice following Oral Immunization1
Stefania Senger,* Francesco Maurano,* Maria F. Mazzeo,* Marcello Gaita,* Olga Fierro,*Chella S. David,† Riccardo Troncone,‡ Salvatore Auricchio,‡ Rosa A. Siciliano,* andMauro Rossi2*
Celiac disease, triggered by wheat gliadin and related prolamins from barley and rye, is characterized by a strong association withHLA-DQ2 and HLA-DQ8 genes. Gliadin is a mixture of many proteins that makes difficult the identification of major immuno-dominant epitopes. To address this issue, we expressed in Escherichia coli a recombinant �-gliadin (r-�-gliadin) showing the mostconserved sequence among the fraction of �-gliadins. HLA-DQ8 mice, on a gluten-free diet, were intragastrically immunized witha chymotryptic digest of r-�-gliadin along with cholera toxin as adjuvant. Spleen and mesenteric lymph node T cell responses wereanalyzed for in vitro proliferative assay using a panel of synthetic peptides encompassing the entire sequence of r-�-gliadin. Twoimmunodominant epitopes corresponding to peptide p13 (aa 120–139) and p23 (aa 220–239) were identified. The response wasrestricted to DQ and mediated by CD4� T cells. In vitro tissue transglutaminase deamidation of both peptides did not increasethe response; furthermore, tissue transglutaminase catalyzed extensive deamidation in vitro along the entire r-�-gliadin molecule,but failed to elicit new immunogenic determinants. Surprisingly, the analysis of the cytokine profile showed that both deamidatedand native peptides induced preferentially IFN-� secretion, despite the use of cholera toxin, a mucosal adjuvant that normallyinduces a Th2 response to bystander Ags. Taken together, these data suggest that, in this model of gluten hypersensitivity,deamidation is not a prerequisite for the initiation of gluten responses. The Journal of Immunology, 2005, 175: 8087–8095.
I mmune responses generally do not recognize all possibleepitopes, but are focused on relatively few immunodominantones. In various murine and human experimental systems, it
has been reported that a large fraction of activated T cells is spe-cific for a single epitope-MHC combination (1). Therefore,epitope-based immunodominance could represent a major issue fortherapeutic or prophylactic intervention in many infective and au-toimmune diseases.
Celiac disease is an inflammatory disease of the small intestinewith an autoimmune component (2, 3) affecting genetically sus-ceptible individuals. A major feature of celiac disease is the inap-propriate intestinal T cell activation in HLA-DQA1*05-DQB1*02(DQ2) and HLA-DQA1*03-DQB1*0302 (DQ8) patients, trig-gered by peptides from wheat gliadin and related prolamins frombarley and rye. Activation of innate immunity mechanisms seemsto precede the initiation of gluten-specific adaptive immune re-sponse (4). To date, several gliadin epitopes have been identified(5). Most of these T cell epitopes were recognized following de-amidation by the tissue transglutaminase enzyme (tTG)3; this en-zyme converts some specific glutamine residues to glutamic acid
(6). Moreover, DQ2-restricted T cell epitopes cluster in regionsthat are rich in proline residues (7); the presence of proline protectsagainst digestive proteolysis and directs tTG-mediated deamida-tion on glutamine (8). However, it still remains questionablewhether tTG-catalyzed deamidation of gluten is a prerequisite forthe development of disease. In fact, there is strong evidence thatnative gluten peptides are also recognized by CD4� T cells (9).
A specific gliadin peptide has been described to be immuno-dominant in DQ2 adults (10). This peptide is part of an immuno-active 33-mer gliadin peptide found to be resistant to digestion bygastric and pancreatic enzymes (11). To date, only one DQ8-re-stricted gliadin epitope has been reported to be recognized by in-testinal T cell clones from celiac disease patients (12). Interest-ingly, this peptide has also been found to activate the immuneresponse in in vitro cultured intestinal biopsies from HLA-DQ8celiac disease patients in both native and deamidated forms (13).
HLA transgenic (tg) models are considered important tools tobetter address immunodominance of epitopes and have also beenfound particularly useful to identify and characterize autoantigensassociated with autoimmune diseases (14). Recently, two HLA tgmice models have been developed based on the expression ofHLA-DQ8 genes in the absence of their endogenous counterparts(15, 16). Results obtained showed a clear correlation between thepresence of DQ8 and acquired sensitivity to gluten in both models;however, the inability to induce a small-bowel pathology suggeststhat further, still undetected mechanisms are involved in the gen-esis of the enteropathy.
In this study, DQ8 mice were used to identify the immunodom-inant components of a recombinant �-gliadin (r-�-gliadin), whose
*Istituto di Scienze dell’Alimentazione, Consiglio Nazionale delle Ricerche, Avel-lino, Italy; †Department of Immunology, Mayo Clinic College of Medicine, Roches-ter, MN 55905; and ‡European Laboratory for Food Induced Diseases and Depart-ment of Pediatrics, University “Federico II” of Naples, Naples, Italy
Received for publication May 27, 2005. Accepted for publication October 12, 2005.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This study was supported by Progetti di Ricerca di Interesse Nazionale funds fromthe Italian Ministry of Education and Research.2 Address correspondence and reprint requests to Dr. Mauro Rossi, Istituto di Scienzedell’Alimentazione, Consiglio Nazionale delle Ricerche, via Roma 52, 83100 Avel-lino, Italy. E-mail address: [email protected] Abbreviations used in this paper: tTG, tissue transglutaminase; CT, cholera toxin;ES/MS, electrospray mass spectrometry; MLN, mesenteric lymph node; NALT, na-
sal-associated lymphoid tissue; r-�-gliadin, recombinant �-gliadin; RP-HPLC, re-versed-phase HPLC; tg, transgenic.
The Journal of Immunology
Copyright © 2005 by The American Association of Immunologists, Inc. 0022-1767/05/$02.00
by guest on April 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

sequence is widely conserved among the various forms of �-gli-adin. To use a physiological route of Ag delivery, we orally ad-ministered the protein along with cholera toxin (CT) as mucosaladjuvant, and the response was examined by T cell proliferativeassay after in vitro stimulation with a panel of synthetic peptidesspanning the whole protein sequence. We found that, among the 26peptides tested, only two were able to induce strong immune re-sponses both in spleen and in mesenteric lymph node (MLN). Thecytokine profile induced by these peptides was Th1-like, despitethe use of CT, which is associated with Th2 or Th1/Th2 mixedresponses. To date, this is the first study in which gliadin immu-nodominant epitopes were identified following oral immunizationin a mouse model.
Materials and MethodsMice
The tg mice expressing the HLA-DQ8 molecule in the absence of endog-enous mouse class II genes, non-tg for human CD4 (17), were maintainedin pathogen-free conditions at the animal facility of the Istituto di Scienzedell’Alimentazione. Mice were reared on a gluten-free diet for several gen-erations. Mice from this colony were used at the age of 8–14 wk.
Production of a r-�-gliadin
A database containing alignments of 34 �-gliadin amino acid sequences,retrieved from Swiss Prot, was developed using Megalign Lasergene Sys-tem software (DNASTAR). On the basis of these data, the most conservedsequence was identified into the �-gliadin encoded by the pTS63 clone(European Molecular Biology Laboratory accession AJ130948) (18). The714- to 1517-nt region of the DNA clone pTS63, corresponding to the ma-ture form of the �-gliadin, was amplified by PCR using the forward primer5�-GATCCATATGGTTAGAGTTCCAGTGC-3� and the reverse primer5�-GATCAAGCTTTCATTAGTTAGTACCGAAGATG-3�. These prim-ers have been designed to introduce a NdeI site and the start codon at the5� end of the PCR product and two stop codons and a HindIII site at its 3�end. Proofreading PCR was performed in a final reaction volume of 50 �lusing 250 ng of digested pTS63 plasmid, 2.5 U of Pfu Turbo DNA poly-merase (Stratagene), 1� cloned PFU DNA polymerase buffer (Stratagene),40 pmol of each primer, and 0.2 mM each deoxyribonucleotide. Amplifi-cation conditions were for 29 cycles at 95°C for 1 min, 48°C for 30 s, and72°C for 1 min, followed by a final step at 72°C for 5 min. The NdeI/HindIII-digested PCR product was cloned into the expression vectorpET17xb vector (Novagen) under control of bacteriophage T7 transcriptionand translation signals. After transformation into Escherichia coli XL-10ultracompetent cells according to the manufacturer’s instructions (Strat-agene), the recombinant colonies were screened for the presence of the�-gliadin fragment. The resulting recombinant plasmid pMR191 waschecked by sequencing and subsequently transformed into competentBL21(DE3)pLysS cells, according to the manufacturer’s instructions (No-vagen). The plasmid pMR191 was expressed in this strain at 37°C. Whenthe OD at 590 nm reached 0.6, 0.4 mM isopropyl �-D-thiogalactoside(Sigma-Aldrich) was added to the culture medium, followed by furtherculturing for 18 h. Harvested bacteria were washed once in 20 mM Tris-HCl (pH 8.0), resuspended in 70% ethanol, and incubated at 60°C for 2 h,according to Arentz-Hansen et al. (19). All of the purification steps there-after were performed at 4°C. After removal of the bacteria cell debris bycentrifugation, proteins were precipitated overnight by adding 2 vol of 1.5M NaCl. The precipitate was collected by centrifugation at 14,000 � g for30 min and washed once with n-butanol for the removal of residual bac-terial lipids. The pellet was solubilized in 0.1 M Tris-HCl (pH 8.5) con-taining 6 M guanidinium chloride and subjected to reversed-phase HPLC(RP-HPLC) purification, as previously described (20), except that the elu-tion step was achieved by means of a linear gradient from 30 to 60% B over30 min. Electrospray mass spectrometry (ES/MS) analysis of the HPLCfraction was performed using a Platform single quadrupole mass spectrom-eter (Micromass), as previously described (20). Gliadin preparations usedfor administration in mice were routinely assayed for endotoxin by usingthe QCL-1000 reagent kit (BioWhittaker) and found to have endotoxinlevels of �0.5 EU/ml.
Production of synthetic peptides
To map the dominant T cell determinants of r-�-gliadin, a set of 26 pep-tides, spanning the entire protein sequence, was synthesized as 20-mers bysolid-phase synthesis using Fmoc chemistry on a PerSeptive Biosystems
Pioneer Peptide Synthesis System GEN600611 (Applied Biosystems), us-ing the manufacturer’s protocol; each peptide overlapped the adjacent pep-tides by 10 aa. Their identity and purity were assessed by means ofRP-HPLC and MALDI-TOF mass spectrometry analyses. Synthetic pep-tides were between 92 and 95% pure.
Enzyme reactions
Chymotryptic digestion of r-�-gliadin was performed in 100 mM ammo-nium bicarbonate (pH 7.5) at 37°C for 16 h (1:100; enzyme:substrate ratio).Deamidation of 0.8 mM synthetic peptides was conducted in 0.125 MTris-HCl (pH 8.5), containing 0.2 mg/ml guinea pig liver tTG, 1 mMcalcium chloride, and 10 mM DTT. The reaction was conducted at 37°Cfor 4 h.
Immunization protocol
Mice were mucosally immunized by intragastric administration of 0.5 mlof PBS containing 500 �g of a chymotryptic digest of r-�-gliadin plus 25�g of CT on days 0, 7, and 14. In some experiments, mice were immunizedby intrafootpad injections into rear footpad with 50 �g of r-�-gliadin in 50�l of distilled water emulsified in 50 �l of CFA.
In vitro proliferation assays
Mucosally immunized mice were sacrificed on day 21; MLN and spleenswere harvested and passed through a stainless steel wire mesh to dissociatecells. Cells were cultured (4 � 105 cells/well) in 0.2 ml of culture mediumin 96-well bottom tissue culture plates at 37°C for 96 h in the presence of125 �g/ml r-�-gliadin or 50 �M peptides. Sixteen hours before harvesting,cultures were pulsed with 1 �Ci/well [3H]thymidine. Results were ex-pressed as stimulation index (mean cpm of triplicate cultures containingAg)/(mean cpm of cells cultured with medium alone).
In some experiments, CD4� cells were purified by magnetic cell sortingusing CD4� (L3T4) microbeads, according to the manufacturer’s instruc-tions (Miltenyi Biotec); cell yield was assessed by FACS analysis. A totalof 2 � 105 purified CD4� cells was incubated with 4 � 105 irradiatedsyngenic spleen cells as APC in the presence of Ag, as described above.
Parenterally immunized mice were sacrificed on day 10 to collect pop-liteal lymph node cells for proliferation assay, as described above.
Cytokine assay
CD4� T cells were cultured at 2.5 � 106 cells/ml in the presence of APC(5 � 106 cells/ml) and subjected to one to three rounds of Ag stimulationin vitro; after 72 h following each Ag incubation, supernatants were col-lected and analyzed for IFN-�, IL-4, and IL-10 protein levels by ELISA.
FACS analysis
The staining of cell markers was performed on ice. All conjugated Abswere purchased from BD Discovery Labware. After preincubation for 5min with Fc block to avoid nonspecific binding, 2 � 105 cells were stainedfor 20 min with FITC-conjugated anti-DQ, FITC-conjugated anti-mouseCD3, FITC-conjugated anti-mouse MHC class II, PE-Cy5-conjugated anti-mouse CD4, or PE-conjugated anti-mouse CD8. For dendritic cell staining,cells were incubated with biotinylated anti-CD11c; after washing, cellswere incubated for 20 min with streptavidin-conjugated R-PE. Cells wereanalyzed by flow cytometry using a FACSCalibur and CellQuest software(BD Biosciences). An isotype control of appropriate subclass was used foreach specific Ab.
ResultsAntigens
Native gliadin is a mixture of many components (21) showingvariability in inducing immune responses (22). This heterogeneitymakes difficult the identification of immunodominant epitopes. Toovercome this issue, we focused our attention on the “a” compo-nent of this mixture and, particularly, on the most conserved se-quence among all the �-gliadins found in the European MolecularBiology Laboratory Nucleotide Sequence Database. A total of 34entries was examined by using a protein alignment software toderive a consensus sequence. Swiss Prot entry Q9ZP09 (Fig. 1A)was the closest in identity (99%) to the consensus sequence withonly two mismatches (K3N; T3A). The cDNA sequence, en-coding for the mature form of the identified protein (European
8088 EPITOPES OF �-GLIADIN IN HLA-DQ8 MICE
by guest on April 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
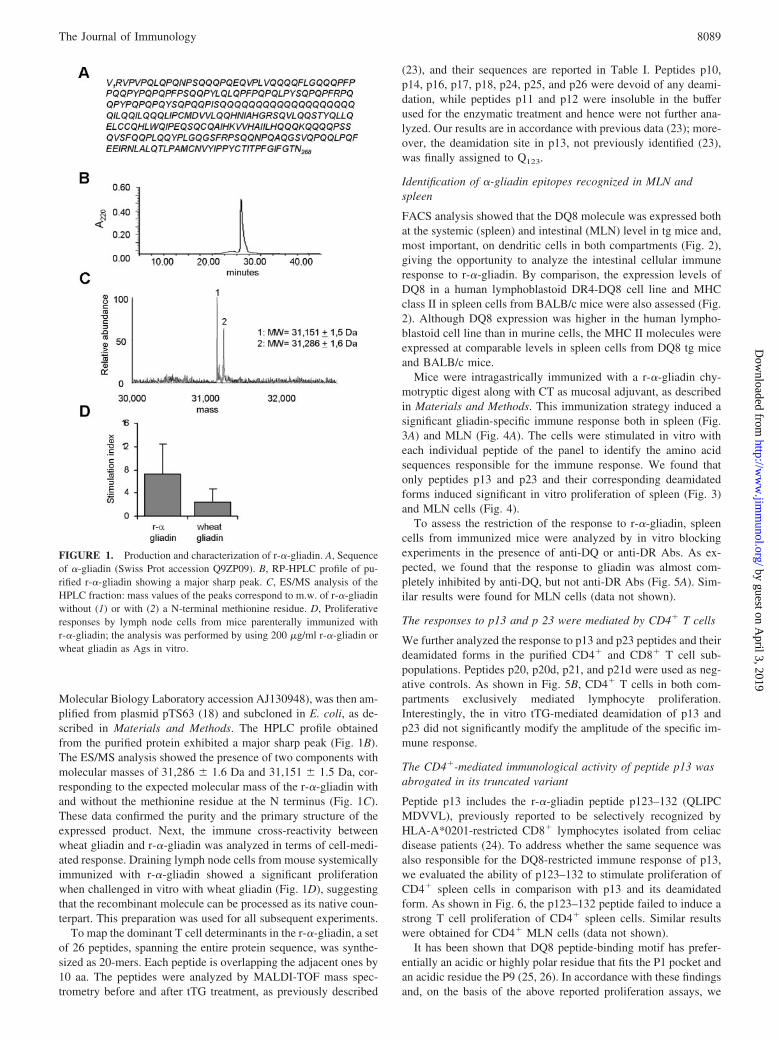
Molecular Biology Laboratory accession AJ130948), was then am-plified from plasmid pTS63 (18) and subcloned in E. coli, as de-scribed in Materials and Methods. The HPLC profile obtainedfrom the purified protein exhibited a major sharp peak (Fig. 1B).The ES/MS analysis showed the presence of two components withmolecular masses of 31,286 � 1.6 Da and 31,151 � 1.5 Da, cor-responding to the expected molecular mass of the r-�-gliadin withand without the methionine residue at the N terminus (Fig. 1C).These data confirmed the purity and the primary structure of theexpressed product. Next, the immune cross-reactivity betweenwheat gliadin and r-�-gliadin was analyzed in terms of cell-medi-ated response. Draining lymph node cells from mouse systemicallyimmunized with r-�-gliadin showed a significant proliferationwhen challenged in vitro with wheat gliadin (Fig. 1D), suggestingthat the recombinant molecule can be processed as its native coun-terpart. This preparation was used for all subsequent experiments.
To map the dominant T cell determinants in the r-�-gliadin, a setof 26 peptides, spanning the entire protein sequence, was synthe-sized as 20-mers. Each peptide is overlapping the adjacent ones by10 aa. The peptides were analyzed by MALDI-TOF mass spec-trometry before and after tTG treatment, as previously described
(23), and their sequences are reported in Table I. Peptides p10,p14, p16, p17, p18, p24, p25, and p26 were devoid of any deami-dation, while peptides p11 and p12 were insoluble in the bufferused for the enzymatic treatment and hence were not further ana-lyzed. Our results are in accordance with previous data (23); more-over, the deamidation site in p13, not previously identified (23),was finally assigned to Q123.
Identification of �-gliadin epitopes recognized in MLN andspleen
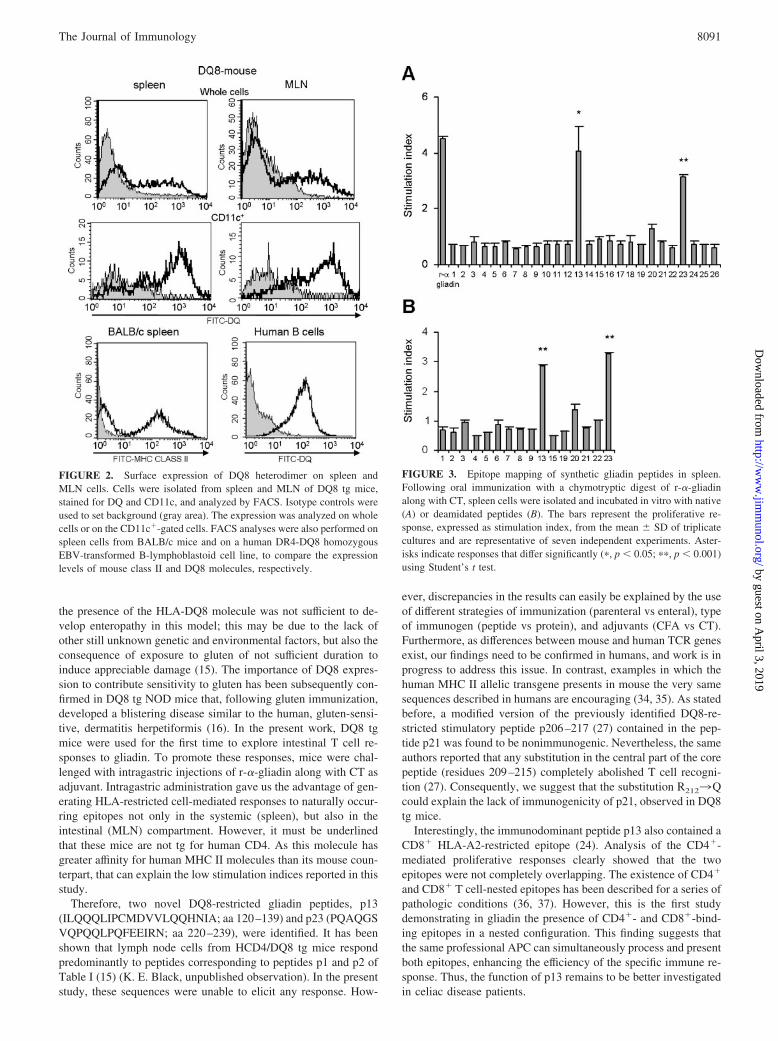
FACS analysis showed that the DQ8 molecule was expressed bothat the systemic (spleen) and intestinal (MLN) level in tg mice and,most important, on dendritic cells in both compartments (Fig. 2),giving the opportunity to analyze the intestinal cellular immuneresponse to r-�-gliadin. By comparison, the expression levels ofDQ8 in a human lymphoblastoid DR4-DQ8 cell line and MHCclass II in spleen cells from BALB/c mice were also assessed (Fig.2). Although DQ8 expression was higher in the human lympho-blastoid cell line than in murine cells, the MHC II molecules wereexpressed at comparable levels in spleen cells from DQ8 tg miceand BALB/c mice.
Mice were intragastrically immunized with a r-�-gliadin chy-motryptic digest along with CT as mucosal adjuvant, as describedin Materials and Methods. This immunization strategy induced asignificant gliadin-specific immune response both in spleen (Fig.3A) and MLN (Fig. 4A). The cells were stimulated in vitro witheach individual peptide of the panel to identify the amino acidsequences responsible for the immune response. We found thatonly peptides p13 and p23 and their corresponding deamidatedforms induced significant in vitro proliferation of spleen (Fig. 3)and MLN cells (Fig. 4).
To assess the restriction of the response to r-�-gliadin, spleencells from immunized mice were analyzed by in vitro blockingexperiments in the presence of anti-DQ or anti-DR Abs. As ex-pected, we found that the response to gliadin was almost com-pletely inhibited by anti-DQ, but not anti-DR Abs (Fig. 5A). Sim-ilar results were found for MLN cells (data not shown).
The responses to p13 and p 23 were mediated by CD4� T cells
We further analyzed the response to p13 and p23 peptides and theirdeamidated forms in the purified CD4� and CD8� T cell sub-populations. Peptides p20, p20d, p21, and p21d were used as neg-ative controls. As shown in Fig. 5B, CD4� T cells in both com-partments exclusively mediated lymphocyte proliferation.Interestingly, the in vitro tTG-mediated deamidation of p13 andp23 did not significantly modify the amplitude of the specific im-mune response.
The CD4�-mediated immunological activity of peptide p13 wasabrogated in its truncated variant
Peptide p13 includes the r-�-gliadin peptide p123–132 (QLIPCMDVVL), previously reported to be selectively recognized byHLA-A*0201-restricted CD8� lymphocytes isolated from celiacdisease patients (24). To address whether the same sequence wasalso responsible for the DQ8-restricted immune response of p13,we evaluated the ability of p123–132 to stimulate proliferation ofCD4� spleen cells in comparison with p13 and its deamidatedform. As shown in Fig. 6, the p123–132 peptide failed to induce astrong T cell proliferation of CD4� spleen cells. Similar resultswere obtained for CD4� MLN cells (data not shown).
It has been shown that DQ8 peptide-binding motif has prefer-entially an acidic or highly polar residue that fits the P1 pocket andan acidic residue the P9 (25, 26). In accordance with these findingsand, on the basis of the above reported proliferation assays, we
FIGURE 1. Production and characterization of r-�-gliadin. A, Sequenceof �-gliadin (Swiss Prot accession Q9ZP09). B, RP-HPLC profile of pu-rified r-�-gliadin showing a major sharp peak. C, ES/MS analysis of theHPLC fraction: mass values of the peaks correspond to m.w. of r-�-gliadinwithout (1) or with (2) a N-terminal methionine residue. D, Proliferativeresponses by lymph node cells from mice parenterally immunized withr-�-gliadin; the analysis was performed by using 200 �g/ml r-�-gliadin orwheat gliadin as Ags in vitro.
8089The Journal of Immunology
by guest on April 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

speculated that p13 residues Q122 and D130 fit respectively in P1and P9 pockets of DQ8, as shown in the inset of Fig. 6. This modelcan explain both our observations, i.e., the failure of p123–132 ininducing a response and a quite similar immune response of p13 inboth native and deamidated form (Fig. 5). In fact, p13 tTG treat-ment deamidates Q123, but not Q122 (Table I), indicating that noamino acidic substitution occurs at P1 pocket. Similarly, gliadinresidues Q227 and E235 of p23 could allocate in P1 and P9 pockets,respectively. A comparative alignment for gliadin peptide 207–213(27) is presented in the inset of Fig. 6. Interestingly, all of the threeexamined peptides (p13, p23, peptide 206–217) showed a prolineresidue that could be allocated in the P6 pocket; no other similarsequences were identified all along the r-�-gliadin.
Analysis of the cytokine pattern induced by the immunodominantp13 and p23 peptides
To study the pattern of cytokine induction, DQ8 tg mice wereimmunized with r-�-gliadin and CD4� T cells were in vitro cul-tured with both r-�-gliadin and the immunodominant peptides p13and p23. The cell culture supernatants were analyzed by quantita-tive ELISA for IFN-�, IL-4, and IL-10 production. As shown inFig. 7, following first stimulation with r-�-gliadin, p13, or p23,spleen CD4� T cells secreted significant levels of IFN-� and notof IL-4 or IL-10; however, these cytokines were undetectable inMLN CD4� T cells. After a second round of in vitro Ag stimu-lation, MLN CD4� T cells also showed a measurable response,characterized by production of both IFN-� and IL-4. Interestingly,after three rounds of Ag stimulation, the peptide-specific T cellresponses were characterized by a predominant IFN-� secretion inboth compartments, whereas r-�-gliadin-derived T cells showedincreased expression of IL-4 and IL-10.
DiscussionGliadin, which has been considered responsible for inducing celiacdisease (28), is a mixture of many proteins (21), with marked dif-ferences in stimulating murine T cell proliferative responses (22).
The gliadin fraction has been grouped in three classes named “�,”“�,” and “�” based on biochemical analysis that revealed a tightstructural relationship among the various constituents (29). Ourprevious studies also showed that purified �-gliadin is able todown-regulate the systemic immune response after a challengewith the whole gliadin fraction (20), suggesting a strong immunecross-reactivity among the different classes. Based on these obser-vations, we focused our attention on the “�” component of gliadin.Upon database searches, it was not possible to retrieve a fullyconserved consensus sequence among all of the 34 entries of�-gliadins; as a consequence, we considered the most conservedentry, Q9ZP09, representing an �-gliadin from spelt wheat (18).This protein includes sequence LGQQQPFPPQQPY correspond-ing to peptide p31–43, previously recognized to induce damageboth in vitro (30) and in vivo in celiac disease (31), and sequenceLQLQPFPQPQLP of the DQ2-restricted �9 peptide p57–68.However, it does not contain DQ2-restricted �2 peptide p62–75(FPQPQLPYPQPQLP) (32) or the multivalent 33-mer peptidep57–89(LQLQPFPQPQLPYPQPQLPYPQPQLPYPQPQPF)(11).The sequence SGQGSFQPSQQN of the gliadin peptide p206–217, previously found to be recognized by T cells isolated fromDQ8 patients (12), with two substitutions (L2063S and R2123Q),is also found in this protein.
Wheat gliadin-stimulated responses in r-�-gliadin-primed mice,as shown by proliferation assays, indicated the existence not onlyof marked structural homologies between native and recombinantproteins, but also of similarities in the processing steps followed byAPC. Our previous findings on induction of systemic tolerancetoward wheat gliadin following nasal administration of r-�-gliadin(33) further strengthened this concept.
Genetics plays a key role in the pathogenesis of celiac disease. Inparticular, alleles that code for HLA-DQ2 or HLA-DQ8 heterodimersare present in virtually all patients with celiac disease (3). Interest-ingly, it has been reported that expression of the human heterodimerDQ8 in tg mice confers sensitivity to gluten, providing a powerfulmodel system to study some aspects of celiac disease (15). However,
Table I. List of synthetic native and deamidated peptides of r-�-gliadin
Code Peptide Sequence Code Peptide Sequence
p1 1–20 VRVPVPQLQPQNPSQQQPQE p1d 1–20 VRVPVPELQPENPSQQQPQEp2 10–29 PQNPSQQQPQEQVPLVQQQQ p2d 10–29 PQNPSQEQPQEEVPLVQQQQp3 20–39 EQVPLVQQQQFLGQQQPFPP p3d 20–39 EEVPLVQEQQFLGQQQPFPPp4 30–49 FLGQQQPFPPQQPYPQPQPF p4d 30–49 FLGQEQPFPPEQPYPQPQPFp5 40–59 QQPYPQPQPFPSQQPYLQLQ p5d 40–59 QQPYPQPQPFPSEQPYLQLQp6 50–69 PSQQPYLQLQPFPQPQLPYS p6d 50–69 PSEQPYLQLQPFPQPELPYSp7 60–79 PFPQPQLPYSQPQPFRPQQP p7d 60–79 PFPQPELPYSQPQPFRPQQPp8 70–89 QPQPFRPQQPYPQPQPQYSQ p8d 70–89 QPQPFRPEQPYPQPQPEYSQp9 80–99 YPQPQPQYSQPQQPISQQQQ p9d 80–99 YPQPQPEYSQPEQPISQQQQp10 90–109 PQQPISQQQQQQQQQQQQQQp11 100–119 QQQQQQQQQQQQQQQQILQQp12 110–129 QQQQQQILQQILQQQLIPCMp13 120–139 ILQQQLIPCMDVVLQQHNIA p13d 120–139 ILQEQLIPCMDVVLQQHNIAp14 130–149 DVVLQQHNIAHGRSQVLQQSp15 141–160 GRSQVLQQSTYQLLQELCCQ p15d 141–160 GRSQVLQESTYQLLQELCCQp16 150–169 TYQLLQELCCQHLWQIPEQSp17 160–179 QHLWQIPEQSQCQAIHKVVHp18 170–189 QCQAIHKVVHAHLHQQQKQp19 180–199 AHLHQQQKQQQQPSSQVSF p19d 180–199 AHLHQQQKQQEQPSSQVSFp20 190–209 QQQPSSQVSFQQPLQQYPLG p20d 190–209 QQQPSSQVSFEQPLQEYPLGp21 200–219 QQPLQQYPLGQGSFRPSQQN p21d 200–219 QQPLQEYPLGQGSFRPSQQNp22 210–229 QGSFRPSQQNPQAQGSVQPQ p22d 210–229 QGSFRPSQQNPQAEGSVQPQp23 220–239 PQAQGSVQPQQLPQFEEIRN p23d 220–239 PQAEGSVQPQELPQFEEIRNp24 230–249 QLPQFEEIRNLALQTLPAMCp25 240–259 LALQTLPAMCNVYIPPYCTIp26 250–268 NVYIPPYCTITPFGIFGTN
8090 EPITOPES OF �-GLIADIN IN HLA-DQ8 MICE
by guest on April 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
the presence of the HLA-DQ8 molecule was not sufficient to de-velop enteropathy in this model; this may be due to the lack ofother still unknown genetic and environmental factors, but also theconsequence of exposure to gluten of not sufficient duration toinduce appreciable damage (15). The importance of DQ8 expres-sion to contribute sensitivity to gluten has been subsequently con-firmed in DQ8 tg NOD mice that, following gluten immunization,developed a blistering disease similar to the human, gluten-sensi-tive, dermatitis herpetiformis (16). In the present work, DQ8 tgmice were used for the first time to explore intestinal T cell re-sponses to gliadin. To promote these responses, mice were chal-lenged with intragastric injections of r-�-gliadin along with CT asadjuvant. Intragastric administration gave us the advantage of gen-erating HLA-restricted cell-mediated responses to naturally occur-ring epitopes not only in the systemic (spleen), but also in theintestinal (MLN) compartment. However, it must be underlinedthat these mice are not tg for human CD4. As this molecule hasgreater affinity for human MHC II molecules than its mouse coun-terpart, that can explain the low stimulation indices reported in thisstudy.
Therefore, two novel DQ8-restricted gliadin peptides, p13(ILQQQLIPCMDVVLQQHNIA; aa 120–139) and p23 (PQAQGSVQPQQLPQFEEIRN; aa 220–239), were identified. It has beenshown that lymph node cells from HCD4/DQ8 tg mice respondpredominantly to peptides corresponding to peptides p1 and p2 ofTable I (15) (K. E. Black, unpublished observation). In the presentstudy, these sequences were unable to elicit any response. How-
ever, discrepancies in the results can easily be explained by the useof different strategies of immunization (parenteral vs enteral), typeof immunogen (peptide vs protein), and adjuvants (CFA vs CT).Furthermore, as differences between mouse and human TCR genesexist, our findings need to be confirmed in humans, and work is inprogress to address this issue. In contrast, examples in which thehuman MHC II allelic transgene presents in mouse the very samesequences described in humans are encouraging (34, 35). As statedbefore, a modified version of the previously identified DQ8-re-stricted stimulatory peptide p206–217 (27) contained in the pep-tide p21 was found to be nonimmunogenic. Nevertheless, the sameauthors reported that any substitution in the central part of the corepeptide (residues 209–215) completely abolished T cell recogni-tion (27). Consequently, we suggest that the substitution R2123Qcould explain the lack of immunogenicity of p21, observed in DQ8tg mice.
Interestingly, the immunodominant peptide p13 also contained aCD8� HLA-A2-restricted epitope (24). Analysis of the CD4�-mediated proliferative responses clearly showed that the twoepitopes were not completely overlapping. The existence of CD4�
and CD8� T cell-nested epitopes has been described for a series ofpathologic conditions (36, 37). However, this is the first studydemonstrating in gliadin the presence of CD4�- and CD8�-bind-ing epitopes in a nested configuration. This finding suggests thatthe same professional APC can simultaneously process and presentboth epitopes, enhancing the efficiency of the specific immune re-sponse. Thus, the function of p13 remains to be better investigatedin celiac disease patients.
FIGURE 2. Surface expression of DQ8 heterodimer on spleen andMLN cells. Cells were isolated from spleen and MLN of DQ8 tg mice,stained for DQ and CD11c, and analyzed by FACS. Isotype controls wereused to set background (gray area). The expression was analyzed on wholecells or on the CD11c�-gated cells. FACS analyses were also performed onspleen cells from BALB/c mice and on a human DR4-DQ8 homozygousEBV-transformed B-lymphoblastoid cell line, to compare the expressionlevels of mouse class II and DQ8 molecules, respectively.
FIGURE 3. Epitope mapping of synthetic gliadin peptides in spleen.Following oral immunization with a chymotryptic digest of r-�-gliadinalong with CT, spleen cells were isolated and incubated in vitro with native(A) or deamidated peptides (B). The bars represent the proliferative re-sponse, expressed as stimulation index, from the mean � SD of triplicatecultures and are representative of seven independent experiments. Aster-isks indicate responses that differ significantly (�, p � 0.05; ��, p � 0.001)using Student’s t test.
8091The Journal of Immunology
by guest on April 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
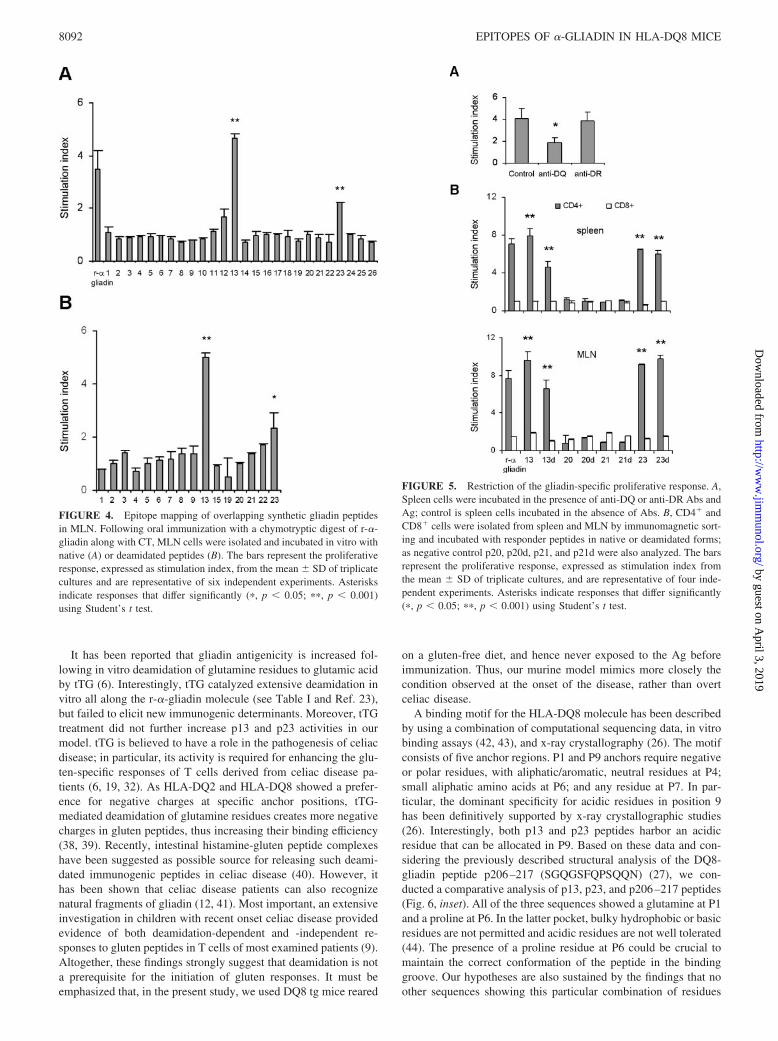
It has been reported that gliadin antigenicity is increased fol-lowing in vitro deamidation of glutamine residues to glutamic acidby tTG (6). Interestingly, tTG catalyzed extensive deamidation invitro all along the r-�-gliadin molecule (see Table I and Ref. 23),but failed to elicit new immunogenic determinants. Moreover, tTGtreatment did not further increase p13 and p23 activities in ourmodel. tTG is believed to have a role in the pathogenesis of celiacdisease; in particular, its activity is required for enhancing the glu-ten-specific responses of T cells derived from celiac disease pa-tients (6, 19, 32). As HLA-DQ2 and HLA-DQ8 showed a prefer-ence for negative charges at specific anchor positions, tTG-mediated deamidation of glutamine residues creates more negativecharges in gluten peptides, thus increasing their binding efficiency(38, 39). Recently, intestinal histamine-gluten peptide complexeshave been suggested as possible source for releasing such deami-dated immunogenic peptides in celiac disease (40). However, ithas been shown that celiac disease patients can also recognizenatural fragments of gliadin (12, 41). Most important, an extensiveinvestigation in children with recent onset celiac disease providedevidence of both deamidation-dependent and -independent re-sponses to gluten peptides in T cells of most examined patients (9).Altogether, these findings strongly suggest that deamidation is nota prerequisite for the initiation of gluten responses. It must beemphasized that, in the present study, we used DQ8 tg mice reared
on a gluten-free diet, and hence never exposed to the Ag beforeimmunization. Thus, our murine model mimics more closely thecondition observed at the onset of the disease, rather than overtceliac disease.
A binding motif for the HLA-DQ8 molecule has been describedby using a combination of computational sequencing data, in vitrobinding assays (42, 43), and x-ray crystallography (26). The motifconsists of five anchor regions. P1 and P9 anchors require negativeor polar residues, with aliphatic/aromatic, neutral residues at P4;small aliphatic amino acids at P6; and any residue at P7. In par-ticular, the dominant specificity for acidic residues in position 9has been definitively supported by x-ray crystallographic studies(26). Interestingly, both p13 and p23 peptides harbor an acidicresidue that can be allocated in P9. Based on these data and con-sidering the previously described structural analysis of the DQ8-gliadin peptide p206–217 (SGQGSFQPSQQN) (27), we con-ducted a comparative analysis of p13, p23, and p206–217 peptides(Fig. 6, inset). All of the three sequences showed a glutamine at P1and a proline at P6. In the latter pocket, bulky hydrophobic or basicresidues are not permitted and acidic residues are not well tolerated(44). The presence of a proline residue at P6 could be crucial tomaintain the correct conformation of the peptide in the bindinggroove. Our hypotheses are also sustained by the findings that noother sequences showing this particular combination of residues
FIGURE 4. Epitope mapping of overlapping synthetic gliadin peptidesin MLN. Following oral immunization with a chymotryptic digest of r-�-gliadin along with CT, MLN cells were isolated and incubated in vitro withnative (A) or deamidated peptides (B). The bars represent the proliferativeresponse, expressed as stimulation index, from the mean � SD of triplicatecultures and are representative of six independent experiments. Asterisksindicate responses that differ significantly (�, p � 0.05; ��, p � 0.001)using Student’s t test.
FIGURE 5. Restriction of the gliadin-specific proliferative response. A,Spleen cells were incubated in the presence of anti-DQ or anti-DR Abs andAg; control is spleen cells incubated in the absence of Abs. B, CD4� andCD8� cells were isolated from spleen and MLN by immunomagnetic sort-ing and incubated with responder peptides in native or deamidated forms;as negative control p20, p20d, p21, and p21d were also analyzed. The barsrepresent the proliferative response, expressed as stimulation index fromthe mean � SD of triplicate cultures, and are representative of four inde-pendent experiments. Asterisks indicate responses that differ significantly(�, p � 0.05; ��, p � 0.001) using Student’s t test.
8092 EPITOPES OF �-GLIADIN IN HLA-DQ8 MICE
by guest on April 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

were present all along the r-�-gliadin. Moreover, the recently iden-tified HLA-A2-restricted gliadin peptide p123–132 (24), whosesequence represents a truncated version of our proposed bindingmotif for p13 (see inset in Fig. 6), also failed to stimulate CD4� Tcells. Studies aimed at the identification of the minimal core re-gions of the stimulatory gliadin p13 and p23 peptides are ongoingin our lab.
It is known that the type of adjuvant and route of administrationinfluence the immune response. In particular, mucosal coimmuni-zation with CT predominantly enhances Th2 responses to coad-ministered Ag (45, 46). Instead, the results reported in this workare indicative of a powerful Th1 cellular response in spleen, fol-lowing in vitro r-�-gliadin stimulation. This characteristic was lostafter subsequent rounds of in vitro stimulation, when the responsewas consistently Th2 in both the examined compartments. The invitro polarization of the immune response has already been de-scribed and related to a possible immunosuppressive function of Tcells generated by immunization with an Ag along with CT (46).These data suggest that gliadin-specific Th2/T regulatory T cellsare involved in the progressive in vitro suppression of Th1 T cells.It is intriguing to note that both the immunodominant peptidesinduced predominantly IFN-� secretion in vitro, as expected in theTh1-like response in celiac disease (47). This result can be ex-plained by the selective in vitro expansion of T cells that recognizep13 and p23 peptides, in the absence of a regulative Th2/T regu-latory activity driven by other gliadin peptides. Further studies areneeded to better address this issue.
In conclusion, our findings showed that only two immunodom-inant nondeamidated peptides from r-�-gliadin could be identifiedin sensitized DQ8 tg mice. These peptides were able to elicit potentCD4�-mediated Th1 responses. Clearly, the strategy of using asingle gliadin molecule, instead of the whole gliadin mixture,could underestimate the real number of epitopes, but this result canbe justified by assuming the existence of epitope spreading. Wesuggest that progression of the disease can be seen as a two-stepprocess: in an early phase, the immune response is directed towarda few natural peptides encompassing, among other crucial resi-dues, a glutamic or aspartic acid, useful for optimal binding toDQ8/DQ2. Subsequently, in situ tTG-mediated deamidation couldcreate new binding sites to the DQ8/DQ2 heterodimer, thus trig-gering the production of novel and, perhaps, more immunogenicepitopes. Epitope spreading has been demonstrated in a number ofanimal models of autoimmunity (48–50) and suggested to be in-volved in the pathogenesis of human autoimmune diseases(51, 52).
The finding that only two peptides may be responsible for ini-tiating DQ8-restricted T cell reactivity represents a key step to-
ward providing reagents for possible interventions in celiac dis-ease. Several approaches could be envisaged to block induction ofgluten intolerance by using these peptides. The administration ofnonstimulatory analogues that are still able to compete for bindingto HLA molecules is one possibility. An alternative strategy isrepresented by the administration of immunodominant peptides astolerogens. As the classical approach to induce tolerance, i.e., Agfeeding, is not applicable in celiac disease, alternative forms ofinduction of mucosal tolerance are mandatory. From this perspec-tive, the nasal-associated lymphoid tissue (NALT), the inductive
FIGURE 7. Cytokine production. CD4� cells, isolated from the spleenand MLN of mice orally immunized with a chymotryptic digest of r-�-gliadin plus CT, were tested for cytokine production after one, two, or threerounds of Ag stimulation in vitro. Data, expressed as picograms per mil-liliter, represent mean � SD of triplicate cultures and are representative ofthree independent experiments.
FIGURE 6. Response of CD4� spleen cells to anHLA-A2-restricted epitope included in p13. Spleen cellswere incubated in the presence of r-�-gliadin, p13,p13d, or A gliadin p123–132 peptide (24). Data repre-sent mean � SD of triplicate cultures and are represen-tative of two independent experiments. ��, p � 0.001.Inset, Suggested binding register of newly identified im-munodominant gliadin peptides to DQ8 anchor posi-tions. For comparative purposes, the sequences of A gli-adin p123–132 and gliadin peptide p206–217 (27) werealso reported.
8093The Journal of Immunology
by guest on April 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

site of the upper respiratory airway in mice, is a possible candidate.In fact, NALT has been shown to be a potent inductive site notonly for respiratory, but also for other distant mucosal districts,including gut (53). In addition, NALT has been shown to be in-volved in the generation of tolerance (54). Moreover, we havealready found that intranasal administration of gliadin in mice candown-regulate the systemic immune response to this Ag (20, 55).Finally, r-�-gliadin induced a similar down-regulatory effect inDQ8 tg mice immunized with a commercial preparation of wheatgliadin (33). Hence, the identification of immunodominantepitopes of this molecule can be seen as a further step toward thedefinition of safer tolerogenic molecules to be administered in pa-tients with celiac disease for blocking the onset of the disease.
AcknowledgmentsWe thank Dr. Renato D’Ovidio (Universita della Tuscia, Viterbo, Italy) forthe generous gift of the clone pTS63, and Dr. Luigi Longobardo (Univer-sity “Federico II” of Naples, Naples, Italy) for assistance in the preparationof synthetic peptides. We also thank Dr. Prafullakumar Tailor andDr. Sunil Gangadharan (National Institute of Child Health and HumanDevelopment, National Institutes of Health, Bethesda, MD) for carefulreading of the manuscript.
DisclosuresThe authors have no financial conflict of interest.
References1. Sette, A., and L. Fikes. 2003. Epitope-based vaccines: an update on epitope
identification, vaccine design and delivery. Curr. Opin. Immunol. 15: 461–470.2. Maki, M., and P. Collin. 1997. Coeliac disease. Lancet 349: 1755–1759.3. Kagnoff, M. F. 2005. Overview and pathogenesis of celiac disease. Gastroenter-
ology 128: S10–S18.4. Maiuri, L., C. Ciacci, I. Ricciardelli, L. Vacca, V. Raia, S. Auricchio, J. Picard,
M. Osman, S. Quaratino, and M. Londei. 2003. Association between innate re-sponse to gliadin and activation of pathogenic T cells in coeliac disease. Lancet362: 30–37.
5. Vader, L. W., A. de Ru, Y. van der Wal, Y. M. C. Kooy, W. Benckhuijsen,M. L. Merin, J. W. Wouter Drijfhout, P. van Veelen, and F. Koning. 2002.Specificity of tissue transglutaminase explains cereal toxicity in celiac disease.J. Exp. Med. 195: 643–649.
6. Molberg, O., S. N. McAdam, R. Korner, H. Quarsten, C. Kristiansen, L. Madsen,L. Fugger, H. Scott, O. Noren, P. Roepstorff, et al. 1998. Tissue transglutaminaseselectively modifies gliadin peptides that are recognized by gut-derived T cells inceliac disease. Nat. Med. 4: 713–717.
7. Arentz-Hansen, H., S. N. McAdam, O. Molberg, B. Fleckenstein, K. E. Lundin,T. J. Jorgensen, G. Jung, P. Roepstorff, and L. M. Sollid. 2002. Celiac lesion Tcells recognize epitopes that cluster in regions of gliadins rich in proline residues.Gastroenterology 123: 803–809.
8. Hausch, F., L. Shan, N. A. Santiago, G. M. Gray, and C. Khosla. 2002. Intestinaldigestive resistance of immunodominant gliadin peptides. Am. J. Physiol. Gas-trointest. Liver Physiol. 283: G996–G1003.
9. Vader, W., Y. Kooy, P. Van Veelen, A. De Ru, D. Harris, W. Benckhuijsen,S. Pena, L. Mearin, J. W. Drijfhout, and F. Koning. 2002. The gluten response inchildren with celiac disease is directed toward multiple gliadin and glutenin pep-tides. Gastroenterology 122: 1729–1737.
10. Anderson, R. P., P. Degano, A. J. Godkin, D. P. Jewell, and A. V. Hill. 2000. Invivo antigen challenge in celiac disease identifies a single transglutaminase-mod-ified peptide as the dominant A-gliadin T-cell epitope. Nat. Med. 6: 337–342.
11. Shan, L., O. Molberg, I. Parrot, F. Hausch, F. Filiz, G. M. Gray, L. M. Sollid, andC. Khosla. 2002. Structural basis for gluten intolerance in celiac sprue. Science297: 2275–2279.
12. Van de Wal, Y., Y. M. Kooy, P. A. van Veelen, S. A. Pena, L. M. Mearin,O. Molberg, K. E. Lundin, L. M. Sollid, T. Mutis, W. E. Benckhuijsen, et al.1998. Small intestinal T cells of celiac disease patients recognize a natural pepsinfragment of gliadin. Proc. Natl. Acad. Sci. USA 95: 10050–10054.
13. Mazzarella, G., M. Maglio, F. Paparo, G. Nardone, R. Stefanile, L. Greco,Y. van de Wal, Y. Kooy, F. Koning, S. Auricchio, and R. Troncone. 2003. Animmunodominant DQ8 restricted gliadin peptide activates small intestinal im-mune response in in vitro cultured mucosa from HLA-DQ8 positive but notHLA-DQ8 negative coeliac patients. Gut 52: 57–62.
14. Abraham, R. S., and C. S. David. 2000. Identification of HLA-class-II-restrictedepitopes of autoantigens in transgenic mice. Curr. Opin. Immunol. 12: 122–129.
15. Black, K. E., J. A. Murray, and C. S. David. 2002. HLA-DQ determines theresponse to exogenous wheat proteins: a model of gluten sensitivity in transgenicknockout mice. J. Immunol. 169: 5595–5600.
16. Marietta, E., K. Black, M. Camilleri, P. Krause, R. S. Rogers III, C. David,M. R. Pittelkow, and J. A. Murray. 2004. A new model for dermatitis herpeti-formis that uses HLA-DQ8 transgenic NOD mice. J. Clin. Invest. 114:1090–1097.
17. Cheng, S., J. Baisch, C. Krco, S. Savarirayan, J. Hanson, K. Hodgson, M. Smart,and C. David. 1996. Expression and function of HLA-DQ8 (DQA1*0301/DQB1*0302) genes in transgenic mice. Eur. J. Immunogenet. 23: 15–20.
18. Kasarda, D. D., and R. D’Ovidio. 1999. Deduced amino acid sequence of an �gliadin gene from spelt wheat (Spelta) includes sequences active in celiac disease.Cereal. Chem. 76: 548–551.
19. Arentz-Hansen, E. H., S. N. McAdam, O. Molberg, C. Kristiansen, andL. M. Sollid. 2000. Production of a panel of recombinant gliadins for the char-acterization of T cell reactivity in coeliac disease. Gut 46: 46–51.
20. Maurano, F., R. A. Siciliano, B. De Giulio, D. Luongo, M. F. Mazzeo,R. Troncone, S. Auricchio, and M. Rossi. 2001. Intranasal administration of one� gliadin can down-regulate the immune response to whole gliadin in mice.Scand. J. Immunol. 53: 290–295.
21. Platt, S. G., and D. D. Kasarda. 1971. Separation and characterization of gliadinfractions. Biochim. Biophys. Acta 243: 407–415.
22. Kagnoff, M. F., R. K. Austin, H. C. L. Johnson, J. E. Bernardin, M. D. Dietler,and D. D. Kasarda. 1982. Coeliac sprue: correlation with murine T cell responsesto wheat gliadin components. J. Immunol. 129: 2693–2697.
23. Mazzeo, M. F., B. De Giulio, S. Senger, M. Rossi, A. Malorni, andR. A. Siciliano. 2003. Identification of transglutaminase-mediated deamidationsites in a recombinant �-gliadin by advanced mass-spectrometric methodologies.Protein Sci. 12: 2434–2442.
24. Gianfrani, C., R. Troncone, P. Mugione, E. Cosentini, M. De Pascale, C. Faruolo,S. Senger, G. Terrazzano, S. Southwood, S. Auricchio, and A. Sette. 2003. Celiacdisease association with CD8� T cell responses: identification of a novel gliadin-derived HLA-A2-restricted epitope. J. Immunol. 170: 2719–2726.
25. Kwok, W. W., G. T. Nepom, and F. C. Raymond. 1995. HLA-DQ polymor-phisms are highly selective for peptide binding interactions. J. Immunol. 155:2468–2476.
26. Lee, K. H., K. W. Wucherpfennig, and D. C. Wiley. 2001. Structure of a humaninsulin peptide-HLA-DQ8 complex and susceptibility to type 1 diabetes. Nat.Immunol. 2: 501–507.
27. Moustakas, A. K., Y. van de Wal, J. Routsias, Y. M. Kooy, P. van Veelen,J. W. Drijfhout, F. Koning, and G. K. Papadopoulos. 2000. Structure of celiacdisease-associated HLA-DQ8 and non-associated HLA-DQ9 alleles in complexwith two disease-specific epitopes. Int. Immunol. 12: 1157–1166.
28. Trier, J. S. 1991. Celiac sprue. N. Engl. J. Med. 325: 1709–1719.29. Wieser, H., A. Modl, W. Seilmeier, and H. D. Belitz. 1987. High-performance
liquid chromatography of gliadins from different wheat varieties: amino acidcomposition and N-terminal amino acid sequence of components. Z. Lebens.Unters. Forsch. 185: 371–378.
30. De Ritis, G., S. Auricchio, H. W. Jones, E. J. Lew, J. E. Bernardin, andD. D. Kasarda. 1988. In vitro (organ culture) studies of the toxicity of specific A-gliadin peptides in celiac disease. Gastroenterology 94: 41–49.
31. Sturgess, R., P. Day, H. J. Ellis, K. E. Lundin, H. A. Gjertsen, M. Kontakou, andP. J. Ciclitira. 1994. Wheat peptide challenge in coeliac disease. Lancet 343:758–761.
32. Arentz-Hansen, H., R. Korner, O. Molberg, H. Quarsten, W. Vader, Y. M. Kooy,K. E. Lundin, F. Koning, P. Roepstorff, L. M. Sollid, and S. N. McAdam. 2000.The intestinal T cell response to �-gliadin in adult celiac disease is focused on asingle deamidated glutamine targeted by tissue transglutaminase. J. Exp. Med.191: 603–612.
33. Senger, S., D. Luongo, F. Maurano, M. F. Mazzeo, R. A. Siciliano, C. Gianfrani,C. David, R. Troncone, S. Auricchio, and M. Rossi. 2003. Intranasal adminis-tration of a recombinant �-gliadin down-regulates the immune response to wheatgliadin in DQ8 transgenic mice. Immunol. Lett. 88: 127–134.
34. Geluk, A., V. Taneja, K. E. van Meijgaarden, E. Zanelli, C. Abou-Zeid,J. E. Thole, R. R. de Vries, C. S. David, and T. H. Ottenhoff. 1998. Identificationof HLA class II-restricted determinants of Mycobacterium tuberculosis-derivedproteins by using HLA-transgenic, class II-deficient mice. Proc. Natl. Acad. Sci.USA 95: 10797–10802.
35. Khare, M., M. Rodriguez, and C. S. David. 2003. HLA class II transgenic miceauthenticate restriction of myelin oligodendrocyte glycoprotein-specific immuneresponse implicated in multiple sclerosis pathogenesis. Int. Immunol. 15:535–546.
36. Bristol, J. A., C. Orsini, P. Lindinger, J. Thalhamer, and S. I. Abrams. 2000.Identification of a ras oncogene peptide that contains both CD4� and CD8� Tcell epitopes in a nested configuration and elicits both T cell subset responses bypeptide or DNA immunization. Cell. Immunol. 205: 73–83.
37. Carreno, B. M., R. V. Turner, W. E. Biddison, and J. E. Coligan. 1992. Over-lapping epitopes that are recognized by CD8� HLA class I-restricted and CD4�
class II-restricted cytotoxic T lymphocytes are contained within an influenzanucleoprotein peptide. J. Immunol. 148: 894–899.
38. Van de Wal, Y., Y. M. Kooy, J. W. Drijfhout, R. Amons, and F. Koning. 1996.Peptide binding characteristics of the coeliac disease-associated DQ(�1*0501,�1*0201) molecule. Immunogenetics 44: 246–253.
39. Sollid, L. M. 2000. Molecular basis of celiac disease. Annu. Rev. Immunol. 18:53–81.
40. Qiao, S. W., J. Piper, G. Haraldsen, I. Oynebraten, B. Fleckenstein, O. Molberg,C. Khosla, and L. M. Sollid. 2005. Tissue transglutaminase-mediated formationand cleavage of histamine-gliadin complexes: biological effects and implicationsfor celiac disease. J. Immunol. 174: 1657–1663.
41. Gjertsen, H. A., K. E. Lundin, L. M. Sollid, J. A. Eriksen, and E. Thorsby. 1994.T cells recognize a peptide derived from �-gliadin presented by the celiac dis-ease-associated HLA-DQ (�1*0501, �1*0201) heterodimer. Hum. Immunol. 39:243–252.
8094 EPITOPES OF �-GLIADIN IN HLA-DQ8 MICE
by guest on April 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

42. Godkin, A. J., M. P. Davenport, A. Willis, D. P. Jewell, and A. V. Hill. 1998. Useof complete eluted peptide sequence data from HLA-DR and -DQ molecules topredict T cell epitopes, and the influence of the nonbinding terminal regions ofligands in epitope selection. J. Immunol. 161: 850–858.
43. Godkin, A., T. Friede, M. Davenport, S. Stevanovic, A. Willis, D. Jewell, A. Hill,and H. G. Rammensee. 1997. Use of eluted peptide sequence data to identify thebinding characteristics of peptides to the insulin-dependent diabetes susceptibilityallele HLA-DQ8 (DQ 3.2). Int. Immunol. 9: 905–911.
44. Kwok, W. W., M. L. Domeier, F. C. Raymond, P. Byers, and G. T. Nepom. 1996.Allele-specific motifs characterize HLA-DQ interactions with a diabetes-associ-ated peptide derived from glutamic acid decarboxylase. J. Immunol. 156:2171–2177.
45. Marinaro, M., H. F. Staats, T. Hiroi, R. J. Jackson, M. Coste, P. N. Boyaka,N. Okahashi, M. Yamamoto, H. Kiyono, H. Bluethmann, et al. 1995. Mucosaladjuvant effect of cholera toxin in mice results from induction of T helper 2 (Th2)cells and IL-4. J. Immunol. 5: 4621–4629.
46. Lavelle, E. C., E. McNeela, M. E. Armstrong, O. Leavy, S. C. Higgins, andK. H. Mills. 2003. Cholera toxin promotes the induction of regulatory T cellsspecific for bystander antigens by modulating dendritic cell activation. J. Immu-nol. 171: 2384–2392.
47. Nilsen, E. M., F. L. Jahnsen, K. E. Lundin, F. E. Johansen, O. Fausa, L. M. Sollid,J. Jahnsen, H. Scott, and P. Brandtzaeg. 1998. Gluten induces an intestinal cy-tokine response strongly dominated by interferon � in patients with celiac dis-ease. Gastroenterology 115: 551–563.
48. McMahon, E. J., S. L. Bailey, C. V. Castenada, H. Waldner, and S. D. Miller.2005. Epitope spreading initiates in the CNS in two mouse models of multiplesclerosis. Nat. Med. 11: 335–339.
49. Klehmet, J., C. Shive, R. Guardia-Wolff, I. Petersen, E. G. Spack, B. O. Boehm,R. Weissert, and T. G. Forsthuber. 2004. T cell epitope spreading to myelinoligodendrocyte glycoprotein in HLA-DR4 transgenic mice during experimentalautoimmune encephalomyelitis. Clin. Immunol. 111: 53–60.
50. McRae, B. L., C. L. Vanderlugt, M. C. Dal Canto, and S. D. Miller. 1995.Functional evidence for epitope spreading in the relapsing pathology of experi-mental autoimmune encephalomyelitis. J. Exp. Med. 182: 75–85.
51. Ott, P. A., M. T. Dittrich, B. A. Herzog, R. Guerkov, P. A. Gottlieb,A. L. Putnam, I. Durinovic-Bello, B. O. Boehm, M. Tary-Lehmann, andP. V. Lehmann. 2004. T cells recognize multiple GAD65 and proinsulin epitopesin human type 1 diabetes, suggesting determinant spreading. J. Clin. Immunol.24: 327–339.
52. Veeraraghavan, S., E. A. Renzoni, H. Jeal, M. Jones, J. Hammer, A. U. Wells,C. M. Black, K. I. Welsh, and R. M. du Bois. 2004. Mapping of the immuno-dominant T cell epitopes of the protein topoisomerase I. Ann. Rheum. Dis. 63:982–987.
53. Zuercher, A. W., S. E. Coffin, M. C. Thurnheer, P. Fundova, and J. J. Cebra.2002. Nasal-associated lymphoid tissue is a mucosal inductive site for virus-specific humoral and cellular immune responses. J. Immunol. 168: 1796–1803.
54. Prakken, B. J., R. van der Zee, S. M. Anderton, P. J. van Kooten, W. Kuis, andW. van Eden. 1997. Peptide-induced nasal tolerance for a mycobacterial heatshock protein 60 T cell epitope in rats suppresses both adjuvant arthritis andnonmicrobially induced experimental arthritis. Proc. Natl. Acad. Sci. USA 94:3284–3289.
55. Rossi, M., F. Maurano, N. Caputo, S. Auricchio, A. Sette, R. Capparelli, andR. Troncone. 1999. Intravenous or intranasal administration of gliadin is able todown-regulate the specific immune response in mice. Scand. J. Immunol. 50:177–182.
8095The Journal of Immunology
by guest on April 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from


![Phosphorylation-dependentepitopes antibodies Alzheimertau · ment antibodies SMI31, SMI34, SMI35, or SMI310 (with phosphorylated epitopes) and SM133 [unphosphorylated epitopes (3)].](https://static.fdocuments.in/doc/165x107/5e62d2f4d3d32f22a55ed9e3/phosphorylation-dependentepitopes-antibodies-alzheimertau-ment-antibodies-smi31.jpg)















