![07/03/18 - UJS Homerecords, Dr. Bell noted: “[Lagler] is in for follow-up on her foot pain. She’s really not doing any better. She’s still complaining of a tremendous amount](https://static.fdocuments.in/doc/165x107/602302fc7197f8380777fa7d/070318-ujs-home-records-dr-bell-noted-aoelagler-is-in-for-follow-up-on.jpg)
Ictiologia. lagler
498
Ictiología PRIMERA EDlCldN EN ESPAfilOL Karl F. Lagler Escuela de Recursos Naturales Universidad de Michigan John E. Bardach Instituto Hawaiano de Biología Marina Universidad de Hawai Robert R. Miller Museo de Zoología Universidad de Michigan Dora R. May Passino Laboratorio de Pesquerías de los Grandes Lagos Dept. de Pesca y Vida Silvestre de EUA m FlGT EDITOR, SA.
description
Transcript of Ictiologia. lagler

Ictiología
PRIMERA EDlCldN EN ESPAfilOL
Karl F. Lagler Escuela de Recursos Naturales
Universidad de Michigan
John E. Bardach Instituto Hawaiano de Biología Marina
Universidad de Hawai
Robert R. Miller Museo de Zoología
Universidad de Michigan
Dora R. May Passino Laboratorio de Pesquerías de los Grandes Lagos
Dept. de Pesca y Vida Silvestre de EUA
m FlGT EDITOR, SA.

Titulo del original en inglés: Ichthyology Copyright @ 1977 (2a. Ed.), John Wiley & Sons, Inc.
Primera edición en español Primera reimpresión 1990
-
@A.G.T. Editor, S.A. Progreso 202 - Planta Alta México 18, D.F.
Traducción: Marcos Arellano Armenta. Biólogo (Escuela Nacional de Ciencias Biológicas del IPN) Licenciado en Ciencias Naturales (Escuela Normal Superior de
Celaya, Gto.). Profesor de Ciencias Naturales a nivel de enseñanza media básica
en la SEP. Profesor de Zoología IV (Cordados) en la Facultad de Ciencias
de la UNAM. Becario del Wildlife Management Institute para estudiar Vida
Silvestre en la Escuela de Recursos Naturales de la Universidad de Michigan.
El cuidado de la presente edición y la portada estuvieron a cargo del Taller Gráfico Macehual.
Reservados todos los derechos. Ninguna parte de este libro puede ser reproducida, almacenada en un sistema de informática o transmitido en cualquier forma o por cualquier medio electrónico, mecánico, fotocopia, grabación u otros métodos, sin previo y expreso permiso del propietario del copyright.
Impreso y hecho en México, 1984 Printed and made in Mexico ISBN: 968-463-017-4

Dedicado a Carl L. Hubbs,
erudito, maestro y amigo

Prefacio
Resumiendo los prefacios de los respectivos autores para la primera y segunda edi- ción en inglés, es de destacarse el esfuerzo hecho por los mismos, para que este texto se adapte fácilmente a los cursos de licenciatura, y que su material sirva como lectura medular tanto en los cursos de introducción como en los más avan- zados sobre ictiología.
“Sin embargo -aclaran sus autores en el prefacio de la primera edición-, se debe poner atención a través de la selección de los capítulos y secciones asignadas para hacerlo también útil en los cursos de anatomía comparada de los vertebrados, historia natural de los vertebrados, fisiología comparada y evolución. Además -agregan-, hemos puesto énfasis en las comparaciones que se pueden hacer entre los grupos más importantes de peces, tanto en lo que respecta a su estructura como a su función.”
A continuación, en sus puntos más destacados especifican: “Los tres primeros capítulos introducen al lector en el conocimiento de la
diversidad de los peces y le muestran la posición filogenética y conformación de los grupos superiores, su clasificación, relaciones entre s í y estructura básica, prin- cipalmente en los peces vivientes. Los capítulos 4 al 11 describen y analizan la anatomía comparada, así como la fisiología también comparada de los diez siste- mas clásicos del cuerpo y su integración en todo el cuerpo del pez. Los principios de genética, evolución, sistemática, ecología e ictiogeografía son abordados en los capítulos 12 al 14. Por lo tanto, un curso a nivel de licenciatura sobre ictiología sistemática y ecológica debe utilizar primordialmente los capítulos 1 al 3 y 12 al 14, mientras que uno dedicado al estudio de la conducta y la fisiología de los peces debe remitirse a los capítulos 4 al 11. ”
“Desde que apareció en 1905 la venerable Guía para el estudio de los peces (Guide to the Study o f Fishes) de David Starr Jordan -apuntan los autores- no se había publicado un texto sobre ictiología cuyos ejemplos consideraran primor- dialmente a la fauna de América del Norte. Nosotros hemos tratado de llenar este vacío” y , en su segundo prefacio, en el cual destacan la incorporación de la Dra. Dora R. May Passino especializada en bioquímica ictiológica ‘gracias a la generosa
ix

autorización de U.S. Fish and Wildlife Service’ enfatizan lo siguiente: “En los años que han pasado desde que la primera edición fue publicada, muchos estudios han mejorado el conocimiento de la ictiología. De éstos, una buena proporción ha sido hecha en distintos lugares del orbe y si bien nosotros hemos aprovechado estos conocimientos, preferimos seguir empleando ejemplos norteamericanos.”
En sus reconocimientos agradecen el auxilio recibido de estudiantes y colegas y , en especial del Dr. Gerald P. Smith por su ayuda en la revisión del capítulo 2, referido a los grupos superiores de peces, y del Dr. Paul R. Webb por sus sugeren- cias sobre la locomoción, tema que aborda el capítulo 6. Asimismo agradecen a: Elsie Goode, Frances Hubbs Miller, Joanna Schmidt, Mary Lou Lagler y Donna Brown, al editor Wiley, Robert L. Rogers y al supervisor de producción, Christine Pines.
EL EDITOR
X

Contenido
CAPfTULO 1 Peces, animales y hombre
Qué son los peces, 1. Los peces como ancestros del hombre, 2. Dónde viven los peces, 3. Cómo viven los peces, 3. Cómo y por qué son estudiados los peces, 5 . Oportunidades en ictiología, 6 . Referen- cias, 7.
CAPfTULO 2 Los grupos superiores de peces
1
9
Alcances de la clasificación, 9. Propósitos de la cla- sificación, 10. Métodos de clasificación, 10. Los ancestros de los vertebrados, 11. Grupos superiores de peces vivientes, 13. Caracterización de los gru- pos superiores de peces vivientes, 15. Grupos supe- riores de peces extinguidos, 17. Relaciones de los grupos superiores de peces, 21. Lista de las familias comunes representativas de peces vivientes, 30. Referencias, 48.
CAP~TULO 3 Anatomía básica de los peces 51
Anatomia externa general,51. Esqueleto,57. Múscu- los, 70. Branquias y vejigas gaseosas, 77. Tracto digestivo, 81. Sistema circulatorio, 85. Riñones, 87. Glándulas reproductoras - Gónadas, 89. drganos endocrinos, 91. Sistema nervioso, 92. Referen- cias, 99.

CAPfTULO 4 Piel 101
Estructura, 101. Las escamas, 102. Barbillas y ale- tas, I 1 1 . Coloración, I I 1. órganos luminosos, 122. Glándulas venenosas, 123. Referencias, 124.
CAPfTULO 5 Alimentos, digestión, nutrición y crecimiento
El alimento natural de los peces, 125. Hábitos ali- menticios, 129. Digestión, absorción y utilización de los alimentos, 145. Nutrición de los peces, 150. Crecimiento de los peces, 160. Referencias, 166.
CAPfTULO 6 Esqueleto, estructura y movimiento
Esqueleto, 167. Locomoción, 174. Forma del cuer- po y locomoción, 176. Aletas y locomoción, 177. Miómeros y natación, 181. Natación, 182. Loco- moción no natatoria, 185. Velocidad de recorrido, 187. Migraciones, 189. Referencias, 191.
CAPfTULO 7 Sangre y circulación
Corazón y vasos circulatorios, 194. Sangre, fluidos tisulares y órganos formadores de la sangre, 207. Temperatura del cuerpo, 21 3. Referencias, 21 3.
CAPfTULO 8 Respiración
Estructura y función de las branquias, 216. La san- gre de los peces como transportadora de gas, 224. Adaptaciones para la respiración del aire entre los peces, 229. La vejiga gaseosa, 233. Referencias, 245.
CAPfTULO 9 Excreción y regulación osmótica
Órganos osmorreguladores y excretores, 247. Con- trol endocrino de la excreción y osmorregulación, 260. Referencias, 262.
CAPfTULO 10 Reproducción
125
167
193
215
247
263
Tipos de reproducción, 263. El sistema reproduc- tor, 264. Los espermatozoides y su formación, 264.

Los óvulos y su formación, 266. Diferencias sexua- les, 269. Maduración sexual, 273. Ciclos reproduc- tores, 274. Reproducción, 275. Cuidado de los huevos y de la cría, 280. Desarrollo, 284. Referen- cias, 303.
CAF'fTULO 11 Integración 305
El papel de los sistemas nervioso y endocrino,305. El sistema nervioso, 306. L a médula espinal, 31 7. Los nervios espinales, 320. El sistema nervioso autó- nomo, 321. Tejidos de sostén del sistema nervioso central, 322. Inteligencia y comportamiento, 323. Órganos endocrinos, 329. Los sentidos (órganos de los sentidos) de los peces, 342. Referencias, 369.
CAPfTULO 12 Genética y evolución 373
La herencia en los peces, 373. Determinación del sexo, 377. Hibridación, 379. Mecanismos de la evolución, 381. Mecanismos de aislamiento, 384. Evolución paralela o convergencia, 386. Duración del proceso evolutivo, 387. Referencias, 387.
CAPfTULO 1 3 Sistemática y nomenclatura 389
Disponibilidad apropiada de los peces, 390. Ante- cedentes históricos, 391. La tarea de los sistemáti- cos, 392. Conceptos taxonómicos, 393. Los datos para la clasificación, 394. Colecciones de estudio, 395. Nomenclatura zoológica, 397. Referencias, 401.
CAPfTULO 14 Ecología y zoogeografía 403
Definiciones, 403. Agua, 404. Productividad orgá- nica en los ecosistemas acuáticos, 404. Clasifica- ciones ecológicas de los peces, 406. Factores ecoló- gicos, 406. Ecosistema marino, 415. Ecosistema estuarino, 421. Ecosistema de agua dulce, 422. Zoogeografía, 428. Geografía de los peces de agua dulce, 432. Geografía de los peces marinos, 442. Bipolaridad, 44 7. Referencias, 44 7.
Índice Sistemático
Índice Temático
451
469

Capítulo 1
Peces, animales, y hombre
QUk SON LOS PECES
Los peces son animales de sangre fría, caracterizados por poseer vértebras, bran- quias y aletas (en lugar de miembros pentadáctilos), y dependen primordialmente del agua, que es el medio en donde viven. Su estudio comprende los aspectos puro y aplicado de la ciencia ictiológica. Obviamente no están incluidos en este campo del conocimiento los mamíferos como las ballenas, las focas y los delfi- nes; reptiles como las tortugas acuáticas, e invertebrados como los moluscos bivalvos, camarones y langostas de mar.
Los peces son los vertebrados más numerosos (Figura l.l), estimando que existen cerca de 20,000 especies vivientes, aunque se piensa que podrían ser hasta 40,000. En contraste, se calcula que el número de especies de aves se acerca a 8,600; de mamíferos a 4,500 (en donde la del hombre es sólo una de ellas); de reptiles a 6,000 y de anfibios a 2,500. No solamente hay muchos peces diferen- tes, sino que también sus especies son de muchos tamaños y formas distintas. Los hay desde verdaderos peces miniatura como Etheostoma microperca,' cuya maduración sexual ocurre cuando su longitud es de solamente 27 mm, y Euiotu sp. del Pacífico que se reproduce sin llegar siquiera a los 15 mm de longitud, hasta gigantes como el tiburón ballena (Rhincodon) del que se ha llegado a supo- ner que alcanza a veces una longitud de hasta 21 m y un peso de 25 o más tonela- das. La mayoría de los peces tienen forma de torpedo, aunque los hay redondos o cilíndricos, otros son planos y algunos angulares.
1 (American percid least darter).
1

2 PECES, ANIMALES Y HOMBRE
Figura 1.1 -Composición porcentual por grupos de las casi 41,600 especies de vertebrados recientes.
LOS PECES COMO ANCESTROS DEL HOMBRE
De acuerdo con la teoría de la evolución de las especies que se basa en evidencias, entre las cuales se incluyen las que dan los fósiles, la anatomía comparada, la embriología y la genética, el origen de los peces está muy distante de los antepa- sados del hombre. Su presencia en la Tierra antecede a la del hombre-mono en cerca de 500 millones de años; a la de los demás vertebrados en poco más de 100 millones. De no haber tenido un ancestro marino el hombre posiblemente nunca hubiera aparecido. Muchos avances fisiológicos notables y adaptaciones anatómicas del hombre fueron originadas, o estaban ya presentes desde hacía ya mucho tiempo, en los antepasados pisciformes. Estos desarrollos se han dado en los planes fundamentales y funciones básicas de los diez sistemas de órganos, incluyendo aquellos tan importantes como la visión, la fertilización interna, el

DdNDE VIVEN LOS PECES 3
desarrollo intrauterino (incluyendo la placenta), la viviparidad y, seguramente, la capacidad de aprender y memorizar.
DONDE VIVEN LOS PECES
Realmente no es sorprendente que haya tanta variedad de peces si tomamos en cuenta la antigüedad del grupo y la gran extensión y variedad que presenta su hábitat. Hasta el presente, más del 70% de la superficie de la Tierra está cubierta de agua (Figura 1.2). Cuando la clase de los peces iniciaba su proceso evolutivo había incluso más agua porque gran parte de la corteza terrestre que hoy es con- tinental, era en aquel tiempo fondo marino. Es de esperar que el aumento de la diversidad de las condiciones que permiten la vida en el agua favorezca una ma- yor especiación.
Aparentemente, los peces fueron capaces de adaptarse a los sucesivos cambios ecológicos que tuvieron lugar en donde residían y ahora pueden vivir casi en cualquier lugar donde haya agua, ya sea superficial o subterránea comu- nicada con el exterior. Habitan en muy diversos lugares, desde las aguas del Antártico cuya temperatura está por debajo del punto de congelación, hasta los manantiales de los que brota agua a más de 40°C; y desde el agua dulce y blanda, hasta en depósitos donde el agua es mucho más salada que la del mar. Están pre- sentes en corrientes fluviales que bajan de montañas soleadas, con aguas tan torrenciales que ni el hombre ni los perros pueden vadearlas o nadar en ellas, y en aguas tan quietas, profundas y oscuras que nunca han sido habitadas por otros vertebrados o exploradas en su totalidad por el hombre. Los límites de su dis- tribución vertical exceden en distancia a los de cualquier otro vertebrado; viven desde aproximadamente 5 km sobre el nivel del mar hasta cerca de los 11 km por debajo de éste (Figura 1.2).
C ~ M O VIVEN LOS PECES
El agua es vía rápida de circulación, calle lateral, medio general de comunicación, guardería infantil, patio de recreo, escuela, recámara, lecho, despensa, bebede- ro, sanitario y tumba para un pez. Todas las funciones vitales de los peces como la alimentación, la digestión, la asimilación, el crecimiento, las respuestas a los estímulos y la reproducción, dependen del agua. Para el pez, los aspectos más importantes y básicos del agua, para su exterminio o supervivencia, son: la pre- sencia de oxígeno disuelto, las sales en solución, la penetración de la luz, la tem- peratura, la presencia de sustancias tóxicas, la concentración de organismos infecciosos y la oportunidad de poder escapar de los enemigos.
Aunque los humanos somos capaces de absorber oxígeno directo del aire a través de paredes vascularizadas de los pulmones, pocos peces tienen pulmones u otros medios para aprovechar el oxígeno del aire. La mayor parte de los peces, incluyendo a los pulmonados, dependen primordialmente de las branquias para utilizar el oxígeno disuelto en el agua. Los peces no pueden vivir largo tiempo en un hábitat deficiente en oxígeno disuelto, como los humanos tampoco pueden sobrevivir en la exósfera o el espacio estelar a menos que lleven consigo un equipo especial que les proporcione dicho gas para respirar.

4 PECES, ANIMALES Y HOMBRE
I OCÉAN t
Figura 1.2 - Área de distribución de los peces y su o,sposición vertical, con respecto esta última a las máximas dibrencias entre la altura y la profundidad del relieve de la Tierra.
La “pastura” que el mar, los lagos y los arroyos proporcionan a los peces, depende en primer lugar de la cantidad de luz que penetre en el agua, así como sucede con el pasto que brota en campo abierto gracias a la luz solar que recibe. El “pasto” de las aguas está formado de vida vegetal microscópica, diatomeas y algas, denominadas en conjunto como fitoplancton.
El primer eslabón de la cadena alimenticia relacionada con la producción de peces consiste generalmente en el cuerpo de cada uno de los componentes del fitoplancton. Éstos utilizan la energía luminosa y el dióxido de carbono disuelto para formar materia orgánica que eventualmente se transforma en alimento para los peces. Además de proporcionar energía para la producción de alimento para to-

COMO Y POR QUg SON ESTUDIADOS LOS PECES 5
dos los peces, se sabe que la luz también es activadora de los mecanismos de reproducción, crecimiento y muchas clases de conductas, incluyendo la relacio- nada con la alimentación.
Los materiales indeseables, como las toxinas producidas en la naturaleza y la contaminación procedente de las actividades humanas, constituyen serias amenazas para la vida de los peces. El hábitat acuático no proporciona vías de escapatoria a los peces de las sustancias daííinas en solución. Los materiales tóxicos que se depo- sitan en el agua son para los peces una amenaza comparable a la que para el ser humano constituye la presencia de los contaminantes suspendidos en el aire. Aunque los peces son capaces de detectar muchos de esos contaminantes quími- cos, a menudo no pueden evitar su acción dañina.
Como ocurre con todos los animales, los peces también tienen que conten- der con buena cantidad de enfermedades. Muchas de éstas son producidas por agentes externos; otras tienen origen interno. Entre los externos se encuentran los virus, hongos, bacterias, protozoarios parásitos, gusanos, crustáceos y lam- preas. Los desórdenes orgánicos y degenerativos que enferman al hombre son los que en los peces aparecen como factores internos que, finalmente, les pueden producir la muerte. Tales son el cáncer, las ricketsias, la degeneración del hígado, la ceguera y una buena variedad de anomalías del desarrollo, como la aparición de gemelos siameses y la flexión de la columna vertebral. Aun cuando no se vean atacados por enfermedades o los desórdenes que se acaban de mencionar, los peces todavía tienen que defenderse contra periódicas condiciones químicas adversas del agua, los predadores y las actividades piscícolas del hombre.
CdMO Y POR QUh SON ESTUDIADOS LOS PECES
El incremento del interés por el conocimiento de la vida de los peces ha sido el resultado del natural deseo que se nos presenta por saber más sobre la naturaleza y de nuestra necesidad de recabar más información relacionada con las especies que nos sirven para el comercio y la recreación. Por lo menos desde el siglo X a.c. ya los chinos estaban tratando de cultivar con éxito los peces. Los antiguos egipcios, griegos y romanos hicieron registros sobre las variedades, hábitos y cua- lidades de varias especies de peces. El símbolo del antiguo movimiento cristiano que tuvo lugar en las catacumbas de Roma fue el pez.
El estudio de los peces, o ictiología, no obtuvo forma de disciplina cien- tífica sino hasta el siglo xVm en Europa. Desde ese entonces se ha desarrollado rápidamente en todo el mundo, en torno a las siguientes especialidades:
Taxonomía: el continuo esfuerzo por ubicar toda clase de pez vivo y fósil en grupos o taxa para determinar sus relaciones naturales.
Anatornia: la estructura de los peces desde el punto de vista de la micros- copia, la embriología y la comparación, incluyendo a los fósiles para llenar la solución de continuidad en las líneas evolutivas, siempre que sea pertinente y haya buenos ejemplares.
Evolución y genética: los diversos orígenes de los peces y la secuencia y modo como los peces modernos se desarrollaron a partir de sus antepasados, así como los mecanismos que dieron lugar a tales cambios evolutivos; especiación, origen, transmisión y cambios en los caracteres mediante los cuales las diversas especies pueden ser identificadas.

6 PECES, ANIMALES Y HOMBRE
Historia rtatural y ecologíu: ciclos de vida y hábitat~, y las interrelaciones de los peces entre s í y con e1 medio ambiente.
Fisiologia y bioquirnica: llunciones de los órganos y sistemas, el metabo- lismo y la int.egración de sistemas a nivel molecular, y tolerancias a los cambios de las condiciones físicas del media ambiente.
Conseruacicin: uso adecuado y administración correcta de los recursos pesqueros para beneficio del hombre mediante estadísticas de pesqueria.s, tec- nologia y mercadotecnia pesqueras, leyes, manejo de poblaciones, cultivo y alma- cenamiento de pescado y mejoramiento del ambiente.
Esas ireas de trabajo están a cargo de organizaciones internacionales, agen- cias gubernamentales, museos, universidades e industrias. La Organización de las Xaciones Unidas para la Alimentación y la Agricultura ( F A O ) tiene una División de Pesca muy activa. La mayor parte de los paises tienen departamentos federa- ?.es de pesca comparables al Departamento de Pesca y Vida Silvestre y al Departa- mento de Pesquería de la Marina Nacional de los Estados Unidos de Norteamérica,* y los estados o provincias a menudo tienen secciones especializadas de pesca en sus oficinas relacionadas con los recursos naturales. También museos y academias que abordan estudios científicos en sus programas de trabajo tienen secciones de pesca. Entre estas instituciones se destacan: el Museo Británico (Historia Natural), el Museo Nacional de los Estados Unidos, la Academia de Ciencias de California y el Museo de Zoología de la Universidad de Michigan. Gran número de institucio- nes de educación superior ofrecen cursos sobre ictiología o pesquerías o las dos disciplinas.
OPORTUNIDADES EN ICTIOLOGÍA
Las oportunidades para estudiar los peces no tienen límite y cualquier persona puede aprovecharlas, sea o no ictiólogo profesional. Muchas contribuciones para el conocimiento de los peces han sido aportadas por filósofos, clérigos, doctores en medicina, pescadores o aficionados a la ictiología que cultivan peces en peque- ños acuarios. Las oportunidades que se presentan para la investigación son enor- mes. Es muchísimo más lo que se desconoce en todos los aspectos de la ictiología, que lo que se sabe.
Las plazas para maestros de ictiología no abundan; pocos son los que inves- tigan y se dedican a la enseñanza en esta rama de la ciencia. Las oportunidades para encontrar colocación como curadores en los museos con el fin de aumentar, cuidar y estudiar las colecciones de peces son todavía menores. Hay solamente unos 20 museos en Norteamérica, por ejemplo, que tienen ictiólogos formando parte de su cuerpo de investigadores, y sólo hay entre 35 y 50 puestos de curador. Las obligaciones consisten en el enriquecimiento, cuidado y estudio de las colec- ciones antiguas y recientes, la supervisión de las exhibiciones públicas, la atenci6n a las consultas y la contestación de la correspondencia enviada por el público, así como la preparación de artículos relacionados con la investigación científica para su publicación. Muchos curadores (todos los de los museos que están asociados con instituciones educativas) colaboran en los programas de entrenamiento para
*Fish and Wildlife Service y National Marine Fisheries Service, respectivamente. (N. del T.)

REFERENCIAS 7
graduados y otros aspectos de la enseñanza. Las plazas para curadores no se pre- sentan con frecuencia; posiblemente su promedio es de una por año* y hay una gran competencia para conseguirlas, para lo cual se requiere generalmente un grado de Doctor en Filosofía. En años recientes, sin embargo, se ha creado un empleo técnico denominado Administrador de Colecciones que exige mucho menos entrenamiento formal.
Las grandes empresas comerciales o pesquerías, la administración profesio- nal de la pesca, la distribución y venta de alimento y todo lo relacionado con los peces usados para el deporte, ornamento y cebo, son vastos campos y oportu- nidades de trabajo para todos los que tengan entrenamiento y experiencia en ictiología. Estos trabajos no requieren más que un grado de Maestro en Ciencias, cuando mucho. El personal debidamente entrenado es necesario en la adminis- tración de pesquerías de las aguas marinas y continentales y también en viveros y estaciones para la reproducción inducida de peces. La ampliación de las áreas de trabajo en la rama de las pesquerías ofrece muchas oportunidades de empleo en esos tópicos que no habían sido previamente investigados. Se sabe que en los años setenta ha habido un gran resurgimiento de puestos en departamentos priva- dos y gubernamentales relacionados con el creciente deterioro del medio ambiente. Y de todas las oportunidades para trabajar con peces, la más popular para los estudiantes es la que se refiere al cultivo en acuarios. Aquí, la ciencia de la ictio- logia tiene mucho que ofrecer para el bienestar de todos.
REFERENCIAS GENERALES
American Fisheries Society. 1970. Fisheries as a profession. Es una guía de carreras para el campo de la ciencia de las pesquerías. Es distribuida por la Sociedad.
American Society of Ichthyologists and Herpethologists. 1973. Oportunidades de carreras para los ictiólogos. Preparado por Victor G. Springer y distribuido por la Sociedad.
Boulenger, G.A. 1904. Fishes (systematic account of Teleostei). En The Cambridge natural history. vol. VII. The Macmillan Company, Nueva York. Reimpresión 1958. Hafner Publishing Company, Nueva York.
Bridge, T.W. 1904. Fishes (exclusive of the systematic account of Teleostei). En The Cam- bridge natural history. vol. VIL The Macmillan Company, Nueva York, Reimpresihn 1958, Hafner Publishing Company, Nueva York.
Brown, M.E. (ed.). 1957. The physiology o f fishes. vol. 1 Metabolism. vol. 2. Behavior. Acade- mic Press, Nueva York.
Curtis, B. 1949. The life story o f the fish his morals and manners. Harcourt, Brace and Com- pany, Nueva York.
Dean, B. 1895. Fishes, living and fossil. The Macmillan Company, Nueva York. Goodrich, E.S. 1909. Cyclostomes and fishes. En A treatise on Zoology. Lám. IX. Fas. 1. A.
Grassé, P.P. (ed.). 1958. Agnathes et poissona anatomie, éthologie, systernatique. Trait6 de
Giinther, A.C.L.G. 1880. A n introduction to the study o f fishes. A. and C . Black. Edimburgo. Herald, E.S. 1961. Living fishes o f the world. Doubleday & Company Inc.. Garden City,
Hoar, W.S., y D.J. Randall. i969-1976. Fish physiology. Academic Press, Nueva York y Idon-
Jardan, D.S. 19CIi. -4 guide fo the s tudy of f ishes. 2 vols. Henry Holt and Company, Nueva
and C. Black, Londres,
Zoologie. Torne XIII, 3 vols. Masson et Cie., París.
Nueva York.
dres. 8 vols.
York.
*En los Estados Unidos de Norteamérica. (N. del T. j

8 PECES, ANIMALES Y HOMBRE
Kyle, H.M. 1926. The biology o f fishes. Sidgwick and Jackson, Londres; Macmillan Company,
Lanham, U. 1962. The fishes. Columbia University Press, Nueva York. Marshall, N.B. 1965. The life o f fishes. Weidenfeld and Nicolson, Londres. Matsubara, K. 1955. Fish morphology and hierarchy. Ishizaki Shoten. Tokio. Myers, G.S. 1970. How to become an ichthyologist. T.F.H. Publications, Jersey City, N.J. Nikolski, G.W. 1957. Spezielle Fischkunde. VEB Deutscher Verlag der Wissenschaften, Berlín.
(También: 1961, Special ichthyology, Programa Israelita para Traducciones Científicas, Jerusalén).
Norman, J.R. 1975. A history of fishes. Tercera edición por P.H. Greenwood. John Wiley and Sons, Nueva York.
Pimher, C. 1948. A s tudy o f f i sh . Duell, Sloan and Pearce, Nueva York. Roule, L. 1935. Fishes and their ways of life. W.W. Norton and Company, Nueva York. Schultz, L.P., y E.M. Stern. 1948. The ways of fishes. D. Van Nostrand Company, Princeton,
Sterba, G. 1959. Süsswasserfische aus aller Welt . Verlag Zimmer & Herzog, Berchtesgaden. Young, J.Z. 1962. The life ofvertebrates. (2da. ed.) Oxford University Press, Londres.
Nueva York.
N.J.

Capítulo 2
Los grupos superiores 1
de peces
La rama de la ictiología ha sido definida como la ciencia que estudia los peces y, a su vez, éstos han sido definidos, en términos generales, vertebrados acuáticos de respiración branquia1 y provistos de aletas. Aparte de esta generalización, la diver- sidad tan grande de formas, tamaños y variedades en los peces, así como su dife- rente distribución en el tiempo y el espacio es tan enorme que se han encontrado muchas dificultades para procurar su clasificación. Además, relativamente pocos científicos contemporáneos tienen el suficiente acervo de conocimientos bastante profundos como para abordar siquiera el problema de la clasificación, La falta de un acuerdo aceptable que relacione los diferentes esquemas de clasificación que se han propuesto en el presente siglo señala, desde luego, que existen muchas lagunas, es decir, que aún se ignora mucho en el conocimiento de los peces; y, además, explica la razón de las controversias que entre los científicos se presentan cuando se habla de esos animales.
ALCANCES DE LA CLASIFICACIdN
El material que debe ser clasificado comprende todos los miles de clases de peces vivos y fósiles que han sido descubiertos y todos los que aún son desconocidos para la ciencia. Es seguro que casi en cualquier lugar de la Tierra se espere encon- trar nuevas especies para ser descritas y clasificadas. Como es sabido, esos peces incluyen algunos sin mandíbulas, de los cuales unos son vivientes y desprote- gidos (lampreas y los del género Myxine' ), y otros son arcaicos y armados, o sea,
Mixinoa
9

10 LOS GRUPOS SUPERIORES DE PECES
con gruesas escamas protectoras; así como los demás peces mandibuiados, algunos extinguidos y otros vivientes (tiburones y peces óseos).
PROPÓSITOS DE LA CLASIFICACION
Con el fin de obtener una visión panorámica de las muchas, muchas clases de peces que han aparecido en la Tierra, el estudiante debe atender al problema de dividirlos en grupos para poderlos estudiar. La solución a este problema re- quiere la formulación de conceptos que faciliten la formación de grupos. La enunciación de estos conceptos es auxiliada por una definición de los posibles medios empleados en la clasificación. Por lo menos existen dos: 1) el práctico, y 2) el ideal.
El medio práctico consiste simplemente en ordenar todas las clases de peces de tal modo que sea posible localizar las que hayan sido manejadas antes, como sucede cuando se disponen en orden alfabético los ejemplares colocados en envases de vidrio que se exhiben en los anaqueles; o la información sobre peces contenida en hojas de papel almacenadas en un fichero. Un método así de funcional podría ser adoptado para facilitar en lo posible la distinción de las diferentes clases de peces. Entonces, el procedimiento básico para una clasifi- cación práctica deberá consistir, fundamentalmente, en un método operativo, pudiendo hacerse distribuyendo los peces por tamaños, alfabéticamente, o de alguna otra manera útil.
Si al aspecto utilitario del método que nosotros sigamos para hacer la cla- sificación, le agregamos nuestra idea de que debe ser tomada en cuenta la historia evolutiva de los peces, habremos llegado al aspecto ideal, al método perfecto. En este caso el sistema es conveniente porque es sistemático y conceptual, e imparte una expresión condensada de la filogenia del grupo más importante de organis- mos, El método ideal, por lo tanto, consiste en que se clasifique a los peces si- guiendo los postulados de la evolución de las especies, la genética, o lo natural por lo que se refiere a los caracteres, en lugar de basarse en algo meramente arti- ficial o mecánico. Siempre existen lagunas en el conocimiento, sin embargo, que hacen difícil cumplir con nuestro ideal. El resultado es que las clasificaciones modernas de los peces, aunque se basen en métodos ideales, siempre son parcial- mente artificiosas.

LOS ANCESTROS DL LOS VERTEBRADOS 11
Existe uniformidad no solamente para las categorías mayores consideradas en la clasificación; también la hay en los acuerdos tomados para formar los nom- bres específicos, de género y familia (ver Capítulo 13). Se carece, sin embargo, de reglas para la denominación de filum, clase y orden. A este nivel es donde mayor confusión existe en relación a los nombres de los grupos (ver Tabla 2.1); la falta de una práctica previamente establecida impide a los alumnos tener una idea inmediata sobre los grupos naturales de peces.
LOS ANCESTROS DE LOS VERTEBRADOS
Los siguientes animales son a menudo considerados como peces en el grupo, o Phylum Chordata, sobre la base de la presencia de la notocuerda como caracte- rística común (de aquí que sean llamados cordados):
Carecen de cráneo (Acrania) y vértebras (Protochordata) Balanoglosos (como Balanoglossus) Ascidias o tunicados (como Molgula) Lancetas (como anfioxus, Brunchiostoma)
Poseen cráneo (Craniata) y vértebras (Vertebrata) Peces (Pisces y otros animales relacionados) Anfibios (Amphibia) - ranas, sapos, salamandras y cecilias. Reptiles (Reptilia) - lagartijas, víboras, tortugas, cocodrilos. Aves (Aves) Mamíferos (Mammalia)
Los grupos de cordados vivientes que nos llevan a los peces y los incluyen, han sido a menudo denominados como sigue:
Phyllum Chordata Subphyllum Hemichordata (balanoglosos)
Subphyllum Cephalochordata (lancetas) Subphyllum Vertebrata (desde los peces
Subphyllum Urochordata (ascidias o tunicados) protocordados colectivamente,
a los mamíferos)
Es posible que los ancestros de los vertebrados se hayan encontrado entre los protocordados y por lo general no tuvieron estructuras esqueléticas duras. El más remoto antepasado metazoario de los protocordados y los vertebrados debe haber sido “una criatura simple y sedentaria compuesta de apenas un tracto diges- tivo y un órgano para capturar partículas alimenticias del agua que le rodeaba. . .” (Romer, 1968). Los tipos más primitivos de cordados, cuyas formas adultas vivían sin duda alguna en el fondo de los mares y las larvas, ciliadas, nadaban libremente, como la tornaria de algunos hemicordados, tienen ese tipo de estructura. El si- guiente paso en el desarrollo hacia uri cordado superior pudo haber consistido en la aparición de un sistema de aberturas branquiaies y alirnentaci6n por filtración que todavía es encontrado en vertebrados vivientes corno la larnprea en su forma larval y en otros animales. Del estado larval amocete debió haber pasado a otro en el que su forma era ya de pez, con desarrollo directo.

I r 1
LOS GRUPOS SUPERIORES DE PECES
I
E

GRUPOS SUPERIORES DE PECES VIVIENTES 13
La pregunta de si los primeros vertebrados fueron habitantes de aguas dulces o marinas ha sido repetidamente debatida. El problema es dónde se origi- naron los Agnatha, desde el momento que es generalmente aceptado que los primeros craneados pertenecían a este grupo. Una reciente confirmación de la teoría del origen marino de los ágnatos es dado por Lehtola (1973), quien des- cribe restos del heterostrácodo Astraspis desiderata de las areniscas de fondos marinos poco profundos del Ordovícico Medio en Ontario, Canadá. Este es el primer registro de un vertebrado del Ordovícico en arenisca, una roca depositada en mar abierto. Su investigación revisa la literatura relacionada con este tema tan controvertido.
Muchas teorías sobre la evolución de los cordados han sido propuestas y la mayoría han sido desechadas. La notable similitud estructural entre ciertos equi- nodermos del Ordovícico (mitrados) y los heterostrácodos ha llevado a una nueva y también controvertida teoría sobre el origen de los vertebrados. Los mitrados son casi simétricos y se cree que tuvieron notocuerda, un cordón nervioso dorsal y un sistema nervioso central complejo parecido al de los ostracodermos, además de aberturas branquiales en la faringe como las de los hemicordados. Los primeros heterostrácodos (Figura 2.2b) pudieron haberse originado directamente de los mitrados.
GRUPOS SUPERIORES DE PECES VIVIENTES
Los peces vivientes probablemente exceden las 20,000 especies, una abundancia y diversidad que no se iguala reuniendo a todos los otros vertebrados (Figura 1.1). Su remoto antepasado, que se remonta hasta los 500 millones de años, ha dado suficiente tiempo para dar lugar a la aparición de una gran divergencia evolutiva y para dar origen, y permitir la extinción, de líneas filéticas superiores. Inclusive los modernos peces óseos, o teleósteos, tuvieron su origen hace unos 200 millones de años, en el periodo Triásico, cuando los grupos superiores (clases y subclases) de peces estaban ya perfectamente establecidas. La evolución e interrelaciones de los peces todavía están sometidas a discusión, a pesar de que hay una intensa y repetida investigación al respecto; esto se debe a la gran diversidad de puntos de vista filosóficos utilizados por los taxónomos, en parte, lo que da como resul- tado que los detalles de las clasificaciones superiores de estos animales perma- nezcan confusos.
Conociendo estas limitaciones, nosotros hemos decidido sin embargo acep- tar sólo una clasificación de los peces con el propósito de proporcionar una base operacional para uso del estudiante y, esperamos, para que tenga un punto de vista más panorárgico sobre este tema tan difícil de resolver. La clasificación que presentamos ha sido elaborada en base a fuentes recientes, todas las cuales son citadas ya sea directa o indirectamente al final del presente capítulo. Las referen- cias bibliográficas pertinentes dadas ahí también proporcionan una base para la revisión histórica de las numerosas clasificaciones de peces que han sido amplia- mente usadas durante el presente siglo; cuatro son comparadas con la nuestra en la Tabla 2.1. La división primordial en la clasificación de los vertebrados queda entre los vertebrados sin mandíbulas (peces primitivos), los Agnatha, y los que sí las poseen, los Gnathostomata.
Nosotros adoptamos aquí el nombre de Pisces como el más conveniente para incluir a todos los peces y a los vertebrados ictiomorfos. Éstos constituyen

14 LOS GRUPOS SUPERIORES DE PECES
- "" Lamprea
Tiburón
Pez de aietas lobuladas

CARACTERIZACION DE LOS GRUPOS SUPERIORES DE PECES VIVIEN'I'ES 15
tres importantes divisiones naturales (Figura 2.1): 1) CEPHALASPIDOMORPHI: los mixinos (Myxine) y las lampreas, que no tienen mandíbulas (ágnatos) y tienen marsipobranquias; 2) CHONDRICHTHYES: las quimeras, tiburones, rayas y formas relacionadas, que presentan mandíbulas verdaderas (gnatostomados), tienen bran- quias que nacen sobre aberturas branquiales situadas entre cámaras respiratorias (una sola cámara en las quimeras), y tienen esqueleto cartilaginoso (que puede estar calcificado pero sin formar verdadero hueso), y 3) OSTEICHTHYES: todos los peces superiores, que son gnatostomados, tienen una cámara branquia1 prote- gida por una serie opercular hioidea, incluyendo la cubierta protectora de las branquias u opérculo, y tienen esqueleto óseo.
Pisces (grupo con representantes vivientes)
I. Agnatha: los ciclóstomos sin mandíbulas, un grupo que como relicto se derivó de ostracodermos extinguidos. 1. Clase Cephalaspidormorphi: comprende la Subclase Cyclostomata-Myxini-
formes, mixinos y lampreas, y los Petromyzoniformes, las verdaderas lampreas.
11. Gnathostomata: peces mandibulados*
(Superclase Elasmobranchiomorphi: los placodermos extintos y los tiburones,
2. Clase Chondrichthyes: las quimeras cartilaginosas, tiburones, rayas, y gru-
(Superclase Teleostomi: los acantodos fósiles y otros, y los peces óseos) 3. Clase Osteichthyes: celacantos, peces pulmonados, y todos los demás peces
óseos, donde quedan incluidas las especies de peces en una proporción de más del 90% de los vivientes.
holocéfalos y otros grupos relacionados)
pos relacionados.
CARACTERIZACIdN DE LOS GRUPOS SUPERIORES DE PECES VIVIENTES
Para caracterizar los representantes vivos de los grupos superiores de peces es necesario generalizar. Debe comprenderse, por lo tanto, que en las siguientes listas de características distintivas se citan las más frecuentes o notables, mientras que las excepciones, aunque sean conocidas, por lo común son omitidas. Además de la caracterización general, son incluidos algunos términos útiles que han sido aplicados a subgrupos o a diversos aspectos anatómicos.
Clase Cephalaspidormophi (mixinos y lampreas)
a. Notocuerda sin constricciones. b. Sin mandíbulas (ágnatos). c. Esqueleto axial principal (vértebras) cartilaginoso o fibroso.
*Como se emplea aquí, el término incluye todos los vertebrados mandibulados, pero sola- mente los peces mandibulados son Pisces.

16 LOS GRUPOS SUPERIORES DE PECES
d. Dos canales semicirculares en el oído en cada lado de la cabeza en las lampreas, pero sólo uno en cada lado en los mixinos.*
e. Sin arcos verdaderos para el soporte de las branquias y su protección; en lugar de bolsa branquial común cada abertura tiene la suya y el conjunto está situado externamente a las branquias, las arterias del órgano respi- ratorio (arterias branquiales y troncos arteriales) y nervios branquiales.
f. El conjunto branquial firmemente unido al cráneo (neurocráneo). g. Aletas pares (pectorales y pélvicas) ausentes. h. Un solo orificio nasal (monorrino).
Clase Chondrichthyes (quimeras, tiburones, rayas, y torpedos, peces guitarra, etc.)
a. Notocuerda constreñida por vértebras. b. Mandibulados (gnatostomados). c. Vértebras cartilaginosas (con alguna calcificación pero no osificación en
d. Tres canales semicirculares del oído en cada lado de la cabeza. e. Arcos branquiales cartilaginosos y situados por dentro de las branquias
f. Arcos branquiales no unidos firmemente al cráneo, pero unidos a él por
g. Aletas pares presentes. h. Un par de orificios nasales (diminos).
las formas vivientes).
con sus arterias y nervios accesorios.
tejido conectivo.
Subclase Holocephali (quimeras)
a. Branquias dispuestas en cuatro pares; un solo par de aberturas branquiales. b. Ausencia de espiráculo. c. Piel desnuda en los adultos. d . Cloaca ausente. e. Machos con órganos pélvicos intromitentes (gonopodios) que pueden
tener prolongaciones prepélvicas, y algunos (Chimaera) tienen una pro- longación cefálica (tenaculum).
Subclase Elasmobranchii (tiburones, rayas, torpedos, etc.)
a. Cinco a siete pares de branquias y aberturas branquiales. b. Espiráculo presente. c. Escamas placoideas (dentículos dérmicos) presentes o ausentes. d. Cloaca presente. e. Generalmente los machos con órganos intromitentes pélvicos (myxopte-
rigia) .
*El Dr. José Alvarez del Villar afirma que son dos en cada lado, &lo que ‘*. . .uno colocado dentro del otro. . .” (Los cordados. Origen, evolución y hábitos de los vertebrados, 3a. ed. 1977, p. 43. C.E.C.S.A., México) (N. del T.).

GRUPOS SUPERIORES DE PECES EXTINGUIDOS 17
Clase Osteichthyes (peces óseos)
a. Notocuerda constreñida o no. b. Mandibulados (gnatostomados). c. Esqueleto óseo. d . Tres canales semicirculares del oído, en cada lado de la cabeza. e. Arcos branquiales osificados, por dentro de las branquias, junto con las
f. Arcos branquiales no firmemente unidos al cráneo. g. Con aletas pares. h. Un par de orificios nasales (dirrinos).
arterias aferentes y nervios.
Subclase Dipnoi (peces pulmonados)
a. Maxila y premaxila ausentes, tres pares de placas dentarias. b. Con coanas (en las formas recientes). c. Sin articulación movible entre las porciones anterior y posterior del cráneo. d. Palatocuadrado fusionado con el cráneo. e. Prolongación de los radios y músculos hasta un lóbulo basal de la aleta
(en formas recientes); aleta dorsal única. f. Cloaca presente.
Subclase Crossopterygii (peces con aletas lobuladas)
a. Maxila ausente (excepto en algunas formas fósiles); premaxila presente;
b. Sin coanas (en formas recientes). c. Articulación móvil entre las porciones anterior y posterior del cráneo. d. Palatocuadrado no fusionado con el cráneo. e. Prolongación de los radios y músculos hasta la base de la aleta; dos aletas
f. Cloaca ausente.
dientes normales.
dorsales separadas.
Subclase Actinopterygii (de aletas con radios, peces óseos superiores)
a. Maxila y premaxila presentes. b. Sin coanas. c. Sin articulación móvil entre las porciones anterior y posterior del cráneo. d. Palatocuadrado no fusionado con el cráneo. e. No se prolongan los radios y músculos hasta la base de la aleta (excepto
f. Cloaca ausente. en Polypteriformes); aleta dorsal única o dividida.
GRUPOS SUPERIORES DE PECES EXTINGUIDOS
Para un paleontólogo los animales vivientes constituyen solamente una pequeña (e incompleta) sección tangencia1 de la historia de los vertebrados. Esto se com- prende fácilmente si consideramos que, aunque puedan existir algo así como 5 o 10 millones de especies animales recientes, las que se han extinguido deben

18 LOS GRUPOS SUPERIORES DE PECES
haber llegado a sumar algo así como medio billón (Mayr, 1969;véaseCapítulo 13). Excepto para los mamíferos, el regist,ro fósil para ios vertebrados vivientes presenta grandes lagunas de conocimiento por no haber los suficientes restos fósiles. Mu- cho material fósil está representado solamente por fragmentos cuya identifica- ción hasta especie es a menudo imposible, principalmente porque la osteología de las especies vivientes y relacionadas, o sea, que tienen cierta afinidad filogené- tica con ellos, es muy poco conocida. Por lo tanto, la delimitación y posición precisa de los grupos extinguidos ha sido especialmente difícil, no solamente porque aún se tienen registros imperfectos, sino también porque muchos depó- sitos de materiales fósiles no han sido hasta la fecha diagnosticados y mucho de lo que hay está regado en colecciones de estudio en el mundo entero.
La interpretación de los fósiles requiere de una mente imaginativa que haya sido cuidadosamente entrenada a través del estudio de la morfología comparada y la sistemática, y que combine estos conocimientos con la capacidad de juzgar intuitivamente. Aunque las inferencias relativas a las interrelaciones de los fósiles están siendo mejoradas mediante la incorporación de nuevos principios y métodos de estudio, persiste todavía un acalorado debate entre los especialistas en relación a la clasificación de los diversos grupos de fósiles.
Todos los peces fósiles y vivientes son seleccionados como ágnatos o gnatos- tomados pero, a pesar de que es razonablemente bien conocida la evolución de estos dos grupos considerados como fundamentales, la filogenia de los peces en su conjunto puede ser reconstruida solamente a grandes rasgos. Hay muchos grupos de peces ágnatos y mandibulados que fallaron en sus intentos de adaptación y ahora están extinguidos, ya que no pudieron sobrevivir al tiempo geológico. Las formas ágnatas más antiguas son los heterostrácodos y telodontos del Paleozoico, que son los primeros vertebrados que han aparecido en los registros fósiles. Éstos y otros ágnatos del Paleozoico han sido frecuentemente relacionados con los ostracodermos, como lo hace Romer (1966), aunque Miles (en Moy-Thomas and Miles, 1971) evitó este término porque é1 cree que representa una especie de ensamble artificial, y Romer no lo emplea en su clasificación formal.
La clasificación para los Agnatha favorecida por Miles es adoptada aquí, ha- ciendo la excepción de los ostracodermos, que sí se toman en cuenta. Dos clases, Cephalaspidomorphi y Pteraspidomorphi, son tratadas con estos nombres (corres- pondiendo a los Monorhina y Diplorhina de Romer). Para los Gnathostomata, los grupos desaparecidos son los Placodermi y los Acanthodii. Aunque no hay un total acuerdo sobre las interrelaciones de los peces del Paleozoico, los materiales básicos con que se cuenta constituyen un récord valioso que explica la evolución temprana de los peces que involucran a los grupos superiores que les siguen.
Clase Cephalaspidomorphi (osteostracodos y anápsidos extinguidos, y los mixinos y lampreas vivientes), Figura 2.2a
En las formas extinguidas,* la cabeza y la porción anterior del tronco están cu- biertas con una coraza ósea continua. El neurocráneo de cualquiera de ellas está formado por una combinación de cartílago y hueso, o (raramente) está osificado en su totalidad. Hay una abertura nasal media y única que no comunica con la
*Las formas vivientes fueron caracterizadas en las páginas 15-17.

GRUPOS SUPERIORES DE PECES EXTINGUIDOS 19
Figura 2.2 - Grupos representativos de peces fósiles. Los que se muestran son (a) un cefalas- pidomorfo (Hemicyclaspis); (6) un pteraspidomorfo (Pteraspis); (c) un placodermo artrodiro (Rernigolepis), restaurado; ( d ) un placodermo antiarcano (Gernuendina), en su vista dorsal; (e) un acantodio (Climatius); ( f l un tiburón del nivel cladodonto, Cladoselache; (S) un tiburón del nivel hibodonto (Hybodus); y (h) el más antiguo tiburón moderno conocido (Paleospinax). (Figuras 2.2f a l a h, redibujadas de Schaeffer, 1967).

20 LOS GRUPOS SUPERIORES DE PECES
cavidad bucal. Hay dos canales semicirculares. Las branquias están en bolsas como ocurre en las actuales lampreas y mixinos. El esqueleto branquial es externo y está en contacto con la musculatura asociada. Hay diez pares de aberturas bran- quiales externas en la superficie inferior de la cabeza. La parte del cuerpo situada detrás de la coraza cefálica está cubierta con escamas imbricadas. Presentan aleta caudal heterocerca; aletas pectorales generalmente presentes, cubiertas con esca- mas y sin radios dérmicos. No hay aletas pélvicas (pero existe un pliegue ventral). Una o dos aletas dorsales. Nueve órdenes son reconocidos por Miles.
Clase Pteraspidomorphi (heterostráceos y telodontos), Figura 2.2b
Tienen la cabeza y parte posterior del tronco cubiertos con una placa de hueso sdido (carapacho) sin células óseas verdaderas. Aberturas nasales externas aparen- temente ausentes, pero una coana comunica con la cavidad bucal. Dos canales semicirculares. Sólo una abertura branquial externa a cada lado. El cuerpo, detrás del carapacho, está cubierto con escamas. Hay aleta caudal hipocerca; no hay más aletas. Nueve órdenes son reconocidos por Miles.
Clase Placodermi (placodermos), Figura 2 . 2 ~ y d
Es un grupo de peces óseos mandibulados muy diversificado y de representantes ya extinguidos; dorsoventralmente comprimidos (deprimidos); con una coraza cefálica movible y articulada con el tronco a una concha que cubre a éste en su porción anterior; la posterior se prolonga hacia atrás hasta terminar en una cola heterocerca en la mayoría de las especies. Los artrodiros, que muestran la mayor radiación adaptativa de todos los grupos de placodermos, han sido estudiados minuciosamente durante casi cuarenta años por Stensio (1969). La clase puede ser polifilética en origen; posteriormente dio lugar a la aparición de los Chondrich- thyes y los Osteichthyes. Miles reconoces seis órdenes.
Clase Acanthodii (acantodios), Figura 2.2e
Los primeros peces verdaderamente mandibulados con hueso verdadero en el endoesqueleto fueron los acantodios. Pequeños, con escamas rómbicas, estriadas y tan pequeñas que son casi granulares, espinas rígidas delante de las aletas dorsal, anal y las pareadas; aleta caudal heterocerca. Son los únicos peces del Paleozoico con espinas en las aletas pares. La posición de este grupo es debatible aunque algunas opiniones recientes sugieren que los acantodios están más estrechamente relacionados con los osteictios que éstos a los condrictios (Miles, 1974).
Dos importantes grupos de fósiles del Mesozoico, los folidofóridos y los lep- tolépidos, corresponden a peces que han sido motivo de discusión principalmente por Patterson (1975), quien ha concluido que ellos “están en la base de los Te- leostei” (p. 278). Más tarde afirmó que “en la evidencia del cráneo de los teleós- teos vivientes se ve que forman un grupo monofilético cuyos orígenes se sitúan dentro del nivel de los leptolépidos, no de los folidofóridos” (p. 562). En la clasi- ficación empleada aquí, hemos enfatizado esta cuestión colocando los peces lep-

RELACIONES DE LOS GRUPOS SUPERIORES DE PECES 21
tolépidos en los Teleostei, aunque se concede que los folidoforoideos son precur- sores de los Teleostei (Gosline, 1971, p. 96).
RELACIONES DE LOS GRUPOS SUPERIORES DE PECES
Las relaciones que los animales tienen entre sí, tomando en cuenta el factor tiem- po, refuerza la teoría de la evolución de las especies. Este respaldo es suficiente- mente fuerte en los grupos donde el registro fósil no es bastante bueno como para aclarar lo que sucedió durante la evolución, es decir, en los grupos cuya evidencia fósil deja mucho que desear. Inclusive, para aquellos grupos cuya evidencia es muy escasa, ninguna persona de buen juicio podría concluir que las formas aisladas o grotescas hayan aparecido de nouo. Por ejemplo, el hecho de que los peces más primitivos que se conocen están fuertemente protegidos con armadura no impide creer que exista la posibilidad de que sus ancestros no lo hayan estado. Parece ser que, más bien, las formas con armadura persistieron como fósiles mientras que sus parientes sin armadura no se fosilizaron. En general, la secuencia de aparición de los grupos de peces sugiere una evolución ordenada, aunque existan lagunas en los registros fósiles entre los grupos superiores; soluciones de continuidad que perma- necen sin explicación o que no han sido comprendidas todavía.
El avance en la comprensión de las interrelaciones de los peces ha sido lento por la carencia de fósiles, la escasez de formas primitivas y un exceso de formas
. modernas, muy evolucionadas, de peces tanto fósiles como vivientes. La situación se ha visto agravada en el pasado reciente porque los peces fósiles y los vivientes han sido estudiados bajo los métodos de dos disciplinas independientes, desde el punto de vista de la investigación científica. Que esta dicotomía está siendo unida en algunas porciones de sus ramas, ha quedado demostrado en un reciente trabajo importante sobre este tópico (Greenwood, Miles, y Patterson, 1974).
El estudio de los peces conocidos como los más primitivos se basa en los fragmentos de huesos de los heterostráceos, que datan de los depósitos del Ordo- vícico medio, hace cerca de 500 millones de años. Hacia el Silúrico los dos grupos superiores de vertebrados -10s peces sin mandíbula, o Agnatha, y los primeros vertebrados mandibulados, o Gnathostomata- ya se habían establecido.
El espacio morfológico y evolutivo que separa los Agnatha de las otras clases de peces es mucho mayor del que existe entre los peces mandibulados y las cuatro clases de tetrápodos. Por lo tanto los peces, o Pisces, tomados en sentido amplio, reúnen a un conjunto de diversas agrupaciones de genealogías primitivas y sin relación entre sí, las que han sido dispuestas por diferentes autoridades en ictiología en cuatro clases o más, sobrepasando a veces una docena de ellas.
Peces primitivos
Los vertebrados vivientes más primitivos son los ciclóstomos cefalaspidomorfos, que comprenden las lampreas y los mixinos, que son, con sus marcadas modifi- caciones, descendientes degenerados de ágnatos con armadura. Además de la ausencia de mandíbulas mordedoras, estas criaturas delicadas, delgadas y de as- pecto superficial viperino, algunas de las cuales se han adaptado para vivir como parásitas o alimentarse de desperdicios, también carecen de apéndices pares a l e - tas o miembros locomotores- que están presentes (a menos que hayan sido

22 LOS GRUPOS SUPERIORES DE PECES
perdidos después) en todos los demás peces de grado evolutivo avanzado. Des- pués adquirieron un esqueleto cartilaginoso, que alguna vez se pensó constituía un carácter primitivo. Ha sido bien demostrado, sin embargo, que el hueso, o el tejido parecido al hueso, y el cartílago, son materiales de cuyo primitivism0 no se puede dudar (Moss, 1964).
La línea ancestral de la que los ciclóstomos se expandieron comprende a los ostracodermos sin mandibulas, de vida bentónica y fuertemente armados con una cubierta protectora, adernás de ser deprimidos, carentes de aletas y con el cuerpo rodeado por una concha ósea. Posiblemente, debido en parte a la posesión de un esqueleto bien osificado, éstos fueron los tipos mis conspicuos de verte- brados durante las etapas mas antiguas de la historia que conocemos de los peces; el Silúrico en sus postrimerías y el Devónico. Qariaban en longitud de solamente unos cuantos centrímetros hasta treinta aproximadamente. Hasta que apareció el estudio revolucionario deí gran paleontólogo sueco Erik Stensio a finales de los años veinte, los ostrocodermos habían sido superficialmente descritos y larga- mente ignorados. Mediante la aplicación de fatigosas y muy elaboradas técnicas, Stensia demostró que había correspondencia anatómica fundamental entre los cefaláspidos (Figura 2.2a; uno de los mayores grupos de ostracodermos) y las lam- preas (Petromyzonidae), especialmente al examinar la larva arnocete. Sin embargo, los mixinos (Myxinidae) difieren de las lampreas por tener la abertura nasal en la extremidad de la cabeza, el canal pituitario abierto hacia la laringe, de 5 a 15 (de preferencia 7 ) bolsas branquiales en cada lado, un esqueleto branquia1 rudi- mentario, un canal semicircular* (con más frecuencia que dos diferentes) y los huevos de mayor tamaño y con desarrollo directo (no hay estados larvariosf. Aunque algunos mixinos son clasificados con las lampreas y los cefalaspidomor- fos aquí, posiblemente derivaron de una diferente línea evolutiva (los pteráspidos; Figura 2.2b). Una tercera linea evolutiva (los telodontos) comprende solamente ostracodermos extinguidos.
Los cambios externos tan sorprendentes, de la línea ancestral, de los cefalás- pidos para terminar con las lampreas vivas de l a actualidad, es parcialmente el resultado de un desarrollo regresivo del esqueleto, que trajo consigo primero algo así como una desintegración y finalmente una pérdida total del esqueleto óseo. Los ostracodermos todavia prosperaron a principios del Devónico pero se extin- guieron a finales del mismo periodo, tiempo durante el cual fueron apareciendo otros vertebrados superiores.
La presencia de tejido óseo cubriendo a los primitivos ágnatos de la cabeza a la cola ha confundido a los investigadores. Se ha sugerido que los jgnatos provis- tos de armadura ósea pudieron defenderse de los euriptéridos, que tenían forma de escorpión y fueron sus potenciales predadores contemporáneos. También se supone que la arrnadura atenuó a estos vertebrados primitivos el peligro de sufrir fuertes intercambios osmbticos en los tejidos de sus cuerpos, cuando se encontra- ban era aguas dulces, pues hay que recordar que su antepasado fue de origen marino.
Primeros peces mandibulados
Los Cnathostomata, que incluyen a los peces provistos de mandíbulas, compren- den a los grupos fundamentales de los que se originaron los tetrápodos (Figura
*Véase la nota de la pigina 36. (N. del T.)

RELACIONES DE LOS GRUPOS SUPERIORES DE PECES 23
2.3). Un poco separados de la línea evolutiva principal que dio lugar a los peces óseos, y cerca de la base o punto de origen de la rama que dio lugar a la aparición de los Elasmobranchiomorphi, están los placodermos (Figura 2.3). Los primeros restos fósiles de estos peces pertenecen al periodo Silúrico, se acrecientan en el periodo Devónico, la “Edad de los Peces”, extinguiéndose en el Carbonifero. Su historia queda confinada a la Era Paleozoica (Moy-Thomas y Miles, 1971). Todos tienen tejido óseo en su esqueleto y aletas pares, aunque la estructura anatómica de algunos se aleja de lo normal.
Los “tiburones con espinas” o acantodios (Figurra 2.2f) formaron un grupo bien representado y exitoso, caracterizado por la posesión de fuertes espinas que reforzaban a todas las aletas excepto la caudal, que estaba muy levantada, como la aleta heterocerca de los tiburones. Su cubierta corpórea consistía en una serie de escamas óseas pequeñas y aplanadas, muy diferentes de los dentículos dérmi- cos de los tiburones. Las grandes espinas, probablemente fijas, o sea sin movimien- to articulado, evidentemente sirvieron como estabilizadores. Eran excepcional- mente chatos, es decir, de hocico corto, como resultado de la reducción de la región del etmoides de la cabeza y la presencia de enormes órbitas. La región branquial estaba aparentemente cubierta por estructuras operculares. Sus cuerpos estaban rodeados de una armadura completa de verdaderas escamas, cuya estruc- tura microscópica es casi idéntica a la de las escamas ganoideas de ciertos actinop- terigios. Los acantodios típicos no crecieron más que unos cuantos centímetros, aunque algunos géneros marinos aberrantes alcanzaron considerable tamaño. En el Devónico Inferior, el grupo había alcanzado la cúspide de su desarrollo en relación al número de individuos y variedad de formas, y abarca los vertebrados más abundantes en los depósitos de esa edad. Estos peces muestran una interesante combinación de características primitivas y avanzadas, y aunque el grupo que forman no aparece a lo largo de la rama directa que dio lugar a la aparición de los peces superiores, claramente muestran que están cerca del ancestro que originó los peces óseos y ha sido colocado por algunos investigadores contemporáneos en o cerca del origen de los Osteichthyes (véase la discusión de Romer, 1966). Acan- thodes bronni (la especie del Pérmico que revela detalles de la estructura interna) y osteictios primitivos muestran mutuas semejanzas fundamentales en el cráneo que no son compartidas por los condrictios (Miles, 1974).
Los placodermos aparecieron casi al mismo tiempo que los acantodios. Sin embargo, a excepción de un género que sobrevivió hasta el Carbonífero, se extin- guieron cuando terminaba el Devónico, hace alrededor de 300 millones de años. Se consideran dos importantes subgrupos: los Arthrodira (Figura 2.2e), y 10s Antiarchi (Figura 2.2d).
LOS llamados peces con carapacho articulado en el cuello, los artrodiros, fueron los que formaron los grupos más grandes de placodermos y sus restos son 10s más comunes entre los vertebrados del Devónico. El nombre vernacular se refiere a la disposición del carapacho óseo en dos porciones rígidas, una cubriendo la cabeza y la región branquial (las placas cefálicas) y la otra cubriendo gran parte del tronco (las placas torácicas) ; la segunda articulada con la primera 0 anterior mediante el sistema de cóndilos articulares (en el carapacho del cuerpo) y fosetas (en el carapacho de la cabeza). Por lo tanto, fue la primera vez que apareció en un pez el movimiento libre de la cabeza, hacia arriba y hacia abajo, en relación al tronco. La forma de su cuerpo, siempre deprimida, sugiere que llevaban un tipo bentónico de vida. Estos peces tan peculiares, cuya longitud variaba desde 10s

r I / I

RELACIONES DE LOS GRUPOS SUPERIORES DE PECES 25
0.5 m hasta, posiblemente, los 10 m, tenían láminas mandibulares muy particula- res y diferentes a las estructuras mandibulares de los vertebrados superiores. La ausencia general de coraza en la parte posterior del cuerpo es indicación de la ad- quisición de mejoras en la locomoción que permitieron a estos peces comenzar a despojarse de su pesada armadura.
Otro grupo aun más aberrante asignado al de los Placodermi es el de los antiarcos, peces pequeños y grotescos con apéndices óseos articulados que no son espinas ni aletas. Estuvieron ampliamente distribuidos en el Devónico y fueron en su mayoría más bien pequeños, pues su longitud no era mayor de 30 cm. Sus características fundamentales recuerdan a las de los artrodiros, de los que difieren sobre todo por las mandíbulas de muy débil constitución o estructura, y en las extraordinarias “aletas” articuladas y de movimientos libres, posiblemente deri- vadas de las espinas fijas encontradas en los dos grupos citados anteriormente y empleadas por el animal para sostenerse en el sustrato de los ríos. Estudios anató- micos detallados han sugerido la posibilidad de que estos animales hayan tenido pulmones.
Tiburones y sus parientes (Chondrichthyes)
El siguiente grupo importante de peces, la clase Chondrichthyes, comprende los tiburones, rayas, quimeras y sus parientes fósiles. Los tiburones y las rayas forman el último grupo superior de peces que aparece en el registro fósil. La fertilización interna es característica de todas las formas vivientes y es conocida en todos los tiburones del Jurásico; es posible que se remonte hasta los tiempos de finales del Devónico. El esqueleto no tiene hueso; sin embargo, como lo hemos hecho notar arriba, se cree que en lugar de ser una condición ancestral, es más bien una carac- terística derivada, y el antepasado de los tiburones debe ser buscado entre los primitivos peces óseos a lo largo de la rama evolutiva que dio lugar a los placo- dermos y otros grupos cercanos o relacionados (Figura 2.3). La línea principal en la evolución de los tiburones se desarrolló en el mar.
Los primeros fósiles de tiburones aparecen en el Devónico Inferior, como los dientes y espinas de un tipo que es característico de cierto número de elas- mobranquios primitivos. A fines del Devónico dos tipos primitivos de tiburón poseían esos dientes. Uno de ellos es el famoso género Cludoseluche (Figura 2.2f) en el cual las aletas pares tienen base ancha y no se le han encontrado siquiera trazas de gonopodios. En el Carbonífero había géneros aparentemente transicio- nales entre estos tipos del Devónico y las relativamente primitivas formas conoci- das de fines del Paleozoic0 al Mesozoico Medio. Puede seguirse hacia atrás hasta el Jurásico el desarrollo evolutivo de los tiburones y rayas modernos, cuando comenzó su intensa radiación adaptativa. Por lo tanto, la historia de los elasmo- branquios, desde sus antepasados fósiles hasta los grupos modernos, vivientes, es bastante bien conocida (Zangerl, 1974). Después de la diversificación ocurrida en el Carbonífero hubo una gran reducción en el número y variedad de estos peces a finales del Pérmico; época que marca la desaparición de muchas formas primitivas. Después de que en el Triásico y el Jurásico hubo de nuevo expansión de grupos, casi todos los modernos linajes de tiburones, rayas y torpedos estaban representados en los mares de principios del Cretácico.
LOS elasmobranquios, en su historia evolutiva, pawon a través de dos nive-

26 LOS GRUPOS SUPERIORES DE PECES
les sucesivos de organización o radiaciones, el “cladodonto” y el “hibodonto”, en su recorrido hacia las formas modernas, en las que obtuvieron una nueva suspen- sión mandibular (la hiostílica) y un esqueleto axial con centra vertebral calcificada (las líneas galeoide, escualoide y batoide). Cladoselache (Figura 2.2f) es un repre- sentante del nivel cladodonto, Hybodus (Figura 2.2g) lo es del nivel hibodonto, y Pdeospinax (Figura. 2.2h) es, de los modernos, el más antiguo conocido.
Las quimeras de nuestros días (Holocephali) son peces marinos grotescos que se alimentan principalmente de moluscos. Poseen esqueleto cartilaginoso y otros atributos escualoides, pero se ha pensado que los dos grupos están muy poco relacionados. Algunos descubrimientos fósiles recientes (Zangerl, 1974) indican, sin embargo, que los precursores de este grupo del Carbonífero, los Iniopterygii (Figura 2.3), que han sido interpretados como holacéfaios, tienen características dentarias que obligan a separarlos de los elasmobranquios primitivos; entonces, la afirmación (mencionada por Zangerl) de que los holockfalos se hayan originado de alguna rama de los placodermos es errónea.
Peces óseos (Osteichthyes)
Los mayores grupos de peces considerados hasta aquí, aunque extremadamente importantes en los estratos del Paleozoico, en gran parte se extinguieron (a excep- ción de los tiburones) hacia la terminación del Pérmico. La clase mayor de formas vivas, la Osteichthyes o de los peces óseos, en contraste, ha incrementado conti- nuamente su importancia desde sus inicios al comenzar el Paleozoico. Aunque el registro de fósiles señala que los peces óseos no aparecieron sino hasta fines del Silúrico, los más importantes grupos estaban ya bien establecidos hacia media- dos del Devónico y, en consecuencia, es razonable inferir que los comienzos del grupo se extienden hasta tiempos tan lejanos como los del Ordovícico, hace más de 500 millones de años.
Aunque los Osteichthyes poseen un esqueleto bseo, hemos visto que esta característica no es exclusiva de ellos. La evidencia fósil claramente apunta a la conclusión de que sus antepasados no solamente tenían hueso; sino que muchos de ellos tenían inclusive mejor osificación que sus actuales descendientes. Tres grupos importantes de peces óseos pueden ser reconocidos (los Acanthodii, des- critos ya, son considerados por algunos investigadores como una subclase de los Osteichthyes): 1) los peces de aletas lobuladas o crosopterigios (Crossopterygii), importantes porque son la línea ancestral que conduce a los tetrápodos; 2) los peces pulmonados o dipnoos (Dipnoi), y 3) los peces con espinas en las aletas (Actinopterygii). Los grupos (1) y (2) equivalen a la subclase Sarcopterygii de Romer (1966) pero no a la de Nelson (1969) quien cree que este grupo debería incluir también los Polypteriformes (Polypterus y Calamoichthys),* aquí situados dentro del grupo Chondrostei bajo el grupo (3).
Los dipnoos se originaron en el Devónico Inferior y sobreviven hasta la actualidad seis especies en tres géneros y dos familias; hay cerca de 35 géneros extinguidos en nueve familias.
Los peces de aletas lobuladas también están distribuidos en el tiempo desde
*El autor da los nombres vernáculos en inglés; nosotros mencionamos los géneros por no existir nombres vulgares en español. (N. del T.)

RELACIONES DE LOS GRUPOS SUPERIORES DE PECES 27
el Devónico Inferior hasta el Reciente con cerca de 55 géneros extinguidos y sola- mente una especie viviente (Latimeria chalumnae, conocida desde 1938).
De los peces con espinas en las aletas, que datan desde el Devónico Inferior, hay cerca de 20,000 especies vivientes y muchos grupos extinguidos.
Los primeros de estos peces con espinas en las aletas fueron en su mayor parte paleoniscoideos del Paleozoico; un vasto grupo de formas sumamente diver- sificadas. La mayoría fue de tamaño moderado y presentaba un parecido muy superficial con sus sucesores actinopterigios; por ejemplo, el pejelagarto (Lepi- sosteidae), las sardinas (Clupeidae) y los pececillos de acuario (Cyprinidae). Hubo, sin embargo, diferencias estructurales fundamentales: las escamas ganoideas grue- sas, la aleta caudal heterocerca, la estructura craneal primitiva y una cintura pectoral con clavícula y cleitro. En Cheirolepis, uno de los géneros más primiti- vos, las pequeñísimas escamas cuadrangulares son notablemente parecidas a las de los acantodios escualiformes (véase arriba); otros rasgos primitivos son una gran abertura bucal y maxila y la presencia de aletas pélvicas con base ancha y mu- chos radios.
Los paleoniscoideos, desde los primitivos a los modernos teleósteos, fueron los peces de agua dulce dominantes desde el Carbonífero hasta los primeros tiem- pos del Triásico. La radiación adaptativa de este grupo extinguido ha de haber competido con la de los actuales peces óseos. Los representantes modernos de los paleoniscoideos son unas cuantas formas especializadas y degeneradas como los es- turiones (Acipenseridae) y los peces espátula (Polyodon, Psephurus), con sus esqueletos internos casi totalmente cartilaginosos, con aletas primitivas y pareci- das a las de los tiburones, escamas reducidas a hileras (formando placas óseas como en los esturiones), etcétera. Actinopterigios de ancestro Mesozoico son los pejelagartos y amiidos, que persisten en la actualidad en solamente dos géneros, Lepisosteus y Amia. Los pejelagartos ancestrales fueron más abundantes en los co- mienzos del Mesozoico, mientras que los peces Amia estaban especialmente bien representados a mediados de la misma era geológica.
Se observa que los antepasados paleoniscoideos, tras su paso por dos etapas intermedias, llegaron al gran florecimiento que se manifiesta en los peces óseos verdaderos de nuestros días. Algunos términos descriptivos, de significación histórica en ictiología, han sido aplicados a estos estados, aunque se reconoce que son arbitrarios y no hay un total acuerdo sobre las familias de peces que deben ser incluidas en cada grupo. Sin embargo, dichos términos son descripti- vos de los representantes vivientes de los grupos. Yendo en orden progresivo, es decir, comenzando con los primitivos, pasando por los intermedios y llegando a los más evoluciocados, ellos son: a) los Chondrostei, donde quedan incluidos los esturiones (Acipenseridae) y los peces espátula (Polyodontidae); b ) los Holostei (pejelagartos, Lepisosteidae; Amiu, Amiidae), y c ) Teleostei, el enorme conjunto de los peces más comunes. Los rumbos ya reconocidos que siguió la evolución a través de estas etapas están resumidos en la Tabla 2.2. Es cada día mayor la indi- cación de que la evolución a través de estos grupos fue desviada (o sea, no fue ortogenética), donde las características más avanzadas derivaron en forma inde- pendiente a lo largo de diferentes líneas, siendo de múltiple origen (polifilético) los Holostei y los Teleostei. En general, en la historia evolutiva de los Actinop- terygii las estructuras esqueléticas se han reducido; esto ha sucedido efectivamente en el endocráneo, los huesos dérmicos y las escamas.
En los peces óseos superiores, más numerosos, los Teleostei (Tabla 2.3),

28 LOS GRWOS SUPERIORES DE PECES
c1 al
6 2 I l a d C
z O
C al

RELACIONES DE LOS GRUPOS SUPERIORES DE PECES
I
I
i
I I
I
7 C

30 LOS GRUPOS SUPERIORES DE PECES
ocurrió una gran divergencia entre los tipos generalizados de peces parecidos a las sardinas y los salmones (algunas veces dominados Isospondyli o, aproximadamen- te, Clupeiformes). Ellos integran una gran porción de los peces de aletas con radios blandos, los “Malacopterygii” o “Malacopteri” de los antiguos autores. Muchas familias marinas y de agua dulce han florecido en este grupo. Ellas son típica- mente óseas pero de cuerpo blando, tienen aletas reforzadas Gnicamente por ra- dios blandos (no por espinas), poseen escamas cicloideas, tienen el hueso maxilar formando una parte de la abertura o borde bucal (borde superior de la abertura bucal), y t,ienen l a vejiga gaseosa (vejiga aérea, vejiga natatoria) comunicada con el esófago (condición fisostoma). Durante el Cretácico los bericoides o Beryciformes participaron en una transicibn a peces be aletas con espinas (más o menos los “Acanthopterygii” o “Acanthopteri” de algunos autores), y ocurrió otro gran florecimiento de familias.
El grupo más grande que resultó de la divergencia del tronco evolutivo es el de los peces percoides o percomorfos, los Perciformes. A este grupo pertenecen miles de peces marinos y de aguas dulces, tipificados por los meros (Serranidae) de distribución mundial y las percas de agua dulce (Percidae) del Hemisferio Nor- te. En las especies vivientes las aletas están reforzadas por espinas y radios blan- dos, las escamas son con frecuencia ctenoideas, las maxilas son excluidas de la abertura bucal por la elongacibn de las premaxilas, 3 7 las aletas pélvicas, cuando están presentes (como es le usual), son de posición torácica o yugular (con una f6rmula reducida, típicamente de una espina y cinco radios blandos); la vejiga gaseosa carece de conducto que l a comunique con el esófago (condiciónfisoclista).
LISTA DE LAS FAMILIAS COMUNES Y REPRESENTATIVAS DE PECES VIVIENTES
Teniendo en consideración que en tiempos recientes se ha presentado una impresionante actividad en la clasificación de los peces (véanse las referen- cias), el sistema de Berg (19401, empleado antes por nosotros, no puede ser sos- tenido por más tiempo. Fishes of the World (Lindberg, 1971) es un libro que contiene claves iitiles de identificación y describe todas las familias de peces (550 en 62 órdenes, que é1 reconoce), pero es una compilación más que una síntesis. El sistema empleado a continuación no sigue los postulados de una auto- ridad, iinica y ampliamente aceptada; representa más bien el esfuerzo concertado para seleccionar lo que nos pareció más razonable.
Agnatha Clase Cephalaspidomorphi
Subclase Cyclostomata Orden Myxiniformes
Orden Petromyzoniformes Familia Myxinidae - mixinos (Figura 2.4)
Familia Petromyzonidae - lampreas (Figuras 2.1 y 2.5) Gnathostomata
Clase Chondrichthyes Subclase Holocephali
Orden Chimaeriformes Familia Chimaeridae - quimeras (Figura 2.9)

LISTA DE LAS FAMILIAS COMUNES REPRESENTATIVAS DE PECES VIVIENTES 31
"- glutinosa. Figura 2.4 (Basado - Mixino en del Bigelow Atlántico, y Schroeder, Myxine <:a 1948).
/""" -Y"-
..',. -
Figura 2.5 - Lamprea marina, Petromyzon marinus. (Basado en Jordan y Evermann, 1900).
Figura 2.6 - Tiburón de seis branquias, Hexanchus griseum. (Basado en Jordan y Evermann, 1900).
A-- Figura 2.7 - Tiburón toro, leucas. (Basado en Jordan 1900).
Carcharhinus y Evermann,
Figura 2.8 - Raya pequeña, Raja erinacea. -a ......- (Basado en Jordan y Evermann, 1900). O&' ,...--...
.. . ..
Figura 2.9 - Pez rata (quimera), Hidrola- gus collie¡. (Basado en Jordan y Ever- mann, 1900).

32 LOS GRUPOS SUPERIORES DE PECES
Subclase Elasmobranchii (Selachii) Orden Heterodontiformes
cabeza de toro Familia Heterodontidae - tiburones cornudos o
Orden Hexanchiformes Familia Hexanchidae -tiburones vaca (Figura 2.6) Familia Chlamydoselachidae tiburones ornados
Familia Odontaspidae - tiburones arenosos Familia Alopiidae - tiburones coludos o zorros Familia Lamnidae - tiburón azul, tiburón bonito, etc. Familia Orectolobidae - tiburón gata Familia Rhincodontidae - tiburones ballena Familia Cetorhinidae - “basking sharks”* Familia Scyliorhinidae - tiburones gata Familia Carcharhinidae - tintorera (hay muchos
Familia Sphyrnidae - cornuda, pez martillo Familia Squalidae - cazón de espina, galludo Familia Squatinidae - diablo, angelote
Familia Pristiophoridae - tiburón sierra
Familia Pristidae - peces sierra Familia Rhinobatidae - peces guitarra Familia Rajidae - rayas Familia Dasyatidae - raya de espina Familia Myliobatidae - raya pinta Familia Mobulidae - manta raya
Familia Torpedinidae - torpedos
Orden Squaliformes
nombres vernáculos)
Orden Pristiophoriformes
Orden Rajiformes (Batoidei)
Orden Torpediniformes
Clase Osteichthyes Subclase Crossopterygii
Orden Coelacanthiformes Familia Coelacanthidae - celacantos (Figuras 2.1 y 2.10)
Subclase Dipnoi Orden Dipteriformes
Familia Ceratodontidae -pez pulmonado australiano Familia Lepidosirenidae - peces pulmonados de América del Sur y África
Subclase Actinopterygii Orden Polypteriformes
Orden Acipenseriformes Familia Polypteridae - “bichirs” y “reedfishes”
Familia Acipenseridae - esturiones Familia Polyodontidae - peces espátula
en español. (N. del T.) *Se conserva el nombre del pez en inglés, entre comillas, cuando no hay equivalente

LISTA DE LAS FAMILIAS COMUNES REPRESENTATIVAS DE PECES VIVIENTES 33
Figura 2.10 - Celacanto, Latimeria cha- lumnae. (Basado en E. London Mus., Sudáfrica).
Figura 2.1 2 - Pol íptero, Polypterus sene- galus. (Basado en Bridge, 1904).
Figura 2.11 - Pez pulmonado australiano, Neoceratodus forsteri. (Basado en Nor- man, 1931).
Figura 2.13 - Esturión del Atlántico, Acipenser oxyrhynchus. (Basado en Jor- dan y Evermann, 1900).
Figura 2.14 - Pez de aleta ondulada, Amia . -- ......-... - ..... calva. (Basado en Jordan Y Evermann, 1900).
Figura 2.15 - Catán de nariz larga, Lepi- sosteus osseus. (Basado en Jordan y Evermann, 1900).
Figura 2.16 - Arenque, Chupea harengus. (Basado en Jordan y Evermann, 1900).

34 LOS GRUPOS SUPERIORES DE PECES
Orden Amiiformes
y 6.2a) Familia Amiidae - pez de aleta dorsal ondulada (Figuras 2.14
Orden Lepisosteiformes
Orden Elopiformes Familia Lepisosteidae - peje lagartos (Figura 2.15)
Suborden Elopoidei Familia Elopidae - lisa Familia Megalopidae .. tarpón, &halo
Familia Albulidae - pez señorita Subordcn Albuloidei
Orden Anguilliformes Suborden Anguilloidei
Familia Anguillidae - anguila Familia Muraenidae - morena Familia Dysommidae - “Arrowtooth eel” Familia Muraenesocidae - congrio Familia Nettastomidae - “duckbill eels” Familia Congridae - congrio Familia Ophichthidae - morena, culebra Familia Simenchelyidae - “snubnose eel” Familia Derichthyidae - “longneck eel” Familia Nernichthyidae - “snipe eel” (Figura 3.1)
Familia Saccopharyngidae - ‘‘swallowers” (Figura 2.22) Familia Eurypharyngidae “gulper”
Familia Halosauridae - anguila halosaurida (Figura 2.26) Familia Notacanthidae -- “spiny eel” (Figura 2.27)
Suborden Saccopharyngoidei
Orden Notacanthiformes
Orden Clupeiformes Suborden Denticipitoidei
Suborden Clupeoidei Familia Denticipitidae “African herring”
Familia Dussumieriidae - “round herrings” Familia Clupeidae - sardinas (Figuras 2.16 y 8.12) Familia Engraulidae -- anchoa, anchoveta
Orden Osteoglossiformes Suborden Osteoglossoidei
Familia Osteoglossidae - “bonytongues”, lengua de hueso* Familia Pantodontidae - “African mudskipper”
Familia Hiodontidae -- - ojos de luna Familia Notopteridae -- lomo emplumado
Familia Morrnyridae - pez elefante (Figuras 2.23 y 5.5a) Familia Gymnarchidae - gimnárchidos
Suborden Notopteroidei
Suborden Mormyroidei
*Para mayor comprensibn de1 estudiante, ya que algunos nombres vernáculos vienen a ser de por sí descriptivos, hacemos la traducción literal de los nombres en inglés, siempre que tengan significado en espaitol. (N. del T.)

LISTA DE LAS FAMILIAS COMUNES REPRESENTATIVAS DE PECES VIVIENTES 36
Figura 2.17 - "Herring" de mar profundo, Bathyclupea argentea. (Basado en Jordan y Evermann, 1900).
Figura 2 1 8 -Trucha de Nueva Zelandia, Galaxias brevipinnis. (Basado en Boulen- ger, 1904).
Figura 2.19 - Pez lagartija de bahía, Syno- dus foetens. @asado en Jordan y Ever- mann, 1900).
Figura 2.20 - Atele6pido (de mar profun- do), Ateleopus japonicus. (Basado en Goode y Bean, 1895).
Figura 2.21 - Gigantúrido (de mar pro- fundo), Giganturn vorax. (Basado en Regan, 1925).
Figura 2.22 - Tragador, Saccopharynx ampullaceus. (Basado en Jordan y Ever- mann, 1900).

36 LOS GRUPOS SUPERIORES DE PECES
Orden Salmoniformes Suborden Salmonoidei
Familia Salmonidae - truchas, salmones, y otros (Figuras 3.15b
Familia Plecoglossidae - “ayu” Familia Osmeridae - “smelt”
Familia Argentinidae - argentinos Familia Opisthoproctidae - “spookfish” Familia Bathylagidae - “deepsea smelt” Familia Alepocephalidae - “deepsea slickhead” Familia Searsidae - “searsid” Familia Bathylaconidae - batilacónidos
Familia Salangidae - salángidos Familia Retropinnidae - “southern smelts” Familia Galaxiidae - galáxidos (trucha de Nueva Zelanda, Figura
y 10.4)
Suborden Argentinoidei
Suborden Galaxiodei
2.18) Suborden Esocoidei
Familia Umbridae -- “mudminnow” Familia Esocidae - lucios
Familia Gonostomatidae - “lightfish” Familia Sternoptychidae - pez hacha de mar profundo Familia Photichthyidae - fotíctidos Familia Astronesthidae - pez de dientes romos de mar profundo
Familia Idiacanthidae -pez de ojos pedunculados y mar profundo Familia Malacosteidae - pez de mandíbulas sueltas y mar
Familia Melanostomiatidae - pez dragón sin escamas Familia Stomiatidae - pez dragón escamado, de las profundidades
Familia Chauliodontidae - pez víbora
Familia Giganturidae - gigantúridos
Suborden Stomiatoidei
(Figura 8.14)
profundo
marinas
Suborden Giganturoidei
Orden Gonorynchiformes Suborden Chanoidei
Familia Chanidae - “milkfishes” Familia Kneriidae - kneridos Familia Phractolaemidae - fractolémidos
Familia Gonorynchidae - gonorrínquidos Suborden Gonorynchoidei
Orden Cypriniformes (Ostariophysi) Suborden Characoidei
Familia Characidae - caracínidos” Familia Gymnotidae - anguila eléctrica (Figura 6.2b)
*Las bases proporcionadas para la identificación de las familias de este Suborden no están lo suficientemente fundamentadas como para garantizar más de las que aquí se reconocen.

LISTA DE LAS FAMILIAS COMUNES REPRESENTATIVAS DE PECES VIVIENTES 37
Figura 2.23 - Pez elefante, Mormyrus caballus. (Basado en Boulenger, 1904).
----""." ."_ _ _ _ _ _ _ "
Figura 2.25 - Anguila americana, Anguilla rostrata. (Basado en Jordan y Evermann, 1900).
Figura 2.24 - Carpa, Cyprinus carpio. (Basado en Hubbs y Lagler, 1941).
Figura 2.26 - Anguila holosáurida, Aldro- vandia macrochir. (Basado en Jordan y Evermann, 1900).
Figura 2.27 - Anguila espinosa de mar profundo, Notacanthus analis. (Basado en Jordan y Evermann, 1900).
Figura 2.28 - Saurio del Atlántico, Scom- beresox saurus. (Basado en Jordan y Evermann, 1900).
Figura 2.29 - Bacalao del Atlántico, Gadus morhua. (Basado en Jordan y Evermann, 1900).

38 LOS GRUPOS SUPERIORES DE PECES
Suborden Cyprinoidei Familia Cyprinidae - carpas común y dorada (Figuras 2.24, 6.4, 10.4 “creek chub”, y 10.6 “bitterling”)
Familia Gyrinocheilidae - girinoqueilidos Familia Catostomidae - carpa hocicona, matalote (Figura 5.3) Familia Homalopteridae - “hillstream loach” Familia Gastromyzonidae - “suckerbelly loach”
Familia Diplomystidae - bagres diplomístidos Familia Ictaluridae - bagres de agua dulce (Figuras 3.1 y 10.6 “cabeza de toro”) Familia Bagridae - bagres Familia Pimelodidae - bagres pimelódidos Familia Siluridae - bagres euroasiáticos, siluro Familia Schilbeidae - bagres schilbeidos Familia Amblycipitidae - amblicipítidos Familia Amphiliidae - anfílidos Familia Akysidae - aquísidos Familia Sisoridae - sisóridos Familia Clariidae - bagres de respiración aérea (Figura 8.10b) Familia Heteropneustidae - bagres ponzoñosos (Figura 8 . 1 0 ~ ) Familia Chacidae - cácidos Familia Olyridae - olíridos Familia Malapteruridae -- bagres eléctricos Familia Mochokidae - bagres de natación invertida Familia Ariidae - bagres marinos Familia Doradidae - bagres dorádidos acorazados Familia Aspredinidae - “banjo” o bagre partero (Figura 10.6) Familia Plotosidae -- bagre plotósido marino Familia Ageneiosidae ”ageneiósidos Familia Hypophthalmidae - hipoftálmidos Familia Helogeneidae - helogeneidos Familia Cetopsidae - cetópsidos Familia Trichomycteridae - bagres parásitos Familia Callichthyidae - bagres calíctidos acorazados Familia Loricariidae - bagres acorazados
Familia Synodontidae - pez iguala, pez lagartija Familia Harpadontidae - “bombay duck” Familia Alepisauridae - peces lanceta Familia Scopelarchidae - pez ojos perla Familia Myctophidae - pez linterna
Suborden Siluroidei
Orden Myctophiformes
Orden Cetomimiformes Suborden Ateleopoidei
Familia Ateleopidae -- ateleópidos de los fondos marinos (Figura 2.20)
Suborden Cetomimoidei Familia Cetomimidae - cetomímidos Familia Mirapinnidae - mirapínidos

LISTA DE LAS FAMILIAS COMUNES REPRESENTATIVAS DE PECES VIVIENTES 39
Figura 2.30 - Espinocha de tres espinas, Gasterosteus aculeatus. (Basado en Jordan y Evermann, 1900).
Figura 2.31 - Pez pipa, Pseudophallus starksi. (Basado en Jordan y Evermann, 1900).
Figura 2.32 - Pez remo, Regalecus glesne. (Basado en Jordan y Evermann, 1900).
Figura 2.33 - Pez sanguinario rayado, Fundulus majalis. (Basado en Jordan y Evermann, 1900).
Figura 2.34 - Pez “Priapium”, Noestethus amaricola. (Basado en Berg, 1940).
Figura 2.35 - Perca trucha, Percopsis omiscomaycus. (Basado en Jordan y Ever- mann, 1900).

40 LOS GRUPOS SUPERIORES DE PECES
Familia Eutaeniophoridae - euteniofóridos Familia Megalomycteridae - megalomictéridos
Familia Gibberichthyidae - “gibberfishes” Familia Barbourisiidae -- barburísidos Familia Rondeletiidae - rondelétidos
Suborden Rondeletioidei
Orden Beloniformes Suborden Scomberesocoidei
Familia Scornberesocidae - “saury” (Figura 2.28) Familia Belonidae - aguja, sierrita (Figuras 3.1 y 5.5b)
Familia Exocoetidae - - peces voladores Familia Hemiramphidae - pajarito (Figura 5 . 5 ~ )
Suborden Exocoetoidei
Orden Cyprinodontiformes Suborden Adrianichthyoidei
Familia Oryziatidae -- “ricefishes” Familia Adrianichthyidae - adrianíctidos Familia Horaichthyidae - horaíctidos
Familia Cyprinodontidae - “killifishes” (Figura 2.33
Familia Goodeidae - peces vivíparos mexicanos Familia Jenynsiidae - jenínsidos Familia Anablepidae - peces cuatro-ojos Familia Poeciliidae - peces vivíparos (Figuras 12.1 y 12.2)
Suborden Cyprinodontoidei
y 10.7)
Orden Gasterosteiformes Suborden Gasterosteoidei
Familia Gasterosteidae - espinochas (Figura 2.30 y 10.6) Familia Aulorhynchidae - “tube-snouts’’
Familia Aulostomidae - peces trompeta Familia Fistulariidae - peces corneta Familia Macrorhamphosidae - “snipefishes” Familia Centriscidae - peces camarón
Familia Syngnathidae - agujones y caballitos de mar (Figuras
Suborden Aulostomoidei
Suborden Syngnathoidei
2.31, 4.6, y 10.6f) Orden Lampridiformes
Suborden Lampridoidei
Suborden Trachipteroidei Familia Lamprididae - “opahs”
Familia Radiicephalidae - radicefálidos Familia Lophotidae - “crestfishes” Familia Trachipteridae - peces cinta Familia Regalecidae - peces remo (Figura 2.32)
Familia Stylephoridae - “tube-eyes” Suborden Stylephoroidei
Orden Beryciformes Suborden Stephanoberycoidei

LISTA DE LAS FAMILIAS COMUNES REPRESENTATIVAS DE PECES VIVIENTES 41
Figura 2.36 - Estefanobericido, Stephano- beryx monae. (Basado en Jordan y Ever- rnann, 1900).
Figura 2.37 - Berícido, Beryx splendens. (Basado en Jordan y Everrnann, 1900).
Figura 2.38 - "John Dory" americano, Zenopsis ocellata. (Basado en Jordan y Everrnann, 1900).
.". ."_"." Figura 2.39 - Pez cabeza de vbora, Channa striata. (Basado en Norman, 1931).
Figura 2.40 - Cuchia, Amphipnopscuchia. (Basado en Norman, 1931).

42 LOS GRUPOS SUPERIORES DE PECES
Familia Stephanoberycidae - peces espinosos de las profundida-
Familia Melamphaeidae - peces de escamas grandes y de las des marinas (Figura 2.36)
profundidades marinas Suborden Polymixioidei
Suborden Berycoidei Familia Polymixiidae - pez barbudo
Familia Diretmidae - dirétmidos Familia Trachichthyidae - traquíctidos Familia Berycidae “berícidos (Figura 2.37) Familia Monocentridae - pez piña de pino Familia Anomalopidae - peces ojo linterna Familia Holocentridae - peces ardilla
Familia Zeidae - “dory” Familia Grammicolepidae - gramicolépidos Familia Caproidae - “boarfish” Familia Antigoniidae - antigónidos
Orden Zeiformes
Orden Mugiliformes Suborden Mugiloidei
Suborden Sphyraenoidei
Suborden Polynemoidei
Suborden Atherinoidei
Familia Mugilidae - lisa
Familia Sphyraenidae - barracuda
Familia Polynemidae - barbudo, ratón
Familia Atherinidae - Peyerrey, pejerrey, gruñones Familia Melanotaeniidae - peces arco iris (de Australia) Familia Isonidae - isónidos Familia Neostethidae - “neostethid priapium fishes” Familia Phallostethidae - “phallostethid priapium fishes”
(Figura 2.34)
Suborden Percoidei Orden Perciformes
Familia Centropomidae - robalo Familia Percichthyidae - “temperate basses” (Figura 2.41) Familia Theraponidae - pez tigre Familia Serranidae - meros (Figura 4 . 6 ~ ~ ) Familia Grammistidae - peces jabón Familia Kuhliidae - “aholeholes” Familia Centrarchidae - peces sol (Figuras 3.1; 3.15; 6.3, y
Familia Percidae - percas (Figuras 2.1; 3.1; 8.3, y 10.6 “darter”) Familia Priacanthidae - ojón Familia Apogonidae -- cardenal Familia Branchiostegidae - “tilefishes” Familia Pomatomidae - anchoa, pez azul Familia Rachycentridae - esmedregal, cobia, bacalao Familia Carangidae -jurel (Figuras 2.42 y 10.11)
10.6)

LISTA DE LAS FAMILIAS COMUNES REPRESENTATIVAS DE PECES VIVIENTES 43
Figura 2.41 - Lobina rayada, MOrOne sa- xatilis. (Basado en Jordan Y Evermaw 1900).
......- ..."".
Figura 2.42 - Papelillo, Selene vomer. (Ba- sado en Jordan y Evermann, 1900).
Figura 2.43 - Pez loro arco iris, Scarus guacamaia. (Basado en Jordan y Ever- mann, 1900).
Figura 2.44 - Lenguado de invierno, Pseudopleuronectes americanus. (Basado en Jordan y Evermann, 1900).

44 LOS GRUPOS SUPERIORES DE PECES
Familia Coryphaenidae - dorado, doradilla (Figura 10.4b) Familia Bramidae - “pomfrets” Familia Lobotidae - chopa, biajaca de la mar, dormilona Familia Lutjanidae - pargos, huachinangos Familia Leiognathidae - “slipmouths” Familia Gerreidae - mojarra Familia Pomadasyidae - burro Familia Sparidae - sargo, pez pluma Familia Sciaenidae - corvina, totoaba Familia Pempheridae - “sweepers” Familia Ephippidae - peluquero, barbero Familia Kyphosidae - chopa, salema Familia Mullidae - salmonete, chivato Familia Monodactylidae - pez dedo Familia Bathyclupeidae - sardina de aguas profundas (Figura
Familia Toxodidae - “archerfishes” Familia Scatophagidae - “scats” Familia Chaetodontidae - pez mariposa (Figuras 4.6d y 5.5f) Familia Nandidae - “leaffishes” (Figura 4.6g) Familia Badidae - bádidos Familia Cichlidae - cíclidos (Figura 5.5d) Familia Embiotocidae - “surfperches” (Figura 10.6) Familia Pomacentridae - chopa, mojarra rayada Familia Cirrhitidae - tigre Familia Latridae - trompetero Familia Labridae - vieja, gallo, perro colorado Familia Scaridae - loro jabonero, pez loro, vieja lora
Familia Trichodontidae - “sandfishes” Familia Opistognathidae - “jawfishes” Familia Bathymasteridae - “ronquils” Familia Zoarcidae - “eelpouts” Familia Pholidae - “gunnels” Familia Mugiloididae - “sandperches” Familia Trachinidae - “weevils” Familia Percophididae - “flatheads” Familia Trichonotidae - “sandivers” Familia Dactyloscopidae - “sand stargazers” Familia Uranoscopidae - pez sapo Familia Chiasmodontidae - tragador de fondos marinos
Familia Echeneidae - rémora, pez pega (chupadores de tiburones;
2.17)
(Figura 2.43)
Suborden Echeneioidei
Figura 2.46) Suborden Ophidioidei
Familia Gadopsidae - pez negro de río Familia Brotulidae - lengua, lengua de vaca, bacalao, brotulas Familia Ophidiidae - lengua Familia Carapidae - pez perla

LISTA DE LAS FAMILIAS COMUNES REPRESENTATIVAS DE PECES VIVIENTES 45
Figura 2.45 - Anguila mastacembklica, Mastacembelus maculatus. (Basado en Boulenger, 1904).
Figura 2.47 - Pez globo del norte, Sphoe- roides maculatus. (Basado en Jordan y Evermann, 1900).
Figura 2.49 - Pez sapo leopardo, Opsanus pardus. (Basado en Jordan y Evermann, 1900).
Figura 2.51 - Mariposa marina, Pegasus umitengu. (Basado en Jordan, 1905).
Figura 2.46 -Tiburón succionador o rk- mora, Echeneis naucrates. (Basado en Jordan y Evermann, 1900).
Figura 2.48 - Pez adhesivo, Gobioesox ma- candricus. (Basado en Jordan y Evermann, 1900).
Figura 2.50 - Pejesapo, Lophius america- nus. (Basado en Goode y Bean, 1895).

46 LOS GRUPOS SUPERIORES DE PECES
Suborden Notothenioidei Familia Nototheniidae - “Antarctic blennies” Familia Bathydraconidae - pez dragón Familia Channichthyidae - pez hielo
Familia Blenniidae - pez de roca Familia Anarhichadidae - pez lobo Familia Clinidae - sargacero Familia Stichaeidae -- “pricklebacks” Familia Ptilichthyidae - “quillfishes” Familia Scytalinidae - cavador de grava Familia Zaproridae - “prowfishes”
Familia Icosteidae - “ragfishes”
Familia Schindleriidae - aquindléridos
Familia Ammodytidae - “sand lances”
Familia Eleotridae - dormilón Familia Gobiidae - gobio Familia Microdesmidae - pez gusano
Familia Kurtidae - “forehead brooders” (Figura 10.6)
Familia Acanthuridae - cirujano, lancero Familia Siganidae - pez conejo
Familia Gempylidae - macarela culebra Familia Trichiuridae - sable, cintilla, listón, machete Familia Scombridae - macarela, bonito, atún, sierra (Figura 3.1)
Familia Xiphiidae - pez espada Familia Luvaridae - luváridos Familia Istiophoridae - “billfishes” (“marlins”, “sailfishes”,
Suborden Blennioidei
Suborden Icosteoidei
Suborden Schindlerioidei
Suborden Ammodytoidei
Suborden Gobioidei
Suborden Kurtoidei
Suborden Acanthuroidei
Suborden Scombroidei
Suborden Xiphioidei
“spearfishes”) Suborden Stromateoidei
Familia Stromateidae - palometa (Figura 3.1 “harvestfish”) Familia Nomeidae - pez pastor Familia Tetragonuridae - “squaretails”
Familia Channidae - “snakeheads” (Figuras 2.39 y 8.10a) Familia Anabantidae - perca trepadora (Figura 10.6 “figh-
Familia Luciocephalidae - “pikeheads”
Suborden Anabantoidei
tingfish”)
Orden Gobiesociformes Suborden Callionymoidei
Suborden Gobiesocoidei Familia Callionymidae - “dragonets”

LISTA DE LAS FAMILIAS COMUNES REPRESENTATIVAS DE PECES VIVIENTES 47
Familia Gobiesocidae - cucharitas (Figura 2.48) Orden Tetraodontiformes (Plectognathi)
Suborden Balistoidei Familia Triacanthodidae - “spikefishes” Familia Triacanthidae - “triplespines” Familia Balistidae - pez puerco Familia Monacanthidae - lija (Figuras 3.1 y 6 . 2 ~ ) Familia Aracanidae - “keeled boxfishes” Familia Ostraciidae - torito, pez cofre
Familia Triodontidae - “pursefishes” Familia Tetraodontidae - pez globo, tambor (Figura 2.47) Familia Canthigasteridae - “sharpnose puffers” Familia Diodontidae - pez erizo, pez puerco Familia Molidae - “ocean sunfishes” (Figura 3.1)
Suborden Tetraodontoidei
Orden Pleuronectiformes Suborden Pleuronectoidei
Familia Bothidae - lenguado (oculado a la izquierda) Familia Pleuronectidae - lenguado (oculado a la derecha) (Figura 2.44)
Suborden Soleoide Familia Soleidae - sol, lenguado de río, tepalcate Familia Cynoglossidae - lengua, lenguado, lengiiita
Orden Scorpaeniformes Suborden Scorpaenoidei
Familia Scorpaenidae - escorpión, rocote Familia Triglidae - soldadito, angelito, lapón, vaca
Familia Platycephalidae - “flatheads”
Familia Hexagrammidae - “greenlings” Familia Anoplopomatidae - pez sable Familia Zaniolepidae - pez peine
Familia Cottidae - “sculpins” Familia Agonidae - “poachers” Familia Cyclopteridae - “lumpfishes” y “snailfishes”
Suborden Platycephaloidei
Suborden Hexagrammoidei
Suborden Cottoidei
Orden Mastacembeliformes Suborden Mastacembeloidei
Suborden Chaudhurioidei Familia Mastacembelidae - anguilas mastacembélidas (Figura 2.45)
Familia Chaudhuriidae - chadhuriidos
Familia Symbranchidae - falsa anguila, culebra de agua Familia Amphipnoidae - “cuchia” (Figura 2.40)
Familia Dactylopteridae - “flying gurnards”
Familia Pegasidae - “seamoths” (Figura 2.51)
Orden Synbranchiformes
Orden Dactylopteriformes
Orden Pegasiformes

48 LOS GRUPOS SUPERIORES DE PECES
Orden Percopsiformes Suborden Amblyopsoidei
Suborden Percopsoidei
Suborden Aphredoderoidei
Familia Amblyopsidae - “cavefishes” peces cavernícolas
Familia Percopsidae - “trout-perches” (Figura 2.35)
Familia Aphredoderidae - “pirate-perches’’ Orden Gadiformes
Suborden Gadoidei Familia Gadidae - merluza (Figura 2.29) Familia Merluciidae - merluza
Familia Melanonidae - “grenadiers” (o “rattails”; originalmente Suborden Melanonoidei
Macruridae) Orden Lophiiformes
Suborden Batrachoidei
Suborden Lophioidei
Suborden Antennarioidei
Familia Batrachoididae - pez sapo, cabezón (Figura 2.49)
Familia Lophiidae - “anglerfishes” (Figuras 2.50 y 3.1)
Familia Antenariidae - pez antenado, sapo, pescador Familia Ogcocephalidae - pez murciélago
Familia Ceratiidae - “seadevils” (Figura 10.4 “deepsea Suborden Ceratioide
anglerfish”)
REFERENCIAS ESPECIALES SOBRE LAS CATEGORfAS SUPERIORES DE LOS PECES
Bailey, R.M., y T.M. Cavender. 1971. (Fishes). En McGraw-Hill Encyclopedia of Science and Technology, 3a. ed., McGraw-Hill Book Company, Nueva York.
Barlow, G.W., K.F. Liem, y W. Wickler. 1968. Badidae, a new fish family-behavioural, osteo- logical, and developmental evidence. Jour. Zool., Londres, 156:415-447.
Berg, L.S. 1940. Classification of fishes both recent and fossil. Trav. Insf. Zool. Acad. Sci. URSS, 5:87-517. Reimpresión, 1947. Edward Brothers, Ann Arbor, Mich.
Birdsong, R.S. 1975. The osteology of Microgobius signatus Poey (Pisces:Gobiidae) with comments on other gobiid fishes. Bull. Florida State Mus., Biol., Sci., 19(3):135-187.
Bohlke, J.E., y C.H. Robins. 1974. Description of a new genus and species of clinid fish from the western Caribbean, with comments on the families of the Blennioidea. Proc. Acad. Nut. Sci. Phila., 126(1):1-8.
Cohen, D.M. 1964. Suborder Argentinoidei. En Bigelow, H.B., ed., Fishes of the Western North Atlantic. Mem. Sers Found. Mar. Res. No. l(4):l-70.
Colbert, E.H. 1955. Evolution of the vertebrates. John Wiley and Sons, Nueva York. Eaton, T.H., Jr. 1970. The stem-tail problem and the ancestry of chordates. Jour. Paleo.,
Gosline, W.A. 1966a. The limits of the fish family Serranidae, with notes on other lower
-. 1966b. Comments on the classification of the percoid fishes. Pac. Sci., 22(4):409-418. -. 1968. The suborders of perciform fishes. Proc. U.S. Nat. Mus., 124:l-78. -. 1971. Functional morphology and classification o f teleostean fishes. Univ. Press of
Greenwood, P.H. 1968. The osteology and relationships of the Denticipitidae, a family of
44~969-979.
percoids. Proc. Calif. Acad. Sci., ser. 4,33(6):91-112.
Hawaii, Honolulu.
clupeomorph fishes. Bull. Brit. Mus. (Nat. Hist.), Zool., 16:215-273.

REFERENCIAS 49
Greenwood, P.H., R.S. Miles, y C. Patterson, eds. 1974. Interrelationships o f fishes. Publicado por la Linnaean Soc. de Londres por la Academic Press, Nueva York y Londres.
Greenwood, P.H., D.E. Rosen, S.H. Weitzman, y G.S. Myers. 1966. Phyletic studies of tele- ostean fishes with a provisional classification of living forms. Bull. Am. Mus. Not. Hist., 131:339-456.
Jarvik, E. 1968. The systematic position o f the Dipnoi. Nobel Symp., 4:223-245. Jefferies, R.P.S. 1968. The subphylum Calcichordata (Jefferies 1967) primitive fossil chordates
Jordan, D.S. 1923. A classification of fishes including families and genera as far as known.
Lehtola, K.A. 1973. Ordovician vertebrates from Ontario, Contrib. Univ. Mich. Mus. Paleo.,
Lindberg, G.U. 1971. (trad. 1974) Fishes of the World, a key to families and a checklist. John Wiley and Sons (Iwael Program for Scientific Translations, Ltd.).
McAllister, D.E. 1968. The evolution of branchiostegals and the classification of teleostome fishes, living and fossil. Bull. Nat. Mus. Canada, 221:l-239. -. 1971. Old Fourlegs. A “Living Fossil”. National Museums of Canada, Nat. Mus. Nat. Soc., Zool. Div., Odyssey Ser. 1:l-25.
Miles, R.S. 1974. Relationships of acanthodians, pp. 63-103. En Interrelationships o f fishes, P.H. Greenwood, R.S. Miles, C. Patterson, eds. Publicado por la Linnaean Society de Londres por la Academic Press, Nueva York.
Monod, T. 1968. Le complexe urophore des poissons téléostéens. Mém. Inst. Fondamental Afrique noire, No. 81,705 pp.
Moss, M.L. 1964. The phylogeny of mineralized tissues. Int. Rev. Gen. and Exptl. Zool.,
Moy-Thomas, J.A., y R.S. Miles. 1971. Paleozoic fishes. 2a. ed. Chapman and Hall, Londres. W.B. Saunders Company, Filadelfia y Toronto.
Nelson, G.J. 1969. Gill arches and the phylogeny of fishes, with notes on the classification of vertebrates. Bull. Am. Mus. Nat. His., 141:475-552.
Nybelin, O. 1971. On the caudal skeleton in Elops with remarks on other teleostean fishes. Acta Reg. Soc. Sc. Lit . Gothob. (Zool.), 7:l-52.
Orvig, T., ed. 1968. Current problems of lower vertebrate phylogeny. Almquist and Wiksell, Estocolmo, 539 pp.
Patterson, C. 1975. The braincase of pholidophorid and leptolepid fishes, with a review of the actinopterygian braincase. Philos. Trans. Roy. Soc. London B. Geol. Sci.,
Roberts, T. 1974. Interrelationships of ostariophysans, pp. 373-395. En Interrelationships of fishes, P.H. Greenwood, R.S. Miles, C. Patterson, eds. Publicado por la Linnaean Society of London por la Academic Press, Nueva York.
with echinoderm affinities. Bull. Brit. Mus. (Nat. Hist.), Geol., 16(6):241-339.
Standford Uni. Publ. Biol. Sci., 3(2):79-243.
24(4):23-30.
1:298-332.
269(899):275-579.
Romer, A.S. 1966. Vertebratepaleontology. 3a. ed. Univ. Chicago Press, Chicago, 468 pp. “ . 1968. Notes and comments on “Vertebrate Paleontology ”. Univ. Chicago Press, Chicago,
304 pp. Schaeffer, B. 1967. Comments on elasmobranch evolution, pp. 3-35. En Sharks, skates, and
rays. Gilbert, P.W., R.F. Mathewson, y D.P. Rall, eds. Johns Hopkins Press, Baltimore, Maryland.
Stensio, E.A. 1969. Elasmobranchiomorphi Placodermata Arthrodires, pp. 71-550. En Traité de Paléontologie, 4( 2), J. Piveteau, ed. Masson et Cie, París.
Thomson, K.S. 1969. The biology of the lobe-finned fishes. Biol. Rev., 44:91-154. Tyler, J.C. 1974. Tetraodontiformes. Encycl. Britannica, 15a. ed., pp. 162-164. Weitzman, S.H. 1974. Osteology and evolutionary relationships of the Sternoptychidae, with
a new classification of stomiatoid families. Bull. Amer. Mus. Not. Hist., 153:327-478. Zangerl, R. 1974. Interrelationships of early Chondrichthyans, pp. 1-14. En Interrelationships
o f fishes. P.H. Greenwood, R.S. Miles, C. Patterson, eds. Publicado por la Linnaean Society de Londres por la Academic Press, Nueva York.


Capítulo 3
Anatomía básica de los peces
Hablando en términos biológicos, un pez está compuesto de diez sistemas forma- dos por Órganos que trabajan en coordinación para formar al individuo como un todo. Estos diez sistemas cubren al pez, manejan su alimento y arrojan al exterior los desperdicios; ellos integran los procesos biológicos del pez y lo relacionan con las condiciones del medio ambiente; ellos cuidan de su respiración y lo pro- tegen contra daños; ellos le sirven de soporte físico al cuerpo y le permiten efec- tuar ciertos movimientos; y, finalmente, su funcionamiento permite al pez per- petuarse como especie y, mediante la evolución, formar categorías de animales superiores.
ANATOMÍA EXTERNA GENERAL (FIGURAS 3.1 Y 3.2)
Forma (Figura 3.1; Capítulo 6 )
Comúnmente el cuerpo del pez tiene forma de torpedo (fusiforme), y en sección transversal es más a menudo ligera o francamente ovoide. En las especies de nata- ción libre la forma se aproxima a la que teóricamente debe tener un cuerpo para na- dar con perfección, es decir, donde la sección mayor se encuentre localizada a la altura del 36 % , si es que dividimos la longitud del cuerpo en cien partes y comenza- mos a contar desde el extremo cefálico; los contornos correrán hacia atrás, hacia el extremo caudal, suavemente, hasta terminar en la base de la aleta caudal. La forma de muchos peces se aparta moderada o totalmente de la típica. Estas varia- ciones hacen que algunos peces tengan forma de globo (globiforrnes- peces globo, Tetraodontidae), colubriformes (anguiliforme-Anguillidae), o hasta filamentosa en su forma general (filiforme - “spine eels”, Nemichthyidae). Algunos son aplanados
51

52 ANATOMf.4 BÁSICA DE LOS PECES
I * IDEAL
r : 1
ATENUADO
rugoso
Figura 3.1 - Variación de la forma del cuerpo entre los peces.
en exageración, de lado a lado (deprimidos peces mariposa, Chaetodontidae y lenguados, Pleuronectidae),* otros aplanados pero muy alargados (trachipterifor- mes, payasitos y obispos, Trachipteridae) y, otros más, aplanados de arriba a abajo (deprimidos - los torpedos, rayas y peces murciélago, Ognocephalidae, etc.).
A pesar del gran número de variaciones morfológicas que presentan, la organización general de la forma de los peces se basa en la simetría bilateral, como sucede en la mayoría de los vertebrados. Las mitades derecha e izquierda del cuer- po se corresponden, como si fuesen imágenes especulares una de la otra. Y, toda- vía más, hay una notable cefalizacihn en los peces, seguida de un cuerpo de cierta solidez que va a terminar, siguiendo líneas delicadas, hacia la cola o urosoma. La cola no es realmente un apéndice del cuerpo, sino parte de él mismo; las aletas, en cambio, son partes periféricas y anatómicas en la mayoría de los peces.
"Aquí consideraríamos a los lenguados como comprimidos, es decir, aplastados o apla- nados en sus lados izquierdo y derecho. (N. del T.)

ANATOMfA EXTERNA GENERAL 53
Cubierta protectora del cuerpo (Capítulo 4)
Un pez ordinario está cubierto con una piel relativamente delgada, lo suficiente como para que sea fácilmente eliminada para el consumo de aquél en la mesa o la disección en el laboratorio. La piel no presenta solución de continuidad cuando se une a las mucosas de los orificios naturales del cuerpo y se vuelve transparente cuando cubre la superficie de los ojos. La diversidad de colores que se observa en los peces se debe al color de las células, y el líquido mucoso que presenta re- sulta de la secreción de las células mucosas.
En muchos peces la piel es lisa, pero en otros está cubierta por escamas derivadas de la misma piel. &stas varían en tamaño, desde las muy pequeñas, casi microscópicas, hasta las muy grandes, visibles a distancia; en grosor varían desde las que son tan delgadas como la misma piel, hasta las que forman verdade- ras placas; en lo relacionado con la ornamentación, desde las simples hasta las complejas; de la extensión del cuerpo que cubren, se distinguen las que lo prote- gen parcialmente y las que lo hacen en forma total; en estructura, se identifican las no óseas de las óseas; y, por su fijación, las deciduas (caducas) de las que están firmemente implantadas en la piel. Las clases de escamas son muy útiles en la tarea de caracterizar a los grupos superiores de peces. La morfología de las esca- mas y el número de hileras que forman a lo largo del cuerpo o alrededor de él, son con frecuencia caracteres específicos y genéricos. Los ágnatos vivientes son des- nudos, es decir, no tienen escamas; los tiburones y especies afines tienen denticu- los, O sea escamas placoideas formadas de dentina; los peces óseos tienen varios tipos de escamas óseas.
Apéndices (Capítulo 5)
Los apéndices en los peces consisten en las aletas y los cirros (elongaciones carno- sas) que aparecen extraordinariamente desarrollados en el pez sargazo (Pterophryne) y el “leafy sea dragon” (Phyllopteryx, Figura 4.6f). Las aletas pueden ser clasifica- das como medias o pareadas.
Aletas medias (Figura 3.2). Las aletas radiadas que están alineadas con el eje ante- roposterior de un pez típico son las del lomo (aleta o aletas dorsales), la cola (aleta caudal) y el borde inferior del cuerpo precisamente detrás del orificio intestinal (aleta anal). Aunque por lo general todas las aletas citadas están presentes, alguna puede faltar en algunas clases de peces. También alineada al eje anteroposte- rior puede estar otra aleta, sin radios y carnosa, llamada aleta adiposa (como en las truchas, Salmonidae) o aletas reducidas a unas cuantas espinas libres (como en las espinochas, Gasterosteidae). La aleta anal puede estar transformada en un órgano intromitente, el gonopodio, que participa en las funciones reproductoras, o sea, la copulación (como en los “livebearers”, Poeciliidae, Figura 10.5b).
Aletas pareadas (Figura 3.2). Las aletas pectorales y las pélvicas (ventrales) están dispuestas por pares. Las pectorales están sostenidas por la cintura pectoral que se articula al cráneo y, en los peces con radios espinosos, están situadas a nivel más elevado en los lados del pez, en relación a las pélvicas. Aunque de ordinario están presentes en los peces, algunas veces las aletas pectorales faltan como en las

54 ANATOMfA BASICA DE LOS PECES
TIPOS DE PECES
Tiburón
Pez óseo
Figura 3.2 - Anatomía externa comparada de los grupos principales de peces vivientes. Los datos que siguen constituyen una l ista para el etiquetado. Aphdices: d l -- primera aleta dorsa¡, d2 segunda aleta dorsa!, c - aleta caudal (cd, caudal dificerca; chc - caudal heterocerca; cho -- caudal horno- cerca), a -- aleta anal, pi -- aleta pectoral, p2 - aleta peivica; drganos los sentid~s: N - orificio nasal, e -.- ojo, I I - línea lateral; Esutieletu: i j - man- dibula inferior, uj -- rnandihula superior, gc - cubierta branquial; Aberturas: m -- boca, s -- espiráculo; ga - abertura branquial, v - ano.

ANATOMfA EXTERNA GENERAL 55
en los clupeoideos, salmónidos, ciprinoideos) pero también pueden estar situa- das en posición anterior, justamente debajo de las pectorales, en posición torúcica (como en muchas especies de radios espinosos, los acantopterigios), o aun bajo la garganta, en posición yugular, como en los “blennies”. Las cinturas pectoral y pélvica están conectadas por ligamento cuando las aletas pélvicas son torácicas o yugulares. Las pélvicas faltan en las lampreas y mixinos (los cefalaspidomorfos vivientes) y han desaparecido en algunos otros peces como las anguilas (Angui- llidae, etc.). En los tiburones y otros peces relacionados están modificadas como gonopterigios (myxopterigia) (10.5), que sirven como órganos intromitentes durante la cópula. En los peces adheribles (Gobiesocidae), “suckerbelly loaches” (Gastromyzon) y otros peces, las aletas pélvicas forman parte de un órgano fijador que recuerda a una ventosa en el vientre.
Aberturas (Figura 3.2)
La boca, las aberturas branquiales (incluyendo el espiráculo en algunas formas primitivas) y la abertura del extremo caudal del intestino (ano) son los orificios más importantes relacionadas con el tracto digestivo en el cuerpo del pez. Las aberturas para los órganos de los sentidos incluyen las del conducto nasobucal o nasal (Figura 3.21) y pequeños poros sensoriales ampliamente distribuidos sobre la superficie del cuerpo.
Boca (Figura 3.2). La boca del pez está situada delante de la cabeza, en posición terminal, pero debido a ciertas adaptaciones su posición puede ser superior (hacia el dorso) o inferior (apenas o marcadamente sobrepasada por el hocico). Su tama- ño relativo puede ser pequeño (como en el “tube-snout” Aulorhynchus o en ciprínidos como Phoxinus) o muy grande (como en los tragadores de los fondos marinos y “gulpers”, Saccopharyngidae y Eurypharingidae, respectivamente). La boca está bordeada por labios que pueden llegar a formar una placa cartilaginosa (Campostoma), puede ser membranosa (como en la mayoría de los peces) o varia- damente carnosa y a menudo muy papilosa (rémora, Catostomidae). En la mayoría de los peces los labios no tienen escamas y están cubiertos con diminutos órganos sensoriales, pero en los tiburones y muchos otros peces relacionados, las escamas placoides que cubren su cuerpo se continúan hacia los labios para desarrollarse abruptamente y transformarse en los enormes dientes que se fijan en las mandíbulas.
Aberturas branquiales (Figuras 3.2 y 3.13). En los numerosos peces que tienen cubiertas branquiales (los peces operculados - los teleósteos en general) hay una abertura única en cada lado de la cabeza. Normalmente esta abertura está enfrente de la aleta pectoral, hacia adelante de la base, pero en los peces murciélago (Ogco- cephalidae), curiosamente, está por detrás. Las aberturas forman un Único par en las quimeras (Holocephali) y en algunos mixinos (como Myxine), ninguno de los cuales, sin embargo, tiene un aparato opercular óseo. En los otros mixinos, las aber- turas varían de 5 a 14 en cada lado. Todas las lampreas (Petromyzonidae) tienen 7. Casi todos los tiburones (Squaliformes) tienen 5, aunque algunos poseen 6 o 7; todas las rayas (Rajiformes y Torpediniformes) tienen 5.
Principalmente en el grupo de los seláceos (Chondrichthyes), hay un orifi- cio suplementario de las aberturas branquiales que va desde el exterior hacia la

56 ANATOMfA BASICA DE LOS PECES
cavidad faríngea. Es el espiráculo que está localizado entre la abertura branquial anterior y el ojo. Se localiza entre el arco hioideo y el cráneo y ha sido citado como evidencia de la evolución de la función del arco hioideo, cambiando de soporte branquial a soporte de la mandíbula, lengua y garganta. Dentro del espi- ráculo hay un pequeño penacho de filamentos branquiales (la pseudobranquia hioidea) que persiste a pesar de no ser funcional. Esta estructura es retenida en los peces óseos superiores aunque no tengan espiráculo.
Ano (Figuras 3.2 y 3.12). El ano u orificio vertedor de excreta de un pez está en la porción media de la línea ventral del cuerpo, generalmente en la segunda mitad, caudal, si tomamos en cuenta la longitud total del cuerpo, detrás de las bases de las aletas pélvicas y justamente en frente de la aleta anal. Raramente es encontrado en la parte anterior, como sucede en la perca pirata adulta (Aphredoderus), que lo tiene en posición yugular. En los tiburones y otros parientes así como en los peces pulmonados, el ano se abre en una depresión ubicada en la superficie ventral del cuerpo, la cloaca, que contiene los orificios de salida de los conductos urina- rio y genital, así como el del intestino. En la mayoría de los peces, sin embargo, las aberturas de los conductos urogenitales están en la superficie, detrás del ano.
Poros abdominales. Los poros abdominales se abren anteriormente en el vientre de algunos peces entre los cuales quedan incluidas las lampreas (Petromyzonidae) y tiburones (Squaliformes, etc.). Puede ser que estas aberturas no tengan origen idéntico en estos dos grupos, pero en ambos producen una comunicación entre la cavidad del cuerpo y el exterior. Normalmente están dispuestos en pares, uno a cada lado de la línea media, aunque en algunos individuos sólo uno está pre- sente. Parece ser que representan vestigios de orificios de salida de huevos, o huevos y esperma, o solamente éste, en formas primitivas. Los grupos superiores de peces los perdieron por entero. En algunas sardinas (Clupeidae) y otros teleós- teos, sin embargo, la vejiga gaseosa se abre posteriormente hacia el exterior me- diante un poro ubicado cerca del ano.
órganos sensoriales
Orificios nasales (Figuras 3.2 y 3.21). Uno o dos orificios nasales (nostrilos) en cada lado del hocico (condición dirrina) que desembocan en un saco ciego, repre- sentan en los peces, externamente, los órganos de la olfación. Sin embargo, en las lampreas y los mixinos (cyclostomos Cephalaspidomorphi) el orificio nasal es Único y en posición media (condición monorrina). La mayoría de los peces tienen las aberturas nasales a los lados del frente del hocico (dorsolateralmente). En otros, como los tiburones, rayas y torpedos (Elasmobranchii) los orificios están en la superficie ventral del hocico. Hay diferencias en la diminuta anatomía de las aberturas nasales que pueden ser una abertura ancha, en forma de surco, como en el “tiburón perro” (Squalus), o una parcial división de la abertura por una aleta de tejido para separar dos poros, uno incurrente (anterior) y otro excurrente (posterior), para llegar finalmente a separar la corriente de agua que penetra o sale, mediante el desarrollo de un poro especial para cada corriente; esto último ocurre en la mayoría de los teleósteos (Figura 3.21). A menudo el tejido carnoso valvular de los orificios nasales es de utilidad taxonómica.

ESQUELETO 57
En los peces pulmonados vivientes (Dipnoi) los orificios nasales incurrentes externos comunican mediante conductos con los orificios excurrentes que se encuentran en la cavidad bucal; esa clase de comunicación también se encuentra, raramente, en ciertos peces óseos superiores (Actinopterygii).
Ojos (Figura 3.22). Los ojos, que no pueden ser cerrados por carecer de párpados, están situados en órbitas, una en cada lado de la línea media de la cabeza del pez. Más a menudo los ojos son laterales, con campos de visión parcialmente inde- pendientes y movimiento relativamente limitado y también independiente. Los ojos son dorsales en muchos residentes de los fondos marinos, entre los cuales se cuentan las rayas (Rajidae), en la mayoría de “sculpins” (Cottidae), y los rape (Lophiidae). En los adultos de los lenguados y peces afines (Pleuronectifomes) los dos ojos están en un mismo lado de la cabeza. Los ojos pueden reducirse o desaparecer en los peces de las cavernas (Amblyopsidae y en el caracínido ciego cavernícola, Anoptichthys, entre otros).
A través de la piel transparente (conjuntiva) que cubre el ojo y a través de la córnea transparente del globo del ojo son visibles: a ) la abertura de la pupila del ojo y, a través de ella, el cristalino, que se encuentran dentro del globo del ojo, y b ) rodeando la pupila, el iris de bordes irregulares y coloreado.
drganos de la piel. Numerosas aberturas microscópicas de los órganos sensoriales de la piel se desarrollan en la superficie del cuerpo del pez. En muchos peces una serie de estos poros, extendiéndose a lo largo de una única serie a cada lado, desde la cabeza hasta la aleta caudal, comprende la línea lateral (Figura 3.2). El sistema de la línea lateral forma ramificaciones en la cabeza, incluyendo una arriba y otra debajo de la órbita de cada ojo (Figura 11.24). En algunos peces (incluyendo al lucio, Esox lucious, y los pescaditos de plata, Atherinidae) los poros y sus órga- nos sensoriales no están dispuestos en hilera sino esparcidos sobre el cuerpo. En los tiburones y especies afines (Elasmobranchii), hay porciones especializadas de la línea lateral, especialmente en la región del hocico. En algunos peces hay boto- nes gustativos en la piel y han sido también descritas estructuras tegumentarias sensibles al tacto; ninguna de éstas es visible a simple vista.
ESQUELETO (CAPfTULO 5)
En el esqueleto de los peces está incluida la notocuerda, los tejidos conectivos, el hueso, el cartílago, las escamas no óseas y componentes de los dientes como el es- malte y la dentina, células de sostén del sistema nervioso (neuroglia) y radios de las aletas (córneos en los tiburones, ceratotriquios, y escamosos y espinosos en los peces superiores, lepidotriquios y actinotriquios, respectivamente).
Las variadas clases de materiales esqueléticos están dispuestos en estructuras externas e internas, ambas con porciones blandas y duras.
Rasgos del esqueleto externo
Una parte del esqueleto del pez es, como ya se ha dicho, evidente mediante una inspección superficial externa. En el esqueleto tegumentario están incluidas las

58 ANATOMfA BASICA DE LOS PECES
escamas óseas de la piel, los radios de las aletas y tejidos conectivos que refuerzan la piel y la unen con la musculatura subyacente, el hueso y el cartílago. Evidentes también son algunas partes del esqueleto interno (endoesqueleto), principalmente los huesos superficiales de la cabeza y la cintura pectoral. fistas y otras partes del esqueleto interno serán descritas a continuación,
Esqueleto membranoso (Figuras 3.7 y 3.18)
Una envoltura de tejido conectivo une la piel y sus apéndices a la musculatura que se encuentra por debajo de aquélla; también a la parte firme o soporte del esque- leto. Esta envoltura es continua hacia las partes que coinciden con la linea media dorsal y línea media ventral, o sea el septo esqueletógeno medio. En la región caudal este septo medio divide al pez en dos mitades laterales y cubre las estruc- turas medias de ambos lados, como las vértebras caudales. A nivel de la cavidad del cuerpo el septo medio o axial, después de dividir al cuerpo en dos mitades laterales a lo largo del dorso, se divide él mismo para envolver las vértebras y se une al peritoneo parietal para continuar hacia el lado ventral siguiendo la pared interior del cuerpo. Hacia la mitad del lado ventral se vuelve visible en forma de una línea blanca de tejido conectivo: la Iinea alba.
El septo esqueletógeno horizontal se extiende, cerca de la parte media de cada lado, desde la envoltura superficial hasta el septo medio. Como resultado de la partición media y horizontal de la cola, ésta se ve formada por cuatro cua- drantes de masa muscular. La separación de los músculos en haces fuertemente unidos se debe a la presencia de los miosepta. Estos septa son fácilmente visibles entre los miómeros; paquetes regulares de músculo que se ven a los lados de un pez al que se le ha quitado la piel. Los términos descriptivos que han sido asigna- dos a las subdivisiones del esqueleto membranoso incluyen los siguientes:
a. Vaina perineural, que rodea al sistema nervioso central, su membrana externa es la duramadre, y la que está en contacto con la masa cerebral y la médula espina1 es la piamadre.
b. Perineruio, envuelve a los nervios. c. Pericondrio, rodea al cartílago, envolviéndolo. d. Periósteo, protege al hueso. e. Perimisio, cubre los músculos. f. Peritoneo, cubre los órganos de la cavidad del cuerpo (peritoneo visce-
g. Pericardio, envuelve el corazón y cubre la cavidad pericardial. h. Tendones y ligamentos, unen ciertos músculos entre sí o al esqueleto
i. Mesenterios, sostienen en posición los órganos en la cavidad del cuerpo.
ral) y delimita la cavidad visceral (peritoneo parietal).
basal.
El esqueleto membranoso situado detrás de la cabeza divide al pez en numerosos “compartimentos” y sirve para sujetar éstos uno contra otro; también para unir los músculos entre sí y con el esqueleto basal. Es obvio que muchas de las divisiones del esqueleto membranoso citadas arriba están unidas por tejido conectivo a una o más divisiones; en consecuencia, kstas son más o menos conti- nuas entre sí.

ESQUELETO 59
Notocuerda
La notocuerda no es esencialmente firme o dura ni membranosa en su consisten- cia. Aparece durante los primeros estadios del desarrollo embrionario del pez (y de los demás cordados) en forma de una cuerda alargada de tejido que se localiza en la línea media y el axis del cuerpo. Cuando ha alcanzado su desarrollo total presenta una vaina pericordal relativamente resistente (parecida a la envoltura resistente de una salchicha), llena de células turgentes por la presencia de un lí- quido viscoso. La notocuerda suele ser cilíndrica a todo lo largo (peces pulmona- dos y condrósteos); en ocasiones es muy larga (un esturión adulto, de poco más de un metro de largo, Acipenser, tiene su notocuerda de casi un metro de longitud y de un centrímetro y medio de diámetro). En otros casos puede estar reducida a una serie de pequeñas masas de material, una entre cada par de vértebras adya- centes (en muchos peces óseos superiores). En algunos peces, como los salmóni- dos, es moniliforme (recordando la disposición de las cuentas en un hilo); en los tiburones y formas afines (Chondrichthyes) aparece esta condición por la invasión de la vaina pericordal por cartílago. En los peces gnatostomados, generalmente la vaina pericordal da lugar a la aparición de variadas formas de vértebras a partir del estado embrionario.
Esqueleto basal axial (Figura 3.3)
El esqueleto basal axial de un pez está compuesto por el cráneo, la columna ver- tebral, las costillas y los huesos intermusculares.
Cráneo (Figura 3.4). Los cefalaspidomorfos vivientes (lampreas y mixinos) tienen esqueleto basal axial relativamente primitivo en algunos aspectos y muy especiali- zado en otros. El esqueleto axial es cartilaginoso e incluye el cráneo, las vértebras y los radios de las aletas. El cráneo de la lamprea (Petromyzonidae) está compuesto de una caja o estuche para el cerebro (neurocráneo) y cápsulas para los órganos de la olfación, visión y audición y equilibrio (son, respectivamente, las cápsulas olfa- tiva, óptica y Ótica). Un canasto branquial sostiene la faringe y las bolsas branquia- les, y otros cartílagos fijan en posición el rostro, el embudo bucal alrededor de la boca y la lengua. El cráneo en los mixinos (Myxinidae) no es tan completo como el de las lampreas; los “arcos” que dan lugar a la compleja estructura branquial en forma de canasto de las lampreas están reducidos a un simple anillo cartilaginoso que rodea el conducto que va desde el esófago hacia el exterior en los mixinos.
La caja craneal de tipo tiburón (elasmobranquio) está compuesta por un cráneo cartilaginoso (condrocráneo) y los arcos viscerales (branquiocráneo) y sus derivados. El condrocráneo está formado de una caja incompleta en la porción dorsal (neurocráneo) y dos pares de cápsulas para los órganos de los sentidos, es decir, para los de la audición y la visión. Debido a la gran abertura (fontanela anterior) que vuelve incompleta la parte dorsal del neurocráneo de estos peces, se dice de ellos que tienen agujeros en sus cabezas. Se piensa que los arcos visce- rales llegaban a ocho primitivamente. El primero se perdió, o posiblemente esté representado por los cartílagos labiales de algunos tiburones. El segundo arco está re- presentado por la mandíbula superior (cartílago pterigocuadrado o maxilar) y la mandíbula inferior (cartílagos de Meckel o mandibular). El tercer arco en esta

ANATOMfA BASICA DE LOS PECES
v) O I- W -1 Lu 3
B W W o

ESQUELETO 61
I
N w a
”’--
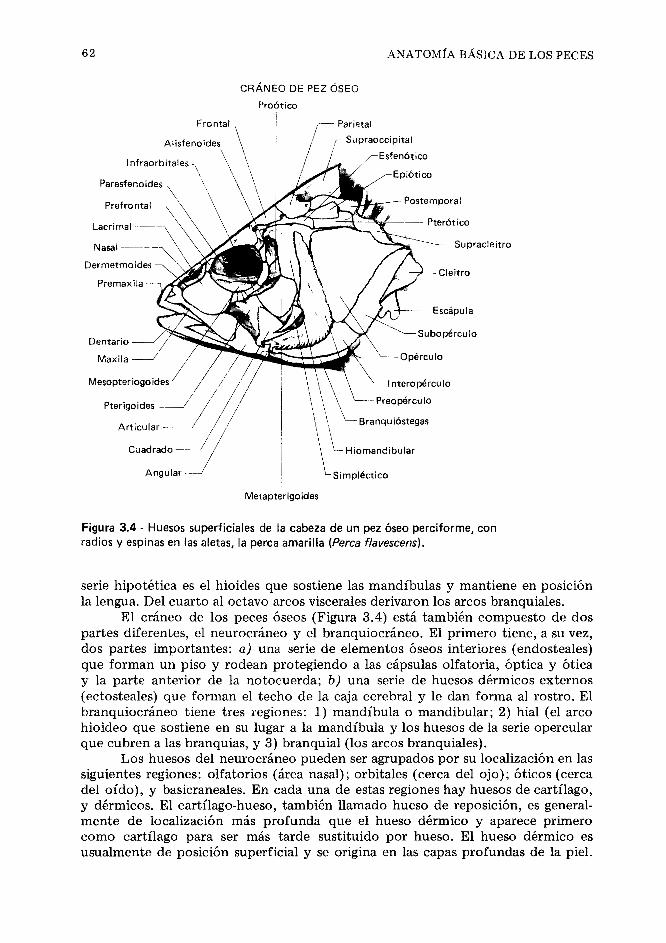
62 ANATOMfA BASICA DE LOS PECES
CRÁNEO DE PEZ ÓSEO
Proótico
Alisfenoides
lnfraorbitales -
Parasfenoides
Lacrimal ~
- Pterótico
Derrnetrnoides
Articular - -~~~
Cuadrado Hiomandibular
I Metapterigoides
Figura 3.4 - Huesos superficiales de la cabeza de un pez óseo perciforme, con radios y espinas en las aletas, la perca amarilla (Perca flavescens).
serie hipotética es el hioides que sostiene las mandíbulas y mantiene en posición la lengua. Del cuarto al octavo arcos viscerales derivaron los arcos branquiales.
El cráneo de los peces óseos (Figura 3.4) está también compuesto de dos partes diferentes, el neurocráneo y el branquiocráneo. El primero tiene, a su vez, dos partes importantes: a) una serie de elementos óseos interiores (endosteales) que forman un piso y rodean protegiendo a las cápsulas olfatoria, óptica y Ótica y la parte anterior de la natocuerda; b ) una serie de huesos dérmicos externos (ectosteales) que forman el techo de la caja cerebral y le dan forma al rostro. El branquiocráneo tiene tres regiones: 1) mandíbula o mandibular; 2) hial (el arco hioideo que sostiene en su lugar a la mandíbula y los huesos de la serie opercular que cubren a las branquias, y 3) branquia1 (los arcos branquiales).
Los huesos del neurocráneo pueden ser agrupados por su localización en las siguientes regiones: olfatorios (área nasal); orbitales (cerca del ojo); óticos (cerca del oído), y basicraneales, En cada una de estas regiones hay huesos de cartílago, y dérmicos. El cartílago-hueso, también llamado hueso de reposición, es general- mente de localización más profunda que el hueso dérmico y aparece primero como cartílago para ser más tarde sustituido por hueso. El hueso dérmico es usualmente de posición superficial y se origina en las capas profundas de la piel.

ESQUELETO 6 3
Estos dos tipos de huesos están representados por piezas pareadas err posición bilateral; solamente unos pocos no son pareados.
La región olfatoria del neurocráneo permanece parcialmente cartilaginosa en el adulto. Sin embargo, el hueso de cartílago que contiene incluye el etmoides o paretmoides pareado y colocado a ambos lados; el preetmoides, también en número par y colocado hacia los dos lados, y el etmoides medio. El paretmoides está cubierto dorsolateralmente por los prefrontales; el etmoides y los preetmoi- des están cubiertos ventralmente por el vómer (prevómer) y dorsolateralmente por los nasales. Los huesos dérmicos de esta región son los prefrontales y nasales en número par cada uno, y el vómer impar que lleva dientes en muchos peces.
La región orbital tiene, como huesos de cartílago, el orbitoesfenoides medio y los alisfenoides pareados. Los frontales cubren estos tres huesos dorsolateral- mente. Los pequeños huesos dérmicos que rodean la región orbital son llamados co- lectivamente circumorbitales o huesos escleróticos. Todos son pareados, incluyendo los huesos frontales que forman parte de la porción dorsal de la cavidad orbital. Los supraorbitales y muchos infraorbitales también están presentes. Aunque los infraorbitales tienen nombres, se los numera convencionalmente, comenzando con los más anteriores: núm. 1, lagrimal o preorbital; núm. 2, yugal; núm. 3, in- fraorbital verdadero; núm. 4, 5 y 6 (dermofenóticos), que llevan la rama infraor- bital del órgano sensorial de la línea lateral (Figura 11.24).
La región Ótica tiene muchos huesos de cartílago, a menudo difíciles de identificar aunque el neurocráneo sea cuidadosamente desarticulado. Están pre- sentes los esfenóticos pareados (autosfenóticos), pteróticos (autopteróticos), proóticos, epióticos, opistóticos, exoccipitales y el supraoccipital Único está situado en posición dorsomesial, extendiéndose en dirección caudal. Huesos aso- ciados de origen dérmico son los parietales (pareados), postemporales y supra- cleitros. En este complejo de piezas óseas el hueso esfenótico está cubierto dorso- lateralmente por dermoesfenóticos y el postemporal conecta la cintura pectoral con el neurocráneo.
En la región basicraneal el Único hueso de cartílago es el basioccipital, con una superficie cóncava posterior que se articula con la primera vértebra. El Único hueso dérmico es el paraesfenoides medio, cuya forma imita vagamente la de un crucifijo. Este hueso largo se extiende desde la región olfatoria hasta el basi- occipital.
La región mandibular del branquiocráneo está compuesta por el oroman- dibular (mandíbulas superior e inferior) y huesos asociados, todos pareados. Los huesos de cartílago que constituyen el mecanismo de la mandíbula son los palati- nos, cuadrados y metapterigoideos. Los palatinos son huesos provistos de dientes. Los cuadrados vienen a ser las “claves” de la articulación del pterigoideo, huesos de la porcibn superior del hioides y la mandíbula inferior. Los huesos dérmi- cos de la mandíbula superior son las premaxilas, las maxilas, los pteriogoideos y los mesopterigoideos. Las premaxilas llevan dientes en la mayor parte de 10s peces dentados, pero las maxilas son típicamente excluidas de la abertura bucal en 10s peces provistos de aletas con espinas y, consecuentemente, no tienen dientes en ellas (condición edentada). En muchos peces hay un pequeño hueso de origen dérmico, la supramaxila, fijada posterodorsalmente a la maxila. En la mandíbula inferior los únicos huesos de cartílago son los articulares, que se articulan poste- riormente con los cuadrados. El extremo posterior de cada articular es algo redon- deado y ancho; el extremo anterior se adelgaza para adaptarse a la hendidura de

64 -4NATOMfA BASICA DE LOS PECES
posición inclinada y en forma de V del dentario. Los huesos dérmicos de la man- díbula inferior son los angulares pareados y los dentarios. Cada angular es un hueso pequeño fijado a la parte posteroventral del articular. Los principales componentes de la mandíbula inferior son los dentarios y tienen forma subtri- angular. Los dentarios de muchos peces llevan dientes.
La región hioidea está compuesta de huesos pares e impares de los tipos cartilaginoso y dérmico. Los huesos pares de cartílago incluyen los hiomandibu- lares, simplécticos, interhiales, epihiales, ceratohiales e hipohiales. El basihial (glosohial, entoglosal o hueso de la lengua) es un hueso de cartílago medio. Los huesos dérmicos de la región son los preopérculos en número par, los opérculos, los subopérculos, los interopérculos y los branquióstegos, y el urohial, Único.
El hiomandibular se articula dorsalmente con el esfenótico, proótico y el pterótico; se une anterodorsalmente con el metapterigoideo. El hiomandibular está conectado al interhial y al simpléctico por tejido conectivo y cartílago, y también se articula con el opérculo. El simpléctico es pequeño y encaja en un angosto surco del hueso cuadrado. El interhial se conecta al epihial y hace que éste se conecte a su vez al hiomandibular y al simpléctico. El epihial es un hueso triangular situado entre el interhial y el ceratohial. El ceratohial y el epihial tienen formaciones óseas que se interconectan produciendo una ligazón entre esos dos huesos. El hipohial consta de dos partes y se articula con el ceratohial. La serie de huesos operculares que, situados en cada lado de la cabeza integran la cubierta protectora de las branquias y limita posteriormente la mejilla, está compuesta del preopérculo, opérculo, interopérculo y subopérculo. El opérculo es el hueso más grande en esta serie y se extiende en dirección posterior. Los radios branquióste- gos sostienen la membrana branquióstega. Se mantienen fijos por su extremo anterior a los huesos hioideos, principalmente los ceratohiales. Su número y tipo de fijación difieren en los diferentes grupos de peces (Tabla 2.3). El urohial es un hueso fuerte que posee componentes planos horizontales y verticales que le dan el aspecto de una T invertida viéndolo en sección transversal. Se sitúa en el septo medio de la garganta, en los fuertes músculos hipobranquiales situados entre las ramas de la mandíbula inferior.
La región branquia1 del esqueleto básico está hecha enteramente de hueso de cartílago. Los componentes pares que preceden ventralmente en un arco bran- quia1 típico son los faringobranquiales, epibranquiales, ceratobranquiales e hipo- branquiales, así como el Único basibranquial. Es típico que en los peces bseos haya tres basibranquiales, tres pares de hipobranquiales y cuatro pares de cerato- branquiales, epibranquiales y faringobranquiales. En muchos peces perciformes hay dientes en las superficies faríngicas de varios de estos huesos branquiales. Los dientes dorsales faríngicos son suprafaríagicos, y los ventrales, infrafaríngicos. Los dientes de la garganta (ventrales faríngicos) que caracterizan los “minnows” (Cyprinidae) y los succionadores (Catostomidae) estSn situados en el quinto par de ceratobranquiales.
Columna vertebral y costillas (Figura 3.5). La espina dorsal de un pez está com- puesta de una serie de segmentos llamados vértebras. Generalmente a cada segmento corporal corresponde una vértebra (diplospondyli, la cola de algunos tiburones, Elasmobranchii). Aunque en algunas especies estas vértebras son más o menos semejantes, están generalmente modificadas de acuerdo a la región del cuerpo donde se encuentren. Una o dos (el atlas y el axis) localizadas en el extremo ante-
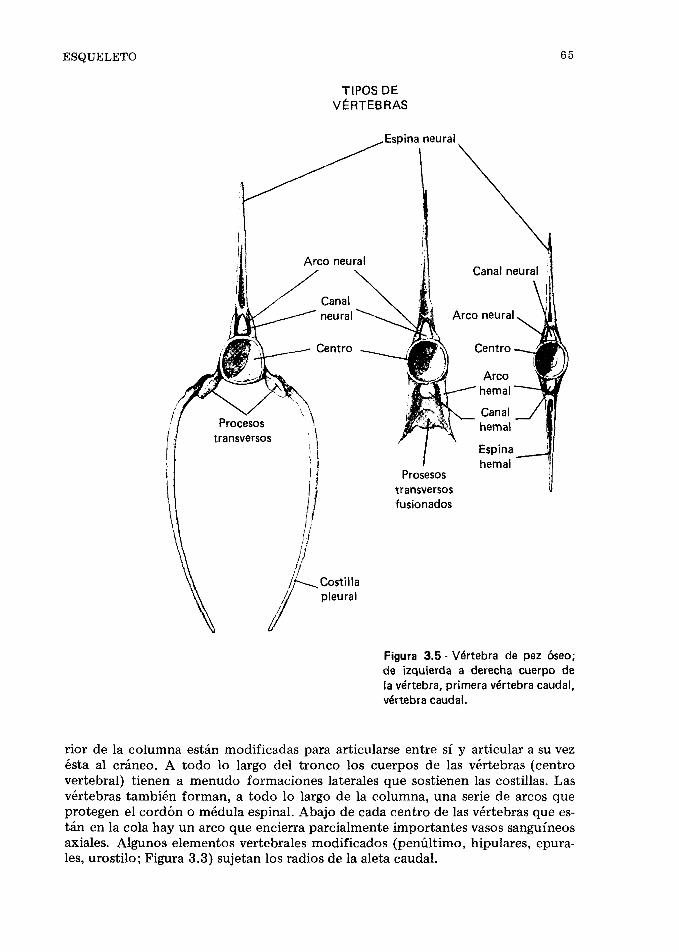
ESQUELETO 65
TIPOS DE VERTEBRAS
Figura 3.5 - Vertebra de pez óseo; de izquierda a derecha cuerpo de la vértebra, primera vertebra caudal, vertebra caudal.
rior de la columna están modificadas para articularse entre sí y articular a su vez ésta al cráneo. A todo lo largo del tronco los cuerpos de las vértebras (centro vertebral) tienen a menudo formaciones laterales que sostienen las costillas. Las vértebras también forman, a todo lo largo de la columna, una serie de arcos que protegen el cordón o médula espinal. Abajo de cada centro de las vértebras que es- tán en la cola hay un arco que encierra parcialmente importantes vasos sanguíneos axiales. Algunos elementos vertebrales modificados (penúltimo, hipulares, epura- les, urostilo; Figura 3.3) sujetan los radios de la aleta caudal.

66 ANATOMfA BASICA DE LOS PECES
En las lampreas vivientes (Petromyzonidae) las vértebras son primitivas, se han vuelto cartilaginosas, carecen de centro, están degeneradas y son menos com- pletas que las de los peces sin mandíbulas, ya extinguidos. Los mixinos vivientes (Myxinidae) ni siquiera tienen esas vértebras incompletas. Los restos de la columna vertebral están formados po: una vaina. cartilaginosa que rodea la notocuerda. En el grupo de los tiburones (Elasmobranchii), los centros de las vértebras del tronco cartilaginoso llevan arcos dorsales (neurales) y procesos transversos jbasip6fisisj donde se unen pequeiías costillas. En este grupo, los centros de las vkrtebras cau- dales llevan arcos neurales y ventrales (hemales). Los holocéfalos, relacionados con los tiburones, o sea las quimeras, tienen centros incompletos en forma de anillos de cartílago calcificado pero no tienen costillas (de aquí que no haya for- maciones que sostengan costillas en el centro, que está muy reducido en tamaño). Las vértebras de los peces óseos (Qsteichthyes) están osificadas.
Por su estructura, las vértebras de los peces son menos complejas que las de los vertebrados terrestes. Es típico que la articulación sea el resultado de la simple aposicibn de los centros, en lugar de existir protuberancias que produzcan una complicada articulación de vértebras entre si. Esto ocurre porque el agua propor- ciona un soporte al cuerpo del pez, cosa que no sucede con los animales terrestres.
Las costillas de 10s peces, de acuerdo a su localización, son de dos tipos: dorsales y ventrales. La presencia de las costillas ventrales se limita a los peces óseos (Qsteichthyes) y se originan en las parapbfisis del tronco ventral. Su desa- rrollo se efectfia en las particiones (,miosepta) de tejido eonectivo de los haces laterales musculares (miómems) en el perit,oneo. Las costillas dorsales también están localizadas en la regi6n del tronco pero se adosan en el septo esqueletógeno horizontal, donde &te se une a los mioseptos; de aquí que a veces sean llamadas “’huesos intermusculares”.
Con la excepción de los c6falospid .,xnorfos y los holocéfalos, otros grupos de peces vivientes (Elasmobranchii y Os$eicht,hyes) tienen todos costillas dorsales. Además, muchas especies poseen también costillas ventrales, la mayoría de los peces 6seos superiores tienen huesos intermuscularrs.
lluesos intermusczllares. Muchos peces tienen huesos pequefios y cielgados, de diversas formas: en l!.)s mioseptos; los grupos de peces que los tienen incluyen a las sardinas (CJupeidae), lucios (Esocidae), algunos salmones y especies afines [Salmonidae), “Suckers” (Catost,omidae) y carpas (Cyprinidae), para nombrar solamente a unos cuantos. En la trucha uei género Salmo los huesos son muy rectos y corren en sentidos lateral y caudal desde la coluxvlna vertebral, cor1 la cual cada hueso interrnuscular tiene u m conexibn ligamentosa. En las sardinas (como en Alosa y Dorosorna) icts huesos de esta clase tienen a menudo forma de C y corren en sentido lateral en los mlosepia desde una csnexibn tendinosa con una espina neural vertebra:. Fn la familia de los lucios (Esocidae) y en algunos de los Catostomidae los huesos intermusculares tienen forma de horqueta al estar hifurcados, es decir, tienen f.’orma de Y. Un brazo de cada Y está conectado por ligamento a una espina neura1 vertebrai, desde donde se dirige hacia afuera. El otro brazo y la base de la Y son m$s o menos paralelos a 13 columna vertebra! en un rniosepto epaxial. La presencia de huesos intermusculares de forma de Y en una especie no impide Ía ocurwxia simultinea de otras formas, Son estos huesos intermusculares los que pruvocan dificuitsdes al consumir comidas hechas con pescado.

ESQUELETO 67
Esqueleto de los apéndices
El soporte esquelético de las aietas medias y pares difiere fundamentalmente en que las aletas pectorales y pélvicas poseen un soporte en forma de cinturbn: en tanto que las primeras carecen de A ,
Soportes de las aletas dorsal y anal (Figura 3.3) . Los soportes esquelkiicos internos de 10s tipos comunes de aleta dorsa; y mal en los peces 6seos (Osteichtiyes) sor: series de tres bursas cada una, Un pterigi?lporo (axonostoj proxima! derivado d.c% an interceural, dorsai, o un interhemal, ve2rltraI, se encuentra hacia adentro, ado. shndose en el septo esqueletógeno medio. En posicibn intermedia, en camino hacia el radio de aleta que va a ser sostenido, se er,cuentra un pterigibforo. Miis afuera de l a serie, articulándose con el radio de aleta, se encuentra un pterigióforo distal (baseostoj. En los tiburones y peces afines (Chondrichtyes) las aletas dorsa- les están sostenidas internamente por cartilagos basales situados en la parte media, y descansan sobre las espinas neurales de las vertebras adyacentes; hacia afuera se extienden cartílagos cartilaginosos radiales para sostener los numerosos radios de aleta, aunque la espina de la aleta (como en el galludo espinoso, Squalus acanthias) puede unirse al cartílago basal directamente. En las lampreas (Petromyzonidae), los radios cartilaginosos de aleta tienen solamente soporte membranoso. En algu- nos peces el pterigióforo proximal se articula con el radio de aleta que tenga más cerca y el pterigióforo distal se articula con el siguiente radio de aleta más caudal; el elemento intermedio se ha perdido.
Soportes de la aleta caudal. Hay suficientes elementos que permiten distinguir los principales tipos de suspensión esquelética para las aletas caudales, de tal modo que no pudieron haber dejado de interesarse los ictiólogos en la clasificación de aquéllos, ni en la teorización sobre su secuencia evolutiva. Ordenados por sus nombres, las principales clases son:
a. Proterocerca (dificerca): probablemente es el tipo original donde la línea axial del cuerpo se continúa hacia la aleta caudal, dividiéndola supuesta- mente en una porción dorsal y otra ventral, ambas iguales. Como ejemplos tenemos a las lampreas y los mixinos (Cephalaspidomorphi, Figura 3.3).
b. Heterocerca: la línea axial del cuerpo se levanta al llegar al extremo cau- dal, sosteniendo a la aleta caudal sobre elementos ventrales; está presente este tipo de aleta en los arcaicos ostracodermos y placodermos, los tibu- rones y especies afines (Chondrichthyes) (Figura 3.3) y peces óseos primi- tivos (Chondrostei y Holostei) (en forma abreviada en el último). Los tipos de aleta proterocerca y heterocerca aparecen transitoriamente durante el desarrollo embrionario de muchos peces que presentan el siguiente tipo de aleta.
c. Homocerca: la línea axial del cuerpo, al terminar en la penúltima vérte- bra, es continuada por un urostilo (el producto de la fusión de varios elementos vertebrales) y típicamente sostiene los radios de aleta cauda- les siguiendo una vertical apenas redondeada, que da término d urostilo y sus elementos óseos relacionados (siendo esta una combinación de pie- zas superficialmente denominada como placa hipural, que raramente consta de un solo elemento, como sucede en los Gasterosteidae). En los

68 ANATOMfA BASICA DE LOS PECES
peces óseos superiores es absolutamente prevalente la presencia de una aleta caudal casi o totalmente homocerca (Teleostei; Figura 3.3).
Soportes de la aleta pectoral (Figura 3.6). Las lampreas y los mixinos (Cephalas- pidomorphi) no solamente carecen de aletas pares, sino también de las cinturas que las sostienen. En los tiburones y especies relacionadas (Chondrichthyes), sin embargo, la cintura pectoral está compuesta básicamente de un resistente cartí- lago coracoescapular que tiene forma de U ancha. Los elementos coracoides pares forman la parte ventral de la U, con articulaciones para aleta en sus esqui- nas, y las extremidades superiores vienen a ser las partes escapulares. En las rayas, torpedos y quimeras los cartílagos escapulares se fijan a las vértebras mediante sus extremos dorsales.
En los peces óseos (Osteichthyes) la cintura pectoral está compuesta de huesos de cartílago y huesos dérmicos. Los elementos presentes en la mayoría de los peces óseos en calidad de huesos de cartílago son el coracoides y los esca- pulares en número par cada uno y cuatro pares de radiales; y, como huesos dérmi- cos en número par, los postemporales, los supracleitros, los cleitros y los postclei- tros. Comenzando a describir desde la articulación dorsal de la cintura pectoral con el neurocrine0 y siguiendo hacia el lado ventral, el postemporal es un hueso en forma de horqueta, bifurcado, con las dos prolongaciones dirigidas hacia ade- lante. Una prolongación se articula con el epiótico y la otra con el opistótico. El postemporal se reúne posteroventralmente al supracleitro. El postcleitro se adapta ventralmente al cleitro y se extiende dentro del tronco muscular lateral; a menudo consta de dos partes. El coracoides es un hueso a primera vista de forma triangular que se adapta a la escápula por arriba y al cleitro mediante tejido conectivo. La escápula es un hueso cuadrangular y presenta una gran abertura o foramen. La ma- yoría de las radiales que sostienen a los radios de aleta y, en ciertos peces, algunos radios de aleta, se articulan con el borde ventral de la escápula. La cintura pectoral está reducida de tamaño o bien ausente entre los peces globo (Tetraodontidae).
La cintura pectoral está conectada a los radios de la aleta pectoral mediante varios elementos esqueléticos intermediarios. En los tiburones y especies afines, están siguiendo una secuencia hacia el exterior a cada lado del pez, siendo así que se pueden nombrar sucesivamente al propterigio, el mesopterigio y el metapteri- gio, además de numerosos radiales que en su turno llevan los radios de las aletas. En los peces de aletas lobuladas (Crossopterygii), el mesopterigio persiste y se alarga para formar, más o menos, un soporte axial para los radios, creando enton- ces las bases lobuladas, en número par, dando lugar así al nombre descriptivo de estos peces. Estos elementos son generalmente reducidos en los peces óseos supe- riores (Actinopterygii), con la excepción del “bichir” primitivo (Polypterus) en el cual existe una condición lobulada distintiva. El pez óseo típico tiene solamente cuatro radiales pequeños en forma de reloj de arena en cada lado.
Soportes de las aletas pélvicas (Figura 3.6). La cintura pélvica en los tiburones y grupos afines (Elasmobranchii) consiste en una simple barra cartilaginosa, deno- minada hueso isquiopúbico, soporta la radialia con sus radios de aleta. En las qui- meras (Holocephali) esta barra es doble, como sucede con su derivado óseo de la cintura pélvica de los peces lobulados primitivos (Crossopterygii) y los peces pul- monados (Dipnoi). En los peces óseos (Osteichthyes) la cintura pélvica consiste en un par de huesos de cartílago, los basipterigios, separados o fusionados en

ESQUELETO 69
CINTURAS DE LAS ALETAS PARES
(cintura p6lvica)
CINTURA PECTORAL
Cleitro 1 (cintura pectoral)
Basipterigio
Aleta p6lvica
CINTURA PÉLVICA
Figura 3.6 - Cinturas pectoral y pblvica de un pez óse0 con espinas y radios, la perca amarilla (Perca flavescens).

5 con una flexión hacia adelante y dos hacia atrás, siendo tres de curvatura
amplia. La inclinación interna de todo el miómero es craneal. El miómero típico
piscino es de la forma: con flexiones abruptas. Las flexiones de miómeros
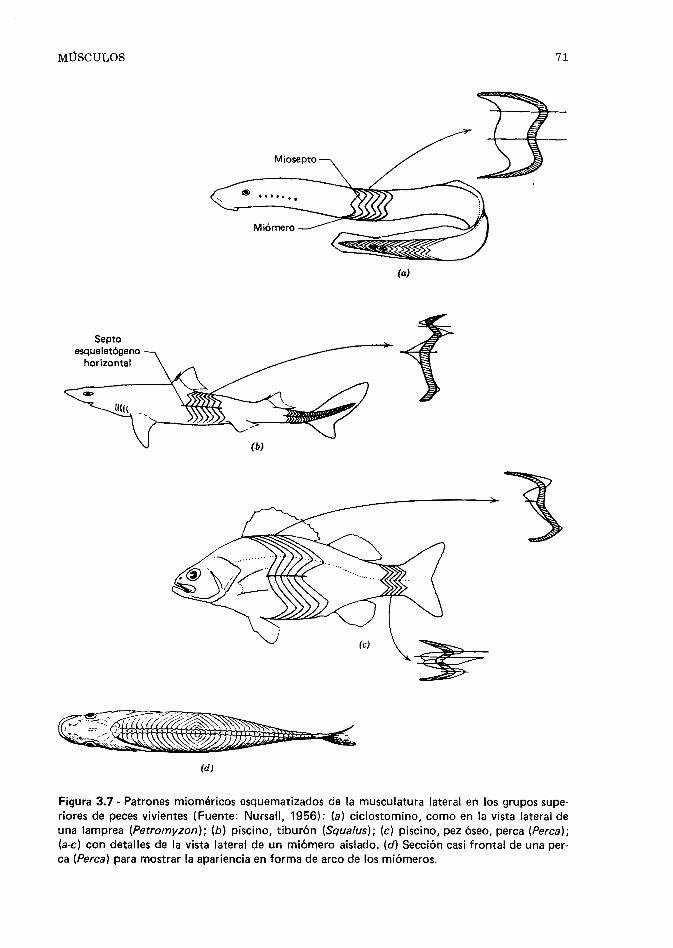
M ~ S C U L O S 71
Septo
Figura 3.7 - Patrones miomdricos esquematizados de la musculatura lateral en los grupos supe- riores de peces vivientes (Fuente: Nursall, 1956): (a) ciclostomino, como en la vista lateral de una lamprea (Perrornyzon); (b) piscino, tiburón (Squalus); (c ) piscino, pez óseo, perca (Perca); (a-c) con detalles de la vista lateral de un miómero aislado. ( d ) Sección casi frontal de una per- ca (Perca) para mostrar la apariencia en forma de arco de los miómeros.

12 ANATOMfA BASICA DE LOS PECES
sucesivos forman, internamente, concavidades cónicas. Las aberturas terminales de los conos miran hacia atrás y sus ápices en dirección craneal. Los extremos abier- tos de las flexiones miran hacia adelante pero apuntan hacia atrás, en dirección caudal. Para el estudio de la anatomía superficial, sin embargo, la clasificación que va de acuerdo a la localización es más conveniente, aunque para propósitos com- parativos, se debe prestar mucha atención al origen embriológico. La siguiente caracterización está basada en la disposición de clases tomando en cuenta la loca- lización.
Es bueno recordar que los músculos individuales, especialmente los esque- léticos, son mejor conocidos por su acción, localización y/o su origen e inserción.
Músculos esqueléticos del tronco (Figuras 3.7 y 3.8)
Los músculos del tronco se insertan firmemente en el esqueleto basal y son de utilidad primordial para efectuar los movimientos de las partes esqueléticas y para la locomoción. Los miotomos, o sea, esos prominentes haces de músculos de los lados del tronco, son visibles y conocidos como carne del pez cuando éste ha sido despojado de la piel o está preparado frente a nosotros en forma de tajada (en sección transversal). Las particiones delgadas que unen a los miotomos han sido ya descritas y denominadas miosepta (partes del esqueleto membranoso). Los míotomos aparecen segmentariamente como miomeros en los embriones. De estos miomeros, en posiciones apropiadas en el cuerpo, derivan:
a. Los músculos oculomotores: tres pares por cada ojo (Figura 3.23). b. Los músculos hipobranquiales: generalmente en el piso de la faringe, las
mandíbulas, el hioides y los arcos branquiales (importantes como exten- sores; Figura 3.8).
c. Los lnúsculos branquioméricos: de la cara, mandíbulas y arcos branquia- les (importantes como constrictores; Figuras 3.8 y 3.9).
d . Músculos apendiculares: músculos radiales en las bases de los radios de aleta y protractores, retractores, elevadores, depresores, aductores y abductores en las aletas pares, etc. (Figuras 3.10 a 3.12).
En un pez adulto los segmentos de la musculatura lateral del tronco se extienden desde la piel hasta el eje del cuerpo y está limitada en la línea media por el septo medio esqueletógeno. Por otro lado, se extiende a todo lo largo desde el occipucio (detrás del cráneo), la cintura pectoral y la garganta, hasta la base de la aleta caudal. Al orientarse y dirigirse hacia la línea media, los haces de múscu- los laterales no se extienden en ángulos rectos en relación a la línea sagital del cuerpo, pues siempre se inclinan en diversos grados hasta interceptar el plano que se encuentra sobre esa línea sagital, formando ángulos agudos con él. Lo que re- sulta de lo anterior es que cuando una aguja de disección es introducida en la cola siguiendo ]la difección de un ángulo recto en relación con el plano sagital, llegará a un miotomo, atravesándolo; luego a través de otro, y partes de otro, y así suce- sivamente, hasta llegar al septo esqueletógeno medie. La consecuencia de la natu- raleza estructural en forma de cufia de los miotomos, que les obliga a traslaparse, es que la sección transversal de un pez siempre muestra las superficies cortadas de muchos miotsmos.

MOSCULOS 73
M ~ S C U L O S DE LA CABEZA D E L T I B U R ~ N
Figura 3.8 - Vistas lateral y ventral de los músculos superficiales de la región cefálica de un tiburón perro espinoso (Squalus acanrhias). (Fuente: Stockard, 1949).
Las dos masas musculares laterales más importantes están separadas por el septo esqueletógeno horizontal (Figura 3.18). La masa superior (epaxial) en cada lado del plano sagita1 se apoya sobre el septo axial horizontal y la ventral (hipaxial) debajo del mismo. Una tercera masa de músculo, relativamente oscuro y en forma de cuña delgada, se encuentra en, cerca o extendida arriba y abajo de la intersec- ción del septo esqueletógeno horizontal y la cápsula de tejido conectivo del cuer- po, bajo la piel. Esta masa muscular ha sido llamada de varios modos, como “carne negra” o “músculo rojo” y, a menudo, cuando se trata de peces que se alimentan en el fondo de los depósitos de agua como la carpa (Cyprinus carpio), “franja lodosa” o “línea lodosa”. Este tejido muscular oscuro es notable en la cola de muchos peces óseos y termina antes de llegar a la cintura pectoral. El nombre técnico M. lateralis superficialis es el que se le ha dado; es a menudo grasoso (por ejemplo en el salmón asiático, Oncorhynchus tshawytscha). Las fibras musculares

7 4 ANATOMfA BASICA DE LOS PECES
difieren bioquímicamente de otros músculos y tienen diferentes propiedades contráctiles.
Los músculos profundos del tronco, de menor masa en proporción a la de los laterales, son los supracarinales y los infracarinales. Los supracarinales se ex- tienden desde la región escapular de la cintura pectoral hacia atrás hasta la aleta caudal, más o menos en una posición media axial (para flexionar el cuerpo dorsal- mente). Los infracarinales son músculos ventrales longitudinales que se apoyan en cualquier lado de la línea media ventral desde la garganta hasta la aleta caudal (para flexionar el cuerpo ventralmente y para mover la cintura pélvica y la aleta anal).
Musculatura esquelética de la cabeza (Figuras 3.8 y 3.9)
En la cabeza de un pez la musculatura está asociada primariamente ya sea con las mandíbulas o con los arcos branquiales. Pero hay componentes superficiales y profundos para ambos que no solamente difieren entre los grupos superiores de peces, sino hasta en las especies afines. Solamente se da aquí una introducción al estudio de los músculos superficiales más evidentes en dos peces; uno de ellos, el tiburón, que tiene las mandíbulas sostenidas por el arco hioideo (condición hiostílica) y el otro, un pez óseo, que no tiene las mandíbulas sostenidas de esa manera (condición autostílica).
Cuando el rostro de un tiburón es despojado de la piel muestra muchos músculos mandibulares (Figura 3.8) entre los cuales el masticador de la mandíbula inferior (M. adductor mandibuluris) es el más evidente. A nivel de las branquias los constrictores externos (dorsal y ventral) son superficialmente más notables. Músculos mandibulares semejantes son visibles en un pez óseo (Figura 3.9), pero
Elevador del arco Dilatador
Prenlaxila
Mandibula
/
mandibular 'v' Aductor mandibular
"-

MOSCULOS 7 5
algunos músculos especiales se han vuelto útiles para el protector de las branquias, el opérculo.
Músculos esqueléticos de las aletas medias (Figuras 3.10 y 3.11)
Los músculos de las aletas medias sirven para que el pez pueda desplazarse al moverlas y para maniobrar en el agua (mientras es impelido mediante la acción de otros músculos). Superficialmente estos músculos, para las aletas dorsal y anal, están dispuestos por pares: el protractor (para levantar) y el retroactor (para deprimir); los inclinadores laterales (para doblar) en cada radio de aleta desde cada lado; y, de la misma manera, para cada uno de los radios de aleta de los que están cercanos al plano medio, un erector hacia adelante y un depresor hacia atrás (Fi- gura 3.10). La aleta caudal tiene las masas laterales de músculo fijadas mediante tendones a su base. Además, de la musculatura superficial lateral nacen interna- mente secciones de músculo que se han especializado para mover a la aleta caudal: para doblarla y levantarla (flexores dorsales), y para bajarla (flexores ventrales), así como para extenderla y estrecharla como si fuese un abanico (flexores interfila- mentales entre los radios de aleta) (Figura 3.11).
Músculos esqueléticos de las aletas pares (Figuras 3.8 y 3.12)
Son de interés aquí las masas musculares relativamente pequeñas pero muy especia- lizadas que nacen como secciones de los miomeros axiales embriónicos para servir a las aletas pares del mismo modo como otros músculos pequeños lo hacen con las aletas medias. Haces superficiales de músculo apendicular son visibles cerca de las bases de las aletas cuando la piel es removida. Ellos son responsables de los movi- mientos de los apéndices que son independientes de los movimientos del tronco.
SUPERFICIALES
Membrana de la aleta", PROFUNDOS
Erector /-"- dorsal
Retractor /" dorsal
Radio blando
Miosepto J' Masa muscular epaxial Miómero
Figura 3.10 - Músculos superficiales y profundos de la aleta media de un pez (aleta dorsal del salmón chinense, Oncorhynchus tshawytscha). (Basado en Greene y Greene, 1914).

76 ANATOMfA BASICA DE LOS PECES
Flexor caudal epaxial dorsal
superior Aductor Espina neural de la vertebra
inferior Flexor -;-z4
-3:
caudal ventral
superficie1
... "
, 3 - " " _- ?" ?!
Flexor caudal ventral inferior
Figura 3.11 - Musculatura profunda de la aleta caudal de un pez óseo (cola del salmón chinense, Oncorhynchus rshawytscha). (Basado en Greene y Greene, 1914).
Protractor
Masa muscular
profundo profundo
Figura 3.12 - Músculos superficiales de las aletas pblvicas y cintura p6lvica en un pez óseo (región p6lvica del salmón chinense, Oncorhynchus rshawytscha). (Basado en Greene y Greene, 1914).
Los músculos de las aletas pares son, básicamente? abductores (producen movimiento hacia afuera) y aductores (hacia adentro), y están provistos de apén- dices como delgadas hojas de músculo entre los radios de aleta (para doblarlas) y otros que sirven para sostener o mover la cintura (Figuras 3.8 y 3.12). Cuando las aletas pares tienen otras funciones, es decir, que no se limitan a maniobrar el cuer- po; se especializan y sirven también para conducir el esperma hacia la hembra (el abrazo o amplexus e introducción como en los machos de tiburón y especies afi- nes? Elasmobranchii).

BRANQUIAS Y VEJIGAS GASEOSAS 77
Músculo cardíaco
Los principales componentes del corazón son el músculo y el tejido conectivo. El músculo cardíaco es de color rojo oscuro, en contraste con el músculo esquelético cuyo color varía del blanquecino al rosado (raramente café rojizp), de acuerdo a las especies. Su contracción es involuntaria. La masa muscular es más gruesa en las paredes del ventriculo; el atrio o auricula tiene paredes relativamente delgadas. La musculatura del corazón (miocardio) está cubierta externamente por el epicardio e internamente por el endocardio, ambos componentes esqueléticos membranosos del pericardio.
Músculos lisos
En los peces, como ocurre generalmente en todos los vertebrados, los músculos involuntarios lisos están localizados en muchos órganos diferentes. Quedan inclui- dos los siguientes:
a. Los del tracto digestivo: fibras longitudinales y circulares, responsables de producir movimientos y desplazamiento del alimento en ei tracto (peristalsis); las vejigas gaseosas también tienen estas dos clases de fibras.
b. Los de las arterias: fibras circulares que mantienen la presión sanguínea. c. Los de los conductos reproductores y excretores: desplazan los productos. d. Los del ojo: acomodación de la visión mediante movimientos en el lente
(cristalino) y regulación de la intensidad de la luz (dilatación o contrac- ción del iris); que difieren en desarrollo y función en los diferentes gru- pos de peces.
BRANQUIAS Y VEJIGAS GASEOSAS (CAPiTULO 8; FIGURAS 3.13 A 3.15)
Las branquias en las lampreas y mixinos (ciclóstomos ágnatos) están contenidas en bolsas (de ahí el término marsipobranquios que significa “branquias en bolsas” y se aplica a los ágnatos). Cada bolsa tiene una abertura interna en la faringe y otra que lleva hacia afuera. Otras partes que conducen el agua en las lampreas durante su estado larvario y en los mixinos incluyen el vestíbulo oral, la boca y la faringe. En los mixinos (Myxinidae) un conducto pituitario conecta el exterior con la fa- ringe por el lado dorsal. En la lamprea adulta (Petromyzonidae) la faringe queda dividida desde su porción anterior dorsal por una membrana de tejido o velo. El tubo ventral respiratorio que así se forma lleva a las bolsas branquiales vía las aberturas branquiales internas. Las lampreas tienen siete pares de aberturas exter- nas para las branquias. Algunos mixinos tienen muchas bolsas branquiales en un lado, cada una con una abertura externa y otra interna (de cinco a catorce en Eptatretus). En otros mixinos (como en Myxine) los seis pares de bolsas bran- quiales se abren por cada lado en un conducto común, el que a su vez se abre ha- cia afuera casi a nivel de la sexta bolsa. Esta disposicibn se hace funcionalmente paralela a las cubiertas operculares de las branquias en las quimeras y los peces óseos. Cada mixino tiene, además, un tubo (el conducto esofagocutáneo) después

78
TIPOS DE BRANQUIAS
ANATOMfA BÁSICA DE LOS PECES
Lamprea Tiburón Pez óseo
Figura 3.13 - Diagrama de la organización branquial entre los peces mostrado en secciones frontales: ac, cartílago anular; bbc, cartílago branquial forman- do un canasto; e, esófago; eba, abertura branquial externa; ga, arco branquial; gc, cubierta branquial (op6rculo); gf, filamento branquial de una hemibran- quia; gp, bolsa branquial; gr, rastrillo branquial; ha, arco hioideo; iba, abertura branquial interna; ibs, septo interbranquial; ma, arco mandibular; ph, faringe; rm, techo de la boca; S , hocico, sd, disco suctor; sp, espiráculo; t, dientes. (Tiburón y pez óseo basados en Kingsley, 1926).
de las bolsas branquiales, pero solamente en el lado izquierdo. Este canal lleva indirectamente del esófago al exterior, a través del conducto respiratorio izquier- do, en los géneros que tienen solamente una abertura branquial, o a través de la última abertura branquial, de mayor tamaño en el lado izquierdo, en los géneros que poseen muchas aberturas branquiales.
En los gnatostomados las branquias continúan siendo los principales órga- nos de la respiración, aunque hay innovaciones especializadas suplementarias que han aparecido en muchos grupos (notablemente en los peces pulmonados que pe- riódicamente, y de acuerdo con cierta estación del año, aprovechan la adaptación de una vejiga gaseosa para el intercambio respiratorio con el aire atmosférico). Los arcos branquiales llevan consigo las branquias en los gnatostomados. En los tibu- rones y especies afines (Elasmobranchii), las branquias están contenidas en cavida- des que recuerdan a las bolsas branquiales de las lampreas y los mixinos. Los septos interbranquiales (Figura 3.13 y 8.1) que separan las cavidades en los Chondrich- thyes están reducidos en grados variables en los Qsteichthyes donde las branquias de todos los arcos yacen en una cámara más o menos común. Los septos son en-

BRANQUTAS Y VEJIGAS GASEOSAS 79

80 ANATOMfA BASICA DE LOS PECES
TIPOS DE ABERTURA DE LA VEJIGA GASEOSA
2 n d u c t o c c - > -
Vejiga gaseosa
pneumático
udigertiuo Tracto
(a) Esturiones y la mayoría de los peces de radios blandos
(b) Catanes y el pez de aleta ondulada
(c ) Caracino predador
(d) Pez pulmonado australiano
(e) Peces de Reed
(f) Peces pulmonados africano y sudamericano
Figura 3.14 - Diagramas que muestran las variaciones de las relaciones de la vejiga gaseosa con el tract0 digestivo en los peces fisóstomos:
esturiones (Acipenser); catanes (Lepisosteus), y pez de aleta ondulada (Arnia) con revesti- miento rugoso; percas trepadoras (frythrinus); pez pulmonado australiano (Neo- cerarodus) con revestimiento al- veolado; los polípteros (Polyprerus) y peces caña (Calamoichrhys); y los peces pulmonados africano y sudamericano (Proroprerus y Le- pidosiren, respectivamente).
(Fuente: Dean, 1895).

TRACTO DIGESTIVO 81
TRACTO DIGESTIVO (CAPfTULO 5; FIGURA 3.15)
La estructura básica del tracto digestivo de los vertebrados fue establecida desde .$os peces. La siguiente es una lista de lo más sobresaliente que encuentra un bo- cado de alimento que se desplaza a lo largo del tracto de un teleósteo ordinario de radios espinosos como uno de los meros (Serranidae), percas (Percidae), peces sol (Centrarchidae), o cíclidos (Cichlidae).
a. Boca bordeada por mandíbulas dentadas y rodeada internamente por válvulas orales.
b. Cavidad oral con dientes vomerinos y palatinos en el techo y lengua con dientecillos linguales en su superficie.
c. Faringe con dientes faríngicos en cojinetes hacia los lados de los arcos branquiales que miran a la garganta y branquispinas que protegen las aberturas branquiales internas.
d . Esófago (gaznate). e. Estómago. f. Píloro (válvula pilórica seguida de aberturas hacia los ciegos pilóricos en
muchos peces). g. Intestino corto, pasando las aberturas de los ductos que vierten al inte-
rior del intestino la bilis y el jugo pancreático, y sigue hacia adelante a lo largo de por lo menos una curva grande en forma de S, para llegar a la si- guiente sección.
h. Intestino largo. i. Ano.
Entre los peces óseos hay, sin embargo, muchas adaptaciones especiales del tracto digestivo (Capitulo 5).
Las lampreas y mixinos (ciclóstomos) se alejan más notablemente de la estructura fundamental del tracto digestivo por la ausencia de mandíbulas, de un estómago bien definido y de curvaturas en el intestino. En una lamprea pará- sita, un par de glándulas “salivales” relativamente grandes vierten al exterior me- diante ductos situados bajo la lengua; la secreción de estas glándulas retarda la coagulación de la sangre del huésped y también disuelve al tejido (efecto histolí- sico). El más sorprendente alejamiento de lo normal en los tiburones y especies afines, es el desarrollo de una válvula espiral (Figura 5.8) cuya presencia sustituye a la curvatura del intestino, para alargar el recorrido del alimento. Una válvula similar también aparece en Latimeria, el Único pez viviente que posee aletas lobu- ladas. También está presente una válvula espiral en algunos teleósteos primitivos como en el tzotzín (“10-libras”), Elops. La estructura básica puede ser visuali- zada si imaginamos un sacacorchos fabricado con alambre plano y bastante ancho, adaptado a las paredes en el interior y a todo lo largo de un cilindro, en este caso el intestino. En este ejemplo, el sacacorchos correspondería a la válvula espiralada. En las lampreas existe un doblez en la pared intestinal (tiflosol) que se extiende en la luz del tubo digestivo y de manera efectiva facilita tanto la digestión como la absorción de los alimentos.

C S
-U al
VI m E 8 .- t

TRACT0 DIGESTIVO
O t O U U
a w D a r o 3 II: t-
83
U Q
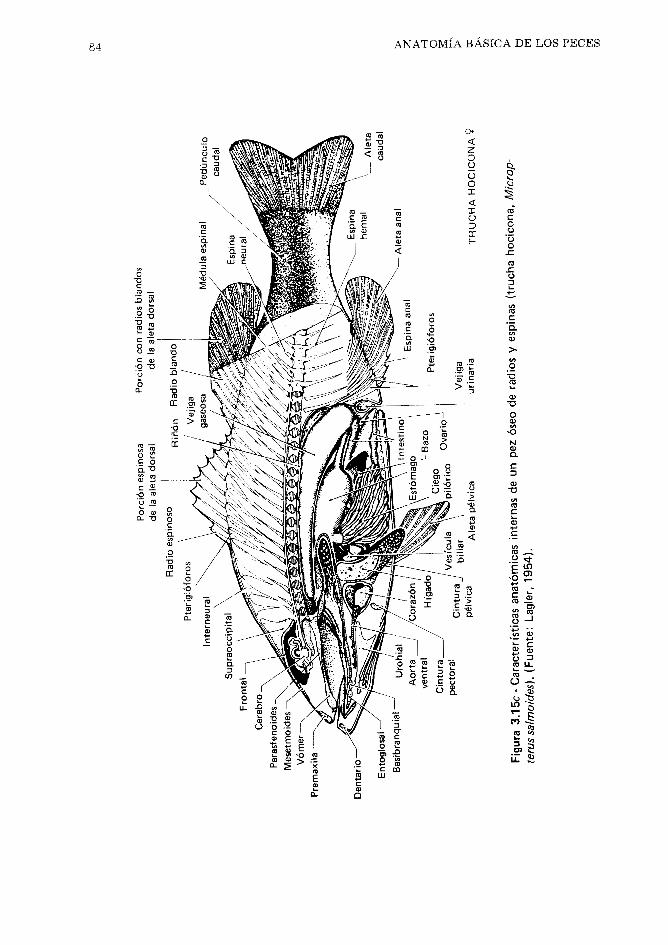
84 ANATOMfA Bk3ICA DE LOS PECES
'O u c w .- Y a
O ._ o, U
C .O IC U
51
E . C .-
O .- P U
Ln m S .- o W
U W
O
.O % N
B S 3
U al
In m E f" al C .-

SISTEMA CIRCULATORIO 86
SISTEMA CIRCULATORIO (CAPÍTULO 7 ; FIGURAS 3.16 U 3.17)
La sangre en los peces circula mediante un sistema tubular, más o menos continuo, de corazón y vasos. El corazón es una bomba con válvulas que impulsan la sangre hacia las branquias para su aireación. Teniendo arcos aórticos al llegar o salir de las branquias, la sangre arterial es finalmente distribuida en los demás tejidos mediante vasos capilares. La sangre venosa procedente de los tejidos regresa al corazón, aunque la que fluye a través de los riñones y el hígado es primero dis- tribuida por medio de capilares en los sistemas renal (excepto en los ciclósto- mos) y portahepático, respectivamente. La sangre sale del corazón a través de las arterias y se introduce a ese órgano por las venas.
Sangre
La sangre de los peces, al igual que ese mismo tejido de los demás vertebrados, consta de dos partes: una fluida, la otra sólida. La parte fluida es el plasma en donde las partes sólidas, o sea, las células sanguíneas, están suspendidas y son transportadas. Las células son rojas (eritrocitos) y blancas (linfocitos, leucocitos). La hemoglobina de los glóbulos rojos aumenta notablemente la capacidad de la sangre para transportar oxígeno. No todos los peces, sin embargo, poseen células rojas y hemoglobina. Ciertos peces del Antártico (Chaenichthyidae; “icefishes” o peces cocodrilo blanco) tienen sangre incolora desprovista de eritrocitos. La sangre de las anguilas jóvenes (larva leptocéfala) es también incolora. Más aún: el pigmento sanguíneo de la lamprea, Petrornyzon, no es la hemoglobina típica de otros vertebrados.
Corazón (Figura 3.16)
El corazón del pez está estructurado para permitir, fundamentalmente, sólo una circulación en lugar de dos, como ocurre en los mamíferos. Es típico que al regre- sar la sangre al corazón se introduzca a un seno venoso, pase a través de la auricula o atrio, y sea impulsada hacia las branquias para su aireación mediante las paredes musculares relativamente gruesas del ventriculo. El anterior es el sistema que apa- rece en los ciclóstomos. En los Chondrichthyes existe además una base muscular contráctil provista de válvulas (el cono arterial) en la aorta ventral, en la porción donde sale del ventriculo. El sistema es igual al de los tiburones y especies afines en los peces óseos superiores, pero la primera sección de la aorta ventral es, típica- mente, de paredes delgadas y provistas de válvulas (el bulbo arterial), y es elástica en lugar de contráctil, agrandándose y encogiéndose alternativamente en respues- ta a los cambios de la presión sanguínea que resulta de la contracción (sístole) y reposo (diástole) que se produce alternadamente en el ventriculo. El cono es redu- cido y reemplazado anteriormente por el bulbo no contráctil, con muchas variacio- nes, en la serie de teleósteos.
La aorta ventral en un pez es de posición media, por debajo de las branquias. De ella se desprenden las arterias branquiales aferentes que irrigan cada arco o bolsa branquial. Dentro de las branquias los vasos aferentes se ramifican en capi- lares y se unen de nuevo (después de bajar la presión sanguínea) para formar vasos

86 XNATOMfA BÁSICA DE LOS PECES
TIPOS DE CORAZdN
)no arterioso 0 -Bulbo arterioso
Particiones parciales
LAMPREA TlBURdN PEZ QSEO PEZ PULMONADO
Figura 3.16 - Esquemas comparativos generalizados del corazón en los grupos supe. riores de peces vivientes.
eferentes que forman finalmente la aorta dorsal, vaso principal para la distribu- ción de la sangre en el cuerpo.
Existe una sorprendente diferencia en la estructura del corazón previamente citada, en los peces pulmonados. Ambos, el atrio y el ventriculo, están parcial- mente divididos en dos cámaras. El efecto de esta división se extiende hacia la aorta ventral reducida, el tronco arterioso, de tal manera que la sangre del lado derecho del corazón se concentra en su flujo hacia los dos últimos arcos hranquia- les. En el pez pulmonado de Australia (Neoceratodus) el pulmón es irrigado por un vaso que proviene del cuarto arco aórtico de cada lado (arterias pulmonares), pero, en cada uno de los otros peces pulmonados, de las dos especies que aun viven, la arteria pulmonar nace de la aorta dorsal. La sangre regresa desde el pul- món mediante una vena pulmonar, al lado izquierdo del seno venoso. Debido a la partición del atrio esa sangre tiende a concentrarse en el lado izquierdo y a ser en- viada hacia el frente, principalmente a los arcos aórticos primero y segundo. Por lo tanto, una separación parcial de las circulaciones pulmonar (menor) y del cuerpo (mayor) fue efectuada en los peces pulmonados (Dipnoi) primitivos, posible-. mente como un anuncio de la condición más avanzada que más tarde habría de aparecer en los anfibios y que condujo a la circulación doble completa de los ver- tehrados superiores: los reptiles, las aves y los mamíferos.
Vasos sanguíneos (Figura 3.17)
L a sangre oxigenada abandona las branquias para irrigar la región cefálica, princi- palmente a través de las arterias earótidas y para llegar al cuerpo mediante ramifica- ciones de ia aorta dorsal, incluyendo a muchos vasos que están distribuidos en los segmentos. La aorta dorsal de la región caudal es denominada arteria caudal y atraviesa los arcos hemales de las vi-rtebras caudales. La sangre de la cola es reci- bida principalmente en la vena caudal tan luego como corre hacia adelante a lravits de los arcos hemales. justamente debajo de la arteria caudal. En los grupos

RIÑONES 87
C I R C U L A C I ~ N
Vena Arteria Arteria Vena caor;,nnl --, subcl T i a Arteria m;;;;:f;t” ,,/- postcardinal cel iaca Arteria
f caudal
/ /- segmental Arteria
Sistema venoso Vena intestinal
Vena subclavia
Figura 3.17 - Plan general de la circulación en un pez.
de peces superiores que siguen a los ciclóstomos, la vena caudal drena en el sis- tema portal renal y después de salir de los riñones la sangre se reúne con la que proviene de la musculatura dorsal en las venas superiores fundamentales o cardi- nales. Las postcardinales siguen su curso hacia adelante y reciben sangre de espa- cios sanguíneos anteriores, como las cardinales anteriores. Las venas cardinales se unen a los conductos de Cuvier (venas cardinales comunes) en cada lado del esófago, casi a nivel de la cintura pectoral. Los conductos de Cuvier reciben san- gre adicional de la porción inferior de la cabeza (vía las venas yugulares) y de las paredes laterales del cuerpo (vía las venas abdominales laterales). Además de la que recibe de los conductos de Cuvier, el seno venoso también colecta sangre del hígado (vía las venas hepáticas). De las vísceras la sangre llega al hígado a través del sistema portal hepático.
Vasos linfáticos
El sistema linfático es mucho menos conocido en los peces que los sistemas arterial y venoso. Los vasos linfáticos son senos de paredes delgadas y no son fácilmente distinguibles en una disección superficial (véase el Capítulo 7 y la Figura 7.7).
RIÑONES (CAPfTULO 9; FIGURAS 3.15,3.18, 3.19 Y DE LA 9.2 A LA 9.5)
Aunque difieren mucho en función, los sistemas excretor renal y reproductor están estrechamente relacionados en los peces (como en otros vertebrados).

ANATOMÍA BASICA DE LOS PECES
TIPOS DE RIN;JdN
1 ,,,-Aleta dorsal
- ~ - Espina neural
Septo esqueletógeno medi0 dorsal
Arco neural ~ - -
Centro de vértebra
Septo esquelet6geno
horizontal Túbulo -.
pronéfrico
Conducto pronéfrico
.~~ ~
Peritoneo
Nefrostoma cuerpo
PRONEFROS MESONEFROS
Figura 3.18 - Digrama de las relaciones estructurales de los tipos de riñón en los peces y localización de las gónadas, tal como se muestra en secciones transversales en dos niveles del tronco. (Basado en Kingsley, 1926).
Los riñones de los peces están dispuestos en número par y en forma de estructuras longitudinales que se adosan arriba de la cavidad del cuerpo, hacia el lado ventral de la columna vertebral y la aorta dorsal, precisamente hacia el exte- rior del peritoneo. Son comúnmente de color café rojizo, pulposos y sanguino-. lentos cuando se abren (corno cuando se extraen al limpiar un pescado para la mesa). Cada riñón drena al exterior a través de un conducto, que puede unirse caudalmente en un solo conducto o en un seno alargado (el seno orogenital en los tiburones y grupos afines o Elasmobranchii, la vejiga urinaria en muchos peces óseos, Actinopterygii). Sin embargo, la vejiga urinaria de los actinopterigios di- fiere de la de los vertebrados superiores en que tiene diferente origen embrionario- mesodérmico en lugar de endodérmico.
Dos tipos anatómicos básicos de riiíones pueden ser distinguidos en los peces: el pronéfrico y el mesonéfrico (Figura 3.18). En el tipo pronéfrico los em- budos anteriores llevan directamente de la cavidad del cuerpo al conducto pro- néfrico por medio de los túbulos pronéfricos. En el tipo mesonéfrico faltan los embudos que se abren en la cavidad del cuerpo, pero en su lugar hay ramificacio- nes del conducto mesonéfrico, los túbulos mesonéfricos, que tienen cada uno su extremo alargado y ciego. Este extremo ciego, llamado cápsula de Bowman (Fi- gura 9.3) recuerda en su contorno a una pelota de hule presionada por un dedo. En la correspondiente concavidad de una cápsula de Bowman se localiza una pelota de vasos capilares (un glomérulo que proviene de una rama de la arteria

GLANDULAS REPRODUCTORAS - GONODAS 89
renal). Los materiales que van a ser excretados pasan de un glomérulo a la cáp- sula y luego al conducto urinario, y de aquí hacia el exterior.
El pronefros es de posición anterior al mesonefros. En la mayoría de los peces es transicional porque las funciones que ocurren en é1 durante sus prime- ras etapas de formación son más tarde transferidas al mesonefros. Sin embargo, un pronefros puede permanecer como un riñón funcional (como en el mixino Myxine) y va cambiando gradualmente hacia atrás en un mesonefros igualmente funcional. Se ha dado en llamarle opistonefros a la parte posterior de los riñones cuando los mismos túbulos mesonéfricos quedan envueltos por capilares y ex- tiende así la parte funcional del sistema más allá del simple tipo mesonéfrico, en donde la acción es concentrada en los glomérulos y sus cápsulas.
En algunos grupos marinos de peces óseos, de los más evolucionados, hay una reducción extrema del número de glomérulos en el riñón. El extremo de esta situación puede ser observado en los peces blenioideos, los peces sapo (Batrachoi- didae), lófidos (Lophiidae) y los peces antenados (Antennariidae), en los que los glomérulos de sus riñones han desaparecido por completo y les han quedado en su lugar riñones puramente tubulares (aglomerulares).
GLANDULAS REPRODUCTORAS-G~NADAS (CAPITULO 10; FIGURAS 3.15 y 3.18)
Las gónadas femeninas son los ovarios y las gónadas masculinas, los testículos. Normalmente aparecen en individuos adultos separados. En muchas especies han sido descritas, como anormalidades, las combinaciones de gónadas masculinas y femeninas (ovotestis), dando evidencia entonces de su origen embrionario común. El hermafroditismo normal ocurre en algunos grupos de peces (como en los me- ros, Serranidae). La autofertilización es, sin embargo, una verdadera excepción.
Testículos
Los testículos en los peces son internos y longitudinales. Se originan como estruc- turas pares y permanecen así en la mayoría de las especies. Están suspendidos por mesenterios (mesorquia) alargados en la sección superior de la cavidad del cuerpo y se les puede localizar hacia los lados, a todo lo largo, o por debajo de la vejiga gaseosa, cuando este órgano está presente. Están compuestos de folículos donde se desarrollan los espermatozoides. El tamaño y color varían de acuerdo al estado de maduración de estos órganos y al desarrollo sexual del pez. El peso puede ser igual al 12% o más del peso del cuerpo. A menudo el color de los testículos es blanco cremoso y de superficie lisa. El aspecto superficial de su contenido es grumoso más que granular. En los testículos maduros los túbulos espermáticos son suficientemente largos en algunas especies, de tal modo que producen una apariencia hasta cierto punto granular; pero un pequeño desgarramiento o aplasta- miento que se haga de algunos fragmentos observados bajo pequeña amplificación del microscopio, hará que puedan distinguirse sus túbulos, del tejido ovárico u ovarígeno de los ovarios.
En los tiburones y grupos afines (Chondrichthyes), los espermatozoides de un testículo hacen primeramente su recorrido hacia el exterior, en la parte ante-

90 ANATOM~A BASICA DE LOS PECES
rior del órgano, a través de pequeños túbulos enrollados (vasos eferentes, túbulos mesonéfricos modificados); en secuencia caudal, siguen por el conducto espermá- tic0 (conducto mesonéfrico modificado), un órgano para el almacenamiento temporal (vesícula seminal), después por el seno urogenital, la papila urogenital y el poro urogenital. A nivel de la parte anterior del testículo, el conducto esper- mático (conducto de Wolff, un conducto mesonéfrico modificado) está enrollado (epidídimo); hacia su parte media es propiamente un vaso deferente, y hacia el extremo caudal se transforma en una vesícula seminal de paredes gruesas. El seno urogenital lleva a los lados sendas bolsitas llamadas sacos espermáticos.
Los peces óseos (Osteichthyes) retienen la estructura básica de los tiburo- nes; sin embargo, las vesículas seminales, y/o los sacos espermáticos, están presentes en sólo unas cuantas clases (como en los 'peces sapo, Opsanus). La relación del vaso deferente con el conducto mesonéfrico difiere entre los grupos de peces superiores; las relaciones evolutivas de los conductos permanecen ignoradas. Los conductos espermáticos (uusu deferentia) faltan en los salmones y las truchas (Salmonidae); los espermatozoides son desparramados en la cavidad del cuerpo y expulsados después por la vía de una abertura espermática situada detrás del ano. En los tiburones y especies afines (Chondrichthyes), los peces pulmonados (Dip- noi), los esturiories (Acipenseriformes) y los pejelagartos (Lepisosteus) se hace un uso variado de los túbulos renales y del conducto mesonéfrico o de Wolff. En Amia hay una condición intermedia en la cual los túbulos testiculares (vasos efe- rentes) evaden el riñón y comunican directamente con el conducto de Wolff. La disposición más avanzada es la del políptero (Polypterus) y la de los peces óseos superiores (Teleostei) en los que, como sucede en el pez sapo, no hay paso a tra- vés del riñón, pero el conducto espermático es nuevo y se origina en los testículos, más que mediante la usurpación del conducto mesonéfrico. El conducto esper- mático funcional de los peces óseos superiores no es entonces homólogo con el de ciertos peces óseos inferiores, los tiburones y sus especies afines.
Ovarios
Como sucede con los testículos, los ovarios también son internos, usualmente longitudinales y se originan como estructuras pares, pero a menudo se fusionan y acortan. Están suspendidos (mesovaria) de la parte superior de la cavidad del cuerpo por un par de mesenterios y, en consecuencia, cuando existe una vejiga natatoria o gaseosa se les puede localizar directamente debajo de este órgano. El tamaño y distribución de los ovarios en la cavidad del cuerpo varía con el estado de maduración sexual de la hembra. Cuando están maduros los ovarios forman algo así como el 70% del peso del cuerpo. Muy a menudo el color de los ova- rios varía del blanquecino en los jóvenes, al verdoso cuando son inmaduros y al amarillo oro (color de yema) en los adultos maduros. Las texturas de estos órganos varían desde la grumosa (casi igual a la de los testículos en el joven), pasando por la granular microscópica en los juveniles, hasta la de granos gruesos, variando con el tamaño de cada huevo en lo individual, en los adultos. Por lo común, justamente antes de ser puestos, los huevos aumentan de tamaño y se vuelven claros.
Los huevos pasan, en los ciclóstomos, del ovario a la cavidad del cuerpo y después a! exterior a través de los poros abdominales que llevan al seno urogenital

6RGANOS ENDOCRINOS 91
en el vientre del pez. En los condrictios, un oviducto (conducto de Müller, que no es una modificación del conducto mesonéfrico) con una entrada en forma de embudo (ostium tubae abdominale) en su extremo cefálico, está situado anterior- mente en la cavidad del cuerpo y conduce los huevos hacia atrás, a la abertura cloacal, vía las aberturas genitales (la cápsula ovárica no es continua con el ovi- ducto; es una condición gimnoaria). Los tiburones y las especies afines son, o bien ovopositores (ovíparos) o bien vivíparos (pueden ser ovovivíparos o simplemente vivíparos). En los ovopositores el tejido del oviducto está modificado hacia ade- lante en una glándula de la cáscara. Entre los vivíparos el oviducto aumenta de tamaño hacia atrás para formar un útero para la retención de la cría durante su desarrollo embrionario. Otros grupos de peces, donde quedan incluidos los pulmo- nados (Dipnoi), esturiones (Acipenseriformes) y un pez óseo primitivo, el pez sapo (Arniu), mantienen la condición condrictina y la gimnoaria, así como el sistema resultante que induce el recorrido del huevo y modifica el mecanismo de expul- sión del producto. La mayor parte de los peces óseos superiores tienen la cápsula ovárica continua con el oviducto (es la condición cistoaria). Los huevos pasan en- tonces directamente desde el interior del ovario al canal ovárico y no a través de la cavidad del cuerpo como sucede en los condrictios. Los oviductos faltan, sin embargo, en las truchas, los salmones y otros peces parecidos donde (como sucede en las lampreas, Petromyzonidae) los huevos irrumpen en la cavidad del cuerpo y encuentran su salida a través de poros adyacentes a las aberturas urinaria y rec- tal. Los poros se abren cuando ocurre la reproducción y pueden representar ovi- ductos cuyo tamaño está reducido en extremo.
6RGANOS ENDOCRINOS (CAP~TULO 11; FIGURAS 3.15,3.18 Y 3.19)
Desde el punto de vista de su estructura (morfología) y su función (fisiología), las glándulas sin conductos (endocrinas), como la pituitaria y la tiroides, son relativa- mente menos conocidas que las que tienen conductos (exocrinas) y secreciones evidentes, como sucede con el hígado. Sin embargo, la mayoría de los órganos
Figura 3.19 - Localización esquemática GLÁNDULAS ENDOCRINAS de los órganos endocrinos Tejido en los peces. renal Tejido
Tejido interrenal Corpúsculos de Stannius
Ultimobranquial
Teiido intestinal

92 AXATOMfA BASICA DE LOS PECES
endocrinos de los vertebrados superiores y aun otros han sido encontrados en los peces. La siguiente lista los nombra (y el diagrama en la Figura 3.19 sugiere su localización): la pituitaria (hipófisis), la urohipófisis, la tiroides, el timo, los islotes pancreáticos (islotes de Langerhans), el tejido cromafínico, el tejido in- terrenal, el tejido intersticial en las gónadas, el cuerpo últimobranquial (¿glándula paratiroides?), el tejido intestinal (productor de secretina), el tejido renal (que produce la renina), e¡ cuerpo pineal y los corpúsculos de Stannius. Entre los grupos de peces hay considerable variación en la ubicación y forma de muchas de las glándulas secretoras y tejidos citados anteriormente. También hay variedad en la organización de las partes de la glándula entre las especies, como puede observarse en la pituitaria (Figura 11.8). La tiroides de los tiburones y grupos afi- nes y de algunos peces óseos es una glándula discreta encapsulada. En la mayoría de los peces óseos, sin embargo, los folículos tiroideos están más o menos difun- didos en la región de la garganta. Detalles adicionales sobre la morfología y fisio- logía de las glándulas endocrinas son dadas en el Capítulo 11.
SISTEMA NERVIOSO (CAPfTULO 11; FIGURAS 3.20 A 3.25)
El sistema nervioso de los vertebrados, bien representados hasta en los grupos más primitivos de peces vivientes, está adecuadamente clasificado sobre bases anató- micas, como se indica a continuación:
Sistema cerebroespinal
División central: cerebro y médula espinal
División periférica: nervios craneales y espinales, órganos especiales de los sentidos
Sistema autónomo Ganglios y fibras, partes simpáticas y parasimpáticas
La anatomía superficial de los componentes nerviosos mencionados, a excepción de los receptores microscópicos para el tacto, gusto y temperatura, son descritos aquí. Los detalles sobre la anatomía y la fisiología de este sistema son dados en el Capítulo 11.
Cerebro y médula espinal (Figura 3.20)
El cerebro del pez es una prolongación del extremo anterior de la médula espinal. Sus partes van progresando linearmente a partir de una región cerebral delantera (los hemisferios cerebrales de relativamente gran tamaño y la conexión del cerebro intermedio), seguida por el cerebro medio con sus turgencias (los lóbulos ópticos), para terminar en el cerebro posterior (el cerebelo y la médula) y hacia la aleta cau- dal en la médula espinal.
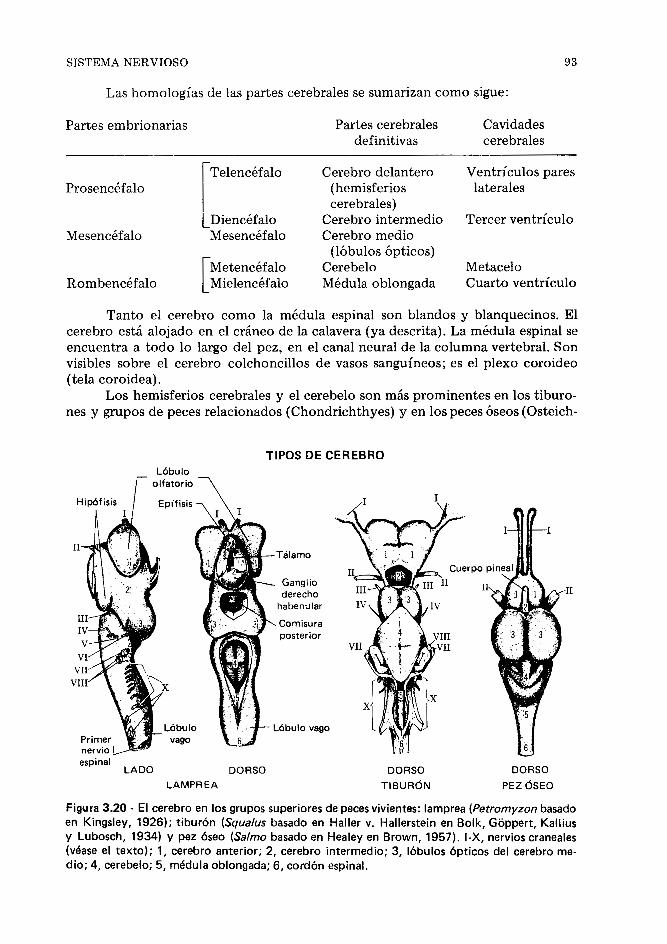
SISTEMA NERVIOSO 93
Las homologías de las partes cerebrales se sumarizan como sigue:
Partes embrionarias
Prosencéfalo r Telencéfalo
LDiencéfalo Mesencéfalo Mesencéfalo
Metencéfalo Rombencéfalo Mielencéfalo
Partes cerebrales Cavidades definitivas cerebrales
Cerebro delantero Ventrículos pares (hemisferios laterales cerebrales)
Cerebro intermedio Tercer ventriculo Cerebro medio
Cerebelo Metacelo Médula oblongada Cuarto ventriculo
(lóbulos ópticos)
Tanto el cerebro como la médula espinal son blandos y blanquecinos. El cerebro está alojado en el cráneo de la calavera (ya descrita). La médula espinal se encuentra a todo lo largo del pez, en el canal neural de la columna vertebral. Son visibles sobre el cerebro colchoncillos de vasos sanguíneos; es el plexo coroideo (tela coroidea).
Los hemisferios cerebrales y el cerebelo son más prominentes en los tiburo- nes y grupos de peces relacionados (Chondrichthyes) y en los peces óseos (Osteich-
TIPOS DE CEREBRO Ldbulo
espinal LADO DORSO DORSO DORSO
LAMPREA T I B U R ~ N PEZ dSE0
Figura 3.20 - El cerebro en los grupos superiores de peces vivientes: lamprea (Petromyzon basado en Kingsley, 1926); tiburón (Sgualus basado en Haller v. Hallerstein en Bolk, Goppert, Kallius y Lubosch, 1934) y pez óseo (Salmo basado en Healey en Brown, 1957). I-X, nervios craneales (véase el texto); 1, cerebro anterior; 2, cerebro intermedio; 3, lóbulos ópticos del cerebro me- dio; 4, cerebelo; 5, medula oblongada; 6, cordón espinal.

94 ANATOMfA BASICA DE LOS PECES
thyes), que en las lampreas y mixinos (ciclóstomos). El cerebro medio, que es prominente en los ciclóstomos, es también prominente en los condrictios. Sin embargo, el de los peces óseos superiores (Actinopterygii) es a menudo muy gran- de. Las cavidades del cerebro se continúan con el canal del epéndimo de la médula espinal.
Nervios craneanos (Figura 3.20; Capítulo 11)
Hay diez pares de nervios craneanos asociados con el cerebro del pez (doce en los vertebrados superiores). Son los siguientes: I, olfatorio; 11, óptico; 111, oculo- motor; IV, troclear; V, trigémino; VI, abductor; VII, facial; VIII, acústico; IX, glosofaríngeo; X, vago. En el Capítulo 11 se describen las funciones de estos nervios.
Nervios espinales
Los nervios pares espinales nacen segmentalmente de la médula, detrás del cerebro, y llevan fibras sensoriales y motoras de y hacia la masa del cuerpo del pez. Exis- ten plexos de mayor tamaño a nivel de las aletas pares. A lo largo de la médula espinal, y dispuestos en relación a segmentos, hay centros de relevo (ganglios) de los nervios espinales.
drganos olfatorios (Figura 3.21)
El órgano del olfato en los peces es una bolsa que se abre al exterior y hace con- tacto con el agua a través de uno o dos orificios. La bolsa es un saco ciego en todos los peces, a excepción de unos cuantos de los óseos vivientes (por ejemplo, en Astroscopus) y los pulmonados (Dipnoi: Neocerutodus, Lepidosiren y Protop- terus), en los cuales los orificios nasales externos llevan el agua a través de la bolsa a la cavidad oral. El saco está cubierto de manera característica por un epitelio laminar sensorial conectado por el nervio olfatorio al cerebro anterior.
ORIFICIOS NASALES
Figura 3.21 - Diagramas de los orificios nasales externos y el órgano olfatorio en los peces óseos: (a) pez blanco redondo (Prosopium cylindraceum); (b) "cisco" de agua poco profunda (Coregonus artedii); (c) recorrido del agua y su relación con el recubrimiento sensorial (epitelio olfatorio) inervado por el nervio olfatorio. (Fuen- te para a y b, Hubbs y Lagler, 1958).
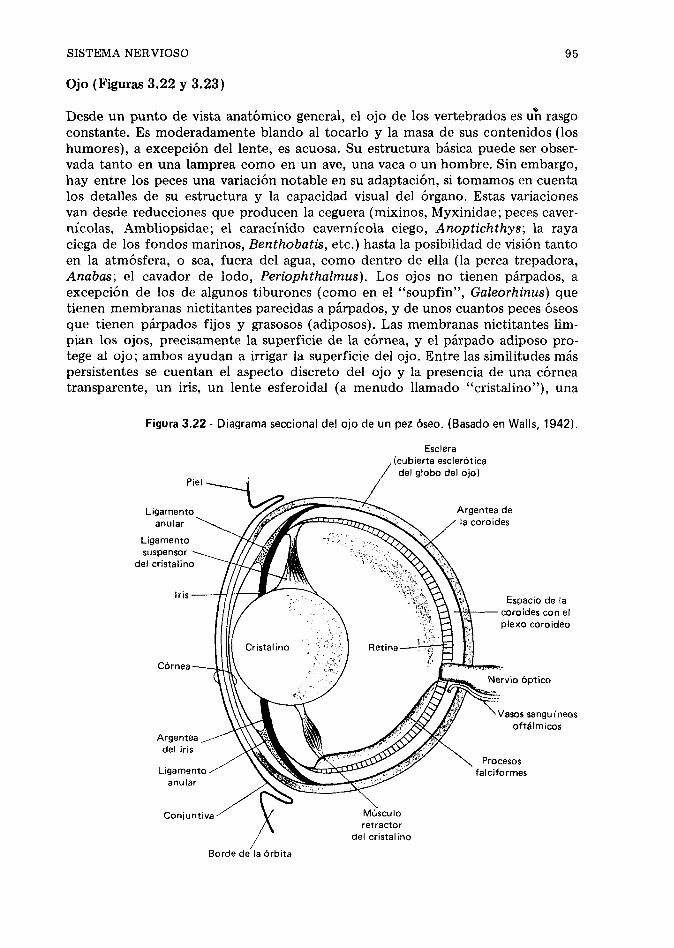
SISTEMA NERVIOSO 95
Ojo (Figuras 3.22 y 3.23)
Desde un punto de vista anatómico general, el ojo de los vertebrados es u’; rasgo constante. Es moderadamente blando al tocarlo y la masa de sus contenidos (los humores), a excepción del lente, es acuosa. Su estructura básica puede ser obser- vada tanto en una lamprea como en un ave, una vaca o un hombre. Sin embargo, hay entre los peces una variación notable en su adaptación, si tomamos en cuenta los detalles de su estructura y la capacidad visual del órgano. Estas variaciones van desde reducciones que producen la ceguera (mixinos, Myxinidae; peces caver- nícolas, Ambliopsidae; el caracínido cavernícola ciego, Anoptichthys; la raya ciega de los fondos marinos, Benthobatis, etc.) hasta la posibilidad de visión tanto en la atmósfera, o sea, fuera del agua, como dentro de ella (la perca trepadora, Anabas; el cavador de lodo, Periophthalnus). Los ojos no tienen párpados, a excepción de los de algunos tiburones (como en el “soupfin”, Galeorhinus) que tienen membranas nictitantes parecidas a párpados, y de unos cuantos peces óseos que tienen párpados fijos y grasosos (adiposos). Las membranas nictitantes lim- pian los ojos, precisamente la superficie de la córnea, y el párpado adiposo pro- tege al ojo; ambos ayudan a irrigar la superficie del ojo. Entre las similitudes más persistentes se cuentan el aspecto discreto del ojo y la presencia de una córnea transparente, un iris, un lente esferoidal (a menudo llamado “cristalino”), una
Figura 3.22 - Diagrama seccional del ojo de un pez óseo. (Basado en Walls, 1942).
Esclera (cubierta esclerótica del globo del ojo)
Borde de la órbita /

retina, una cápsula escierbtica con humor vitreo en su cavidad y tres pares de músculos oculomotores (Figura 3.23).
Más allá de estas semejanzas superficiales que se han señalado para los ojos de los peces, existen, sin embargo, diferencias. En los ciclóstomos, la piel y la córnea no se han fusionado y un epitelio corneal se ha desarrollado solamente en los grupos superiores de peces. Una capa de tejido interno muy peculiar (capa autóctona) está variablemente presente en la córnea de los teleósteos. En el ojo de los ciclóstomos no hay musculos que cierren el iris (esfínter) o que lo abran (dilatador); algunos pece; parecidos a los tiburones (Chondrichthyes) y peces óseos primitivos, tampoco tienen esos músculos en los ojos; cuando están presen- tes en los de los peces óseos superiores, están mal definidos. En los ciclbstomos, el globo ocular está compuesto de tejido conectivo, pero está reforzado con cartí- lago en todos los grupos superiores, siendo de notar que en los teleósteos el globo está protegido por placas óseas delgadas. Es peculiar en el ojo de los teleósteos, con algunas variaciones, la presencia de ligamento anular, ligamento falciforme y glándula coroidal. El ligamento anular es una conexih circular situada entre Ia córnea y el borde exterior del iris. El ligamento (proceso) falciforme es un cor- doncillo situado en la parte inferior del globo del ojo, que está conectado de diversas formas con el músculo retractor (retractor lentis) del lente; en los peces de aletas lobuladas y en los pulmonados, que no tienen estructuras para el aco- modamiento del ojo, faltan el proceso y su ligamento. La glándula coroidal es una parte característica de la coroides (situada entre la retina y la esclerótica). Además de todo lo anterior, hay en la retina muchas diferencias (y capacidades) en su estructura. Y, todavía más, los dos músculos oculares oblicuos no se en- cuentran en el pez pulmonado de América del Sur (Lepidosiren); solamente se han desarrollado en é1 los dos pares de músculos rectos.
Oblicuo - / O bl icuo superior
Borde anterior
de la brbita
Globo del ojo
Borde posterior de la
6rbita
Figura 3.23 - Diagrama de los tres pares de músculos oculomo- tores en un pez, vistos desde arriba. (Fuente: Walls, 1942).
Recto interno
I Recto
externo superior

SISTEMA NERVIOSO 97
drganos de la audición y del equilibrio (Figuras 3.24 y 3.25)
Como sucede en todos los vertebrados, los receptores de la audición y del equili- brio están concentrados en el oído. En los peces, sin embargo, solamente un oído interno *está presente; faltan las otras dos regiones bien conocidas del oído hu- mano, por ejemplo, el externo (que termina internamente en el tímpano) y el medio (que contiene la cadena de huecesillos: martillo, yunque y estribo). La có- clea del oído interno de los demás vertebrados falta también en los peces. Debido a que no hay membrana del tímpano, las vibraciones que debe recibir el oído del pez son conducidas a este órgano a través de los tejidos del cuerpo. A excepción de las piedrecillas del oído (otolitos), que este órgano contiene, el oído interno es un laberinto membranoso muy delicado, lleno de un fluido (la endolinfa) y sumergido o bañado por la perilinfa. Está encapsulado en los ángulos posteriores del cráneo (posterolateralmente). Están incluidos en el órgano los receptores para las vibraciones y los estímulos que afectan el mantenimiento del equilibrio.
Desde el punto de vista evolutivo el oído interno es una parte especializada del sistema sensorial de la línea lateral, y ambos componen el “sistema acústico- lateral”. El sistema de la línea lateral está contenido, en muchos peces, en el canal de la línea lateral, prominente y visible a lo largo de los lados del cuerpo y la ca- beza. En las lampreas (Petromyzonidae), en algunos pececillos de acuario (Cypri- nidae), en las sardinas (Clupeidae) y en otros peces, sin embargo, los órganos de los extremos de la línea lateral están desparramados sobre el cuerpo. Se piensa que su función es la de detectar los movimientos del agua y, en consecuencia, participar en la orientación del pez, siendo responsables del trabajo de manteni- miento del equilibrio del laberinto membranoso y los otolitos del oído interno. En los tiburones y grupos afines (Chondrichthyes), la presencia de un conducto y poro endolinfáticos que comunican el medio externo del pez y el interior del
Figura 3.24 - Oído interno de los peces. Dos tipos principales de laberintos mem- branosos como aparecen viendo a los órganos del lado derecho desde la I ínea media. (Fuente: von Frish en Brown, 1957).
TIPOS DE OíDO Crus
común Crus
común
vertical ante
TIPO COMúN (SALMÓNIDO) TIPO ClPRlNOlDEO

98 ANATOMfA BASICA DE LOS PECES
laberinto membranoso del oído interno, ha sido señalada como una evidencia de la intima y obvia relación entre el órgano del oído y los órganos de la linea lateral.
El laberinto membranoso de los peces puede ser comparado con una bolsa (Figura 3.24). Unos conductos curvos (los canales semicirculares) que se originan en la cavidad principal y una bolsa lateral (sacculus) que tiene a su vez o’tra aber- tura (lagena), son considerados como partes accesorias. La cavidad principal es llamada “utriculus”. Los otolitos están presentes en las tres bolsas principales; en los peces óseos estas estructuras son calcáreas y a menudo muestran zonas de crecimiento (anillos anuales); los del utrículo son los “lapillus”, los del sáculo, los “sagitta”, y los de la lagena, los “asteriscus”. En los tiburones y especies afi- nes, los otolitos son pequeños, numerosos y difusos. Su número relativamente corto e individualidad específica hacen que los otolitos de los teleósteos sean de suma utilidad en taxonomía. Cada canal semicircular es tubular; ambos extre- mos comunican con la cavidad (vestíbulo) del sistema de bolsas que ya ha sido descrito. Cerca de uno de sus extremos, precisamente antes de abrirse en el ves- tíbulo, tiene una ampliación o ámpula. Cada ámpula contiene terminaciones nerviosas sensoriales (células neuromástiles) en una cápsula gelatinosa terminal, como sucede con los otolitos y otras regiones de la vesícula compuesta que for- man el oído interno. Los peces superiores tienen tres canales semicirculares en cada lado de la cabeza; el horizontal, el anterior vertical y el posterior vertical. El sistema, en ambos lados de la cabeza, puede ser conectado por un canal trans- verso que une los dos sáculos. Cada lado de las lampreas (Petromyzonidae) tiene solamente dos canales semicirculares y en el grupo de los mixinos (Myxinidae) sola- mente uno.
En algunos peces, especialmente en los de acuario, las rémoras y los peces gato (Cypriniformes = Ostariophysi), hay una conexión elaborada de ligamentos alternados y pequeños huesos (osículos) entre la vejiga gaseosa y la cápsula Ótica (Figura 3.25). Esta estructura (aparato de Weber) no solamente es característica en todo el orden de los cipriniformes; también las diferencias individuales de los osículos son de utilidad para la identificación de las especies. Los osículos son derivaciones de las vértebras anteriores.
Sistema nervioso autónomo
Tal como sucede generalmente en los vertebrados, el sistema nervioso autónomo de los peces es menos conocido, en cuanto a su anatomía y función, que el sistema nervioso central. No hay concentraciones bien definidas (ganglia) de célu- las nerviosas del sistema autónomo en los ciclóstomos. Más bien, las células están ampliamente distribuidas y oscuramente conectadas a los demás componentes nerviosos. Los tiburones y especies afines muestran una mejor serie regular de ganglios simpáticos en el cuerpo, con fibras pregangliónicas procedentes de la médula espina1 que van hacia ellos, y fibras postganglionares que salen de ellos para ir a inervar los músculos del tubo digestivo y de las arterias. El vago (nervio craneal X) también manda fibras nerviosas involuntarias al tubo digestivo y al corazón. Los peces bseos superiores (telebsteos) muestran mejor una cadena de ganglios simpáticos en la región del tronco y hacia l a cabeza, hasta donde se en- cuentra el nervio craneal \’, el trigémino. A diferencia de los tiburones y grupos relacionados, los peces óseos tienen un suplemento de fibras postganglionares

REFERENCIAS 99
horizontal
Canal semicircular vertical posterior
Canal transverso entre el sáculo
Figura 3.25 - Disecci6n diagramática de la cabeza de un "minnow" (Cyprinidae) mostrando la posici6n de los canales semicirculares del oído interno, y sus relacio- nes con el aparato de Weber y la vejiga gaseosa en los cipriniformes. (Basado en von Frish en Young, 1950).
que van a la piel (y al tubo digestivo y a las arterias también). A las fibras sim- páticas del iris se oponen en su acción las fibras parasimpáticas.
La mayor parte del sistema autónomo está intimamente conectado, ya sea al cerebro o a la médula espinal. El hecho de que sea tomado en cuenta como una división del sistema nervioso es por mera conveniencia, no por un carácter fun- cional verdadero.
REFERENCIAS ESPECIALES EN ANATOMfA
Bolk, L., E. Goppert, E. Kallius, y W. Lubosch. 1931-1939. Handbuch der Vergleichenden
Bronn, H.G. 1940. Klassen und Ordnungen des Tierreichs. Band 6. Abteilung 1. Pisces. 2
Daniel, J.F. 1934. The elasmobranch fishes. University of California Press, Berkeley, California. Greene, C.W., y C.H. Greene. 1914. The skeletal musculature of the king salmon. Bull. U.S.
Gregory, W.K. 1933. Fish skulls. Trans. Amer. Philos. Soc., 23(2):76-481. (Reimpresión en
Kingsley, J.S. 1926. Outlines of comparative anatomy of vertebrates. The Blakiston Company
Mujib, K.A. 1967. The cranial osteology of the Gadidae. Jour. Fish. Res. Bd. Canada, 24(6):
Anatomie der Wirbeltiere. Urban und Schwarzenberg, Berlín.
Buchs. Akademie Verlagsgessellschaft MBH, Leipzing.
Bur. Fish., 33:21-60.
offset, Eric Lunderberg Publ., Laurel, Fla., 1959).
(McGraw-Hill Book Company, Nueva York).
1315-1376.

100 ANATOMfA BASICA DE LOS PECES
Neal, H.V., y H.W. Rand. 1939. Comparative anatomy. The Blakiston Company (McGraw-Hill
Nursall, J.R. 1956. The lateral musculature and the swimming of fish. Proc. 2002. Soc. London,
Stockard, A.H. 1949. A laboratory manual of comparative anatomy of the chordates. Edwards Brothers, Ann Arbor, Mich.
Tominaga, S. 1965-67. Anatomical sketches of 500 fishes. 3 Volcmenes (5 libros). Kadokawa- Shoten, Tokio.
Walls, G.L. 1942. The vertebrates eye and its adaptive radiation. Bull. Cranbrook Inst. Sci., 19:785 p.
Winterbottom, R. 1974. The familial phylogeny of the Tetraodontiformes (Acanthopterygii: Pisces) as evidenced by their conlparative miology. Smithsonian Contributions, Zoology, 155, 201 p.
Wunder, W.W. 1936. Physiologie der Süsswasserfische Mitteleuropas. Band 11. B. Handbuch der Binnenfischerei Mitteleuropas. E. Schweizerbart'sche Verlagsbuchhandlung, Stuttgart.
Book Company, Nueva York).
126(1):127-143.

Capítulo 4
Piel I
Como en los demás vertebrados, la piel de un pez es la envoltura de su cuerpo y constituye la primera capa de defensa contra las enfermedades. También propor- ciona protección y adaptación respecto de los factores ambientales que influyen en su vida, porque contiene receptores sensoriales ajustados al medio en que vive. Además, la piel tiene funciones respiratorias, excretoras y osmorreguladoras. Tam- bién en la piel se encuentran los mecanismos que proporcionan al pez la colora- ción y opacidad o brillantez que le sirven para ocultarlo, advertir su presencia o facilitar su reconocimiento sexual. También está presente en la piel del pez gato eléctrico (Mulupterurus), un conjunto de órganos eléctricos que atontan a las presas cuando producen su descarga. La piel también contiene glándulas venenosas, como en los peces gato (Noturus), así como glándulas mucosas cuyas secreciones le dan la característica mucosidad y olor a los peces.
ESTRUCTURA
La piel de un pez consta de dos capas (Figura 4.1). La externa es la epidermis y la interna la dermis o corion. La epidermis del pez es en muchos aspectos simi- lar a la mucosidad de la boca humana. Generalmente está compuesta superficial- mente de muchas capas de células epiteliales, planas y húmedas. Las capas más profundas conforman una zona de crecimiento celular activo y de multiplicación (stratum gerrninatiuurn). La multiplicación celular es constante para reponer desde dentro a la capa más externa de células, a medida que se van perdiendo, y asegurar el crecimiento del pez. Estas células epiteliales de la epidermis son las primeras que ayudan a cicatrizar una herida superficial.
101

102
Borde dseo (circulus) en la escama
PIEL
Vaso
Figura 4.1 - Sección de la piel de pez. (Fuente: General Biological Supply House).
La capa dérmica de la piel contiene vasos sanguíneos, nervios y órganos sensoriales cutáneos, además de tejido conectivo. Cuando se despoja a un pez de su piel las fibras del tejido conectivo (esqueleto membranoso) que la unen al músculo y esqueleto que están inmediatamente por debajo, son muy evidentes. La dermis juega un papel muy importante en la formación de escamas y estruc- turas integumentarias relacionadas con ella.
Hay en la piel numerosas aberturas que corresponden a las células mucosas glandulares, en forma de recipiente de cuello delgado que se extienden hasta den- tro de la dermis, y se encuentran ampliamente distribuidas entre las células planas de la epidermis. Estas células secretan el moco resbaloso que cubre a la mayo- ría de los peces. En algunos, como en un mixino de 30 cm de longitud (Myxine, algunas veces llamado anguila de lodo) la función secretora de estas células es tan efectiva que se dice que un solo individuo es capaz de producir una cubeta llena de sustancia viscosa. Es presumible que el mucus permite a un pez desplazarse con mayor facilidad cuando nada en el agua. Como la mucosidad se va eliminando puede ser que tenga la misión de expulsar microorganismos y sustancias irritan- tes que pueden ser dañinas si son acumuladas. En algunas especies, la mucosidad coagula y precipita el lodo u otros sólidos suspendidos en el agua. El olor típico a pez está contenido en el mucus. Las células mucosas pueden ser también, pro- bablemente, un recurso de comunicación química entre los peces (por ejemplo, la sustancia-alarma o “Schreck-stoff”).
LAS ESCAMAS
Entre las características especiales más notables de la piel contamos los apéndices, y todavía más notables entre aquellas están, por supuesto, las escamas que la mayoría de los peces exhiben. Algunos grupos están, sin embargo, “desnudos”, en el sentido técnico de no tener escamas. Los ejemplos incluyen a las lampreas (Petromyzonidae) y los bagres de agua dulce de Norte América (Ictaluridae). Una

LAS ESCAMAS 103
categoría estructural intermedia comprende a los peces que están casi desnudos y muestran escamas solamente en algunas regiones del cuerpo, o tienen las esca- mas reducidas a unas cuantas espinas. Algunas de estas espinas tienen localización precisa. Ejemplos de peces que tienen pocas escamas son el pez espátula (Polyo- don), un pariente de los esturiones que habita en los arroyos del centro de Norte América y tiene también un pariente cercano, el pico-espada de agua dulce de China (Psephurus). Polyodon está esencialmente desnudo, a excepción de las re- giones de la garganta, la cintura pectoral y sobre la base volteada de su cola, donde se pueden encontrar algunas escamas. Algunas espinochas (Gasterosteidae), son des- nudas, incluyendo a Culaea y Pungitius; otras, notablemente la espinocha de tres espinas, Gasterosteus aculeatus, tienen placas óseas (raramente ausentes). Es inte- resante ver cómo la superficie mayor o menor de piel cubierta por placas óseas está relacionada con el grado de salinidad del agua donde vive este pez.
Los esculpinos (Cottidae) del Hemisferio Norte que viven en los arroyos, algunos en los lagos de gran amplitud como en los Grandes Lagos, y en los mares del Artico, están esencialmente desnudos, pero pueden estar adornados en forma variable. Es común que entre estos cótidos las axilas de las aletas pectorales y la cabeza tengan pequeñas espinas desparramadas, las que parecen ser vestigios de escamas. Otras partes derivadas de las escamas incluyen a los dientes, el estilete espinoso de las rayas de espina (Dasyatidae) y las placas óseas armadas como las de los caballitos de mar y peces pipa (Syngnathidae) y los bagres neotropicales armados de América del Sur (como en la familia Loricariidae). Una clase de la carpa común (Cyprinus carpio), la variedad espejo, tiene solamente unas cuantas escamas grandes que están a menudo aisladas por amplias Breas de piel. Hay tam- bién unos cuantos peces que tienen las escamas tan pequeñas y/o profundamente embebidas, que parecen estar desnudos, aunque no sea así; las anguilas de agua dulce (Anguilla) nos ofrecen un ejemplo legendario. Otros ejemplos incluyen a la trucha de arroyo (Subelinus fontinalis) y el “burbot” (Lota), un miembro de agua dulce de la familia de los bacalaos (Gadidae).
Los modelos de escama (Figura 4.2) están fundamentalmente asociados con la segmentación del cuerpo, como se ha manifestado inicialmente en el desarrollo embrionario de las vértebras y los miómeros. En relación a su implantación las escamas están más a menudo imbricadas y, en consecuencia, traslapadas como las tejas en un techo, dirigiendo su margen libre hacia la cola, de tal manera que disminuyen así la fricción con el agua. Ocurre raramente que algunos individuos aberrantes muestran total o parcialmente invertida esta disposición. En algunos peces, como el “burbot” (Lota) y las anguilas de agua dulce (Anguilla), el modelo es un mosaico; más que traslaparse una con otra, las escamas están separadas ape- nas, o se tocan unas con otras solamente en sus márgenes.
Formas de las escamas
Aunque solamente hay unos cuantos tipos estructurales básicos, las escamas exhi- ben tantas modificaciones que a menudo caracterizan a grupos Q especies. Cla- sifiquemos primero las escamas de los peces sobre la base de !as lormas dominan- tes, y luego hagámoslo sobre la base de la estructura. Sobre !a base de la forma un tipo será el placoideo, con una pequeña cúspide sobre cada placa, tan común entre los tiburones (Eiasmobranchii). Un segundo tipo es el de la forma de diamante

104 PIEL
Figura 4.2 - Diversidad de la disposición de las escamas entre los peces.
(rómbico) y caracteriza el integument0 de los peje-lagartos de Norte América (Le- pisosteidae) y de los polípteros (Polypterus) del Nilo. Esas escamas también apa- recen en la cola de los esturiones (Acipenseridae) del Hemisferio Norte y del pez espátula norteamericano (Polyodon). Un tercer tipo de escama de pez considerado en base a su forma es el cicloideo (Figura 4.3). Toma su nombre por su forma típica de disco suave en su curvatura, de borde más o menos circular (la superficie o margen caudal descubierto de esta escama no está dentado, como sucede en el siguiente tipo). En el cuarto tipo, al que pertenecen las escamas ctenoideas (Figu- ra 4.4), la superficie o margen posterior tiene dientes o forma de peine. Las esca- mas cicloideas se encuentran en l a mayoria de los peces de radios suaves (Malacop-

LAS ESCAMAS 106
1 Pez pulmonado (Cicloidea)
(Placoidea) 3 Tiburón
5 Bagre acorazado (Placa &ea)
7 Perca (Ctenoidea)
2 Catán (Rómbica, Ganoidea)
(Cicloidea)
6 Carpa (Cicloidea)
Figura 4.2 - Continuación
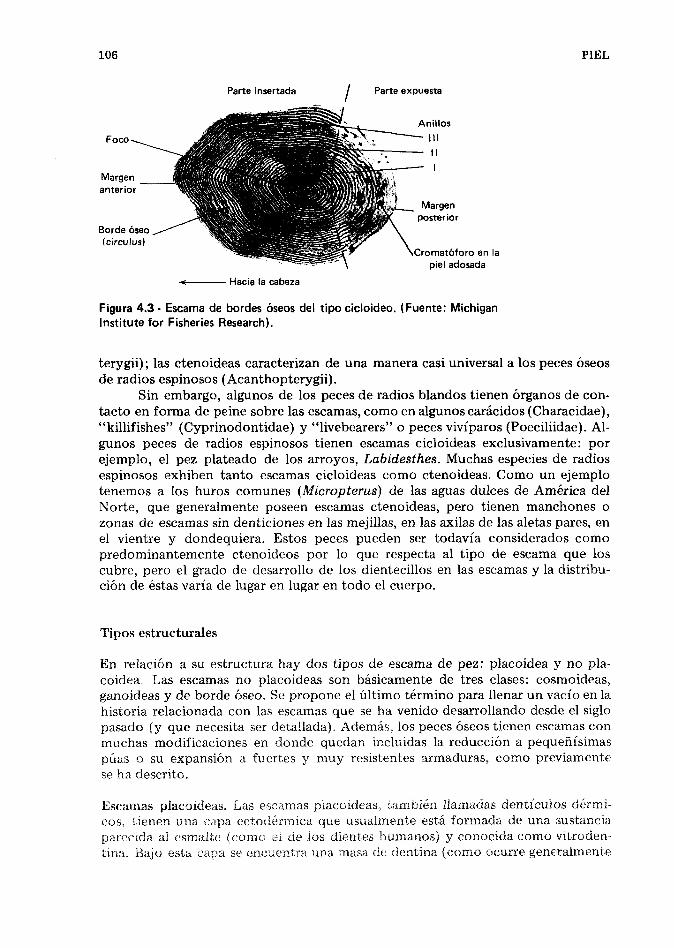
106
Parte insertada / Parte expuesta
PIEL
”
Cromat6foro en la
+“-- Hacia la cabeza
Figura 4.3 - Escama de bordes óseos del tipo cicloideo. (Fuente: Michigan Institute for Fisheries Research).
terygii); las ctenoideas caracterizan de una manera casi universal a los peces óseos de radios espinosos (Acanthopterygii).
Sin embargo, algunos de los peces de radios blandos tienen órganos de con- tacto en forma de peine sobre las escamas, como en algunos carácidos (Characidae), “killifishes” (Cyprinodontidae) y “livebearers” o peces vivíparos (Poeciliidae). Al- gunos peces de radios espinosos tienen escamas cicloideas exclusivamente: por ejemplo, el pez plateado de los arroyos, Labidesthes. Muchas especies de radios espinosos exhiben tanto escamas cicloideas como ctenoideas. Como un ejemplo tenemos a los huros comunes (Micropterus) de las aguas dulces de América del Norte, que generalmente poseen escamas ctenoideas, pero tienen manchones o zonas de escamas sin denticiones en las mejillas, en las axilas de las aletas pares, en el vientre y dondequiera. Estos peces pueden ser todavía considerados como predominantemente ctenoideos por lo que respecta al tipo de escama que los cubre, pero el grado de desarrollo de los dientecillos en las escamas y la distribu- ción de éstas varía de lugar en lugar en todo el cuerpo.
Tipos estructurales
En relación a su estructura hay dos tipos de escama de pez: placoidea y no pla- coidea. Las escamas no placoideas son básicamente de tres clases: cosmoideas, ganoideas y de borde &eo. Se propone el último término para llenar un vacío en la historia relacionada con las escamas que se ha venido desarrollando desde el siglo pasado (y que necesita ser detallada). Ademh, los peces óseos tienen escamas con muchas modificaciones en donde quedan incluidas la reducción a pequeñísimas púas o su expansión a fuertes y muy resistentes armaduras, como previamente se ha descrito.

LAS ESCAMAS
Parte insertada / Parte expuesta
107
Anillo I
Dientes
Cromat6foro en la dermis
Figura 4.4 - Escama de bordes óseos del tipo ctenoideo. (Fuente: Michigan Institute for Fisheries Research).
en los dientes de los vertebrados) y en su interior una cámara pulpar y túbulos dentinales que proceden de ésta. Cada escama tiene una placa basal en forma de disco en la dermis y una cúspide que se proyecta hacia afuera a través de la epi- dermis. Las escamas placoideas se encuentran en los tiburones y especies relacio- nadas (Chondrichthyes) (Figura 4.23 ).
Escamas cosmoideas. Las escamas cosmoideas tienen una capa externa más del- gada, más dura y más separada hacia el exterior que las placoideas. Aunque el material externo del cual están formadas las escamas cosmoideas es ligeramente diferente en su aspecto cristalográfico al de las escamas placoideas, de todas maneras se le llama vitrodentina. La capa particular que está debajo de este esmalte es dura, sin células y ha sido llamada cosmina. Sigue hacia adentro una capa media vascularizada que constituye una zona ósea perforada (llamada isopedina). Se ha reportado que otro carácter distintivo de este tipo de escama se refiere a que el crecimiento ocurre en el borde de la escama desde abajo -no desde fuera- por- que no hay células vivas que cubran la superficie. Las escamas cosmoideas se encuentran en los peces de aletas lobuladas vivientes (Latimeria) y extinguidos. Las escamas tienen en Latimeria una superficie externa denticulada, casi ctenoidea a la observación general. Los peces pulmonados (Dipnoi) tienen escamas que parecen ser cicloideas porque la estructura cosmoidal básica está muy modificada.
Escamas ganoideas. En una escama ganoidea la capa externa está formada por una sustancia inorgánica dura (ganoína) que difiere de la vitrodentina. Debajo de la capa de ganoína hay una capa parecida a l a cosmina. La capa ósea laminar más profunda está formada por isopedina. Además de diferir por su estructura de una escama cosrnoidea, se afirma que una escama ganoidea crece no solamente por los bordes y debajo; tamhikn por la superficie. Entre los peces vivientes las escamas ganoideas están mejor representadas en los polípteros (Polypterus) y los pejes- lagarto (Lepisosteidae) en los que cubren todo el cuerpo. Su forma rómbica también está representada en el lóbulo volteado de la cola de los condrosteos,

108 PIEL
como los esturiones (Acipenseridae), y en los peces espátula (Polyodontidae), co- mo se ha mostrado previamente.
Escamas de borde óseo. Las escamas de borde óseo son típicamente delgadas y translúcidas y les faltan las capas densas de esmalte y dentina que tiene las otras tres clases de escamas. Las escamas de borde óseo caracterizan a las numerosas es- pecies de peces óseos vivientes (Osteichthyes) que tienen escamas cicloideas (Fi- gura 4.3) o ctenoideas (Figura 4.4). La superficie externa de esta clase de escama está marcada con bordes óseos que alternan con depresiones en forma de pequeños valles. La parte interna o placa de la escama está formada de capas de tejido conectivo fibroso que se entrecruzan. El crecimiento de estas escamas se produce tanto en la superficie externa como por debajo.
Para describir la estructura y desarrollo de la escama de borde óseo es nece- sario normalizar los nombres de las partes para evitar las inconsistencias del pasado. El uso de dos términos parece ser permisible para nombrar los bordes de las esca- mas: bordes y círculos. Los cambios en el patrón de crecimiento del individuo pueden estar reflejados en el carácter y distribución de los bordes. En muchas especies han sido identificadas, sobre esta base, marcas natales, marcas meta- mórficas, marcas de reproducción y marcas anuales (annuli) (Figuras 4.3 y 4.4). En las escamas ctenoideas o cicloideas, que son las más ordinarias, puede recono- cerse una zona central o nuclear. Esta zona debe ser llamada con más propiedad el foco de la escama. Es la primera parte en desarrollarse y es a menudo de posi6Ón central, aunque debido al crecimiento diferencial posterior que ocurre hacia el frente o por detrás de la escama se observa un aparente o relativo desplazamiento del foco hacia el margen posterior o el anterior de la escama. En las escamas de muchas especies aparecen unos surcos (radii), que nacen en el foco o cerca de é1 e irradian hacia uno o más márgenes de la escama. Para las denticulaciones de las escamas ctenoideas el término más comúnmente usado y aceptable es el de dien- tes, los que se encuentran localizados en el borde posterior.
Si usamos las escamas para la clasificación e identificación de los peces o para el estudio de sus hábitos de vida, es a veces deseable describir varias regiones en la escama. En la mayoría de las escamas ctenoideas y cicloideas hay un campo anterior fácilmente reconocible que en muchas escamas corresponde burdamente a la porción de su inserción. También hay un campo posterior que a menudo se aproxima a la porción de la escama que puede ser vista sin necesidad de removerla del cuerpo del pez. ¿Mediante qué características puede uno de ordinario recono- cer, si se nos da una escama de pez, el campo anterior o de inserción del expuesto o posterior? En las escamas ctenoideas, como ya se ha señalado, los dientes son partes características del campo posterior. En las escamas cicloideas, el campo posterior puede ser a menudo identificado por el hecho de que los bordes son menos diferentes aquí, pareciendo a menudo como si hubieran sido desgastados. También, con frecuencia, las células pigmentarias (cromatóforos) se adhieren en esta zona.
Respecto a su desarrollo, las escamas de bordes óseos aparecen primero en la dermis en forma de pequeñas agregaciones de células; más a menudo se for- man antes en el pedúnculo caudal, extendiéndose desde ahí. Esa agregación de células pronto forma una plaquita, el foco de la escama definitiva. Estas plaquitas hacen su primera aparición en individuos de distinto tamaño y de especies dife- rentes; en los peces comunes la medida ordinaria de las primeras escamas puede

LAS ESCAMAS 109
ser algo así como dos centrímetros y medio. Pronto los bordes son depositados en la superficie de la escama en crecimiento, un hecho que contradice una idea anticuada en el sentido de que los bordes son los márgenes de láminas o capas sobrepuestas, estructuras de las cuales se presume que las escamas estaban com- puestas, Los únicos bordes, entonces, que aparecen en la superficie de las esca- mas ordinarias de los peces, se generaron durante el desarrollo y aparecen de al- guna manera en el crecimiento. En las aguas de las zonas templadas la deposición se realiza anualmente de acuerdo con los cambios de estación y este fenómeno hace que los círculos se vuelvan indicadores parciales de los anillos que aparecen en la escama del pez. La parte más interna de la escama, la placa (también llamada “placa basal”, “placa laminar”, etc.) está formada de capas sucesivas de fibras paralelas; las de una capa forman con las de la siguiente cruzamientos abruptos. Puede presentarse alguna calcificación de esta capa fibrosa de tal manera que refuerce la escama. En algunos peces, sin embargo, falta la placa fibrosa, como sucede en la anguila (Anguilla), cuyas escamas están compuestas de pequeñas zonas calcáreas dispuestas en forma más o menos concéntrica.
Derivaciones
Se opina que las escamas constituyen el origen de muchas estructuras de los pe- ces. Los dientes de las mandíbulas en los tiburones y especies afines, así como los de los vertebrados superiores, son modificaciones de las escamas placoideas; también lo son las espinas como las de las aletas dorsales del cazón de espina (Squalus) y los quimeroides (Chimaera, Hidrokzgus, etc.) y el “pinchador” en la cola de las rayas de espina (Dasyatidae). Los dientes en sierra del tiburón sierra (Pristis) tienen el mismo origen. Las lancetas de los peces cirujano (Acanthurus) se originaron obviamente de las escamas de bordes óseos, como sucedió también con la escutelación en forma de serrucho del vientre de las sardinas (Clupeidae). Si se hiciera caso omiso de las escamas como partes ancestrales que dieron origen a otras, los radios blandos (lepidotriquia) nunca hubieran aparecido para sostener en posición las aletas en los Osteichthyes, ni los huesos superficiales que cubren el cráneo en estos peces y sus descendientes vertebrados. Los lepidotriquia y los huesos dérmicos son derivaciones de las escamas óseas. Algunas escamas especial- mente modificadas con perforaciones y estructuras en forma de tubo, forman un canal de paredes duras para la línea sensorial del canal lateral que aparece en muchos peces óseos. La armadura también varía, yendo desde las placas óseas espaciadas en los esturiones (Acipenseridae) a los estuches semirrígidos de los peces pipa y los caballitos de mar (Syngnathidae), y finalmente las cubiertas que rivalizan con las de las tortugas mejor protegidas, en los peces cofre (Ostraciidae), por ejemplo el pez torito (Lactophrys quadricornis). En el pez erizo (Diodon hys trh) se pueden observar extensiones de la armadura dérmica más allá de la superficie regular del cuerpo.
Su empleo en la clasificación y en la historia natural
Las escamas de los peces, que son de utilidad para el pez en muchos aspectos, algunos de los cuales han sido sugeridos ya, son también de utilidad para los ictió-

110 PIEL
logos en sus trabajos de clasificación e investigación biológica. ¿Cómo es que las escamas son de utilidad en la clasificación? Haciendo observaciones en los grupos superiores de peces vivientes encontramos que las lampreas y los mixinos (Cyclos- tomata) está desprovistos de escamas; los tiburones y las especies relacionadas (Chondrichthyes) se caracterizan por la posesión de escamas de tipo placoideo; los peces de aletas lobuladas (Crossopterygii) y los pulmonados (Dipnoi) tienen esca- mas cosmoideas; los peces óseos primitivos están dotados de escamas ganoideas (Ganoidei de Agassiz); los peces óseos superiores tienen a menudo escamas de borde óseo. Además de ser de utilidad para hacer una clasificación superficial de los peces, las escamas presentan características que se pueden emplear para separar órdenes y familias. Si alguien es lo suficientemente afortunado de encon- trarse en una región donde buena parte de la fauna está formada por familias representadas por sólo una especie (familias monotípicas), como en los amiidos (Amiidae) y el bacalao (Gadidae), familias que habitan en la región de los Gran- des Lagos de Norteamérica, é1 o ella podrá identificar las especies muy fácilmente basándose solamente en una escama como si fuese el individuo entero. También las escamas tienen utilidad en la clasificación de los restos de peces que se encuen- tran en los basureros (desechos de cocina) que dejó abandonados el hombre pre- histórico y en los depósitos de fósiles. También se ha comprobado que es valiosa la posibilidad de identificar a los peces mediante el examen de las escamas, cuando se trata de estudiar los hábitos alimenticios de los que son carnívoros y devoran a otros peces.
El valor diagnóstico de la morfología de las escamas para la separación de las especies varía entre las familias o géneros. En los grupos que tienen especies estrechamente relacionadas, posiblemente de origen reciente, la estructura de las escamas se parece tanto entre sí como el cuerpo de los peces. Por ejemplo, en los peces blancos (Coregonus), la multitud de formas demasiado parecidas del subgé- nero Leucichthys no pueden ser distinguidas de manera satisfactoria sobre la base del examen de las escamas. Lo mismo sucede con el enorme grupo de las carpas (Cyprinidae), tanto de Asia como de América del Norte. Por lo anterior, se entien- de que la morfología de las escamas no es útil para efectuar separaciones precisas, aunque pueden indicar afinidades O interrelaciones en los grupos superiores. Pero muchísima utilidad tienen las escamas para los trabajos taxonómicos cuando se les cuenta a lo largo de la línea lateral o cuando se determina su número en las hileras que van a lo largo o alrededor del cuerpo (Capítulo 3).
Además del valor que tienen para la identificación y la clasificación, las escamas tienen alguna utilidad en el estudio del ciclo de vida de los peces. Recor- demos, por ejemplo, que mediante los anillos de las escamas (así como vdliéndo- nos de otras estructuras bseas, Figura 5.11) podemos determinar ia edad en años. A partir del espacio que existe entre los anillos anuales y el conocimiento que se tenga de la longitud del pez en el momento de su captura, podernos determinar ia longitud que tenía el pez en relacibn a tal o cual espacio, siempre que hayamos determinado ia proporcionalidad entre el crecimiento de l a escama y el dei pez. ,Qunus veces tarnRIC;n las e s ~ m m s s no5 permiten descubrir las veces que el pez ha yeajizado ::j :5",esove. Z7n ei .:aimfin del A t l h t i ~ c : jSalm+o saiace.r\, por ~jemplo, se .;~uec.iep. c j ~ ~ , e r v a : - -nx;~~:;ks de 1: :.sr,csj.ciiirz ex?. Las est.:unas. Por io tante, w x j puede
decir ia edad que cenia 21 pez ::uandi: !hizo ;u .primer viajo a i mar: su ~ i a c i m c! :nczme~?io d e ia, captura, su edod ce;ando hizo la ?rimera ovoposicir?n y las veces ?que i-ia llegado a poner huevecillos. L a &oca en que el individuo fue por primera

BARBILLAS Y ALETAS 111
vez al mar puede ser discernida por el cambio abrupto que se observa en la distan- cia que hay entre los círculos de la escama, porque cuando el crecimiento se ace- lera en el mar los círculos están más espaciados.
BARBILLAS Y ALETAS (FIGURAS 4.6f y g)
Además de la desnudez que les da cierto aspecto liso y algunas divergencias de relativamente poca importancia en la disposición básica de las escamas, los peces han desarrollado también una gran variedad de extensiones tegumentarias. Entre ellas se cuentan unas extensiones extrañas de la piel para formar barbillas y aletas. Las barbillas k q n aparecido en forma independiente en muchos de los grupos supe- riores de peces, como estructuras accesorias que sirven para la alimentación y llevan consigo órganos sensoriales. Barbillas de diferente estructura y ubicación se pueden encontras en los esturiones (Acipenser), los peces gato de agua dulce o marinos (Cypriniformes nematognatos o siluroides), los peces barbones (Mullidae) y muchos otros. El pez sargazo (Pterophryne) y el dragón marino que semeja un alga (Phyllopteryx) son los ejemplos citados con mayor frecuencia por la exten- sión de su piel en lengüetas. La función que tienen éstas en las dos clases de peces es la imitación protectora; o, por lo menos, no puede negarse que los peces con lengüetas se confunden con esos adornos con su medio ambiente.
Ciertas generalizaciones pueden ser hechas a partir de la gran variedad de colora- ciones que exhiben los peces, si tomamos en cuenta el modelo básico relacionado con su tipo de vida. Los peces de vida libre que habitan en las aguas abiertas, como las diversas clases de sardinas y especies afines (Clupeidae) y los peces de los fondos marinos durante su juventud, cuando viven en forma planctónica, como los lenguados (Pleuronectidae), presentan generalmente una coloración simple que va de un vientre blanquecino, a través de los lados inferiores plateados, a los lados superiores y el dorso de color azul o verde iridiscente. Los cavadores de los fondos y los peces que viven entre la vegetación acuática tienen a menudo marcas muy contrastadas y de figuras complicadas en el dorso, y una coloración pálida por debajo. Las especies que frecuentan los arrecifes de coral tropicales son las que tienen los colores más brillantes, elegantes y adornados. Entre éstos se cuentan los peces cardenal (Apogonidae), los peces mariposa y los peces ángel (Chaetodontidae), los peces gallo (Labridae), las mojarras rayadas (Pomacentri- dae), los peces cirujano (Acanthuridae), los peces guacamaya (Scaridae) y los peces cocuyo (Balistidae). Los machos de muchas de las percas (Percidae: Etheos- tomatinae), peces nativos de los ríos del oeste de América del Norte, están bella- mente coloreados, como sucede también con los pececillos tropicales que se venden para los acuarios. Tres clases de color predominan en los peces oceánicos: plateado en las zonas cercanas a la superficie; rojo en las intermedias; y violeta o negro en las grandes profundidades.
La coloración de los peces se debe en primer lugar a los pigmentos de la piel. El color de fondo o aspecto se debe, desde luego, a los tejidos subcutáneos, a los fluidos del cuerpo e inclusive al contenido del tubo digestivo. El color de

112 PIEL
fondo es el matiz esencial de los peces cavernícolas ciegos (Amblyopsidae). En otros peces los colores varían desde brillante a opaco, enmascaran el color de fondo y son de origen dérmico. La coloración básica más común en los peces es la brillantez que prevalece en la superficie ventral del cuerpo, la oscuridad en la parte dorsal y un cambio gradual de intensidad hacia los lados, desde brillante en el viente hasta oscuro en el dorso, como ya se dijo. Este plan ilustra el primer principio del ocultamiento mediante la anulación de la sombra contraria; los estu- diosos de la coloración animal llaman a esto principio de Thayer. Más allá de este plan general existen muchos rasgos extraordinarios en la coloración de los peces. Uno de éstos es la uniformidad del color, debida ya sea a la falta o exceso del pigmento en los albinos y los negros, respectivamente.
Jóvenes pelágicos y de vida acuática libre de muchas clases de peces carecen de pigmento y, en consecuencia, son transparentes. Otra característica extraor- dinaria que se presenta es la coloración casi uniforme debida a la presencia de uno u otro colorante. Un ejemplo de esto podría ser el color dorado uniforme que caracteriza a ciertas razas genéticas de la carpa dorada (Carassius) o el magnífico color naranja mostrado por el garibaldi (Hypsypops) de la costa sur de California. Todavía otros ejemplos de aproximación a la coloración uniforme podrían incluir el melanism0 que caracteriza a muchos de los peces de las profundidades mari- nas, aunque algunos han revertido la anulación de la sombra contraria. Otras especies de peces, sin embargo, rivalizan en su coloración con las más hermosas mariposas o aves, por las combinaciones de los muchos tintes que aparecen en sólo un individuo.
Orígenes del color
La coloración de los peces se debe a los esquemacromos (colores que resultan de la configuración física) y a los biocromos o pigmentos verdaderos (Fox, 1953). Los esquemacromos blancos pueden ser observados en el esqueleto, la vejiga gaseosa, las escamas y los testículos; los azules de Tyndall y los violetas aparecen en el iris; y los colores iridiscentes pueden ser vistos en las escamas, los ojos y las mem- branas intestinales. El integument0 también contiene los esquemacromos ya mencionados. Los biocromos incluyen a los carotenoides (amarillo, rojo y otros tonos), cromolipoides (de amarillo a café), indigoides (azul, rojo y verde), mela- ninas (generalmente de negro a café), porfirinas y pigmentos biliares (rojo, ama- rillo, verde, azul y café), flavinas (amarillo, a menudo con fluorescencia verdosa), purinas (blanco o plateado) y pterinas (blanco, amarillo, rojo y naranja). Los carotenoides, las melaninas, las flavinas y las purinas se encuentran en la piel del pez. El hígado, huevos y ojos contienen carotenoides. Las melaninas aparecen en el endoderm0 y la piel. El músculo y la sangre contienen porfirinas, mientras que el esqueleto y la bilis contienen pigmentos biliares. Las flavinas est&, amplia- mente distribuidas en la sangre, el músculo, el bazo, las branquias, el corazón, los riñones, los huevos, el hígado y los ojos. Las purinas están en las escamas y los ojos. Las pterinas se encuentran en los ojos, la sangre, el hígado, los riñones y el estómago.
Las células especializadas que dan color a los peces son de dos clases, los cromatóforos (Figura 4.5) y los iridocitos. Los cromatóforos están ordenados por tonos y dan el verdadero color. Están localizados en la dermis, ya sea por fuera o

113
Xantóforos vlelal nóforos
Guandforos
Figura 4.5 - CBlulas de color de la piel de un lenguado, Paralich- rhys. (8asado en Kuntz, 1917).
bajo las escamas. También a menudo se les encuentra en el peritoneo e, inclusive, muy profundamente, alrededor del cerebro y la médula espinal. Los gránulos pigmentarios, que son inclusiones citoplásmicas de los cromatóforos, son los cor- púsculos que proporcionan el color. Muchos de los cambios de coloración que exhiben los peces se deben a la migración de los pigmentos dentro de los croma- tóforos, ya que los gránulos se pueden dispersar a través de la célula, o concentrar en la misma. Los pigmentos de los gránulos reflejan algunas longitudes de onda de la luz y absorben otras; las reflejadas son las que se pueden ver, las absorbidas no pueden ser vistas. De acuerdo a los colores de sus gránulos pigmentarios los cromatóforos básicos son rojo y naranja (eritróforos), amarillo (xantóforos), ne- gro (melanóforos) y blanco (leucóforos). Los pigmentos rojo, amarillo y naranja son más a menudo carotenoides que han sido asimilados a través del alimento y están relacionados con la vitamina A. Los pigmentos negro o café, las melaninas, son compuestos de polimerización elevada derivados del aminoácido tirosina y otros fenoles. Además de estos colores los peces exhiben otros como el verde y el café. Estos y otros tonos se originan de varias asociaciones de las tres clases bási- cas de cromatóforos. Los cromatóforos negro y amarillo esparcidos entre sí dan, por ejemplo, vistosos tonos verdes. El amarillo y el negro dan café, pero el naranja y el azul pueden también asociarse para dar el mismo color.
Los iridocitos bien podrían ser llamados células especulares porque contie- nen materiales reflectantes que reflejan los colores del medio ambiente donde vive el pez. Los leucóforos y los iridóforos contienen purinas, principalmente guanina, pero los primeros contienen pequeños cristales que pueden desplazarse hacia to- dos lados dentro del citoplasma, mientras que en los segundos hay cristales grandes que no se pueden mover y usualmente están amontonados formando capas. La guanina misma es un producto derivado de los ácidos nucleicos (material genético).

114 PIEL
Significado
Las funciones de la coloración y otras señales visuales (como la bioluminiscencia) sirven para la comunicación con otros miembros de la misma especie (señales intraespecíficas) y con otras clases de animales (señales interespecíficas). Las señales intraespecíficas tienen utilidad social (para el reconocimiento, la amenaza, la pelea) y sexual, y pueden también ser empleadas por el pez huésped para atraer a los pececillos hacia su cuerpo con propósitos de limpieza. Los cíclidos jóvenes nadan en grupos cerca de sus padres y se acercan rápidamente a ellos cuando el peligro se presenta. El reconocimiento de sus padres se basa parcialmente en el co- lor. Por ejemplo, en Hemichromis, que es rojo brillante durante el periodo en que cuida de su cría, el pececillo tiene una preferencia pronunciada, entre todos los colores, por el rojo y al anaranjado. Las señales interespecíficas pueden ser em- pleadas para la advertencia o la intimidación de predadores potenciales y otros asaltantes, o como cebo o propósito de enmascaramiento dirigidos ya sea a la presa o al predador. Cott (1940) agrupó principales funciones de la coloración bajo tres encabezados: críptico o de ocultación, de enmascaramiento y apose- mútico o de advertencia (Figura 4.6). Se sugiere que las diferentes clases de ocul- tamiento mediante el color son: 1) la semejanza del color en general; 2) la seme- janza variable del color; 3) el sombreado que destruye el contorno; 4) la colora- ción disruptiva, y 5) la coloración disruptiva coincidente.
Semejanza del color. La semejanza general del color entre un pez y el medio am- biente es la característica básica de los peces que imitan las sombras y los colores de los hábitats que frecuentan. Como se ha establecido previamente, muchos peces de los arrecifes de coral son tan coloridos como los corales que frecuentan, pero algunos pueden destacar su color para advertir su presencia a los pececillos que acostumbran limpiarlos de parásitos, o para poder ser reconocidos por sus congéneres o sus parejas. Los peces que viven en zonas donde abunda la vegeta- ción acuática, donde predominan los claros y sombras, son generalmente motea- dos. Los peces de los litorales que viven sobre fondos de sombra clara tienen colores brillantes, pero las mismas especies son oscuras cuando viven sobre fondos muy sombreados (o a grandes profundidades o durante la noche).
Semejanza variable del color. Es la capacidad de un pez (o cualquier otro animal) para cambiar el color, gradual o rápidamente, con el ostensible propósito de imitar su medio ambiente a la mayor perfección. Algunas de esas variaciones en la seme- janza del color ocurren durante el ciclo biológico. La fase de residencia en los arroyos de la trucha arco iris migratoria (Salmo gairdneri) es multicoloreada, con manchas negras en los jóvenes, y lados rosados en los adultos. Pero, en el mar, la coloración del mismo individuo cambia de azul metálico en el dorso a blanco en el vientre, pasando por plateado. También hay variación de la coloración entre los peces con los cambios de estación, del día y la noche, e inclusive con los mo- mentáneos del hábitat. La trucha (Salmo), viviendo en un medio acuático bri- llante o con pequeñas zonas sombreadas, durante el verano, tiene un aspecto moteado. Pero cuando vive en estanques o en aguas cubiertas por hielo durante el invierno, será siempre totalmente oscura.
Una forma de semejanza variable del color hace que se vea un cambio en la forma o estructura del cuerpo, o sea, hay ruptura del contorno visual. Esto es
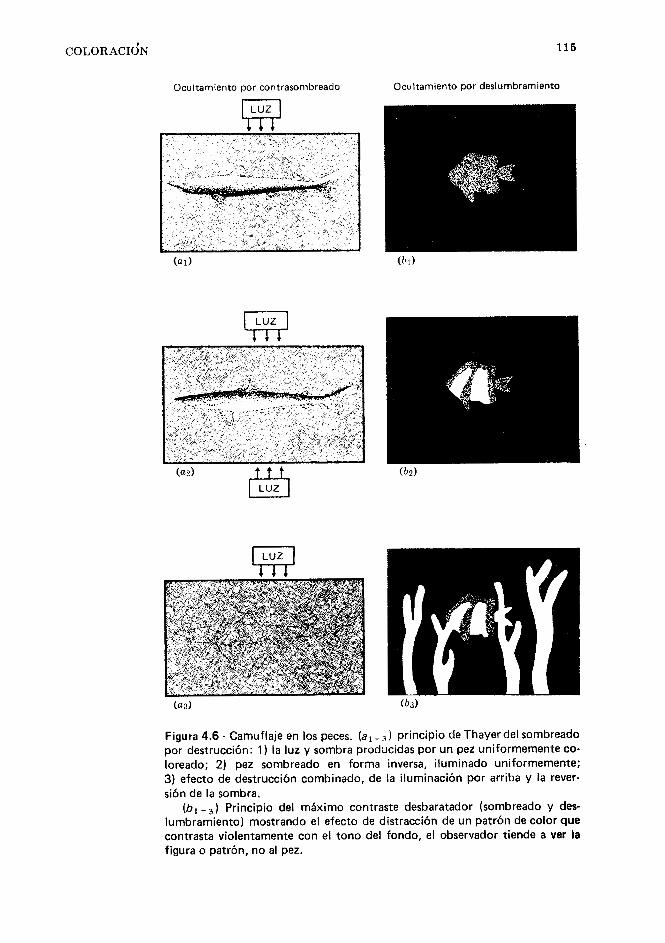
116
Ocultamiento por contrasombreado Ocultamiento por deslumbramiento
Figura 4.6 - Camuflaje en los peces. (al - 3 ) principio de Thayer del sombreado por destrucción: 1) la luz y sombra producidas por un pez uniformemente co- loreado; 2) pez sombreado en forma inversa, iluminado uniformemente; 3) efecto de destrucción combinado, de la iluminación por arriba y la rever- sión de la sombra.
(b, - 3 ) Principio del máximo contraste desbaratador (sombreado y des- lumbramiento) mostrando el efecto de distracción de un patrón de color que contrasta violentamente con el tono del fondo, el observador tiende a ver la figura o patrón, no al pez.

116
Ocultamiento mediante la coloración disruptiva o de rompimiento
PIEL
Marca deflectiva protectora
Figura 4.6 - Continuación. (c, - 2 ) Ocultamiento mediante la coloración de rompimiento: 1) en un
mero de Nassau (Epinephelus stfiatus), y 2) en un joven pez hacha (Equetus lanceolatus).
(d) Coloración de rompimiento y marca que pueden desviar los ataques del predador en un pez ángel joven (Pornacanthus).

117
Aviso mediante órganos luminosos
Pez linterna
Parecido protector mediante la imitación
Dragón marino Pez imitador de hoja
Figura 4.6 - Conrinuación. (e) Anunciamiento a traves de la posible protección o reconocimiento da-
dos por los caracteres en los órganos luminosos (lunares brillantemente coloreados) en un pez linterna (Mycrophum). (4 Imitación protectora tanto en el color como en la forma de una alga
marina, en un drag6n marino (Phylopreryx). (g) Imitación protectora del color (note "venación") y la forma de una
hoja en el pez imitador de hoja (Monocirrhus). Parte (e) basada en Jordan y Evermann, 1900; todos los demás en Cott,
1940.


C O L O R A C I ~ N 119
vés de su vida desde el momento en que hay cambios a corto y largo plazo. Esos cambios o respuestas pigmentarias son morfológicas y fisiológicas. En el caso de los melanóforos el cambio morfológico es lento y exhibe la formación de pig- mento y células. El cambio fisiológico puede ser rápido (a menudo en unos cuan- tos minutos) e involucra una redistribución de los gránulos pigmentarios contenidos en los cromatóforos. Estos dos tipos de cambio son respuestas a estímulos, sean visuales o no visuales; y, en el segundo caso, puede ser coordinado por nervios y hormonas, o no coordinado, en el cual el melanóforo es su propio receptor y efec- tor (no conocido para los peces). En los tiburones sin ojos (Mustelus) y los bagres ciegos (Ictalurus) el oscurecimiento tiene lugar en la luz y el empalidecimiento en la oscuridad, mientras que los individuos normales pueden oscurecerse por dispersión melánica cuando son colocados en un medio oscuro. Igualmente, to- mando en cuenta las respuestas hormonales de la pituitaria, la extraccih de esta glándula da como resultado la palidez de otro tiburón (Scyliiurn) y la implanta- ción de la misma resulta en el oscurecimiento de una raya (Raja) . El papel que desempeñan los impulsos nerviosos en la regulación cromática ha sido demostrado mediante la sección de los nervios periféricos para estimularlos después mediante corriente eléctrica cuando el área denervada se mantenga oscurecida. En Scyllium, por lo menos, el oscurecimiento considerado como una respuesta en forma de fondo negro involucra al MDH (hormona dispersora de melanina) y el empalideci- miento en fondo blando al MDH o la secreción de MAH (hormona acumuladora de melanina), ambas procedentes de la pituitaria. También hay buena evidencia de la acción del MDH en algunos teleósteos (Anguilla, Ictalurus y Fundulus). En las especies de teleósteos que se han estudiado, relativamente pocas, hay información irregular sobre el mecanismo para el cambio de color, pero hay concenso general en el sentido de que la coordinación de ese cambio se debe a la interacción que existe entre los controles nervioso y hormonal.
Sombreado que destruye el contorno (Figura 4.6a). Hay todavía otra clase de co- loración para el ocultamiento y consiste en el sombreado que destruye el contorno del animal. El principio óptico en el que se basa este tipo de coloración para el ocultamiento depende de la inversión de la sombra. La luz y la sombra dan a un observador la tercera dimensión del objeto que ve. La sombra, por lo tanto, cons- tituye un serio problema por resolver cuando se trata de reducir la visibilidad de cualquier objeto, como sucede en los tiempos de guerra, en que se procura oscu- recer un vehículo, un barco o un aeroplano. La inversión práctica de la sombra queda evidenciada en la mayoría de los peces -también en muchas aves, mamí- feros, reptiles y anfibios- porque tienen la superficie dorsal oscura y el vientre claro, tendiendo a volverlos ópticamente planos, como una sombra (es el principio de Thayer cl que se aplica). Las superficies expuestas normalmente a la luz son in- vertidas en su sombra mediante el oscurecimiento, mientras que las normalmente expuestas a la sombra son invertidas mediante el aclaramiento; y si a lo anterior se agrega que hay tonos intermedios y graduales entre las dos superficies, el ob- jeto entonces se verá plano y reducida su visibilidad. La inversión de las sombras ha desaparecido en los peces cavernícolas (Amblyopsis y Anoptichthys) y se ha revertido en las grandes profundidades marinas, como ya se dijo.
Coloración disruptiva (Figura 4.6b, c, d). Otro mecanismo de ocultamiento que han desarrollado los peces es la coloración disruptiva, un nuevo sistema de enmascara-

120 PIEL
miento. Desde el momento en que una superficie cualquiera, limitada por un con- torno definido, nos permite reconocer el objeto al que pertenece, porque la figura nos es familiar, la destrucción de ese contorno nos puede hacer titubear, porque tiende a ocultar al animal. Por lo tanto, para que el ocultamiento sea efectivo es necesario que la figura familiar representada por la superficie y su contorno sea destruida. La función de la desfiguración, benéfica para un pez, puede pensarse, entonces, que consiste en la prevención, o por lo menos en la demora del recono- cimiento visual. Cuando la superficie del pez está cubierta por manchas irregulares de colores y tonos contrastados, estas manchas tienden a desviar la atención del observador y a alejarla de la superficie que las contiene. Al contradecir la forma, las manchas hacen que la atención se concentre en ellas mismas, de tal manera que las figuras que forman esas manchas hacen que el objeto que las contiene -en este caso el cuerpo del pez- parezca por ilusión óptica que es parte del medio ambiente general.
Coloración disruptiva coincidente. Es una clase especial de enmascaramiento. Puede aparecer como la unión de partes separadas y no relacionadas del cuerpo con el fin de evitar el fácil reconocimiento. Los apéndices y los ojos son ocultados mediante este procedimiento. Aunque la forma del ojo puede ser cambiada, es la pupila negra en especial la que sufre modificaciones en su contorno mediante varias formas de máscaras disruptivas coincidentes y, por lo tanto, unida la figura con la de cualquier otra parte del cuerpo, deja de parecer un ojo. Sin embargo, también se ha propuesto que la línea del ojo, ya sea la horizontal o la vertical, cons- tituye una línea visual para dirigir el ataque (Figura 4.6, c1 , cz ).
Formas relacionadas de ocultamiento y disfrazamiento (Figura 4.6d, f, g)
También asociados con el ocultamiento mediante el empleo del color hay otros mecanismos de enmascaramiento que a menudo relacionan la estructura con el hábito de vida, como bien lo ha mostrado Breder (1946). Algunos hábitos, como las actitudes crípticas, son empleados para ocultar la posición y pueden completar el papel desempeñado por el enmascaramiento mediante el color.
Los peje lagartos (Lepisosteus) cuando jóvenes y adultos tienen el hábito de asolearse, permaneciendo inmóviles cerca de la superficie del agua. Por su pos- tura, forma y coloración imitan notablemente a ramas y troncos que a veces llegan a flotar en el agua. Strongylura, un género de peces aguja (Belonidae) se parece mucho a los tallos de las plantas de su hábitat. Los jóvenes de algunos peces paja- rito (Hemiramphus) y peces pipa (Syngnathidae), cuando son vistos de perfil, se parecen a las hojas del pasto marino (Zostera). Otros peces que en estado juvenil o siendo ya adultos recuerdan a las hojas de las plantas son el jurel (Oligoplites scaurus), la chopa (Lobotes surinamensis), el pez lija (Monacanthus polycanthus), algunos peces espada (Ephippidae; Platax) y la cochinita cimarrona (Alutera schoepfi). Los jóvenes de algunos peces voladores (Cypselurus) se parecen mucho a los botones de una planta (Barringtonia) que vive en su hábitat. Hay muchos peces, como los dragones marinos (Syngnathidae; Phyllopteryx), el “dwarf wrasse” (Doratonotus megalepsis) y el sargacero (Heterostichus rostratus), que recuer- dan, porque las imitan, a las frondas de algas (por ejemplo el talo) o sus fragmen- tos. Además, las chabelas del Atlántico (Chaetodipterus faber) recuerdan vainas de

C O L O R A C I ~ N 121
una planta (Rhizophora) y el “lumpfish” (Cyclopterus lumpus) se ve como si fuese una cápsula de algas en flotación.
El disfrazamiento es también realizado por varios caracteres conspicuos y bien localizados, los que simplemente tienden a reducir el parecido de un pez a lo que es, o sea, ;un pez! Las marcas deflectivas y directivas son importantes aquí. Se piensa que las marcas deflectivas son las que desvían el ataque del enemigo de una parte más o menos vital del cuerpo a otra que no lo sea. Un ejemplo muy simple es la mancha negra que aparece en la cola de Amia; otro es el joven de ciertas galline- tas (Pomacanthus, Figura 4.6d). Durante el ritual en el cual el pez ángel (Ptero- phyllum scalare) defiende su territorio aparecen cambios en la intensidad del color oscuro de la figura en forma de barra vertical que presenta su cuerpo y en la mancha que figura un ojo sobre cada opérculo. Se ha pensado que, por el con- trario, una marca directiva tiene el fin de distraer la atención de la presa de la parte más peligrosa del cuerpo del predador. Los peces sapo (Uranoscopidae) tienen bocas rodeadas por flecos que pueden muy bien ocultar el órgano cuando el pez está casi cubierto por la arena. Las víctimas nadan hacia la región de la boca, hasta cierto punto oscura, y lo hacen atraídas posiblemente por unas aletas que imitan alimento y son devoradas (se ha pensado inclusive que la función pri- mordial de esas aletas sea la de retirar la arena de la boca del predador). La tenta- ción de la presa para dirigirse hacia la región bucal es otra función que realizan las marcas directivas y constan usualmente de una combinación de estructura y color. Los peces antenados (Lophiiformes) tienen uno o dos apéndices delgados y pedunculados, siendo cada uno una “varilla-cebo” virtual (illicium) ; se extien- den hacia adelante sobre la nariz y algunas veces poseen un pequeño bulbo lumi- noso o una especie de borla que provoca la tentación, en la punta de la estructura. Se dice que un pez sapo (Uranoscopus) del Mediterráneo tiene un pequeño “gu- sano” rojo en su boca y sirve de tentación a la presa para provocar su muerte.
La advertencia o aviso (Figura 4.6e)
Algunas formas de coloración de los peces parece ser que sirven para anunciar o revelar su presencia, más que para ocultarla. Ejemplos muy notables de este caso se encuentran entre los eteostomátidos (Etheostomatinae) de los ríos americanos. Entre estos miembros pequeños de la familia de las percas (Percidae) están los peces de agua dulce más brillantemente coloreados de este continente. Este tipo de coloración puede ser de significación para la atracción sexual; por lo tanto, es una forma de aviso. Algunos experimentos que se han realizado con la espinocha (Gasterosteus) sugieren que el color tiene valor para el reconocimiento sexual en algunos peces, pero los intentos que se han hecho con otras especies han mostrado que el color no es discriminado o puede que no tenga valor para este fin.
Su empleo en la clasificación
El diseño colorido en los peces es usado a menudo como un carácter para distin- guir las categorías taxonómicas inferiores como son las especies y subespecies (Capítulo 13). A menudo la disposición individual exacta de los cromatóforos que resulta en el patrón o diseño fundamental de color, está bajo control genético, a

122 FIEL
pesar de la variación que existe entre los individuos de un tamaño y clase, y la variabilidad en relación al sexo, edad, habitat, etc. (Capitulo 12) . N o es poco co- mún, por lo tanto, que los caracteres de color aparezcan en ¡a descripción de los grupos taxonómicos de menor nivel.
dRGANOS LUMINOSOS
La luz en los seres vivos (bioiuminiscencia), aunque no aparece en los peces de agua dulce, no es rara en los oc6anos y en muchos grupos de animales, incluyendo a numerosos peces marinos. Es parecida a la. que producen las luciérnagas y 10s chupiros (Lampyridae). La producci6n de luz se ha desarrollado más extensamente en muchas de las especies que viven a media agua o como cavadoras en los fondos profundos. Se sospecha que los brganos luminosos les sirven a los propios peces para reconocer a las especies y a sus parejas. Ciertos órganos luminosos de la ca- beza quedan implicados para actuar corno cebos que permiten atraer a las presas. La luminiscencia produce generalmente luz de calor azul o verde. Los órganos tienen utilidad como caracteres taxonórnicos; por ejemplo, en los peces linterna y especies afines (Myctophiformes).
La luminiscencia en los peces es de dos tipos (Harvey, 1957): a) la que resulta de la presencia de bacterias luminosas que viven en el pez en una relación simbiótica; b ) la que nace de chlulas por sí mismas luminosas, propias del pez, los fotóforos. La luminiscencia propia puede ser luz generada por el animal en sus tejidos (luminiscencia intracelular) o mediante la descarga de una secreción lumi- nosa (luminiscencia extracelular). Un ejemplo del segundo caso lo tenemos en el searsiido, Searsia schnakenbecki.
Las familias de peces en las que la luminiscencia es de origen bacteriano están representadas en las aguas donde hay bancos de arena y en grandes profun- didades; como ejemplos tenemos a los granaderos (Melanonidae), los bacalaos (Gadidae), los peces cono de pino (Monocentridae), los anomalópidos (Anoma- lopidae), los peces cardenal luminoso (Rcropomatidae), los “slipmouths” (Leiog- nathidae), los serranos (Serranidae), los cardenales (Apogonidae), los tragadores (Saccopharyngidae) y algunos peces de las profundidades marinas que pertenecen al orden Lophiiformes. Los elasmobranquios que tienen luminiscencia propia incluyen a ciertos tiburones (Spinax, Centroscyllium, Etmoptems) y una raya eléctrica (Benthobatis rnoresbyi). El mayor número de especies con luminosidad propia entre los peces óseos está representado o formado por los peces dragón con escamas que viven en las grandes profundidades marinas (Stomiatidae) y los peces linterna (Myctophiformes). Los fotóforos también están presentes en los pe- ces sapo y los “midshipmen” (Ratrachoididae).
Se han dado a conocer muchas hipótesis para explicar el valor de la biolu- miniscencia entre los peces. La evidencia respalda la importancia de los órganos luminosos de especies tales como el pez linterna Gonichthys y los estomiatoideos para alumbrar las aguas oscuras donde viven. También la bioluminiscencia cumple un importante papel durante el cortejo en el pez “midshipman”. Otras hipótesis que esperan la confirmación explican el uso de la luz como cebo para la presa, para confundir a los predadores, como aviso o para desvanecer la silueta del pez. Muchas otras posibilidades también incluyen la territoriaiidad de los peces que viven a media agua. La persistencia de los órganos luminosos en el lado ventral

GLANDULAS VENENOSAS 123
de estos peces sugiere que los protegen contra los predadores que viven nadando arriba de ellos.
GLÁNDULAS VENENOSAS
En muchos peces existen glándulas venenosas como formaciones derivadas de la piel, y más precisamente como adaptaciones de las glándulas mucosas. Estas glán- dulas secretan un veneno que, inyectado en el hombre, puede ser muy doloroso, pero rara vez es letal. El estudio de estas glándulas y sus secreciones es abordado por la ictiotoxicología, tan adelantada por el trabajo de Halstead (P.e., 1959 y 1970). El campo de la ictiotoxicología incluye el estudio de las diversas formas de intoxicación que resultan de la ingestión de peces venenosos (ictiosarcotoxis- mo) o por la picadura con las espinas venenosas de algunos peces (ictioacantoto- xismo) .
Siendo derivaciones del integumento, las glándulas venenosas se han desa- rrollado de una manera independiente multitud de veces en familias de peces ampliamente separadas. Se sabe que el ictioacantotoxismo resulta de la picadura de muchos peces; en la siguiente lista se dan algunos ejemplos, para lo cual se des- cribe el aparato picador y se citan los venenos. Los ejemplos de peces han sido seleccionados de tal modo que representen a muchos brdenes muy separados des- de el punto de vista filogenético.
a. Tiburones: espina en la aleta y epitelio glandular venenoso en un surco; en el tiburón cornudo (Heterodontus francisci) y en el galludo (SquaZus acanthias).
b. Rayas (Dasyatidae y Myliobatidae): espina caudal con piel glandular en su vaina; raya de espina (Dasyatis), raya pinta (Aetobatus narinari) y en la raya murciélago de espina (Myliobatis californicus).
c. Quimeras (Chimaeridae) : espina de la aleta dorsal con veneno procedente del epitelio glandular que forma su vaina y cubre el surco en la Chimaera y en el Hidrolagus (el pez ratón).
d . Peces escorpión; peces león y rocotes (Scorpaenidae): espinas de las ale- tas dorsal, anal y pélvica con glándulas venenosas en sus surcos; quedan incluidas especies de pez escorpión (Scorpaena), “hullrouts” (Notesthes), peces león (Pterois) y peces piedra (Synanceja).
e. Peces araña (Trachinidae): espina en el opérculo y otras en la aleta dor- sal, todas con glándulas venenosas en sus surcos; Trachinus.
f . Peces sapo (Batrachoididae): espina en el opérculo y otras en la aleta dorsal, todas con glándulas venenosas en sus bases; Batrachoides, Tha- lassophyryne y Opsanus.
g . Bagres: las espinas de las aletas dorsal y pectorales con glándulas bajo la piel, vertiendo al exterior a través de poros en la base de las espinas; “stonecats” y “rnadtoms” (Noturus), los bagres (Ictalurus), otros bagres y el bagre bandera (Galeichthys felis y Bagre marinus) y bag-res del Indo- Pacífico (Heteropneustes, Clarias y Plotosus).
h. Peces cirujano (Acanthuridae): aguijón en cada lado del pedúnculo cau- dal, presumiblemente con glándulas venenosas en la vaina de la espina; Acanthurus, Nuso.

124 PIEL
i. Dragonetes (Callionymidae): espinas en las aletas con glándulas veneno-
j . Peces conejo (Siganidae): las espinas de las aletas dorsal, anal y pélvicas
h. Peces sapo (Uranoscopidae): espinas en los hombros con glándulas vene-
sas; Callionymus.
con glándulas venenosas; Siganus.
nosas en sus bases.
El veneno producido en las púas de las especies de peces que se menciona- rán a continuación puede producir síntomas graves y dolorosos y aún causar la muerte al hombre: rayas, quimeras, peces escorpión, peces araña y peces sapo. Veneno no fatal pero, sin embargo, doloroso para los humanos, a veces muy dolorosos: tiburones, peces sapo, bagres, peces cirujano, peces conejo y dragonetes. Todas las heridas producidas por las espinas pueden estar expuestas a una infección secundaria donde la gangrena y el tétanos serían las más importantes. Nada se sabe del significado biológico del aparato venenoso de los peces, aunque es fácil aducir que juega un importante papel en la alimentación, el ataque para lograrla y la defensa.
REFERENCIAS ESPECIALES SOBRE LA PIEL
Breder, C.M. 1946. An analysis of the deceptive resemblances of fishes to plant parts, with critical remarks on protective coloration, mimicry, and adaptation. Bull. Bingham Ocea- nogr. Coll., lO(2):l-49.
Cott, H.B. 1940. Adaptive coloration in animals. Oxford University Press, Nueva York. Crozier, G.F. 1974. Pigments of fishes. En Chemical Zoology, 8:509-521. Academic Press,
Nueva York. Fox, D.L. 1953. Animal biochromes and structural colors. Cambridge University Press, Cam-
bridge. Fox, D.L. 1957. The pigments of fishes. En The Physiology o f fishes., 2:367-385. Academic
Press, Nueva York. Halstead, B.W., L.S. Kuninobu, y H.G. Hebard. 1953. Catfish stings and the venom appara-
tus of the Mexican catfish, Galeichthys felis (Linnaeus). Trans. Amer. Microsc. Soc., 72(4):297-314.
Halstead, B.W. 1959. Dangerous marine animals. Cornel1 Maritime Press, Cambridge, Md. -, 1970. Poisonous and venomous marine animals o f the world. U.S. Gov't. Printing Office,
Harvey, EN. 1957. The luminous organs of fishes. En The physiology of fishes, 2:345-366.
Nicol, J.A.C. 1969. Bioluminescence. En Fish Physiology, 3:355-400. Academic Press, Nueva
Washington, D.C. 3 volúmenes. I
Academic Press, Nueva York.
York. Odiorne, J.M. 1957. Color changes. En The physiology of fishes, 2:387-401. Academic Press,
Nueva York.
mic Press, Nueva York.
Rept. Inst. Freshwater Res. Drottningholm, 38~385-447.
York.
Van Oosten, John. 1957. The skin and scales. En The physiology of fishes, 1:207-244. Acade-
Wallin, O. 1957. On the growth structure and developmental physiology of the scale of fishes.
Waring, H. 1963. Color change mechanisms of cold-blooded vertebrates. Academic Press, Nueva

Capítulo 5
Alimentos, digestión, I O o r
nutrición O O 1 y crecimiento
Para los peces, como sucede con todos los animales, es indispensable una nutri- ción adecuada para poder crecer y sobrevivir. A través de la observación en el campo y la identificación minuciosa de los contenidos del tracto digestivo, y a través también de los estudios fisiológicos en el laboratorio, los investigadores han aprendido mucho en relación a los hábitos alimenticios, las clases de organismos que comen y los mecanismos que han desarrollado para realizar la digestión. Los experimentos también han dado a conocer valores dietéticos de varias clases de alimentos y han analizado los factores que afectan al crecimiento. Ha sido parti- cularmente significativa la información que sobre la nutrición se ha obtenido, como un logro del hombre en sus intentos por ayudar a propagar los peces de la manera más eficiente que sea posible.
La anatomia general del tracto digestivo es descrita en el Capítulo 3.
EL ALIMENTO NATURAL DE LOS PECES
Variedades de alimentos
Una gran variedad de alimentos ofrece a los peces la Naturaleza, entre los cuales quedan incluidos los nutrientes en solución y huéspedes de diferentes plantas y animales. Poco es lo que se conoce sobre la utilización directa de los nutrientes solubles por los peces, aunque hay alguna evidencia del aprovechamiento que hacen de la glucosa disuelta en el agua ambiente. Muchos compuestos necesarios e innecesarios, y iones del agua, son absorbidos directamente, a través de las branquias por ejemplo, o son deglutidos con el alimento y después absorbidos
125

en e¡ tracto digestivo. Por ejemplo, existe la absorción extra-enteral para el calcio, t,an necesario para la formaci6n de las escamas y e! hueso.
Es de esperarse que un grupo tan dlversificatlo como el de los peces se haya adaptaado a una gran variedad de alimentos. Algunos peces se alimentan exclusiva- mtmte de plantas, otros de animales solamente, 121ientras que existe un tercer grupo que se alimenta de las dos fuentes. Peces parkitos especializad.os como la lamprea marina (Yelromyaon rnarinus) viven de la sangre y fluidos producidos en los teji- dos de otros peces. Muchos peces jbvenes, c o n h x a s pequeñas y mostrando a menudo todavia el saco de la yema, comienzan por alimeniarse de plancton, plan- tas y animales microscbpicos que viven en el agua, incluyendo bacterias. Algunos peces provistos de muchas efongaciones que !‘orman especies de rastrillos, COMO
Ics “gizzard shads” (Durosorna) se alimentan durante toda su v d a de fitoplanc- ton. Otros peces que tienen dientes cxwtantes, como ios peces loro (Scaridae) y los peces cirujano (Acarcthurus), 1as tilapias de agua duice (Tilapia melartopleura) y la carpa de los pastos acuáticos (Ctenopharyngodon idella): cortan partes de las plantas p x a devorarlas como alimento, rnordisquearldo las hojas, las frondas y !os tallos pequeños. Quienes est,%n meJjor adaptados para este ghnero de aiirnen- tacibn son los p e c c ~ loro que tienen uncx dientecillos parecidos a incisivos, los cuales han llegado a fusionarse pera formar en ei pico un borde cortante. Mien- tras que el ).,arde cortante sirve para separar ptdazos d e alga, en ei interior de 12 Faringe existe un conjurrt-o de dientecillos Inoldores que rompen !as células y dejan que salga el colatenitlo de las mismas para que sufra l a accibn de ios jugos gk,tricos (Figura 5.6e). Los dientecillos tarnbi6n lifgan a moler fragmeilhs de coral.
Una gran diversidad de clases y tamaños de animales es necesario que exista en las cadenas alimenticias de los peces (Figura 5.1). Entre los alimentos animales primordiales consumidos están los planctónicos, mejor conocidos corno zooplanc- ton. El zooplancton incluye muchas especies diferentes de protozoarios, micro- crustiíceos y otros invertebrados nicroscbpicos y los huevos y larvas de muchos animales, incluyendo a los de los propios peces.
Entre los animales invertebrados de mayor tamaiio que sirven de alimento a los peces tienen extraordinaria importancia los anélidos (Annelida), los caracoles y bivalvos (Mollusca) y los crustáceos e insectos (Arthropoda). En lo que respecta a los vertebrados, todos pueden ser presa para los peces: aves, mamíferos, reptiles, anfibios y, por supuesto, los peces mismos. No es extraño encontrar un roedor pequeño, una vibora o una tortuga en el estbmago de un huro (Micropterus), un lucio (Esox), un peje-lagarto (Lepisosteus) o un amiido (Arnia). Algunas veces un patito o un ave canora desafortunados pueden también ser encontrados en uno de los peces anteriores o era un pejesapo (Lophius). Cualquier pescador sabe que las ranas son el alimento natural de los peces predadores de agua duice; por lo tanto, constituyen un cebo magnifico para muchas especies que son de irnportan- cia para la caza deportiva, Los humanos, inclusive, no se escapan, desde el mo- mento en que son atacados por muchos peces, entre ellos la barracuda (Sphyraena), ciertos tiburones y las pirañas (Serrasalrnus) de América del Sur.
Abundancia de alimento
Cualquier organismo que pueda servir de alimento a los peces no siempre esti disponible en forma constante, desde el punto de vista numérico, debido a que

EL ALIMENTO NATURAL DE LOS PECES 127
ALIMENTO DEL ARENQUE
Joven Adulto-
Figura 5.1 " Resumen diagramático de las relaciones alimenticias del arenque (Clupea harengus) de diferentes tamaños. (Fuente: Hardy, 1959).
hay fluctuaciones naturales en su abundancia. Esas fluctuaciones de los organis- mos que sirven de forraje son a menudo cíclicas y se deben a factores propios de su desarrollo biológico, a condiciones climáticas o a otras relacionadas con el medio ambiente. Las migraciones de peces reflejan a menudo la necesidad de la búsqueda de una fuente particularmente abundante de comida. El arenque (Clupea harengus) del Mar del Norte, por ejemplo, sigue a las concentraciones de plancton, y las predadores del arenque, como ciertas macarelas (Scombridae), siguen al pez a su vez cerca del mismo mar. Se ha demostrado repetidamente que el crecimiento y la sobrevivencia de los peces no es más que un reflejo de la abundancia de los organismos que sirven de alimento. La existencia de plancton suficiente, precisa- mente durante la temporada de nacimiento de los peces, es muy importante para lograr la sobrevivencia de la camada de ese año. Las truchas (Salmo; Sulvelinus) de los ríos de Norteamérica realizan la mayor parte del crecimiento correspon- diente a un año en unas cuantas semanas, de las postrimerías de la primavera a las primeros días del verano, que es cuando se presenta una etapa de emergencia para devorar insectos. Esta emergencia tiene lugar a un tiempo cuando los estados acuiticos de muchas clases diferentes de insectos se metamorfosean y abandonan el agua para iniciar la reproducción. Durante este intervalo la presa se concentra, como lo hace siempre en esa temporada del año, y es particularmente vulnerable para la predación.
La abundancia de una especie potencialmente buena como alimento deter- mina a menudo que sea o no devorada por los peces, porque su disponibilidad es un factor clave para determinar lo que un pez debe comer. La mayoría de los peces se adaptan muy bien en sus hábitos alimenticios y aprovechan los alimentos que tengan más a la mano. Relativamente pocas clases de peces son estrictamente herbívoras o carnívoras, y posiblemente ninguna de ellas se alimenta de un sólo

128 ALIMENTO, DIGESTION, NUTRICI6N Y CRECIMIENTO
organismo. Algunos, como el pez ángel de las Bermudas (Holacanthus berrnuden- sis) van aún más lejos, porque pueden cambiar su dieta de acuerdo con la estación, siendo francamente herbívoros en invierno y primavera, y predominantemente carnívoros en el verano y principios de otoño.
Cadenas alimenticias, tramas alimenticias y pirámides alimenticias o de masas (Figura 5.1)
Por las tramas alimenticias, los peces están prácticamente atados a otras formas de vida en su medio ambiente. Si nosotros consideramos las posiciones relativas de los comedores y los comidos, los herbívoros y los carnívoros estarán en diferentes posiciones verticales (niveles tróficos) de una pirámide alimenticia. Usualmente los carnívoros más grandes o predadores tope pueden ser colocados en el vértice de la pirámide. Una pirámide así, representa los números descendentes de estos animales en una comunidad en la misma medida en que su tamaño aumenta; tam- bién puede representar la disminución de la biomasa en los diferentes niveles de la pirámide, estando la menor en la punta (el predador más grande).
Las plantas verdes que aprovechan la energía solar para que sea utilizada más tarde a través del mundo viviente ocupan, desde luego, el primer nivel o base de la pirámide. Viene después un nivel intermedio compuesto por los herbívoros: moluscos, ciertas larvas de insectos, muchos crustáceos y peces. Finalmente se encuentra el nivel trófico más alto ocupado por carnívoros, en donde deberá haber numerosas hileras de peces que devoran a otros peces que a su vez devoran a otros, y así sucesivamente.
Para completar la circularidad de la rueda de la vida, o sea, para cerrar el ciclo alimenticio, las excreciones de los peces vivos y de otros organismos, y sus restos cuando mueren, son atacados por las bacterias. Sus materiales son entonces disueltos en el agua para dar de nuevo a las plantas sustancias primas que devuel- van la energía radiante para formar nuevo protoplasma durante el proceso de la fotosíntesis.
Las interrelaciones alimenticias pueden parecer relativamente simples si observamos solamente a los adultos, digamos un huro (Micropterus) o una mojarra (Lepomis macrochirus); pero cuando es considerado el ciclo biológico completo de un pez, desde que está recién nacido hasta el adulto, pasando por el joven, po- dremos darnos cuenta que las tramas alimenticias son sumamente complejas. Aparecen como una red de interrelaciones de los que comen y los que son comi- dos, en la que quedan involucrados muchos phyla de organismos acuáticos a me- dida que van apareciendo cambios de alimentación paralelos a los cambios del ciclo de vida.
Hay una fase de síntesis en las aguas superficiales, soleadas, de los lagos, los ríos y los océanos, la que contiene eslabones de consumidores y se apropia los ma- teriales de desecho a través de excreta, restos de organismos y sustancias en pu- trefacción que pueden ser usados otra vez; por lo tanto, el ciclo alimenticio es fácilmente observable en esos lugares. Pero, ¿qué sucede en las zonas abisales de los océanos, las que están muy oscuras y profundas?, ¿qué sucede en las profun- didades de los grandes lagos continentales como el Superior y el Baikal? Aquí los organismos, incluyendo a los peces, dependen de organismos que se mueven hacia las profundidadea derde arriba, ya sea por migración o en forma de lluvia de des-

HABITOS ALIMENTICIOS 129
pojos. Y todavía más, en la zona de aguas profundas de las aguas estancadas, la transferencia puede involucrar a una gran serie de capas a través de las cuales los nutrientes son transmitidos por organismos de la zona de producción de las aguas superiores a la puramente consumidora que se encuentra en las aguas profundas. Conectadas de esta manera, debemos pensar en la existencia de una capa superior donde se realiza la síntesis, seguida hacia abajo por otra zona donde viven crustá- ceos y otros invertebrados y peces que se nutren de la vegetación de la parte infe- rior de la zona de síntesis. Debajo de esa zona de los crustáceos pelágicos que se alimentan de la vegetación que aún se produce en la parte inferior de la zona de síntesis, debe haber otra con peces que se alimentan de los crustáceos y que sirve como puente de unión de las zonas tróficas, iluminadas, y la oscura, y así sucesiva- mente continúan las capas de peces y otros animales hasta llegar a las grandes profundidades (Capítulo 14). Es en este último lugar donde toda la materia orgá- nica que se asienta se acumula formando concentraciones de cieno que sirve de alimento a las bacterias de las profundidades marinas, y nuevas tramas alimenticias se vuelven a formar. Yendo hacia abajo, en cada nivel sucesivo (excluyendo el fon- do del mar), la biomasa total se vuelve cada vez más y más pequeña.
HABITOS ALIMENTICIOS
La manera de comer y el estímulo para hacerlo deben ser incluidos en cualquier discusión que haya sobre la alimentación de los peces. Los hábitos alimenticios o la conducta relacionada con la alimentación en los peces no se relaciona más que con la búsqueda y la ingestión de alimentos, es decir, la manera de alimentarse. Éstos deberán distinguirse de los hábitos de alimento y la dieta, que corresponden al estudio de los materiales que habitualmente o fortuitamente llegan a comer.
Tipos importantes de alimentación
Por lo que se refiere a la manera de alimentarse, la más común es sólo una: cuando el alimento es tomado por la boca. Los otros hábitos y adaptaciones para la ali- mentación de los peces, que no sean el de la boca, son muy diversos. Sin embargo, hay algunos tipos de alimentación muy especializados pero también muy exten- didos y se pueden observar ya sea en forma específica o en ciertas etapas de los ciclos biológicos. A partir de esta base, los peces pueden ser de acuerdo a sus hábi- tos de alimentación: predadores, ramoneadores, coladores de alimento, succiona- dores de alimento y parásitos.
Predadores. Los peces que se alimentan de animales macroscópicos tienen algunas adaptaciones en común. Usualmente tienen dientes muy bien desarrollados para asir, como sucede en los tiburones (Elasmobranchii), las barracudas Sphyruenu), los lucios (Esox), o los catanes (Lepisosteus). Los peces predad es tienen un estómago bien definido con secreciones ácidas muy fuertes y el intestino muy corto comparado con el de los herbívoros del mismo tamaño. Muchos predadores, como el voraz pez azul (Pornutcrmus sultutrk), y muchos peces de las profundida- des marinas cazan su presa con mucha actividad mientras que otros, como las cabrillas (Epinephelus), a menudo permanecen quietos esperando que pase cerca
d

130 ALIMENTO, DIGESTION, NUTRICIQN Y CRECIMIENTO
de ellos un animal y luego se lanzan rápidamente para capturarlo. Los peces ante- nados (Lophiidae y Antennariidae) han desarrollado un radio anterior de la primera aleta dorsal de tal manera que, teniendo forma de señuelo, atrae a las presas. El pez arquero (Toxotes jaculator) del sureste de Asia llega hasta a escupir a los in- sectos que están parados sobre las plantas, a la orilla del agua, para derribarlos y devorarlos. %a punteria tan precisa de est8e pez demuestra que tiene un sent,itlo de la vision a h a muy bien desarrollado. Algunos peces predadores cazan ayudán- dose con la vista, mientras que otros, donde quedan incluidos muchos tiburones (SqualiCormes), peces nocturrros (como los bagres, Ictulurras) y ¡as morenas (Mu- raenidae) se atienen mucho al olfato, el gusto y el tacto y probablemente localizan y capturan su. presa ayudándose de los 6rga.nos sensoriales de ia linea lateral.
Ramoneadores. Mediante pequeiios mordiscos los peces ramoneadores toman su alimento vegetal. Algunas veces los peces ramonean solos, pero en otras io hacen en pequeños grupos a semejanza de las reses o los carneros en un pastizal. El ra- m ~ n e o caracteriza a muchos peces gue se alimentan de plancton o de organismos clek fondo. Por ejemplo, tenemos a la mojarrita (Lepomis rnacrochirus) que se dimenla a lo largo del fondc dr lago, tomando de aqui y de alla, a menudo crn form;a individual, ptquefias larvas de díptero. Muchos peces que, cuando add- tos serán predadores de otros peces, se alirnentan dc Los planctbnicos a quienes cazan er, forma individual.
Los peces loro (Scaridae) o 1.0s ‘.wes mariposa (Chaetodontidae), cuando ramonean en un arrecife de coral pueden raspar partículas de coral para ingerir ios pcilipos o morder trozos de las frondas de las algas, como ya se ha indicado. May una clase especial de ramoneo en la cual los peces se xnordisquean partes entre sí. La parte mis inlportante de la alimentacihn de un bagre de la India (Schilbcidae) y de un cíclido africano esta constituida por las escamas que arran- can de otros peces. Y ‘iodavla m&, no es raro ver en las aguas congestionadas de los criaderos de pescado a las truchas (Salmo; Sultielinu.~) mordisqueando partes d.:. las aletas de los dernk ptces.
Coladores. La separticiljn de los organismos del agua que los contiene mediante el colado, es un tipo generalizado de alimentacicin, ya que los materiales alimen- ticios son seh?ccionddos por su tamaiio y no por su clase. La filtracibn del planc- ton y ia deglución de un concentrado parecido a la sopa de chícharo ilustra esta clase de alimentacihn, come !a practican muct-~os peces parecidos al arenque (clupeoicieos), entre los cuales podríamos incluir los “gizzard shads” (Dorosoma), como ya se ha señalado, Las lachas (Brevoortia tyrannus) que se reúnen en gran- des bancos R io largo de Ja costa atlhntica d.2 NortearnGrica, nadan con la boca abierta a travks de l as ~iyuisi.rr.as masas planct,Ónicas. Una Iacha adulta es capaz de filtrar de 5 a 40 litros de agua por minuto a través de los rastrillos de sus bran- quids. Por lo tanto, el pez puede deglutir durante ese corto tiempo varios centí- metros cClhicos de concentrado de plancton, formado principalmente por diato- meas y crustáceos. Otros peces, algunos de gran tamaño como el pez espátula (Polyodon), el marrajo gigailte (Cetorhinusj y el tiburón ballena (Rhincodon), también tienen adaptaciones muy eficientes para ef colado o la filtración del alimento. El desarrollo de numerosos peines formados por branquispinas largas y muy cercanas entre si es la principal caracteristica de !os peces coladores (Fi- gura 5.2).

HÁBITOS ALIMENTICIOS 131
Chupadores. L a introducción a la boca de alimento o materiales que lo contengan, mediante la succión, es a menudo practicada por peces que viven en los fondos para alimentarse en estos lugares, como sucede con los esturiones (Acipenseridae) y las carpas hociconas (Catostomidae) (Figura 5 .3 ) . Hábitos similares tienen los pececillos del Viejo Mundo (como los de labios con orlas, Labeo y Osteochilus), que tienen sus bocas hacia abajo y los labios adaptados para la succión. En unas cuantas especies de carpa (Cyprinidae) del sureste de Asia la respuesta a la succión depende en gran medida al estímulo del tacto en los labios carnosos y sensibles; al grado de que es necesario para el pescador extirpar los labios de estos peces antes de juntarlos en los tanques con otras especies. El peligro consiste en que los demás peces pueden ser lastimados al ser confundidos con las algas o con las piedras cubiertas de musgo, que es donde los pececillos generalmente raspan y chupan para obtener su alimento.
Los peces que succionan en el lodo para extraer los organismos que hay en él, pueden o no obtener un buen bocado de alimento en cada ingestión. Algunos separan las partículas alimenticias entre el sedimento antes de tragarlas, pero otros,
RASTRILLOS BRANQUIALES
Rastrillo
branquiales ( 4
Figura 5.2 - Relaciones de los rastrillos branquiales con los hábitos alimenticios, mostrando el mejoramiento de la eficiencia del potencial de tamizado de izquierda a derecha: (a) pez blanco redondo, Prosopium cylindraceum; (6) pez blanco lacus- tre, Coregonus clupeaformis; (c) "cisco" de aleta negra, Coregonus nigripinnis. (Fuente: Hubbs y Lagler, 1958).
BOCAS CHUPADORAS
Figura 5.3 - Variaciones en el desarrollo de los labios chupadores y la boca en tres carpas hociconas (Moxosroma); (a) carpa hocicona de cabeza corta, M. macrolepi- dorum; (6) carpa hocicona dorada, M. erythrurum; (c) carpa hocicona plateada, M. anisurum. (Fuente: Hubbs y Lagler, 1958).

132 ALIMENTO, DIGESTI6N, NUTRICI6N Y CRECIMIENTO
como algunos bagres orientales (Schilbeidae, Siluridae), acumulan en su tracto digestivo pequeñas porciones del sedimento junto con elevadas concentraciones de organismos del fondo. El hábito alimenticio de estos peces daña indirectamente a la maquinaria empleada en la extracción del petróleo, porque los desperdicios de los peces hacen que la parte metálica se cubra de una emulsión pegajosa for- mada por grasas de pez y barro.
Parásitos. El parasitismo* es posiblemente el hábito alimenticio más raro y espe- cializado que se haya desarrollado entre los animales. Un notable ejemplo en el mundo de los peces es representado por las lampreas parásitas (algunos Petromy- zonidae) y los mixinos (Myxinidae) que chupan los fluidos del cuerpo de los hués- pedes después de haberles raspado la piel. Una especie que sirve de ejemplo es la lamprea marina (Petromyzon murinus; Figura 5.4) que vive hacia el oeste del Atlántico del Norte y en aguas continentales adyacentes. Otro es la lamprea del Pa- cífico (Lumpetru tridentuta) que incluye a las ballenas entre sus huéspedes. Una anguila de los mares profundos (Simenchelys parusiticus) es también parásita. Los machos de algunos pejesapos (Ceratias) de aguas muy profundas son parásitos obligatorios en las hembras de las mismas especies (Figura 10.4). Apenas acabado de nacer el macho busca una hembra y al encontrarla se fija a su cuerpo mediante la boca. La hembra, complaciente, responde desarrollando una papila carnosa por donde el pez macho puede absorber nutrientes, desde el momento en que como consecuencia de la fijación no toma ningún alimento de vida libre. El macho per- manece relativamente pequeño, un poco más que una gónada animada. La absor- ción intrauterina que se realiza durante la gestación (y el intercambio respiratorio), por los embriones de algunos peces vivíparos, representa una especialización nutricional todavía mayor, si la comparamos con el caso del parasitismo extremo que citamos anteriormente.
Adaptaciones para la alimentación
La diversidad de los hábitos alimenticios que los peces exhiben es el resultado de la evolución que conduce a las adaptaciones estructurales que sirven para ob- tener el alimento, a partir de las muy diversas situaciones que se han desenvuelto en el medio ambiente.
Labios. La aparición de verdaderas mandíbulas para bordear la abertura de la boca fue un avance significativo de la evolución de los vertebrados. La boca equipada con mandíbulas tiene más bien una función mordedora y los peces que degluten grandes bocados de alimento usualmente tienen labios relativamente delgados y sin modificar. Los peces que se alimentan mediante la succión tienen una boca ventral y modificaciones carnosas en los labios (Figuras 5.3 y 5.5d). Entre éstos son notables los esturiones (Acipenseridae) de la zona templada del Norte y las carpas hociconas (Catostomidae; Figura 5.3) de América del Norte y Asia. Los labios de los esturiones y las carpas hociconas son movibles y pueden ser descritos
*Los significados de las palabras parásito y predador no connotan con precisión los há- bitos sarcofágicos de las lampreas o mixinos “parásitos”. En consecuencia, empleamos el término parásito en sentido amplio (N. de los As.).

HABITOS ALIMENTICIOS
?!
m
L i .-
4 w
.- a
2 5
a z a
133
9 C =I a) U

Modificaciones en la forma de la boca (Figura 5.5). Entre los peces ramoneadores y los que se alimentan mediante la succión no hay solamente labios especialmente desarrollados, sino también adaptaciones de otras partes de la boca. Los peces trompeta (Aulostomidae), los peces corneta (Fistulariidae), los peces pipa (Syng- nathidae) y muchos peces mariposa (Chaetodontidae) de los arrecifes de coral, tienen bocas que recuerdan picos largos. Esta adaptación es adquirida mediante la protracción del hueso hiomandibular más bien que por el alargamiento de los huesos de la mandíbula inferior (los dentarios).
Entre los peces el método seguido para la alimentación puede ser realizado por succión como si se hiciese con una jeringa, como en el caso de los peces trom- peta, los peces corneta y los peces pipa; o puede efectuarse mediante el ramoneo selectivo para lo cual se hace uso de filosos dientes en donde la trompa larga del pez mariposa le permite a éste buscar en las cavidades pequeñas de los corales.
El alargamiento de las mandíbulas debido a una articulación especial de los premaxilares y otros huesos de la cabeza permite que algunos predadores, como los "dory" (Zeidae), algunos peces gallo (Labridae) y el ciprínido europeo (Abru- mis brama, Cyprinidae), formen una especie de tubo temporal con el que engolfan su presa desde corta distancia. En Chuuliodus, el pez víbora de las grandes profun- didades, se produce un aumento de la abertura bucal debido a otras adaptaciones, también de los huesos de la cabeza y sus articulaciones.

HABITOS ALIMENTICIOS 135
BOCAS
Figura 5.5 - Algunas adaptaciones estructurales de la boca entre los peces: (a) pez elefante, Gnathonemas elephas; (b ) pez aguja, Strongylura longirostris; (c) pajarito, Hyporhamphus unifasciatus; (d) mojarra de labios gruesos, Cichla- soma lobochilus; (e ) astrónomo, Zalescopus tosae; ( f ) pez mariposa de nariz larga, Forcipiger longirostris; (g) pez espátula, Polyodon spathula. (Fuente: Nor- man, 1931).
En los peces pajarito existe una estructura peculiar que modifica a la boca, y se debe a la proyección hacia adelante de la mandíbula inferior, hasta una dis- tancia de un tercio de la longitud misma del pez, de tal manera que se forma una especie de pico pero con la boca hacia arriba. Los peces pajarito (Hemiramphidae) son generalmente peces que se alimentan en la superficie y se ha pensado que el “pico”, además de servir para la captura del alimento, sirve de timón y para man- tener el equilibrio, si su función se asocia con la de una aleta dorsal insertada en el extremo posterior.
Dientes (Figura 5.6). Los dientes son estructuras de lo más notable que se han formado en los peces como adaptaciones para la alimentación. Se piensa que se

136 ALIMENTO, DIGESTION, NUTRICION Y CRECIMIENTO
originaron de las escamas que cubren los labios, como puede observarse que sucede en los tiburones vivientes (Squaliformes), en los que sin mucho pensarlo se ve cómo las escamas placoideas de la piel se van transformando hasta volverse dientes en las mandíbulas. En los peces óseos (Osteichthyes) los dientes son de tres cla- ses, si tomamos en cuenta el lugar donde se encuentran: mandibulares, bucales y faríngeos. Los dientes mandibulares son variados y se les encuentra en los huesos maxilar y premaxilar, arriba, y en los dentarios, abajo. Los dientes nacen de varias maneras por el vómer medio, por el palatino y los huesos ectopterigoideos en cada lado, en el techo de la cavidad oral. La lengua, ubicada en el piso de la cavidad bucal, tiene a menudo dientes. En varios elementos de los arcos branquiales de muchas especies se encuentran los dientes faríngicos en forma de cojincillos. Los únicos dientes que se encuentran en las carpas (Cyprinidae) y las carpas hociconas (Catostomidae) están muy adentro de la faringe, y tiene su origen en las modifi- caciones de los elementos inferiores del último arco branquial. Las modificaciones parecidas a dientes de los peines o rastrillos branquiales y los huesos dérmicos (posiblemente escamas) no son adornos suplementarios extraños ubicados en las superficies internas de los arcos faríngeos de muchos peces predadores como el lucio (Esox lucius).
De acuerdo con su forma, podemos considerar las siguientes clases de dien- tes mandibulares: cardiformes, viliformes, caninos, incisivos y molariformes. Los dientes cardiformes son numerosos, cortos, finos y puntiagudos. Un conjunto de ellos en un hueso recuerda a la herramienta con multitud de púas que llegó a emplearse hace muchos años, cuando se cardaba la lana y se fabricaba hilo en la rueca; de ahí su nombre cardiforme. Esa clase de dentición, con algunas variacio- nes, se encuentra en muchísimos peces que tienen los dientes dispuestos en varias hileras; podemos citar como ejemplos a los bagres de Norteamérica (Icta- luridae), las percas (Percidae) y muchos serranos (Serranidae). Los dientes vili- formes son realmente cardiformes más o menos alargados, en donde la relación longitud-diámetro recuerda la que tienen las vellosidades intestinales (como su- cede en el pez aguja, Belone, y en los peces león, Pterois). Los dientes caninos se parecen a los de los perros e, inclusive, exageran ese parecido; es decir, son más colmillos que los de los perros. Son alargados y subcónicos, derechos o cur- vos y están adaptados para clavarse y sujetar; los “walleyes” (Stizostedion) po- seen, entre muchos otros peces, esta clase de dientes. En ciertos peces como las morenas (Muraenidae) y el predador pelágico marino, el llamado “handsawfish” (Alepisaurus), los caninos están articulados como en bisagra, ceden a una presión dirigida hacia atrás, pero cuando son empujados hacia la abertura bucal se ende- rezan bruscamente y adquieren una posición rígida; obviamente es una adaptación para retener viva a la presa que se está moviendo. Incisivos son los dientes que tienen los extremos cortados en bisel. En algunos peces como el sargo (Archosur- gus) parecen casi humanos; en otros tienen los bordes serrados, y todavía en otros más se han fusionado para formar bordes cortantes como en los peces loro (Sca- ridae). Los dientes molariformes sirven para machacar y moler y, en consecuencia, son algo aplanados, a menudo con amplias superficies rellenas; son generalmente característicos en las rayas moradoras de los fondos marinos (queda incluida Ruja), las quimeras (Holocephaii) y muchos tambores o corvinas (Sciaenidae).
Hay una gran correlación entre el tipo de denticibn, los hábitos alimenti- cios y el alimento consumido. Los peces predadores como el lucio (Esox), los catanes (Lepisosteus) y los sacofaringoideos (Saccopharyngoidei) tienen dientes

HABITOS ALIMENTICIOS 137
DIENTES DE PECES dSEOS
ientes cardiformes
, Die can
(a) Bagre (b) Pez víbora de mares profundos
molariformes
(d ) Raya águila
Hipohi Músculo
Dientes
Cavidad bucal
raspar, desgarrar y moler (e) Pez loro (g) "Minnows"
Figura 5.6 - Algunas variaciones de los dientes en los peces, por su estructura y ubicación: (a) mandíbula superior del pez gato de cabeza aplanada, Pylodictis olivaris (fuente: Hubbs y Lagler, 1958); (b) pez víbora de mar profundo, Chaw liodus sloani (basado en Goode and Bean, 1895); (c) mandíbula inferior de un pez pluma, Sparidae, ( d ) dientes pavimentarios en el techo y piso de la boca de una raya águila, Myliobatis; (e) sección sagita1 de un pez loro (Scaridae) (basado sobre Monod en Grassh, 1957) para mostrar el "pico" por la fusión de los dien- tes maxilares y para mostrar los dientes faríngicos; (9 dientes faríngicos de una carpa hocicona dorada, Moxostorna erythrururn (basado en Hubbs y Lagler, 1958); (S) tipos de dientes faríngicos de "minnow", de izquierda a derecha, mostrando dientes en punta para prehender, de un pez barba (Barbus); dientes almenados, propios para desgarrar, en un "rudd" (Scardinius), y dientes molari- formes propios para remover, en una tenca (Tinca) (basado en Grassé, 1958).

138 ALIMENTO, DIGEST16N, NUTRICIdN Y CRECIMIENTO
puntiagudos que aparentemente les son de mucha utilidad para raspar, punzar y detener a la presa. Los peces que se alimentan de plancton y son raspadores del perifiton que se incrusta en partes sólidas acuáticas tienen como caracteris- tica mandíbulas sin dientes, aunque debido a una adaptación especial la mandí- bula puede tener un borde cortante, como sucede en el pececillo rodador de piedras (Campostoma unomulum) que tiene en los dentarios sendos bordes prin- cipales, de cartílago endurecido, que es empleado para raspar superficies en busca de alimento. En las rayas (Rajidae) y en los tambores o corvinas (Scianidae) hay dientes moledores (molariformes) en las cavidades oral y faríngea en asociación con un tipo de dieta que incluye caracoles, almejas y crustáceos de caparazón duro. En peces predadores como la formidable piraña (Serrusulrnus) del río Ama- zonas y la barracuda (Sphyraena) de los mares cálidos, se han desarrollado dientes parecidos a hojas de rasurar por su enorme filo. Algunos consumidores de vegeta- les, como sucede en muchos peces loro (Scaridae), hacen una verdadera ensalada picando su alimento con filosas herramientas cortadoras que resultan de la fusión de dientes individuales para formar bordes filosos en las mandíbulas. Los cortes iniciales son seguidos por una trituración fina que se realiza por medio de las pla- cas trituradoras de la faringe.
Dentro de un solo grupo, la dentición se puede presentar con tanta diversi- dad en huesos idénticos que puede abarcar en principio la gama citada anterior- mente. En las carpas y demás pececillos (Cyprinidae), los dientes faríngeos com- prenden desde los filosos en los carnívoros, como en el “chub” de los arroyos (Sernotilus), a los molariformes en la carpa común (Cyprinus curpio) y casi han desaparecido en algunas especies. En éstos y en muchos otros peces los dientes tienen importancia para la clasificación. Muchas especies de tiburones fósiles han sido descritas a partir únicamente de los dientes.
Por lo general, los dientes están ausentes en los consumidores de plancton y en algunos de los omnívoros más generalizados, pero el número de piezas va siempre en aumento a medida que los peces se van volviendo más y más predadores. Los huesos premaxilares son dentados en la mayoría de los peces que tienen algún diente mandibular por lo menos. Esto que se dice es cierto en muchas especies de radios blandos como Arniu, los catanes o peje-lagartos (Lepisosteidae), los salmones y truchas (Salmoninae) y en los peces dentados de radios espinosos, en general (por ejemplo, el numeroso orden de los Perciformes).
Las maxilas son típicamente dentadas en los peces de radios blandos que presentan dientes en los premaxilares, Sin embargo, es característica de las maxilas carecer de dientes en los peces que llevan dientes y radios espinosos en las aletas. Siguiendo el proceso de la evolución de peces con radios blandos a los que tienen radios rígidos, vemos que las maxilas son huesos dentados solamente en algunas clases de peces de radios blandos. La maxila no es dentada en los peces con radios espinosos y no aparece en la abertura bucal, porque ya no forma parte del mar- gen de la boca.
Branquispinas. Además de proteger los filamentos branquiales tiernos contra la abrasión que producen los materiales de textura tosca que son ingeridos, las bran- yuispinas también están especializadas en todo lo relacionado con los alimentos y los hábitos alimenticios de los peces, como ya se dijo anteriormente. Están muy espaciadas y sin adornos en los omnívoros como la mojarrita verde (Lepomis cya- nellus) y la mojarrita semilla de calabaza (L. gibbosus). Muchos comedores de

HABITOS ALIMENTICIOS 139
plancton tienen las branquispinas alargadas, son numerosas y lameladas u orna- mentadas de una manera variable, posiblemente para aumentar la eficiencia de la filtración. Dorosorna y el pez espátula (Polyodon) las tienen muy numerosas pero muy simples. Las de estructuras ornamentadas, sin embargo, se encuentran en los lenguados (Pleuronectidae) y son de utilidad taxonómica. En este caso, cada branquispina recuerda una pluma por tener un axis con alargamientos laterales (y aun ramificaciones en cada elemento lateral). Una coladera o cedazo más fino se forma cuando los alargamientos de branquispinas vecinas se entrecruzan; pue- den lograrse, entonces, colados más limpios.
El tubo digestivo. La gran dilatabilidad del esófago es otra adaptación que tienen los peces para su alimentación. Nunca ha llegado a suceder que un individuo se ahogue hasta morir porque no pueda deglutir algo que haya tomado con su boca. Sólo ocasionalmente ha llegado a pasar que un pez predador se haya encontrado en zozobra mortal porque una presa se le alojó en la garganta. Quienes frecuente- mente dañan a sus predadores de esta manera son los bagres (Ictaluridae) y la espinocha (Gasterosteus). Sus radios espinosos de la aleta, cuando están erectos, hacen que las presas se atoren. Pero, si están adosados al cuerpo de la presa, ésta resbalará fácilmente a lo largo del esófago, lo que es muy frecuente. El esófago es tan dilatable que puede acomodar en su interior cualquier cosa que el pez haya tomado con la boca e inclusive puede acomodarla en el caso de que al ser capturada haya sufrido dos o tres dobleces sobre sí misma en su recorrido hacia el estómago.
Una de las adaptaciones tan variadas que puede mostrar el estómago se refiere a la forma (Figura 5.7). En los peces ictiófagos el estómago es típicamente alargado, como en los peje-lagartos (Lepisosteus), Arniu, el lucio (Esox), las barra- cudas (Sphyraena) y la lobina rayada (Morone suxatilis). Las especies omnívoras tienen el estómago en forma de saco, similar al de los humanos. Los esturiones (Acipenser), las sardinitas (Dorosoma) y las lisas (Mugil) (Figura 5.7) tienen trans- formado el estómago en un órgano moledor, como una adaptación muy especial. En este caso el estómago es de tamaño reducido, pero sus paredes son muy gruesas y musculosas. El tejido interior también está muy reforzado con tejido conectivo y la luz, o sea, el espacio que queda dentro, está muy reducido. El órgano está muy lejos de ser un simple depósito de alimento donde las paredes musculares lo re- vuelven y se realiza la digestión primaria; es más bien un órgano moledor, como la molleja de las gallinas y otras aves de corral. En los tragadores predadores de las profundidades marinas (Saccopharyngidae) y los “gulpers” (Eurypharyngidae) la adaptación del estómago que les permite a estos peces tomar presas de gran tamaño, es esta notable dilatación.
Una modificación muy notable del estómago puede ser observada en el pez tambor (Tetraodontidae) y en el pez erizo (Diodontidae), que pueden inflarlo con agua o aire hasta tomar la forma de un globo. El estómago mismo o bien una eva- ginación de su porción anterior, se llenan mediante la acción de los esfinteres cardíaco y pilórico y por otro esfínter, durante la evaginación misma. Cuando están inflados, esta evaginación del saco gástrico está tan distendida que sus pare- des son tan delgadas como el papel. Cuando el saco se vacía actúan de nuevo los esfinteres, auxiliados esta vez por los rn.úsculos abdominales. Se puede observar que la presencia de un estómago dilatable en estas familias va de la mano con la ausencia de aletas pélvicas; también faltan los huesos pélvicos. Probablemente

140 ALIMENTO, DIGESTION, NUTRICIdN Y CRECIMIENTO
EST~MAGOS
(u) Pez gato
Capa dentada Abertura hacia el conducto neumático Píloro
Conducto biliar Intestino 1’
( b ) Lucio
Cieoo ,/r Esdfago
(c) Mújol
Figura 5.7 - Variaciones en la forma, aphdices y covertura de la porción anterior del tracto digestivo en tres peces: (a) un bagre omnívoro, el siluro europeo (Silurusglanis); (b) un pez carnívoro, el lucio del norte (€sox lucius); (c) un pez ramoneador de los fondos, el mugílido rayado (Mugil cephalus). (Basado en Pernkopf en Boik, Goppert, Kallius, y Lubosch, 1937).

HABITOS ALIMENTICIOS 141
el principal valor adaptativo de esta modificación del tracto digestivo tenga que ver con la defensa ante los predadores, ya que muchos peces tambor y erizo tienen el cuerpo cubierto de espinas que pueden ponerse erectas. Es necesario mencionar que el saco gástrico de los peces tambor, erizo y especies afines, ni tienen relación ni se derivan de la vejiga natatoria.
Esa porción del tubo digestivo con una secreción típicamente ácida y una cubierta interior epitelial diferente a la del intestino, es el estómago; y no todos los peces lo tienen. En un pez devorador de plantas (Rutilus rutilus), ciprínido del Viejo Mundo, por ejemplo, el tejido epitelial del esófago se transforma dircc- tamente en el intestinal. En otros ramoneadores como los peces loro (Scaridae) tam- bién se puede encontrar la misma condición. Algunos carnívoros también han perdido el estómago, como es el caso de los peces “saury” (Scornberesox), y como sucede también en ciertos peces devoradores de plancton de la familia de los peces pipa-caballitos de mar (Syngnathidae). El criterio primario para considerar a estos peces sin estómago no es el que sean hervíboros o carnívoros sino la presencia de órganos accesorios para la trituración y molienda de los alimentos, ya sea en forma de dientes o de una bolsa musculosa parecida a una molleja. Cuando el estómago existe como es el caso notable de los carnívoros, tiene la característica principal de mostrar un pH bajo y la presencia de jugos digestivos, principalmente pepsina.
El intestino también tiene muchas variaciones (Figura 5.8). En los peces exclusivamente carnívoros, como el lucio (Esox Zucius; Figura 5.8c), es acortado, posiblemente porque es más fácil que los alimentos formados por tejido animal se digieran, a diferencia de lo que ocurre con los vegetales. Por lo contrario, es muy alargado y con varios dobleces en los peces exclusivamente herbívoros, como sucede en ciertos bagres loricáridos (Figura 5.8a). Los tiburones y especies afines (Chondrichthyes), y algunos otros peces, han adquirido una válvula espiral (Fi- gura 5.8b) o un doblez largo formado por tejido absorbente y enrollado con cierta soltura, en lugar de los dobleces ordinarios del intestino; mediante esta innova- ción anatómica el intestino se ha vuelto más compacto, ha mejorado la eficiencia de la digestión y la absorción, o sólo una de esas funciones. Parece ser que en los peces que cesan de alimentarse, porque así lo exige su maduración sexual y la llegada de las actividades reproductoras, el intestino se digiere a sí mismo (auto- lisis) hasta desaparecer. El tracto digestivo que alguna vez fue funcional en una lamprea adulta marina (Petromyzon marinus), se transforma en un simple fila- mento sin lumen por la época en que la puesta de huevos ha terminado y la muerte se aproxima. Una reducción similar también aparece en los salmones migratorios del Pacífico (Oncorhynchus) cuando han llegado a los arroyos de agua dulce a desovar.
Los estímulos para la alimentación
Cualquier consideracih que se haga en relación con los estímulos indispensables para la alimentación de los peces tiene interés tanto para los ictiólogos como para los pescadores. Sin embargo, el mecanismo de la conducta de la alimentación es muy complicado, como veremos más adelante. Los estímulos para la alimentación son de dos clases: a) factores que afectan la motivación interna o conducen hacia la alimentación, entre los cuales se encuentran la estación del año, la hora, la in- tensidad luminosa, el tiempo transcurrido desde la ingestión del último alimento

.4LTMENTO. I)IGESTI6N, NUTRICI6N Y CRECIMIENTO
c 2 o c W U
O

HABITOS ALIMENTICIOS 143
y la naturaleza del mismo, la temperatura y cualquier ritmo interno que pueda existir, y b ) estímulo para el acto de la alimentación percibido por los senti- dos como el olfato, el gusto, la vista y los que percibe la línea lateral, que li-, bera y controla el acto momentáneo de 1.a alimentación. La interacción de estos dos grupos de factores determina cuándo y cómo debe comer un pez y lo que debe comer.
Un factor importante que influye en la alimentación es la hora del día. Algunos peces, como los bagres (Zctalurus), que encuentran su alimento mediante el olfato y el gusto, son predominantemente comedores nocturnos. Y todavía otros, como el lucio (Esox) y otros predadores que se alimentan principalmente ayudándose con la vista, son más activos durante las horas diurnas. Las estaciones del año, que influyen con gran fuerza en la temperatura del agua en las áreas tem- pladas y frías, y los niveles del agua en la región tropical, parece que también tienen algo que ver con la alimentación de los peces. Algunos peces, como los salmones (Salmo, Oncorhynchus) y las lampreas (Petromyzonidae), dejan todos de comer durante su temporada de puesta de huevos o desove. Otros, como las anguilas (Symbranchidae), de los pantanos del sudeste de Asia, mientras pasan el estío en concavidades húmedas en el lodo, viven durante semanas a expensas de la grasa que han llegado a acumular en su cuerpo. La mayoría de los peces de las zonas templadas se alimentan activamente mientras las condiciones del medio ambiente cambian durante la primavera, casi al comenzar una nueva estación de crecimiento. El acervo de conocimientos sobre los peces está repleto de afirma- ciones contradictorias en relación a las condiciones climáticas o los estímulos que determinan que el pez muerda. Sin embargo, lo más que se puede decir con toda seguridad y honestidad es que aún queda mucho por descubrir en relación a la actividad de la alimentación regulada por los factores climáticos.
Al considerar que los peces pueden oler y gustar los alimentos, muchos investigadores han ensayado los factores químicos como determinantes de la alimentación. Para muchas especies, los sentidos que perciben a las sustancias químicas (el gusto y el olfato) son verdaderamente los que dan la primera señal para realizar el acto de la alimentación. Los cazones de espina (Squalus) y algu- nos de los tiburones devoradores de hombres (Carcharodon) se alimentan ayudán- dose de una manera muy importante mediante la atracción química, y se ha visto que las morenas manchadas (Gymnothorax) buscan y seleccionan su alimento mediante el olfato. Los bagres (Ictaluridae) y los salmonetes (Mullidae) se alimen- tan principalmente guiados por el sentido del gusto y también el tacto.
Los estímulos visuales impulsan la acción de la alimentación en los peces a través del movimiento, el color y la forma de los objetos percibidos, jugando cada uno su papel como disparadores del acto fisiológico. Cuando un individuo encuentra alimento, despierta entre los demás peces que forman el grupo o ban- co, ya sean de la misma especie o de varias asociadas en un mismo medio eco- lógico y en ayunas, el apetito devorador habitual en estos animales. Es realmente un hecho, según se ha sugerido, que el valor de adaptación que tiene entre los peces el instinto de asociación en grupos, es el de facilitar la alimentación común. Los estímulos de la audición, la olfación y la visión pueden asociarse experimen- talmente en los peces de acuario debidamente entrenados. Por otro lado, los peces pueden también aprender a evadir algunos alimentos si es que no son atrac- tivos por su textura, sabor o cualquier otra cualidad. Un lucio (Eson), por ejem- plo, aprende muy pronto a evitar a la espinocha (Gasterosteus), a pesar de que en

144 ALIMENTO, DIGESTION, NUTRICIdN Y CRECIMIENTO
forma innata el primero de los mencionados tiene fija la respuesta ante el estímulo dimensión, movimiento y color de la espinocha, para devorarla.
Los cambios rápidos de la intensidad luminosa son también importantes para la alimentación. La perca amarilla (Perca flauescens) y muchos otros peces presentan ritmos circádicos y movimiento con brincos durante el ocaso y en la noche, mientras están comiendo. Hay muchas modificaciones sutiles en el medio ambiente debido a los cambios que se suceden entre día y noche y a las condicio- nes que resultan por las mareas; quedan incluidos no solamente la luz, también la temperatura, la salinidad, el pH y la corriente. Las variaciones de cualquiera de estos factores por sí mismos o en combinación con los demás es desde luego notada por los peces y puede influir en la manera de alimentarse. En los peces pueden existir mecanismos innatos para controlar el tiempo, llamados relojes fisiológicos, y deben ser independientes o no relacionados con los estímulos del medio ambiente. En la naturaleza, sin embargo, existen factores como la luz diur- na, el pH, la temperatura y otros que raramente faltan y generalmente determinan niveles de actividad.
La sensación que resulta de tocar algo y percibir su textura es uno de los medios que inducen al pez a tomar sus alimentos. Por ejemplo, en los acuarios, cuando se les ofrece caracoles a la mojarrita (Lepomis rnacrochirus) y a la moja- rrita semilla de calabaza (L. gibbosus), solamente la segunda tritura las conchas, elimina los fragmentos a través de las aberturas branquiales y deglute las porciones blandas. Sin embargo, si únicamente las partes blandas de los caracoles se le ofre- cen a la primera, las comerá. Estas observaciones nos dan muy buena evidencia sobre la importancia de la textura de las presas en la alimentación y el sentido del tacto en la boca. Hay que hacer notar, sin embargo, que aunque en las dos moja- rritas los dientes mandibulares son casi idénticos, no sucede lo mismo con los dientes faríngeos, ya que la llamada “semilla de calabaza” los tiene truncados y, por lo tanto, adaptados para triturar.
Además de los factores ya mencionados hay todavía otros que sin duda alguna conducen a los peces a la alimentación. Estos incluyen el hambre, la gloto- nería, la curiosidad y otros menos definidos. Entre las espinochas puede obser- varse que, después de un largo ayuno forzado, la primera comida que le sigue es más rápida en muchos aspectos, sugiriendo esto que hay un incremento en la motivación. Puede ser que el hambre y la saciedad sean controlados por el hipo- tálamo. Por lo que respecta a la glotonería, que consiste en la inclinación para comer mucho más de lo que el cuerpo normalmente puede almacenar, ha sido descrita en los peces Amia de los acuarios. En el curso de algunos experimentos sobre la utilización del alimento, pequeños serránidos (Epinephelus guttatus) se alimentaron por sí mismos, ingiriendo mucho más alimento del que realmente necesitaban para lograr su óptimo crecimiento. En los casos de glotonería se ha observado que son arrojados con el excremento materiales no totalmente dige- ridos y en mayor cantidad que lo normal. Este acto fisiológico puede ser, por supuesto, acelerado, debido a la escasez de algún nutriente en el alimento.
Las variaciones en la alimentación y en el metabolismo pueden ser obser- vadas en las fluctuaciones cíclicas de los porcentajes de crecimiento anual, aún bajo condiciones ambientales constantes, en la trucha café experimental (Salmo trutta). El crecimiento tiene un máximo en primavera, cierta rapidez en verano, al- guna lentitud o rapidez casi nula en otoño e invierno. En esta trucha hay otro aspecto relacionado con los cambios en el metabolismo, aparentemente determi-

DIGESTIdN. ABSORCI6N Y UTILIZ.ACI6N DE LOS ALIMENTOS 145
nados por factores internos, y se refiere a la ocurrencia de períodos de crecimiento rápido en longitud que se alternan con periodos de rápido crecimiento en peso. Estos periodos duran de 4 a 6 semanas, pero todavía no se sabe a qué se deben estas fluctuaciones.
Hay una relación entre la motivación interna que induce a la alimentación del pez y el grado de selectividad que controla el acto apropiaao para alimentarse como, por ejemplo, la captura de la presa; y esa relación induce a la acción de todos los estímulos para realizar la alimentación. Por ejemplo, cuanto menos activo sea el pez deben ser más numerosos los estímulos emanados de la misma presa, entre los cuales deben quedar comprendidos los que resultan de la forma, dimensiones, color y movimiento, y que deben corresponder a los de una presa idea- lizada por la experiencia, por una concepción innata, o por ambas.
DIGESTION, ABSORCI6N Y UTILIZACION DE LOS ALIMENTOS
La función básica del aparato digestivo consiste en disolver los alimentos hacién- dolos solubles para poder ser más tarde absorbidos y utilizados en los procesos metabólicos del pez. La función también sirve para eliminar las propiedades tóxi- cas peligrosas de ciertas sustancias alimenticias.
Movimiento del alimento en el tracto
El desplazamiento de los materiales alimenticios y sus restos sin degerir, a lo largo del tracto digestivo, es acompañado como sucede en el hombre y otros animales vertebrados, por ondas peristálticas producidas por contracciones musculares. El movimiento en la parte anterior del tracto digestivo es, por lo menos, voluntario, debido a la presencia de músculos esqueléticos en las paredes del mismo. Los movimientos en el resto del tracto son involuntarios debido a la presencia de músculos lisos. Por lo que respecta a la lengua, en contraste con la de los vertebra- dos superiores, no es móvil por s í misma, no tiene componentes musculares esque- léticos y, en consecuencia, se mueve solamente cuando el pez mueve sus piezas esqueléticas bucales inferiores. El recorrido del bolo alimenticio a lo largo del tracto digestivo de los peces es semejante al de los vertebrados superiores, como ya se ha explicado anteriormente.
Muchos peces predadores regurgitan grandes cantidades de alimento, va- ciando su estómago con suma facilidad. Se ha sugerido que esto es posible porque en el tracto digestivo, desde el esófago hasta el estómago, hay míisculos estriados muy bien desarrolados.
Superficies intestinales
Existe una membrana mucosa y blanda que cubre la cavidad bucal, el esófago y el estómago, así como el resto del tracto digestivo. No hay glándulas salivales (a excepción de las que existen en algunos peces especializados, como las lam- preas parásitas de la familia Petromyzonidae); sin embargo, hay glándulas que secretan moco en abundancia, protegen con este material la superficie interna del

Glándulas y enzimas digestivas
Ya se h a hecho mención, en forma general, a l a amplia distribución que tienen las glrindulas mucosas, tanto en el interior como en el exterior del pez. Además, por lo menos en el estómago de los peces predadores, aparecen las glándulas gástricas. És- tas secretan ácido clorhídrico y pepsinógeno, sustancias químicas que en combina- ción son efectivas para desdoblar las enormes moléculas proteínicas. En los peces típicamente carnívoros, como en el lucio neártico (Esox Eucius), se ha determi- nado la acidez gástrica que oscila entre pH 2.4 a 3.6. En consecuencia, hay en el estómago un sustrato apropiado para esas enzimas o peptidasas desdobladoras de las proteínas, cuya presencia en el estómago de muchos peces ha sido ya de- mostrada. No se ha establecido claramente que haya otras enzimas, además de las peptidasas, en el estómago de los peces. Algunos pececillos (Cyprinidae) carecen de glándulas gástricas y, por lo tanto, no tienen un estómago verdadero. Otro tanto ocurre con la molleja (Figura 5.7c), que no tiene glándulas digestivas.
Cuando está presente el ciego pilórico (Figura 5.8) puede tener una fun- ción digestiva o una absorbente, o las dos. En realidad hay varios ciegos pilóricos (Figura 5.8) como bolsas en forma de dedo y aparecen como extensiones del intestino al nivel del píloro; en los ciegos de la trucha (Salmoninae) se ha encon- trado la enzima lactasa. En los ciegos pilóricos y en el intestino de la carpa y la mojarrita, comparados con los del lucio, se ha detectado una gran actividad de la sacarasa (invertasa). Los primeros ingieren una considerable cantidad de ma- teria vegetal, mientras que el lucio prácticamente no la consume. Los ciegos piló- ricos y la mucosa intestinal son fuentes de la lipasa que desdobla las grasas en ácidos grasos y glicerina. La absorción de las grasas puede ocurrir en la porción anterior del intestino. Los copépodos que sirven de alimento a las anchovetas, las sardinas y los arenques contienen más del 50% de cera. Los ciegos pilóricos de estos peces producen una lipasa activada por la bilis para la digestión de la cera, o sea, cataliza el metabolismo de esta sustancia. La morfología del ciego pilórico, ya sea que se presente como un saco Único o dividido (bífido) y ramificado, así como el número de las ramificaciones, son características de utilidad para la clasi- ficación de algunos peces.
Hay algunas glándulas discretas o difusas que están asociadas con el tracto digestivo. Son de función endocrina la tiroides, el timo y la últimobranquial, pero se han originado del tracto digestivo durante su desarrollo embrionario (Capí- tulos 10 y 11) y no tienen relación directa con la digestión.
El hígado, como sucede en los demás vertebrados, se origina en el embrión

DIGESTIdN. ABSORCIdN Y UTILIZACIdN DE LOS ALIMENTOS 147
como una evaginación del intestino durante el desarrollo de éste. La porción anterior da lugar al hígado propiamente dicho, mientras que la posterior origina la vesícula biliar y sus conductos, Algunos peces holósteos y otros teleósteos tienen más de dos lóbulos hepáticos, pero solamente dos conductos hepáticos que comunican el hígado con el conducto cístico que, a su vez, va a terminar en la vesícula biliar. La vesícula biliar es un reservorio temporal de las secrecio- nes del hígado. El conducto biliar común, que vierte en la primera porción del intestino, cerca de la región pilórica, no es más que la unión de los conductos cístico y hepático.
Algunos peces de las grandes profundidades marinas tienen solamente una vesícula biliar vestigial mientras que otros, como el “burbot” (Lotu lotu), no la tienen. El hígado ajusta su forma a la de la cavidad del cuerpo y consta de lóbu- los formados por glándulas tubulares en donde la sangre baña hileras de células, por un lado, mientras que por el otro la bilis secretada vierte en los canales bilia- res. La bilis contiene las sales biliares que emulsifican las grasas, y los pigmentos biliales biliverdina y bilirrubina, que se originan de los eritrocitos y de la hemo- globina, al ser desdoblados sus componentes. Las sales biliares no solamente ayudan a hidrolizar las grasas; también ajustan la alcalinidad de los jugos diges- tivos del intestino para que la acción de las enzimas sea más efectiva.
Además del papel que juega en el proceso de la digestión, el hígado tam- bién sirve como un órgano de almacenamiento de grasas y carbohidratos (glucó- geno). Más tarde desempeña importantes funciones en la destrucción de las células de la sangre y la bioquímica sanguínea, y en otras funciones metabólicas como la producción de urea y compuestos relacionados con la excreción del nitrógeno (Capítulo 9). Los peces pueden ser divididos en dos tipos de acuerdo a su capaci- dad para almacenar la grasa en el hígado. En los peces planos (Pleuronectiformes) y los bacalaos (Gadidae), por ejemplo, la grasa es almacenada principalmente en el hígado. Los atunes (Scombridae) y los arenques (Clupeidae) almacenan en sus músculos grandes cantidades de grasa. Los tiburones y peces óseos difieren mu- cho por las cantidades relativas de ácidos grasos no saturados que contienen los aceites de sus hígados. En algunos tiburones el peso de su hígado llega a alcanzar el 20% del de su cuerpo. No solamente almacenan grasa los hígados de los peces; también conservan vitaminas A y D. Es, por ejemplo, tan elevado el contenido de estas vitaminas en los atunes que el consumo persistente de sus hígados puede acarrear disturbios tan severos en el proceso metabólico humano como los corres- pondientes a una hipervitaminosis.
El páncreas está formado, en realidad, por dos órganos en lugar de uno; tiene funciones endocrinas y exocrinas (Capítulo 11). Entre los diversos grupos de peces tiene mucha variación por su posición y tipo de desarrollo. En los tiburones y rayas (Elasmobranchii) el páncreas es relativamente compacto y a menudo hi- lobulado, pero en los peces óseos (Osteichthyes) es muchas veces difuso (Tabla 2.3). Hay nódulos pequeños o grandes de tejido pancreático en el omento, con- junto de tejidos suspensores del intestino, además de la dispersión del páncreas en el hígado para formar un hepatopáncreas, entre los numerosos peces de radios espinosos (Acanthopterygii). La disposición de las células endocrinas que secretan la insulina, los islotes pancreáticos o de Langerhans, también varían en los diversos grupos de peces que poseen un páncreas discreto.
En la mayoría de los peces los dobleces de la pared intestinal, en el interior del mismo, muestran figuras rectangulares, como si fuesen, aparentemente, un

148 ALIMENTO, DIGESTI6N, NUTRICI6N Y CRECIMIENTO
dispositivo mecánico para retardar el desplazamiento del alimento. Las enzimas secretadas por el intestino delgado, así como las secreciones pancreaticas y biliares que vierten en esta parte del tubo digestivo, trabajan mejor en un margen de pH que oscila entre neutral y alcalino. Las diferentes proteasas que afectan las ligaduras terminales e internas que unen a los aminoácidos de las proteínas, son secretadas, ya sea por la mucosa intestinal, el páncreas o el ciego pilórico. La proteasa alcalina ha sido encontrada en el intestino de la trucha arco iris, y en el estómago de esta misma especie la proteasa ácida. Las enzimas intestinales son secretadas en una for- ma inactiva como zimógenos, un término general referido a las enzimas inactivas, antes que los cambios químicos que se realizan en el lumen o la luz del intestino las vuelvan activas durante la digestión. Este cambio es realizado por otra enzima como la enteroquinasa. Esta adaptación previene la autodigestión (autólisis) de la mucosa intestinal.
En el intestino y en el jugo pancreático de los peces se han llegado a encon- trar varias enzimas que digieren carbohidratos específicos. En Tilapia, pez predo- minantemente herbívoro, la actividad amilásica estaba distribuida a lo largo del tracto gastrointestinal, pero en la perca carnívora el páncreas constituía la única fuente de actividad amilásica. Existe evidencia en varios peces como la lacha (Brevoortia), el pez de flancos plateados (Menidia) y la corvina (Bairdiellu) de la presencia de bacterias viviendo en calidad de comensales que poseen celulasa, la cual puede desintegrar los materiales vegetales celulósicos, y aprovechar para sí mismas los materiales nutritivos, aunque sea en pequeña cantidad, como para po- nerlos a disposición del propio pez. Y todavía muchos peces herbívoros confían en exceso en el desmenuzamiento mecánico de las paredes celulares de las plantas, de aquí el desarrollo extensivo entre ellos de los dientes para cortar, desgarrar y triturar, y el hábito adquirido de alimentarse casi constantemente durante el día. El examen de las heces de un pez herbívoro, como la carpa de los “pastos” asiá- ticos (Ctenopharyngodon idella), revela que las células que no han sido trituradas por los dientes aparecen intactas y aún con los pigmentos verdes retenidos. Las células de las algas también pasan enteras e intactas a lo largo del tracto digestivo de algunos peces devoradores de plancton, como el llamado molleja (Borosorna cepedianum).
La inervación de los órganos digestivos proviene del sistema simpático a través de ganglios pares situados a los lados de la espina dorsal, y del parasimpá- tic0 a través de ramas del nervio vago (Capítulo 11). Algunos experimentos reali- zados en tiburones y rayas han demostrado que la luz y el olor del alimento no producen secreción gástrica. Esto va en contra de lo observado en los mamíferos, en los que se produce el flujo de los jugos digestivos mediante reflejos condicio- nados a través del nervio vago. La secreción pancreática y, probablemente, tam- bién la intestinal y el flujo biliar, están bajo el control hormonal y nervioso. Muchas hormonas que han sido aisladas de los tejidos intestinales de los peces para su estudio, han demostrado que actúan en el flujo de sustancias zimógenas y de otros fluidos como la bilis y la secreción pancreática amortiguadora del pH.
Absorción de los materiales digeridos
Procesos de la absorción. Para que los alimentos digeridos sean absorbidos, deben estar en una solución acuosa; por lo tanto, deben ser solubles. Los componentes

DIGESTI6N, ABSORCI6N Y UTILIZACION DE LOS ALIMENTOS 149
rnoleculares deben ser, más adelante, de un tamaiio tal que les permita atravesar las membranas que cubren el tracto digestivo, entrar al sistema circulatorio y final- mente ser llevados por la sangre hasta las células, penetrando en las que requieren nutrirse o simplemente almacenarlos. Es interesante hacer notar en este contexto que la absorción de la grasa es intensificada en el estómago pilórico de algunos peces (como en el tiburón colador, Cetorhinus) y los ciegos pilóricos de otros (co- mo en el género Salmo). Se ha visto que las grasas penetran en los conductos linfáticos en estas regiones sin haber sido desdobladas en las moléculas de ácidos grasos y glicerol, paso éste del que depende la absorción intestinal.
Aunque la mayoría de las proteínas digeridas son absorbidas en los intesti- nos, se ha visto que la absorción de algunos de los derivados de ellas ocurre en el estómago del tiburón. Se ha visto también que las unidades más pequeñas en que se llegan a desdoblar las proteínas en los intestinos son dipéptidos (compuestos que consisten de dos aminoácidos). Este paso intermedio hace pensar que el final, o sea, el último desdoblamiento de las proteínas hasta llegar a los aminoácidos, se realiza mediante la digestión intracelular; de las piedras milares (los aminoácidos) son resintetizadas las proteínas de los peces. Por lo tanto, la evidencia se acumula para darle menos crédito a la antigua creencia de que existía una diferencia entre los invertebrados inferiores y los vertebrados, basándose en la presencia de la diges- tión intracelular en el primer grupo, y su ausencia en el segundo.
Toxificación de la carne. Aparentemente, debido a la absorción de sustancias a través de los alimentos, la carne de muchos peces es tóxica en el hombre (ictio- sarcotoxismo). Esto sucede en algunas especies de peces, mientras que otras pue- den ser consumidas cientos de veces porque se ven inofensivas, pero, sorpresiva- mente, causan una grave enfermedad e inclusive la muerte, cuando son consumidas de nuevo. La evidencia tan notable que hay en cuanto al origen del ictiosarcoto- xismo apunta ahora hacia los hábitos alimenticios de los peces. Se piensa que las plantas marinas producen el veneno, el cual es transferido al hombre cuando el pez es comido, posiblemente después de su acumulación, concentración e inclusive alteración química en el propio pez. Los peces herbívoros pueden transmitir el veneno a los peces predadores, acumulándose la toxina en la carne de éstos, pero sin que haya ningún efecto dañino. Sin embargo, cuando estos peces predadores son consumidos por el hombre, entonces sí aparece la intoxicación. Aunque están ampliamente distribuidos en el mundo, la mayor parte de los peces venenosos conocidos son de aguas tropicales. Las aguas templadas tienen, sin embargo, mu- chos peces globo (Tetraodontidae), entre otros, los cuales son muy ponzoñosos debido a las sustancias que ellos mismos producen. Algunas veces es venenoso el tiburón de Groelandia (Somniosus microcephalus), que habita en los mares árticos.
Se sabe que tienen carne ponzoiíosa muchos tiburones entre los cuales se incluyen algunos tiburones “requiem” (Carcharhinidae), tiburones vaca (Hexan- chidae), tiburones perro (Squalidae) y tiburones macarela (Lamnidae). Entre los peces óseos frecuentemente son venenosos los que habitan en los arrecifes. Las familias que tienen representantes tóxicos conocidos son las que incluyen a las mo- renas (Muraenidae), las macarelas y atunes (Scombridae), los peces cofre o toritos (Ostraciidae), los peces globo (Tetraodontidae) y los peces erizo (Diodontidae). Más de trescientas clases de peces han sido incriminadas en el tipo “ciguatera” de toxicidad ictiológica, que es fatal para cerca del 7% de los humanos que se arries- gan a comer la carne de estos peces. No hay un tratamiento específico, aunque

150 ALIMENTO, DIGESTION, NUTRICI6N Y CRECIMIENTO
actualmente se está desarrollando un anticuerpo “ciguatera”. Las reacciones son en su mayor parte nerviosas, con sensaciones reversibles de calor y frío, siendo comunes éstas en los casos severos. Las familias de peces que periódicamente presentan la toxicidad incluyen a los quijos (Albulidae), las sardinas (Clupeidae), las anchoas (Engraulidae), los salmonetes (Mullidae), los serranos (Serranidae), los pargos o huachinangos (Lutjanidae), los jureles (Carangidae), los peces pluma (Sparidae), los peces señorita o viejita (Labridae), los peces loro (Scaridae), las barracudas (Sphyraenidae), los cirujanos (Acanthuridae) y los peces torito o cofre (Ostraciidae). Desgraciadamente, la aparición de la “ciguatera” es impredecible y puede llegar de repente.
NUTRICIdN DE LOS PECES
Una vez que el alimento ha sido ingerido y digerido, puede participar en varias fun- ciones en el cuerpo del pez. Puede proporcionar energía para los procesos bioló- gicos o ceder material para la restauración o reposición de los componentes celulares destruidos o desgastados, o para el crecimiento y la reproducción. Unos cuantos peces (los atunes) pueden regular la temperatura de su cuerpo; en los de- más fluctúa con la del medio ambiente (condición poiquiloterma). Por lo tanto, su grado de metabolismo y, como consecuencia, sus requerimientos nutricionales dependen, sobre todo, de la temperatura que tenga el agua en donde viven. Desde el momento en que las aguas de las zonas templadas presentan fluctuaciones ma- yores de 25°C (77”F),* puede ocurrir un amplio margen de grado de metabolismo y consumo de alimento en un solo individuo a lo largo del año.
La mayor parte de nuestro conocimiento relacionado con los requerimien- tos alimenticios de los peces se basa en la esperanza del hombre de criarlos en lagos y ríos, con fines alimenticios y de reserva. La información detallada está restrin- gida, por lo tanto, a sólo unas cuantas especies, principalmente las truchas y los salmones de la familia Salmonidae. Solamente aquí y all6 aparecen, ocasional- mente, estudios sobre el medio ambiente de las aguas dulces naturales y las ma- rinas, que complementan los datos nutricionales básicos que existen sobre estos salmónidos.
Conversión y eficiencias del alimento
Los peces, como los demás animales, pueden ser comparados a máquinas comple- jas en relación a su eficiencia en el aprovechamiento del alimento, o sea, a su rendimiento. Uno puede simplemente medir el peso del alimento que incrementa en una unidad del peso de1 pez (sea tomado en libras, gramos o kilogramos), para obtener asi un “factor de conversión”. Por ejemplo, una. trucha arco iris (Salmo
*El original indica: 2 5 ° C (45”F), lo que evidentemente es un error, ya que la fórmula de conversión es la siguiente:
“F -32
1.8 “C =
y, en caso inverso ‘F= “C X 1.8+ 32. Entonces, 25°C equivalen a ‘77 ’F. En los casos posteriores hemos indicado las equivalencias correctas (N. del T. ).

NUTRICIdN DE LOS PECES 151
guirdneri) obtenida en un vivero piscícola, bajo ciertas condiciones y recibiendo determinados alimentos, requiere 1,589 gramos de alimento por cada 454 gramos de incremento de peso. Esta clase particular de alimento daría un factor de con- versión de 3.5.
Los valores de los factores de conversión difieren con la naturaleza de la dieta, la especie y tamaño del pez, la temperatura y otras variables. El margen de los factores de conversión se extiende desde 1.5, cuando ciertas dietas secas arti- ficiales son proporcionadas a la trucha (Salmonidae) en los viveros, a 8 o más cuando se le da a la carpa (Cyprinus carpio) o a otras especies de peces omnívoros una dieta rica en componentes vegetales como el maíz y leguminosas del género Lupinus. El factor de conversión es de 2.5 a 3.0 en la mayoría de las dietas para los viveros piscícolas. Los factores de conversión-peso tienden a menudo a ser altos en la naturaleza, mucho más de los que se miden en los viveros; ello debido a la presencia de mucho desperdicio no digerible como los esqueletos de los crus- táceos e insectos y, combinado esto con el empleo de grandes cantidades de ener- gía debido a la desusada actividad del pez cuando está en libertad a la que realiza cuando vive en cautiverio en los estanques de un vivero piscícola. Tambiénse puede expresar la relación que hay entre el alimento consumido y el peso ganado como un coeficiente de conversión de peso. Siguiendo al ejemplo que se dio en el pá- rrago anterior, tendremos que
1 unidad de peso ganada 3.5 unidades de peso de alimento
= 0.2857
o una eficiencia de conversión de peso del 28.57%, si multiplicamos por cien el resultado anterior.
Para el nutriólogo resulta muy útil el conocimiento de la eficiencia de la conversión del alimento; también para el fisiólogo o el ecólogo. Pero esta eficien- cia de conversión del alimento, para serle útil a estos especialistas, debe estar ex- presada en equivalentes de calorías relativas al peso del alimento y al de la carne del pescado, respectivamente. Los carbohidratos, grasas y proteínas pueden ser quemados para obtener de ellos energía calórica cuando se combinan con el oxí- geno durante el proceso de la combustión. Se puede pensar que los procesos meta- bólicos son combustiones similares, pero lentas, o cualquier otra transformación química donde se hace derivar la energía de los carbohidratos, las grasas, las pro- teínas y otros compuestos. La energía calórica es expresada y medida en calorías. Una caloria es la cantidad de calor necesaria para elevar un grado centígrado de temperatura a un gramo de agua en la escala centígrada o Celsius. Otras formas de energía, como la química, la mecánica o la eléctrica, pueden también ser expre- sadas en términos de calorías después de haber hecho las transformaciones o con- versiones matemáticas correspondientes.
Un pez transforma un compuesto en otro cuando produce carne, escamas y huesos, y cuando realiza sus movimientos y otras funciones vitales. La energía empleada para estos fines deriva del desdoblamiento de varios compuestos, los que tienen diferentes equivalentes de energía, dependiendo de su composición química y de su estructura. Por ejemplo, 454 gramos de carne ordinaria de pescado tienen cerca de 600 kilocalorías; el azúcar tiene 1,695 y la mantequilla, grasa con un elevado equivalente energético, tiene 3,260 kilocalorías.
Se sabe bien que parte de la energía de la gasolina que explota en el cilindro

be un motor. de aut,ombvil se disipa p ~ r el frotmniento que oeurre en las diferentes piezas que conforman el motor y se pierde en calidad de efectivo propelente des- perdiciado. Por io tanto, la eficiencia de una máquina nunca llega a; 3.00%. Pur una simp!" ana.iogía nosotros podernos comparar esas pkrdicias corn las que suce- den en los animales cuando una porcibn de !a energía que ellos liberan d i j r m k sus procesos metab6,licos se gasta en el simpk mantenimiento de procesos biolb- gicos tales cemo la respiraci6n7 digestih, excrecilin y otms funciones vitales, y no se le encuentra en forma de aumento de peso.
Nosotros podemos medir o pesar l a cantidad de Inaterial nuevo que el pez agrega a su propio cuerpo durante e1 crecimiento y tambidn podemos medir c! pesar la cantidad de alimento guc consume. Podemos tambikn expresar esas can- tidades en ca1oría.s en lugar de kilogramos o gramos, y compararlas, a. saber:
calorías contenidas en la carne agregadas por unidad de tiempo
calorías contenidas en varios alimentos comidos por unidad de tiempo ..____-__ "" x 100
Esta fracción, expresada en porcentajes, es conocida como coeficiente de eficiencia en primer grado. En !a carne del lucio se ha llegado a encontrar entre el 3-4 y el 33% de las calorías consumidas. Woscrtros podemos ir todavía más lejos y rebajar el denominador mediante la determinación de las calorias empleadas en el Inantenimiento celular, Ha excreción, l a respiración, y otros procesos metabblicos que se suceden mientras que los tejidos nuevos son agregados a los ya existent.es y, en consecuencia, representan parte del consumo de energía que no es empleada para el crecimiento (si consideramos al crecimiento COMO la transformación del material ingerido, en tejido nuevo, aparte del que se requiere para la reparación o la reposición). Por lo tanto,
calorias agregadas en la carne, por unidad de tiempo (calorías contenidas en el alimento consumido, por unidad de tiempcd)
x 100
- (calorías empleadas en el metabolismo, mantenimiento, etc.)
se transforma en el coeficiente de eficiencia de segundo orden que siempre es mayor que el de primer orden, y proporciona una medida más exacta de las pro- piedades nutritivas de varios alimentos, que la que proporciona el coeficiente de eficiencia de primer orden.
Por lo tanto, el coeficiente de primer orden, o coeficientes totales, deben ser distinguidos de los coeficientes o eficiencias de segundo orden. En las eficiencias de segundo orden la ganancia en peso, la energía potencial o la agregación de proteína, es comparada con las cantidades de material o energía que produce esa ganancia. El término asimilado debe ser sustituido por comido, en el denominador, para tomar en cuenta solamente al alimento que pasa a lo largo del tubo digestivo y excluir la defecación de los materiales no digeridos. Las comparaciones, sobre la base de los equivalentes calóricos, permiten la tasación de los efectos de la utili- zacibn de los alimentos, de variables como la edad de los peces, la temperatura, las clases y cantidades de alimento y otras.
Las eficiencias totales de energía declinan con la edad y tamaíío del pez. Varían desde el 58% en los embriones de pez (como en el "mummichog", Fundu- lus heteroclitus) al 10% o menos como en los peces grandes y viejos (como en los

NUTRICIdN DE LOS PECES 153
lenguados, Pleuronectes). La mayor parte de los coeficientes de energla queda comprendida entre el 15% para los peces viejos y el 45% para los jóvenes. Es posi- ble que aparezcan valores negativos para periodos cortos; como sucede en el in- vierno, cuando muchos peces de la zona templada pierden peso.
La formación de proteínas en el cuerpo es influenciada por la clase de alimento que el pez consume. Esto ha sido demostrado en las truchas (Salmoninae) mediante la comparación entre lotes que han sido alimentados con alimento pre- parado o con alimento natural. A pesar del hecho de que la dieta artificial era superior a la natural por su contenido de proteínas y vitaminas, la trucha que fue alimentada con alimentos naturales (insectos, etc.) fue doblemente eficiente que la alimentada con una dieta artificial, en el vivero piscícola, al convertir las proteí- nas del alimento en carne; además, la trucha mantenida con alimento natural, tenía carne con un bajo contenido de agua y mucho de proteína.
Los valores para la asimilación, expresados como porcentajes de la utiliza- ción de los alimentos retenidos en el cuerpo, tienen un valor entre el 80 y el 90% ; las tensiones que resultan del medio ambiente y la baja temperatura del agua pueden reducir estas eficiencias. Considerando que las dietas tengan la misma composición, los experimentos de alimentación en la trucha café (Salmo trutta) y en el lucio del Norte (Esox lucius) indicaron que el pez utiliza el alimento con más eficiencia después de una abstinencia voluntaria que cuando está sa- ciado, y también que el alimento es menos absorbido cuando el pez es sobre- alimentado.
Cantidad de alimento consumido
Los peces predominantemente ramoneadores de los arrecifes, como los peces loro (Scaridae) y los cirujano (Acanthuridae), se alimentan a lo largo del día en forma más o menos ininterrumpida. De esta manera, ellos, y también todos los que se alimentan de plancton, a la inversa de los carnívoros, ingieren cantidades mucho más grandes de alimento. Sin embargo, la mayor parte de las algas que comple- mentan la dieta alimenticia tienen solamente la mitad del valor calórico y menos de la cuarta parte del contenido proteínico de la carne del animal. Y, todavía más: a pesar del hecho de que las algas y diatomeas unicelulares y planctónicas, así como pequeñas porciones de las plantas acuáticas enraizadas puedan tener valores nutritivos adecuados, contienen un elevado porcentaje de agua. Por estas razones, usualmente los herbívoros consumen mayores cantidades de alimento, en peso, que los carnívoros.
Los peces carnívoros que tienen diferentes hábitos alimenticios también consumen cantidades variables de alimento porque sus presas varían, a su vez, en composición y tamaño. Los que se alimentan de macrocrustáceos consumen relativamente más materiales no digeribles que los piscívoros. Algunos peces pue- den, a veces, tomar una cantidad tal de alimento que les puede dejar satisfechos por una semana o más. Un lucio del norte (Esox lucius), después de haber tragado un pez casi de su mismo tamaño, puede estar haciendo la digestión de la cabeza de la presa mientras exhibe la cola de la misma, por tenerla fuera del hocico. Los bien llamados tragadores (Saccopharyngidae) y, con el mismo nombre vernáculo, pero de otra familia (Eurypharyngidae) de las grandes profundidades marinas, a menudo capturan presas que son mucho más grandes, sabiendo que cuentan
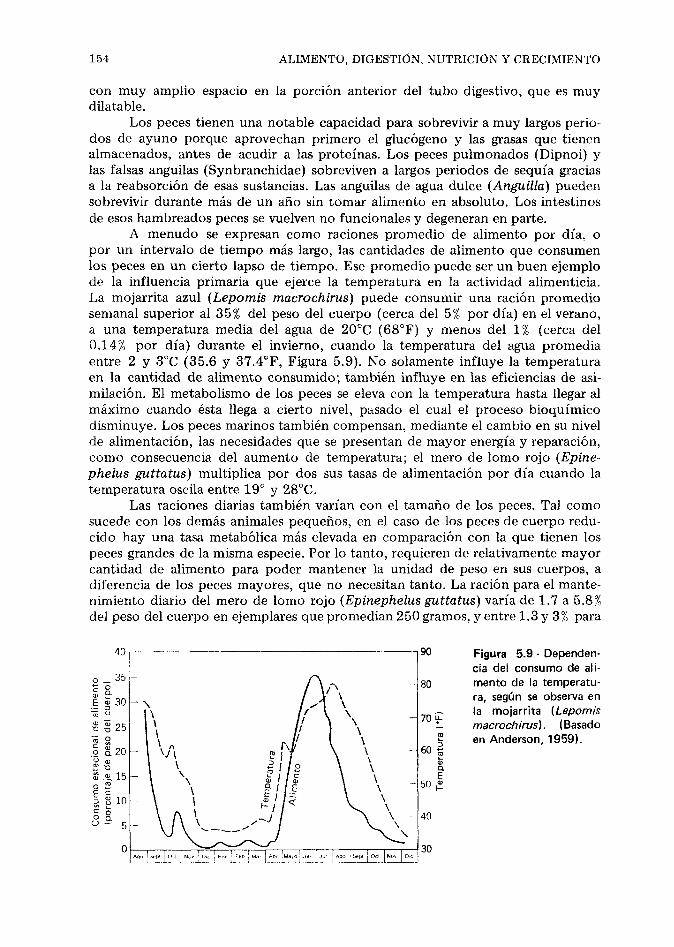
154 ALIMENTO, DIGESTI6N, NUTRICION Y CRECIMIENTO
con muy amplio espacio en la porción anterior del tubo digestivo, que es muy dilatable.
Los peces tienen una notable capacidad para sobrevivir a muy largos perio- dos de ayuno porque aprovechan primero el glucógeno y las grasas que tienen almacenados, antes de acudir a las proteínas. Los peces pulmonados (Dipnoi) y las falsas anguilas (Synbranchidae) sobreviven a largos periodos de sequía gracias a la reabsorción de esas sustancias. Las anguilas de agua dulce (Anguilla) pueden sobrevivir durante más de un año sin tomar alimento en absoluto. Los intestinos de esos hambreados peces se vuelven no funcionales y degeneran en parte.
A menudo se expresan como raciones promedio de alimento por día, o por un intervalo de tiempo más largo, las cantidades de alimento que consumen los peces en un cierto lapso de tiempo. Ese promedio puede ser un buen ejemplo de la influencia primaria que ejerce la temperatura en la actividad alimenticia. La mojarrita azul (Lepomis macrochirus) puede consumir una ración promedio semanal superior al 35% del peso del cuerpo (cerca del 5% por día) en el verano, a una temperatura media del agua de 20°C (68°F) y menos del 1% (cerca del 0.14% por día) durante el invierno, cuando la temperatura del agua promedia entre 2 y 3°C (35.6 y 37.4"F, Figura 5.9). No solamente influye la temperatura en la cantidad de alimento consumido; también influye en las eficiencias de asi- milación. El metabolismo de los peces se eleva con la temperatura hasta llegar al máximo cuando ésta llega a cierto nivel, pasado el cual el proceso bioquímico disminuye. Los peces marinos también compensan, mediante el cambio en su nivel de alimentación, las necesidades que se presentan de mayor energía y reparación, como consecuencia del aumento de temperatura; el mero de lomo rojo (Epine- phelus guttatus) multiplica por dos sus tasas de alimentación por día cuando la temperatura oscila entre 19" y 28°C.
Las raciones diarias también varían con el tamaño de los peces. Tal como sucede con los demás animales pequeños, en el caso de los peces de cuerpo redu- cido hay una tasa metabólica más elevada en comparación con la que tienen los peces grandes de la misma especie. Por lo tanto, requieren de relativamente mayor cantidad de alimento para poder mantener la unidad de peso en sus cuerpos, a diferencia de los peces mayores, que no necesitan tanto. La ración para el mante- nimiento diario del mero de lomo rojo (Epinephelus guttatus) varía de 1.7 a 5.804 del peso del cuerpo en ejemplares que promedian 250 gramos, y entre 1.3 y 3% para
90 Figura 5.9 - Dependen- cia del consumo de ali-
80 mento de la temperatu- ra, según se observa en la mojarrita (Lepornis - rnacrochirus). (Basado 70
40
30

NUTRICI6N DE LOS PECES 155
los peces cuyo promedio es de 600 gramos, cuando la temperatura del agua se eleva de 19" a 28°C. Los individuos pertenecientes a una especie cuyo promedio sea menor en relación al tamaño, al del mero de lomo rojo, deberán tener todavía mayores requerimientos de ración alimenticia diaria que los citados anteriormente.
La disponibilidad de alimento para el pez también influye en las cantidades consumidas. El mero de lomo rojo se engulle a sí mismo por los consumos diarios de alimento que hace, que sobrepasan el 10% del peso del cuerpo en los experi- mentos de alimentación para mostrar los arrebatos de saciedad, pero pronto vuelve a mostrar un hábito alimenticio con tasa más conservadora. Carpas (Cyprinus carpio) de un año de edad conservadas en estanques pueden tener una ración diaria tan elevada como el 16% del peso de su cuerpo, cuando comienzan a comer al iniciarse la estación de crecimiento. A diferencia de lo que ocurre en la mayoría de los animales terrestres, casi todos los peces experimentan un agotamiento severo en una etapa de cada año de su vida.
Calidad de alimento
El verdadero crecimiento corresponde a la agregación de carne (proteínas) y tejido óseo; no incluye los depósitos menos permanentes de grasa y agua en los te- jidos del pez. Sin embargo, la dieta de los peces debe ser balanceada y contener los componentes alimenticios primarios o básicos -proteínas, carbohidratos y lípidos (grasas)- como un requisito, aunque se encuentren en diferentes propor- ciones en las diversas especies de peces. También son requeridas las vitaminas y los minerales para el crecimiento, sostenimiento y reposición de los tejidos, así como para mantener el metabolismo normal.
Alimentos básicos. Las adaptaciones evolutivas que se han presentado entre los peces les han llevado a diferir en cuanto a sus capacidades para utilizar alimen- tos con cantidades variables de proteínas, carbohidratos y grasas. La trucha (Sal- moninae), por ejemplo, desarrolla hígados con grasa infiltrada cuando el conte- nido de grasa de su dieta excede al 8% (en peso seco), mientras que la lacha del Atlántico (Breuoortiu tyrannus) se alimenta normalmente de algas que contienen por lo menos un 25% de grasa. La carpa común (Cyprinus carpio) y las tilapias de la familia Cichlidae pueden ser cultivadas con una dieta que contenga inclusive una cantidad mucho mayor de lípido. No solamente las cantidades relativas de grasa contenidas en el alimento afectan la utilización de éste; también la compo- sición de la grasa tiene su efecto: los ácidos grasos no saturados con un bajo punto de fusión (número de yodos elevado) son más fácilmente digeribles que los ácidos grasos saturados.
El contenido permisible de carbohidrato en las dietas artificiales que, des- pués de todo, al ser fabricadas resultan iguales si no es que superiores en calidad al alimento natural de los peces, varía igualmente para las diferentes especies. Cuando las truchas son alimentadas con una dieta que contenga más del 12% de carbohidrato digerible (base húmeda) se obtienen como resultados una disposición anormal de glucógeno hepático y mortalidad excesiva. Como contraste, los pece- cillos de agua dulce de la familia Cyprinidae, criados en todo el mundo para ali- mento, a menudo crecen muy bien con mezclas alimenticias que contengan un 50% o más de fécula. Las facultades de la trucha para digerir los diferentes carbo-

hidratos difieren para cada uno de 6stos; por ejemplo solamente el 99% de la glu- cosa fue digerido, y ~610 el 73q, de la sucrasa. Los requerimientos más rigurosos, de !os tres elementos nutritivos básicos, corresponden probablemente a las proteinas. La dieta de cualquier animal debe contener necesariamente cierta cantidad de prot,eína. El requerimiento de proteina varia con los cambios que aparecen @:I el ciclo de vida del pez, siendo grande cuando &e es pequeño, crece ripido y cuando se prepara para realizar la forrnnclbn de óvulos y esperma. EI alimento para peces formado por invertebrados contiene corno promedio un 11.5% de proteína, aproximadamente. Parece ser que cuando el alimento tiene menos del 6 % de prn- teina (en peso húmedo) no nut.re suficientemente a ningGn pez y de hecha frena el crecimiento.
No son las cantidades de proteina ingeridas las únicas importantes; tam- bién lo son sus contenidos relativos de aminoácidos, segiín queda insinuado por las eficiencias de conversicin tan elevadas que se obtienen cuando un pez se come a otros de su misma clase. Ta.l como sucede con todos los animales, los peces pue- den fácilmente sintetizar ciertos aminoieidos muy rápidamente, pero otros s& pueden hacerlo muy lentamente para lograr la reposición satisfactoria. Por lo tanto, éstos tienen que recibir una dieta suplementaria. Y todavía un tercer grupo de aminoácidos es absolutamente esencial y tiene que ser agregado al alimento, porque los peces no lo pueden sintetizar. Algunos estudios detallados sobre este aspecto de la nutrición de los peces han sido hechos en el salmón chinense (Oncorhynchus tshawytscha). Este salmón no sintetizó la arginina, histidina, isoleucina, leucina, lisina, rnetionina, fenilalanina, teonina, triptofano y valina, ami- noácidos que, de todas maneras, de haber estado presentes, pudieron haber sido sintetizados en tan pequefías cantidades, que no habrían influido en el crecimiento del animal, por estar prácticamente ausentes en la dieta alimenticia. En este aspec- to, el salmón chinense recuerda a la rata (Rattus) más que al hombre, porque éste puede .pasarla con una dieta en la que falten dos de esos aminoácidos (la his- tidina y la arginina) dado que pueden ser sintetizados en el cuerpo humano.
El agua, a pesar de no ser propiamente un alimento básico, es necesaria en los peces, como en los demás animales, para facilitar los procesos metabólicos en los que queda incluida la digestión extracelular. Un animal puede perder prác- ticamente toda su grasa y la mitad de sus proteínas y seguir viviendo; pero una pérdida de solamente el 10% de su agua le produce la muerte. La cantidad de agua que es tomada incidentalmente por los peces en el momento de comer no se conoce con precisión. La presencia en algunos peces de un juego de esfínteres en la entrada del estómago, y las adaptaciones concentradas y reforzadas en la boca y la faringe de otros, como los que se alimentan de plancton, sugiere que debe ser insignificante la entrada accidental de agua al tracto digestivo. Los pe- ces de agua dulce absorben agua a través de las membranas semipermeables de las branquias y de otras superficies expuestas. Los peces marinos contrarrestan las pbrdidas de agua por exbsmosis (Capítulo 9; Figura 9.1), en parte tragando agua de mar y en parte adquiriéndola de los alimentos. Es probable que mucha del agua requerida para los procesos digestivos en los peces de agua dulce sea también extraída de los alimentos. Es entre un 70 y un 90% la cantidad de agua, calculado por peso húmedo de los alimentos, que se encuentra en los que consu- men los peces; no se toman en cuenta para el cálculo de estos porcentajes las partes duras, como las conchas y los huesos.

NUTRICIdN DE LOS PECES 157
Vitaminas. El conocimiento que se tiene actualmente del papel que desempeñan las vitaminas en la nutrición del pez se deriva primeramente de los experimentos que sobre su alimentación se han realizado en varias especies de truchas y salmo- nes (Salmoninae), en bagres y carpas. Las vitaminas aparecen en cantidades sufi- cientes en la dieta natural de los animales, de tal manera que satisfacen las necesi- dades de cantidades muy pequeñas para cumplir con las funciones rnetabólicas normales. Sin embargo, cuando los peces son cultivados con dietas artificiales pueden presentarse deficiencias específicas por la selección de los constituyentes alimenticios, la propiedad negativa que tienen algunas vitaminas de perder su vita- lidad en corto tiempo, y otros factores, con los daños consecuentes en las funcio- nes del cuerpo. La trucha y el salmón requieren de diez componentes del complejo vitamínico B (Tabla 5.1). Para la carpa se ha establecido bien que se requiere piridoxina, riboflavina y ácido pantoténico; y para el bagre, algunas de las vitami- nas solubles en agua y grasas. La trucha necesita vitamina C soluble en agua. Se ha establecido para algunos peces la necesidad de las vitaminas solubles en grasas: A, K y E. Pero no para la vitamina D.
Minerales. Los minerales, como sucede con las vitaminas, aparecen en casi todas las dietas naturales en cantidades suficientes para satisfacer los requerimientos
Tabla 5.1 - Efecto de las deficiencias vitamínicas en los peces.a
Sustancia Síntomas por la deficiencia
Vitamina B1 (tiamina) PBrdida del apetito; atrofia muscular; convulsiones; malfuncionamiento nervioso; muerte.
Vitamina Bz (riboflavina) Los ojos se vuelven opacos; cesa el crecimiento.
Vitamina B6 (piridoxina)
Vitamina BIZ (cobalamina)
Desórdenes nerviosos; ataxia; anemia; reducción del crecimiento; muerte (de 4 a 6 semanas en las truchas).
Hemoglobina y cantidades de eritrocitos erráticas; reducción del crecimiento.
(Innumerables vitaminas en el complejo B) Biotina Falta de correspondencia en la producción de mu-
cosidad; convulsiones; "enfermedad de la visco- sidad azul".
Colina Reducción del crecimiento.
Ácido fólico Riñón e intestino hemorrágicos; reducción del crecimiento; anemia.
Inositol Reducción delcrecimientoyprobablernente anemia.
Niacina Incremento en la sensibilidad a la luz ultravioleta; "quemadura solar".
Ácido pantoténico Enfermedad de las branquias; falta de correspon- dencia en la producción de mucus; "enfermedad de la viscosidad azul"; pereza.
ase han adscrito ciertos efectos a otros factores menores pero se ha resuelto que son acciones sinergisticas de muchas vitaminas del complejo B y a la falta de algunos aminoácidos esenciales.

158 ALIMENTO, DIGESTION, NUTRICI6N Y CRECIMIENTO
metabólicos de los peces; inclusive en los criaderos las deficiencias minerales son raras y, cuando aparecen, son fácilmente solucionables.
Los minerales y otros elementos pueden ser clasificados de acuerdo a las tres funciones más importantes que desempeñan en el cuerpo: u ) estructural: calcio, fósforo, flúor y magnesio (para la formación de dientes y huesos); b ) respi- ratoria: hierro, cobre y cobalto (para la formación y función de la hemoglobina), y e) metabolismo en general: muchos elementos (para las funciones de las células y el cuerpo). En la última categoría se encuentran incluidos el sodio, potasio, cloro y calcio que regulan el equilibrio osmótico y la turgencia celular. En este caso también son requeridos el boro, aluminio, cinc, arsénico y trazas de otros minerales; los iones de cloro son necesarios para la elaboración del jugo gástrico; y el magnesio, fósforo y cloro, en forma de iones también, son necesarios para activar las enzimas digestivas. Para algunas enzimas son cofactores indispensables el cobre y el cinc, y el azufre y el fósforo intervienen en forma importante en el trifosfato de adenosín (ATP) y otros compuestos de alto contenido energético. El bromo interviene en la maduración de las gónadas, mientras que el yodo es un constituyente de las hormonas tiroideas (Capítulo 11). Una deficiencia de yodo puede producir malestares parecidos a las paperas en las truchas cultivadas en las pesquerías, y puede ser contrarrestada mediante su agregación en los alimentos previamente elaborados. Los isótopos radioactivos de calcio contenidos en el agua son absorbidos directamente en cantidades cada vez más elevadas, a medida que el mineral va faltando en las dietas artificiales. También a través de las branquias pueden penetrar al cuerpo el litio, sodio, cloro y bromo, pero la mayoría de los elementos se encuentran debidamente representados en los alimentos naturales de los peces, y parece ser que realmente a través de la comida es como son con- sumidos.
La composición química de los peces. La mayor cantidad de datos que se cono- cen sobre la composición química elemental de los peces se refiere a los valores nutritivos y a la presencia de componentes químicos en la carne de las especies de importancia comercial, cuyo número llega a ser de unas 350 a 400, la mayor parte formada por peces óseos marinos. Los datos relacionan el agua, la grasa, las proteínas, los carbohidratos y los minerales por unidad de peso.
El contenido de agua de la carne de los peces promedia un SO%, un poco más del que se encuentra en las aves y mamíferos. En los peces se han encontrado valores extremos de 53 y SS%, para ciertas especies, en ciertas estaciones del año y en ciertas localidades.
El contenido de agua de la carne de los peces varía con la especie y las eta- pas de su desarrollo vital. Las diferencias que pueda haber en el contenido de agua de acuerdo con el sexo son pequeñas, según se ha reportado para unas cuantas especies, como en el arenque, Clupea harengus. Los machos de este arenque con- tienen a menudo más agua que las hembras, en una proporción de 1 a 2 % . Esta diferencia puede estar relacionada con el alto contenido de agua de los testículos, siempre mayor que el de los ovarios. Las crías con saco (larvas que aún tienen adherido el saco de la yema) tienen un porcentaje de agua más bajo que el de los adultos maduros. La larva leptocéfala de las anguilas de agua dulce (Anguilla) contiene, sin embargo, más agua en este estado que cuando se ha transformado en individuo con estructura y forma de adulto. Entre los adultos, sin embargo, todavía continúa una deshidratación que prosigue a medida que avanza la edad.

NUTRICI6N DE LOS PECES 159
Las especies anadromas, como las de los salmones (Oncorhynchus y Salmo salar) y las razas de la lamprea marina (Petromyzon marinus) que hacen largos recorri- dos migratorios, aumentan su contenido de agua en forma perceptible en los momentos de la puesta debido a que reponen con agua las grandes cantidades de grasa y proteína, cuya energía emplean para alcanzar los ríos para el desove. Las bacalaos (Gadidae), los lenguados (Pleuronectidae) y los lófidos (Lophiidae) pue- den contener más del 80% de agua, mientras que las anguilas de agua dulce (An- guilla), estando en el mar, los arenques y sardinas (Clupeidae) y los atunes y maca- relas (Scombridae), tienen generalmente un contenido de agua menor al 75%. Este primer grupo, representado por las merluzas, los lenguados y los lófidos, es ben- tónico y sus miembros tienen una gravedad específica apenas un poco mayor que la del agua marina; para poder elevarse requieren del empleo de la vejiga gaseosa. El segundo grupo, representado por las familias donde se incluyen las sardinas y las macarelas, es principalmente pelágico, con carne de un alto contenido graso, mayor que el del primer grupo, y con algunos representantes cuya vejiga gaseosa se ha reducido en tamaño o aparece en forma vestigial. La gravedad específica de los individuos del segundo grupo se aproxima a la del agua marina, debido 2. la pre- sencia de grandes masas grasosas en el tejido muscular.
El contenido de grasa de la carne de los peces varía no solamente en rela- ción con las familias; también con las estaciones del año. Los devoradores de planc- ton de las zonas templadas, como los arenques y sardinas (Clupeidae), contienen más grasa y menos agua en sus tejidos después de los pulsos planctónicos de primave- ra que después de la ovoposición, que es su etapa de mayor debilidad o decaimiento. Los contenidos de grasa y agua en los tejidos de un pez, tomado como individuo, varían inversamente entre una y otra; y, por otra parte, el contenido de grasa varía en el mismo pez, de un lugar a otro. El músculo del arenque (Clupea harengus) tiene un contenido de grasa inusualmente alto en la región abdominal; como contrapeso, posee una concentración diferente de esa sustancia en una región cercana a la aleta dorsal.
Los peces de agua dulce también muestran diferencias de grupo en su con- tenido relativo de grasa y, generalmente, ésta tiene características químicas dife- rentes a las que presenta la grasa de las especies marinas. Los bagres tropicales de agua dulce, como los siluros laberintinos (Clariidae) y los esquilbeídos (Schilbei- dae), contienen más grasas que los demás grupos de la misma región, mostrando así que poseen ciertos hábitos y adaptaciones específicos para la alimentación, du- rante la cual sólo utilizan ciertos alimentos. La mayoría de los peces de agua dulce de las zonas templadas acumulan grasa como reserva para el invierno y/o para su gas- to energético durante la ovoposición. Los sitios más importantes donde se realiza la acumulación corresponden al tejido conectivo y a los mesenterios de los intestinos.
Solamente ácidos grasos en número invariable aparecen formando parte de las grasas y aceites de los peces. Estos ácidos grasos están menos saturados que los de los animales terrestres y de los que intervienen en la formación de los aceites vegetales. Su contenido de ácido oleico, un precursor del colesterol, es relativa- mente bajo. Por esta razón han sido considerados como benéficos al formar parte de la dieta humana, ya que no influyen mucho en el endurecimiento patológico de las arterias.
Las proteínas representan del 14 al 23% del peso húmedo de los peces. Y aquí, otra vez, las variaciones aparecen por especies, estaciones del año y etapas en el ciclo biológico. Usualmente, entre el 2 y el 3% del peso del cuerpo del pez vivo

I60 ALIMENTO, DlGEST16Ni NUTRIC16N Y CRECIMIENTO
corresponden a la totalidad del nitrógeno que contiene el animal. Los peces seme- jantes al ti.burón (Elasmobranchii) contienen más nitrógeno que los peces 6seos (Osteichthyes) porque los tejidos de los elasmobranquios contienen mucha urea osmorreguladora, a diferencia de los tejidos de los peces 6seos que no la presentan (Capítulo 9). Sobre la base del peso seco el nitrógeno está más concentrado en el hígado, menos en la piel y aún en menor proporción en la carne.
Los carbohidratos que están presentes en el músculo y proporcionan ener- gía para su uso inmediato forman menos del 1% del peso húmedo de los peces; están más concentrados en el hígado, donde permanecen en reserva como glucógeno.
Algunos de los numerosos elementos químicos que se encuentran en los peces están más concentrados en las especies marinas que en las de agua dulce. Se piensa que el elevado contenido de estos elementos refleja su abundancia en el agua de mar y su correspondiente incorporación pasiva por los peces marinos a través de su concentración en todos los eslabones de sus cadenas alimenticias. El contenido de cenizas a partir de los tejidos blandos de los peces llega a ser apro- ximadamente el 1% de su materia viva. Cuando en el análisis son incluidos los huesos y las escamas, las cantidades de ceniza llegan a corresponder al 3% o más, debido a la presencia del fosfato de calcio, Ca3(P04)2, y al carbonato de calcio, CaCO,, compuestos que forman casi toda la masa de estas partes duras de los peces. Las escamas forman casi el 30 a 35% de la ceniza, sobre la base del peso seco del material.
CRECIMIENTO DE LOS PECES
El modelo de crecimiento de los peces es notable, desde el momento en que la mayoría de ellos tiene la capacidad de sostener el ritmo de crecimiento, a lo Iargo de sus vidas, aunque a veces disminuya (crecimiento indeterminado), si hay dis- ponible suficiente cantidad de alimento. Por lo tanto, los miembros de una espe- cie, siendo de la misma edad, pueden ser de tamaños variables, en contraste con los tamaños más definidos, a determinada edad (crecimiento determinado), que aparecen en unos cuantos peces, como sucede en los machos de los gupies (Lebis- tes) y generalmente entre los individuos de los grupos de vertebrados terrestres. En otras palabras, la mayoría de los peces, en contraste con lo que sucede en las aves y mamíferos, no cesan de crecer después de que han alcanzado su madura- ción sexual. Mientras que el crecimiento de los peces es, por lo tanto, mucho más variable y flexible que el de las aves o mamíferos, todavía existe una última limi- tación genética para el crecimiento de una especie, y este factor es más evidente en las clases más pequeñas. Se ha propuesto, como una explicación al contraste citado anteriormente, que los peces que viven en un medio fluido que los sos- tiene mecánicamente, pueden continuar creciendo a lo largo de sus vidas porque hay más factores limitantes bióticos que mecánicos que se imponen a sus tama- ños máximos.
Se ha definido el c.recimiento en los animales como la agregación de ele- mentos estructurales o carnosos; por eso se habla de las proteínas. El crecimiento de los peces, en lo particda, es de gran importancia para el hombre por ser el explotador y administrador potencial de las poblaciones de peces, ya sea para la obtención de alimento o simplemente como deporte. Es un hecho que ha quedado demostrado por medio de experimentos de calorimetría controlada realizados con

CRECIMIENTO DE LOS PECES 161
truchas y aves domésticas, que los peces son mucho mejores que las aves de corral en la formación de proteínas. Esto se debe a que las aves y mamíferos tienen que emplear parte de sus calorías en el mantenimiento del calor de su cuerpo y para mantenerse a sí mismos vivos; en contraste, los peces son poiquilotermos y obtie- nen su sustento del medio ambiente. Algunos aspectos especiales relacionados con el crecimiento y los factores que influyen en él serán discutidos más adelante.
Determinación de la edad y el crecimiento
Cuando los registros del aumento de longitud (o peso) de un pez son confrontados con el tiempo a lo largo de su vida, se obtiene la tasa de crecimiento que se repre- senta con una curva (Figura 5.10). Las fluctuaciones que aparecen con respecto al crecimiento a medida que transcurren las estaciones del año, son a menudo soslayadas y solamente en los periodos postembrionarios e, inclusive, en los post- juveniles, es cuando usualmente se emplean datos empíricos relacionados con el crecimiento.
Dado que muchos peces de importancia comercial sólo pueden ser medidos una o pocas veces en su ciclo de vida, el hombre ha procurado la manera de re- construir sus tasas y modelos de crecimiento. El procedimiento más útil con este propósito -por lo menos para los peces de las zonas templadas-, es el de las mar- cas anuales de las partes óseas (Capítulo 4), como las escamas, los otolitos, las espinas y los huesos operculares (Figura 5.11). Las zonas de crecimiento anual en estas partes se deben a la disminución durante el invierno de los procesos que dependen de la temperatura y a la reanudación de un metabolismo rápido en cierta época de la primavera. Algunos peces de agua dulce de los trbpicos, donde domi- nan los monzones, también muestran crecimiento relacionado con el cambio de estación, el cual está señalado por marcas de crecimiento que corresponden al final de las estaciones de sequía. Las zonas de crecimiento pueden ser medidas y, por lo tanto, puede ser establecida la correspondencia entre el crecimiento del cuerpo y el de las partes óseas. Por ejemplo, en ciertas regiones del pez las esca- mas crecen paralelamente al cuerpo (crecimiento isométrico), por lo menos en
Figura 5.10 - Curva del crecimiento en longitud como se observa en el "cisco" de aguas poco profundas (Coregonus artedii). (Basado en Hile, 1936). Z 300
.- E 250
2 200 a
"
2 3 4 5 6 7 8 9 1 0 1 1 1 2 Años de vida
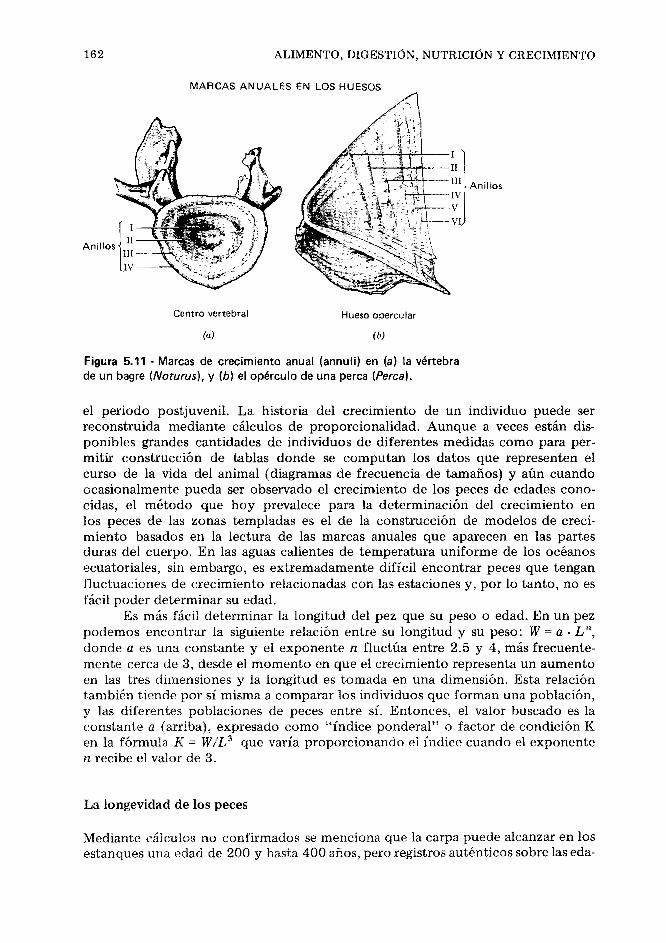
162 ALIMENTO, DIGESTION, NUTRICION Y CRECIMIENTO
MARCAS ANUALES EN LOS HUESOS R
Centro vertebral
(a)
J Anillos
Hueso opercular
( b )
Figura 5.11 - Marcas de crecimiento anual (annuli) en (a) la vertebra de un bagre (Noturus), y (b) el opdrculo de una perca (Perca).
el periodo postjuvenil. La historia del crecimiento de un individuo puede ser reconstruida mediante cálculos de proporcionalidad. Aunque a veces están dis- ponibles grandes cantidades de individuos de diferentes medidas como para per- mitir construcción de tablas donde se computan los datos que representen el curso de la vida del animal (diagramas de frecuencia de tamaños) y aún cuando ocasionalmente pueda ser observado el crecimiento de los peces de edades cono- cidas, el método que hoy prevalece para la determinación del crecimiento en los peces de las zonas templadas es el de la construcción de modelos de creci- miento basados en la lectura de las marcas anuales que aparecen en las partes duras del cuerpo. En las aguas calientes de temperatura uniforme de los océanos ecuatoriales, sin embargo, es extremadamente difícil encontrar peces que tengan fluctuaciones de crecimiento relacionadas con las estaciones y, por lo tanto, no es fácil poder determinar su edad.
Es más fácil determinar la longitud del pez que su peso o edad. En un pez podemos encontrar la siguiente relación entre su longitud y su peso: W = a - L”, donde a es una constante y el exponente FZ fluctila entre 2.5 y 4, más frecuente- mente cerca de 3, desde el momento en que el crecimiento representa un aumento en las tres dimensiones y la longitud es tomada en una dimensión. Esta relación t.ambién tiende por sí misma a comparar !os individuos que forman una población, y las diferentes poblaciones de peces entre sí. Entonces, el valor buscado es la constante a (arriba), expresado como ‘“indice ponderal” o factor de condición K en la fbrmula K = W/L3 que varía proporcionando el índice cuando el exponente I E recibe el valor de 3 .
La longevidad de los peces
Mediante cálculm no confirmados se menciona que la carpa puede alcanzar en los estanques una edad de 200 y hasta 400 anos, pero registros auténticos sobre las eda-

CRECIMIENTO DE LOS PECES 163
des de peces cautivos sugieren que la vieja carpa más venerada no llega a los 5 0 afios.
Es así como unos cuantos peces grandes, en estado silvestre, no pueden vivir más de 1 2 a 20 años e inclusive en un acuario, donde viven protegidos, rara- mmte exceden los 30 años. Generalmente se puede esperar que las especies repre- sentadas por individuos grandes lleguen a alcanzar mayor edad que las compuestas por individuos pequeños. La mayor parte de los peces cuya longitud llega a ser mayor de los 30 centímetros tiene un periodo de vida de por lo menos 4 o 5 años. Muchos pececillos de acuario (Cyprinidae) de las zonas templadas y otros pececi- llos de las zonas templada y tropical, con frecuencia tienen un periodo de vida menor a 2 años, incluyendo algunos que son de vida anual.
Factores genéticos (Capítulo 12)
La presencia de variaciones genéticas en el potencial de crecimiento en la naturaleza y en diferentes poblaciones de la misma especie, es usualmente enmascarada por los factores ambientales como la temperatura y la disponibilidad de alimento. Sin em- bargo, algunas razas selectas de la trucha de arroyo (Saluelinus fontinalis), la trucha arco iris (Salmo gairdneri) y la carpa (Cyprinus carpio) tienen un crecimiento que es decididamente superior al de otras razas.
El crecimiento del pez usualmente disminuye después de haber comenzado la maduración sexual, cuando grandes cantidades de material nutritivo son indu- cidas periódicamente a formar parte de los óvulos y el esperma. Las gónadas de la perca amarilla (Perca flauescens) forman más del 20% del peso del cuerpo de la hembra y más del 8% del peso del cuerpo del macho, antes de la temporada de reproducción; después de la puesta, los ovarios o los testículos se reducen a cerca del 1% o menos del peso del cuerpo.
El inicio de la maduración sexual y su consecuente cambio en la tasa de crecimiento en el ciclo biológico de un pez, puede también ser genéticamente determinado, como se ha hecho en la trucha café (Salmo trutta). El tamaño de los huevos también varía de acuerdo a las instrucciones genéticas y, como consecuen- cia, varía asimismo el tamaño inicial del pez al momento de nacer. Los huevos grandes producen larvas grandes que también pueden tener en el lote la mayor potencialidad de crecimiento. Las ventajas que el gran tamaño de huevo presenta como modelo de crecimiento, son detectables durante corto tiempo, mientras no intervengan otros factores como la disponibilidad de alimento; pero esas ventajas, sin embargo, perdurarán durante toda la vida.
Con frecuencia hay diferencias de tamaño entre los peces macho y hem- bra maduros, dentro de la misma especie. Las diferencias de tamaño entre los sexos, que afectan las tasas de crecimiento y los modelos de crecimiento, pueden deberse a la acción de factores genéticos, a menudo correlacionados con patro- nes de conducta heredados. Entre los pececillos de acuario (Cyprinidae) de Amé- rica del Norte, por ejemplo, la mayoría de las especies tienen hembras más grandes que los machos, pero en algunas especies, como el pez con cabeza cubierta de cuernos (Nocomis biguttrrtus) y los pececillos de acuario de cabeza grasa (Pime- phalinae), el macho es de mayor tamaño. Es muy posible que los hábitos rela- cionados con la reproducción, como la construcción de nidos, produjeran esas diferencias de tamaño entre los sexos. El mayor tamaño aparece más a menudo asociado con el padre más “importante”.

164 ALIMENTO, DIGESTI6N, NUTRICI6N Y CRECIMIENTO
Factores estacionales y de temperatura
Los peces que tienen una distribución latitudinal muy amplia, como el huro (Mi- cropterus salmoides), por lo común tienen diferencias en la tasa de crecimiento y en la edad en la cual alcanzan la maduración sexual, contrastando los casos qde ocurren en las aguas del norte, contra los de las aguas del sur, Por ejemplo, en las aguas de Lousiana y Arkansas, el huro o trucha de Pátzcuaro puede desovar después de un año, pero al sur de Michigan o en Wisconsin solamente lo pueden hacer a partir de los tres años como promedio. La maduración sexual de este huro puede ser retardada hasta el cuarto o quinto año en Ontario y otras lati- tudes y climas comparables. El tamaño y edad finales del pez pueden no diferir entre los representantes del norte y del sur de la especie, pero la forma de la curva que representa el registro del peso contra la edad en el modelo de crecimiento, reflejará una diferencia dependiente de la temperatura. La temperatura también induce a que haya diferencias estacionales en el crecimiento, desde el momento en que la temperatura afecta el metabolismo y el consumo de alimento.
La elevación estaciona1 de la temperatura de las aguas dulces coincide con un incremento del alimento natural para los peces, pero también hay variaciones en actividad que dependen de las estaciones y la temperatura, las que influyen en la cantidad de alimento que es desviado hacia los requerimientos relacionados con el crecimieto y el mantenimiento de las demás funciones biológicas, o sólo para alguno de los dos. En un experimento realizado por Brown (1957), la trucha café (Salmo trutta), creció óptimamente entre los 7" y 9"C, y también entre los 16" y 19°C. El apetito mejoró tan luego como el agua se calentó alos 10°C. Losre- querimientos para el mantenimiento también se incrementaron rápidamente entre los 9" y 11°C; luego su gráfica formó un dibujo parecido a una cinta y se volvió casi constante cuando se llegó a los 20°C. Por lo tanto, las temperaturas óptimas para lograr el crecimiento rápido son las que coinciden con un apetito alto y requerimientos de mantenimiento bajos, mientras que el crecimiento mínimo ocurre a temperaturas intermedias, cuando los requerimientos de mantenimiento son elevados porque el pez es más activo. Cuando la temperatura es menor a los 7°C los requerimientos de mantenimiento son bajos, pero también así de bajo es el apetito; sobre los 19°C el apetito vuelve a descender, mientras que los requeri- mientos de mantenimiento se mantienen al mismo nivel. Por lo tanto, la trucha café logra tener un crecimiento óptimo estando fuera del margen de temperatura en el que el pez muestra máxima actividad.
La luz, principalmente a lo largo del día solar, puede afectar la tasa y el modelo de crecimiento del pez. Desde el momento en que los cambios de la lon- gitud del día coinciden con la elevación o descenso estacionales de la temperatura, es difícil evaluar los respectivos efectos de estos factores ambientales. Se sabe que la iluminación actúa indirectamente en el metabolismo tiroideo y que los picos de la actividad tiroidea coinciden con la máxima duración del día, pero estos picos también incrementan la actividad natatoria, lo que resulta en una reducción de la tasa de crecimiento, según los experimentos que se han realizado con la tru- cha café (Salmo truttu) hacia mediados del verano. Si el alimento es abundante, los días cortos serán más provechosos para lograr el crecimiento rápido, debido a la existencia de modelos de descanso inducidos por la oscuridad; pero en donde hay fuerte competencia y la alimentación se realiza con el auxilio de la visión, los días largos pueden ser de utilidad para el pez.

CRECIMIENTO DE LOS PECES 165
Factores bióticos
Estos factores han sido llamados variables dependientes de la densidad, pero tam- bién es válido llamarlos con el término “social”. Entre ellos queda incluida la com- petencia, o sea, la influencia del número de individuos en la cantidad de alimento disponible, los sitios para anidar, la conducta de la alimentación, etc. Desde el momento en que los efectos de los factores bióticos son más estudiados en la naturaleza, es más fácil confundirlos con los efectos del medio ambiente (físicos y químicos), o sea, los factores independientes de la densidad. Son relativamente pocos los experimentos que se han realizado, en forma controlada, donde los factores dependientes de la densidad han sido segregados con éxito mediante es- crutinio, ya sea uno por uno o en combinación.
El efecto indirecto de pez sobre pez. Los experimentos realizados con la carpa dorada (Carussius auratus) que ha crecido en agua donde previamente ha vivido otra carpa dorada, han mostrado que ese pez experimental creció mejor que sus compañeros-control mantenidos en agua “no condicionada”. Puede haber suce- dido que el crecimiento mejorado se haya debido a la utilización de alimento regurgitado contenido en el agua condicionada. Más claramente, sin embargo, ello se debió a la presencia de una sustancia promotora del crecimiento que pudo haber sido aislada del fango contenido en los tanques con agua condicionada. Se encontró que la sustancia era efectiva para estimular el crecimiento en dilu- ciones tan bajas de concentración como 1.2 partes por millón (ppm).
Efectos directos de peces sobre peces (efectos de grupo). El orden de dominan- cia, u orden de picoteo, ha sido observado como factor de crecimiento entre los peces cautivos, donde se incluyen los huros (Centrarchidae), los peces semejantes a las truchas (Salmonidae) y los cíclidos (Cichlidae). Un individuo es el primero en el orden (a menudo el más grande) y generalmente asegura para sí el mejor bocado o la mayor cantidad de éstos, cuando el alimento es proporcionado al grupo. Cuando sólo hay unos cuantos peces en el tanque, estos efectos jerárqui- cos son a menudo rotos, pero hay suficientes indicios de que persisten en la natu- raleza. La trucha de lago (Saluelinus mmaycush), de los lagos y ríos del norte, entre otros peces, ha mostrado dominancia definida de un individuo sobre los demás. No se sabe todavía si las diferencias en el desarrollo se deban directamente al aumento del consumo de alimento como resultado del orden de picoteo, o indirectamente a través de la tensión psicológica que aparece en los miembros in- feriores del orden.
También se ha sugerido que puede haber un efecto de grupo en el creci- miento, por arriba y por abajo de la jerarquía entre peces considerados como individuos. La sugerencia se basa en el siguiente experimento. Cuando números diferentes de truchas fueron colocados en tanques de la misma medida y distribui- das en grupos de 25, 50, 80, 100 y 150 individuos, el grupo con 80 peces fue el que más sobresalió, cuando a cada grupo se le dio una sobresalimentación. Parece ser, entonces, que la tasa de crecimiento de los individuos fue influenciada por el tamaño del grupo, así como por el número presente de peces más grandes y más pequeños.
El más notable efecto de grupo en el crecimiento se encuentra en las pobla- ciones de peces aglomerados, Cuando el pez crece en condiciones de aglomeración

REFKRENCIAS ESPECIALES SOBRE ALIMENTOS, DIGESTION, NUTRIC16N Y CRECIMIENTO
hi-Itussaini, A.M. 1949. On the functional morphology of t,he alimentary tract of some fishes in relation t,o differences in t,her feeding habits. Anatomy and histology. Quart Jour Mirr. Sr¿., 90:109-139.
to differences in their feeding habits. Cytology and physiology. Quart. Jour. Micr. Sei., ” , 1949. On the functional morphology of the alimentary tract of some fishes in relation
90:323-354. eagenal, T.R., ed. 1.974. Proc. Intern. Symp. Aging o f Fish. Unwin Bros., Inglaterra. Barrington, E.J.W. 1957. The alimentary canal and digestion. The physiology o f fishes,
Brown, M.E. 3957. Experimental studies on growth. The physiology of fishes, 1:361-400.
Gerking, S.D. 1954. Food turnover of a bluegill population. Ecology, 35(4):490-498. Gregory, W.K. 1933. Fish skulls. Trans. Amer, Phil. Sor . , 23(2):75-481. (Reimpresión en
offset por Eric Lundberg Pub]., Laurel, Fla., 1959.) Halver, J.E. 1972. Fish nutrition. Academic Press, Nueva York. Hile, R. 1941. Age and growth of the rock bass, Arnbloplites rupestris (Rafinesque), in Nebish
Ivlev, V.S. 1961. Experimental ecology o f the feeding of fishes. Yale University Press, New
Love, R.M. 1970. Chemical bilogy o f fishes. Academic Press, Nueva York. Phillips, A.M., Jr., 1969. Nutrition, digestion, and energyutiiization. Fishphysiology, 1:391-432.
nicker, W.F. 1946. Production and utilization of fish populations. Ecol. Monogr., 16:373-391. Vonk, H.J. 1927. Die Verdauung bei Fischen. Zeitschr. Vergl. Physiol. , 5:445-546.
1:109-161. Academic Press, Nueva York.
Academic Press, Nueva York.
Lake, Wisconsin. Trans. Wisconsin Acad. Sci., Arts and Let t . , 33~189-337.
Haven, Conn.
Academic Press, Nueva York.

Capítulo 6
Esqueleto, estructura e 1
y movimiento
La forma del cuerpo y la locomoción son en los peces el resultado de la interacción del esqueleto y la musculatura. Con el fin de adaptarse a la multitud de hábitats que se han formado en el mundo acuático, se ha desarrollado una gran variedad de formas corporales, con su correspondiente modificación del esqueleto y los músculos. Y el modelo básico de locomoción por serpenteo, que complementa la adaptación de la forma del cuerpo, también se ha diferenciado en mucho otros tipos que, aunque están especializados, son de menor significación.
ESQUELETO
Las características superficiales del esqueleto interno y externo de los peces ya han sido descritas, y ahora vamos a considerar algunos aspectos del esqueleto como principio básico de la forma y el movimiento de los peces, junto con las tendencias adaptativas y evolutivas relacionadas.
Funciones
Una de las funciones del esqueleto es la de proporcionar soporte a las partes y, en consecuencia, determinar indirectamente la forma de los peces, como ya se ha señalado. Otra función es la de dar ventaja mecánica, y posiblemente la de tomar un papel importante en la formación de los glóbulos rojos (hematopoiesis). Ade- más, en algunos peces hay modificaciones de las aletas que permiten la colocación del esperma en el tracto reproductivo de la hembra. Y en este caso también el esqueleto de la aleta ha sufrido una especialización.
167

168 ESQUELETO, ESTRUCTURA Y MOVIMIENTO
Simetría
El plan estructural básico que presenta la organización esquelética de los peces consiste en una simetría bilateral fundamental; esto es, que los lados derecho e izquierdo de un individuo son realmente imágenes especulares uno respecto al otro. Aunque de ese plan estructural del cuerpo de los peces (como sucede en otros vertebrados) hay algunas excepciones principales como lo son ciertos órganos digestivos, la simple mención de unas cuantas desviaciones esqueléticas puede servir otra vez para ilustrar la gran variación que se encuentra en los detalles ana- tómicos de los peces. Un ejemplo clásico de la desviación de la simetría bilateral de los vertebrados es la de los lenguados (Pleuronectiformes). Tanto las fosas orbitales como los ojos están situados en un lado, a donde el ojo del lado ciego ha migrado a lo largo de los sucesivos estados de crecimiento, desde una larva de simetría bilateral hasta el adulto asimétrico.
Mandíbulas y cara
El sistema esquelético de los peces ejemplifica muchas de las adaptaciones y ten- dencias evolutivas. Un grupo de estas tendencias y adaptaciones se relaciona con la formación de las mandíbulas y sus soportes y, por consiguiente, determina en gran parte la apariencia facial. Los peces ágnatos no poseyeron verdaderas man- díbulas, pero desarrollaron por primera vez algunas estructuras esqueléticas para sostener las branquias. Más tarde, durante el proceso de la evolución las mandíbu- las de los peces fueron modificadas a partir de los componentes del arco branquia1 anterior que poseían sus ancestros amandibulados. De manera similar, también se ha propuesto que el arco visceral o faríngeo que sigue al que dio lugar, a través de su especialización, a las mandíbulas, se transformó en el apartato hioideo de los peces modernos. El arco hioideo sostiene las mandíbulas, la lengua y los arcos branquiales en casi todos los peces mandibulados.
Las mandíbulas de los peces se han transformado en muchos tipos diferen- tes, tomando en consideración su posición y los detalles de su estructura; todo aparentemente en relación con sus hábitos. Una variación extrema de la condición general donde la boca es mandibulada y terminal, corresponde al aparato suctor ventral u órgano de fijación formado por la boca de esos peces, como algunos de los bagres armados (Plecostornus y otros géneros), cuyos labios gruesos y protrác- tiles situados en las mandíbulas verdaderas forman aparatos succionadores eficien- tes. Frecuentemente las mandíbulas son inferiores (ventrales), especialmente entre los peces que se alimentan en el fondo, como las rayas (Rajidae; Figura 2.8) y los esturiones (Acipenseridae; Figura 2.13). Otros peces tienen mandíbulas superiores (dorsales), adaptadas para la alimentación en la superficie del agua, o para comer en dirección hacia arriba. Las bocas son superiores en la mayoría de los “top- minnows” (Cyprinodontiformes; Figura 2.33), incluyendo a las gambusias (Gurn- bush) que se alimenta efectivamente de los maromeros, o sea, las inquietas larvas de mosquito, en la superficie del agua. Los “anglerfishes” (Lophiiformes; Figura 2.50) presentan mandíbulas situadas en el dorso que les permiten descansar en el fondo y capturar las presas desde arriba.
En algunos peces, como las agujitas (Hemiramphidae; Figura 5.5c), las mandíbulas inferiores están muy alargadas. Esta modificación se relaciona también

ESQUELETO 169
con el hábito de la alimentación en la superficie del agua, como ya se describió. En otros peces, la mandíbula superior se ha alargado para formar un pico o un rostro largo y aplanado. Los peces espada (Xiphiidae) y los merlines y peces vela (Istiophoridae) de los océanos, así como los peces espátula (Polyodon; Figura 5.5g) de las aguas continentales norteamericanas, muestran este tipo de elonga- ción, pero es difícil asignarle una función relacionada con la alimentación más o menos fácil. Por ejemplo, el pez espada (Xiphias) y otros peces con una lanza similar, aparentemente no hieren a menudo a su presa, pero se sabe de casos en los que han atacado las estructuras de los cascos de los botes de madera. Los peces sierra (Pristis), sin embargo, mutilan a su presa mediante golpes dados con los bordes dentados de su rostro alargado. En el pez espátula (Polyodon) la cara se ha originado de un órgano que protege gran número de dispositivos sensoriales que posiblemente sirven para detectar poblaciones de plancton, las cuales son recogidas mediante colado por los rastrillos que en gran número tiene el pez, mientras éste nada lentamente hacia adelante y hacia atrás por entre el plancton, y con la boca desmesuradamente abierta. Es posible que la “espátula” sirva como órgano estabilizador o cucharón, o ambas cosas, cuando el pez nada con la boca abierta y se alimenta del plancton.
Incluso existen peces que tienen muy alargadas las mandíbulas superior e inferior. Entre éstos podemos contar a los peje-lagartos (Lepisosteidae; Figura 2.15) de las aguas dulces de América del Norte. También aquí podemos incluir a los peces aguja oceánicos (Belonidae) y a los peces mariposa de nariz larga (Forci- piger; Figura 5.5f), que tienen un pico alargado provisto de mandíbulas que, como pinzas en el extremo, son usadas para recoger pólipos pequeños e invertebrados diminutos que se esconden entre los corales. Concluimos, entonces, en que la forma de la cabeza y las mandíbulas determina muchas de las cosas que un pez puede o no hacer en lo referente a su alimentación y a la defensa o el ataque durante el combate.
Aletas
Podemos considerar a las aletas de los peces en dos grupos importantes: medias y pareadas. Tanto la forma como la posición de estas aletas se relacionan con la forma del cuerpo y la localización del centro de gravedad del pez (y por consi- guiente también se relacionan con la ubicación y forma de la vejiga gaseosa, los hábitos y el carácter y velocidad de la natación).
Orígenes. Se ha teorizado que las aletas de los peces se originaron todas de ances- tros que no las tenían. Es de presumir que para el pez era ventajoso responder a la turbulencia del agua para lograr la estabilización, junto con los problemas de esta- bilidad durante la natación; de lo anterior resultó la aparición de un borde medio o quilla que se desarrolló en el cuerpo, desde cerca de la cabeza hasta el ano, pa- sando por una porción terminal de la cola. Este borde, que posiblemente en un principio no tuvo radios que lo soportaran, dio lugar a la aparición de las aletas medias. Bordes similares aparecieron en los lados y luego se coligaron precisa- mente en los sitios donde se presume que aparecieron los antecedentes ancestrales de las actuales aletas pares. Los dobleces de las aletas del anfioxo (Bmnchiosto- ma), posiblemente uno de los ancestros de los vertebrados, y los dobleces de las

170 ESQUELETO, ESTRUC>TUHA Y MOWMIENTO
aletas que aparecen en muchos embriones de pez, tienden a dar credit0 a la teoría ya citada de los dobleces en las aletas. Se puede esperar, sin embargo: que haya peces sin aletas, y así sucede en efecto en peces que inclusive tienen hábitos especializados (por ejemplo, la anguila simbránquida, Typhlosym branchus, que tiene forma de gusano y cava en el fango).
Diferenciación. Las diferencias que se pueden observar en las aletas medias y pares de los peces, muchas de las cuales parecen poseer funciones adaptativas para la locomoción y las diversas maniobras que el pez debe hacer, han resultado de las va- riaciones del propio esqueleto. Tanto el esqueleto intern9 de la aleta caudal como algunas de sus modificaciones, han sido ya descritos. Más superficialmente, el lóbulo inferior de la aleta caudal está reducido en tamaño en muchos peces cava- dores del fondo marino, como sucede en las rayas (Rajiformes; Figuras 2.8 y 6.2), pero está alargado en los peces voladores (Exocoetidae) que pueden usar la cola como un órgano impulsor en la superficie del agua, justamente antes de salir al medio atmosférico. Y todavía en otros, la aleta caudal original está sustituida por ios radios procedentes de las aletas dorsal y anal, que han tenido un crecimiento exagerado en dirección caudal (como en el bacalao, Gadus; Figura 2.29); con poca frecuencia está ausente, como se puede ver en los parientes de la anguila (Anguilliformes), como Ophisurus y Sphugebrunchus, y en el caballito de mar (Hippocampus; Figura 10.6). Y todavía más rara aún es la ausencia de las aletas caudal y anal, como se observa en Gymnurchus (Mormyridae).
Las aletas dorsales varían mucho en lo referente al soporte y, en consecuen- cia, en el tamaño, extensión y localización. La aleta puede faltar (Xenomystus), ser sólo rudimentaria (Notopterus) o extenderse a casi todo lo largo del dorso, como en el amiído Amiu (Figuras 2.14 y 6.2), la doradilla (Coryphuem), los peces cinta (Trachipteridae) y los peces remo (Regulecus). O la aleta puede continuarse con la caudal y la anal, como en las lampreas (Petromyzonidae; Figuras 2.1 y 2.5), las anguilas de agua dulce (Anguillidae; Figura 2.25) y las anguilas falsas (Sym- branchidae). Es usual, para un pez de radios espinosos, tener dos aletas dorsales. Algunos peces de aguas templadas (Percichthyidae; Figura 2.41) y los pércidos (Percidae) presentan muchos buenos ejemplos de esto. Sin embargo, algunas espe- cies tienen tres o más aletas dorsales. Los bacalaos (Gadus; Figura 2.29), por ejemplo, tienen tres. La condición múltiple se reduce finalmente a una serie de pequeñas aletas más o menos separadas y aisladas (espinas separadas con su propia membrana) en la espinocha (Gasterosteidae; Figura 2.30) y en los poliptéridos (Polypterus; Figura 2.12). En algunos peces, como en las truchas y sus parien- tes (Salmonidae; Figura 6.6) y en los bagres neárticos (Ictaluridae), una de las aletas dorsales no tiene radios y está transformada en una estructura carnosa lla- mada aleta adiposa.
Entre las modificaciones poco usuales de la aleta dorsal está la elongación de sus últimos radios en los peces saltadores como el sábalo (Megalops) y la sardi- nita (Dorosom). Está fuertemente modificada en la rémora o chupador de tibu- rones (Echeneidae; Figura 2.46), al comprender el disco de succión con el cual este famoso polizonte se fija a otros peces, a tortugas marinas y a los barcos. En los lófidos (Lophiiformes; Figura 2.50) la aleta dorsal proporciona los elementos para la atracción de la presa, el llamado “bastón y señuelo” que adorna la parte superior de la cabeza. Solamente en unos cuantos casos, muy raros, llega a estar la aleta dorsal muy reducida en tamaño o francamente ausente, como se ve en las

ESQUELETO 171
anguilas gimnótidas de América del Sur (Gyrnnotidae; Figura 6.2b), en algunas rayas (Trygonidae) y en las rayas gavilán (Myliobatidae).
La aleta anal es, posiblemente, entre las aletas medias, la más estable, porque es la que muestra menos variaciones. En todas las familias de peces de agua dulce de América del Norte, a excepción de una, la aleta anal es de ordinario uniforme; sirva lo anterior como un ejemplo. En los machos de los pececillos viviparos (Poeciliidae) sirve, sin embargo, como un órgano intromitente llamado gonopodio (Figura 10.5b). Esta estructura es usada para guiar a los espermatozoi- des (ya sea como esperma o como pelotas de esperma, llamadas técnicamente espermatóforos) del vientre del macho al de la hembra, para realizar la fertiliza- ción interna durante la época de procreación. Muchos peces también muestran diferencias sexuales de la aleta anal, con respecto al tamaño; uno de ellos es el succionador blanco (Catostornus comrnersoni). La aleta anal falta en algunos peces, como en las rayas modernas y sus parientes (Rajiformes).
Las aletas pectorales de los peces son un poco más variables que la aleta anal ya descrita. Y tienen como principal función el maniobraje durante la loco- moción. Algunos ejemplos de la función locomotora primaria de las aletas pec- torales pueden encontrarse en los pércidos (Etheostomatinae), las espinochas (Gasterosteidae), “sculpins” (Cottidae) y los peces pipa y caballitos de mar (Syng- nathidae), así como en los peces loro (Scaridae), los sargos (Sparidae) y otros peces que tienen la cintura pectoral reforzada y, por consiguiente, modificada. Por lo menos un pececillo (Gambusia) tiene la aleta pectoral modificada en forma de clásper, que el pez macho emplea para guiar su gonopodio hacia el vientre de la hembra. Otras variaciones de las aletas pect,orales incluyen las relacionadas con el tamaño; comúnmente son de mayor tamaño en los peces cavadores de los fon- dos marinos como los “darters”, “sculpins” y rayas (Rajidae). Están demasiado agrandadas en los peces voladores (Exocoetidae), porque las usan para sostenerse en el aire durante la planeación. Faltan en las lampreas vivientes (Petromyzoni- dae), como resultado de una variación extrema.
Además de ser empleadas en la locomoción, en algunos peces las aletas pectorales están adaptadas en forma de una estructura para el ataque y la defensa. En los bagres de agua dulce (Ictaluridae) de América del Norte el radio endure- cido y filoso que se halla en el borde delantero de estas aletas está articulado al cuerpo por medio de una estructura como bisagra que le permite al pez levantarlo y dejarlo en posición erecta, posiblemente como un instrumento de combate. En los machos malos (Noturus), este radio espinoso tiene una glándula especial en su base. La secreción de la glándula, inyectada por la espina como si fuese una aguja, tiene un fortísimo efecto doloroso y paralizante en el hombre.
Se considera que las aletas pélvicas son más bien órganos accesorios para facilitar las maniobras durante la natación, si consideramos a las pectorales como las que desarrollan el principal papel en este aspecto. Las aletas pélvicas están ausentes, desde luego, en algunos peces (lampreas, Petromyzonidae; anguilas, Anguillidae; ciertos “killifishes”, Cyprinodontidae-Orestias, Tellia; cabezas de víbora, Channa); en otros está sólo reducida a una espina (espinocha, Gasteros- teidae; Figura 2.30) o a un simple manojo de unos cuantos radios en muchos otros peces (pez vela, Istiophoridae). Ocasionalmente las aletas pélvicas no apa- recen en un pez debido a una anormalidad individual en el desarrollo. En Etheos- toma microperca, sin embargo, las aletas pélvicas son de mayor tamaño en los machos y constituyen un notable carácter sexual secundario, aunque no se le

172 ESQUELETO, ESTRUCTURA Y MOVIMIENTO
ha encontrado una función biológica determinada. Los tiburones machos tienen las aletas pélvicas modificadas en forma de estructuras ganchosas, llamadas mixop- terigios. Cada mixopterigio tiene tres partes principales (Figura 10.5a) además de la parte con radios. Los ganchos (“claspers”) funcionan, como los gonopodios, guiando los espermatozoides hacia el tracto reproductor de la hembra. Algunas variaciones extremas de las aletas pélvicas pueden ser posiblemente mejor expli- cadas, dando como ejemplo el disco suctor que forman en algunos peces, como los de las corrientes fluviales de las montañas de la India (GZyptosternurn) y algunos de los góbidos (Gobiidae), que habitan las zonas agitadas de las aguas costeras en todos los continentes del mundo.
Hay una característica común de las aletas pélvicas de los peces provistos de radios espinosos, y consiste en que cada una posee sólo una espina. El número de radios que aparece en las aletas pares es más constante que el de las aletas me- dias o verticales. Esto explica cómo los taxónomos nunca encuentran aquí las suficientes características para separar las especies o los grupos más pequeños de peces, pero sí las logran para formar las categorías más elevadas. Por ejemplo, la fórmula típica de la aleta pélvica en los peces de radios espinosos es una espina y cinco radios blandos (I$), y a partir de ella por lo general solamente se encuen- tran variaciones menores.
Radios de las aletas
En los peces óseos (Osteichthyes) las estructuras que sostienen en posición a las aletas pueden ser divididas en dos clases principales (Figura 6.1): espinas (radios únicos) y radios blandos (radios segmentados). Las espinas de las aletas de los pe- ces son estructuras uniseriadas y sin segmentar, como se ve en la primera aleta dorsal de los meros marinos o serranos (Serranidae) y la perca y sus parientes (Percidae), entre otros. Los radios blandos son típicamente segmentados, a me- nudo ramificados, y siempre biseriales (dos componentes laterales pareados en la línea media); pueden estar presentes en cualquier aleta, pero siempre forman parte de todas las aletas caudales de los peces óseos (Osteichthyes). El carácter de los radios de las aletas, ya fuesen blandos o espinosos, muy pronto dio lugar al nacimiento del concepto de dos grupos correspondientes de peces óseos, los Malacopterygii y los Acanthopterygii, respectivamente. Como representantes de las familias de peces pertenecientes a la fauna de agua dulce de América del Nor- te, y con radios exclusivamente blandos, tenemos al esturión (Acipenseridae), el pez espátula (Polyodontidae), pez de aleta ondulada (Amiidae), el peje-lagarto (Lepisosteidae), los clupeidos (Clupeidae), los osméridos (Osmeridae), las tru- chas (Salmonidae), el pez ojo de luna (Hiodontidae), el “mudminnow” (Umbri- dae), el lucio (Esocidae), la carpa hocicona (Catostomidae), la carpa común (Cy- prinidae), los bagres de agua dulce de América del Norte (Ictaluridae), la anguila (Anguillidae), el “killifish” (Cyprinodontidae), los pececillos vivíparos (Poecilli- dae) y el bacalao (Gadidae).
De todos los ejemplos citados anteriormente, la única excepción se nos pre- senta cuando nos referimos a las familias de la carpa y el bagre. En estos dos grupos, algunos de los radios blandos se han fusionado desde el estado embriona- rio del pez para dar lugar a un radio espinoso (que no corresponde morfológica- mente a un radio espinoso como anteriormente se describió). La carpa (Cyprinus)

ESQUELETO
RADIOS DE LAS ALETAS
Lado Frente Lado Frente Lado (a) Radio blando ( 6 ) Radio blando espinoso (e) Espina
Frente
Figura 6.1 - Radios de aleta de peces óseos: (a) radio blando y (6) radio blando espinoso de la aleta anal de una carpa (Cyprinus carpio) donde en a y 6 muestran simetría bilateral y solamen- te en a ramificación y segmentaci6n; (c) espina anal del pez tambor de agua dulce (Aplodinotus grunniens) demostrando que es un elemento medio, Único.
y el pez dorado o carpa dorada (Carassius), presentan sendos manojos de esos radios originalmente blandos pero ahora endurecidos, en las aletas dorsal y anal. Los bagres de agua dulce de América del Norte tienen un radio espinoso en el borde anterior de cada aleta pectoral.
Para describir brevemente los elementos que sostienen en posición a las aletas de los grupos más numerosos de peces, consideraremos a los Agnatha como uno de ellos, y después a los Chondrichthyes y Osteichthyes como otros.
En las lampreas y los mixinos (Cyclostomata), los soportes de las aietas aparecen como radios cartilaginosos y sin segmentar. Estos radios se conectan a través de tejidos membranosos esqueléticos a la columna vertebral aún imper- fecta y sostienen débilmente a las aletas medias.
En los tiburones y sus parientes y en los peces óseos las aletas pares y las medias tienen soportes internos esqueléticos (Capítulo 3) y radios de aleta dérmi- cos. Los radios de aleta dérmicos son llamados, en conjunto, dermatotriquia. Los dermatotriquia son generalmente de tres clases: a) ceratotriquia: córneos y a menudo sin articular ni ramificar, se les encuentra en los tiburones y sus pa- rientes; b ) actinotriquia (Figura 6.1): radios córneos con forma de espina, se desarrollan, por lo menos embrionariamente, en las aletas de todos los peces óseos y pueden persistir como las espinas en los peces de radios espinosos (Acan- thopterygii), o c) lepidotriquia (Figura 6.1): que en los peces de radios blandos sir- ven para reponer a los actionotriquia, o que en las aletas de radios blandos tam- bién pueden formar parte de las aletas de los peces de radios espinosos. Los lepidotriquia son radios blandos que, como ya se indicó anteriormente, están típicamente ramificados y unidos y su desarrollo es más superficial que el de los radios espinosos. Se puede pensar de ellos que han adquirido sus compo-

rientes superficiales de los elementos escamosos. Es casi asombroso ver qu i corno hay una migración embrionaria de las escamas sobre las espinas embribnicas, para cubrirlas y, finalmente, terminar por reemplazarlas. El término camptotri- quia ha sido propuesto para los radios de las aletas de los peces pulmonados (Dipnoi), de los que se ha pensado que difieren microscópicamente de los de las aletas de todos los demás grupos de peces.
Para la clasificación de los peces es de mucha utilidad conocer el número de los radios, particularmente de los de las aletas verticales. Los radios de las aletas dorsal y anal son llamados merísticos (de acuerdo al número de hileras de esca- mas que se encuentran a los lados del cuerpo) (Capítulos 10 y 13) y su número está relacionado con el número de segmentos embrionarios del animal en des- arrollo. La enumeración de los radios de aleta (y otros elementos merísticos) ha sido de mucha utilidad para la caracterización de la especies y en el estudio de los problemas relacionados con las subespecies y las razas.
Singularidades
Son numerosas las singularidades del esqueleto que se han llegado a encontrar. Algunas han aparecido en s610 una especie mientras que otras caracterizan a un grupo entero. Por ejemplo, en Amia hay una placa gular ósea, ubicada bajo la barba y entre los dos lados (rami) de la mandíbula inferior; en unos polípteros (Polypterus) existe un par de esas placas gulares. En los peces salmónidos (Salmo- ninae, Coregoninae, Osmeridae) hay unas protuberancias parecidas a espolones en los ángulos (axillae) de las aletas pélvicas. En Amia hay una protuberancia similar (es la protuberancia gular) en cada lado de la garganta. No se les ha encon- trado hasta hoy, a esas dos estructuras divergentes, una significación funcional.
Entre los casos de anormalidades del esqueleto quedan incluidas las anoma- lías del desarrollo y las patológicas. Estas aberraciones estructurales son menos frecuentes en la naturaleza que en los peces que se crían en los viveros piscícolas; porque muchas de estas deformaciones no les permiten sobrevivir. A menudo, esas anormalidades impiden a los peces tener la motilidad necesaria para poder escapar de sus predadores. Entre esas condiciones se encuentran la duplicidad cefálica, la reversión del patrón o modelo de escama, la ausencia de una o más aletas, una flexión sigmoidea de la columna vertebral, etcétera.
LOCOMOCIbN
Nuestro interés se centra aquí en la locomoción que permite que haya desplaza- miento o progresión, Esta locornocibn puede ser pasiva o activa.
Recorrido pasivo
Los huevos y crías pelágicos de muchos peces son a menudo transportados pasiva- mente por las corrientes. Esto sucede err los océanos, lagos y arroyos donde esos huevos y los pequeños son realmente parte del plancton y a menudo aparecen en las muestras de la fauna que se obtienen con las redes. Los peces viviparos han encon-

trado cn el vie:lt,re rle l a madre un buen vehículo pasivo para sus pequeños (Fi- gura -10.6). Puede considerarse que ha~7 tambikn wtxe los peces algiín medio de transport,c: protector para los iOvenes, dcspu&a de SU nacirnicr,to O la eclosión de huevos, rnpdiante la proteccibn temporal representatla por la ~onsC~aCi6 )n temporal en la boca de uno de los padres, cornu se ha visto que sucedo en algunas tilapias ( c o r n o en Tzlapia nmwamblea), !os bagres (suborden cipriniforme, Silu- roidea; por ejemplo, el bagre nektico cafi., Ictalirus nebulasus), las espinochas (Gasterosteidaej y otras especies que protegen a sus crias con mucho celo. Un ma.rsupio o bolsa para Ia cria (Figura 10.6) es tambikn una especie de cabina donde los pcquellos iladan y se desplazan, como sucede en los peces aguja (Syng- ~athu.s) y los cabaliitos de mar (Hippocampus).
El viaje pasivo puede ser realizado tambittn mediante la fijación a otro organismo u objeto. Los peces fijadores más renombrados son los succionadores marinos como las rémoras (Echeneidae). que se adhieren a los tiburones y otros peces, las tortugas de mar, e inclusive a las ballenas, sin producirles dario. El ór- gano de fijación está situado arriba de la cabeza y es una modificación de la aleta dorsal. Los peces fijadores más extravagantes son los machos parásitos de ciertos pejesapos (Ceratiidae) que se fijan por la boca a la superficie del cuerpo de las hembras (Figura 10.4). Los parásitos adultos de muchas clases de lampreas (Pe- tromyzonidae) también son acarreados por sus huéspedes. La mayoría parasita solamente a los peces, pero la lamprea del Pacífico (Lampetra tridentata) tam- bién ataca a las ballenas. Se sabe que la lamprea marina (Petrornyzon marinus) se fija por sí misma al casco de los botes. Con respecto a la anguila del fango (Simenchelys parasiticus) y algunas de las anguilas culebra (Pisodonophis, Ophich- thidae), se sospecha que también son transportadas por el huésped durante la alimentación parasitaria.
Una clase muy poco usual de transporte pasivo entre los peces ocurre cuando las masas acuriticas que conOienen peces son levantadas al aire por fuertes corrien- tes de viento, produciendo columnas de agua en forma de chorro. Los peces caen a tierra más tarde como si fuesen lluvia. Hay registros auténticos de estas lluvias, aunque el método seguido para obtenerlos parece ser hasta cierto punto impro- bable. No hay registros conocidos del transporte de huevos de pez por las aves acuáticas; sin embargo, se ha observado que algunas aves devoradoras de peces, como las garzas (ardeidae), han dejado caer de sus picos peces vivos mientras van volando.
El hombre es responsable del considerable transporte pasivo de peces en la tierra, el mar y el aire. Las actividades piscícolas envuelven inevitablemente en todos los continentes el desplazamiento de especies indígenas. Y, todavía más, en la mayoría de los continentes han sido introducidas especies exóticas, algunas veces teniendo su origen en la región de las antípodas.
Natación
Los movimientos más característicos de los peces se efectúan por medio de la natación. Estos movimientos se realizan para la alimentación y la reproducción, para el ataque y la defensa y para el transporte activo. Tienen como consecuencia las capturas comerciales hechas en la gran variedad de dispositivos estacionarios distribuidos por todo el mundo, donde se aplica de diversas maneras el encierro de

176 ESQUELETO, ESTRUCTURA Y MOVIMIENTO
los peces. h t o s pueden nadar para hacer largos viajes de hasta 1,000 kilómetros y aún más.
FORMA DEL CUERPO Y LOCOMOCI~N
La forma del cuerpo es en los peces el resultado de la interacción entre el esque- leto, la masa muscular y la adaptación evolutiva de estos dos sistemas a los sistemas de vida individual de estos animales. Las adaptaciones se han realizado, pri- meramente, para lograr varias clases de natación, aunque algunos peces también se han adaptado a la locomoción limitada en la tierra.
Como previamente hemos visto, la forma generalizada del cuerpo es fusi- forme en la mayoría de los animales acuáticos de natación libre, sean ellos peces, mamíferos o invertebrados. Esta forma es una adaptación al sentido de la corriente para el movimiento a través de un medio fluido. La redondez de la porción ante- rior y el adelgazamiento en forma de cinta de la porción posterior disminuye la oposición del fluido. Además, en la mayoría de los peces el cuerpo está cubierto por una sustancia mucosa que reduce el frenado al atenuar las irregularidades del cuerpo y reducir la viscosidad.
Si hemos hecho la suposición de que la forma primitiva del cuerpo de los peces es más o menos fusiforme, las desviaciones de esta forma tipo vienen a ser adaptaciones a los modos especializados de vida que llevan esos animales. Muchos peces pelágicos todavía retienen formas muy parecidas a la fusiforme ideal (Fi- gura 3.1), como en los nadadores rápidos: los atunes y las macarelas (Scombridae) y muchos tiburones como la tintorera (Carcharhinidae). La forma ideal de los peces diverge según tres direcciones: por compresión, depresión y elongación. Cada una de estas tres desviaciones se puede extender, a su vez, hacia adaptacio- nes extremas.
La forma del cuerpo comprimida lateralmente (Figura 3.1) prevalece en los hábitats de aguas quietas y con cobertura relativamente densa. Muchos habitan- tes típicos de los fondos herbáceos de los lagos, estanques y partes elevadas de los ríos son de esta naturaleza. Quedan incluidos en este grupo las mojarritas (Lepornis), y las tilapias africanas (Tilapia), algunos ciprínidos del Viejo y Nuevo Mundo, como el “bream” europeo (Abramis) y el brillador dorado (Noternigonus), respectivamente, y muchos cíclidos, como los peces ángel del Amazonas (Ptero- phyllum). También es común entre los habitantes de los “bosques” de coral en los arrecifes tropicales marinos, como sucede por ejemplo con los peces mariposa (Chaetodontidae) y muchos otros. Sin embargo, la fuerte compresión lateral en la forma del cuerpo, con la utilidad que ello presta para la rápida maniobra en los regresos violentos, de ninguna manera está confinada a los peces que viven bajo la cobertura densa. Esto sucede frecuentemente, como severo contraste, entre los peces de mar abierto y los gregarios, como los arenques y las sardinas (Clupeidae). Los habitantes de los “pastizales” marinos son, sin embargo, relativamente más cortos y menos comprimidos que los arenques. Los peces sol o molas de las aguas oceánicas abiertas (Molidae), también están aplanados de lado a lado pero nadan empleando pares de aletas medias. Los peces planos (Pleuronectiformes), que son batipelágicos par excellence, son, entre los peces, los que muestran con mayor exageración la compresión lateral.
Sin embargo, es la depresión del cuerpo la que prevalece generalmente entre

ALETAS Y LOCOMOCIdN 177
los habitantes de los fondos. Las rayas y especies afines (Rajiformes), que son altamente silenciosas, no solamente descansan a menudo en e l fondo, también ex- cavan en 61 a poca profundidad ayudándose con mucha ventaja de su cuerpo extraordinariamente plano. Entre los peces óseos, este tipo de forma aplanada está tipificada por dos de las familias más especializadas del grupo: los lófidos (Lophiidae) y los peces murciélago (Ogcocephalidae). El aplastamiento dorso- ventral en diversos grados es característico de muchos peces de los arroyos, como un medio ostensible para resistir el empuje de la corriente y mantenerse en posi- ción, así como de los que se protegen colocándose debajo de las piedras y otros objetos en el fondo. Como ejemplos comunes de ese aplastamiento, principal- mente de su región cefálica, tenemos a los cótidos (Cottidae) del Hemisferio Norte y a los bagres (ciprinimorfos del suborden Siluroidea), de muy amplia dis- tribución en todos los continentes.
Las pérdidas de la eficiencia natatoria debidas a la depresión del cuerpo son posiblemente compensadas por las ventajas obtenidas con la imitación mi- mética del fondo, la eficiencia de ciertos hábitos de reserva o por otras adapta- ciones adquiridas para vivir en el fondo. Hay modificaciones extremas de la forma del cuerpo que son acompañadas por la pérdida de la motilidad y abundan entre los peces. Podemos citar como ejemplos a los peces globo (Tetraodontidae) y los peces erizo (Diodontidae), asi como al que imita una hoja (Monocirrhus; Figura 4.6) y los caballitos de mar, entre los que incluimos al dragón marino, con pro- longaciones en forma de alga (Phyllopteryx; Figura 4.6), los caballitos de mar mismos y los peces aguja (Syngnathidae; Figura 10.6) y los peces cofre (Ostra- ciidae).
ALETAS Y LOCOMOCIdN
Aunque las aletas son partes muy características de los peces, muchas especies pueden prescindir con éxito de ellas. Y todavía más: los individuos a quienes se les han quitado todas las aletas pueden nadar eficientemente, aunque sea a bajas velocidades. Su habilidad para iniciar con violencia el movimiento (aceleración) es obviamente dañada. Con este daño su competencia como predadores se ve dis- minuida, y aumentada su susceptibilidad como presa. A pesar de esto, las aletas son importantes para lograr la locomoción del pez. Un gran número de peces puede desplazarse o hacer movimientos especiales solamente mediante el empleo de sus aletas, aunque la mayoría tiene que hacer uso de los movimientos del cuerpo para poder nadar mejor. En la mayoría de los peces, la aleta que tiene más importancia es la caudal para la natación a velocidad baja o sostenida, pero cuando se quiere obtener muy altas velocidades o una actividad que podríamos llamar “explosiva”, el empleo de esa aleta es imprescindible. Unas cuantas clases de peces muy especializados se desplazan ayudándose únicamente por sus aletas, como, por ejemplo, los caballitos de mar (Hippocampus) y los peces cofre (Ostraciidae).
De las variadas clases de aletas, todas, a excepción de las pélvicas, son em- pleadas por algunas especies como órganos primarios para la natación (Figura 6.2), aunque no necesariamente para obtener los impulsos más fuertes de los que el pez es capaz. La aleta dorsal de base larga de Amia puede ser movida en ondulaciones para darle al pez desplazamientos hacia adelante y hacia atrás. La aleta anal es similar por lo que respecta a su elongación y función en los peces de lomo plumo-

( d ) Mantarrava

ALETAS Y LOCOMOCI6N 179
so (Notopterus) y en la anguila eléctrica (Electrophorus), en los cuales se extiende en dirección caudal lo suficiente para fusionarse con la caudal. Tanto la aleta dorsal como la anal tienen bases moderadamente largas en los peces puerco (Bs- listidae), y son onduladas, con lo que dan al pez movimientos relativamente len- tos hacia atrás y hacia adelante, aun cuando la aleta de la cola esté en posición de descanso. En unos cuantos peces, y notablemente en los cofre (Ostraciidae), la aleta caudal es el órgano primario para la locomoción y funciona como remo. Las aletas pectorales son las estructuras locomotoras básicas que trabajan median- te la ondulación en las rayas, las mantarrayas y sus demás parientes (Rajiformes). Estas aletas son también capaces de dar movimiento, trabajando como remos, a grupos tan diversos como los caballitos de mar (Hippocampus), los cótidos (Cotti- dae) y los pércidos (Etheostomatinae).
Las aletas pares y medias son de mucha utilidad en los peces típicos, como órganos para la conservación de la estabilidad y para lograr la capacidad de ma- niobra (Figura 6.3). Las aletas medias desempeñan un papel obvio como quillas, para ayudar al individuo a mantenerse derecho, en posición vertical. Debido a que la musculatura intrínseca localizada en la base de las aletas (Figuras 3.10 y 3.11) permite que todas las porciones de las mismas presenten movimientos delicados y diferenciados, todas las aletas medias tienen también valor como órganos para maniobrar en el líquido. Las aletas pélvicas y pectorales también pueden servir como quillas de las porciones más voluminosas del pez, para retardar o frenar el movimiento giratorio de éste, aunque parece ser que sus principales funciones son las de permitir maniobras tales como trepar y bucear (Figura 6.4), posarse en al- guna roca dentro del agua, girar y permanecer quieto en el agua. Las aletas pecto- rales son las más importantes para lograr estas acciones y las pélvicas desarrollan un papel secundario, según se ha aprendido en la remoción experimental de las aletas en peces vivos.
Si quisiéramos comparar un dispositivo aerodinámico fabricado por el hom- bre con uno natural, parecido a un planeador, el cuerpo del pez sería el fuselaje, la aleta caudal el timón y las aletas pares las alas, los alerones, las aletas, los estabili- zadores y los elevadores. Las aletas medias comparten la función guiadora de la aleta caudal y la vejiga gaseosa actúa en parte como un estabilizador adicional.
Las aletas pares actúan para elevar la porción anterior del cuerpd de los tiburones (Squaliformes). La parte posterior del cuerpo es elevada mediante un
Figura 6.3 - Aletas medias curvadas hacia lados opuestos para servir como anclas marinas en la lobina de hocico gran- de (Micropterus salmoides). (Basado en Breder, 1926).

180 ESQUELETO, ESTRUCTURA Y MOVIMIENTO
A
Figura 6.4 - Empleo de las pectorales para dirigir el ascenso (a) o el descenso (b) en el agua por el brillador dorado (Nofe- migonus crysuleucas). Las flechas curvas indican la dirección del recorrido y las angulosas, vectores de fuerza. (Fuente: Breder, 1926).
levantamiento originado por la cola heterocerca. Las fuerzas elevadoras de las aletas pectorales y caudales se contraponen a la flotación negativa del cuerpo que carece de una vejiga natatoria. En los escómbridos, que también carecen de una vejiga gaseosa, las aletas pectorales y el pedúnculo caudal también generan fuerzas similares de levantamiento. Como contraste, la mayor parte de los peces óseos (Osteichthyes) tienen vejiga gaseosa y, al estar en equilibrio, tienen, en consecuen- cia, una gravedad específica igual a la del agua. Como resultado de lo anterior el mantenimiento de la posición a cualquier profundidad habitual presenta poco problema. Esa estabilidad, sin embargo, es dinámica y cuando un pez trepa o bucea en la columna de agua la vejiga gaseosa proporciona la suficiente compensación para logar el equilibrio dinámico entre la propia vejiga y las aletas.
Hay una función adicional en las aletas pectorales porque sirven para fre- nar. En los teleósteos relativamente largos como los salmones (Oncorhynchus), las aletas pectorales están situadas enfrente del centro de gravedad y las posteriores (abdominales) o aletas pélvicas, evitan que el extremo caudal del cuerpo se levante cuando el pez detiene su marcha. En muchos peces de cuerpo corto (Lepornis) las aletas pectorales están cerca del centro de gravedad, pero las pélvicas se hallan muy por delante y se oponen a la fuerza que hacen hacia arriba las pectorales cuando el pez se detiene.
Aunque todas las aletas juegan un papel importante en la función estabili- zadora, ésta no está completamente determinada para cualquiera de esos órganos. Cuando una o más aletas son removidas en plan experimental (o accidentalmente), aparece una acción compensatoria en las otras, de tal modo que todavía proporciona un equilibrio razonable. Solamente en el caso en que todas las aletas hayan sido cortadas es cuando aparece la evidencia de un serio desequilibrio. De todas las

MI6MEROS Y NATACI6N 181
aletas son las pélvicas las que tienen menor importancia en la estabilización. Se ha hecho notar, sin embargo, que los peces que tienen aletas pélvicas abdominales (generalmente los de radios blandos) tienden a tener aletas dorsales y anales cor- tas, mientras que los que tienen las aletas pélvicas en posición torácica o yugular (generalmente los peces con radios espinosos) tienen a menudo aletas dorsales y anales largas como compensación. En los mecanismos del equilibrio intervienen el sentido de la vista y el del equilibrio, este último alojado en el laberinto mem- branoso del oído interno (Capítulo 11).
Las aletas de los peces también sirven para muchas otras funciones, además de las de la locomoción o equilibrio.
MI~MEROS Y NATACI~N
Cuando las fibras musculares de un miómero son estimuladas, se acortan. Si los miómeros de un lado del pez se acortan, la cabeza y la cola se doblan hacia ese lado y los miómeros del lado opuesto se alargan. Las contracciones seriadas y alternadas de los miómeros de los dos lados hacen que el pez se flexione, o que reaccionen porciones sucesivas del animal, formando una especie de ondulación.
La onda que resulta de las contracciones seriadas recorre suavemente el cuerpo a todo lo largo, desde la cabeza hasta la cola, debido a que los músculos segmentarios típicos de los gnatostomados se traslapan. La cola se agita con cierta frecuencia (según sean las ondas que pasen sobre el cuerpo), y varía con la veloci- dad de nado, el tamaño del pez y la clase de animal de que se trate. Las contrac- ciones musculares que dan nacimiento a una onda propulsiva que se dirige hacia atrás del cuerpo resultan en un movimiento de natación hacia adelante, cuando la propia onda lleva una velocidad hacia atrás mayor que la del movimiento de natación hacia adelante. La expresión de fuerzas como éstas en los miosepta deriva en el fortalecimiento de huesos intermusculares en muchos teleósteos. Si la onda del cuerpo se desplaza de la cola a la cabeza, el pez nadará hacia atrás, Esta capacidad es notable en las anguilas y otros peces alargados que tienen locomoción anguili- forme. Las lampreas son una excepción, desde el momento en que ellas no pueden producir ondulaciones hacia atrás; pero tienen su compensación en la gran flexibi- lidad que presentan, la cual les permite girar con mucha facilidad en un espacio reducidísimo.
El principio de la acción de las fuerzas que es aplicado por la contracción de un miómero puede ser representado por medio del diagrama de la Figura 6.5. En esta figura, AB representa la línea media del cuerpo, estando A dirigida hacia la cabeza. Los romboides abcd y efgh representan respectivamente secciones hori- zontales a través de las flexiones hacia adelante y hacia atrás de un miómero. Un miómero aplica su fuerza para doblar el cuerpo en esas flexiones. Los miómeros se fijan a la línea media del cuerpo, a la altura de las regiones de flexión, formando los ángulos Odc y Oef, mientras que las fibras musculares en el miómero son más o menos paralelas a AB. Cuando las fibras musculares se contraen tiran el septo anterior de la región delantera de flexión y el septo posterior de la región trasera de flexión. P representa el estiramiento de las fibras musculares en la línea media. La contracción muscular acerca las flexiones entre sí, mientras que la línea me- dia se curva en las inmediaciones del punto central O. Hecha la flexión, los mió- meros y la línea media AB se doblan formando el arco mostrado por la línea a

182 ESQUELETO, ESTRUCTURA Y MOVIMIENTO
,/\\ , \
Figura 6.5 - Acción de un miómero al doblar el cuerpo de un pez. Para su explicación vdase el
o "- .' I texto. (Fuente: Nursall,
e h 1956).
rayas. Las contracciones seriadas de los miómeros producen una onda de curva- turas suaves a lo largo del cuerpo, como resultado de la inserción y entrecruza- miento de losmiómeros, como ya se ha descrito (Figura 3.7).
NATACION
La natación muestra los aspectos meciinico y energético, y los veremos conden- sados en las dos siguientes subsecciones.
Natación y movimiento del cuerpo
El principal procedimiento empleado por los peces para lograr el movimiento hacia adelante consiste en la flexión seriada y alternada de los músculos del cuer- po. Existe suficiente variación en las manifestaciones externas de esta clase de acción muscular como para haber permitido a los observadores clasificar esos movimientos natatorios en tres amplias categorias: anguiliformes o parecidos al de las anguilas, ostraciformes o parecidos al de los peces cofre y carangiformes o parecidos a los del jurel.
El tipo anguiliforme de locomoción es por naturaleza serpentiforme, como puede observarse en una víbora que serpentea al arrastrarse o en una anguila cuando nada. En la anguila se produce este movimiento por las contracciones secuenciales o alternadas de los miotomos en cada lado del cuerpo.
La locomoción ostraciforme consiste en un movimiento de vaivén que puede ser visto especialmente en la accibn de remar de la cola. Es inducido por la simple contracción alternada de todos los segmentos musculares de un lado del cuerpo, y luego la de los del lado opuesto. Estas contracciones alternadas hacen que la cola corta se mueva rápidamente hacia uno y otro lado, como si fuese un remo colocado detrás del tronco relativamente rígido del pez. El cuerpo se mueve siguiendo una serie de arcos cortos y cruzados en el agua, a medida que el pez se desplaza hacia adelante. Sería el mismo efecto que guiar un bote, haciéndolo avanzar llevando el remo sobre la borda, por la popa.
El tercer tipo de locomoción, que es el más común, es el carangiforme, donde el pez nada por sí mismo hacia adelante mediante movimientos de ba- rrido que van de lado a lado, producidos por la región caudal. Este caso es, en un sentido, intermedio entre los tipos anguiliforrne y ostraciforme. Es produ- cido por las contracciones alternadas de los mitomos, primero los de un lado

NATACIdN 183
del cuerpo y luego los del otro, comenzando en la cabeza y corriendo luego ha- cia atrás.
Las vértebras forman una cadena de palancas articuladas que se mueven solamente en sentido horizontal. Por lo tanto, en los tipos de natación carangi- forme y anguiliforme la columna vertebral azota u ondula de lado a lado, en con- traste con lo que sucede en las ballenas y animales relacionados con ellas, en los que se mueve de arriba a abajo. Esto hace que el cuerpo del pez se doble formando curvas cortas que se alternan de un lado al otro, y aumentando su amplitud a medida que se acercan a la cola. Los movimientos laterales amplios de la extensa área de la aleta caudal generan fuerzas laterales que hacen mover a la cabeza como en rebote, Este movimiento obviamente se resiste debido a la proporcio- nalmente gran cantidad de masa que hay en la parte anterior del pez, con res- pecto a la de la región caudal. Uno puede pensar que estos movimientos consisten realmente en una serie de pequeñas porciones del movimiento continuo del tipo anguiliforme. Cuando son consideradas las fuerzas relacionadas con la natación de un pez del tipo carangiforme, se ve que como la región caudal es doblada por una onda de contracciones musculares que corre hacia atrás sobre el cuerpo, las fuerzas resultantes del agua sobre el cuerpo arrastran al pez hacia adelante (Figura 6.6). Las fuerzas locomotoras anguiliformes son simplemente múl- tiplos del tipo carangiforme, donde cada una de las ondulaciones seriadas provee la fuerza de sólo una curvatura carangiforme. El ciclo de los movimientos laterales de la aleta caudal o cualquiera otra porción corta del cuerpo, ha sido comparado con el de una hélice de impulso variable, la que periódicamente podría variar su direc- ción de rotación y, simultáneamente, cambiaría su dirección de desplazamiento.
Gastos de energía en la natación
Aunque todavía es incompleto el conocimiento que tenemos sobre el gasto ener- gético que se produce durante la propulsión del pez, la hoja que muestra las bases del equilibrio energético requiere la medida de la energía metabólica puesta a disposición de los músculos locomotores, la proporción de energía metabólica convertida en energía mecánica, y la cantidad de energía mecánica muscular empleada para desplazar al pez. Un ejemplo de esas hojas de equilibrio es mos- trado en la Figura 6.7, para la natación relativamente lenta de la trucha arco iris (Salmo gairdneri).
El pez permanece en equilibrio con su reserva de energía, cuando son está- ticos los niveles de actividad y, por lo tanto, el metabolismo es aeróbico. Los
Figura 6.6 - Contorno del 10 cuerpo durante un ciclo propulsor para el movi- miento subcarangiforme. Dibujado sobre los trazos 3
hechos por la trucha arco iris (Salmo gairdneri) en intervalos sucesivos de 0.03 segundos. (Propor- , cionado por P.W. Webb).

184 ESQUELETO, ESTRUCTURA Y MOVIMIENTO
100 I I I 1
IO
Figura 6.7 - Relaciones entre la eficiencia aeróbica total,v,bh, eficiencia muscular, vm, y eficiencia propulsora de la aleta cau- dal, qp, como una función de la velocidad de natación expresada como porcentaje de la velocidad crítica de natacibn, para Salmo gairdneri. Los valores para vp (medidos) fueron calculados a partir de la medición del consumo de oxígeno y el esfuerzo de natación y las eficiencias musculares esperadas. Los valores para qp (calculados) fueron obtenidos a partir del modelo de Light- hill. (Basado en Webb, 1975).
niveles de natación violentos o acelerados requieren de la utilización de grandes cantidades de energía en periodos de tiempo muy cortos. Esta energía es propor- cionada de inmediato por el ATP (adenosintrifosfato) de los músculos, los cuales son repuestos de sus pérdidas a través de otros medios metabólicos anaeróbicos (principalmente por la glichlisis), además de recibir otros compuestos ricos en energía.
En la locomoción intervienen dos sistemas de fibras musculares, usualmente en la forma de bloques homogéneos de músculo rojo y blanco. El músculo rojo representa algo así como el 1 al 20% de los miotomos en los peces, dependiendo de su hábitat. Su velocidad es baja y funciona principalmente en forma aeróbica

LOCOMOCI6N NO NATATORIA 186
cuando los niveles de actividad son mantenidos sin cambios. El músculo blanco (músculo violento) tiene una velocidad más elevada y funciona principalmente en forma anaeróbica; es usado para el arranque y la aceleración durante la natación.
Los músculos que actúan en el sistema propulsor caudal generan una fuerza de empuje que es igual a la fuerza de arrastre. La fuerza requerida para nadar va- ría desde algo así como cinco veces la necesaria para un vehículo equivalente como un submarino, hasta un poco más el valor citado anteriormente.
LOCOMOCIdN NO NATATORIA
Aunque la forma típica de locomoción en los peces es la natación, ellos también cavan, se arrastran, saltan, se elevan sobre el agua y avanzan por sí mismos ayu- dándose de chorros de agua. Los peces como las rayas (Rajidae) y los lenguados (Pleuronectidae y Soleidae) cavan superficialmente sobre el fondo, arrojando el fango sobre su cuerpo para cubrirse, a excepción de la boca y los ojos. El pececillo del fango (Umbra) y el pez climático japonés (Misgurnus) hacen las excavaciones más profundas, como lo hacen algunos otros peces cuando perciben una alarma; rápidamente nadan y se introducen en el material blando del fondo. Los peces pulmonados africanos (Protopterus) excavan un túnel y permanecen en é1 para protegerse de la desecación mientras respiran aire atmosférico durante el largo periodo de estiaje. También las lampreas (Petromyzonidae), cuando son jóve- nes, permanecen en un túnel que construyen en el fondo durante los tres años o más de su vida larvaria; sólo sacan la cabeza, pero la introducen en la arena o el lodo cuando son asustadas. Los peces lanza de la arena (Ammodytidae) y los peces de la arena (Trichodontidae) son habitantes persistentes de la arena suelta hasta profundidades de 15 cm. Algunas morenas (Ophichthidae), de 50 a 90 cm de longitud, cuando son asustadas cavan rápidamente e introducen primero la cola, hasta profundidades de un metro. Sus parientes más cercanos, las anguilas gusano (Echelidae) comparten a menudo estos hábitos. Una de las anguilas culebra (Pisodonophis) remontan las aguas dulces para cavar en busca de alimento en los campos de arroz de la India y han desarrollado en los orificios nasales unos tubos de extensión especializados para cerrarlos y evitar así que el lodo penetre y les mo- leste cuando están construyendo el túnel.
Mediante la combinación de movimientos del cuerpo y las aletas el pez puede arrastrarse en el fondo o en tierra firme. Los peces que se arrastran en el fondo parecen tener, ya sea forma de anguila, como las morenas trepadoras de las grietas (Muraenidae), o poseer aletas pectorales modificadas, como se ve en los pe- ces soldadito o angelito (Triglidae), que tienen radios parecidos a dedos en el margen anterior de sus aletas, las cuales son muy alargadas. Las aletas pectorales han desarrollado porciones basales más o menos parecidas a brazos para sus puntas digitiformes, como sucede en otros peces batipelágicos como los peces sapo (Ba- trochoididae), los lófidos (Lophiidae), los peces antenados (Antennariidae), y en forma más pronunciada en los peces murciélago (Ogcocephalidae). La perca tre- padora oriental (Anabas) se arrastra por la tierra usando sus aletas pectorales y las coberturas espinosas de las branquias, y puede inclusive trepar por sobre las raíces de los árboles o los desperdicios que están en la playa, para alimentarse, respi- rando aire atmosférico mientras está fuera del agua. El saltarín del fango (Perio- phthahus) que habita en Australia, Asia y África, juguetea y brinca sobre 10s

186 ESQUELETO, ESTRUCTURA Y MOVIMIENTO
lodazales después de haber comido. El pez con mandíbula en forma de formón (Pantodon) de Africa es capaz de recorrer considerables distancias sobre la tierra, al trepar de una manera muy peculiar con la ayuda de sus aletas pares. En el curso de su migración hacia el océano para desovar, se ha visto a la anguila (Anguilla) que “corta camino” en las esquinas de los arroyos durante las mañanas húmedas. Posiblemente de todos los que caminan en tierra es el bagre de la especie Clarius batrachus el más notorio.
Algunos peces, literalmente hablando, se han posesionado del aire con muy diversos propósitos como son los de escapar, capturar la presa (como en el caso de los insectos aéreos) y posiblemente hasta para jugar. El salto del sábalo (Mega- lops), el del pez vela (Istiophorus) o el de cualquier otro pez aprovechado en la pesca deportiva, cuando es ejecutado al ser atrapado por el anzuelo, consiste real- mente en un nado rápido hacia y a través de la superficie lanzando el cuerpo hacia el aire, como sucede cuando un submarino lanza un proyectil hacia fuera del agua. El pez vela puede realizar saltos de 1 5 metros de extensión. Tanto la sardinita (Dorosoma cepedianum) y el pescadito de lados plateados (Labidesthes sicculus) representan a los saltadores de los bancos de peces; a este hábito, mostrado por estas dos especies, se debe que localmente se les haya aplicado el nombre de “skipjack”. Las lisas (Mugil) y las carpas (Cyprinus) escapan saltanto rápidamente sobre los aros de las redes de los pescadores que trabajan en las playas. Son sor- prendentes los saltos que hacen los salmones (Oncorhynchus y Salmo salar), a menudo frente a fuertes corrientes, para salvar los obstáculos o los saltos de agua, en su ruta para desovar por las áreas de nacimiento de las corrientes. Diversos peces saltadores, entre los que pueden quedar incluidos los sábalos, las sardinitas y muchos otros, tienen una elongación peculiar de los últimos radios de la aleta dorsal que bien puede estar relacionada con el hábito del salto, de una manera que no ha sido todavía determinada en forma experimental.
Mediante la aplicación de una velocidad adecuada a los saltos extendidos puede el pez remontar el vuelo siempre que tenga bien desarrolladas estructuras semejantes a alas. Pero éste es más bien un planeo que un vuelo verdadero. Con el fin de alcanzar las velocidades necesarias para remontarse o planear, el pez nada hacia arriba a través de la superficie del agua, llevando mucha velocidad y un án- gulo muy agudo con respecto a aquella, y entonces se remonta apoyándose en las aletas extendidas. Algunas clases de peces voladores (Exocoetidae), después de salir del agua y antes de remontarse, aumentan la velocidad para iniciar el vuelo, mediante la propulsión en la superficie del agua con la ayuda de la cola que vibra muy rápido, y en donde juega muy importante papel el lóbulo inferior de la aleta caudal, Único que hace contacto con el agua durante esta maniobra. El vuelo en planeo ha sido desarrollado en forma independiente en varios grupos de peces, entre los cuales quedan incluidos los peces voladores mismos (Exocoetidae) y los dactilópteros (Dactylopteridae), los gasteropelecinos (Gasteropelecinae) de América del Sur y los peces con mandíbula en bisel, Pantodon buchhoki de Áfri- ca. Aparentemente, las aletas pélvicas están alargadas para aumentar la capacidad de remontamiento en los peces voladores del género CypseZurus, haciendo de ellos verdaderos biplanos, cuando se les considera en conjunto con las aletas pectora- les, también alargadas.
Cuando las cámaras branquiales se contraen el agua es lanzada hacia l a cola y, como consecuencia, resulta un empuje hacia adelante. Muchos peces, cuando están en descanso, compensan continuamente este empuje mediante los

VELOCIDAD DE RECORRIDO 187
movimientos de las aletas pectorales. Parece ser que la propulsión a chorro forma parte importante del empuje necesario para adquirir la velocidad de despegue a partir de la posición de descanso, como puede ser fácilmente observado en los peces predadores como el lucio (Esox).
VELOCIDAD DE RECORRIDO
Puede pensarse que la velocidad de recorrido de los peces corresponde a una velo- cidad de crucero (o de actividad rutinaria), otra máxima sostenida y, finalmente, una velocidad de ímpetu violento. Las velocidades de crucero son las que corres- ponden a los viajes ordinarios. La mayor cantidad de información que se ha obtenido sobre las velocidades de crucero en los peces se debe a la subsecuente recaptura de peces marcados, en diferentes épocas y lugares, de aquél en que fueron marcados y liberados. Es obvio comprender que esas estimaciones sobre la velocidad de recorrido son necesariamente bajas y no corresponden a las velo- cidades reales porque no se cuenta con la información sobre los rodeos y las in- movilizaciones temporales de los peces. Las velocidades máximas sostenidas son las que un pez puede mantener durante intervalos muy largos. Han sido medidas experimentalmente en muchos peces pequeños mediante la determinación del tiempo que ellos pueden durar nadando en un lugar, contra la corriente de las aguas de un río, cuando la velocidad de éstas es conocida. Estas velocidades má- ximas sostenidas son intermedias entre las de crucero y las violentas. Éstas co- rresponden a un empuje máximo efectuado en un tiempo mínimo. Puede ocurrir cuando el pez, estando en una posición de descaL,*o, se ievanta violentamente; o bien, puede ser el avance sorpresivo de un pez de caza deportiva.
Las velocidades de crucero varían por especies, por el tamaño del individuo y por la temperatura del agua. Las velocidades de viaje rutinario, en millas por hora y kilómetros por hora, han sido determinadas para las siguientes especies:
Pez Velocidad de travesía*
millas/hora kilómetros/hora
“blenny” (Zoarces) gobio (Gobius) “rock gunnel” (Pholis gunnellus)
0.8 0.5 1.0 0.6 1.1 0.7
-c
*Debido, seguramente, a la falta de familiaridad en el empleo del Sistema Métrico Deci- mal, los autores proporcionan datos equivocados sobre el desplazamiento de los peces en km/h. Considerando que la información dada en la primera columna es la correcta, si 1 milla terrestre es igual a 1,609 m, en el caso del pez Zoarces, por ejemplo, 0.8 m/h equivalen a 1.287 k/h (o sea 1,287 m), y la misma ecuación es válida en todos los casos. Por lo tanto, la relación es la siguiente:
millas/hora 0.8 1.0 1.1 2.3 2.6 3.9 4.2
kilómetros/hora 1.287 1.609 1.769 3.700 4.183 6.275 6.757
millas/hora kilómetros/hora 4.7 7.562 5.0 8.045 5.8 9.332 7.6 12.228 8.2 13.193 11.0 17.699

188 ESQUELETO, ESTRUCTURA Y MOVIMIENTO
Pez Velocidad de travesía millas/hora kilómetros/hora
arenque (Clupeu sprattus) espinocha (Spinuchiu spinuchia) lenguado (Pleuronectes flesus) anguila (Anguilla rostmta) lenguado (Pleuronectes platessu) pez ángel ( T r i g l a ) carpa dorada (Carassius) lucio (Esox Zucius) trucha café (SuZmo truttu) macarela (Scomber scombrus)
2.3 2.6 3.9 4.2 4.7 5.0 5.8 7.6 8.2
11.0
1.4 1.6 2.4 2.6 2.9 3.1 3.6 4.7 5.3 6.8
Se han registrado los promedios de velocidades de recorrido de los salmones del Pacífico, cuando remontan los ríos, quedando entre 11.2 y 16.0 kilómetros por día* (aproximadamente 7 y 10 millas, respectivamente).
En la naturaleza, como contraste, también se han reportado algunas veloci- dades extremas. La perca rayada (Morone saxatilk) y las percas negras han alcan- zado, según los registros, velocidades de hasta 19.2 kilómetros por hora (12 mph). El registro para las macarelas (Scomber) es de 32.8 kilómetros por hora (20.5 mph) y el de la barracuda (Sphyruem) es de 43.2 kilómetros por hora (27 mph); tam- bién se han registrado valores parecidos a éstos en el salmón y el tiburón azul. Los peces voladores (Exocoetidae) han alcanzado a mostrar velocidades de hasta 56 kilómetros por hora (35 mph) y el dorado (Coryphaena), 59.2 (37 mph). El merlín (Maltuira), el atún, (Thunnus), el bonito (Sarda) y el albacora (Thunnus alalunga) han llegado a alcanzar, según los reportes, 64.0 a 80.0 kilómetros por hora (40 a 50 mph). A la cabeza de todos, con cerca de 96 kilómetros por hora (60 mph), se encuentran el pez vela (Istiophorus) y el pez espada (Xiphius). Estos valores no se han confirmado experimentalmente y, por lo que respecta al del pez vela, es por lo menos irreal desde el punto de vista de la cantidad de caballos de fuerza requeridos para alcanzar esa velocidad.
Las velocidades en donde el animal despliega su máximo esfuerzo, son de gran importancia para que el pez escape de sus enemigos, incluyendo las redes y otros dispositivos ideados por el hombre para pescar. Para los predadores, durante la captura de su alimento representado por las presas, y para los que desovan en los ríos, como sucede con el salmón del Pacífico (Oncorhynchus) que tiene que brincar sobre cascadas o embalses cuyas cortinas tienen dos metros de altura y cuatro metros de grosor, las velocidades máximas de ímpetu violento son vitales. Para realizar esta hazaña el salmón necesita salir del agua a una velocidad mínima estimada de 22.4 kilómetros por hora (14 mph). Las velocidades máximas soste- nidas y los arranques violentos son, sin embargo, la excepción más bien que la regla en el movimiento ordinario de travesía. Los salmones alcanzan una velocidad máxima sostenida cercana a los 12.8 kilómetros por hora (8 mph), no la de 22.4 que se requiere para lograr un salto elevado.
*En este caso al igual que en los posteriores hemos indicado las equivalencias correctas según la nota anterior, tomando como válidos los valores en millas terrestres.

MIGRACIONES 189
MIGRACIONES
Partiendo de lo que se conoce acerca de los recorridos que hacen los peces puede concluirse que la mayoría de las especies mantienen áreas territoriales relativa- mente reducidas, donde realizan casi todos sus movimientos. Algunos peces son, sin embargo, grandes viajeros y parece ser que están continuamente en movimien- to. Muchos peces pequeños de los arroyos, lagos y aguas marinas costeras, aunque activos, son muy estacionarios porque no salen de su territorio. Los peces pelá- gicos y otros que a menudo viajan entre las aguas dulces y las marinas, se despla- zan mucho. En esta categoría quedan incluidos los salmones (Oncorhynchus y Salmo) que se alimentan principalmente en el mar pero desovan en las aguas dul- ces, y las anguilas que hacen precisamente lo contrario. Entre estos dos extremos hay variaciones graduales. No hay una línea precisa que separe tajantemente el empleo de los Grminos movimiento y migración. Ambos pueden incluir al pez en sus recorridos verticales como los que parten de las profundidades a la super- ficie de las aguas, o los que suben y bajan por las laderas sumergidas. También pueden incluir los movimientos en direcciones horizontales, como cuando se alejan de la playa o se acercan a ella, o cuando remontan o descienden la corriente. Durante el curso de su movimiento deben estar en agua dulce (potamódromos o limnódromos), en el océano (oceanódromos), o en aguas intermedias (diádromos), desplazándose de las aguas marinas hacia las dulces (anádromos) o de las aguas dulces a las del mar (catádromos).
A los recorridos que hacen los peces en los ríos y corrientes de agua dulce en general se les llama potamódromos y son muy diversos. Estos recorridos son muy espectaculares entre los nadadores de los ríos, como el bocachica (Prochi- lodus), que viaja con otros peces (Mylinae y Leporinus) grandes distancias aguas arriba en las cuencas fluviales de América del Sur durante las temporadas de inun- daciones. La mayor parte de las carpas hociconas (Catostomidae) y las lampreas (Petromyzonidae) ascienden los ríos tributarios para desovar, como lo hacen tam- bién algunas poblaciones del “walleye” (Stizostedion). Es típico que los habitantes de los ríos, como las carpas hociconas, las lampreas, las carpas ordinarias (Cypri- nidae) y las truchas (Salmoninae), viajen río arriba para desovar. Los movimientos limnódromos, que se realizan hacia la playa cuando comienza a oscurecer y hacia el centro del lago cuando amanece, caracterizan a los peces como la trucha-perca (Percopsis), la perca amarilla (Perca fluuescens) y el pez de la roca (Ambloplites), entre muchos otros. En muchas especies de peces se ha visto claramente cómo hay movimientos masivos en los lagos, cuando andan en busca de la multitud de orga- nismos que constituyen muy buen alimento cuando están agrupados; entre esas especies queda dramáticamente incluida la del pescado blanco (Morone chrysops). La trucha de lago (Suluelinus nulnuycush) y muchos otros peces blancos y “cis- coes” (Coregonus) se dirigen hacia los bancos lacustres de arena para desovar, como lo hacen las poblaciones lacustres del “walleye” (Stizostedion uitreum).
Los peces viajeros marinos, oceanódromos, tienen hábitos de movimiento tan diversos como los que presentan los de aguas dulces. Los que viven en las zonas templadas del Norte, tienen la tendencia de dirigirse hacia el norte cuando el agua se calienta y hacia el sur cuando se enfría. Varios arenques entre los que se consideran los verdaderos (Clupeu) y las lachas (Breuoortia) viajan a lo largo de las costas siguiendo el rumbo citado, como lo hacen también muchos otros peces marinos costeros y pelágicos como las macarelas (Scomber) y 10s atunes (!lYzunnus).

190 ESQUELETO, ESTRUCTURA Y MOVIMIENTO
Los peces diádromos que migran libremente entre las aguas dulces y mari- nas incluyen a los anádromos y catádromos previamente descritos. Como buenos ejemplos de peces anádromos podemos citar al pez rayado (Morone saxatilis), los salmones del Pacífico (Oncorhynchus), los salmones del Atlántico (Salmo salar) y las lampreas marinas (Petromyzon). De todos estos peces citados, son los salmo- nes del Pacífico los que han registrado mayores distancias de recorrido, pues pueden desovar a cientos de kilómetros tierra adentro después de haber atravesado miles de kilómetros de mar durante su desarrollo. Las anguilas de agua dulce (Anguillu) de América del Norte y el oeste de Europa son el ejemplo clásico de los peces catádromos. Los adultos comienzan en el otoño a dirigirse hacia el mar desde algo así como unos 250 kilómetros tierra adentro y desde elevaciones que llegan a ser de unos 2,000 metros. Algunos individuos viajan en el mar más de 1,500 kilómetros para llegar a encontrar, mediante un sistema de navegación ad- mirable, regiones del Mar de los Sargazos, apropiadas para desovar. Los jóvenes se reúnen y nadan de regreso hacia las aguas continentales, en un viaje que llega a durar hasta tres años. Tan luego llegan a las aguas dulces sufren una metamorfo- sis, cambiando su estado larvario leptocéfalo y luego remontan los ríos donde completan su desarrollo hasta llegar a la maduración sexual, a lo largo de un pe- riodo de tiempo que puede llegar a ser de doce años o más, antes de regresar de nuevo al mar. La orientación oceánica de estos peces, seguida por el reconoci- miento de los ríos que les vieron nacer, fenómeno demostrado por los muchísi- mos salmones (Oncorhynchus y Salmo salar) que han sido marcados para estudiar sus migraciones, es una de las maravillas de la navegación animal.
El movimiento completamente libre que realizan los peces entre las aguas dulces y las marinas, no precisamente con el fin de reproducirse, ha sido llamado anfídromo. Algunos gobies (Sicydium) de los ríos tropicales desvían su reco- rrido hacia el mar cuando están en estado larvario pero regresan al agua dulce cuando son todavía pequeños, Los pescadores deportistas del sureste de los Esta- dos Unidos conocen bien los hábitos del sábalo (Megalops) que penetra esporá- dicamente a los ríos calmados y en los lagos de agua dulce. Los movimientos irregulares del chiro (Chanos) nos obligan a considerarlo como anfidromo, ya que frecuentemente entra al agua dulce y sale de ella.
Por lo general, una migración consiste en un movimiento directo y más o menos continuo, durante el cual el pez se desplaza de un lugar a otro. Ese movi- miento está bajo el control del pez, genéticamente determinado e influenciado por los factores ambientales. Una migración incluye, típicamente, uno o más regresos hacia el lugar de salida (tendencia de regreso al hogar); estos modelos de migra- ción específicos para una especie dada a menudo producen la agregación de los peces y el establecimiento de un sentido de la dirección. Las migraciones se pro- ducen usualmente para desovar o alimentarse, y generalmente están orientadas por el tiempo, aunque algunas veces tienen lugar cuando los peces huyen franca- mente de las condiciones adversas.
Hay muchos factores que influyen en los movimientos de migración, entre los que podemos considerar la tendencia a regresar al lugar de origen; pueden ser agrupados, para nuestra conveniencia, en factores físicos, químicos y biológicos. Los factores físicos incluyen a los materiales que forman el fondo, la profundi- dad del agua, la presión (atmosférica y del agua), la corriente y la marea, la tur- bidez, la topografía, el declive, la temperatura y la luz; intensidad, fotoperiodo y cualidad. Entre los factores químicos tenemos la salinidad, la alcalinidad, la con-

REFERENCIAS 191
centración de iones de hidrógeno, la cantidad de gases disueltos, los olores, los sabores y las sustancias contaminantes. Se reconocen como factores biológicos que influyen en la migración la presión sanguínea, el desarrollo sexual, la fotota- xis, la respuesta social, los predadores, los contendientes, el hambre, el alimento, la memoria, el marcador fisiológico del tiempo y la situación endocrina. Los factores citados arriba interactúan de diversas maneras para continuar, dirigir y reprimir las migraciones y sus movimientos, Y, por sobre todo, los mecanismos nerviosos y endocrinos son de gran importancia para la orientación y la determi- nación del tiempo que debe imperar en la migración.
Los hábitos que los peces tienen para viajar pueden variar con la latitud. Algunos esculpinos marinos del norte (Myoxocephalus) penetran más a menudo a las aguas dulces de las partes frías de la zona que habitan mientras que los peces sierra tropicales (Pristidae) se mueven en la misma dirección en las porciones más calientes de su zona de distribución. Por el contrario, la espinocha de tres espinas (Gasterosteus uculeutus) es marino en las aguas frías del Norte, pero en el Sur se vuelve habitante predominante de agua dulce.
REFERENCIAS ESPECIALES SOBRE LA LOCOMOCION
Bainbridge, R. 1958. The speed of swimming fish as related to size and to the frequency and
-. 1958. The locomotion of fish. New Scientist, 4:476-478.
Breder, C.M. 1926. The locomotion of fishes. Zoologica, 4(5):159-297. Gerking, S.D. 1959. The restricted movements of fish populations. Biol. Rev., 34:221-242. Gero, D.R. 1952. The hydrodynamic aspects of fish propulsion. Amer. Mus. Novitates, 1601,
Gray, J. 1953. The locomotion of fishes. En Essays in Marine Biology being the Richard
-.1957. How fishes swimm. Sci. Amer., 197(2):48-54. Lighthill, M.J. 1969. Hydromechanics of aquatic animal propulsion. Annual Review of Fluid
Nursall, J.R. 1956. The lateral musculature and the swimming of fish. Proc. 2001. Soc. London,
Weeb, P.W. 1974. Pisces (zoology). En McGraw Hill, Yearbook of Science and Technology,
Webb, P.W. 1975. Hydrodynamics and energetics of fish propulsion. Bull. Fish. Res. Bd. Canada,
amplitude of the tail beat. Jour. Exptl. Biol., 35:109-133.
- . 1961. Problems of fish locomotion. Symp. 2001. Soc. London, 5:13-32.
32 p.
Elmhirst Memorial Lectures. Oliver and Boyd, Londres y Edimburgo, pp. 1-16.
Mechanics, 1 :413-446.
126(1):127-143.
PP. 333-336.
190, 158p.
REFERENCIAS ESPECIALES SOBRE EL ESQUELETO
Goodrich, E.S. 1913. Studies on the structure and development of vertebrates. Macmillan and
Gregory, W.K. 1933. Fish skulls, Trans. Amer. Phil. Soc., 23:75-481. (Reimpresión en Offset,
Harrington R.W., Jr. 1955. The osteology of the American cyprinid fish Notropis bifrenatus, with and annotated synonymy of teleost skull bones. Copeia, 1955(4):267-290.
Hyman, L.H. 1942. Comparative oertebrate anatomy. 2nd. Ed., Univ. Chicago Press, Chicago.
Co., Ltd., Londres.
Eric Lundberg Publ., Laurel, Fla., 1959.)

192 ESQUELETO, ESTRUCTURA Y MOVWIENTO
Norden, C.R. 1961. Comparative osteology of representative salmonid fishes, with particular reference to the grayling (Thymallus arcticus) and its phylogeny. Jour. Fish. Res. Bd. Canada, 18(5):679-791.
Shufeldt, R.W. 1899. The skeleton of the black bass. Bull. U.S. Fish Comm. , 19:311-320.

Capítulo 7
Sangre y circulación
Las funciones de la respiración, el equilibrio de los fluidos, la excreción e inclu- sive la digestión, están muy relacionadas con la sangre y los vasos sanguíneos que forman parte de un sistema circulatorio cerrado que acarrea gases respiratorios, desperdicios celulares, metabolitos excretables y minerales y sustancias nutritivas en solución o suspensión. Aunque se describe en capítulos separados, debe ser recordada en su totalidad la interdependencia de los diversos sistemas de órganos. Un área extensa de tejido semipermeable queda expuesta al agua en las branquias de los peces, y la función del complejo formado por la sangre y las branquias no consiste solamente en el intercambio de gases en ese tejido, sino también en la excreción de ciertos compuestos nitrogenados y la adquisición o eliminación de minerales solubles.
Los peces, a excepción de los pulmonados (Dipnoi), tienen una circulación única definida que va del cuerpo a las branquias para regresar de nuevo a aquél (Capítulo 3; Figuras 3.16 y 3.17). El corazón bombea en esta única ruta circula- toria la sangre con bajo contenido de oxígeno y contenido elevado de dióxido de carbono. El volumen de sangre involucrado en esta parte del proceso circula- torio es muy bajo, correspondiendo al 1.5-3% del peso del cuerpo en los peces teleósteos superiores (Teleostei), si lo comparamos al 6% o más de los mamíferos. Sin embargo, un tiburón, el cazón espinoso (Squalus acanthias), tiene un volumen de sangre igual al 5% del peso de su cuerpo. El plasma sanguíneo y las células son renovables en un mayor número de órganos y sistemas entre los peces que en los mamíferos. Hay otra característica muy notable del aparato circulatorio de los pe- ces y consiste en el paso de la sangre a través de cierto número de sistemas espe- ciales, capilares o sinusoidales, situados sobre el curso de las circulaciones arterial o venosa. Estos sistemas especiales de disposición capilar son llamados sistemas
193

194 SANGRE Y CIRCULACION
porta. Aparecen en las branquias, el hígado (el sistema porta-hepático) y en los riñones (sistema porta-renal). Además, entre los peces fisoclistos hay todavía otra red capilar, la “red maravillosa” (rete rnirabile) que forma parte de la vejiga ga- seosa. Los teleósteos tienen una disposición similar en la glándula coroides de sus ojos; en algunos tiburones (Lamnidae) rápidos para nadar y en los atunes y maca- relas (Scombridae), aparecen formaciones capilares de tipo especial en otros órga- nos, especialmente los músculos, dando lugar probablemente a que exista un mayor intercambio gaseoso entre la sangre y los tejidos.
CORAZ6N Y VASOS CIRCULATORIOS
El plan básico del sistema circulatorio ha sido ya explicado, teniendo como com- ponentes al corazón, los vasos sanguíneos y linfáticos y los tejidos fuidos entre los que se incluyen las células sanguíneas.
El corazón
En la mayoría de los peces el corazón está ubicado inmediatamente hacia atrás de las branquias (Figura 3.17). Entre los peces óseos superiores (Teleostei), sin embargo, que tienen coberturas branquiales (condición operculada), el corazón está situado muy por delante en el cuerpo, en comparación con los tiburones y grupos relacionados (Elasmobranchii), que son peces sin opérculo. En unos cuan- tos peces se puede observar un gran desplazamiento del corazón hacia atrás, como sucede en el pez pulmonado (Lepidosiren) de América del Sur y en los “swam- peels” (Monopterus) de la familia Synbranchidae, que viven en Asia. Por lo gene- ral, el corazón está bien protegido por la cintura pectoral.
El saco pericárdico membranoso que contiene al corazón es de capacidad amplia en los tiburones y especies emparentadas (Elasmobranchii) y más adherido en los peces óseos (Osteichthyes). El pericardio parietal, que cubre al saco peri- cárdico, se adhiere al pericardio visceral que cubre al músculo cardiaco, en las porciones donde las venas más importantes se unen al seno venoso y donde la ar- teria más importante, el tronco arterial (aorta ventral), sale del corazón para irri- gar las branquias. Una coalescencia similar de los pericardia parietal y visceral aparece donde los vasos sanguíneos del corazón mismo, las arterias y venas coro- narias, entran al corazón y salen de éI, respectivamente. El espacio que existe entre el pericardio parietal y el pericardio visceral del corazón deriva de la por- ción craneal del celoma embriónico o cavidad del cuerpo. En el pez adulto la cavidad pericárdica está cerrada hacia atrás, a excepción de los tiburones y espe- cies emparentadas (Elasmobranchii), así como en los esturiones y especies afines (Acipenseriformes) en donde debe haber una o dos conexiones secundarias entre las cavidades pericárdica y peritoneal.
El corazón varía considerablemente en lo que respecta a su desarrollo y tamaño relativos (Figuras 3.16 y 7.1). El tamaño del corazón, en relación con el bajo volumen y la baja presión sanguíneos de los peces, cuando se expresa en par- tes por millar del peso del cuerpo, es mis bajo en los peces que en otros grupos de vertebrados. Los peces de movimiento lento y relativamente sedentarios tienen pesos cardíacos relativos menores a 1 parte por millar de peso total del cuerpo,


196
CORAZONES
SANGRE Y CIRCULACI6N
D C - q (a) Tiburón (ventral)
VCA VCP I
AHA
(6) Catán (dorsal)
V
(c) Trucha (ventral)
((1) Pez pulmonado ídorsal)
Figura 7.1 - Variación de las pro- porciones entre los corazones de los peces y los vasos sanguíneos asocia- dos: (a ) pez perro manchado (Scy- liorhinus caniculus), vista ventral; (6) catán de nariz larga (Lepisosteus osseus), vista dorsal; (c ) trucha arco iris (Salmo gairdneri), vista ventral; (d) pez pulmonado africano ( fro- topterus annectens), vista dorsal. A, atrio; ABA, arteria branquia1 aferente; ACA, arteria coronaria derecha; ACM, arteria comisural; AHA, arteria aferente hioidea; AHM, arteria hipobranquial me- dia; BA, bulbo arterioso; C, cono arterioso; DC, conducto de Cuvier; PC, pericardio; SV, seno venoso; T, tronco arterial; V, ventriculo; VCA, vena cardinal anterior; VCP, vena cardinal posterior; VC, venas coro- narias; VE, vena epigástrica; VH, vena hepática; VJ, vena yugular; VP, vena pulmonar; VS, vena sub- clavia. (Fuentes: (a ) y (cl Grodzins- ki; (6) Rose; (d) Goodrich; todos en Bronn, 1938).
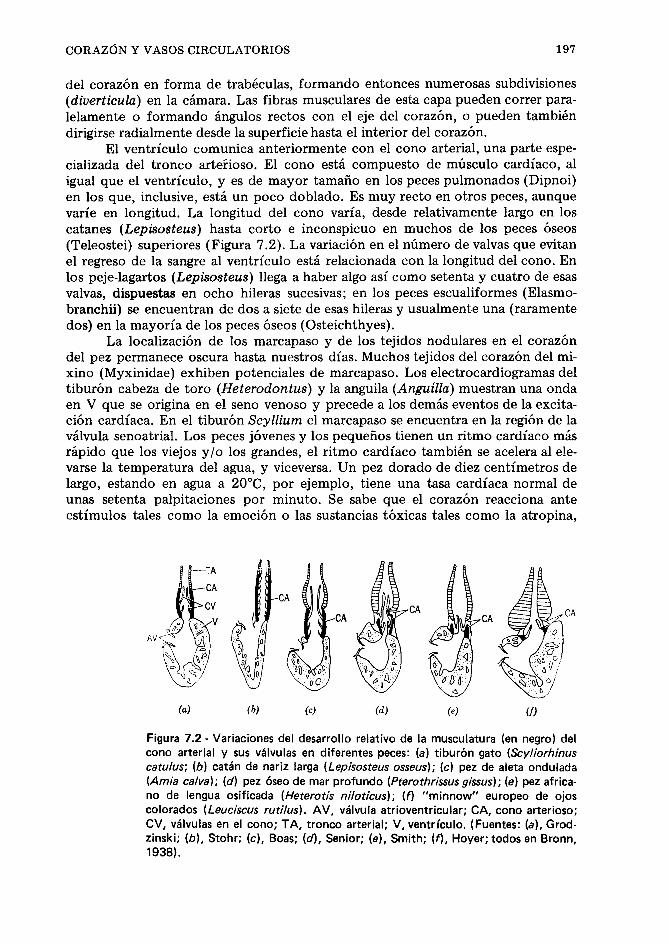
CORAZ6N Y VASOS CIRCULATORIOS 197
del corazón en forma de trabéculas, formando entonces numerosas subdivisiones (diverticula) en la cámara. Las fibras musculares de esta capa pueden correr para- lelamente o formando ángulos rectos con el eje del corazón, o pueden también dirigirse radialmente desde la superficie hasta el interior del corazón.
El ventriculo comunica anteriormente con el cono arterial, una parte espe- cializada del tronco arteiioso. El cono está compuesto de músculo cardíaco, al igual que el ventriculo, y es de mayor tamaño en los peces pulmonados (Dipnoi) en los que, inclusive, está un poco doblado. Es muy recto en otros peces, aunque varíe en longitud. La longitud del cono varía, desde relativamente largo en los catanes (Lepisosteus) hasta corto e inconspicuo en muchos de los peces óseos (Teleostei) superiores (Figura 7.2). La variación en el número de valvas que evitan el regreso de la sangre al ventriculo está relacionada con la longitud del cono. En los peje-lagartos (Lepisosteus) llega a haber algo así como setenta y cuatro de esas valvas, dispuestas en ocho hileras sucesivas; en los peces escualiformes (Elasmo- branchii) se encuentran de dos a siete de esas hileras y usualmente una (raramente dos) en la mayoría de los peces óseos (Osteichthyes).
La localización de los marcapaso y de los tejidos nodulares en el corazón del pez permanece oscura hasta nuestros días. Muchos tejidos del corazón del mi- xino (Myxinidae) exhiben potenciales de marcapaso. Los electrocardiogramas del tiburón cabeza de toro (Heterodontus) y la anguila (Anguillu) muestran una onda en V que se origina en el seno venoso y precede a los demás eventos de la excita- ción cardíaca. En el tiburón Scyllium el marcapaso se encuentra en la región de la válvula senoatrial. Los peces jóvenes y los pequeños tienen un ritmo cardíaco más rápido que los viejos y/o los grandes, el ritmo cardíaco también se acelera al ele- varse la temperatura del agua, y viceversa. Un pez dorado de diez centímetros de largo, estando en agua a 20°C, por ejemplo, tiene una tasa cardíaca normal de unas setenta palpitaciones por minuto. Se sabe que el corazón reacciona ante estímulos tales como la emoción o las sustancias tóxicas tales como la atropina,
CA CA CA CA
(a) (b) (4 ( 4 (4 (f)
Figura 7.2 - Variaciones del desarrollo relativo de la musculatura (en negro) del cono arterial y sus válvulas en diferentes peces: (a ) tiburón gato (Scyliorhinus carulus; (b) catán de nariz larga (Lepisosteus osseus); (c ) pez de aleta ondulada (Arnia calva); (u‘) pez óseo de mar profundo (Prerorhrissusgissus); (e) pez africa- no de lengua osificada (Hererotis niloricus); ( f l “minnow” europeo de ojos colorados (Leuciscus rurilus). A V , válvula atrioventricular; CA, cono arterioso; CV, válvulas en el cono; TA, tronco arterial; V , ventriculo. (Fuentes: (a) , Grod- zinski; (b), Stohr; (c ) , Boas; (u‘), Senior; (e), Smith; (0, Hoyer; todos en Bronn, 1938).


!as l&minas hranquiales, que es donde t,iene iugar e1 k!,ercw1h10 de gases y ot,~,rs srist,ancias en sc~iució~. entre !a sangre y ~1 agua mlhiente.
De esas siete a catorce arterias branyuiales aferentci; ei m q ~ o r numero aparece en el ghpo de las lampreas-mixinos (Cyclostomata) y depecde del nlj- mero de bolsas branquiales que deban ser irrigadas. La mayoria de \os escualifor- mes (Elasmobranchii) tienen cuatro (Figura 7.3) , con las excepciones del tiburon de seis branquias (Hexanchus) que tiene cinco, y el tiburbn de siete branquias (Notorynchus) que tiene seis. Los peces pulmonados (Dipnoi) tienen cuatro o cinco pares de arteri.as branquiales aferentes; los otros peces óseos generalmente tienen cuatro (Figura 7.3), aunque en los embriones de estos grupos se localizan hasta seis arcos branquiales. En cierto número de peces como las quimeras (Chi- maera), el bacalao (Gadus), algunos gobios (Gobius), peces globo o tambor (Tetra- odontidae) y los “mudskippers” (Periophthalmus), las arterias que van a las bran- quias nacen separadamente del tronco principal de la aorta ventra:. En la mayoría de los tiburones, sin embargo, el vaso miis anterior que nace dol tronco da lugar a la arteria branquial aferente y a la artería hinidea. Las arteriss branqu.iales IV y TI1 muestran una raiz común en la mayoría de los peces bseos, pero en algunos, como la zarpa c o m h (Cyprinus carpio) y otros pececillos de acuario (Cyprln:l- dae), los vasos hrnnyuiales aferentes I1 y I tienen un origen conriln.
Una r.amificaci6n de la arteria aferente va a dar a cada filamento branquial donde a su vez se desdobla en capilares y lagunas en las laminillas do los filam.en- tos. La sangre que se oxigena en el filamento es eoleclada !ateralmente en una asa eferente que se localiza alrededor de cada abertura branquia1 interna. Las asas van a comunicar con las arterias eferentes branquiales que forman m& tavde la aorta dorsal. Las arterias branquiales aferentes, por lo tanto, llevan a las branquias sangre venosa pobre en oxígeno y rica en bióxido de carbono. El flujo sanguineo se hace m6s lento en la vasta superficie capilar de las branquias. En las laminillas branquiales el flujo sanguineo es tal que I a reoxigenación y la remoción del CO, ocurre gradualmente del borde aboral al oral, con el objeto de hacer uso bptirno de las corrientes de agua que bañan a las branquias. La caída de la presión san- guínea en el lado de colecta de los vasos branquiales comienza en las laminillas de las branquias. Las presiones diastólicas de la aorta dorsal han sido registradas en forma de porcentajes, de acuerdo a la presih sanguínea de la aorta ventral; como se indica a continuación: “skate” (Raja binoculatu), 15; pez perro (Squalus suckleyi), 26; trucha (Salmo gairdneri) y carpa c o m h (Cyprinus carpio), entre 50 y 60. La presión sanguínea y el flujo en la aorta dorsal del bacalao (,Gadus rnorhuu) son pulsátiles, a pesar de que la sangre fluye a lo largo de los capilares de las branquias antes de llegar a ese vaso.
Las arterias branquiales eferentes pueden ser únicas, como se observa en los peces Óseos (Teleostei) superiores; o pareadas, una para cada hemibranquia del arco branquial, como se ve en los tiburones, rayas y “skates” (Elasmobranchii), con la excepción del arco anterior, el hioideo, que tiene solamente una arteria branquial eferent.e. En los peces pulmonados (Dipnoi) prevalecen condiciones intermedias, como sucede también en el pez espátula (Polyodon), los esturiones (Acipenser) y los catanes (Lepisosteus).
En los peces de los grupos de las lampreas y tiburones los vasos branquiales eferentes aparecen dorsalmente de uno en uno para unirse en la aorta. media. La primera arteria branquial eferente proporciona al cerebro y otras regiones cefá- licas gran parte de la sangre, aunque algunos vasos arteriales, principalmente la

200
CIRCULACI~N BRANQUIAL
SANGRE Y CIRCULACIdN
hioidea / / //
Arteria branquial eferente
Arteria subclavia Arteria Arteria cerebral basal
eferente pseudo branquial Arteria dorsal Asa colectora
lateral izquierda Arteria
Anrta dorsal
ciptica
Arteria oftálmica
Arteria orbital
Arteria - Círculo
pseudobranquial
Arteria celíaca
rteria mesentérica
rteria de la vejiga gaseosa
Arteria subclavia 4. arco branquial con
su holobranquia Filamento branquial Conductos de Cuvier izquierdo y derecho
Pseudobranquia
mandibular
aferente / Arteria Branquia' hipobranquial Aorta aferente 2
I coronaria
secundaria Cono
ventral arterioso
(b) Pez óseo
Figura 7.3 - Diagrama generalizado de la circulación branquial (lado izquierdo) en (a) el grupo de los tiburones (Elasmobranch¡¡) y (b) el grupo de los peces ÓSeos (Osteichthyes) como se observan en un bacalao, Gadus. (Basado en Goodrich, 1930).

CORAZdN Y VASOS CIRCULATORIOS 201
arteria hioidea, nacen del primer vaso branquial aferente. En los peces óseos supe- riores (Actinopterygii) las arterias branquiales eferentes de cada lado aparecen y se reúnen en un vaso troncal en cada lado. De esto resulta que sobre la región branquial la aorta dorsal está pareada y no se transforma en vaso medio sino hasta que llega justamente detrás de la región branquial.
Una disposición especial se encuentra en el sistema que proporciona sangre al espiráculo de los tiburones, las rayas y los “skates” (Elasmobranchii) y a su estructura homóloga en los peces óseos (Actinopterygii), la pseudobranquia hioi- dea. Ambas son alimentadas con sangre oxigenada procedente de una arteria mandibular o su derivada; en los peces óseos un vaso pseudobranquial eferente conduce la sangre que ha pasado por los capilares pseudobranquiales a través de la arteria oftálmica de la glándula coroidal del ojo. Por lo tanto, las conexiones vasculares de la pseudobranquia pueden sugerir que tiene una función primaria no respiratoria al guardar para sí otras propiedades anatómicas y fisiológicas (Ca- pítulo 8). La pseudobranquia puede contener receptores sensibles a la tensión del oxígeno y el bióxido de carbono.
Las venas de la región cefálica dorsal (Figura 7.4) comienzan siendo venas faciales, orbitales, postorbitales y cerebrales, y se unen en las cardinales anterio- res pareadas que van a dar a la cardinal común o ducto de Cuvier y, desde aquí, al corazón. La vena yugular colecta sangre venosa procedente de la región cefálica inferior y es tributaria de la vena cardinal, común en los peces óseos (Actinop- terygii). En los tiburones y especies afines (Elasmobranchii) la vena yugular se une con la subclavia procedente de las aletas pectorales y el lomo, antes de aso- ciarse con el ducto de Cuvier. Los elasmobranquios tienen muchas venas con espacios interiores (lumen) relativamente grandes, en comparación a las que tie- nen los peces óseos; los tiburones, rayas y “skates” tienen también numerosos receptáculos sinusoidales venosos de corriente sanguínea lenta, como se ve en el seno orbital, y también anastomosis venosas complejas, como se puede observar en las regiones condrocraneal e hipobranquial.
L o s principales vasos sanguíneos del cuerpo (Figura 7.5)
En lo que respecta al tamaño, los vasos sanguíneos del cuerpo de un pez son prin- cipalmente de dos clases: a) las principales arterias y venas que corren paralejas al axis en toda su longitud y b ) ramificaciones de los vasos principales que van a y proceden de la piel, el esqueleto y los músculos, y que también irrigan el cerebro, la médula espina1 y los intestinos.
La aorta dorsal constituye la ruta principal para el transporte de la sangre, desde las branquias al cuerpo; en todos los peces es única por detrás de las bran- quias. La aorta dorsal yace directamente en la región del tronco, por debajo de la columna vertebral de los peces mandibulados (gnatostomados) o bajo la noto- cuerda de los peces amandibulados (Cyclostomata). Después de desprender una serie de vasos dorsales, ventrales y laterales (véase abajo) la aorta se reduce y es conocida entonces con el nombre de arteria caudal, al atravesar el canal hemal que forman las vértebras caudales. El canal hemal es un túnel rígido e indeformable que sirve de protección para la aorta dorsal y la vena caudal, ya que de otra ma- nera sufrirían las presiones en forma de ondas producidas por la tensión muscular, como sucede cuando se presentan las flexiones que corren a lo largo del ccerpo

2i) 2
i

durante la natacihn. L a vena csu:jai se dobla hacia la región caudal, siguiendo el curso de la arteria eaudal y regresa la sangre ai eorazbn. E n ~ G S tiburones hay un sistema df: bombeo que ha sido descrito en Hetetodontus, en el cual ese sistema de bombeo depende de los movimientos de la cola; y también en Mustelus, en donde las ondas producidas por l a contraccibn de los músculos comprimen los de- pósitos venosos de la cola cuando el tiburón está descansando.
Las numerosas ramificaciones de los vasos que nacen de la aorta y de las venas correspondientes están dispuestas para adaptarse a los segmentos del cuerpo. Por cada segmento del embrión aparece un par de arterias que se dividen en rami- ficaciones dorsales, leterales y ventrales; es solamente en las lampreas y en los mixinos (Cyclostomata) donde aparecen estas arterias provistas de valvas que evitan el regreso de la sangre hacia la aorta. Modificaciones en la disposición de 10s segmentos del embribn aparecen cuando e! pez ha llegado a ser adulto. Las ramas dorsales segmentarias, sin embargo, todavía surten a la musculatura dorsal y a la aleta dorsal, o aletas dorsales si es que hag más de una, pero tienen ramifi- caciones que se introducen entre !os paquetes musculares (conos mioméricos) de la musculatura epaxial de los lados. Las arterias segmentarias laterales siguen la línea. divisoria (septo horizontal esqueletógeno), entre las regiones epaxial e hipa- xial. Las arterias segmentarias ventrales del embrión nutren a los músculos y aletas de la regibtn caudal, pero también pierden su disposición segmentaria y sufren muchas modificaciones, como se ve en ciertos vasos del tract0 intest,inal, princi- palmente el sistema celíaco-mesentérico con sus ramificaciones y las arterias ilíacas (Figuras 7.5 y 7.6).
Los sistemas venosos de muchos peces (Cyclostomata; Elasmobranchii) presentan senos elaborados; el sistema en los mixinos (Myxine) tiene tambi6n “cora~ones’~ secundarios en forma de bulbos contráctiles situados a todo lo largo, como se observa en las venas de la cola. Además, la sangre venosa que retorna al corazón fluye, a través de áreas que semejan senos, de los sistemas porta hepático y renal. Las venas del sistema portal renal que se originan en la vena caudal del embrión se descomponen, en el riñón en desarrollo, en una red de capilares cada vez más pequeños que luego se reúnen en las venas cardinal posterior o post- cardinal. La post,cardinal de cada lado termina en el correspondiente seno cardinal común o conducto de Cuvier, del lado donde se halla, el que a su vez continúa en el seno venoso. En el tiburón (Heterodontus) hay un gradiente de presión san- guínea venosa a lo largo de la vena lateral cutánea (Figura 7.5), comenzando con una presión positiva y constante en el extremo periférico, para llegar a tener una presión negativa y fluctuante en relación a las contracciones cardíacas, hacia el extremo central.
En la circulación del intestino de las lampreas (Petromyzonidae) existe una característica muy curiosa porque en la parte posterior del tract0 digestivo aparecen vasos que tienen una arteria colocada en el lumen de una vena. Y, todavía más: la sangre del intestino regresa al corazón, no a través de un sistema porta hepático, sino que alcanza las venas cardinales a través de un seno venoso suprarrenal.
Las venas del estómago y el intestino de los peces mandibulados se reúnen en la vena portal hepática para llegar al hígado. En este órgano la vena porta da lugar a miríadas de sinusoides que facilitan la transferencia al tejido hepático de las sustancias alimenticias disueltas. La sangre es entonces colectada por las venas hepáticas que penetran al seno venoso. La mayoría de 10s peces tienen dos venas hepáticas, pero los pececillos de acuario (Cyprinidae) y el bacalao (Ga-

SANGRE Y CIRCULACI6N

CORAZ6N Y VASOS CIRCULATORIOS 205
O
v) I- w I-
z
z -I w D v)
v) O
> a

I! I
Tronco 1
Tronco linfático linfáticn yugular lateral
I Vaso l inf i t ica
ventral longitudinal
Figura 7.7 - Vista lateral de los vasos linfaticos superficiales y el coraz6n linfático de una trucha caf6 joven (Salmo ;rum). (Basado en Hoyer en Bronn, 1938)-

SANGRE, FLUIDOS TISULARES Y ÓRGANOS FORMADORES DE LA SANGRE
La sangre de los peces consta de plasma fluido y contiene particulas especiales que sun en su mayoria células sanguineas y sus restos. La linfa, parte del plasma

208 SANGRE Y CIRCULACI6N
que atraviesa los vasos sanguíneos para bañar los tejidos, es fluida, pero puede contener algunos de los componentes celulares de la sangre; los glóbulos rojos (eritrocitos) aparecen con más frecuencia en la linfa de los peces que en la de los vertebrados superiores. En los mixinos (Myxinidae) los espacios de los senos pueden estar divididos en linfáticos rojos y blancos, teniendo los primeros un contenido relativamente elevado de glóbulos rojos.
En los peces también existen diferentes tipos sanguíneos, como ocurre en los humanos. Algunos se parecen al sistema Rh de los humanos y al B de las reses.
Plasma del pez
El plasma es un líquido claro en el que se encuentran, además de las células san- guíneas, minerales en solución, productos absorbidos durante la digestión, pro- ductos de desecho procedentes de los tejidos, secreciones especiales, enzimas, anticuerpos y gases disueltos. Las diferencias específicas incluyen a las diferencias de los coeficientes de sedimentación de las principales proteínas del plasma. Para el bacalao (Gadus callarias) la composición electrolítica de la sangre en mmoles/litro es como sigue: sodio (Na), 180; potasio (K), 4.9; magnesio (Mg), 3.8; calcio (Ca), 5.0; cloro (Cl), 158; y fosfato (PO,), 3.1. Las concentraciones de sodio y cloro son generalmente más bajas en los teleósteos de agua dulce. La sange de los tibu- rones (Squaliformes) posee un contenido más elevado de Mg, pero tiene una re- serva alcalina más débil de la que tiene la sangre de los peces óseos superiores (Actinopterygii).
La cantidad de soluto contenido en una solución es señalada por la dismi- nución del punto de congelación ( A ) , que corresponde también a la medida de la presión osmótica. Al incrementarse la presión osmótica se presenta un aumento en la tendencia del agua a difundirse a través de una membrana semipermeable para diluir la solución. Las disminuciones de los puntos de congelación del plasma en los peces quedan comprendidos desde 0.5"C para los peces óseos de agua dul- ce, hasta 1.O"C para los contados tiburones y especies afines (Elasmobranchii) de agua dulce; y entre 0.6"C y 1.O"C para los peces óseos marinos, hasta llegar a alcanzar una cúspide de 2.17"C en los elasmobranquios marinos. El valor de A para el agua de mar es de 208°C (Capítulo 9).
Los peces tienen bajos niveles de proteína de plasma comparados con los vertebrados superiores. Las principales proteínas del plasma sanguíneo del pez son la albúmina (que controla la presión osmótica), las lipoproteínas (que transportan los lípidos), las globulinas ("bind heme")*, la ceruloplasmina (que liga al cobre), el fibrinógeno (coagula la sangre) y el ioduroforino (Único en los peces, que liga al yodo inorgánico). Los niveles relativamente bajos de fibrinógeno y proteínas se- mejantes a la protrombina no se correlacionan con la muy conocida coagulación tan rápida de la sangre de los peces. La sangre de la trucha arco iris (Salmo gaird- neri), que es capaz de vivir a temperaturas un poco mayores a O"C, se coagula rápidamente a bajas temperaturas. Contrasta con esto el caso de la sangre de la carpa común (Cyprinus carpio), un pez de agua caliente, que no se coagula rápidamente hasta que la temperatura del agua llega a alcanzar aproximadamente los 15°C.
*No se encontró traducción (N. del T.).

SANGRE, FLUIDOS TISULARES Y 6RGANOS FORMADORES DE LA SANGRE 209
Los peces antárticos que viven en aguas cuya temperatura llega a bajar hasta los -1.9”C resisten la congelación, en parte por la presencia en su suero de un grupo Único de glicoproteínas. Los únicos aminoácidos presentes en estas glico- proteínas anticongelantes son la alanina y la treonina, en una proporción que va de 2 a 1. El peso molecular de las glicoproteínas anticongelantes queda compren- dido entre 2,600 a 33,000. Estas glicoproteínas anticongelantes aparentemente bajan el punto de congelación de la sangre del pez al interferir con el crecimiento de cristales de hielo.
Cierto número de enzimas han podido ser aisladas del plasma sanguíneo de los peces, contándose entre ellas la lipasa y la anhidrasa carbónica que son más abundantes en los peces marinos que en los de agua dulce. Se ha observado que el suero de ciertos teleósteos, entre los que se incluyen las anguilas de agua dulce (Anguillu), algunos bagres (Siluridae) y los atunes (Thunnus), es tóxico cuando se inyecta en la corriente sanguínea de los mamíferos. El suero de las anguilas de agua dulce destruye las células sanguíneas mediante la hemolisis, e influye en el tono y la permeabilidad de los vasos sanguíneos en los mamíferos.
Glóbulos rojos
Se han encontrado eritrocitos nucleados y de color amarillo rojizo en la sangre de todos los peces (Figura 7.8), a excepción de tres especies pequeñas del Antár- tico que tienen un metabolismo bajo, adaptado al frío y viven en aguas ricas en oxígeno; y, también, en la larva leptocéfala en forma de listón de las anguilas (Anguillu) y ciertos peces de los fondos marinos. Estas formas excepcionales pue- den, aparentemente, producir sus cambios gaseosos mediante la difusión. Los glóbulos rojos maduros de los peces tienen forma oval, son pequeños y su diá- metro mayor queda comprendido entre las 7 micras en muchos peces, como ciertos “wrasses” (Crenilubrus), a 36 micras en el pez pulmonado de Africa (Pro- topterus); una micra equivale a 0.001 de milímetro y el eritrocito humano, redondo, tiene un diámetro de 7.9 micras. Los glóbulos rojos anucleados son raros, pero la proporción de eritrocitos inmaduros y redondeados varía consi- derablemente en los peces, La sangre de los tiburones (Squaliformes), como la de las anguilas (Anguillidae) y las macarelas (Scombridae), puede contener algo así como un 20% de células inmaduras, mientras que la sangre de otros peces óseos activos puede tener un porcentaje mucho más bajo, como sucede en las rémoras (Echeneis) y en los peces pipa (Syngnuthus).
El transporte del oxígeno en la sangre depende del compuesto de hierro, la hemoglobina, que es el pigmento respiratorio de la sangre. El contenido de hemoglobina en la sangre del pez varía de acuerdo al número de eritrocitos pre- sente; expresados como porcentajes de peso seco de eritrocitos encontramos valores que van del 14 al 19% en el tiburón-perro (Mustelus canis) y del 37 al 79% en cierto número de teleósteos marinos y de agua dulce. A menudo van aso- ciados en los peces la capacidad para transportar oxígeno (Capítulo 8) al 95% de saturación, el contenido de hierro de la sangre y el número de glóbulos rojos; y lo anterior varía de acuerdo con la etapa de desarrollo, los hábitos y las condi- ciones ambientales (Tabla 7.1). Esto ha sido bien demostrado por Grodzinski (1938), de cuyo trabajo ha sido tomada la Tabla 7.1.

Tambih aparecen variaciones circ2idicas en el nGmero de eritrocitos , corno lo muestran un blenio (Blenniusj y un crenilabm (Crenilabrus). Los peces, entre otras reacciones; responden a l a conmoción ("shock"') bajando el nimero de gl6bulos rojos. Sin embargo, cuando sufren pérdidas de sangre puede aumentar rápidamente el nhrnero de eritrocitos jóvenes en la sangre; se trata de un efecto comparable al que aparece en los mamíferos con el aumento de l a altitud.
La sangre de los peces contiene, aparte de los eritrocitos con su pigmento respira- torio, varios tipos de c6iulas incoloras o blancas (Ieucocitos), todas de forma ovoide a esferoide. Mientras el niimero de los glbbulos rojos queda comprendido entre 2O,000 a 3,000,000 por milirne.tro citbico de sangre, el número de tos g16-
TabDa 7.1 - Comparación de ciertas características de los glóbulos rojos de los peces. _I_ _ll_"___ I__ "ll."""_ ___
Especies Capacidad de retención Número de eritrocitos Contenido de Fe de oxígeno ( 7 6 de (por mm") (mg por 100 cc)
sangre por volumen)
Pejesapo (Lophius)
Macarela (Scomber) (sedentario) 5
(activa) 16
867,000
3,000,000
13.4
37.1

SANGRE, FLUIDOS TISULARES Y 6RGANOS FORMADORES DE LA SANGRE 211
bulos blancos comprende entre 20,000 y 150,000 por milímetro cúbico en los diferentes grupos de peces. Entre los glóbulos blancos se cuentan los granulocitos que forman entre el 4 y el 40% de todos los corpúsculos blancos. Su diámetro promedio es de casi 10 micras pero queda comprendido entre 24 a 33 micras en el pez pulmonado de Africa (Protopterus). Los granulocitos se dividen, de acuerdo a sus propiedades de tinción, en neutrófilos, que son los más comunes, acidófilos (eosinófilos), y basófilos, que son raros en la sangre de los peces. Algunos elasmo- branquios poseen un cuarto tipo de granulocito, el heterófilo. También están presentes linfocitos y monocitos agranulares, así como trombocitos ovales, gene- ralmente pequeños. Los leucocitos agranulares son los componentes celulares blancos más numerosos en la sangre de los peces. Los monocitos tienen una fun- ción macrófaga. Los linfocitos parecen diferenciarse en dos grandes poblaciones, una dedicada a la producción de anticuerpos y la otra a la inmunidad celular. Los trombocitos equivalen a cerca de la mitad de todos los leucocitos del pez y están encargados de coagular la sangre.
Formación de la sangre
En los vertebrados de sangre caliente la formación de las células sanguíneas está restringida a la médula de los huesos, el bazo, y los nódulos linfáticos; muchos otros órganos participan en la formación de los glóbulos (hematopoiesis) en peces y anfibios.
En las primeras etapas embriológicas de los peces se diferencian los vasos sanguíneos y, a partir de éstos, las células sanguíneas también. Las células san- guíneas pueden aún ser formadas, en el adulto, a partir de los tegumentos de los vasos sanguíneos, pero han aparecido otros centros formadores de células san- guíneas, muy distintos. Todas las diferentes clases de células sanguíneas son for- madas en el bazo difuso que se encuentra en la submucosa del tracto digestivo de las lampreas y los mixinos (Cyclostomata); pero otras regiones hematopoié- ticas difusas también se encuentran, probablemente, en varios vasos sanguíneos. Los eritrocitos y los granulocitos se forman en el arco protovertebral, dorsal a la notocuerda.
En los peces gnatostomados hay un bazo diferente; está más o menos divi- dido en una corteza externa de color rojo y una médula interna, blanquecina. El bazo fabrica los eritrocitos y los trombocitos en su zona cortical, y los linfocitos y algunos granulocitos en su región medular. Los glóbulos rojos también son destruidos en el bazo de los peces óseos superiores (Actinopterygii); no se sabe si otros órganos también tienen la función de descomponer la sangre o cómo se pro- duce su descomposición en los peces amandibulados (Agnatha) o en los tiburones y rayas (Elasmobranchii). Los trombocitos se forman en el riñón mesonéfrico de los peces (Capítulo 9) y los granulocitos nacen en la submucosa del tracto diges- tivo, el hígado, las gónadas y el riñón mesonéfrico. Los tiburones, rayas y quime- ras (Chondrichthyes) tienen, bajo la submucosa del esófago, adherencias planas y amarillas de tejido conectivo suelto llenas de leucocitos que ahí mismo son produ- cidos; en caso de que el bazo sea extirpado esta estructura, llamada órgano de Leydig, se apropia la función de producir eritrocitos. La válvula espiral del intes- tino de los Chondrichthyes y los peces pulmonados (Dipnoi) produce muchos ti- pos de leucocitos. En los esturiones (Acipenser), en los peces espátula (Polyodon)

212 SANGRE Y CIRCULACI6N
y en el pez pulmonado de América del Sur (Lepidosiren), el corazón está rodeado de tejido lobular, café rojizo y de carácter esponjoso, donde se producen linfoci- tos y granulocitos. Los elementos celulares de los cartílagos craneales de algunos tiburones (Squaliformes), de las quimeras (Chimaeridae), de los huesos cefálicos grandes de los peje lagartos (Lepisosteus) y de los peces de aleta ondulada (Arnia), también producen todos los tipos de células sanguíneas, de tal modo que se anti- cipan a la función hematopoiética de la médula ósea de los vertebrados superiores.
Sangre y enfermedades
En los peces, tal como sucede en los humanos y otros vertebrados, el aparato circulatorio juega un papel muy importante en el combate de las enfermedades infecciosas. Probablemente los anticuerpos que se forman contra los antígenos específicos aparecen en el hígado, riñón, bazo y timo, pero en las lampreas (Petro- myzonidae) no se forman porque no tienen un bazo y un timo bien definidos. La inmunoglobulina de las lampreas es una molécula, única entre las imnunoglobulinas que se han descrito para los vertebrados. Tiene una estructura tetramérica, no posee ligaduras covalentes entre las cadenas, carece de una estructura en cadena, pesada y liviana, y tiene una hélix alfa prominente. En general, los peces, después de sufrir una estimulación antigénica forman fagos neutralizadores y anticuerpos aglutinantes. Se ha demostrado que sólo dos especies de tiburones y dos de ho- lósteos, el pez de aleta ondulada (Arnia calva) y el catán de Florida (Lepisosteus platyrhincus) producen precipitina. Se sabe, asimismo, que existen antitoxinas y lisinas en la sangre de los peces.
También en un pez existen diferencias individuales en lo que respecta a la resistencia a las enfermedades, siendo más resistente a ciertas bacterias que a otras. Estas diferencias están basadas en las variaciones genéticamente inducidas en el sistema de producción de anticuerpos y, probablemente, sobre todo, en otras dife- rencias leves en la bioquímica individual del cuerpo. Por lo tanto, han sido produ- cidas varias especies de truchas (Salmoninae) resistentes a las enfermedades, pero también es posible inocular a peces especialmente valiosos, contra enfermedades específicas. Pueden ser empleadas cepas débiles (atenuadas) de bacterias vivas o, como se hace con más frecuencia, aplicarse inyecciones repetidas de suero que con- tiene bacterias muertas y sus antígenos. El pez que ha recibido este tratamiento adquiere entonces una inmunidad pasiva temporal.
Otro aspecto de la infección consiste en el cambio que ocurre en el espectro sanguíneo, tan útil en el hombre para diagnosticar algunas enfermedades. Cuando los peces están infectados, ocurren cambios en el contenido de la hemoglobina de la sangre, así como en el número de corpúsculos sanguíneos blancos y rojos. Por ejemplo, en el caso de que la carpa (Cyprinus carpio) adquiera una enferme- dad común con inflamación debida a la bacteria Pseudomonas punctata, la sangre puede mostrar los siguientes cambios: los linfocitos decrecen en número, de 92.0 a 49.4% en relación a todos los corpúsculos blancos; los monocitos aumentan de 5.7 a 38% y los leucocitos polimorfonucleares del número normal, 2.3, al 12.6%; el número de eritrocitos decrece desde más de dos millones por milímetro cúbico de sangre a menos de un millón, y el contenido de hemoglobina en la sangre se re- duce considerablemente.

TEMPERATURA DEL CUERPO 213
TEMPERATURA DEL CUERPO
Los peces, como los anfibios y reptiles, son vertebrados poiquilotermos de sangre fría. La temperatura del cuerpo se ajusta pasivamente a la del agua ambiente. El calentamiento o enfriamiento rápidos pueden ser fatales para algunos peces, especialmente para las especies delicadas que viven en aguas frías, como las tru- chas (Salmo) y las truchas de arroyo (Suluelinus). Las dificultades que encuen- tran los peces para ajustarse a los cambios de temperatura se deben no solamente a la baja tolerancia heredada para los cambios termales, sino, especialmente, por lo que respecta a la elevación de la temperatura, a la tensión metabólica o a la respiratoria.
Los cambios de la temperatura ambiente de ligeros a moderados se trans- miten rápidamente a través de todo el cuerpo del pez mediante la transmisión positiva o negativa del calor realizada por medio de los capilares de la piel y la gran capa de capilares de las branquias. Los peces jóvenes, que tienen un radio branquias-superficie del cuerpo muy elevado, se ajustan más rápidamente que los más grandes.
En algunos peces hay un control limitado de la temperatura del cuerpo. Las percas (Perca), la mojarrita (Lepomis) y algunos bagres (fctalurus) reaccionan ante los estímulos externos de frío con unligero incremento de la temperatura delcuerpo.
Los atunes y los tiburones lámnidos pueden mantener la temperatura muscu- lar muy por arriba de la ambiental. El atún grande de aleta azul (Thunnus thymus) puede mantener la temperatura de un músculo rojo entre los 26°C y 32”C, mien- tras que la temperatura ambiente varía entre los 6°C y 30°C. El músculo rojo, que es la parte más caliente del músculo, recibe energía mediante la oxidación de las grasas, y es empleado para mantener la continuidad de la natación. El músculo blanco es empleado para iniciar con violencia la natación, o realizar ésta durante corta duración, y la energía es proporcionada mediante la glucólisis anaeróbica. El atún tiene la capacidad de termorregular debido a la presencia de una red mara- villosa (rete mirubile) o “wondernet”, formada por arterias y venas que nacen de un par de vasos cutáneos que alimentan a los músculos rojos. Los retia actúan para intercambiar calor en contracorriente, de tal manera que la sangre venosa calienta a la arterial penetrando en el músculo y reduciendo la pérdida de calor. Los retia forman una barrera térmica, reteniendo el calor metabólico en los teji- dos y evitando su pérdida en las branquias.
REFERENCIAS ESPECIALES SOBRE SANGRE Y CIRCULACI6N
Black, E.C. 1940. The transport of oxygen by the blood of freshwater fish. Biol. Bull., 79:
Carey, F.G., y J.M. Teal. 1969. Regulation of body temperature by the bluefin tuna. Comp.
Catton, W.T. 1951. Blood cell formation in certain teleost fishes. Blood, Jour. Hematol.,
Good, R.A., J. Finstad, y G.W. Litman. 1972. Inmunology. The biologyof lampreys, 2:405-432.
Grodzinski, Z. 1938. Das Blutgefaessystem. En Bronn ‘s Klassen und Ordnungen des Tierreichs.
215-229.
Biochem. Physiol., 28:205-213.
6 :39-60.
Academic Press, Nueva York.
Band 6, Abt. 1, Buch 2, Teil 2, Leiferung, 1: l -77 .

214 SANGRE Y CIRCULACIdN
Holmes, W.N., y E.M. Donaldson. 1969. The body compartments and the distribution of
&yer, H. 1938. Das Lymphgefaessystem. En Bronn’s Klassen und Ordnungen des Tierreichs.
Randall, D.J. 1970. The circulatory system. En Fish Physiology, Academic Press, Nueva Y01-k.
Root, R.W. 1931. The respiratory function of the blood of marine fishes. Bol. Bull . , 61~427-
Satchell, G.H. 1971. Circulation in fishes. Cambridge University Press. Londres. Skramlik, E. von. 1935. Ueber den Kreislauf bei den Fischen. Ergeh. d. Biol., 2:1-130.
electrolytes. En Fish Physiology, Academic Press, Nueva York. 1:1-89.
Band 6, Abt. 1, Buch 2, Teil 2, Lieferung, 1:78-101.
4:133-172.
456.

Capítulo 8
Respiración
Tal como le sucede a otros animales, una de las necesidades fundamentales de un pez consiste en contar con una provisión adecuada de oxígeno en los tejidos, de tal manera que pueda realizarse la oxidación que le proporcione la energía nece- saria para mantener la vida, El éxito del pez para sobrevivir depende de su capaci- dad para obtener el oxígeno del medio externo con el auxilio de las branquias vascularizadas, los pulmones o la piel, transportar este oxígeno a los tejidos y descargarlo en los mismos. De igual manera, el dióxido de carbono que resulta de la oxidación celular de los compuestos que contienen carbono, debe ser transpor- tado en la sangre y eliminado a través de las branquias u otras estructuras respi- ratorias. En algunos peces sin escamas el intercambio gaseoso con el agua también tiene lugar a través de la piel, y en los embriones de los peces varios tejidos llegan a servir como estructuras respiratorias temporales. Entre estas estructuras pode- mos contar al saco vitelino, que en sus primeras etapas de desarrollo tiene una vascularización elevada y algunas funciones respiratorias. Después de la eclosión, la vascularización transitoria de las aletas pectorales en desarrollo puede auxiliar en su función respiratoria a las branquias. También puede aparecer la vasculari- zación temporal del opérculo para el intercambio de gases, como sucede en el pez de aleta ondulada (Arnia). En los peces pueden aparecer muchas adaptaciones para absorber el oxígeno directamente del aire. Incluso hay modificaciones de las branquias, la cavidad bucal, el intestino y la vejiga gaseosa.
Por lo que respecta a los grupos más importantes de peces, se ha descrito previamente la anatomía de las branquias en forma superficial, pero en este capí- tulo se da énfasis a la función respiratoria de esos órganos, a sus modificaciones para realizar con mayor perfección esa función y a las evaginaciones pneumáticas del tracto digestivo. También se tratará con cierta brevedad la función que realiza la sangre de los peces en el intercambio de gases.
215

<, ' .. : t..
; , O S pare:+ t d r " S Z K < * S de las agalias o ii<;lsat. br-a.ntpjalss de lamprea n~ar i t~a ~ P ~ ~ K I I B ; ~ z u ~ I W ~ ~ ~ W S ) repr~:senta~~ Is condicibn il:ranqEial de todas ]ampreas (PC?. i,r~~;.iyzor~idae). L O S sacos. de las agallas se abrer; hack ia iuz del trac'io digestivo. ("da LUIO est& ceparada del siguiente mediante un diafragtm delgado adherido 2. l,i pared del cuerpo. 11 excepcibn del borde ;nterr,o de: diafragma, ei interior de r.>ada saco de a g d h está cubierto de filamentos dispuestos radialmente y provistos de dobleces securldarios y cruz8dos, de tal manera que aumentan la superficie respiratoria. J,as bolsas branquiales, sostenidas en posici6n por una estructura cartilaginosa branquial en forma de canasto de eelosia o enrejado (Capit.ulo 3 ; Fi- gura 3.131, comunican con el exterior a travks de atrios más o menos pronunciados y cubiertos de epitelio, mediante los poros de agalla o las aberturas branquiales externas. Entre cada atrio branquial y su poro de agalla externo correspondien- te: existe un canal branquia1 corto, oblicuo y dirigido hacia atrás.
Las lampreas raramente usan la boca suctorial para la inspiración, incltisive en los casos en que esta libre, o sea, que no está siendo empleada para la fija- ci6n en a l g h olJjeto o pez huésped. Se han llegado a contar desde cincuenta a setenta contracciones de las bolsas de agalla por minuto en la lamprea de mar (Petrornyzonj cuando está fija a la presa; pero en la lamprea de río (Lampetra ,Fl'uuiatilis) se han registrado desde ciento veinte hasta doscientas contracciones por minuto, siendo necesario recordar que esta especie de lamprea nada con mucha rapidez. El agua respirable penetra a las bolsas siguiendo la corriente, pero es expelida mediante las contracciones de los músculos compresores branquiales que se insertan en el canasto branquial y en las paredes que separan las bolsas; cada poro que da salida a las agallas está rodeado por un esfínter. Tambih es auxiliada la penetración del agua por el alargamiento y contracción del saco nasal (bolsa nasofaringica) que funciona como si fuese un sistema hidrkdico. Por el contrario, el llenado y vaciado alternados de las bolsas branquiales sirve tanibién para promover el intercambio de agua en el saco nasal.
El tipo de "copa de succión", o sea, la estructura representada por la boca succionadora (embudo bucal) de las lampreas, implica que las desigualdades de presiin que aparecen debido a la respiración no se transmiten hacia la cavidad bucal. Cuando la lengua raspadora está trabajando, el embudo bucal es cerrado en su lado posterior por el músculo semianular y las aberturas interiores de las bolsas brmqtiiales son protegidas por el velo. En consecuencia, este es un caso Único entre los vertebrados de respiración branquial, ya que existe una separación casi completa de las vias de recorrido del agua. Entre las bolsas branquiales exis- ten septos que contienen senos venosos sanguíneos, soportes cartilaginosos que provienen del canasto branquia1 y músculos. Cada uno de estos septos interbran- quiales recibe una arteria aferente que proviene, también cada una, del tronco arterial. La arteria aferente se divide en las arterias de la bolsa branquial, una ante- rior y otra posterior, y luego en las arterias de los filamentos. Las arterias de los filamentos se desparraman en capilares y lagunas en las láminas de las agallas, lugar en donde se produce el intercambio de gases. La sangre oxigenada se aleja de las branquias de la lamprea y se dirige hacia la aorta dorsal mediante un sistema de vasos dispuestos en los septos paralelos a las arterias aferentes de las bolsas. La

ESTRUCTURA Y FUNCI6N DE LAS BRANQTJIAS 217
fisiologia del intercambio de gases en los ciclóstomos es en esencia similar a la de los peces superiores.
Los mixinos (Myxiniformes) tienen dos hábitos de respiración diferentes. Cuando un individuo no se está alimentando puede permanecer sumergido en el fango, a excepción de la porción anterior de su cabeza. Cuando el animal se en- cuentra en esta posición, el agua llega a las bolsas branquiales por la cavidad naso- faríngica. Cuando la boca está dentro del alimento, el agua respirable fluye alter- nadamente hacia adentro y hacia afuera de las bolsas branquiales, a través del conducto esofagocutSneo. Este ducto se abre hacia el exterior por detrás de la última bolsa branquial.
Branquias de los tiburones y rayas (Figura 8.1)
Cinco, más raramente seis o siete, son las aberturas branquiales externas que pre- sentan en series ventrales en cada lado, las rayas y especies afines (Rajiformes), y en series laterales los tiburones (Squaliformes). A esta serie de aberturas se le agrega otra por delante, el espiráculo, tanto en los tiburones como en las rayas (Elasmobranchii), y que corresponde a los vestigios de una abertura branquial, la primera y más primitiva de la serie. Hacia dentro del espiraeulo puede persistir un grupo de laminillas branquiales formando una pseudobranquia hioidea que recibe sangre oxigenada, y que contiene células dispuestas de tal modo que sugieren que la pseudobranquia de los elasmobranquios puede no tener función respiratoria, pero sí secretora, corno sucede en los peces óseos (Osteichthyes), o puede tomar parte en la formación de células sanguíneas. En los tiburones, el agua respirable pe- netra a través de la boca, pero en las rayas cavadoras de los fondos marinos el espiráculo admite la mayor parte del agua que fluye subsecuentemente sobre las branquias.
Cada holobranquia está caracterizada por tener septos bien desarrollados provistos de soportes cartilaginosos y músculos de los arcos branquiales indivi- duales; cada lado oral y aboral de los septos presenta una hemibranquia compuesta del tejido propiamente dicho de la branquia (Figura 8.1). Los filamentos bran- quiales primarios y secundarios están presentes en cada hemibranquia. Los extre- mos distales de los filamentos branquiales primarios se desprenden del septo (Figura 8.1) de tal manera que las dos hemibranquias en oposición pueden for- mar una barrera efectiva que obliga al agua a penetrar entre todos los filamentos, cuando busca su salida, como resultado de la succión y presión que ocasionan los movimientos respiratorios. El ciclo respiratorio de los tiburones y de las rayas puede ser dividido en tres fases consecutivas. En la primera fase, los músculos coracohioideos y coracobranquiales se contraen y amplían el ángulo formado por los arcos branquiales y agrandan la cavidad orofaríngea; el agua penetra mediante succión a través de la cavidad bucal y/o por el espiráculo; durante esta fase, las laminillas branquiales son adosadas a la piel por la presión exterior del agua; por lo tanto, las aberturas branquiales exteriores permanecen cerradas. En la segunda fase, los músculos abductores de la mandíbula inferior y los arcos branquiales se relajan, pero los músculos aductores (aductores interarcuales) ubicados entre las porciones superior e inferior de cada arco branquial, se contraen, y la cavidad bucal comienza a funcionar como una bomba de presión. Mientras la válvula oral previene el flujo del agua hacia adelante y fuera de la boca, el agua es desviada

218
O
'O B
RESPIRACI6N
w I
O t. Q ar v)

ESTRUCTURA Y FUNCIdN DE LAS BRANQUIAS 219
para atrás, hacia las hendiduras branquiales internas. La contracción de los múscu- los aductores interarcuales comba la porción oral de los espacios interseptales; la presión hidrostática en las superficies branquiales internas es entonces reducida y el agua es desviada hacia las cavidades branquiales, las que aún permanecen cerradas hacia afuera. En la tercera fase, los músculos aductores interarcuales se relajan, otro grupo de músculos se contrae para estrechar las aberturas bran- quiales internas y el agua es forzada hacia y a través de las laminillas branquiales. Entonces, las aletas de las aberturas branquiales externas se abren pasivamente y permiten que el agua fluya hacia afuera. Sin embargo, muchos tiburones, como los tiburones macarela (Lamnidae), no muestran esos movimientos respiratorios tan pronunciados y solamente mientras nadan pueden utilizar suficientemente agua respirable; se sofocan rápidamente cuando son inmovilizados por la captura u otras causas.
Branquias en los peces óseos (Figura 8.2)
Los peces óseos muestran, en general, la misma disposición básica de sólo una abertura branquial externa, localizada a cada lado de la cabeza y bajo un opérculo que cubre las branquias. Sin embargo, la desaparición del espiráculo, la reducción de las hendiduras branquiales a sólo cuatro y la presencia de una dentición gra- dual entre las dos hemibranquias de cada holobranquia (Figuras 8.1 y 8.2), forman una transición que va desde los peces óseos inferiores de aletas con radios (Chon- drostei) a los superiores (Teleostei). Asimismo, la arteria branquial eferente, bi- furcada, de cada arco aórtico de los tiburones, que hasta cierto punto persiste en los condrosteos, es sustituida por un vaso eferente Único de los teleósteos. En todos los peces óseos los músculos mueven las bases de las hemibranquias, de tal
BRANQUIAS
eferente
(u) Abductado ( b ) Aductado
Figura 8.2 - Secciones transversales diagramáticas a traves de las holobranquias adyacentes de un pez óseo, mostrando los elementos de soporte y musculares que permiten (a) la abducción y (b) la aducción. (Basado de Bitjel en Grass& 1958).

manera que la presión que va hacia l a cavidad oral, y posiblemente también e! flujo de agua que sale de ella, es debidamente regulada (Figura 8.2). Cuando el pez quiere liberar las iaminillas de l a presencia de detritus acumulados produce fuertes aducciones de las dos helnibranquias de una holobranquia, de tal manera que, a: juntarse, producen una especie de “tos” y, como consecuencia, un barrido violento con agua sobre las laminillas Dranquiales.
Aunque hayan perdido el espiritculo, los peces bseos de aletas con radios (Actinopterygii) retienen una pseudobranquia hioidea (Figura 8.3), que es libre en algunos peces, pero en otros está cubierta por la piel. Esta porción de la región branquial es la primera que se desarrolla en el embrión y tiene probablemente una temprana funci6n respiratoria. En los adultos de la mayoría de los peces óseos, sin embargo, la pseudobranquia recibe sangre oxigenada a través de una provisión sanguínea establecida en forma secundaria por la aorta y vasos relacionados, te- niendo también una conexión vascular directa con la glándula coroidea del ojo. Las pseudobranquias no se encuentran en las anguilas y peces relacionados (An- guilliformes), ni en los peces elefante (Mormyridae), los peces gato (Siluroidei), los de lomo plumoso (Notopteridae) y otros grupos especializados en los que también falta la glandula coroidea del ojo.
Cada filamento de la pseudobranquia tiene un soporte en forma de varilla cartilaginosa delgada y vasos sanguíneos en su porción medular, alrededor de la cual se adosan capas de células pseudobranquiales y espacios sanguineos sinusoida- les. Las células pseudobranquiales grandes y granulares, que predoniman en el epi- telio, esttín cubiertas en sus bordes externos por células epiteliares escamosas que forman una especie de pavimento (Figura 8.5). El espacio sanguíneo es sostenido por células en columna cuyas extensiones delgadas forman la cubierta endotelial del espacio propiamente dicho. Dentro de las células de la pseudobranquia hay rnitocondrias en grandes cantidades y estructuras dispuestas en forma de membra- nas tubulares a muy corta distancia. Estos túbulos son extensiones de la mem- brana de plasma y muestran actividad secretora. Los túbulos pueder, corresponder al lugar donde se produce la anhidrasa carbónica que facilita la formación de ácida carbónico a partir del dióxido de carbono y del agua.
Pseudobranquia
Mandíbula’ inferior Radio
branquióstego \ branquial Mlisculo aductor
del opkrcula
Figura 8.3 - Posíci6n de la pseudobranquia como lo muestra el pez europeo de ojo lateral (Stizostetedion lucioperca), familia Percidae. (Fuente: Rauther en Bronn, 1940).

ESTRUCTURA Y FUNCX6N DE LAS BRANQUlAS 221
Debido a la existencia de la conexión vascular directa con el ojo, la elevada tensión de oxígeno intraocular, la similitud de la disposición capilar parecida a la rete mirubile en la glándula coroidea y el complejo secretor de gas de la vejiga gaseosa qus contiene también mucha anhidrasa carbónica, se sospecha que la pseudobranquia participa en el intercambio metabólico de gas en la retina. El tejido que forma la retina es el que, de todos los que forman el cuerpo, demanda más oxígeno para realizar sus funciones. L a pseudobranquia puede tener también una relación funcional, a través de !a producción de anhidrasa carbónica, para llenar la vejiga gaseosa; el ciprinodóntido Fundulus, por ejemplo, no puede llenar ese órgano cuando le son extirpadas las pseudobranquias. Asimismo, ciertos peces de los fondos marinos (Gonostoma bathyphilum, Aphyonus gelatinosus) con las vejigas llenas de grasa en lugar de gas carecen de pseudobranquias, aunque sus parientes más próximos presenten la vejiga normal llena de gas y las pseudobran- quias. La pseudobranquia ha sido también implicada en la regulación de la presión intraocular debido a su conexión vascular directa con la glándula coroidea del globo del ojo.
Cada filamento branquial lleva consigo muchas subdivisiones o laminillas (Figura 8.4) que son el asiento principal del intercambio gaseoso. En algunos pe- ces como las anguilas (Anguilliformes), existen células secretoras acidófilas locali- zadas en las bases de las laminillas. Los bordes libres de las laminillas son extrema- damente delgados, cubiertos con epitelio, y contienen una extensa red de vasos capilares sostenidos en posición por células en columna (Figura 8.5). El número y tamaño relativos de las laminillas determinan el área respiratoria de las bran- quias. El área respiratoria varía mucho de acuerdo con los hábitos de los peces. Los peces marinos pelágicos y de rápido desplazamiento mediante la natación, com.0 las macarelas (Scomber) y las lachas (Brevoortiu), no solamente tienen más laminillas por milímetro de filamento branquial; también poseen un área bran- quia1 cinco veces más grande (en milímetros cuadrados por gramo de peso del cuerpoj que la que se encuentra en los peces sedentarios, como los peces sapo (Opsunus).
El aprovechamiento del oxígeno del agua respirable es mejorado no sola- mente por la subdivisión de los filamentos branquiales en laminillas; también lo incrementa la dirección de la corriente sanguínea y la circulación del agua. Estas circulaciones son realmente un sistema de contracorriente, donde el gra- diente de oxigeno del agua, fluyendo desde el lado oral al aboral de las branquias, decrece a medida que la sangre en las laminillas corre desde los vasos sanguíneos aferentes de las laminillas aborales a los eferentes de las laminillas orales (Figura 8.4). La contracorriente de la sangre y el agua respirable mantienen un gradiente de difusión suave en las branquias para que penetre el oxígeno y se desprenda el dióxido de carbono en el pez. La eficiencia de este sistema de contracorriente en la tenca (Tinca) le da una utilización promedio de 51% del contenido de oxí- geno del agua, mientras que la reversión experimental de la dirección del flujo sobre las branquias, desde la abertura branquial hacia la boca, le da una utilización de sólo el 9%.
El ciclo respiratorio acuático de los peces operculados (Osteichthyes) recuerda burdamente al de los tiburones y rayas (Elasmobranchii), teniendo en cuenta que tanto la succión como la presión empujan al agua a través de las bran- quias (Figura 8.6).
Al comenzar la inspiración, precisamente después de que las cubiertas pro-
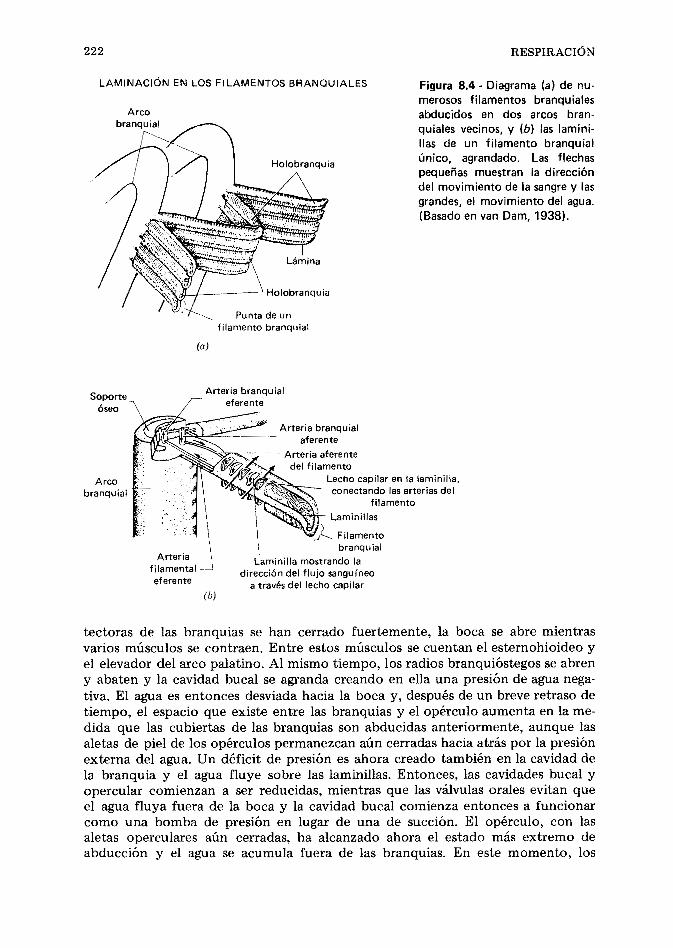
222 RESPIRACIdN
LAMlNAClÓN EN LOS FILAMENTOS BRANQUIALES
Arco branquia!
Soporte bseo
Arco branquial
~ 4 Punta de un filamento branquial
(a)
Figura 8.4 - Diagrama (a ) de nu- merosos filamentos branquiales abducidos en dos arcos bran- quiales vecinos, y (b) las lamini- llas de un filamento branquial Único, agrandado. Las flechas pequeñas muestran la dirección del movimiento de la sangre y las grandes, el movimiento del agua. (Basado en van Dam, 1938).
Arteria branquial
'/ Arteria branquial z- Arteria aferente
/- eferente
aferente
-?A del filamento 7& Lecho capilar en la laminilla,
i? conectando las arterias del filamento
Arteria i, eferente
I branquial
Laminilla mostrando la
a través del lecho capilar filamenta' "' dirección del flujo sanguíneo
(b)
tectoras de las branquias se han cerrado fuertemente, la boca se abre mientras varios músculos se contraen. Entre estos músculos se cuentan el esternohioideo y el elevador del arco palatino. Al mismo tiempo, los radios branquióstegos se abren y abaten y la cavidad bucal se agranda creando en ella una presión de agua nega- tiva. El agua es entonces desviada hacia la boca y, después de un breve retraso de tiempo, el espacio que existe entre las branquias y el opérculo aumenta en la me- dida que las cubiertas de las branquias son abducidas anteriormente, aunque las aletas de piel de los opérculos permanezcan aún cerradas hacia atrás por la presión externa del agua. Un déficit de presión es ahora creado también en la cavidad de la branquia y el agua fluye sobre las laminillas. Entonces, las cavidades bucal y opercular comienzan a ser reducidas, mientras que las válvulas orales evitan que el agua fluya fuera de la boca y la cavidad bucal comienza entonces a funcionar como una bomba de presión en lugar de una de succión. El opérculo, con las aletas operculares a h cerradas, ha alcanzado ahora el estado más extremo de abducción y el agua se acumula fuera de las branquias. En este momento, los

ESTRUCTURA Y FUNCIdN DE LAS BRANQUIAS 223
CBlulas sanguíneas en los capilares
Células sanguí en vasos laminares
Figura 8.5 - Sección a traves de dos laminillas adyacentes de un filamento branquial. (Fuente: Keys y Willrner en Grassd, 1958).
Figura 8.6 - Diagrama del mecanismo para la ventilación branquial por medio de dos bombas; la etapa que se observa ocurre inmediatamente despues de la inspiración. (Basado en Hughes, 1960).
opérculos son rápidamente adosados al cuerpo, las aletas branquiales se abren y el agua es expelida, evitándose su regreso mediante la excesiva presión que existe en la cavidad bucal, comparada a la de la cavidad branquial.
El proceso respiratorio básico descrito anteriormente es modificado de acuerdo a los diferentes hábitos de vida. Los peces de natación rápida como las macarelas (Scombridae), y las numerosas truchas y salmones (Salmoninae), pue- den dejar su boca y las aletas branquiales abiertas para permitir que sus branquias sean bañadas por la corriente de agua producida durante la natación. Por lo gene- ral, las cavidades branquiales de los peces de natación rápida son más pequeñas que las de los peces sedentarios. Los peces del fondo, como los lenguados (Pleu- ronectidae) y los pejesapos (Lophiidae), tienen una cavidad opercular muy agran- dada y distensible; en ellos la boca no se abre mucho durante la inspiración, pero los movimientos de la respiración son lentos y profundos. Los efectos de avance y reduccibn de la fricción producidos por el lanzamiento del agua pueden alguna

224 RESPIRACldN
vez auxiliar en l a locomoción del pez (Capítulo 6 ) o en otras ocasiones contra- rrestar los movimientos de frenado y retroceso, especialmente entre 10s peces cavadores de los fondos marinos que tienden a nadar hacia atrás, mucho más que otros peces.
La salida enérgica del agua a través de la boca mediante el empleo del me- canismo respiratorio como bomba de presión, con la inversión de la dirección del flujo del agua, aparece en el pez chile (Albula uulpes) y los salmonetes (Mullidae). Cuando el pez encuentra un gusano o molusco en sus comederos arenosos, deberá destaparlo, para devorarlo, mediante un chorro de agua que salga por la boca como resultado de la rápida aducción de las estructuras cobertoras de las bran- quias. Entre los cavadores de los fondos de los arrecifes de coral se encuentran las morenas (Muraenidae) que llevan casi continuamente la boca abierta mientras respiran. El aparato opercular que presenta efectos de bomba es pequeño, el meca- nismo branquióstego está muy reducido y el agua respirable es en gran parte impul- sada por el efecto de la bomba bucal. El flujo del agua hacia afuera de la boca en las morenas es evitado gracias a la presencia de pequeños esfínteres ubicados en las aberturas branquiales internas. Estas aberturas llevan a una gran cámara bran- quial carnosa que va a terminar en una abertura branquial externa muy pequeña.
El esqueleto rígido de peces como el cofre (Ostraciidae) y el globo o tam- bor (Tetraodontidae) restringe la acción respiratoria de la cubierta branquid. Sin embargo, estos peces gozan de una elevada tasa compensatoria de intercambio de agua respirable, adquirida mediante una respiración rápida (mayor de ciento ochenta movimientos respiratorios por minuto).
Los peces que se fijan temporalmente por medio de la boca a las piedras de los arroyos torrenciales, como los peces gato de los arroyos asiáticos (Sisoridae y Akysidae), han solucionado el problema del aprovechamiento del agua mientras se encuentran fijos al sustrato, ya sea suspendiendo la inhalación o bien desarro- llando surcos‘protegidos por barbillas que permiten la absorción del agua con sola- mente una pequeña reducción en la fuerza de succión. La ventilación es realizada casi únicamente mediante la acción opercular. En los girinoquélidos (Gyrinochei- lus) y en los bagres de los Andes (Arges) se encuentra una adaptación extrema para respirar en los arroyos torrenciales. Estos peces pueden tomar y arrojar el agua respirable a través de una hendidura inhalante ubicada en la cubierta bran- quial dividida horizontalmente, mientras se encuentran fijos al sustrato con sus bocas succionadoras (Capítulo 5). Las larvas de algunos peces tienen “branquias externas” transitorias que sobresalen de las aberturas branquiales externas o de la región opercular (Figura 8.7); algunos ejemplos incluyen a las larvas del pez pulmonado de América del Sur (Lepidosiren) y las del pez de los juncos (Polyp- terns). En éstos la provisión sanguínea proviene, para las branquias externas de las larvas, de la arteria hioidea, y el Único pliegue branquial es sostenido en posi- ción por un elemento cartilaginoso; la estructura completa se pierde cuando las branquias verdaderas aparecen al transformarse las larvas en formas adultas du- rante la metamorfosis.
LA SANGRE DE LOS PECES COMO TRANSPORTADORA DE GAS
El oxígeno, cuando pasa de un líquido a otro, se difunde muy lentamente. Por lo tanto, los peces, como ocurre con otros vertebrados, han desarrollado en sus
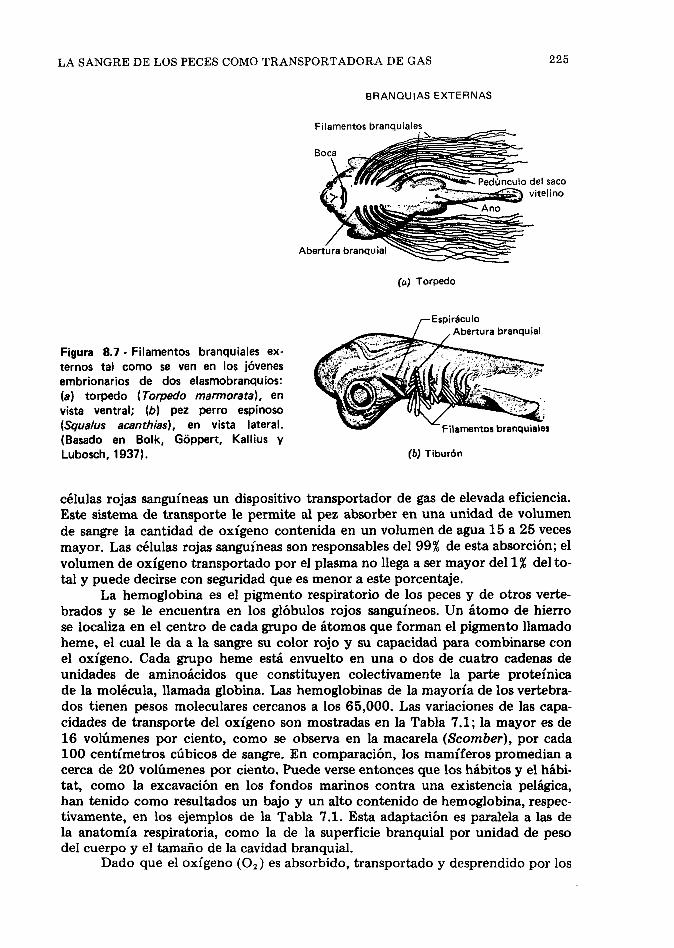
LA SANGRE DE LOS PECES COMO TRANSPORTADORA DE GAS 225
Figura 8.7 - Filamentos branquiales ex- ternos tal como se ven en los j6venes embrionarios de dos elasmobranquios: (a) torpedo (Torpedo marmorara), en vista ventral; (6) pez perro espinoso (Sgualus acanrhias), en vista lateral. (Basado en Bolk, Goppert, Kallius y
BRANQUIAS EXTERNAS
[U) Torpedo
f ,Abertura branquial Espiráculo
W f ¡lamentos branpuiales
Lubosch, 1937). (b) Tibur6n
células rojas sanguíneas un dispositivo transportador de gas de elevada eficiencia. Este sistema de transporte le permite al pez absorber en una unidad de volumen de sangre la cantidad de oxígeno contenida en un volumen de agua 15 a 25 veces mayor. Las células rojas sanguíneas son responsables del 99% de esta absorción; el volumen de oxígeno transportado por el plasma no llega a ser mayor del 1% del to- tal y puede decirse con seguridad que es menor a este porcentaje.
La hemoglobina es el pigmento respiratorio de los peces y de otros verte- brados y se le encuentra en los glóbulos rojos sanguíneos. Un átomo de hierro se localiza en el centro de cada grupo de átomos que forman el pigmento llamado heme, el cual le da a la sangre su color rojo y su capacidad para combinarse con el oxígeno. Cada grupo heme está envuelto en una o dos de cuatro cadenas de unidades de aminoácidos que constituyen colectivamente la parte proteínica de la molécula, llamada globina. Las hemoglobinas de la mayoría de los vertebra- dos tienen pesos moleculares cercanos a los 65,000. Las variaciones de las capa- cidades de transporte del oxígeno son mostradas en la Tabla 7.1; la mayor es de 16 volúmenes por ciento, como se observa en la macarela (Scornber), por cada 100 centímetros cúbicos de sangre. En comparación, los mamíferos promedian a cerca de 20 volúmenes por ciento. Puede verse entonces que los hábitos y el hábi- tat, como la excavación en los fondos marinos contra una existencia pelágica, han tenido como resultados un bajo y un alto contenido de hemoglobina, respec- tivamente, en los ejemplos de la Tabla 7.1. Esta adaptación es paralela a las de la anatomía respiratoria, como la de la superficie branquial por unidad de peso del cuerpo y el tamaño de la cavidad branquial.
Dado que el oxígeno (O,) es absorbido, transportado y desprendido por los

226 RESPIRACI6N
glóbulos rojos sanguíneos, podemos nosotros hablar de los procesos de “carga” y “descarga” del oxígeno y de las correspondientes tensiones que ocurren en esos procesos. Con el fin de contar con las medidas adecuadas para realizar compara- ciones, se han seleccionado dos pasos: 1) T, o Tsat, o sea, la tensión de carga de la sangre, esa presión parcial del oxígeno (O,) a la que la hemoglobina de una espe- cie en particular está saturada en un 95% con oxígeno; 2) Tu o TI/, sat , o sea, la tensión de descarga de la sangre, esa presión parcial del oxígeno (O,) a la que la hemoglobina está saturada en un 50% o, en otras palabras, la tensión de oxíge- no a la cual la mitad de la hemoglobina de la sangre está en el estado oxigenado y la otra mitad en el estado de desoxigenación. La tensión de saturación media es una medida de la afinidad de la hemoglobina por el oxígeno. Si la hemoglobina tiene un T I / * sat bajo es que tiene una elevada afinidad, y viceversa.
La curva de disociación de oxígeno (Figura 8.8) describe el equilibrio del oxígeno y la hemoglobina. La curva puede ser hiperbólica, como la muestra la anguila (Anguillu), o sigmoidea, como sucede en los humanos. La forma de la curva de equilibrio es influenciada por el grado de interacción de las cuatro cade- nas de polipéptidos y sus grupos heme. La estructura tridimensional de las cadenas determina qué residuos de aminoácidos deberá haber en la superficie de las molé- culas y estar disponibles para su agregación subunitaria. La falta de interacción entre hemes origina una curva hiperbólica, mientras que la cooperación de ellos da una curva con tendencia sigmoidal. Una curva hiperbólica con elevada afinidad de oxígeno es característica del pez que puede vivir en el agua con una baja con- centración de oxígeno, como sucede en la anguila. La hemoglobina de la anguila se satura cuando las tensiones de oxígeno son mucho más bajas que las requeridas por los mamíferos, mientras que otros peces, como la macarela del Atlántico (Scomber scornbrus), recuerdan con regular aproximación a los mamíferos en ese aspecto y requieren elevadas tensiones de oxígeno para lograr la saturación. La diferencia entre la Tsat y la sat determina la cantidad total de oxígeno liberada para los tejidos. La sangre, descrita por una curva sigmoidea, puede proporcionar más oxígeno para los tejidos que cuando está descrita por una curva hiperbólica. Por lo tanto, es fácil comprender por qué un pez activo como la macarela ha desa- rrollado una hemoglobina descrita por una curva, de disociación sigmoidea.
100
C ‘O .G ao E
3 60 m U al ‘E 40
*”
S (u
a 20 ; O
Tensión de oxígeno en milímetros
Figura 8.8 - Comparacibn de las curvas de saturación de la hemo- globina como función de la presion de oxígeno. (Basado en Prosser y Brown, 1961).

LA SANGRE DE LOS PECES COMO TRANSPORTADORA DE GAS 227
A medida que aumenta la presión parcial del dióxido de carbono (a menudo señalada como Pco2, como se ve en la Figura 8.8), se requiere mayor tensión del oxígeno para alcanzar el valor Tl y, en consecuencia, disminuye el valor Tu. Este fenómeno, llamado en honor de su descubridor “efecto Bohr”, es a menudo más pronunciado en los peces que en los demás vertebrados y facilita la descarga del oxígeno a las células de los tejidos donde la tensión del CO, es relativamente ele- vada. El grado de protonación de los grupos carboxilos y d-aminos de los amino- ácidos en las cadenas de polipéptidos está determinado por la oxigenación y el enlace del COZ a estos grupos. Se han encontrado grandes diferencias en el efecto Bohr para diferentes especies (Figura 8.9). La macarela (Scornber scornbrus) es un pez de mar adentro que nunca encuentra niveles bajos de O2 y vive bajo tensiones uniformes, pero bajas, de COZ. Su sangre es muy susceptible a los pequeños cam- bios en el dióxido de carbono, mientras que la sangre de la carpa común (Cypri- nus curpio) y especialmente de los bagres (Ictulurus), que viven a menudo en aguas estancadas, es poco afectada por los cambios de nivel del dióxido de carbono. Un incremento en la concentración iónica del hidrógeno (H) (o sea, el descenso en los valores de la escala acidez-alcalinidad del pH) tiene efectos similares para un incre- mento en la tensión del COZ de la capacidad de acarreo de oxígeno de la hemoglo- bina. La forma de la curva de disociación del oxígeno de la hemoglobina del atún cambia con el pH; la cooperación de los grupos heme se pierde cuando el pH baja de 9 a 6. Cuando prevalecen las tensiones elevadas de COZ, siecdo las presiones parciales del oxígeno tan extremas como de 100 atmósferas, no pueden saturar por completo a la sangre (el llamado efecto Root). Un incremento de la tempe- ratura eleva la presión parcial del oxígeno requerida para saturar la sangre; ade- más, la capacidad absoluta de transporte de oxígeno de la sangre del pez es algo más baja a temperaturas elevadas, que a temperaturas bajas.
El metabolismo puede ser definido como el conjunto de cambios químicos en las células vivas, mediante los cuales la energía es puesta a disposición para 10s procesos vitales, y de las actividades mediante las cuales se asimilan nuevos mate-
I I I I
5 10 15 20 pco2, en milímetros Hg
Figura 8.9 - El efecto Bohr en peces de aguas ricas en oxígeno (A) y pobres en el mismo gas (6). (Fuente: Fry en Brown, 1957).

228 RESPIRACIdN
riales para reponer los que se pierden. La tasa metabólica patrón del pez es apro- ximadamente equivalente a la tasa metabólica basal que se mide en los estudios clínicos de los humanos. La tasa de consumo de oxígeno del pez es una medida de su metabolismo.
El consumo de oxígeno puede ser controlado de muchos modos: 1) me- diante la intensidad del metabolismo oxidador a nivel celular; 2) mediante la tasa de agitación que controla el movimiento del agua sobre las branquias y, en conse- cuencia, el gradiente de difusión en esos órganos; 3) por convección interna, o sea, por la velocidad de circulación de la sangre y el volumen que llega a las branquias, y 4) mediante los ajustes a largo plazo en la extensión de las superficies donde se realiza el intercambio respiratorio, o bien por la afinidad de la hemoglobina por el oxígeno.
La tasa de consumo de oxígeno es independiente de la concentración ex- terna de este gas, coincidente con la concentración crítica, o sea, la más baja. Por debajo de la concentracibn o tensión crítica de oxígeno, la absorción de este gas depende de la baja concentración, a un nivel incipiente y letal en el que el pez no puede sobrevivir indefinidamente. La tensión crítica de oxígeno, T,, varía con las especies. Para el “mummichog” (Fundulus heteroclitus), T, vale 16 mm Hg mientras que para el pez globo o tambor (Sphoeroides maculutus) T, es de 100 mm Hg.
La absorción del oxígeno durante el descanso varía con las especies: para el tiburón Scyliorhinus a 15°C llega a ser de 54 cc de oxígenolkg de peso húmedo por hora, y para el “killifish” de California (Fundulus puruipinnis) a 18-21”C, es de 130-230 cc de oxígeno/kg de peso húmedo por hora. En general, el consu- mo de oxígeno aumenta con la elevación de la temperatura a cierto valor crítico, más allá del cual se hacen evidentes algunos efectos daiíinos y la tasa se desploma rápidamente. En algunos peces tiene lugar la aclimatación estacional; por lo tanto, en la zona templada el pez puede tener una tasa más elevada de absorción a 15°C en el invierno que en el verano.
La tasa de consumo de oxígeno es de ordinario más baja en los individuos más grandes de una especie dada. La interrelación entre el consumo de oxígeno y el peso del cuerpo es una función exponencial,
R = kW”
donde R = consumo de oxígeno en volumen de oxígeno consumido por unidad de tiempo, k = una constante, W = peso del cuerpo, y x = un exponente cuyo va- lor varía de 0.6 a 1.0.
Otros factores que pueden influir en el consumo de oxígeno incluyen la activi- dad, la edad y estados fisiológicos como la reproducción, la nutrición, la enfermedad y los mecanismos reguladores intrínsecos (el control nervioso y hormonal).
Los aumentos en las tensiones de dióxido de carbono o las disminuciones en el pH conducen a la disminución del consumo de oxígeno, pero aumentan la tasa de intercambio. N o solamente las facilidades de transporte del oxígeno por la sangre se vuelven menos eficientes cuando hay tensiones elevadas de COZ (efec- to Bohr), el calentamiento del agua aumenta más tarde la respiración debido a que los tejidos demandan más oxígeno cuando las temperaturas son elevadas que cuando son bajas. Este círculo “vicioso” de la inflación respiratoria (cuanto más activa sea para el pez la respiración más elevado será el costo en términos de re-

ADAPTACIONES PARA LA RESPIRACION DEL AIRE ENTRE LOS PECES 229
querimientos de oxígeno) puede conducir a la mortalidad de los peces por asfi- xia. Es en los lugares donde la utilización del agua por el hombre contribuye a elevar la temperatura de ésta y a obtener grandes concentraciones de dióxido de carbono, como sucede cerca de los efluentes de los manantiales termales y de las alcanta- rillas, que en forma muy especial provocan mortandades de peces, las que pueden ocurrir aun cuando la situación del medio esté por arriba de las concentraciones absolutas y letales más bajas de oxígeno disuelto.
El dióxido de carbono es considerablemente más soluble en el agua que el O2 como se muestra por el hecho de que, a 15"C, 1 litro de COZ puede ser disuelto en 1 litro de agua. Por lo tanto, la baja cantidad de dióxido de carbono libre en las aguas naturales favorece la eliminación del gas de desperdicio mediante su difusión en las branquias.
El dióxido de carbono es transportado primeramente en la sangre venosa de los peces en forma de bicarbonatos, pero también puede ir en solución en el plasma. La carbamino hemoglobina de la sangre venosa humana lleva consigo cerca del 7 % de dióxido de carbono. Aunque la carbamino hemoglobina puede estar pre- sente en algunos peces (por ejemplo en la trucha, Salmo), hay evidencias indi- rectas que señalan su ausencia en el tiburón mamón o tiburón gato (Mustelus) y en la carpa (Cyprinus). El desdoblamiento de los bicarbonatos en COZ y agua es catalizado por la enzima anhidrasa carbónica que se encuentra en las células acidó- filas de las branquias, en los glóbulos rojos y en otros tejidos.
ADAPTACIONES PARA LA RESPIRACIdN DEL AIRE ENTRE LOS PECES
Los peces han desarrollado muchos mecanismos para la respiración del aire. Res- piradores del aire, habituales o naturales, aparecen en más de dos docenas de géneros pertenecientes a veinte familias diversas de peces vivientes.
Es generalmente aceptado que la respiración aérea de los peces apareció en asociación con la eficiencia de oxígeno en el medio ambiente; sin embargo, el pez pulmonado de Australia (Neocerutodus), los peces holósteos Amiu y Lepi- sosteus y muchos teleósteos, respiran aire aún en los casos en que haya suficiente cantidad de oxígeno disuelta en el agua. Las deficiencias internas de oxígeno que resultan de las actividades metabólicas elevadas pueden haber producido también una presión selectiva para el desarrollo de la respiración del aire. Las adaptaciones estructurales son de dos clases principales: aquéllas en las que no interviene la vejiga gaseosa y las que cuentan con esa vejiga en calidad de pulmón.
Otras estructuras diferentes a las de los pulmones
Muchos peces de las aguas dulces tropicales y algunos de las saladas, asumen tam- bién un hábito temporal de respiración aérea que les permite salir del agua durante cortos periodos de tiempo o les hace posible tolerar el agotamiento del oxígeno del agua.
Más abajo se enlistan algunas adaptaciones para la respiración aérea en las que no interviene la vejiga gaseosa en forma de pulmón. Branquias no modificadas morfológicamente pueden secretar una sustancia mucosa protectora que es aún capaz de permitir la difusión gaseosa en las anguilas espinosas mastacembélidas

230 R E S P I R A C I ~ N
asioafricanas (Mastacernbelus). La pared interna del opérculo y la cámara bran- quial están provistas de dobleces y muy vascularizadas en los saltarines del fango (Periophthalmus). En estos peces y en los de cabeza de víbora (Channa) se desa- rrollan divertículos en las cavidades bucal y fa.ríngica (Figura 8.10). Los diver- ticulos presentan un epitelio respiratorio con dobleces, muy rico en provisión sanguínea, usualmente procedente de la circulación aferente branquial.
El tracto intestinal propiamente dicho está modificado en un estómago de paredes delgadas que ayuda a la respiración en los peces de armadura, como los Loricariidae. En los “loaches” (Cobitidae) las porciones media y posterior del intestino sirven por sus funciones, tanto digestivas como respiratorias. En algunos de los “loaches” cobítidos la fase digestiva intestinal alterna con la respiracibn en intervalos cortos, teniendo a todas las estructuras formadoras de sangre y 3.
las homeostáticas participando en el ajuste de las fases. En otros de estos “lua- ches” el tracto digestivo no es respiratorio durante el invierno y solamente parti- cipa de l a respiración cuando las aguas están estancadas durante el verano.
El tegumento de la cavidad branquial se extiende en un saco alargado que llega a alcanzar las regiones de la cola, con acceso a la cavidad branquia1 entre el segundo y tercer arcos branquiales, en un pez como Saccobranchus, un miembro de la familia asiática de bagres Saccobranchidae (Figura 8.10). Las blanquias se f’und.en para formar una válvula sobre el orificio de los sacos aéreos. La sangre es proporcionada a través de la arteria V branquial afesente, de aquí que !a estruc- tura no sea homóloga con la de los pulmones de los anfibios, a pesar de que hay cierto parecido superficial con ellos. Otro caso de agrandamiento de la cavid.acl branquial se encuentra en Clarias, de ia familia Clariidae, y en otros géneros rela- cionados de bagres en los que la abertura situada entre los arcos ixanquiales se- gundo y tercero lleva a dos sacos provistos de extensiones arborescentes muy vascularizadas en soportes cartilaginosos que se originan de las branquias I1 y IV. Las paredes de los sacos participan en la respiración aérea y t,oda la estr?.lctura recibe su aporte de sangre de las cuatro arterias branquiales aferentes; la sangre oxigenada regresa a los correspondientes vasos branquiales eferentes (Figura 8.10). Las percas trepadoras (Anabantidae) tienen una estructura respiratoria de aire ligeramente diferente conocida como un laberinto, si tomamos en cuenta que también es arborescente (ramificada). Está cubierta de pliegues de epitelio respi- ratorio y se deriva del primer arco branquial. El laberinto está situado en una bolsa húmeda que ocupa mucho de la región preopercular dorsolateral de la cabeza (Figura 8.10).
La mayoría de los peces con tejidos adaptados para la respiración del aire, como los citados anteriormente, pueden permanecer muchos horas fuera del agua y se les puede ver trepando en la arena seca mediante el auxilio de sus aletas pec- torales, de sus opérculos espinosos o de ambos órganos. Por ejemplo, un pez ca- beza de víbora (Channa) puede vivir fuera del agua durante más de 24 horas y regresar a su vida normal aún después de tener reseca la piel. Algunas especies deben tener acceso al aire atmosférico, aún cuando tengan la suficiente agua para respirar. Es algo todavía desconocido si sus branquias son incapaces de cumplir con su función respiratoria en forma completa o deben emplear sus téjidos adap- tados para la respiración aérea, a fin de deshacerse por sí mismas del dióxido de carbono.

ADAPTACIONES PARA LA RESPIRACI6N DEL AIRE ENTRE LOS PECES 231
Z
u 'O a u a m w [r
Is a a IT
a v) w 2
U
I- a o Q
o a a
n L
O
a

23 2 RESPIRACIdN
Pulmones
Aunque los pulmones originados en la vejiga gaseosa, refiriéndonos a los peces pul- monados (Dipnoi), tienen la misma función respiratoria que la de los pulmones de los vertebrados superiores, las homologías de ambos órganos no es clara. Sin embargo, la mayoría de los especialistas en anatomía comparada creen que los pulmones de los peces pulmonados y la vejiga gaseosa de los peces superiores están relacionados desde el punto de vista evolutivo. Además de la función respi- ratoria, la vejiga gaseosa ha desarrollado muchas otras funciones en la diversifica- ción de los peces, como la adaptación a la gravedad, el equilibrio hidrostático, la producción de sonido y su participación en la percepción del mismo.
Es únicamente en los peces pulmonados (Dipnoi) donde la vejiga gaseosa se ha desarrollado estructuralmente para jugar el papel de pulmón, tal como existe en otros vertebrados (Figura 8.11). Los septos internos, las rugosidades y los pilares dividen los espacios de aire en compartimientos más pequeños que comunican con una cavidad media. Otras subdivisiones de estos compartimien- tos, que consisten en septos reticulados que se van haciendo progresivamente más pequeños, terminan en bolsitas alveolares ricamente cubiertas de vasos sanguí- neos. El conducto pneumático corto y el parénquima pulmonar están ricamente provistos de músculo liso, de gran importancia para el mecanismo respiratorio y puede proporcionar influencia adicional en la distribución del aire en el interior del pulmón. La estructura fina del pulmón, tal como es revelada por el micros- copio electrónico, no difiere básicamente de la que tienen los órganos correspon- dientes de los vertebrados superiores.
Los pulmones de los peces pulmonados se han originado, como los de los demás vertebrados, en una evaginación media ventral del tubo digestivo ante- rior del embrión. El pez pulmonado de Australia (Neocerutodus) tiene un pulmón que se adosa a la pared dorsal del tubo digestivo pero presenta un tubo pneumá- tic0 que se abre en la pared ventral de aquél. En los peces pulmonados africanos (Protopterus) y en los de América del Sur (Lepidosiren), el pulmón es pareado y bilobulado, pero situado en la pared ventral del tubo digestivo.
El aporte de sangre para los pulmones proviene de la última arteria bran- quia1 eferente (la tercera arteria branquia1 fusionada en Protopterus y la cuarta en los otros peces pulmonados), y la sangre oxigenada en los pulmones regresa al corazón directamente a través de una vena pulmonar para verter en el lado iz-
Saculaciones, Trabeculas
Figura 8.11 - Saculacion, ta l como se ve en una porción de la pared de la vejiga gaseosa en un pez pulmonado africano (Proroprerus). (Basado de
¿."- Borde del corte de la vejiga gaseosa ' Spencer en Bronn, 1940).

LA VEJIGA GASEOSA 233
quierdo del atrio (Figura 3.16). También una válvula “espiral” divide el cono arterioso y separa parcialmente las sangres oxigenada y no oxigenada en su dis- tribución hacia los vasos branquiales.
El pez pulmonado africano (Protopterus unnectens) permanece durante el verano dentro de un capullo formado por lodo endurecido de color café y aperga- minado. El animal descansa enroscado en el capullo y con la boca hacia arriba, teniendo sus labios conectados mediante un embudo a un conducto que comu- nica con el exterior. Un pez pulmonado viviendo esta situación durante el verano por lo general permanece, en su capullo rodeado de tierra endurecida, solamente los tres o cuatro meses de la sequía, pero se han registrado casos de hasta un año de duración de la permanencia en el capullo. Durante el veraneo ya mencionado se reduce el metabolismo y el consumo de oxígeno disminuye por consiguiente. Cuando comienza la temporada de lluvias el tubo que comunica el exterior con la boca del pez se llena de agua, el aire no puede llegar más a los pulmones y el animal es “despertado” como resultado de una asfixia incipiente; el capullo es roto posteriormente y a medida que el suelo comienza a ser ablandado por las aguas el pez emerge y emplea sus branquias por el resto del año, hasta que se pre- senta otra vez la temporada seca. Se presume que el pez pulmonado de América del Sur pasa el verano de la misma manera que el africano, pero las especies austra- lianas sobreviven solamente cuando hay agua. Nunca excavan ni forman un capullo.
LA VEJIGA GASEOSA
La vejiga gaseosa, llamada con menos propiedad “vejiga natatoria” o “vejiga aérea”, es un órgano característico de los peces verdaderos y funcionando como un ór- gano hidrostático alcanza su máximo desarrollo entre los teleósteos de radios espinosos (Acanthopterygii). La vejiga gaseosa funciona también como un órgano accesorio para la respiración, la producción de sonido y el desempeño del papel de resonador durante la percepción de éste. Existe la posibilidad de que la fun- ción original de la vejiga gaseosa haya sido la de un pulmón. En ciertas especies emparentadas con el “mouthfish” de las profundidades marinas (por ejemplo el gonostomátido Yurrellu), la vejiga puede servir también como un órgano para el almacenamiento de grasa y posiblemente con funciones de flotación. En la pared de la vejiga hay una estructura especial, el complejo secretor de gas con su rete mirubile o “red maravillosa” de vasos circulatorios y una glándula gaseosa, que permite realizar la función hidrostática. En muchos peces de radios espinosos (Actinopterygii) la reabsorción del gas es también realizada por una estructura especial, el órgano oval situado en la porción posterior de la vejiga gaseosa, en lugar de hacerlo a través de los vasos sanguíneos situados sobre toda la pared superior de la vejiga.
La vejiga gaseosa como órgano respiratorio
Muchos peces fisóstomos emplean la vejiga gaseosa como un órgano suplementa- rio temporal para la respiración. El pirarocú (Arupuimu gigas) del Amazonas es uno de los más grandes peces óseos de agua dulce y llega a medir hasta 5 metros de longitud y a pesar 200 kilogramos. Vive en las ciénegas y tiene la vejiga gaseosa

234 RESPIRACI6N
particularmente bien desarrollada para respirar el aire atmosférico (Figura 8.11). Los peje lagartos (Lepisosteus), los peces de aleta ondulada (Arniu ccaluu), algu- nos mormíridos (Mormyridae) y el pez de cola plumosa (Notopterus) son ejemplos adicionales de respiradores facultativos de aire atmosférico con un tegumento repleto de saquillos o alvéolos en la vejiga gaseosa. Estos peces pueden sobrevivir en el agua desprovista de oxígeno si son capaces de tragar aire y en este caso puede pasar a la vejiga gaseosa vía el conducto pneumático (y en algunos puede salir de la vejiga gaseosa por una abertura ubicada en el ano; Figura 8.12). La importancia relativa de la respiración del aire atmosférico por el pez de aleta ondulada, que vive en las regiones templadas de Norteamérica, depende también de la temperatura del agua. Cuando la temperatura del agua llega a ser de 20 a 25°C el pez regularmente asciende a la superficie en busca de aire, aunque el agua esté bien aireada. Desde el momento en que la vejiga gaseosa de los peces fisós- tomos siempre contiene más dióxido de carbono que el aire, se ha sugerido que la eliminación de este gas de desperdicio se realiza en ese órgano, pero no se sabe hasta qué grado se cumple esta función.
A la vejiga gaseosa le llega sangre proveniente de la aorta dorsal, como sucede en los catanes (Lepisosteus), o de los vasos branquiales, como sucede en el pez de aleta ondulada (Amia). Sin embargo, en la mayoría de los peces la san- gre proviene de la arteria celíacomesentérica; es también común que haya duali- dad en la función realizada por los vasos sanguíneos. La sangre regresa al corazón proveniente de la vejiga gaseosa al verter en una de las venas postcardinales, como sucede en los catanes; o a través de venas pareadas que se fusionan en un solo vaso común antes de penetrar al seno venoso, como ocurre en el pez de aleta ondulada.
Los pececillos del fango (Umbra), que son considerados como respirado- res facultativos de aire atmosférico de aguas temporalmente estancadas, tienen tanto los órganos alveolares como los que absorben y secretan gas, en sus vejigas gaseosas.
Figura 8.12 - Extensión de la vejiga gaseosa hacia regiones del oído interno y el ano en un arenque (Clupea), así como en muchos clupeidos. (Basado de Maier y Scheuring en Bronn, 1940).

LA VEJIGA GASEOSA 23 5
La vejiga gaseosa en la recepción del sonido
La densidad de la carne del pez es tan parecida a la del agua de mar que el animal podría ser virtualmente “transparente” al sonido, si no fuese porque el hueso y la vejiga gaseosa constituyen discontinuidades acústicas por sus diferencias de densidad en relación con la del agua; tanto el hueso como la vejiga gaseosa actúan, por lo tanto, como conductores del sonido o resonadores. En ciertos grupos de peces la vejiga gaseosa puede extenderse hasta la región del oído interno y las variaciones en la presión debidas a las ondas del sonido pueden ser transmitidas directamente a la perilinfa. Es así entonces como en los bacalaos (Gadidae) y los peces pluma (Sparidae) hay una extensión anterior de la vejiga gaseosa que toca los huesos de la cabeza cerca del sáculo del oído interno. En las sardinas (Clu- peidae) hay extensiones de la vejiga gaseosa que crecen dentro de cápsulas carti- laginosas (elementos proótico y pterótico) en aposición cercana a los espacios perilinfáticos de las porciones superior e inferior del oído interno (Figura 8.12); una condición similar es encontrada en el cíclido Etroplus.
La transmisión al oído interno de los cambios en el volumen de la vejiga gaseosa se lleva a cabo de una manera muy especial en el orden Cypriniformes (= Ostariophysi). Gran número de familias, predominantemente de agua dulce, son incluidas aquí. Tienen en común un juego peculiar de huesos pequeños pa- reados u osículos que constituyen el aparato de Weber (Figura 3.25) que conecta la vejiga gaseosa con el oído. Estos osículos derivan de la apófisis de las vértebras anteriores; entre ellas, la más posterior, el tripus, toca la pared anterior de la vejiga gaseosa y está conectada al siguiente hueso mediante un ligamento, el inter- calar o, cuando éste falta, al escafio, el que, a su vez, se fija al diminuto claustro. El claustro de cada lado toca a la ventana membranosa del seno impar que se adosa en el hueso basioccipital de la cabeza y es una extensión del sistema peri- linfático del oído interno. Si el claustro falta, como sucede en los gimnótidos (Gymnotidae), el escafio toca en su lugar la ventana membranosa del seno im- par. Los cambios en el volumen de la vejiga gaseosa provocan que los osículos weberianos se muevan de tal manera que los cambios de presión son transmitidos a la perilinfa y de aquí a las células sensoriales de la porción inferior del laberinto, que es el asiento de la recepción auditiva, En algunas especies, sin embargo, la vejiga gaseosa se encuentra incluida en una cápsula ósea o de tejido conectivo; en estos peces la compresión rítmica de la vejiga gaseosa disminuye momentánea- mente, más que aumenta, la presión en la perilinfa.
Entre los peces cipriniformes, la forma y medida de la vejiga gaseosa varía de acuerdo con la familia. En los pececillos de acuario (Cyprinidae), por ejemplo, la vejiga gaseosa está dividida anteroposteriormente en dos cámaras, con una Cons- tricción entre las dos, y constituida por una acumulación en forma de esfínter del músculo liso que se encuentra en ambas porciones de la vejiga, cerca de la constricción. En los grupos cipriniformes predominantemente cavadores de 10s fondos marinos, como los “loaches” y especies relacionadas (Homalopteridae, Gastromyzonidae y Cobitidae), la porción posterior de la vejiga gaseosa casi des- aparece Y la porción anterior se adosa a una cápsula ósea, y es amortiguado su movimiento por un colchoncillo formado por un líquido gelatinoso. En algunos bagres (por ejemplo, Sisoridae) sólo una cámara anterior de la vejiga persiste, la cual puede estar encapsulada y desplazada hacia afuera, de tal manera que toca a la piel a través de una abertura lateral de la cápsula. Aquí, la disposición me&-

236 RESPIRACION
nica forma una especie de estructura registradora del sonido que recuerda a los registros del tipo de membrana vibradora de los fonógrafos antiguos.
Los peces cipriniformes poseen un amplio margen de percepción del sonido y mucha mejor discriminación del mismo que aquellos peces que carecen del apa- rato de Weber. La desaparición de la vejiga gaseosa en los pececillos de acuario (Cyprinidae) reduce mucho el margen de audición. En los peces que carecen de los osiculos weberianos, los elementos óseos de la cintura pectoral o del cráneo pueden adosarse a la vejiga. En el pez soldado (Holocentrus) y en los peces puer- co (&distes) estas conexiones óseas probablemente desempeñan un papel en la conducción del sonido.
Se piensa que muchos peces son sensibles a los cambios de la presión at- mosférica. Un “loach” (Misgurnus fossilis) es sensible a esos cambios. Sin em- bargo, todavía no se ha demostrado que esos pequeños pero graduales cambios de presión sean percibidos a través de la vejiga gaseosa, los osículos weberianos y el oido interno, o que el pez reaccione a los cambios en otras variables que usualmente marcan una elevación o caída de la presión barométrica, como lo son la lluvia o la cesación de la misma, y cambios en la concentración del oxígeno disuelto. Durante largo tiempo ha existido una controversia sobre si efectiva- mente sucede o no que la vejiga gaseosa y el oído interno, juntos, vuelven a un pez sensible a los cambios pequeños pero rápidos de la presión del agua. Sin embargo, muchas especies de pececillos de acuario (Cyprinidae) pueden detec- tar cambios rápidos tan pequeños como los que corresponden a unos cuantos centímetros de presión. La participación del oído interno en la percepción de esta clase de cambio de presión es anulada mediante la eliminación del órgano por medio de una delicada operación; por lo tanto, la vejiga gaseosa y, más precisamente, su pared, está implicada cor.10 el órgano sensorial.
La vejiga gaseosa en la producción de sonido
El concepto de que los peces son mudos se ha disipado gracias a las grabaciones hidrofónicas que se han realizado a diferentes profundidades y mediante las cua- les se ha demostrado que producen sonido muchas especies variadas de peces y algunos de los organismos invertebrados que les sirven de alimento. De las poco más de 20,000 especies de peces sólo unos cuantos centenares han sido clara- mente identificadas como productoras de sonido. Hay en los peces tres tipos generales de mecanismos sonoros: 1) estriduladores, que producen el sonido mediante la fricción de los dientes (por ejemplo, los roncos, Pomadassyidae), las espinas de las aletas o los huesos; 2) hidrodinúmicos, que resultan de los movi- mientos natatorios, especialmente cuando hay cambios rápidos en la dirección o la velocidad, y 3) la vejiga gaseosa. La vejiga gaseosa es señalada directamente como productora de sonidos en peces como el pez tambor (Sciaenidae), los gra- naderos (Melanonidae) y los angelitos (Triglidae), en los que los músculos estria- dos que se originan en la pared dorsal del cuerpo y se insertan en la vejiga gaseosa, vibran (Figura 8.13). En los peces sapo (Batrachoididae), músculos estriados de la misma vejiga producen cambios rápidos de volumen que desencadenan señales audibles. La vejiga gaseosa muscular que produce sonidos en los peces sapo, es tan efectiva que puede ser extraída del cuerpo para hacerla producir sonido me- diante la estimulación eléctrica. Los sonidos producidos por la vejiga gaseosa son

LA VEJIGA GASEOSA 237
f-- Hacia la cabeza r Pared del cuerpo
Figura 8.13 - La vejiga gaseosa y los músculos sonoríficos que la hacen vibrar para que produzca sonidos, en la trucha marina manchada (Cynoscioh nebulosus), un miem- bro de la familia de los peces tam- bor (Sciaenidae). (Fuente: Tower en Bolk, Goppert, Kallius y Lubosch, 1937). gaseosa
generalmente de bajas frecuencias, mientras que los producidos por los dientes u otras partes óseas tienden a ser de tono elevado.
En la mayoría de las especies cuya vejiga gaseosa emite sonidos, ya sea a través de su propia modificación especial o de estructuras anexas, éstos tienen significación en las actividades reproductoras o en la defensa de sus territorios. Sin embargo, los sonidos de significación problemática pueden suscitarse (general- mente bajo los efectos del stress) en muchísimas más especies de peces en las que se ha visto que tienen una conducta reproductora o de cuidado territorial muy elaborados. Es así como, por ejemplo, los pececillos de acuario (Cyprinidae), los “loaches” (Cobitidae) y las anguilas (Anguillidae), muy excitados, producen soni- dos de tono elevado mediante la expulsión de aire de la vejiga a través del con- ducto pneumático. Se ha llegado a escuchar a las sardinas (Clupeidae) en sus nidos expulsar gas de su vejiga gaseosa a través de un conducto que abre al exterior en el ano (Figura 8.12). Las circunstancias bajo las cuales estos sonidos son produ- cidos señalan que son sonidos de alarma. En los peces de los fondos marinos los sonidos específicos para cada especie pueden ser de significación poco usual por- que la luz, excluyendo a la bioluminiscencia, no existe.
La vejiga gaseosa como órgano hidrostático
La densidad de casi 1,076 @/cm3) de la carne del pez es más grande que la del agua, cuyo valor es de aproximadamente 1.0005 (gr/cm3) para la dulce y 1.026 para la marina. Con el fin de perder por completo el peso en el agua y gastar el mínimo de energía al mantener la posición, el pez deberá acumular grasas y acei- tes en el músculo y en el hígado, o bien deberá usar una inclusión gaseosa. La vejiga gaseosa de los peces óseos (Osteichthyes) es precisamente este órgano y tiene como una de sus funciones la de acercar la densidad total del pez a la del agua que le rodea. Las lampreas y mixinos (Cyclostomata), los tiburones, rayas y especies afines (Elasmobranchii), y los celacantos (Latimeria) vivient,es de aletas lobuladas no tienen vejiga gaseosa. Se sospecha que los tiburones y rayas regulan el equilibrio del cuerpo por medio del ajuste del “lastre de agua” conte- nido en la cavidad del cuerpo y operado a través de los poros abdominales. Entre los peces, la vejiga gaseosa puede formar del 4 al 11% del volumen del cuerpo, 4 a

238 RESPIRACION
6 % en las especies marinas y 7 a 11% en las de agua dulce; la diferencia se debe a las distintas densidades de los dos medios.
La división de los peces en fisóstomos (vejiga con abertura) y fisoclistos (vejiga cerrada) tiene una base tanto funcional como morfológica. Hay un cambio gradual, pero no abrupto, de una condición a otra por lo que concierne a las estructuras secretoras y reabsorbedoras de gas y en los adultos de muchas espe- cies fisóstomas la vejiga gaseosa pierde el conducto pneumático que la conectaba con el exterior en los estados juveniles del pez. Esta condición es llamada parafi- soclista y se le encuentra, por ejemplo, en los peces linterna de las profundidades medias (Myctophidae). Sin embargo, los peces de radios blandos (Malacopterygii) son prevalentemente fisóstomos y los de radios espinosos (Acanthopterygii), fiso- clistos. Los teleósteos fisoclistos verdaderos, con los pércidos (Perciformes) como ejemplo típico, ajustan por entero la presión de sus vejigas gaseosas a través de la secreción o reabsorción de gases adquiriéndolos de o vertiéndolos a la sangre.
La posición de la vejiga gaseosa en relación al centro de gravedad del pez tiene importancia para la natación y el mantenimiento de la posición (Capítulo 6). La perca amarilla (Perca fluuescens), por ejemplo, tiene el centro de gravedad por debajo de la vejiga gaseosa, una disposición que tiende a eliminar el esfuerzo del pez para tomar la posición normal de natación.
El bagre invertido (“upside-down”) (Synodontis batensodu) emplea una vejiga gaseosa desplazada, asegurando así la desusada posición de natación de esta especie.
El llenado y vaciado de la vejiga gaseosa. Aunque en las aguas más naturales y en la sangre arterial de los peces las presiones parciales del oxígeno y el nitrógeno son de cerca de 0.2 y 0.8 atmósferas, respectivamente, las presiones parciales del oxí- geno y el nitrógeno en la vejiga pueden ser, también respectivamente, de 100 y 20 atmósferas. Esta capacidad de la vejiga para concentrar el O2 y el N2 unas 500 y 30 veces, respectivamente, es la única propiedad que tiene el órgano.
En la mayoría de los fisóstomos la vejiga gaseosa debe ser llenada tragando aire al tiempo que se pierde el saco vitelino. Aunque estos peces son capaces de secretar y absorber gas por medio de la sangre cuando son adultos, no pueden llenar la vejiga gaseosa inicialmente sin tener acceso a la atmósfera; los ejemplos incluyen a las truchas y salmones (Salmoninae). Muchos peces fisoclistos tienen conductos pneumáticos durante su vida larval y pueden también tragar aire atmos- férico para llenar por primera vez la vejiga gaseosa; los ejemplos incluyen a las espinochas (Gasterosteus), el “guppy” (Lebistes) y los caballitos de mar (Hippo- campus). La evidencia fósil en los peces de aletas lobuladas (Crossopterygii), la presencia del conducto pneumático larval entre los fisoclistos y la prevalencia de la verdadera condición fisoclista entre muchos de los teleósteos superiores, se ha interpretado como un indicio de que los primeros peces fueron fisóstomos mLs que fisoclistos.
Cualquiera de las formas de los fondos marinos que haya retenido una ve- jiga gaseosa funcional y presumiblemente vive por siempre en las aguas profundas, como los granaderos (Melanonidae) de los abismos, debe haber desarrollado un mecanismo diferente para el llenado inicial de la vejiga gaseosa, a menos que haya pasado cierta etapa de su vida en las aguas superiores accesibles al aire, como sucede con la mayoría de los peces linterna (Myctophidae).

LA VEJIGA GASEOSA 239
El ajuste a las variadas condiciones hidrostáticas, como las que se encuen- tran cuando un pez con vejiga gaseosa realiza migraciones verticales, implica la existencia de estructuras que aumentan o reducen la presión y el volumen de la ve- jiga gaseosa, de acuerdo a las necesidades del animal. Los peces pueden variar el contenido gaseoso de tal modo que el volumen de gas es casi constante a pesar de la presión hidrostática. La ley de Boyle, que establece que el volumen de los gases varía inversamente con la presión, obviamente también se aplica a la vejiga gaseosa. Desde el momento en que la secreción y absorción de gas son procesos fisiológi- cos, hay limitaciones para la velocidad a la que los peces provistos de vejiga gaseosa pueden cambiar su nivel de profundidad. La perca europea (Perca fluuiutilis), por ejemplo, estando a 20 metros de profundidad puede elevarse y sondear rápida- mente y sin molestias, pero sólo dentro de un margen de 16 y 24 metros, ya que el límite de movimiento rápido vertical para esta especie es del 20% de la profun- didad total, sea hacia arriba o hacia abajo de su propio nivel. Las migraciones ver- ticales nocturnas hacia la superficie del océano, mucho más espectaculares, de los peces linterna (Myctophidae) y otras especies de las profundidades medias, que abarcan los 400 metros o más, se deben a las estructuras notablemente agran- dadas y muy eficientes para la secreción y reabsorción de gas de sus vejigas gaseosas.
Otro acontecimiento usual ilustra las limitaciones que una vejiga gaseosa cerrada establece en los movimientos verticales de un pez. Cualquiera que haya enganchado a un pez que haya estado situado a unos cuantos metros de profun- didad habri notado la evaginación del estómago que muestra el animal al ser llevado a la superficie. La explicación consiste en que la vejiga gaseosa se ha ex- pandido rápidarnente al emerger el pez, mucho más rápidamente que la capacidad del mecanismo de reabsorción del gas permitida en esa situación, y una porción del tracto digestivo es empujada hacia afuera a través de la boca del animal.
La transferencia a la luz de la vejiga gaseosa de los gases acarreados en la sangre es lograda a travks de regiones altamente vascularizadas, situadas en la pa- red de la misma vejiga. Algunas partes de la pared de la vejiga, junto con los vasos sanguíneos especiales, forman las dos partes de este complejo secretor de gas (Figura 8.14); ellas son: 1) la glándula del gas, y 2) la red maravillosa. La glándula del gas es una región del epitelio de la vejiga que puede estar formada de una sola capa, doblada, o compuesta de epitelio estratificado y engrosado por muchas ca- pas de células. En este lugar muchos vasos sanguíneos pequeños procedentes de la red maravillosa, cubierta de otros tejidos, invaden las células epiteliales. La red maravillosa o rete mirubile está compuesta de muchos vasos capilares y venillas dispuestas perpendicularmente a la pared de la vejiga. En las vejigas fisoclistas la red maravillosa y el epitelio secretor mantienen posiciones que recuerdan al pe- dúnculo y sombrerillo de un hongo. La sangre es proporcionada por la aorta dorsal a través de una rama de la arteria celíaco-mesentérica y es extraída por una ramificación de la vena porta renal. En una anguila (AnguiZZu), más de 100,000 arteriolas y una cantidad un poco menor de venillas le dan a la red una extensión total de más de 2 metros cuadrados donde los vasos sanguíneos se oponen entre sí. La sangre arterial que va a la vejiga y la venosa que sale de ella están en intimo contacto difuso entre sí. Los sistemas de esta clase, llamados amplificadores de contracorriente, tienen la capacidad de formar diferencias de concentración de varias sustancias desde un lado al otro del órgano (Figura 8.15). En la rete mirubile es principalmente el oxígeno, el gas que va siendo modificado a lo largo de los capilares, hasta llegar a presentar elevadísimas tensiones en las paredes de la vejiga.

240
VEJIGA GASEOSA
RESPIRACIbN
Glándula del Red milagrosa ._
red de la red
Vena Complejo
ejiga seosa
celiacomesenterica porta renal ( 4
Figura 8.14 - L a vejiga gaseosa: (a) posición de la vejiga y la red milagrosa en un pez de mar profundo, y con los dientes oblicuos (Asfronesthes); (b) detalles de la vejiga gaseosa en Astronesthes; (c) provisión sanguínea ge- neralizada de la vejiga gaseosa en peces óseos fisoclistos. (a y b basados en Marshall, 1960, y c en Goodrich, 1930).
Algunos experimentos en los que se detectó la ruta seguida por un isó- topo del oxígeno, desde el agua exterior hasta la vejiga gaseosa, mostraron que la red maravillosa es capaz de concentrar oxígeno en el epitelio de la glán- dula del gas hasta tensiones muchísimo más elevadas de las que persisten en el agua alrededor del pez. Por lo tanto, las células epiteliales de la glándula del gas transfieren los gases disueltos, desde los capilares hasta la vejiga gaseosa. El pro- ceso de transferencia se apoya en la formación intracelular de diminutas bur-

LA VEJIGA GASEOSA 241
Figura 8.15 - Principios de la secreción gaseosa en la vejiga gaseosa de los peces. (a) Ilustración del principio de contracorriente. (b) Un capilar de contracorriente en la red milagrosa. (c) Disposición de los vasos sanguíneos en la red milagrosa. (Sección transversal). (Fuente: Scholander, 1957).
bujas de gas que penetran en la vejiga. Además, las células de la glándula del gas son impermeables a los gases disueltos, aunque por sí mismas pueden secretar burbujas; ellas pueden, en consecuencia, actuar como una barrera para la difu- sión del gas, previniendo el desplazamiento del gas hacia afuera de la vejiga ga- seosa y su entrada a los capilares del epitelio y a la red maravillosa.
El pez blanco (Coregonus), por ejemplo, que vive en las aguas profundas, tiene nitrógeno casi puro en su vejiga gaseosa. Las concentraciones de ese gas, que difieren radicalmente de las que presenta la sangre, confirman que hay una secre- ción celular activa de gas que se origina en la glándula y va a dar a la vejiga ga- seosa, en lugar de haber una acumulación de gas debida a la disociación en los capilares de la rete mirabile del oxígeno procedente de la sangre arterial.
Los peces de las aguas profundas llenan en general la vejiga gaseosa, princi- palmente con oxígeno; cuando los peces soldadito (Trigla) son bajados desde las aguas superficiales a las más profundas, aumenta el porcentaje de oxígeno en sus vejigas gaseosas desde 16% a un metro de profundidad hasta 50% a 8 metros; a 175 metros aparece un 87% de oxígeno en la vejiga gaseosa de un congrio (Conger).
La reabsorción de gas en la vejiga gaseosa se realiza de varias maneras:
a. A través de la difusión hacia los vasos sanguíneos sobre toda la pared de la vejiga gaseosa, fuera de los del complejo secretor de gas, como su-

242 RESPIRACIdN
Figura 8.16 - Aspectos estructurales de la vejiga gaseosa: (a) diagrama del lado ventral de la vejiga gaseosa en un pez boca de mar profundo (Mauro- licus) con detalles sobre la alimentación sanguínea; las venas son negras, las arterias blancas; (6) la vejiga gaseosa en un ”berycoid” de mar profundo (Melamphesf como se ve desde el lado ventral, y (c) el ”oval”, en detalle. (Fuente: Marshall, 1960).
cede en los “saury” (Scombresocidae)* y los “killifishes” (Cyprino- dontidae).
b. A travks de una red capilar reabsorbente especializada, conectada algu- nas veces con el complejo secretor de gas (Figura 8.16), de tal manera que aparece expandida y totalmente activa cuando el pez se eleva. Du- rante ese tiempo la glándula del gas se colapsa y permite una rápida reabsorción del gas y la subsiguiente liberación por encima, a través de las branquias. Cuando el pez desciende la glándula del gas se expande y la red capilar resorbente se colapsa de tal modo que aumenta el área de contacto de la glándula del gas para la secreción rápida de gases den- tro de la vejiga, como se puede ver en los peces hacha (Sternoptychidae).
c. Usualmente, pero no invariablemente -hacia atrás de las vejigas gaseosas de una sola cámara o en la porción posterior de las que tienen dos-,
*Scomberesocidae, según el Catálogo de Peces Marinos Mexicanos. (Secretaría de Industria y Comercio, Subsecretaría de Pesca, Instituto Nacional de Pesca. México. 1976, pp. 74 y 460). (N. del T.).

LA VEJIGA GASEOSA 243
hay en la pared de la vejiga m a región muy delgada donde existe un complejo de capilares separado de la luz de la vejiga por sólo una capa de células epiteliales. Esta área de vasos sanguíneos que provienen de las arterias intercostales y drenan en las venas cardinales, es llamada el órgano oval, porque usualmente está rodeado por un esfínter que con- trola el gasto de reabsorción de gas, mediante la expansión o concentra- ción de un orificio oval en la región de los capilares (Figura 8.16). Al órgano oval se le encuentra típicamente en los bacalaos y especies rela- cionadas (Gadiformes) y en los peces de radios espinosos (Acanthopte- rygii). La evidencia embriológica señala una homología del conducto pneumático de los fisóstomos con el órgano oval de los fisoclistos.
La vejiga gaseosa es inervada por ramificaciones del nervio vago y de los ganglios celíacos. Han sido observadas terminaciones nerviosas en la región re- absorbente, en el óvalo, en la rete y en el epitelio secretor. Cerca del corazón polar de la rete hay numerosas células ganglionares grandes. La capa muscular de la vejiga gaseosa, localizada en la pared, está bien provista por nervios. El fun- cionamiento de este aporte nervioso todavía no se ha comprendido completa- mente. Parece ser que la reabsorción de gas es estimulada por catecolaminas, mientras que la deposición es aumentada por la falta de ese estímulo químico.
La vejiga gaseosa en la distribución y ecología de los peces. Durante la evolu- ción, la vejiga gaseosa de los peces ha asumido varias funciones que ya fueron discutidas, pero en los peces de nuestros días la función más ampliamente repar- tida de este órgano consiste en la regulación de la flotación. La inclusión de gas y la reducción de peso pueden ser en algunos hábitats un riesgo más que una ventaja. Por ejemplo, los peces de los rápidos pueden gastar energía extra en el mantenimiento de su posición preferida, si es que son de la misma densidad del agua, en lugar de ser más pesados. Muchos peces dardo de los arroyos (Etheosto- matinae) han perdido la vejiga gaseosa, mientras que un miembro lacustre de la misma subfamilia de los pércidos, el “logperch” (Percinu cuprodes), retiene el órgano. Los “loaches” del sur de Asia (Cobitidae) que viven en arroyos rápidos de las montañas, comparados con los de los lagos, se ve que han perdido sola- mente la porción posterior de la vejiga gaseosa tripartita, típica del grupo; las dos cámaras anteriores que están conectadas con el oído interno persisten en todos los “loaches” y tienen participación instrumental en la audición (Capí- tulo 11). Estas relaciones sugieren que la adaptación evolutiva a las corrientes rápidas de agua, repetida e independientemente, ha conducido a la pérdida de la vejiga gaseosa.
Posiblemente los ejemplos más sorprendentes sobre la pérdida de la vejiga gaseosa, por un lado, y sus modificaciones, incluyendo su extraordinario desa- rrollo, por el otro, aparecen en los peces de las aguas más profundas de los océa- nos. Aunque se ha muestreado solamente aquí y allá, como consecuencia de las dificultades que se presentan para rastrear con redes en los fondos marinos, hay indicios de que entre los 200 y 1,000 metros de profundidad hay de hecho cantidades extremadamente grandes de peces pequeños mesopelágicos. Su nú- mero es tan alto que son en gran medida responsables de producir las zonas dispersas de sonido que pueden ser detectadas mediante el sonar. Tal como sucede con los peces, la zona dispersa de sonido se encuentra cerca de la superficie de las

244 RESPIRACION
aguas en la noche y a grandes profundidades durante el día. El eco tiene las ca- racterísticas de un reflejo producido por pequeñas burbujas de gas y se cree que son las vejigas gaseosas de los peces las que reflejan las ondas de sonido.
Los representantes más conocidos del complejo de especies mesopekigicas de peces son los peces linterna (Myctophidae); ellos realizan largas migraciones verticales nocturnas hacia las aguas superiores del océano para alimentarse de plancton. Naturalmente que sus predadores, como los peces víbora de los fondos (Chuuliodus) y los tragadores de los fondos marinos (Chiusmodon), también rea- lizan extensas migraciones verticales. Muchas especies mesopelágicas poseen muy grandes y altamente eficientes complejos secretores de gas y redes capilares re- absorbentes o también órganos ovales. De hecho, las vejigas gaseosas de los peces mesopelágicos muestran mucho mejor que las de cualquier otro grupo que la rete mirubile y la glándula del gas son dos partes diferentes del complejo secretor de gas. La extensión relativa del complejo secretor de gas en la vejiga gaseosa de esos peces nadadores tan activos de la superficie de las aguas, como los peces voladores (Exocoetidae), corresponde a un décimo del área de la vejiga gaseosa, pero en los peces linterna (Myctophidae) y otros grupos de las profundidades medias es mucho más grande en extensión. Son estas estructuras agrandadas para el intercambio gaseoso las que permiten las migraciones tan extensas de esos peces de las profundidades medias, las cuales a menudo se extienden más allá de los 3 a 400 metros, hacia arriba durante la noche y otra vez hacia abajo en la mañana.
Las vejigas gaseosas de ciertos peces, como los de la familia Melamphaeidae de la zona baja mesopelágica (500 metros y más abajo), se llenan de grasa; proba- blemente ellos no migran tan extensamente como sucede con algunos otros gru- pos como el de los peces linterna (Myctophidae), y aunque la grasa es más liviana que el músculo o hueso del pez, un espacio del tamaño de la vejiga gaseosa lleno de grasa no puede realmente contribuir para aligerar al pez en su totalidad.
Cuando uno prosigue con la investigación hacia las partes profundas del océano, hasta la zona batipelágica situada entre los 1,000 y 4,000 metros, encon- trará condiciones más precarias para la vida. El alimento está esparcido y, en consecuencia, también los peces. Éstos muestran las adaptaciones típicas que ocurren cuando las condiciones tróficas (alimenticias) son desfavorables. La vejiga gaseosa desaparece, los huesos son reducidos y los músculos son blandos, de consistencia aguada. Las observaciones desde el batiscafo muestran animales letárgicos y de muy pocos movimientos, comparados con los de los peces de la zona mesopelágica, muy activos y rápidos como saetas. Debido a la presión de la columna de agua que se halla sobre estos profundísimos fondos, los gases se han vuelto notablemente más densos que los de la superficie y una vejiga gaseosa debería ser relativamente grande para ser útil, a menos que pudiera generar pre- siones excesivas en su interior; el inflado y el mantenimiento de la presión en esa vejiga gaseosa requeriría mucho trabajo y, por lo tanto, un consumo abundante de alimento como recurso energético. La adaptación evolutiva ha seguido otro camino, aligerando los huesos y la carne del pez en lugar de incluir gas en su cuerpo.
Finalmente, al llegar al fondo del océano, en los dominios batial, abisal y hadal, en cualquier lugar situado entre los 2,000 y 5,000 metros o más, se en- cuentran de nuevo zonas de concentración de alimento, esta vez formado por invertebrados bentónicos y peces que finalmente sobreviven gracias a la acumu- lación de productos originados en la superficie. Entre los peces reaparecen los que están provistos de vejiga gaseosa, constituyendo el 50% de las pocas especies

REFERENCIAS 245
hasta ahora conocidas. Con cierta uniformidad, los peces también adquieren hue- sos más pesados y tejidos blandos más compactos, a diferencia de los que viven so- bre ellos, nadando en aguas poco menos profundas. Los vasos sanguíneos de la rete mirabile, en relación al tamaño de la vejiga gaseosa, de las especies bénticas de las aguas profundas son los más grandes que se conocen. No es de sorprender el alargamiento de esta porción del complejo secretor de gas si recordamos que la función de inflamiento de la vejiga descansa en un principio de contracorriente, aumentando su eficiencia a medida que aumenta la longitud de los vasos opues- tos. Existen suposiciones en el sentido de que una vejiga gaseosa auxilia a muchos de estos peces de los fondos de las aguas marinas más profundas a mantenerse ligeramente sobre el fango mientras buscan su alimento.
Los peces fisoclistos predominan en el ambiente marino donde, inclusive, las familias de peces de las profundidades medias, como la Argentinidae y la Mi- crostomidae, que corresponde a parientes de los fisóstomos de agua dulce como los salmones (Salmonidae), han perdido gradualmente la conexión original entre la vejiga gaseosa y el esófago. A muchos fisoclistos se les encuentra entre los pe- ces de agua dulce, pero los fisóstomos de conductos pneumáticos bien desarro- llados predominan entre la fauna piscícola de agua dulce en la zona de corrientes de agua.
REFERENCIAS ESPECIALES SOBRE RESPIRACION
Black, E.C. 1951. Respiration in fishes. Univ. Toronto Stud. Biol. Ser., 59 (Publ. Ont. Fish. Res. Lab. 71):91-111.
Dendy, L.A., R.L. Deter, y C.W. Philpott. 1973. Localization of Na', K+-ATPase and other enzymes in teleost pseudobranch. I. Biochemical characterization of subcellular fractions. J. Cell Biol., 57:675-688.
Dendy, L.A., C.W. Philpott, y R.L. Deter. 1973. Localization of Na', K+-ATPase and other enzymes in teleost pseudobranch. 11. Morphological characterization of intact pseudo- branch, subcellular fractions, and plasma membrane substructure. Jour. Cell. Biol.,
Fry, F.E.J. 1947. Effects of the environment on animal activity. Univ. Toronto Stud. Biol. Ser.,
Hughes, G.M. 1960. A comparative study of gill ventilation in marine teleost. Jour. Exptl.
Hughes, G.M., y M. Morgan. 1973. The structure of fish gills in relation to their respiratory
Johansen, K. 1970. Air Breathing in fishes. En Fish physiology, Academic Press, Nueva York.
Jones, F.R.H., y N.B. Marshall. 1953. The structure and function of the teleostean swimbladder.
Leiner, M. 1938. Die Physiologie der Fischatmung. Akademishe Verlagsgellschaft, Leipzig. Maren, T.H. 1967. Carbonic anhydrase: chemistry, physiology and inhibition. Physiol. Reu.,
Randall, D.J. 1970. Gas exchange in fish. En Fish physiology, Academic Press, Nueva York.
Steen, J.B. 1970. The swim bladder as a hydrostatic organ. En Fish physiology, Academic
van Dam, L. 1938. On the utilization of oxygen and regulation of breathing in some aquatic
57 ~689-703.
55 (Publ. Ont. Fish. Res. Lab. 68):l-62.
Biol., 37(1):28-45.
function. Biol. Rev., 48:419-475.
4~361-411.
Biol. Rev. Cambridge Philos. Soc., 28:16-83.
47:595-781.
4~253-292.
Press, Nueva York. 4:413-443.
animals. Drukkerij Vollharding Groeningen, Holanda.

246 RESPIRACION
Wittemberg, J., y B.A. Wittemberg. 1961. The secretion of oxygen into the swim-bladder of fish. 11. The simultaneous transport of carbon monoxide and oxygen. Jour. Gen. Physiol.,
Woskoboinikoff, M. 1932. Der Apparat der Kiemenatmung be¡ Fishen. 2001. Juhrb. Abt. Anat. 44(3):527-542.
u Ontog. Tiere, 55:315-488.

Capítulo 9
y regulación osmótica
dRGANOS OSMORREGULADORES Y EXCRETORES (FIGURAS 3.15 Y 3.18)
Los vertebrados eliminan algunos desperdicios metabólicos a través del intestino y la piel, pero la mayor parte es eliminada a través de órganos excretores especia- les: los riñones. Los peces y otros animales acuáticos enfrentan un problema particular en la eliminación, ya que sus branquias y membranas orales son per- meables tanto para el agua como para las sales. La salinidad del agua en el océano es más concentrada que la de los fluidos corporales del pez, y el agua es eliminada, pero las sales tienden a difundirse, concentrándose en el interior; de aquí que los peces marinos beban agua de mar. Por el contrario, tratándose de los peces de agua dulce, éstos pierden sales y absorben agua a través de las branquias porque su concentración salina interna es mucho mayor que la de las aguas circundantes (Figura 9.1).
Muchos desperdicios nitrogenados de los peces pasan a través de los riño- nes, órganos que también participan en el mantenimiento del equilibrio agua-sal (homeostasis) mediante la excreción o retención de ciertos minerales. Las bran- quias también toman parte prominente en la excreción de las sustancias de dese- cho, eliminando principalmente el amoníaco.
El riñón típico de un pez está formado de muchas unidades individuales o nefronas, constando cada una de un corpúsculo renal (cuerpo de Malpighi; Figura 9.3) y un túbulo del riñón. Los túbulos se unen a conductos colectores que final- mente vierten hacia afuera a través del ducto mesonéfrico y sus variadas modifi- caciones terminales. El cuerpo de Malpighi está formado de glomérulos, un vaso sanguíneo fuertemente enrollado, provisto de arteriolas aferentes y eferentes, y
247

, .. ,
i $\ 4 Sangre

6RGANOS OSMORREGULADORES Y EXCRETORES 249
Figura 9.1 - Principales procesos conectados con la osmorregulación en (a) peces de agua dulce, (b) entrada \J salida de las sales y agua en un pez de agua dulce, y (c) regulacibn osrnótica en peces óseos marinos. (Fuente: (a) , Wikgren, 1953, ( b ) y (c), Baldwin, en Brown, 1957). (b) Re- presentacibn esquerniitica de 10s intercambios ibnicos en la c6lula branquia1 de la carpa dorada, Carassius aurarus. Dase, enzimas de dearnidacih y deaminacih; (C.A.) anhidrasa carboni- ea; M a + transporte activo de los ¡ones que requieren energía. (Fuente: Maetz y Garcia R o r n e ~ , 19643.
encapsulado por cblulas rellales deIga;lzs (capsula de Bowmam). Algunos t e l eh - teos tienen rifiones habitualmente con pequeños giomérulos, o sólo unos cua.ntus de tamaiío mediano; ot,ros muestran una tendencia de los glornérulos 3 V Q ~ W ~ W
miís pequefios, menos frecuentes y pobremente vascularizados, como se observa en los “sculpins” cornudos (Myo.xocephaEusj, e incluso existen otros que trenen muy poccrs giomérulos y no son funcionales cuando el animal es adulto; por ejem- plo, el pejesapo (Lophius) pierde Ir, conexión entre los glomérulos y los tiibulos del riñi6n cuando el pez madura. Hay finalmente un número regular de peces marinos, incluyendo a los peces óseos del Antártico, que carecen por entero de los glomérulos (por ejemplo, los peces sapo, Opsanus). Los glomérulos y ¡a cáp- sula actúan en común como ultrafiltros donde la presión de la sangre produce una diálisis no selectiva de moléculas con un peso molecular de hasta 70,000; esto es, las proteínas del suero son retenidas en la sangre. El fluido excretable sufre alteración durante su recorrido a lo largo de los túbulos donde la glucosa, varios minerales, otros solutos y, en algunos casos (“sculpins”, Cottidae), el agua, es reabsorbida hacia la sangre mediante un proceso consumidor de energía. La pre- sencia de glucoproteínas anticongelantes y el aglomerulismo de los riñones jugaron probablemente papeles importantes al permitir a los peces del Antártico adap- tarse a las temperaturas casi congelantes del agua. Desde el momento en que estas glucoproteínas de bajo peso molecular no son filtradas hacia la orina en forma- ción, el pez evita los gastos de energía que serían necesarios para su reabsorción tu- bular. El control de la filtración y la reabsorción tiene lugar a través de la acción hormonal; en los peces la corteza adrenal, la glándula tiroides, la glándula supra- rrenal, las gónadas, el hipotálamo y posiblemente la glándula pituitaria, están relacionados en este proceso regulador (Capítulo 11).
Debe ser puesto énfasis aqui sobre los aspectos funcionales de la excreción del nitrógeno y otros procesos relacionados; los peces de agua dulce, los del mar y los diadromos (los que migran hacia y del mar) son tratados separadamente -como los tiburones, las mantarrayas y las rayas (E1asmobranchii)- debido a que han desarrollado una elevada tolerancia para la urea y, en consecuencia, a través de ella han adquirido una homeostasis osmótica diferente a la de otros peces.
Lampreas de agua dulce y peces óseos
La presión osmótica de los fluidos del cuerpo depende del contenido de compues- tos minerales y orgánicos. En todos los peces de agua dulce esta presión es más elevada que la del agua de los alrededores y existe en el agua la tendencia a difun- dirse en el animal, por cualquier lugar donde tenga una membrana permeable para ese líquido, esto es, las branquias, las membranas orales y la superficie intestinal. La piel y el moco que la recubre reducen mucho la permeabilidad para el agua aunque

260 EXCRECI6N Y REGULACIdN OSM6TICA
hayan pequeñas cantidades que se introduzcan en el pez a través de la piel. La depresión al punto de congelación, A , de la sangre de las lampreas de agua dulce (algunos Petromyzonidae) es de -0.38” a --0.46”C, y la de los teleósteos dulce- acuícolas varía alrededor de -0.57”C cuando el valor de A en el agua del exterior es virtualmente igual a cero.
Para competir con la penetración constante de agua que resulta de las diferencias de tonicidad que hay entre los medios interno y externo, los peces dulceacuícolas producen una cantidad copiosa y muy diluida de orina que es hipotónica en relación al pez. Las lampreas en agua dulce presentan una elimi- nación de orina que normalmente varía entre 15 y 36% del peso del cuerpo por día. Los valores comparables en los peces óseos de agua dulce (Osteichthyes) varían entre el 5 y el 12%. La orina tiene una depresión del punto de congelación ( A ) cercano a -0.025”C para los peces dulceacuícolas; mayores valores de A son raros, aunque se ha registrado el de -0.09”C para un pez pulmonado de África (Protopterus). El trabajo principal del riñón en los peces de agua dulce consiste, por consiguiente, en la excreción del agua; ciertos compuestos nitrogenados que usualmente suman solamente una fracción de la totalidad del nitrógeno excre- tado, también vierten al exterior por intermedio de los riñones y sus conductos asociados. Otras funciones del riñón son la retención de los azúcares y otros solu- tos vitales. Ocurren también algunas pérdidas de sal porque los peces en el agua dulce son hipertónicos para el medio externo. Estas pérdidas tienen lugar al ha- cerse la absorción selectiva de sal a través de las branquias, contra el gradiente natural de difusión.
Los riñones de los peces de agua dulce. Las larvas de las lampreas (Petromyzoni- dae) tienen riñones pronéfricos funcionales hasta la edad en que han alcanzado una longitud de 1 2 a 15 milímetros. Los nefrostomas tienen embudos que abren en el celoma pericardial. La composición de los fluidos del cuerpo está determi- nada por la permeabilidad de la superficie del cuerpo y por el desarrollo del meca- nismo branquia1 para la regularización del equilibrio iónico. Los fluidos celómicos son extendidos en el nefrostoma, a la altura del glomus, o sea, del componente circulatorio del túbulo pronéfrico; las sustancias que se deben conservar son re- absorbidas hacia la sangre y el resto del filtrado acuoso es conducido al seno urogenital (Figura 9.2). Durante la metamorfosis de la larva el riñón mesonéfrico persiste y las aberturas pronéfricas que se abren dentro del celoma desaparecen. La sangre arterial es filtrada a través de los glomérulos sin retención selectiva de
Túbulos F p r o d f r i c o s Túbulos
,r meson6fricos
Nefrostomo GlornBrulos
Figura 9.2 - Riñón de una larva de lamprea (Petrornyzon) de 22 milí- metros mostrando las relaciones de las porciones pronéfrica y meson6- frica. (Basado de Wheeler en Bolk, Goppert, Kallius y Lubosch, 1938).

6RGANOS OSMORREGULADORES Y EXCRETORES 251
Arteriola eferente
Arteriola aferente
Tibulo Glombrulo mesonbfrico
” Aorta
Figura 9.3 - Diagrama de un corpúsculo renal (el glOm6rUlO más la CáP- sula) en el extremo de un túbulo urinífero como se ve en un meSOnefrOS de pez. (Fuente: Smith, 1953).
los solutos, mientras que el túbulo enrollado forma parte instrumental en la re- absorción de las sales y otras sustancias (Figura 9.3).
Con algunas excepciones, como el pez de aleta ondulada (Amiu) y los cata- nes (Lepisosteus) que tienen nefrostomas que se abren en el celoma anterior (Figura 9.4), los peces adultos de agua dulce con aletas de radios (Actinopterygii), tienen un riñón mesonéfrico aislado de la cavidad del cuerpo. Aquí otra vez el exceso de agua de la sangre es filtrado a través de los glomérulos, mientras que la sal y los azúcares son reabsorbidos hacia la corriente sanguínea a través del epitelio de los túbulos del riñón y regresados a los capilares circundantes.
El riñón típico de los peces dulceacuícolas de aletas con radios (Actinop- terygii) tiene muchos glomérulos de mayor tamaño que los del riñón de los peces marinos de aletas con radios, típicos y con riñón glomerulado. El proveedor de sangre está también bien desarrollado. El riñón de los peces de agua dulce es a me- nudo de mayor tamaño, en relación al peso del cuerpo, que el de los peces marinos y las medidas obtenidas atestiguan el paso de grandes cantidades de agua a través del órgano. La cantidad de glomérulos existentes en un riñón calculada a partir de cierto número de especies de agua dulce a menudo excede los 10,000, mien- tras que muchas especies verdaderamente marinas tienen un número muchísimo menor. Estas caracteristicas están relacionadas con el contenido de sal del medio ambiente y por consiguiente con el volumen de orina excretado, como se de- muestra mediante una comparación de los riñones de los salmones (Oncorhin- chus) retenidos en agua dulce contra los de la misma edad en el mar; los prime- ros tienen indiscutiblemente más glomérulos que los segundos. La comparación más reveladora puede ser hecha, sin embargo, observando el diámetro de los pro- pios glomérulos; en muchas especies de agua dulce la medida va de 48 a 104 mi- tras (como promedio 71), mientras que en muchas especies marinas el prome- dio (48) queda dentro de un margen que va de 27 a 94 micras. Llegan a existir algunas variaciones glomerulares extremas; un pez pipa cavador de los ríos tropi- cales (Microphis bouju) no los presenta, y tampoco los tienen los rapes (Lophiidae) que también penetran en las aguas dulces. Los túbulos que drenan las cápsulas de Bowman de los peces dulceacuícolas tienen un segmento anular abierto, por- ciones enrolladas próximas y distales y, en algunas formas, uns región colectora inicial (Figura 9.5).

. ... .-
Conducto meson6frico / /
Abertura A' "Testículo derecho urogenita!
Macho
Figura 9.4 - Diagrama de las relaciones entre riñón, gbnadas, conductos urinarios, seno urogenital, y los vasos sanguíneos de un macho y una hembra de pez holósteo, el catán (Lepisosteus). (Basado en Pfeiffer (macho) y Balfour y Parker (hembra) de Bolk, Goppert, Kallius y LU- bosch, 1938).

Ei r:-GCn del pez cs surtido de sangre por dos vias (Figura 3,1'7; Capitulo 7) : l a arteria renal 3' ias venas poria renales. L a arteria renal manda sangre a los gio- rniirulos rnlentras que 1.2 elevaclo p r e s i h artwia! sanguínea ayuda a producir la filtracibr; g!omwu?ar. L a 5 vena:; pori,a renales es& conectadas 3 la red capilar que rodea !os tG:aaios ,enales y su sangre se u.ne a la que proviene df.1 lecho capilar lo~aliza:l~-t d t - ~ i t r ~ j d e los glorndrulus; toda l a sang" sale de l a regiót-; renal a travks dc: i : t c v"-r:;ts ps!cardma!e!: !.,os rlnones dr. las I a r n p ~ x y ICs rn i .~ i~? :x jl:yclosl.o- mata.) c%:'ecen, sin embargo, ctc la circulac:bn porta ret~a!
. -
!,a oril;a de ](:)S p t . < * e i : aglia <xui<zi r;~~iil~'-ic? ~ , y f : : ~ t i ~ i ~ ~ ::ri+atir:i~~a, COI??-
;>u<:G~o~ n i~~ ;g :?n~ : f~>s 31ri id~n;.if::;ar; p<! ie~ldo W Y i>.igl.ixl;ls cie CGUS i ~ m i r ~ & o i d ~ > s , j :
- 3
. .
un poco de urea y ~ . I E C ~ ~ ~ C C I . L;xs c;.rtntidades de nitrbgei~a urinar:o varian entrt- c\l 2 5 ; (:I ZLi;,; del ~ c r L 3 1 nit~s>.ge:~c excretado poY ¡os p~:cej: ci&xacuicolas. Y ~i vol~xnela ;;ale a l cxt.erior y t r avk de las branquias en forma dc arnoní;:co (56% (3
m i s (in la carpa, Cyprinusj. Ej resto t:stri foranado proba5iemt:nte por urea y otros compu~st,us simpies nitrogcnados que tambikn salen 2 travks t ie braaquias. El voiurnen de orina, corno ya se ha dichr;, depende mucho de l a permeabilidad a! agua de la superficie del cuerpc, las branquias y la mucosa bucal. No todos los peces tienen un moco de l a misma inrpermeabilidad; el agua deberá difundirse hasta veinte veces más rápido en una larnprea (Lampetraj que como lo hace en una anguila (Anguilla), pero solamente hasta dos veces más rápido en la carpa dorada (Carassius); por consiguiente, los volúmenes de orina varían.
El aspecto más importante para la difusión hacia el exterior de las sustan- cias del cuerpo de un pez consiste en la relación que hay entre la branquia y la superficie del cuerpo; los peces pequeños, con una elevada demanda metabólica, tienen superficies branquiales relativamente más grandes que las de sus mayores compañeros de especie. Diferencias pronunciadas en las proporciones de super- ficie branquial a superficie del cuerpo también existen entre las especies y se reflejan en el volumen de orina producido por unidades de peso del cuerpo y tiempo. La relación más desfavorable de superficie branquial a superficie del cuerpo en cualquier pez es la que hay en las lampreas (Petromyzonidae).
La absorción pasiva del agua a través de las branquias y la boca y, en con- secuencia, también la producción de orina, dependen de la temperatura; las lam- preas (Petromyzon) mantenidas entre los 2" y 3°C "achicaron" los excedentes de agua a través de la producción de orina a una tasa de 60 mililitros por kilo- gramo de peso del cuerpo por día, mientras que estando a 18°C la pérdida llegó a ser mayor de 500 mililitros por kilogramo por día. El margen tan extremo de estos valores está comprendido entre el 6 y el 50% del peso del cuerpo: realmente se trata de una corriente copiosa de orina, si consideramos que los peces óseos (Osteichthyes) de agua dulce raramente llegan a acumular en un día más del 20% del peso de su cuerpo de orina, y los mamíferos terrestres sólo el 1.5%.
En conexión con la secreción tan copiosa de orina, muchos peces óseos dulceacuícolas han desarrollado una evaginación posterior, algunas veces promi- nente, de los conductos mesonéfricos que sirven como depósito de la orina; la estructura es frecuentemente considerada como una vejiga urinaria (Figura 3.15).
Algunas sales, especialmente los cloruros, también se pierden en la orina y a través de la mucosidad de los peces. Las branquias y la boca son también otros lugares por donde se pierden los cloruros, y se estima que las pérdidas en estos Ór- ganos igualan a las de las otras zonas del cuerpo. La proporción de las pérdidas urinarias con las del total de los cloruros varía de acuerdo con las diferentes

2 54 EXCRECIdN Y REGULACION OSM6TICA
especies, siendo de 1 : l O O en una lamprea Petromyzon) y ciertos salmones (Sal- moninae), y 1:35 en la carpa (Cyprinus), ya que hay una reabsorción del cloro filtrado en los túbulos renales. Las pérdidas totales de cloruros también varían considerablemente; las pérdidas de la lamprea (Petromyzon) llegan a ser siete veces mayores a las de la carpa (Cyprinus) cuando la salida del cloruro del cuerpo de estos peces es comparada en 100 gramos de tejido por hora.
El papel desempeñado por las branquias de los peces de agua dulce en la reposición de las sales perdidas. En los animales de agua dulce el control osmótico es nece- sario para el mantenimiento de una concentración fija interna de sal, un compo- nente importante de la homeostasis. Las pérdidas de sal varían de acuerdo con las *especies; el salmón del Atlántico (Salmo salar), por ejemplo, puede perder en el agua dulce más del 17% de los cloruros de su cuerpo en un día mediante la difu- sión, mientras que la carpa dorada (Carussius auratus) pierde solamente el 5%. Los peces pueden también abstenerse de beber agua durante considerables periodos, mientras que las especies de agua dulce rara vez lo hacen. Se ha observado, sin embargo, que algunas especies, como la carpa dorada, la consumen en pequeñas cantidades; la carpa (Cyprinus), un pez mucho más resistente a las pérdidas de sal, puede vivir durante algunos días sin recibirla del exterior.
Las branquias y las membranas orales son los lugares donde se realiza la absorción iónica activa necesaria para reponer la sal perdida y suplementar los minerales del alimento. Se ha comprobado que los siguientes iones son absorbi- dos por estas estructuras: litio (Li'), sodio (Na'), cobalto (Co"), estroncio (Sr++), calcio (Ca++), cloro (C1 -), bromo (Br -), fosfato ácido (HP04-) y sulfato (SO,=). Cada uno de estos iones tiene su propio umbral de absorción, pero los datos cuantitativos que se tienen sobre su aprovechamiento son escasos a excep- ción de los del C1- . La lamprea (Petromyzon), la carpa (Cyprinus) y el lucio (Leuciscus, familia Cyprinidae) toman los iones de cloro del agua que contiene menos de 0.05 milimoles de C1 por litro (2 ppm); otros peces de agua dulce, como la perca (Perca), requieren concentraciones más elevadas de cloruros antes de que tenga lugar la absorción activa a través de las branquias. No solamente hay diferen- cias específicas entre las especies de peces que absorben cloro y otros iones, sino que también es muy probable que esas diferencias se extiendan a las razas fisioló- gicas, dependiendo de las características químicas del agua en donde viven. La tasa de absorción corresponde a la tasa de pérdidas por difusión; la lamprea (Pe- trornyzon) absorbe 90 micromoles de C1 por cada 100 gramos de tejido corporal por hora; el salmón del Atlántico (Salmo salar), 30, y muchos pececillos de acua- rio (Cyprinidae), entre 4 y 30. Estos valores muestran que la reposición activa de los cloruros mediante la absorción a través de las branquias puede exceder consi- derablemente a las pérdidas pasivas. Sin duda alguna que muchos minerales son repuestos en cantidades suficientes con el alimento, pero algunos deben contar con mecanismos de reposición similares a los investigados para los cloruros.
En la Figura 9.ld se ilustra un mecanismo para la absorción de Na' y C1- por las branquias de un pez de agua dulce. Los iones hidrógeno (H') producidos por la reacción catalizada por la anhidrasa carbónica reaccionan con el amoníaco (NH3 +) para producir amonib (NH, +). La mayor cantidad de amoníaco se pro- duce aparentemente por la desamidación y desaminación oxidativas en las bran- quias, pero en la sangre deberán existir rastros de amoníaco como resultado de estas reacciones que ocurren en el hígado. Los iones amonio se difunden pasi-

6RGANOS OSMORREGULADORES Y EXCRETORES 255
vamente hacia afuera de las branquias, procedentes de las células de estos órganos respiratorios. En calidad de intercambio y mediante un proceso que requiere de energía, una cantidad equivalente de sodio cargado positivamente (Na’) es in- troducido activamente a través de las branquias. El anión bicarbonato (HC03 -), que resulta de la reacción de la anhidrasa carbónica, se difunde hacia afuera de las células branquiales, mientras que los de cloro (C1 -) son transportados activamente a través de las branquias. El resultado neto consiste en un intercambio entre sodio y amonio, y otro entre cloruro y bicarbonato. Las características de las células encargadas del transporte activo de las sales consisten en tener abundancia de mi- tocondrias y un laberinto muy desarrollado de membranas agranulares (retículo endoplasmático). Los centros autónomos del cerebro localizados en el tallo cere- bral y en el tálamo ejercen una influencia poderosa en la absorción de la sal y se sabe que los peces con médulas espinales seccionadas muestran una reducción severa en la capacidad reguladora de la sal y el agua. El control endocrino del equi- librio de la sal y el agua también ha sido demostrado en los peces. El epitelio bran- quia1 y el de los túbulos renales, apelotonados y proximales, muestran ciertas similitudes fisiológicas, ya que agentes tales como el ion mercurio (Hg) perturban la absorción del Na y del C1 en ambos órganos, posiblemente debido al deterioro de la enzima dehidrogenasa succínica.
Hay muchas diferencias entre los peces de agua dulce en relación a la tole- rancia de la sal, de tal manera que pueden ser dispuestos en grupos, como los “estenohalinos” (relativamente intolerantes a los cambios de salinidad), a los que corresponden generalmente los peces de agua dulce, o “euryhalinos” (tolerantes a los cambios salinos). Entre los segundos están comprendidas muchas especies diádromas como la anguila (Anguillu), el salmón del Atlántico (Salmo salar) y también ciertas espinochas (Gasterosteus) y el ciprinodóntido Fundulus. Entre los peces “estenohalinos” hay variación en la capacidad para ajustarse a una sali- nidad más elevada que la normal; la carpa (Cyprinus) y la carpa dorada (Curussius) pueden tolerar salinidades superiores a 17 partes por millar, equivalentes a un descenso del punto de congelación ( A ) de -0.9”C y también equivalente a una concentración de sales considerablemente mayor que la de sus propios cuerpos. Las concentraciones salinas, tanto del tejido como de la orina, se elevan bajo esas condiciones y el flujo de orina se reduce en consecuencia. Cuanto más leve sea la imposición sobre el pez de esas poderosas condiciones externas, mayor será su tolerancia al cambio. Las capacidades de los peces de agua dulce para adaptarse a las concentraciones anormalmente elevadas de sales no tóxicas dependen tam- bién de factores específicos en las especies, como la relación que hay entre las branquias y la superficie del cuerpo, la histología branquial, el control neurose- cretor y/u hormonal de la permeabilidad de la membrana y los niveles de oxígeno y temperatura. También existen en esta función reguladora variaciones fisiológicas determinadas racialmente en las especies,
Peces marinos
En contraste con lo que sucede en el medio dulceacuícola, los peces marinos viven en un medio que es hipertónico en relación a los fluidos y tejidos del cuerpo y, por lo tanto, ellos tienden a perder agua y obtener sales a través de sus membranas osmóticas (Figuras 9. lc y 9.6). Para contrarrestar las pérdidas de agua, los peces

2 56 E X C R E C I ~ N Y REGULACI6N OSMOTICA
Grupo animal
\ Mamífero acuático 1 Mamífero terrestre \ Ave marina 1 Ave terrestre ! Tortuga marina I Tortuga de agua dulce ! Bacalao marino f Bacalao de agua dulce \ Pez perro marino
Pez sierra de agua dulce ! Mixino marino / Lamprea de agua dulce
Bivalvo marino Bivalvo de agua dulce
j Cangrejo marino I Cangrejo de agua dulce
1
PRESIONES O S M ~ T I C A S
A Mar normal I I
I ! I I I I I I
I I l l I ! ! ! I O 0.4 0.8 1.2 1.6 A
Depresión del punto de congelamiento
( a )
Figura 9.6 - Presiones os- móticas de la sangre y fluidos corporales (a) y va- riaciones en la concentra- ción sanguínea de tres peces en aguas de diferentes concentraciones (b). El tiburón marino es un tibu- rón gato (Scyliorhinus), la anguila marina es un con- grio (Conger), y la anguila de agua dulce (eurihalina)
A Medio externo Anguila. (Fuente: Nicol,
-0.5 O O. 5 1 .O 1.5 2.0 2.5 es la especie europea de
( b ) 1960).
marinos beben agua del mar y, en consecuencia, incrementan el contenido salino de los fluidos del cuerpo. Mientras que la deshidratación es prevenida mediante este proceso, el exceso de sales debe ser eliminado. Este mecanismo homeostático es un circuito consumidor de energía.
La función renal y el equilibrio del agus en los peces marinos. Desde el momento en que los peces marinos son forzados por las condiciones osmóticas a conservar agua, el volumen de orina se ve muy reducido en comparación con el de las espe- cies de agua dulce. Se ha llegado a medir una cantidad tan pequeña como 3 milili- tros de orina por kilogramo de peso del cuerpo por cada 24 horas, con una tonicidad comparable a la de la sangre. Los túbulos renales son aparentemente capaces de retener el agua, según se sabe por los cótidos marinos (Cottidae), porque el fil- trado glomerular es por lo general cinco veces mayor que la cantidad de orina final-

6RGANOS OSMORREGULADORES Y EXCRETORES 257
mente excretada. Los riñones de los peces marinos varían mucho en su estructura; los mixinos (Myxine) tienen un mesonefros primitivo, aunque el pronefros persiste en el adulto, No es clara la función del pronefros. Puede servir en alguna función reguladora iónica, pero no se han llegado a realizar experimentos críticos para examinar esta posibilidad. Como en las especies de lampreas (Petromyzonidae) de agua dulce la circulación a la porción glomerular y tubular de la nefrona (unidad renal) es únicamente arterial, no existe la circulación porta renal venosa. Algunos ciprinodóntidos (Fundulus), cótidos (Cottus) y el saltarín del fango (Periophthal- mus) tienen pronefros funcionales, según se ha reportado, pero el riñón de los peces de aletas con radios (Actinopterygii) es en general verdaderamente mesonéfrico.
Más del 90% de los desperdicios nitrogenados de los peces marinos puede ser eliminado a través de las branquias, principalmente en forma de amoníaco y pequeñas cantidades de urea, pero la orina también contiene trazas de estos com- puestos. Parece ser que el amoníaco está formado de sustancias no tóxicas en el lugar donde se produce la excreción, debido a que es tóxico para los tejidos y solamente aparecen rastros en la sangre. La orina de los peces óseos (Osteichthyes) marinos contiene creatina, creatinina, algunos compuestos nitrogenados sin iden- tificar y óxido de trimetilamina (TMO). Desde el punto de vista metabólico, aún no se ha podido determinar el origen del TMO en los peces. Debido a su presencia, es clara la ventaja osmorreguladora en los fluidos del cuerpo de las formas mari- nas. La posible explicación que se puede dar a la presencia de niveles elevadísi- mos de TMO en las especies marinas, si se las compara con las de agua dulce, po- dría deberse a la presencia de contenidos elevados de TMO en las dietas de los peces marinos, la síntesis del TMO por microorganismos intestinales y las diferen- cias que existen en las capacidades de retención de las branquias y el riñón, así como en la posibilidad de que pueda haber alguna síntesis endógena del TMO.
El equilibrio salino en los peces marinos (Figura 9.6). Los mixinos (Myxinidae) ocupan, entre los ciclóstomos marinos, una posición peculiar en vista de que su sangre tiene una concentración osmótica muy similar a la del agua marina, aunque con un contenido de cloro más bajo; también se ha visto que varían las concen- traciones de urea en la sangre, dependiendo probablemente de que el mixino sea alimentado, antes de la prueba de análisis, con un pez óseo (Osteichthyes) o un miembro del grupo de los tiburones (Elasmobranchii). Ya que son casi o muy iso-osmóticos en relación al agua circundante, los mixinos no necesitan beber agua, y utilizan cualquier cantidad de la que se encuentra en los alimentos para la formación de su propia orina. La sustancia viscosa producida por la piel del mixino es rica en Mg+ + , Ca' + y K + , y puede servir como un mecanismo primitivo para la secreción de estos cationes. Los peces óseos marinos eliminan sus sales exce- dentes, que provienen principalmente del agua de mar y de los alimentos deglu- tidos, a través de las branquias y del tubo digestivo; solamente por medio de la orina se eliminan rastros. Iones univalentes, especialmente los cloruros exceden- tes, pasan del intestino a la corriente sanguínea y son eliminados a través de las branquias, mientras que los bivalentes, como el magnesio (Mg" ) y el calcio (Ca"), por ejemplo, permanecen en el intestino donde la naturaleza alcalina de los fluidos promueve su combinación con los óxidos e hidróxidos, para formar compuestos insolubles que son entonces eliminados con las heces fecales. Las células clorosas o salinas eliminan el exceso de iones de cloro en los peces marinos, mientras que esos iones son absorbidos por las mismas células en los peces de agua

258 EXCRECI6N Y REGULACIdN OSM6TICA
dulce. Estas células han sido localizadas en las bases de las laminillas branquiales como se puede observar en una anguila (Anguillu), y también en las membranas orales, como se ve en ciertos ciprinodóntidos (Fundulus). Hay incertidumbre acerca de la participación en la excreción y absorción iónica de toda la superficie epitelial oral, en contraste con la función de las células secretoras especiales.
Entre las especies de peces óseos marinos, así como en las especies óseas de agua dulce, existen algunas que son más estenohalinas y otras que lo son menos, Puede ser hecho un ajuste de las concentraciones salinas más elevadas o más bajas de lo normal. Los saltarines del fango (Periophthalrnus) a menudo encuentran una salinidad del 45 por millar en esteros aislados y muchas especies marinas pueden ajustarse gradualmente a vivir en aguas salobres e inclusive en aguas casi dulces; la transferencia directa, siguiendo el margen completo de salinidad, desde las 35 par- tes por millar hasta llegar a las O partes, a menudo falla, inclusive en las especies que en otras ocasiones son tolerantes, como el chiro (Chanos). Un factor impor- tante en el ajuste de los peces marinos a las aguas de baja salinidad lo constituye la presencia de una concentración elevada de iones de calcio. Desde el momento en que el calcio baja la permeabilidad celular, tanto para las sales como para el agua, el pez señorita (Halichoeres bivittutus), un pez habitante de los arrecifes (Labridae), puede ser mantenido en forma saludable y alimentarse en una mezcla de un décimo de sal en nueve décimos de agua dulce filtrada a través de arena calcífera de origen coralino.
Muchos experimentos en osmorregulación han sido complicados debido a diuresis temporal de laboratorio. La condición es causada por la simple manipu- lación o cualquier otra alteración del pez y se manifiesta por sí misma por el fun- cionamiento desparejo de las branquias y el riñón, un aumento temporal del flujo de orina y pérdida de iones como el lnagnesio a través de los riñones.
Peces diádromos
Cuando una anguila (Anguillu) alcanza el hábitat de agua dulce en su migración ha- cía los arroyos se encara con los problemas de la disminución de la concentración salina y la hiperhidratación, comparadas con la tendencia hacia la deshidrata- ción y el exceso de sal en el océano. Un salmón joven (Oncorhynchus) migrando corriente abajo hacia el hábitat oceánico se encuentra, por el contrario, con esas condiciones invertidas. Se comprende entonces que, tanto los peces aná- dromos como los catádromos deben ser versátiles en sus ajustes osmóticos, tener riñones glomerulaes que puedan ajustarse a las diferencias en los volúmenes de orina debido a la presencia de diferentes salinidades y poseer membranas bran- quiales y orales capaces de enfrentarse en la absorción y la secreción de ciertos iones, contra los gradientes de difusión existentes. Algunos experimentos realiza- dos con la anguila (Anguilla) han sugerido que las células del cloro pueden tener funciones tanto secretoras como absorbedoras, en forma de unidades. Debe ha- cerse notar que los peces diádromos están ajustados para soportar las fases de agua dulce o de agua salada de sus ciclos de vida, y que dichos ajustes dependen prime- ramente de los cambios fisiológicos determinados genéticamente. La lamprea marina (Petrornyzon), por ejemplo, no puede ajustarse a las salinidades mayores de 17 partes por millar después de haber penetrado al agua dulce para desovar y los salmones jóvenes (Oncorhynchus; Salmo salar) no pueden introducirse con

6RGANOS OSMORREGULADORES Y EXCRETORES 259
éxito al mar antes de que sus células cloradas estén bien desarrolladas. La facilidad de la transición depende de peculiaridades anatómicas como la relación superficie branquia1 a superficie del cuerpo, bien ilustrada en las diferentes razas de la espi- nocha (Gusterosteus) que muestran diferentes tasas de ajuste al agua dulce en sus migraciones para desovar a lo largo de las costas del oeste de Europa.
Los cambios en la actividad endocrina preceden o son usualmente simul- táneos con los cambios en los mecanismos de equilibrio de la sal y el agua; la pituita- ria, la tiroides y las gónadas son glándulas que están primordialmente relacionadas con los cambios en el ajuste fisiológico que ocurren antes y durante la migración. Un incremento en la actividad de la tiroides ha sido reportado para el salmón (Oncorhynchus) durante la migración hacia la parte baja de los ríos, y posible- mente tiene el fin de facilitar el proceso de gasto de energía para la excreción de la sal en el agua del mar, entre algunas otras funciones. Los cambios pituitarios y gonadales a menudo conducen a presentar una conducta de deseo, como sucede con las espinochas (Gusterosteus) durante su estado de maduración, que muestran preferencia por el agua dulce y son, en consecuencia, importantes para llevar el cómputo de las migraciones.
Tiburones, rayas y mantarrayas
Los tiburones, rayas y mantarrayas (Elasmobranchii) ajustan su presión osmótica interna de tal manera que poca o ninguna agua pasa a través de sus membranas permeables; pueden, por lo tanto, evitar algunos de los choques o tensiones osmó- ticos que sufren otros peces. La sangre y los fluidos de los tejidos de los elas- mobranquios marinos tienen una depresióñ del punto de congelación, A , de más de "2.O0C, valor un poco más elevado del que posee el agua de mar. La sangre de los elasmobranquios es, por lo tanto, hipertónica en relación al agua circundante. Esta hipertonicidad se debe en parte a un contenido elevado del ion cloro, más elevado del que se encuentra en la sangre de los peces óseos de aletas con radios (Actinopterygii). Sin embargo, entre el 42 y el 55% de los solutos de la sangre osmóticamente activos son compuestos nitrogenados, principalmente urea. Los tejidos de los elasmobranquios marinos tienen un contenido elevado de urea, algo así como 2 a 2.5%. Poco o nada de urea se pierde en las branquias de los tiburo- nes, un hecho repetido solamente por los túbulos renales de los mamíferos, entre todos los demás órganos de los vertebrados. En los elasmobranquios se presenta una pequeña absorción de agua a través de las membranas branquiales y orales, pero el resto de la superficie del cuerpo es relativamente impermeable al agua. Consecuentemente, sólo una cantidad reducida de orina hipotónica se produce con cierto contenido de urea. Medidas representativas del flujo de orina en los tiburones varían de 2 a 24 mililitros por kilogramo de peso del cuerpo por cada 24 horas, pero el volumen del filtrado glomerular es de cerca de 80 mililitros por kilogramo por día; la reabsorción del agua tiene lugar en los túbulos enrollados proximales. El óxido de trimetilamina (TMO) es el responsable de un 7 a 12% o más de la parte de la presión osmótica producida por los compuestos orgánicos que aparece en los tiburones, las rayas y las mantarrayas. El TMO es importante en la osmorregulación de los elasmobranquios, porque, como sucede con la urea, es reabsorbido del filtrado glomerular en los túbulos renales.
La dieta de agua de los elasmobranquios marinos es regulada primero por

260 EXCRECI6N Y REGULACION OSM6TICA
la absorción de la misma a través de las branquias, de tal manera que la sangre se diluye y aumenta la excreción urinaria. En consecuencia, el contenido de urea en la sangre y en los tejidos disminuye y la excreción de orina se reduce. Esta retro- alimentación negativa puede siempre proveer a los animales de suficiente agua dulce, a fin de resolver sus requerimientos de ese líquido para llevar a cabo sus funciones urinarias, así como otras de carácter metabólico. Las sales son primor- dialmente excretadas por los riñones y la glándula rectal (Figura 3.15a). Los volú- menes de la orina y el fluido de la glándula rectal son casi iguales; la composición de los dos fluidos difiere, sin embargo, porque comparado con la orina, el primer fluido es esencialmente una solución de cloruro de sodio hipertónica con un poco de magnesio (Mg + + ). Menores cantidades de Na + son excretadas por las bran- quias de los elasmobranquios. Alguna reabsorción de sales se realiza en los túbulos renales. Los individuos jóvenes y las especies de agua dulce de los elasmobran- quios tienen problemas osmóticos especiales.
Los tiburones, las rayas y las mantarrayas son vivíparos y protegen sus hue- vos en envolturas impermeables y córneas que contienen urea. En cualquiera de esas situaciones el desarrollo tiene lugar en un medio con elevada concentración de urea y la cría nace con los mecanismos para la retención de ésta. El suero tiene un A de casi -l.O"C, y las sales recuerdan por su composición porcentual a las de los peces óseos (Actinopterygii) de agua dulce, con un A sanguíneo de casi -0.57"C. Las diferencias de -0.43"C o más son el resultado de la presencia de urea en la sangre y los tejidos, y sucede entonces que los elasmobranquios de agua dulce deben tener riñones con abundantes glomérulos y un flujo copioso de orina hipotónica. El nitrógeno es excretado principalmente en forma de amo- níaco a través de las branquias. La urea es también un producto de excreción de los elasmobranquios dulceacuícolas; cerca de un tercio de ella sale a través de los riñones y sus conductos, pero la mayor parte es excretada a través de las bran- quias. Las pérdidas de cloro mediante la difusión y la orina son relativamente elevadas pero no se sabe si los miembros de agua dulce del grupo de los tiburones pueden aprovechar este ion, tomándolo del agua mediante la absorción branquial.
CONTROL ENDOCRINO DE LA EXCRECI6N Y LA OSMORREGULACI6N
El volumen de la orina excretada y el equilibrio de sales es regulado en los peces, como ocurre en los demás vertebrados, mediante las secreciones endocrinas.
Las hormonas pueden influir en el riñón ya sea aumentando o disminuyendo la presión sanguínea de tal modo que se altera el grado de filtración dentro de la cápsula del corpúsculo renal y, en consecuencia, la cantidad de excreción del flui- do. Las hormonas pueden también afectar la excreción renal mediante su acción específica en las células de los túbulos, cambiando las capacidades de permeabi- lidad y reabsorción para determinadas sustancias. En los peces, cuyas branquias y riñones comparten los procesos osmorreguladores, las hormonas pueden tam- bién influir en los procesos de filtración o absorción en las branquias.
Los extractos de la pituitaria posterior contienen en el hombre y en los anfibios fracciones que influyen directamente en la producción de orina mediante la constricción de las arteriolas glomemlares aferentes. Los experimentos que se han realizado con anguilas (AnguilZu) no muestran ese efecto en el equilibrio hí-

CONTROL ENDOCRINO DE LA EXCRECI6N Y OSMORREGULACI6N 26 1
drico, aunque el cipronodóntido Fundulus, habiendo sufrido la hipofisectomía, puede ser mantenido en forma saludable solamente en agua dulce mediante la inyección de extractos de glándula pituitaria. No se sabe hasta qué grado este tratamiento influye en el equilibrio hidrológico, desde el punto de vista de la opo- sición del agua al equilibrio electrolítico.
La información sobre los efectos de los órganos y secreciones endocrinas de los peces en el equilibrio mineral es algo más amplia que la de la producción de orina, aunque todavía falta mucho por investigar. Las hormonas corticales adrenales que influyen en el equilibrio del sodio y cloro en los vertebrados supe- riores juegan también un papel muy importante en la regulación de las funciones renal y branquial de los peces. La sangre de las truchas (Suluelinus; Salmo) con- tiene hidrocortisona y corticosterona, así como otras hormonas corticales y compuestos relacionados. Los corticoesteroides administrados a la trucha dismi- nuyen la eliminación de sodio a través de los riñones y aumentan su excreción a través de las branquias, ya sea mediante un cambio en la permeabilidad del epi- telio branquial o afectando el grado de su absorción. En los peces marinos, que viven en un medio rico en sodio, no es necesario contar con ese mecanismo de conservación del sodio en los riñones, aunque deberá ser benéfico un control de la permeabilidad de las branquias para graduar el movimiento de los iones de sodio. Hay evidencias de que esta función en las especies marinas es, hasta ahora, escasa y no concluyente.
Otros tejidos endocrinos (Tabla 11.3 y 11.4) del cuerpo del pez, implicados en la función relacionada con el equilibrio mineral, incluyen: el hipotálamo, la tiroides, los cuerpos suprarrenales, las gónadas y la urohipófisis.
Las células neurosecretoras localizadas en el núcleo preóptico del hipotá- lam0 pierden rápidamente, en muchos peces marinos, sus secreciones; entre estos peces quedan incluidos los dragonetos (Cullionymus), los lancetas de la arena (Ammodytes) y los moledores (Mugilidae), cuando son colocados temporalmente en agua marina hipertónica. Una hora después de haber sido regresados al agua marina normal las células neurosecretoras hipotalámicas se vuelven a llenar; sigue después, también, una recuperación de las células secretoras en la neurohipófisis.
Las larvas estrictamente acuáticas del saltarían del fango (Periophthulmus) tienen una glándula tiroides menos activa que la de los adultos, quienes llevan un tipo de vida semiacuático. La inyección de tiroides a este pez puede inducirle a dejar el agua por periodos de tiempo mucho más largos. Parece ser que la hormona tiroidea, zparte de influir en otros cambios durante la metamorfosis, afecta en los peces el equilibrio del agua y los minerales, no sólo en la adaptación a la vida tem- poral fuera del agua, sino también durante los cambios del agua dulce a la salada, como ocurre en muchas especies de peces migratorios. Los jóvenes migrantes del salmón, SuZmo salar, que nadan por los ríos hacia el mar, desarrollan células secre- toras de cloro coincidente con, o posiblemente como una consecuencia de, la hiperactividad tiroidea que ocurre durante su desarrollo y por el tiempo en que comienzan con su migración hacia el mar. Todavía no se han hecho investigacio- nes detalladas sobre la manera en que actGan las secreciones tiroideas en los tejidos osmorreguladores de los peces.
Algunos experimentos realizados en anguilas (Anguillu) con branquias uni- das han revelado que la adrenalina, o sea, la hormona producida por las glándulas suprarrenales, tiene un fuerte efecto vasodilatador en 10s Vasos de las propias branquias y reduce, o de plano detiene, la secreción de cloro, función que nor-

26 2 EXCRECI6N Y REGULACI6N OSM6TICA
malmente ocurre en ese lugar. Sin embargo, se desconoce si la adrenalina influye también en el intercambio de cloro en el animal intacto, o sea, el normal.
Tratándose de una especie, hay diferencias entre el macho y la hembra en relación a los niveles de calcio y cloro en la sangre y en el volumen de agua con- tenido en sus tejidos, como sucede en los bacalaos (Gadus), los peces tambor (Tetruoclon) y en el salmón (Salmo). Y pueden ser el resultado de la acción de las hormonas gonadales, desde el momento en que se sabe, por lo que se ha obser- vado en los mamíferos, que las hormonas sexuales femeninas efectúan una acción hidratante por medio de la retención del sodio.
Finalmente, la acetilcolinesterasa, enzima que destruye la acetilcolina, sus- tancia neurohumoral, influye en la absorción del sodio en la piel de la rana y tiene también la misma función en las papilas anales de las larvas de los insectos dípteros. En vista de la distribución tan amplia de la sustancia y de sus efectos com- parables en grupos muy diferentes de animales, puede uno esperar que la acetil- colinesterasa pudo también jugar un papel importante en el equilibrio del sodio en los peces.
REFERENCIAS ESPECIALES SOBRE LA EXCRECI6N Y LA OSMORREGULACIdN
Black, V.S. 1951. Osmotic regulation in teleost fishes. Univ. Toronto Stud. Biol. Ser., 59 (Publ.
Conte, F.P. 1969. Salt secretion. En Fish physiology, 1:241-292. Academic Press, Nueva York. Dobbs, G.H., 111, Y. Lin, y A.L. DeVries, 1974. Aglomerularism in antarctic fish. Science,
Fontaine, M. 1956. The hormonal control of water and salt-electrolyte metabolism in fishes. Mem. Soc. Endocrinol. (Cambridge University Press), 5:69-82.
Forster, R.P., y L. Goldstein. 1969. Formation of excretory products. En Fish physiology, 1:313-350. Academic Press, Nueva York.
Hickman, C.P., Jr., y B.F. Trump. 1969. The kidney. En Fish physiology, 1:91-239. Academic Press, Nueva York.
Krogh, A. 1937. Osmotic regulation in freshwater fishes by active absorption of chloride ions. Zeifschr. Vergl. Physiol., 24:256-266.
Marshall, E.K. 1934, The comparative physiology of the kidney in relation to theories of renal secretion. Physiol. Revs., 14:133-159.
Maetz, J., y F. Garcia Romeu. 1964. The mechanism of sodium and chloride uptake by the gills of a fresh-water fish, Curassius aurutus. 11. Evidence for NH:/Na+ and HC03-/CI- exchanges. Jour. Gen. Physiol., 47:1209-1227.
Pantin, C.F.A. 1931. The origin of the composition of the body fluids in animals. Biol. Revs.,
Robertson, J.D. 1974. Osmotic and ionic regulation in cyclostomes. En Chemical zoology,
Shewan, J.M. 1951. The biochemistry of fish. Biochemical Soc. Symposia (Cambridge, Ingla-
Smith, H.W. 1932. Water regulation and its evolution in fishes. Quart. Rev. Biol., 7:l-26.
Ont. Fish Res. Lab., 71):53-89.
185:793-794.
6:459-482.
8:149-193. Academic Press, Nueva York.
terra), 6 :28-48.
.__ . 1936. The retention and physiological role of urea in the Elasmobranchii. Biol. Reus.,
__ . 1953. From fish to philosopher. Little, Brown and Company, Boston. 11 :49-82.
Wikgren, Bo-J. 1953. Osmotic regulation in some aquatic animals with special reference to the influence of temperature. Acta Zool. Fennica, 71:l-102.

Capítulo 10
Reproducción
La reproducción (Figura 10.1) es el proceso biológico por el cual las especies se perpetúan y, en combinación con los cambios genéticos, aparecen por primera vez características para las nuevas especies.
TIPOS DE REPRODUCCIdN
Son posibles por lo menos tres tipos de reproducción: bisexual, hermafrodita y partenogenética. En la primera, que es la clase que prevalece, los espermatozoides y los óvulos se desarrollan en individuos masculinos y femeninos separados. En el hermafroditismo (un tipo de intersexualidad), los dos sexos se encuentran en un mismo individuo y, como sucede en ciertos serránidos y en una docena o más de familias, existe la autofertilización o verdadero hermafroditismo funcional. Este hermafroditismo sincrónico, visto desde un punto de vista evolutivo, puede ser la forma más ventajosa de reproducción.
Las glándulas sexuales hermafroditas son conocidas en muchas especies entre las que se encuentran las truchas (salmónidos), percas (Perca), “walleyes” (Stizostedion), y el pércido Etheostoma y algunos de los huros (Micropterus). Algunos serranos (Serranidae) son hermafroditas protándricos, dando comienzo como machos con esa función, y luego como hembras.
La partenogénesis consiste en el desarrollo del óvulo sin fertilización, y esa condición que realmente debería llamarse ginegénesis ocurre en un pez tropical, pecílido vivíparo del Amazonas (Poecilia formom); también es conocida en Poeciliopsis. Requiere su apareamiento con un macho, pero el esperma sirve sólo para una de sus dos funciones, la de provocar el desarrollo del óvulo, y no parti-
263

26 4 REPRODUCCI6N
v \ E N T E S 0
Figura 10.1 - Papel de la reproducción en la supervivencia y evolución de las especies.
cipa en ninguna forma en el proceso de la herencia. La cria resultante está for- mada siempre por hembras que no presentan siquiera trazas de caracteres paternos.
EL SISTEMA REPRODUCTOR
La función reproductora en los peces corresponde primordialmente al sistema reproductor. Ya se han descrito previamente los componentes del sistema, las glándulas sexuales o gónadas, los ovarios en la hembra y los testículos en el ma- cho, y sus conductos. El sistema endocrino juega un importante papel en la regu- lación de la reproducción.
LOS ESPERMATOZOIDES Y su FORMACI~N
Al proceso de formación de las células espermáticas en los testículos se le llama espermatogénesis o espermiogénesis. En este proceso es donde quedan bioquímica- mente listos, para su unión con sus correspondientes del óvulo, los materiales que determinan las características que heredará la cria de parte de sus padres. N o solamente el esperma o los espermatozoides de diferentes peces contiene mate- riales hereditarios distintos; ellos difieren también en la forma (Figura 10.2a). Asimismo, es durante este proceso cuando las células espermáticas obtienen un apéndice caudal en forma de látigo que les permite desplazarse hacia el óvulo y penetrar en él para realizar la fertilización. Más aún, durante su desarrollo, los esyermatozoides se diferencian por su forma, de tal manera que esta modifica-

LOS ESPERMATOZOIDES Y SU FORMACIdN 26 5
ESPERMATOZOIDES
Cabeza
m- (Q) LUCiO ( b ) Perca
( c ) Arenque
( d ) Trucha arco iris (e) Trucha cafe - (g) Anguila
Figura 10.2a - Variaciones de la forma en espermatozoides (muy amplifi- cados) de ciertos peces óseos: (a ) lucio del norte (€sox lucius); (6) perca europea (Perca fluviatilis); (c) arenque del Atlántico (Clupea harengus); ( d ) trucha arco iris (Salmo gairdneri); (e) trucha cafe (Salmo trutta); (4 "guppy" (Lebistes reticulatus), y @) anguila europea (Anguilla anguilla). (Basado de Tuzet, Ballowitz, Fontaine y Vaupel en G r a d , 1958).
HUEVOS D E G O B I O
"
Figura 10.2b - Huevos no esfbricos de pez (muy amplificados) tal como se observan en gobios (Gobiidae) seleccionados. (Fuente: Breder, 1943).

26 6 REPRODUCCION
ción resulta útil como característica específica. Con el fin de asegurar la fertili- zación, cada pez macho, como lo hacen los de otras especies de vertebrados, produce enormes cantidades de espermatozoides, tan pequeños que varios millones pueden caber en una pequeña gota. Los espermatozoides y las secreciones de los conductos espermáticos forman el esperma que el pez macho expulsa durante la temporada de ovoposición (como sucede en la eyaculación del semen en el hombre). Los espermatozoides son inactivos e inmóviles hasta que se reúnen con la secreción de los conductos espermáticos. Una vez que se han vuelto activos y han comenzado a desplazarse mediante la agitación de sus colas pronto perecen, a menos que se encuentren con un óvulo para fertilizarlo. El margen de vida de los espermatozoides varía considerablemente de acuerdo a las especies y al sus- trato en que han sido despositados. Si las células espermáticas han sido deposita- das en agua, viven entonces un tiempo mucho más corto, a diferencia del tiempo relativamente largo que llegan a vivir en el interior de los órganos sexuales de la hembra. En el caso de que el agua tenga casi la misma concentración salina que hay en los fluidos del cuerpo del pez, las células espermáticas viven más tiempo, lo que no sucede si el contenido salino de los fluidos del cuerpo es diferente al del medio, ya sea porque éste tenga mayor o menor cantidad de sales.
La 'vitalidad de los espermatozoides es también afectada por la temperatura; por lo general, viven más tiempo cuando las temperaturas son bajas que cuando son elevadas y, tal como sucede con los espermatozoides humanos, pueden ser conservados bajo refrigeración para ser empleados más tarde en la inseminación artificial; en consecuencia, es posible conservar el esperma de los peces para su utilización posterior. Los espermatozoides de los peces, como ha ocurrido con los del hombre, toro, gallo y rana macho, han sido preservados con éxito mediante la congelación; por ejemplo, en la carpa (Cyprinus), en la sardina del Atlántico (Clupea) y en el salmón (Oncorhynchus), sus espermatozoides han permanecido vivos después de su congelación.
LOS ~ V U L O S Y su FORMACION
La ovogénesis es el proceso de desarrollo del óvulo en los ovarios, paralelo al de la formación de los espermatozoides en los testículos. De este proceso no solamente resulta la más grande de las células que se pueda encontrar en los peces (la mayo- ría de 0.5 a 5.0 milímetros de diámetro, pero algunas del tamaño de una bola de golf y aún mayores), sino también una célula provista de la especial peculiaridad de poder producir, después de su fertilización, un nuevo individuo de la misma especie. Durante la ovogénesis, las células epiteliales (granulosas) envolventes pro- veen al ovocito en desarrollo de cantidades relativamente grandes de material ali- menticio conservado en forma de yema granular (proteína) y grasa, generalmente en forma de pequeñas gotas de aceite.
Fecundidad es un término generalmente empleado para describir el número de huevos producidos. La fecundidad de una hembra varía de acuerdo a muchos factores entre los que se incluyen la edad, el tamaño (Figura 10.3), la especie y las condiciones ambientales (disponibilidad de alimento, temperatura del agua, temporada del año).
Parece ser que la fecundidad conlleva alguna interrelación amplia con el cuidado o protección de los huevos. Es así como el pez sol del océano (Mola

LOS OVULOS Y SU FORMACI6N 267
Figura 10.3 - La relación en una especie entre la fecundidad y la lon- gitud. 1. Acipenser stellatus (Nikolsky, 1963); 2. Salmo salar (Pope et al., 1961); 3. Cyprinus carpi0 (Nikolsky, 1963); 4. Pleuronectes platessa (Clyde) (Bagenal, 1966); 5. Clupea harengus (norte del Mar del Norte) (Baxter, 1959); 6. Melanogramus aeglefinus (v6ase Parrish, 1956); 7. Osmerus eperlanus (Lillelund, 1961); 8. Salvelinus fonrina- lis (Vladykov, 1956); 9. Sardinops caerulea (MacGregor, 19571, y 10. Sebastes marinus (Isla Faroe) (Raitt y Hall, 1967). (Fuente: Blaxter, 1969).
rnola) puede producir cantidades enormes de óvulos, pero son simplemente ex- pulsados por la hembra en mar abierto mientras nada acompañada por los ma- chos. Un pez de esta especie, de 24.5 kg de peso, llega a producir, según se dice, 28,000,000 o más óvulos por temporada. El bacalao (Gadus), que también es un desovador pelágico produce algo así como 9,000,000 de óvulos por hembra y por temporada. Estas especies no se preocupan por el cuidado de sus huevos, a no ser que desoven cuando los machos están en las cercanías o cuando éstos las acom- pañan, regando los espermatozoides. Las espinochas (Culaea y Gasterosteus), por el contrario, fabrican nidos muy elaborados y cuidan paternalmente la cría cuando ésta se está desarrollando. En este caso, el número usual de huevos varía entre treinta y cien. El Lebistes, que porta a su cría viva dentro del saco ovárico y le proporciona en consecuencia un gran cuidado (esto es, hay incubación interna), el número usual de individuos por camada es menor a dos docenas. Algunos pe-

26 8 REPRODUCCI6N
ces, inclusive, solamente llegan a tener un huevo en cada temporada, o uno en cada ovario.
Los huevos de la mayoría de los peces están envueltos por cáscaras. La célula huevo, por sí misma, tiene solamente una membrana delgada cuando está por desprenderse del ovario. Cuando los óvulos de las especies ovíparas pasan a través de los oviductos atraviesan glándulas que los envuelven con sus secreciones, formando la cáscara. Ésta, en los peces que cuidan de su cría, ya sea que los pa- dres permanezcan en guardia a su lado o que cubran los huevos con grava 0 arena, es a menudo más delicada y menos cargada de caliza o córnea que la de los huevos expuestos a las olas, corrientes de agua u otros peligros. En los huevos de muchas mantarrayas y en los de algunos tiburones, por ejemplo, las cáscaras son córneas y casi tan duras como las uñas de los dedos. Estos huevos resisten los daños, inclu- sive la sequía, al grado de que cuando son arrastrados por el agua hacia la playa, pueden permanecer ahí durante algún tiempo y aún hacer eclosión cuando son arrastrados de nuevo al mar. Un huevo de la raya pequeña (Raja erinacea) fue una vez remitido sin empaque a nuestro laboratorio de Ann Arbor, metido simple- mente en un sobre ordinario para carta, desde la costa de New Jersey. Estuvo por lo tanto fuera del agua y en tránsito durante algunos días, recorriendo algo así como mil ciento veintiséis kilómetros. Pudo, sin embargo, hacer eclosión con todo éxito y se volvió para nosotros una mascota durante algunos meses.
Aparte de su cubierta protectora, los huevos de los peces tienen otras carac- terísticas, muchas, muy interesantes. Algunos son errantes (pelágicos), flotadores y tienen una gravedad específica casi igual a la del agua dulce. Aunque la mayoría de los peces marinos comestibles, como el bacalao (Gadus), tienen huevos flota- dores, algunas especies marinas, como el arenque (Clupea harengus), no los tienen así, por lo que se mantienen fijos al sustr2,to. L a mayoría de los peces de los arro- yos tienen huevos pesados que se sumergen con una gravedad específica mayor que la del agua dulce. Posiblemente ésta es una adaptación que evita que estos huevos sean arrastrados corriente abajo al tiempo del desove, función biológica que puede tener lugar sobre lechos de grava en lugares donde siempre hay corrien- tes rápidas, preferidas por las truchas de arroyo (Saluelinus), las truchas, el salmón del Atlántico del género Salmo y las especies de los salmones del Pacífico (On- corhynchus).
Los huevos también varían por lo que respecta a sus características adhesi- vas. La mayoría de los huevos flotantes no se adhieren y normalmente no se pegan a ningún otro cuerpo con el fin de fijarse. Pero la mayoría de los huevos que se sumergen son, sin embargo, pegajosos, por lo menos temporalmente. Los huevos del salmón y de la trucha (Salmoninae) son pegajosos temporalmente durante el proceso de endurecimiento en el agua que ocurre en los primeros minutos después del desove, mientras están absorbiendo agua en grandes cantidades. Se puede casi distinguir los huevos como “chupadores” de agua, por la rapidez con que se adhieren a otros cuerpos, u otros cuerpos se les adhieren o se adhieren entre sí. Durante este proceso, los huevos de las truchas y salmones que desovan en las corrientes de agua recogen partículas pequeñas de arena que les permiten sumer- girse hasta el fondo y los fijan ahí para cubrirlos y facilitar su desarrollo. La adherencia de los huevos de los peces, como el “walleye” (Stizostedion), tiene el propósito dc evitar que se sumerjan en el fango del fondo y se sofoquen. Los huevos pegajosos, demersales, se adhieren a ramas y otros restos orgánicos del fondo, sin llegar a las partes más profundas donde puede haber materiales más

DIFERENCIAS SEXUALES 269
blandos. La adherencia puede ser debida temporalmente al proceso de endureci- miento por el agua, o puede presentarse en forma más durable por la presencia de un depósito pastoso, mucoso, sobre el huevo, que lo vuelve pegajoso durante todo su desarrollo.
Los huevos también varían en su forma (Figura 10.2b), pero los de la ma- yoría de las especies de peces son esféricos. Parece ser que la forma esférica es la natural, porque así es el contorno básico de la mayoría de los huevos de los ani- males. En muchas anchoas de la familia sardinoide Engraulidae la forma del huevo es elíptica, ocasionalmente esférica. Pero en el género Rhodeus, la bermejuela europea, y en muchos otros peces donde quedan incluidos los cíclidos, los huevos son de perfil oval (ovoides). En los gobios varían de ovoides a la forma de una lágrima.
Los huevos de los peces difieren también considerablemente de acuerdo a los apéndices que llevan consigo. El huevo del mixino (Myxine), por ejemplo, es un ovoide alargado y con algunos apéndices en forma de zarcillos en cada extremo. Aparentemente, esta especie de zarcillos fijan los huevos a la vegetación o a cualquier otro material. En el eperlano americano (Osmerus mordax) sucede que sus huevos tienen hacia abajo un pedúnculo pegajoso que los fija al fondo rocoso de los arroyos donde las especies más a menudo desovan. En el pez de lado plateado de los arroyos (Labidesthes sicculus) hay un filamento Único que sirve primordialmente para la flotación temporal y después para la fijación. Los huevos de algunas rayas (Rajidae) y tiburones gato (Scyliorhinidae) tienen pro- longaciones en forma de zarcillos que nacen en cada una de las cuatro esquinas de la cáscara, la cual tiene forma parecida a una caja. La mayoría de los peces voladores (Exocoetidae) tienen huevos provistos de numerosos filamentos muy parecidos a cabellos largos, que nacen de la superficie del huevo esférico. Los huevos de unos cuantos peces voladores, sin embargo, no presentan filamentos; algunos tienen los filamentos confinados a los dos polos del huevo y otros sola- mente a uno.
Así como los espermatozoides presentan diferencias en la vitalidad de acuerdo a las diversas situaciones a las que se enfrentan, los huevos tienen tam- bién varias características de supervivencia. Es interesante el hecho de que no todos los huevos maduros son puestos durante un periodo de desove, cualquiera que é1 sea. Algunos permanecen en el ovario y son normalmente reabsorbidos. Parece ser, sin embargo, que cuando cantidades desproporcionadamente grandes de huevos no son puestos, el ovario es incapaz de reabsorberlos. El individuo puede quedar repleto de huevos y la boca del oviducto taponada por una masa de tejido fibroso. El paso futuro de los huevos puede quedar obstaculizado.
DIFERENCIAS SEXUALES (FIGURA 10.4)
Cualquiera puede distinguir a un pez macho de uno hembra cuando la lecha o los óvulos son expulsados. Mediante una presión ligera en el vientre de la mayoría de los peces maduros se hace aparecer cerca del ano a la lecha blanquecina o a los óvulos. Tanto en los humanos como en los peces, sin embargo, se emplean también otras características para poder distinguir los sexos. Las características de la diferencia sexual, o el dimorfismo sexual que permiten la identificación de los sexos, son clasificados como primarias y secundarias. Los caracteres sexuales

270
DIMORFISMO SEXUAL
REPRODUCCIdN
Figura 10.4 - El dimorfismo sexual entre los peces es mostrado por: (a) el dorso giboso y las mandíbulas ganchudas del macho del salmón rosado (Oncorhynchus gorbuscha); (6) el frente de la cabeza como domo y la posición anterior de la aleta dorsal en el dorado (Coryphaena hippurus); (c) macho parásitodel pez ángel de los mares profundos (Photocorynus spiniceps); ( d ) tubbrculos nupciales en el hocico y el frente de la cabeza del “chub” de los arroyos (Semotilusatroma- culatus).
primarios tienen la responsabilidad directa del proceso reproductor. Los testicu- los y sus conductos en el macho, y los ovarios y sus conductos en la hembra, constituyen los Caracteres sexuales primarios. Los caracteres sexuales primarios a menudo requieren de la disección para poder ser distinguidos, por lo que mu- chas veces los caracteres sexuales secundarios se vuelven de más utilidad, aunque en ocasiones con poco éxito. astos, por sí mismos, corresponden a dos clases: los que realmente no tienen ninguna relacibn con el acto reproductor y los que definitivamente tienen función auxiliar en el desove. A continuación se indican algunos aspectos diferenciales importantes de naturaleza dimbrfica entre los sexos.

DIFERENCIAS SEXUALES 271
Una papila genital distingue a los machos de peces como las lampreas (Petro- myzonidae), el pércido Etheostoma nigrum y la lobina blanca (Morone chrysops). En un grupo de peces, los Oligocottinae, la papila es un pene verdadero. La forma del cuerpo constituye un importante carácter sexual secundario. Por lo general, el vientre de la hembra es más prominente que el del macho, principalmente cuando ha madurado o está próxima a desovar, debido a la cantidad relativamente grande de productos sexuales que contiene en su interior. En los machos de mu- chos peces aparecen órganos aperlados o tubérculos nupciales que permiten dis- tinguir el sexo; como ejemplos podemos mencionar al eperlano (Osmerus), la mayoría de los pececillos de la familia Cyprinidae (Figura 10.4) y las carpas hoci- conas (Catostomidae). Estos tubérculos son pequeñas excrecencias córneas que se vuelven conspicuas precisamente antes de la temporada de la puesta o desove y desaparecen muy poco tiempo después, bajo la influencia de secreciones hormo- nales. Con frecuencia las aletas pueden caracterizar a los machos; en promedio, son de mayor tamaño que las de las hembras. Y a propósito de esto, en un pez de América del Norte, el pércido Etheostoma flabellare, con la caudal en forma de abanico, el macho tiene las puntas de las espinas de la aleta dorsal, durante la época de reproducción, transformadas en perillas carnosas, de tal manera que se ven como cachiporras, mientras que la hembra tiene por lo general radios ordina- rios. En algunas otras especies, los machos tienen radios carnosos en su totalidad, como en el pececillo de nariz roma (Pirnephales notatus), mientras que los radios de las aletas de la hembra son precisamente al revés, es decir, de aspecto normal. En algunos peces, la aleta caudal puede mostrar dimorfismo sexual; el lóbulo infe- rior está muy alargado en los machos del pececillo gallito o cola de espada (Xipho- phorus helleri) muy conocido por el aficionado a los acuarios, y es algo alargado en la carpa chupadora (Catostornus commersoni).
Un carácter sexual accesorio distingue a los machos de muchas especies en los cuales la aleta anal aparece alargada formando un órgano intromitente, de utilidad para la cópula. Este órgano, llamado gonopodio (Figura 10.5), puede ser visto en pececillos como la gambusia o pez mosquito (Gambusia affinis), el pece- cillo de acuario Lebistes (“guppy”) y en otros pecílidos viviparos. En algunas percas (Embiotocidae) vivíparas, la aleta anal lleva algunas estructuras glandulares que facilitan la intromisión. En el macho de la carpa hocicona (Catostomus com- mersoni), la aleta anal, alargada, sirve para conducir la lecha acuosa hacia el fondo del nido perturbado por las corrientes en los arroyos, mejorando en con- secuencia las oportunidades de poner en contacto los espermatozoides y los óvu- los. En el ciprinodonto abrazador (Xenodexia), la aleta pectoral modificada, o heteroquiridio, es usada evidentemente durante la cópula para sostener el gono- podio en posición para su inserción en el oviducto de la hembra. Las aletas pélvi- cas se encuentran modificadas de diversas maneras en los tiburones y especies afines (Elasmobranchii) para formar estructuras intromitentes, los mixopterigios (Figura 10.5), que aseguran la fertilización interna, tan extendida en este grupo de peces.
Sin duda alguna que la coloración de los peces también constituye una característica para la distinción y el reconocimiento sexuales; se le llama dicro- matismo sexual. Por lo general, los machos presentan colores más brillantes 0 intensos que las hembras. Esto también ocurre en las aves, pero, tal como sucede en éstas, también hay excepciones a la regla. Un ejemplo notable de dicromatismo sexual que aparece en la fauna ictiológica del centro de América del Norte corres-

272 REPRODUCCIdN
ÓRGANOS INTROMITENTES
(a) Clasper de tiburón (mixopterigio)
Base de la
Radios blandos anales
modificados
( b ) Gonopodio de Gambusia
Figura 10.5 - drganos intromitentes del macho en dos peces: (a) aleta pélvica izquierda de un tiburón cornudo (Heterodontus francisci) mostrando su mo- dificación en un mixopterigio; vista ventral; (b) aleta anal modificada para formar un gonopodio en el pez mosquito (Gambusia affinis), vista lateral. Parte (a) basada en Daniel, 1934, y (6). en Turner, 1941).
ponde a la mojarrita de manchas anaranjadas (Leponis hurnilis). Las manchas naranja del cuerpo de esta mojanita son en el macho más numerosas y brillantes que en la hembra. En el pez de aleta ondulada (Arniu calva) hay una mancha oscura, parecida a un ojo, en la región caudal de los jóvenes de los dos sexos, pero desaparece gradualmente, aunque no totalmente, y queda reducida en tamaño, en las hembras adultas, mientras que en el macho se vuelve brillante y se trans- forma en un ocelo, con un anillo de color atractivo a su alrededor. Los peces señorita (Labridae) y loro (Scaridae) del mundo marino, pueden ser mencionados como ejemplos primarios de dicromatismo sexual y, en algunos casos, también de dimorfismo.
Muchas características diferentes de la cabeza también sirven para distin- guir los sexos entre los peces. El macho de las quimeras (Chimeridae) desarrolla una protuberancia fuerte, espinosa y retráctil, el clásper frontal, en la parte supe-

MADURACI6N SEXUAL 213
rior de la cabeza; una estructura similar caracteriza la porción frontal de la cabeza de los incubadores frontales (Kurtidae; Figura 10.6). En el caso de los salmones y las truchas (Salmoninae), los machos criadores desarrollan típicamente una protuberancia ganchuda cerca de los extremos de las mandíbulas superior e inferior.
Unas cuantas estructuras reproductoras accesorias sirven entre las hembras como características sexuales. Un ejemplo notable de esto puede ser observado en el tubo ovopositor de las hembras de la bermejuela de Europa, Rhodeus ama- rus y en un pez de Asia, Careproctus. En unas cuantas especies de peces se han llegado a descubrir diferencias entre machos y hembras por el número de radios en las aletas y por el de vértebras en la columna dorsal. El indicio de la madurez sexual en las larvas de algunas especies, o sea, la neotenia, es un caso de dimorfis- mo sexual entre los machos de los peces de ojos pedunculados de las grandes pro- fundidades marinas, como en Idiacanthus y entre ciertas lampreas europeas (Pe- tromyzonidae).
Probablemente la forma más extrema de dimorfismo sexual ocurre en el pez pescador de los fondos marinos profundos (Figura 10.4c), cuyo macho es muy pequeño y no más que una fabriquita parásita de espermatozoides, fijado al cuerpo comparativamente mucho mayor de la hembra.
MADURACIdN SEXUAL
Muchos factores influyen en los peces a fin de que adquieran por primera vez la capacidad para reproducirse, o sea, la maduración sexual. Así, deben tomarse en cuenta la edad, tamaño del animal y la fisiología. Por lo general, las especies cuyo tamaño máximo es pequeño y su promedio de vida corto, maduran a temprana edad, si comparamos el fenómeno con el de los peces cuyo tamaño máximo es mayor. Algunos peces alcanzan la madurez sexual al momento de nacer; los ma- chos de la perca de los arrecifes (Micrometrus aurora) y de la perca enana ( M . minirnus) se reproducen poco tiempo después de haber nacido. Aunque las hem- bras de estas especies son impregnadas por el esperma muy pronto después de haber nacido, no producen cría hasta después de un año. Otros peces, como el de acuario (Lebktes) y el pez mosquito (Gambusia affinis), alcanzan su madurez sexual cuando tienen edades menores de un año y longitudes menores a 2.5 cm o menos, aunque hay que tomar en consideración que no nacen en estado adulto. Muchos peces maduran a la edad de un año. Hay algunos ejemplos, tomados de la fauna de las aguas dulces norteamericanas, que ilustran este caso, como ocurre con el pequeño pércido Etheostoma microperca de apenas 2.5 cm de longitud, y otras especies menores a 1 5 cm, como el pez plateado de los arroyos (Labidesthes sicculus); un ejemplo entre los peces marinos, de los muchos que hay, es el pejerrey de California (Leuresthes tenuis). Muchas especies alcanzan por primera vez la madurez sexual a edades de 2 a 5 años, con longitudes de 7.5 a 30 cm o más. Entre ellas se cuentan las truchas (Salmo), las lobinas negras (Micropterus) y la mojarrita (Lepomis). Las anguilas (Anguilla) se vuelven sexualmente maduras desde los 10 a los 14 años de edad y teniendo 60 o más centímetros de longitud, y los esturiones (Acipenser) a los 15 o más años de edad, con longitudes de 1 metro o más. Tanto la edad como el tamaño, combinados, constituyen factores determi- nantes de mucha significación, en relación al estado adulto.
Una vez que el pez ha madurado sexualmente debe todavía esperar a que

274 REPRODUCCI~N
10s productos de las gónadas maduren para poder realizar el acto reproductor. Hay muchos factores que intervienen para que tengan lugar estos hechos. Y pueden ser reunidos en dos grandes grupos: los que aparecen en el propio pez (in- trínsecos; Capítulo 11) y los que se encuentran en el medio ambiente (extrinse- COS). El pez como individuo independientemente del grupo al que pertenezca, no tiene realmente control sobre esos factores. Entre los factores intrínsecos que determinan la maduración sexual y los subsecuentes desoves y maduraciones se cuentan la especie y su juego cromosómico como factor hereditario, los ali- mentos seleccionados y, finalmente, el todo fisiológico del individuo que, por supuesto, podría ser muy difícil de separar de su herencia. Dentro de una especie, el sexo del individuo es muy importante para determinar sus reacciones ante los factores que conducen a la ovoposición o desove. En muchas clases de peces, los machos maduran y están listos para la reproducción más pronto que las hem- bras, ya sea en su ciclo completo de vida o durante el curso de sólo una tempo- rada de desove. Por ejemplo, las glándulas sexuales de algunos machos de ciertos salmones del Pacífico (Oncorhynchus) pueden madurar, y los individuos pueden fertilizar, aunque la especie normalmente expulse la lecha por primera vez 2 o 3 años más tarde. Algunos de los machos de maduración temprana, los llamados “grilse”, que a menudo se aparean con las hembras, adelantan su maduración un año en relación a las hembras de la misma edad. En muchos peces de agua dulce, como las truchas (Salmo), carpas chupadoras (Catostomus) y “chubs” de los arro- yos (SemotiZus), los machos llegan primero a los sitios de desove que, para las diversas clases registradas corresponden a los rápidos o corrientes, que es donde acostumbran desovar. En este caso el macho comienza con la construcción de un nido, o simplemente espera la llegada de las hembras. Se presume que esta tem- prana maduración en los machos se debe al nivel elevado del metabolismo, posi- blemente a través de la influencia endocrina que se produce en respuesta a factores genéticos y ambientales.
El ritmo fisiológico interno de la maduración gonadal, que envuelve predo- minantemente las interacciones pituitario-gonadales, es ajustado de tal modo que asegure la aparición de las actividades sexuales cuando las condiciones ambientales sean más favorables para la supervivencia de la cría. Los cambios que ocurren en la duración del día solar (fotoperiodo) y la temperatura afectan el ritmo de la maduración gonadal y el desove del pez en la zona templada y las latitudes altas. Cerca del Ecuador, donde la duración del día no afecta la reproducción estricta- mente de temporada, la actividad reproductora de muchos peces tropicales coincide a menudo con las extensas inundaciones que ocurren durante la temporada de llu- vias, según los numerosos ejemplos conocidos. Otros factores extrínsecos que pueden afectar a la reproducción son la presencia del sexo opuesto, la corriente, la marea, el aspecto de la luna y las facilidades para desovar. En la bermejuela (Rhodeus amarus) se ofrece un caso de estimulación para la maduración, debido a la presencia de un mejillón de agua dulce que sirve para recibir a los huevos durante el desove. Las corrientes producidas por el sifón exhalante del mejillón estimulan finalmente al pez para que realice el desove.
CICLOS REPRODUCTORES
La conducta reproductora en la mayoría de los animales es cíclica, en periodos más o menos regulares, y esto ocurre en casi todos los peces. El acto reproductor

REPRODUCCI~N 276
ocurre en algunos peces solamente una vez durante su corta vida; un ejemplo de esto lo tenemos en el pez de lado plateado de los arroyos (Lubidesthes sicculus). Incluso en otros peces acontece una vez en un espacio de vida moderadamente largo: en los salmones del Pacífico (Oncorhynchus), una sola vez en un periodo de 2 a 5 o más años; en la lamprea marina (Petromyzon murinus), una vez en 5 o 6 años, y en las anguilas de agua dulce (Anguillu), una vez en 10 a 14 años. La mayoría de los peces, sin embargo, tiene un ciclo anual de reproducción y, una vez que han comenzado con é1, lo continúan teniendo hasta que mueren. Muchas otras especies desovan más de una vez en un año y más o menos continuamente. El “guppy” (Lebktes) puede producir cría a intervalos de aproximadamente 4 semanas; también, puede haher crías en buen número por año en ciertas tilapias (TiZupiu). El pez de aleta filamentosa (Polyductilus sexfilis) de los alrededores de Hawai desova m a vez por mes entre mayo y octubre. El gruñón (Leuresthes) de la costa de California puede desovar cada dos semanas en la estación durante la temporada de desove hasta que ésta termina. En todos los actos reproductores existe el objetivo de lograr la unión del espermatozoide y el óvulo. Se presentan después otras adaptaciones que permiten la supervivencia del óvulo fertilizado.
Los peces han desarrollado diversos medios para lograr la unión del espermatozoide con el óvulo. Entre estos medios es muy notable el de la impresionante relación que hay, con respecto al tiempo, entre las diversas funciones reproductoras. En los desovadores prolíficos de las aguas abiertas, por ejemplo, no todos los millo- nes de huevecillos que son puestos maduran al mismo tiempo, sino progresiva- mente, en cada ovario, de tal manera que al llegar a su término el correspondiente crecimiento, la hembra no es distendida en forma exagerada por los productos sexuales agrandados. Si todos maduraran en forma simultánea, ocuparían un espa- cio mucho más grande del que podría permitir el cuerpo de la madre; por lo tanto, las emisiones de óvulos se hacen cronológicamente, así como su maduración, para lograr el acomodo en el cuerpo del pez hembra. Muchos peces tienen un periodo muy corto de reproducción, ocurriendo solamente una vez al año según hemos visto, y todavía así los machos y las hembras están en buenas condiciones de madurar al mismo tiempo y, en consecuencia, son capaces de exhibir sus reaccio- nes complejas de apareamiento. Las características sexuales secundarias y las estructuras accesorias que son empleadas durante el apareamiento -el cortejo, el abrazo o la intromisión- se desarrollan simultáneamente con la maduración de los productos sexuales primarios.
Posiblemente el caso más famoso de interrelación reproductora realizado con precisión cronométrica es el del gusano “palolo”*, de los arrecifes de coral del Pacífico sur, que surge en multitudes a la superficie de los mares abiertos una vez por año, para reproducirse, cuando existe cierta fase lunar. Unos cuantos peces muestran también una maravillosa interrelación para reproducirse, cronoló- gicamente dispuesta. El caso mejor conocido es el del gruñón (Leuresthes tenuis) de la costa de California. Este pez desova precisamente durante el retroceso del agua en la marea alta en ciertas ocasiones del año. Puede decirse que desova fuera
*Anélido poliqueto (N. del T.).

dei agua, tan alejado de la playa conlt> lo pueda permitir la mas grancle Q C ~ [as ajas de la marea aka que lo empuje. Este pez deposita en l a arena hizmeda bolsas huevecillos y esperma, en cierto tiempo y posicibn precisos, de t a l modo que 10s huevos no sean barridos por las aguas sino hasta dos semana.s O un mes m& tarde, cuando se presente la siguiente marea alta. En ese momento, las olas regre- san y bañan la arena de nuevo, y cuando tocan los lugares donde se encuentran los huevos, depositados en nidos, la revuelven; es en ese momento cuando la cría sale del huevo casi instantáneamente y se aleja con las olas antes de que retroceda la marea y le sea después imposible aprovechar la oportunidad de hacerlo. Sin duda alguna que este fenómeno es un ejemplo notable de reproducción cronoló- gicamente establecida así como de desarrollo y eclosión. Con relación a las interre- laciones cronometradas, también deberíamos pensar que en algún lugar de un continente como América del Norte, hay peces que están desovando casi a cual- quier hora debido a la influencia de la latitud, la altitud y a la individualidad de las especies y razas o variedades que las componen.
La mayoría de las especies tienen estaciones definidas para el desove como parte de sus interrelaciones reproductoras cronometradas, y son generalmente agrupadas como se indica a continuación. Los peces de aguas cálidas desovan durante el verano y los de aguas frías durante el otoño e invierno. Las especies que toleran las temperaturas intermedias desovan generalmente durante la pri- mavera. Algunas especies tropicales desovan durante todo el año. Las temporadas fijas de desove están correlacionadas en forma superficial con el periodo de desa- rrollo que requiere el pez. Los peces de aguas cálidas (como las lobinas negras y los peces sol, Centrarchidae) emplean solamente algunos días para hacer eclosión y emerger en un medio generalmente favorable para el crecimiento y la supervi- vencia. Muchos meses pueden ser requeridos para una trucha (Salvelinus) o un pescado blanco (Coregonus). Algunas truchas (Salmo), a una temperatura media de 50°F (lO°C), pueden promediar alrededor de 50 días para su periodo de desa- rrollo. El pescado blanco (Coregonus clupeaformis), estando a una temperatura inferior a los 30°F (-l"C), puede emplear alrededor de 130 a 150 días para su desarrollo hasta la eclosión. Parece que las dos clases de peces han sido crono- metradas para producir cría en condiciones favorables para ella. La cronometri- zación de la reproducción permite también a más de una especie utilizar la misma área de reproducción en un año calendario. Es así como en los arroyos norte- americanos se ha observado que conviven normalmente los desovadores de pri- mavera, o sea, las carpas chupadoras (Catostomus), que pueden usar la misma área ("redd") de nidación, como lo hace también la trucha de arroyo desovadora de otoño (Salvelinus fontinalis).
Las interrelaciones cronometradas, no importa cuán buenas sean, podrían no ser de mucha utilidad para el pez si éste no pudiera reconocer una pareja del sexo opuesto cuando estuviera presente. Cuando un científico está identificando el sexo de un pez se fija en caracteres como la estructura, la forma y el color, como se ha dicho ya. Con el pez vivo puede también aprovechar su conducta. Los mismos peces pueden reconocer la forma, el color y la conducta de sus parejas. Timbergen realizó un notable estudio experimental para reconocer el valor de estos atributos que le permiten al pez hacer el reconocimiento sexual; demostró todos estos detalles mediante la observación de las espinochas (Gasterosteus) en acuarios.
Además de la evolución de las interrelaciones cronometradas, y de las capa-

cidades para X C Q ~ C C ~ ~ ni sex(. opuestr>, se har;. des;:::roJiadc~ enzre loz peces mu- chos dispositivos de utilidad para asegurar la fsrtilizacion, y a sea externa o interns (Figura 40.5). El medio m& com6nmente empleado para consumar l a fertilización externa consiste en la simple proximidad de dos individuos de S ~ X Q S opuestos mientras se arrojan al agua los espermatozoides y los óvulos, respectivamente. En ocasiones es empleado el apareamiento y alguna forma de sujeción (ample- xus) como adaptación especial para la aproximación de los sexos. Ciertos peces, al aparearse, se ponen en contacto lado a lado y emiten simultáneamente óvulos y espermatozoides; en los peces de forma muy alargada como las lampreas (Pe- tromyzonidae), el macho enrosca su cuerpo alrededor del de la hembra, en un semicírculo, e inclusive en forma de espiral como si fuese sacacorchos.
El macho enano parásito del pez batial (Photocorynus) se adhiere a la piel de la hembra mediante su boca (Figura 10.4). En este caso, el macho tiene sola- mente una función reproductora y ha perdido virtualmente el tracto digestivo como consecuencia de su adaptación parasitaria. La concentración de huevos en masas constituye un hecho que asegura la fertilización externa. Podemos ver hue- vos unidos con material gelatinoso en las percas (Perca; Figura 10.6). Los huevos son agrupados debajo de las piedras por peces como el pércido de Johnny (Etheos- toma nigmm; Figura 10.6) y los pececillos de nariz roma y cabeza grasa (Pirne- phales). Los peces escorpión marinos (Scorpaerzu) forman huevos en un globo de dos lóbulos. Esto se relaciona con la vitalidad y la gravedad específica de los esper-
DESTINO DE LOS HUEVOS
PUESTO A L A Z A R
" A la deriva
/ ._ \. '\ I
' ~. t . 1- Pez volador
Figura 10.6 - Diversidad de la disposición de los huevos y la cría en los peces.
PUESTOS EN MASAS
PUESTOS EN OTRO ANIMAL
. .
Continúa

278
Figura 10.6 - Continuación.
DESTINO DE LOS HUEVOS
PUESTOS EN NIDO
Sin cuidado paternal subsecuente
I c;- "" Corriente +
I ""I Con cuidado paternal subsecuente
REPRODUCCI6N

279
Figura 10.6 - Continuación
DESTINO DE LOS HUEVOS
TRANSPORTADOS POR LOS PADRES
Externamente -
Bagre criador en la boca 9
Pez criador en el frente cefálico d
Internamente

La produccihn efectiva de Gvulos y espermatozoides, únicamente, no seria S u f i - ciente para lograr la supervivencia de la especie. Mientras que muchos peces, especialmente los marinos, se hasan en “’la seguridad por fa cantidad”, se han desarrollado varios procedimientm para lograr el cuidado de los huevos y de la

&ir, PO: uno o 10s dos sexos (Tabla 10.1). Uno de esos medios ha sido el de llevar los huevos ai sitio preciso, el lugar que corresponde a la adaptación de su estruc- tura de desarrollo. Asi es como los peces anádrornos corno l a lamprea marina (Petromyxon marinus), los esturiones migratorios (Acipenser) y los salmones (Sal- mo, Oncorhynchus) ascienden hacia los arroyos de agua dulce para desovar. Mu-
Tabla 10.1 - Diversidad de lugares para ¡os nidos y el desove, como lo muestran ejemplos selec- cionados de las aguas dulces de America del Norte.
I . Especies que construyen nidos.
A. Las que proporcionan cuidados paternales.
1. El nido es una depresión circular. a. En un lecho de lodo, aluvión o arena, a menudo en y entre raíces de plantas acuá-
ticas fanerógamas. Peces sol (Centrarchidae), incluyendo las mojarritas propiamente dichas (Lepomis), huro (Micropterus salmoides), "crappies" (Pomoxis), "rock basses" (Ambloplites) y el "warmouth" (Chaerlobryttus); el pez de aleta ondulada (Amia); la mayoría de los bagres (Ictalurus).
b. En un fondo de grava. La perca de boca pequeña (Micropterus dolomieui); rara- mente, la perca de boca grande; los "rock basses".
2. Nido excavado bajo una piedra o cualquier otro objeto sumergido, con los huevos adheridos por debajo del techo del nido. El pércido de Johnny (Etheostoma nigrurn), el percido de aleta caudal en abanico (Etheostoma flabellare), los "sculpins" de las moradas en los arroyos (Cottus), los pececillos de nariz roma y cabeza grasa (Pime- phales) .
3. Nido fabricado con materiales vegetales y de forma esférica o de montículo. Espino-
4. Nido en forma de túnel. En un banco, bagre de canal (Ictalurus punctatus); en el fondo,
chas (Culaea; Gasterosteus),
bagre amarillo (Ictalurus natalis).
B. Peces que abandonan el nido despues de la reproducción; nido en el fondo de grava, con una estructura muy bien definida. Lampreas (Petromyzonidae); truchas de arroyo, inclu- yendo a Salvelinus fontinalis, trucha de cuello cortado (Salmo clarki), trucha arco iris (S. gairdneri) y trucha café (S. trutta); salmones (Oncorhynchus y Salmo salar); gordo de arroyo (Semotilus atromaculatus); pez "fallfish" (S. corporalis); gordo de río (Hybopsis micropogon); pez de cabeza córnea (H. biguttata); pez lanza arco iris (Etheostoma cae- rulum ).
I I . Especies que no construyen nidos.
A. Huevos dispersos, generalmente sobre plantas acuáticas, en sus raíces o restos. Lucio (Esox lucius); carpa (Cyprinus carpio); carpa dorada (Carassius auratus); brillo dora- do (Notemigonus crysoleucas).
B. Huevos desparramados sobre bajos arenosos, grava o cantos rodados. Peces blancos-y ciscos (Coregonus); trucha de lago (Salvelinus namaycush); chupadores (Catostomus); "walleyes" (Stizostedion).
C. Huevos depositados en masas aisladas de forma definida. Una "cuerda" de huevos, perca amarilla (Perca flavescens).
D. Vertiendo productos sexuales cerca de la superficie sin tomar en cuenta la vegetación o tipo de fondo. Pez de lado plateado de los arroyuelos (Labidesthes sícculus); "alewife" (Alosa pseudoharengus) .

282 REPRODUCCION
chos salmones adultos se alojan precisamente en los arroyos en donde nacieron y vivieron su juventud, después de haber viajado miles de millas durante periodos de tiempos de 5 años o más. Las anguilas de agua dulce (Anguilla) tienen un hábito catádromo, descendiendo hacia el océano para depositar los huevos en el hábitat de agua salada para el cual están adaptadas y lograr así su desarrollo.
Encontrándose ya en los sitios de desove se prosigue con la selección del lugar apropiado para realizar esa función. En los peces que construyen nidos este proceso se hace con mucho cuidado y es de gran importancia para lograr una reproducción exitosa. Una vez que el sitio ha sido escogido, algunos peces pelea- rán hasta la muerte, si es preciso, para defenderlo. Un ejemplo notable lo tenemos en el pez luchador siamés (Betta splendens), bien conocido de los aficionados a los acuarios como una especie que defiende su territorio hasta morir. Muchos peces, sin embargo, no le proporcionan especiales cuidados a los huevos. En este caso, los huevos, por sí mismos, están adaptados para sobrevivir. Ellos deben flo- tar, ya sea con o sin un pequeño apéndice filamentoso, que les permita fijarse durante su desarrollo. Los huevos pelágicos, transparentes, se hacen invisibles para los predadores potenciales. Los huevos pueden estar dispuestos en grupos como sucede con las hileras huecas de huevos de la perca (Perca), de tal modo que no están desparramados, no caen al lodo del fondo y tienen cierta protección por estar agrupados. El huevo-globo transparente del pez escorpión (Scorpaena) ya mencionado, podría tener algún valor para obtener protección, así como para asegurar su dispersión. Los diversos dispositivos de fijación, así como su adheren- cia, también ayudan a la supervivencia de los huevos que son abandonados por sus padres.
Los peces que construyen nidos pueden ser agrupados en dos grandes cla- ses: los que construyen un nido y luego lo abandonan, y los que lo construyen y permanecen a su lado para proteger el lugar, los huevos y/o la cría. Podemos con- siderar, como unos cuantos ejemplos de peces que excavan para construir sus nidos y luego los protegen llenándolos con huevos revueltos con piedras, al pez gordo de los arroyos (Semotilus), las truchas (Salmo) y los salmones (Oncorhyn- chus). Algunos de los peces que excavan un nido y luego lo defienden contra los invasores son el pececillo de nariz roma y cabeza grasa (Pimephales), los peces sol y las lobinas negras (Centrarchidae). Esta familia, formada por peces de agua dulce de América del Norte, se caracteriza por realizar la construcción de nidos y por el hábito defensivo en el cual el macho es el agresor; excepto un género, la perca de Sacramento (Archoplites), que no construye nido. Los bagres (Icta- lurus) excavan un nido y permanecen a su lado en guardia. El pez luchador siamés (Betta) produce un nido de burbujas en la superficie y lo defiende, como las espinochas (Culaea y Gasterosteus) defienden el suyo, que está hecho de palitos, hojas y detritus en un sustrato. Los huevos de la bermejuela (Rhodeus) son deposita- dos en la cavidad del manto de un mejillón y los del “lumpsucker” (Careproctus) debajo del cuerpo del cangrejo de Kamchatka.
Para llevar al máximo la protección de los huevos algunos peces han desarro- llado varios tipos de incubación o gestación interna. Algunas veces la protección proporcionada por el cuerpo progenitor es concedida a la cría por el macho, como sucede en los caballitos de mar (Hippocampus) y en los peces agujón (Syngna- thus), en los que los huevos son colocados, por la hembra, dentro de una bolsa que posee el macho. En un pez gato o bagre neotropical del Brasil (Loricaria typus) el padre desarrolla un labio inferior, agrandándolo para formar una bolsa,

CUIDADO DE LOS HUEVOS Y DE LA CRfA 283
donde tiene lugar una incubación labial de los huevos. Los bagres marinos (Arii- dae), incluyendo al bagre bandera (Bagre marinus) y al del Atlántico (Galeichthys felis), emplean la boca como una cámara incubadora oral. Cada progenitor macho lleva consigo de diez a treinta huevos en desarrollo en su cavidad bucal, donde la cría puede vivir aún después de haber comenzado a alimentarse en forma inde- pendiente. La incubación oral también se presenta en los peces cardenal (Apogo- nidae), entre otros, pudiendo por lo tanto acarrear cien o más huevos.
La verdadera incubación interna es la que ocurre en los peces vivíparos. La viviparidad, con todas sus variaciones de intercambio embrionario y nutricional, se ha desarrollado independientemente en muchos grupos de peces. La viviparidad se produce entre los tiburones, en más de una docena de familias. La mayoría no son placentarios u ovovivíparos, pero hay unas cuantas especies que pre- sentan estructuras bien definidas para el intercambio nutritivo y respiratorio entre los géneros Triakis, Carcharhinus y Mustelus de los tiburones “requiem” (Carchar- hinidae), y Sphyrna de los tiburones martillo. Entre las rayas, mantarrayas y gru- pos afines (Rajiformes), la ovoviviparidad es la regla, siendo las mantarrayas ovíparas (Rajidae), la excepción. Es interesante ver cómo los tiburones ovíparos y especies afines son cavadores de los fondos de aguas relativamente poco profun- das y nunca son grandes, mientras que los vivíparos van de pequeños a gigantescos y ampliamente variables en sus hábitos y distribución.
La viviparidad, entre los peces óseos, está más desarrollada en los “top- minnows” (Cyprinodontiformes), especialmente en las familias de los “guppy” (Lebistes) y de los peces mosquito (Gambusia affinis), Poeciliidae, los vivíparos mexicanos (Goodeidae), los peces cuatro ojos (Anablepidae) y los Jenynsiidae. También aparece en las agujitas (Hemiramphidae) y en muchas familias de los peces en forma de perca (Perciformes): “surfperches” (Embiotocidae), clíni- dos (Clinidae), “eelpouts” (Zoarcidae), brotulos (Brotulidae), peces escorpión y peces roca (Scorpaenidae). En esta última familia la viviparidad es universal entre más de cincuenta especies del género Sebastodes de las aguas costeras de Califor- nia. El celacanto viviente, Latimeria, es también ovovivíparo.
En la literatura sobre ictiología se ha mencionado a menudo que el desarro- llo de los peces vivíparos tiene lugar en un útero. Es necesario precisar esta gene- ralización. Si se acepta que un útero es fundamentalmente un oviducto alargado, el órgano está presente en los tiburones vivíparos (Squaliformes). Entre los peces óseos (Osteichthyes) vivíparos, sin embargo, el desarrollo tiene lugar en los ovarios. En algunas especies, el desarrollo del embrión es intrafolicular. En este caso, el folícu- lo está vascularizado, dilatado y lleno de fluido. El embrión, que carece de corion envolvente, se desarrolla dentro del folículo correspondiente al óvulo del que nació, siendo la ovulación y el nacimiento simultáneos. Este es el caso de los pe- cecillos vivíparos (Poecillidae) y de otras familias entre las que quedan comprendi- dos las agujitas (Hemiramphidae) y los peces cuatro ojos (Anablepidae). En algunos peces, sin embargo, el óvulo debe ser fertilizado cuando aún está en su folículo, pero completa su desarrollo dentro de la cavidad del ovario después de haberlo aban- donado; de casos como éste podemos citar a la perca de las rompientes marinas (Embiotocidae) y las familias Jenynsiidae y Goodeidae. En algunos ciprinodon- tiformes, más de una cría puede estar presente simultáneamente en un ovario; condición denominada superfetación.
Además del cuidado de los huevos, nosotros debernos pensar por un mo- mento sobre el cuidado de la cría que deriva de aquéllos. La cría es ardientemente

DESARROLLO
Hace cien aiíos los aspectos relacionados con la descripción del desarrollo emo- cionaron a los estudiosos de las ciencias naturales. Ahora, aunque todavía no se conoce la embriología y el desarrollo postnatal de la mayoría de las especies de vertebrados y, en consecuencia: no se ha hecho la descripción -y esto es cierto para la mayor parte de los peces-, lo emocionante del descubrimiento se centra en el conocimiento de los mecanismos que permiten que los procesos de desa- rrollo se realicen, quedando incluidas las interacciones de las fuerzas geneticas y del medio ambiente que influyen en la determinación de varias características del individuo.
El desarrollo es, en su sentido más amplio, un proceso infinito? que se con- tinúa de generación en generación. Para el individuo se inicia en la fertilización del óvulo; la maduración y la reproducción aseguran la continuidad, y la muerte termina con el desarrollo. La espermatogénesis y la ovogénesis son, en consecuen- cia, estadios subterminales, que sirven de preparación de los procesos para ser utilizados en la generación próxima. Por conveniencia, sin embargo, pueden ser con- siderados los periodos sucesivos del desarrollo como: a) embrionarios iniciales; b ) embrionarios o larvales transicionales, y e ) postembrionarios. Debido a la acción de los predadores, las limitaciones de la reserva alimenticia y la gran sensibilidad de los embriones y las larvas hacia los cambios de las propiedades físicas y químicas del medio ambiente, estas etapas del desarrollo sufren notables detrimentos por mortalidad, mucho mayores que los sufridos en los estadios más tardíos, en los peces que producen huevos, o sea, los ovíparos. Los periodos embrionarios inicia- les comienzan cuando el óvulo es fertilizado por un espermatozoide y termina cuando el embrión ha adquirido los sistemas orgánicos generalizados, como apa- recen comúnmente en todos los embriones de los peces.
La fase transicional del desarrollo embrionario incluye la transformación de los órganos que forman sistemas generalizados y la forma del cuerpo del embrión en su periodo inicial, la que recuerda al adulto, Por lo tanto, la forma definitiva del cuerpo es obtenida en o cerca del final de este periodo. Durante esta fase del desarrollo están representadas, entre los peces, dos clases de larvas, las de vida li-
*"Criadores en la boca", sería la especificación en español (N. del T ~ ) .

.- , ,I - ;,; YI:> f;brc:~ i s 6 'laxa: dt. vicia litre se ~:a:^ac::c:izí~~~ POT trner exlij- -,.; , , , ,-/ " I , i ,- .. , .
; T J ? X lcii-;?c, L.e las F _ ' S L T ~ I C \ L S Y R ~ prokctoras eirlbr:ionarias. Las 1:xvas de vida no libre x b m p e i ~ r ; SL t:amici6n dentro de las membranas del huevc 3 dentro del cuerpo ae i i ?,I;.L?T~. r> c:' padre. En las especies que presentan desarrollo indirecto, inclu- yun;lc i a ntetkmmrfosis, es 6st.a l a que da tkrmino a la etapa. TJna fase embrionaria tardis cabre, en algunos peces, esos cambios de desarrollo que ocurren después de que ha aparecido la forma definitiva del cuerpo, El periodo transcurre dentro del tracto reproductor de la madre en los peces viviparos. En las especies que pro- ducen larvas de vida libre la forma adulta es obtenida ya sea apenas después de l a eclosión o después de pasar por varios estados de desarrollo en los cuales el pece- cillo se parece muy poco al adulto (por ejemplo, el pez cirujano, Acanthurus triostegus).
El desarrollo postembrionario comprende las fases juvenil, la del adulto sexualmente maduro y la de senectud. En un pez joven, 40s sistemas de órganos, particularmente los del sistema reproductor, completan su desarrollo. La forma del cuerpo es esencialmente la del adulto, aunque debe haber ciertas diferencias en el crecimiento de algunas partes. En el estado adulto, el individuo tiene la forma madura típica de su especie y es capaz de reproducirse. De la fase de adulto pasa a la de senectud y degeneración. En la senectud, el potencial reproductor declina y/o termina, otras actividades se atenúan y varias partes del cuerpo dege- neran. El periodo termina con la muerte.
Desarrollo embrionario
Impregnación. Para nuestro propósito debemos considerar que el periodo embrio- nario comienza desde el momento de la impregnación. En el proceso de impregna- ción, un espermatozoide penetra en el óvulo. Como resultado de la penetración aparece la acción del huevo. Casi inmediatamente después se presenta una re- acción cortical en el huevo, como consecuencia de la penetración del esperma- tozoide, y se bloquea la entrada para cualquier otro. En algunos huevos, el mi- cropilo, que es el puerto de entrada principal del espermatozoide, a través de la membrana vitelina (a menudo llamada corion) que rodea los huevos, es sellado por material secretado por las vesículas grandes, los alveolos corticales de la cor- teza del huevo. Toda la membrana vitelina o corion debe ser también impermea- bilizada para evitar la entrada de nuevos espermatozoides. Como resultado de esto se sabe que no es común en los peces el proceso de la polispermia, tan común en otros animales. Después de terminada la reacción cortical, la membrana vitelina se transforma en la membrana de la fertilización y prosigue el proceso con el endurecimiento mediante el agua en los huevos diseminados por los peces ovípa- ros. Independientemente de la impregnación, el huevo absorbe agua y es por lo menos temporalmente transformado el corion en sustancia adhesiva, debido a los procesos osmóticos involucrados. Cuando termina la absorción del agua, el huevo se vuelve turgente y firme al tacto.
Durante varios estados de desarrollo, los huevos del pez son especialmente sensibles al manoseo. Son a menudo muy delicados y este hecho persiste hasta que aparecen los ojos. Las sacudidas y los cambios rápidos en la temperatura les son dañinos durante este periodo inicial, muy sensible. Una vez que el pigmento hace SU aparición en el iris de los ojos, los embriones son denominados huevos

286 REPRODUCCIdN
oculados. Estando en esta condición son muy duros y pueden ser inclusive trans- portados hasta distancias muy grandes siempre que las condiciones sean apropia- das, como las relacionadas con la humedad y la temperatura. Permanecen así en buenas condiciones hasta unos días antes de la eclosión. Para ese entonces, posi- blemente debido al reblandecimiento de la cáscara como preparación para casos de una emergencia, los embriones se vuelven de nuevo delicados.
Fertilización. La fertilización de un huevo no se completa sino hasta que el núcleo del óvulo y el del espermatozoide han realizado su unión en el citoplasma. La fusión de los pronúcleos masculino y femenino procedentes del espermatozoide y del óvulo, respectivamente, completa el proceso de la fertilización. Entonces, los cromosomas que traen consigo los factores hereditarios, llamados genes, son juntados durante un proceso llamado anfimixis, y los genes de los dos padres co- mienzan a producir sus efectos durante el desarrollo del embrión.
División (Figura 10.7). Tanto los procesos como los resultados del desarrollo em- brionario son similares entre los grupos más importantes de peces, aunque la información detallada que se tiene de aquéllos sea reducida. Para todos los grupos, los procesos incluyen la segmentación, que consiste en la división del huevo en células sucesivamente más pequeñas llamadas blastómeros. La segmentación resulta finalmente en la formación de un estado de blástula transitoria que con- siste en una capa única de células llamadas blastómeros, dispuesta como la super- ficie de una esfera que contiene una cavidad central, el blastocele; o se extiende en forma de placa aplanada en la superficie superior de una masa relativamente grande de vitelo. Algunas superficies celulares de la blástula están por anticipado destinadas a formar ciertos órganos. La gastrulación sigue a la blastulación y trans- forma al embrión en un estado que consiste de dos cayps, una exterior o epiblast0 y una interior o hipoblasto. Se establece entonces el eje anteroposterior. Ense- guida la gástrula se alarga y las más importantes áreas formadoras de órganos se extienden. Los procesos anteriores establecen la primera forma generalizada del cuerpo. Su velocidad está controlada por los genes y el medio ambiente, y los detalles difieren entre los grupos más importantes de peces, de acuerdo a la cantidad y distribución del vitelo, en relación al citoplasma contenido en la cé- lula huevo.
La segmentación de los huevos de los cordados es típicamente holoblás- tica o meroblástica, pero existen muchas modalidades intermedias. En el tipo holoblástico sucede que la célula entera se divide en dos, y luego en cuatro, si- guiendo cada división un plano meridional. El tercer plano de división es latitu- dinal, resultando en ocho células de casi el mismo tamaño. En la segmentación meroblástica, solamente una pequeña parte del huevo, en forma de disco, se divide para producir el blastodermo. El material vitelino permanece sin dividir para ser más tarde envuelto por los tejidos en crecimiento. Puede verse un ejem- plo clásico de segmentación holoblástica en un cefalocordado, el anfioxo (Bran- chiostorna). En este caso las células se dividen casi por igual resultando en una blástula perfecta que consiste en una bola hueca formada por células; la cavidad del interior de la blástula corresponde a la que resulta de la segmentación, o sea, el blastocele. La blástula es transformada en gástrula mediante la simple invagina- ción de un lado, al igual que una pelota hueca a la que se le ha presionado profun- damente uno de sus lados. En los huevos de las lampreas (Petromyzonidae) hay
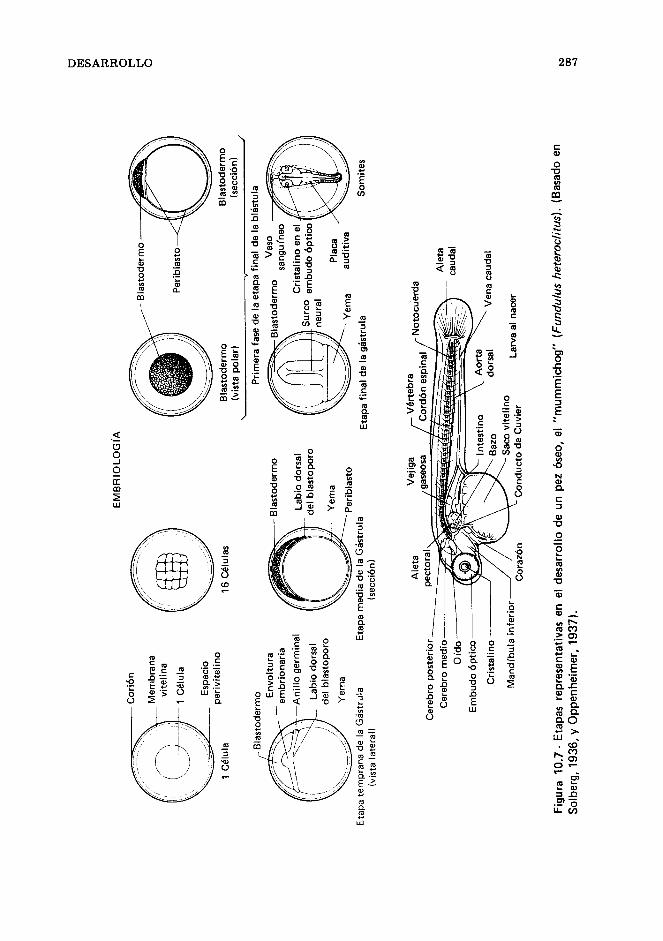
DESARROLLO 287
\ \
C W

c;:& O!&f+lC, .itje ,'lh i:.,; :!{y: <<::i::,i<:.8 :,, S > ;:ikk:-;(>s ?;":-Ti y;: C:?-.L,* ,:i.;{,&j<.i.J;; "2 q p , :
-:ai,ejo e~ ei p(:~l::; :n$t:p-,vy .; "r:$gt,,:,a;,i,~(., ,I '11: J-; lap~;;.- .A<. ?!yl& ;;Ay;. ,.:. ;e &a;.; :"i:j<2 , :
cato. Cumbo ro~ui~ado de: ia resistencia c i d vitejo! ia aegmentae;bri cc.. es ideaimera; holoblAstica como is de3 anfioxo, pero aGtn asi se ie clasifica como de tipo hulo- blástico. Las células libres de vitelo del po1.0 superior o animal (los micri?meros,a, se dividen con mayor rapidez que las del inferior o vegetal (los macrómeros) y llegan a. cubrir a éstos. La gastrulación se realiza por convergencia de células hacia e¡ blastoporo, ubicado en el extremo posterior del embrión y por la subsecuente introducción de los micrómeros sobre la línea que los separa de los macrómeros. La segmentación es algo holoblástica en el pez pulmonado de América del Sur (Lepidosiren), en los esturiones (Acipenser), en los catanes (Lepisosteus) y en el pez de aleta ondulada (Arnia), pero exhibe algunas características meroblásticas. El desarrollo inicial de estos peces es, por lo tanto, intermedio entre los tipos de segmentación holoblástico y meroblástico.
La segmentación meroblástica es la forma que prevalece cuando se dividen los huevos de los peces y de otros vertebrados. El huevo es predominantemente telolecito en los primitivos mixinos (Myxinidae) y en los tiburones y especies afines (Chondrichthyes), y su segmentación queda restringida a un disco pequeño de citoplasma en el polo animal. Lo mismo se cumple en los peces óseos (Osteich- thyes) que en los descritos arriba. La segmentación da como resultado dos tipos de células, blastodérmicas y periblásticas. Las células blastodérmicas son diferen- tes y producen el embrión. Las periblásticas o trofoblásticas se encuentran entre el vitelo y las células del blastodermo, cubren por entero a la masa vitelina y se originan de los blastómeros exteriores y más marginales. Se vuelven sinciciales y son responsables de la movilización de las reservas de vitelo. Las blástulas de los mixinos, de los tiburones y especies afines, y de los teleósteos (Actinopterygii) tienen forma de disco y por tal motivo se les llama blastodiscos. El blastocele o cavidad segmentaria se encuentra entre el blastodermo exterior y el periblast0 central que cubre al vitelo, por debajo del blastodermo.
A través de la embriología experimental ha sido posible identificar, desde la fase de la blástula, regiones destinadas a dar lugar a órganos específicos. Se sabe cuáles son las áreas de las que se presume se forman el tracto digestivo, la notocuerda y las células neurales y epidérmicas, y se ha establecido también la re- gi6n del mesodermo potencial (Figura 10.8). El término mesoderrno se refiere a la capa celular germinal media del embrión. Se forma durante la gastrulación cuando el hipoblasto se subdivide en mesoderrno y endodermo. La capa germinal externa de los primeros estadios de la gástrula, el epiblasto, es llamado ectoderms cuando el embrión está ya compuesto por tres capas.
. " .
Gastrulación. El proceso de la gastrulación es generalmente similar en la mayoría de los peces que presentan segmentación meroblástica. Hay, sin embargo, algunas diferencias dentro de los peces óseos, por ejemplo en el de aleta ondulada (Arnia), así como entre los peces óseos, los tiburones y especies afines (Chondrichthyes). Una breve descripción del proceso básico será suficiente aquí.
Cuando llega el momento de realizarse la gastrulación, el borde inferior del blastodisc0 sufre un engrosamiento y forma el "randwulst" o borde marginal al que se le agrega una capa interna llamada anillo germinal. El anillo es más grueso en dirección caudal y es reconocido como el escudo embrionario. Al dar comienzo la gastrulación las presuntas células endodérmicas del borde caudal del escudo em-

289
Ectoderm0 neural
brionario se introducen por debajo del blastoderm0 y luego prosiguen hacia ade- lante, por debajo del mismo, para transformarse en la porción endodérmica del hipoblasto. Para dar lugar a este movimiento de células se abre el blastoporo, una abertura que aparece hacia el extremo caudal del escudo embrionario. Moviéndose también hacia adentro, sobre el labio dorsal del blastoporo, hay células de la placa precordal y de la notocuerda, las que establecen el eje embrionario. Se supone que las células mesodérmicas también migran hacia adentro sobre el labio del blastoporo y se acomodan por sí mismas en ambos lados del axis embrionario, por debajo del ectodermo. La masa celular involucionada es denominada inicialmente hipoblasto. Durante la subsecuente proliferación y laminación los componentes endodérmicos y mesodérmicos del hipoblasto se organizan para formar los primordios de los sis- temas orgánicos internos.
Mientras los procesos de rellenado avanzan, como resultado de la involución de las células, el sobrecrecimiento celular del vitelo (epibolia) también continúa. En la epibolia el “randwulst” y las células del anillo germinal, que no participan en la involución pero son acompañadas por células que se presume son ectodérmi- cas, crecen hasta cubrir por fuera la masa vitelina y forman el epiblasto; la misma masa vitelina es cuhierta en su interior por el periblasto, aprovechando su propio crecimiento. Las primeras células del periblasto y el epiblasto, al cubrir al vitelo, forman el saco vitelino. Al continuar en proceso la epibolia la presunta placa neu- ral, precursora del sistema nervioso central, se transforma en una especie de cresta dispuesta a lo largo de la línea media dorsal ectodérmica. Esta quilla sólida, al estar en contacto con las células notocordales inferiores, es gradualmente cubierta por la epidermis y se vuelve eventualmente tubular hasta formar el tubo neural hueco.
El proceso de la gastrulación es completo en los peces óseos cuando la masa de vitelo se ha desarrollado mucho (Figura 10.9). Durante este proceso, algo del tejido mesodérmico situado a ambos lados y a lo largo de la notocuerda se orga-

290 REPRODUCCION
Figura 10.9 - Sección transversal diagramática del embrión de un pez removido del cascarón durante la últirna etapa embrionaria, para mos- trar cómo es que está cubierto el saco viteti- no. (Fuente: Lagler, 1956).
niza en segmentos, los somites. Es posible distinguir externamente la separación que existe entre el embrión y el saco vitelino.
Organogénesis y eclosión. En los peces, después de la gastrulación prosigue el desa- rrollo con los primeros pasos del desenvolvimiento embrionario hasta que aparece un cuerpo con forma primitiva de vertebrado (Figura 10.7). En esta etapa el cuer- po es más o menos cilíndrico y de simetría básicamente bilateral. Las regiones cefálica y faríngica se proyectan anteriormente, a partir de la masa de vitelo; el tronco se apoya en el vitelo y la cola se proyecta posteriormente. Cinco tubos fundamentales formadores de órganos han aparecido y tienen relaciones consis- tentes con la notocuerda axial.
La tubulación es responsable de la formación de cinco conductos de tejido: epidérmico (que cubre al cuerpo), neural, endodérmico (tract0 digestivo primi- tivo) y dos mesodérmicos (las cavidades somíticas y celómicas). Estos cinco tubos de tejido, más las células mesenquimáticas no especializadas cuyo origen es prin- cipalmente mesodérmico, juegan importantes papeles en la organogénesis.
El tubo ectodérmico deviene la cubierta del cuerpo o epidermis. En los peces, quedan incluidos entre sus partes derivadas la capa externa de los dientes, el epitelio y nervio olfatorios, el cristalino y el oído interno. El tubo neural, que se desarrolla a partir del hundimiento de la cresta neural, es también de origen ectodérmico. Da lugar al cerebro, a la médula espina1 y a algunas partes del ojo, incluyendo a la retina y al nervio óptico. Formando parte del mesénquima hay células asociadas, de la cresta neural, que entre otras cosas contribuyen con gan- g l io~ del sistema nervioso. Las células pigmentadas conocidas como melanóforos se originan, según parece, de la cresta neural.
Los tubos mescdérmicas del tronco se dividen inicialmente en las partes dorsal, intermedia y lateral. Mug pronto se divide el mesodermo dorsal en los somites en forma de bloc, ya mencionados. Cada somite queda subdividido en tres regiones llamadas esclerotomo, miotomo y dermatomo. El esclerotomo par-

DESARROLLO 291
ticipa en la formación del esqueleto axial. El miotomo se transforma en la mus- culatura del tronco, el esqueleto apendicular y los apéndices con sus músculos. El dermatomo da origen al tejido conectivo de la dermis de la piel y sus derivacio- nes, incluyendo a las escamas.
El mesodermo intermedio da lugar a los riñones, parte de las gónadas y sus conductos. El mesodermo lateral se separa en las capas interna y externa para limitar a la cavidad celómica. Los tejidos que cubren o limitan a las cavidades pericardial y peritoneal se originan del mesodermo lateral, como sucede también con el corazón, la sangre, los vasos sanguíneos del tronco y todas las capas del tracto digestivo, a excepción de su cubierta epitelial, que proviene del endoder- mo. El mesénquima contribuye en la cabeza a la formación de las capas exteriores del ojo, los huesos del cráneo y cara, la musculatura y las capas de dentina de los dientes.
En los peces, el endoderm0 contribuye con las células sexuales primordia- les (el tracto germinal) para las gónadas y forma el epitelio interno de todo el tracto digestivo y sus glándulas asociadas. También contribuye al desarrollo de muchas glándulas endocrinas que son derivaciones del tubo digestivo primitivo, y quedan incluidas entre ellas la tiroides y los cuerpos últimobranquiales. La eclosión resulta del reblandecimiento del corion mediante enzimas u otras sus- tancias químicas que son secretadas por glándulas ectodérmicas, situadas usual- mente en la superficie anterior, o a partir de glándulas endodérmicas que se ubi- can en la faringe. El embrión participa activamente en el desgarramiento del corion para salir del huevo. Debido a que no se conoce bien la composición química del corion, se duda todavía sobre la manera en que actúan las enzimas (corionasas). Una parte considerable del material nutritivo en el corion puede ser utilizado por el embrión a través del fluido perivitelino.
Recapitulación por etapas. Cuando son comparados los embriones y larvas de diferentes especies, del mismo tamaño o de la misma edad, sucede que se observa muy a menudo que las etapas de desarrollo son diferentes. Las comparaciones son de mayor utilidad cuando se basan en las etapas del mismo desarrollo. Y aún así, aparecen los problemas debido a las dificultades que se tienen para definir estas etapas de desarrollo. Al revisar el desarrollo embrionario, sin embargo, pode- mos nosotros seguir una secuencia de etapas, de utilidad demostrada, para hacer comparaciones entre las especies, por lo menos entre las ovíparas, como las regis- tradas para el pez “mummichog” (Fundulus heteroclitus), un ciprinodóntido (Cyprinodontidae; Figura 10.7). La edad aproximada en horas que se cita en el estudio del “mummichog” corresponde a huevos conservados a 27°C (80.6” F):
Etapa Etapa
Etapa Etapa Etapa Etapa Etapa Etapa
1. El huevo sin fertilizar. 2. Embrión unicelular (Figura 10.7), el huevo recientemente fertilizado
3. Embrión de dos células (1112 horas). 4. Embrión de cuatro células (2 horas). 5. Embrión de ocho células (2112 horas). 6. Embrión de dieciséis células (3 horas, Figura 10.7). 7. Embrión de treinta y dos células (31/2 horas). 8. Primera blástula máxima. El blastoderm0 se eleva formando una estruc-
(1 hora).

29 2
Etapa 9.
Etapa 10.
Etapa 11.
Etapa 12.
Etapa 13.
Etapa 14.
Etapa 15.
Etapa 16.
Etapa 17.
Etapa 18.
Etapa 19.
Etapa 20.
Etapa 21.
Etapa 22.
Etapa 23.
Etapa 24. Etapa 25.
Etapa 26. Etapa 27.
REPRODUCCI6N
tura como domo o sombrerillo, y se forma el periblast0 (5112 horas, Figura 10.7). Última blástula máxima. Las células del blastodermo son más pequeñas que las observadas en la etapa 8. Blástula plana. El blastodermo se ha transformado en un disco plano, en lugar de permanecer como una comba elevada, según se vio anterior- mente (8 horas). Expansión de la blástula. El blastodermo ha comenzado a crecer sobre el vitelo. Primera gástrula. El anillo germinal y el escudo embrionario son formados y comienza a abrirse el blastoporo (16 horas, Figura 10.7). Gástrula media. Cerca de la mitad del vitelo cubierto por blastodermo, la cresta neural visible justamente en la línea media del escudo em- brionario (19 horas, Figura 10.7). Última gástrula. Más de la mitad del vitelo cubierto por blastodermo, el escudo embrionario se estrecha, la cresta neural más claramente visible (22 horas, Figura 10.7). Final del blastoporo. Pequeña diferencia embrionaria a excepción de la formación de rudimentos del sistema nervioso central y posiblemente de las vesículas ópticas y primeros somites (26 horas). Comienza la expansión del cerebro anterior (para la formación de las vesículas ópticas). Las tres vesículas primarias son visibles en el cerebro. Formación de la cavidad en las vesículas ópticas, la segmentación mesodérmica produce de uno a cuatro somites. Formación de la plácoda auditiva. El número de somites es de cuatro a catorce y aparece un celoma extra-embrionario como una cavidad desarrollada por el epitelio del saco vitelino (36 horas). La cavidad aparece detrás del cerebro, en la médula espinal. Hay engrosamiento del ectoderm0 para formar el cristalino del ojo y la fosa nasal. El número de somites llega a ser de catorce a veinte (42 horas). Expansión del cerebro medio para formar los lóbulos ópticos. El número de somites queda comprendido entre veinte y veinticinco. Los melanóforos aparecen cerca de la médula espinal y se hacen presentes sobre el vitelo. Aparece el pericardio. El corazón tubular y pulsátil se vuelve visible. La localización de la aleta pectoral puede notarse debido a la concentración celular. Hay motilidad. Cerca de veintiocho somites presentan contracciones musculares. Circulación. La circulación comienza en unos treinta y cinco somites. Las paredes del cerebro anterior forman los hemisferios cerebrales. Los otolitos aparecen en el oído. Los melanóforos son conspicuos en el pericardio y la sangre fluye en los vasos vitelinos. Se observa ya el primordio de la aleta pectoral. Formación de la vesícula urinaria (como una evaginación del tract0 digestivo posterior). Formación del hígado y la cavidad peritoneal. La aleta pectoral se redondea. Hav Digmentación en las Daredes peritoneales.

DESARROLLO 293
Etapa 29. Se establece la circulación en la aleta pectoral. La aleta adquiere mo-
Etapa 30. Los radios aparecen en la aleta caudal. Se forma la mandíbula inferior. Etapa 31. Formación de la vejiga gaseosa (como undivertículo deltractodigestivo).
Etapa 32. Eclosión. Quedan todavía residuos del vitelo (11 días, Figura 10.7). Etapa 33. Pigmentación y crecimiento de la vejiga gaseosa. Etapa 34. Se completa la absorción del vitelo (12 días o más).
vimientos.
Adquieren movimientos los ojos y las mandíbulas.
En el desarrollo larvario subsecuente la piel y las escamas no han terminado todavía su diferenciación, el esqueleto axial no ha llegado todavía a su término, tienen que formarse todavía las aletas dorsal y anal y los riñones propios de un adulto, y se tienen que realizar todavía las proporciones corporales definitivas y la pigmentación.
Interrelaciones nutritivas y respiratorias. Hay diversas relaciones nutritivas y res- piratorias en los embriones de las diferentes clases de peces, las cuales dependen de que los jóvenes hayan nacido vivos o hayan resultado de la eclosión de huevos puestos por peces ovíparos.
Los embriones de huevos de peces ovíparos dependen totalmente del vitelo, y este material es indispensable para su alimentación. Muchas especies presentan una o más gotas de aceite en o cerca de la masa vitelina, que sirve tanto como un recurso potencial de energía como un órgano para la flotación o el equilibrio. El material orgánico suplementario disuelto y las sales minerales pueden ser absor- bidas directamente a partir del agua. La respiración y excreción de los huevos depositados en el agua se hacen a través de membranas semipermeables del huevo y la cáscara.
Hay intercambio nutritivo y respiratorio en los embriones de todos los pe- ces vivíparos, in utero o in ovario, inclusive cuando falta una disposición placen- taria, como sucede en los tiburones o peces óseos ovovivíparos. En estos grupos las secreciones uterinas u ováricas realizan una unión fisiológica entre la madre y el embrión. El intercambio es más intimo, sin embargo, entre los peces placentarios vivíparos. La cubierta uterina varía, histológicamente, desde una capa secretora de mucus formada por células epiteliales de forma cúbica en las especies que de- penden del vitelo del huevo para su alimentación (Acanthias vulgaris), a través de formas zon capas cobertoras de tejido secretor-seroso moderadamente dobladas o plegadas (Torpedo), hasta los que tienen cubiertas uterinas rodeadas de vellosi- dades (trophonemata) de varias longitudes y complejidades, que secretan grasa en abundancia (Trygon violacea). El vitelo es digerido en su mayor parte dentro del intestino de los embriones que dependen de aquél para su nutrición (Squalus acanthias). La verdadera viviparidad con una placenta formada por un saco vite- lino está limitada a ciertas especies de dos familias de tiburones (Carcharhinidae y Sphyrnidae). Los embriones de Sphyrnu tiburo desarrollan redes circulatorias elaboradas en las paredes de sus sacos vitelinos, que se pliegan mucho. La mucosa uterina se pliega o se arruga y la interdigitación de estas dos series de pliegues con un adelgazamiento de sus epitelios acercan íntimamente a las circulaciones ma- terna y fetal. El embrión se fija a la placenta mediante un conducto vitelino modificado (cordón umbilical).
Entre los peces óseos vivíparos la nutrición embrionaria y la respiración

294 REPRODUCCI6N
pueden relacionar con éxito muchos recursos y órganos, algunos de ellos muy especiales. En lus peces vivíparos mexicanos (Goodeidae) el saco vitelino embrio- nario, el saco pericárdico y las trofotenias se hacen cargo de diversas formas del trabajo. Las trofotenias son las extensiones en forma de dedo del tejido rectal. En los pececillos de la familia Jenynsidae existe una relación similar entre el embrión y la cavidad ovárica y los tejidos, pero las trofotenias son sustituidas por trofonemas, ext,ensiones en forma de gusano de la pared del ovario, que crecen en las cimaras branquiales del embrión. Cuando se realiza la nutrición embrio- naria las brotulas (Brotulidae) relacionan el saco vitelino, la cubierta del cuerpo, las branquias y finalmente el tubo digestivo propiamente dicho del embrión a través de la ingestión del fluido intraovárico y mediante el consumo de los tejidos ováricos, incluyendo a las células del huevo. Algunos embriones jenínsidos practican el canibalismo en detrimento de otros dentro de la misma cavidad intraovárica.
En los huevos de las truchas y salmones (Salmoninaej, el nivel de carbo- hidrato aumenta gradualmente durante el desarrollo, aunque el nivel de glucosa desciende temporalmente al momento de la eclosión. El glucógeno es probable- mente sintetizado casi al hacer eclosión el huevo y es almacenado en el hígado del embr ih al término de la etapa del saco vitelino. La grasa es empleada como combustible, y posiblemente un 70 a 80% de ella es consumida durante todo el periodo de desarrollo. Se estima en 68 calorías la contribución de la grasa por huevo en los momentos en que el vitelo es absorbido. La proteína contribuye con unas 45.9 calorías para realizar el proceso. Y en él también participa la proteína del vitelo como, por ejemplo, en varias especies de truchas, donde se emplea de un 60 a un 63% ; la grasa también proporciona energía en un 16 a un 40%. El re- querimiento energético del desarrollo es de unas 80 calorías y la combustión metabólica emplea 114 calorías. La eficiencia es elevada; durante el crecimiento del embrión, cerca del 40X de la energía originalmente incorporada al huevo es convertida; el resto, cerca del 60% , se emplea en actividades tales como la osmo- rregulación, la secreción, la circulación y los movimientos.
Desarrollo transicional o lawario (Figuras 10.10 y 10.11)
Luego de la etapa embrionaria, en los primeros días de su vida libre, el pez tiene distintos grados de desarrollo, que varían de acuerdo a la especie. Generalmente, las etapas de desarrollo que se presentan antes de llegar al estado adulto aunque después del nacimiento o la salida del huevo, son denominadas larvarias, y los jóvenes son llamados larvas durante este periodo. Los peces larvarios son también llamados cría. La longitud del periodo larvario puede comprender sólo unos mo- mentos o varios años. La larva amocete de la lamprea marina (Petromyzon rnari- nus), por ejemplo, requiere de cinco o más años antes de sufrir su metamorfosis.
En los peces, es muy común que exista el desarrollo larvario y, aunque no de carácter universal, éste es divisible en dos estados, el prelarval y el postlarval. La prelarva se distingue por la presencia del saco vitelino y es llamada común- mente cría vitelina por los cultivadores de peces. Cuando el saco vitelino desapa- rece, en algunas especies el pececillo viene a ser un adulto pequeño, denominado comúnmente como alevino o cría mayor, por algunos cultivadores. El desarrollo directo es característico, por ejemplo, en las truchas y salmones (Salmoninae), los bagres de agua dulce de América del Norte (Ictaluridae), los mixinos (Myxi-

DESARROLLO 295
nidae) y los tiburones y sus especies afines (Chondrichthyes). En otros peces, un estado postlarval diferente le sigue al prelarval, libre de saco vitelino. Una post- larva debe sufrir una metamorfosis para perder sus estructuras larvarias y obte- ner las características de adulto. El criterio para reconocer el estado postlarval en muchas especies, es muy subjetivo. Ha sido reconocido un desarrollo indirecto en muchas familias, incluyendo, entre otras, a los peces pulmonados de América del Sur (Lepidosirenidae), lampreas (Petromyzonidae; Figura 10.10), el pez de aleta ondulada (Amiidae), sardinas (Clupeidae), carpas chupadoras (Catostomi- dae), carpas (Cyprinidae), peces sol (Centrarchidae), lófidos (Lophiidae), mólidos (Molidae), peces ángel de las profundidades (Ceratiidae), pez escorpión (Scorpae- nidae) y anguilas (Anguillidae). Son las diferencias entre las postlarvas y los adul- tos en algunos peces tan notables que han dado lugar, de vez en cuando, a ciertas confusiones taxonómicas. ¡Es así como las larvas de la anguila (Anguilla) fueron descritas una vez como del género Leptocephalus y las de las lampreas (Petromy- zonidae) en el género separado Ammocetes!
Las características de las postlarvas son muchas y muy diversas. Incluyen desde una precisa pigmentación, como ocurre en muchas carpas (Cyprinidae), y transparencia, como sucede en la postlarva leptocéfala de la anguila (AnguiZla), hasta características anatómicas profundas. Muchos peces marinos tienen una transición muy tajante, desde una postlarva pelágica hasta un cavador, juvenil o adulto, del fondo marino. En ninguna parte está mejor demostrado este cambio que en la transformación que sufre el joven pelágico del grupo de los peces planos (Pleuronectiformes), de posición vertical y de simetría bilateral para volverse habitante de los fondos después de la metamorfosis, y presentar los dos ojos en un sólo lado de la cabeza y nadar con el lado oculado hacia arriba. Otras post- larvas extraordinarias han sido dotadas de elaborados dispositivos para la flota- ción. Por ejemplo, las aletas pélvicas de los 16fidos larvarios (Lophius) son alarga-
METAMORFOSIS DE LA LAMPREA
Piel cubriendo al ojo Capuch6n oral carnoso, sin dientes
(u) Larva
Figura 10.10 - Meta- morfosis en la lamprea marina (Petromyzon marinus); (a) larva casi a t6rrnino; (b) adulto re- cientemente metamor- foseado. Ambos dibu- jos están a la misma escala.
oral - Diente &neo
(b ) Adulto recientemente metamorfoseado

296 REPRODUCCION
das, y las molas (Molidae) tienen espinas corporales (así como una aleta caudal muy evidente) que se pierde durante la metamorfosis. En algunos miembros de la familia de los peces ángel (Ceratiidae) de los fondos marinos y en la fami- lia de los peces escorpión (Scorpaenidae);la piel se infla hacia afuera. La larva leptocéfala, en forma de hoja, de las anguilas (Anguilliformes), y las larvas de los peces machete (Hops) , los peces de hueso (Albulidae) y sus parientes, y muchas sardinas (Clupeidae), tienen todas una sustancia mucosa, acuosa, en los músculos, de tal modo que los vuelve muy transparentes y los hace flotar. El agrandamiento en las larvas de los radios anteriores de la aleta dorsal para formar estructuras flotadoras son visibles en un pez marino (Diploprion; Serranidae), el lenguado (Laeops), el “dealfish” (Trachipterus), los peces perla (Curupus) y los lófi- dos (Lophius). Las larvas ammocetes de las lampreas (Petromyzonidae) son excava- doras antes de sufrir la metamorfosis, esencialmente ciegas y equipadas con una cubierta en forma de cofia, oral, y células ciliadas especializadas en la faringe para llevar partículas alimenticias hacia el esófago. Cuando las larvas ammocetes asu- men la condición adulta, cambian su conducta y morfología. Los adultos son nadadores libres, no presentan la cubierta oral o aparato larvario para la alimen- tación, y desarrollan una boca en forma de disco chupador, un embudo bucal con dientes y, como es evidente, ojos funcionales. Los dientes son fuertes en especies parásitas como la lamprea de mar (Petrornyzon marinus), pero son débi- les en las especies no parásitas, como en la lamprea americana de los arroyos (Lumpetru Zumottei), que no se alimenta después de la metamorfosis.
Las aletas externas en forma de pluma son características premetamórficas de los peces pulmonados de Africa y América del Sur, Protopterus y Lepidosiren, respectivamente, y también de los polípteros (Polypterus) de Africa, y de ciertos elasmobranquios (Figura 8.7). Las postlarvas de los catanes (Lepisosteus) y el pez de aleta ondulada (Arniu) tienen órganos adhesivos en sus narices. En los “deal- fish” (Trachipterus) los radios de aleta que son modestamente alargados en el adulto difieren tanto en su gran número como en su alargamiento en el joven antes de sufrir la metamorfosis. Los peces espada (Xiphiidae) y sus parientes más cer- canos los merlines, los peces vela y peces lanza (Istiophoridae) carecen todos, cuando son jóvenes, de la enorme prolongación en forma de pico de la nariz. El alargamiento de la mandíbula inferior en los peces aguja (Belonidae) tiene lugar mucho antes de que la mandíbula superior crezca hasta igualarlo.
Unos cuantos peces han eliminado con mucho éxito, ya sea la etapa larva- ria de vida libre (mediante un desarrollo más directo) o raramente la etapa adulta (mediante la neotenia). Los machos de algunos embiotócidos vivíparos (Embio- tocidae) son sexualmente maduros al momento de nacer, eliminando entonces los peligros de una vida larvaria pelágica. Los peces neoténicos que maduran se- xualmente mientras retienen sus características larvarias y más tarde las conser- van, incluyen a ciertos gobios (Gobiidae), peces hielo del Asia oriental (Salangidae) y las larvas del género Schindleriu que deben alcanzar la madurez mientras son aún una postlarva de solamente 1 2 milímetros de longitud. Un pariente de los pe- ces boca de los fondos marinos (estomiatoides), el pez de ojos pedunculados (Idiacanthus), presenta neotenia sexualmente dimórfica y muy marcada: el ta- maño de los machos maduros es menor de un sexto del que presentan las hembras, y son larvoideos.

DESARROLLO 297
El desarrollo de juvenil a adulto
La metamorfosis típica o terminación del desarrollo embrionario es seguida por un periodo de crecimiento en tamaño acompañado por la maduración de las gó- nadas. Las hembras usualmente retienen las mayores dimensiones, pero general- mente los machos maduran primero. Un pez pasa, como individuo, de joven o pequeño, con forma de adulto en miniatura, a través de una condición juvenil, a la etapa de la maduración sexual. El tiempo empleado puede ser corto, de 4 a 10 semanas (variando con la temperatura) requeridas para Lebistes, o más largo, como los 6 a 12 años requeridos por las anguilas de agua dulce (Anguilla), o los 15 o más años que requieren los esturiones (Acipenser). En las larvas de algunos pe- ces, unos cuantos, la mayor parte del crecimiento es logrado antes de la metamor- fosis. Aparece, por ejemplo, en las lampreas inofensivas, no parásitas (Lampetra y algunas especies de Ichthyomyzon). Estas lampreas no consumen alimento después de la metamorfosis, de aquí que no haya crecimiento sino sólo madura- ción de las gónadas, una estación de desove y la muerte.
Cualquier consideración sobre el desarrollo después de la eclosión debe relacionarse con el inicio de la alimentación. Como ya se ha dicho, es en las pri- meras etapas de la vida de los peces cuando ocurren las grandes mortandades (excediendo algunas veces el 99% ). Es en este periodo cuando el alimento es un factor crítico para la supervivencia. Los peces pequeños, al momento de la eclo- sión del huevo, requieren de alimentos pequeños. De aquí que la primera dieta alimenticia esté formada para la mayoría de los peces de plancton, bacterias o partículas pequeñas de desperdicios o restos orgánicos, ya sea que se encuentren en el fondo o estén libres en el agua, dependiendo del lugar donde la larva viva (Figura 5.1).
Otra característica propia de los primeros estadios de la vida del pez es su pigmentación (Figura 10.11). Muchas clases de peces, en etapas particulares del desarrollo, presentan cromatóforos, especialmente melanóforos, distribuidos en patrones que permiten fácilmente la identificación. La mayoría de los peces con huevos sumergibles pasan por un periodo, después de hacer eclosión el huevo, en el cual sus movimientos son torpes como resultado del enorme peso relativo que tienen que soportar al cargar con el saco vitelino. A medida que el vitelo es absor- bido, los pececillos que salieron del huevo en el fondo pueden pronto comenzar a levantarse y moverse con movimientos directos, y se vuelven más y más inclina- dos al medio ambiente para alimentarse.
Senectud
La senectud se refiere a los cambios degenerativos que van de la mano con la edad en un organismo. En los peces se caracteriza por la detención paulatina del creci- miento longitudinal, un aumento de la tasa de mortalidad, la pérdida gradual de la capacidad reproductora, un aumento de las anormalidades en relación a la cría y, algunas veces, una relativa elevación del nivel de obesidad. Por lo general, los ma- chos envejecen y mueren a más temprana edad que las hembras. El periodo de la senectud muestra también típicamente una disminución de la actividad acompa- ñada de cambios en la alimentación, distribución y otros hábitos. El periodo puede ser largo cuando el pez vive en su medio ambiente natural, en libertad,

REPRODUCCIdN
Figura 10.11 - Desarrollo posterior al nacimiento de un pez óseo, la macarela (Trachurus symmetricus). (a) Larva de 4.9 mm a la que le falta la aleta p6I- vica y el desarrollo de los radios en las aletas dorsal y anal. (b) Larva de 10 mm con la aleta pélvica justamente apareciendo pero con los radios ya presentes en las otras aletas; nótense los cambios en la pigmentación sobre (a). (c ) Juve- nil de 28 mm mostrando el establecimiento de la forma adulta y el desarrollo posterior de la pigmentación sobre (a) y (6). (Fuente: Ahlstrom y Ball, 1954).
pero puede ser prolongado viviendo en cautiverio. Por ejemplo, el lucio septen- trional (Esox lucius) ha alcanzado la edad de 25 años en su medio natural, según se ha registrado, pero en cautiverio ha llegado a vivir hasta 75 años. La senectud puede comenzar cuando el individuo es muy pequeño, como sucede en el delicado “goby” de Filipinas (Mistichthys Euzonensis) cuya longitud nunca excede del centímetro y medio. También puede aparecer cuando el individuo es relativamente joven, como sucede también en otro “goby” (Latrunculus pellucidus), cuya vida media es de solamente un año.
En las poblaciones silvestres de peces la muerte sobreviene tan rápido como el crecimiento individual.
Tasas y factores de desarrollo
Al hacer el cultivo práctico de los peces se considera que el periodo de desarrollo embrionario, o sea, el periodo de desarrollo, se extiende desde la impregnación

DESARROLLO 299
hasta la fecha promedio de la eclosión de un grupo de huevos, o el nacimiento de una cría. Este intervalo de tiempo varía considerablemente de acuerdo a la espe- cie e, inclusive, un poco entre la misma cría de padres individuales dentro de la especie. Su duración es duramente influenciada por las condiciones ambientales. Por definición, el periodo de desarrollo termina cuando ha hecho eclosión la mitad de los huevos en un grupo considerado. La eclosión (o nacimiento), con- siderada como periodo, es el intervalo de tiempo en el cual un conjunto de huevos hace eclosión (o los pequeños emergen de la hembra madre), desde el primero has- ta el último.
Muchos factores afectan al proceso total de desarrollo. Ellos influyen en el éxito y determinan la falla. También afectan la tasa y determinan la forma y la estructura. Entre los factores del medio ambiente es muy notable la temperatura. Por ejemplo, la temperatura afecta la tasa de desarrollo o la fracción del proceso de desarrollo realizado por día. Este grado o tasa ha sido expresado simplemente como el recíproco del periodo de desarrollo en días. Cuanto más grande sea la fracción, más rápido será el desarrollo. Por lo tanto, un pez con un periodo de desarrollo de 88 días tiene una tasa de 1/88, mientras que uno que se desarrolla en 9 días tendrá 1/9. Para permitir una comparación entre las especies, esta tasa puede ser también expresada con valores decimales (por ejemplo, 0.011 o 0.111 respectivamente), para las fracciones consideradas.
Temperatura. Tanto el periodo de desarrollo como el de eclosión son generalmente más cortos cuando las temperaturas son elevadas que cuando son bajas. Es inte- resante ver cómo muchas especies normalmente se desarrollan en la naturaleza bajo condiciones de temperatura que no son las óptimas, según se ha determinado mediante experimentos de laboratorio. Las especies difieren en cuanto a la tem- peratura óptima y las tolerancias requeridas durante su desarrollo; pero para todas las especies hay temperaturas que son demasiado bajas y demasiado elevadas para que se lleve al cabo el desarrollo. Los cambios extremos o súbitos pueden ser leta- les. El margen de temperatura dentro del cual puede realizarse el desarrollo nor- mal puede ser amplio. El ciprinodóntido Fundulus heteroclitus, por ejemplo, se desarrolla a temperaturas que van desde los 12°C (53.6"F) a los 27°C (80.6"F) con un 2% de anormalidades menores.
A continuación mostramos cómo difiere la tasa de desarrollo entre las espe- cies. A una temperatura promedio de 6.1"C (43°F) durante el periodo de desa- rrollo, la trucha café (Salmo trutta) requiere de 88 días; la trucha de arroyo (Sal- velinus fontinulis), 80; el salmón "sockeye" (Oncorhynchus nerka), 75; la trucha arco iris (Salmo guirdneri), 61; la perca blanca (Morone americana), 20; el bacalao (Gadus morhua), 14; y el pez "pollock" (Pollachius virens), 9.
Ha habido durante largo tiempo interés en estudiar la relación directa de la temperatura con el desarrollo de los individuos dentro de una especie. Esa relación fue determinada hace más de cien años para la trucha de arroyo (Salvelinus fonti- nalis). En este pez el periodo de desarrollo decrece con la elevación de la tempe- ratura promedio del agua (en grados Fahrenheit) como sigue: 37", 165 días; 41", 103 días; 48", 56 días; 50", 47 días, y 54", 32 días.* Una generalización que se estableció primeramente para 10s huevos de la trucha de arroyo fue que el periodo de desarrollo era de 50 días para una temperatura promedio del agua de
*2.8"C, 5"C, 8.9"C, 10°C y 12"C, respectivamente (N. del T.).

300 REPRODUCCI6N
50°F (lO"C), y que deberían requerirse por cada grado de ascenso o descenso de la temperatura, más o menos 5 días, respectivamente. Durante largo tiempo este esquema sirvió en los viveros para hacer las correspondientes estimaciones.
Se ha reconocido subsecuentemente que el tiempo indispensable para al- canzar un estado definitivo de desarrollo, multiplicado por la temperatura, da una constante ( K ) en la fórmula y T = K que de varias maneras se ha propuesto en el pasado. Cuando el periodo de desarrollo es anotado en el eje de las abcisas y la temperatura en el de las ordenadas, la figura resultante es una hipérbola rectan- gular (Figura 10.12). En la ecuación de la temperatura anterior, y viene a ser el tiempo necesario para alcanzar cierto estado de desarrollo siguiendo a la fertili- zación (por ejemplo, al momento de hacer eclosión), y T la temperatura a la cual el desarrollo está teniendo lugar. La correspondiente ecuación de intercepciones para el grado de desarrollo es u = KO + K1 T donde u es la velocidad o grado (esto es, 1,000 veces el recíproco de y ) y KO y K1 son las constantes para la intercep- ción con la ordenada (cero biológico) y la pendiente, respectivamente. Una grá- fica de la tasa o grado (Figura 10.13) contra la temperatura da una línea recta útil para predecir las desviaciones; sin embargo, esta gráfica es curvilínea sobre amplios márgenes de temperatura, la simple linearidad se aplica sobre un margen muy estrecho.
La temperatura influye en la longitud del periodo de eclosión, así como en el intervalo total de desarrollo. Se indica a continuación un ejemplo de esta in- fluencia de la temperatura en el periodo de eclosión, a partir de diferentes mues- tras tomadas de un solo lote de huevos de la trucha arco iris (Salmo guírdneri) y mantenidas a diferentes temperaturas.
- 160 2 140 Y) Fig. 10.12
- o 120
a
U 1
- F loo
80
60
.- O 40 U
2 20 O 2 4 6 8 ?O 12 14 16
Temperatura
Fig. 10.13 Temperatura
Figura 10.12 - Efecto de la temperatura promedio del agua en el periodo del desarrollo (la fertilización significando fecha de la eclosión del huevo) en la trucha de arroyo (Salvelinus fontinalis). (Basado en Hayes, 1949).
Figura 10.13 - Grado de desarrollo (l,OOO/periodo del desarrollo en dias) en relaci6n a la temperatura promedio durante el periodo de desarrollo. Mismos datos como los de la Figura 10.12. (Fuente: Hayes, 1949).

DESARROLLO 301
Temperatura Periodo de eclosión al día más cercano
"F "C
42.3 5.7 14 46.8 8.2 8 55.2 12.9 6 63.7 17.6 3
La tasa o grado de eclosión dentro del periodo de eclosión puede esperarse que siga la figura de una curva de distribución en forma de campana. El punto o cresta de esta curva, bajo situaciones ordinarias, corresponde a la eclosión del 50% de los huevos y a la terminación del periodo de desarrollo.
La temperatura afecta a la eficiencia con la cual el alimento almacenado en el huevo es convertido en peso del cuerpo del embrión. En un experimento realizado con recién nacidos mediante la eclosión de huevos del salmón del Atlán- tico (Salmo salar), fueron comparadas las ganancias en el peso de las larvas con las pérdidas en peso del saco vitelino en los peces guardados en una serie de tanques que diferían en temperatura, de 0.2 a 16°C. La eficiencia en la conversión del vitelo en carne de pescado era baja y constante, alrededor del 42%, en los tanques más fríos. Comenzó a incrementarse a los 5°C y alcanzó un máximo de eficiencia hasta cerca del 60% en el tanque más caliente, cuando se hizo el cálculo a lo largo de un intervalo de 10 días.
Luz. La luz puede influir en el desarrollo del embrión del pez, pero los datos disponibles son ambiguos. Aunque algunos experimentos realizados con huevos de trucha y salmón (Salmoninae) han indicado que su exposición a la luz fluores- cente ha tenido como resultado una elevada mortalidad, otros experimentos no han mostrado efectos en los embriones expuestos a bajos niveles de luz fluores- cente (que es normalmente empleada en los viveros). Y aún así, la trucha de gar- ganta cortada (Salmo clarki), en su estado juvenil, mostró el más rápido creci- miento bajo la luz.
Gases disueltos. Los gases disueltos en el agua constituyen también factores de desarrollo, especialmente de los huevos de los peces ovíparos. Puede esperarse que tanto las concentraciones óptimas como los extremos intolerables del oxí- geno disuelto existen y varían de acuerdo a las especies. El margen generalmente aceptado para el desarrollo queda comprendido entre 4 y 12 partes por millón (ppm) de oxígeno disuelto. Las especies de aguas frías, desovadoras en las co- rrientes de agua dulce, tienen requerimientos más elevados que los que tienen las perezosas, de aguas calientes. Las condiciones y los requerimientos de los peces de las aguas profundas son casi totalmente desconocidos, primordialmente de- bido a las dificultades para obtener peces saludables de lo más profundo del océa- no, además de mantenerlos y experimentar con ellos bajo muy elevadas presio- nes barométricas.
La presión del oxígeno puede influenciar el número de los elementos merís- ticos. La disminución de la presión de oxígeno durante el desarrollo embrionario

produce un aumento en el nfimero de vbrtebras de !a trucha café (Salmo tru- tZa 1
Por lo menos dos gases disueltos en el agua, el dióxido de carbono y el amoníaco, son tóxicos para los peces y sus embriones. La información de que se dispone sugiere que bajo las condiciones ordinarias de oxígeno disuelto, .las con- centraciones de dióxido de carbono superiores a 8 o 9 ppm, tienen poco efecto en el desarrollo. En concentraciones de 10 a 30 ppm puede esperarse un gradual impedimento del proceso, mientras que las concentraciones mayores de 30 ppm pueden impedir el desarrollo y acarrear la muerte. Es interesante ver cómo al ele- varse las presiones del dióxido de carbono durante el desarrollo embrionario, decrece el número de vértebras en la trucha café (Salmo trutta). Niveles muy elevados, entre 55 y 80 ppm, aumentan la mortalidad de los embriones oculados y también el número de larvas deformadas. El amoníaco puede ser tóxico a ba- jas concentraciones. Los especialistas en la cualidad del agua reconocen al 1.5 ppm como tolerable para la vida acuática, pero expresan dudas sobre las grandes con- centraciones. El agua supersaturada con nitrógeno, como la que existe bajo algu- nos embalses, puede provocar en la cría una enfermedad por burbujeo gaseoso.
Salinidad. La salinidad puede ser dañina para los huevos de los peces de agua dulce y, por el contrario, el agua dulce puede ser nociva para los huevos de las especies marinas. Esto es claramente visible en las barreras que las aguas dulces o marinas constituyen para los peces cuya vida debe quedar restringida precisa- mente para el medio contrario (Capítulo 14). Si el contenido salino del agua es intolerablemente elevado, los huevos de peces de agua dulce que en ella son su- mergidos podrían perder agua y morir por encogimiento. En forma similar, si los huevos de peces marinos son sumergidos en agua dulce pueden absorber agua y reventar. La salinidad juega también un papel selectivo en el desarrollo de algunas estructuras, como se ve experimentalmente en el efecto de su variación en el nú- mero de placas óseas que se desarrollan a los lados de la espinocha de tres espinas (Gasterosteus aculeatus).
Glándulas endocrinas. Se reconoce bien la importancia de los factores endo- crinos en el desarrollo. Son de particular interés los papeles que juegan la pituitaria y la tiroides en la metamorfosis (Capítulo 11).
Cantidad de vitelo. La cantidad de vitelo presente en el huevo tiene relación con la velocidad o grado de desarrollo. Parece común en los peces que cuanto ma- yor sea la cantidad de vitelo, más lento será el grado de desarrollo. Es muy común entre los animales que la gran dependencia hacia los padres parezca estar asociada con la relativa lentitud en el desarrollo, pero no se sabe si lo anterior se pueda aplicar a los peces.
Fijación de los elementos merísticos
El grado de desarrollo tarnbibn está involucrado en la determinación de los ele- mentos merísticos como las vértebras, los radios de las aletas, las branquispinas y los números de hileras de escamas. Por ejemplo, el pececillo de brillo dorado (Notemigonus crysoleucas) de América del Norte tiene, cuando crece en el norte,

REFERENCIAS. 303
más elementos merísticos que cuando crece en el sur, hacia el borde de su área de distribución, cerca del Golfo de México. Debido a que se emplea menos energía durante el metabolismo en las regiones de baja temperatura de las latitudes sep- tentrionales, mayor cantidad del material nutritivo del huevo puede quedar dis- ponible para la síntesis de los elementos merísticos. Pareciera ser que los factores ambientales cambian la relación entre el crecimiento y la diferenciación. Si la diferenciación es tardía, más tejido quedará disponible para su diferenciación, resultando en un número mayor. Analizado experimentalmente, el mecanismo de ese control es más complicado que cuando se le juzga a partir de observacio- nes de campo. Submuestras de huevos fertilizados de un par de padres de la trucha cafe (Salmo trutta), ambos con el mismo número de vértebras, fueron tratados con temperaturas bajas, intermedias y elevadas, constantes todas, durante su desa- rrollo. El número de vértebras que resultó fue el más bajo para la temperatura intermedia y el más alto para las temperaturas bajas y elevadas. El más alto número de radios apareció en las aletas dorsal, anal y pectoral a la temperatura inter- media, pero fue menor tanto en las más bajas y elevadas temperaturas. En el mismo grupo de experimentos se comprobó que los números de vértebras y los elementos de la aleta anal son fijados en las etapas embrionarias antes de que el ojo esté totalmente formado, o sea, mucho antes de hacer eclosión; los radios de las aletas dorsal y pectoral aparecen más tarde, como sucede también con el número de escamas en la linea lateral de la trucha arco iris (Salmo gairdneri).
Teoría biogenética
Ninguna discusión de la embriología de los peces podría ser completa sin hacer mención de la evidencia que los embriones ofrecen para reforzar la teoría bioge- nética, cuando afirma que la ontogenia es la recapitulación de la filogenia. El posible soporte para la biogénesis queda bien ilustrado con el desarrollo de la condición heterocerca del esqueleto caudal, entre otras características de los pe- ces. La terminación heterocerca de las vértebras aparece en los embriones de muchas clases de peces, no importando que la cola definitiva sea o no heterocerca, o de algún otro tipo como la homocerca, que predomina. Esta secuencia en el desarrollo ha sido adoptada como evidencia de que la condición heterocerca es la más primitiva, sobre la base de que la ontogenia o desarrollo embrionario es la recapitulación de la filogenia o evaluación de la estructura.
REFERENCIAS ESPECIALES EN REPRODUCCION Y DESARROLLO
Ahlstrom, E.H., y O.P. Ball. 1954. Description of eggs and larvae of jack mackerel (Trachurus symmetricus) and distribution and abundance of larvae in 1950 and 1951. U.S. Fish and Wildlife Service Fish Bull., 56(97):209-245.
Armstrong, C.N., y A.J. Marshall. 1964. Intersexuality in vertebrates including man. Academic Press, Nueva York.
Battle, H. 1940. The embriology and larval development of the goldfish (Carassius auratus L. ) from Lake Erie. Ohio Jour. Sci., 40(2):82-93.
Blaxter, J.H.S. 1969. Development: Eggs and larvae. En Fish physiology, 3 :177-252. Academic Press. Nueva York.

304 REPRODUCCI6N
Breder, C.M., Jr. y D.E. Rosen. 1966. Modes of reproduction in fishes. Natural History Press,
Echelle, A.A. 1972. Developmental rates and tolerances of the Red River pupfish, Cyprinodon
Gerking, S.D. 1957. Evidence of aging in natural populations of fishes. Gerontologia,
Hayes, F.R. 1949. The growth, general chemistry, and temperature relations of salmonoid
Hoar, W.S. 1969, Reproduction. En Fishphysiology, 3:l-72. Academic Press, Nueva York. Hubbs, Clark. 1967. Analysis of phylogenetic relationship using hybridization techniques.
Symposium of newer trends in taxonomy. Bull. Nat. Inst. Sei. India, 34:48-59. Hubbs, Clark, y D.F. Burnside. 1972. Developmental sequences of Zygonectes notatus at
several temperatures. Copeia, 1972(4):862-865. Kerr, J.G. 1907. The development of Polypterus senegalus Cub. En Budgett memorial volume,
Garden City, N.Y.
rubrofuviatilis. Southwestern Nat., 17(1):55-60.
1(5):287-305.
eggs. Quart. Rev. Bioi., 24(4):281-308.
PP. 195-284. - . 1909. Normal plates of the development of Lepidosiren paradoxa and Protopterus annectens. Normeltafeln z. Entw. Gesch. der Wirbelt., 1O:l-163.
Copeia, 1965(2):224-228.
McGraw-Hill Book Company, Nueva York.
Mathews, S.B. 1965. Reproductive behavior of the Sacramento perch, Archoplites interruptus.
Nelson, O.E. 1953. Comparative embryology of the vertebrates. The Blakiston Company.
Oppenheimer, J.M. 1937. The normal stages of Fundulus heteroclitus. Anat. Rec., 68(1):1-16. Orton, G.L. 1953. The systematics of vertebrate larvae. Systematic Zool., 2(2):63-75. Schultz, R.J. 1971. Special adaptive problems associated with unisexual fishes. A m . Zool.,
11:351-560. - . 1973. Unisexual fish: laboratory synthesis of a “species”. Sci., 179(69):180-181. Solberg, A.N. 1938. The development of a bony fish. Progr. Fish-Culturist, 4O:l-19. Taning, A.V. 1952. Experimental study of meristic characters in fishes. Biol. Rev., 27:169-193. Tinbergen, N. 1953. Social behaviour in animals. Methuen and Co., Londres, John Wiley and
van Tienhoven, A. 1968. Reproductive physiology of vertebrates. W. B. Saunders Company,
Wilson, H.V. 1891. The embryology of the sea bass (Serranus atrarius). Bull. U.S. Fish Comm. ,
Sons, Nueva York.
Filadelfia.
9(1889):209-278.

Capítulo 11
Integración
EL PAPEL DE LOS SISTEMAS NERVIOSO Y ENDOCRINO
Los vertebrados responden a los estímulos del medio ambiente a través de los órganos de los sentidos, el cerebro y/o la médula espina1 que desvían los impul- sos a los músculos o las glándulas. En los peces, las respuestas musculares resultan frecuentemente en movimientos de todo el cuerpo más que de un sólo apén- dice. Ciertos cambios lentos y cíclicos del medio ambiente, como son las eleva- ciones de temperatura de acuerdo a las estaciones y los cambios en la longitud del día, también afectan a una o más glándulas endocrinas, por medio del sistema nervioso. Las secreciones hormonales de las glándulas endocrinas (Tabla 11.3) actúan sobre órganos específicos, los cuales a su vez pueden ser también glándulas endocrinas, o bien las secreciones tienen un efecto difuso en el metabolismo general.
Los sistemas nervioso y endocrino son muy interdependientes y a menudo actúan juntos. La conducta reproductora de muchos peces, por ejemplo, es in- fluenciada por los sentidos a través de su percepción de los cambios del medio ambiente que aparecen cuando se aproxima la primavera; muchas glándulas endo- crinas son afectadas y actúan a su vez sobre los ovarios y los testículos. Más ade- lante, hay interacción entre los órganos de los sentidos, el sistema nervioso central y las glándulas endocrinas, al tiempo que las condiciones fisiológicas están listas para el acto reproductor. En el momento de la reproducción, muchas manifesta- ciones de conducta más o menos complejas se hacen patentes a través de señales visuales, químicas y auditivas. El mismo acto reproductor, a través de impulsos nerviosos, reajusta el estado de los tejidos endocrinos.
La dificultad para describir brevemente la complejidad de la interacción de los componentes de los sistemas nervioso y endocrino en los peces, es aumentada
306

306 INTEGRACI6N
por la enorme cantidad de especies de peces que existen y sus diversas adaptacio- nes a la gran variedad de hábitats que ocupan. Los peces, ma8 que cualquier otro grupo de vertebrados, tienen hábitats muy variados y, en consecuencia, mayores diferencias anatómicas que cualquier otro grupo de vertebrados. Existen revisio- nes extensas sobre la información, con frecuencia controvertida, sobre los siste- mas nervioso y endocrino de los peces; esas revisiones, así como artículos especí- ficos selectos, son citados al final de este capítulo o aparecen en la lista junto con las referencias generales, al final del Capítulo 1.
EL SISTEMA NERVIOSO
El modelo básico y el progreso de la cefalización, la concentración de la recepción de estimulos y las unidades integradoras en la cabeza pueden ser seguidos mediante el estudio del grupo de las lampreas-mixinos (Cyclostomata) y a través de los pe- ces en forma de tiburón (Chondrichthyes), hasta llegar a los peces óseos (Osteich- thyes). L a organización general del sistema nervioso en cada uno de estos tres grandes grupos de peces ha sido descrita en el Capítulo 3 (Figura 3.20). La anato- mía general y la localización de los órganos de percepción especial y de las glán- dulas endocrinas han sido dadas en el mismo capítulo. Debemos considerar ahora ciertos detalles de la anatomía y fisiología del sistema nervioso, los órganos de los sentidos y las glándulas endocrinas.
Las conexiones de las diversas partes del sistema nervioso central están resumidas en las Figuras 3.20 y 11.1 y en la Tabla 11.1.
El telencéfalo (área de la olfación en el cerebro; Figura 11.1; Tabla 11.1)
La región más anterior del cerebro de los peces, el telencéfalo o cerebro anterior, está dedicado principalmente a la recepción, elaboración y conducción de los impulsos olfativos. Su tamaño relativo varía de acuerdo con el grado en que la olfación juega un papel en la vida de la clase de pez examinado. El nervio olfato- rio, o nervio craneal I, penetra al cerebro a partir del epitelio sensorial de las lami- nillas olfatorias o plácoda que está ubicada en el orificio o fosa nasal. En los tibu- rones y especies afines (Elasmobranchii) y en los peces óseos (Osteichthyes) los nervios olfatorios derecho e izquierdo están acompañados por otro par de nervios, los nervios pequeiíos terminales (O). Se cree que ellos tienen funciones vasomoto- ras más que sensoriales. Aunque el órgano de la olfación de las lampreas, que es impar, ha asumido en forma secundaria una posición media, las estructuras cere- brales subyacentes son pareadas, como ocurre en los demás vertebrados. Hacia adelante y a cada lado el telencéfalo está compuesto de un bulbo olfatorio se- guido en dirección caudal por un lóbulo olfatorio (Figura 3.20); sus dos cavidades internas corresponden a los ventrículos I y 11. Los elasmobranquios, especialmente, y los peces óseos que dependen principalmente de la olfación para su alimenta- ción o de las relaciones sociales, tienen lóbulos olfatorios muy grandes. Estos lóbulos, en su lado ventrolateral, contienen un ganglio grande, el cuerpo estriado, que viene a ser un centro de correlación, principalmente para retransmitir los impulsos olfatorios a los centros nerviosos sensoriales posteriores o somáticos. El manojo más anterior de fibras nerviosas que conecta los dos lados del cerebro, es

EL SISTEMA NERVIOSO 307
Istmo ganglionar Área de transici6n
Núcleo dorsal
Núcleo olfatorio
Comisura anterior
Ventriculo I I I Comisura ansular
Núcleo anterior Saco vascular
Núcleo ventral del tálamo
Figura 11.1 - Sección diagramática del cerebro de un pez óseo con los principales tractos fibrosos, tal como se ve en una carpa (Cyprinus). Tractos nerviosos: A, olfato- rio; B, olfatohabenular; C, hipocampotalámico; O, striotalámico; €, striohipotalámico; F, preópticohabenular; G, olfatohipotalámico; U, palial; /, espinohipotalámico; J, espinotalámico; K, ópticotectal; L, tectobulbar; M, fasciculus retroflexus; N, teg- mentocerebelar; O, dienchfalocerebelar; P, espinocerebelar; O, mesenchfalocerebelar; R, lobopedúnculoespinal; S, cerebelotegmental; T, facialicerebelar; U, lateralicerebe- lar; V, vestíbulocerebelar. Los nervios craneales son: I, olfatorio; II, óptico; Ill, oculo- motor y IV, troclear. (Basado en Neal y Rand, 1939).
la comisura anterior. Esta comisura está localizada en el telencéfalo, y recorre, en parte, a través de la llamada lámina terminal, la pared anterior media de todo el sistema del canal neural.
Aunque la olfación es una función obvia, no es ella la única función que tiene el telencéfalo de los peces con espinas en las aletas (Actinopterygii), ya que se piensa que tiene una función adicional generalizada, facilitando las acti- vidades de los centros y mecanismos del cerebro inferior. El desarrollo morfo- genético del cerebro anterior de los peces con radios en las aletas es enteramente diferente del de otros vertebrados, en el sentido de que se forma mediante la eversión de las paredes laterales del área dorsal (palial), en contraposición a la evagi- nación que da lugar a los ventrículos laterales. En consecuencia, el cerebro ante- rior de los actinopterigios retiene un ventriculo ventral (subpalio) único y medio, en contraste con los ventrículos ventrales (basales) pareados de los peces inferiores y otros vertebrados.

Tabla 1 1 .I - Sistemas importantes de fibras nerviosas en el cerebro y en Ba médula espinalabs de los peces tele6stoos. t
Divisiones del sistema nervioso central
Regiones del cerebro o ¡a médula espinal conectadas por las fibras
Telendfalo Telencéfalo
Diencefalo
Diencefalo
Diencéfalo
Telendfalo
Telencdfalo
Telendfalo
Diendfalo
Diencéfalo Telencdfalo
Telencefalo
Diendfalo Diencéfalo
Mesencefalo Diendfalo
Mesendfalo Mesendfalo
Diendfalo Mesencifalo
Diendfalo Mesendfalo
Metencdfalo MetencBfalo
Mesendfalo MesencBfalo
Mielendfalo Mesendfalo
Maula espinal Mesendfalo
Mielendfalo Mesendfalo
Mbdula espinal Mielendfalo
Mesendfalo MBdula espinal
DiencBfalo Metendfalo
Mielendfalo Metendfalo
Mielendfalo Mielendfalo
Mbdula espinal Di-, Mes- y Metendfalo
Telenc6falo * Telendfalo Paredes media y lateral del lóbulo olfatorio + Núcleo
~ ~~~ ~
habenular
Area preóptica + Núcleo habenular
Area somática del cuerpo estriado + Hipotálamo
Area olfato-somática del cerebro anterior -+ Tálamo ventral
Area olfato-somáttica del cerebro anterior ++ Tálamo dorsal
Hipotálamo f, Hipocampo primordial y áreas septales precomisurales del cerebro anterior
Núcleos laterales geniculados ++ Tecto óptico
Núcleo habenular -+ Núcleo interpeduncular en la base del
Tecto óptico +> Tálamo cerebro medio
Tecto óptico + Hipotálamo
Tecto óptico + Cerebelo
Cerebelo -+ Tegmento
Núcleo del nervio V + Tecto óptico
Tecto 6ptico -+ MBdula espinal
Núcleo de la I ínea lateral de la medula +Torus semicircularis
Medula espinal +. Tegmento y valvular del núcleo en el tegmento
Lóbulos gustativos de la medula + Núcleos gustativos
Medula espinal + Tálamo secundarios del cerebro medio y tambihn del hipotálamo
Núcleo vestibular y fibras del nervio Vl l l+ Cerebelo
Núcleo de la I ínea lateral de la medula ++ Cerebelo
Núcleo de los nervios VI I a X en la mklula + MBdula
Muchas partes del cerebro +. Medula espinal del tronco y espinal
de la cola Medula espinal

c "____
Nombre del tracto Función del tracto
Comisura anterior Olfactohabenular
medio y lateral
Preóptico-habenular
Strio-hipotalámico
Pedúnculo ventral del bulto lateral del cerebro anterior
Pedúnculo dorsal del bulto
Bulto medio del cerebro lateral del cerebro anterior
anterior
Genículo-tectal tectogeniculada
Fascículo retroflex0
Tálamo-tectal-tectotalámica
Tecto-hipotalámico
Tectocerebelar
Cerebelo-tegmental
Bulbotectal
Tecto-espina1
Lemnisco acústico-lateral
Espino-tegmental (probablemente multisináptico)
Sistemas ascendentes y gustativos secundario y terciario
Espino-talámico
(rama del colícolo superior)
(habbnulo-interpeduncular)
Vestíbulocerebelar y fibras
Láterocerebelar
Vestibuloespinal
nerviosas directas vestibulares
Cerebelo-lateral
Glulas gigantes de Mauthner
Conecta los dos lados del cerebro anterior Proyecta los impulsos de la olfación al núcleo habenular
como estaciones de paso para los reflejos de la alimentación
alimentación
hipotalámicos
las respuestas motoras
Tracto olfatorio terciario, una ruta refleja de la
Conduce los impulsos del cerebro anterior a los centros
Principio evolutivo, por el cerebro anterior, del control de
Ruta primitiva de descarga entre el tálamo y los ganglios
Conduce las sensaciones viscerales, incluyendo la basales del cerebro anterior
gustación, hacia y de la región olfatoria del cerebro anterior
Retransmite los impulsos ópticos
Parte de las rutas de descarga de las áreas olfativo-somáticas a los centros motores (reflejos de la alimentación)
Relaciona el tecto óptico muy estrechamente con el tálamo dorsal en desarrollo
Conecta los impulsos de la visión con los centros autónomos y con la región pituitaria
Relaciona la visión y otros impulsos sensoriales con el tono muscular para la locomoción
Porción del sistema de retransmisión que va a los centros motores, regulando el tono muscular
Coordina los órganos del sentido del tacto de la cabeza con la visión
Coordina la visión y otros impulsos sensoriales con la locomoción
Proyecta los impulsos de la I ínea lateral al cerebro medio
Controla la locomoción
Coordina los estímulos gustativos y olfatorios para la
Dirige las sensaciones generales del cuerpo al tálamo ( s e
Parte del sistema del equilibrio
alimentación, especialmente en los bagres (Siluroidei)
ha dudado de la presencia de este sistema en los peces)
Coordina los impulsos locomotores y los de la línea lateral
Retransmite los impulsos vestibulares y los de la línea lateral a los centros inferiores medulares y espinales
Coordina el tronco y la agitacibn de la cola durante la natacibn

310 INTEGRACIdN
El telencéfalo de los peces está extensamente inervado por fibras olfatorias secundarias del bulbo olfatorio. Estas fibras se proyectan a través de la porción subpalial de cada hemisferio y también se proyectan a las áreas basolaterales del palio. El resto del palio del telencéfalo de los actinopterigios parece que no recibe fibras olfatorias secundarias. Hay un incremento en la diferenciación y en la sub- división del área dorsal a través de la serie filogenética, de los Polypteriformes a los Teleostei.
Mediante la ablación del cerebro anterior se ha descubierto que está rela- cionado con otras actividades, además de la función olfatoria. La carpa dorada (Carassius auratus) a la que se le ha extirpado el cerebro anterior, muestra menos iniciativa y expontaneidad, al explorar su medio ambiente, que los animales in- tactos. Ciertas tilapias (Tilapia) desatienden a sus crías cuando su cerebro anterior es extirpado, y ciertos pececillos (Cyprinidae) que sufren esta operación son menos cautelosos y recelosos de una nueva situación que sus compañeros norma- les. Las ablaciones del cerebro anterior resultan en una supresión de la conducta agresiva, sexual y paternal en la espinocha de tres espinas (Gasterosteus aculeatus). Ellas suprimen también en el pez paraíso (Macropodus opercularis) la conducta de reproducción y de construcción de nidos. La extirpación del cerebro anterior en la carpa dorada da lugar a una supresión selectiva de la discriminación en la visión de los colores.
El diencéfalo (área I de la visión; Figura 11.1; Tabla 11.1)
Una constricción dorsoventral, y a menudo también una lateral pronunciada, marca la transición entre el telencéfalo del pez y el diencéfalo o cerebro intermedio. Ha- cia la porción dorsal, el parencéfalo, o el saco delgado dorsal, forma el techo de la cavidad dentro del diencéfalo, también conocido como tercer ventriculo. En algunos peces como las lampreas (Lampetra), catanes (Lepisosteus) y en el pez de aleta dorsal ondulada (Amia), el saco dorsal está agrandado y se extiende
aEl sistema nervioso central de la lamprea posee un cerebro anterior olfatorio de gran tamaño con orígenes sencilios de centros no olfatorios, un diencefalo dorsal sensible a la luz y una epífisis que, comparado con el de los peces óseos, tiene centros tactiles y gustativos menos desarrollados. Hay conexiones prominentes de los centros inferiores (medulares) al tecto óptico, que deben tener otras funciones además de las visuales.
Correspondiendo a las c6lulas de Mauthner de los peces óseos, existen en las lampreas los manojos de Mueller con cuerpos celulares en la medula y fibras eferentes que van a la musculatura del cuerpo. Tanto las c6iulas de Mauthner como las de Mueller se encuentran en la medula espinal de las lampreas. Las dendritas vienen de diferentes centros del cerebra superior y e l sistema entero coordina la entrada sensorial con los movimientos sinuosos de !a natación de los animales.
bLos cerebros y las medulas espinales de los elasmobranquios, siendo los tiburones mejor conocidos que las rayas en este aspecto, tienen un área olfatoria (cerebro anterior) más pronunciada que los de muchos peces óseos. En correspondencia, hay muchos tractos fibrosos prominentes que llevan hacia y traen de esta área los impulsos.
Las rayas nadan mediante la ondulación de sus enormes aletas pectorales en farma de ala; en las regiones correspondientes, no está la medula espinal bien diferenciada para permitir la producción de reflejos multi- segmentarios, pues las conexiones con la medula han aumentado tambien en importancia, comparada con la de los tiburones de forma más alargada, cuyas aletas pectorales funcionan solamente como estabilizadores. CDeseamos dar las gracias a la Dra. Elizabeth C. Crosby, quien nos ayudó en la preparación de esta tabla. dLas fibras nerviosas que siguen sólo un camino se indican con la flecha: -+. Las fibras ascendentes y des- cendentes. con la flecha: *.

EL SISTEMA NERVIOSO 311
lateral y frontalmente, inclusive sobre el telencéfalo. Hay dos sitios destacados a lo largo de la línea media del diencéfalo en las lampreas (Petromyzonidae): los órganos parapineal y pineal, también conocidos como órganos epifisiarios, loca- lizados ligeramente hacia la izquierda y hacia la derecha de la línea media, res- pectivamente. Sin embargo, en los tiburones adultos y en especies afines (Elas- mobranchii), y en peces óseos superiores (Actinopterygii), solamente se desarrolla el órgano pineal. Los embriones de algunos peces óseos, como los peces blancos (Coregonus), muestran ambos órganos durante las primeras etapas del desarrollo embrionario, pero el órgano parapineal se pierde poco después, persistiendo sola- mente el pineal. Ya sea que las dos o sólo una de estas evaginaciones persista, su estructura y función ha sido conectada a la recepción de un estímulo luminoso más o menos difuso.
En las lampreas y en los mixinos (Cyclostomata), el órgano pineal está conectado al ganglio habenular derecho (véase abajo), tiene una retina, células pigmentadas y una estructura en forma de lente. Hay un orificio en el techo del cráneo, arriba de é1, precisamente por detrás de la abertura nasal, y una mancha depigmentada en la piel, de tal manera que el estímulo luminoso puede llegar a los órganos pineal y parapineal. Ciertos tiburones (Squaliformes) tienen una man- cha, o sea, una pequeña zona sin pigmentar sobre la piel de esta región de la cabeza y algunos de ellos poseen también una perforación del techo cartilaginoso del cráneo. Sin embargo, hay generalmente en los tiburones un órgano pineal menos desarrollado que entre los ciclóstomos y algunos peces óseos. Zonas pineales par- ticularmente bien desarrolladas se encuentran en los peces de profundidades me- dias cuyas migraciones se realizan en sentido vertical, como sucede con los peces hacha (Argyropelecus), muchos bagres (como Arius y Macrones) y también en los aguja (Hemiramphus).
A pesar de los estudios tan numerosos que se han hecho sobre el cuerpo pineal de los peces, no se ha podido demostrar, sin caer en equivocaciones, que tenga otra función. La hipótesis actual puede ser dividida en dos categorías: una que pone énfasis primordialmente en el papel sensorial y la otra en una función secretora. La siguiente hipótesis se relaciona con la sensorialidad: el cuerpo pineal es una estructura sensorial; actúa como un baro- o quimiorreceptor para el fluido cerebroespinal; o actúa como un mediador en la respuesta olfatoria a las exohormo- nas. En relación a las funciones secretoras, la glándula pineal puede ser primaria- mente una glándula de secreción externa relacionada con la composición química del fluido cerebroespinal o el metabolismo del tejido cerebral o, alternativamente, el cuerpo pineal puede ser una glándula de secreción interna con una función endocrina.
El resto del diencéfalo puede ser dividido en una región epitalámica con su ganglio habenular, un tálamo, y un hipotálamo como su más grande e importante componente. Los nervios ópticos penetran al cerebro y cruzan hacia adelante hasta el diencéfalo, y esta región del cerebro se une posteriormente a la porción ventral del mesencéfalo a través del saco vascular de paredes delgadas y con cir” cunvoluciones. Debajo del hipotálamo se adosa la hipófisis o glándula pituitaria (Figura 11.1). Esta glándula está estrechamente unida a la base del cerebro en las lampreas, pero entre los tiburones y en los peces óseos superiores es sostenida por un pedúnculo o infundíbulo (Figura 11.8). El tálamo sirve como un centro de relevo para la transmisión de los impulsos olfatorios y los del cuerpo estriado a los tractos tálamo-medular y tálamo-espinal. En su región ventral, hacia la base del

cerebro y del hipotilarno, se apoya e¡ ntlcleo genlculado o ganglio (Figura Il.1) que recibe algunas conexiones del nervio Gpt,icu (11) antes de que sea alcanzado til tecto bptico por los axones de este nervio craneal. Estos nccleos están bien desarrollados en 10s tiburones, melusive hasta el grado de que justificadamente pueden ser llamados í6bulos geniculados; sin embargo, nada cierto se sabe acercz de su funcih. En el quiasma óptico de todos los peces las fibras nerviosas de la retina de un ojo cruzan el tecto óptico para ir a dar al otro lado del mesencéfalo. pasando la intersección o cruce de las fibras de un ojo por sobre e inclusive 2 través del manojo de fibras del otro ojo.
Parece ser que el diencéfalo de los peces es un importante centro de corre- lación para la llegada y la salida de mensajes relacionados con la homeostasis interna. El hipotálamo afecta especialmente al sistema endocrino a través de la glándula pituitaria. Células neurosecretoras han sido descritas en algunos núcleos diencefálicos y han sido encontradas vías especiales que llevan las secreciones del diencéfalo a la hipófisis.
El mesencéfalo (área II de la visión; Figura 11.1 ; Tabla 11.1 )
El mesencéfalo o cerebro medio de los peces es relativamente grande; consta del tecto óptico dorsal -que aparece a la visión dorsal como los dos lóbulos Ópticos y del tegmento ventral. El tecto esta formado de un cierto número de zonas de células nerviosas de diferentes tamaños o neuronas. La mayoría de las fibras del nervio óptico van a terminar en el tecto, de tal manera que la porción fron- tal de la retina se proyecta en la porción caudal contralateral del tecto, en el otro lado del pez. LOS peces, al igual que los demás vertebrados, tienen en sus ojos lentes convexos que forman una imagen invertida en la retina; a través de la con- figuración tectal ya descrita la imagen puede proyectarse tal como es en la natu- raleza. En las lampreas (Petromyzonidae) el cerebro medio está cubierto con un plexo coroideo vascular que ayuda en la nutrición del cerebro y comunica con los ventrículos llenos de fluido y con el canal central. Entre los peces óseos supe- riores (Actinopterygii), la materia gris tectal se extiende hacia el ventriculo como un torus longitudinalis pareado y se une a la comisura posterior entre los dos hemisferios que descansan en el borde del diencéfalo. El torus está presumible- mente relacionado con la visión desde el momento en que es muy pequeño en un pececillo ciego (Trypauchen).
La estimulación eléctrica y la extirpación de porciones del cerebro medio indican que el tecto contiene unidades neuronales funcionales que correlacionan las impresiones visuales con las respuestas musculares, como podría ocurrir, por ejemplo, al mirar la presa o al dirigir la natación hacia objetos que se mueven den- tro del campo visual; en pocas palabras, con varios movimientos complejos desti- nados a lograr ciertos objetivos. La carpa “crucian” (Carussius carussius) no pudo distinguir las posiciones de la luz (orientación espacial) después de que los hemis- ferios tectales fueron separados mediante una incisión longitudinal media. Esta carpa también mostró dificultades para localizar la fuente del estímulo sonoro. Los experimentos realizados con un pez globo (Sphoeroides) en el cual un ojo fue desviado de adelante hacia atrás y el otro fue cegado, lo llevaron a realizar mOVi- mientos forzados, circulares y persistentes, aún cuando otras partes del cerebro, a excepción de la médula, habían sido removidas. Por lo tanto, hay fuerte evidencia

EL SISTEMA NERVIOSO 313
de que el tecto óptico es un centro coordinador del ojo-cuerpo. La estimulación eléctrica del tegmento, sin embargo, conduce a la obtención de respuestas loco- motoras generalizadas, pero relativamente descontroladas. El tecto óptico también realiza alguna función relacionada con el aprendizaje. Las respuestas al estímulo olfativo han sido asociadas con las señales visuales; después de la extirpación del cerebro medio desaparecen las respuestas a la olfación.
En vista del cruzamiento total de las fibras nerviosas ópticas es evidente que la carpa dorada (Carassius auratus) y el “goby” (Buthygobius) podrían ser entrenados para realizar una transferencia sensorial óptica. Las respuestas somá- ticas y autónomas mostraron que el ojo sin entrenar reconoció al estímulo apren- dido con el otro ojo, pero que las respuestas motoras a la transferencia fueron di- fíciles de obtener.
El tecto óptico ha sido comparado muy a la ligera con la corteza cerebral de los mamíferos debido a su importancia funcional en el sistema nervioso cen- tral de muchos peces y a sus extensas conexiones, y debido también a las nume- rosas capas de células nerviosas que contiene.
El metencéfalo (cerebelo, equilibrio, tono muscular; Figura 11.1; Tabla 11.1)
El cerebelo se desarrolla en el metencéfalo a partir del aumento de volumen de la médula subyacente, como un crecimiento externo dorsal del borde superior del &o. ventriculo del cerebro. Se ha reportado que sus principales funciones se relacionan con el equilibrio durante la natación, el mantenimiento del tono mus- cular y la orientación en el espacio; es el mayor componente del cerebro de algu- nos peces. En el cerebelo pueden distinguirse muchas capas diferentes formadas por células nerviosas de diferentes formas, que incluyen a la capa celular receptora o molecular y a las células llamadas de Purkinje, que son de gran importancia en el cerebelo de los vertebrados superiores.
El cerebelo forma solamente un puente dorsal pequeño en las lampreas (Petromyzonidae), ubicado entre las áreas nerviosas de la línea acústica lateral. El cerebelo está representado en los mixinos (Myxiniformes) por una comisura pequeña que consta de ocho fibras nerviosas y de la línea lateral, o sea, el sistema octavo-lateral. En los tiburones y en las rayas (Elasmobranchii) el cerebelo aumen- ta bruscamente con el tamaño de las especies. Tiburones relativamente pequeños, como los peces perro (Squalidae), tienen un cerebelo simple bilobulado, mientras que en el grande, rápido y predador, como el tiburón macarela (Lamnidae), el cerebelo se ha transformado en la parte de mayor tamaño del cerebro, presenta circunvoluciones y tiene un área superficial muy ampliada (Figura 11.2). Pueden formarse también en el cerebelo crecimientos prominentes laterales o auriculares y que están conectados principalmente a la línea lateral y a los órganos sensoria- les vestibulares.
El cerebelo adquiere en los peces óseos superiores (Actinopterygii) una característica proyección hacia adelante, la válvula cerebelosa (Figura 11.3), que se extiende por debajo del tecto óptico. En los peces elefante (Mormyridae), muchos de los cuales tienen trompas alargadas y muy movibles, las válvulas cere- belosas se enarcan mucho más allá del cerebro anterior y se transforman en el mayor componente del cerebro; entre los silClridos (Siluroidei), la estructura está tam- bién muy desarrollada. Desde el momento en que los mormíridos emiten, y re-

314 INTEGRACION
accionan a, corrientes eléctricas muy débiles y desde el momento en que existe también por lo menos un bagre que produce descargas eléctricas (el bagre eléc- trico, Malapterurus), se ha sugerido que el cerebelo de gran tamaño en estos grupos tiene que ver con los impulsos eléctricos.
El ventriculo del cerebelo, aún prominente en los tiburones y en rayas (Elasmobranchii), ha desaparecido casi por completo en el cerebelo de los peces
CEREBROS DE TIBURÓN
Comisuras transversas
T e a o óptico
rdón c Hacia el hocico espina1
(a)
Comisuras transversas
Cerebelo A
Cordón espina1
""
Tecto óptico
transversas
Figura 11.2 - Tamaño relativo y desa- rrollo del cerebro entre los tiburones como se ve en secciones diagramáticas: (a) un tiburón gato (Pristiurus); (6) un pez perro liso (Mustelus); (c) un ti- burón de arena (Carcharias). (Fuente: Voorhoeve en Grad, 1958).
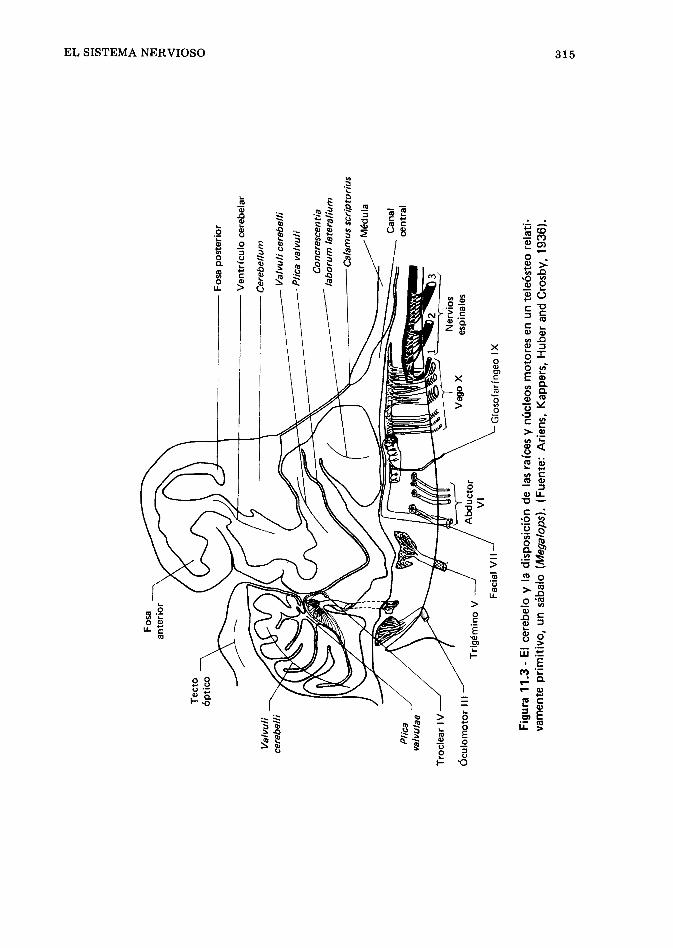
EL SISTEMA NERVIOSO 315

iiseos superiores, como ha sucedido tambikn con las “sulci”, cs pliegues transver- sales, tan prominentes en algunos tiburones grandes. Las auriculas laterales y las fibras interauriculares aumentan en importancia., especialmente en h s peces que tienen cjrganos de la línea lateral bien desarrollados.
IJna función aparentemente importante del cerebelo corlsiste en la coordi- nación del tono muscular y l a natación.
El mielencefalo (área bisica del cerebro, tallo cerebral y médula; Figura 11.1; Tabla 11.1)
Las divisiones cerebrales desde el cerebelo al telencéfalo son, en su más amplio sentido, centros sensoriales y centros coordinadores de las sensaciones que se co- nectan con el tallo cerebral, o mielencéfalo, mediante varios tractos neurales. Estas cuatro divisiones situadas en la porción más anterior han aumentado en importancia como integradores sensoriales y se han vuelto más y más perfeccio- nados en el curso de la evolución. El mielencéfalo, con la médula oblongada como su componente más importante, es el centro a donde llegan los nervios sen- soriales, a excepción de los del olfato (I) y la visión (11). La distinción entre la medula oblongata y la médula espina1 de los peces es imprecisa. La m6dula puede estar dividida en columnas de fibras nerviosas basadas en el tipo de información transmitida. Por lo tanto, hay columnas sensoriales somáticas y viscerales y tam- bién columnas motoras somáticas y viscerales.
En el tallo cerebral, los núcleos de los nervios craneales 111 a X están dis- puestos anteroposteriormente (Figura 11.1); los tractos de los nervios I11 y IV cruzan hacia el lado contralateral. Los componentes de los nervios aferentes del tallo cerebral pueden estar divididos en nervios sensoriales somáticos y especiales que entran con el nervio craneal VI1 (facial), VI11 (estato-acústico), IX (gloso- faríngeo) y X (vago), la mayoría sensoriales. Hay más fibras sensoriales somáticas de los nervios precedentes en las lampreas (Petromyzonidae) que en los peces como los tiburones (Elasmobranchii). Las fibras sensoriales somáticas se reducen más adelante en los peces óseos (Osteichthyes). Los detalles sobre la distribución de los nervios craneales que están, en su mayor parte, conectados al tallo cerebral, pueden ser encontrados en la Tabla 11.2.
Diferentes porciones del tallo cerebral están agrandadas, dependiendo de la importancia relativa de los diferentes sentidos. Peces como la sardina del Pacífico (Clupea pullasi), la lisa (Mugil cephalus) y ciertos carángidos como Truchurus tie- nen protuberancias prominentes pareadas, en el borde anterolateral del cuarto ventriculo, designadas cristae cerebelli. Las conexiones nerviosas del cristae no son conocidas, pero su función puede ser relacionada con la fuerte tendencia de estas especies a agruparse. Ciertos peces chupadores, como Curpiodes ueEifer o carpa chupadora de aleta levantada, y pececillos como la carpa dorada (Carussius aurutus), tiene lóbulos vagales prominentes posteriores a la región de los cristae cerebelli de donde emergen los nervios craneales IX y X. Esos peces también han desarrollado un órgano en el techo de la boca (el órgano palatal) donde es pro- bado el alimento mediante los sentidos del gusto y el tacto. Otra característica importante que se encuentra entre pececillos (Cypriniformes) es el tubérculo im- par, o lóbulo facial, una pequeña giba central posterior al cerebelo y algunas ve- ces parcialmente cubierto por él. El tubérculo impar se origina de una fusión de

LA MfiDULA ESPINAL 317
elementos radicales del nervio facial (VII) y del nervio vagal (X), donde los impulsos gustativo y tactil están correlacionados con los sensoriales de las vísceras, como se observa en un pez europeo (Barbus fluuiatilis) y en la carpa común (Cyprinus carpio; Figura 11.4).
La médula de los peces óseos superiores (Actinopterygii) contiene un par de neuronas grandes. Las células gigantes de Mauthner, al nivel del nervio craneal VIII. Las dendritas laterales de estas células gigantes se conectan con las fibras de los nervios craneaies V, VII, IX y X, al cerebelo y al tecto óptico. Sus axones pasan a través de la médula espinal hacia los músculos de la cola y están mejor desarrollados entre los peces que son buenos nadadores, como el salmón del Atlán- tico y ciertas truchas (Salmo), mientras que son menos prominentes en los peces que habitan los fondos, como en algunos gobios (Gobius) y peces escorpión (Scor- paena). En los anguilas (Anguilliformes) y en los molas (Moja), las células de Mauthner no aparecen. Aparentemente, las células son coordinadoras motoras para retransmitir impulsos sensoriales múltiples, principalmente de los centros de la línea lateral, hacia la musculatura caudal y abdominal destinada a la natación. Un sistema coordinador comparable conecta en las lampreas (Petromyzonidae) los niveles oculomotores del cerebro con la musculatura corporal y la de la cola a través de las neuronas llamadas de Mueller y también de las células de Mau- thner.
Además de ser un área de relevo entre la médula espinal y el cerebro, la médula tiene también centros que controlan ciertas funciones somáticas y visce- rales. Éstas incluyen, entre los peces óseos, los centros respiratorios, los de la palidez (actuando a través de la agregación de melanóforos) y los osmorregula- dores. Los núcleos medulares también están relacionados para el mantenimiento del equilibrio durante la natación. Las pruebas que se han realizado sobre el control medular del movimiento de las aletas en algunos peces que las emplean principalmente para la locomoción, como los peces señorita o viejita (Labridae), muestran que el ritmo de agitación de las aletas es dirigido por un centro que se localiza en la cuerda anterior o tallo cerebral.
LA MfiDULA ESPINAL (FIGURA 11.1; TABLA 11.1)
El sistema nervioso central de los peces pierde mucho de su complejidad estruc- tural en la transición que hay del cerebro a la médula espinal. Esto ocurre ya en las lampreas (Petromyzonidae), pero en grado mucho mayor en los tiburones y en las rayas (Elasmobranchii) ya que, al realizarse un corte transversal en la mé- dula (Figura 11.5) se observa una región central de sustancia gris que consta pri- mordialmente de células nerviosas, y un área exterior de sustancia blanca, las fibras nerviosas. Estas fibras están envainadas, o mielinizadas, y forman manojos de acuerdo a su función y conexiones. En el centro de la materia gris de la mé- dula espinal corre el canal central. Como sucede en los demás vertebrados, la materia gris de la médula espinal de los peces óseos (Osteichthyes) recuerda a primera vista una letra X, con cuernos pares dorsales y ventrales. Las astas dorsa- les reciben fibras sensoriales somáticas y viscerales mientras que las astas ventrales contienen núcleos motores, con centros para las musculaturas dorsal o ventral, adosándose en posiciones media y lateral, respectivamente.

Tabla 11.2 - Nervios craneales de los peces y sus funciones "t
Nervio Número
Nombre Conexión periferal
O Terminal I Olfatorio
II dptico I!! Oculomotor
IV Troclear
V I
VI1
V Trigdmino dividido en ramas oftálmica profunda, maxilar y mandibular
Abductor
V l l l
IX
X
Faciala Y
Acústicoa
G losofar íngeo
a menudo fusionado con el
Vago
Eulfo olfatorio Cdlulas mitrales; bulbo olfatorio; núcleo
Retina del ojo Músculos del ojo: superior, inferior, anterior,
Músculo superior oblicuo del ojo
olfatorio anterior
recto y oblicuo inferior
Porción anterior de la cabeza, regiones mandibulares superior e inferior
Músculo recto posterior
1.
2.
3.
4.
1.
2.
1.
2.
3.
4.
5.
Grupo dorsal: canal de la línea lateral postorbital; botones gustativos en el cuerpo, donde estdn presentes Grupo orbital: canales de la línea lateral supra e infra orbitales; botones gustati- vos en el hocico Grupo acústico: oído interno; canales de la I ínea lateral temporal Grupo branquial: canales de la línea op6rculo-mandibular, oral y yugal; sen- tido del gusto dentro de la boca; sentidos del gusto y tacto de las regiones opercular hioidea y mandibular; músculos de la cabeza antagónicos con los que están bajo el control de la rama mandibular del V Grupo dorsal: parte del canal de la línea lateral temporal; sensibilidad y propio- cepción de la piel dorsolateral, región de la primera abertura branquial Grupo branquial: órganos de la gustación en, la mucosa faríngica dorsal y ventral, músculos de la primera abertura branquial (tercer arco branquial) Rama supratemporal: piel y órganos de la linea lateral de la región supratemporal Rama recurrente dorsal: se une con la inervación de los botones gustativos del cuerpo, del V I I Rama de la línea lateral del cuerpo: de los órganos de la línea lateral del cuerpo Rama visceral: nervios que van hacia y salen de los órganos internos Ramas branquiales: van hacia o salen de las cuatro aberturas branquiales poste- riores o regiones equivalentes
aNervios craneales VI1 y VIII, considerados mejor juntos como nervio acústico-facial, dividido en grupos nerviosos dorsal, orbital, acústico y branquial.

Función
Cerebro anterior; lámina terminal Centros de la olfación del cerebro
Tecto óptico del cerebro medio Tallo cerebral anterior bajo el tecto
anterior
óptico (Mesencdfalo)
Columna lateral de la región de la md- dula oblongata del tallo cerebral, frecuentemente se entrecruza con los centros del VI1 (Mielendfalo)
Tallo cerebral ventral detrás del ori- gen del V
Serie alargada de núcleos conectados hacia la mitad del tallo cerebral (mé- dula oblongata; mielencéfalo)
Sensorial somática: ¿punta del hocico? ¿vasomotor? Sensorial especial, conduce los impulsos de la ol-
Sensorial especial, conduce los impulsos visuales Motor somático, inerva cuatro de los seis músculos
estriados del ojo y los músculos dentro del ojo
fación
Funciones sensoriales somáticas y motoras com- binadas. Sensibilidad termal y táctil de la piel en la porción anterior de la cabeza; propioceptores de la musculatura cefálica
lnerva seis músculos estriados que mueven al globo del ojo
Un nervio realmente combinado; funciones senso- riales viscerales y motoras especiales y somáticas. Trae a los centros cerebrales los impulsos acús- ticos de la línea lateral y gravitatorios, así como los impulsos de la gustación y del tacto de la cabeza y la boca; propiocepción e inervación de algunos músculos cefálicos
Series de ganglios, o raíces, principal- Un nervio combinado; sentidos del gusto y de la mente en la región terminal dorsal y línea lateral en componentes aferentes; inerva media del tallo cerebral (mddula los músculos branquiales anteriores (primera oblongata; mielencéfalo) abertura branquial)
Series de ganglios posterior a, pero Un nervio combinado; control motor del paquete unidas con, las raíces del nervio IX; de músculos branquiales, fibras especiales de los a menudo se entrecruza con las raíces órganos de la línea lateral y del gusto, motor de los primeros nervios occipitoespi- visceral y sensorial hacia y desde los órganos in- nales hacia el extremo posterior del ternos; sistema nervioso parasimpático autónomo tallo cerebral (mMdula oblongata, mielencdfalo)

3 20 INTEGRACION
ióbulo ómico
I . Nervio olfatorio
Lóbulo olfatorio
I Cerebelo
de la rn6dula
Figura 11.4 - Vista dorsal del cerebro de la carpa (Cyprinus car- p i ~ ) mostrando el desarrollo de los lóbulos facial y vago. (Basado en Uchihashi, 1953).
LOS NERVIOS ESPINALES
Los nervios espinales están dispuestos segmentariamente. Las raíces ventrales aparecen en las lampreas (Petromyzonidae) entre los niveles de las raíces dorsa- les, de tal manera que este grupo de peces no tiene nervios espinales realmente combinados; solamente entre los mixinos (Myxinidae) comienzan a aparecer los nervios espinales realmente mezclados como los que están presentes, con unas pocas excepciones, a lo largo de la serie de peces mandibulados, los tiburones y rayas (Chondrichthyes) y los peces 6seos (Osteichthyes). Los peces mandibula- dos tienen por lo general ganglios espinales para las neuronas de las raíces ner- viosas sensoriales o dorsales, aunque en muchas familias, como en los pececillos de acuario (Cyprinidae), bacalaos (Gadidae), percas (Percidae) y peces tambor (Sciaenidae), algunas fibras aferentes vienen también a la médula desde los gan- glios supramedular e inframedular.
A partir de los tiburones (Squaliformes) y hasta los peces óseos (Osteich- thyes) hay una diferenciación progresiva, positiva y mucho mejor, y una sub- división de tractos fibrosos ascendentes y descendentes entre el cerebro y la médula espinal. En los peces soldadito o angelito (Triglidae), los radios anterio- res largos y separados de las aletas pectorales llevan receptores especiales para los indicios táctiles y químicos; los nervios sensoriales de estos radios están marca- dos en la médula espinal por protuberancias separadas que corresponden a las raíces de sus nervios dorsales, y sus proyecciones también alcanzan a la médula y al cerebro medio.
Los reflejos ipsilaterales y contralaterales de los nervios espinales en un segmento de la médula pueden estar presentes en los elasmobranquios; los refle- jos plurisegmentales son comunes en los tiburones y en los teleósteos. En vista del papel tan importante que desempeña el tronco en la locomoción del pez, ha recibido considerable importancia el estudio sobre el control nervioso de los movimientos de la natación. Hay controversia sobre si un pez requiere de estimu- lación externa para hacer movimientos coordinados de natación o si la natación es debida a la actividad automática de la médula espinal y los centros medulares, para ser coordinada solamente por estímulos periféricos. Von Holst, después de

EL SISTEMA NERVIOSO AUT6NOMO 321
Fibras de la Tracto raíz dorsal del fibroso nervio espinal dorsal
Tractos \
Tracto / I I raíz ventral del fibroso ”-’ ventral Canal
central ventral
.~
nervio espinal
Figura 11.5 - Sección transversal del cordón espinal del grupo de los tiburones (Elasmobranchii). (Basado de Keenan en Grass& 1958).
haber realizado numerosos experimentos con tiburones perro (Squalidae), angui- las (Anguilla), cobítidos (Cobitidae) y pececillos de acuario (Cyprinidae), ha sugerido que la natación se debe al ritmo endógeno de la médula posterior, que pasa enseguida al cordón espinal, pero influido en todos los niveles por los estí- mulos exteriores del medio ambiente. Lissmann y Gray, basándose principalmente en los experimentos realizados con ciertos tiburones gato (Scyliorhinidae), opinan que no existe ningún centro automático de la natación, sino que los movimientos natatorios son de naturaleza refleja y dependen de la coordinación de impulsos d e - rentes de la médula espinal. Los movimientos de las jóvenes larvas ammocoetes de Lumpetru (una lamprea) son interpretados como enteramente reflexogénicos.
EL SISTEMA NERVIOSO AUTdNOMO
En el sistema nervioso autónomo de las lampreas y de los mixinos (Cyclostomata) no hay cadena de ganglios simpáticos. Sin embargo, se han descrito en estos peces un sistema difuso de nervios y algunos plexos de función simpática. En las lam- preas y los mixinos, tanto el corazón como los órganos de la digestión son inerva- dos por el vago. Los plexos nerviosos subcutáneos que son inervados por fibras nerviosas espinales, aparentemente por el sistema simpático, se encuentran con-

322 INTEGRACION
centrados en las paredes y en los orificios de las glándulas mucosas de Myxine (mixino). Los ganglios simpáticos se sitúan, en los tiburones y en las rayas (Elas- mobranchii), a los lados y a todo lo largo de la columna vertebral; son irregulares y no se extienden hasta la región cefálica. Los músculos lisos del tubo digestivo y las paredes arteriales son inervados por ellos. Sin embargo, no hay conexión entre ellos y los nervios que inervan la piel, como sucede con las ramificaciones que están mezcladas y se comunican en los peces (Osteichthyes) y en los vertebrados superiores. En todos los peces parecidos a los tiburones los corazones y los órga- nos internos de la digestión son abundantemente inervados por fibras vagales. En los peces óseos, los ganglios dispuestos segmentariamente forman una cadena simpática que llega hacia adelante hasta donde se encuentra el nervio trigémino (nervio craneal V); la mayoría de las fibras preganglionares para los ganglios sim- páticos de la cabeza nacen del cordón espinal de la región del tronco y siguen hacia adelante. Las conexiones simpáticas de los nervios de la piel también se encuentran en los peces óseos superiores más evolucionados (Teleostei).
El sistema parasimpático está casi exclusivamente representado por porcio- nes de las amplias ramificaciones del vago (nervio craneal X). Una inervación pa- rasimpática de algunas partes del ojo (en el nervio craneal 111) ha sido ya mencio- nada. Un sistema parasimpático sacro similar al de los mamíferos está insinuado en las mantarrayas, por la respuesta de los “claspers” u órganos para la cópula a la estimulación de los nervios espinales posteriores. Las fibras simpáticas ace- leradoras del tejido muscular cardíaco de los vertebrados superiores faltan en todos los peces y, en consecuencia, la estimulación vagal tiene una acción podero- samente inhibidora en el seno venoso y en la auricula, pero no así en el ventriculo. La estimulación de las ramas del vago que van hacia otros órganos internos, así como al control de las drogas colinérgica (parasimpatomimética) y adrenérgica (simpatomimética), ha producido resultados conflictivos, especialmente en la acción de la vejiga gaseosa y en ciertas porciones del canal alimenticio.
Los nervios del centro de la palidez corren en el cordón espinal y lo van dejando en varios niveles en diferentes peces (en el “minnow” Phoxinus, los segmentos 1 2 a 18; en el salmón-trucha del género Salmo, segmento 26; y en los peces gallo, Trigla, segmento 3). Los nervios melanóforos van, entonces, a la cadena simpática subvertebral donde se dividen en un manojo anterior que se dirige a las células del color de la cabeza y la nuca, y un manojo posterior que va hacia el tronco y la cola. Poco es lo que se sabe acerca del control melanofó- rico en los ciclóstomos; sin embargo, ciertos elasmobranquios tienen por lo me- nos algunos melanóforos inervados por el simpático. Se ha llegado a postular la presencia de un centro oscurecedor en el cerebro medio y de los nervios para- simpáticos dispersores de la melanina.
TEJIDOS DE SOSTBN DEL SISTEMA NERVIOSO CENTRAL
Algunas derivaciones del tejido embrionario ectodérmico y algunos tejidos meso- dérmicos en la región del cerebro y del cordón espinal asumen las funciones de soporte, revestimiento y nutrición de las células nerviosas propiamente dichas. Las células ectodérmicas del epéndimo cubren los canales cerebroespinales de todos los peces, vuelven permeable el cuerpo del cordón y el cerebro, y se con- centran especialmente a lo largo de las paredes ventriculares del tálamo de los

INTELIGENCIA Y COMPORTAMIENTO 323
peces óseos (Osteichthyes), donde están provistas de cilios. La histología de es- tos conjuntos celulares sugiere que tienen función secretora. Las células de neuro- glia de forma y función especiales han sido observadas en la médula espinal de la lamprea marina (Petrornyzon). En las cuerdas espinales de los tiburones y de las rayas (Elasmobranchii) aparecen muchas células de neuroglia ramificadas, asteroideas o aracnoideas. Varios elementos neurogliales también están diferen- ciados en el cerebelo y en el cordón espinal de los peces óseos.
El plexo coroideo, una área cerebral altamente vascularizada, especialmente prominente en los catanes y en especies relacionadas (Holostei) y en los elasmo- branquios, consta principalmente de tejido mesodérmico a menudo dispuesto en varias capas y, por lo tanto, denominado leptomeninge. El plexo coroideo o tela coroidea está lleno de fluido cerebroespinal, está ricamente irrigado por vasos sanguíneos y participa en la nutrición del tejido nervioso, al que rodea y cubre.
INTELIGENCIA Y COMPORTAMIENTO
LOS componentes sensoriales del sistema nervioso del pez son muy discriminativos y la sensibilidad de las células sensoriales es a menudo sorpresivamente grande como, por ejemplo, en la olfación, en la visión y en la percepción de la tempera- tura. La mayoría de los mecanismos sensoriales trabajan mediante la adición, y la eficiencia total al nivel normal de estimulación puede ser lograda solamente con un umbral más bajo que ese con el cual normalmente sería recibida. Esos umbrales bajos implican la existencia de procesos centrales que permiten al animal ignorar al azar (“white noiseyy*) en la mayoría o en todas las modalidades sensoriales, pero ello también implica que el animal puede también hacer selecciones con un grado elevado de exactitud y certeza, logrando el éxito en la meta que se pro- puso alcanzar.
Muchos acontecimientos de discriminación o selección en la naturaleza están basados en el condicionamiento del pez a uno o a muchos estímulos e, inclusive, sin hacer experimentos, no podría dudarse de que los peces pueden ser fácilmente condicionados. La capacidad para asociar el estímulo y la respuesta está presente inclusive en los invertebrados más simples; pero los niveles relativos de “inteligencia” pueden ser distinguidos, entre otros criterios, en base a la rapidez para aprender a realizar tareas sencillas y en el margen de retención de las respues- tas aprendidas. Los análogos acuáticos de la “caja de Skinner” y de la “shuttle box”** son dispositivos que cada día se están volviendo más populares para el estudio instrumental del aprendizaje en el pez.
El condicionamiento mediante descargas eléctricas de la carpa dorada (Cura- ssius aurntus), para el estímulo visual, y del pececillo de cabeza adiposa (Pimephules promelas), entre otros, para el estímulo olfatorio, conduce a lograr respuestas de aprendizaje. Los resultados son detectados mediante la palpitación alterada del corazón (una respuesta autónoma) después de treinta y cinco tentativas, o me- diante movimientos de natación (una respuesta motora), después de cincuenta o
*Diriamos en español “quedar en blanco” (N. del T.)
**Para más detalles sobre estas cajas, véase Cohen, Josef, Aprendizaje Complejo. Cómo Aprenden los Animales. Ser. Temas de Psicología, No. 6. Ed. en Español. 1974. Edit. Trillas, México. (N. del T.)

a24 INTEGRACION
más ensayos. Existen grandes diferencias individuales en las respuestas. Algunos peces de un grupo experimental son más lentos para aprender que otros, y algu- nos parecen ser incapaces de aprender, inclusive cuando se les somete a pruebas “simples”.
El condicionamiento mediante una recompensa como el alimento, puede ser establecido más rápidamente que mediante el castigo; dependiendo de la tarea a cumplir, son suficientes de diez a veinte pruebas para lograr respuestas visuales en un pececillo del género Phoxinus o para obtener en Tilapia macrocephaiú el aprendizaje del regreso espacial para la localización del alimento. Los salmones (Oncorhynchus) que migran aguas abajo aprenden rápidamente a nadar una ruta constante de agua en un canal circular. Estas respuestas son probablemente esta- blecidas rápidamente, debido a que el pez está listo psicológicamente para su mi- gración hacia el océano, para la cual se reúne en grupos y nada grandes distancias. El aprendizaje mediante un ensayo ha sido mostrado con la carpa común (Cyprinus curpio), la cual, después del experimento, se volvió huidiza durante meses des- pués de haber sido capturada sólo una vez.
No solamente los peces óseos superiores (Actinopterygii), como los citados anteriormente, son capaces de aprender mediante los reflejos, sino también lo puede hacer el tiburón tigre (Galeocerdo cuvieri). Este tiburón aprende a golpear un blanco y a nadar a lo largo de un sendero señalado por estacas con el fin de obtener alimento como recompensa. El tiburón (Curcharhinus leucus) puede ser entrenado para asociar al alimento sonidos de 400 a 600 ciclos por segundo.
También los peces establecen reflejos condicionados de segundo arden. La carpa dorada puede, después de considerable entrenamiento, transferir a indicios visuales una respuesta aprendida sobre la base de señales olfatorias. Puede tam- bién hacer una transferencia similar de indicios auditivos a térmicos.
Los experimentos de discriminación sensorial que por primera vez y en forma más avanzada prueban la agudeza sensorial, se confían a la generalización, des- pués de que una respuesta condicionada ha sido establecida; esto es, ellos dependen de la capacidad del animal para responder a estímulos similares a, pero no idén- ticos con, los empleados en el procedimiento original para el entrenamiento. La generalización de diferentes valores de brillo y matiz, como componentes de la vi- sión, son fácilmente logrados por varias especies en muchas familias de peces; tam- bién se ha observado la generalización de los indicios olfatorios en los pececillos de acuario (Pirnephales).
Otro criterio que considere la inteligencia relativa puede consistir en la re- tención de las respuestas aprendidas sin necesidad de que haya un reforzamiento subsecuente (esto es, una memoria de gran duración). Los salmones (Qncorhyn- chus y Salmo) pueden retener los olores de los arroyos, aprendizaje que realizaron durante su periodo juvenil crucial, a lo largo de toda su vida de 2 a 6 años o más La memoria del olor de las aguas de las corrientes nativas es adquirida mediante un proceso de impronta. Debido a que las aguas de las corrientes donde los peces nacieron se encuentran muy diluidas para el periodo en que tardan en llegar a la desembocadura del rio, se ha propuesto la hipótesis de que existe una impronta múltiple. l,Jn salmón puede recordar los diferentes olores de cierto número de lugares durante su migración aguas abajo. El viaje de retorno podría consistir en- tonces en una serie de subviajes, en los cuales el salmón hace una especie de lec- tura de las series de 10s olores fijados mediante la impronta, pero en orden inverso, como si fuesen submetas. Mientras existe una fuerte evidencia para considerar

INTELIGENCIA Y COMPORTAMIENTO 325
a los olores fijados mediante la impronta como los principales recursos para la orientación durante el maravilloso recorrido que hace el salmón hacia su lugar de origen, es muy posible también que haga uso de indicios adicionales, como los visuales.
La percepción y uso conceptual de las relaciones espaciales son atributos de la visión, solo realizados, a juicio del hombre ordinario, por los mamíferos su- periores, porque esas habilidades son más fácilmente demostrables en los animales con miembros prehensiles que en otros vertebrados. Sin embargo, la percepción del fenómeno espacial y el aprendizaje perspicaz existe en la guaseta (Bathygobius soporator) y probablemente en muchos otros peces. La guaseta nada sobre pozas durante la marea alta, pero más tarde, cuando el nivel del agua ha bajado, salta de poza en poza con un extraño propósito y sin fijarse en la próxima poza al estar en el sitio de despegue. Peces trasplantados, que no han llegado a saltar en el área y son colocados en las pozas durante la marea baja no saltan o, cuando son empu- jados violentamente, pueden eventualmente hacerlo pero sin rumbo fijo. Después de 12 horas, durante la noche y habiendo marea alta, en una serie de pozas expe- rimentales, los mismos individuos saltaron indirectamente a las pozas de mayor tamaño. La orientación efectiva fue retenida durante cuarenta días sin necesidad de recibir experiencia adicional de “marea alta”. Realizaciones comparables, aun- que menos elaboradas, han sido reportadas para el pez sol verde (Lepomiscyanellus).
Respuestas retardadas y aprendidas con éxito, han sido consideradas como indicación de que el sujeto experimental en cuestión está respaldado por cierta dosis de perspicacia. Pececillos de acuario (Phorinus), después de habérseles per- mitido familiarizarse con una cámara de tres puertas para nadar libremente en su interior durante cierto tiempo, fueron más tarde obligados a coger un bocado de alimento que previamente se les mostró, nadando hacia adelante y regresado a través de uno o dos caminos para alcanzar el premio que para ese entonces estaba fuera del campo visual. Después de muchos ensayos, varios cientos, la actitud del pez no mejoró sobre un promedio de seis sobre diez soluciones correctas. Los sujetos también mostraron una conducta errática y vicaria de ensayo y error en el punto seleccionado. También desarrollaron actividades de desplazamiento, como la selección de piezas regadas de arena a la entrada del canal, conducta que signi- ficaría la toma de una decisión. Estas observaciones sugieren que el estímulo saciedad, una “actitud” negativa para el trabajo o montaje experimental, después de repetir varias veces los experimentos, pueden volver tormentosos estos experi- mentos en lugar de ser un medio inequívoco para demostrar que los peces son incapaces de aprender a resolver experimentos sobre respuestas retardadas.
Las observaciones de campo que corroboran los descubrimientos que se han hecho mediante los experimentos sobre las reacciones retardadas, a pesar de la precaución tomada con anticipación mediante el estímulo de saciedad, se han realizado al escapar de su trampa los pargos (Lutjunus, familia Lutjanidae) y los meros (Epinephelus, familia Serranidae). Los peces que han penetrado a las trampas deberán hacer cien o más intentos, mediante el procedimiento de ensayo-error, para encontrar una salida a través del alambre o malla antes de que la casualidad les pueda ayudar a localizar la salida. Una hora de encierro en la trampa no permitió a ninguno de los peces, aparentemente, realizar una exploración sistemática para encontrar una ruta o puerta de escape. Sin embargo, algunos peces de las familias ya citadas y aun de otras, como la del pez sol (Centrarchidae), hacen de las tram- pas SUS casas, según se sabe, y nadan entrando y saliendo libremente de ellas.

3 26 INTEGRACION
Otra aproximación, posiblemente indirecta, a la visión o a la habilidad con- ceptualizada de los peces proviene de la comparación entre las ratas y un pez (Tilapia macrocephala) en relación a la actuación sobre el reforzamiento incon- sistente del estímulo. En veinte ensayos por día, demandando discriminación visual entre barras horizontales y barras verticales, las respuestas al primer estí- mulo fueron reforzadas en un 70% y al segundo en 30% del tiempo. Después de 30 días de entrenamiento el estímulo previo al 70% fue reforzado consistente- mente en un 100% . Los registros de las respuestas de cada día a los estímulos del 70% mostraron que las ratas participaron activamente y que su preferencia por el estímulo más frecuentemente reforzado se elevó gradualmente de un nivel situado en 70.30 hasta alcanzar el 90% en el día 30. El pez, sin embargo, después de haber alcanzado el nivel 70.30 en el décimo día, aproximadamente, correspondiendo al estímulo reforzado con mayor frecuencia, permaneció en este nivel de actuación por el resto del experimento.
El entrenamiento de posición, que exige la selección entre dos posiciones de la colocación del alimento, fue realizado de una manera similar al experimento anterior, y dio lugar a resultados comparables. La deficiencia sensorial fue elimi- nada como una causa que produce la diferencia de conducta entre el mamífero y el pez, y se sugirió que la rata, en tales situaciones de prueba, hace una selección para responder todo o nada, lo cual constituye una indicación entonces de que no solamente tiene un elevado nivel de versatilidad en los procesos cerebrales, sino también cierta capacidad de abstracción. El pez, por el contrario, jugó con- tinuamente la probabilidad de ganar o perder mediante la selección de su res- puesta e indicó, como en el caso de la reacción retardada, que tiende a actuar en una forma más estereotipada y rígida que la del mamífero.
Otro aspecto de la conducta del pez, prominentemente involucrada con el sistema nervioso, es la presencia de un comportamiento reproductor pronunciado, a menudo espectacular, muy especializado y estereotipado (Capítulo lo), el cual ha sido objeto de numerosas investigaciones etológicas (conducta animal). La base de los componentes de la conducta son modelos de coordinación neuromus- cular que conducen a producir movimientos en el despertar de estímulos externos o internos. Estos modelos están genéticamente determinados y los movimientos que forman la serie conductual durante las actividades reproductoras aparecen, con sorpresiva homogeneidad, en todos los individuos de la especie. Ha sido posi- ble estudiar bien a los peces en este aspecto, debido a que es fácil guardarlos y se comportan con mucha normalidad en cautiverio. Estudios detallados sobre la con- ducta reproductora han sido realizados en la espinocha de tres espinas (Gasteros- teus aculeatus), varios cíclidos, en Cottus, el pez luchador (Betta) y otros.
Los peces producen movimientos típicos para cada especie durante el cortejo y/o construcción de nido, inclusive cuando son mantenidos en pleno aislamiento hasta su maduración. Por lo tanto, el modelo y ausencia de estos movimientos es justamente parte de la estructura genética, como lo es también la forma del cuerpo y la coloración. La literatura etológica antigua hacía caso omiso del papel del aprendizaje en la conducta heredada, pero actualmente se concluye que el apren- dizaje también juega un importante papel y que la modificación, y posiblemente el refinamiento, de los componentes de la conducta es posible, e incluso puede considerársela como la regla (como se sabe, por ejemplo, de un cíclido, Cichla- soma meeki).
Los movimientos que complementan las actividades de cortejo de las espi-

INTELIGENCIA Y COMPORTAMIENTO 3 27
nochas (Gasterosteidae), por ejemplo, pueden ser analizados hasta el detalle; así, por ejemplo, la actividad de músculos aislados o grupos de músculos, como los que mueven a una sola aleta. Esos movimientos localizados forman parte de gran- des unidades y están coordinados de acuerdo a grupos ordenados con cierta jerar- quía para formar, en su conjunto, los instintos. Se infiere, en consecuencia, que estos movimientos pertenecen a distintos sistemas causales y funcionales que ac- túan como unidades. Cuando ellos están trabajando reducen la actividad de otros sistemas, diferentes funcionalmente pero equivalentes. El instinto también im- plica una serie de actividades que, más que realizadas al azar, siguen cierto patrón. Estos patrones de conducta operan a través de la función ordinaria de los órganos de los sentidos, el sistema nervioso central, y los efectores, de tal manera que nosotros encontramos una secuencia más o menos establecida y obligatoria de estímulos y respuestas para toda la actividad, hasta llegar a su conclusión. Entre las secuencias que siguen un cierto patrón, la más estudiada corresponde a la conducta reproductora de la espinocha de tres espinas. Se ha mostrado repetida- mente en este pez que un movimiento particular del tren de conducta actúa como un estímulo para el siguiente (Figura 11.6). También se ha mostrado cómo uno puede representarse todo el instinto reproductor de este pez subdividiéndolo en fases y organizándolas jerárquicamente en componentes que consisten de movi- mientos individuales.
La interacción entre las fuerzas genbticas y las del medio ambiente durante la evolución le ha dado forma de tal manera al modelo y a las respuestas del sis- tema nervioso, incluyendo a los receptores, que el estímulo simplificado, breve, o parte del conjunto, a menudo es suficiente para “disparar” movimientos posteriores formando una secuencia. Por ejemplo, el color plateado o el perfil hinchado de una espinocha hembra madura provocan la danza en zigzag del macho al comienzo de las actividades del cortejo. Se ha demostrado que muchos patrones prominen- tes de colores de los peces y otros animales, especialmente manchas que pueden
Macho
Danza en
Muestra la entrada -+-”- al nido \
Fertiliza
Figura 11.6 - Secuencia de los
Hembra
Aparece
-
Cortejo
Prosigue
Penetra al nido
Se aparea
movimientos de- cortejo mostrados por el macho, y las reacciones de la hembra, en la espinocha de tres espinas (Gasterosteus aculeatus). (Fuente: Timbergen, 1951).

328 INTEGRACION
ser exhibidas, poseen esas funciones disparadoras como, por ejemplo, las marcas blancas y negras de la cola y los flancos del pececillo Lebistes. Muchas señales liberadoras pueden ser usadas en determinada situación, sumándose cada una a la fuerza de la anterior, de tal manera que uno puede obtener un modelo supernormal con un valor liberador mucho mayor del normal (principio de la adición heterogé- nea); inclusive un modelo burdo de una espinocha hembra que está muy plateada y tiene un abdomen excesivamente grande asume esa función. El principio de la adición del estímulo también se aplica a las variables hidrográficas y a otras del medio ambiente que actúan como liberadoras en la conducta reproductora de pe- ces como el lucio septentrional (Esox lucius), los peces blancos (Coregonus) y la trucha café (Salmo trutta).
La primera teoría sobre los mecanismos liberadores le dio probablemente excesiva importancia a los componentes del sistema nervioso central. Debemos señalar, sin embargo, que el término “mecanismo liberador” es operativo antes que indicativo de un lugar preciso en el animal. También debemos notar que el concepto ha sido forjado para el complejo sensorial órgano-nervio sistema-efector, antes que para un modelo central o dispositivo patr6n.
El curso de la conducta estereotipada está influido tanto por factores inter- nos como externos. Entre los factores externos se cuentan los estímulos liberadores en forma de signo, como los colores y la forma específicos para cada especie, como ya se ha mencionado. Entre los factores internos que intervienen en la con- ducta reproductora se cuentan primordialmente los niveles hormonales que determinan la intensidad y, también, hasta cierto punto, el curso seguido por la conducta. En las actividades no reproductoras, como la alimentación, el ham- bre, como un factor interno, puede determinar la extensión y secuencia de los movimientos requeridos para satisfacer el apetito. En el análisis de los patrones y de la motivación de los movimientos aparecen ciertas tendencias diferentes, como el sexo, el vuelo y la agresión, que influyen en el curso del comportamiento ins- tintivo. La danza en zig-zag es determinada por el macho y varía de acuerdo a su acercamiento a la hembra y al nivel de sus tendencias sexual y agresiva, lo que determina en esta el movimiento en “zag”.
En el curso de una cadena de comportamiento integrada como el cortejo, un pez puede interponer ciertos movimientos como los de la alimentación, irri- tarse, u otros que muestren confort y que pertenecen a actividades enteramente diferentes o a instintos muy diversos. Entonces, por ejemplo, si dos espinochas machos se encuentran en el lindero de sus respectivos territorios, por lo general no pelean, pero exhiben movimientos de excavación que pertenecen a la conducta relacionada con la construcción del nido, que podrá observarse mucho más tarde durante el comportamiento reproductor y no durante el periodo en que estable- cen sus territorios. Estos “movimientos de desplazamiento” resultan de un con- flicto entre tendencias diferentes, en este caso, de la agresión y la fuga y quedan fuera de la secuencia normal de actividades.
Las actividades que no son liberadas por sus correspondientes estímulos pueden ser exhibidas durante un tiempo considerable sin que esté presente nin- guno de los estímulos adecuados, u.n fenómeno llamado “actividad vacia”. Ma- chos de pececillos de acuario (Lebistes), mantenidos en aislamiento, pueden, cuando se vuelven sexualmente maduros, realizar movimientos de exhibición sigmoideos que muestran en una situacibn normal estando a la vista de la hembra. Cíclidos (Cich- lidae) que no han tenido la oportunidad de cuidar su propia cría pueden comenzar

6RGANOS ENDOCRINOS 329
a proteger un grupo de pulgas de agua (Daphnia) como una reacción “overflow” (de desbordamiento).
El sistema nervioso ha jugado un papel clave en la evolución de los patrones de conducta específicos de las especies. Al mismo tiempo, el estudio de la conduc- ta de las especies relacionadas revela considerable plasticidad en el cambio y mo- dificación del patrón, de tal manera que el comportamiento derivado de una acti- vidad puede ser utilizado para otra y adquirir nuevo significado (ritualización). Las “manchas oculares” que aparecen comúnmente en los opérculos, colas o aletas de los peces, son aparatosamente exhibidas cuando se comportan con miedo o se li- beran y huyen, o cuando buscan para su protección a un compañero de la misma especie, pero también pueden ser empleadas para las actividades del cortejo. Los liberadores sociales, esto es, los patrones especialmente adaptados para tener una función de señal, juegan cierto papel en muchas clases de comportamiento. Ayu- dan al joven a encontrar a su padre, sirven para mantener un orden jerárquico entre los individuos y son instrumentales al ritualizar el comportamiento de pe- lea en el que le permiten al animal golpeado mostrar signos de inferioridad antes de ser lastimado. Son muy importantes en la sincronización de las actividades sexuales y pueden ser inclusive operativos en las relaciones interespecíficas. Mu- chos peces territoriales, como los cirujanos (Acanthurus) y los chopa o señorita (Eupomacentrus), realizan simples exhibiciones amenazadoras, así como el coml portamiento de persecución y empuje para defender su reserva alimepticia for- mada por algas bentónicas, de la invasión de otros herbívoros. Los peces de los arrecifes, en todo el mundo, muestran exhibiciones de solicitación más o menos ritualizadas para atraer la atención de organismos que les auxilien en la limpieza. Estas solicitaciones y exhibiciones muestran un grado notable de variabilidad entre las familias, pero las especies dentro de muchas familias muestran casi idén- tica conducta simbiótica, inclusive a pesar de que han sido separados de sus con- géneres en los diferentes océanos durante muchos miles de años.
La falta de información sobre la organización y función detallada del sis- tema nervioso central retarda el análisis del asiento de la coordinación del com- portamiento instintivo. La extirpación del cerebro anterior en la carpa dorada o la tilapia resultó en defectos masivos en la adquisición y retención del comporta- miento de escape. Otros experimentos indican que la remoción del cerebro anterior disminuye muchas funciones, pero elimina unas cuantas. Puede considerarse a los peces entre los sujetos mejor investigados por los etólogos en sus tres objetivos de estudio: la explicación de la conducta en términos fisiológicos, el estudio de su significado biológico en conjunción con la ecología, y la comprensión del desa- rrollo de la conducta en el curso de la evolución.
~ R G A N O S ENDOCRINOS
El sistema endocrino de los vertebrados está ya presente en su esencia en las lam- preas y los mixinos (Cyclostomata); las similitudes posteriores con los vertebrados superiores aparecen tanto en los tiburones (Elasmobranchii) como en los pe- ces óseos (Osteichthyes). Sin embargo, las diferencias entre las glángulas endocri- nas del pez y del mamífero son todavía considerables. Estas diferencias glandulares pueden, probablemente, ser correlacionadas con las diferencias en la evolución de otros sistemas corporales en los dos grupos y con las exigencias que se presentan

330 INTEGRACI~N
al vivir en el agua. Aunque la endocrinología de los mamíferos está muy avan- zada, solamente ciertos aspectos de las funciones controladas hormonalmente han sido investigados en los peces. Los que han sido estudiados se refieren a la influen- cia de las secreciones endocrinas en las células que producen color, la acción recíproca entre las células sexuales y la pituitaria, y la función tiroidea y la adapta- ción para la migración, pero los detalles minuciosos, los más finos, sobre la acción e interrelaciones de las glándulas endocrinas del pez, esperan todavía su elucidación.
El sistema endocrino, en contraste con el sistema nervioso, concierne pri- mariamente a los procesos relativamente lentos de una naturaleza metabólica, como el metabolismo del nitrógeno (tejido cortical adrenal, glándula tiroides), metabo- lismo de los carbohidratos y el agua (tejido cortical adrenal), la maduración de las célu1asb;exuales y el comportamiento reproductor (la glándula pituitaria y las hor- monas gonadales).
A continuación se consigna una lista (Tablas 11.3 y 11.4) de las glándulas endocrinas de los peces, con la mención de los órganos que posiblemente tienen funciones endocrinas; asimismo, las glándulas individuales y su acción, a excep- ción de las que tienen función dudosa que serán tratadas en secciones separadas; para la localización de las glándulas en el cuerpo del pez véase la Figura 3.19, y, para conocer su interacción, véase el diagrama en la Figura 11.7.
La glándula pituitaria (hipófisis)
Esta glándula maestra endocrina se encuentra en toda la línea evolutiva de los ver- tebrados y se desarrolla embrionariamente a partir de un elemento neural que crece hacia abajo, desde el diencéfalo, para encontrarse con un elemento epitelial (la bolsa de Rathke) que crece hacia arriba a partir de la cavidad bucal. Estos dos elementos son entonces de origen ectodérmico y encierran entre sí al mesodermo, del cual se origina la futura y copiosa irrigación sanguínea de la pituitaria. Esta alimentación sanguínea se deriva de la arteria carótida interna. Hacia adelante de la glándula pituitaria se encuentra el quiasma óptico y hacia atrás el saco vascular. El tamaño del pedúnculo o infundíbulo que fija la glándula al piso del diencéfalo aumenta del grupo de las lampreas (Cyclostomata) al de los peces óseos (Osteich- thyes); simultáneamente, el hueco situado en la base de la caja craneal, donde la glándula está alojada, se vuelve más pronunciado (Figuras 3.20, 11.1 y 11.8).
Anatomía e histología de la glándula. La glándula pituitaria o hipófisis posee en los peces una adenohipófisis tripartita y una neurohipófisis (con variaciones en la terminología de las partes). La división de la adenohipófisis en pro-, meso- y meta- adenohipófisis está brsada en sus posiciones anteroposteriores relativas y en los diferentes tipos de sus células secretoras. Estas divisiones de la adenohipófisis son también denominadas la rostral pars distalis, la proxirrlal pars distalis y la pars intermedia, para enfatizar su homología con la glándula en los tetrápodos. La porción posterior, meta-adenohipofisal, se encuentra por debajo de la delgada neurohipófisis; ella recibe las prolongaciones de las células neurosecretoras hipo- talámicas. En los tiburones y en las rayas (Elasmobranchii) la hipófisis se proyecta lejos en dirección ventral y tiene una extensión hacia adelante, el lóbulo rostral, que contiene componentes pro-adenohipofisarios de diferenciación histológica similar a la de los ciclóstomos. Un crecimiento hacia abajo en los elasmobranquios

6RGANOS ENDOCRINOS 331
es llamado lóbulo ventral. La porción más anterior de la adenohipófisis es su división vesicular y se ha demostrado que en ella ocurren cambios cíclicos en su actividad secretora con la gestación y el crecimiento. En el celacanto de aletas lobuladas (Latimeria) la división rostral de la pituitaria (proadenohipófisis) se deriva ventralmente y está independientemente vascularizada, una característica organizativa que recuerda con mucho a los elasmobranquios.
La glándula pituitaria en los peces óseos superiores (Actinopterygii) se ha vuelto más compacta que en las lampreas y en los tiburones, acarreando grandes dificultades para poder establecer una división clara entre sus partes. La actividad secretora diferencial puede, sin embargo, ser demostrada mediante técnicas ordi- narias de tinción. Células ganglionares de gran tamaño aparecen en la adenohipó- fisis y en la neurohipófisis de peces como las truchas y el salmón (Salmo) y en los bacalaos (Gadus). Hay evidencias de que las hormonas neurohipofisarias son se- cretadas en los tejidos neurosecretores del hipotálamo y que se acumulan en la neurohipófisis desde donde encuentran su camino hacia el torrente circulatorio. Parece ser que el control de la adenohipófisis, por lo menos en parte, está basado en la neurosecreción. La hipófisis del pez de aletas lobuladas (Latimeria) difiere marcadamente de la de otros peces. Consiste en una estructura en forma de cuer- da, de alargamiento poco usual.
Las hormonas pituitarias. Muchas de las hormonas de la pituitaria del pez produ- cen los mismos o análogos efectos que las de la incomparablemente mejor estudiada glándula de los mamíferos. Todas las hormonas pituitarias conocidas son proteí- nas o polipéptidos y en muchos casos se han llegado a usar extractos d,o laglándula de mamíferos para establecer el papel de algunas de ellas en los peces. Es de ha- cerse notar que se han establecido pequeñas diferencias en la composición quí- mica para la misma hormona en diferentes familias de mamíferos (cerdos y reses); es por lo tanto de esperarse que existan diferencias similares entre las hormonas de los diferentes grupos de peces. La especificidad de las hormonas de la pituita- ria del pez ha sido ya mostrada mediante experimentos y aplicaciones prácticas con inyecciones e implantación para forzar el desove de ciertos peces de impor- tancia económica, como los esturiones (Acipenseridae), truchas (Salmoninae), bagres (Ictaluridae) y lisas (Mugilidae).
Las hormonas de la pituitaria de los peces pueden ser divididas en dos gru- pos, uno que controla recíprocamente la función de otras glándulas endocrinas y el otro que influye en procesos enzimáticos selectos en uno o varios tipos de célu- las somáticas. En el primer grupo se cuentan la adreno-corticotrópica, la tirotrópica y las hormonas gonadotrópicas que son secretadas en la pro o mesoadenohipófi- sis, dependiendo de la especie de pez. En el segundo grupo hay varias hormonas, descritas abajo, que afectan a los melanóforos.
Se sospecha que la pro-adenohipófisis es el origen de una hormona melá- nica (MAH) que actúa en las células portadoras de melanina de la piel, para pro- ducir una adaptación de fondo más permanente de la que se podría obtener a través de los impulsos nerviosos únicamente. La hormona estimuladora de los melanóforos (MSH o intermedina) de la meta-adenohipófisis, actúa antagónica- mente al MAH y expande el pigmento en las células de color, contribuyendo entonces a producir un ajuste de fondo de gran amplitud; esta hormona sintetiza posteriormente a la melanina.

332 INTEGRACION
Tabla 11.3 ~ Partes, secreciones y acci6n de las secreciones del sistema endocrino en los peces. *
Glándula Localizacibn Origen embrionario
Pituitaria Base del cerebro
Lbbulo posterior (Neuro- hipbfisis)
Lbbulo o zona intermedia (Metaadenohip6fisis)
Ldbulo o zona transicional (Mesoadenohip6fisis)
Lóbulo o zona anterior (Pro-adenohipbfisis)
Tiroides
Últimobranquiales
Peces bseos, difusa, o en unos cuantos, Endodermo (con mesoder- discreta, a lo largo de la aorta ventral. mo agregado despu6s) Peces cartilaginosos, línea media entre los músculos linguales
Estructuras en forma de saco entre la Endodermo (?) pared ventral del esbfago y el seno ve- noso (en el caracínido Astyanax y en muchos otros peces bseos superiores)
Suprarrenales (tejido cro- Dorsalmente a lo largo de los riñones Mesodermo mafínico)
Tejido cortical adrenal (Te- Una de las tres capas celulares que se Mesodermo jido de Giacomini; hornb- hallan a lo largo de las venas cardinales logo a la corteza adrenal en la regibn de la cabeza hematopoib- de los mamíferos) tica del riñbn
Islotes pancreáticos Paredes del tubo digestivo en las lampreas Mesodermo larvarias: hepatopáncreas, la mayoría de los peces bseos; páncreas discreto, algunos peces.
Glándulas sexuales Cavidad del cuerpo Mesodermo
Mucosa intestinal Intestino pequeño Endodermo
Corpúsculos de Stannius Adosados o incluidos en los riñones de Mesodermo los holbsteos y los telebsteos

6RGANOS ENDOCRINOS 333
Secreción Accibn
Vasotocina arginina Oxitocina
lntermedina (MSH)
Hormona de crecimiento (GH)
Tirotropina (TH)
Gonadotropina(s) (LH, FSH)
Prolactina
Sustancia productora de exoftalrnia O?). Hormona concentradora de los rne- lanbforos ( ? I
Prolactina
Corticotropina (ACTH)
Hormona concentradora ds los mela- nóforos (?)
Hormona(s) tiroidea(s1
Calcitonina
Epinefrina, Norepinefrina
Esteroides corticales adrenales
lnsulina
Andrbgeno(s?) en los machos
Estr6geno(s?) en las hembras
Secretina (Pancreozimina)
Desconocida
Osrnorregulacibn y equilibrio del agua salada Unidn sexual y desove
Melanogenesis (fundulus); dispersión melánica
Aumento de tamaño; efectos metabólicos (?I
Controla la secrecibn tiroidea
Controla la secreción de las hormonas gonadales
Melanoghesis junto con MSH
¿Palidez a traves de la contracción melánica?
Regulación de electrolitos y melanoghesis
Controla la secrecibn cortical y adrenal; melanoghesis (Care- ssius)
¿Palidez a traves de la contracción melánica?
Apresura la maduración; controla el metabolismo, pero aparentemente no tiene control directo del metabolismo oxidativo; relacionada con los cambios metambrficos y posi- blemente con la osmorregulacibn
Metabolismo del calcio (funcibn parecida a la de la parati- roides)
Concentra los gránulos de pigmento en los rnelanbforos; con- trola la elevacibn de la presibn sanguínea y la dilatacibn pupilar; afecta el ritmo del pulso, etc. (efectos comparables a los producidos por el sistema nervioso simpático)
Promueven la utilización de las reservas de grasa; metabolis- mo de los carbohidratos; metabolismo del agua;catabolismo de las proteínas; retención del sodio; metabolismo elec- trolítico
Metabolismo de los carbohidratos
Desarrollo de los caracteres sexuales secundarios; comporta- miento reproductor; maduracidn de los garnetos
Desarrollo de los caracteres sexuales secundarios; comporta- miento reproductor; maduración de los gametos
Estimula la secreci6n pancreática
Metabolismo del calcio

334 INTEGRACIdN
Tabla 11.4 drganos endocrinos del pez con funciones dudosas y desconocidas.
Glándula Localización Evidencia de funcibn endocrina
”-
Timo AI dorso de los arcos branquia- Crecimiento (?) les (y, en algunos, en el lado ventral)
Cuerpo pineal Dienc6falo Acciona en los melanóforos Pseudobranquias Cavidad branquia1 en el hioides Equilibrio del cloro Sistema neurose- Extremo de la médula espina1 Metabolismo del agua y de cretor caudal en la cola; células similares a los iones; intercambio (Urohipófisis) las células secretoras del siste- de sodio ( ? I ; metabolis-
ma hipotálamo-hipofisial neu- mo de los gases (?) rosecretor
Estímulos externos
/Luz h
Temperatura COmpOSiCiÓn‘
\ J , química del agua I(
Centros diencefálicos (tálamo)
Glándula pituitaria
d- T e j i L iuprarrenal Tejido Tiroides Islotes ‘u x
Gónadas
I \ I
.1 Reproducción
(cromafínico) i n t e r y a k pancreáticos
Metabol
Conducta Pigmentación
l i m o
Figura 11.7 - Interacción de algunas glándulas endocrinas de 10s Peces. (Basado en Vivien en Grass6, 1958).
Se ha demostrado que la prolactina, similar a la hormona que influye en la lactación de los mamiferos, es producida en la proadenohipófisis. En unión con la intermedina, la prolactina promueve la formación de nueva melanina en los melanóforos de la piel de algunos peces como el ciprinodóntido Fundulus hete- roclitus. La prolactina es una de tantas hormonas implicadas en la regulación electrolítica en los teleósteos, pero su importancia para mantener la homeostasis parece variar con las especies. L a hormona adrenocórtico-trópica (ACTH, cuya naturaleza será discutida más adelante), aparte de su acción directa en el tejido productor de hormona de la corteza adrenal (localizado difusamente en el riñón de los peces), tiene también un efecto estimulante en la producción melánica del

6RGANOS ENDOCRINOS
GLÁNDULAS PITUITARIAS
335
(u) Lamprea . ..
MA
Figura 11.8 - Diagramas de la glándula pituitaria de los peces: (a ) lamprea (Petromyzon); (6) tiburbn perro (Squalus); (c) trucha (Salmo); (d) per- ca (Perca). HL, luz de la hipbfisis; IN, infundíbulo; MA, mesoadenohipbfi- sis; ME, metaadenohipbfisis; NE, neurohipbfisis; PA, proadenohip6fisis; SV, saccus vasculosus; VL, 16bulo ventral. (Basado en Pickford y Atz, 1957).

336 INTEGRACIdN
pez dorado amarillento (Carassius auratus). Investigaciones posteriores deberán explicar o reconciliar estas diferencias que hay sobre la causa de la melanogéne- sis. Una hormona de crecimiento de la meso-adenohipófisis promueve el creci- miento en longitud del cuerpo de los peces. No se ha explicado el modo en que actGa en la división celular y en la síntesis de las proteínas.
El lóbulo ventral secreta probablemente gonadotropinas en el tiburón perro y en la mantarraya. También se ha encontrado que el lóbulo ventral tiene una función tirotrófica. La neurohipófisis produce o almacena, a partir de las células neurosecretoras hipotalbmicas, sustancias endocrinas que son bien conocidas por los efectos que producen en el metabolismo de los mamíferos. Las hormonas vasopresoras y antidiuréticas contraen los vasos sanguíneos de los mamíferos y promueven la retención del agua a través de su acción en el riñón. L a oxitocina estimula al músculo uterino de los mamíferos, aumenta la producción de leche en las mamas iactantes y disminuye la presión sanguínea en las aves. El material pituitario del pez produce esos efectos en animales o tejidos testigo de vertebra- dos superiores, pero se puede presumir que los órganos blanco o los lugares espe- cíficos de acción de las hormonas neurohipofisarias, en los peces, podrian diferir de las de los vertebrados superiores donde nosotros encontramos problemas radi- calmente diferentes de osmorregulaci6n y, en consecuencia, del equilibrio del agua y la sal. Si se comprueba que la sangre del pez carece de hormonas neurohipofi- sarias, se entenderá entonces que ellas actúan como “hormonas locales” en la adenohipófisis.
La glándula tiroides
La hormona de la glándula tiroides contiene yodo inorgánico extraído del to- rrente circulatorio y almacenado en los folículos de la glándula, el cual es liberado en el sistema circulatorio bajo las demandas metabólicas específicas. El nivel de actividad de los folículos está bajo la influencia de la hormona tirotrópica de la pituitaria que, oportunamente, es gobernada por lo menos en parte por una com- binación de procesos de maduración genéticamente determinados, en conjunción con ciertas variables del medio ambiente como el fotoperiodo, la temperatura y la salinidad. En ciertos tiburones (Elasmobranchii) y en la mayoría de los peces óseos superiores (Actinopterygii), con algunas excepciones como los peces loro (Scaridae), la glándula tiroides posee un tejido difuso desparramado a lo largo de la base de la aorta ventral (Figura 3.19); la extirpación o inactivación de la glán- dula, un prerrequisito para hacer estudios sobre deficiencia, es en consecuencia difícil de hacer, pero se ha podido realizar mediante eli bloqueo fisiologico o la radiotiroidectornia usando Poco es lo que se sabe sobre la acción de las hor- monas tiroideas en los ciclóstomos o en los elasmobsanquios. La estimulación respiratoria mediante la tiroxina es la acción mejor conocida en los mamíferos, y aun así la mayor evidencia indica que la tiroxina no produce ese efecto en los teleósteos. Pequeñas dosis de tiroxina y triyodotironina causan, fisiológicamente, engrosamiento de la epidermis y palidez en la carpa dorada (Carassius auratus). Dosis elevadas de tiroides retardan el crecimiento de hs larvas de los salmones (Oncorhynchus), pero es acelerada la transformación al estado juvenil latente en el mismo género mediante la inducción de la hiperactividad tiroidea. En el cava- dor de fango (Periophthulmus), un pez que permanece parcialmente fuera del

ÓRGANOS ENDOCRINOS 337
agua mucho tiempo, la inducción de la hiperactividad tiroidea conduce al animal a una existencia todavía más terrestre, seguida de cambios morfológicos y meta- bólicos. Parece ser que la glándula tiroides influye en parte en la osmorregulación de los salmones (Oncorhynchus) y de las espinochas (Gasterosteus) y la hiperacti- vidad tiroidea ha sido observada durante las migraciones de los salmones para su reproducción. Por último, aunque no de menor importancia, son las indicaciones que hay sobre el efecto de la tiroides en la eficiencia del crecimiento y el metabo- lismo del nitrógeno en la carpa dorada, expresado a través de la elevación de la excreción del amoníaco.
Parece ser que en los peces las funciones de la tiroides están sujetas a una amplia variedad de procesos, que algunos de ellos están relacionados con el creci- miento y con los fenómenos de la maduración y que la tiroides, junto con otras glándulas endocrinas, juega un importante papel en las migraciones diádromas de aquéllos.
Las glándulas últimobranquiales
Las glándulas últimobranquiales pequeñas y pareadas tienen su origen embriona- rio cerca del quinto arco branquial, pero finalmente se alojan cerca de la glándula tiroides. La hormona calcitonina se encuentra en las glándulas últimobranquiales del pez y en las glándulas tiroides y paratiroides internas de los mamíferos, en donde produce hipocalcemia. Los experimentos realizados con el ciprinodóntido Fundu- lus heteroclitus y con las anguilas europea y japonesa (Anguilla anguilla y A. japo- nica) han fracasado al intentarse conformar un efecto hipocalcémico de la calci- tonina. La calcitonina puede estar relacionada con la osmorregulación. Se ha mostrado que la calcitonina de la anguila disminuye la osmolaridad del suero, el sodio y el cloro en las anguilas japonesas. Las glándulas últimobranquiales pue- den estar sujetas al control pituitario.
Tejido cromafínico (cuerpos suprarrenales; tejido medular)
Los tejidos cromafínicos reciben su nombre a causa de una reacción de tinción característica. En el grupo de las lampreas (Cyclostomata), las células cromafí- nicas están dispuestas como filamentos a lo largo de la aorta dorsal y en el ven- triculo y la vena porta del corazón, pero en los tiburones y en las rayas (Elasmo- branchii) el tejido se adosa en asociación con la cadena simpática de los ganglios nerviosos. En los peces óseos (Actinopterygii), la localización y concentración del tejido cromafínico muestra considerable variación, comprendiendo desde un modelo elasmobranquial, por un lado, como sucede en los lenguados (Pleuronec- tes), hasta una disposición verdaderamente adrenal, como ocurre en los peces del género Cottus, donde la cromafina y el tejido cortical adrenal están unidos en un órgano, comparable a la glándula adrenal de los mamíferos.
Los extractos de tejido cromafínico de los peces son ricos en adrenalina y noradrenalina. La administración exógena de las catecolaminas (adrenalina y no- radrenalina) da como resultado las siguientes respuestas fisiológicas: bradicardia, cambios en la presión sanguínea, vasodilatación branquial, incremento en los niveles de glucosa en la sangre, diuresis en los teleósteos glomerulares e hiperven-

338 INTEGRACI~N
tilación. Estas reacciones “pelea o huye” de los peces son análogas a las de los vertebrados superiores y señalan a los cuerpos suprarrenales desempeñando una función similar a la de la médula adrenal de los mamíferos.
Tejido cortical adrenal (tejido interrenal)
Las células endocrinas que producen compuestos de esteroides adrenocorticales están dispuestas a través de la cavidad del cuerpo, cerca de las venas postcardina- les en el grupo de las lampreas (Cyclostomata). Entre los tiburones (Squaliformes), descansan entre los riñones. En las rayas (Rajiformes) asumen una asociación más o menos estrecha con el tejido renal posterior, teniendo algunas de sus especies el tejido cortical adrenal agregado cerca de la izquierda y otras cerca de la dere- cha del borde central de ese órgano. En los peces óseos (Actinopterygii) hay una o varias capas de células interrenales que se encuentran a lo largo de las venas postcardinales, cuando pasan a través de la cabeza renal (pronefros).
La secreción de las hormonas córticoadrenales está bajo el control de la hormona adrenocorticotrópica (ACTH) de la hipófisis. En la platilla o sardinita mexicana (Astyanux mexicanus) la hiperfunción de las células interrenales puede ser producida experimentalmente, así como una condición, que recuerda el sín- drome de tensión adrenal, bien descrito en los mamíferos. El cortisól, la corti- sona, la corticosterona y la aldosterona, conocidas en las glándulas adrenales de los mamíferos, así como algunos compuestos desconocidos, han sido aislados del plasma sanguíneo del salmón “sockeye” (Oncorhynchus nerka). Estos hallaz- gos sugieren, como es evidente en los mamíferos, que dos tipos de secreciones son producidos por el tejido cortical adrenal del pez: a) corticoides minerales que influyen en ciertos aspectos de la osmorregulación del pez; b ) glucocorticoides que influyen en el metabolismo de los carbohidratos, especialmente a nivel de los azúcares sanguíneos. La evidencia experimental para la primera función proviene de la trucha arco iris (Salmo gairdneri) que excreta cantidades mucho mayores a la normal de iones de sodio a través de sus branquias, pero retiene más de la can- tidad normal de sodio en los riñones cuando está sometido a cargas internas de sal bajo el tratamiento corticoide mineral. La segunda función fue verificada en el pez sapo (Opsanus tau) donde los niveles de azúcar de la sangre fueron elevados mediante la inyección intramuscular de compuestos corticoesteroidales.
La concentración de cortisona en el plasma sanguíneo de los salmones (On- corhynchus) es elevada durante las etapas de desove de su ciclo biológico, pero baja durante las etapas más sedentarias. En Oncorhynchus, el 60% de la proteína del cuerpo es catabolizada durante la fase de migración-desove, y este catabolism0 está asociado a un incremento sextuplicado de los corticoesteroides en el plasma y a un aumento del glucógeno del hígado. La inyección de hormonas, cortico- adrenales promueve la liberación de linfocitos en Astyanux o la de anticuerpos en la perca europea (Perca fluuiutilis), haciendo patente la similitud en la función de los tejidos corticoadrenales en los vertebrados superiores e inferiores. Los corticoesteroides recuerdan por SL: estructura a los andrógenos y presentan efec- tos laterales androgénicos. Estos efectos laterales sugieren alguna equivalencia funcional en los peces. Los ritmos diarios de las concentraciones de corticoides adrenales del plasma han sido reportados para muchos teleósteos, y estos ritmos pueden desempañar un papel importante en la conducta de estos peces.

6RGANOS ENDOCRINOS 339
Corpúsculos de Stannius
Los corpúsculos de Stannius son órganos endocrinos adosados a los riñones de los holósteos y de los teleósteos, o embutidos en ellos. Su número varía con las espe- cies y recuerdan histológicamente a las células corticales adrenales. Los corpúscu- los de Stannius bajan los niveles del calcio en el suero del ciprinodóntido Fundulus heteroclitus mantenido en medio de elevadas concentraciones de calcio, como el agua de mar. Parece ser que los corpúsculos trabajan conjuntamente con la glándula pituitaria, que produce un efecto hipercalcémico para mantener un nivel relativamente constante de calcio en el suero.
Las glándulas sexuales como órganos endocrinos
En los peces, la maduración y posiblemente también la elaboración de gametos requieren la presencia de las hormonas sexuales; además, las características sexua- les secundarias, como la coloración, los tubérculos reproductores y la maduración del gonopodio, dependen de la presencia de las sustancias endocrinas originadas en las glándulas sexuales. Las hormonas sexuales son producidas por células especia- lizadas de los ovarios y de los testículos a niveles determinados por la producción de las hormonas gonadotrópicas provenientes de parte de la glándula pituitaria (meso-adenohipófisis).
La hormona masculina, testosterona, ha sido medida en el plasma sanguí- neo de un elasmobranquio (Ruju radiata) y en el salmón, en donde hay una corre- lación entre los niveles del plasma y el ciclo reproductor. Otro esteroide gonadal, la 11-ketotestosterona, es fisiológicamente diez veces más androgénica que la testosterona en Oryzias latipes (“medaka”) y en el samón “sockeye” (Oncorhyr- hynchus nerha). De los estrógenos femeninos, el estradiol-170 ha sido encontrado en muchas especies, a menudo acompañado de cantidades variables de estrona y estriol. La progesterona está presente en el pez, pero no se sabe si este compuesto tiene una función hormonal.
Ha habido unas cuantas demostraciones inequívocas para los peces en el sentido de que el comportamiento reproductor es inducido o regulado por este- roides gonadales. Muchos investigadores han demostrado los efectos de los esteroi- des exógenos en la determinación del sexo y en el desarrollo de caracteres sexuales secundarios en el pez, pero en la mayoría de los casos se menciona muy poco sobre los efectos de estos tratamientos en el comportamiento reproductor.
Después de una inyección de testosterona y estrona de mamíferos, una lamprea (Lampetru fluuiatilis) desarrolló sus labios cloacales y los poros celómi- cos, ambas estructuras relacionadas con el proceso reproductor. Las pruebas realizadas en tiburones y rayas (Elasmobranchii) para encontrar una similitud entre las hormonas sexuales de diferentes clases de vertebrados no han mostrado claros resultados, pero entre los peces óseos (Osteichthyes) esa equivalencia parece existir, aunque los experimentadores mantienen ciertas reservas. Por ejemplo, la etinil testosterona (pregneninolona), que tiene efectos ligeros parecidos a los de la hormona androgénica y la progesterona en los mamíferos y en las aves, parece ser altamente androgénica en los peces. Las inyecciones de testosterona en peces loro (Scaridae) inmaduros y femeninos evocan por igual la apariencia del macho, por lo que respecta a su patrón de coloración, en el sujeto sometido a la experi-

340 INTEGRACXON
mentación. Las hormonas sexuales femeninas de los peces son menos parecidas a las de los otros vertebrados, que las hormonas testiculares que también pueden afectar fuertemente el desarrolio ovárico, como se ha visto en el pez climático japonés (Misgurnus fossilis). La evidencia hasta la fecha señala que hay menos especificidad bioquímica en las hormonas androgénicas y estrogénicas de los peces que en los mamíferos y en las aves, y un mayor grado de acción en las gónadas por hormonas sexuales análogas, como, por ejemplo, ciertos corticoesteroides.
Las prostaglandinas, que son sustancias muy potentes parecidas a las hor- monas, han sido recientemente descubiertas en los testículos y en el semen del lenguado (Paralichthys oliuaceus), del atún de aleta azul (Thunnus thynnus) y del salmón compañero (Oncorhynchus ketu) . En los mamíferos, las prostaglandi- nas bajan la presión arterial sanguínea y estimulan la contracción de los músculos lisos, pero su función en el pez no ha sido estudiada.
La acción de las gonadotropinas en las gónadas. La extirpación de la pituitaria (hipofisectomía) realizada en muchas especies de los tres grupos más importantes de peces vivientes, reduce o detiene el desarrollo gonadal, tanto en el periodo de transición desde la etapa juvenil a la de maduración sexual, como durante el ciclo estaciona1 de desove. La dependencia en la glándula pituitaria para la producción de hormonas sexuales por las mismas gónadas, ha sido, por lo tanto, claramente establecida en los peces. Las investigaciones actuales reconocen la existencia de sólo una gonadotropina funcional en los peces, para la cual ha sido propuesto el término “Piscine Pituitary Gonadotropin (PPG)” (Gonadotropina pituitaria ictiológica). La única gonadotropina de los teleósteos posee propiedades que en los mamíferos están distribuidas entre dos hormonas, LH y FSH. En una eviden- cia menos completa de la que existe para los mamíferos descansa la presunción de que la mesoadenohipófisis de los peces produce dos diferentes hormonas esti- muladoras de las gónadas.
La hormona luteinizante (LH) de los mamíferos promueve la liberación de los gametos de las gónadas casi maduras en los peces y estimula la aparición de los caracteres sexuales secundarios. De esto se infiere que hay una hormona similar en los peces mismos. Las gonadotropinas puras de la pituitaria del salmón recuer- dan al LH. Y, además, las gonadotropinas coriónicas humanas y la orina de las yeguas preñadas, que tienen efectos como los del LH, promueven la liberación de los óvulos en los peces hembra. La evidencia de la existencia de una hormona estimuladora del folículo en los peces (FSH), la segunda hormona gonadotrópica conocida de la glándula pituitaria del mamífero, descansa, hasta donde se sabe, en el ensayo patrón para el FSH de los mamíferos, realizado con extractos de la pituitaria del pez. Estos ensayos, sin embargo, no proporcionan una prueba de la existencia de una segunda hormona gonadotrópica en la pituitaria de los peces.
Las gonadotropinas o las preparaciones de la pituitaria entera de los mamí- feros, aves y reptiles, son efectivas en algunas especies de peces para apresurar el desove o para influir en las glándulas sexuales y en el comportamiento. Cuando son empleados materiales obtenidos de los anfibios, los éxitos son más frecuentes que cuando se usan los de los mamíferos. Los mejores resultados, sin embargo, se obtienen usando glándulas o extractos de las glándulas de la misma especie de pez; no obstante lo anterior, las diferencias bioquímicas interfamiliares e inter- específicas de las gonadotropinas de los peces parecen ser, en muchos casos, toda- vía más grandes que las que se observan entre los peces y los anfibios.

6RGANOS ENDOCRINOS 341
El empleo de las gonadotropinas en el cultivo de peces. En las operaciones de cul- tivo de peces puede resultar muy costoso retener reproductores potenciales du- rante largos periodos de tiempo solamente para tenerlos disponibles en cualquier momento, sin dejarlos madurar. Las implantaciones de pituitaria o las inyecciones de sus extractos son ahora ampliamente usadas para producir la liberación crono- métrica de los óvulos y del esperma. Los países en los que se practica más activa- mente la reproducción artificial mediante la inducción con pituitaria son Brasil y Rusia. La principal cosecha de peces de agua dulce en el Brasil es de caracínidos (Characidae). La técnica es aplicada en Rusia para los esturiones (Acipenser), con el propósito de obtener el caviar. El método tiene cierto empleo también para el cultivo de peces variados, incluyendo a las truchas y a los salmones (Salmoninae), y a las carpas (Cyprinidae), en Europa, Norteamética y Asia.
Para inducir la reproducción forzada, la glándula pituitaria fresca es inyec- tada intraperitonealmente después de deshacerla en una solución al 0.7% de sal (NaCl), o bien injertada in totu. Las glándulas pueden ser congeladas y utilizadas dentro de los seis meses siguientes o, como se hace con más frecuencia, pueden ser deshidratadas en acetona o alcohol, luego desecadas, en seguida reducidas a polvo, para después almacenarlas durante muchos años en contenedores cerrados herméticamente que evitan la entrada del aire. Con el polvo se hace una suspensión en solución salina acidificada para su inyección intramuscular o intraperitoneal.
Los mejores resultados se logran con glándulas obtenidas de donadores maduros o casi maduros de la misma especie, aunque también pueden ser emplea- das las de otras especies. Las dosificaciones dependen tanto del estado de madu- rez del donador como del receptor. Por lo general, los peces maduros solamente necesitan una dosis, mientras que los individuos que están por madurar pueden requerir la inyección de material a partir de toda una glándula cada segundo o tercer día, durante un mes. Es promisorio el empleo en el cultivo práctico de los peces o acuacultura del principio de estimulación de las gónadas mediante las go- nadotropinas de la glándula pituitaria. Es de esperarse que en lo futuro se logre un refinamiento de los procedimientos, luego de que la información se acumule sobre las fórmulas estructurales de las gonadotropinas de los peces y de las hormo- nas sexuales, tal como ya fue señalado por los resultados tan efectivos obtenidos al inducir la reproducción en muchas especies de peces mediante la gonadotropins altamente purificada del salmón.
La mucosa intestinal
El recubrimiento intestinal (mucosa) produce hormonas que se unen a los con- troles nerviosos para regular las secreciones pancreáticas. Las hormonas intestina- les son la secretina y la pancreocimina (originalmente las dos eran confundidas con el nombre de secretina). La secretina produce el flujo de líquidos que aca- rrean enzimas desde el páncreas, y la pancreocimina acrecenta el flujo de los zimógenos. Ambas hormonas son formadas principalmente en la porción anterior del intestino delgado. Su producción puede ser llevada a cabo mediante la intro- ducción de carne de pescado hornogeneizada y acidificada en la región pilórica del estómago de los peces carnívoros, o a través de la inyección de secretina en la vena gástrica, que estimula una secreción del páncreas. Por lo tanto, las pruebas clásicas para conocer la actividad endocrina son satisfechas por el intestino del-

342 INTEGRACION
gad0 del pez. En los vertebrados superiores existe otra hormona intestinal, la colecistocinina, que promueve el flujo de la vejiga gaseosa. Un polipéptido con actividad parecida a la de la pancreociminacolecistocinina ha sido encontrada en las células del intestino de las lampreas.
LOS SENTIDOS (ÓRGANOS DE LOS SENTIDOS) DE LOS PECES
Los órganos de los sentidos reciben estímulos físicos y químicos del medio am- biente. Los cambios físicos en el flujo del calor o del tacto son percibidos a través de los receptores de la piel; los estímulos visuales implican cambios en la inten- sidad y cualidad de la luz; y los acústicos son recibidos a través del oído interno o de la línea lateral. Los estímulos químicos son los que se experimentan a través de los órganos del olfato o del gusto. Probablemente el dolor no es experimentado por los peces, como una sensación fuerte, aunque los estímulos físicos o químicos enérgicos o nocivos producen reacciones violentas.
Los sentidos cutáneos (temperatura, tacto y gusto; Figuras 11.9 - 11.12)
La percepción de la temperatura puede ser empleada por el pez para permanecer en un medio con la temperatura de su preferencia y contribuir a la orientación cuando realiza desplazamientos largos o cortos. Sin embargo, la percepción de la temperatura solamente ha sido ampliamente investigada entre los peces óseos superiores (Actinopterygii). Mediante el entrenamiento con los reflejos condicio- nados muchas especies de peces óseos marinos y de agua dulce han mostrado capacidad para detectar cambios de temperatura de unos O.O3"C, cuidando que la cantidad de flujo de calor sea moderadamente grande (en exceso de 0.1"C por minuto), y la detección de pequeñas diferencias de temperatura se basa en la suma por área de los impulsos que resultan de las terminaciones nerviosas de las finas ramificaciones de la epidermis (Figura 11.9). Los peces pueden distinguir una elevación de temperatura a partir de un nivel bajo, pero se desconoce el me- canismo con el que logra esa discriminación. Hay evidencia de que algunos peces pueden ser más sensibles a los cambios de temperatura en sus porciones anteriores y que la sensación de calor está íntimamente ligada a la percepción del estímulo táctil o débil.
La ampolla de Lorenzini de los tiburones y rayas (Elasmobranchii), un componente del sistema de la línea lateral, responde al enfriamiento mediante una elevación y al calentamiento mediante una disminución en el grado de des- carga a lo largo de sus nervios (Figura 11.10). No se sabe si sirven como receptores de la temperatura en el animal intacto, o si está o no presente también un sentido cutáneo difuso de la temperatura.
Algunos peces son sensibles al tacto, como sucede en algunos bagres (Silu- ridae). Sin embargo, todavía hay poca evidencia sobre los receptores del tacto dérmicos y especializados, como los de los mamíferos y las aves (corpúsculos de Vater-Pacini, Krause y Herbst). En estos vertebrados superiores, cada corpúsculo táctil tiene una red nerviosa en forma de madeja incluida en una cápsula de tejido conectivo. Estructuras comparables han sido hasta este momento descritas sola- mente en las aletas de los tiburones (Squaliformes), donde actúan como propio-

LOS SENTIDOS (6RGANOS DE LOS SENTIDOS) DE LOS PECES 343
Terminací6n nerviosa libre
CBlulas de - sosten en papila gustativa
CBlula sensorial . en papila
gustativa
Figura 11.9 - Terminaciones nerviosas libres que rodean a las papilas gusta- tivas de una barbilla de esturión (Acipenser). Basado en Dogie1 en Bolk, Goppert, Kallius y Lubosch, 1938).
ceptores cuando la aleta realiza movimientos de aducción o de abducción, y en la región del hocico de algunas anguilas morenas (Gymnothorax), en las que sirven para la discriminación fina del tacto durante la selección del alimento (Figura 11.11). Algunas estructuras similares también aparecen en los radios sepa- rados de las aletas pectorales anteriores de los peces soldadito o ángel (Triglidae).
Las observaciones de la conducta y los registros eléctricos de las descargas nerviosas han mostrado que los bagres neárticos (Ictalurus) son altamente sensi- bles al tacto en la cabeza y barbas. Estas regiones poseen una red de nervios finos, delgados, profusamente ramificados, con terminaciones libres, derivada de los nervios craneales V y VII, pero no se han encontrado estructuras especializadas para el tacto en pececillos de la familia Cyprinidae, donde parece que reaccionan unidos ante los estímulos táctil y térmico. En la superficie del cuerpo hay termi- naciones libres de las raíces sensoriales de los nervios espinales, que reciben el estímulo del tacto.
En general, los peces tienen órganos especializados del gusto en la epider- mis de la boca y de los labios, en la faringe y en el hocico (Figura 11.9). Ciertos grupos de peces que viven a menudo en agua lodosa o que no se auxilian mucho de la luz para alimentarse, como los bagres (Siluroidei), los bacalaos (Gadidae) Y 10s cobítidos (Cobitidae), tienen botones gustativos adicionales en otros lugares

INTEGRACION
Figura 11.10 - Cambios de las frecuencias de impulso sobre los cambios de temperatura en una ampolla de Lorenzini (AL), órgano de la I ina late- ral (LL), y un &gano ter- minal intramuscular nervio- so (OSI). (Fuente: Sand en Grassé, 1958).
"-1"- ""
0 20 40 60 80 Segundos
del cuerpo; se ha estimado que existen más de 100,000 de estas unidades en el cuerpo de un bagre neártico (Ictalurus). Los botones son inervados principal- mente por una rama (ramus recurrens) del nervio craneal VII, pero también parti- cipan elementos de los nervios craneales IV y X en la inervación de los receptores del gusto que se encuentran en las partes exteriores del cuerpo (Figura 11.12j.
El botón gustativo del vertebrado está típicamente compuesto por células sensoriales alargadas dispuest.as como los gajos de una naranja (Figura 11.9). Los tres tipos diferentes de células de los botones gustativos del pez son células recep- toras, células de soporte y células basales. Además, hay formas transicionales o intermedias de células que pueden transformarse, ya sea en células sensoriales o de soporte. Las células sensoriales tienen extensiones cortas, en forma de pelos, que están en contacto con el agua y se extienden desde la punta del botón gustativo hasta la dermis interna donde pueden presentar una expansión en forma de botella y donde se localiza el núcleo. En las regiones especialmente sensibles, como las barbas de los bagres (Ictalurus) y de los esturiones (Acipenser), y en los labios de las anguilas congrio (Congridae) y de los anfibios, los nervios que alcanzan los bo- tones gustativos dan también lugar a una red de terminaciones nerviosas libres que se localizan en las regiones aledañas. Parece ser que el gusto y el tacto pueden tra- bajar en estrecha cooperación para la selección del alimento antes de ser ingerido.
Los botones gustativos han sido encontrados en la faringe, en la cavidad bran- quia1 y en la cabeza de las lampreas (Cyclostomata). En los tiburones y en las rayas (Elasmobranchii) los botones gustativos se encuentran en la boca y en la faringe, y estructuras parecidas a los botones gustativos, probablemente también para la percepción química, están presentes en depresiones de la superficie del cuerpo.

LOS SENTIDOS (6RGANOS DE LOS SENTIDOS) DE LOS PECES 345
Figura 11.11 - Diagrama de un presunto sensor táctil del labio inferior de una anguila morena (Gymnothorax vicinus). (Basado en Bardach y Loewen- thal, 1961).
Figura 11.12 - Distribución de la rama recurrente inervando las papilas del gusto en el cuerpo de un "loach" (Noemacheilus). (Fuente: Dijkgraaf en Grass6, 1958).
Cerebro
Barbas

3 46 I N T E G R A C I ~ N
La carpa común (Cyprinus carpio) experimenta sensaciones gustativas simi- lares a las de los mamíferos; puede percibir lo salado y lo dulce, lo amargo y lo ácido. También saborea extractos de piel de pez y otras sustancias de las llamadas sápidas. Los aminoácidos aislados o en mezclas son particularmente efectivos como estimulantes del gusto en los peces. Los peces marinos (peces angelito o vaca, Prionotus; merluza, Urophycis) con receptores del gusto o químicos en sus aletas pares modificadas también perciben las sustancias sápidas, por ejemplo, los extractos de almeja o gusanos. Al parecer perciben por el gusto los compuestos que normalmente aparecen en sus alimentos, pero sus receptores no responden a las sustancias saladas, dulces o amargas, aunque sí reaccionan ante los sabores ácidos. El examen histológico de la piel de un pececillo (Phoxinus) revela la existencia de un plexo nervioso subepidérmico que envía ramificaciones hacia la epidermis donde las fibras nerviosas se dividen y se prolongan para formar terminaciones finas. Algunas células únicas especializadas, presumiblemente quimiosensoriales, han sido también encontradas en el mismo pez (Phoxinus). Por lo tanto, las sen- saciones táctiles, las de temperatura y las químicas, están estrechamente integra- das en la piel del pez. Entre estas divisiones las más superficiales o simples son probablemente las táctiles, mientras que se cree que las más finas atienden a las sensaciones químicas y a las de la temperatura.
Visión
Las aguas naturales raramente son puras; por lo tanto, en ellas los rayos luminosos sufren múltiples dispersiones en todas direcciones. Y todavía más, las aguas son a menudo turbias, casi opacas, y en consecuencia es difícil de obtener una imagen nítida cuando se mira a través de ellas. Sin embargo, es de gran importancia bio- lógica para los peces detectar ligeros movimientos y cambios de color que signi- fiquen presa, predador, acompañamiento en un cardumen o presencia de un com- pañero. Por lo tanto, parece como si el mecanismo retinal de los peces se hubiera desarrollado hacia la percepción del movimiento y el mejoramiento del contraste, además de la formación de una imagen claramente enfocada. Las características anatómicas básicas de los ojos de los peces han sido ya descritas (Figura 3.22);aquí se agregarán ciertos detalles sobre la anatomía funcional y la visión del color y de la forma.
La córnea del ojo. En los grupos de las lampreas y de los tiburones (Cyclostomata y Elasmobranchii, respectivamente) la córnea es transparente, sin pigmentación, como ocurre en la mayoría de los peces óseos (Osteichthyes). Algunos peces óseos, como el de aleta ondulada (Amia) , las percas (Perca) y los peces viejita (La- bridae), tienen membranas corneales amarillas o verdes. La lamprea marina (Petrornyzon) tiene una córnea compuesta de dos capas de tejido separadas por una sustancia gelatinosa. Esta condición primitiva también se encuentra en los embriones de los peces superiores y en los adultos de ciertas clases con visión pobre, como las anguilas congrios (Congridae). La capa interna de la córnea se continúa en un músculo corneal que, cuando está contraído, comprime al ojo y produce una distancia de acomodación modesta. No existe en los ciclóstomos un mecanismo fino de acomodación intraocular. Una córnea de dos capas, diferente, se encuentra en ciertos lenguados (Soleu), posiblemente para proteger los ojos

LOS SENTIDOS (6RGANOS DE LOS SENTIDOS) DE LOS PECES 347
de esos peces cavadores de los fondos contra la arena y los detritos. El saltarín del fango (Periophthalmus) sale del agua durante considerables periodos de tiempo; y también posee una córnea formada de dos capas y pliegues de la piel por debajo del ojo que retienen agua y ayudan a prevenir la desecación del órgano. En los peces pulmonados (Dipnoi) la córnea está arrugada pero no hay estructuras para el humedecimiento.
El recubrimiento esclerótico. El recubrimiento esclerótico, esclera, o capa fuerte del globo del ojo, está reforzado de diversas maneras en todos los grupos de peces. En las lampreas (Petromyzonidae) la esclera es fibrosa y dura, pero en los tiburo- nes y rayas (Elasmobranchii) hay elementos cartilaginosos y también placas cal- cificadas en esta capa, para su reforzamiento adicional. La capa esclerótica en un elasmobranquio está muy vascularizada y tiene una base de tejido conectivo fibroso. El esturión y las especies afines (Chondrostei) llevan un soporte cartila- ginoso en el borde corneal de la esclerótica, así como algunas estructuras óseas. En los esturiones (Acipenser) un hueso en forma de media luna, el hueso conjunti- val de Mueller, rodea en parte a la córnea. Los peces óseos superiores (Teleos- tei) tienen recubrimientos escleróticos fibrosos y flexibles dentro de la órbita y alrededor del nervio óptico, pero debe haber elementos de soporte cartilaginoso e inclusive óseos en la porción exterior del ojo que pueden rodear a la córnea por entero. El celacanto viviente de aletas lobuladas (Latimeria) lleva reforzamientos cartilaginosos en la esclera que se engrosan hacia el nervio óptico; también hay placas esclerales calcificadas que forman un anillo pericorneal.
El recubrimiento coroideo (Figura 11.13). Los ciclóstomos tienen mucha mela- nina en el recubrimiento coroideo, especialmente hacia el polo posterior del ojo. La naturaleza brillante de la mayoría de los ojos de los tiburones se debe al ma- terial reflectante que hay en una cubierta coroidea bien desarrollada o tapetum Zucidum. El tapetum sirve para absorber una cantidad máxima de luz y devolverla después a la retina mediante la flexión dispersora de los cristales de guanina que contiene.
En los esturiones y peces relacionados (Chondrostei) los elementos vascu- lares se vuelven más prominentes en la capa coroidal del ojo, que en peces como los tiburones (Elasmobranchii), y también están presentes las capas de guanóforos.
La mayoría de los peces óseos holósteos y teleósteos, con la excepción de las anguilas (Anguilliformes) y de los grupos relacionados con ellas, tienen los vasos sanguíneos del recubrimiento coroidal reunidos en una glándula coroidal en forma de herradura, descansando entre las cubiertas coroidal y esclerótica del ojo (Fi- gura 11.13) y dispuesta alrededor de la salida del nervio óptico. La glándula tiene sus vasos sanguíneos dispuestos igual que el rete mirabile en la vejiga gaseosa (Ca- pítulo 8), para la secreción del oxígeno con tensiones más elevadas de las que se encuentran en la sangre, a fin de proveer oxígeno en abundancia a los tejidos en la vecindad de la glándula. Debe hacerse notar que la retina está cerca, que tiene la demanda de oxígeno más elevada que cualquier tejido del cuerpo pueda presentar y que la circulación retinal propiamente dicha no está especialmente bien desa- rrollada en esos peces óseos que tienen una glándula coroidal grande. Las medi- ciones hechas con electrodos polarográficos en el humor vítreo han mostrado que los peces con una rete o red coroidal bien desarrollada, como el pez azul (Poma- tornus sultatrix) y las rémoras (Remora), tienen tensiones de oxígeno intraoculeres

348
Arteria oftálmica Capilares arteriales
del cuerpo lentiforme ramificaciones
coroideo Vena retinal falciforrnes
Figura 11.13 - Disposición de los vasos sanguíneos del cuerpo coroideo en el ojo de una trucha (Salmo). (Basado de Barnett en Grad, 1958).
de 450 a 780 mm Hg, que son muchas veces más que las que tiene su sangre arte- rial. Las tensiones de oxígeno (210 mm Hg) en el ojo, más bajas que las de la san- gre arterial, son encontradas en los peces con una red coroidal pequeña, como se observa en el tautoga (Tuutogu onitis), y también en la anguila (Anguillu conger) y en los tiburones y rayas (Elasmobranchii), los cuales no tienen red coroidal y presentan tensiones de oxígeno intraoculares (10 a 20 mm Hg) menores que las del agua marina que les rodea. Por lo tanto, la glándula coroidal puede proveer a la retina de una buena reserva de oxígeno, aparte de cualquier otra función que pueda tener; se ha sospechado también que actúa como un cojín protector contra la compresión del globo del ojo.
Los guanóforos brillantes aparecen en la cubierta coroidal de muchos peces óseos (Actinopterygii) para aumentar la cantidad de luz que llega a la retina. La coroides del pez de aletas lobuladas viviente (Latimeria) tiene también una cubierta brillante de guanina.
El iris. El iris forma la pupila y controla la cantidad de luz que llega a la retina. En el ciclóstomo Morduciu m o r d a el iris carece de músculos, pero los tiburones y las rayas (Elasmobranchii) tienen elementos musculares en sus iris salpicados de gua- nina y, por lo tanto, pueden ajustar la forma de la pupila. Los grupos restantes de peces, con algunas excepciones entre los lenguados (Pleuronectiformes) y las anguilas (Anguillidae) tienen una pupila fija, ya sea circular u oval. Sus iris no presentan músculos, pero tienen algo de guanina y melanina. La pupila del pez de aletas lobuladas (Latimeria) es grande y no contráctil. Algunos teleósteos de los mares profundos carecen de iris.

LOS SENTIDOS (6RGANOS DE LOS SENTIDOS) DE LOS PECES 349
El cristalino y su acomodación. El cristalino del ojo del pez es una bolsa dura y transparente hecha de proteína no colágena. Desde el momento en que el índice de refracción de la córnea y de los humores oculares son casi los mismos que los del agua (1.33), la refracción y la formación de imágenes depende casi entera- mente del cristalino, que tiene un índice de refracción (1.67) muy elevado y efectivo. El cristalino del pez no es homogéneo pero tiene un índice refractivo actual de 1.53 en el centro a 1.33 cerca de los bordes. Una característica casi constante de los ojos del pez es la distancia desde el centro del cristalino a la re- tina, dividida por el radio del cristalino. Esta medida es de casi 2.55. El cristalino es a primera vista esférico en la mayoría de los peces óseos (Osteichthyes) y en las lampreas (Petromyzonidae). Sin embargo, los tiburones y las rayas (Elasmobran- chii) tienen un cristalino comprimido horizontalmente y en los peces que salen algunas veces del agua aparecen con modificaciones espectaculares; en el pez cua- tro ojos (Anablepidae), por ejemplo, el cristalino es piriforme para permitir la visión acuática y aérea. El cristalino está sostenido desde arriba por el ligamento suspensor y por abajo está unido a la prolongación falciforme con su músculo retractor del cristalino.
En la mayoría de los peces, la acomodación, que es el ajuste de la visión, de cerca a lejos, resulta del cambio de la distancia entre el cristalino y la retina, más que a la alteración de la forma del lente. Algunos tiburones y rayas (Elasmo- branchii) hacen el acomodamiento mediante pequeños cambios en la convexidad del cristalino. Otros son capeces de mover el cristalino por medio de una papila muscular localizada en el cuerpo ciliar y, por lo tanto, logran hacer un pequeño ajuste. Los peces de aletas con radios (Actinopterygii) muestran varios grados de acomodación del cristalino, dependiendo del desarrollo del músculo retractor del lente. De acuerdo con Pumphrey, los peces que se alimentan ayudándose de la visión, como la trucha café (Salmo trutta), tienen este músculo bien desarro- llado y poseen otras características notables en el cristalino y en la retina, las cuales sugieren que estos peces tienen una visión aguda sobre un campo visual muy amplio. Las truchas (Salmoninae) tienen de hecho la capacidad de enfocar simultáneamente en diferentes porciones de la retina objetos que están, unos distantes y otros cercanos. El cristalino de estos peces es ligeramente elipsoidal, con el diámetro mayor paralelo al largo eje del pez. El cristalino tiene en conse- cuencia dos longitudes focales. Uno es para los rayos luminosos reflejados por objetos distantes que están hacia los lados del pez y se enfocan en el centro de la retina. El otro, corto, es para objetos cercanos situados por delante, cuya luz es proyectada en la retina posterior (temporal). Y aún más, durante la acomodación el cristalino es movido sobre un plano paralelo al eje largo del pez en lugar de ha- cerlo hacia adentro. En consecuencia, la distancia entre el cristalino y la retina cen- tral permanece sin cambio, y el foco se mantiene ajustado para la visión a distancia. Al mismo tiempo, la contracción del músculo retractor del lente trae al cristalino más cerca de la retina posterior, que representa el campo de visión frontal del pez. El pez es entonces capaz, en esta porción del campo visual, de obtener imá- genes nítidas dentro de márgenes de distancias que van de unos cuantos centíme- tros hasta el infinito, o hasta el límite de la transparencia del agua.
La retinoscopía ha mostrado que muchas especies de peces óseos (Osteich- thyes) son présbitas. Los errores refractivos cambian por muchas dioptrías durante esas mediciones en los peces que forman cardúmenes, como el de lados plateados (Menidia), ciertos jureles (Carangidae) y otros, sugiriendo que esos peces pueden

350 INTEGRACIdN
desde luego acercar o alejar sus cristalinos de la retina o deformar su globo ocular para la acomsdación. Algunos peces de los fondos, como los lenguados (Parulich- thys) , y de los arrecifes, como el pez candil (Holocentrus) y los peces mariposa (Chaetodontidae), son emétropos, esto es, que las imágenes enfocan en la super- ficie de la retina. Las pruebas en las que no se ha empleado la retinoscopía se han llevado a cabo en peces de arroyos y lagos pequeños, como ciertos peces sol (Cen- trarchidae), pero las analogías que se han encontrado con los peces de los arreci- fes, por lo que respecta a la conducta y al hábitat, sugieren que también ellos podrían ser emétropos. En el “alewife” (Alosa pseudohurengus) el plano focal del ojo se encuentra a una distancia promedio de 0.2 milímetro por detrás de la retina, sugiriendo que las imágenes estáticas que este pez ve pueden no ser tan nítidas como las que observan los animales con visión aérea.
En los peces de los fondos marinos se observan cristalinos especialmente largos y tubulares y ojos dirigidos hacia arriba, como ocurre en el pez ojos de perla (Scopelarchus), que posiblemente les ayudan a recoger en la retina inferior seííales provenientes de los fotóforos ventrales de las especies compañeras, de la presa o de los predadores.
Frente al cristalino existe un pequeño espacio lleno de un humor claro y salino. La cavidad principal del globo del ojo está llena del humor vítreo transpa- rente que es secretado por el cuerpo ciliar y contiene reforzamientos tendino- sos transparentes.
La retina. LOS peces tienen retinas típicas de vertebrado, estando así formadas de muchas capas celulares parcialmente imbricadas (Figura 11.14) o unidas. Al chocar la luz con la retina después de haber viajado a través del cristalino y de los humores, las capas retinales que se le enfrentan van en el siguiente orden: a ) fibras que se dirigen al nervio óptico, b ) sus células ganglionares, c ) células bipolares que, en su turno, quedan en contacto con d ) las células fotorreceptoras propiamente dichas, conocidas como bastones y conos de la retina. En la retina de las lampreas (Petromyzonidae) ninguna distinción puede ser hecha entre los bastones y los co- nos, pero en algunos tiburones y rayas (Elasmobranchii) y en todos los peces óseos superiores (Teleostei) los bastones y los conos están bien diferenciados. En cier- tos peces óseos hay conos gemelos y se han llegado a reportar conos triples e inclusive cuádruples; no se sabe si estas agregaciones de conos, peculiares en los peces, poseen alguna función especial. En los teleósteos, pero no en los elasmo- branquios, sus retinas presentan una quinta capa e ) de células retinales, el epitelio, que contiene melanina y bordes en la cubierta coroidal.
Cuando hay luz brillante, el pigmento de melanina de la capa epitelial de la retina sombrea los bastones sensibles; en luz débil, sin embargo, la melanina se concentra cerca del borde coroidal, de tal manera que las células fotosensibles quedan completamente expuestas a cualquier cantidad de luz que esté disponi- ble (Figura 11.15). Al mismo tiempo, los elementos mioideos contráctiles situados en la base de los bastones y conos mueven los extremos sensibles de las células de tal manera que los bastones se extienden lejos del cristalino cuando la luz es brillan- te, para ser engolfados por la melanina epitelial, mientras que en la oscuridad los mioides de los bastones se contraen y acercan a las células hacia el lumen del globo del ojo. Los conos se mueven en la dirección opuesta, hacia el cristalino, cuando la luz es brillante y hacia el epitelio externo cuando es débil (Figura 11.15). Los ojos de los peces muestran estas reacciones fotomecánicas o retinomotoras en

LOS SENTIDOS (6RGANOS DE LOS SENTIDOS) DE LOS PECES 351
CURSO DE L A L U Z "------.-"----
Membrana Célula limitante bipolar
Membrana limitante
externa "Difusa"
Núcleo del cono Célula
amacrina
Fibra nerviosa
Célula ganglionar "parasol"
Célula bipolar
"centrífuga"
Célula - bipolar
"Midget"
CBlula bipolar
"Horizontal"
Capa Capa Capa Capa celular celular celular de cBlulas visual bipolar ganglionar nerviosas
Figura 11 .I4 - Diagrama de la disposición de los componentes neurol6gicos de la retina de un salmón (Oncorhynchus). (Fuente: Al¡, 19591.
forma más pronunciada que como lo hacen los ojos de los otros grupos de verte- brados. La cantidad de luz que llega a la retina puede también ser controlada mediante la contracción de la pupila. Con excepción de unos cuantos grupos, como las anguilas y los peces planos, este proceso está pobremente desarrollado en los peces. En los tiburones y especies relacionadas (Elasmobranchii) la capa fotorreceptora puede ser oscurecida por la melanina que se mueve hacia aden- tro o hacia afuera en la capa coroidal, ya que la retina carece de un epitelio pigmentado.
El número de bastones y conos conectados a un ganglio y, en consecuencia, a una fibra del nervio óptico, varía mucho en diferentes clases de peces. Los que se alimentan viendo con luz crepuscular y algunos de los mares profundos tienen muchos bastones concentrados en una fibra óptica, como una adaptación para la visión en luz tenue (adición de impulsos); en otros grupos, como en los lucios (Esocidae), se presume que confían en la buena luz para alimentarse, y tienen muchos menos receptores retinianos conectados a una neurona óptica. Los nú- meros relativos de bastones y conos también varían considerablemente en dife- rentes especies. Las proporciones cono/bastón más elevadas están en los peces que se alimentan ayudándose de la vista y son activos durante el día, mientras que mu- chas especies crepusculares han aumentado aparentemente el número de bastones, en comparación con el de los conos. Ciertos peces de las aguas medias (por ejemplo,

352
RETINA Membrana
Pigmento 1
Cono Bastón limitante r I i externa
INTEGRACI6N
A a la
,daptada oscuridad
Figura 11.15 - Diagramas de las adaptaciones de los elementos visuales a la luz y la oscuridad, de la retina de un pez óseo. (Fuente: Wunder en Herter, 1953).
Winteria y Opisthoproctus), además de tener grandes ojos con órganos luminosos adyacentes, también tienen cantidades muy grandes de bastones en las retinas.
Unos cuantos peces óseos (Osteichthyes) tienen una fovea (área retiniana de visión clara) donde los conos y las células ganglionares son más numerosos que en otras regiones de la retina. La fovea aparece restringida a los peces que viven en aguas claras y especialmente a los que pueden mover sus ojos en forma independiente o hacerlos converger para dominar un ángulo amplio dentro del cual puedan tener una visión binocular; los ejemplos incluyen a los caballitos de mar (Hippocampus) y a muchos miembros de la familia marina Serranidae.
Las retinas de los peces producen dos clases de pigmentos sensibles a la luz, la rodopsina y la porfiropsina. La rodopsina, de color púrpura, ha sido aislada de los ojos de muchos peces marinos de aletas con radios (Actinopterygii), mien- tras que las retinas de los mismos peces, pero de agua dulce, y los peces señorita o viejita, marinos (Labridae), contienen una sustancia sensible a la luz y de color rosado, la porfiropsina. Los peces diádromos como la lamprea marina (Petromy- zon marinus), las anguilas (Anmilla) y el salmón del Atlántico (Salmo salar), poseen rodopsina y porfiropsina; la lamprea (Petrornyzon), cuando migra hacia el mar, tiene retinas que producen más rodopsina, mientras que la porfiropsina predomina en los ojos de los reproductores de las aguas dulces; cambios opuestos ocurren en la anguila cuando viaja de los lagos y ríos hacia el Mar de los Sarga- z o ~ . La rodopsina y la porfiropsina son sintetizadas en la oscuridad a partir de la vitamina A. En la exposición a la luz aparece en los bastones el pigmento reti- niano amarillo que puede, sin embargo, ser empleado para reconstruir los pig- mentos púrpura o rosado sensibles a la luz. El cambio químico del pigmento, de púrpura a amarillo en los bastones, inducido por la luz, es transformado en la retina como impulsos nerviosos que viajan a lo largo del nervio óptico. Los pro- cesos bioquímicos y fisiológicos que tienen lugar en los conos y, por lo tanto,

LOS SENTIDOS (6RGANOS DE LOS SENTIDOS) DE LOS PECES 3 53
están relacionados con la visión del color, no son bien conocidos. Algunos pig- mentas con diferente sensibilidad máxima han sido aislados de las retinas de los vertebrados y han sido realizados registros eléctricos a partir de la capa de células bipolares de muchas familias de peces marinos con aletas provistas de radios. Estos hallazgos sugieren que los conos en las retinas de las especies de aguas poco pro- fundas tienen respuestas pareadas ante los estímulos luminosos de diferentes longitudes de onda. Hay una familia de elementos sensibles a la combinación de estímulos para el amarillo y el azul y otra para el rojo y el verde. Cuando los bastones son eliminados de una de esas retinas, los conos responden a la luz blan- ca, sugiriendo entonces que la visión del color en los peces está basada en un sistema de tres pigmentos. Los pargos o huachinangos (Lutjanidae) solamente responden a la luz blanca. Ellos pueden ser ciegos al color, pero debe hacerse no- tar que viven en profundidades comprendidas entre los 30 y 70 metros (cerca de 100 a 200 pies), donde la gama luminosa queda comprendida principalmente en la porción azul-verde del espectro y donde inclusive al hombre, con su visión del color bien desarrollada, le aparecen las cosas con diferentes sombras en lugar de diferentes colores.
El primer paso en la visión es fotoquímico e incluye reacciones en los pigmentos sensibles a la luz de los bastones y conos, como ya se indicó. No se sabe cómo es que estos cambios químicos se transforman en los impulsos eléctri- cos que pueden ser recogidos en la retina, ni tampoco cómo es que se transforman en señales que salen del ojo por el nervio óptico y viajan hacia el cerebro. Las señales en el nervio óptico, como sucede en otros aferentes, son de tres clases: “encendido”, “apagado” y “encendido-apagado”. Se caracterizan por presentar cambios en la frecuencia de descarga a lo largo del nervio. La retina de la carpa dorada (Carassius) tiene su umbral más bajo adaptado para responder al “encen- dido” en el azul, y para responder al “apagado” en la porción roja del espectro, indicando que este tipo de código para la función receptora permite la visión aguda de contraste. La buena discriminación del movimiento y contraste está también implicada para este pez por las siguientes observaciones. Cuando un haz de luz monocromática de 153 micras de diámetro es empleado para probar el campo receptor de un ganglio nervioso óptico que cubre un campo de cerca de 4 milímetros cuadrados, los tres tipos de respuestas ya mencionados transmiten mensajes muy detallados acerca de la dirección y avance del estímulo luminoso que se mueve (Figura 11.16). El avance diagonal desde la esquina inferior izquierda a la superior derecha da registros de la respuesta correspondiente a “encendido” como un incremento en la descarga de los impulsos nerviosos, cuando la luz apa- rece. Entonces, cuando el haz luminoso continúa su recorrido es anunciado en una forma “encendido-apagado” por descargas, ya sea que aparezca o desaparezca de cierta área del campo. Los receptores retinianos provocan en el centro un incre- mento en el grado de descarga (respuesta “apagado”) a lo largo del nervio, so- lamente cuando la luz desaparece, esto es, cuando se mueve hacia arriba. La mitad superior del recorrido diagonal muestra la secuencia inversa de los mensajes codi- ficados, a diferencia de los observados en la mitad inferior.
Los ojos de los peces pulmonados (Dipnoi) recuerdan a los de los anfibios, y no tienen poder (o es muy pobre) en sus retinas para la acomodación de la visión a distancia; tienen pocos bastones y muchos conos, así como algunos conos geme- los. El pez de aletas lobuladas viviente (Latimeria) tiene ojos grandes, retinas sin melanina y capas receptoras que constan casi exclusivamente de bastones. Y toda-

3 54 INTEGRACI6N
Figura 11.16 - Campo receptor de una cklula ganglionar aislada. La región central clara indica la parte donde solamente pueden ser obtenidas respuestas "no". El área rayada indica la región donde se obtienen las respuestas "sí-no". Las respuestas "sí" son obtenidas solamente en el área perifbrica que aparece en negro intenso. Los estímulos para la prueba tuvieron un diá- metro de 153 micras, una longitud de onda de 600 milimicras, y la intensidad de 18 micro- watts/cm2. (Fuente: Wagner, MacNichol y
+-___lmm____l Wolbarsht, 1960).
vía más, relativamente pocos bastones están conectados a cada fibra nerviosa óptica, y los ojos recuerdan del todo a los de otros peces de las aguas medias que viven en una zona océanica algo oscura.
Los mensajes codificados transmitidos por el nervio óptico son más tarde elaborados en el cerebro. Estos procesos, de los que en cualquier vertebrado in- cluyendo al hombre, se conoce muy poco, deben finalmente transformarse en un estímulo visual con algún significado para el organismo.
La visión en la vida de los peces. Muchos peces confían en la vista para capturar a sus presas, para recibir señales que le traigan o completen su conducta de unión con un compañero, y para encontrar y relocalizar su albergue, siendo éstos sola- mente algunos de los aspectos del comportamiento del pez que dependen de la vista o son influidos por ella.
El entrenamiento experimental ha mostrado que los peces de las aguas poco profundas tienen visión de los colores muy parecida a la del hombre; pueden dis- tinguir hasta vehticuatro tonalidades diferentes del espectro de banda angosta. Algunos peces, inclusive, superan el poder del ser humano para ver la porción violeta del espectro. El blanco es para los peces una sensación básicamente dife- rente a la de los colores y, además ven colores complementarios. Es muy impor- tante señalar que estos peces entrenados, en su mayoría pececillos de acuario (Cyprinidae, incluyendo al piscardo, Phoxinus phoxinus), son capaces de percibir simultáneamente el brillo y los contrastes de color. Hay también buena discrimi- nación para diferentes grados de sombras en una serie de grises. La diferencia de las respuestas eléctricas en las retinas de los pargos o huachinangos (Lutjanidae) de las aguas profundas, comparada con la de los peces de aguas poco profundas, ha sido ya mencionada, y bien puede ser que no todos, sino más bien únicamente los peces de las aguas de poca profundidad tengan desarrollada la visión para el color. Esa diferencia entre los peces podría no ser sorpresiva si consideramos que los mamíferos, siendo menor su número de especies y viviendo en un medio igual- mente variado aunque diferente, tienen también grupos con capacidad unos, y sin capacidad otros, para la visión de los colores.
A los peces que se alimentan auxiliándose de la vista, como el piscardo (Phoxinus phoxinus) y el huro (Micropterus), cuando se les da entrenamiento

LOS SENTIDOS (6RGANOS DE LOS SENTIDOS) DE LOS PECES 355
mediante reflejos condicionados en diferentes condiciones y sin significado bioló- gico inherente, pero acoplado éste en el entrenamiento con un premio o un cas- tigo, muestran una forma de discriminación de regular a buena. Los peces apren- den más rápidamente y cometen pocos errores si los objetos se mueven en lugar de permanecer estáticos.
A este respecto nosotros debemos considerar muy de cerca el comporta- miento alimentario de ciertos peces que se auxilian de la vista para comer, como los salmones del Pacífico (Oncorhynchus). En estos peces, así como en los del entrenamiento ya citado, se hace buen uso del mecanismo retiniano que es adap- tado para detectar el contraste y el movimiento. El salmón, igual que la trucha (Salmo; Saluelinus) cazan usando sus ojos y cambian la visión de los conos a la de los bastones según se va extinguiendo la luz (Figura 11.17). Dentro de un mar- gen considerado entre 0.1 y 100 o más bujías-pie de luz el pez depende de los conos para recibir su mayor información sobre los colores y sombras de su presa que se mueve; cuando han sido localizados son atrapadas y llevadas e introduci- das de cabeza en la boca. Por debajo de 0.1 y cerca de 0.0001 bujías-pie (la canti- dad de luz equivalente a la producida por una luna nueva, clara, durante la noche), los bastones intervienen, la alimentación se detiene y los cardúmenes se rompen. La reacción ante el estímulo luminoso en estas intensidades tan bajas sugiere más tarde que los peces pueden usar su vista todavía a profundidades más bajas, tan considerables como los 700 metros o más en aguas oceánicas claras. En los expe- rimentos en los que las pulgas de agua (Daphnia) son dadas como alimento de los salmones jóvenes en intensidades bajas de luz, los peces toman una posición obli- cua por debajo de su presa para verla entonces contrastando al máximo con la pantalla formada por la superficie del agua, la observan en sus movimientos du- rante unos momentos y luego las cogen una por una.
La visión binocular que existe sobre ángulos variados en diferentes espe- cies es importante para juzgar la distancia y la profundidad. Algunos peces, como los lucios (Esox) , tienen verdaderos surcos visuales en sus hocicos frente a sus ojos. Dos líneas imaginarias proyectadas desde los surcos para cruzar la cabeza en frente de sus ojos deberán incluir el ángulo dentro del cual una especie particular
Umbral de los bastones los cardúmenes se dispersan
se alimentan al mínimo la luz más tenue a la que se
NO se I
Umbral de los conos
Bastones produce el máximo de ,dscóptica,,) alimentación
alimenta I,-*-, ,"-, I Desplazamiento hacia abajo Conos (visión fotóptica)
h h , , I / I \/ I I \ ,
IO-^ 10-4 I 10-3 1 IO-' loo lo1 Intensidad de la luz en bujías-pie I
Noche con luz de luna nueva - I
Ocaso y penumbra Día brillante de verano
Noche de luna llena Día nublado
Figura 11.17 - Diagrama que sumariza la visión por los bastones y conos bajo la acción de diferentes intensidades luminosas junto con algunas otras respuestas en un salmón (Oncorhynchus) sometido a diferentes intensidades luminosas. Las intensidades de la luz bajo condiciones naturales tambih son indicadas para hacer la comparación. (Fuente: Ali, 1959).

3 56 INTEGRACION
tiene imbricado el campo visual y, por lo tanto, presumiblemente el campo en el que el animal tiene la visión tridimensional. Muchos peces pueden mover sus ojos independientemente y ajustar esta porción de su área visual. Los peces de las co- rrientes, como la mayoría de las truchas (Salmoninae) y muchos pececillos (Cy- prinidae), fijan su posición en la corriente marcando visualmente una piedra, leño u otro objeto.
La luz incidente y la reflejada por el fondo, yendo a dar a las porciones supe- rior e inferior de la retina, inicia el ajuste cromatofórico del color del cuerpo contra el fondo. La luz que llega al ojo desde arriba, junto con los receptores de la grave- dad localizados en el oído interno, mantiene al pez en la posición apropiada para nadar o descansar. Además del tratamiento anterior de la visión debe hacerse mención sobre las superficies de la piel sensibles a la luz en las lampreas y los mi- xinos (Cyclostomata). En la piel de la región dorsal caudal de la larva ammocetes de las lampreas (Petromyzonidae) y de la región cloacal de los mixinos (Myxine) hay células sensitivas redondas con pigmentos sensibles a la luz, de reactividad máxima a las longitudes de onda de 500 a 530 milimicrones. Estas células están inervadas en las lampreas mediante ramificaciones de los nervios de la línea lateral y su función consiste en aumentar el valor de las reacciones fototrópicas negativas del animal. La epífisis y la región dorsal del cerebro son también sensibles a la luz en varios peces.
El sentido del olfato
La mayoría de las sustancias odoríferas que desempeñan un papel biológico im- portante en la vida de los peces son de naturaleza orgánica. Ellas son detectadas a través del epitelio olfatorio, conectado al nervio olfatorio del saco nasal (Figu- ras 3.21, 11.18 y 11.19), aunque algunas de ellas son percibidas a través de los botones gustativos. Sin embargo, los espectros de la sensibilidad para el gusto y el olfato son diferentes para las mismas sustancias y algunas moléculas orgánicas son olidas solamente mientras que otras son únicamente gustadas. La sensibilidad del sentido del olfato sirve también para la orientación, cuando es seguido el aumento de la intensidad odorifera.
Los orificios nasales, así como toda la estructura nasal, presentan entre los peces amplias variaciones en sus adaptaciones. El Único orificio nasal medio de las lampreas y de los mixinos (Cyclostomata) conduce a un tubo nasal corto que se dobla hacia atrás y termina en una bolsa nasofaríngea al nivel de la segunda abertura branquia1 interna. La bolsa es sometida a la acción de los movimientos respiratorios rítmicos que producen un intercambio de agua, en ella y en su saco nasal tributario. En los mixinos (Myxiniformes) la bolsa nasofaríngea se comu- nica con el tubo digestivo, pero en las lampreas (Petromyzonidae) termina en un ciego. El saco nasal, ramificándose a partir del tubo nasal, está protegido por una válvula que dirige un chorro de agua hacia adentro durante la inspiración, pero permite que se produzca un €lujo en sentido inverso cuando se vacía la bolsa nasal como respuesta al movimiento expiratorio de las branquias. La cámara nasal propiamente dicha está dividida en veinticinco pliegues en la lamprea marina (Petrornyzon marinus) y cubierta con epitelio olfatorio en el que un pliegue se enfrenta a otro; la porción restante de la superficie de cada pliegue está cubierta con epitelio indiferente, es decir, no sensitivo. Coma sucede en todos los demás

LOS SENTIDOS (6RGANOS DE LOS SENTIDOS) DE LOS PECES 357
Borde del Orificio nasal -, orificio nasal
olfatorio \ " Bolsa
nasofaríngea
Figura 11.18 - Boceto de la restauración en cera del órgano olfatorio de una lamprea marina (Petromyzon marinus). (Basado en Kleere- koper y van Erkel, 1960).
Figura 11.19 - (a) Posición y extensión, y ( b ) sección, del órgano olfatorio de una anguila (Anguilla). Las flechas indican la dirección del flujo del agua. El pedúnculo del orificio nasal introductor es erbc- til. (Fuentes: (a) von Frisch en Grassé, 1958; (6) Wunder, 1936).
Orificio nasal expulsor
Orificio nasal introductor
"
/ Y Cápsula
Pliegues del
nasal epitelio olfatorio sensorial

358 INTEGRACION
vertebrados, las células sensitivas del epitelio olfatorio dan nacimiento a las fi- bras del nervio olfatorio.
En los tiburones y en las rayas (Elasmobranchii) los dos orificios que con- ducen al saco nasal plegado internamente están usualmente en el lado ventral del hocico y divididos por un pliegue de la piel en una entrada y una d i d a ; la tihima lleva algunas veces hacia la boca por medio de un surco nasal cubierto de piel y a medida que el pez nada y respira, el agua llega al epitelio olfatorio. Ciertos tiburones dependen en gran medida de su sentido del olfato para encontrar el alimento y persiguen a los aumentos de grado de olor nadando en circunferencia o siguiendo un recorrido en forma de ocho.
Los órganos olfatorios pares de los peces de aletas con radios (Actinopte- rygii) son relativamente más pequeños que los de los elasmobranquios pero tienen una más amplia variedad de adaptaciones para la entrada y el intercambio del agua. Las aberturas nasales anteriores están generalmente por delante y algo por enmedio de los ojos. Las aberturas llevan directamente, o a través de un tubo corto, a la cavidad nasal con su roseta de epitelio olfatorio plegado. En la mayoria de los peces de aletas con radios las aberturas nasales posteriores están normal- mente muy cerca de las anteriores; sin embargo, en las anguilas (Anguilla; Fi- gura 11.19) y en las morenas (Gymnothorax), las aberturas están relativamente muy separadas. Algunos peces, como los del género Cottus, y las espinochas (Gmterosteus), tienen solamente una abertura nasal y la bolsa nasal es alternati- vamente llenada y vaciada por los movimientos de la respiración. Una anguila (Anguilla) tiene un tubo ciliado proyector que conduce el agua de la abertura nasal anterior, sobre los pliegues del saco nasal ovoideo, y hacia afuera de las aberturas nasales posteriores, cerca de los ojos (Figura 11.19). En las sardinas (Clupeidae) y en los bagres (Siluroidei) existen senos linfáticos cerca del saco nasal que actúan como un sistema hidráulico, y llenan y vacían el saco nasal a medida que la presión es transferida hacia las bolsas accesorias durante la respi- ración, gracias a los movimientos de muchos de los huesos de la cabeza. En mu- chos peces, teniendo al lucio (Esox) y al bacalao (Gadus) como ejemplos, el agua es desviada hacia la cavidad nasal mediante los movimientos de natacih. La sepa- ración entre nariz anterior y nariz posterior está dada por un pliegue eréctil que conduce el agua hacia adentro, hasta el centro de la roseta olfatoria.
El mecanismo fisiológico de la recepción del olor no se ha entendido por completo. Los registros hechos con electrodos colocados en unidades únicas del tracto olfatorio de los peces Abrumis brama (brema europea) y Carassius auratus (carpa dorada) muestran lo siguiente: una fibra exhibe diferentes modelos de actividad cuando son ensayados diferentes olores, diferentes fibras responden al mismo aroma de diferentes modos (codificación espacial), y la mayor parte de las fibras exhiben adaptación al seguir la estimulación continua con el mismo olor.
Muchos experimentos han mostrado la sensibilidad y gran importancia del sentido del olfato para los peces cuando buscan su alimento y se orientan. LOS pececillos de nariz roma y cabeza grasa (Pimephales) pueden distinguir mediante el olfato entre los lavados de especies relacionadas de invertebrados y los de plantas acuáticas. Los peces que se alimentan auxiliándose del olfato, como los bagres (Ictalurus), algunos tiburones (por ejemplo, Squalus) y las morenas (Gym- nothorax), no pueden encontrar su alimento cuando tienen sus narices tapadas. Las feromonas, “sustancias especiales” producidas por la presa y percibidas por el sentido del gusto del pez, incluyen a los glicerofosfolípidos y también a los

LOS SENTIDOS (6RGANOS DE LOS SENTIDOS) DE LOS PECES 359
mono y difosfatos nucleótidos. Los pececillos de nariz roma y cabeza grasa pue- den ser entrenados para detectar clorofenoles en el agua, a una concentración muy por debajo de la que puede ser ensayada por procedimientos químicos. Se ha reportado a una anguila (Anguilla) detectando una sustancia pura orgánica a una dilución de 2 X 10 -’ ’ cuando solamente 3 o 4 moléculas del material pudie- ron haber encontrado su camino hacia el saco nasal del pez. Por consiguiente, no es de sorprender que las mojarritas (Lepornis) desplazadas de su territorio nativo en una corriente, puedan encontrarlo de nuevo auxiliándose de su sentido del olfato, y que los olores de las corrientes parecen desempañar un papel muy im- portante y decisivo para guiar en su viaje de retorno a los salmones (Oncorhyn- chus) a sus corrientes nativas. Los pececillos de acuario (Cyprinidae) producen una sustancia de alarma, del grupo de las feromonas (Schreckstoff), cuando son asustados o molestados. Cuando esta sustancia es olida por los otros miembros de un cardumen, produce su dispersión. Los peces con olfación aguda como los bagres (Ictaluridae) emplean su sentido del olfato en sus relaciones sociales, haciendo la diferenciación entre los individuos de un grupo o jerarquía social por medio del olor específico de la mucosa de cualquier pez.
El oído interno (equilibrio y audición; Figuras 3.24,3.25 y 8.12)
Los órganos del equilibrio y de la audición han sido divididos en un par superior y un par inferior. El par superior está compuesto de los canales semicirculares y sus ámpulas, y una vesícula en forma de saco, el utrículo. El par inferior, o estruc- tura para la recepción propiamente dicha del sonido, consta de otras dos vesícu- las, el sáculo y la lagena. Las ámpulas contienen “manchas” de tejido sensitivo, los cristales estáticos, con células sensitivas que recuerdan a las de la línea lateral y hasta cierto punto también a las de los botones gustativos. Sin embargo, los cris- tales tienen pelos sensitivos largos en los ápices de sus células, más largos que los de los botones gustativos y están cubiertos por una cofia gelatinosa (Figura 11.20). El desplazamiento del cuerpo en cualquier dirección produce fuerzas barredoras a las que las células sensitivas de las ámpulas reaccionan. Las células tienen descar- gas expontáneas y rítmicas. Cuando la cúpula o cofia se dobla en una dirección se produce una disminución en la actividad rítmica del receptor, mientras que cuando se dobla en la dirección opuesta, se acelera el grado de descarga. Este es el mecanismo para la detección del desplazamiento angular.
El utrículo contiene, en los peces óseos (Osteichthyes), un otolito, o piedra auricular, llamada el lapillus (Figura 3.24). Este y los otolitos del sáculo y de la lagena están formados de carbonato de calcio y crecen discontinuamente, por lo menos en los peces de las zonas templadas, y pueden por lo tanto servir para calcular la edad del pez. El lapillus descansa horizontalmente sobre los pelos de las células sensitivas de los cristales de los utrículos. El lapillus responde a la fuerza de gravedad y activa las células sensitivas de los cristales. Ellas trabajan en unión de la porción inferior de la retina para el mantenimiento del equilibrio. Por lo tanto, teniendo a los ojos y al utrículo intactos, la luz desde arriba y la gravedad desde abajo empujando y jalando, como si así fuere, el pez se mantiene en una posición derecha. Después de la extirpación de un utrículo, o después de hacer brillar un fuerte rayo de luz hacia el ojo de un pez, formando ángulos rectos en lugar de dirigirlos desde arriba, puede lograrse que el pez se incline hacia un lado 0 hacia el otro. Cuando a un pez se le ha eliminado el par superior de los dos oídos inter-

360
EPITELIO SENSORIAL
INTEGRACI6N
Canai de la línea
lateral
Neuromasto libre
Neuromasto en la línea
lateral
Cresta en una ampolla de un
canal semicircular
/" Otolito
Células sensitivas bajo un otoi i to
en el oído interno
Figura 11.20 - Comparación diagramática de los diferentes epitelios sensoriales del sistema actisticolateral. (Fuente: Dijkgraaf en Grassé, 1958).
nos, se le puede hacer nadar en un ángulo de 90" e inclusive invertido, es decir, con el vientre arriba y el lomo por debajo, si se le ilumina un lado o la porción inferior (Figura 11.21). El peso total del lapillus, descansando sobre las células que poseen los pelos sensitivos, y sin molestarlos o doblarlos, viene a ser el esti- mulo utricular adecuado para la natación en posición derecha. Un desplazamiento de 90" del pez hará perder peso al lapillus pero producirá un movimiento rasante máximo en los pelos de las células sensitivas, bajo la acción del obolito. El oto- lito lagenar y ei tejido sensitivo que se encuentra por debajo, son, en un grado pequeño, relacionados también con los reflejos gravitatorios o de la posición derecha, principalmente a través de su influencia en el tono muscular.
En los tiburones y rayas [Elasmobranchii) hay un conducto endolinfático que comunica el oído interno con el exterior. A través de este conducto, la arena llega a las &pulas gelatinosas, sobre las células sensitivas del oído interno. Sin embargo, no se sabe hasta qué grado esas partículas que provienen del exterior colaboran en la estimulaciór, de las cklulas sensitivas, si se toma en cuenta que también existen en el oido interno de los tiburones concreciones calcáreas en €orma de otolito, de origen end6geno.
El tamaño y la disposición de los conductos semicirculares y de las vesículas son considerablemente diferentes entre varios peces. Los canales son relativamente grandes en los peces elefante (Mormyridae) y en ciertos peces cavernícolas ciegos (Amblyopsidae). El sáculo y la lagena están más ampliamente unidos uno al otro en los tiburones (Squaliformes) quo en los peces óseos (Osteichthyes).

LOS SENTIDOS (6RGANOS DE LOS SENTIDOS) DE LOS PECES 361
Figura 11.21 - Efectos de la retina del ojo y el /api//us del utrículo del oído interno en el equilibrio de los peces. Cuando los lapilli son removidos, el pez ya no se llega a orientar con relaci6n a la gravedad y la luz juntas; solamente lo hace de acuerdo con la luz presente. (Basado en von Holst, 1950).
El desarrollo relativo de las porciones superior e inferior del oído interno también difieren en los peces de aletas con radios (Actinopterygii) de hábitos diferentes. Los peces voladores y los pelágicos de rápida natación que requieren de una orientación tridimensional superior, tienen un par superior mucho me- jor desarrollado que las especies que se sumergen, donde por turno la porción inferior alcanza su más alto grado de desarrollo.
El par inferior del oído interno, la lagena y el sáculo con sus otolitos, lla- mados asteriscus y sagitta, respectivamente, son el asiento de la recepción del sonido. Estas dos estructuras, así como el canal semicircular posterior, son iner- vadas por la rama posterior del nervio auditivo (nervio craneal VIII) mientras que la parte superior del oído interno y los otros dos canales semicirculares est6n conectados a l a rama anterior del mismo nervio. Los otolitos de la lagena y del sáculo estan sostenidos verticalmente sobre la mancha sensitiva acústica, mediante ligamentos tendinosos, a las paredes internas de las vesículas.
El mecanismo fisiológico para la discriminación de las diferentes frecuen- cias de sonido no es conocido, pero su principal asiento está en el utrículo y uno debe distinguir entre dos tipos de estructuras auditivas entre los peces óseos, los tipos común y el cipriniforme. Entre los pececillos de acuario, los bagres y las car- pas chupadoras (Cypriniformes), el oído interno está conectado en forma única a la vejiga gaseosa a través de una cadena de huesecillos, los osículos de Weber (Figura 3.25; Capítulo 8). Los Cypriniformes tienen un margen más amplio y fino para la audición que los peces del tipo normal; tienen su lagena y sáculo divididos por una estrecha constricción y su sagitta lleva hacia la endolinfa un ala grande, aparentemente mucho mejor para recoger las vibraciones que son trans- feridas a ella desde la vejiga gaseosa que actúa como resonador.
Nada se sabe sobre la amplitud y agudeza de la recepción del sonido en las

36 2 INTEGRACIdN
lampreas y en los mixinos (Cyclostomata). Los tiburones (Squaliformes) reaccio- nan ante varios ruidos y sonidos, algunos de frecuencias puras comprendidas entre los 400 y 600 ciclos por segundo. Los peces de aletas con radios (Actinopterygii) pueden percibir tanto las diferencias de intensidad como un margen de vibracio- nes que va de los 16 a los 13,000 ciclos por segundo (cps), como ocurre en los bagres (Ictalurus). Las frecuencias bajas, comprendidas entre los 16 y 100 ciclos son probablemente sentidas a través de la piel y/o los receptores de la línea late- ral. El nivel más agudo de percepción del sonido en el pez gordo de los arroyos (Semotilus atromaculatus), queda cerca de los 28 cps; este pez también es capaz de distinguir intervalos de cuarto de tono entre los 400 y 800 cps. Sin embargo, la verdadera audición direccional parece estar limitada a sólo unas cuantas doce- nas de metros. El margen superior de la percepción del sonido en los peces que carecen de órganos accesorios queda cerca de los 5,000 cps, como sucede en las percas trepadoras (Anabantidae), y ellas tienen menos capacidad para distinguir los intervalos entre los tonos.
La línea lateral (Figuras 11.22 - 11.24) El sistema de la línea lateral está formado por un juego de órganos sensitivos restringidos a los peces, y a los anfibios en sus etapas acuáticas de crecimiento. Aunque tiene conexiones en el cerebro con los recorridos nerviosos aferentes del sistema auditivo del oído interno, es en realidad un sistema diferente. Ya existía en los ostracodermos fósiles, y se piensa que se originó de células sensitivas cutá- neas primitivas. L a evidencia fósil desde Pteruspis (Agnatha: Pteraspidomorphi)
Canal de la Poro linea lateral
I / Epidermis -
/ \ \ i Músculo
Miosepto Escama
Neuromasto
-- Nervio
Figura 11.22 - Neuromastos en el sistema de la línea lateral de una anguila japonesa (Rhynchocymba). (Basado en Katsuki, Mizuhira y Yoshino, en Grad , 1958).

LOS SENTIDOS (6RGANOS DE LOS SENTIDOS) DE LOS PECES 363
hacia adelante, permite saber algo sobre la ruta seguida por la evolución de la línea lateral, pero la evidencia embriológica es menos clara debido a que en algu- nos peces la línea lateral se origina en la plácoda Ótica o auditiva mientras que en otros, como por ejemplo en un pececillo de Europa (Leuciscus), conocido con el nombre de cacho, se forman en la cabeza y el cuerpo muchas plácodas separa- das para sendas líneas laterales.
Los peces óseos (Osteichthyes), además de tener los canales típicos de la línea lateral (Figura 11.22) con sus poros en la piel o escamas en la cabeza y el cuerpo, sólo se les consideran las siguientes estructuras sensitivas ampliamente diferen- ciadas, tentativamente comprendidas en el complejo de la línea lateral:
a. Criptas sensoriales o cavidades y papilas entre o por debajo de las esca- mas placoideas de los tiburones y rayas (Elasmobranchii), dispuestas en grupos o hileras, usualmente en las regiones anterior y dorsal del cuerpo; ciertas peculiaridades histológicas de sus células sensitivas sugieren que ellas son receptoras de la gustación (Figura 11.9) más que una parte del sistema de la línea lateral propiamente dicho.
b. Las ampollas de Lorenzini (Figura 11.23) son estructuras que tienen más o menos forma de saco en la región cefálica de los tiburones y rayas (Elas- mobranchii) y también de Plotosus, un bagre del sureste de Asia, donde varios grupos de poros (supraorbital, bucal, hioideo y mandibular) dan acceso a través de un canal. Las estructuras están llenas de un material gelatinoso y pueden tener desde unos cuantos hasta muchos divertículos cubiertos de epitelio sensitivo. Las células sensitivas están inervadas por ramificaciones del nervio facial (VII). Las preparaciones aisladas de esas ampollas dan descargas nerviosas rítmicas que son aceleradas con el frío y menguadas con el calor. También son sensibles a los estímulos mecánicos y los eléctricos débiles, así como a los cambios pequeños en la salinidad. Su mayor sensibilidad corresponde, sin embargo, a las diferencias en el potencial eléctrico, como el que podría ser generado por los movimientos musculares; por ejemplo, al abrir y cerrar las aberturas branquiales. Actualmente se cree que las ampollas de Lorenzini son principalmente receptoras eléctricas que ayudan en la localización de la presa, así como para llevar a cabo las funciones propioceptivas durante la natación. Su sensibilidad es extremadamente elevada, reaccionando a un cambio de 0.01 microvoltios/cm de agua de mar, como lo muestra la raya europea con espinas en el lomo (Ruja cluuatu) en la que la proporción de palpita- ciones cambia cuando es estimulada con esos pequeños cambios del potencial eléctrico. Los tiburones gato (Scyliorhinus) presentan una sensibilidad similar, indicando que ellos, y probablemente otros elasmo- branquios, son capaces de detectar la presencia de presa potencial si los músculos de estos animales que merodean en los alrededores presentan cambios en su potencial eléctrico.
c. Las vesículas de Savi, que se encuentran en varios géneros de rayas (Raji- formes), incluyendo a ciertas rayas eléctricas (Torpedo), principalmente por debajo del hocico como series lineales de bolsitas llenas de material gelatinoso que comprende corpúsculos grises, granulares y amorfos. Estas vesículas son inervadas por una ramificación del nervio craneal trigémino (V), pero su función es desconocida.

364
m E Y X al
INTEGRACIdN
3 c
U oi

LOS SENTIDOS (ORGANOS DE LOS SENTIDOS) DE LOS PECES 36 5
d . Los órganos de Fahrenholz son orificios que se encuentran en la región cefálica de los peces pulmonados larvarios (Dipnoi). Estos orificios tie- nen células sensitivas con pelos y recuerdan a los de los órganos de la línea lateral; su función, sin embargo, es desconocida.
e. Los espiráculos, estructuras sensoriales del tipo básico de la línea lateral, son de función desconocida en los Chondrichthyes, los Holostei y los Chondrosbei.
f . Las células sensoriales cutáneas de los peces elefante y las anguilas gim- nótidas (Mormyridae y Gymnotidae, respectivamente), de las que se presume son electrosensitivas.
Entre todos los animales, solamente algunos peces tienen órganos que gene- ran una descarga eléctrica. El voltaje de esta descarga varía considerablemente en los diferentes grupos de peces, como sucede también con el origen y la naturaleza de la estructura que genera la corriente. Se pueden distinguir dos grupos: 1) los que producen una corriente tan fuerte que aturde, como la anguila eléctrica (Elec- trophorus), las rayas eléctricas (Torpedo) y el bagre eléctrico (Mulupterurus); y 2) los que producen corrientes de bajo voltaje (Mormyridae y Gimnotidae, con la excepción de Electrophorus). El segundo grupo emite una serie continua de pulsos que forman un campo eléctrico alrededor del animal. El campo se distor- siona cuando las presas entran en él o cuando lo obstruyen los obstáculos inani- mados, como sucede durante la natación. El dispositivo permite la localización de objetos en el hábitat turbio de estos peces. El número de descargas por segundo varía con la especie, pero en los gimnótidos (Gymnotidae) de América del Sur y en los peces elefante de África (Mormyridae), ciertas especies producen las descar- gas con frecuencias relativamente bajas de hasta 200 ciclos por segundo, mientras que otras las emiten hasta de 1,600 impulsos de bajo voltaje, por segundo. Los grados de descarga específica para cada especie puede desempeñar un papel muy importante en el reconocimiento de los compañeros.
La generación de un campo eléctrico para la percepción de los objetos de los alrededores implica la existencia de receptores eléctricos. En los peces elefante hay poros, concentrados en la región cefálica, con canales angostos que condu- cen hacia orificios aislados, únicos. Cada orificio tiene una pequeña superficie de tejido que recuerda a los receptores de la línea lateral. Estas estructuras son reno- vadas a lo largo de la vida del animal, como sucede con las demás células de la piel. Ellas pueden responder a los estímulos químicos y a los de la presión, ade- más de ser sensibles a los impulsos eléctricos. La sensibilidad del mormírido Gym- narchus niloticus a las cargas de electricidad estática es de 0.15 microvoltios por centímetro cuadrado. Los gimnótidos tienen órganos de la piel semejantes, com- puestos de muchas células sensoriales que están contenidas en una vesicula que se abre hacia el exterior a través de un canal lleno de gelatina. Las células no sensoriales pueden también aparecer en estas estructuras que pertenecen al sistema de la línea lateral. Se presume que las estructuras actúan como receptores eléctricos. LOS bagres (Ictulurus), como se ha podido ver, también responden a la estimula- ción eléctrica débil, estando localizada su sensibilidad en los pequeños órganos de los orificios del sistema de la línea lateral. En los restantes grupos de peces, 10s órganos del sistema de la línea lateral propiamente dicho pueden estar estructu- ralmente dispuestos como se indica a continuación:

366 INTEGRACIdN
1. Libres, dispuestos en grupos en la cabeza y alineados a lo largo del cuerpo (el piscardo, Phoxinus; el “loach”, Cobitis).
2. Sumidos en agujeros, en grupos, o en varias hileras en la cabeza o a lo largo del cuerpo (muchos elasmobranquios, ciclóstomos) o en bandas que cru- zan la superficie ventral (elasmobranquios, ciclóstomos).
3. En canales parcialmente abiertos o cerrados, desarrollados de diversos modos (Figura 11.24), localizados a. en la cabeza, donde hay típicamente tres canales principales, el su-
praorbital, el infraorbital y el hiomandibular, con ramas, extensiones o conexiones, como el canal comisural supratemporal que conecta el sistema de canales a los dos lados del cuerpo;
b. en el cuerpo, donde hay típicamente un canal que se extiende desde el cleitro de la cintura pectoral a la base de la aleta caudal.
Tanto el canal de la línea lateral del cuerpo como el de la cabeza exhiben muchas modificaciones. Por ejemplo, el canal del cuerpo puede ser extremadamente corto, como ocurre en la bermejuela europea (Rhodeus amarus); de longitud intermedia, como en el pececillo de cabeza grasa (Pimephales promelas); interrum- pida, como en el pez loro (Scams), teniéndola fuertemente curvada hacia el dorso o descurvada hacia el vientre. Puede haber también muchas líneas laterales dis- puestas burdamente en paralelo, como en el pez verdoso (Hexagammus) y en la lisa (Mugil), o algunas distribuidas en segmentos cortos sobre la mayor parte del lado del cuerpo, como en el lucio (Esox) .
Las diferentes partes de la línea lateral están inervadas por los nervios cra- neales VII, IX y X y tienen asociaciones centrales muy cercanas con los centros auditivos surcados por el nervio VIII. Los centros en la médula oblongada del cerebro están divididos en un grupo dorsal para las líneas laterales de la cabeza y en un ganglio ventral para el sistema de la línea lateral del cuerpo. Conexiones secundarias en el cerebro desvían impulsos del sistema de la línea lateral al tála- mo, el hipotálamo, el tecto óptico (especialmente en los tiburones, Squaliformes) y el cerebelo (particularmente en los Mormyridae). Las células gigantes de Mauth- ner, con su recorrido final común en calidad de activadores grupales de la mus- culatura del tronco y de la cola, tienen prolongaciones axonales que las ponen en contacto directo con las ramificaciones de la línea lateral de los nervios VII, IX y X, así como con el tecto óptico, el sáculo y los centros locomotores del cerebelo.
La unidad receptora del sistema de la línea lateral es el neuromasto, una superficie de tejido sensorial formado por células receptoras piriformes, cada una con una extensión en forma de pelo en su ápice, que llega hasta una cúpula ge- latinosa (Figura 11.22). A diferencia de las células sensitivas de los botones gusta- tivos, a las que se les parecen, las células piriformes no alcanzan a llegar a la base del neuromasto pero están acojinadas por debajo por células de soporte, no sensitivas.
Igual que como sucede con los órganos terminales del equilibrio en los canales semicirculares, los neuromastos presentan una descarga rítmica continua (Figura 11.25). En las mantarrayas (Raja) pueden ser aislados dos tipos de recep- tores mediante la perfusión o baño con un chorro de agua de unas cuantas unida- des funcionales. Un grupo responde con un fuerte incremento en la frecuencia de la descarga a un chorro lento de agua dirigido hacia la cabeza, mientras que la perfusión dirigida hacia la cola inhibe la actividad de la descarga. El final de la per-

LOS SENTIDOS (6RGANOS DE LOS SENTIDOS) DE LOS PECES 367

368 INTEGRACI6N
l i l l l l l l l l l l l l l l l l l l l 0.2 sec “ 3
Figura 11.25 - Descargas eléctricas en un nervio de la 1 ínea lateral de una mantarraya (Raja c ha t a ) con flechas que indican el principio de la esti- mulación lograda con un chorro de agua: (a) chorro de agua dirigido a la cabeza; (b) corriente de agua hacia la cola. (Basado en Sand y Dijkgraaf en Grassé, 1958).
fusión dirigida a la cabeza es marcado por un periodo de silencio, pero la interrup- ción del chorro del agua hacia la cola produce una reanudación vigorosa de la actividad nerviosa; el segundo grupo de receptores se comporta a la inversa. Los receptores antagónicos y que producen descargas rítmicas están mezclados al azar en cada grupo de células sensoriales, pero también se han encontrado en las rayas, unidades de “silencio expontáneo” en el sistema de la línea lateral que pueden ser inducidas a emitir impulsos durante la perfusión en una dirección y, al terminar el baño, en la otra. En algunas anguilas japonesas (Anguillu japonica y Rhyncho- cymba nystrani) las células sensitivas del centro de los neuromastos no presentan descarga de reposo y se adaptan rápidamente, en contraste con las células perife- rales que tienen un grado de adaptación muy lento y una descarga expontánea. Las células centrales están conectadas con fibras nerviosas sustanciales de 15 mi- tras de diámetro, mientras que las células periféricas están conectadas a fibras pequeñas de 4 micras de diámetro.
Los mecanismos anatómicos y fisiológicos que refuerzan la detección de los desplazamientos hidrodinámicos de la presa, predadores o acompañantes sociales, son: las células sensitivas de los neuromastos, que tienen sus cilios en forma de pelo y constan de un kinocilio elástico y numerosos estereocilios más blandos (de 40 a 50 en L ~ t a , el “burbot”), y las células sensitivas adyacentes o con vellosidades, que tienen sus kinocilios orientados en direcciones opuestas, alternándose hacia la cabeza o la cola del pez, pero siempre dispuestas a 10 largo del canal de la línea lateral. Cuando la flexión de la cúpula se dirige hacia la cabeza o la cola afectará en forma distinta a los kinocilios de diferentes células con “pelos” y nosotros podremos distinguir la forma en que se producen las descargas del nervio de la línea lateral para conducir al cerebro información exacta sobre la dirección y/o procedencia del desplazamiento del agua en las cercanías del pez (Figura 11.25).
Los experimentos para el entrenamiento, las operaciones para eliminar la recepción en la línea lateral y las investigaciones fisiológicas, muestran que el sis- tema responde a una gran variedad de estímulos. La presión irregular de las ondas y las vibraciones de baja frecuencia, de 50 a 100 ciclos por segundo (cps), excitan a los receptores de las fibras más largas de la línea lateral de los bagres (Ictalurus), mientras que un flujo de agua débil, pero regular, o vibraciones de 20 a 50 cps, ac- túan en los receptores conectados a los nervios más delgados. Muchos peces han

REFERENCIAS 369
sido entrenados para localizar pequeños bastones o discos vibratorios cerca de sus cabezas y lados, así como para reaccionar ante pequeñas corrientes de agua loca- lizadas. Después de hacer la difícil operación de seccionar solamente los nervios de la línea lateral en la cabeza y en el tronco, y no los nervios del sentido del tacto, se pierde la capacidad de discriminación fina de las vibraciones pequeñas o de las alteraciones de la corriente.
Se puede concluir que el sistema de la línea lateral informa al pez sobre las alteraciones localizadas causadas por corrientes pequeñas, por vibraciones mecá- nicas por debajo de los 100 cps, o por el desplazamiento calmado pero irregular del ag;a circundante. El sistema está también relacionado con la localización “tacto a distancia” de los objetos móviles, como los predadores o la presa, o en la percepción de los objetos fijos que reflejan los movimientos del agua traídos por el pez hacia sí mismo mediante la natación. Los órganos de la línea lateral tam- bién reaccionan ante los cationes monovalentes (por ejemplo, Na’ ) pero no ha sido evaluada la significación funcional de esta sensibilidad.
REFERENCIAS ESPECIALES SOBRE INTEGRACION
A . El sistema nervioso
Ariens-Kappers, C.U., C. Huber, y E. Crosby. 1936. The comparative anatomy o f the nervous system of vertebrates, including man. The Macmillan Company, Nueva York.
Bernstein, J.J. 1970. Anatomy and physiology of the central nervous system. En Fishphysiology, 4:l-90. Academic Press, Nueva York.
Eddy, J.M.P. 1972. The pineal complex. En The biology of lampreys, 2:91-103. Academic Press, Nueva York.
Evans, H.M. 1940. Brain and body of fish. The Blakiston Company (McGraw-Hill Book Com- pany, Nueva York).
Fenwick, J.C. 1970. The pineal organ, En Fish physiology, 4:91-108. Academic Press, Nueva York.
Ingle, D. (Ed.) 1968. The central nervous system and fish behavior. University of Chicago Press, Chicago, 111.
Lissmann, H.W. 1946. The neurological basis of the locomotory rhythm in the spiny dogfish (Scillium canicula, Acanthias vulgaris). I Reflex behavior, I1 The effect of de-afferentation. Jour. Exptl. Biol. 23:143-176.
Nicol, J.A.C. 1952. The autonomic nervous system in lower chordates. Biol. Rev. Cambr.
Uchihashi, K. 1953. Ecological study of Japanese teleosts in relation to the brain morphology.
von Holst, E. 1934. Weitere Reflexstudien an spinalen Fischen. Zeitschr. uergl. Physiol. 21:
Philos. SOC., 27(1):1-49.
Bull. Japanese Sea Reg. Fish Res. Lab., 2(2):1-180.
658-665.
B. El sistema endocrino
Ball, J.N., y B.I. Baker. 1969. The pituitary gland: anatomy and histophysiology. En Fish physiology, 2:l-110. Academic Press, Nueva York.
Bardach, J.E., R. Ryther, y W. McLarney. 1972. Aquaculture: the farming and husbandry o f freshwater and marine organisms. Wiley Interscience. John Wiley and Sons. Nueva York.
Bretschneider, L.H., y J.J. Duyvene de Wit. 1947. Sexual endocrinology of non-mammalian Vertebrates. J. Elsevier Publishing Company, Amsterdam.

370 INTEGRACION
Gorbman, A. (ed.). 1959. Comparative endocrinology. John Wiley and Sons, Nueva York. Gorbman, A., y H.A. Bern. 1962. A textbook o f comparative endocrinology. John Wiley and
Hoar, W.S., y D.J. Randall (eds.) 1969. The endocrine system. vol. 2 , Fish physiology, Academic
Lagios, M.D. 1975. The pituitary gland of the coelacanth Latimeria chalumnae Smith. Gen.
Nomura, T., H. Ogata, y M. Ito. 1973. Occurrence of prostaglandins in fish testis. Tohoku
Pickford, G., y J.W. Atz. 1957. The physiology of the pituitary gland of fishes. New York
Sons, Nueva York.
Press, Nueva York.
Comp. Endocrinol., 25:126-146.
Jour. Agric. Res., 24:138-144.
Zoological Society, Nueva York.
C. Órganos de los sentidos
I. Los Sentidos Cutáneos
Bardach, J.E., y J. Atema. 1971. The sense of taste in fishes. pp. 293-336. En Handbook of sensory physiology, vol. IV (Z), L. Beidler, ed. Springer Verlag, Berlin, Heidelberg, Nueva York.
Bull, H.O. 1936. Studies on conditioned responses in fishes. Part VII, Temperature discrimi- nation in teleosts. Jour. Marine Biol. Assoc. United Kindom, 21(1):1-27.
Dijkgraaf, S. 1940. Untersuchungen ueber den Temperatursinn der Fische. Zeitschr. vergl. Physiol., 27:587-605.
Sand, A. 1938. The funct.ion of the ampullae of Lorenzini with some observations on the effect of temperature sensory rhythms. Proc. Roy. Soc. London Ser. B. Biol. Sci.,
Spath, M. 1974. On the processing of the mechanical and thermal information of the receptors in fish skin. En Symposium on Mechanoreception, Rheinisch-Westfaelishe Akad. d. Wissenshaften. 53:251-262. Bochum.
Sullivan, C.M. 1954. Temperature reception and responses in fish. Jour. Fish. Res. Bd. Canada,
Whitear, M. 1952. The innervation of the skin of teleost fishes. Quart. Jour. Microsc. Sci.,
125:524-553.
11(1):153-170.
93:289-305.
11. Visión
Ali, M.A. 1959. The ocular structure, retinomotor and photobehavioral responses of juvenile
Feuerwerker, E., y M.A. Ali. 1975. La vision chez les poissons historique et bibliographie
Granit, R. 1947. Sensory mechanisms of the retina. Oxford University Press, Londres. Kleerekoper, H. 1972. The sense organs. En The biology of lampreys, 2:373-404. Academic
MacNichol, E.J., Jr., y J. Svaetichin. 1958. Electric responses of fish retinas. Amer. Jour.
Munz, F.W. 1971. Vision: visual pigments. En Fish physiology, 5:l-32. Academic Press, Nueva
Pumphrey, R.J. 1961. Concerning vision. En The ceZZ and the organism, pp. 193-208. Cam-
Wagner, H.G., E.J. MacNichol, Jr., y M.L. Wolbarsht. 1960. Opponent color responses in retinal
Wald, G. 1945. The chemical evolution of vision. The Harvey Lectures, 41:117-160. Walls, G. 1942. The vertebrate eye and its adaptive radiation. Cranbrook Inst. Sci. Bull. núm.
Pacific salmon. Canadian Jour. Zool., 37:965-996.
analytique. Rev. Can. Biol., 34(4):221-285.
Press, Nueva York.
Ophthalmol., 46(3 II):26-46.
York.
bridge University Press, Cambridge.
ganglion cells. Science, 131:1314.
19, Cranbrook Institute of Science, Bloomfield Hills, Mich.
111. Olfación
Bardach, J.E. y T. Villars. 1974. The chemical senses of fishes. pp. 49-104. En Perspectives in chemoreception by marine organisms. Academic Press, Nueva York.

REFERENCIAS 371
Hasler, A.D. 1954. Odour perception and orientation in fishes. Jour. Fish. Res. Bd. Canada,
Kleerekoper, H. 1969. Olfaction in fishes. Indiana University Press, Bloomington, Ind. Kleerekoper, H., y G.A. van Erkel. 1960. The olfactory apparatus of Petromyton marinus.
11(2):107-129.
Canadian Jour. Zool., 38:209-223.
IV. El Oído Interno
Kleerekoper, H., y E.C. Chagnon. 1954. Hearing in fish with special reference to Semotilus atromaculatus atromaculatus (Mitchill). Jour. Fish. Res. Bd. Canada, 11(2):130-152.
Lowenstein, D. 1950. Labyrinth and equilibrium. Symp. Soc. Exp. Biol., IV:60-82. von Holst, E. 1950. Die Arbeitsweise des Statolithenapparates bei Fischen. Zeitschr. vergl.
Physiol., 32:60-120.
V. La Línea Lateral
Dijkgraaf, S. 1933. Untersuchungen ueber die Funktion der Seitenorgane bei. Fischen. vergl. Physiol., 20:162-214.
Dijkgraaf, S. y A.J. Kalmijn. 1966. Versuche zur biologischen Bedeutung der Lorenzinischen Ampuller bei den Elasmobranchiern. Zeitschr. vergl. Physiol. 53:187-194.
Flock, A. 1965. Electromicroscopic and electrophysiological studies on the lateral line canal organ. Acta Oto-laryngologica, Suppl. 199:l-90.
Freihofer, W.C. 1972. Trunk lateral line nerves, hyoid arch gill rakers, and olfactory bulb location in atheriniform, mugilid, and percoid fishes. Ocass. Papers Cal. Acad. Sei., 95, 31 p.
Hoagland, H. 1933. Electrical responses from the lateral line nerves of catfishes. Jour. Gen. Physiol., 16:695-731.
Katsuki, Y., T. Hashimoto, y T.T. Kendall. 1971. The chemoreception in the lateral line organs of teleosts. Japanese Jour. Physiol., 21:99-118.
Katsuki, Y., S. Yoshino, y J. Chen. 1950. Action currents of the single lateral line nerve fiber of fish. Japanese Jour. Physiol., 1:87-99; 179-194; 264-268.
Murray, R.W. 1962. The response of the ampullae of Lorenzini of elasmobranchs to electrical stimulation. Jour. Exptl. Biol., 39!1):119-128.


Capítulo 12
Genética y evolución
Lo que nosotros conocemos sobre los mecanismos de la herencia en los peces está basado primordialmente en los estudios intensivos que se han hecho de unos cuantos miembros de un sólo orden, los Cyprinodontiformes, particularmente el “guppy” (Lebistes reticulatus), los peces ancho y los cola de espada (Xiphophorus), el “medaka” o pez asiático de los arrozales (Oryzias latipes), el “molly” del Ama- zonas (Poeciliu forrnosa) y especies mexicanas del género Poeciliopsis. El conoci- miento de este campo se ha ampliado en los años recientes gracias a las investiga- ciones que se han estado realizando en la carpa común y en la carpa dorada (Cyprinidae), los ciprinodóntidos de Anatolia, a los experimentos con truchas y “chars” (Salmoninae), y especímenes representativos tomados al azar de algunos otros grupos de peces. Mucho se sabe sobre la genética de la carpa dorada (Cara- ssius aumtus), principalmente por lo que se ha dado a conocer en la literatura japonesa. Pero queda pendiente la gran mayoría de peces, incluyendo a los verte- brados primitivos, sin mandíbulas (Cyclostomata), de casi todos los cuales se carece de información al respecto. En otras palabras, apenas hemos indagado superficialmente en esta gran cantidad de datos potenciales sobre la herencia, que nos tiene reservada este grupo tan grande y diverso de animales.
LA HERENCIA EN LOS PECES
La herencia simple, mendeliana, está bien demostrada por la aparición de dorado, un carácter mutante recesivo del “guppy” (Lebistes) que resulta de la reducción de cerca de un 50% de los melanóforos característicos de las poblaciones libres. La pérdida de las células con pigmento negro descubre y vuelve visibles a las células
373

374 GENfiTICA Y EVOLUCI6N
con pigmento amarillo que se encuentra por debajo, en la piel de todos los gup- pies.
Al cruzar un tipo libre o silvestre con uno dorado se produce una genera- ción F1 que se asemeja mucho en el color al padre silvestre. Pero al cruzar a dos de esta primera generación de grises (hermano-hermana o pareja de parientes) resulta una nueva generación que va en la proporción de tres silvestres y uno do- rado. Esta proporción típicamente mendeliana es la que se espera cuando se fija la atención en un simple carácter recesivo mendeliano.
Pueden aparecer desviaciones significativas de la proporción 3:l a partir de complicaciones secundarias. Por ejemplo, el "guppy" (Lebistes) albino, caracteri- zado por la falta de melanóforos y la presencia de ojos rosados, cuando es cruzado con el tipo silvestre produce una generación F1 igual a la del padre silvestre (Fi- gura 12.1). Pero en la generación F,, que resulta, como ya se mencionó, del cruzamiento de un hermano y una hermana de la generación F, , la proporción que resultará de silvestre (ordinario) a albino será de 53:l. Esta discrepancia tan
Albino 9 p1 Silvestre d X
I 1 Albino? 1 Albinod

LA HERENCIA EN LOS PECES 375
amplia en relación a la proporción 3:1, que se esperaba, es atribuible a una debi- lidad inherente al grupo del “guppy” albino, siendo el albinism0 una mutación recesiva, semiletal, que da como resultado la mortalidad de un elevado porcentaje de la cría albina, aun antes de su nacimiento.
Otro carácter del “guppy” que muestra el mecanismo de la herencia men- deliana simple es el “cebra” o patrón de dos a cinco barras situadas en la parte posterior del cuerpo de los machos (Figura 12.2). Aunque incapaces de mostrar el patrón, las hembras pueden sin embargo transmitir el factor hereditario a sus hijos. Los caracteres como éste, que son transmitidos por los dos sexos, pero solamente mostrados por el macho (o la hembra), son limitados al sexo. Al cruzar una cebra macho con una hembra no cebra resultan hijos heterocigóticos para ese carácter “esto es, cada uno tiene un gene “cebra” dominante y un gene “cebra” recesivo-, pero desde el momento en que el carácter está limitado a los machos, solamente los hijos lo muestran en el fenotipo (las hijas son portadoras). Cuando se hace un cruzamiento con hermanos o parientes de la generación F 1 , la cría resultante, tomando en cuenta solamente a los hijos, muestra una proporción de 3:l en favor del patrón cebra. Esto indica que el carácter representa un factor mendeliano simple, dominante.
No-cebra ? p1 Cebra d
I (Portador de cebra) ? F I Cebra d I
Retro-cruzamiento
(Portador de cebra) ? Nocebra d
I (Portadores de cebra) Cebras d d No cebras d 6 I y notebras 9 ?
Figura 12.2 - La herencia de la ”cebra” en el “guppy“ (Lebistes) muestra una simple herencia Mendeliana de sexo limitado (Fuente: Gordon, 1953).

376 GENfiTICA Y EVOLUCIdN
La reversión al tipo silvestre de las razas domesticadas de plantas y animales puede ser observada comúnmente. En la carpa dorada (Curussius uurutus), el pri- mer pez que se cree haya sido domesticado en China hace casi mil años, con fre- cuencia ocurre la reversión al color ancestral verde-olivo. Estos retrocesos son fácilmente explicados en términos de la herencia mendeliana ordinaria. La re- aparición de las razas ancestrales es usualment,e el resultado de la reunión de caracteres hereditarios precisos que se han mantenido separados durante el pro- ceso de la domesticación.
La herencia no mendeliana o la aparición de resultados no predecibles en los experimentos de cruzamiento puede deberse a fenómenos como la combina- ción de caracteres o herencia matroclina (que es transmitida solamente por la madre, véase abajo). Por ejemplo, cuando el gallito dorado (un mutante recesivo de Xiphophorus rnuculutus) domesticado es cruzado con el gallito cometa silvestre (el cual es gris y tiene un margen negro alargado en los lóbulos superior e inferior de la aleta caudal), la filial I?, no es ni gris ni cometa (como podría esperarse, desde el momento en que estos caracteres son dominantes sobre los dorado y no co- meta), sino que resultan grises y con “cola abundante”, en la cual n o solamente toda la aleta caudal es negra, sino también las demás (Figura 12.3). En otras pala- bras, la característica del gris se condujo como era de esperarse en la herencia, pero la forma de cometa no. La razón para que haya aparecido este resultado inesperado en la filial F1 es que el gene que determina la condicibn cometa es afectado por otro gene. Este gene adicional, denominado modificador específico, alteró radicalmente el efecto usual del gene para el carácter cometa, causando una respuesta manifestada por la aparición de pigmento mucho más ampliamente distribuido.
Un tipo de herencia que fue desconocido para Mendel es llamado alelismo múltiple. Un alelo representa a una de las muchas fases alternadas de un gene. En
X
Gris, cometa 9
Primera generación
Platidorado d
Todos grises, cola meneadora
Figura 12.3 - Resultado genético de la cruza entre cometa gris y el pez platidorado, mostrando el efecto dado por un gene específico modificador en el efecto usual del gene cometa (Fuente: Gordon, 1953).

DETERMINACION DEL SEXO 311
un cruzamiento que comprenda dos pares de caracteres contrastantes indepen- dientes, como sucede con un gallito (Xiphophorus) cometa (CoCooo) y otro con una mancha (cocoOO), todos los miembros de la primera filial F1 son cometas y con una mancha, debido a que tanto el carácter de cometa como el de una man- cha son dominantes. Cuando una hembra F1 se cruza con su hermano, los resul- tados esperados sobre la base de la herencia de dos factores deberán corresponder a la proporción de 9:3:3:1 (9 cometas, una mancha: 3 cometas, solamente: 3 de una mancha, solamente: 1 sin ningún carácter heredado). Los dieciséis fenotipos observados se produjeron, sin embargo, en la proporción de 1:2:1 (4 cometas solamente: 8 cometas; una mancha: 4 de una mancha, solamente). Estas desvia- ciones de lo esperado, aunque no grandes, persisten con los cruzamientos repe- tidos y con el nacimiento de segundas generaciones adicionales. Las proporciones sorpresivas o no esperadas son significativas y pueden ser explicadas de la siguiente manera. En lugar de tener un gene justamente dos fases, una dominante y otra recesiva, puede tener dos o más expresiones en su fase dominante, esto es, O y Co pueden ser las dos dominantes ante un recesivo común. El término que aplican los genetistas para este tipo interesante de herencia que comprende a una serie de genes es el de los “alelos dominantes múltiples”. De aquí que en el ejemplo justa- mente dado de los modelos o patrones de color contrastante del gallito, no esta- mos considerando a dos genes simples dominantes y a dos genes simples recesivos, sino a una serie de alelos múltiples. En consecuencia, la generación F1 no está propiamente expresada por la fórmula CocoCo, desde el momento en que Co y O son alelos; el cruzamiento deberá ser expresado por las fórmulas siguientes:
PI Cometa X Un-Lunar COlCO o/o
F1 Todos cometa, Un-Lunar co/o
Al reproducirse la generación F1, Col0 X Col0 produce una generación con la proporción de 1:2: 1 (1 cometa: 2 cometas, una mancha o lunar: una man- cha). Resultan tres pececillos gallo presentando el modelo de cometa, contra uno sin el patrón cometa.
De la misma manera hay tres pececillos gallo con el patrón de una mancha o lunar contra uno al que le falta el patrón una mancha.
Aunque estos resultados no corresponden a las predicciones de Mendel (sobre la base de la interpretación de dos factores), son sin embargo explicables desde el punto de vista de sus principios básicos. La proporción 1:2: 1 es mera- mente una ligera variación de la proporción fundamental 3 : 1.
DETERMINACI6N DEL SEXO
Casi todos los tipos de reproducción sexual conocidos en los animales pueden ser encontrados en los peces (Capítulo lo) , incluyendo ciertas especializaciones sin paralelo entre todos los vertebrados. Sin embargo, en su conjunto, el mecanismo de la determinación del sexo en la mayoría de los peces óseos está todavía en una condición lábil, primitiva. Los cromosomas sexuales están tan poco diferenciados de los autosomas que apenas se pueden distinguir unos de los otros. La determina-

378 GENBTICA Y EVOLUCIdN
ción del sexo puede ser controlada en parte, si no completamente, por los genes dis- persos entre los autosomas. El potencial determinante del sexo puede diferir entre varias especies, como cuando, al realizarse cruzamientos entre híbridos, el macho de una especie, más fuerte y productor de genes, se combina con la hembra de la otra especie, más débil y también productora de genes; ello da como resultado un desequilibrio muy fuerte, e inclusive total, en las proporciones del sexo, favore- ciendo al macho.
Los datos disponibles sobre la determinación del sexo en poblaciones de peces, tanto libres o silvestres como de laboratorio, son más extensos en compa- ración a las de otros vertebrados. Mientras que en los mamíferos la hembra es homocigótica (XX) y el macho heterocigótico (XY), y en las aves prevalece pre- cisamente lo opuesto (hembra, WY; macho, YY), en los peces estos dos sistemas están representados.
Debe insistirse mucho en que los cromosomas sexuales, como tales, son excesivamente difíciles de identificar en los peces por medios citológicos, porque los cromosomas de éstos son delicados, usualmente numerosos y difíciles de ob- servar. En muchos de los ejemplos en que los investigadores han asegurado que han podido distinguir morfológicamente a los cromosomas sexuales, posiblemente se ha tratado solamente de artefactos lo que se ha creído que son estructuras, el resultado de una fijación inadecuada. Un par muy nítido de cromosomas sexuales ha sido encontrado, sin embargo, en el macho del pez de los mares profundos, Bathylagus wesethi; el mayor en el kariotipo es presumiblemente, X, y el más pequeño, Y. Otras dos especies de este género, y algunas especies de Fundulus, tienen un par similar de cromosomas sexuales, como en Gambusia affinis y otros. En Megupsilon uporus, un ciprinodóntido mexicano, hay un mecanismo cromo- sómico sexual múltiple XI Xz Y con un Y metacéntrico muy grande. El tipo XY de cromosoma sexual fue demostrado recientemente para un anfioxo, Brunchios- toma belcheri. La discusión que sigue está basada, por lo tanto, en experimentos de genética que dan una evidencia fuertemente teórica sobre los cromosomas sexuales, pero no prueban su existencia.
La herencia ligada al sexo es aquélla en la que el carácter en cuestión está determinado por un gene (o genes) localizado en los cromosomas sexuales. La herencia limitada al sexo o “de un lado’’ (Figura 12.2) resulta cuando un carácter es cedido solamente de un macho a un macho o de una hembra a una hembra; el gene de que se trata se encuentra en el cromosoma impar. Un ejemplo de carácter ligado al sexo, herencia limitada al sexo, es la de un carácter de color, dominante, el lunar negro en la aleta dorsal del “guppy” macho (Lebistes). Esto también ilustra que Lebistes tiene un tipo de determinación sexual genética que se iden- tifica con el modelo XX para la hembra y XY para el macho, conocido en los mamíferos. En las fórmulas siguientes los símbolos para los cromosomas sexuales están escritos con mayúsculas y situados dentro de paréntesis, mientras que los genes que ellos portan están italizados (el lunar negro es llamado macuhtus y está representado por M a ) :
Hembras Machos

HIBRIDACIdN 379
Este tipo de herencia limitada al sexo ha sido también demostrado para el “medaka” (Oryzius) y el gallito (Xiphophorus).
Investigaciones citológicas
L a cantidad de cromosomas, junto con el criterio morfológico convencional, los datos proporcionados por la Paleontología, los modelos de comportamiento, la ecología y los experimentos de genética, proporcionan una herramienta adicio- nal para descifrar la filogenia de los peces. Los más recientes inventarios muestran que el número diploide varía de 18 a 104, pero los investigadores pueden discrepar sobre el número preciso para una especie dada. Los números de cromosomas de- terminados a partir de las figuras mitóticas cromosómicas que se ven tan bien durante las etapas de la segmentación del huevo, pueden diferir radicalmente dentro de una sola especie, de los determinados mediante el estudio de las células germinativas del macho durante la espermatogénesis. Se dice que el segundo mé- todo es el más seguro. Es de esperarse que haya una correlación estrecha entre la morfología cromosómica y la organización estructural, desde el momento en que los cambios evolutivos son iniciados por cambios en los cromosomas.
Los exámenes realizados sobre el número de cromosomas son menos com- pletos para los peces que para los demás grupos de animales. Debe tenerse pre- sente, sin embargo, que los peces forman un grupo mucho más numeroso y diverso de especies que todos los demás vertebrados combinados. El trabajo serio sobre los cromosomas de los peces fue iniciado en 1930, y este campo de investigación ha recibido mucho impulso a partir de los trabajos publicados dentro de la década pasada. Aproximadamente cuatrocientas especies vivientes, de un número esti- mado en veinte mil, han sido estudiadas por los citólogos interesados en los cro- mosomas. El interés que se está incrementando por el estudio del número de cromosomas y la morfología en estos animales promete producir importantes contribuciones para el mejor conocimiento de la teoría evolutiva, así como de la filogenia de los peces.
Hay una tendencia general para mostrar similitud del número y forma en los cromosomas de las especies y géneros relacionados, pero nuestro conocimiento de ese aspecto es todavía tan incompleto que las conclusiones definitivas relativas a los cromosomas y su clasificación, apenas están comenzando a cristalizarse.
HIBRIDACIdN
La hibridación natural interespecífica es de lo más común entre los peces, espe- cialmente para los que habitan en las aguas dulces continentales (Holárticas), a diferencia de cualquier otra clase de vertebrado. Es un fenómeno particularmente recurrente en los salmónidos, lucios (Esocidae), “minnows” (Cyprinidae), carpas chupadoras (Catostomidae), peces sol (Centrarchidae) y en los peces flecha (Etheos- tomatinae, familia Percidae). Son conocidas (1972) 56 familias de peces híbri- dos dentro de la fauna mundial. Más de doscientos son conocidos entre los peces de agua dulce de Norteamérica. Para nuestros propósitos, un híbrido es definido como el resultado biológico del cruzamiento de padres de dos especies (o géne- ros) diferentes. El factor más importante que influye en la producción de los hí-

380 GENgTICA Y EVOLUCI6N
bridos es el medio ambiente alterado, pero de ninguna manera el imito. Debido a la continua modificación del medio acuático por el hombre, y a su alteración de la distribución de los peces, la hibridación está aumentando entre los peces de las aguas dulces.
Ya que la fertilización es primordialmente externa, debe esperarse que los peces se hibridicen con más frecuencia que los grupos superiores como las aves y los mamíferos. Las familias, en las que un gran porcentaje de especies se hibridi- zan se caracterizan por tener numerosas formas estrechamente relacionadas, con hábitos similares e iguales requerimientos de hábitat.
Hablando genéricamente, los híbridos F1 poseen un espectro de caracteres que está comprendida aproximadamente entre los que distinguen en forma signi- ficativa a las especies padres. La uniformidad de los caracteres intermedios puede ser tomada como una poderosa indicación de la esterilidad de los híbridos. Por el contrario, la variación amplia que pueda observarse en el producto híbrido será una indicación de que hay por lo menos fertilidad parcial. En los ejemplos que han sido estudiados citológica o experimentalmente, los cruzamientos interespe- cíficos muestran por lo común más tarde proporciones del sexo alteradas, a me- nudo exhiben heterosis o “vigor híbrido”, y esterilidad parcial o completa en por lo menas un sexo.
La evidencia de la hibridación natural debe, usualmente, ser en gran parte circunstancial. Esta evidencia, cuando ha sido cuidadosamente documentada y valorada por alguien que esté completamente familiarizado con la fauna, puede ser tomada sin embargo como una indicación confiable para la interpretación del híbrido. En los numerosos grupos que han sido ensayados experimentalmente, los presuntos híbridos naturales han sido convincentemente confirmados.
El trabajo experimental que se ha hecho con los peces sol (Lepomis) ha mostrado que los híbridos interespecificos crecen más rápido que cualquiera de las especies progenitoras y que ellos dominan en la jerarquía social que corres- ponde al orden de picoteo en las aves. Por ejemplo, cuando se le da de comer a un grupo mezclado, los híbridos -de mayor tamafío y más activos que las especies puras- son los primeros en tomar el alimento. El vigor incrementado de los hí- bridos también se ha visto que aparece en los “minnows” (Cyprinidae) y en las truchas (Salmoninae), y en los métodos de pesca en donde se emplean anzuelos en línea logran que se capturen relativamente más individuos híbridos que cual- quiera de sus supuestos padres.
En la gran mayoría de los peces sol (Lepomis) híbridos existe preponderancia del sexo masculino, a menudo en una relación de 4 a 1 o más. Estos machos, cuando maduran, construyen, abanican y protegen sus nidos de grava con gran vigor durante largos periodos, pero la energía en muchos de esos híbridos es desperdiciada, desde el momento en que la esterilidad puede ser completa, o casi, debido a la tremenda anormalidad de la espermatogénesis, entre otras po- sibilidades.
Uno de los ejemplos más ampliamente conocidos de reproducción aberrante en los vertebrados es ilustrado por el llamado “molly” del Amazonas (Poeciliu formosu) del que se sabe que existe en la naturaleza con el sexo femenino sola- mente (se han visto ocasionalmente machos fenotípicos que parecen representar hembras con la reversión del sexo). Se cree que este pez ha aparecido mediante la hibridación entre dos especies estrechamente relacionadas de Poeciliu y que se trata de un diploide permanentemente fijo. Ella persiste en la naturaleza mediante

MECANISMOS DE LA EVOLUCIdN 381
su apareamiento con el macho de cualquier especie de Poeciliu con el que simpa- tiza; considerando el 100% de su población, alrededor del 50% está formado por hembras reproductoras y parece ser que esta fecundidad incrementada le da una ventaja selectiva definida sobre las especies bisexuales con las que vive. El desa- rrollo embrionario es iniciado solamente mediante la fertilización (interna en este ejemplo); pero, aunque los espermatozoides son esenciales, la fusión gamé- tica (nuclear) no ocurre. De aquí que el fenotipo del producto sea totalmente dependiente del genotipo de la madre. Los jóvenes son invariablemente hijas y son genéticamente idénticos a sus madres. La producción consistente de crías femeninas de P. forrnosu ocurre no solamente en los cruzamientos de la primera generación, sino también en todos los subsiguientes cruzamientos con individuos de generaciones anteriores (lineales) o con los de otras madres (laterales). Este pro- ceso reproductor es llamado ginogénesis y el mecanismo hereditario es referido como herencia rnatroclina, si consideramos que solamente la cromatina de la madre es transferida a su cría, que viene a estar formada de puras hermanas.
Recientemente han sido demostrados experimentalmente otros tipos de hembra unisexual en la misma familia, los vivíparos (Poeciliidae), pero pertene- cientes al género Poeciliopsis. Sin embargo, a diferencia de la situación del “molly” del Amazonas (Poeciliu forrnosa), la cromatina del macho es transmitida al pro- ducto que es todo femenino, de tal manera que la primera generación de jóvenes peces manifiesta caracteres de ambos progenitores. De aquí que haya un meca- nismo genético diferente, según se piensa, para explicar satisfactoriamente la monosexualidad en Poeciliopsis. Se ha postulado que un proceso de maduración selectiva elimina los cromosomas del padre durante la ovogénesis. Todas las for- mas unisexuales descritas arriba pueden asegurarse como híbridos perpetuos.
Otros productos formados únicamente de hembras, de otros peces vivípa- ros, son conocidos solamente en los cultivos de laboratorio. Con el auxilio de genes marcados se han encontrado machos con el sexo invertido entre pececillos “guppy” (Lebistes), presumiblemente llevando consigo cromosomas XX; su sexo genético era hembra, su sexo real, macho. Los machos normales en Lebistes con- forman el tipo XY. Cuando uno de los machos excepcionales XX fue cruzado con una hembra normal XX, todos los individuos componentes del producto re- sultaron hembras, como era de esperarse. La primera suerte de fenómeno ha sido encontrado en el gallito (Xiphophorus) domesticado y en el pez asiático de los arrozales (Oryzias latipes).
Existe una correlación positiva entre el éxito de la hibridación y la proxi- midad de las relaciones, de tal manera que la supervivencia más grande de los híbridos ocurre cuando las especies progenitoras están más íntimamente relacio- nadas. Esto es de esperarse sobre la base de la hipótesis que explica que las espe- cies estrechamente relacionadas poseen cromosomas de igual morfología, número y características bioquímicas.
MECANISMOS DE LA EVOLUCIdN
La evolución “la transformación continua y gradual de la línea de descendencia a partir de un ancestro común- es un hecho perfecto. El prolongado proceso de la evolución, extendiéndose a lo largo de dos billones de años, ha sido el resultado de causas fundamentales que están todavía en actividad. Mientras que Darwin

382 GENOTICA Y EVOLUCIdN
tuvo que deducir su teoría principalmente a partir de evidencias indirectas, noso- tros podemos estudiar en nuestros días y aún producir cambios evolutivos, a nuestro gusto, en el laboratorio. La manera en que se ha producido la evolución, que aún está realizándose, es el tema de esta sección.
Debido a que las modificaciones hereditarias proporcionan los materiales básicos para que se produzca la evolución, la manera como estas variaciones here- dables se originan es algo básico que debemos conocer para la comprensión de los mecanismos evolutivos.
La mutación, junto con la recombinación y la selección, es esencial para que se produzca la evolución continuada de la mayoría, si no es que en todos, los tipos de organismos. Los materiales genéticos primordiales proporcionados por la mutación dan lugar a los bloques de construcción a partir de los cuales los cam- bios evolutivos pueden ser realizados mediante la selección natural. El origen de la variación es un problema enteramente diferente al de la acción de la selección.
Mutación del gene
El gene es la unidad de la herencia mendeliana. Los genes aparecen ante todo en los cromosomas de los núcleos de las células y, desde el momento en que forman parte de los cromosomas, existen por pares, justamente como ocurre con los cromosomas. Un cambio permanente y espontáneo en un gene es llamado muta- ción genética. Esos cambios heredables proporcionan un importante material para que tenga lugar la variación orgánica, y la variabilidad heredable es esen- cial para la evolución. Contrariamente a las ideas expresadas por los críticos de la genética, cuando esta ciencia estaba en su infancia, ya estaba bien establecido que la mutación es un fenómeno normal en la naturaleza. Los grados de muta- ción probablemente varían mucho de acuerdo a los diferentes genes mutando en diversos grados, inclusive dentro de la misma población. Los grados de mu- tación pueden ser incrementados en las poblaciones silvestres mediante los lla- mados genes mutadores.
Tanto en los medios experimentales como en los naturales se ha encon- trado que la mayoría de las mutaciones son desventajosas, y por consiguiente son eliminadas por selección natural. Pero quedan muchas mutaciones que son “neu- trales” o ventajosas para el organismo que las posee, y algunas que aparentemente son perjudiciales podrían ser valiosas estando en medios modificados. Desde el momento en que la mayoría de las mutaciones alteran únicamente a un solo gene a un tiempo, es la acumulación de pequeños cambios (no un cambio grande, sú- bito) en un periodo de tiempo largo la que contribuye al cambio de constitucio- nes genéticas. La radiación, por ejemplo con rayos X, acelera los grados de muta- ción, según se sabe, y es probable que la mutación sea parcialmente causada en la naturaleza por la acción química (como, por ejemplo, por el peróxido de hidró- geno). En general, la mutación espontánea ocurre al azar o de una manera indi- recta; esto es cierto para todos los tipos de mutación.
Mutación cromosómica
Los estudios intensivos de los cromosomas de la glándula salival gigante de las moscas de la fruta (Drosophila) han demostrado que los cromosomas están dife-

MECANISMOS DE LA EVOLUCIdN 383
renciados longitudinalmente y que sus genes aparecen en loci o posiciones defi- nidas. Por lo tanto, se han llegado a hacer “mapas” cromosómicos de los cromo- somas de las glándulas salivales, e inclusive son identificables pequeños segmentos mediante la referencia a un mapa modelo. La redistribución de los cromosomas puede ocurrir siguiendo cuatro procesos: u) escisión: la pérdida de un segmento de un cromosoma; b ) duplicación: la duplicación de un segmento cromosómico; c ) inversión: la inversión de un segmento de un cromosoma debido a su rotura en dos puntos; y d) truslocación: la transferencia de un segmento de un cromo- soma a un cromosoma no homólogo. Estos cambios arquitectónicos de los cro- mosomas pueden ser producidos experimentalmente mediante radiaciones de energía elevada, y es seguro inferir que la radiación que ocurre de manera natural, así como también los mutágenos químicos, pueden jugar un papel importante en la producción de cambios que han llegado a ser observados en forma experi- mental. Cada uno de los tipos de rearreglo ya mencionados se comportan como si fuesen una mutación genética, según se ha observado, produciendo entonces un fenotipo diferente y hereditario. Muchos son letales cuando son homozigó- ticos, pero otros no. La traslocación de los segmentos cromosómicos puede con- ducir a un cambio en el efecto del gene, por ejemplo de recesivo a dominante. También pueden afectar las traslocaciones al número de cromosomas, como sucede en los peces de la familia Cobitidae: las lochas japonesas (Misgurnus y Barbatula) tienen de 24 a 26 pares de cromosomas, respectivamente, la mayoría de los cuales tienen forma de bastón. Sin embargo, dos pares de cromosomas de Barbatula tienen forma de V y la reducción (de 26) es atribuida a las traslocacio- nes que resultaron al unirse originalmente diferentes pares de cromosomas.
Selección natural
El más grande descubrimiento de Darwin, la teoría de la selección natural, cons- tituye un tema central en todo el pensamiento biológico. Es el proceso evolutivo más importante y es esencialmente un intento de explicación de la adaptación de los organismos a su medio ambiente. El meollo de la selección es que los portadores de los diferentes genotipos en una población contribuyen en forma diferente a formar el acervo genético de las generaciones sucesivas. Algunos genotipos hacen contribuciones relativamente grandes en promedio, en comparación con otros, en el mismo medio ambiente. La relativa capacidad de los portadores para trans- mitir sus genes constituye el valor adaptativo, o buena condición darwiniana, de ese genotipo. Las poblaciones mendelianas (no los individuos) son las unidades de la selección natural y de la adaptación.
Como resultado de la selección natural los organismos que gradualmente van cambiando se adaptan a las condiciones bajo las cuales deben existir, debido a que algunas variaciones producen más progenie apta para sobrevivir que otras. La adaptación -afectada a través de la selección natural- es por lo tanto un proceso de importancia primordial para la planta o animal que se está desenvolviendo. La paleontología pone especial énfasis en este hecho, desde el momento en que el registro de las adaptaciones progresivas que están teniendo lugar a través de perio- dos definidos de tiempo geológico no deja dudas sobre la importancia de la adap- tación y de la selección en la evolución.
En ciertas especies se han producido cambios importantes como resultado

384 GENBTICA Y EVOLUCI6N
de la selección natural dentro de los tiempos históricos. Así es como la fase de color melanística (negra) de las mariposas, generalmente de aparición esporádica, se ha vuelto notable en áreas afectadas por la revolución industrial (de aquí el fenómeno que ha sido denominado “melanismo industrial”). Los centros de dis- tribución de las formas melanísticas son las grandes ciudades industriales. La explicación de este acontecimiento, de que haya una fase oscura sobre otra clara, y con verdadero éxito, es la de que las mariposas melanísticas constituyen presas menos conspicuas. Sin embargo, la coloración más oscura puede ser una adapta- ción secundaria a los cambios fisiológicos, como la resistencia a las sustancias químicas venenosas contenidas en el humo industrial. Se encontró que las dos cualidades eran preadaptaciones para una condición de cambio del medio am- biente y eran de poco valor para el animal, hasta que prosperó la revolución industrial.
Poliploidía
La existencia en organismos muy relacionados (un género Único, por ejemplo) de cierto número de cromosomas en algunas especies, que es múltiplo de otros en otras, es llamada poliploidía. Por ejemplo, varias especies de trigo tienen 14, 28 y 42 cromosomas; los dos juegos de mayor valor son múltiplos del que tiene menor valor, Los poliploides pueden resultar de la duplicación del genoma dentro de una especie (autopoliploidía), o un incremento en el complemento cromosómico puede aparecer a través de la hibridación y producir genomas diferentes (alo- poliploidía).
Aunque es muy importante en el reino vegetal, la poliploidía es un fenó- meno raro o, cuando menos, poco común en los animales. Por lo general, se ha señalado a la reproducción bisexual como la base para esta diferencia entre los dos reinos. Esta sugerencia es apoyada por el hecho de que la poliploidía es rara en las plantas que han llegado a tener sexos separados (dioicas). Mientras que la hi- bridación es importante en la evolución vegetal y se le responsabiliza por la fre- cuencia de aparición de alopoliploides en las plantas, la producción y manteni- miento con éxito de híbridos parece haber desempeñado papeles de relativamente menor impartancia en la evolución animal. Ha sido demostrado que las especies poliploides que se reproducen sexualmente se encuentran entre los ciprínidos y salmónidos, en los que los juegos de cromosomas de algunas especies parecen ser múltiplos de los de otras. La mayoría de los animales reportados como po- liploides, sin embargo, son partenogenéticos y, curiosamente, todos ellos son artrópodos. El camarón, Arterniu salina, aparece en formas tetraploides y octo- ploides, partenogenéticas, pero también hay una raza tetraploide que se reproduce sexualmente.
MECANISMOS DE AISLAMIENTO
A menudo aparecen juntas especies muy relacionadas, completamente distintas, sin perder su identidad. Ellas no se entrecruzan porque permanecen más o menos separadas mediante una variedad de mecanismos de aislamiento. Desde luego que la formación de especies sin haber previa forma de aislamiento es considerada

MECANISMOS DE AISLAMIENTO 385
como imposible. En general, estas barreras que se presentan para el cruzamiento caen en dos grupos principales: geográfico y genético, o extrínseco e intrínseco.
El aislamiento geográfico comprende la separacibn espacial de las poblacio- nes en diferentes regiones geográficas, ya sea dentro de un área habitada sin solu- ción de continuidad, o separada por brechas distribucionales. Es independiente de cualquier diferencia genética que haya entre las poblaciones. El intercambio directo de genes entre las poblaciones cuyas áreas de distribución no se superponen o en- trecruzan es evitado simplemente porque ellas son alopátricas. Las poblaciones alopátricas pueden o no exhibir aislamiento reproductor si son cruzadas artificial- mente o, como resultado de un segundo entrecruzamiento, cuando son removidas sus barreras distribucionales. Por el contrario, algunas especies simpátricas -por ejemplo, Xiphophorus helleri y X . maculatus, el gallito y el “platyfish’” que se sabe no se cruzan en la naturaleza, son fácilmente inducidos para cruzarse en los acuarios.
Los mecanismos de aislamiento intrínsecos comprende un gran conjunto de medios. Y todavía más, el entrecruzamiento de un par de especies es usual- mente prevenido no solamente por un mecanismo, sino por muchos que forman una especie de engranaje, colaborando mutuamente.
Las características conductuales, incluyendo la sexual, la psicológica o la etológica, consideradas como barreras para el cruzamiento, son, probablemente, operativas en la promoción de la segregación de especies en la mayoría de los peces que son sexualmente dimórficos y/o dicromáticos. Los peces dardo (Etheos- tomatinae), los peces vivíparos (como los Poeciliidae), las espinochas (Gasterostei- dae) y los de los arrecifes de coral, incluyen muchos ejemplos en los cuales esos mecanismos desempeñan un papel importante en el mantenimiento de las especies como poblaciones diferentes.
Las diferencias que existen con respecto a la temporada de reproducción constituyen barreras temporales contra el cruzamiento. El “Dolly Varden” (Salue- linus malma) se reproduce en el otoño, la trucha arco iris (Salmo gairdneri) en las postrimerías del invierno o en la primavera. Por lo tanto, cuando estas dos especies de peces conviven, aun así no se pueden cruzar.
Las adaptaciones para los diferentes nichos ecológicos previenen el colapso de las barreras genéticas entre las especies relacionadas, Por ejemplo, el pez saeta de Johnny (Etheostoma nigrum) desova debajo de las rocas en una posición inver- tida, mientras que el pez saeta de lados verdes (Etheostoma blennioides) desova, mientras está en una posición derecha, normal, sobre algas y musgo.
Las adaptaciones fisiológicas pueden colaborar con las ecológicas en la prevención del trastorno genético. La espinocha de tres espinas (Gasterosteus aculeatus) aparece en Bélgica, ecológica y fisiológicamente, en dos poblaciones diferentes. Una habita solamente en agua dulce, usualmente pequeños arroyos, mientras que la otra vive en el mar durante el invierno, pero migra hacia los estua- rios en la primavera y en el verano para desovar. Aunque las dos formas difieren en promedio en cierto número de características morfológicas, las variaciones se entrecruzan ampliamente. La forma de agua dulce tiene la capacidad osmorre- guladora para mantener el contenido de cloruro en la sangre a un nivel siempre constante en las aguas de salinidad baja. La forma migratoria, sin embargo, pierde iones de cloro en las aguas que contienen una concentración salina menor a la cuarta parte de la del océano. Esto conduce rápidamente a los peces a la muerte. Los intergrados morfológicos que aparecen en los hábitats naturales no son fi-

3 86 GENfiTICA Y EVOLUCI6N
siológicamente intermedios sino que pertenecen a uno u otro tipo fisiológico. Aunque las dos formas son fácilmente cruzadas mediante la inseminación artificial de los óvulos, los híbridos raramente aparecen en la naturaleza. Su falla para so- brevivir ahí es debida a la existencia de un medio inadecuado, desde el momento en que son escasas las aguas de salinidad apropiada.
La incompatibilidad física de los genitalia debido a las diferencias estructu- rales de estos órganos produce una barrera mecánica de aislamiento. Esto, por ejemplo, previene virtualmente la fertilización cruzada de muchas especies de pececillos vivíparos (Poeciliidae).
Los factores bioquímicos pueden prevenir la unión del espermatozoide con el óvulo o, en los peces vivíparos, el esperma puede tener una vitalidad reducida en los conductos sexuales de la otra especie. Además, la competencia espermática puede ser un factor en la falla de los gametos masculinos extraños para penetrar en el óvulo.
En el caso de que ocurra la hibridación, y esto sucede repetidamente en los peces de agua dulce del Hemisferio Norte, hay muchos mecanismos que se con- ducen contra el establecimiento exitoso de los híbridos. Los zigstos híbridos pueden ser no vitales o menos adaptados que los zigotos de cualquiera de las es- pecies progenitoras, como sucede en los híbridos de la mayoría de los peces sol. Los híbridos pueden fallar para producir células sexuales funcionales; esta esteri- lidad híbrida es un mecanismo de aislamiento común y efectivo y se presume que es un factor importante en el fracaso de la mayoría de los peces híbridos de interespecificidad natural, para sobrevivir más allá de la primera generación. Pero, en caso de que los híbridos de la generación F1 produzcan gametos normales, toda o parte de la generación F2, o los cruzamientos regresivos, pueden ser no vitales o adaptativamente inferiores a los de los híbridos de la generación F 1 .
EVOLUCI6N PARALELA O CONVERGENCIA
Cuando los animales de diferente origen o historia se desarrollan en medios muy parecidos y en regiones bien aisladas, pueden desarrollarse formas funcionalmente similares pero sólo cuando están lejanamente relacionadas. La evolución paralela produce el parecido superficial, un fenómeno no claramente reconocido por Dar- win, pero ya sabido y muy común desde mucho tiempo atrás. También es llamada evolución convergente o, simplemente, convergencia.
Es un ejemplo clásico la distribución de las grandes aves no voladoras, las Ratites (kiwis, avestruces, casuarios, emús y ñandtíes) de Africa, Madagascar, América del Sur, Australia, Nueva Guinea y Nueva Zelanda. Antiguamente se pensó que estaban íntimamente relacionadas y fueron clasificadas en un grupo Único; actualmente se sabe que provienen de ancestros divergentes. Un ejemplo notable entre los peces puede ser observado en los extraordinariamente abundantes y diversos caracínidos (familia Characidae), un grupo dominante entre los peces de agua dulce de América del Sur. Al adaptarse por sí mismos a los variados nichos ecológicos, especies de esta única familia han llegado a parecerse mucho a peces, sin que medie parentesco, como los de las familias norteamericanas de peje-lagartos (Lepisosteidae), peces de aleta ondulada (Amiidae), lucios (Esocidae), pececillos de acuario (Cyprinidae) y peces saeta (Etheostomatinae). La pérdida de los ojos y el pigmento en diferentes familias de peces que habitan en las cavernas proporciona otro buen ejemplo de convergencia.

DURACION DEL PROCESO EVOLUTIVO 387
DURACION DEL PROCESO EVOLUTIVO
¿Qué tan rápido se desarrollan los organismos? Esta pregunta evolutiva básica no puede ser contestada con facilidad, porque los cambios evolutivos, por lo que respecta al tiempo, no solamente varían mucho en los diferentes grupos de anima- les, sino que también hay una gran fluctuación dentro de las clases, familias o géneros. Los accidentes geográficos, como esos que permiten a una forma ancestral vivir en un medio saturado (un lado aislado, por ejemplo), pueden dar como resul- tado una evolución “explosiva” relativamente rápida. Este tipo de diferenciación ha ocurrido repetidamente en los lagos, especialmente en los de los viejos trópicos.
Una de las mayores dificultades que se presentan al intentar estimar los grados de especiación en los peces es determinar si las diferencias observadas entre las poblaciones son fenotípicas o tienen, en parte o en su totalidad, una base genética. Otro problema consiste en medir el tiempo geológico, pero este impedi- mento está siendo constantemente disminuido mediante la introducción de téc- nicas mucho más precisas.
En general, para los peces, el periodo de tiempo de unos 10,000 a 12,000 años que ha transcurrido desde la terminación de la última glaciación Wisconsin es usualmente asociado con la diferenciación a un nivel no mayor del de subespecie. La mayoría de las especies tienen por lo menos una antigüedad de un millón de años, y muchas la tienen mayor. Sin embargo, la adquisición del rango de especie verdadera, bajo especiales circunstancias, debe haber tenido lugar dentro de los pasados 30,000 a 60,000 años, o inclusive en sólo 4,000 años. La especiación puede ser especialmente rápida en las poblaciones pequeñas, aisladas. Muchos de los géne- ros ahora vivientes probablemente se desarrollaron por el Mioceno o el Plioceno, otros posiblemente poco tiempo después de que terminó el Pleistoceno. El registro fósil de la Era Cenozoica es todavía muy inadecuado para servir de mucha ayuda en la estimación de las velocidades evolutivas. Muchos fósiles de las postrimerías del Pleistoceno, originalmente interpretados como especies diferentes, difieren muy poco, según se ha visto, o de plano nada, de sus descendientes vivientes. Por lo general, los periodos de mayor cambio geológico y los periodos de evolu- ción rápida están correlacionados, mientras que las etapas muy largas de uniformi- dad geológica muestran cambios evolutivos lentos. Hubo una explosión evolutiva en las eras Silúrica y Devónica, cuando la Tierra estaba siendo colonizada por plantas y animales. Estas evoluciones explosivas, aunque en una escala mucho me- nor, muchas veces han sido recurrentes. La congregación de especies -la evolución de muchas especies de un género ímico a través de la radiación adaptiva en un área restringida- proporciona un ejemplo relativamente reciente en los lagos africanos.
REFERENCIAS ESPECIALES SOBRE GENETICA Y EVOLUCIdN
Blair, W. Frank (ed.). 1961. Vertebrate speciation. University of Texas Press, Austin. Brooks, J.L. 1950. Speciation in ancient lakes. Quart. Rev. Biol., 26:30-60 y 131-176. Chen, T.R., y A.W. Ebeling. 1966. Probable male heterogamety in the deep-sea fish Bathylagus
Chen, T.R., y F.H. Ruddle. 1970. A chromosome study of four species and a hybrid of the
Dobzhansky, Theodosius. 1951. Genetics and the origin of species, 3ra. ed. Columbia Univer-
wesethi (Te1eostei:Bathylagidae). Chromosoma (Berl.), 18:88-96.
killifish genus Fundulus (Cyprinodontidae). Chromosoma (Berl.), 29:265-267.
sity Press, Nueva York.

3 88 GENBTICA Y EVOLUCION
Ebeling, A.W., y T.R. Chen. 1970. Heterogamety in teleostean fishes. Trans. Amer. Fish, Soc.,
Fryer, Geoffrey. 1959. Some aspects of evolution in Lake Nyassa. Euolution, 13:440-451. Gordon, Myron. 1957. Physiological genetics of fishes. En The physiology offishes. Academic
Press, Nueva York, 2:431-501, figs. 1-16. Greenwood, P.H. 1965. The cichlid fishes of Lake Nabugabo, Uganda. Bull. Brit. Mus. Nat.
Hist., Zool. 12(9):315-357. "-. 1973. The cichlid fishes of Lake Victoria, East Africa: The biology and evolution of a
species flock. Bull. Brit. Mus. Nal. , Zoo., suppl. 6:l-134. Myers, George S. 1960. The endemic fish fauna of Lake Lanao, and the evolution of higher
taxonomic categories. Evolution, 14:323-333. Nogusa, Shyunsaku. 1960. A comparative study of the chromosomes in fishes with particular
considerations on taxonomy and evolution. Mem. Hyogo Univ. Agr., 3(1):1-62, lams. 1-6. Sadoglu, Perihan, 1957. Mendelian inheritance in the hybrids between the Mexican blind
cave fishes and their overground ancester. Verh. Deutsch. Zool. Gesell. Graz, 1957:
Schultz, R. Jack. 1969. Hybridization, unisexuality, and polyploidy in the teleost Poeciliopsis
-. 1971. Special adaptive problemsassociated with unisexual fishes.Amer. Zool., 11,351-360. Schwartz, Frank J. 1972. World literature to fish hybrids with and analysis by family, species,
Tax, Sol (ed.). 1960. Evolution after Darwin. vols. 1-111 University of Chicago Press, Chicago. Uyeno, Teruya, y Robert Rush Miller. 1971. Multiple sex chromosomes in a Mexican cyprin-
White, M.J.D. 1973. Animal cytology and evolution, 3ra. ed. Cambridge University Press,
99(1):131-138.
431-439.
(Poeciliidae) and other vertebrates. Amer. Nat . , 103(934):605-619.
and hybrid. Gulf Coast Res. Lab., Publ. 3 :1-328.
odontid fish. Nature, 231(5303):452-453.
Londres.

Capítulo 13
Sistemática y nomenclatura
Sistemática es la ciencia relacionada con las clases y diversidad de los seres vivien- tes y con su disposición en una clasificación natural. Siendo básicamente un estu- dio de las relaciones evolutivas de los organismos, la sistemática reposa sobre los fundamentos de la biología al asegurar una estructura confiable, y es elemental para comprenderlos. Un sistema de clasificación proporciona los medios para esclarecer el problema del origen y evolución de la vida. Es erróneo suponer, como muchos lo hacen, que la clasificación es una etapa ya pasada de moda en la histo- ria natural. Cada uno es, aun involuntariamente, un clasificador, ya sea debido a la necesidad o simplemente por mera curiosidad. El origen de la información necesaria para hacer la clasificación está en la taxonomía, o sea, en el estudio de las bases teóricas, principios y procedimientos que deben seguirse para obtener un mejor entendimiento de las relaciones. La sistemática incluye tanto a la taxo- nomía como a la clasificación.
Todas las relaciones que hay entre los seres vivos son los resultados directos de lo que les ha sucedido durante su historia evolutiva; esa historia es un fenó- meno real, ya sea que hayamos tenido o no la habilidad de reconstruirla en forma precisa. Todas las aproximaciones al entendimiento de la sistemática de los orga- nismos quedan limitadas al estudio de las similitudes y diferencias y son en prin- cipio básicamente comparativas. El interés renovado por los métodos y la teoría de las clasificación ha recibido gran realce por el desarrollo de las computadoras y el advenimiento de nuevas áreas de investigación designadas en forma global como taxonomía numérica, quedando incluida entre ellas la cladística. La taxonomía es la teoría y práctica de la clasificación de los organismos. Es importante distinguir desde un principio entre clasificar y nombrar o identificar las cosas, porque estas dos actividades son totalmente diferentes. La clasificación comprende la filosofía
389 i

390 SISTEMATICA Y NOMENCLATURA
científica que utiliza procedimientos inductivos, mientras que la identificación emplea procedimientos deductivos que nos permiten ubicar a los individuos en taxa previamente establecidos. La nomenclatura zoológica comprende la aplica- ción de los diferentes nombres dados de acuerdo a reglas ya establecidas.
La función primordial de la clasificación es la de crear el orden donde se encuentra el caos, mediante la identificación correcta de los individuos y su ubi- cación o disposición en varios taxa, desde el momento en que es imposible discu- tir o pensar en los organismos sin haberles asignado primero nombres. Para hacer esto también se requiere de la aplicación de la nomenclatura biológica a los grupos (taxa) que san reconocidos. En auxilio de l a clasificacibn está la conveniencia. Las categoría del sistemático están basadas en el grado de simili.tud, de tal ma- nera que cuanto más estrecha sea la relación de dos organismos, usualmente serán los caracteres que compartan. Mucho antes de 1735 fueron aplicados los nom- bres latinos a las plantas y los animales, pero esa fecha corresponde al año en que Linneo procuró por primera vez catalogar a todas las clases conocidas; y sin duda se hizo aparente, inclusive en las sociedades más primitivas, que era inútil hacer observaciones en una planta o animal a menos que uno supiera su nombre. Los aztecas tenían considerable conocimiento de las especies que habitaban su región antes de la conquista espafiiola; y todavía más, ellos empleaban nombres que, me- diante la agregaci6n de los prefijos apropiados, servían para indicar sus relacio- nes o parentesco.
Esto conduce a una segunda función de la clasificación, la que sirve como una guía para establecer los parentescos. La clasificación moderna está basada en la filogenia, pero la clasificación y la filogenia son cosas distintas y no deben ser confundidas. La filogenia es la historia evolutiva actual de los organismos y es natural, continua y dinámica. La clasificación es el resultado de los esfuerzos humanos para interpretar o reconstruir la filogenia, y es arbitraria en el sentido de que muchas clasificaciones pueden ser elaboradas a partir de la misma filoge- nia. La historia evolutiva de un grupo vincula a una, y sólo a una, filogenia; pero, al desarrollar una interpretación confiable sobre el origen y evolución de ese grupo, los científicos proponen varios esquemas de clasificación que sufren cam- bios a medida que llegan nuevas luces en el conocimiento del curso de la evolución.
El problema fundamental de la sistemática es, como Huxley certeramente lo dijo, “el de detectar a la evolución en acción”. El sistemático moderno utiliza la información que le llega de una gran vuiedad de fuentes o disciplinas (genética, ecología, paleontología, fisiología comparada, comportamiento animal, citolo- gía, zoogeografía, bioquímica, biometría y otros campos relacionados). La infor- mación panorámica proporcionada por esas disciplinas es esencial para obtener la solución de muchos problemas taxonómicos. La sistemática se ha transformado en el punto focal para muchos campos de la biología, desde el momento en que estudia la diversidad orgánica, una rama integral superior de la biología.
DISPONIBILIDAD APROPIADA DE LOS PECES
Los peces son animales particularmente apropiados para hacer estudios sobre sistemática, porque: a) son abundantes en especies y número, de distribución muy amplia y fácilmente disponibles; b ) poseen muchos caracteres que son muy apro- piados para hacer análisis taxonómicos y tratamientos estadísticos; c) las correla-

ANTECEDENTES HISTdRICOS 391
ciones de su medio ambiente son bien marcadas; d ) muchos han probado ser apropiados para realizar análisis experimentales bajo condiciones de laboratorio, y e ) la intergradación y la hibridación son relativamente comunes y proporcionan comprobaciones de las relaciones genéticas.
ANTECEDENTES HISTdRICOS
No se duda que los comienzos de la taxonomía anteceden a los registros históri- cos, desde el momento en que una de las características del hombre consiste en nombrar y ordenar las cosas. Aristóteles (383-322 a.c.) sintetizó el conocimiento de su tiempo y lo formuló en lo que consideramos los principios de una ciencia. Aunque no propuso una clasificación formal, Aristóteles proporcionó una base para lograrla cuando afirmó que “los animales pueden ser caracterizados de acuer- do a su manera de vivir, sus acciones, sus hábitos y sus partes corporales”. Su filosofía prevaleció durante cerca de 2,000 años. De los zoólogos que vivieron un poco antes de Linneo, John Ray (1627-1705), el hijo de un forjador britá- nico, propuso un sistema de clasificación más natural, que el de sus predecesores y su trabajo influyó notablemente en Linneo (1707-1778). Este gran naturalista sueco fue el primero en aplicar con firmeza lo que hoy conocemos como sistema binomial de nomenclatura (véase abajo), en la décima edición de su monumental Systernu Nutume (1758), que marca el punto de partida de la nomenclatura zoo- lógica. Sus contribuciones influyeron tanto en los estudiosos que le sucedieron que, con justa razón, es llamado padre de la taxonomía. Linneo también propor- cionó una jerarquía de categorías: variedad, especie, género, orden, clase y reino. Los principales cambios que ocurrieron después consistieron en agregar la familia y el phylum. Era tan práctico su sistema que pronto fue adoptado, difundido y mejorado, y dominó el campo de estudio durante la siguiente centuria, en la cual se consideró a las especies como inmutables.
La revolucionaria teoría de la evolución de Carlos Darwin produjo, un siglo más tarde, un estímulo tremendo en el pensamiento y en la labor de investigación biológicos. Los hombres de ciencia de las décadas que siguieron de inmediato a la publicación de El Origen de las Especies (1859) se interesaron principalmente por saber si realmente los organismos vivientes son descendientes de ancestros comu- nes. La filogenia era, por consiguiente, la preocupación clave de este periodo. El ár- bol filogenético fue introducido por Haeckel (1866), y probó ser un método útil y estimulante, auxiliando a los taxónomos con un medio gráfico en el que se expresan las presuntas relaciones de las especies. Este era un periodo produc- tivo y excitante en la historia de la taxonomía, y algunas de las mentes más inteli- gentes fueron atraídas a este campo ante la perspectiva de ser premiadas con los descubrimientos que casi a diario se hacían de nuevas especies y géneros y, con no poca frecuencia, de nuevas familias u órdenes. Sin embargo, hacia las postrime- rías del siglo diecinueve o antes, se había terminado el periodo de los grandes descubrimientos entre los animales superiores. Quienes estaban ansiosos por des- cribir nuevos órdenes, familias y géneros, acudieron ahora al afinamiento de la clasificación y a dividir las categorías ya existentes. Como era de esperarse, algunas de estas divisiones fueron necesarias y benéficas, pero en otros casos produjeron una desintegración de las categorías naturales mediante el ocultamiento de las ver- daderas afinidades. Parte del descrédito en que cayó la taxonomía al finalizar el siglo

392 SISTEMATICA Y NOMENCLATURA
diecinueve y en los primeros años del actual es atribuible a esa excesiva digre- gación.
La fase más reciente en el desarrollo de la sistemática se ha caracterizado por el estudio de la evolución de las especies. Han ampliado notablemente las pers- pectivas de la sistemática hasta hacerla avanzar a un grado superior, como ciencia genuina y vigorosa, el redescubrimiento de las leyes de Mendel, el surgimiento de la genética y más tarde el de la genética de las poblaciones, el empleo de la biolo- gia experimental, la introducción de la biometría y, en consecuencia, el de la taxonomía numérica, ¡os avances de la ecología, el estudio del comportamiento comparado y el desarrollo de nuevas herramientas de trabajo. Los antecedentes de estas ciencias son esenciales para iograr la solución de muchos problemas de sistemática. El reconocimiento que finalmente surgió señala que los organismos con los que el sistemático debe trabajar “no son una jaula llena de animales de laboratorio ni una hilera de especímenes muertos en un museo, sino en lugar de elfo, las poblaciones naturales de seres vivientes en el campo” (Myers, 1941). El sistemático moderno se ha vuelto inclusive más interesado en Eormular genera- lizaciones, para lo cual la nominación y descripción de nuevos taxa es meramente el paso inicial.
LA TAREA DE LOS SXSTEMÁTICOS
Ei trabajo final del sistemático consiste no solamente en reconocer y describir las disposiciones infinitas de la vida que nos rodea, sino en contribuir también a su comprensjiin. Es critica la necesidad de cont.ar con identificaciones correctas para facilitar el trabajo de los administradores de la vida silvestre, los geólogos, los eciiiogos y los biólogos experimentales, para hablar solamente de unos cuantos, Para ser competente en el estudio de un grupo particular de animales se requiere realizar un trabajo difícil y tener una gran experiencia, y puede que no se logren conseguir algunas determinaciones correctas de organismos porque simplemente no haya disponible un especialista en el grupo de investigación. El sistemático moderno es uu naturalista de campo competente quien estudia la ecología y conducta de las especies en su medio natural. La gran amplitud de la sistemática proporciona oportunidades para hacer estudios, de acuerdo a sus intereses y talen- tos, aunque sean de los más diversos.
Después de reconocer y describir con minuciosidad a la especie, el sistemá- tico deberá ubicar enseguida a la especie en el orden que le corresponde, colocán- dola o adscribiéndola en la hilera del taxón relacionado, y elaborar un esquema que oriente sobre las relaciones de la especie con las categorías superiores. Este paso comprende más especulacion que el de dar nombre a la especie y es más tei;rico, desde el momento en que el taxónomo debe decidir si las similitudes señalan una convergencia o una verdadera relación filogenética. & I ha pasado ahora de la etapa analítica a la sintética.
La tercera tarea del sistemático consiste en estudiar cómo es que se han formado las especies, y esto conduce a la consideración de los factores de la evo- luciiin: jcómo es que se originan las especies, cbmo están relacionados y qué significa esta relación? El sistemático, en esta fase de su trabajo, se pone en mris intimo contacto con las demás disciplinas biológicas como la genética y la fisio- logía, la ecologia y la citología, y la biogeografía y la paleontología.

CONCEPTOS TAXON6MICOS 393
Estas diversas etapas a través de las cuales pasa la clasificación de un grupo particular han sido denominadas taxonomía alfa, beta y gamma. La taxonomía alfa se refiere al nivel en el cual la especie ha sido reconocida, caracterizada y nominada; la taxonomía beta, a la colocación de la especie en un sistema natural de clasificación; y la taxonomía gamma, al análisis de la variación intraespecífica y los estudios de la evolución. Hay entrecruzamiento e intergradación entre las tres, pero el curso del estudio es real. Aunque el taxónomo dotado de un método biológico puede aspirar a concentrarse en la taxonomía gamma, hay todavía una necesidad real de conducir estudios todavía más detallados en los niveles alfa y beta, especialmente de los grupos menos conocidos, incluyendo a los peces.
CONCEPTOS TAXONOMICOS
En la taxonomía clásica los organismos fueron dispuestos desde las especies hasta las categorías más elevadas, basándose en un conjunto de caracteres presentes presumiblemente en cada miembro de un grupo dado. En la taxonomía moderna los conceptos de variación dentro de un grupo en un tiempo dado y en diferentes periodos del tiempo geológico están volviéndose decisivos en la caracterización de las categorías.
La clasificación comienza con el individuo. Los individuos que mantienen relaciones muy estrechas entre sí y se cruzan con éxito son agrupados como espe- cie. Las especies son agrupadas en géneros, que pueden contener una o más de aquéllas. Los géneros relacionados son entonces agrupados en familias, cada fami- lia conteniendo uno o más géneros. Enseguida las familias constituyen órdenes, y los órdenes son agrupados en clases.
La presente evidencia señala que las especies estrechamente relacionadas difieren marcadamente entre sí en los detalles de su funcionamiento bioquímico como en su morfología externa o interna. Las investigaciones más recientes para descifrar las relaciones interespecíficas están enfocadas a los estudios cromosómi- cos, en las secuencias de los aminoácidos y en el comportamiento comparativo, entre otros. Aunque el estudio de los ejemplares preservados proporciona valiosos datos sistemáticos, es imperativo obtener una visión más amplia mediante el cono- cimiento del organismo en su hábitat natural.
Por lo tanto, aunque las especies son más a menudo definidas mediante el empleo de grupos de caracteres morfológicos, y por su variación geográfica, en algunos casos puede haber disponible considerable información biológica (como en la conducta, la genética, la fisiología y la ecología), inclusive considerando al elemento tiempo. La especie es un concepto basado en el de población, y deberá ser definida de acuerdo a un grupo particular de circunstancias. En consecuencia, debe haber diferentes clases de especies en diferentes grupos de animales o in- clusive en el mismo grupo. Las especies monotípicas muestran características uniformes en toda la serie, mientras que las politípicas están formadas de sub- especies diferenciadas. Especies hermanas (crípticas) son las que no se pueden distinguir morfológicamente o son muy parecidas, pero genéticamente están muy diferenciadas, así como por los aspectos fisiológico, ecológico o conductal, qu? implica al aislamiento reproductor. Las subespecies constituyen una pobla- ción geográficamente segregada, o grupo de poblaciones locales que difieren taxonómicamente de otras subdivisiones de una especie.

394 SISTEMATICA Y NOMENCLATURA
El género tiende, más que cualquier otra categoría, a retener su status clá- sico y a ser definido en la práctica como un grupo de especies que comparten ciertos caracteres.
La familia como categoría incluye, por lo menos teóricamente, muchas lí- neas filéticas diferentes y de aquí que su valoración tome en consideración la variación con respecto al tiempo y también al espacio.
LOS DATOS PARA LA CLASIFICACI6N (FIGURA 13.1)
¿Qué clase de caracteres son utilizados por los ictiólogos en la identificación de los peces o al proponer una clasificación? Los caracteres son tantos y tan varia- dos y, como el oro, que está donde usted lo encuentra, Y a sea que ellos sean rasgos merísticos -como el número de escamas, los radios de las aletas o vértebras-, proporciones del cuerpo, colores del animal vivo, características osteológicas, pa- trones de conducta o número de cromosomas, tienen una característica común: es probable que cualquiera de ellos tenga diferente significado en los diferentes grupos de peces. Por ejemplo, el número de radios caudales es virtualmente cons- tante en muchas familias y, en consecuencia, no puede ser usado en los grupos situados por debajo del nivel de familia, pero en los Cyprinodontidae su número puede distinguir razas, subespecies o especies, aún sin nombre. El número de vértebras, muy consistente en la mayoría de los peces con espinas y radios -por consiguiente un carácter ordinal- puede ser empleado para distinguir categorías desde la subespecie hasta el género en los grupos más primitivos. La ausencia de aletas pélvicas puede ser una característica de familia o una variación individual (como ocurre en algunos ciprinodóntidos desérticos). En los pececillos de acuario (Cyprinidae) el tamaño relativo de la cabeza puede ser empleado para dis- tinguir entre una y otra especie; y aún así, en los peces sol (Centrarchidae) el tamaño de la cabeza tiene importancia para determinar una familia. Por lo tanto, no puede haber un concepto fijo sobre el nivel taxonómico al que un carácter dado puede pertenecer.
Por lo general, los caracteres que pueden ser vistos externa y fácilmente son los que se utilizan para separar especies e inclusive hasta géneros. Estos carac- teres son, sin embargo, muy a menudo los más superficiales y los más variables, y de aquí que deban ser empleados con precaución. Los rasgos empleados para dis- tinguir las categorías superiores a los géneros, familias etc., son generalmente más profundos y menos variables. Ciertos caracteres osteológicos, por ejemplo, mues- tran poca variación y parecen estar menos sujetos a las influencias del medio (como los efectos de la temperatura) que esos caracteres merísticos, como por ejemplo el número de escamas. El color tiene, a menudo, valor de carácter espe- cífico, pero no puede caracterizar a los grupos superiores, como lo hace el color rojo de los peces ardilla (Holocentridae).
Cuanto más diversos sean los datos empleados en la clasificación, más rea- lista resultará la disposición de las especies. El sistemático moderno puede tener la facilidad de adquirir información de campos tan variados como la bioquímica, la genética, el comportamiento, la ecología, la fisiología, la distribución geográfica, la paleontología y la citología para suplementar y reforzar los datos de laborato- rio más convencionales sobre la morfología y la anatomía. La mayor parte de la clasificación es realizada, sin embargo, sin tener un conocimiento definido sobre

COLECCIONES DE ESTUDIO 395
Figura 13.1 - Un pez con espinas y radios ilustrando las partes y metodos del conteo y medi- ci6n: 1 -interorbital; 2-occipital; 3-nuca; 4-longitud de la cabeza; 5-longitud predorsal; 6-longitud estándar; 7-longitud al escote; 8-longitud total; 9-longitud de la base de la primera aleta dorsal o aleta espinosa; 10-una de las espinas de la primera aleta dorsal; 11 -espi- na de la segunda aleta dorsal; 12-altura de la segunda aleta dorsal; 13-longitud del borde distal, externo o libre, de la segunda aleta dorsal; 14-uno de los radios blandos de la segunda aleta dorsal; 15-longitud del hocico; 16-longitud del ojo; 17-longitud cefllica postorbital; 18-escamas sobre la linea lateral o series laterales que son contadas; 19-altura máxima del cuerpo; 20-uno de los poros de la línea lateral en una línea lateral completa; 21 -una de las escamas laterales que con las demás forma la serie de la línea lateral; 22-longitud de la base de la segunda aleta dorsal, o aleta blanda; 23-altura mínima o del pedúnculo caudal; 24-la aleta pectoral; 25-uno de los radios blandos de la aleta pectoral; 26-regi6n abdominal (vientre); 27-escamas bajo la línea lateral o series laterales que son contadas; 28-longitud de la base de la aleta anal; 29-longitud del pedúnculo caudal; 30-longitud de la mandíbula superior; 31 -istmo; 32-pecho; 33-altura de la espina pélvica; 34-altura de la aleta pblvica; 35-uno de los radios blandos de la aleta p6lvica; 36-espinas de la aleta anal; 37-radios blandos de la aleta anal; 38-radios rudimentarios de la aleta caudal; 39-uno de los principales radios blan- dos sin ramificar de la aleta caudal; 40-radio blando ramificado de la aleta caudal; 41 -la aleta caudal con el número en lo más profundo del escote de la misma. (Fuente: Trautman, 1957).
los antepasados de los grupos tratados, su potencial reproductor, la conducta y mucha otra información útil. Lamentablemente esos son datos por lo general olvida- dos, pero continuamos clasificando sin ellos y hacemos un trabajo razonablemente bueno, por lo menos hasta los niveles de especie y género.
COLECCIONES DE ESTUDIO
Las colecciones de museo constituyen el depósito de la materia prima, en su ma- yor parte, en la que se basan las conclusiones de los sistemáticos. Esas colecciones son similares a las grandes bibliotecas, en el sentido de que deben estar eficiente-

3 96 SISTEMATICA Y NOMENCLATURA
mente catalogadas para ser útiles y siempre siguen en continuo crecimiento. A diferencia de una biblioteca, los avances en los conocimientos de 10s grupos de seres vivientes obligan a los museos modernos y eficientes a efectuar una recatalo- gación continua con el fin de actualizar la clasificación de un género en particular, una familia u otra categoría. Mientras que al principio los colectores preservaban solamente unos cuantos de los llamados representantes típicos de cada especie, la práctica moderna pone especial énfasis en la reunión de grandes series de espe- címenes colectados dentro del área de distribución conocida de la especie, porque el taxónomo moderno es un estudioso de las poblaciones. Las muestras deben ser adecuadas para dar una imagen clara de la variación, el crecimiento, etc., y permi- tir el análisis estadístico de los datos.
Las colecciones de los museos pueden poner énfasis en los peces de una región particular -por ejemplo, las aguas dulces de Norte y Centroamérica- o pueden ser cosmopolitas, esforzándose por tener algunos ejemplares representati- vos de todas las especies que sea posible adquirir, de diversas regiones del mundo. Las colecciones de peces están formadas principalmente por ejemplares fijados er, formol y preservados en alcohol, pero una tendencia reciente por investigar en el campo de la clasificación superior está resultando en un inter66 creciente por conservar material esquelético y ejemplares transparentados y teñidos. Muchos museos tienen ahora aparatos de rayos X para colaborar en la localización de caracteres sistemáticos, así como para estudiar los efectos de la radiación en las poblaciones vivientes.
Cierta información esencial debe acompañar a cada colección o ejemplar. El dat,o más importante consiste en la localización precisa, ya que un ejemplar que no esté correctamente etiquetado no tendrá virtualmente utilidad para el siste- mático. Si la localidad se refiere a un lugar desconocido, su posición relativa con respecto a un lugar bien conocido deberá ser dada en la etiqueta original. El estado o país, el condado o distrito, la fecha de la colecta y el nombre del o los colecto- res, también deben ser registrados. Es de gran utilidad citar la elevación (aún cuando sólo sea estimativa), y es de mucho valor la información ecológica adicio- nal. La etiqueta deberá ser escrita en el campo, en el momento mismo de hacer la colecta, deberá ser colocada en el baño fijador (usualmente formol al l o%, lo que equivale a una solución acussa de formaldehído al 4%) junto con los ejempla- res, y no deberá ser repuesta más tarde. Es necesario emplear una etiqueta de buena calidad, porque no importa el cuidado con el que se registraron los datos, pues si la etiqueta se deshace se perderá el valor científico de la colecta. Es mejor escribir en las etiquetas con tint,a china indeleble negra, pero un lápiz blando produce resultados satisfactorios. Los ejemplares que tienen varios centímetros de longi- tud deberán ser cortados a lo largo del lado derecho del abdomen antes de ser sumergidos en el formol; los peces de varios kilogramos deberán también ser cor- tados en los músculos de cada lado de la columna vertebral. Si los ejemplares van a ser conservados durante algunas semanas en la solucijn de formol, deberá agregárseie bórax (una cucharada cafetera por cada dos litros de solución fija- dora = 5 gramos12 litros) para neutralizar el efecto ácido, retardando en conse- cuencia la contraccihn de los ejemplares, endureciendo sus partes blandas, Y pre- viniendo la descalcificación de los tejidos óseos.
NO todos los peces pueden ser preservados por entero; a los ejemplares grandes se les puede quitar la piel para secarla o colocarla en un liquido preserva- dor. El material que se pretenda armar como esqueleto en el laboratorio (mediante

NOMENCLATURA ZOOLdGICA 397
el auxilio de los escarabajos derméstidos) puede ser preparado en el campo conser- vándolo bajo refrigeración o, si no hay disponible hielo, mediante su preparación en seco. ÉSta se logra descarnando, eviscerando (asegurándose de registrar el sexo y la longitud) y colocando entonces el ejemplar en una caja de tela de alambre, sellada contra las moscas, para que se seque. Al regresar al laboratorio se le dará al ejemplar un breve remojón en amoníaco, el cual reblandecerá al pez seco lo suficiente para que los derméstidos realicen su trabajo.
NOMENCLATURA ZOOL6GICA
Es esencial distinguir entre clasificación y nomenclatura, ya que sus correspon- dientes papeles son a menudo mal comprendidos. La clasificación es un asunto biológico, mientras que la nomenclatura es la herramienta por medio de la cual son proporcionadas las etiquetas para las categorías taxonómicas, de tal modo que los biólogos puedan comunicarse entre sí. La nomenclatura es un procedi- miento destinado a un fin más, que un fin en sí mismo. Desde el momento en que los zoólogos trabajan con animales y emplean sus nombres, es esencial que se familiaricen por sí mismos con los principios generales de la nomenclatura zooló- gica -en el lenguaje del zoólogo-, sea sistemática o no.
Con el fin de que sea de utilidad general, la nomenclatura debe no solamente ser de amplia aplicación, sino que las mismas palabras que emplea deben ser enten- didas por todos. La mira principal de la nomenclatura es lograr la estabilidad y la uniformidad. Sin embargo, el progreso en la sistemática ha llevado al cambio en los conceptos de las categorías taxonómicas y sus relaciones; en consecuencia, los cambios de nombres son inevitables.
La nomenclatura científica moderna comprende un sistema binomial, uni- formado por primera vez por Linneo. Antes de é1 los organismos fueron nom- brados, generalmente en latín, mediante frases descriptivas, oraciones o párrafos, lo que probó ser un sistema incómodo e impráctico. Linneo condensó esos nombres en sólo dos palabras latinas: una, el género, para indicar la clase general, la otra, la especie, para designar la clase particular. Por lo tanto, el sistema binomial es un sistema formado por dos nombres. Desde el momento en que Linneo aplicó por primera vez, en forma consistente, el sistema binomial en las plantas y animales en su décima edición del Systerna Naturae (1758), este trabajo es designado en el Código Internacional como el punto inicial para la nomenclatura zoológica (y botánica). Los nombres publicados desde entonces deben adaptarse a los princi- pios del sistema binomial para que sean aceptados.
El sistema binomial se ha ampliado incluyendo entonces a las subespecies, de aquí que ahora contemos con un sistema trinomial. Muchos de los géneros linneanos constituyen ahora nuestras familias, e inclusive órdenes, porque Linneo conoció sólo menos de diez mil especies de animales, mientras que nosotros cono- cemos bien más de un millón. Sin embargo, el sistema de Linneo todavía tiene aplicación; por tal motivo ha sido universalmente aceptado.
Aunque el sistema es básicamente simple pueden surgir muchas complica- ciones en su aplicación. La confusión se manifestó muy pronto debido a la ausencia de un código de procedimientos aceptado, ya que no fue sino hasta 1842 cuando fue propuesto una suerte de código “oficial”; en ese año, un comité de la Aso- ciación Británica para el Avance de la Ciencia publicó el “Stricklandian Code”.

398 SISTEMATICA Y NOMENCLATURA
Con los agregados de 1865 éste sentó la base para gran parte de la codificación de las reglas de la nomenclatura zoológica. En años subsecuentes fueron propues- tos otros códigos (francés, norteamericano y alemán) y la situación se volvió caótica hasta que se llegó a un acuerdo general en 1901, fecha en que se adoptó uno y el comité que lo elaboró se transformó en una organización permanente, la Comisión Internacional de Nomenclatura Zoológica.
En la actualidad, la Comisión Internacional no tiene un número fijo de miembros, pero el mínimo es de dieciocho; ellos representan diferentes disciplinas y regiones geográficas y son elegidos cada 10 años en el Congreso Internacional de Zoología (que se reúne cada 5 años), en clases que terminan en intervalos de 3 años. Las tareas generales de la Comisión consisten en “estudiar el caso general de la teoría y práctica de la nomenclatura zoológica”. Sus poderes se han mani- festado de cinco importantes maneras: a) dando recomendaciones al Congreso Internacional para hacer correcciones al Código Internacional, o para la adición de “Recomendaciones” a determinados artículos del Código en particular; b ) pro- porcionar (desde 1907) “Opiniones” a las consultas que se le hacen en relación a la nomenclatura zoológica (pero las mismas, desde 1939, constituyen decisiones con relación a la situación de un nombre en particular); c ) mediante la compilación (desde 1910) de la “Lista Oficial de los Nombres Genéricos en Zoología (nomina conservanda) y otras listas e indices como esa; d ) publicando (desde 1939) “De- claraciones” que constituyen interpretaciones de las Reglas, y e ) mediante el em- pleo (desde 1913) de los “Poderes Plenarios” (nombres retenidos por suspensión de las reglas).
Desde 1939, a instancias de Francis Hemming (Inglaterra), Secretario de la comisión desde 1936 hasta 1957, la Comisión ha creado el Bulletin of Zoological Nomenclature (1943) y una serie de Opinions and Declarations Rendered by the International Commission on Zoological Nomenclature (1939). Las propuestas enviadas a la Comisión para su resolución, los comentarios de los zoólogos y la correspondencia con los mismos en relación a estas propuestas, así como los ar- tículos sobre las implicaciones de la nomenclatura en los desarrollos de la teoría y práctica taxonómicas, son publicados en el Bulletin.
Una revisión de las reglas apareció como Código Internacional de Nomen- clatura Zoológica (1961. Int. Trust. Zool. Nomen., Londres), adoptado por el XV Congreso Internacional de Zoología, en su reunión que tuvo lugar en Lon- dres en 1958. La revisión fue publicada paralelamente en inglés y francés.
La jerarquía convencional de la nomenclatura puede ser resumida, como se indica para la perca amarilla, Perca flauescens:
Phylum Chordata Clase Osteichthyes Orden Perciformes Familia Percidae Género Perca Especie Perca fluuescens (Mitchill)
Mediante el empleo de los prefijos “sub” y “super”, los grupos anteriores pueden ser ampliados en quince categorías que cubren la mayor parte de las nece- sidades para expresar los conceptos de grupo en ictiología: phylum, subphylum, superclase, clase, subclase, superorden, orden, suborden, superfamilia, familia, sub-

NOMENCLATURA ZOOL6GICA 399
familia, género, subgénero, especie y subespecie. En algunos grupos grandes donde la clasificación es comparativamente bien conocida y las diversas líneas están re- presentadas, puede estar justificado el empleo de tribus (formadas agregando la terminación “ini” al tronco del género típico, por ejemplo, Cyprinini). Otra cate- goría útil es el grupo específico, algunas veces llamado superespecie, que dentro de la jerarquía encaja entre la especie y el subgénero. Puede ser empleado en lu- gar del subgénero cuando toda la clasificación de un grupo no ha sido todavía bien trabajada, pero se tienen agrupaciones naturales de especies estrechamente relacionadas.
El nombre genérico es siempre un nombre en singular, caso nominativo, y está escrito con la letra inicial en mayGscula. La parte específica del nombre cien- tífico está escrita con minúsculas; por ejemplo, Perca flauescens. Puede ser: 1) un adjetivo, el cual deberá tener el mismo género del nombre genérico; por ejemplo Gasterosteus aculeatus; 2) un nombre en aposición (un sustantivo) que siempre esté en el caso nominativo, pero no necesariamente debe corresponder con el género del nombre genérico, por ejemplo Cichlasoma rnaculicauda; 3) un nombre en el genitivo singular como ocurre en un patronímico, Etheostoma jordani (por David Starr Jordan), o Epinephelus clarkae (por Eugenie Clark), o 4) un nombre común en el genitivo plural, indicando usualmente algo relacionado con el hábitat, por ejemplo Riuulus paludium (significando “de los pantanos”).
Algunos de los nombres de los taxa que se encuentran sobre el nivel genérico también son formados de acuerdo con los principios generalmente aceptados. Por ejemplo, la tribu es una subdivisión de la familia, subordinada a la subfamilia, y formada mediante la adición de “ini” al tronco de un nombre genérico incluido (como Fundulini de Fundulus); la subfamilia, una subdivisión superior de la fami- lia, puede ser empleado en familias grandes, y se forma mediante la adición de “inae” al tronco genérico (por ejemplo Epinephelinae); la familia, comprendiendo un grupo de géneros relacionados o, excepcionalmente, un sólo género, es for- mada mediante la agregación de “idae” al tronco de un género incluido (por ejem- plo Percidae); y el orden, el taxón mayor situado inmediatamente arriba del nivel de la familia, está en este libro formado por la adición de “iformes” al tronco de un género incluido y válido (por ejemplo Clupeiformes). Lamentablemente los ictiólogos no están de acuerdo en la interpretación o aplicación de los nombres ordinales y el Código no especifica cómo formar los nombres que puedan estar sobre el nivel de familia.
Sabiendo que un individuo, u ocasionalmente dos o más personas, es el responsable de la propuesta y descripción de cada taxón, el nombre de la persona o personas es colocado después del nombre científico y es conocido como el autor del mismo. El nombre es escrito generalmente completo, pero se puede, cuando se trata de autores muy conocidos (como Linneo), ser abreviado. El he- cho de citar al autor no significa que se le quiera dar crédito por su trabajo, más bien se le señala la responsabilidad por el nombre que asignó y ayuda en la loca- lización de la descripción original. Cuando una especie es transferida a un género diferente al que se le asignó en la descripción, el nombre (o nombres) original del autor es colocado entre paréntesis.
La ortografía original de un nombre debe ser conservada a menos que haya habido un error de transliteración, un “lapsus calami”, o un error tipográfico. El nombre válido de un género, especie o subespecie puede ser el nombre más anti- guo que llene los requerimientos de la Ley de Prioridad. Esta importante regla fue

400 SISTEMATICA Y NOMENCLATURA
convenida para evitar confusión en la aplicación de los nombres cientificos y para eliminar la duplicación; por lo general, los nombres publicados primero adquieren precedencia sobre los nombres del mismo rango publicados más tarde. Si otros nombres son publicados en forma subsecuente para el mismo taxón, se vuelven sin& nimos, o nombres no válidos (el mismo organismo puede tener solamente un nom- bre legítimo, aunque en la literatura aparezca con muchos nombres sinónimos).
La nomenclatura zoológica es independiente de la nomenclatura botánica en el sentido de que el nombre de un animal no puede ser rechazado simplemente porque es idéntico al nombre de una planta. Los nombres genéricos y subgenéri- cos son, de acuerdo a la nomenclatura iguales y sujetos a las mismas reglas; las especies tipo del género y el subgénero típico deben ser lo mismo. De igual modo, los nombres específicos y subespecíficos tienen equivalencias por su nomenclatura y estlin sujetos a las mismas reglas.
En el caso de que dos o más nombres para la misma unidad taxonómica sean publicados en la misma composición (de aqui que sean simultáneos), es pri- vilegio del primer revisor seleccionar uno de estos nombres para considerarlos como válidos y colocar los otros en la sinonimia,
Desde el momento en que la identificación de una especie puede no ser fácilmente hecha a partir de la descripción original, se ha ideado un sistema cono- cido como método de tipo para confinar los nombres científicos a las entidades taxonómicas objetivas. El ejemplar tipo funciona como un “poseedor del nom- bre”. El autor de una nueva especie puede designar a determinado ejemplar como el tipo de esa especie. Solamente los tipos primarios son poseedores del nombre. Estos son: a) el hoZotipo ( o simplemente t ipo) , un ejemplar Único designado por el autor original de la descripción original, o el Único ejemplar conocido cuando la especie fue descrita; b ) el lectotipo, un ejemplar tipo seleccionado después de haber sido hecha la descripción original, de uno de los sintipos que forman la base de la descripción original; c ) el sintipo (a menudo llamado originalmente cotipo), uno de todos los ejemplares que forman !a base de la descripción original, cuando ningún ejemplar haya sido designado como tipo, y d ) el neotipo, un ejemplar se- leccionado para tipo después de haber sido hecha la descripción original, cuando se sabe definitivamente que el ejemplar ( o ejemplares) tipo ha sido perdido o destruido.
Otros tipos de uso común son: a ) el paratipo (que a menudo era llamado cotipo), cualquier ejemplar que no sea el holotipo empleado por el autor original para describir su nuevo taxón; b ) el alotipo, un paratipo del sexo opuesto al del holotipo; c ) el topotipo, cualquier ejemplar de la localidad típica colectado ahí después de publicada la descripción original, y d ) el paratopotipo o isotipo, cual- quier ejemplar que no sea el holotipo capturado en el mismo lugar que el holotipo e incluido en la descripción original. Los tipos no requieren ser “ejemplares típicos” y no pocas veces son muy aberrantes. El tipo nomenclatural de un género no es un ejemplar sino una especie, aquélla en la que el nombre genérico se basó. Origi- nalmente fue llamado genotipo, pero el nombre de la especie tipo es recomendado con el propósito de evitar conflictos al emplear el término genotipo también en genética.

REFERENCIAS 401
REFERENCIAS ESPECIALES EN SISTEMATICA Y NOMENCLATURA
Bailey, R.M., et al . 1970. A list of common and scientific names o f fishes from the United States and Canada. 3ra. ed. Special Publication núm. 6. American Fisheries Society, Washington, D.C.
Crowson, R.A. 1970. Classification and biology, Aldine Publ. Co., Chicago. Dobzhansky, T. 1970. Genetics o f the evolutionary process. Columbia University Press, Nueva
Hennig, W. 1966. Phylogenetic systematics. University of Illinois Press, Urbana. Huxley, J.S. (ed.). 1940. The new systematics. Ciarendon Press, Oxford. Mayr, E. 1942. Systematics and the origin of species. Columbia University Press, Nueva York. ”-. 1969. Principles of systematic zoology. McGraw-Hill Book Company, Nueva York. Mayr, E., E.G. Linsley, y R.L. Usinger. 1953. Methods and principles of systematic zoology.
Myers,,G.S. 1941. (Review of Huxley’s, The new systematics). Copeia, 1941(1):61, Schenk, E.T., y J.H. McMasters. 1956. Procedure in taxonomy, 3ra. ed., Stanford University
Simpson, G.G. 1961. Principles of animal taxonomy. Columbia University Press, Nueva York.
York.
McGraw-Hill Book Company, Nueva York.
Press, California.


Capítulo 14
E cología y zoogeograIía
La abundancia y distribución de los peces en las aguas de la Tierra son los produc- tos de la interacción entre los peces y los medios químicos, físicos y biológicos que les rodean. El estudio de las relaciones entre un organismo y su medio am- biente es el tema tratado por la ecología. El estudio de la distribución ictiológica que resulta de las relaciones ecológicas de los peces es la zoogeografía, o más específicamente, la ictiogeografía.
DEFINICIONES
La unidad básica de estudio en ecología es el individuo. Las aplicaciones de la ecología, sin embargo, se refieren comúnmente a las especies, que son grupos de individuos que se parecen más a sí mismos que a cualquier otra cosa y que produ- cen lo de su misma clase (reproducción verdadera) después de aparearse. Los individuos de una especie o los de todas las especies que ocupan un hábitat deter- minado forman una población. Los miembros de las poblaciones ocupan uno o más nichos ecológicos. Un nicho puede ser concebido como lo hace biológicamente un organismo tomando en cuenta todos los factores físicos, químicos, fisiológicos y bióticos que requiere para vivir; es, por lo tanto, una abstracción y no precisa- mente un espacio limitado físicamente. Su habitat es donde vive (aire, suelo, agua). El medio ambiente es todo lo que rodea al organismo ya sea biótico (viviente o muerto) y abiótico. Las poblaciones de peces y otros organismos que viven en el agua ocurren en comunidades en diferentes hdbitats. Finalmente, la suma de la comunidad más el hábitat que la sostiene puede ser considerada como un ecosis- tema abierto. Obviamente el concepto de ecosistema puede comprender a la
403

404 ECOLOG~A Y ZOQGEOGRAFÍA
'Tierra o ecósfera, o puede simplemente ser un estanque pequefio o estanque de marea o, más extensamente, otra vez, todas las aguas de !a Tierra, la hidrósfera. Por lo tanto un ecólogo está entonces interesado en las cantidades de materia y energía que pasan a través de un ecosistema dado.
AGUA
Todos los hábitats donde viven los peces y todas las comunidades en estos hábitats están cambiando continuamente. Todos son diferentes de un instante al otro, y todos tienden a desenvolverse gradualmente en direcciones reconocidas, a travks de la sucesión ecológica. La dinámica del ecosistema acuático depende primordial- mente de las propiedades del agua, entre las cuales se cuentan las siguientes. Su elevada capacidad calorífica previene las fluctuaciones extremadamente rápidas de la temperatura. Tanto las aguas dulces como las marinas alcanzan sus máxi- mas densidades cuando sus temperaturas están por encima del punto de congela- ción. Si no fuese así, muchas áreas acuáticas se congelarían hasta solidificarse, por lo menos en cada estación invernal, eliminando sus poblaciones de peces y de muchos otros organismos. El poder disolvente del agua es mayor, que el de cual- quier otro líquido, y este fenómeno físico permite el transporte rápido a buen número de materiales de los cuales depende finalmente la vida del pez.
La transparencia del agua es de primordial importancia para la cantidad de radiación solar (energía luminosa) que puede ser aprovechada por el fitoplancton y otras plantas acuáticas. En el ecosistema acuático total es el Sol la principal fuente de energía para los productores primarios. La transparencia, junto con la disponibilidad de los nutrientes disueltos para el fitoplancton, son características que básicamente pero indirectamente determinan la cantidad de peces que cualquier hábitat puede mantener.
PRODUCTIVIDAD ORGÁNICA EN LOS ECOSISTEMAS ACUÁTICOS
Transferencia de energia
La interacción continua de los componentes del ecosistema acuático se funda en la transferencia de energía. La luz es la fuente de la energía, de la cual solamente una fracción en los ecosistemas acuáticos aparece en la carne del pez. La energía es aprovechada primero por las plantas acuáticas, algunas grandes, otras pequeñas. La clave de la producción piscícola es el aprovechamiento hecho por algas micros- cópicas, desmidiáceas y diatomáceas, el fitoplancton. El fitoplancton ha sido con- siderado como la verdadera pastura en las aguas, proporcionando materiales ali- menticios que son directa o indirectamente convertidos en peces. La fotosíntesis es el proceso por medio del cual la energía luminosa es aprovechada para la fabri- cación de material orgánico. Durante este proceso las plantas verdes toman dióxido de carbono del agua y forman carbohidratos y otros componentes moleculares. Es posible que cada año sea fijada una cantidad mayor de carbono en la produc- ción primaria (primer nivel trófico) en los ecosistemas acuáticos que en los terres- tres. Continúa después el consumo por los animales y la transformación de los materiales vegetales. Los herbívoros, incluyendo a algunos peces, son los consu-

PRODUCTIVIDAD ORGANICA EN LOS ECOSISTEMAS ACUATICOS 405
midores primarios. Los grados subsecuentes de consumidores son predadores. Ya que hay únicamente un 10% de eficiencia en la conversión de la energía de un nivel trófico al siguiente, con solamente un pequeño incremento en la eficiencia en los niveles tróficos superiores, se comprende que cuanto más pequeña sea la cadena alimenticia, mayor será la probabilidad de que una especie de pez sobre- viva y abunde. Los ecólogos han comparado este concepto de producción con una pirámide, ya sea de biomasa o número de individuos. Los productores están en la base y exceden en masa y número; los consumidores primarios, secundarios, terciarios, etc., están en varios niveles intermedios de la pirámide; y los consumi- dores que tienen la cadena alimenticia más larga, como los más grandes predadores y/o los peces de los fondos marinos, están en la cúspide de la pirámide y son los que representan menos masa y menor número. En las cadenas alimenticias no parásitas, no solamente decrecen en número los predadores a lo largo de los esla- bones; también crecen mucho más en tamaño que su presa en eslabones sucesivos.
Ciclos biogeoquímicos
Los componentes clave disponibles para la síntesis dentro del protoplasma en la hidrósfera circulan en ciclos biogeoquímicos. Los ciclos reconocidos incluyen los del carbono, nitrógeno y fósforo. Las fuentes iniciales de los elementos clave, así como sus concentraciones, difieren a lo largo del año. Es así como en los ciclos de producción la atmósfera es la fuente del carbono (del dióxido de carbono) y del nitrógeno, en forma de gases. El fósforo se origina del suelo a través de la ero- sión o la filtración. La fase productiva de los ciclos biogeoquímicos es la fijación de los elementos en el protoplasma por los seres vivientes. El fitoplancton, por ejemplo, es capaz de fijar el carbono, el nitrógeno y el fósforo. Los peces que se alimentan directa o indirectamente del fitoplancton convierten en carne los com- puestos de estos tres elementos, fijados por las plantas, y otros. Los productos de desperdicio y los despojos de quienes consumen plancton y de los consumido- res en general, regresan los elementos clave directamente o a través de la descom- posición bacteriana a sus ciclos biogeoquímicos para ser vueltos a usar. Este proceso de regreso puede ser muy rápido o muy lento. Los consumidores de plancton, los peces y otros animales acuáticos pueden devolver muy pronto el dióxido de car- bono al ecosistema mediante la respiración. El fósforo, aprisionado en el esqueleto de un pez, puede tomar años para su liberación después de que el pez ha muerto y caído al fondo. La escena entera, sin embargo, corresponde a una continuidad, con las fuerzas de la producción alimentadas desde tierra y la atmósfera adminis- trando consistentemente aquellos que se liberan. La muerte es indispensable para guardar el equilibrio, pero es la cantidad de los materiales nutritivos originales que están disponibles en forma natural en un ecosistema la que limita el volumen de la biomasa que el sistema puede mantener. Por esta razón, es práctica común en acuacultura (la producción intensiva de peces y otros organismos útiles, más a menudo en estanques), como en agricultura, utilizar los fertilizantes para propor- cionar más sustancias nutritivas de las que el hábitat produce en forma natural.
El medio ambiente de los peces está influido por muchos factores además de los nutrientes, que pueden limitar las poblaciones o modificar la distribución geográfica. Cualquiera de estos factores puede ser limitante cuando está presente en cantidades ya sea demasiado pequeñas o demasiado grandes.

406 ECOLOGfA Y ZOOGEOGRAFÍA
CLASIFICACIONES ECOLdGICAS DE LOS PECES
Los organismos acuáticos, incluyendo a los peces, pueden ser clasificados ecológica- mente de diferentes maneras. De acuerdo a las tolerancias ambientales, pueden ser agrupados como estrecha o ampliamente tolerantes. A la expresión correspondiente se le asigna el respectivo prefijo, ya sea “esteno” (angosto) o “euri” (amplio). La clasificación para la temperatura es, por lo tanto, estenotermal o euritermal, para la salinidad es estenohalina o eurihalina, etcétera. A los peces también se les puede asignar su categoría sobre la base de su ubicación en los ecosistemas acuáticos: bénticos (cavadores de los fondos o peces de piso marino), pelágicos (de nado libre) o planctónicos (que dependen de las corrientes para realizar sus movimien- tos, como sucede con las larvas jóvenes de muchas especies). En los lagos y estan- ques, los peces de la zona litoral son los que viven en las entrantes de la playa, donde la luz penetra hasta el fondo y a menudo están presentes las plantas con raíces; los peces de la zona limnética son los que viven aguas adentro donde no hay plantas con raíces, y se extienden hacia el fondo hasta el nivel de compensación luminosa donde la iluminación es inadecuada para sostener la vida del fitoplanc- ton; y los peces de la zona profunda son los que viven en las aguas oscuras, por debajo del nivel de compensación de la luz. Las categorizaciones anteriores ilustran, incidentalmente, la estratificación de las comunidades en ecosistemas. Sin embargo, puede esperarse que varíe geográficamente la composición es especies de las zonas. En las corrientes de agua tenemos la división más obvia del hábitat y sus ocupantes, si consideramos dos zonas siempre subjetivas: los estanques y los rápidos. La corriente de los segundos es en general lo suficientemente rápida para mover la arena y el cieno, los cuales son después depositados en los estanques.
FACTORES ECOL6GICOS
Los factores del medio ambiente que inciden en las vidas de los peces son muchos, complejos e interrelacionados en sus efectos. Seleccionaremos unos cuantos como ilustración.
Temperatura
La temperatura es un factor de significado muy amplio y variado, como ya se ha mencionado en el Capítulo 5, sobre la alimentación y el crecimiento, y en el Capitulo 10, sobre el desarrollo, entre otros capítulos en que también se men- ciona. Por lo general, dentro del margen de temperaturas que pueden ser toleradas por el pez, los efectos se manifiestan en la aceleración de los procesos vitales por las temperaturas calientes y la desaceleración por las frías. Las temperaturas ex- tremas o los cambios súbitos son a menudo letales. Las temperaturas elevadas pero subletales pueden inducir a la estivación, mientras que las bajas, a la hibernación. Dentro del ciclo hidrológico la precipitación y la evaporación son fenómenos térmicos. Los peces de agua dulce, las especies marinas que emplean el hábitat dulceacuícola para su crecimiento, y los peces de las aguas salobres, dependen de este ciclo para obtener el agua suficiente para vivir.
La temperatura puede determinar, dentro de los cuerpos de agua, el éxito de

FACTORES ECOL6GICOS 407
una especie, así como su distribución. Dentro del área de distribución geográfica de las truchas de la familia Salmonidae, las aguas son por lo común clasificadas desde el punto de vista térmico como frías (habitadas por la trucha) o calientes (intole- rables para la trucha). Cuando las aguas están térmicamente estratificadas a me- diados del verano en los lagos de las zonas templadas, los peces de aguas frías como las truchas (Salmo), “chars” (Suluelinus) y los peces blandos (Coregonus) permanecen en el agua profunda o por debajo de la termoclina (el hipolimnion) y los peces de aguas cálidas quedan restringidos a la delgada capa superior de agua caliente (el epilimnion). La termoclina es una capa delgada con declinación rápida de temperatura demarcada en las zonas templadas por un cambio de la tempera- tura a razón de 1°C por cada metro de incremento en la profundidad. Los lagos de las zonas templada y ártica se cubren durante el invierno de hielo por las tem- peraturas tan bajas, y es durante esta estación del año cuando el crecimiento de los peces se hace más lento o de plano se detiene. Precisamente antes de que el hielo cubra las aguas de un lago en la zona templada durante el otoño y, de nuevo, justamente cuando termine el deshielo en la primavera, aparece una condición de homotermia, desde la superficie hasta el fondo. Esta condición permite que haya una circulación profunda térmica e inducida por el viento en ese lago (y tam- bién en algunas partes de los océanos), con lo cual se facilita la distribución de los nutrientes y de los materiales floculados del fondo, desde los fondos hasta la superficie. El oxígeno disuelto en la superficie de las aguas es, durante este perio- do, mezclado a todo lo largo de la columna de agua. En los arroyos superenfriados el hielo se puede formar en el fondo y destruye o restrega, eliminándolos, a los organismos que les sirven de alimento a los peces.
La temperatura del agua puede tener una influencia directiva importante en la migración y en los movimientos de los peces. Como resultado de la correlación que existe entre la temperatura y la distribución de los peces, los zoogeógrafos han desarrollado teorías isotérmicas para explicar la distribución de los peces. Desde hace mucho que los limnólogos han procurado clasificar los lagos sobre la base de sus características térmicas. La clasificación más ampliamente aceptada tiene tres importantes categorías: lagos polares, que tienen temperaturas no ma- yores de 4°C (39.2”F) en su superficie; lagos templados con la temperatura de su superficie oscilando sobre y bajo los 4°C; y lagos tropicales con la temperatura de su superficie siempre mayor a los 4°C.
Debe concluirse que los peces no solamente son sensibles a la temperatura (Capítulo ll), ya que los que son capaces de moverse buscan generalmente lo que puede llamarse preferencia de temperatura. Dentro de un margen tolerable de temperturas los peces se mueven continuamente como si buscaran la temperatura óptima para la actividad vital en su momento.
Luz
La luz es otro factor ecológico de importancia en la vida de los peces. Los efectos directos se manifiestan a través de la visión, pero hay también muchos efectos indi- rectos. La coloración del integument0 en un momento dado es una función directa de la calidad y cantidad de la luz (Capítulo 4). La luz también impulsa y dirige las migraciones y los movimientos (Capítulo 6) , desempeña el papel de cronómetro en la reproducción (Capítulo lo), e influye en la velocidad y en las características

408 ECOLOGÍA Y ZOOGEOGRAFÍA
del crecimiento. La luz también determina las clases y cantidades de alimento dis- ponibles para- los peces y es, por supuesto, la fuente directa de energía para el primer eslabón, el fotosintético, en la cadena alimenticia de todos los peces. La región donde la intensidad luminosa es suficiente para que se realice la fotosínte- sis es conocida como la zona trofogénica o eufótica, y se extiende hacia abajo hasta algunos 200 metros, si es en agua marina transparente.
Corrientes
Las corrientes constituyen factores físicos en las vidas de los peces, tanto de las aguas en movimiento como de las estancadas. Los movimientos del agua tienden a equilibrar, en la superficie, las temperaturas del aire y el agua. A través de la cir- culación también tienden a homogeneizar la temperatura y los factores químicos del agua. La mezcla forzada por los movimientos del viento por sobre la superficie, mueve las moléculas del agua y produce turbulencia u ondas. Cuando la turbulencia en la superficie entra en contacto con la capa de agua situada por debajo, la capa inferior de agua forma una corriente en la dirección opuesta. Esas corrientes pro- ducen remolinos turbulentos internos que conducen a un intercambio vertical de las partículas del agua (intermezcla). Este intercambio es conocido como difusión de remolino.
La acción erosiva de las corrientes a 1s largo de los bancos de los arroyos y de las márgenes altera continuamente el hábitat local, y los materiales despren- didos son transportados y depositados por dondequiera, produciendo diversos cambios allí. La turbidez que resulta de la erosión puede afectar a las adaptacio- nes (como las de los ojos) de los peces pero también puede ser letal o puede redu- cir notablemente las oportunidades que tienen los peces para su sobrevivencia, al sedimentarse los materiales en los lugares donde se alimentan o se reproducen. También acelera la velocidad con la que el agua absorbe calor a partir del sol.
Dentro de un sistema Lidrológico las corrientes cambian típicamente, desde muy rápidas en sus orígenes, o sea, las porciones elevadas, hasta lentas al nivel de su desembocadura, con un correspondiente cambio en las especies de peces. Por lo general, cuanto más fuerte es la corriente, más depauperada se en- cuentra la fauna piscícola. El desarrollo de un perfil hidrodinámico y de órganos para nadar con mayor velocidad, es, entre los peces, su obvia respuesta a la mag- nitud de la corriente. Sin embargo, así como el declive de las corrientes constituye un factor ecológico para los peces, también lo son la forma, profundidad, configu- ración y la composición del fondo de las aguas estancadas.
Oxígeno disuelto
El oxígeno disuelto en el agua es necesario para la respiración, o sea, el desprendi- miento de la energía por el alimento. En las aguas limpias, no contaminadas, de los arroyos, y en la superficie de los lagos, estanques y de los océanos, el agua está normalmente saturada de oxígeno para una temperatura dada. Puede, sin embargo, descender por debajo del requerimiento mínimo de unas cuantas partes por mi- llón para los peces en el nacimiento de las corrientes, y estacionalmente en las aguas estancadas del fondo de los cuerpos de agua quietos. La deficiencia de oxí-

FACTORES ECOL6GICOS 409
geno también puede observarse en los depósitos con mucha materia orgánica o en las aguas contaminadas con productos orgánicos de desperdicio, como sucede en los albañales domésticos. Puede presentarse una considerable mortandad de peces (muerte de verano) debido a la descomposición que emplea mucho oxígeno, cuando grandes cantidades de algas mueren durante los días nublados en que se oculta la luz solar. Cuando la nieve cubre las capas de hielo y la luz solar no penetra a las aguas, el fitoplancton no produce oxígeno, el poco que existe es empleado en la respiración y la descomposición orgánica y, en consecuencia, aparece una mortan- dad de peces, invernal, cuando la demanda de oxígeno excede a la producción.
Alimento
El alimento es uno de los muchos factores biológicos importantes del medio am- biente de los peces. Su abundancia y variedad son determinantes tanto en la com- posición de especies como en las magnitudes de las poblaciones de peces. Los alimentos clave son el plancton de flotación libre, el bentos compuesto de orga- nismos que se mueven sobre o cavan en el fondo, y el perifiton y el perizoon, los “Aufwuchs”, formados por los organismos que crecen en las superficies libres de los objetos sumergidos.
Los factores químicos clave o nutrientes en el primer eslabón de las cadenas alimenticias de los peces incluyen el dióxido de carbono para la fosotíntesis y elementos comunes como el hidrógeno, el nitrógeno, el fósforo, el potasio y el calcio, así como trazas de elementos (oligoelementos) como manganeso, boro, azufre, magnesio, zinc y molibdeno.
Muchos son los orígenes de los elementos y de los gases en el agua; los pro- cesos principales por medio de los cuales se hacen presentes en el agua son tres: la difusión lenta, la mezcla rápida y la transformación interna. El oxígeno, el dióxido de carbono y el nitrógeno, pero especialmente el primero, pueden difundirse con cierta lentitud relativa en el agua a partir de la atmósfera. Estos mismos gases, más el amoníaco, pueden mezclarse rápidamente en la hidrósfera durante las lluvias, las nevadas y las ventiscas, o cuando aparecen corrientes térmicas y de pendiente. La transformación interna de los nutrientes por los organismos vivos, proporciona al agua el dióxido de carbono por la respiración, oxígeno por la foto- síntesis y sulfur0 de hidrógeno y sales orgánicas por los desperdicios de los cuerpos o por los restos de los organismos muertos.
Factores sociales
Entre todos los factores del medio ambiente los más sutiles en su acción son las interacciones que existen entre los individuos de la misma o de diferentes especies. De estas interacciones son buenos ejemplos la competencia y la predación. Y de estos dos, la predación es más fácilmente observable desde el momento en que considera a un organismo presa devorado por un predador. En los ecosistemas acuáticos que afectan a los peces, cualquiera, ninguno, o los dos organismos en una relación de predación, puede ser un pez. Por lo general, la predación en los peces no elimina la presa. La reposición es posible debido al crecimiento de los demás individuos (un proceso llamado aprovisionamiento). Más bien sirve para

410 ECOLOGfA Y ZOOGEOGRAFÍA
incrementar el tamaño medio del resto de los individuos a través de la reducción de la competencia intraespecífica. La predación también favorece la superviven- cia de los mejor dotados, tanto entre los predadores como entre la presa. Ordina- riamente los primeros en desaparecer son los peces enanos, los más débiles, los albinos y los enfermos.
En un concepto modelo de competencia entre los peces, como sucede en otros animales, los dos competidores pueden estar en una lucha permanente. En la naturaleza, sin embargo, uno de los competidores puede triunfar y eliminar finalmente a su contrincante; uno puede causar en el otro su alejamiento o desapa- rición, o las fuerzas evolutivas pueden conducir a un cambio adaptativo. La com- petencia puede producirse entre individuos de la misma especie (intraespecífica) o entre los de dos o más especies (interespecíficas). Cualquier tipo de competencia puede ser un factor dependiente de la densidad en la determinación del tamaño de la población y de su estructura. Las reacciones competitivas más comunes entre los peces se producen cuando andan en busca de un lugar para desovar, de alimento, de espacio y de protección. El amontonamiento de los peces desovado- res, como sucede con los salmones del Pacífico (Oncorhynchus), conduce a me- nudo a la destrucción de los primeros nidos por los últimos desovadores y puede tener consecuencias psicológicas adversas en los propios desovadores. Y más aún, la Competencia que aparece por los sitios de nidación constituye un factor funda- mental en el surgimiento común de híbridos entre los peces sol o mojarritas (Le- pornis). Los híbridos resultantes son predominantemente machos y aparentemente estériles o de baja fertilidad, más baja que la de los padres, dependiendo esto del cruzamiento. Además, desde el momento en que ellos tienen también una tasa de crecimiento incrementada, puede razonarse que tienen una esperanza de vida mucho más corta que la de las especies progenitoras.
La introducción de peces en aguas donde no son nativos puede producir el desplazamiento competitivo de las especies propias del lugar. Por ejemplo, la introducción en Nueva Zelanda de peces salmónidos, predadores y de importancia deportiva, que tienen una capacidad competitiva superior a la de las especies endé- micas, está terminando con éstas.
Además de la predación y la competencia los peces exhiben una variedad de relaciones simbióticas, o sea los casos en que por lo menos una de las especies es afectada por la presencia de la otra. La simbiosis para la limpieza es una de las más notables y de más amplia distribución. Los peces y camarones limpiadores se alimentan de ectoparásitos, tejido necrosado, escamas y posiblemente moco en- contrado en las superficies del cuerpo, cámaras bucal y branquiales, de sus peces huéspedes. En las zonas tropicales y templadas se han encontrado docenas de especies de peces marinos y de agua dulce con hábitos limpiadores. Los mejor conocidos provienen de los peces señorita o viejita (Labridae), peces ángel y ma- riposa (Chaetodontidae) y los gobios (Gobiidae). Muchos de estos peces limpiado- res están brillantemente coloreados y parecen anunciar su presencia para atraer a posibles huéspedes a sus estaciones de limpieza. Sus huéspedes incluyen a casi todos los peces de los arrecifes de tamaño no mayor a unos cuantos centímetros de longitud, a predadores tan grandes como los meros (Serranidae) y las barracu- das (Sphyraenidae), e inclusive a algunos de los peces pelágicos, como ciertos atunes (Scombridae). También los tiburones reciben servicios de limpieza de las rémoras (Remora). En algunas áreas la simbiosis de limpieza aparece como un verdadero mutualismo, en el sentido de que ambas partes se benefician: el pez

FACTORES ECOLdGICOS 411
limpiador recibe su alimento, “remitido” a su estación de limpieza por los hués- pedes que solicitan los servicios de limpieza, mientras éstos son liberados de los ectoparásitos potencialmente dañinos. Pero en otras áreas, los ectoparásitos pare- cen ser relativamente raros y los peces limpiadores se alimentan activamente de las escamas y del moco de sus huéspedes, convirtiéndose en comensales e inclusive animales parásitos por su propio derecho. ¿Por qué los huéspedes continúan soli- citando los servicios de limpieza de estos peces limpiadores comensales o parásitos?, sería una pregunta a una aparente paradoja, que, sin embargo, ha sido resuelta mediante el descubrimiento de que los peces limpiadores premian continuamente a sus huéspedes. Este premio o reforzamiento positivo se realiza bajo la forma de “cosquilleo” o estimulación táctil suave producida por las aletas y el hocico de los peces limpiadores especializados, y de las antenas y apéndices locomotores de los camarones limpiadores. De esta manera, las relaciones de limpieza continúan bajo casi todas las condiciones y están disponibles para dejar satisfecho el papel del mutualismo en la remoción de los ectoparásitos, cuando los peces están some- tidos a incrementos de infección.
El comensalismo está presente entre muchos celenterados y peces. Parece ser que los peces son, o bien inmunes a las células pinchadoras (nematocistos) de los celenterados, o inmunes inclusive a sus descargas. La carabela portuguesa o agua mala (Physulia), por ejemplo, acoge sin hacerle daño al llamado pez del agua mala (Nomeus gronoui). Algunas medusas de aspecto gelatinoso, como Crumbessu, dan protección a peces pequeños, incluyendo a cardúmenes de jureles (Curunx). Al- gunos peces señorita (peces anémona, Amphiprion; Premnus) están a menudo asociados con grandes anémonas de mar (Stoichuctis). Estos peces pueden inclu- sive acarrear o traer alimento para la anémona y también ahuyentar a otros peces. Algunos peces cardenal (Apogonidae) permanecen en la cavidad paleal de una concha grande (Strombus), y algunos peces perla (Curupus) consiguen refugio en la cámara cloacal de los pepinos de mar (Holothuroidea) u, ocasionalmente, en el estómago de las estrellas de mar grandes (Asteroidea). Una especie de pez perla hace su morada en el manto de una madreperla y de este hecho se origina el nom- bre común de este género de pez. El pez piloto (Naucrutes ductor) forma cardú- menes en frente y por debajo de los grandes tiburones y mantarrayas (Mobulidae) y, teóricamente, correspondiendo a las dádivas de sobras de alimento que recibe (así como por la oportunidad que tienen de limpiar al pez huésped de ectopará- sitos), puede defenderlos de sus enemigos, así como alertarlos cuando hay posibles presas en las cercanías. Cuando las rémoras (Echeneidae) viajan de polizón en las tortugas de mar, ballenas, tiburones y otros peces, también devoran los restos ali- menticios que dejan sus huéspedes. En los peces espada (Xiphius), peces vela (Istiophorus) y en los molas (Molu), hay reciprocidad porque las rémoras limpian a su vez a estos grandes peces de parásitos y tejido enfermo, no solamente en la porción externa, sino también por las cavidades branquia1 y bucal. Un pez ciego (Typhlogobius), cavando en el lecho de un cangrejo (Calliamsu), se ha vuelto muy dependiente de este animal, tanto por el alimento como por el resguardo que re- cibe, y es un ejemplo de adaptación extrema al comensalismo.
En las relaciones de comensalismo un pez se beneficia, pero su huésped no es esencialmente afectado. La cooperación primordial en los peces corresponde aparentemente a la formación de cardúmenes de tiburones predadores para el acorralamiento de presas para matarlas, así como en la conducta de reunirse en cardúmenes de pececillos para lograr la protección contra los predadores. La pro-

41 2 ECOLOGfA Y ZOOGEOGRAFÍA
tocooperación es definida como la situación por medio de la cual ambas especies, o los numerosos individuos de una especie, se benefician de la cooperacibn, aun- que, por otra parte, ésta no sea indispensable.
Llegamos finalmente al parasitismo, condición en la que una especie o individuo (generalmente el más pequeño) vive sobre el otro y depende de é1 para su subsistencia. Muchos organismos, incluyendo a los hongos, las bacterias, gusa- nos, crustáceos, moluscos larvarios, y los mismos peces, son parásitos de los peces. Parece ser que sólo la sobreinfestación por parásitos es tremendamente dañina para el huésped, así de perfeccionado está el sistema parasitario en los animales. El pequeño “candiru”, un bagre sudamericano de la familia Trichomycteridae, que vive en forma natural en las branquias de los peces grandes y les succiona sangre, es un pez que puede ser posiblemente considerado como un verdadero parásito. Este pez es peligroso para el hombre y para otros animales debido a su habilidad para penetrar y alojarse en la uretra (como la de los humanos del sexo masculino) o en la vejiga de la orina (como ocurre en los humanos del sexo femenino). Tam- bién penetra en otros orificios del cuerpo.
Cualquiera de los anteriores factores sociales en la vida de los peces puede variar e interactuar de acuerdo a determinada etapa de su desarrollo, estación del año y ubicación. Como ejemplo sencillo, consideremos a la cría de la trucha café (Salmo trutta) que constituye una presa temprana de los “minnows7’ (Cypri- nidae) con los que convive. Cuando la cría de la trucha pasa a la etapa juvenil, también se vuelve competitiva de sus predadores originales. Más tarde, comple- tando la inversión de las circunstancias, las truchas adultas vienen a terminar como predadoras de los “minnows”.
Densidad de población
Las dinámicas de los medios ambientes específicos se reflejan en el cambio conti- nuo dentro de las poblaciones de peces que los habitan. La densidad, tomada como función del tiempo, volumen o espacio, constituye un parámetro básico de la población. Por lo tanto, la densidad puede corresponder al número de peces en un lago, o números o peso por unidad de área o por su volumen. Los cambios en la densidad de población en los peces son a menudo descritos en términos de su relativa abundancia debido a la dificultad de medir con exactitud el tamaño de la población en cualquier cuerpo de agua que no sea pequeño. Se ha visto que muchos aspectos del comportamiento de los peces y de su fisiología son depen- dientes de la densidad.
La densidad de población en un momento dado es la función que hay de la natalidad contra la mortalidad. En una especie exitosa el potencial de la natali- dad excede normalmente al potencial de la mortalidad y le da a la especie la opor- tunidad de multiplicarse si las circunstancias son favorables. Si la natalidad actual es superior a la mortalidad real, la población aumenta; sin embargo, si las dos son iguales, la población se mantiene estacionaria. Donde la mortalidad real excede a la natalidad real, la población merma. La historia de la dinámica de la población de una especie puede ser resumida como sigue:
a. Colonización y expansión. b. Adquisición de la estabilidad con algunas fluctuaciones en la densidad,
pero con una natalidad real que se iguala a la mortalidad real.

FACTORES ECOL6GICOS 413
c. Regresión, en la cual la mortalidad excede a la natalidad, con las más gran- des pérdidas, como es lo típico, entre los peces cuando son muy jóvenes.
d . Se da cabida a otras especies. e. Desaparición; sustitución por las poblaciones de otras especies.
Estructura de la población
La estructura de la población de una especie de pez es un indicador de su estado evolutivo, ya sea que se esté expandiendo, se mantenga estática o esté retroce- diendo. Puede conocerse la estructura mediante la determinación de la composi- ción por edad de la población. En las aguas de las zonas templadas la estimación de la edad de los individuos de peces es facilitada por las marcas anulares anuales que se han identificado en las escamas, los huesos, las piedrecillas del oído interno (oto- litos) y en las secciones transversales de los radios de las aletas. Se ha llegado a aprender a través de las aplicaciones de esos procedimientos para la determina- ción de la edad, que los jóvenes predominan en las poblaciones en expansión, mientras que los mayores dominan en las que están en retroceso. Las fluctuaciones en la abundancia de individuos, que se manifiestan en una población estable, se deben a la dominancia de la clase anual. En un modelo de dominancia por la clase anual, el mismo número de huevos puede ser producido año tras año, pero sola- mente en ciertos años habrá realmente una sobrevivencia sustancial. Como resul- tado, las pesquerías pueden a menudo depender para su éxito de la clase dominante en el año; en su declinación, del ocaso de esa clase anual; y levantarse de nuevo cuando una nueva clase anual dominante aparece con mucho éxito.
Las especies ictiológicas presentan diferentes características en el agrupa- miento. Algunas, como los predadores errantes, pueden ser relativamente solitarios y agruparse sólo por casualidad. En tales especies no gregarias el crecimiento y la supervivencia son mayores, mientras que la densidad de población es menor. En los peces gregarios como las sardinas (Clupeu) y las lisas (Mugil), la distribución en una población es típicamente azarosa por cardúmenes, pequeños o grandes. La densidad de los grupos, así como los cambios en el medio ambiente, puede favorecer la supervivencia y ser necesaria para la reproducción y para proporcionar las variaciones que pueden favorecer la evolución.
Algunas poblaciones de peces son afectadas de manera notable por la inmi- gración o emigración de los individuos. Las pérdidas o incrementos moderados en el número de individuos que migran hacia o desde la población deberán ser compensados por los correspondientes cambios en la tasa de mortalidad. Por ejem- plo, la inmigración masiva puede reducir el crecimiento de todos los individuos debido al incremento de la competencia. Sin embargo, este efecto negativo puede ser equilibrado por el incremento de la mortalidad. La migración masiva puede in- crementar el crecimiento y la supervivencia, pero también puede causar que la población de una especie pierda su nicho. Aunque los números pueden cambiar en cualquier clase de movimiento, la capacidad fija del hábitat tiende a mantener la misma biomasa.
Las comunidades de peces se caracterizan por la presencia de especies do- minantes. Las comunidades de las partes altas y las partes bajas de un río tienen diferentes dominantes. Es característico que la zona de transición o ecotono en- tre las dos debería tener representantes de las dos zonas, tanto de la alta como de

414 ECOLOG~A Y ZOOGEOGRAFÍA
la baja, y debería esperarse que contuviera más especies de las que aparecen en las dos zonas que separa. Corrientes similares de cualquier lado pueden tener las mis- mas o diferentes combinaciones de especies. Sin embargo, ya sea que las especies sean las mismas o no, los nichos ecológicos de los ocupantes deberán ser esencial- mente los mismos; en consecuencia, estos ocupantes deberán exhibir similitudes muy cercanas en sus relaciones con la cadena alimenticia.
Sucesión
En cualquier parte dada de la Tierra, las aguas y sus habitantes tienden a desarro- llarse en direcciones predecibles. Al hacerlo aparece una sucesión de condiciones dependientes del medio, de la flora y fauna. Por ejemplo, el proceso evolutivo de los lagos glaciales está con mayor frecuencia ordenado (Figura 14.1) y es deno- minado eutroficación, en la cual la situación original oligotrófica y pobre en nu- trientes se vuelve más y más rica en sustancias nutritivas, llenando y eliminando finalmente al lago por entero. Mientras que el medio ambiente está cambiando, los primeros peces de agua fría son gradualmente reemplazados por peces tolerantes de aguas cálidas, pero finalmente desaparecen todos los peces. En la evolución de los arroyos la corriente de agua corta siempre hacia abajo hasta el nivel de la desembocadura (Figura 14.2). A medida que la inclinación disminuye los prime- ros peces de las partes altas desaparecen y son sucesivamente reemplazados por otros mejor adaptados a los cambios del medio ambiente.
La evolución adaptativa de los peces, siguiendo a la evolución de sus hábi- tats, ha dejado unos cuantos nichos desocupados en el ecosistema acuático. En
""
Mucho oxígeno Poco dióxido de carbono Mucho dibxido de carbono Suelo inorgánico Suelo orgánico Vegetación dispersa Abundante vegetación Alimento disperso (principalmente Abundante alimento
pelágico) Poca transparencia Mucha transparencia Elevada fertilidad Baja fertilidad Acidez en aumento Usualmente alcalina Peces "de agua caliente" Peces "de agua fría"
> Poco oxígeno
(Hipotbtico)
Figura 14.1 - Evolución hipotetica de un lago de origen glacial en la por- ci6n central de Norteamhrica. (Basado en Eschmeyer, 1936).

ECOSISTEMA MARINO 415
Oeste de Europa "Minnows" de
Lucio, perca v ciprinidos de
Trucha Cafe "Grayling" agua corriente y Lucio aguas estancadas
En forma de "V" Valle En forma de "U" ancha
Mucho
Claro
Gradiente (corriente)
Turbidez
Pequeño
Nebuloso
De rocoso a cubierto de Fondo De arenoso a lodoso
Poco grava
Materia orgánica (fertilidad) Mucho
Fr ío Temperatura Caliente
I ntraespecíf ico Competencia lnterespecífico
De ninguno a pocos Peces predadores Muchos
Direcci6n del cambio con el tiempo
America del Norte "Minnows" y Lucio Bagres, carpas hociconas, y "Grayling" "Stream char" de agua corriente Loynas "Minnows" de aguas estancadas
L
Figura 14.2 - Sucesibn hipotktica de las condiciones ecol6gicas y los peces en las corrientes de truchas de Norteambrica y Europa Occidental.
las antiguas aguas de historia evolutiva muy larga esto es extraordinariamente cierto, pero inclusive en las aguas relativamente recientes, como las de los lagos de grietas del África, la evolución virtualmente explosiva llena rápidamente los ni- chos disponibles
ECOSISTEMA MARINO
Para el observador casual, y especialmente para el estudiante cuya mayor expe- riencia ha tenido lugar en las aguas continentales, el mar es un medio uniforme, quieto y estable. Sin embargo, el mar es en la actualidad un ecosistema en conti- nuo movimiento y cambio, y con una gran variedad de biotopos. Un biotopo puede ser considerado como una área en donde las condiciones del hábitat y los organismos presentes son muy uniformes. Los biotopos oceánicos difieren de acuerdo a la geología regional, el sustrato, la profundidad, las corrientes y los nu- trientes. También difieren, de lugar en lugar, en la salinidad, los gases disueltos, la luz, la temperatura, el sonido y, desde luego, en la habitación por los peces y otros organismos vivos. Debido a las diferencias que existen entre los biotopos marinos, los oceanógrafos han sentido desde hace largo tiempo la necesidad de contar con un amplio sistema fundamental de clasificación que podría ser desa- rrollado para facilitar la organización y la comprensión de la información detallada, en continuo aumento. Esta necesidad ha conducido a la propuesta de muchos siste- mas de división del ecosistema marino; el de Hedgpeth (1957) sirve bien a nuestras necesidades. Una clasificación así está interesada tanto por el agua (la división pelágica de los mares) como por el fondo (la división bentónica; Figura 14.3).

416 ECOLOGÍA Y ZOQGEQGRAF~A
Supra-
Profundidad aproximada
Litoral lentre los niveles de las mareas)
Figura 14.3 - Clasificación de los medios ambientes marinos. (Fuente: Hedgepeth, 1957).
Todos los componentes de los medios pelágico y béntico anteriormente citados, pueden contener uno o más biotopos, como ya ha sido sugerido. La uni- formidad de condiciones para la vida de la fauna ictiológica que hay en ellos, decrece con la cercanía a la tierra y a la superficie del agua. En consecuencia, los biotopos son más numerosos en las aguas de las costas.
Medio ambiente pelágico
El medio ambiente pelágico está dividido en dos partes importantes: 1) la zona nerítica, o masa de agua situada sobre la plataforma continental, extendiéndose lejos de la costa hasta una profundidad de aproximadamente 200 metros; 2) la zona oceánica, que comprende a todas las aguas que se encuentran más allá de la plataforma continental; por lo tanto, estas aguas están en zonas con una profun- didad mayor a los 200 metros. En este medio ambiente escasea por lo general el abrigo y los peces pelágicos, y aun otros, desde pequeños hasta grandes, se reúnen con frecuencia por debajo de despojos flotantes, rodeando a todo lo que vaya a la deriva, como algas marinas, troncos de árboles, piedra pómez, medusas gran- des y tortugas y ballenas muertas. Dentro de la zona oceánica se reconocen las siguientes divisiones verticales, aunque obviamente no hay separaciones definidas de demarcación entre ellas, y su extensión vertical difiere en los diferentes oceános:
U.
b.
C.
d.
Epipelágica: aguas superficiales alejadas de la plataforma continental pero de unos 200 metros de profundidad. Mesopelágica: aproximadamente entre los 200 y 1,000 metros de pro- fundidad. Batipelágica: aproximadamente entre los 1,000 y 4,000 metros de pro- fundidad. Abisopelágica: por debajo de los 4,000 metros.

ECOSISTEMA MARINO 41 7
Zona nerítica. La zona nerítica está bien iluminada pero tiene variaciones estacio- nales en la iluminación así como en la temperatura, la salinidad, la cantidad de oxígeno disuelto, los nutrientes, la acción de las olas, las corrientes y su biota. Algunas veces puede cambiar drásticamente la abundancia de ciertos componentes del biota nerítico y producir un cataclismo en la población ictiológica. Por ejem- plo, los crecimientos masivos del alga de dinoflagelados, Gymnodinium breve (conocida como “la marea roja”) produce mortandades masivas en la costa oeste de Florida. La muerte de los peces es debida a la liberación, por el alga, de un metabolito de elevada toxicidad. Esta zona también contiene una cantidad muy considerable de detritus, flotando a la deriva. Es la más rica en variedad de bio- topos y, en consecuencia, de todas las aguas es la que contiene mayor abundancia y variedad de fauna ictiológica. Los hábitats representativos incluyen las aguas costeras, bancos de algas estacionarios y a la deriva, y estuarios, así como los que están sobre los arrecifes de coral, bancos o plataformas submarinos elevados, y porciones angostas o amplias de la plataforma continental. Los peces más carac- terísticos son las sardinas (Clupeidae). Entre todos los demás peces de la zona nerítica se cuentan las barracudas (Sphyraenidae), muchos tiburones y algunas macarelas (Scombridae), el pez azul (Pomatomus), los peces aguja (Belonidae), el sábalo (Megulops), las anguilas (Anguillu), los atunes (Thunnus), el merlín (Muhuiru), el pez vela (Istiophorus), los peces palometa (Stromateidae), la cabrilla de sargazo (Purulubrux), los meros (Epinephelus), los pargos (Lutjanidae), los gru- ñidores (Pomadasyidae), los sargos o peces pluma (Sparidae) y la trucha marina (Cynoscion). En base a los análisis de la enorme cantidad de datos obtenidos me- diante el rastreo, los peces de aguas profundas de los bancos de arena continenta- les están organizados en comunidades con preferencias hacia las profundidades.
Zona oceánica. La zona oceánica es menos productiva que la nerítica, pero preva- lecen condiciones de vida de amplia distribución. Hay en esta zona poca materia detrítica y pocas variaciones estacionales en la temperatura, la composición quí- mica del agua y otros factores del medio ambiente, es decir, no tan grandes como las de la zona nerítica.
División epipelágica. La división epipelágica de la zona oceánica es real- mente una extensión a distancia de la costa de la zona nerítica, relativamente delgada, pero con su fondo formado por agua y no por un sustrato sólido. Esta división está bien iluminada en la superficie pero se va oscureciendo hacia su lí- mite inferior situado a unos 200 metros de profundidad. Todas las ondas lumino- sas del espectro penetran en el agua por la superficie, pero los azules y violetas son retenidos en los márgenes más bajos de la profundidad mayor. Las variaciones estacionales son manifestadas por la temperatura, la luz, la salinidad, el oxígeno, los nutrientes y las poblaciones de plantas y animales. De todas éstas, parecen ser la luz y la temperatura las más importantes en la determinación de la distribución animal. Por ejemplo, ningún pez boca (Stornius) ha sido capturado sobre los 300 metros en los muestreos hechos con red de arrastre durante el día, pero la mayor parte de las especies migran hasta por encima de los 100 o 200 metros durante la noche. La fauna ictiológica incluye a la que aprovecha los recursos del océano a partir de la zona nerítica, así como a algunas macarelas, bonitos, albaco- ras y atunes (Scombridae), algunos tiburones, dorados (Coryphaem), peces vola- dores (Exocoetidae), mantas (Mobulidae), anguilas (Anguillu), merlines (Mukuira),

418 ECOLOGfA Y ZOOGEOGRAFfA
peces vela (Istiophorus), pez azul (Pornatomus), molas (Mola ) , peces linterna (Myctophidae), el pez boca pariente (Cyclosthone) y los peces boca (Stomias).
División mesopelágica. Los ocupantes de la división mesopelágica de la zona oceánica dependen para su alimentación de una “lluvia” de plancton, cadáveres y excrementos procedentes de la división epipelágica situada inmediatamente arriba, así como de las relaciones de predación que hay entre uno y otros. Hay una pe- queña variación estaciona1 de la temperatura; el agua está casi siempre fría, con una temperatura que oscila en los 10°C. La presión del agua es elevada y la luz extremadamente débil y en las frecuencias del azul y el violeta. Esta división con- tiene las aguas más afóticas de los océanos y está habitada por animales adaptados a la oscuridad o escotofílicos. Muchos de los peces son negros o rojos y migran hacia la división epipelágica para alimentarse, algunas veces durante la noche en o cerca de la superficie. Durante los estadios larvarios estos invasores han habitado, generalmente, en las aguas epipelágicas. Como ejemplos de estos peces se pueden mencionar los peces linterna (Myctophidae), las anguilas de los fondos marinos (Synaphobranchus), los peces boca parientes (estomiatoideos: Stomias, Cyclo- thone, Argyropelecus y Chauliodus), el pez de ojos pedunculados (Idiacanthus) y el tragador de los fondos marinos (Chiasmodus).
División batipelágica. En la división batipelágica de la zona oceánica los principales alimentos también caen desde las aguas superiores. No hay esencial- mente variaciones estacionales en los factores físicos del medio ambiente; el agua es muy fría (entre los 2” y 4°C a los 2,000 metros de profundidad), las presiones son muy grandes y la oscuridad prevalece, excepción hecha de la bioluminiscencia producida por los órganos luminosos de muchos de sus pobladores. El número de peces es muy reducido, lo mismo que el de clases en relación al de las aguas supe- riores, pero incluye a la mayoría de los peces pescadores ceratioideos de los fondos marinos, “dories” (Zeidae), algunos peces escorpión (Scorpaenidae), el tragador de los fondos marinos (Chiasmodus), el pez boca (estomiatoideo) pariente (Chau- liodus y Malacosteus), los engullidores (Eurypharym), los tragadores (Saccopha- rynx) y las anguilas de los fondos marinos como Cyema.
División abisopelágica. Las grandes profundidades de la zona oceánica, las abisales, son extensas y ocupan más de la mitad de la superficie de la Tierra. En esta división llamada abisopelágica la presión es muy elevada (comprendiendo desde las 200 a más de 1,000 atmósferas), el frío (menos de 4°C) y la oscuridad -a excepción de la bioluminiscencia, que es rara- prevalecen siempre y no hay cambios estacionales de las condiciones física.s. Los animales de la sima son casi ciegos o no tienen ojos; la luz tiene poca importancia ecológica. Las necesidades de alimento primarias son satisfechas mediante la lluvia de materiales proceden- tes de las aguas inmediatamente superiores o bien de las cadenas alimenticias conectadas hacia arriba. Las condiciones y los peces del límite inferior son poco co- nocidos, y nada se sabe esencialmente de lo que ocurre en las profundidades mayores de los 6,000 metros. Los pocos peces que ahí se encuentran están repre- sentados por especies como el pez boca pariente, ChauEiodus, los engullidores (Eurypharynx), una anguila de aguas profundas (Cyema) y los peces pescado- res de los fondos marinos, Melanocetus y Borophryne. Desde una profundidad de 3,590 metros viene Gallatheathauma, un pez pescador del Africa Occiden-

ECOSISTEMA MARINO 419
tal tropical, con un curioso órgano luminoso en su boca, un posible cebo para cierta presa.
Medio ambiente bentónico
En las subsecciones previas hemos examinado peces de las aguas abiertas de las zonas nerítica y oceánica. En el fondo, por debajo de estas zonas, hay peces cavadores de los fondos, característicos, que forman parte del bentos. El medio ambiente bentónico es dividido como se indica a continuación:
a. Supralitoral: desde la marca de la marea alta hasta el borde de la playa o zona de riego.
b. Litoral: entre la marca de la marea alta y el nivel al que llega la marea baja, esto es, intermedio a los niveles de las mareas alta y baja.
c. Sublitoral: entre la marca de la marea baja y el borde de la plataforma continental (hasta cerca de los 200 metros de profundidad); la porción sublitoral interior es subdividida a partir de afuera (mar afuera, a gran distancia de la costa) hasta cerca de los 50 metros.
d. Batial: el fondo está situado aproximadamente desde una profundidad del agua de 200 metros hasta la de 4,000 metros.
e. Abisal: comprendida entre los 4,000 y 6,000 metros. f. Hadal: la más profunda, a más de 6,000 metros aproximadamente.
Zona supralitoral. La zona supralitoral o bañada del medio ambiente bentónico está compuesta típicamente por superficies húmedas y pozas pequeñas en el sustrato. Hay pocos peces aquí debido a las Condiciones de vida, especialmente la dotación de agua y la temperatura, que pueden ser extremadamente variables de uno a otro momento, quedando a merced de las estaciones. En la zona más difícil para ser habitada por plantas y animales y requiere de considerable adaptación. Los pocos peces que han sido capaces de establecer nichos en este medio ambiente tan lábil, incluyen algunos gobios (Gobiidae), anguilas (Anguilliformes) y peces adheribles (Gobiesocidae).
Zona litoral. En las áreas comprendidas entre los niveles de las mareas alta y baja de la zona litoral del medio ambiente béntico, hay también grandes varia- ciones en las condiciones apropiadas para la vida, detectadas por hora, día, mes y estación anual, especialmente en lo que respecta a la humedad. A pesar de esto la productividad es elevada, dado que los peces llegan y se alejan con las mareas. A la mayoría de los peces se les puede encontrar también, por lo tanto, en la división béntica sublitoral. Peces representativos son las rayas de espina (Dasyatidae), mu- chos lenguados de las familias Bothidae y Pleuronectidae, soles (Soleidae), el pez hueso (Albula), anguilas (Anguilla), morenas (Muraenidae), peces adheribles (Go- biesocidae), cótidos (Cottidae), soldaditos (Triglidae), “snailfishes” y “lump- fishes” (Cyclopteridae), peces sapo o cabezón (Porichthys), peces de roca (Blennii- dae), gobios (Gobiidae), peces pipa y caballitos de mar (Syngnathidae) y peces lengua (Ophidiidae).
Zona sublitoral interior. En esta zona sublitoral la productividad permanece ele- vada, las variaciones estacionales están cerca del máximo y la iluminación es buena

420 ECOLOGfA Y ZOOGEOGRAFfA
con los amarillos y rojos cerca de la orilla y los azules mar adentro. Como ejem- plos de peces que habitan esta zona pueden mencionarse a las carpas de las rom- pientes (Embiotocidae), la mayoría de las rayas (Rajidae) y las rayas de espina (Dasyatidae), la mayoría de los lenguados y soles (Pleuronectiformes), soldadi- tos (Triglidae), tiburones perro (Squalidae), peces hueso (Albulidae), anguilas (Anguillidae), morenas (Muraenidae), caballitos de mar y peces pipa (Syngnathi- dae), roncachos, roncadores blancos y tambores (Sciaenidae), merluzas de la fami- lia Gadidae, peces roca y especies relacionadas (Scorpaenidae), peces señorita (Labridae), peces mariposa y peces ángel (Chaetodontidae), peces sapo o cabe- zón (Porichthys) y Gibbonsia.
Los arrecifes de coral unen las zonas sublitoral y litoral en los mares cálidos. Sus peces son en su mayoría muy vistosos por sus colores brillantes. Cerca de estos arrecifes viven la mayoría de algo así como doscientas especies de los peces mariposa (Chaetodontidae). También viven ahí los ídolos moros (Zanclidae), los peces cirujano (Acanthurus), los peces puerco (Balistidae) y peces loro como Scams. También hay muchas especies de peces señorita (Labridae), junto con chopas (Pomacentridae), peces de roca (Blenniidae) y miembros de la familia Serranidae.
Zona sublitoral exterior. La zona sublitoral exterior permanece muy productiva con algunas variaciones estacionales de las condiciones físicas de vida. La ilumi- nación del fondo queda aquí comprendida entre el azul y el violeta. Mucha de la fauna se parece a la batial; hay también peces advenedizos procedentes de la zona sublitoral interior. Los peces incluyen al bacalao de Escocia (Melanogrammus), el bacalao (Gadus) y merluzas (Merluccius y Urophycis) de la familia Gadidae, “halibuts” (Hippoglossus), quimeras (Chimaera), lampreas glutinosas o mixinos (Myxinidae), anguilas (Anguilla) y “pollock” (Pollachius). Ocasionalmente los peces habitantes de los fondos de esta y otras zonas de la costa pueden ser desplazados e inclusive lanzados a tierra por olas enormes.
Zonas batial y abisal. En la zona batial del medio béntico, como en todas las subsecuentes, no hay variación esencial en los factores físicos del medio ambiente. El fondo es frío y oscuro, a excepción de la luminiscencia, rara entre los ocupam- tes, y de la débil luz que llega hasta los 1,000 metros de profundidad. Los alimentos primarios caen desde arriba y son más densos en el fondo que en el agua que está in- mediatamente por arriba. Los peces batiales incluyen a los halibuts (Hippoglossus), quimeras (Chimaera), bacalaos (Gadus) y mixinos o lampreas glutinosas (Myxi- nidae). Los peces abisales están representados por la anguila (Synaphobranchus), los peces cola de rata o granaderos (Melanonidae), las brotulas (Brotulidae), los peces linterna parientes, Ipnops y Bathypterois, y algunos peces oceánicos mesopelági- cos y abisopelágicos que probablemente también llegan al fondo para comer.
Zona hadal. Unos cuantos peces están presentes en la zona hadal. Es cuestionable la inclusión de los peces cola de rata (Macrouridae), las anguilas de las aguas pro- fundas, Synuphobranchus, el brotúlido Bassogigas y los peces caracol, entre los que se incluye Pseudoliparis. Solamente existen bosquejos de información sobre las comunidades hadales de las siete grandes trincheras de los mares.

ECOSISTEMA ESTUARINO 421
ECOSISTEMA ESTUARINO
Un estuario o estero es típicamente un cuerpo de agua donde el agua dulce (o sea en la que la sal no puede ser detectada por el sentido del gusto) procedente de las co- rrientes se mezcla con el agua salada de un océano (la cual es una solución salina sofocante con su promedio de 35 partes de sales por cada mil de agua, o sea, ). Desde el momento en que es realmente una zona de transición entre dos medios, se le denomina, con toda propiedad, ecotono. La mayoría de los esteros están formados por desembocaduras sumergidas de ríos, y varían en tamaño, desde el pequeño que recibe sólo una corriente de agua hasta el muy grande, como Chesa- peake Bay de la costa atlántica norteamericana, que recibe grandes corrientes y muchos tributarios también pequeños. Conocido de todos es el carácter impre- sionante de los esteros en las costas rocosas, llamados “fjords”, como en Alaska y Noruega. Los esteros se caracterizan por la extremosa fluctuación en la salini- dad, la turbulencia producida por las mareas y las corrientes de los ríos, la turbidez y el azolvamiento; las fluctuaciones estacionales que allí se presentan son las mayo- res registradas para cualquier medio ambiente conocido. Los esteros circundados por ciudades o industrias pueden tener también grados extremos de contaminación. La base primordial para la evolución de los esteros consiste en el rellenado. La formación del delta corresponde a una etapa de este proceso en muchos cuerpos de agua de las costas bajas. El efecto de azolvamiento se observa cuando no hay ma- rea y aparecen llanuras de lodo con olores picantes que nunca se olvidan. La for- mación de tierra deltaica en los trópicos es a menudo acelerada por los crecimien- tos densos de los árboles y arbustos de mangle (especialmente Rhizophoru). Las orillas de los esteros están formadas a menudo de arena, aluvión y lodo. Pueden estar bordeados por pasto (Panicum), que aparece con mayor densidad hacia la marca más elevada de la marea alta. Entre la marca de la marea alta normal y la marca de la marea alta extrema, el nivel inferior puede también estar bien cu- bierto de vegetación con plantas como juncos (Juncus), y pastos de las ciénagas (Spurtinu). Entre las marcas de los niveles de las mareas alta y baja normales puede la vegetación desaparecer rápidamente desde las masas de pasto de las cié- nagas para descubrir el fondo lodoso y muy rico en materia orgánica. Un carácter dinámico clave de estos cuerpos de agua consiste en el grado de flujo o renovación del agua. A pesar del flujo y otros factores limitantes hay en los esteros una muy elevada productividad biológica.
Los esteros que se encuentran en la desembocadura de los ríos exhiben, como una característica de ellos, la formación de capas, con un lente de agua dulce sobrepuesto al agua salada que se introduce por debajo. Las dos capas pue- den tener diferentes faunas ictiológicas. Los esteros constituyen rutas de migra- ción de los peces diádromos y son también lugares de estancia de las poblaciones de muchas especies costeras, tanto marinas como de agua dulce. También pueden ser lugares apropiados para la reproducción y la alimentación de otras. Los peces residentes o visitantes muestran sorpresivas tolerancias para los cambios de salini- dad y otros rigores del medio ambiente de la vida estuarina. Los residentes Cornu- nes de los esteros incluyen a muchos arenques y especies afines (clupeoideos) y muchos ciprinodontiformes.

422 ECOLOGfA Y ZOOGEOGRAFfA
ECOSISTEMA DE AGUA DULCE
Aunque el medio marino es muchas veces más extenso que el de agua dulce, se conoce más sobre los peces de agua dulce y sobre los detalles de su historia natu- ral, ecología y distribución. Los tipos de vida y las condiciones que se encuentran en el agua dulce han sido más accesibles para su estudio. Las masas de agua dulce ocupadas por los peces aparecen tanto en la superficie de la Tierra como en las aguas subterráneas de las cavernas y de los ríos que forman las corrientes freáti- cas. Los peces subterráneos se han desenvuelto a partir de los de las aguas super- ficiales, y dependen de la vida de las aguas de la superficie, ya sea directa o indi- rectamente, para obtener su alimento. En consecuencia, las consideraciones que aquí trataremos se centrarán en las aguas superficiales.
Las aguas dulces de la superficie de la Tierra son separadas en dos grandes grupos de. medios: estancadas, o lénticas, y corrientes, o lóticas. Los hábitats lénticos incluyen a los de los estanques y lagos naturales, junto con los numerosos embalses construidos por el hombre desde tiempo inmemorial. Los medios lóti- cos son los de las corrientes. A éstos se les deben agregar las condiciones espe- ciales encontradas en a ) los manantiales, que pueden ser tributarios de los lagos o las corrientes, y b ) los esteros y las “colas” de los grandes embalses donde las condiciones lóticas de las corrientes van volviéndose gradualmente lénticas.
Desde el momento en que los embalses artificiales son de naturaleza lacus- tre, incluyendo áreas como los depósitos de agua situados bajo el nivel del mar en los Países Bajos, no se les da consideración especial aquí. Sin embargo, la cons- trucción de muchos de ellos ha alterado significativamente la ecología regional y la distribución de los peces. Cuando una presa es construida sobre un río cambia completamente la composición química y la temperatura del agua. En consecuen- cia, la fauna local cambia y diferentes especies se vuelven dominantes. Las presas constituyen barreras para los peces migratorios, como es notable ver en el sistema del río Columbia del oeste norteamericano. La ubicación frecuente de presas en las corrientes fluviales que erosionan activamente ha dado lugar a que esos depó- sitos de agua sufran un azolvamiento relativamente rápido y un cambio ecológico, mucho más de lo que ocurre en los lagos naturales; también se sufren las conse- cuencias río abajo.
Lagos
Aunque para el observador casual los lagos pueden diferir primeramente en el ta- maño y en la profundidad, el estudio científico ha descubierto innumerables pun- tos de diferencia. Muchas de estas diferencias constituyen determinantes críticos de la presencia, ausencia o abundancia de los peces. Por ejemplo, la capacidad de un lago para sostener una población ictiológica varía de acuerdo al tipo de su cuenca, la fertilidad del agua, la edad del cuerpo de agua, el empleo de la tierra en la cuenca y los cambios en las clases de peces o en la relativa abundancia de ciertas clases y tamaños de peces. La capacidad de mantenimiento ha sido relacio- nada con un índice morfo-edifico y una potencia conductora específica de agua.
Orígenes de los lagos. Entre los factores que distinguen a los lagos se cuentan sus orígenes naturales. Muchos han aparecido por la acción de los glaciares, espe-

ECOSISTEMA DE AGUA DULCE 423
cialmente los grandes glaciares continentales que cubrieron la corteza terrestre y dejaron como rastros morrenas formadoras de depósitos de agua. En las regio- nes que sufrieron la acción de los glaciares los bloques de hielo formaron muchas cavidades circulares en forma de cazo al quedar enterrados, se derritieron después y quedó el líquido encerrado. Otras se han formado por deslaves que taponan los valles y también debido a movimientos de la corteza terrestre que llegaron a for- mar depresiones colectoras de agua. La disolución de rocas subterráneas ha dado lugar a la formación de algunos depósitos lacustres, particularmente en regiones donde existe la caliza. Unos cuantos lagos pueden encontrarse en los cráteres de volcanes extinguidos. Los situados en la costa se han formado a partir de los que se encontraban detrás de las barras de arena o coral que se originan lejos de la costa; sus aguas son endulzadas por la lluvia cuando las barras eliminan más y más a las aguas marinas. No debe ser pasado por alto el origen de un lago por la acción de un río; hay lagos que se originan de la separación de porciones de mean- dros en llanuras bajas. Cualquiera que sea el origen de un lago tiene que sufrir una cierta evolución. A pesar del origen del lago, sus peces constituyen el producto de las conexiones originales del drenaje y de la supervivencia o adaptación al encontrar condiciones cambiantes. A través de la formación de reservas y otros procedimientos administrativos de la fauna ictiológica, y a través también de los usos conflictivos de la tierra y el agua, el hombre ha llegado a alterar notable- mente la evolución natural de la población ictiológica lacustre (y de la de los ríos).
Clasificación. Los lagos han sido clasificados de varias maneras de acuerdo a su estado evolutivo y a la dirección que sigue la evolución. Aunque no incluyen a todos, hay cuatro tipos básicos de lagos que son generalmente reconocidos en la literatura de la limnología: oligotróficos, eutróficos, mesotróficos y distróficos.
Los lagos oligotróficos son más a menudo profundos, fríos y pobres en nutrientes y plantas, especialmente gramíneas. Hay muchas especies diferentes pero pocos individuos y, en consecuencia, resulta una diversidad elevada. Estas especies están distribuidas hasta gran profundidad a lo largo de la columna de agua. Los peces más característicos de estos lagos, en la zona templada septen- trional, son los salmones de aguas frías y profundas, truchas y peces blancos (Sal- monidae). Los lagos de este tipo sufren una sucesión ecológica hacia el tipo eutró- fico; este proceso de envejecimiento se llama eutroficación. Los lagos eutróficos se transforman en estanques, pantanos y ciénagas. Los salmónidos de las aguas profundas faltan o están desapareciendo, a medida que los lagos se vuelven me- nos profundos, más cálidos y progresivamente más orgánicos y ocupados por plan- tas acuáticas de mayor tamaño. En la zona templada del norte los peces comunes de los lagos eutróficos incluyen a muchas clases diferentes de ciprínidos y a pe- ces de ojos laterales (Stizostedion), lucios (Esox), percas (Perca) y algunos bagres (Ictulurus; Silurus). Un lago mesotrófico está en una etapa media entre la del eutrófico y la del distrófico, cuando la cantidad de material orgánico reciclado en el lago está aumentando. Los lagos distróficos también son poco profundos y están condenados a transformarse en pantanos de turba. Su naturaleza elevada- mente orgánica es el motivo de que por lo menos en ciertas estaciones del año, falte el oxígeno y, en consecuencia, los peces. Representante de los peces que pue- den vivir ahí es el pececillo del fango (Umbridae).
Independientemente de que un grupo de lagos pertenezca temporalmente a cualquiera de las cuatro categorías citadas anteriomente, o a cualquier otra, cada

424 ECOLOGfA Y ZOOGEOGRAFfA
uno puede todavía diferir más en su área, profundidad, volumen, contorno del fondo, regularidad de su margen, así como en otras características. Todas estas características físicas tienen alguna relación con las clases como con la abundancia de los peces que pueden estar presentes.
Zonificación. Los medios ambientes del pez en los lagos pueden ser muy diversos. La diversidad es, sin embargo, agrupada convencionalmente en tres zonas principales: litoral, limnética y béntica. Estos tres términos corresponden aproximadamente, en el ecosistema marino, al nerítico, oceánico ( o pelágico) y béntico, como ya se ha mencionado (Figura 14.3).
Zona litoral. L a zona litoral de un lago se extiende desde las márgenes hasta el límite en donde crecen las plantas con raíces, que de ordinario dominan en la zona. Esta zona es singularmente importante porque es en donde desovan y se ali- mentan muchos peces típicos de los lagos. Más a menudo, a lo largo de las márge- nes de las aguas templadas hay plantas emergentes que incluyen a clases como el tule (Scirpus) y tifa (Typha). Más all6 de las plantas emergentes hay a menudo una región de plantas con hojas flotadoras como las ninfeas y nenúfares (Nymphaea, Nuphar), Brasenia y algunas algas (Potamogeton). Más allá de las plantas de hojas flotantes se encuentran las sumergidas que pueden cubrir el fondo hasta el límite exterior de la zona litoral. Como ejemplos de plantas americanas sumergidas hay muchas algas (Potamogeton, Myriophyllum, Ceratophyllum y Anacharis). En los lados de los lagos, hacia donde se dirige el viento, la vegetación con raíces puede ser escasa o faltar, y las mrirgenes pueden ser variablemente arenosas, erosionadas o formadas de roca desnuda, dependiendo de la conformación geológica del lugar. La zona litoral está bien iluminada pero sujeta a fluctuaciones, cortas en duración, de temperaturas máximas, turbulencias debida a la acción del viento y turbidez, aun cuando ésta sea orgánica; los materiales del fondo tienden a ser los más burdos cerca de la orilla. Los peces que requieren de las aguas más cálidas del lago se en- cuentran usualmente en la zona litoral. Muchos tienen el cuerpo relativamente corto y comprimido como, por ejemplo, la mojarrita (Lepomis) y los “crappies” (Pomoxis); esta forma del cuerpo es una adaptación al gran poder que tienen, durante la natación, de darse vuelta y recorrer distancias cortas (Capítulo 6).
Zona limnética. La zona limnética en un lago corresponde al agua abierta, o sea, a la extensión que de la zona litoral mira hacia el interior del lago. Las aguas limnkticas superiores están bien iluminadas y sujetas a un cambio relativamente rápido en sus condiciones fisicas; el nivel de compensación luminosa corresponde a su límite hacia abajo. Las aguas profundas limnéticas inferiores presentan con- diciones físicas comparativamente estables, o sea, con cambios lentos, con excep- ciones notables al cambiar de estaciones.
La zona limnética es el criadero de muchos peces desovadores de las aguas abiertas, incluyendo a grupos de ciertas sardinas de agua dulce (Clupeidae)* y charales (Atherinidae). Los peces más abundantes de la zona limnética son devo- radores de plancton, como el brillador esmeralda (Notropis atherinoides) y grupos de “alewife” (Alosa pseudoharengus) de agua dulce de los Grandes Lagos de Norte- américa. Los peces son más a menudo plateados, algo más oscuros por arriba que
*Como podría ser Signalosa mexicana, abundante en el lago de Catemaco. (N. del T.).

ECOSISTEMA DE AGUA DULCE 425
por abajo y de cuerpo hidrodinárnico o alargado, pero aplanado por los lados. Las aguas profundas que están situadas por debajo de la termoclina están pobremente iluminadas y habitadas enteramente por organismos heterotróficos. En estos luga- res los organismos dependen para obtener su energía, de la “lluvia” de material orgánico procedente de las aguas superiores. Algunos devoradores de plancton pueden vivir en esta zona, pero migran hacia la superficie para alimentarse. La diversidad de las especies ictiológicas es usualmente reducida.
Zona béntica. La zona béntica declina desde la orilla de un lago, por debajo de las aguas litorales, hacia las grandes profundidades de esta región. Al hacerlo puede cambiar el carácter del sustrato, desde el formado por un lecho rocoso, pasando por uno formado de material inorgánico y orgánico, y llegar finalmente a otros de fango orgánico muy floculado. Las condiciones químicas y físicas que afectan la vida en y sobre el fondo cambian de acuerdo con las condiciones de las aguas inmediatamente superiores. Los organismos del fondo (bentos) viven en la región donde hay mayor cantidad de nutrientes de un lago, pero están expuestos a sufrir los más grandes riesgos de eliminación debido a la aparición de una deficiencia de oxígeno, como resultado de la descomposición de los materiales del fondo. En las latitudes más septentrionales, los peces lacustres típicos de la región béntica inclu- yen a los cótidos (Cottidae), el “burbot” (Lotu), algunos bagres (Ictaluridae) y peces muy adaptados, que se alimentan en los fondos, como las carpas hociconas (Catostomidae) y los esturiones (Acipenser). La cabeza es deprimida en muchos de esos peces y, con frecuencia, también el cuerpo lo es. El hábitat béntico es también visitado por muchos otros peces lacustres que buscan alimentarse y de- sovar o construir sus nidos.
Corrientes
Las corrientes naturales son cursos de agua que se desarrollan a partir de las co- rrientes superficiales o de manantiales que se originan de aguas subterráneas. Sus rutas y gradientes sufren cambios continuos a lo largo del tiempo, a través de fenómenos como la disolución, la erosión y la acumulación o azolvamiento. Tal como ocurre con los lagos, también tienen las corrientes un patrón evolutivo (Figura 14.2), con cambios asociados con los peces que las recorren; parten de una corriente recientemente formada, de cauce recortado y con caídas de agua, hasta llegar a un estado de senectud caracterizado por constar de aguas len- tas, más la presencia de meandros cerca del nivel de la desembocadura. Los humanos han alterado en varios aspectos el patrón natural del drenaje de las porciones de tierra, sobre todo mediante la construcción de canales y desviaciones de agua. Me- diante la contaminación y el cambio de procedimientos en el uso de la tierra han cambiado mucho, también, las condiciones de vida en las corrientes, produciendo a menudo la escasez de peces en grandes extensiones de la cuenca.
Características. Las corrientes o las diversas ramificaciones de un río difieren de una a otra en muchos aspectos. Entre los parámetros de las diferencias físicas se cu3ntan la profundidad, la longitud, la anchura, el área, el volumen de corriente o la descarga, la regularidad de las márgenes, el declive, el tipo de fondo y la tempe- ratura. Las corrientes con estas diferencias pueden ser permanentes o intermiten-

426 ECOLOGfA Y ZOOGEOGRAFfA
tes. La corriente de algunos es interrumpida, vertiendo alternativamente por encima de los materiales del fondo, o a través de ellos. La mayoría presenta fluc- tuaciones en su descarga, con el cambio de las estaciones del año, y cerca de su desembocadura pueden inundar ampliamente las tierras bajas. Desde luego que en los arroyos que se forman en todos los hábitats acuáticos ocurren los cambios más inesperados y abruptos en las condiciones de vida.
Las corrientes son generalmente menos profundos que los lagos. Tienen flujos de agua que se mueven predominantemente en una dirección, sus materiales del fondo tienen menos estabilidad y hay más erosión. Su temperatura puede fluc- tuar rápidamente, pero se va haciendo más uniforme desde la superficie hacia el fondo. Tienden a saturarse de oxígeno desde la superficie hacia el fondo. La tur- bidez inorgánica es mayor en las corrientes y la penetración luminosa, a menudo mínima. El plancton escasea pero el perifiton es la base esencial de la productivi- dad. Las variaciones en la disponibilidad de nutrientes, de acuerdo al sustrato, son mayores en las corrientes que en los lagos, debido ostensiblemente a la imposibili- dad de acumular reservas orgánicas en el fondo. A causa de la fuerza del flujo hay en las corrientes un arrastre interminable de nutrientes.
Zonación. El reconocimiento de las zonas de vida en las corrientes comprende dos clases principales. En una, todo el curso de la corriente es separado en gran parte sobre la base del desnivel en extensiones superior, media e inferior. En la otra, recorre una extensión de rápidos, charcos, flujos, aguas en retroceso y desborda- mientos. Entre los hábitats que han sido reconocidos, son éstos los más importantes.
Extensiones. Las extensiones superiores de una corriente tienen los más grandes desniveles, la mayoría como valles en forma de V. Un ejemplo podría ser el arroyo de una montaña, posiblemente originado en un glaciar. La corriente es rápida, calmándose solamente en ocasiones al llegar a los charcos o pozas. De to- das las extensiones, las aguas son aquí las más frías. Estas son extensiones habitadas por peces tan activos como las truchas de arroyo (Salmo clarki y Salvelinus fonti- nalis) que encuentra refugio detrás de las rocas o en sus bordes durante el curso de la corriente. Las corrientes superiores bajan lentamente hacia las extensiones medias que tienen menor desnivel, corriente más lenta, casi el mismo número de lechos escabrosos y charcos o pozas, y algo de aluvión en las pozas quietas, así como aguas ocasionales en retroceso, todas con valles en forma de U. Las inunda- ciones que ocurren en determinadas estaciones del año están confinadas al canal de la corriente. En los manantiales los peces aumentan tanto en clases como en número; la fauna se vuelve más diversificada, lo que conduce a la estabilidad. A menudo varias clases de pececillos (Cyprinidae) se vuelven partes conspicuas de la fauna. En las extensiones bajas, la corriente se aproxima a las condiciones pro- pias del nivel basal de su desembocadura y el valle toma la forma de una U an- cha, usualmente con extensas llanuras inundables. Los lechos escabrosos desapa- recen y los charcos se hacen más grandes para formar desbordamientos extensos y quietos. Las orillas se pueden volver cenagosas y aparecen las aguas en retroceso, lodazales y corrientes cenagosas. El azolvamiento tiende a ser grave y la corriente en temporadas estacionales abandona su cauce para desbordarse, dejando a me- nudo varados a los peces cuando la inundación baja de nivel. Las aguas son las más cálidas de todas las extensiones. Los peces de las aguas lentas aparecen, con mayor tolerancia al medio ambiente y hábitos alimenticios generalmente menos

ECOSISTEMA DE AGUA DULCE 427
restringidos. Dentro de su área pueden abundar las carpas hociconas (Catostomi- dae), los más grandes pececillos o “minnows” (Cyprinidae), incluyendo la carpa (Cyprinus), y los bagres (Siluroidei).
Dependiendo del continente, una de tres familias de peces de agua dulce tiene por lo menos algunos representantes en practicamente todas las zonas bio- lógicas fluviales. Estos son los “minnows” (Cyprinidae), caracinos (Characidae) y cíclidos (Cichlidae). Australia es la excepción, porque no tiene ninguno de estos tres.
Corrientes rúpidas y charcos. Una masa de agua con lecho escabroso en las corrientes es a menudo relativamente poco profunda y corre sobre grava y pe- druzcos. Su superficie es irregular, en contraste con el aspecto liso de un estanque o un cuerpo de aguas en retroceso. Los fondos de estos cuerpos de agua constitu- yen las porciones donde desovan peces de las corrientes, los cuales tienen el hábito de enterrar sus huevos en la grava; entre éstos quedan incluidos las lampreas (Petromyzonidae), las truchas de arroyo y los salmones de la familia Salmonidae, y muchos pececillos “minnows” (Cyprinidae), carpas hociconas (Catostomidae) y pércidos como Etheostoma (Etheostomatinae). Debido a la riqueza fisiológica del área los fondos de estas porciones de agua son también territorios importantes para la producción de insectos como alimento de los peces, especialmente mu- chos tricópteros (Trichoptera), efeméridos (Ephemeroptera), plecópteros (Plecop- tera) y langostas de río (Decapoda).
Las pozas o charcos tienden a ser más amplios y profundos que el prome- dio de la corriente. En ellos la corriente es reducida, puede ocurrir algún azolva- miento pequeño y las plantas acuáticas con semilla pueden formar lechos. Los charcos son importantes para los peces de los ríos como áreas para la crianza y como sitios para la alimentación y la confección de sus nidos. Los charcos que están cubiertos por bancos volados o cualquier otro tipo de protección sostienen, según parece, un número mayor de peces de tamaño más grande, que en los que no tienen alguna cobertura. El alimento de los peces en los charcos incluye mate- riales de los rápidos, junto con insectos terrestres y acuáticos. Estos insectos acuá- ticos incluyen a ninfas de efeméridos cavadores (Hemgenia) y ninfas de caballitos del diablo (Odonata). Cerca de los bordes de los lechos de plantas acuáticas pueden abundar los anfípodos (Amphipoda). En los charcos más quietos y en las aguas en retroceso, donde se acumula el cieno rico en materia orgánica, los organis- mos importantes como alimento para los peces incluyen a las larvas de moscas de dos alas, como los tendipédidos (Tendipedidae) y los gusanos tubifícidos (Tubifi- cidae). Los gusanos del fango están presentes en la mayoría de las áreas con fondos llenos de materia orgánica, pero son más abundantes en las localidades con elevada contaminación orgánica. Cuando la abundancia alcanza niveles altos esos gusanos son indicio de una gran contaminación orgánica.
Fuentes termales
Las fuentes termales vierten tanto en los lagos como en los ríos. Esas aguas ter- males están ampliamente distribuidas sobre la tierra y son calientes, cristalinas, muy mineralizadas y, generalmente, desprovistas de o con muy poco oxígeno. No se sabe si los peces puedan establecerse en aguas de temperatura superior a los 40”C, aunque pueden sobrevivir durante cortos periodos en aguas de más de 45°C.

428 ECOLOGIA Y ZOOGEOGRAFfA
Aunque existen algunos reportes sobre peces que viven en aguas de 48" a 70"C, este hecho es insostenible pues las observaciones que se han hecho no consiguen respal- darlos. Hay numerosos ejemplos de peces que viven en aguas calientes en el sur- oeste de Norteamérica. Por ejemplo, tres especies son del Valle de la Muerte y viven a más de 45°C. La mayoría de estos peces de aguas cálidas son del género Cyprinodon y de unos cuantos géneros relacionados. Por otra parte, quedan in- cluidos ciertos "minnows" (Cyprinidae) y cíclidos (Cichlidae).
ZOOGEOGRAFÍA
La distribución geográfica y la ecología están estrechamente relacionadas entre sí y con la evolución. Algunas de las primeras evidencias críticas que convencieron a Darwin y Wallace sobre el hecho de la evolución fueron proporcionadas por la zoogeografía. Esta ciencia interdisciplinaria, a través de contribuciones del regis- tro fósil, también pretende reconstruir y entender la historia evolutiva de los organismos desde los puntos de vista del espacio y del tiempo, y proporcionar una prueba independiente para las conclusiones a las que han llegado los geólogos acerca de la excitante teoría, hoy en boga, de la tectónica de placas (véase abajo). Si nosotros pretendemos escudriñar el pasado geológico estudiaremos entonces el campo de la biogeografía histórica; si nuestro interés es por lo contemporáneo, entonces se volverá de nuestro mayor interés la ecología geográfica. Estos dos acercamientos han avanzado mucho durante la pasada década, el primero debido a los notables descubrimientos hechos en las ciencias de la Tierra, y el segundo gracias a las portentosas ideas nuevas en la teoría biológica (MacArthur, 1972; MacArthur y Wilson, 1967; Vuilleumier, 1974).
Las conclusiones que se derivan de los estudios biogeográficos son tan váli- das como lo sea la taxonomía de los organismos en la que ellos se basan, y es necesario hacer un trabajo penetrante sobre la zoogeografía de los peces. Todavía no tenemos los suficientes conocimientos acerca de las relaciones filogenéticas de algunos grupos animales como para estar en condiciones de distinguir entre lo que es un hecho y nuestra simple opinión.
La ciencia de la zoogeografía está relacionada con la distribución geográfica de los animales sobre la Tierra y con la historia que respalda a estas distribuciones. Los animales son organismos vivientes que continuamente están evolucionando y multiplicándose en algunos lugares, extendiéndose en otros sitios y desvanecién- dose en otros más, todo lo cual contribuye a la formación de nuevos patrones geográficos. Las áreas de distribución de las especies y de los grupos son, por lo tanto, pulsátiles y dinámicas, no estáticas, como nosotros estamos forzados a visualizarlas en los mapas de distribución. Las intenciones del zoogeógrafo no son solamente las de describir los patrones distributivos actuales, sino explicar cómo y por qué se han formado esos patrones. Y para esto tanto el tiempo como el espacio deben ser computados. Para lograrlo, la zoogeografía debe delinearse sobre la ecología, la evolución y la geología (incluyendo la paleontología y la paleoclimatología). La distribución animal puede revelar nuevos aspectos sobre cómo tiene lugar la evolución; puede auxiliar al geólogo para interpretar el pa- sado; y, como muy atinadamente lo recalca Darlington, es por sí mismo un es- tudio fascinante.
Por mera conveniencia, la zoogeografía puede ser dividida en tres catego-

ZOOGEOGRAFfA 429
rías: las distribuciones de carácter mundial, que revelan patrones muy amplios; las distribuciones regionales, que conducen a grupos particulares en partes selectas del mundo; y las distribuciones locales, que están relacionadas con la geografía y la ecología de las especies. Las tres son tratadas aquí, pero se enfatiza sobre el aspecto más amplio.
La deriva continental
Hace apenas poco más de dos décadas la mayoría de los geólogos consideraba rígida a la Tierra y creía que los continentes se habían formado donde ahora se encuentran. Una nueva teoría sobre deriva continental fue propuesta a principios del presente siglo, pero la ignorancia sobre la existencia de un mecanismo que pudiese mover a los continentes aseguró su casi universal rechazo. Tal como ha sucedido en la historia del pensamiento, cualquier visión heterodoxa se ha vuelto con el tiempo correcta.
Los geofísicos y los geomorfólogos, estudiando el fondo de los océanos, han descubierto un mecanismo en la actual teoría de la tectónica de placas de la litósfera (también denominada de nueva tectónica global) y la ampliación del fondo marino, la cual propone que la superficie de la Tierra está dividida en un número pequeño de placas bordeadas por cordilleras y zonas de subducción. El nuevo fondo oceánico es formado hacia las cordilleras tan luego como las por- ciones más antiguas de las placas son empujadas lejos. Con el fin de equilibrar esta agregación las partes antiguas de las placas son hundidas en el interior de la tierra en la zona de subducción. La tectónica de placas se basa esencialmente en el océano, es decir, el paquete más grueso de la intensa actividad térmica y sís- mica está asociado con los bordes de las placas bajo el océano y en los bordes de los continentes. Los conceptos básicos de la tectónica de placas y la ampliación del fondo marino se fundan seguramente en los datos magnéticos oceánicos, en investigaciones sismológicas y en la perforación de los fondos marinos que ha sido (y todavía lo es) proseguida sin descanso por el buque de investigaciones Glo- mar Challenger.
“La naturaleza de la ampliación del fondo marino y la deriva continental en el Pre-Jurásico, hace más de 200 millones de años, todavía no ha sido descu- bierta. El registro fósil puede ayudar mucho en este problema, ahora que estamos aprendiendo a interpretar la interacción entre la evolución y la deriva continen- tal” (Scientific American, 1974: 159).
Los continentes descansan pasivamente sobre las placas y se desplazan con ellas, más que avanzar sobre un fondo oceánico sólido, como antes se pensaba. Por lo tanto, la deriva continental es una de las consecuencias de la tectónica de placas. Otras consecuencias importantes consisten en la ocurrencia de terremotos a lo largo de las fronteras de las placas (por ejemplo, la famosa falla de San Andrés en California) y en el desarrollo de cadenas montañosas donde chocan dos placas portadoras de continentes (como cuando el subcontinente indio chocó con Asia y formó la cordillera del Himalaya).
Al tratar de contestar a preguntas sobre si hay patrones distribucionales generales comunes a una diversidad de organismos y, de ser así, cuáles son esos pa- trones y cómo es que han cambiado a lo largo del tiempo, los primeros estudiosos de la zoogeografía histórica (por ejemplo Wallace, Matthew, Simpson y Darlington)

430 ECOLOGfA Y ZOOGEOGRAFfA
han asumido que los continentes y las cuencas oceánicas han permanecido relati- vamente estables, por lo menos a lo largo del Cenozoico y Mesozoico, por no decir todo el tiempo geológico.
De acuerdo a los conceptos actuales de los especialistas en las ciencias de la Tierra, todos los continentes estaban fusionados hace unos 225 millones de años (Era Paleozoica) formando un supercontinente llamado Pangea, que después se fragmentó (hace unos 200 millones de aiios) dando lugar a la formación de una gran masa continental hacia el norte, la Laurasia (Eurasia y América del Norte) y una porción continental hacia el sur, la Gondwana (Antártida, los continentes del sur y el subcontinente de la India). La Gondwana después se fragmentó dando lugar al continente africano y a la India, después a la porción este de la Antártida y Australia, seguido por Nueva Zelanda, y luego, hacia el oeste, con la Antártida y América del Sur (Figura 14.4).
Para determinar la biogeografía histórica de un grupo como el de los verte- brados, se ha desarrollado un conjunto de principios de trabajo dentro de una teoría biogeográfica. Esta teoría especifica que una meta primaria consiste en elaborar la hipótesis más parsimoniosa sobre la localización de las especies ances- trales, basada en la clasificación filogenética (Capítulo 13). Por lo tanto, la re- construcción de los patrones de distribución es una inferencia deductiva basada en un previo análisis filético (Nelson, 1969).
Guías para la historia geográfica
Al pretender explicar el origen y dispersión de los organismos, se han formulado ciertos criterios que pueden proporcionar indicios valiosos para su historia:
a. La aparición del fósil más antiguo es u.n indicio importante para ubicar el origen de un grupo, principalmente cuando el registro fósil es compa- rativamente bien conocido (como en los mamíferos). Sin embargo, un registro fósil deficiente puede ser peor que no tener nada.
b. Probablemente un grupo surgió en la área donde se muestra la mayor diferenciación taxonómica. Los géneros y las especies deberán revelar di- ferencias más grandes, mayor diversidad y un número más grande de formas endémicas en una región donde el grupo ha estado durante largo tiempo, más que aquel a donde ha llegado. Los “minnows” (Cyprinidae) del sureste de Asia proporcionan un buen ejemplo.
Figura 14.4 - Algunos efectos del movimiento de las placas continentales (tectónica de placas o deriva continental) e invasiones cambiantes de los continentes por mares de poca profundidad en la geografía y el clima del globo y, en consecuencia, en la dis- tribución de los peces. (a) Placas continentales más o menos unidas para formar una masa continental única, hacia fines del Paleozoico. (6) Los mares separan las placas continentales en las postrimerías del Mesozoico, y los mares poco profundos se han retirado de los continentes. (c) AI comenzar el Cenozoico se restablecen en algunas Breas los mares poco profundos pero las áreas continentales son de mayor extensión en relación a las del Mesozoico; el clima se vuelve extremoso pero la distribución de las palmas fósiles muestra que el área de distribución de los organismos tropicales y subtropicales permanecía muy extensa. (Adaptado de Newell, 1972).

ZOOGEOGRAFfA 431
(a) Finales del Paleozoico
(b) Finales del Mesozoic0
( c ) Comienzos del Cenozoico
de agua cálida
-" coral y rudistas f6siles
f6siles

432 ECOLOGfA Y ZOOGEOGRAFfA
c. El lugar que contiene el mayor número de géneros y especies puede ser considerado como el área de origen del grupo. Sin embargo, considerando que la mayoría de los animales están probablemente disminuyendo y extinguiéndose como, por lo común, grupos nuevos están aumentando y extendiéndose, los datos sobre su n6mero a menudo conducirán a obtener conclusiones erróneas acerca de la historia de un grupo. Por ejemplo, los lucios y los “minnows” del fango (Esocidae y Umbridae, respectivamente) están mejor representados en Norteamérica (siete espe- cies), pero aparecieron por primera vez en Europa (tres especies) y, en consecuencia, probablemente se originaron en el Viejo Mundo (Figu- ras 14.5 y 14.6).
d. Otro indicio es la extensión del área, en donde se supone que la edad de un grupo es proporcional al área ocupada y se presume que el lugar de origen corresponde al centro de la superficie, la hipótesis de la “edad y kea” del botánico Willis. Los estudios de los ciclos de dispersión y concentración revelados por los registros de vertebrados fósiles no per- miten sostener este punto de vista. Pero este indicio cuenta con algún valor si se combina con otros.
e. Un indicio importante consiste en la continuidad del área. En el grupo de las carpas hociconas (Catostomidae), la carpa hocicona de nariz larga (Catostomus catostomus) aparece fuera de Norteamérica solamente en el este de Siberia, y esto es consistente con la idea de que invadió esa área recientemente. En contraste con lo anterior, la carpa hocicona china, muy diferente y del género Myxocyprinus, está bien aislada del resto del área de distribución de la familia y esto respalda la creencia de que se trata del relicto de una fauna tempranera de carpas hociconas chinas que, desde entonces, se ha reducido hasta llegar casi a la extinción en el sur- este de Asia (Figura 14.7).
f. La distribución de familias relacionadas, competidoras y asociadas, pro- porciona un indicio de mucha utilidad para las historias de la familia. Los parientes más cercanos de las carpas hociconas son los “minnows” (Cy- prinidae), de los cuales probablemente derivaron en el este de Asia; el grupo en forma migró subsecuentemente hacia América del Norte, mostrando entonces una mayor diferenciación en una región donde sus más cercanos competidores son comparativamente pocos en número. La historia probable de las carpas hociconas (Catostomidae) sugiere que los centros de evolución y dispersión también parecen ser centros de extinción.
GEOGRAF~A DE LOS PECES DE AGUA DULCE
Los peces de agua dulce, cuyo número excede las seis mil especies, son de gran importancia en la zoogeografía, pensando incluso que no siempre puede ser hecha una distinción consistentemente clara entre los grupos, basada en su tolerancia a la sal. Por lo general, ellos pueden pasar de un sistema fluvial a otro, solamente a través del cambio fisiográfico lento de la propia tierra. Ellos poseen típicamente una incapacidad fisiológica para sobrevivir en plena agua de mar. Como resultado de esto, su dispersión sobre el mundo es lenta y, por lo tanto, es muy seguro que preserven sus antiguos patrones de distribución (Figura 148) .

GEOGRAFfA DE LOS PECES DE AGUA DULCE 433
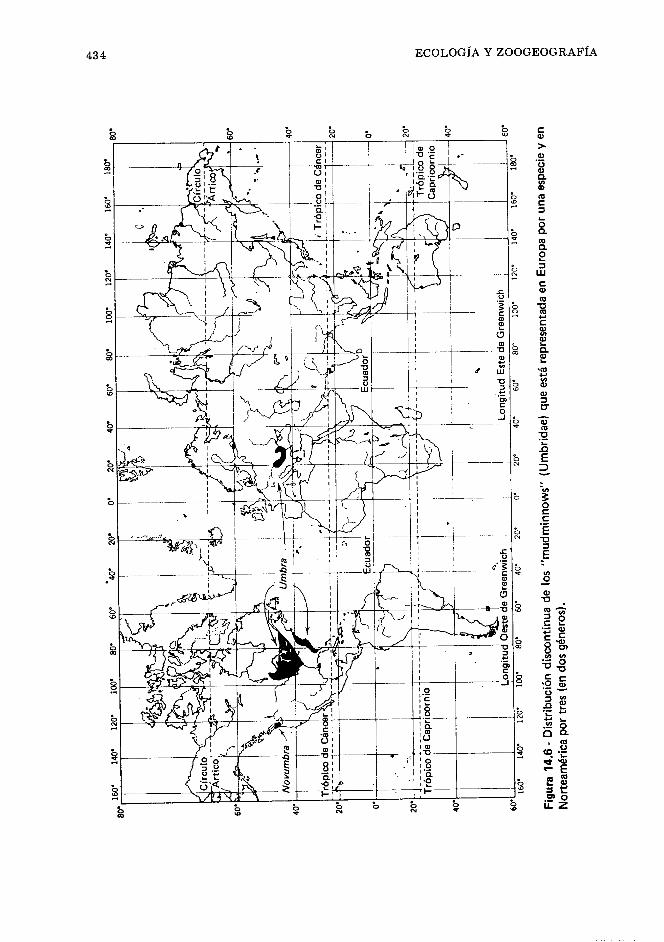
434 ECOLOGÍA Y ZOOGEOGRAFfA
C W
m C 7
O P
L

GEOGRAFfA DE LOS PECES DE AGUA DULCE 43 6

436 ECOLOGfA Y ZOOGEOGRAF~A

GEOGRAFfA DE LOS PECES DE AGUA DULCE 437
Aunque los peces de agua dulce estrictos o primarios son únicos por su valor excepcional para los estudios zoogeográficos, también tienen importancia otros grupos para explicar los patrones de distribución regionales y locales. Mu- chos de los peces de agua dulce secundarios, especialmente los del orden Cyprino- dontiformes (por ejemplo, la familia mexicana Goodeidae), están restringidos al agua dulce y han probado ser de valor en el trazado de las primeras conexiones hidrográficas entre los desagües aislados de la actualidad.
Grupo de peces de agua dulce
Los peces óseos (Osteichthyes) constituyen los grupos más importantes de peces de agua dulce. fistos pueden ser divididos en los no teleósteos (Holostei y Chon- drostei) y los teleósteos (Teleostei); además, el segundo puede ser separado en los Ostariophysi o Cypriniformes (“minnows”, carpas hociconas, bagres, caraci- nos, “loaches”, etc.), y los no Ostariophysi. Los peces primitivos no teleósteos incluyen a grupos de todos los continentes, por ejemplo los “bichirs” africanos o poliptéridos, los peces espátula chinos y de América del Norte (Psephurus y Po- Zyodon) y los peces pulmonados de América del Sur, Africa y Australia (Dipnoi) que comprenden dos líneas evolutivas diferentes. Los teleósteos son numerosos, de origen diverso y de amplia distribución; incluyendo, en América del Norte, los peces ojo de luna (Hiodon), los lucios y “mudminnows” (Esox, y Umbra y No- uumbru, respectivamente), muchos cipriniformes, perca-truchas (Percopsidae) y percas pirata (Aphredoderidae), peces sol y mojarritas (Centrarchidae) y las percas y peces saeta (Percidae).
Los cipriniformes, cuyo número de especies llega a ser de cinco mil aproxi- madamente (o cerca de una cuarta parte del número total de peces conocido), incluyen a las hordas de peces que predominan en todos los continentes, a excep- ción de Australia. Solamente los peces parecidos a las percas (Perciformes) con cerca de ocho mil especies los sobrepasan. De las aproximadamente treinta y cinco familias que constituyen el orden Cypriniformes, solamente dos, los bagres de las familias Ariidae y Plotosidae, son marinos, y sin duda alguna que han deri- vado de ancestros de agua dulce; ellos solos representan al orden en Australia y Madagascar, donde algunos han llegado a reinvadir las aguas dulces.
Patrones continentales
La diferenciación de las faunas ictiológicas en los diferentes continentes es uno de los más importantes patrones distributivos de los peces de agua dulce.
La fauna ictiológica dulceacuícola primaria de América del Norte incluye veintiún familias que comprenden cerca de seiscientas especies. Su centro de abundancia y diversificación se ubica en la cuenca del río Mississippi (con cerca de doscientas sesenta especies en trece familias), hacia el sureste de Estados Uni- dos y en el drenaje de los Grandes Lagos, el último enriquecido por la invasión de la fauna del Mississippi durante las edades glacial y postglacial. La fauna se empobrece hacia el oeste de las Montañas Rocallosas y hacia el sur del Istmo de Tehuantepec, y también escasea marcadamente en Canadá, México y Centroamé- rica, pero se enriquece hacia el este de Panamá, debido a la intromisión faunística

438 ECOLOGfA Y ZOOGEOGRAFfA
procedente del vasto territorio sudamericano. No hay verdaderos peces de agua dulce en las islas de las Indias Occidentales, pero sí en cinco de Alaska.
Hacia el norte del margen meridional de la Altiplanicie Mexicana la fauna es dominada por los “minnows”, carpas hociconas y los peces dardo, que juntos comprenden algo así como cuatrocientas cincuenta especies 0 el 90% de la fauna estrictamente de agua dulce.
Cerca del 30% de todos los peces de agua dulce de América del Norte tienen su origen en la propia América del Norte. Quedan incluidos los peces ojo de luna (Hiodontidae), peces cavernícolas (Amblyopsidae), truchas-perca (Percopsidae), percas pirata (Aphredoderidae), bagres ictalúridos (Ictaluridae), peces sol (Cen- trarchidae) y peces dardo (Etheostomatinae). Cerca del 55% tiene ancestros euroasiáticos, incluyendo a los lucios (Esocidae), “mudminnows” (Umbridae), “minnows” (Cyprinidae), carpas hociconas (Catostomidae) y percas (Percidae), aparte de los peces dardo. Aproximadamente el 15% tiene afinidades sudameri- canas e incluye a los caracinos (Characidae), gimnótidos (Gymnotidae) y muchas familias del grupo de los bagres, pero principalmente a los Pimelodidae.
Los peces de agua dulce primarios de América del Sur están extraordinaria- mente limitados en número de grupos pero, al mismo tiempo, constituyen, de todas las faunas continentales, la más rica y más variada, comprendiendo proba- blemente cerca de dos mil especies. Éstas se derivaron de unos cuantos ancestros, desde el momento en que solamente incluyen a dos grupos principales, los caraci- nos (con los gimnótidos como derivados) y los bagres. Solamente hay otros tres grupos primarios (no ostariofisios): los viejos peces pulmonados (Lepidosiren), con una especie; los peces de lengua de hueso (Osteoglossidae), con dos especies; y dos especies representando al pez de radios espinosos, de la familia Nandidae. Estas tres familias también aparecen en África, y las dos últimas también en el Oriente. La fauna sudamericana se desarrolló en el aislamiento durante la mayor parte del periodo Terciario, cuando este continente se separó de América del Norte. La fauna es arrolladoramente dominada por los Ostariophysi, más de la mitad de las cuales está representada por doce familias endémicas o autóctonas de bagres, y es primordialmente africana en sus relaciones.
Las relaciones directas entre África y América del Sur son justa y fuerte- mente respaldadas por las afinidades de las familias secundarias, los “killifishes” (Cyprinodontidae; Figura 14.9) y los cíclidos (Cichlidae; Figura 14.10). El proble- ma de la derivación de esta fauna está por ser resuelto todavía. Algunos han argüido enérgicamente que los peces vinieron de África por medio de un pasaje de tierra; otros señalan la ausencia de muchos grupos importantes de peces africanos en América del Sur y creen que la migración se produjo a través de América del Nor- te, con la subsecuente extinción en el norte. El registro fósil es muy fragmentario como para poder proporcionar un indicio seguro para resolver el enigma.
La fauna ictiológica de agua dulce del África se caracteriza por su gran diversidad, así como por su riqueza, y también por el desarrollo de extraordina- rios enjambres de especies y el elevado endemismo que hay entre los peces cíclidos (Cichlidae) de los grandes lagos formados por fallas geológicas. Hay numerosas formas arcaicas y generalizadas; por ejemplo, los peces pulmonados (Protopteri- dae) y los polípteros (Polypteridae), y una variedad de grupos isospóndilos, inclu- yendo a los mormíridos (Mormyridae). Hay veintitrés familias primarias que comprenden cerca de mil cuatrocientas especies. Los “minnows”, caracinos y peces gato están ampliamente distribuidos, y también hay grupos de peces de

GEOGRAFfA DE LOS PECES DE AGUA DULCE 439

440 ECOLOGfA Y ZOOGEOGRAFfA

GEOGRAFfA DE LOS PECES DE AGUA DULCE 441
radios espinosos, superiores; por ejemplo, las percas arborícolas (Anabantidae) y las anguilas mastacembélidas (Mastacembelidae). La gran fauna de ciprínidos comprende diversificaciones menores, incluyendo a una forma ciega, procedente de unos cuantos tipos, muy probablemente de reciente introducción de Asia. El gran desierto del Sahara, hacia el norte, y el desierto Kalahari hacia el suro- este, restringen las áreas disponibles para la habitación de los peces, pero la mayor parte del continente africano representa una masa primitiva de tierra que ha estado continuamente sobre el nivel del mar desde los tiempos precámbricos, permi- tiendo entonces que durante largo tiempo se realizara la evolución de su fauna característica. La muy larga estabilidad de esta masa continental también permi- tió la supervivencia de grupos primitivos de peces. El África tropical occidental es el área más rica y tiene una gran diversidad de troncos primitivos no igualada en ninguna parte del mundo. La porción aislada hacia el noroeste del Africa con- tiene una fauna ictiológica empobrecida y muy relacionada con la del sur de Europa.
Poco es lo que se conoce sobre el origen y la dispersión de los grupos más antiguos de los peces primarios del África, pero se presume que ocho familias son autóctonas: los poliptéridos (Polypteridae), los peces elefante (Mormyridae), los Gymnarchidae, los saltarines del fango africanos (Pantodontidae); los Phractole- midae, Kneriidae, Cromeriidae y los bagres eléctricos (Malapteruridae). Las familias que incluyen a los “minnows” (Cyprinidae) y a los cobítidos (Cobitidae), y probablemente algunas de las formas de radios espinosos, aparentemente pene- traron al África procedentes de Asia, durante el Plioceno y el Pleistoceno.
La fauna ictiológica de Eurasia está caracterizada por la escasez de relictos filogenéticos (solamente Psephurus, el pez espátula oriental, y Myxocyprinus, la carpa hocicona (Figura 14.7) del río Yangtze y sus vecindades) y gran riqueza y diversidad de los peces ciprinoideos hacia el este de Asia, representada por los “minnows” (Cyprinidae), cobítidos (Cobitidae) y los peces de los arroyos de las montañas (Homalopteridae). En China, por ejemplo, cerca del 80% de los casi seiscientos peces primarios pertenece a los Cypriniformes (ciprinoideos y cuatro familias de bagres o siluroides). Hacia el sur, a través de China tropical, la fauna se fusiona o confunde de una manera compleja con la fauna ictiológica de la re- gión oriental. Europa, que sufrió recientemente una glaciación, tiene una fauna empobrecida, de la cual solamente seis familias, comprendiendo ciento dieciocho especies, forman los peces de agua dulce primarios. Ninguno de los grupos es en- démico y tres familias (“mudminnows”, Umbridae, lucios, Escocidae, y percas, Per- cidae) comprenden solamente trece especies; los “minnows” (Cyprinidae) y los cobítidos (Cobitidae) forman el grueso de la fauna, y solamente hay dos bagres (Siluridae). Hay una abundancia relativa de grupos como el de las lampreas (Petromyzonidae), esturiones (Acipenseridae), arenques (Clupeidae), truchas (Sal- moninae) y gobios (Gobiidae), que toleran o gastan regularmente una parte de su ciclo de vida en el agua marina; también éstos incluyen a las especies tolerantes a las bajas temperaturas. La fauna es bien conocida en Europa, menos en Rusia, y pobremente conocida en China, a pesar de tener el mayor número de especies.
La Región Oriental incluye la parte principal del Asia tropical, estre- chándose desde la India y el Sur de China, hacia el sur, a través del Archipiélago Indo-Australiano (Sumatra, Java y Borneo) hasta la Línea dewallace (Figura 14.8). Es notable por la riqueza y diversidad de los peces ciprinoideos y de los bagres (nueve familias, seis endémicas) y por la ausencia de grupos arcaicos, y casi to- das las familias de peces, parecidos a los arenques, del Africa; es moderadamente

442 ECOLOGÍA Y ZOOGEOGRAFÍA
rica en peces de radios espinosos. Queda incluida una gran variedad de peces de tierras bajas, algunos de los cuales (como el Chaudhuriidae de la India) son exclu- sivos de la región, y también algunos grupos altamente especializados para vivir en los torrentes montañosos (en particular los homaloptéridos, Homalopteridae, y los chupadores de vientres, Gastromyzonidae).
Australia casi carece de peces de agua dulce verdaderos; solamente pueden ser colocados en esta categoria el arcaico pez pulmonado (Neoceratodus) y un pez de lengua osificada (osteoglósido) (Scleropages). El resto de la fauna dulce- acuícola empobrecida comprende a grupos que han derivado de otros marinos, cerca de ciento ochenta clases, excluyendo a las especies introducidas y a las vagabundas. Los principales grupos marinos representados en los ríos de Austra- lia son las lampreas (Petromyzonidae), peces sierra (histis), arenques (Clupeidae), “smelt” (Retropinna), galáxidos (Galaxiidae), bagres marinos (Ariidae), anguilas (Anguilla), peces plateados (Atherinidae), lisas (Mugilidae), serránidos (Serrani- dae), terapónidos (Theraponidae), peces cardenal (Apogonidae) y gobios (Gobiidae y Eleotridae).
GEOGRAFfA DE LOS PECES MARINOS
A diferencia de los peces que viven en las aguas dulces continentales, los peces marinos no están primariamente restringidos en su distribución por barreras físi- cas, no considerando como tales a los bordes continentales. El factor importante que limita su distribución es la temperatura. Otras influencias distribucionales mayores corresponden a las grandes expansiones de las aguas oceánicas profundas (como las hay entre América y Polinesia), la naturaleza de la configuración cos- tera y los contornos submarinos, la variación en la salinidad del agua y los efectos de las corrientes oceánicas (no solamente por lo que respecta a la dirección del flujo, sino a su temperatura). La dispersión al azar de organismos que están vi- viendo etapas pelágicas de su ciclo biológico por medio de la contracorriente ecua- torial, por ejemplo, puede ser responsable de la existencia común de especies idénticas o muy relacionadas en las costas del Atlántico, frente al Africa y las Américas.
Con propósitos zoogeográficos los peces marinos pueden ser clasificados en tres principales categorías: a) la fauna de la costa o de los bancos de arena, com- prendiendo especies que viven dentro de la costa, a lo largo de los arrecifes conti- nentales, a profundidades de 100 brazas (600 pies, o cerca de 200 metros) o menos; b) los peces pelágicos o de mar abierto, que viven generalmente cerca de la superficie de los mares tropicales o de aguas templadas, a menudo lejos de la costa, a 100 brazas o menos, y c) formas de los mares profundos o abisales, vivien- do a profundidades mayores de las 100 brazas, que también pueden ser divididas en formas batipelágicas y bénticas, las segundas peces de las profundidades, que viven a casi 6,000 brazas (Fosa Challenger, cerca de Guam).
Las formas costeras, que muy bien incluyen a más de la mitad de las espe- cies conocidas de peces marinos, ocupan los bordes continentales (incluyendo al fondo marino), desde la zona comprendida entre los niveles mínimo y máximo de las mareas (la fauna litoral, tal como ha sido restringida por muchos autores) hasta sus bordes exteriores. Dentro de los límites fijados por la temperatura, sus áreas de distribución están muy limitadas por la extensión de la línea costera

GEOGRAFfA DE LOS PECES MARINOS 443
disponible y por los archipiélagos. En contraste, el mar abierto es generalmente una vía de dispersión para los peces pelágicos, muchos de los cuales pueden cruzar áreas que tienen aguas con temperaturas marcadamente diferentes, por ejemplo, el tiburón ballena (Rhincodon), el dorado (Coryphaena), el pez espada (Xiphias) y los atunes (Thunnus). Los peces circunglobales quedan comprendidos desde los lanceta o anfioxus (Branchiostomidae) y tiburones (Squaliformes) primitivos hasta el muy especializado pez tambor o globo (Tetraodontidae) y los cerátidos (Cera- tiidae), con cerca del 85% de esas especies ampliamente distribuidas, típicas del mar abierto. Sin embargo, la barrera constituida por la separación entre la Poline- sia y América ha evitado en más de una tercera parte la extensión territorial de los peces pelágicos y batipelágicos, como consecuencia de su incapacidad para cruzar hacia la porción oriental del océano Pacífico.
Nosotros sabemos menos acerca de la fauna de las profundidades marinas, y las ideas hasta hoy aceptadas sobre su distribución deberán ser modificadas tan pronto como la información nueva sea puesta a disposición a través de los avances en la investigación oceanográfica. Parece ser que muchas de las especies obedecen a sus propias leyes de distribución. Y no con poca frecuencia ellas están amplia- mente distribuidas, debido evidentemente a que las barreras que se oponen a la dispersión, están en el presente caso ausentes por no encontrarse en las regiones poco profundas, como generalmente sucede. O, visto de otra manera, la enorme conformidad física dentro de las masas de agua oceánica -comparadas con las aguas costeras- ha permitido que las especies se distribuyan ampliamente. Aun- que nosotros tenemos mucho que aprender acerca de las formas de los fondos marinos bénticos, puede generalmente decirse que su distribución es siempre res- tringida; se sabe que solamente tres de esos peces tienen distribución circuntropi- cal. La fauna de los fondos marinos está empobrecida, en evidente correlación con la severidad de las condiciones del medio ambiente y la escasez que hay de mate- riales alimenticios en las grandes profundidades. En general, puede decirse que con el incremento de la distancia de los continentes el pico de las profundidades ma- rinas tiende a ser ocupado por menor cantidad de individuos pertenecientes a menor cantidad de especies.
Aunque se piensa generalmente que las faunas ictiológicas de las aguas pro- fundas no están diversificadas, la exploración de la Fosa Kuriles-Kamchatka ha revelado cerca de cincuenta especies que pertenecen a veintidós familias; quince de estas familias eran previamente desconocidas para el territorio ruso. Se cree que estos peces de las aguas profundas representan a especies antiguas que difie- ren en su origen y distribución de las poblaciones jóvenes de peces de las capas de agua superiores.
Las faunas tropicales de la costa son las más ricas y más diversificadas, no solamente por lo que respecta a los peces, sino también a otros organismos mari- nos. Debido a su necesidad especial por el agua caliente (promedio anual de tern- peraturas en la superficie, de 20°C o más), estos peces difícilmente penetran a las regiones subtropicales del norte y del sur. En consecuencia, las faunas tropicales exhiben un elevado grado de endemia. Las faunas subtropicales pueden estar gene- ralmente caracterizadas como faunas tropicales desleídas, limitadas por una tern- peratura anual mínima de unos 16" a 18°C; ellas tienen mucho en común con la fauna tropical, con mucho menor parecido a la fauna vecina de aguas templadas. Muchos peces tropicales aparecen a través de los trópicos en ambos lados de 10s océanos Atlántico y Pacífico, y se dice de ellos que son circuntropicales.

444 ECOLOGfA Y ZOOGEOGRAFfA
Las faunas costeras tropicales pueden ser separadas en cuatro regiones superiores: del Indopacífico, del Africa Occidental, de las Indias Occidentales y del Pacífico Americano o Panameña, Todas son relativamente diferentes en espe- cies y tienen por 10 menos algunos géneros característicos, pero hay similitudes muy amplias que unen a las cuatro más íntimamente, que como lo está cada una por separado de las faunas costeras de las zonas templadas, ubicadas inmediata- mente al norte y al sur.
La fauna de la región Indo-Pacífica es la más rica y contiene esencialmente a todas las familias y a un número considerable de los géneros que se encuentran en las otras tres regiones. También agrupa familias y géneros que no se hallan en ningún otro lado. La configuración compleja de las costas continentales y las numerosas islas ha proporcionado una gran oportunidad para que se realice la evolución al ofrecer una gran diversidad de hábitats. Esta fauna cubre una vasta extensión, distribuyéndose desde el Africa Oriental y el Mar Rojo hasta el norte de Australia, sur del Japón, las islas Hawaianas, y las demás islas de la Polinesia; en total, casi la mitad alrededor del mundo. Las otras tres regiones son, en efecto, segregaciones geográficas empobrecidas de esta gran fauna ictiológica mundial. La distribución de los arrecifes de coral o de los peces loro (Scaridae) deben ser considerados para delimitar las fronteras de la fauna de la región Indo-Pacífica. Las islas hawaianas comprenden el elemento más característico de la fauna ya que, aunque solamente hay tres géneros endémicos conocidos, más del 30% de las especies coralinas es endémico. Una característica notable de esta fauna de aguas marinas interiores consiste en la ausencia virtual de meros (subfamilia Epi- nephelinae, familia Serranidae) y de pargos (Lutjanidae); estos dos importantes grupos de peces son dominantes en los arrecifes de cualquier parte del océano Pacífico tropical.
La fauna tropical del África Occidental es comparativamente reducida y muestra relaciones con los peces tropicales del Mediterráneo y también con los de las Indias Occidentales.
La fauna de las Indias Occidentales o del Caribe, distribuida desde el sur de Florida y la costa de la porción continental de Centroamérica, al sur de Yuca- tán y hacia el sur hasta Brasil (Bahía), es muy rica, conteniendo cerca de los dos tercios de las verdaderas familias de las costas tropicales, que habitan en la región Indo-Pacífica. La mayoría de las especies de las dos regiones son diferentes. Algu- nos elementos del norte, en la porción septentrional del Golfo de México, repre- sentan relictos glaciales que fueron atrapados después del retroceso de los glacia- res continentales. Que la Corriente del Golfo también forma una barrera, es pro- bado por la restricción de ciertos peces al Golfo y al Caribe, respectivamente.
La fauna del Pacífico Americano o Panameña se extiende desde el Golfo de California hasta el Perú; en la porción septentrional del Golfo las condiciones han conducido a la aparición de un endemismo limitado y a algún entrampamiento de elementos de la fauna de las regiones frías del norte. Los peces pertenecientes a esta fauna tienen muchas características propias del Nuevo Mundo y también afinidades correspondientes, bien separadas de las de la fauna de la gran región Indo-Pacífica como consecuencia de la existencia de una gran extensión de agua oceánica profunda, la barrera del Pacífico Oriental. Ellos son esencialmente de las Indias Occidentales, con numerosas especies comunes a los dos lados del Istmo de Panamá, el cual estuvo bajo el mar a fines del Terciario. La distinción que hay entre las faunas del este del Pacífico y la región Indo-Pacífica ha quedado redu-

GEOGRAFfA DE LOS PECES MARINOS 445
cida por las invasiones muy recientes (hablando geológicamente) de las formas de la región Indo-Pacífica, a lo largo de las contracorrientes.
Hay cuatro barreras principales que evitan la dispersión de los peces de las costas tropicales:
a. La del Pacífico Este, que no es cruzada por el 86% de las especies. Unas cincuenta y seis especies son transpacíficas, apareciendo en ambos lados de esta gran barrera; pero trece de estas especies tienen distribución mundial.
b. La porción continental del Nuevo Mundo, cuya importancia como ba- rrera faunística parece incrementarse a medida que son comparados más grupos del este del Pacífico y del oeste del Atlántico; esto es, parecen existir menos especies en común en ambos lados de lo que se había pen- sado, de tal manera que la barrera (al nivel de especie) es efectiva para cerca de un 98%.
c. El Atlántico Medio, que es cruzado por cerca de ciento cuarenta especies tropicales y muestra una efectividad estimativa como barrera de algo así como el 72%.
d. La masa continental del Viejo Mundo, conectando a Eurasia y Africa, eliminó al antiguo Mar de Tetis. El corte del Canal de Suez está permi- tiendo la entrada de peces del Mar Rojo hacia el Mediterráneo (siete especies registradas en 1976), y algunas de éste (cinco hasta la fecha)* al Mar Rojo.
Puede observarse, en consecuencia, que la barrera del Pacífico del Este es solamente un impedimento, y posiblemente no el más importante, para la mezcla de los peces de las costas tropicales.
Las faunas de las zonas templada y boreal quedan ubicadas hacia el norte y el sur de las correspondientes zonas tropicales y se caracterizan por contar con una diversidad mucho menor y menor número de especies si se les compara con la fauna tropical, y a menudo por el número espectacular de individuos, como ocurre con los bacalaos (Gadidae), sardinas y arenques (Clupeidae) y lenguados (Pleuronectiformes). En estas regiones están situadas las pesquerías comerciales más importantes del mundo. Sin embargo, las complicaciones muy numerosas debidas a las corrientes oceánicas, las temperaturas y los cambios estacionales, obligan a que la separación entre las regiones templada y boreal sea extremada- mente arbitraria.
La región norte del Pacífico es por esto definida como el área del océano situada entre el Asia Oriental y la América, por su lado poniente, hacia el norte de los trópicos, California y Japón forman zonas de transición, pero la situación es complicada por la corriente fría de California y la cálida de Kuroshio. Por lo gene- ral, sin embargo, la fauna de agua fría se extiende muy lejos hacia el sur, más bien a lo largo de la costa americana que de la asiática. Esta extensión hacia el sur de los elementos boreales es inducida más tarde, hacia el norte de Baja California, mediante la emergencia de aguas frías y profundas a lo largo de la costa, de tal manera que las especies que no habitan en California, más al sur de Punta Con-
*Aunque más recientemente se hayan introducido más especies del Mediterráneo al Mar
(N. del T.). Rojo, habría que esperar todavía muchos años para ver que pudieran lograr su adaptación.

446 ECOLOGfA Y ZOOGEOGRAFfA
cepción, reaparecen en Baja California, después de una solución de continuidad en su área de distribución, de casi 483 km. La fauna del norte del Pacífico está representada por una gran diversidad de rocotes (Scorpaenidae), “sculpins” (Cotti- dae), peces caracol (Cyclopteridae) e intrusos (Agonidae), y muchas familias endémicas, incluyendo a las percas de las rompientes, vivíparas (Embiotocidae) y los verdosos (Hexagrammidae). Hay numerosos géneros endémicos. Ella posee la fauna más rica, exceptuando la de la región Indo-Pacífica. Esto puede ser re- sultado de la extensión y variedad del hábitat, las conexiones primitivas entre los elementos americanos y asiáticos, los contactos del pasado entre el elemento occidental y el Indo-Pacífico, y la supervivencia de la fauna de agua fría durante los periodos interglaciales. La fauna del norte del Pacífico muestra solamente un débil parecido con la fauna del Artico, con unas cuantas especies en común, mientras que las faunas del norte del Atlántico y del Artico tienen muchas simi- litudes. Esto se puede explicar por la existencia de la estrecha conexión entre el Pacífico y el Ártico, la pasada historia de los puentes continentales de Behring, que aisló frecuentemente a las dos faunas, y la relativa diferencia de las condicio- nes oceanográficas de las dos regiones.
La región templada del sur del Pacífico incluye el sur de Australia, Nueva Zelanda, porción sur de América del Sur y África del Sur. En todas estas áreas hay muchos géneros de la zona templada norte y algunas veces también aparecen especies de más al norte; las últimas son citadas como evidencias de la distribu- ción bipolar (véase el tratamiento que se le da más adelante a la bipolaridad). Las corrientes al sur de la región tropical no son desviadas por las masas continen- tales en la misma magnitud, como sucede en el norte; de aquí que las zonas de distribución sean más fáciles de definir y las isotermas sean más paralelas. La fauna de Australia-Nueva Zelanda contiene cierto número de géneros endémicos y unas cuantas familias peculiares, pero las relaciones con las formas chilenas y antkticas están claramente cortadas, especialmente en Nueva Zelanda.
La fría región Antártica está limitada hacia el norte por la isoterma de 6°C. Sus aguas son capaces de producir grandes poblaciones, en términos de nú- mero de individuos, durante los meses del verano. La fauna es limitada pero peculiar, constituida en su mayor parte por “blennies” del Antártico (Notothe- niidae) y sus parientes. La escasez relativa de la fauna refleja la severidad del medio ambiente (como ocurre con los peces de la zona béntica de los mares pro- fundos), con temperaturas bajas, reducida productividad de las aguas, influencia del hielo en la región litoral y una virtual ausencia de la macroflora, dada la exis- tencia de factores limitantes.
En el Atlántico del Norte las variaciones estacionales de la temperatura del mar son muy marcadas; las grandes corrientes oceánicas están muy extendidas y son causa de que las isotermas estén muy separadas. El encuentro de la Corriente del Labrador, fría, con la Corriente del Golfo, cálida, produce un cambio abrup- to, desde las condiciones frías a las subtropicales a lo largo de la costa de América del Norte, con un correspondiente cambio en la fauna ictiológica. Por ejemplo, la isoterma de 6°C corre desde cerca de los 45” de latitud Norte en nuestra costa hasta cerca de los 68” de latitud Norte en la costa de Noruega, dando entonces una diferencia de latitud de más de 1,200 millas náuticas. El límite sur del Atlán- tico Norte templado queda señalado por la isoterma de 12”C, la cual corre des- de la boca del Canal Inglés, hacia el oeste y el sur hasta Cabo Hatteras. Aquí ocurre una transición de los tipos septentrionales típicos, como los arenques

BIPOLARIDAD 447
(Clupeidae), bacalaos (Gadidae) y lenguados (Pleuronectiformes) a las formas de aguas cálidas como las sardinas (SardineZla) y la anchoa (Anchoa). Muchas especies mediterráneas alcanzan a tener su límite septentrional casi en esta latitud, la cual también señala aproximadamente el límite meridional, en calidad de especies marinas, de esos grupos que tienen atracción por las aguas frías, como la familia de los salmones (Salmonidae). La fauna tiene muchos aspectos en común en am- bos lados del Atlántico del Norte.
En el Pacífico Sur la fauna chilena tiene unas cuantas formas endémicas, un elemento australiano notable (por ejemplo, entre las lampreas, Petromyzoni- dae, y los galáxidos, Galaxiidae), y una mezcla de formas tropicales hacia el Norte, y de la Patagonia hacia el sur, además de esos grupos que son comunes a todas las faunas de la zona templada del sur.
La fauna Ártica, situada hacia el norte de la isoterma de los 6”C, está muy empobrecida por las mismas razones por las cuales la fauna Antártica también lo está (véase arriba). Los grupos importantes están representados por los cótidos (Cottidae), bacalaos (Gadidae), lenguados (Pleuronectiformes) y anguilas ceñu- das (Zoarcidae), estando también representado el último grupo en el Antártico, pero con géneros característicos en cada región.
BIPOLARIDAD
Se dice que las especies que habitan en los hemisferios Norte y Sur, pero con una amplia solución de continuidad en su área de distribución separando a éstas, muestran el fenómeno de bipolaridad. Por ejemplo, el pez “angler” de los mares profundos, Ceratias holboelli, ha sido capturado en Groelandia y también en el Antártico, pero no en las aguas tropicales intermedias. Ese tipo de submergencia también ocurre a lo largo de la costa del África Occidental. Cuando las tempera- turas de las aguas marinas eran muy bajas en el Pleistoceno, las formas de super- ficie, según se presume, tuvieron la oportunidad de pasar sobre la barrera. Estas dos regiones, sin embargo, todavía están insuficientemente investigadas, y las espe- cies de las que se piensa que son bipolares pueden, de hecho, habitar en una u otra (o en ambas) de las regiones intermedias. La bipolaridad es algo más elevada entre los géneros y las categorías taxonómicas que se encuentran sobre éstos, que entre las especies; por ejemplo, la familia de las lampreas (Petromyzonidae) exhibe distribución bipolar. La emergencia de agua fría al este del Pacífico ha favorecido el paso, a través de los trópicos, de las formas septentrionales y me- ridionales.
REFERENCIAS ESPECIALES SOBRE ECOLOGfA Y ZOOGEOGRAFIA
A . Ecología de agua dulce
Carpenter, K.E. 1928. Life in inland waters. Sidgwick and Jackson, Londres. Chapman, R.N. 1931. Animal ecology. McGraw-Hill Book Company, Nueva York. Coker, R.E. 1954. Streams, lakes, ponds. University of North Carolina Press, Chapel Hill, N.C.

448 ECOLOGfA Y ZOOGEOGRAFfA
Hutchinson, G.E. 1957. A treatise on limnology. vol. 1: Geography, physics, and chemistry.
Needham, J.G., y J.T. Lloyd. 1930. The life o f inland waters. The Comstock Publishing Com-
Reid, G.K., y R.D. Wood. 1976. Ecology o f inland waters and estuaries. D. van Nostrand
Ruttner, F. 1953. Fundamentals of limnology. University of Toronto Press, Toronto. (Tradu-
Ward, H.B., y G.C. Whipple. 1918. Fresh-water biology. John Wiley and Sons, Nueva York.
Welch, P.S. 1951. Limnology. McGraw-Hill Book Company, Nueva York.
John Wjley and Sons, Nueva York.
pany, Ithaca, N.Y.
Company, Nueva York.
cido por Frey y Fry.)
Segunda edición, 1959, editada por W.T. Edmondson.
H . Ecología marina
Coker, R.E. 1954. This great and wide sea. University of North Carolina Press, Chapel Hill, N.C. Colman, J.S. 1950. The sea and its mysteries. G. Bell and Sons, Londres. Halstead, Bruce, W. 1959. Dangerous marine animals. Cornel1 Maritime Press, Cambridge, Md. Harvey, H.W. 1928. Biological chemistry and physics of the sea. Cambridge University Press,
Hedgpeth, J.W. (editor y coautor). 1957. Treatise on marine ecology and paleoecology. Mem.
Idyll, C.P. 1964. Abyss: the deep sea and the creatures that live in it. Thomas Y. Crowell Com-
Jonstone, J. 1928. A n introduction to oceanography. Hodder and Stoughten, Londres. Kuenen, P.H. 1955. Realms o f water. John Wiley and Sons, Nueva York. MacGinity, G.E., y N. MacGinity. 1949. Natural history o f marine animals. McGraw-Hill Book
Moore, Hilary B. 1958. Marine ecology. John Wiley and Sons, Nueva York. Nicol, J.A.C. 1967. The biology of marine animals. Interscience Publishers, Nueva York. Pettersson, H. 1954. The oceun floor. Yale University Press, Nueva Haven, Conn. Romanovsky, V. 1953. La mer. Librairie Larousse, París. Russell, F.S. 1928. The seas. F. Warne and Company, Londres. Sears, Mary. (Ed.). 1961. Oceanography. Publ. 67, Amer. Assoc. Adv. Sci., Washington, D.C. Smith, F.G.W. 1954. The ocean. Charles Scribner’s Sons, Nueva York. Sverdrup, H.U., M.W. Johnson, y R.H. Fleming. 1942. The oceans. Prptice-Hall, Englewood
Sverdrup, H.U. 1942. Oceanography for meteorologists. Prentice-Hall, Englewood Cliffs, N.J. Tait, J.B. 1952. Hydrography in relation to fisheries. E. Arnold, Londres.
Cambridge.
Geol. Soc. Amer., 67.
pany, Nueva York.
Company, Nueva York.
Cliffs, N.J.
C. Geografía ictiológica
Barbour, C.D., y J.H. Brown. 1974. Fish species diversity in lakes. Amer. Nut., 108:473-489. Briggs, J.S. 1974. Marine zoogeography. McGraw-Hill Book Company, Nueva York. Darlington, P.J., Jr. 1957. Zoogeography: The geographical distribution of animals. John Wiley
de Beaufort, L.F. 1951. Zoogeography of the land and inland waters. Sidgwick and Jackson,
Ekman, S. 1953. Zoogeography o f the sea. Sidgwick and Jackson, Londres. Gosline, W.A. 1975. A reexamination of the similarities between the freshwater fishes of Africa
and South America. Mém. Mus. Nat. D’Hist. Nut., ns., Ser. A , , Zool., 88:146-155. Gosline, W.A., y V.E. Brock. 1960. Handbook o f Hawaiian fishes. University of Hawaii Press,
Honolulu. Grey, M. 1956. The distribution of fishes found beIow a depth of 2000 meters. Chicago Nat.
Hist. Mus. Fieldiana: Zool., 36( 2):75-337. Hubbs, C.L., y R.R. Miller. 1948. Correlation between fish distribution and hydrographic
history in the desert basins of western United States: En: The Great Basin, with emphasis on glacial and postglacial times. Bull. Univ. Utah, 38:17-166.
and Sons, Nueva York.
Londres.
MacArthur, R.H. 1972. Geographical ecology. Harper and Row, Nueva York.

REFERENCIAS 449
MacArthur, R.H., y E.O. Wilson. 1967. The theory of island biogeography. Princeton Univer-
Marshall, N.B. 1954. Aspects o f deep sea biology. Philosophical Library, Nueva York. Miller, R.R. 1959. Origin and affinities of the freshwater fish fauna of western North America.
En: Zoogeography. Amer. Assoc. Adv. Sci., Publ. 51(1958):187-222. Myers, G.S. 1938. Fresh-water fishes and West Indian zoogeography. Ann. Rept. Smithsonian
Inst., 1937:339-364. -. 1940. The fish fauna of the Pacific Ocean, with especial reference to zoogeographical
regions and distribution as they affect the international aspects of the fisheries. Sixth Pacific Sei. Cong., 3:201-210.
Bijd. Tot Dierk., 28:315-322.
sity Press, Princeton, N.J.
- . 1949. Salt-tolerance of fresh-water fish groups in relation to zoogeographic problems.
-. 1951. Fresh-water fishes and East Indian zoogeography. Stanford Ichthyol. Bull., 4 : l l - 2 1 . Nelson, G.J. 1969. The problem of historical biogeography. Syst. Zool., 18(3):243-246). Scientific American, Inc. 1972. Continents adrift. Readings from Scientific American, with
introductions by J. Tuzo Wilson. W.H. Freeman and Co., San Francisco. Scientific American, Inc. 1974. Planet Earth. Readings from Scientific American, with intro-
ductions by Frank Press and Raymond Siever. W.H. Freeman and Co., San Francisco. Seyfert, C.K., y -L.A. Sirkin. 1973. Earth history and plate tectonics. An introduction to his-
torical geology. Harper and Row, Nueva York. Sterba, G. 1959. Siisswasserfische aus aller Weit. Verlag Zimmer und Herzog, Berchtesgaden. Vuilleumier, F. 1974. Zoogeography. vol. 4.


fndice Sistemático
Abramis, 176 Abramis brama, 134, 358 Acanthias vulgaris, 293 Acanthodii, 18, 20, 24, 26 Acanthopteri, 30 Acanthopterygii, 26, 29, 30,106,147,172,
Acanthuridae, 46, 111, 123, 150, 153 Acanthuroidei, 46 Acanthurus, 109,123, 126,329,420
triostegus, 285 Acantodios, 15, 18, 19, 20, 23, 27 Acantopterigios, 55 Acipenser, 59, 80, 111, 134, 139, 195, 199,
173,233, 238,243
211, 273, 281, 288, 297, 341, 343, 344,347,425
oxyrhynchus, 33 stellatus, 267
Acipenseridae, 27, 32, 104, 108, 109, 131,
Acipenseriformes, 32, 90, 91, 194 Acrania, 11 Acropomatidae, 122 Actinopteri, 12 Actinopterigios, 14, 26, 27 Actinopterygii, 11, 14, 17, 24, 26, 27, 28,
132,168,172,331,441
29, 32, 57,68, 94, 201, 207, 208, 211,220,233,251,257,259,288, 307,311, 314,317,324,331, 336, 337,342,352,358,361,362
271, 273, 275, 283, 597, 328, 373, 374,375,378,381
Acuario, pececillos de, 160, 238, 265, 267,
Adherentes, peces, 43,47, 54, 55, 419 Adrianictidos, 40 Adrianichthyidae, 40 Adrianichthyoidei, 40 Aetobatis narinari, 123 Ageneiosidae, 38 Ageneiósidos, 38
Agnatha, 13, 15, 18, 21, 24, 30, 211, 362 Agnatos, 70, 77, 173 Agonidae, 47, 446 Agua mala, 41 1 Agujitas, 40, 120, 135, 168, 283, 311 Aholeholes, 42 Akysidae, 38, 224 Albula, 419
vulpes, 224 Albulidae, 34, 150, 296, 420 Albuloidei, 34 Aldrovandia macrochir, 37 Alepisauridae, 38 Alepisaurus, 136 Alepocephalidae, 36 Alewife, 281, 350, 424 Alopiidae, 32 Alosa, 66
Alutera schoepfi, 120 Ambloplites, 189, 281 Amblicipítidos, 38 Amblycipitidae, 38 Amblyopsidae, 47, 57, 95, 111, 280, 360,
Amblyopsis, 119 Amblyopsoidei, 47 Amia, 27,80,90,121,126,138,139,144,
pseudoharengus, 281, 350, 424
43 8
170,174,178,206,215, 227,229, 234,251,281,284,288,296,311, 346,367
calva, 33, 179, 195, 197, 212, 223, 234 Amiidae, 27, 35, 110, 172, 286, 295 Amiidos, 28 Amiiformes, 34 Ammocetes, 22, 295 Ammodytes, 127, 261 Ammodytidae, 46, 185 Ammodytoidei, 46 Amphiliidae, 38
451

462 ÍNDICE S I S T E M ~ C O
Amphiliidos, 38 Amphioxus, 11, 169, 288 Amphipnopidae, 47 Amphipnops cuchia, 41 Amphipnon, 411 Anabantidae, 46, 230, 262, 441 Anabantoidei, 46 Anabas, 95,185, 231 Anablepidae, 40, 283, 349 Anarhichadidae, 46 Anáspidos, 18 Anchoa, 447 Anchoa, 34,146,150, 269,447 Anchoas, 42,129,347,417 Anfioxus, 11, 378,443 Angelitos, 47, 185, 188, 236, 241, 320, 323,
346,419,420
Anguila viscosa, 102 Anguilas, 51, 52, 55, 85, 109, 118, 171, 172,
voladores, 47, 54, 186
182,186,187,197,209, 221, 237, 239, 253, 255,256,257,258,261, 273, 295,296,297,317,320,337, 348,350,351,357,359,417,418, 419.419.420.442
agua dÚlce, de, 34, 103, 154, 158, 170, 190.207
aguas profundas, de, 420 americana, 37 congrio, 34,347 congrio lucio, 34 cuello largo, de, 34 culebra, 34, 175, 185 dientes de flecha, 34 eléctrica, 177, 365 espinosa, 34, 230 espinosa de mares profundos, 37 europea, 265 gimnótida, 36, 171, 179, 365 gusano, 184 holosáurida, 34, 37 japonesa, 363,368 limo, de, 15, 175 mares profundos, de, 132 mastacembélida, 43, 47 morena, 143,343,345 nariz chata, de, 34 pantano, del, 47, 143 pequeña, 34, 51, 52 pico de pato, 34 simbránquida, 170
Anguilas ceñudas, 44,283,447 Anguilas de los pantanos, 154,170,194 Anguilas lengua. 44.419 Anguilla, 103,109,.118, 119,154, 158,
159.186.190.197.207.209.226. 239; 253; 255; 256; 258; 260; 273; 275,282,295,297,321,348,352, 357,358,359,417,420,442
anguilla, 265, 337 japonica, 337, 368 rostrata, 37, 188
Anguillidae, 34, 51, 55, 170, 171,172, 209,237,295,348,420
Anguilliformes, 34, 51,138, 170, 220, 221,296,317,347,419
Anguilloidei, 34 Anomalopidae, 42, 122 Anomalópidos, 122 Anoplopomatidae, 47 Anoptichthys, 57,95, 119 Antennariidae, 48, 89, 130, 185 Antennarioidei, 48 Antiarcos, 25 Antiarchi, 23 Antigoniidae, 42 Antigóniidos, 42 Aphredoderidae, 48,437,438 Aphredoderoidei, 48 Aphredoderus, 56 Aphyonus gelatinosus, 221 Aplodinotus grunniens, 173 Apogonidae, 42, 111,122, 283, 411, 442 Aracanidae, 47 Arapaima gigas, 233 Archoplites, 282 Archosargus, 136 Areneros, 44, 185 Argentinos, 36 Argentinidae, 36, 245 Argentinoidei, 36 Arges, 224 Argyropelecus, 311,418 Ariidae, 38, 283, 437, 442 Arius, 311 Arthrodira, 23 Artrodiro, 19 Artrodiros, 20, 23 Aspredinidae, 38 Astraspis desiderata, 13 Astronesthes, 240 Astronesthidae, 36 Astrónomos, 44, 121, 124, 135
arena, de la, 44 Astroscopus, 94 Astyanax, 332,338
Ateleopidae, 38 Ateleópidos, de los mares profundos, 35, 38 Ateleopoidei, 38 Ateleopus japonicus, 35 Atherinidae, 42, 57, 424, 442 Atherinoidei, 42 Atunes, 46, 52,147,149,159,176,188,
189,194,195,209,213,410,417, 442
mexicanus, 338
aleta azul, de, 213, 340 Aulorhynchidae, 40 Aulorhynchus, 55 Aulostomidae, 40, 134 Aulostomoidei, 40 Ayu, 36
Bacalao, 70, 110, 122, 147, 159,170, 172, 199, 200, 206, 208, 235, 243, 262, 268,277,299,320,331, 343, 358, 420,445,447
Atlántico, 37

ÍNDICE SISTEMÁTICO 453
Bacalao, familia del, 103 Bacalaos, 48, 195 Badidae, 44 Bádidos, 44 Bagre marinus, 123, 283 Bagres cabeza de toro, 38,123, 130,143,
278, 281, 282, 343, 344, 358, 362, 368,414
amarillo, 281 café, 174
Bagridae, 38 Bairdiella, 148 Balanoglossus, 11 Balistes, 236 Balistidae, 47, 111, 179, 420 Balistoidei, 47 Barbatula, 383 Barbo, 137 Barbo europeo, 317 Barbourísidos, 40 Barbourisiidae, 40 Barbudos (ratón), 42, 275 Barbus, 137
Barracudas, 42, 126, 129, 138, 139, 150,
Barrenderos, 44 Bassogigas, 420 Bathyclupea argentea, 35 Bathyclupeidae, 44 Bathydraconidae, 46 Bathygobius, 313
soporator, 325 Bathylaconidae, 36 Bathylagidae, 36 Bathylagus wesethi, 378 Bathymasteridae, 44 Bathypterois, 420 Batilacónidos, 36 Batoidei, 32 Batrachoidei, 48 Batrachoides, 123 Batrachoididae, 48, 89, 122, 123, 185, 236 Belone, 136 Belonidae, 40, 120, 169, 296, 417 Beloniformes, 40 Benthobatk, 95
moresbyi, 122 Berícidos, 41, 42 Bericoideos, 30, 242 Bermejuela, 38, 269, 272, 274, 277, 282, 366 Berrugato, 420 Beryciformes, 29, 30, 40 Berycoidei, 42 Berycidae, 42 Beryx splendens, 41 Betta, 282, 326
splendens, 282 “Billfishes”, 46, 296 Blenniidae, 46, 419, 420 Blennioidei, 46 Blennius, 210 “Boarfishes”, 42 Boca belicosa, 281 Bocachica, 189 Bonitos, 188, 417
fluviatilis, 317
188,416,417
Borophryne, 418 Bothidae, 46,419 Brachiopterygii, 12, 24 Bradyodonti, 12 Bramidae, 44 Branchiostegidae, 42 Branchiostoma, 11,169, 286
belcheri, 378 Branchiostomidae, 443 Brema europea, 134,176,359
marina, 136 Brevoortia, 148, 189, 221
tyrannus, 130, 154 Brillador esmeralda, 424
dorado, 176,180,281,302 Brotulas, 44, 283, 294,420 Brotulidae, 44, 283, 294,420 Brotúlido, 421 “Bullrouts”, 123 “Burbot”, 103, 105, 147, 368, 425
Caballitos de mar, 40,103, 109, 139, 170, 171,175,177,179,239,279,282, 352,419,420
Calamoichthys, 80 Callichthyidae, 38 Callionymidae, 48, 124 Callionymoidei, 46 Callionymus, 124, 261 Campostoma, 55
anomalum, 138 Candiru, 412 Canthigasteridae, 47 Caproidae, 42 Caracinos, 36. 106, 118, 332, 341, 386,
427,437,438,441 ciegos cavernícolas, 57, 95 voladores, 54
Carangidae, 42,150,349 Carángidos, 316 Caranx, 411 Carapidae, 44 Carapus, 296, 411 Carassius, 112, 173, 188, 253, 255, 333,
3 54
316,323,336,358,373,376 aumtus, 165, 248, 254, 281, 310, 313,
Carcharhinidae, 32, 149, 176, 195, 283, 293 carassius, 313
Carcharhinus, 283 leucas, 31, 324
Carcharias, 314 Carcharodon, 143 Careproctus, 273, 282 Carpa, 37,38,66, 73, 105, 131,136, 138,
146,151,157,172,173,186,189, 199,209,210,212,227,228,254, 255,266, 281,295,307,317,320, 324,341,346,373,427
comiin, 103,138,154 Crucian, 313 pasto, Carpa de, 126, 149 Véase también Pececillos de acuario
Carpa chupadora de aleta elevada, 316

c;>,vp: <ir,r:i(ya, !:t i ,, ;hC ; " . I;$&, .:ay" 2:8:13 ? 5 4 , !.'fs:i, 28;, %Ií/. 313, :< .j ;J " 4 '?. ;; , 3 :: i: ~ :; 6 , 3 5 3 , ;?, 7 3 7 F
CL'rpii:? ?llscic.onas;, C.? cnhpzs 8 m ~ t : ~ , ¿:e, (25
pla.ieada., 181 d o r a 6 : i v !ST., I37
C;t!-I,-iode? L'elifcr, 316 Gatostmnidae, 3 8 , 55, 64, 66, 131, 132,
136, i l2 , , 189, 211, 295, 379, 425, 427.332, 435,438
C l c ~ l o s t í ~ m ~ s , 227, 275, 276 uutostamus, 432, 435 cornmersoni, 171, 271
Cacdhúridos, 47 Cavadures de la arena, 44 Cavilats, 47, 57, 103, 171, 177, 179, 249,
256, 2 8 l , 326, 331, 358,419, 425, 446,447
cuernos con, 249 marinos del norte (charrasco, cuervo
marino), 190 Cefalaspidomori'os, 19, 22, 55, 58 Cefaláspidos, 22 Celacantos, 15, 32, 33, 237, 283 Centrarchidae, 42, 84, 165, 166, 276, 281,
284, 295, 325, 350, 379, 395, 437, 438,440
Centriscidae, 40 Centropomidae, 42 Centroscyllium, 122 Cephalaspidomorphi, 12, 14, 15, 18, 30, 56,
68 Cephalochordata, 11 Ceratias, 132
holboelli, 447 Ceratiidae, 48, 175, 295, 296, 443 Ceratioidei, 48 Ceratioideos, 418 Ceratodontidae, 3 2 Cetomimidae, 38 Cetomímidos, 38 Cetomimiformes, 38 Cetomimoidei, 38 Cetopsidae, 38 Cetópsidos, 38 Cetorhinidae, 32 Ceforhinus, 130, 149
maximus, 280 Cíclidos, 44, 84, 130, 165, 177, 235, 269
Ciclóstomos, 15, 21, 56, 77, 83, 84, 85, 91, Cichlasoma, lobochilus, 135
326 ,329 ,427 ,438 ,440
maculicauda, 399 meeki, 326
Cichlidae, 44, 84, 155, 165, 166, 329, 427,
Ciprínidos, 55, 141, 176,415, 441 Cipriniformes, 99, 437 Ciprinodóntidos, 373, 394 Ciprinoideos, 54,97, 441 Cirrhinus mrigala, 195 Cirrhitidae, 44 Cladodonto, 19, 25 Cladoselache, 19, 25, 26
428 ,438 ,440
<.;[a:.i(i:< j "7'2, 230 , rkziruchtis, 1.36
í;!m%&?&!. 3s. s59, 23(?
Cltnidae, 46, 285 . : ; i ¿ , m t i t d : : > I 9
Giinidos, 4 6 ~ 283 Clupea, 189, 234: 266, 413
horengus, 33 : 1 2 7 , 158, 159, 265, 267, 26 8
pnllasi, 316
Clupeidae, 27, 3 4 , 56, 6 6 , 83, 97, 109, 112, .sprallus, 188
141, 150, 1 7 2 , 176, 235, 231, 295,
446 Clupeiformes, 30, 34, 399 Clupeoidei, 34 Clupeoidos, 54, 130, 235, 421 Cobias, 42 Cobitidae, 38, 230, 235, 231, 243, 321,
Co bitis, 366 Coelacanthidae, 32 Coelacanthiformes, 24, 32 Cola de espada, 271, 373, 385 Cometa, 377
Condrosteanos, 58, 59, 107 Congriclae, 34, 344, 346 Condrictianos, 21, 23, 83, 91 Congrios, 207, 241, 256, 348 Coregonidae, 174 Coregonus, 110, 189, 241, 276, 281, 311,
296 ,358 ,417 , 424 ,441 ,442 ,445 ,
343 ,383 ,441
gris, 376
328,407 artedii, 94, 161 clupeaformis, 131, 276 nigripinnis, 131
Corvina, 148 Coryphaena, 170, 188, 417, 443
hippurus, 210 Coryphaenidae, 44 Cottidae, 47, 57, 103, 171, 177, 179, 249,
Cottoidei, 47 Cottus, 257, 281, 326, 337, 358 Craniata, 11 "Crappies", 281, 424 Crenilabrus, 210 Cresta, peces coil, 40 Criador de frente cefálico, 46, 278, 279 Cromeriidae, 441 Crosopterigios, 26 Crossopterygii, 12, 14, 17, 24, 26, 32,68,
110, 238 Ctenopharyngodon idella, 126, 148 Cuchia, 41, 47 Culaea, 103, 277, 281, 282 Cyclopteridae, 47, 419, 446 Cyclopterus lumpus, 120 Cyclostomata, 15, 30, 70, 110, 173, 206,
211,237, 253, 306,311, 321,329, 3 3 0 , 3 3 7 , 3 4 4 , 3 5 6 , 3 6 2 , 3 7 3
256 ,419 ,425 ,446 ,447
Cyclostomi, 12 Cyclothone, 418 Cyema, 418

iNDICE SISTEMÁTICO 455
Cynoglossidae, 47 Cynoscion, 417
nebulosus, 237 Cyprinidae, 27, 38, 64, 66, 97, 98, 110,
131,134, 136, 138,146,155,163, 172, 189, 199, 203, 235, 236, 237, 254, 271, 295,310, 320, 321, 341, 343, 354, 356, 359, 373, 379, 380, 386,394,412,423,426,427,428, 432,436,438,441
Cypriniformes, 36, 98, 111, 235, 316, 361, 437,441
Cyprinini, 399 Cyprinodon, 428 Cyprinodontidae, 40, 106, 171, 172, 242,
Cyprinodontidos, 373, 394 Cyprinodontiformes, 40, 54, 168, 283, 373,
421,437 Cyprinodontoidei, 40 Cyprinoidei, 36 Cyprinus, 172, 186, 229, 253, 254, 255,
291,394,438,439
266,307,427 carpio, 39,73,103,138,151,154,155,
163, 173, 199, 208, 210, 212, 227, 267,281,317,320,324,346
Cypselurus, 120, 186
Chabelitas, 44, 120 Atlántico, del (Chabela), 120
176,350,410,420 Chacidae, 38 Chácidos, 38 Chaenichthyidae, 85 - Chaenobryttus, 281 Chaetodipterus faber, 120 Chaetodontidae, 44, 52, 111, 130, 134,
Chanidae, 38 Chnnna, 171, 230, 231
striata, 41 Channichthyidae, 46 Channidae, 46 Chanoidei, 36 Chanos, 190, 258 Chaqueta de cuero, 120 Characidae, 36, 106, 118, 341, 386, 427,
438 Characoidei, 36 Chaudhuriidae, 47, 443 Chaudhurioidei, 47 Chauliodontidae, 36 Chauliodus, 134, 244, 418
sloani, 13 7 Cheirolepis, 27 Chiasmodon, 244 Chiasmodontidae, 44 Chiosmodus, 418 Chimaeridae, 30, 123, 212, 272 Chimaeriformes, 30 Chiros, 36,190, 258 Chlamydoselachidae, 32 Chondrichthyes, 12, 14, 15, 16, 20, 24, 25,
94,98,311,348,366
30 ,55 ,59 ,67 ,70 ,78 ,83 ,85 ,89 , go, 94,96,97, 107, 141,173, 211, 288, 296,306,317,320,365
365,437 Chondrostei, 26, 27, 28, 67, 207, 219, 347,
Chopas, 44 Chordata, 11 Chupadoras, carpas, 38, 55, 64, 66, 69, 131,
132, 136, 172, 189, 271, 274, 276, 281, 295, 361, 379, 415, 425, 427, 432, 435, 438
blanca, 171, 271 China de, 432 nariz larga, de, 432 “redhorse”, 83 Véase también Carpas hociconas
Dactylopteridae, 47, 54, 1% Dactylopteriformes, 47 Dactyloscopidae, 44 Dasyatidae, 32, 103, 109, 123, 419, 420 Dasyatis, 123 “Dealfish”, 296 Denticipitidae, 34 Denticipitoidei, 34 Derichthyidae, 34 Diablos de mar, 48 Diodon hystrix, 109 Diodontidae, 47, 139, 149, 177, 420 Diplomystidae, 38 Diploprion, 296 Diplorhina, 18 Dipneusti, 12 Dipnoi, 12, 14, 17, 24, 26, 32, 57, 68, 86,
90, 91, 94, 107, 110, 154, 174, 193, 195,197,198,199, 211, 232,347, 353,365,437
Dipnoos, 26 Dipteriformes, 32 Diretmidae, 42 Dirétmidos, 42 “Dolly Varden”, 385 Doradidae, 38 Dorados o doradillas, 44,170, 188, 270,
417,443 Doratonotus megalepis, 120 “Dories”, 42, 134, 418
Dorosoma, 66,126,130,139,170 Dormidores, 46
cepedianum, 148,186 Dragones de mar, 111,117,120
filamentoso, 53, 177 “Dragonets”, 46, 114, 261 Dussumieriidae, 34 Dysommidae, 34
Echelidae, 184 Echeneidae, 44,170,175,411 Echeneioidei, 34 Echeneis, 209
Elasmobranquio, 26, 59, 83, 122, 306 naucrates, 43

456 ÍNDICE SISTEMÁTICO
Elasmobranchii, 12, 16, 24, 32, 56, 57, 64, 66, 78, 88.103, 129, 147, 160, 194, 195,197,198,199,201, 206, 208, 217,221,237,249,259, 271, 306, 311, 314, 316, 317,320, 323, 329, 330,336. 337,339,342,344, 347, 348,352,358,360,363,366
Elasmobranchiomorphi, 15, 23 Electrophorus, 179, 365 Eleotridae, 46,442 Elopidae, 34 Elopiformes, 29, 34 Elopoidei, 34 Elops, 84 Embiotocidae, 44, 271, 283, 296, 420, 446 Engraulidae, 34, 150, 269 Engullidores, 34, 55, 136, 139, 153 Eperlanos, .36, 172, 271, 442
americano, 269 mares profundos, de, 36 sur, del, 36
Ephippidae, 44, 120 Epinephelinae, 399, 444 Epinephelus, 129, 417
clarkae, 399 guttatus, 144, 540 striatus, 116
Eptatretus, 77 Equetus lanceolatus, 116 Erythrinus, 80 Escómbridos, 179 Esocidae, 36, 66, 172, 350, 379, 386,432,
Esocoidei, 36 Esox, 126,129,136,139,143, 188,207,
lucius, 57,136,140,141,142,146,
433,438,441
355,366,437
153,188,281,298,328 Espinas en el dorso, peces con, 46 Espinochas, 40,53,103,121,139,143,170
171,175,187,238,255,259, 267, 276,278,281,282,284,337,359
tres espinas, de, 39,103, 191,302, 311, 326,327,385
Esquilbeidos, 159 Esquindlhridos, 46 Estefanoberícido, 41 Estomiatoideos, 122,418 Esturiones, 27,32,58,80,90,91,103,104,
108, 109, 112,131, 132, 139, 168, 172,194,195,201,273, 281, 288, 297,331, 341,343,344,425,441
Atlántico, del, 33
blenniodes, 385 caeruleum, 281 flabellare, 27, 281 jordani, 399 microperca, 1, 171, 273 nigrum, 27, 277, 281, 385
Etheostoma, 263
Etheostomatinae, 83, 111, 121, 171, 177,
Etmopterus, 123 Etroplus, 235 Eupomacentrus, 329
244,379,384,386,427,438
Euriptéridos, 22 Eurypharyngidae, 34, 55,139,153 Eurypharynx, 418 Euselachii, 12 Eutaeniophoridae, 40 Euteniofóridos, 40 Eviota, 1 Exocoetidae, 40, 54, 170, 171, 186, 188,
Exocoetoidei, 40
Fistulariidae, 40, 134 Folidofóridos, 20 Forcipiger, 169
Fotíctidos, 36 Fractolémidos, 36 Fundulini, 399 Fundulus, 119, 221, 255, 257, 258, 261,
heteroclitus, 152, 228, 287, 289, 291,
majalis, 39 parvipinnis, 228
195,244, 269,417
longirostris, 135
333,378,399
299,334,337,339
Gadidae, 48, 70, 103, 110, 122, 147, 159,
Gadiformes, 48, 195, 243 Gadoidei, 48
172,235,320,343,420,445,447
Gadopsidae, 44 Gadus, 170, 199, 200, 203, 262, 267, 269,
331.358 callarias, 208 morhua, 37,199, 299
Galaxias brevipinnis, 35 Galáxidos, 36 Galaxiidae, 36 Galaxioidei, 36 Galeichthys felis, 123, 283 Galeocerdo cuvieri, 324 Galeorhinus, 95 Gallatheathauma, 418 Gambusia y Gambusia, 168, 170
Garibaldi, 112 Gasteropelecinae, 54, 186 Gasterosteiformes, 40 Gasterosteleidae, 40, 53,68, 103, 139, 170,
Gasterosteoidei, 40 Gasterosteus, 121, 143, 238, 255, 259, 267,
aculeatus, 39, 106, 191, 302, 311, 326,
affinis, 271, 272, 273, 283, 378
171,175, 284,326,385
276, 281, 282, 337,358
327,385,399 Gastromyzon, 54, 55 Gastromyzonidae, 38, 325, 442 Gempylidae, 46 Gemuendina, 19 Gerreidae, 44 Gibberichthyidae, 40 Gibbonsia, 420 Gigantura uorax, 35 Giganturidae, 36 Gigantdridos, de mares profundos, 35, 36 Giganturoidei, 36

ÍNDICE SISTEMÁTICO 457
Gimnárquidos, 34 Gimnótidos, 235,438 Girinoqueilidos, 38, 134 Glyptosternum, 134, 172 Gnathonemas elephas, 135 Gnathostomata, 15, 18, 21, 22, 24, 30 Gnatóstomos, 59, 78 Gobiesocidae, 46, 54, 55, 419 Gobiesociformes, 46 Gobiesocoidei, 46 Gobiesox maeandricus, 45 Gobiidae, 46, 172, 265, 296, 410, 419, 441,
Gobioidei, 46 Gobios, 46,172,187, 190,199,202,265,
442
269,296,297,313,317,410,419, 441
aleta volante, de, 425 ciegos, 312,411 enano pigmeo, 1 fiiipino, 298
Gobius, 187, 199, 317 panizzae, 202
Gonichthys, 122 Gonorrínquidos, 36 Gonorynchidae, 36 Gonorynchiformes, 36 Gonorynchoidei, 36 Gonostoma bathyphilum, 221 Gonostomatidae, 36 Gonostomátidos, 221 Goodeidae, 40, 283, 291, 437 Gordos, arroyo, 38, 138, 270, 274, 281,
282,362 cabeza córnea, de, 163, 281 río, 281
Grammicolepidae, 42 Grammicolépidos, 42 Grammistidae, 42 Granaderos, 48, 122, 236, 238,420 Grandes señores, 47, 419, 420, 446 “Graylings”, 36, 415 “Greenlings”, 47, 366, 446 Gruñón, 275
Guachinangos (pargos), 44,150,353,354, 417,444
“Gunnels”, 44 Gusanos bellota, 11 Gymnarchidae, 34, 441 Gymnarchus, 170
niloticus, 365 Gymnothorax, 143,343
vicinus, 345 Gymnotidae, 36, 171, 235, 365, 438 Gymnotus, 179 Gyrinocheilidae, 38 Gynnocheilus, 134, 224
“Haddock”, 420 Halibuts, 420 Halichoeres bivittatus, 258 Halosauridae, 34 Harpadontidae, 38 Helogeneidae, 38
California, de, 273
Helogeneidos, 3 8 Hemicyclaspis, 19 Hemichordata, 11 Hemichromis, 114 Hemiramphidae, 40,135,168, 283 Hemimmphus, 120,311 Heterodontidae, 32 Heterodontiformes, 32 Heterodontus, 197, 203
francisci, 123, 272 Heteropneustes, 123 Heteropneustidae, 38 Heterostichus rostratus, 120 Heterostracos, 12, 18, 20, 21 Heterotis niloticus, 197 Hexagrammidae, 47, 446 Hexagrammoidei, 47 Hexagrammus, 366 Hexanchidae, 32, 149 Hexanchiformes, 3 2 Hexanchus, 199
griseum, 31 Hiodon, 437 Hiodontidae, 172, 438 Hipoftálmidos, 3 8 Hippocampus, 170,175,177,238,282, 352 Hippoglossus, 420 Holacanthus bermudensis, 128 Holocéfalos, 15, 26, 66 Holocentridae, 42, 394 Holocentrus, 236 Holocephali, 12, 16, 24, 26, 32, 55,68,136 Holostei, 27, 28, 67, 70, 207, 323, 365, 437 Holósteo, 347 Homalopteridae, 38, 134, 235, 441, 442 Horaictidos, 40 Horaichthyidae, 40 Horaichthys, 280 Hybodonto, 19, 26 Hybodus, 19, 26 Hybopsis, biguttata, 281
micropogon, 281 Hydrolagus, 109, 123
colliei, 31 Hypophthalmidae, 38 Hyporhamphus unifasciatus, 135 HYPSYPOPS, 112
Icosteidae, 46 Icosteoidei, 46 Ictaluridae, 38, 102, 136, 139, 143, 170,
438 171,172, 284,294,331, 359, 425,
281, 282, 343,344, 358,362,368, 423
Ictalurus, 119, 123, 130, 143, 213, 225,
natalis, 281 nebulosus, 175 punctatus, 281
Idiacanthidae, 36 Zdiacanthus, 273, 296, 418 fdolos moriscos, 420 Inopterygii, 26 Invasores, 47, 446 Ipnops, 420

‘15% � NU ICE SISTEMÁTICO
Imnidae, 42 Isinidos, 42- isospondyli, 30 Istiophoridae, 46, 168,.171, 296 Istiophorus, 186, 188, 411, 417, 418
Jawfishes, 44 denynsiidae, 40, 283, 284, 294 John Dory, 4 1 dureles, 42, 150, 349, 411
“Killifishes”, 40, 106, 171, 172, 221, 242, 255, 257, 258,261, 291,422, 428, 438,439
California, de, 228 rayados, 39
Knéridos, 36 Kneriidae, 36, 441 Kuhliidae, 42 Kurtidae, 46, 278 Kurtoidei, 46 Kyphosidae, 44
Labeo. 131 iabidesthes, 106
Labridae. 44, 111, 134, 150, 258, 272, 317, sicculus, 186, 269, 273, 274, 281
346,352,410, 420 Lactophrys quadricornis, 109 Lachas, 130, 148, 155, 189, 221 Laeops, 296 Lamnidae, 32, 149, 194, 219, 314 Lampefra, 253, 311, 321
fluviatilis, 216, 329 lamottei, 296 tridentata, 132, 175
Lampreas, 5, 9, 11, 12, 14, 15, 16, 19, 21, 22, 24, 30, 54, 59, 60,66, 67, 70, 71, 77, 78, 84, 85,86, 91, 93, 94,97, 98, 102,110,132, 134,141,146,170, 171,173,175,181,185,190, 206, 250, 254, 257,258, 271,277, 278, 281, 286, 295, 296,306, 311, 312, 313, 315, 316, 320, 335,338, 339, 344,349,351,352,427,441,447
agua dulce, de, 250 americana, de arroyo, 296 marina, 14, 31, 61, 126, 132, 133, 141,
159,175, 190, 216, 275, 281, 294, 296, 323,346, 352,356, 357
Pacífico, del, 132, 175 río, de, 216
Lamprididae, 40 Lampridiformes, 40 Lampridoidei, 40 Latimeria, 84, 107, 237, 283, 331, 347,
348, 353 chalumnae, 14, 27, 33
Latridae, 44 Latrunculus pellucidus, 298 Lebistes, 160, 238, 267, 271, 273, 275, 283,
297,328,378,381 reticulatus, 265, 373, 374, 375
Leiognathidae, 44, 122
Lenguados, 47, 52, 57, ill, 113,152,159, 1.85, 224, 296, 337, 340: 346, 348, 350.419.420
invierno, de, 45 ojo derecho, de, 47 oio izauierdo. de. 47
- 1 I
Lep&xirkn, 80,’83,’94, 96, 194, 206, 211, 224, 288,296,438
Leuidosirenidae. 32. 295 Lepisosteidae, 26, 34, 84, 104, 107, 169,
172,386 LeDisosteiformes. 34 Lebisosteus, 27, 80, 90, 120, 126, 129, 136,
138,195,197,199,206,212, 229, 234 ,251 ,288,296,311
osseus. 33. 196. 197
Lepomis, 176,180, 213, 273, 281, 359, 380.410.424
cyanellus, 138, 325 gibbosus, 138, 144 humilis, 272 macrochirus, 128, 130, 144, 154, 166
Leporinus, 189 Leptocephali, 118 Leptocephalus, 85, 118, 159, 295 Leptolépidos, 20 Leucichthys, 110 Leucisco, 141, 254 Leuciscos, 110, 189, 281, 414
aguas someras, de, 94, 161 aleta negra, de, 131
Leuciscus, 254 rutilus, 197
Leuresthes, 276 tenuis, 273
Lisa (chiro), 34, 84 Lisas, 42, 139, 146,186, 261, 331, 336,
413.442 rasadas, 140. 316
Loaches, 38, 230, 235, 236, 243, 317, 343,345,383,441 -
arroyos montañosos, de, 38, 134, 441,
ventosa ventral, de, 38, 54, 55, 442
aguas templadas, de, 42 algas marinas grandes, de las, 417 blanca, 189
442.
Lobinas, 106, 126, 150,170, 415
hocico grande, de, 82, 164, 180, 281, 283,414
hocico’pequeiio, de, 281,414 marina, 30, 42, 84, 89, 122, 138, 144,
163,172,263, 296,352,420 negra, 128,188,263, 273, 276, 281,
353,437 rayada, 41,139,188,190 rocas, de las, 189, 281
Lobotes surinamensis, 120 Lobotidae, 44 Lophiidae, 48, 57, 89, 130, 159, 177, 185,
Lophiiformes, 48, 121, 122, 168, 170 Lophioidei, 50
223, 251,295

ÍNDICE SISTEMÁTICO 459
Lophius, 126, 216, 249, 295, 296
Lophotidae, 40 Loricaria, 142
typus, 283 Loricariidae, 38, 103, 134, 230 Lota, 103, 368, 425
Lucio, 129 Luciocephalidae, 46 Lucios, 129, 146 Lutjanidae, 44, 150, 352, 354, 417, 444 Luvaridae, 46 Luváridos, 46
arnericanus, 45
lota, 147
Macarelas, 46, 127, 149, 159, 176, 188, 189, 194, 195, 209, 210, 221, 223, 225,417
Atlántico, del, 226 “Jack”, 298 víbora, 46
Mucrones, 311 Macropodes opercularis, 311 Macrorhamphosidae, 40 Macruridae, 48 “Madtoms”, 123, 171 Makaira, 188, 417 Malacopteri, 30 Malacopterigios, 54 Malacopterygii, 29, 30, 104, 172, 173 Malacosteidae, 36 Malacosteus, 418 Malapteruridae, 38, 441 Malapterurus, 101, 314, 365 Mandibulados en cincel, 185, 186 Mantarrayas, 12, 14, 16, 25, 52, 54, 56, 57,
68, 119, 138, 168, 170, 171, 177, 178, 179, 185, 199, 259, 268, 269, 368, 420
pequeña, 31, 268 Mantas, 32, 179, 411, 417 Marsipobranchii, 12 Mastacembelidae, 47, 441 Mastacembeliformes, 47 Mastacembeloidei, 47 Mastacembelus, 230
maculatus, 43 Maurolicus, 242 Medaka, 339,373,379 Megalomycteridae, 40 Megalomictéridos, 40 Megalopidae, 34 Megalops, 170, 186, 190, 315, 417 Megupsilon aporus, 378 Melamphaeidae, 42, 244 Melamphaes, 242 Melanocetus, 418 Melanogrammus, 420
aeglefinus, 267 Melanonidae, 48, 122, 236, 238, 420 Melanonoidei, 48 Melanostomiatidae, 36 Melanotaeniidae, 42 Menidia, 148, 349 Merlines, 46, 169, 188, 296, 417
Merluccius, 420 Merluciidae, 48 Merluzas, 48, 346, 420 Mero, 154, 155 Meros, 129, 410, 417, 444
Nassau, de, 116 Microdesmidae, 46 Micrornetrus, aurora, 273
rninirnus, 273 Microphis boajn, 251 Micropterus, 106, 126, 128, 263, 273, 354
dolomieui, 281 salmoides, 82, 164, 180, 281, 284
Microstomidae, 244 Minnows, 27, 38, 55, 64, 97, 99, 110, 137,
138, 146, 155, 163, 199, 236, 254, 271, 310, 316, 320, 321, 322, 343, 346, 354, 356, 359, 361, 379, 386,
432,434,436,438,441 394, 412, 415, 423, 427, 428, 430:
cabeza gorda, de, 163, 277, 281, 324, 358,366
nariz roma, de, 271, 277, 281, 358 ojo rojo, europeo de, 197 rodapiedras, 138 Viejo Mundo, del, 131 Véase también Carpa
Mirapínidos, 38 Mirapinnidae, 38 Misgurnus, 185, 383
fossilis, 236, 340 Mistichthys luzonensis, 298 Mixinos, 9, 12, 14, 15, 16, 18, 20, 22, 24, 30,
54, 55, 56, 58, 59, 66, 68, 70, 77, 78, 84, 89, 94, 95, 98, 102, 110, 132, 134, 173, 197, 203, 208, 216, 217, 253, 257, 269, 288, 294, 311, 313, 320, 321,356,362,420
Atlántico, del, 31 Mobulidae, 32, 411, 417 Mochokidae, 38 Mojarras, 44
Mojarrita gibosa (semilla de calabaza). labios gruesos, de, 135
138r144 ’
41 4
,I
Mojarritas, 128, 130, 144, 147, 154, 166,
Molas y M o l a , 52, 176, 266, 317, 411 Molgula, 11 Molidae, 47, 176, 295, 296 “Molly” del Amazonas, 263, 373, 380 Monacanthidae, 47 Monacanthus, 179
polycanthus, 120 Monocentridae, 42, 122 Monocirrhus, 117, 177 Monodactylidae, 44 Monopterus, 194 Monorhina, 18 Mordacia rnordax, 348 Mormíridos, 170 Mormyridae, 34, 220, 234, 314, 360, 365,
366,438 Mormyroidei, 34 Morrnyrus caballus, 37

460 ÍNDICE SISTEMÁTICO
Morone, americana, 299 Chrysops, 271 saxatilis, 41, 139, 188, 190
Moxostoma, 83, 131 anisurum, 131 erythrurum, 131, 137 macrolepidotum, 131
Mrigal, 195 Mugil, 139, 146, 186,366, 413
Mugilidae, 42, 261, 331, 442 Mugiliformes, 42 Mugiloidei, 42 Mugiloididae, 44 Mullidae, 44, 111, 143, 150, 224 Mummichog, 152, 228,287,289, 291,299,
Muraena, 207 Muraenesocidae, 34 Muraenidae, 34, 130, 136, 149, 185, 224,
419,420 Murenas, 34, 130, 136, 149, 185, 207, 224,
358,418,420 Mustelus, 119, 203, 229, 283, 314
antarcticus, 204, 205 canis, 209
cephalus, 140, 315
334,337,339
Myctophidae, 38, 238, 239, 244, 418 Myctophiformes, 38, 122 Myctophum, 117 Mylinae, 189 Myliobatidae, 32, 123, 171 Myliobatis, 137
Myoxocephalus, 191 Myxine, 55, 77, 89, 102, 203, 322, 356
Myxini, 12, 24 Myxinidae, 22, 30, 54, 69,66, 77, 95, 98,
132, 134, 197, 208, 257, 288, 294, 420
californicus, 123
glutinosa, 3 1
Myxiniformes, 15, 30, 217, 313, 314, 356 Myxocyprinus, 432, 441
asiaticus, 435
Nandidae, 44, 438 Naso, 123 Naucrates, 411 Nemichthyidae, 34, 51 Neoceratodus, 80, 83, 86, 94, 229, 232, 442
forsteri, 33 Neopterygii, 12, 83 Neostethidae, 42 Nettastomidae, 34 Nocomis biguttatus, 163 Noemacheilus, 345 Noestethus amaricola, 39 Nomeidae, 46 Nomeus gronovi, 411 Notacanthidae, 34 Notacanthiformes, 29, 34 Notacanthus analis, 37 Notemigonus, 176
Notesthes, 123 Notopteridae, 220
crysoleucas, 180, 281, 302
Notopteroidei, 34 Notopterus, 170,177, 234 Notorhynchus, 199 Nototheniidae, 46, 446 Notothenioidei, 46 Notropis atherinoides, 424 Noturus, 101, 123, 162, 171 Novumbra, 437
Odontaspidae, 32 Ogcocephalidae, 48, 52, 56, 177, 185 Ojo de luna, 34, 172,437, 438 Ojos grandes (“Bigeyes”), 42 Ojos parietales, 136, 189, 263, 268, 281, 423
europeo, 220 Ojos-tubo, 40 Oligocottinae, 271 Oligoplites saurus, 120 Olfridos, 38 Olyridae, 38 Oncorhynchus, 141, 159,180,186,189,
190, 251, 258, 266, 268, 274, 275, 281, 324, 336, 337,338, 351, 355, 359,410
gorbuscha, 270 keto, 340 nerka, 299, 338, 339 tshawytscha, 73, 74, 75, 76, 156
“Opahs”, 40 Ophichthidae, 34, 175, 185 Ophidiidae, 44, 419 Ophidioidei, 44 Ophisurus, 170 Opistognathidae, 44 Opisthoproctidae, 36 Opisthoproctus, 352 Opsanus, 90, 123, 221, 249
pardus, 45 tau, 338
Orectolobidae, 32 Orestias, 171 Oryzias, 379
latipes, 339, 373, 381 Oryziatidae, 40 Osmeridae, 36, 172, 174 Osmerus, 271
eperlanus. 267 mordax, 269
Ostariophysi, 36, 98, 235, 437,438 Osteictios. 20. 23 Osteichthyes,’l2, 14, 15, 17, 20, 23, 24, 26,
32, 66,67, 68, 70, 83, 90, 94, 108, 109, 136, 147, 160, 172, 173, 179, 194, 195, 197, 198, 200, 217, 221, 250, 288, 306, 317, 320, 329, 330, 352,359,360,398,437,438
Osteochilus, 131 Osteoglósidos, 442 Osteoglossidae, 34, 438 Osteoglossiformes, 34 Osteoglossoidei, 34 Osteostrácodos, 18 Ostraciidae, 47, 109, 150, 177, 179, 224,
Ostracodermos, 12, 15, 19, 21, 22, 24 420

ÍNDICE SISTEMÁTICO 461
Paleopterygii, 12 Paleospinax, 18, 26 Palometas, 46, 417 Pámpanos, 42 Pantodon, 186
Pantodontidae, 34, 441 Paralabrax, 417 Paralichthys, 113, 350
olivaceus, 340 Patos de Bombay, 38 Pececillos del fango, 36, 172, 185, 234, 423,
Peces acechadores, 40 Peces aguililla, 44 Pecesaguja, 40, 52, 120, 135, 136, 170,
296,417 Pecesagujón, 39, 40, 103, 109, 120, 134,
139, 171, 175, 177, 209, 251, 279, 282,419,420
Peces ángel, 111, 116, 121, 176, 410, 420 Bermuda, 128
Peces araña, 123 Peces arco iris, australianos, 42 Peces arquero, 44, 130 Peces azulejo, 42 Peces barba, 42 Peces blancos, 36, 110, 189, 241, 276, 281,
buchholzi, 186
434,437,441
311,328,407,423 lago, de, 131 redondos, 94,131
Peces blenioideos, 89 Peces boca, 242,417,418 Peces bolsa, 47 Peces burlones (¿harapos?), 46 Peces camarón, 40 Peces cardenal, 42, 111, 122, 283, 411, 442 Peces cavadores de grava, 46 Peces cavernícolas, 48, 57, 95, 120, 360, 438
ciegos, 111, 280 Peces cinta, 40, 52, 170 Peces cirujano, 46, 109, 111, 123, 126, 150,
153,329,420
Peces cocodrilo, blanco, 85 Peces cofre, 47 Peces cola de rata (Granaderos), 48, 420 Peces con aletas con radios, 12, 14, 17, 24,
26, 27, 28, 349, 361 Peces con armadura y cuello articulado, 23 Peces con grumos, 47, 120, 419 Peces con púas, 46 Peces conejo, 46, 124 Peces cono de pino, 42,122 Peces corneta, 40, 134 Peces cuatro ojos, 40, 283, 284, 349 Peces de aleta ondulada, 27, 28, 33, 34, 80,
90,91,110,121,126,138,139,170, 172,174, 178, 179,195, 199, 206, 212, 215, 234, 251, 272, 281, 284, 288,295,311,346, 367, 386
Peces de aletas lobuladas, 12, 14, 17, 24, 26,68, 84, 96, 107, 110, 237, 238, 331,347, 348, 353
convicto, 285
celacanto, 14, 347
Peces de algas marinas grandes, 420
Peces de arrecife, 350 Peces de boca delgada, 44, 122 Peces de cabeza lustrosa, de mares profundos,
Peces de cabeza plana, 44,46 Peces de cola cuadrada, 46 Peces dedo, 44 Peces de dientes oblicuos, de mar profundo,
240 Peces de escama grande, de mares
Peces de hueso, 34,150,224,296,419,420
Peces de lengua ósea, 34, 197, 438,442 Peces de lomo emplumado, 34, 177,220 Peces de los arrozales, 40, 373, 381 Peces de ojos pedunculados, 36, 273, 296,
Peces de puntas, 47 Peces de Shepherd, 46 Peces dragón, 46
122
365,441
296,317,418
gigante, 120
36
profundos, 42
mares profundos, de los, 197
418
escamosos de los mares profundos, 36,
Peces elefante, 34, 37, 135, 220, 314, 360,
Peces escorpión, 47, 123, 124, 277, 283,
Peces espada, 46, 169, 188, 296, 411, 443 Peces espátula, 27, 32, 103, 104, 108, 130,
135,139, 169, 172, 195, 199, 211, 437,441
Peces espinosos de mares profundos, 42 Peces fantasma, 36 Peces flechadores, 42, 111, 121, 171, 179,
243, 263, 379, 385, 386, 427, 437, 438
abanico, con cola en, 271, 281 arco iris, 281 Johnny, de, 271, 277,278,281,385 menor, 1,171, 273 verde, de lado, 385
Peces gato (Bagres), 52, 99, 123, 139, 140, 143, 162, 170, 175, 177, 209, 213, 220, 224, 230, 235, 284, 311, 314, 331, 342, 343, 358, 359, 361, 363, 365, 412, 415, 423, 425, 427, 437, 438,441
acorazado, 38, 105, 132, 168, 230 agua dulce, de, 111 América del Norte, agua dulce, 38, 102,
136 arrido, 437 arroyos montañosos, de los, 224 Asiático, 363 bágrido, 38 bandera, 283 banjo, 38 Brasileño, 283 cabeza de toro, 227 cabeza plana, de, 137 calíctido acorazado, 38 caminante, 186 Véase también Respiración aérea, de

canal, 157, 281 criador en el hocico, 279 diplomístido, 38 dorádidos acorazados, 38 eléctrico, 38, 101, 314, 365, 441 espinoso, 38 esquilbeido, 38 Eurásico, 38 herbívoro, 142 ictalúrido, 438 Indio, 130 Indopacífico, 123 invertido, 38, 238 laberíntico, 231 laberintino, 158 loricárido acorazado, 141 “madtom”, 101 marino, 38, 111, 124, 283 obstétrico, 38, 279 oriental, 132 parásito, 38 pimelódido, 38 plotósido marino, 38 respiración aérea, 38 Véase también Caminando saco aéreo, 231 sisórido, 134 Sudamericano acorazado, 103
Peces gato (glanos), 140 Peces gato-piedra, 123 Peces “gibber”, 40 Peces globo, 47, 51, 68, 141, 149, 177, 199,
228, 262,313,420,443 nariz afilada, de, 47 norte, del, 43
Peces gusano, 46 Peces hielo, 46, 85, 296 Peces hinchables chupadores, 273, 282 Peces hocico-tubo, 40, 55 Peces hoja, 44 Peces incubadores, 242, 311
Peces jabón, 42 Peces laberinto, 362 Peces lagartija, 38
bahía, de, 35 Peces lanceta, 38 Peces lanza areneros, 46, 185, 261 Peces lengua, 47 Peces linterna, 38, 116, 122, 238, 239, 244,
418,420 Peces lobo, 46 Pecesloro, 44, 111, 126, 130, 137, 138,
mares profundos, de, 36
141, 150, 153, 171, 272, 336, 339, 366,420
arco iris, 41 Peces luchadores, 278, 282, 326 Peces machete, 46 Peces mandibulados, 15, 24 Pecesmariposa, 44, 49, 111, 130, 134, 176,
350,411,420 nariz larga, 135, 169
Peces murciélago, 48, 53, 55, 177, 185 Peces negros de río, 44 Peces ojos de linterna, 42
Peces ojos de perla, 38, 350 Feces bseus, 9, 12, 13? 14, 15, 17, 24, 26,
29: 54, 55, 57, 61, 62, 54, 66, 67, 69, 70, 61, 73, 74, 75, 86, 90, 91, 93, 94, 95, 106, 107, 108, 219, 248, 346, 349,351, 352,359,360, 361,437
Peces peine, 47 Peces perciformes, 61, 62, 64 Peces perlados, 44, 296, 411 Peces piedra, 123 Peces planos, 118, 139, 147, 168, 176, 295,
352.445. 446 Peces proa, 46 Pecespuerco, 47, 52, 111, 120, 178, 179,
236.420 naranja‘, 120
Peces puerco espín, 47, 109, 139, 149, 177, 420
Pecespulmonados, 12, 14, 15, 17, 24, 26, 56, 57, 59, 68, 86, 90, 91, 94, 96, 105, 107, 110, 154, 174, 193, 195, 197, 198, 199, 211, 232, 233, 347, 353,365,437,438,441,442
africanos, 32, 80,83, 185, 196, 209, 211,232,250,296
australianos, 32, 33,80,83,86,229,232 sudamericanos, 32, 80,83,96, 194,206,
211,224,232,288,295,296 Peces rana (o sapo), 48, 89, 185 Peces relampagueantes, 36 Peces remo, 39, 40, 170 Peces sable, 47 Peces sapo, 48, 89, 90, 123, 185, 221, 236,
249,338 leopardo, 45
Peces señorita, 44, 111, 329, 411, 420 Peces sierra, 32, 169, 442
Peces sin mandibulas, 24 Peces tigre, 42 Peces tres-espinas, 47 Peces triple-cola, 44, 120 Peces trompeta, 40, 134
tropicales, 191
Pecesvela, 46, 169, 171, 186, 188, 296, 411.417.418
Peces víbora, 36
Peces vieja (señorita), 44, 111, 134, 150, 209, 210, 258, 272, 317, 346, 352, 410,420
mar profundo, de, 134, 137, 244
enano, 120 Peces vivíparos, 40, 53, 106, 171, 172, 284,
381, 385 mexicanos, 40, 283, 294
Peces voladores, 40, 54, 120, 170, 171,
Pegasidae, 47 Pegasiformes, 45 Pegasos, 45 Pegasus umitengu, 45 Pejelagartos (catanes), 27, 28, 34, 80, 84, 90,
186,187, 188,244,269, 277,417
104, 105, 107,120,126,129, 138, 139,169,172,195,197,199, 206, 212, 234, 251,288, 296, 311, 323, 387

ÍNDICE SISTEMÁTICO 463
Florida, de, 212 nariz larga, de, 33, 196, 197
Pejesapos, 45, 48, 52, 57, 88, 121, 127, 129 169, 168, 170, 177, 185, 210, 223, 249, 251, 295, 296, 297,443
mares profundos, de los, 48, 122, 132, 175, 270, 271, 273, 280, 295, 296, 418, 419, 447
Pempheridae, 44 Perca, 71, 142, 162, 213, 254, 263, 277,
282, 335, 346,423 flavescens, 14, 61, 62, 69, 144, 163,
166, 189, 238, 281, 398, 399 fluuiatilis, 166, 239, 265, 338
Perca de arrecife, 283 Perca de las rompientes, 44, 279, 283, 296,
420, 446 Perca de tronco, 243 Percas, 42, 52, 71, 84, 105, 121, 136, 142,
162, 172, 213, 254, 263, 277, 282, 320, 335, 346, 415, 423, 437, 438, 441
agua dulce, de, 30 amarilla, 14, 61, 62, 69, 144, 163, 166,
189, 238, 281.398. 414 arrecife, de,’273 . blanca, 299 enana, 273 europea, 166, 239, 265, 338 Sacramento, de, 282
Percas areneras, 44 Percas pirata, 48, 56, 437, 438 Percas trepadoras, 46, 80, 95, 185, 230,
Percas-trucha, 39, 48, 105, 189, 437, 438 Percichthyidae, 42, 170 Percidae, 30, 42, 84, 111, 121, 136, 170,
172,320, 379,398, 399,437,438 Pércidos, 170 Perciformes, 30, 42, 138, 238, 283, 398,
Percina caprodes, 243 Percoidei, 42 Percophididae, 44 Percopsidae, 48, 437, 438 Percopsiformes, 48 Percopsis, 189
Percopsoidei, 48 Periophthalmus, 95, 185, 199, 206, 257,
Petromyzon, 71, 85, 93, 190, 250, 253,
239, 245
437
omiscomaycus, 39
258,261, 336, 347
254, 258,323,335,346 marinus, 14, 31, 61, 126, 132, 133, 141,
159, 175, 216, 275, 281, 294, 296, 352, 356, 357
66, 67, 77, 91, 97, 98, 102, 132, 134, 143, 146, 170, 171, 175, 185, 189, 203, 206, 212, 216, 250, 253, 257, 271, 273, 277, 281, 286, 295, 296, 311, 313, 316, 317, 320, 347, 349, 351 ,356 ,427 ,442 ,447
Petromyzonidae, 22, 30, 54, 55, 56, 59,
Petromyzoniformes, 15, 30, 312 Petromyzontes, 12, 24
Pez adherente, 410
Pez anémona, 41 1 Pez cabeza de víbora, 41, 46, 171, 230,
Pez cola-de-espada, 373, 379, 381, 385
Véase también Rémora
230, 231
cometa silvestre, 376 dorado, 376
Pez de cola emplumada, 234 Pez de labios marginales, 131 Pez de lados plateados, 42, 57, 148, 348,
424,442 arroyo, de, 106, 186, 269, 273, 274, 281
Pez de los sargazos, 53, 111 Pez de mandíbulas sueltas, de mar
Pez de otoño, 281 Pez de roca, 55, 187, 210, 419, 420
profundo, 36
Antártico, 46, 446 dientes en peine, 46
Pez guitarra, 32 Pez iluminador, 46 Pez imitador de hoja, 122, 177 Pez meteorológico, 185, 340 Pez mira abajo, 41 Pez mosquito, 271, 272, 273, 282
Pez salto de carpa, 116 Pez sapo, 122,419 ,420 Pez señorita, 84 Pez soldado (candil), 42, 236, 394 Pez paraíso, 311
Véase también Gambusias y Gambusia
Pez perro, 199, 229,314 espinoso, 14, 61, 67, 73, 80, 109, 123,
193, 225 liso, 204, 205, 314 manchado, 196, 202
Pez piloto, 411 Pez sierra de mano, 136 Pez rata, 31,123 ,311 Pez rodador de piedras, 55 Phallostethidae, 42 Pholidae, 44 Pholis gunnellus, 187 Photichthyidae, 36 Photocorynus, 277
spiniceps, 270 Phoxinus, 55, 322, 346, 366
phoxinus, 354 Phractolaemidae, 36, 441 Phyllopteryx, 53, 111, 117, 120, 177 Picnodontos, 28 Pico de espada, de agua dulce, 103 “Pikeheads” (Lucios), 46 “Pikes” (Lucios), 36, 66, 126, 136, 139,
141, 143, 152, 172, 187, 207, 352,
433.437.441 355, 358, 379, 387, 415, 423, 432,
~ > ~~- norte, del, 57, 136, 140, 142, 146, 153,
187,265, 281, 298,328,414 Pimelodidae. 38. 438 Pimephales, 277; 28¡, 324, 358
notatus, 271 promelas, 323, 366
Pimephalinae, 163

464 ÍNDICE SISTEMÁTICO
Piraña, 126,138 Pirarocú, 233 Piscardo, 354,366 Pisces, 11, 12, 16, 21 Pisodonophis, 175,184 Placodermi, 19, 20, 23 Placodermo antiarco, 19 Placodermos, 15, 20, 22, 24, 26
Platija, 187 Platax, 120 Platilla (sardinita), 33 8 Platycephalidae, 47 Platycephaloidei, 47 Plecoglossidae, 36 Plecostomus, 168 Plectognathi, 46 Pleuronectes, 152, 153, 337
flesus, 188 Pleuronectes platessa, 188, 267
antiarcanos, 18
Pleuronectidae, 47, 52, 111, 118, 139, 159, 185. 223.419
Pleuronectiformes, 47, 57, 118, 147, 168, 176, 295,348,420,445,447
Pleuronectoidei, 47 Plotosidae, 38, 437 Plotosus, 123 Poecilia formosa, 263, 373, 380 Poeciliidae, 40, 53, 106, 171, 172, 271,
Poeciliopsis, 263, 373, 380 Poliptéridos, 24, 25, 32, 33, 68, 80, 86, 90,
280, 283,381,384,385
104, 107, 170, 174, 195, 224, 296, 338,438
Polípteros, 12, 26, 34, 80, 224 Polymixiidae, 42 Polymixioidei, 42 Polynemidae, 42 Polynemoidei, 42 Polyodon, 27, 103, 104, 130, 139, 169, 199,
211,437 spathula, 135, 195
Polyodontidae, 27, 34, 108, 171 Polypteridae, 32, 438 Polypteriformes, 24, 26, 35, 307 Polypterus, 68, 80, 83, 90, 104, 107, 170,
174,195,296 senegalus, 33
“Pollacks”, 299, 420 Pollachius, 420
Pomacanthus, 116, 121 Pomacentridae, 44, 111, 420 Pomadasyidae, 44, 236, 417 Pomatomidae, 42 Pomatomus, 417
“Pomfrets”, 44 Pomoxis, 424 Porichthys, 419, 420 Premnas, 411 Priacanthidae, 42 Priapium, peces, 39
virens, 299
saltatrix, 129, 347
falostétidos, 42 neostétidos, 42
Prionotus, 346 Pristidae, 32, 191 Pristiophoridae, 32 Pristiophoriformes, 32 Pristis, 109, 170 Pristiurus, 3 14 Prochilodus, 189 Prosopium cylindraceum, 93,131 Protocordados, 11 Protochordata, 11 Protopteridae, 438 Protopterus, 80, 83, 94, 185, 209, 211, 233,
250,296 annectens, 196
Psephurus, 27, 103, 437, 441 Pseudoliparis, 420 Pseudophallus starksi, 39 Pseudopleuronectes americanus, 43 Pteraspidomorfo, 18 Pteraspidomorphi, 12, 19, 20 Pteráspidos, 22 Pteraspis, 18, 362 Pterois, 123, 136 Pterois voladores, 123, 136 Pterophryne, 53, 111 Pterophyllum, 176
scalare, 121 Pterothrissus gissus, 197 Ptilichthyidae, 46 Pujeques, 54, 168, 171, 283, 437 Pungitius, 103 Pylodictis olivaris, 137
Quimeras y Chimaera, 12, 15, 16, 24, 25, 26,31, 33, 55,66,68, 109, 123, 136, 199,212, 272,420
Quimeroides, 109
Rachycentridae, 42 Radiados, 12, 14, 15.16, 25, 26, 70, 122,
124, 147, 148, 170, 171, 217, 259, 337, 342, 344, 347, 361, 358, 360, 363
águila, 32,137,171,179 águila manchada, 123 ciegos de.los mares profundos, 95 eléctricos, 32, 122, 363 púa en el lomo, con, 363
Radiicefálidos, 40 Radiicephalidae, 40 Raja, 119, 136, 179
binoculata, 199 clavata, 363 erinacea, 31, 68 radiata, 339
269, 283
179,217, 283,363
Rajidae, 32, 52, 57, 138, 168, 171, 185,
Rajiformes, 32, 55, 57, 138, 168, 171, 177,
Ramoneador, 46, 52 Rayas de espina, 32, 103, 109, 123, 171,
178,419,420 murciélago, 123
Regalecidae, 40 Regalecus, 170

ÍNDICE SISTEMÁTICO 466
glesne, 39 Remigolepis, 19 Rémoras y Remora, 44,171, 209, 348,
Retropinna, 442 Retropinnidae, 36 Rhincodon, 1, 130,443 Rhincodontidae, 32 Rhinobatidae, 32 Rhodeus, 269, 282
Rhynchocymba, 362
Riuulus paludium, 399 Robalos, 42 Roccus, Chrysops, 189
410,412
amarus, 272, 274, 366
nystrani, 368
saxatilis, 43, 146, 198, 199 Véase también Morone
“Rock gunnel”, 187 Rocotes, 47, 123, 283, 295, 296, 420, 446 Roncachos, 420 Roncadores, 44, 236,417 Rondelétidos, 40 Rondeletiidae, 40 Rondeletioidei, 40 Ronquilos, 44 “Rudd”, 137 Rutilus rutilus, 141
Sábalos, 34 Saccobranchus, 230 Saccopharyngidae, 34, 55,122,139,153 Saccopharyngoidei, 34 Saccopharynx, 418
ampullaceus, 35 Salangidae, 36, 296 Salángidos, 36 Salmo, 66, 93, 108, 127, 130, 131, 143,
149, 189, 207, 213, 229. 261. 262. 268, 273, 274, 281; 282; 317; 322; 324,331.335.355.407
clarki, 281, 301, 426 ’
gairdneri, 114, 150, 163, 183, 184, 196, 199, 204, 205, 208, 265, 281, 299, 300,338,385
salar, 110, 159, 186, 190, 254, 255, 258, 261,267,301,352
trutta, 144, 153, 163, 164, 188, 206, 265, 281, 299, 302, 303, 329, 349, 412
180, 186, 189, 190, 223, 238, 244, 251, 254, 258, 266, 281, 294, 299, 324, 331, 337, 338, 351, 355, 359, 423,447
Atlántico, del, 110, 190, 254, 255, 268, 301,317,352
Salmón, 30, 36, 66, 90, 138, 143, 158, 159,
Salmonetes, 44, 111,143,150, 224 Chinense, 73, 74,75, 76,156 “chum”, 340 Pacífico, del, 141, 188, 189, 268, 274 rosado, 270 “sockeye“, 338, 339
Salmonidae, 36, 53, 66, 90, 151, 165, 170, 172, 244,407,423,447
Salmoniformes, 36 Salmoninae, 138, 146, 150, 151, 153, 155,
158, 174, 189, 212, 238, 254, 268, 272,294,331,341,356,373,441
Salmonoidei, 36 Salmonoideos, 55, 59, 97, 174, 263, 379 Saltarines del fango, 95, 185, 199, 206, 230,
257, 258, 261,336,347 Africanos, 34
355,407
299,300,426
Saluelinus, 127,130, 213, 261, 268, 276,
fontinalis, 103, 163, 267, 276, 281,
malma, 385 namaycush, 165, 189, 281
Véase también Pejesapos Sapos, 121
Sarcopterygii, 26 Sarda, 188 Sardinas (Arenques), 27, 30,33,34, 56,
66,83,97,111,127,146,147, 150,158,159,172,176,189, 234,237,268,295,296,358, 413,417,421,424,441,445, 446,447
Africanos, 34 Atlántico, del, 265, 266
Pacífico, del, 315 mares profundos, de, 35, 44
redondas, 34 Sardinella, 447 Sardinops caerulea, 267 Sargo, 44, 137, 150, 171, 235, 417 “Saury”, 40, 141, 241
Atlántico, del 37 Scardinius, 137 Scaridae, 44, 111, 126, 130, 137, 138, 141,
Scarus, 366, 420 guacamaia, 41
Scatophagidae, 44 “Scats”, 44 Sciaenidae, 44, 136, 236, 237, 320, 420 Scleropages, 442 Scomber, 188, 210, 221
Scomberesocidae, 40, 241 Scomberesocoidei, 40 Scomberesox, 141
Scombridae, 46, 127, 147, 149, 159, 176,
Scombroidei, 46 Scopelarchidae, 40 Scopelarchus, 350 Scorpaena, 123,277,282, 317 Scorpaenidae, 47, 123, 283, 295, 418, 420 Scorpaeniformes, 47 Scorpaenoidei, 47 Scorpidae, 42 Scyliorhinidae, 32, 269, 321 Scyliorhinus, 228, 256, 363
150,153,171, 272, 336,339,420
scombrus, 188, 226, 227
saurus, 37
194,199, 223,410,417
caniculus, 196, 202 catulus, 197
Scyllium, 119, 142, 197

46 6
cunicula, 21 o tieytalinidae, 46 Schilbeidae, 38, 130, 131, 159 Schindleria, 296 Sehindleriidae, 46 Schindlerioidei, 46 Searsia schnakenbecki, ’I 22 Seársido, 3 5, 122 Searsiidae, 36 Sebastes marinus, 267 Sebastodes, 283 Selene vomer, 41 Semotilus, 138, 274, 282
atromaculatus, 270, 281, 362 corporalis, 281
Serranidae, 30, 42, 84, 89, 122, 136, 150, 172, 263, 296, 352, 410, 420, 442, 444
Serranidos, 144, 263, 442 Serrasalmus, 126, 138 Sicydium, 190 Siganidae, 46, 124 Siganus, 124 Siluridae, 38, 132, 209, 342, 441 Siluris glanis, 140 Siluroidea, 175, 177 Siluroidei, 38, 220, 314, 343, 358, 426 Siluroideos, 441 Simenchelyidae, 34 Simenchelys parasiticus, 135, 175 Sinanceja, 123 Sisoridae, 38, 224, 235 Sidridos, 38 “Skipjack”, 186 Solea, 340 Soleidae, 47, 185, 419 Soleoidei, 47 Somniosus microcephalus, 149 “Soupfin”, 95 Sparidae, 44, 137, 150, 171, 235, 417 Sphagebranchus, 170 Sphoeroides, 313
Sphyraena, 126, 129, 138,139, 188 Sphyraenidae, 42, 150, 410, 417 Sphyraenoiaei, 4 2 Sphyrna, 283
tiburo, 293 Sphyrnidae, 32, 293 Spinachia spinachia, 188 Spinax, 122 ”Sprat”, 188 Squalidae, 32, 140, 314, 321, 420 Squaliformes, 32, 55, 56, 130, 136, 179,
208, 209, 212, 217, 283, 311, 320, 328,342,360,362,366,443
maculatus, 43, 228
Squalus, 56, 71, 93, 109, 143, 335, 358 acanthias, 14, 61, 67, 73, 80, 123, 193,
suckleyi, 199 Squatinidae, 32 Stephanoberycidae, 42 Stephanoberycoidei, 40 Stephanoberyx monae, 41 Sternoptychidae, 36, 242
225, 293
Stichaeldae, 46 Stizosledion, 136, 189, 263, 268, 281, 423
lacioperca, 220 vitreum, 169
Stomius, 417, 418 Stomiatidae, 36, 122 Stomiatoidei, 36 Stromateidae, 46, 417 Stromatoidei, 46 Strongylura, 120
longirostris, 135 Stylephoridae, 40 Stylephoroidei, 40 Synaphobranchus, 418,420 Synbranchidae, 47, 143, 154, 170, 194 Synbranchiforrnes, 47 Syngnathidae, 40, 103, 109, 120, 134, 141,
Syngnathoidei, 40 Syngnathus, 175, 207, 282 Synodontidae, 38 Synodontis batensoda, 238 Synodus foetens, 35
171,177,419,420
Tambores, 44, 138, 236, 237, 320, 321
Tautoga onitis, 348 Tautogo, 34, 84, 170, 186,190, 315,417 Tejedores, 44 Teleostei, 21, 27, 28, 29,68, 70, 90, 193,
194, 195, 197, 207, 219, 310, 347, 351,437
Teleósteo, 347 Teleósteos, 84, 98, 119, 194 Teleostomi, 12, 15 Telodontos, 18. 20, 22
agua dulce, de, 173
Tellia, 171 Tenca. 137. 221
. .
Tetragonuridae, 46 Tetragonuroidei, 45 Tetraodon, 262 Tetraodontidae, 47, 51, 68, 139, 149, 177,
Tetraodontiformes, 45 Tetraodontoidei, 47 Thalassophryne, 123 Theraponidae, 42, 442 Thunnus, 188,189, 209,417,443
199,224,420,443
alalunga, 188 thynnus, 213
Tiburón sierra, 109 Tiburones, 9, 12, 14, 15, 16, 24, 25, 26, 54,
55, 57, 59,60, 66.68, 71, 74, 78, 86, 88, 90, 91, 93, 94, 95, 97, 98, 103, 106, 107, 110, 119, 122, 123, 124, 126, 129, 130, 136, 141, 142, 148, 172, 174, 179, 194, 195, 198, 200, 217, 259, 268, 288, 311, 320,321, 334, 336, 338, 342, 346, 347,348, 350, 351, 358, 369, 360, 363,364, 366,411,417,443
angel, 32 arena, 32,314

ÍNDICE SISTEMÁTICO 467
azul, 188 ballena, 1, 32, 130, 149, 280, 443 blanco, 143 cabeza de toro, 32, 197 catiabota, 32,149 cazón, 32, 56, 143, 149, 321, 335, 336,
420 coludo, 32 cornudo, 123, 272 espinoso o acantodio, 23 gata manchada, 32 Groelandia, de, 149 lagarto, 32, 52 lámnido, 213 macarela. 32. 149. 194. 219. 314 , . , mamón, 209 martillo, 32, 294 pintarroja o gato de mar, 32, 197, 256,
269,314,321.363 requiem, 32; 176; 195 seis branquias, de, 31, 198, 199 sierra, 32 siete branquias, de, 198, 199 tigre, 324 toro, 324
Tiburones adherentes, 43, 44 Tilapias y Tilapia, 126, 148, 155, 166, 175,
176, 275, 284,310,329 macrocephala, 324, 326 melanopleura, 126 mossambica, 166, 175
Tinca, 137, 221 Torito, 109 Torpedinidae, 32 Torpediniformes, 32, 55 Torpedo, 225,363, 365
Toxocotes jaculator, 130 Toxotidae, 44 Trachichthyidae, 40 Trachinidae, 44, 123 Trachinus, 123 Trachipteridae, 40, 52, 170 Trachipteroidei, 40 Trachipterus, 296 Trachurus, 316
Tragadores, 35, 122, 153, 418
Traquíctidos, 42 Triacanthidae, 47 Triacanthodidae, 47 Triakis, 283 Trichiuridae, 46 Trichodontidae, 44, 185 Trichomycteridae, 38 Trichonotidae, 44 Trigla, 188, 241, 322 Triglidae, 47, 185, 236, 320, 419, 420 Triodontidae, 47 Trucha de Pátzcuaro, 42, 52, 84,165,166,
marmorata, 225
symmetricus, 298
mar profundo, de, 44, 55, 136, 139, 244
176, 180, 213, 273, 276, 278, 281, 284, 295, 325, 350, 359, 380, 395, 410,424,437,438,440
común, 414
manchas naranjadas, de, 272 oceánica, 47, 177, 266 verde, 138, 227
138, 146, 151, 153, 155, 157, 158, Truchas, 36, 53,66, 90, 114, 127, 130,
170, 172, 189, 199, 213, 223, 228, 238, 261, 268, 273, 274, 282, 294, 317, 331, 335, 341, 355, 356, 373, 380,414,423,426, 441
204, 205, 208, 265, 281, 299, 300, 303,338,385
café, 144, 153, 163, 164, 188, 206, 265, 281, 299, 201, 302, 303, 328, 349, 412,415
arco iris, 118, 148, 150, 163, 183, 196,
garganta cortada, 281, 301 lago, de, 165, 189, 281 marina, 236,417 Nueva Zelanda, de, 35, 36
Truchas Thar”, 213, 268, 276, 373, 407, 415
Toritos (Peces cofre), 109, 150, 177, 224, 420
Trompeteros, 44 Trygon uiolacea, 293 Trygonidae, 17 1 Trypauchen, 313 Tunicados, 11 Typhlogobius, 411 Typhlosymbranchus, 170
Umbra, 185, 234, 437 Umbridae, 36, 172, 423, 432, 434, 438,
Uranoscopidae, 44, 121, 124 Uranoscopus, 121 Urochordata, 11 Urophycis, 346, 420
441
Vertebrata, 11
Winteria, 351
Xenodexia, 271 Xenomystus, 170 Xiphias, 169,188,411,443 Xiphiidae, 46, 169, 296 Xiphioidei, 46 Xiphophorus, 373, 379, 381
helleri, 271, 385 maculatus, 373, 385
Yarrella, 233 Yenínsidos, 40
Zalescopus tosae, 135 Zanclidae, 420 Zaniolepidae, 47 Zaproridae, 46 Zeidae, 42, 134, 418 Zeiformes, 42 Zenopsis ocellata, 41 Zoarces, 187 Zoarcidae, 44, 283, 447


Índice Temático
Abastecimiento de peces, 410 Abertura rectal, 91 Aberturas branquiales externas,77 Aberturas branquiales internas, 77 Ablación del cerebro anterior, carpa
dorada, 310 espinocha, 311 pez del paraíso, 311 tilapia, 310
Abrazador (clasper) frontal, 272 Absorción, de materiales digeridos, 148
Acciones de discriminación, 323 Acido carbónico, 224 Acido hidroclórico, 146 Acido nucleico, 113 Acido sulfhídrico, 409 Acidos grams, 146, 149, 159 Acidófilos, 211 Acetilcolina, 262 Acetilcolinesterasa, 262 Actinotriquia, 173 Actitudes crípticas, 120 Acuacultura, 405 Adaptación, 383 Adenohipófisis, 330, 331, 335 Adenosín trifosfato, 158 Adrenalina, 337
África, 430, 442 efecto vasodilatador, 261
lagos, 415 lagos de falla, 387
capacidad calórica, 404 composición química, 334 consumo, 156, 251, 253 contenido en el pez, 158 culebras, 175 densidad máxima, 404 en reposo, 422
extraenteral, 125
Agua, 404
equilibrio, 257, 259 flujo, 422 Iéntica, 422 Iótica, 422 para el metabolismo, 156 subterránea, 422 transparencia, 404
Aguas rezagadas en las corrientes, 427 Agujeramiento, 185
Agotamiento salino, 258 Aislamiento, bioquímico, 386
ninfas de efeméridos, 427
ecológico, 385 físico, 386 fisiológico, 386 genitalia, 386 geográfico, 3 85 mecanismos, 385 reproducción, 386
Alanina en plasma, 209 Alaska, 431 Albinismo, pececillos de acuario, 374
Albinos, 41 O Albúmina en plasma, 208 Aldosterona, 338 Alelismo mtiltiple, 377 Aleta, ausencia, 170
herencia, 374
adiposa, 53 altura, 395 anal, 53,170,177, 395 caudal, 53,170,177 con radios, 53, 172, 394 diferenciación, 170 diferencias sexuales, 171
dorsal, 53,171 disco suctor pblvico, 170
larvaria, 287 longitud, 395 media, 53, 169, 173, 177
46 9

470 ÍNDICE TEMÁTICO
orígenes, 169 pareadas, 53, 169, 177 pectoral, 53,171,395 pélvica, 53,171,395 ventral, 53
Aleta caudal, dificerca, 68 bifurcada, 395 eficiencia, 184 heterocerca, 67 homocerca, 67 proterocerca, 67
Aleta pectoral, soportes, 68, 69 Aleta pélvica, ausencia, 394
cintura, 68,69 clasper, 123 , disco adhesivo, 172 soportes, 68
Algas, 404 alimento, 155
Algales, células, 148 cápsulas de flotación, 120 como alimento, 130 frondas (talos), 121
condiciones climáticas, 143 estímulo visual, 343, 354 hábitos, 129 olor, 385
Alimentación sarcofágica, 132 Alimento, 125
Alimentación, adaptaciones, 132, 133, 134
absorción, 145 bacteria, 126 básico, 155 bolo, 145 calidad, 155 carbohidratos, 126 consumo, 153 factor de conversión, 150 invertebrado, 126 microcrustáceos, 126 minerales, 126 movimiento en el tracto, 145 pirámides, 128 plancton, 126 primero (primario), 297 proteinas, 126 protozoarios, 126 toma, 154 tramas, 128 utilización, 145 vertebrados, 126 vitaminas, 126
Alimento áspero, 153 Alimentos naturales, 125 Alopoliploidía, 384 Alotipo, 400 América, 442 América Central, 437 América del Norte, 430 América del Sur (Sudamérica), 430 Aminoácidos, 1 4 8
en plasma, 209 síntesis, 156
Amoníaco, 253 excreción, 254, 255
Ámpula (S), 98
Anacaris, 424 Anacharis, 424 Anadejas, como alimento, 126 Anadromia, 189 Anádromos, 159, 258 Anatomía, covertura del cuerpo, 52
de Lorenzini, 342, 344, 363, 364
aberturas, 55 comparada de los grupos principales, 54 corazón, 185 definición, 5 esqueleto, 57,70 forma del cuerpo, 53, 54, 55 interna, 79, 81, 82 interna del tiburón, 83 interna de la trucha de arroyo, 81 interna de la trucha de boca grande, 82 sistema circulatorio, 185 superficial externa, 51 vértebras, 65
Andrógenos, 3 3 3 Anfidromia, 190 Anhidrasa carbónica, 220, 229
en el plasma, 209 Anillos en la escama, 102 Anillos, en huesos, 162
en escamas, 108 Ano, 84 Antártida, 430 Anticuerpo ciguatera, 150 Anticuerpos, 212 Anuncio, 121 Aorta dorsal, 201, 204 Aorta ventral, 194, 196, 198 Aparato de Weber, 98, 236 Apetito, 164 Aprehensión, 325 Aprendizaje, reflejo, 323 Arbol filogenético, 24,391 Arco neural, 65 Archipiélago Indo-Australiano, 441 Ardeidae, 175 Area cerebral básica, 316 Arginina, 156
Aristóteles, 391 Armadura, 103 Artemia salina, 384 Asimilación, 152, 153 Asociación Británica para el Avance de la
Asteriscus, 98, 361 Asteroidea, 411 Atlántico del Norte, 447 Atrio del corazón, 195,196 Audición, 97, 235, 359
direecional, 362 tnargen, 236 órganos, 97 papel de la vejiga gaseosa, 235
‘‘Aufwuchs”, 409 Australia, 430, 442, 446 Autolisis, 148 Autopoliploidía, 384
vasotoxina, 333
Ciencia, 397

ÍNDICE TEMÁTICO 471
Autosomas, 377 Aves, 161
como presa, 127 no voladoras, 386 número de especies, 1, 2
Avestruces, 387 Aztecas, 390
Bacteria, 412
Baja California, 446 Balanceo, 3 6 1 Ballenas, 1 ,132 ,411 , 416 Barbilla(s), 111, 343
esturión, 343 “loach”, 345
endocomensal, 148
Barreras para la dispersión de los peces
Basiapófisis, 66 Basiósteo, 67 Basófilos, 211 Bazo, 287
formación de células sanguíneas, 211 Béntico, 406
medio ambiente, 416 zona, 424
Bentos, 409 Bilirmbina, 147 Bilis, 147, 148
costeros, 445
conducto, 140, 142 pigmentos, 147 sales, 147
Biliverdina, 147 Biocromos, 112 Biogeografía, 428 Bioluminiscencia, 122 Biomasa, 128, 129 Biomasa piramidal, 405 Biotopos, 415, 416 Bioquímica, definición, 6 Boca, 54, 55, 84
inferior, 55 succionadora, 131 terminal, 55
Bocio (papera), 127 Bolsa de la cría, 175 Bolsa de Ratke, 330 Bolsa nasofaríngica, 356, 357 Bomba bucal, 222 Borneo, 441 Bulbo arterioso, 185, 198 Blastodermo, 287
Blastodisco, 289 Blastoporo, 287 Bradicardia, 337 Branquia(s), 77
aberturas, 55 arcos, 218 bacalao, 200 bolsas, 77 circulación, 200 circulación del agua, 221 curso sanguíneo, 218 estructura, 216
Fundulus heteroclitus, 289
excreción, 257, 260 externa, 225 filamentos, 221, 222 función, 216 hendedura, 364 laminillas, 221, 222 musculatura, 217 peces óseos, 200, 218, 219 proporción con la superficie del cuerpo,
253 radios, 217 rastrillo, 84, 131, 220 restitución de las pérdidas de sal, 254 septo, 218 tiburones, 200, 217, 218
Branquiocráneo, 59 Brasenia, 424 Brasenia, 424 Brasil, 444
Cabeza, longitud, 395 tamaño relativo, 394
Cadena(s) alimenticias, 128 relaciones, 414
Caja de Skinner, 323 Calcio, 126
carbonato, 160 fosfato, 160
Calcitonina, 333, 337 Calcitonina paratiroidea, 337 California, 445 Callianasa, 411 Caloría, definición, 151
equivalenes, 151 Calorimetría, 160 Camarón, 384 Camarón limpiador, 410 Camouflage, 112 Camptotrichia, 174 Canadá, 437 Canal, cefálico, 367
de la línea lateral, 364 hiomandibular, 364,367 infraorbital, 364, 367 mandibular, 364, 367 suborbital, 364 supraorbital, 364
Canal central, 314 Canal de Suez, 445 Canal Inglés, 446 Canales semicirculares, 97, 99, 359, 360
Canales sensoriales, 364
Canasto branquial, 59 Cangrejo, 411 Cantidades piramidales, 405 Cápsula de Bowman, 80, 249 Capucha oral, 295, 296 Carabela portuguesa, 411 Caracteres osteológicos, 394 Características sexuales, secundarias, 339 Carbaminohemoglobina, 229 Carbonífero, 24
ampolla, 360
cabeza de tiburón, 364

472 ÍNDICE TEMÁTICO
Carne venenosa, 149 Carnívoros, 128, 140,142 Carotenoides, 112 Carpa, 98 Casuarios, 386 Catadromía, 189 Catadromos, 282 Catecolaminas, 337 Cavidad oral, 84 Cavidad peritoneal, 194 Cefalización, 306 Celenterados, 41 1 Celoma, 290 Célula amacrina, 350 CéluIa ganglionar “Parasol”, 351 Celulasa, 148 Células de Mauthner, 317, 366 Células de Purkinje, 313 Células neurosecretoras, 261 Células pilastra, 221 CQlulas sanguíneas, carpa, 210
tiburón, 207 Células sensoriales, 362 Células sensoriales cutáneas, 365 Cenozoico, 24, 387, 430
Centro, 66 Centro de gravedad, 239 Centro de palidecimiento, 322 Ceratophyllum, 424 Cerebelo, 313, 314, 315, 320, 412
natación, 315 raíces motoras, 315 tarpon (sábalo), 315 tiburones, 314
área olfativa, 306 área visual, 311 área visual 11, 312 canal central, 312 carpa, 320 cavidades, 93 cuarto ventriculo, 313 diencéfalo, 31 1 estimulación eléctrica, 313 homología de las partes, 93 lamprea, 93 “loach”, 345 mielencéfalo, 316 pez óseo, 93, 307 subpalio, 307 tallo, 316 telencéfalo, 310 tiburón, 93 tipos, 93 tractos fibrosos, 308 ventrículos laterales, 307 ventriculo ventral, 307
Cerebro anterior, 92, 306 extirpación, 329
Cerebro gemelo, 92 diencéfalo, 311
Cerebro medio, 93,95,96, 297,326 extirpación, 313
Cerebro posterior, 92 Ceruloplasmia en el plasma, 208
temprano, 431
Cerebro, 92, 287
Ciclos biogeoquímicos, 405 Ciclos carbono, 405
fósforo, 405 nitrógeno, 405
Ciclos reproductores, 274 ciclo respiratorio, 217 Ciego pilórico, 84, 140,146, 148, 149 Ciénaga(s), 423
Cintura pectoral, 69 Circulación, 87, 193 Circulación en el sistema, 86 Circulación portal renal, 253
CIadistica, 389 Circulación pulmonar, 86
Clase, 398 Clasificación, 389
pastos, 421
datos de la, 394 definición de la, 5 , 389 ecológica, 406 métodos, 10 objetivos, 10, 390 propósitos, 9 terminología comparativa para el pez, 1 2
Clasificación ecológica, peces, 406 Claustro, 2 3 5 Cloro, 253 Cloruro, 253
células, 258 células secretoras, 261 pérdida, 251
Cobertura de hielo, 407 Cobertura embrionaria, 287 Cobertura esclerótica (capa), 348 Cóclea, 97 Código Strickland, 397 Coladores, 130 Cola, frecuencia de agitación, 181
heterocerca, 20, 28, 54,179, 303 homocerca, 28, 54, 303
Colecistoquinina, 342 Colesterol, 160 Colgajos de piel, 111 Color, ceguera, 353
células, 11 2 fuentes, 112 visión, 354
Coloración, 111 ajustamiento, 356 aposemática o de advertencia, 117,
cambio ajustable del color, 118 cambio fisiológico del color, 118 críptica o de ocultamiento, 114 de contraste destructor, 115, 116 de ruptura coincidente, 119 desfiguración, 114 de estructura, 120 empleo en clasificación, 121 enmascaramiento o desfiguración, 115 imitación del color, 114 imitación protectora, 117 imitación variable del color, 114 marcas deflectivas, 121 marcas directivas, 121
121,173

ÍNDICE TEMÁTICO 473
oscurecimiento, 119 palidecimiento, 119 principio de Thayer, 115 significado, 114 sombreado desvanecedor, 11 5, 11 9 sombreado para la ruptura, 115
Comensalismo, 411 Comisión Internacional de Nomenclatura
Comisura anterior, 306 Competencia, 409 Composición química de los peces, 158 Comunicación, química, 102 Comunidad, 413
Conceptos taxonómicos, 393 Concha, 411 Condrocráneo, 59 Conducta (comportamiento), 323, 334
Zoológica, 398
definición, 403
actividad vacío, 328 condicionamiento, 324 cortejo, 327 mecanismos disparadores, 329 patrones, 329 reproductora, 326
Conducta condicionante, 324 Conducto biliar común, 147 Conducto cístico, 147 Conducto endolinfático, 360 Conducto pneumático, 83,232
abertura, 140 producción de sonido, 236
Conducto pronéfrico, 88 Conductos hepáticos, 147 Conductos urinarios, 252 Conductos urogenitales, 56 Congregación en cardúmenes, 143 Congresos zoológicos internacionales, 395 Cono arterioso, 185,195, 196 Conservación, definición, 6 Consumidores, 405 Consumidores con la vista, 130 Contaminación, organismos índice, 427
Contenido de ácido oléico, 159 Contenido de carbohidrato en peces, 160 Contenido de ceniza de un pez, 160 Continental, deriva, 429, 430
Contrasombreado, 119 Conversión de alimento, 150
eficiencia, 150 Copa óptica, 287 Copépodos, 146 Coral, 130
arrecifes, 420 Corazón, 185,194, 195, 196, 287
atrio, 185 auricula, 185 lamprea, 86 latido, 198 marcapasos, 197 peces óseos, 86 peces pulmonados, 86 ,198 secundario, 203 seno venoso, 185
orgánica, 427
placas, 430
tamatio, 194 tiburón, 89 válvulas, 197 variaciones, 196
asta dorsal, 320 astas ventrales, 320 canal central (epéndimo), 320 raíz ventral, 320 tiburón, 320 tractos fibrosos, 320 tractos nerviosos, 308
Cordón espinal (médula espinal), 317
Corion, 287 Corium, 101 Córnea, 35,346 Corpus striatum (cuerpo estriado), 306 Corpúsculo renal, 251 Corpúsculos de Herbst, 342 Corpúsculos de Krause, 342 Corpúsculos de Stannius, 339 Corpúsculos de Vater-Paccini, 342 Corriente, 408 Corriente del Golfo, 444, 446 Corriente del Labrador, 446 Corrientes(s), fondos, 415
áreas de crianza, 427 características, 425 competencia, 415 fertilidad, 415 gradiente, 415 materia orgánica, 415 peces predadores, 415 pozas, 427 rápidos, 427 saltos, 426 super-enfriada, 407 temperatura, 42 5 valle, 415 zonación, 426
Corrientes en sistemas fluviales, 427 Cortejo, 336
espinocha, 327 Corteza adrenal, 249 Corticoesteroides, 261 Corticoesteronas, 261, 338 Corticoides, gluco, 338
Cortisol, 338 Cortisona, 338 Costillas, 64
dorsal, 66 ventral, 66
Crambessa, 411 Cráneo, 59
perca, 62 Creatina, 253, 257 Creatinina, 257 Crecimiento, 125, 145
determinaciones, 161 determinado, 160 efectos de la aglomeración, 165 en cisco, 164 estacional, 164 factores bióticos, 165 factores estacionales, 164 factores genéticos, 163
mineral, 338

474 ÍNDICE TEMÁTICO
factores luminosos, 164 factores de temperatura, 164 hormona (GH), 333 indeterminado, 160 isométrico, 162 verdadero, 155
Cretáceo, 24 Cría, cuidado de la, 280, 281 Crianza bucal, 292 Criptas sensoriales, 363 Cristu (e) (cresta), 360
stuticue (estáticas), 359 utriculi (utrículo), 359
Cromatóforos, 102, 112 Cromolipoides, 11 2 Cromosomas, autosomas, 378
diploide , 3 81 duplicación, 383 inversión, 383 morfología, 379 número, 379 octopIoide, 384 poliploidía, 384 sexo, 378 supresión, 383 tetraploide, 384 traslocación, 383
Cromosomas del sexo, 377 Cronómetros fisiológicos, 144 Crustáceos, 128
como alimento, 130 parásitos, 412
Cuaternario, 24 Cuerpo, altura, 395
forma, 176 proporciones, 394
Cuerpo comprimido, 176 Cuerpo coroideo, 348 , . cobertura del ojo, 347
b glándula, 220, 348 grado, 348 plexos, 312, 323, 348
Cuerpo de Malpigio, 247 Cuerpo fusiforme, 176 Cuerpo globiforme, 177 Cuerpo lentiforme, 348 Cuerpo pineal, 118, 312, 334
fotosensible, 312 Cuerpos suprarrenales, 249, 261, 337 Cúpula, 359, 360, 362
Chupadores, 131
Darlington, 428, 429 Darwin, 391,428 Deamidación , 2 54 Débiles, 410 Dentículos dérmicos, 106 Dentina, 107 Dependencia de la densidad, 412 Deriva continental, 429 Dermatotriquia, 173 Dermis, 101 ,102 ,345 Desarrollo embrionario, amoníaco, 302
cantidad de vitelo, 302
efectos del gas disuelto, 302 efectos de la salinidad, 302 efectos de la temperatura, 299, 300 etapas, 291 Fundulus hetemclifus, 287 grado, 298,300 nutrición, 293 mganogénesis, 301 respiración, 293
Desarrollo, pez óseo, 298 aletas, 298 embrionario, 283 grado, 299,300 indirecto, 295 larvario, 294 macarela, 296 periodos, 299, 300 posteclosión, 298 transicional, 294
Descenso del punto de congelación, 422 Desfiguración, 120 Deshidratación, 255, 258 Desierto Kalahari, 441 Desierto de1 Sahara, 441 Deslizamiento, 185 Dedizamieno de tierra, 423 Desmidiáceas, 404 Desovadores, 410
lugar, 282 Desperdicios nitrogenados, 247, 257 Despojos flotantes, 416 Detección del clorofenol, 358 Determinación del sexo, 377
aves, 378 mamíferos, 378 peces, 378
Devónico, 24,387 Devoradores de macrocrustáceos, 154 Diadromía, 189 Diadromo, 249, 255, 421 Diástole, 183 Diatomeas, 404
como alimento, 130 Dicromatismo, 271 Diencéfalo, 92, 311,312
tractos nerviosos, 309 Dientes faríngicos, 64
almohadilla, 84,140 Dientes, huesos que presentan, 135, 136,
137,138 caninos, 136 cardiformes, 136 clases, 136 cortadores, 126 faríngicos, 136, 138 garganta, 64 incisivo, 1 3 6 mandíbula, 134 molariforme, 136 moledores, 126 villiformes, 136
Diferencias de sexo, 269 Difusión remolino, 408 Digestión, 125,145 Dimorfismo sexual, gordo de las

ÍNDICE TEMATICO 476
quebradas, 270 dicromatismo, 272 dorado, 270 pez anguloso de mar profundo, 270 salmón rosado, 270 tamaño, 168
Dinoflagelado, 417 Dioico, 384 Dioptrías, 360 Dipéptidos, 149 Diplospondilia, 64 Dipneumoniano, 83 Dióxido de carbono, 221
en la sangre, 229 presión parcial, 227 solubilidad, 229 tensión, 2 2 8
Discriminación sensorial, 324 Disponibilidad de alimento, 166 Distribución bipolar, 447 Distribución, bipolaridad, 447
carpas hociconas, 436 Catostomidae, 436 Cichlidae, 440 Cyprinidae, 439 Cyprinodontidae, 439 discontinua, 434 Esocidae, 433 “killifishes”, 439 local, 429 mundial, 429 pececillos de acuario, 436 pececillos de acuario del lodo, 434 peces marinos, 442 regional, 429 secundaria de los peces de agua dulce.
Umbridae, 434 Distribución geogrbfica, 243,428 Diuresis, 268,337 División abisopeKgica, 418 División epipel6gica. 417,418 Divisi6n mesopelfigica, 418 Divisiones del mar, abisopelbgico, 416
437
batipelágico, 416 béntico, 416 epipelágico, 4 16 mesopelágico, 416 pelbgico, 416
Dominancia de la clase del año, 413 Dominio hadal (infernal), 419,420 Drogas adrenérgicas, 322 Drogas colinérgicas, 322 Drosophila, 382 Duendecillos, 118 Duramadre, 68
Eclosión, 290
Ecoesfera, definid6n, 404 Ecologk, 403
definicionea, 6,403 geografia, 428 sucesión en las corrientes, 41 6
periodo, 299
Ecosistema de agua dulce, 422 Ecosistema, defiicibn, 403
de agua dulce, 422 estuarino, 421
Ecosistema marino, 41 6 Ecotono, 413 Ectodermo, 289, 290 Ectoparásitos, 411 Edad, valoración, 110
composición, 413 determinación, 161
Efecto Bohr, 227 Efecto hipercalcémico ,3 39 Efeméridos, 427 Electrocardiogramas, 197 Elemento proótico, 236 Elemento pterótico, 236 Elementos meristicos, 174
brillo dorado, pececillo, 303 características, 394 fijación, 302 temperatura, 303 trucha arco iris, 303
Elementos nutritivos, 409 Embalses, 422 Embudo oral, 296 Emergencia, 447 Emigración, 413 Emus, 386 Enanos, 410 Endocrinas, control de excreción, 260
secreciones, 260 Endodermo, 289,290 Endolinfa, 97 Energia, calorífica, 161 Energía, conversión, 406
eficiencia, 164 en nutrición, 160
Enfermadades, 410 bacteria, 2 11
Enmascaramiento, 120 Endocrinas, control de excreción, 260
secreciones, 260 Endodermo, 289,290 Endolinfa, 97 Energía calorífica, 161 Energía, conversión, 406
eficiencia, 164 en nutrición, 160
Enfermedades, 410 bacteria, 211
Enmascaramiento, 120 Enteroquinasa, 148 Envenenamiento de la carne, 149 Enzimas digestivas, 146 Enzimas, intestinal, 148 Eosinófilos, 211 Ephemeroptera, 427 Epidermis, 101, 102, 346,362 Epífisis, 367 Epilimnion, 407 Epinefrino, 333 Epitelio, sensorial, 360 Epurales, 63 Equilibrio, 313,369 Equilibrio salino, 269
Equinodennos, 13 peces marinos, 267

476 ÍNDICE TEMÁTICO
Eritrocitos, 185, 208, 210 capacidad de oxigenación, 207 contenido de hierro, 207 en varios peces, 209 número, 207 variaciones circadianas, 210
Eritróforos, 113 Erosión, 408 Escala del tiempo geoIógico, 24 Escama ganoide, 106,107 Escamación, 102,104 Escamas, 362
bajo la línea lateral, 395 caducas, 53 como alimento, 130 deciduas, 53 derivados, 109 determinación de la edad, 110 diversidad, 104, 105 en la piel, 102 en mosaico, 103 formas, 103 historia natural, 109 imbricadas, 103 óseas, 53 patrones, 103 placoidea, 53 usos, clasificación, 109
desarrollo, 1 O 8 estructura, 108 partes, 107, 108 usos, 108
Escamas cicloides, 104 Escamas cosmoideas, 106,107 Escamas ctenoides, 104, 107 Escamas placoideas, 103, 106 Escarabajos derméstidos, 396 Esófago, 84,139,140 Espacio perivitelino, 287 Especies, críptica, 394
agrupamiento, 413 congregaciones, 387 dominancia, 413 hermanas, 393 monotípico, 393 nombre, 396 politípico, 393
Esperma, actividad, 264 congelamiento, 266 espacio de vida, 266 movilidad, 266 refrigeración, 266 saco, 90 viabilidad, 266
anormal, 3 80
Escamas de bordes óseos, 106,110, 108
Espermatogenesis, 264
Espermatozoo, 264,265 Espermiogénesis, 264 Espina, 395 Espina pectoral, 171 Espina de las rayas de espina, 103 Espiráculo, 201, 219, 364 Esqueletización, 396 Esqueleto, 167
aleta, 169
anomalías, 174 apendicular, 67, 69 cintura pectoral, 58 externo, 57 funciones, 167 lamprea, 59 Iamprea marina, 60 mandíbulas, 168 membranoso, 58 Petromyzon marinus, 60 pez óseo, 61 radios de aleta, 172, 173 rarezas, 174 rostro, 168 simetría, 168 Squalus acanthias, 60 tiburón, 60
Esquemacromos, 112 Estanques, 423 Estenohalino, 255, 257,406 Estenotermal, 406 Esteroides córticoadrenales, 333 Estímulo ambiental, 305 Estivación, 141,154 Estómago, 139
distensibilidad, 139 inflable, 139 variaciones, 140
Estradiol, 174, 339 Estrato germinativo, 101 Estrellas de mar, 411 Estriol, 339 Estrógenos, 333,339 Estrona, 339 Estructura, 167 Estructuras sensoriales táctiles, 57 Estuarios, definición, 421 Etapa postlarvaria, 295 Etapas acuáticas de los anfibios, 362
número de especies, 1, 2 Etiniltestosterona, 339 Etiquetado de colecciones, 396 Eurasia, 430,445 Eurihalina, 255,406 Euritermal, 406 Eutroficación, 423 Evolución, 373
adaptativa, 414 convergente, 386 Darwin, 383 definición de la, 5, 383 explosiva, 387 lagos, 414 mecanismos, 381 paralela, 386
Excreción, 247 amoníaco, 260,337 branquias, 257 control endocrino, 249,260 lampreas de agua dulce, 249 nitrógeno, 249 peces marinos, 255 peces óseos, 249
Exócrinos, 147
Facies de agua corriente, 406

ÍNDICE TEMÁTICO 477
Factor condicional, 162 Factores ecológicos, 406
alimento, 409 corriente, 408 densidad poblacional, 412 luz (iluminación), 407 oxígeno disuelto, 408 social, 409 temperatura, 406
Factores socioecológicos, 409 Falla de San Andrés, 429 Familia, 398
Faringe, 84 Fauna antártica, 445,447
Fauna ártica, 445, 446,447 Fauna chilena, 447 Fauna de las costas tropicales, Caribeña, 444
definición, 393
peces, 249
Africa Occidental, 444 Indias Occidentales, 444 Indo-Pacífico, 444 Pacífico-Americana, 444 Panameña, 444
Fauna tropical, barreras, 445 Fecundidad, 267 Fenilalanina, 156 Fenotipo, 383 Feromonas, 358,359 Fertilización, 286 Fertilizantes, 405 Fibras parasimpáticas, 99
sistema, 321 Filamentos branquiales, 221,222
abducción, 219 aducción, 219 embrionario, 225 lamprea, 7 8 mixino, 78 pez óseo, 78 tiburón, 78
hipotgtica, 24 Filogenia, 303,379,390
Fisiología, definición, 6 Fisoclisto, 83, 194, 237 Fidstomo, 83,234,238 Fisóstomos, dominancia en el agua corriente,
246
Fitoplancton, 404, 405 Flavinas, 112 Florida, 446 Fluidos celómicos, 250 Fluidos de los tejidos, 207 Fondo, habitantes, 406
escardador, 140 Formación Delta, 421 Formaldehido, 396 Formalina, 396 Fosa Challenger, 442 Fotóforos, 122 Fotoperiodo, 336 Fotosíntesis, 128,409 Frenos, 180
Ganglio habenular, 311,312
dominancia marina, 245
Ganglios geniculados, 312 Ganglios simpáticos, 98, 322 Gangrena, 124 Ganoina, 107 Garganta, 84 Garzas, 175 Gástrula, 287 Gastrulación, 289 Gene modificador específico, 376 Generación F1, 374 Generación toda femenina, 381 Género, 398
definición, 393 nombre, 397
definición, 5 hibridización, 379 platidorado, 376 población, 392
Genitalia, 386 Genotipos, 383,400 Geografía, peces de agua dulce, 432
peces marinos, 442 Ginogénesis, 26 3, 3 81 Glándula adrenal, 338 Glándula del gas, 239 Glándula pituitaria, pez óseo, 330
Genética, 373
infundfiulo, 330 lamprea, 335 pedúnculo, 330 perca, 335 pez de aletas lobuladas, 330 tiburón perro, 335 tiburones, 330 trucha, 335
Glándulas reproductoras, 89 Glándulas salivares, 145 Glándulas sin conductos, 91 Glándulas suprarrenales, 261 Glándulas venenosas, 123 Glándula ultimobranquial, 146,337 Glicerina, 147 Glicerol, 149 Glicofosfolípidos, 368 Glicógeno, 147 Glicoproteínas, 249
Globina, 225 en plasma, 208
Globo del ojo, 96 Globulinas en plasma, 209 Glóbulos rojos, 147,234
formación, 167 macarela, 209 rape, 209
211 Glóbulos sanguíneos blancos, clases, 210,
ndmeros, 211 Véase también Leucocitos
Glomar Challenger, 429 GlomBrulos, 88, 247, 249,260, 262, 263 Glomus, 250 Glotonería, 144 Glucosa, 249 Golfo de California, 444 Gónadas, 89,249,262,269,261,264,334
catán (peje-lagarto), 262

478 ÍNDICE TEMÁTICO
Gonadotropinas, acción, 340 empleo en el cultivo de peces, 341 FSH, 340 LH, 340
Gonadotropina (PPG) pituitaria ictiológica, 3 40
Gondwana, 430 Gonopodio, 172, 271,272 Guanina, 113, 347 Guanóforos, 113,348 Grandes Lagos, drenaje, 437 Granulocitos, 210, 211 Grasa, 147
absorción, 148 alimentos, 126 almacenamiento, 147 contenido, 158
Gravedad, 361 Gravedad específica, 179 Grupo heme, 226 Gusano de luz, 122 Gusanos del fango, 427 Gusanos luminosos, 122 Gusanos, parásitos, 412 Gustación, 143, 342
estímulos, 346 órganos, 343 papilas, 57,343,344,345,356,358 sensaciones, 346
Gymnodinium breve, 417
Hábitat(s), definición, 403 Haeckel, 391 Hambriento, 144 Heces, 144,248 Hedgepeth, 415 Hematopoiesis, 167,211 Hemibranquia, 217 Hemisferios cerebrales, 92 Hemoglobina, 83,147,207,225
contenido, 213 saturación, 226
Hemolisis, 209 Hepatopáncreas, 147 Herbfvoros, 128,404
tilapia, 148 Herencia, 373
albinismo, 374 estudios citológicos, 379 “guppy”, 375 matroclina, 381 mendeliana, 376 no mendeliana, 374 ligada al cromosoma sexual, 377 limitada al sexo, 375,376
Hermafroditismo, 263 prottindrico, 263
Hermano(a), 375 Heterocigosidad, 375 Heterófilos, 211 HeterogamBtico, 378 Hetemquiridio, 271 Heterosis, 380 Hexagenia, 427 Hibernación, 406
Hibridación, 379, 386
Híbrido, cruzas, 378 éxito, 381
natural, 4 1 O vigor, 380
Hidrocortisona, 252 Hidrosfera, 404,405 Hidrostasis, 237, 238 Hierba acuática córnea, 424 Hierbas de estanque, 424 Hígado, 146 Himalaya, 429 Hinchamiento, al alimentarse, 130 Hipertónico, 255
medio, 248 urea, 259
Hiperventilación, 338 Hipocalcemia, 337 Hipocerca, 20 Hipofisectomía, 261, 340 Hipófisis, 312, 330,331, 335
Hipolimnion, 407 Hipotálamo, 249,261, 312
Véase también Glándula pituitaria
alimentación, 144 tejidos neurosecretores, 331
Hipural( es), 6 5 Histidina, 156 Historia evolutiva, 390 Historia geográfica, 430 Historia natural, definición, 6 Hocico, longitud, 395 Holobranquia, 217, 219
musculatura, 219 Holothuroidea, 411 Holotipo, 400 Homeostasis, 247,312
mecanismo, 256 Homocigótico, 383 HomogamBtico, 378 Hongos, 412 Hormona adrenocorticotrópica, 338 Hormona folfculo-estimulante (FSH), 340 Hormona luteinizante (LH), 340 Hormonas, 118,148
acción, 249 actuadoras en la melanina (MAH), 331 adrenocorticotrtipica (ACTH), 338 antidiurética, 336 córticoadrenal, 261 estimuladora de la melanina (MSH), 331 gonadotropina, 339 intermedina, 334 mucosa intestinal, 341 neurohipofisiaria, 336 sexuales, 272 tiroides, 336 tubérculos nupciales, 271 vasopresora, 336 Véase también Sistema endocrino
Huesecillos de Weber, 361 Hueso maxilar, 30 Huesos intermusculares, 66 Huevos, adherentes, 269,277
apéndices, 269

ÍNDICE TEMATICO 479
áreas formadoras de órganos, 289 bacalao, 277 cáscaras, 268 cordón, 282 cuidado de, 267,279,280 flotantes, 268,277 formación, 266 globo, 282 gobio, 265 masas, 282 no esféricos, 265 pelágico, 174, 268, 282 pez volador, 277 reabsorción, 269 sumergibles, 269 transporte paternal, 279 viabilidad, 269
Humor acuoso, 350 Humor vítreo, 350
1131 *, 336 Ictioacantotoxicidad, 123 Ictiogeografía, 403 Ictiología, definición, 9 Ictiosarcotoxicidad, 123, 149 Ictiotoxicidad, 123 Illicium, 121 Impregnación, 285 Ión, absorción, 254
difusión, 248 equilibrio, 250 transferencia, 248
Inanición, 154 India, 430
subcontinente, 430 Indias Occidentales, 438 Indice ponderal, 172 Indigoides, 112 Indo-Pacífico, 444 Información de la colección, 396 Infundíbulo, 335 Ingestión de agua, 247
agua marina, 250 Inmigración, 413 Inmunoglobulina, 212 Insectos de piedra, 427 Instinto, reproductor, 327 Insulina, 147,333 Integración, 305 Integumentario, 57 Inteligencia, 323 Intermedina, 333 Interorbital, 395 Intersexualidad, 263 Intestino, carnívoro, 142
hervlloro, 142 longitud, 142 perca, 142 pez gato herbívoro, 142 superficies, 144 tiburón, 142 variaciones, 142
Intestino grande, 84 Intestino pequeño, 84 Intraocular, acomodación, 346
presión, 221 tensión de oxígeno, 348
Introducción de peces, 410 Invertasa, 146 Iridocitos, 11 2 Islotes de Langerhans, 147 Islotes pancreáticos, 148, 334 Isoleucina, 156 Iso-osmótico, 257 Isotipo, 400 Istmo de Panaml, 444 Istmo de Tehuantepec, 437
Japón, 445 Java, 441 Juncos, 421 Juncus, 421 Jurásico, 23,24
Ketosterona, 339 Kinocilia, 368 Kiwis, 386 “Kype”, 272
Labios, 55 carnosos, 131 en la alimentación, 130 sobresalientes, 168 suctores, 131
Lactasa, 146 Lagena, 97,360,361 Lago Baikal, 4,128 Lago Superior, 128 Lagos, 422
alcalinidad, 41 5 alimento, 414 clasificación, 423 cráter, 423 dióxido de carbono, 414 distrófico, 423 en arco, 423 eutrófico, 423 evolución, 414 fertilidad, 422 glacial, 423 intercambio ascendente, 407 mesotrófico, 423 Norteamérica, 414 oligotrófico, 422 orígenes, 423 oxígeno, 414 oxígeno disuelto, 423 peces de agua caliente, 414 peces de agua fría, 414 sucesión, 423 suelo, 422 temperatura, 44 transparencia, 414 vegetación, 414 zonación, 424
Lagos templados, 406 Lagos tropicales, 407 Lagunas, 423

480 ÍNDXCE TEMÁTICO
Lámina krminal, 307 Lamprea marina adulta, 295 Lampreas parásitas, 175 Lampyridae, 122 Langostas de río, 427 Lapillus, 98, 359, 361 Lapsus calami, 399 Larva, ammocetes, 296
anguila, 307 eclosión, 287 lamprea, 9 lamprea marina, 295 saltarín del fango, 261
Larva leptocéfala, 118, 158, 190, 209 Larvas de insecto, 128
díptero, 262 Laurasia, 430 Lectotipo, 400 Lecha, 266 Lengua, 146 Léntico, 422 Lepidofagia, 130 Lepidotriquia, 173 Leucina, 156 Leucocitos, 85, 210 Leucocitos polimorfonucleares, 212 Ley de Boyle, 239 Ley de Prioridad, 399 Ligamentos, 58 Limnodromia, 189 Línea alba, 58 Línea lateral, 54, 57,362, 367
anguila, 362 cabeza de pez de aleta ondulada, 363 cabeza de pez óseo, 367 canal, 362, 363, 366 descargas nerviosas, 368 distribución de los canales, 367 órgano, 344, 365 plácodas, 363 poros, 367 recepción, 368 sistema, 97, 369
Linfa, 206, 207 conductos, 149 corazón, 206 nódulos, 206 senos, 206 sistema, 206, 207 troncos, 206 vasos, 205
Linfocitos, 185, 210, 211 Linnaeus, 390, 391, 395 Lipasa, 209 Lipoproteínas, 208 Líquido cefalorraquídeo, 312 Lisina, 156 Litosfera, 429 Lóbulo(s) óptico, 312, 320 Locomoción, 174,176
aletas, 178,179,181 anguiliforme, 181, 182 costos energéticos de la natacibn, 183 migraciones, 189 miómeros, 181
movimiento del cuerpo, 182,183 natación, 175, 181, 182 no natatoria, 188 pasiva, 174 velocidad, 187
Longevidad de los peces, 162 Longitud cefálica postorbital, 395 Longitud de la escotadura caudal, 395 Longitud del ojo, 395 Longitud patrón (estándar), 395 Longitud predorsal, 395 Longitud total, 394 Lótico, 422 Lucha o vuelo, 338 Luminiscencia, 122 Luminoso, bacteria, 122
células, 122 Luz biológica, 122 Luz, coloración, 407
alimentación, 355 alimento, 408 crecimiento, 407 equilibramiento, 361 glándulas endocrinas, 334 migraciones, 407 movimientos, 407 nivel de compensación, 424 órganos, 408 visión, 355
Lluvia de peces, 174
Mac Arthur, RH., 428 Machos parásitos, 175, 271 Maduración, sexual, 273 Madurez, 60 Madurez sexual, 89,163, 273 Mamíferos, 161
número de especies, 1, 2 presa, 126 tiempo geológico, 24
Manantiales, 422 caliente, 427
Manantiales de agua caliente, 427 Mancha sensorial acústica, 361 Mandíbula superior, 54
longitud, 394 Mandibulas, 168
elongación, 168, 169 evolución, 168 inferior, 168 más superior, 30 superior, 168
Mangle, 421 Maniobra, 179 Mar, alga, 117
pepinos, 411 tortugas, 175, 411 trincheras, 411
Mar Mediterráneo, 445 Mar Rojo, 445 Mar de Tetis, 445 Marcas anuales, 161,162
escamas, 107 Marcas de reposo, 114

ÍNDICE TEMÁTICO 481
Marea roja, 417 Mariposas melanóticas, 384 Materiales hereditarios, 264 Matthew, 429 Maxila(s), 29, 30 Mecanismo de respiración, 224 Mecanismos liberadores, 329 Medio ambiente, béntico, 419
definición, 403 pelágico, 416
Medio hipotónico, 248 Médula, 313, 315, 316,317, 357
lóbulo facial, 320 lóbulo vago, 320
Medusa, 416 Melanina(s), 112
hormona concentradora (MAH),
hormona dispersante (MDH), 119 síntesis, 336
Melanóforo(s), 113 nervios, 313
Melanogenesis, 336 Membrana oral, 254 Memoria, 324
migración, 324 olor, 324
Mendel, 392 Mendeliana, dominante, 376
119
descendencia, 373 herencia, 382 herencia limitada al sexo, 375 radio, 374 recesivo , 3 74
Mesencéfalo, 312 tractos nerviosos, 309
Mesoadenohipófisis, 330 Mesocoracoides, 29 Mesodermo, 290 Mesonéfrico, conducto, 247, 252
Mesonefros, 89 Mesozoico, 20,24,25,27,430
tardío, 431 Metaadenohipófisis, 330 Metabolismo, 144
aeróbico , 184 agua, 334 carbohidratos, 334 desperdicios, 247 grado, 151,228 grasa, 334 mineral, 334 proteína, 334
Metamorfosis, 190 Metencéfalo, 93, 313
Metionina, 156 Métodos, conteo, 396
México, 437 “Midges”, 427 Mielencáfalo, 93, 316
túbulo, 250, 251
tractos nerviosos, 308
medición, 395
miómero, 58,71,345 tractos nerviosos, 308
Miembros pentadáctilos, 1
Migraciones, impronta de los olores, 3 24
factores influyentes, 190 memoria, 324 sincronización, 259
Mineral(es), equilibrio, 261 función estrucural, 158 función metabólica, 157 función respiratoria, 158 nutrición, 157 rastro, 158
Mioceno, 387 Miosepto, 58, 70, 71, 345, 362 Miotomo(s), 70 Miriofilo, 432 “Mitrates”, 13 Molleja, 140,146 Mollusca, 128 Monocitos, 211,212 Monopulmonado, 83 Monorrino, 16,56 Monstruosidades, 174 Montañas Rocosas, 437 Monte Everest, 4 Mortalidad, 412 Mortandad de invierno, 409 Mortandad de peces, 409 Mortandad de verano, 409 Moscas de la fruta, 382 Movimientos, cortejo, 327 Mucosa, intestinal, 148 Mucoso, células, 53
glándula, 101,123 Mucus, 102 Muerte, 298, 405, 409 Multiplicador de contracorriente, 239 MÚsculo(s), 70
aleta caudal, 76 aleta media, 75 aleta par, 75 aleta pélvica, 76 apendicular, 72 branquiomérico, 72 cabeza, 74 corazón, 77 cardíaco, 70, 77 eficiencia, 184 lamprea, 71 lateral, 71 liso, 70 oculomotor, 72 perca, 71 salmón chinense, 74,75, 76 segmental, 70 tiburón, 71 tono, 313
Músculo retractor del cristalino, 349 Músculos lisos, contracción, 340 Músculos oculomotores, 96 Museo(s), Británico, 6
Academia de Ciencias de California, 6 colecciones, 395 Museo de Zooloaia de la Universidad de
Michigan, Nacional de los Estados Unidos. 6
Mutación, 375, 382

482 ÍNDICE TEMÁTICO
cromosoma, 382 velocidades, 382
Mutación genética, 282 Mutágenos, 383 Myriophyllum, 424 Myxopterygia, 16, 55,172, 271, 272
Nado libre, 406 Nasales, orificios, 54, 55, 56, 57,94,
excurrente, 56, 57,345, 357 incurrente, 56, 57, 357 interno, 17 posterior, 367
Nasal, cápsula, 357, 364 saco, 356 tubo, 356,357,367
Natación anguiliforme, 182 Natación carangiforme, 182,183 Natación, velocidad crítica, 184
distancias, 176 eficiencia, 177 esfuerzo de natación, 184 inversión de posición ("de cabeza"),
238,360 Natalidad, 412 Navegación, 177 Nefrostoma, 251 Nelson, 430 Nematocistos, 411 Neotenia, 296 Neotipo, 399 Nervio auditivo, 361
Nervio vago, 317 Nervios craneales, 93, 307, 315,364
placoda, 287
conexión central, 319 conexión periférica, 324 cutáneos, 345 descargas, 343,368 espinal, 317 función, 3 19 nombre, 318 número, 319 olfatorio, 345 óptico, 312, 348 terminaciones libres, 343 terminal, 306
Nervios espinales, 94, 315, 317 diferenciación, 317 disposición, 3 17
Neurocráneo, 16,58 Neuroglia, 323 Neurohipófisis, 261,330, 331 Neuromasto(s), 362,366,360 Neuronas de Mauthner, 317 Neutrófilos, 211 Newell, 430 Nicho(s), 421
Nidos, cabeza de toro, 27 8 definición, 403
burbuja, 283 construcciones, 281,326 constructores, 281, 326 desertores, 281 diversidad, 281
espinocha, 278 lamprea, 278 pez belicoso, 278 pez lanceta Johnny, 278 pez sol, 278 protegidos, 281, 282
Ninfas de caballito del diablo, 427 Ninfeas, 424 Nitrógeno, excreción, 147 Nivel(s) trófico, 128
primario, 404 Nombre genGrico, 399 Nomenclatura, 389
binomial, 397 botánica, 400 Código Internacional, 397 definición, 397 jerarquía, 398 Ley de Prioridad, 399 objetivos, 397 primer revisor, 400 zoológico, 397 Véase también Sistemática
Noradrenalina, 337 Norepinefrina, 333 Notocuerda, 16,58,287, 290 Nuca, 395 Núcleo preóptico, 261 Nucleótido, difosfato, 358
monofosfatos, 358 Nueva Zelanda, 430,446 Nuevo Mundo, 444 Nuphar, 424 Nutrición, embrionaria, 293
requerimientos, 152 Nutrición intrauterina, 132 Nutrientes solubles, 125 Nutrimentos disueltos, 412 Nymphaea, 424
Occipital, 395 Oceanodromía, 189 Ocultamiento, 120 Odonata, 427 Oído, 97
interno, 97,359, 360 otolitos, 97 perilinfa, 235 tímpano, 97
Oído interno, 99,333 ciprinoideo, 99 laberinto membranoso, 99 sáculo, 234 salmonoide, 99
Ojo, 93, 95, 346,367 acomodación, 349 anatomfa seccional, 95 bastón, 351,352,355 célula bipolar, 351 cobertura coroidea, 347 cobertura esclerótica, 347 cono, 351,352,355 córnea, 95,346 cristalino, 95, 312,349 esclera, 347

ÍNDICE TEMATICO 483
humores, 95 iris, 95,348 ligamento suspensor, 349 longitud focal, 349 mdsculo, 95
retina, 312, 350,351 peces pulmonados, 353
trucha, 318 Olas, 408 Olfación, 143,307, 356
impulsos, 306 órgano, 94 sentido de, 358
Olfatorio, bulbo, 310, 357 anguila, órgano de la, 357 discriminación, 358 epitelio, 356, 358 fibras, 311
lóbulo, 320 lamprea, órgano de la, 357
nervio, 356 órgano, 94,358 placoda, 306
Olores, de pez, 102 Omento, 147 Omnfvoro, 138 Ontogenia, 303 Opercular, anillos, 162
bomba, 222 válvula, 222
Opérculo, perca, 162 Orden, 398 Orden de picoteo, 165 Ordovícico, 21, 24, 26 Organismos heterotróficos, 425 Organización de las Naciones Unidas para la
Órgano copulador, 271 6rgano(s) intromitente, 53, 55, 271
gambusia, 272 tiburón cornudo, 272
@gano palatal, 317 Organo parapineal, 311 &gano pineal, 311 Qrganogénesis, 290 Qrganos adherentes, 54, 55 Qrganos aperlados, 271 @ganas de Fahrenholz, 365 Organos del equilibrio, 97 drganos eléctricos, 365 @ganas endocrinos, 91,92,261,329,334 Organos epifísicos, 311 drganos espiraculares, 365 Órganos excretores, 247 Órganos formadores de sangre, 207 Orientación en el espacio, 313 Orientación espacial, 313 Orificios nasales, 16, 17, 56, 94, 357 Orina, 248
Alimentación y la Agricultura (FAO), 6
amoníaco, 257 composición, 251, 257 contenidos, 253 flujo, 259 hipertónica, 248,250,259 hipotónica, 248, 250, 259
producción, anfibios, 260 producción, hombre, 261 urea, 257 volumen, 250, 251,253, 256, 259
Ortografía original, 399 Osmorregulación (regulación osmótica), 247
carpa dorada, 248 lampreas de agua dulce, 248 mantarrayas, 259 órganos, 247 peces de agua dulce, 248 peces diádromos, 258 peces óseos, 248 peces óseos marinos, 248,255 rayas, 259 tiburones, 259
Ósmosis, 248 Osteichthyes, grupos superiores, 26, 27, 28,
Otolito(s), 97,359, 360, 361
Ovario cistoario, 91 Ovario gimnoario, 91 Ovarios, 90,91, 252,264
función endocrina, 339 Oviductos, 91 Ovopositor, 272 Ovopositores, 91,277 Ovotestículos, 89 Óxido trimetilamino, 257,259 Oxígeno, capacidad sanguínea, 225,226,
adquisición, (carga), 226 adquisición en las branquias, 228 consumo en el metabolismo, 228 curva de disociación, 226 curva de equilibrio, 226 deficiencia, 408 disuelto, 408 eliminación (descarga), 226 nivel letal, 228 presión, 226 presión parcial, 226 requerimientos, 408 saturación, 408 tensión, 228 transporte, 207
Oxitocina, 333,336
Pacífico del Norte, 445 Paleozoico, 24,430,431 Panamá, 437 Páncreas, 147
Pancreozimina, 333,341 Pangea, 430 Pantanos, 428 Parásitos, 132,412
Paratipo, 400 Paratopotipo, 400 Parencéfalo, 3 11 Párpados, 95 Partenogénesis, 363,384 Partes, peces, con espinas y radios, 395 Pececillo(s) de acuario, oído interno, 98
29,30
determinación de la edad, 359
secreciones, 148
obligatorios, 132

484 ~ N D I C E TEMÁTICO
Peces cavernícolas, 386 Peces comerciales, 445 Peces de agua caliente, 407, 414 Peces de agua dulce, 422
Africa, 438 América del Norte, 438 distribución continental, 437 Eurasia, 441 Europa, 441 geografía, 432 grupos, 437 primario, 437,438 Región Oriental, 441 Rusia, 441 secundario, 437 Sudamericano, 437,438
Peces de agua fría, 407, 414 Peces del fondo, 406 Peces marinos, ab ides , 442
bancos de arena, 442 batipelágicos, 442 bénticos, 442 de mar abierto, 442 de mar profundo, 442 pelágicos, 442 ribera, 442 ribera tropical (costa tropical), 442
Peces operculados, 221 Peces óseos, evolución de teleósteos, 29
Peces ovíparos, 91,268 Peces ovovivíparos, 91 Peces parafiioclistos, 238 Peces parásitos, 126 Peces venenosos, 123,149 Peces voladores, 361 Pedúnculo caudal, altura mínima, 395
longitud, 395 Pelágico, 406
medio ambiente, 416 Penúltimo, 64 Pepsinógeno, 146 Peptidasas, 146 Pérdida de sal, 251 Periblasto, 290 Pericardio, 58, 194 Pericondrio, 59 Perifiton, 409 Perirnisio, 58 Perineum, 58 Peritoneo, 58 Peritoneo parietal, 58 Perizoon, 409 Pérmico, 24 Perú, 444 Peso, factores de conversión, 150 Pesquería, 445 Pez(ces), anatomía, 51
ancestros de, 11 ancestros del hombre, 2 caracterización de los grupos vivientes,
cultivo, 341 definición, 1 de los peces, 410
grupos superiores, 26, 30
15, 16, 17
distribución vertical, 3, 4 evolución, de los peces óseos, 26,27, 28
29,30 del grupo de los tiburones, 25, 26 de los Chondrichthyes, 25,26 de los de aletas con radios, 28 de los Osteichthyes, 26,27, 28,29, 30
familias de los vivientes, 30-48 filogenia, 24 grupos de los vivientes, 15, 16, 17 grupos extinguidos, 17 grupos superiores, 12 grupos superiores representativos, 14 número de, 1, 2 primeros mandibulados, 22 primitivos, 21 tiempo geológico, 24
Pez migratorio, 261 Pez pulmonado, capullo, 233
estivación, 233 pulmón, 232
en el intestino, 147 PH, 228
Phoxinus, 324 Phylum, 398 Physalia, 411 Piamadre, 58 Piel, 102
estructura, 101, 102 receptores, 342
Pigmentación, 334 Pigmentos fotosensibles, 356 Píloro, 140 Piridoxina, 157 Piscívoros, 153 Pituitaria, 118, 249, 259, 334, 339, 357
desove inducido, 341 extirpación, 340 hormonas, 331 implantaciones, 341
Placa-branquid, 79 Placa gular, 174 Placas óseas, 103 Plancton, 409
consumidores, 148,159,424 filtración, 127
Planeo, 185 Plano focal, 350 Plantas, emergentes, 424
flotantes, 424 sumergidas, 424
coeficientes de sedimentación, 208 composición electrolítica, 208 disminución del punto de congelación,
enzimas, 209 peces del Antártico, 209
Plasma, 85, 225
208
Plasma sanguíneo, 207 Plecoptera, 427 Pleistoceno, 387 Plica valvulae, 315 Plioceno, 387 Población, colonización, 413
definición, 403

ÍNDICE TEMÁTICO 48 5
densidad, 412 estabilidad, 412 estructura, 412 regresión, 412
Poiquilotermo, 150,161, 213 Polinesia, 442 Polipéptidos, 331 Poliploidía, 384 Polizontes, 175, 411 Pómez, 416 Porfirinas, 112 Porfiropsina, 352 Poros abdominales, 90 Potamodromía, 189 Potamogeton, 424 Pozas, 406
Predación, 409 Predadores, 130 Pregneninolona, 339 Presa, 409 Presas, 422 Presente, 24 Preservando peces, 396 Presión osmótica, anguila, 256
anguila marina, 256 tiburón, 256
337,340
en corriente, 426
Presión sanguínea, 185,186,199,260,
F’roadenohipófisis, 331 Proceso falciforme, 348, 349 Procesos pélvicos axilares, 174 Producción de sonido, 235, 236
estridulador, 236 hidrodinámica, 236 peces de mares profundos, 237 vejiga gaseosa, 236
Producción primaria, 404 Progesterona, 339 Prolactina, 333, 334 Pronefros, 88, 252 Propioceptores, 343 Proporción de sexos, 380 Propulsión a chorro, 185,187 Prosencéfalo, 93 Prostaglandinas, 340 Proteasas, 148 Proteína(s), 149
Protocooperación, 411 Protocordados, 11 Prudente, 324 Pseudobranquia, 217,219,220
función endocrina, 334 pez de ojos parietales, 220
contenidos en los peces, 160
Pterigióforos, 67 Petrinas, 11 2 Pulmón, 83,229,232
bilobulado, 232 origen embrionario, 233 vejiga gaseosa, 232
Purinas, 112, 113
Quiasma óptico, 312
Ración diaria de alimento, 154
Ración semanal, 154 Radiación adaptativa, 387 Radio, 391 Radiotiroidectomía, 336 Radios blandos, 395 Radios branquióstegos, 220, 222 Radios de aleta, varillas cartilaginosas,
ritmos, 338
173 dermatotriquia, 173 dérmico, 174 espinas, 172, 173 números, 394 radios blandos, 172, 173 radios espinosos, 138, 172, 173 radios segmentarios, 172, 173
Radios rudimentarios, 395 Ramo recurrente, 345 Ramoneadores, 130 Ranas, como alimento, 126 Rápidos, 406
Ratites, 387 Rayos X, 383 Reabsorción de gas, 238 Recepción del estímulo, acústico, 342
en corrientes, 427
calorífico, 342 del dolor, 342 del gusto, 342 físico, 342 olfativo, 342 químico, 342 táctil, 342 visual, 342
Recepción térmica, 344 Reclutamiento, 409 Reconocimiento sexual, 121 Reflejos, 320
condicionados, 324 de enderezamiento, 360
Región abdominal, 395 Región del Titicaca, 4 Regurgitación, 146 Relación longitud-peso, 162 Reposición de la sal perdida, 251 Reposo, 338 Reproducción, 280,384
bisexual, 112,384 papel de la, 264 partenogenética, 263 tipos de, 263
como presa, 126 Reptiles, número de especies, 1, 2
Resacas en corrientes, 426 Respiración, 2 15
embrionaria, 293 Respiración aérea, 229
adaptaciones, 231 anguilas espinosas, 230 branquias, 229 cabeza de vibora, 230, 231 cobítidos, 230 divertículo oral, 230 facultativa, 234

486 ÍNDICE TEMÁTICO
obligada, 229 pared de la cámara branquial, 230 peces gato acorazados, 230,231 peces gato marchadores (laberínticos),
230,231 peces gato sacobranquiales, 230, 231 perca trepadora, 231 tracto intestinal, 230
Respuesta, condicionada, 323 Rete mirabile (red maravillosa), 194, 239,
240,241 área de los vasos sanguíneos, 239
Retina, 221, 350, 361 adaptaciones a la luz-oscuridad, 359 capas, 351,352 componentes neurológicos, 351 pez óseo, 352 salmón, 351
Retinoscopía, 349 Reversión genética, 376 Rheas, 387 Rhizophora, 421 Riboflavina, 157 Rifión(es), 87,88,252
catán (peje lagarto), 252 función, 256 formación de sangre, 211 lamprea, 250 mesonéfrico, 250 nefrón, 252 peces de agua dulce, 251 pez marino, 256 pronéfrico, 250 túbulo, 247 unidad, 251
Río Mississippi, 437 Roedores como alimento, 126 Rodopsina, 352 Rombencéfalo, 93 Rusia, 443
Saciedad en la alimentación, 144 Saco, vascular, 335 Sáculo, 97, 146, 359, 360
dorsal, 311,320 Sagitta , 3 3 3 Salinidad, 336 Saltadores, 186 Salto, 185 Sangre, 185,193,207
calcio, 262 células, 207 como transportadora de gas, 224 como dióxido de carbono, 228 contenido de hemoglobina, 207 formación, 211 glóbulos blancos, 210 glóbulos rojos, 208 glucosa, 338 plasma, 208 presión osmótica, 256 sistema B, 208 sistema Rh, 208 tipos, 208 y enfermedades, 212
Sapos, 24 Scaphium, 226 Scientific American, 429 Scirpus, 424 Schreck stoff(e), 102, 358 Secretina, 333,341 Sedimentación, 426
en las corrientes, 427 Segmentación, 286 Selección natural, 382, 383 Seno urogenital, 88,250,252 Seno, venas oftálmicas, 348
Sensibilidad a la presión atmosférica, 236 Sentidos, 342
cápsulas para los órganos de los, 69 cutáneo, 342 epitelio sensorial, 360 olfato, 356 órgano, 342
venoso, 195,196
Sentidos químicos, 143 Septo esqueletógeno, 58,71 Septos interbranquiales, 78 Serpenteo, natación por (crawling), 184 Servicio Marino Nacional de Pesquería, 6 Siberia, 432 Silúrico, 24, 387 Simbiosis, 410 Simbiosis de limpieza, 410 Simetría, bilateral, 168 Simpson, 429 Síndrome de tensión adrenal, 338 Sintipo, 400 Sistema acústicolateral, 100, 360 Sistema endocrino, 290,305,329
acciones, 259 glándulas, 332 interacción glandular, 334 orígenes embrionarios, 332 partes, 332 secreciones, 332,333
Sistema hemolinfático, 206 Sistema nervioso, 305
automático, 321 divisiones, 91 tejidos de soporte, 322
Sistema nervioso au thorno , 99,321 Sistema portal, hepático, 194
Sistema reproductor, 263 Sistemdtica, definición, 389
Sistemático, 390,392 Sístole, 85 Sobrehidratación, 258 Sodio, retención, 262
presencia, 262 Sol, energía, 404 Somites, 287,290 Sonido, percepción, margen superior
audible, 361 recepción, 361
renal, 194
conformidad en los peces, 390
Soportes de aleta, dorsal, 67 Spartina, 421 Strombus, 411

ÍNDICE TEMÁTICO 481
Subclase, 398 Subcutis, 102 Subespecie, 399 Subgénero, 399 Sublitoral, zona interior, 419
Suborden, 398 Subphylum, 398 Suero, 209,260
calcio, 339 osmolalidad, 337 proteínas, 249
zona exterior, 420
Sumatra, 441 Sumergible, 159 Superclase, 398 Superfamilia, 398 Superorden, 398 Supresión, 261 Sureste de Asia, 432 Sustancia de aglomeración, 166 Sustancia de alarma, 102 Sustancias promotoras del crecimiento, 166 Systema Naturae, 391
Tacto, discriminación, 343 crianza, 130 receptores, 342 sensor, morena, 345 sentido del, 342
Tálamo, 312,334 Talos, 120 Taxonomía, computadoras, 389
definición, 389 histórica, 389 numérica, 389
Tecto óptico, 312,314,315 Tectónica, global, 429
Tectónica de placas, 431 Tejido corticoadrenal, 338 Tejido cromafínico, 334, 337 Tejido inter-renal, 334, 338 Tejido medular, 338 Tejido suprarrenal, 334 Tela coroidea, 323 Telencéfalo, 93, 306, 315
placa, 429
funcioues, 307 tractos nerviosos, 308
Temperatura, 406 cuerpo, 213 desarrollo, 298 desarrollo embrionario, 298 dispersión de los peces, 407 endocrinas, 334 ingestión de alimento, 154 logro de alimento, 154 metabolismo, 154 órganos en terminaciones nerviosas, 344 percepción, 342 ración de mantenimiento, 154 sentido, 342
control, 213 sobre la ambiental, 213
Temperatura del cuerpo, 150,213
Tenaculum, 16
Tendones, 58 Tentipedidae, 427 Teonina, 156
Teoría del pliegue-aleta, 170 Terciario, 24 Termoclina, 407 Terremotos, 429 Territorio doméstico, 312 Testículos, 89, 252,264
prepélvico, 16
en el plasma, 208
conductos, 90 funciones endocrinas. 339
Testosterqna, 339 Tétanos, 123 Tiflosole, 84,146 Timo, 146,334 Tipo, 400 Tiroides, 147,249,259,261,334
calcitonina, 337 funciones, 337 hiperactividad, 261,337 hormona, 158,260,333 hormona tirotrópica, 333
Tiiosina, 113 Tiotropina (TH), 333 Tiroxina, 336 Tolerancia salina, 255, 258 Tolerancia térmica, 428 Tornaria, 11 Tortugas, 1,416
como alimento, 126 Tracto digestivo, 138,140
tiburón, 205 trucha, 205
Transferencia óptica, 313 Triásico, 13,24,25,27 Tribu, 399 Tricópteros, 427 Trichoptera, 427 Trinchera Kuriles-Kamchatka, 443 Triptofano, 156 Tripus, 235 Triyodotironina, 336 Trombocitos, 211 Tronco arterioso, 16,194 Trucha, corazón linfático, 206
corrientes, 415 Tubérculo impar, 316 Tubérculos nupciales, 270 Tubificidae, 427 Tubo neural, 290 Túbulo convulsionado, 250 Túbulo pronefrico, 250 Túbulos uriníferos, 251 Tule, 424 Turbidez, 408 Typha, 424
Urea, 147,253,257,259 Uretra, humana, 412 Urohipófisis, 261,334 Urosoma, 52 Urostilo, 64

ÍNDICE TEMÁTICO
. T L.S. Fish and Wildlife Service
de EUA), 6 (Departamento de Pesca y Vida Silvestre
1J.S. National Marine Fisheries Service (Departamento Nacional de Pesauerías
Marinas de EUA), 6 U.S. National Museum (Museo Nacional de
EUA), 6 UtrícuIo, 98, 359, 361
Vaina pericordal, 58 Vaina perineural, 58 Valina, 156 Valvula cerebelli, 314, 315 Válvula espiral, 142 Válvula oral, 222 Válvula pilórica, 84 Valle de la Muerte, 428 Variación, 382
geográfica, 393 Vas deferens, 90 Vasa efferentia, 252 Vasodilatación, 338 Vasos circulatorios, 194 vasos sanguíneos, 86,287
branquia, 198,199,200 cabeza, 198 cuerpo, 201,204,205 plan, 87 tiburón, 204 tracto digestivo de tiburón, 205 tracto digestivo de trucha, 205 trucha, 204
Vejiga gaseosa, 83 vejiga(s) gaseosa, 77,180, 194,233, 287
almacenamiento de grasa, 244 Amia, 80 audición, 235, 362 ausencia, 243 catán (peje-lagarto), 80 conductor del sonido, 235 contenidos, 239, 240 distribución en los peces, 243 ecología de los peces, 243 estructura, 242 esturión, 80 extensiones, 234 fidstomos, 80 forma, 235 funciones, 233 glándula de gas, 242 llenado, 238 nitrógeno, 238 órgano oval, 243 órgano respiratorio, 233 oxígeno, 268 perca trepadora, 80 pez boca, 242 pez de dientes oblicuos, 240 pez pulmonado, 80, 232 posición, 240 pulmón, 232 pulmón de pez pulmonado, 232 producción de sonido, 236 recepción de sonido, 235
resonador, 235 saculaciones, 23 2 sardina, 234 secreción gaseosa, 241 suministro sanguíneo, 234, 238, 242 tamaño, 235 vaciado, 238 vestigial, 159
Vejiga natatoria, 83 Vejiga urinaria, 88, 253 Velocidad de especiación, 387 Velocidad, navegación, 187
máxima, 187 recorrido, 187
retinal, 348 sistema, 203
Veneno, peces, 123 escape, 53, 54, 55, 56 glándulas, 123 Véase también Ano
Ventriculo, 195, 196 Ventriculo cerebelar, 315 Vértebra(s),,anillo, 162
Venas, cabeza, 201, 202
centro de, 26 números, 394 pez gato, 162 tronco, 65
Vértebras caudales, 64, 65 Vesícula biliar, 47 VesícuIa seminal, 90 Vesículas de Savi, 363 Vmoras, 126 Viejo Mundo, 432 Visión, 346, 354
bastones y conos, 355 binocular, 355 color, 325 foco, 349 fotoquímica, 353 luz, 355 salmón, 355
almacenamiento, 147 B, 157 C, 157 Complejo B, 157 D, 147,157 E, 157 K, 157 nutrición, 157 síntomas de deficiencia, 157
Vitamina(s), A, 113, 147, 157, 352
vivíparos, 91 Vuilleumier, 428
Wallace, 428, 429 Willis, 432
Xantóforos, 113
Yema, 289,290 Yodo, hormona tiroidea, 336 Yoduroforina, 208 Yucatain, 444

ÍNUICE TEMÁTICQ
Zimógenos, 148 ,149 ,341 Zona abisal, 419 ,420 Zona batid, 419,420 Zona eufótica, 408 Zona limnética, 406 Zona litoral, 419, 424 Zona nerítica, 416, 417 Zona oceánica, 416 ,417
489
Zona profunda, 406 Zona supralitoral, 419 Zona trofogénica, 408 Zoogeografía, 403, 428
definición, 428 indicios históricos, 430 Véase también Distribución

Se terminó de imprimir esta primera reimpresihn de 2 O00 ejemplares, en mayo de 1990, en loa
talleres de Editorial Calypso, S.A. Oculistas 43, Col. Sifón, 09400 Mexico. D.F.














