
Humoral Immune Responses to Select Bacterial Pathogens …
61
Clemson University TigerPrints All eses eses 5-2019 Humoral Immune Responses to Select Bacterial Pathogens in the American Alligator, Alligator mississippiensis Bailey Marie Alston Clemson University, [email protected] Follow this and additional works at: hps://tigerprints.clemson.edu/all_theses is esis is brought to you for free and open access by the eses at TigerPrints. It has been accepted for inclusion in All eses by an authorized administrator of TigerPrints. For more information, please contact [email protected]. Recommended Citation Alston, Bailey Marie, "Humoral Immune Responses to Select Bacterial Pathogens in the American Alligator, Alligator mississippiensis" (2019). All eses. 3135. hps://tigerprints.clemson.edu/all_theses/3135
Transcript of Humoral Immune Responses to Select Bacterial Pathogens …

Clemson UniversityTigerPrints
All Theses Theses
5-2019
Humoral Immune Responses to Select BacterialPathogens in the American Alligator, AlligatormississippiensisBailey Marie AlstonClemson University, [email protected]
Follow this and additional works at: https://tigerprints.clemson.edu/all_theses
This Thesis is brought to you for free and open access by the Theses at TigerPrints. It has been accepted for inclusion in All Theses by an authorizedadministrator of TigerPrints. For more information, please contact [email protected].
Recommended CitationAlston, Bailey Marie, "Humoral Immune Responses to Select Bacterial Pathogens in the American Alligator, Alligator mississippiensis"(2019). All Theses. 3135.https://tigerprints.clemson.edu/all_theses/3135

i
HUMORAL IMMUNE RESPONSES TO SELECT BACTERIAL PATHOGENS IN
THE AMERICAN ALLIGATOR, Alligator mississippiensis
A Thesis
Presented to
the Graduate School of
Clemson University
In Partial Fulfillment
of the Requirements for the Degree
Master of Science
Environmental Toxicology
by
Bailey M. Alston
May 2019
Accepted by:
Dr. Charles D. Rice, committee chair
Dr. Thomas R. Rainwater
Dr. Thomas E. Schwedler

ii
Abstract
The American alligator, Alligator mississippiensis, is widely distributed and
abundant throughout the southeastern United States. Despite their abundance, these
reptiles have not been examined for their role in environmental pathogen distribution, as
a sentinel for the presence of various pathogens, or other humoral immune responses in
individuals from different habitats. This study investigates the humoral immune
responses of alligators to select bacterial pathogens. Immunoglobulin Y (IgY), an
immunoglobulin molecule found in amphibians, birds and reptiles, similar to IgG in
higher vertebrates. IgY was purified from pooled alligator serum collected in coastal
South Carolina. Mouse polyclonal antisera (pAb) was then generated against IgY to
develop a sensitive ELISA to quantify serum antibody responses and relative titers.
Serum samples from alligators collected from multiple localities in Florida were screened
for bacteria-specific antibodies to the following nine aquatic bacteria: Vibrio cholera,
Escherichia coli, V. anguillarum, V. vulnificus, V. parahaemolyticus, Brevundimonas
vesicularis, Mycobacterium marinum, Erysipelothrix rhusiopathiae and Streptococcus
agalactiae. C-reactive protein (CRP) is an ancient acute phase protein, participating in
complement activation and opsonisation of pathogens, and is usually indicative of
relative levels of systemic inflammation. Alligator CRP was purified and used to
generate a specific monoclonal antibody to develop ELISA-based approaches to semi-
quantifying circulating CRP in individuals. Finally, serum lysozyme enzymatic activity
was also quantified in individual samples. Humoral immune responses to various
pathogens, along with CRP and lysozyme activity, may be correlated with environmental

iii
health. This study provides the first baseline data and proof of concept, to compare
responses of wild alligators to pathogens throughout their distribution. This information
will allow A. mississippiensis to be used as a sentinel of pathogen occurrence and
environmental quality in future studies.

iv
Dedication
To the Academy of the Holy Names, Ms. Clancy and Mr. Triller, for instilling a
love of science in me at an early age. To my grandfather, “Daddy Max” for his
inspirational and lifelong pursuit of education.

v
Acknowledgements
First, I would like to thank Dr. Rice for providing me with such a wonderful
opportunity to get my Master’s in his lab and for being so supportive along the way.
Thank you for your mentorship and for setting me up for success. I came out of this
program having learned not only about research, but also important life lessons. Two of
the most important life lessons I have learned from Dr. Rice are that you shouldn’t take
yourself too seriously and that not every experimental error is the end of the world.
Thank you to Dr. Schwedler and Dr. Rainwater for serving on my committee. I really
appreciate your time, expertise, guidance and understanding throughout this process.
Thank you to Dr. Rainwater for providing samples and putting us in touch with Dr. John
Bowden at the University of Florida, and to Dr. Bowden for providing alligator samples.
To Kursten and Delia – the “angels on my shoulder”. Thank you for being such
supportive lab-mates and friends. I know I can always count on you two to have my
back, help clean up messes (both in my life and in the lab (i.e. the centrifuge incident))
and celebrate achievements. To my “ride or die”, Lindsey Louise Barnett Jr. - thank you
for always keeping it real and for hyping me up when I needed it (and when I don’t).
Natalie, thank you for being such a supportive and understanding roomie, cat mom and
friend this past year.
Last but not least, I would also like to thank my family. My parents, George and
Jennifer, for the opportunities and life experiences you’ve given me. For teaching me the
importance of education, always pushing me to do my best and the continuous support
and unconditional love over the past 25 years. Ashley, for showing me the importance of

vi
always being yourself. Grace, for being my voice of reason and best friend, and Will and
Christian for being the best little brothers (making me look good) and always having my
back.

vii
Table of Contents
Page
Title Page ...........................................................................................................................i
Abstract ............................................................................................................................ ii
Dedication ........................................................................................................................iv
Acknowledgements ........................................................................................................... v
List of Figures ............................................................................................................... viii
Chapters
I. Literature Review............................................................................................ 1
Alligator mississippiensis: Species background ............................................. 1
Effects of temperature ..................................................................................... 3
Threats to the American alligator ................................................................... 4
Alligator immunology ..................................................................................... 4
Reptilian innate immunity............................................................................... 5
Reptilian adaptive immunity ........................................................................... 8
Potential role as sentinel species and amplifier host ..................................... 13
II. Chapter One .................................................................................................. 15
Introduction ................................................................................................... 15
Materials and Methods .................................................................................. 20
Results ........................................................................................................... 28
Discussion ..................................................................................................... 41
References ....................................................................................................................... 45

viii
List of Figures
Figure Page
1. American Alligator Habitat Range ....................................................................... 2
2. IgY Structure ....................................................................................................... 12
3. Alligator serum average lysozyme activity ......................................................... 30
4. SDS PAGE Gel of American Alligator Purified IgY ......................................... 31
5. Western Blot of Bacterial Lysates Probed with Alligator Whole Serum ........... 32
6. E.coli titers in serum ........................................................................................... 33
7. M.marinum titers in serum .................................................................................. 34
8. S.agalactae titers in serum .................................................................................. 35
9. V.vulnificus titers in serum .................................................................................. 36
10. V.anguillarium titers in serum ............................................................................ 37
11. V.parahaemoliticus titers in serum ..................................................................... 38
12. V.cholerae titers in serum ................................................................................... 39
13. Alligator relative CRP levels in serum ............................................................... 40

1
Chapter I: Literature Review
Alligator mississippiensis: Species Background
The American alligator, Alligator mississippiensis, is the largest reptile and only
alligator native to the United States. These carnivorous reptiles are found primarily
throughout the Southeast U.S., from the Virginia-North Carolina border to South Florida
and as far west as Texas (Figure 1) (United States Geographical Services). Once
considered an endangered species, A. mississippiensis has made a full recovery as a result
of conservation efforts required by the Endangered Species Preservation Act (US Fish
and Wildlife Service).
Alligators inhabit freshwater marshes, swamps, rivers, and lakes and can
sometimes be found in brackish water. They play a large role in the ecosystem and are
considered to be both keystone species and ecosystem engineers (Schechter and Street
2000, Rice and Mazzotti 2004). As a keystone species and top predator, alligators affect
most aquatic life in their environment (Beard 1938). Using their snouts and tails, they
create ‘alligator holes’ which are used for shelter and hibernation. These burrows serve
an important role, especially in the everglades, as they retain water during the dry season,
providing refuge to other organisms (Craighead 1969).

2
Figure 1. Map of the southeastern U.S showing the geographic range of A.
mississippiensis (United States Geographical Services, 2000).

3
Effects of temperature
As poikilothermic ectotherms, alligators’ body temperatures adjust with the
environment and therefore must be regulated through behaviors. Basking in the sun
raises body temperature, while submerging underwater and allowing heat to be released
through the mouth lowers body temperature (Smith 1975). Physiological processes such
as immune function, feeding behavior, reproduction, growth and development are all
directly related to environmental temperature (Lance 2003, Lang and Andrews 1994). As
a result, there are seasonal changes in behavior that affect how resources are partitioned
(Zimmerman, Vogel, and Bowden 2010).
When exposed to extreme temperatures, reptiles undergo a physiological stress
response. Studies have found that immunoglobulin production decreases in response to
cold weather (Zapata, Varas, and Torroba 1992). Water temperature has an effect on
haematological values in other crocodilians, further demonstrating how temperature can
play a role in immunosuppression (Turton et al. 1997). Phagocytic activity of
macrophages and the amount of histamine released by basophils can also be affected by
temperature (Mondal and Rai 2001, Zimmerman, Vogel, and Bowden 2010).
Alligators are active year-round, but most active during the warmer months. The
mating season begins in May and ends mid-July when females lay their eggs. They lay
one clutch of 20-60 eggs every one to two years (Brandt 1989). Alligators demonstrate
temperature dependent sex determination; the sex is determined by the incubation
temperature of the nest. There is variation in the temperature depending on where the nest
is built, as well as the position of the egg in the nest (Ferguson and Joanen 1982).

4
Temperatures of 32.2 oC – 33.9 oC produce mostly male alligators, while 27.8 oC - 30 oC
results in mostly females (Schechter & Street, 2000).
Threats to the American alligator
Although A. mississippiensis is no longer endangered, the Fish and Wildlife
Service classifies this species as ‘threatened’ because it resembles the endangered
American crocodile (US Fish and Wildlife Service, 2008). The biggest threat is habitat
loss from the development and drainage of wetland (National Wildlife Federation).
Global climate change may affect population dynamics because of the American
alligators’ reliance on temperature for sex determination; as temperature increases, the
male to female sex ratio may be skewed (Valenzuela et al. 2019).
Anthropogenic activities resulting in contamination of aquatic habitats pose
another serious threat to aquatic life. Reptiles commonly inhabit contaminated areas, and
because alligators have long lifespans, there is an increased likelihood that contaminants
will accumulate, increasing the risk of disease and other adverse effects (Guillette et al.
1994). Environmental contaminants are known to affect lymphocyte proliferation
specifically (Zimmerman, Vogel, and Bowden 2010).
Alligator immunology
The immune system protects against infection by microorganisms. Crocodilian
immune systems are similar to those of higher vertebrates in that the lymphoid tissues
include the thymus, spleen, gut associated lymphoid tissue (GALT) and bone marrow
(Kvell et al. 2007). The thymus is where T-cell maturation occurs. The spleen removes

5
old red blood cells and is the site of antibody synthesis. GALT traps and concentrates
antigens, allowing for interaction with lymphocytes. Bone marrow is the site of
hematopoiesis and B-cell maturation (Keller et al. 2005). Crocodilians and other reptiles
do not have lymph nodes, germinal centers or Peyer’s patches, as these structures are
only found in warm blooded vertebrates (Flajnik 2018).
Reptiles demonstrate species-specific, seasonal variation in lymphoid tissues,
analogous to that of hibernating mammals. The thymus and white pulp of the spleen
become less developed during the winter in reptiles and species-specific differences can
be seen in structures during the summer (Zimmerman, Vogel, and Bowden 2010). As the
thymus begins to redevelop in the spring, T-cell populations are reestablished, making T-
cell proliferation strongly dependent on seasonal cycles (Kruman 1992).
The reptilian immune system is comprised of both innate and adaptive immunity,
as seen in all gnathostomes (Dzik 2010). While the innate immune system can be traced
back to more primitive organisms, such as invertebrates, the adaptive immune system is
only found in vertebrates (Song, Sarrias, and Lambris 2000). The innate immune system
acts as the first line of defense with a non-specific response, in attempt to prevent the
spread of infection. Following the innate response, the adaptive response is activated.
The type of adaptive response, cell mediated or humoral, depends on both the type of
innate response and the pathogen (Zimmerman, Vogel, and Bowden 2010).
Reptilian innate immunity
The reptilian innate immune system includes physical barriers, antimicrobial

6
peptides (AMPs), enzymes, proteins (including complement) and non-specific leukocytes
(Finger 2012, Zimmerman, Vogel, and Bowden 2010). Physical barriers include skin and
gut mucosa, which act to keep microorganisms out (Finger 2012). Antimicrobial
peptides (AMPs) are similar in structure and function to beta defensins found in
mammals. Lysozymes, also found in mammals, are antimicrobial enzymes that
hydrolyze the membrane of both Gram-positive and Gram-negative bacteria, resulting in
osmotic lysis. Other antimicrobial enzymes found in crocodilians include phospholipase
A2 (PLA2) and dipeptidyl peptidase IV (DPP2, also known as CD26); both have been
confirmed in A. mississippiensis. PLA2 functions to destroy bacteria by increasing the
permeability of the cellular membrane (G Buckland and C Wilton 2000). The specific
function of DPP2/CD26 has not been determined in reptiles (Finger 2012). In mammals,
DPP2/CD26 plays an important role in numerous parts of humoral immunity, including
role in cellular development, migration and cytokine production (Yan et al. 2003).
The complement system plays a large role in regulating the adaptive immune
response through at least three different pathways: classical (CP), alternative (AP) and
lectin (LP) (Trouw and Daha 2011). It is comprised of around 30 proteins that either
directly lyse or opsonize bacteria in the plasma. Microbial opsonization results in
increased phagocytic efficiency by coating bacteria with proteins, allowing macrophages
to identify and phagocytose the foreign bacteria (Zimmerman, Vogel, and Bowden 2010).
The complement system is more diverse and well developed in poikilothermic species
than in higher vertebrates, yet it is not well characterized (Sunyer, Zarkadis, and Lambris
1998, Zarkadis, Mastellos, and Lambris 2001, Merchant et al. 2005). Studies have found

7
that serum complement from A.mississippiensis is effective against different strains of
Gram-positive bacteria, three Naegleria species and four Acanthamoeba species
(Merchant et al. 2003, Merchant et al. 2004).
Acute phase proteins (APP) are important in innate immunity of all animals
(Cray, Zaias, and Altman 2009). They are produced by hepatocytes as part of the acute
phase response (APR). APR is activated in response to many attacks on the body,
including infection, trauma, stress and inflammation, in attempts to prevent further
infection, initiate inflammation and reestablish homeostasis. C-reactive protein (CRP) is
an important APP in all vertebrates (Lee et al. 2017). CRP is produced by hepatocytes
and is made up of five monomers, each 25 kDa. Although the structure is evolutionarily
conserved, there are species dependent variations in the function of CRP and ligand
binding specificity. In humans, CRP is involved in many parts of the APR; the most
important function seems to be in defense against bacterial infection and clearance of
necrotic and apoptotic cells (Volanakis 2001). It is also used as a marker for many health
complications including infection, trauma and autoimmune diseases (Pepys and
Hirschfield 2003).
The leukocytes of the crocodilian innate immune system include heterophils,
basophils, eosinophils and monocytes. Heterophils are the first cells at the site of
infection (Jacobson 2007). They suppress invasion of microbes and are involved in the
inflammatory response, serving the same function as neutrophils in mammals (Montali
1988). These cells account for over 50% of peripheral leukocytes in A.mississippiensis
and can be found in damaged tissue within hours (Mateo, Roberts, and Enright 1984b, a).

8
This percentage may vary as a result of other factors, such as seasonal differences, age or
infection (Glassman, Bennett, and Hazen 1981, Mateo, Roberts, and Enright 1984b).
Basophils comprise ~12% and eosinophils make up ~10% of circulating leukocytes in
crocodilians (Mateo, Roberts, and Enright 1984b). Basophils are involved in allergic
reactions (Sullivan and Locksley 2009). They have antigen-specific immunoglobulins on
the surface and release histamines when triggered by an antigen. The amount of
histamine released is dependent on the concentration of the antigen and the temperature
(Zimmerman, Vogel, and Bowden 2010). The function of eosinophils is most likely
similar to the function in mammals: producing free radicals and peroxide in response to
parasites (Coico and Sunshine 2015). In A.mississippiensis, eosinophils have been
detected in response to infection by leeches (Glassman, Holbrook, and Bennett 1979).
Lastly, monocytes make up ~1% (1.5+SD 0.3) of peripheral leukocytes in
A.mississippiensis (Mateo, Roberts, and Enright 1984b). In mammals, monocytes can
further differentiate into macrophages and myeloid dendritic cells (DCs) (Owen 2013).
All of these phagocytic cells release cytokines and serve an important function as antigen
presenting cells (APCs). APCs process then present antigens to T cells, serving as a link
between the innate and adaptive immune responses (Coico and Sunshine 2015, Finger
2012).
Reptilian adaptive immunity
As previously mentioned, the adaptive immune response depends on the innate
response. The initial, primary adaptive response ultimately results in clearance of the

9
pathogens and formation of an immunological memory. Memory cells are important in
the secondary response that occurs in the case of re-exposure to the same pathogen
(Owen et al., p 17). The two parts of the adaptive immune system are cell mediated
immunity and humoral immunity.
Reptilian adaptive immunity: cell-mediated
Reptiles are known to have functional T cells, the lymphocytes involved in cell-
mediated immunity (Burnham et al., 2005). T cells are divided into two major types:
cytotoxic T cells (TC) and T helper cells (TH) (Owen et al., p 40). Cytotoxic-like T cells
have been identified in reptiles; they function to kill infected host cells and abnormal
cells (Pitchappan & Muthukkaruppan, 1977; Coico et al., 2003). Studies have suggested
the presence of regulator TH cells in reptiles, however the exact function is unknown
(Pitchappan & Muthukkaruppan, 1977). In mammals, TH cells secrete cytokines which
recruit B cells (Owen et al., p 40). Reptiles do not have germinal centers and lymph
nodes, which provide a location for TH immune cell interactions, so these cells may have
a different function than seen in mammals (Zimmerman et al., 2009). Variation in the
strength of cell-mediated responses are seen between the sexes; females typically have a
stronger response (Ahmed et al., 1985).
Reptilian adaptive immunity: humoral
B cells, another type of lymphocyte, are responsible for antibody production
(Zimmerman, Vogel, and Bowden 2010). B cells are able to recognize antigens in their

10
natural state. When an antigen binds to a B cell, highly specific antibodies, with antigen
binding sites identical to the ones on the surface of the B cell, are secreted (Owen 2013).
The general antibody structure is the same in all jawed vertebrates, consisting of two
identical heavy chains joined to two identical light chains by a disulfide bond. The heavy
chain and the light chain both contain a constant region and a variable region. Antigen
binding sites are found on the variable regions of both the heavy and light chains and are
therefore important for protection against different pathogens (Zimmerman, Vogel, and
Bowden 2010). The variable region contains different gene segments that undergo
rearrangement, allowing for tremendous genetic diversity. The variable (V), diversity (D)
and joining (J) gene segments rearrange to make different variable regions on the heavy
chain; the V and D gene segments rearrange on the light chain (Owen 2013).
Crocodilians produce at least four immunoglobulins (Ig): IgM, IgD, IgY and IgA.
IgM is found in all jawed vertebrates (Pettinello and Dooley 2014). Found in high
concentrations in the blood and spleen, it is the first antibody produced by B cells and
activates complement (Pettinello and Dooley 2014, Zhang et al. 2017). IgM, IgA and
IgD are thought to be involved in mucosal immunity in reptiles, as in mammals (Iwata et
al. 2002, Pettinello and Dooley 2014). IgY is the main antibody of secondary defense and
is believed to be the precursor to IgG and IgE found in mammals (Flajnik 2018). IgY is
produced in larger quantities than IgM and is the main defense against infection (Warr,
Magor, and Higgins 1995).
There are two isoforms found in crocodilians (and some other reptiles): IgY and a
truncated form, IgY(∆Fc) (Figure 2) (Zhang et al. 2017). Both forms have two light

11
chain domains, with a molecular weight of 23.1kDa. IgY heavy chains have one variable
region and four constant regions, with a total molecular weight of 63kDa. IgY(∆Fc) has
one variable region and two constant regions, with a total molecular weight of 33.8kDa.
The total molecular weight of IgY is 180kDa and IgY(∆Fc) is 120kDa. The two isoforms
do not always serve the same function and not all organisms that have IgY have
IgY(∆Fc) (Warr, Magor, and Higgins 1995). IgY(∆Fc) lacks the receptor binding sites;
evidence suggests it is able to induce pathogen neutralization without inflammation (Li et
al. 2012).

12
Figure 2. The structure of the two isoforms of IgY. IgY(∆Fc) is a truncated form, with
only two constant regions on the heavy chain, versus the four constant regions on the IgY
heavy chain. Image taken from Warr, Magor, and Higgins 1995.

13
Potential role as sentinel species and amplifier host
The American alligator serves as a sentinel for habitat quality, specifically related
to environmental pollution in areas affected by anthropogenic activities (Tipton et al.
2017). Previous studies have confirmed that certain crocodilian responses directly relate
to environmental conditions (Mazzotti et al. 2009, Rice and Mazzotti 2004). Alligator
indicators rely on relative density determined by encounter rate, body condition and rate
of occupancy in alligator holes (Mazzotti et al. 2009).
Alligators have not been examined for their role in environmental pathogen
distribution or as a sentinel for the presence of various pathogens. They inhabit
environments known to harbor opportunistic pathogenic bacteria, yet even serious
injuries, including deep wounds and missing limbs, heal without signs of infections
(Merchant et al. 2003). Studies have found that crocodilians are able to live with
microbial infections with no physiological effects (Madsen 1993, Madsen et al. 1998,
Manolis et al. 1991).
There is also evidence that the American alligator may serve as an amplifying
host for West Nile Virus (WNV) (Jacobson et al. 2005). This is significant because there
have been multiple confirmed WNV outbreaks in farmed crocodilians worldwide
(Steinman et al. 2003, Debra et al. 2003, Jacobson et al. 2005). Most mammals serve as
dead end hosts, meaning the virus in the blood will not reach high enough levels to be
picked back up by other mosquitos (CDC, n.d.). However, samples from infected
alligators have had viremic levels above the infectious threshold for Culex pipiens and C.
quinquefasciatus mosquitoes; Culex mosquitos being the main disease vector (Jacobson

14
et al. 2005, Turell, O'Guinn, and Oliver 2000, Sardelis et al. 2001). Further studies are
necessary to fully understand the American alligator’s role in the spread of both bacterial
and viral diseases.

15
Chapter II: Humoral immune responses in the American alligator samples near
NASA, Florida
Introduction
Although the American alligator is the most studied of the extant crocodilians, the
reptilian immune system is not well understood (Brisbin 1986). Alligators have not been
examined for their role in environmental pathogen distribution, or as a sentinel for the
presence of various pathogens. They inhabit environments known to harbor opportunistic
pathogenic bacteria, yet they are often times able to heal serious injuries, including deep
wounds and missing limbs, without signs of infections (Merchant et al. 2003). Studies
have found that crocodilians living in their natural environment are able to live with
microbial infections with no physiological effects (Madsen 1993, Madsen et al. 1998,
Manolis et al. 1991)
This study investigates humoral immune responses in the Alligator
mississippiensis as an environmental monitor for emerging infectious diseases. The intent
is to correlate immune responses to various pathogens with the health of the environment
and establish a baseline of antibody titers against pathogens. This baseline will be used
to compare with responses in alligators throughout their distribution. This information
will allow A.mississippiensis to be used as a sentinel, for not only the overall health of the
environment, but also as a sentinel for the presence of various pathogens.
Lysozymes are antimicrobial enzymes that hydrolyze the membrane of both
Gram-positive and Gram-negative bacteria, resulting in osmotic lysis (G Buckland and C
Wilton 2000). Lysozyme activity can be measured through an enzymatic assay of

16
lysozyme activity. In this assay, Micrococcus lysodeikticus is added to the serum
samples. The lysozymes in the serum will lyse the bacterial cells, resulting in decreased
absorbance. The higher the lysozyme activity, the faster the absorbance will decrease.
C-reactive protein (CRP) is an important APP in all vertebrates (Lee et al. 2017). It is an
ancient acute phase protein, participating in complement activation and opsonisation of
pathogens, and is usually indicative of relative levels of systemic inflammation. CRP
levels can be semi-quantified using an ELISA based approach. Both lysozymes and
CRPs play an important role in immune response to bacteria. This study provides
baseline data that can be used to compare with levels in alligators throughout their
distribution.
IgY is the main immunoglobulin involved in defense against infection (Warr,
Magor, and Higgins 1995) in birds and reptiles. There are two isoforms found in
crocodilians (and some other reptiles) and birds: IgY and a truncated form, IgY(∆Fc)
(Figure 2) (Zhang et al. 2017). Both forms have two light chain domains, with a
molecular weight of 23.1kDa. IgY heavy chains have one variable region and four
constant regions, with a total molecular weight of 63kDa. IgY(∆Fc) has one variable
region and two constant regions, with a total molecular weight of 33.8kDa. The total
molecular weight of IgY is 180kDa and IgY(∆Fc) is 120kDa. The two isoforms do not
always serve the same function and not all organisms that have IgY, have IgY(∆Fc)
(Warr, Magor, and Higgins 1995). ELISAs can be used to quantify the level of
circulating antibodies in serum against pathogens. If the alligator has come into contact
with any of the pathogens that are immunogenic, there will be memory immune cells

17
against the pathogen from the adaptive immune response. Serum samples from 110
alligators collected throughout Florida, were screened for bacteria-specific antibodies to
select, ubiquitous, aquatic bacteria. It is important to look at serum titer levels for
potential correlation between location, gender, and size/age.
The nine bacteria used in this study were Vibrio cholera, V. vulnificus, V.
parahaemolyticus, V. anguillarum, Escherichia coli, Brevundimonas vesicularis,
Mycobacterium marinum, Erysipelothrix rhusiopathiae and Streptococcus agalactiae.
These bacteria were selected because they are abundant in aquatic environments and are
known human pathogens. V.cholera is the most infectious of the vibrio species.
V.cholera and V.parahaemolyticus attach copepods; one copepod can carry an infectious
dose V.cholera (Percival and Williams 2014, Huq et al. 1983). V.parahaemolyticus is
found in temperate coastal waters. It is the leading cause of foodborne illnesses and is
associated with raw shellfish and contaminated seafood consumption (Joseph, Colwell,
and Kaper 1982). V.anguillarum causes vibrosis in organisms inhabiting aquatic
environments (Frans et al. 2011). E.coli are a diverse group of bacteria that can be found
in aquatic environments; only some strains are pathogenic. Both E.coli and
V.parahaemolyticus have been isolated from A.mississippiensis (Russell and Herman
1970, Brown et al. 2001). B. vesicularis causes infection in immunocompromised
humans and is multi drug resistant (Shang et al. 2012).
M.marinum is able to infect all poikilothermic species and has been isolated from
crocodilians (Clark and Shepard 1963, Huchzermeyer and Van Wyk 2003). Many
reptiles are extremely susceptible to M.marinum infection, which often times become

18
systemic. Studies have found that the rate of infection is temperature dependent (Clark
and Shepard 1963). In humans, M.marinum causes a skin infection called “fish tank
granuloma” (Bouricha et al. 2014). E. rhusiopathiae can be a commensal or pathogenic
organism found in humans, domestic and wild animals (Wang, Chang, and Riley 2010).
It is possibly an environmental pathogen of concern near slaughterhouses and processing
plants on the water. S.agalactiae causes septicemia in a variety of aquatic organisms,
including crocodilians (Amborski et al. 1983, Bishop et al. 2007, Evans, Klesius, and
Shoemaker 2006, A. Plumb et al. 1974).
The objectives of this study were as follows:
1. Quantify lysozyme enzymatic activity in individual samples
2. Purify IgY from pooled alligator serum
3. Generate mouse polyclonal antisera against IgY
4. Western blot analysis to determine if whole serum recognized pathogen
antigens
5. Use enzyme linked immunosorbent assay (ELISA) to quantify the level of
circulating antibodies in serum against pathogens and relative titers
6. Purify CRP and generate mouse monoclonal antibody against CRP
7. Use ELISA based approach to semi-quantify circulating CRP in individual
samples
Humoral immune responses to various pathogens, along with CRP and lysozyme
activity can be correlated with the health of the environment. This study provides the

19
first baseline data and a proof of concept, to then compare responses throughout their
distribution. This information will allow A. mississippiensis to be used as a sentinel in
future studies.

20
Materials and Methods
Alligators serum samples
Blood samples were collected from 142 American alligators from sites in South
Carolina, including Yawkey Island. These samples were sent from Dr. Thomas
Rainwater and used for IgY purification. Blood samples were collected from 110
American alligators from sites near NASA, including Cape Canaveral and Merritt Island
National Wildlife Refuge in Florida. These samples were sent from Dr. John Bowden at
University of Florida. The samples used in this study were collected from various month
between 2009 and 2011 and include 74 males and 36 females.
Serum lysozyme activity
Modified, previously published methods were used to determine serum lysozyme
activity (Parry 1965, Burton et al. 2002). Micrococcus lysodeikticus solutions were
prepared immediately prior to use, using 50 mg of lyophilized cells and 100 mL of 0.1M
phosphate buffer (pH 5.9). Next, 25 μL of each plasma sample was added in
quadruplicates to a 96-well plate and 175 μL of Micrococcus lysodeikticus was added to
each sample. Absorbance was measured at 450nm immediately and after a 5 minute
incubation at room temperature. The change in absorbance was quantified by subtracting
the absorbance at 5 minutes from the immediate absorbance. It was previously
determined that 1 unit of activity is a change in optical density (OD) of 0.001 per minute
(Parry 1965).

21
Purification of IgY
Previous studies have indicated that IgY can be purified using the Protein-G from
streptococcal bacteria, which work by binding immunoglobulins (Work et al. 2015,
Rodgers, Toline, and Rice 2018). One Protein-G columns (Thermo-Fisher) were used to
purify the alligator serum in this study. The columns were equilibrated by running 10mL
of 0.10M tris (pH 8.0) through, followed by 10mL of 0.01M tris (pH 8.0). Serum
samples from each of the 12 individual alligators were compiled into one 15 mL tube
(1000 μL from each individual) followed by the addition of 1.2 ml of of 1 M tris (pH
8.0). The sample was centrifuged for 10 minutes at 1,000 x g then passed twice through
the Protein G column. The column was washed with 20 mL of 0.10 M tris (pH 8.0), then
20 mL of 0.01 M tris (pH 8.0). Lastly, 0.05 M glycine (pH 2.5) was used to elute IgY
into seven separate microfuge tubes containing 100 μL aliquots of 1.0 M tris (pH 8).
Generation of polyclonal antisera against immunoglobulin Y
Purified IgY was used to immunize six-week old female Balb/c mice, housed at
the Godley-Snell Animal Facility at Clemson University. All protocols used are in
accordance to Institutional Animal Care and Use Committee (IACUC) regulations. On
day 1, mice were administered a sub-cutaneous (s.c.) injection of 100 ug purified IgY in
0.9% saline containing TiterMax Gold adjuvant. Two weeks later, on day 14, the mice
received a second s.c. injection of 50 μg of purified IgY with Freud’s incomplete
adjuvant (FIA). Boosters of 50 μg of purified IgY were given at 21 day intervals - on
days 35, in saline via s.c. immunizations, on day 56 intraperitoneally. Five days after the

22
last immunization, slow lethal CO2 asphyxiation and bilaterial pneumothorax were used
to sacrifice the mice. After being sacrificed, the mice were bled to collect blood, which
was stored overnight at 4°C to allow time for clotting. The sample was then centrifuged
for 20 minutes at 10,000 x g, the serum was collected to be used as the crude polyclonal
antisera source. The specificity of the mouse antiserum against IgY was determined by
western blotting. Procedures previously described in other studies in our lab (Rodgers,
Toline, and Rice 2018).
Growth and preparation of bacteria
Samples of Vibrio cholera, Escherichia coli, V. anguillarum, V. vulnificus, V.
parahaemolyticus, and Mycobacterium marinum, were acquired from the American Type
Culture Collection and grown according to recommended protocols. Streptococcus
agalactiae cultures were obtained from John Hawke (Louisiana State University) and
were also grown as directed. 20 μL of each of the nine bacteria were inoculated into 5
mL of their specific growth medium. V.cholera and V.parahaemolyticus were inoculated
in nutrient broth with 3% NaCl, E.coli in Luria broth, V.angullarum in marine broth,
M.marinum in Middlebrook broth, and S.agalactiae into tryptic soy broth. The bacteria
were grown in separate 25 mL tubes at 37oC then centrifuged at 2,500 x g for 15 minutes.
The supernatant was poured off and 25 mL of 0.01 M PBS was added to the pellets, to
allow for re-suspension. The samples were centrifuged again at 2,500 x g for 15 minutes,
then washed again by resuspension with 25 mL of 0.01 M PBS and centrifuged under the
same conditions. Cell pellets were re-suspended in 5mL of PBS a final time. Thirty μL

23
of 5× Laemmli sample buffer with 2-mercaptoethanol was added to 120 μL of each
bacterial suspension, then boiled for 8 min. This was done to create reducing conditions
and generate bacterial lysates.
Serum Y reactivity with bacterial antigens
Twenty μL of each lysate were subjected to western blot analysis. A 4-20% Mini-
PROTEAN® TGX Stain-FreeTM Protein Gel (BioRad) was used for the SDS-PAGE.
First, 13 μL of Precision Plus ProteinTM KaleidoscopeTM Prestained Protein Standards
molecular weight marker (BioRad) was added to well 1. Next, 20 μL of the lysate
samples were added to wells 2-10 in the following order: V.cholerae, E.coli,
V.anguillarum, V.vulnificus, V.parahaemolyticus, M.marinum, and S.agalactiae. The gel
was run at 200V for approximately 40 minutes. The proteins on the gel were transferred
to a Immun-Blot® PVDF membrane at 100V for 1 hour. The blots were blocked for 16
hours with 10% fetal calf serum in 0.01M PBS at 4°C. Next, 50 μL of serum from the
pooled serum resource, was put in a 50 mL tube and diluted 1:500, in 25 mL of PBS.
The blot was incubated for 1 hour at room temperature with the diluted alligator serum
then washed three times for 5 minutes with PBS-TW20. Mouse polyclonal IgY antisera
was diluted 1:1000. The blots incubated in this solution for 1 hour at room temperature
followed by three, 5 minutes washes with PBS-TW20. The final incubation was with
goat anti-mouse IgG- AP (1:1,000 in PBS; Thermo-Fisher), following the same
incubation and wash procedure as the previous two steps. The blot was developed with
BCIP and NBT in AP buffer, allowing the AP activity to be visualized and recorded.

24
Development of enzyme linked immunosorbent assays (ELISA) to quantify relative
pathogen specific immunoglobulin Y titers in individual alligators
The seven aquatic bacteria mentioned previously were grown under the specific
conditions explained above. The supernatant was removed from each tube and 25 mL of
0.01 M PBS was added to re-suspend the pellet, this was repeated a second time. Once
the bacteria were re-suspended, the samples were diluted with PBS to optimal ODs that
had been previously established for 15 mL of total bacterial suspension. The optimal ODs
were: 0.200 for V.cholerae, E.coli, V.anguillarum, V.vulnificus, S.agalactiae, 0.1000 for
V.parahaemolyticus, and 0.050 for M.marinum.
High bonding 96-well plates (Medi0sorb, ThermoFisher) were coated with 75 μL
of 1 mg/mL solution of poly-D-lysine in distilled water, for 16 hours at 4 oC, then washed
with PBS-TW20. The optimized bacterial suspensions were then added to the plates; 50
μL of one bacterium was added to every well of three plates. This was repeated for all
nine bacteria, resulting in 27 coated plates, each coated with a single species. The plates
were centrifuged at 2,000 x g for 5 minutes, 50 μL of 0.5% glutaraldehyde was carefully
added to each well, then spun down again under the same settings. The plates incubated
at room temperature for 15 minutes then carefully washed (submerged) in PBS two times.
Next, 100 μL of 10 % BSA blocking buffer was added to each of the wells, followed by a
two hour incubation at room temperature. Each plate was washed three more times with
PBS-TW20. The plates were left out to dry for two hours then stored in plastic wrap at
room temperature for 36 hours.

25
Serum suspensions were prepared using a 100-fold dilution of serum from each
individual alligator into PBS in individual 1.5 mL microfuge tubes. A reference sample
for calculating relative titers was made from a composite sample, comprised of 50 μL of
serum from 6 randomly selected individuals in 2.7 mL of PBS. The composite serum
was added in duplicate to each plate, as a reference for variation from plate-to-plate.
Diluted serums were added to the wells, 75 μL in duplicate. The plates incubated for 16
hours at 4 oC, then washed three times with PBS-TW20.
Mouse polyclonal anti-alligator IgY was added to the wells in 75 μL volumes and
incubated overnight at 4 degrees C. The plates were then washed three times with PBS-
TW20, then 75 μL of goat anti-mouse IgG-AP (1:1,000) was added to the respective
wells and incubated for 2 hours at room temperature. The plates were washed again, four
times, with PBS-TW20. Lastly, 75 μL of 1mg/mL of p-nitrophenol (Fisher) in AP buffer
was pipetted into the wells and left to develop for 30 minutes. The plates were read at 405
nm and the data for each plate was recorded.
Generation of monoclonal antibody against CRP
To isolate CRP, plasma samples from several alligators were pooled and
centrifuged at 1000 rpm at 4 oC for 10 min. The supernatant was centrifuged again at
12,000g at 4 oC for 15 min. Following the methods described by Robey et al., and
modified by Karsten and Rice, the supernatant was brought to room temperature and
passed though an AH-Sepharose 4B-PC column (equilibrated in a 0.15 M NaCl, 0.05 M
Tris, pH 7.4, 0.05 M CaCl2 buffer solution) (Robey, Tanaka, and Liu 1983, Karsten and

26
Rice 2004). Alligator CRP and serum amyloid-P component (SAP) were eluted from the
column using a 0.15 M NaCl, 0.05 M Tris, 0.1 M EDTA, pH 7.4, buffer solution. To
separate alligator CRP from SAP, the eluent was dialyzed against the 0.15 M NaCl, 0.05
M Tris, pH 7.4, CaCl2 buffer solution and applied to a Sepharose CL-4B column
(equilibrated in a 0.15 M NaCl, 0.05 M Tris, pH 7.4, 0.05 M CaCl2 buffer solution). CRP
was collected and subjected to SDS–PAGE and subsequent Coomassie blue staining to
check for purity. Coomassie blue staining of the gel showed a band corresponding to
alligator CRP (at 25 kDa) in the CRP purified sample. Balb/c mice were immunized with
the putative CRP sample over a 51-day period and bled for polyclonal anti-sera, as well
as to collect spleens for subsequent steps in fusion steps for making mAbs. Fusion of
splenic plasma cells with Sp02-14 myelomas, screening, cloning, purification and
isotyping of the antibodies was done according to methods described elsewhere
(Margiotta, Bain, and Rice 2017, Rice and Weeks 1989). Western plot analysis
confirmed that the mouse anti-alligator mAb recognizes and is specific to putative CRP.
Semi quantification of circulating CRP levels using ELISA based approach
Methods previously developed in the lab (a quantitative capture ELISA) were
used to quantify the circulating CRP in alligator serum (Karsten and Rice 2004). This
involved the use of purified CRP and mouse anti-alligator mAb.

27
Statistical analysis
Lysozyme enzymatic activity, antibody titers against bacteria and CRP data were
analyzed using Analysis of Variance (ANOVA), followed by Bonferroni post hoc,
multiple comparison test using GraphPad7.

28
Results
An increase in lysozyme activity was seen during summer months. There were
significant differences (P<0.05, P<0.01, P<0.001) in serum lysozyme activity between
samples collected in in March / April, May, June, July, August and September of 2010
(Figure 3).
The Protein G purification technique was successful in producing pure
A.mississippiensis IgY (Figure 4). Clear bands are present at approximately 63 kDa,
33.8 kDa and 23 kDa. The band at 63 kDa corresponds to the heavy chain of IgY, 7S
IgY, and the band at 33.8 kDa corresponds to 5.7S IgY, the heavy chain of IgY(∆Fc).
The band near 25 kDa corresponds to the light chain of both isoforms of IgY, which is
23.1 kDa. No other strong bands appear on the gel. This indicates that IgY was
successfully purified.
Using pooled serum samples as the probe and the pAb anti-IgY for detection
source of antibodies, distinct bands were seen for E.coli, V.vulnificus,
V.parahaemolyticus, and S.agalactiae (Figure 5). Minor bands of proteins were
recognized in other bacterial lysates as well. This indicates that there are circulating
antibodies against the bacteria in the American alligators used in this study.
Statistical analysis of the serum titer levels against seven bacteria indicate
significant differences between titer levels collected during different months. There was
a statistically significant difference (P <0.001) in the quantity of titers against E.coli in
the summer months compared to the winter and spring (Figure 6). Titer levels against
E.coli are increased in the summer and decrease in the winter. M.marinum titers

29
decreased in serum samples from August and September 2010. Significant differences
(P<0.05, P<0.001) were seen between samples collected in July 2010, August 2010,
September 2010, the Sum of 2009 and the rest of the samples collected earlier in 2010
(Figure 7). S.agalactae titers were highest in January 2010. There was a significant
decrease (p<0.01) in titer levels against this bacteria in months sampled between
February 2010 and August 2010 when compared to titer levels in January 2010 (Figure
8). There were no significant differences in titer levels against V. vulnificus and V.
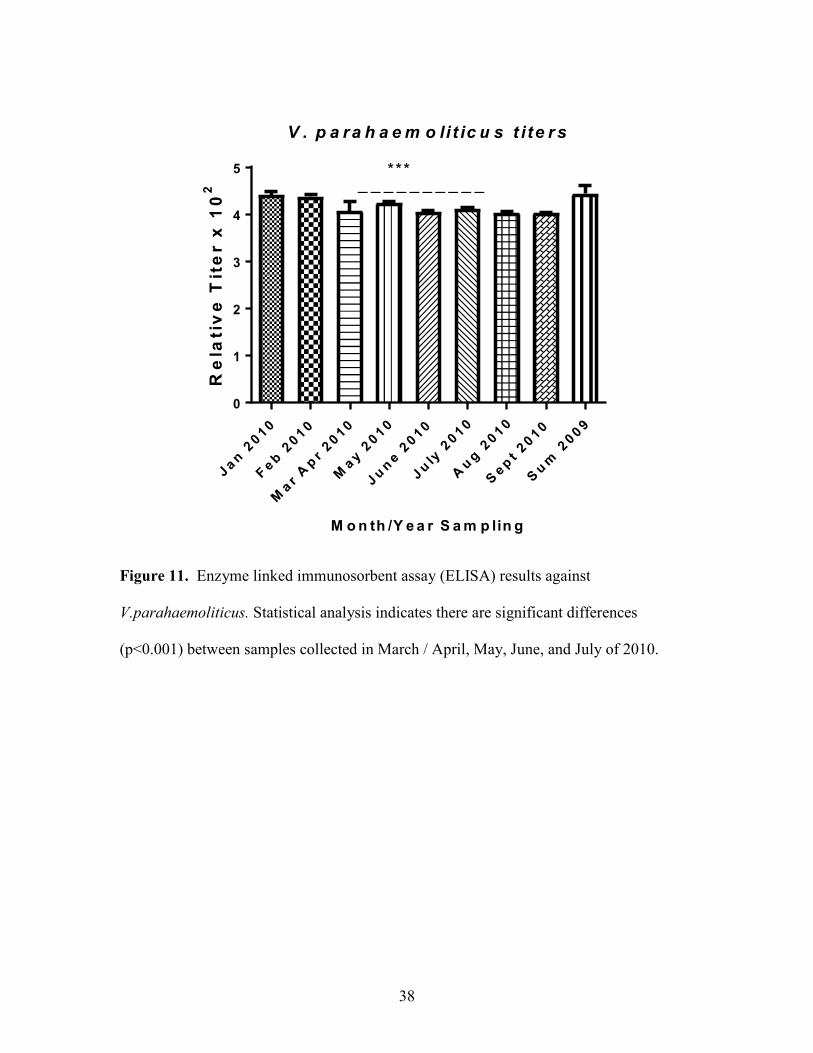
anguillarium between sampling months (Figure 9, Figure 10). V.parahaemoliticus titers
also increased during summer months. There is a significant difference (P<0.001)
between titers in samples collected in March / April 2010, May 2010, June 2010, July
2010 and the rest of the sampling months (Figure 11). V.cholera titers were significantly
different (P<0.001) in June 2010, July 2010 and August 2010 compared to the rest of the
months sampled (Figure 12).
Analysis of Enzyme linked immunosorbent assay (ELISA) for relative putative
CRP levels for the American alligator indicates a significant increase (P<0.05) in samples
collected in June 2010 compared to those collected in March / April 2010 (Figure 13).
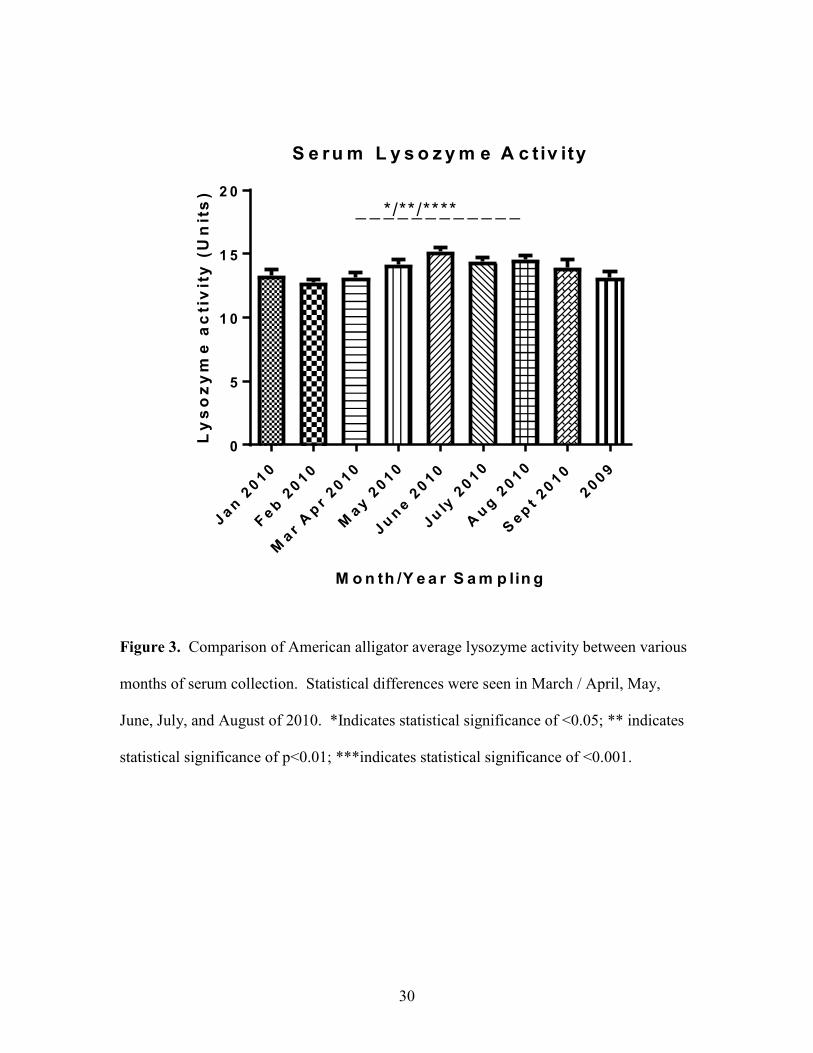
30
Figure 3. Comparison of American alligator average lysozyme activity between various
months of serum collection. Statistical differences were seen in March / April, May,
June, July, and August of 2010. *Indicates statistical significance of <0.05; ** indicates
statistical significance of p<0.01; ***indicates statistical significance of <0.001.
S e ru m L y s o z y m e A c t iv ity
M o n th /Y e a r S a m p lin g
Ly
so
zy
me
ac
tiv
ity
(U
nit
s)
Jan
2010
Feb
2010
Mar
Ap
r 2010
May 2
010
Ju
ne 2
010
Ju
ly 2
010
Au
g 2
010
Sep
t 2010
2009
0
5
1 0
1 5
2 0
_ _ _ _ _ _ _ _ _ _ _ _*/**/****

31
Figure 4. SDS-PAGE gel with purified IgY from the American alligator. Lane 1 shows
the molecular weight marker (Kaleidoscope, BioRad), lane 2 is the sample. Clear bands
are seen around 63 kDa, 33.8 kDa and 23 kDa; 63 kDa corresponds to the IgY heavy
chain, 7S IgY. The band around 33.8 kDa corresponds to the heavy chain of IgY(∆Fc),
5.7S IgY. The band near 25 kDa corresponds to the light chain of both isoforms of IgY,
which is 23 kDa.
MW IgY

32
Figure 5. Western blot analysis showing A.mississippiensis whole serum (diluted 1:500)
activity against bacteria lysates using pAB anti-IgY. Lane 1 contains the molecular
weight marker (Kaleidoscope, BioRad). Lanes 2-9 contain the bacterial lysates in the
following order: V.cholerae, E.choli, V.anguillarum, V.vulnificus, V.parahaemolyticus,
M.marinum, E.rhusiopathiae, S.agalactiae, B.vesicularis.
MW Vc Ec Va Vv Vp Mm Er Sa Bv

33
Figure 6. Enzyme linked immunosorbent assay (ELISA) results against E.coli.
Statistical analysis indicates significant differences (p<0.001) between samples collected
in June 2010, July 2010, August 2010 and September 2010.
E c o li t ite r s
M o n th /Y e a r S a m p lin g
Re
lati
ve
Tit
er x
10
2
Jan
2010
Feb
2010
Mar
Ap
r 2010
May 2
010
Ju
ne 2
010
Ju
ly 2
010
Au
g 2
010
Sep
t 2010
Su
m 2
009
0
1
2
3 _ _ _ _ _ _ _ _ _ _***

34
Figure 7. Enzyme linked immunosorbent assay (ELISA) results against M.marinum.
There is a significant difference between samples collected in July, August, September
from 2010 and the sum of 2009. *Indicates statistical significance of <0.05; ***indicates
statistical significance of <0.001.
M . m a r in u m t it e r s
M o n th /Y e a r S a m p lin g
Re
lati
ve
Tit
er x
10
2
Jan
2010
Feb
2010
Mar
Ap
r 2010
May 2
010
Ju
ne 2
010
Ju
ly 2
010
Au
g 2
010
Sep
t 2010
Su
m 2
009
0
1
2
3
4
5
_ _ _ _ _ _ _ _*/***

35
Figure 8. Enzyme linked immunosorbent assay (ELISA) results against S.agalactae.
Statistical analysis indicates significant differences between serum titers against bacteria
collected in January, March / April, May, June, July and August 2010. *Indicates
statistical significance of <0.05; ** indicates statistical significance of <0.01.
S . a g a la c ta e t i te r s
M o n th /Y e a r S a m p lin g
Re
lati
ve
Tit
er x
10
2
Jan
2010
Feb
2010
Mar
Ap
r 2010
May 2
010
Ju
ne 2
010
Ju
ly 2
010
Au
g 2
010
Sep
t 2010
Su
m 2
009
0
1
2
3
4_ _ _
_ _ _ _ _ _ _ _ _ _ _ _ _ _
*
**

36
Figure 9. Enzyme linked immunosorbent assay (ELISA) results against V.vulnificus. No
significant differences were seen between months of collection.
V . v u ln if ic u s t ite rs
M o n th /Y e a r S a m p lin g
Re
lati
ve
Tit
er x
10
2
Jan
2010
Feb
2010
Mar
Ap
r 2010
May 2
010
Ju
ne 2
010
Ju
ly 2
010
Au
g 2
010
Sep
t 2010
Su
m 2
009
0
1
2
3
4
5
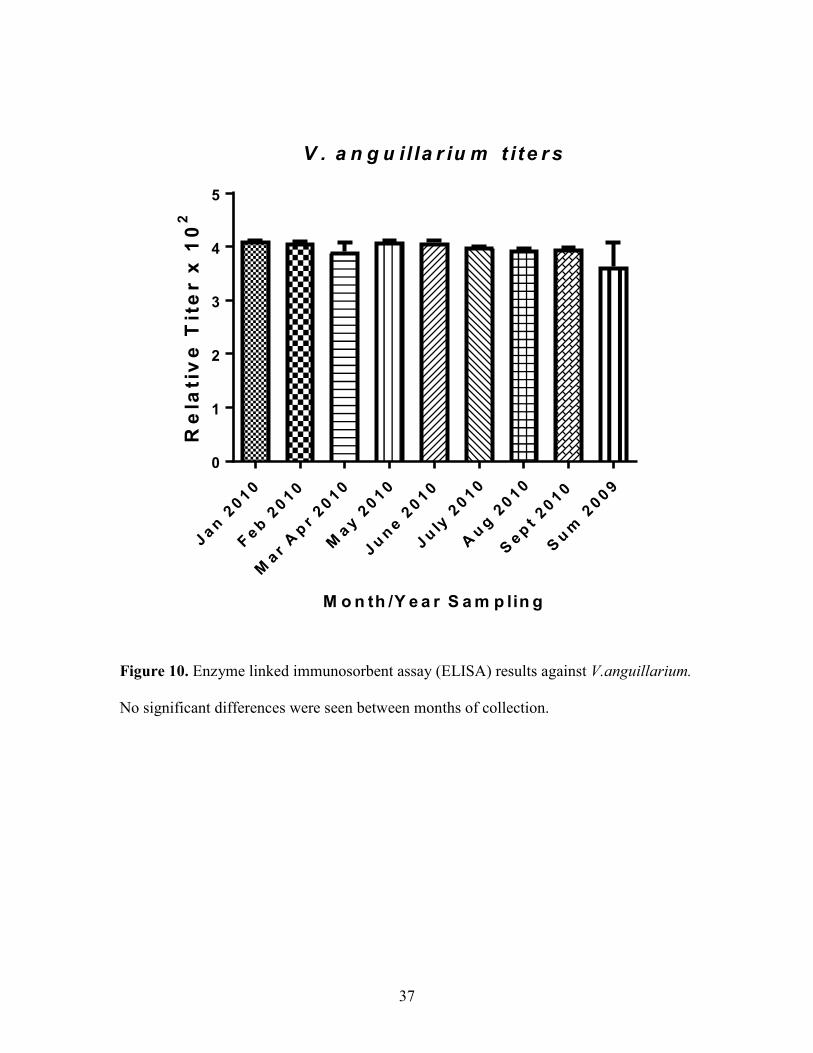
37
Figure 10. Enzyme linked immunosorbent assay (ELISA) results against V.anguillarium.
No significant differences were seen between months of collection.
V . a n g u il la r iu m t i te r s
M o n th /Y e a r S a m p lin g
Re
lati
ve
Tit
er x
10
2
Jan
2010
Feb
2010
Mar
Ap
r 2010
May 2
010
Ju
ne 2
010
Ju
ly 2
010
Au
g 2
010
Sep
t 2010
Su
m 2
009
0
1
2
3
4
5
38
Figure 11. Enzyme linked immunosorbent assay (ELISA) results against
V.parahaemoliticus. Statistical analysis indicates there are significant differences
(p<0.001) between samples collected in March / April, May, June, and July of 2010.
V . p a ra h a e m o li t ic u s t i te r s
M o n th /Y e a r S a m p lin g
Re
lati
ve
Tit
er x
10
2
Jan
2010
Feb
2010
Mar
Ap
r 2010
May 2
010
Ju
ne 2
010
Ju
ly 2
010
Au
g 2
010
Sep
t 2010
Su
m 2
009
0
1
2
3
4
5
_ _ _ _ _ _ _ _ _ _***

39
Figure 12. Enzyme linked immunosorbent assay (ELISA) results against V.cholerae.
Statistical analysis indicates there are significant differences (p<0.001) between samples
collected in June, July, and August of 2010.
V . c h o le ra e t i te rs
M o n th /Y e a r S a m p lin g
Re
lati
ve
Tit
er x
10
2
Jan
2010
Feb
2010
Mar
Ap
r 2010
May 2
010
Ju
ne 2
010
Ju
ly 2
010
Au
g 2
010
Sep
t 2010
Su
m 2
009
0
1
2
3
4 _ _ _ _ _ _ _ _***

40
Figure 13. Enzyme linked immunosorbent assay (ELISA) showing relative CRP levels
for the American alligator. Statistical analysis indicates significant differences (p< 0.05)
between samples collected in March / April, and May of 2010.
C R P
M o n th /Y e a r S a m p lin g
Re
lati
ve
CR
P l
ev
els
Jan
2010
Feb
2010
Mar
Ap
r 2010
May 2
010
Ju
ne 2
010
Ju
ly 2
010
Au
g 2
010
Sep
t 2010
2009
0 .0
0 .2
0 .4
0 .6
0 .8
1 .0
_ _ _ _ _ _*

41
Discussion
This study provides the first baseline data for serum parameters in the American
alligator. Lysozyme activity and CRP levels increase in alligators during summer months.
Seasonal variation is also seen between serum titers against the seven select pathogens
during the sampling months. Lysozyme activity, CRP levels and circulating serum titers
levels against bacteria can be used to compare responses in alligators throughout their
distribution. This information will allow A.mississippiensis to be used as a sentinel for
various pathogens in future studies.
Comparison of American alligator average lysozyme activity between various
months of serum collection indicated that there are seasonal differences. There was a
significant difference between lysozyme activity in summer months compared to the rest
of the year. During summer months, water temperatures increases with the increase in air
temperature, often resulting in increased pathogen loads in the environment. It is
possible that the increase in lysozyme activity is in response to the increase in bacteria in
their environment, as an increase in lysozyme activity is associated with increased ability
to lyse bacteria. Lysozymes play an important role in the innate response to bacteria, as
they are able to hydrolyze the membrane of both Gram-positive and Gram-negative
bacteria, resulting in osmotic lysis.
The results of this study also show that it is possible to purify IgY from alligator
serum through a Protein G purification process. Our lab has previously purified IgY
from sea turtle serum with protein A/G columns (Rodgers, Toline, and Rice 2018). The
purity of the IgY was confirmed; the sample contained the heavy chain, and light chain of

42
both IgY and IgY(∆Fc). The Protein G purification technique is simple and inexpensive
compared to previous methods used to purify IgY, which involved purifying IgY from
eggs instead of from serum. The ability to be able to purify IgY from alligator serum is
important because it can be used to generate both polyclonal antisera and monoclonal
antibodies against these different parts of alligator IgY.
The alligator anti-IgY pAb was used as the probe in a western blot analysis of
alligator serum response to bacteria. The results indicate that the alligators produced
circulating antibodies against the seven different bacteria. This indicates that the
individuals were exposed to the bacteria at high enough levels to elicit an immune
response, at some point in their lifetime. The range of banding seen on the blot indicates
that the polyclonal antisera can successfully be used for detection.
Significant differences were observed in the serum titers against the bacteria
between sampling months, as indicated in the data from the ELISAs used to quantify the
levels. There was an increase in the quantity of serum titers in the spring and the summer
and a decrease in the winter in E.coli, V.parahaemoliticus and V.cholera. S.agalactae
titers were highest in January 2010; the titers remained relatively consistent in the other
months sampled. M.marinum titers decreased in serum samples from August and
September 2010. There were no significant differences in titer levels against V. vulnificus
and V. anguillarium from month to month. These results indicate that there are changes in
titers against various bacteria depending on the sampling month. This study serves as a
proof of concept, showing that that it is possible to successfully quantify circulating
antibodies in alligator serum using the reagents made in this lab.

43
The ability to generate monoclonal antibodies against alligator CRP and
polyclonal antisera against alligator IgY allows for immunological analyses, such as
Western blotting and ELISAs, to be performed on alligator serum samples. The
production of a mAb against CRP allows for an unlimited supply of antibodies that can
continue to be used in these immunological assays because the hybridoma will continue
to produce the mAbs indefinitely. This will allow for further studies to be done on
alligators in other laboratories.
Analysis of circulating CRP levels (using an ELISA) showed an increase during
summer months. There were significant differences between CRP levels in samples
collected in March / April 2010, May 2010 and June 2010. CRP is important in the acute
phase response, participating in complement activation and opsonisation of pathogens,
and is usually indicative of relative levels of systemic inflammation. Increased levels of
CRP during warmer months is likely in response to increased pathogen loads in the
warmer waters.
The observed seasonal differences in immune responses are likely a result of
temperature changes. Previous studies have found that immune function in reptiles is
directly related to environmental temperature (Lance 2003, Lang and Andrews 1994).
When exposed to extreme temperatures, reptiles undergo a physiological stress response.
Studies have found that immunoglobulin production decreases in response to cold
weather (Zapata, Varas, and Torroba 1992). Water temperature has an effect on
haematological values in other crocodilians, further demonstrating how temperature can
play a role in immunosuppression (Turton et al. 1997). Phagocytic activity of

44
macrophages and the amount of histamine released by basophils can also be affected by
temperature (Mondal and Rai 2001, Zimmerman, Vogel, and Bowden 2010).
These findings serve as a proof of concept that the generated reagents are
functional and the immunological assays can be used to determine CRP, lysozyme and
circulating serum titer levels against bacteria using serum samples. The results provide
the first baseline data for serum parameters in the American alligator. This study is
important because it adds to our understanding of the reptilian immune system, by
showing that there are seasonal changes in CRP, lysozyme and serum titers to various
bacteria in the A.mississippiensis. This study should be continued with a larger sample
size, including samples from various geographic locations and size / age groups. Once a
larger sample size is obtained, analysis of immune responses between location, time of
year collected, sex and size of organism (using snout vent length) should be further
investigated. This will provide enough data to determine set baseline parameters for
comparison throughout the alligator’s distribution in the United States. This information
will allow for better understanding of the reptilian immune system, which is essential in
understanding the evolution of vertebrate immune system function.

45
References
A. Plumb, J., J. H. Schachte, J. L. Gaines, W. Peltier, and B. Carroll. 1974. Streptococcus
sp. from Marine Fishes Along the Alabama and Northwest Florida Coast of the
Gulf of Mexico. Vol. 103.
Amborski, R. L., T. G. Snider, 3rd, R. L. Thune, and D. D. Culley, Jr. 1983. "A non-
hemolytic, group B Streptococcus infection of cultured bullfrogs, Rana
catesbeiana, in Brazil." J Wildl Dis 19 (3):180-4.
Beard, DB. 1938. "Everglades National Park Project: Wildlife Reconnaissance." US
Department of Interior, National Park Service. Washington, DC 106.
Bishop, E. J., C. Shilton, S. Benedict, F. Kong, G. L. Gilbert, D. Gal, D. Godoy, B. G.
Spratt, and B. J. Currie. 2007. "Necrotizing fasciitis in captive juvenile
Crocodylus porosus caused by Streptococcus agalactiae: an outbreak and review
of the animal and human literature." Epidemiology and infection 135 (8):1248-
1255. doi: 10.1017/S0950268807008515.
Bouricha, Mehdi, Bernard Castan, Elisabeth Duchene-Parisi, and Michel Drancourt.
2014. "Mycobacterium marinum infection following contact with reptiles:
vivarium granuloma." International Journal of Infectious Diseases 21:17-18. doi:
https://doi.org/10.1016/j.ijid.2013.11.020.
Brandt, Laura A. . 1989. "The status and ecology of the American alligator (Alligator
mississippiensis) in Par Pond, Savannah River site."Dissertation, Biological
Sciences, FIU.
Brisbin, I. Lehr. 1986. A bibliography of the American alligator (Alligator
mississippiensis). Edited by Savannah River Plant National Environmental
Research Park.
Brown, D R, J M Farley, L A Zacher, J M Carlton, T L Clippinger, J G Tully, and M B
Brown. 2001. "Mycoplasma alligatoris sp. nov., from American alligators."
International Journal of Systematic and Evolutionary Microbiology 51 (2):419-
424. doi: doi:10.1099/00207713-51-2-419.
Burton, J. E., I. R. Dorociak, T. E. Schwedler, and C. D. Rice. 2002. "Circulating
lysozyme and hepatic CYP1A activities during a chronic dietary exposure to
tributyltin (TBT) and 3,3',4,4',5-pentachlorobiphenyl (PCB-126) mixtures in
channel catfish, Ictalurus punctatus." J Toxicol Environ Health A 65 (8):589-602.
doi: 10.1080/152873902317349745.
Clark, H. F., and C. C. Shepard. 1963. "EFFECT OF ENVIRONMENTAL
TEMPERATURES ON INFECTION WITH MYCOBACTERIUM MARINUM

46
(BALNEI) OF MICE AND A NUMBER OF POIKILOTHERMIC SPECIES." J
Bacteriol 86:1057-69.
Coico, R., and G. Sunshine. 2015. Immunology: A Short Course: Wiley.
Craighead, Frank Cooper. 1969. The role of the alligator in shaping plant communities
and maintaining wildlife in the southern Everglades: Florida Audubon Society.
Cray, Carolyn, Julia Zaias, and Norman H. Altman. 2009. "Acute phase response in
animals: a review." Comparative medicine 59 (6):517-526.
Debra, L. Miller, J. Mauel Michael, Baldwin Charles, Burtle Gary, Ingram Dallas, E.
Hines Murray, and S. Frazier Kendal. 2003. "West Nile Virus in Farmed
Alligators." Emerging Infectious Disease journal 9 (7):794. doi:
10.3201/eid0907.030085.
Dzik, J. M. 2010. "The ancestry and cumulative evolution of immune reactions." Acta
Biochim Pol 57 (4):443-66.
Evans, Joyce J., Phillip H. Klesius, and Craig A. Shoemaker. 2006. "Therapeutic and
prophylactic immunization against Streptococcus iniae infection in hybrid striped
bass (Morone chrysops x Morone saxatilis)." Aquaculture research 2006 v.37
no.7 (no. 7):pp. 742-750. doi: 10.1111/j.1365-2109.2006.01487.x.
Ferguson, Mark W. J., and Ted Joanen. 1982. "Temperature of egg incubation determines
sex in Alligator mississippiensis." Nature 296 (5860):850-853. doi:
10.1038/296850a0.
Finger, J. 2012. "A review of innate immune functions in crocodilians." CAB Reviews:
Perspectives in Agriculture, Veterinary Science, Nutrition and Natural Resources
7 (067). doi: 10.1079/pavsnnr20127067.
Flajnik, M. F. 2018. "A cold-blooded view of adaptive immunity." Nat Rev Immunol 18
(7):438-453. doi: 10.1038/s41577-018-0003-9.
Frans, I., C. W. Michiels, P. Bossier, K. A. Willems, B. Lievens, and H. Rediers. 2011.
"Vibrio anguillarum as a fish pathogen: virulence factors, diagnosis and
prevention." J Fish Dis 34 (9):643-61. doi: 10.1111/j.1365-2761.2011.01279.x.
G Buckland, Andrew, and David C Wilton. 2000. The antibacterial properties of secreted
phospholipases A2. Vol. 1488.
Glassman, A. B., T. W. Holbrook, and C. E. Bennett. 1979. "Correlation of leech
infestation and eosinophilia in alligators." J Parasitol 65 (2):323-4.

47
Glassman, Armand B., Carol E. Bennett, and Terry C. Hazen. 1981. "Peripheral Blood
Components in Alligator mississippiensis." Transactions of the American
Microscopical Society 100 (2):210-215. doi: 10.2307/3225803.
Guillette, L. J., Jr., T. S. Gross, G. R. Masson, J. M. Matter, H. F. Percival, and A. R.
Woodward. 1994. "Developmental abnormalities of the gonad and abnormal sex
hormone concentrations in juvenile alligators from contaminated and control
lakes in Florida." Environmental health perspectives 102 (8):680-688. doi:
10.1289/ehp.94102680.
Huchzermeyer, F., and W. Van Wyk. 2003. Crocodiles – Biology, husbandry and
diseases. Vol. 74.
Huq, A., E. B. Small, P. A. West, M. I. Huq, R. Rahman, and R. R. Colwell. 1983.
"Ecological relationships between Vibrio cholerae and planktonic crustacean
copepods." Appl Environ Microbiol 45 (1):275-83.
Iwata, A., T. Iwase, Y. Ogura, T. Takahashi, N. Matsumoto, T. Yoshida, N. Kamei, K.
Kobayashi, J. Mestecky, and I. Moro. 2002. "Cloning and expression of the turtle
(Trachemys scripta) immunoglobulin joining (J)-chain cDNA." Immunogenetics
54 (7):513-9. doi: 10.1007/s00251-002-0492-2.
Jacobson, E. R., P. E. Ginn, J. M. Troutman, L. Farina, L. Stark, K. Klenk, K. L.
Burkhalter, and N. Komar. 2005. "West Nile virus infection in farmed American
alligators (Alligator mississippiensis) in Florida." J Wildl Dis 41 (1):96-106. doi:
10.7589/0090-3558-41.1.96.
Jacobson, Elliott. 2007. Infectious Diseases and Pathology of Reptiles: A Color Atlas and
Text.
Joseph, Sam W., Rita R. Colwell, and James B. Kaper. 1982. "Vibrio Parahaemolyticus
and Related Halophilic Vibrios." CRC Critical Reviews in Microbiology 10
(1):77-124. doi: 10.3109/10408418209113506.
Karsten, A. H., and C. D. Rice. 2004. "c-Reactive protein levels as a biomarker of
inflammation and stress in the Atlantic sharpnose shark (Rhizoprionodon
terraenovae) from three southeastern USA estuaries." Mar Environ Res 58 (2-
5):747-51. doi: 10.1016/j.marenvres.2004.03.089.
Keller, J. M., P. D. McClellan-Green, A. M. Lee, M. D. Arendt, P. P. Maier, A. L.
Segars, J. D. Whitaker, D. E. Keil, and M. M. Peden-Adams. 2005. "Mitogen-
induced lymphocyte proliferation in loggerhead sea turtles: comparison of
methods and effects of gender, plasma testosterone concentration, and body
condition on immunity." Vet Immunol Immunopathol 103 (3-4):269-81. doi:
10.1016/j.vetimm.2004.09.029.

48
Kruman, II. 1992. "Comparative analysis of cell replacement in hibernators." Comp
Biochem Physiol A Comp Physiol 101 (1):11-8.
Kvell, K., E. L. Cooper, P. Engelmann, J. Bovari, and P. Nemeth. 2007. "Blurring
borders: innate immunity with adaptive features." Clin Dev Immunol
2007:83671. doi: 10.1155/2007/83671.
Lance, V. A. 2003. "Alligator physiology and life history: the importance of
temperature." Exp Gerontol 38 (7):801-5.
Lang, Jeffrey W., and Harry V. Andrews. 1994. "Temperature-dependent sex
determination in crocodilians." Journal of Experimental Zoology 270 (1):28-44.
doi: 10.1002/jez.1402700105.
Lee, P. T., S. Bird, J. Zou, and S. A. M. Martin. 2017. "Phylogeny and expression
analysis of C-reactive protein (CRP) and serum amyloid-P (SAP) like genes
reveal two distinct groups in fish." Fish Shellfish Immunol 65:42-51. doi:
10.1016/j.fsi.2017.03.037.
Li, Lingxiao, Wang Tao, Yi Sun, Gang Cheng, Hui Yang, Zhiguo Wei, Ping Wang,
Xiaoxiang hu, Liming Ren, Qingyong Meng, Ran Zhang, Ying Guo, Lennart
Hammarström, Ning Li, and Yaofeng Zhao. 2012. Extensive Diversification of
IgD-, IgY-, and Truncated IgY(Delta Fc)-Encoding Genes in the Red-Eared Turtle
(Trachemys scripta elegans). Vol. 189.
Madsen, M., P. Hangartner, K. West, and P. Kelly. 1998. "Recovery rates, serotypes, and
antimicrobial susceptibility patterns of salmonellae isolated from cloacal swabs of
wild Nile crocodiles (Crocodylus niloticus) in Zimbabwe." J Zoo Wildl Med 29
(1):31-4.
Madsen, Mogens. 1993. "Microbial flora of frozen tail meat from captive Nile crocodiles
(Crocodylus niloticus)." International Journal of Food Microbiology 18 (1):71-
76. doi: https://doi.org/10.1016/0168-1605(93)90009-6.
Manolis, S. C., G. J. Webb, D. Pinch, L. Melville, and G. Hollis. 1991. "Salmonella in
captive crocodiles (Crocodylus johnstoni and C. porosus)." Aust Vet J 68 (3):102-
5.
Margiotta, A. L., L. J. Bain, and C. D. Rice. 2017. "Expression of the Major Vault
Protein (MVP) and Cellular Vault Particles in Fish." Anat Rec (Hoboken) 300
(11):1981-1992. doi: 10.1002/ar.23645.
Mateo, M. R., E. D. Roberts, and F. M. Enright. 1984a. "Inflammation induced by
subcutaneous turpentine inoculation of young American alligators (Alligator
mississippiensis)." Am J Vet Res 45 (9):1870-5.

49
Mateo, M. R., E. D. Roberts, and F. M. Enright. 1984b. "Morphologic, cytochemical, and
functional studies of peripheral blood cells of young healthy American alligators
(Alligator mississippiensis)." Am J Vet Res 45 (5):1046-53.
Mazzotti, Frank J., G. Ronnie Best, Laura A. Brandt, Michael S. Cherkiss, Brian M.
Jeffery, and Kenneth G. Rice. 2009. "Alligators and crocodiles as indicators for
restoration of Everglades ecosystems." Ecological Indicators 9 (6,
Supplement):S137-S149. doi: https://doi.org/10.1016/j.ecolind.2008.06.008.
Merchant, M. E., C. M. Roche, D. Thibodeaux, and R. M. Elsey. 2005. "Identification of
alternative pathway serum complement activity in the blood of the American
alligator (Alligator mississippiensis)." Comp Biochem Physiol B Biochem Mol
Biol 141 (3):281-8. doi: 10.1016/j.cbpc.2005.03.009.
Merchant, M., D. Thibodeaux, K. Loubser, and R. M. Elsey. 2004. "Amoebacidal effects
of serum from the American alligator (Alligator mississippiensis)." J Parasitol
90 (6):1480-3. doi: 10.1645/ge-3382.
Merchant, Mark E., Cherie Roche, Ruth M. Elsey, and Jan Prudhomme. 2003.
"Antibacterial properties of serum from the American alligator (Alligator
mississippiensis)." Comparative Biochemistry and Physiology Part B:
Biochemistry and Molecular Biology 136 (3):505-513. doi: 10.1016/s1096-
4959(03)00256-2.
Mondal, S., and U. Rai. 2001. "In vitro effect of temperature on phagocytic and cytotoxic
activities of splenic phagocytes of the wall lizard, Hemidactylus flaviviridis."
Comp Biochem Physiol A Mol Integr Physiol 129 (2-3):391-8.
Montali, R. J. 1988. "Comparative pathology of inflammation in the higher vertebrates
(reptiles, birds and mammals)." J Comp Pathol 99 (1):1-26.
Owen, J. 2013. Immunology: W. H. Freeman.
Parry, R. M., R. C. Chandan, and K. M. Shahani. . 1965. "A rapid and sensitive assay of
muramidase. Experimental Biology and Medicine " Experimental Biology and
Medicine 119:384–386.
Pepys, M. B., and G. M. Hirschfield. 2003. "C-reactive protein: a critical update." J Clin
Invest 111 (12):1805-12. doi: 10.1172/JCI18921.
Percival, Steven L., and David W. Williams. 2014. "Chapter Twelve - Vibrio." In
Microbiology of Waterborne Diseases (Second Edition), edited by Steven L.
Percival, Marylynn V. Yates, David W. Williams, Rachel M. Chalmers and
Nicholas F. Gray, 237-248. London: Academic Press.

50
Pettinello, R., and H. Dooley. 2014. "The immunoglobulins of cold-blooded vertebrates."
Biomolecules 4 (4):1045-69. doi: 10.3390/biom4041045.
Rice, Charles D., and Beverly Anne Weeks. 1989. "Influence of Tributyltin on in Vitro
Activation of Oyster Toadfish Macrophages." Journal of Aquatic Animal Health
1 (1):62-68. doi: 10.1577/1548-8667(1989)001<0062:IOTOIV>2.3.CO;2.
Rice, Kenneth G., and Frank Mazzotti. 2004. American alligator ecology and monitoring
for the Comprehensive Everglades Restoration Plan. In Fact Sheet.
Robey, F A, T Tanaka, and T Y Liu. 1983. "Isolation and characterization of two major
serum proteins from the dogfish, Mustelus canis, C-reactive protein and amyloid
P component." Journal of Biological Chemistry 258 (6):3889-3894.
Rodgers, M. L., C. A. Toline, and C. D. Rice. 2018. "Humoral Immune Responses to
Select Marine Bacteria in Loggerhead Sea Turtles Caretta caretta and Kemp's
Ridley Sea Turtles Lepidochelys kempii from the Southeastern United States." J
Aquat Anim Health 30 (1):20-30. doi: 10.1002/aah.10012.
Russell, WC , and KL Herman. 1970. "Colibacillosis in captive wild animals." Journal
of Zoo Animal Medicine (1):17-21.
Sardelis, M. R., M. J. Turell, D. J. Dohm, and M. L. O'Guinn. 2001. "Vector competence
of selected North American Culex and Coquillettidia mosquitoes for West Nile
virus." Emerg Infect Dis 7 (6):1018-22. doi: 10.3201/eid0706.010617.
Schechter, B., and R. Street. 2000. ""Alligator mississippiensis"." University of
Michigan, accessed March 1.
Shang, Shih-Ta, Sheng-Kang Chiu, Ming-Ching Chan, Ning-Chi Wang, Ya-Sung Yang,
Jung-Chung Lin, and Feng-Yee Chang. 2012. "Invasive Brevundimonas
vesicularis bacteremia: Two case reports and review of the literature." Journal of
Microbiology, Immunology and Infection 45 (6):468-472. doi:
https://doi.org/10.1016/j.jmii.2011.12.021.
Smith, E. Norbert. 1975. "Thermoregulation of the American Alligator, Alligator
mississippiensis." Physiological Zoology 48 (2):177-194.
Song, W. C., M. R. Sarrias, and J. D. Lambris. 2000. "Complement and innate
immunity." Immunopharmacology 49 (1-2):187-98.
Steinman, Amir, Caroline Banet-Noach, Shlomit Tal, Ohad Levi, Lubov Simanov,
Shimon Perk, Mertyn Malkinson, and Nahum Shpigel. 2003. "West Nile virus
infection in crocodiles." Emerging infectious diseases 9 (7):887-889. doi:
10.3201/eid0907.020816.

51
Sullivan, B. M., and R. M. Locksley. 2009. "Basophils: a nonredundant contributor to
host immunity." Immunity 30 (1):12-20. doi: 10.1016/j.immuni.2008.12.006.
Sunyer, J. O., I. K. Zarkadis, and J. D. Lambris. 1998. "Complement diversity: a
mechanism for generating immune diversity?" Immunol Today 19 (11):519-23.
Tipton, Jessica J., Louis J. Guillette, Susan Lovelace, Benjamin B. Parrott, Thomas R.
Rainwater, and Jessica L. Reiner. 2017. "Analysis of PFAAs in American
alligators part 2: Potential dietary exposure of South Carolina hunters from
recreationally harvested alligator meat." Journal of Environmental Sciences
61:31-38. doi: https://doi.org/10.1016/j.jes.2017.05.046.
Trouw, L. A., and M. R. Daha. 2011. "Role of complement in innate immunity and host
defense." Immunol Lett 138 (1):35-7. doi: 10.1016/j.imlet.2011.02.014.
Turell, M. J., M. O'Guinn, and J. Oliver. 2000. "Potential for New York mosquitoes to
transmit West Nile virus." Am J Trop Med Hyg 62 (3):413-4.
Turton, J. A., P. W. Ladds, S. C. Manolis, and G. J. Webb. 1997. "Relationship of blood
corticosterone, immunoglobulin and haematological values in young crocodiles
(Crocodylus porosus) to water temperature, clutch of origin and body weight."
Aust Vet J 75 (2):114-9.
Valenzuela, N., R. Literman, J. L. Neuwald, B. Mizoguchi, J. B. Iverson, J. L. Riley, and
J. D. Litzgus. 2019. "Extreme thermal fluctuations from climate change
unexpectedly accelerate demographic collapse of vertebrates with temperature-
dependent sex determination." Sci Rep 9 (1):4254. doi: 10.1038/s41598-019-
40597-4.
Volanakis, J. E. 2001. "Human C-reactive protein: expression, structure, and function."
Mol Immunol 38 (2-3):189-97.
Wang, Q., B. J. Chang, and T. V. Riley. 2010. "Erysipelothrix rhusiopathiae." Vet
Microbiol 140 (3-4):405-17. doi: 10.1016/j.vetmic.2009.08.012.
Warr, G. W., K. E. Magor, and D. A. Higgins. 1995. "IgY: clues to the origins of modern
antibodies." Immunol Today 16 (8):392-8. doi: 10.1016/0167-5699(95)80008-5.
Work, Thierry M., Julie Dagenais, Renee Breeden, Anette Schneemann, Joyce Sung,
Brian Hew, George H. Balazs, and John M. Berestecky. 2015. "Green Turtles
Have Novel Asymmetrical Antibodies." The Journal of Immunology 195
(11):5452. doi: 10.4049/jimmunol.1501332.

52
Yan, S., D. Marguet, J. Dobers, W. Reutter, and H. Fan. 2003. "Deficiency of CD26
results in a change of cytokine and immunoglobulin secretion after stimulation by
pokeweed mitogen." Eur J Immunol 33 (6):1519-27. doi: 10.1002/eji.200323469.
Zapata, A. G., A. Varas, and M. Torroba. 1992. "Seasonal variations in the immune
system of lower vertebrates." Immunol Today 13 (4):142-7. doi: 10.1016/0167-
5699(92)90112-k.
Zarkadis, I. K., D. Mastellos, and J. D. Lambris. 2001. "Phylogenetic aspects of the
complement system." Dev Comp Immunol 25 (8-9):745-62.
Zhang, X., R. A. Calvert, B. J. Sutton, and K. A. Dore. 2017. "IgY: a key isotype in
antibody evolution." Biol Rev Camb Philos Soc 92 (4):2144-2156. doi:
10.1111/brv.12325.
Zimmerman, L. M., L. A. Vogel, and R. M. Bowden. 2010. "Understanding the
vertebrate immune system: insights from the reptilian perspective." J Exp Biol
213 (5):661-71. doi: 10.1242/jeb.038315.


















