
Human Molecular Genetics - Emory University · PDF fileHuman Molecular Genetics ... Anne-Marie...
12
Human Molecular Genetics doi:10.1093/hmg/ddg220 12:2229-2239, 2003. First published 8 Jul 2003; Hum. Mol. Genet. Anne-Marie Laurent, Meizhang Li, Stephanie Sherman, Gérard Roizès and Jérôme Buard Recombination across the centromere of disjoined and non-disjoined chromosome 21 http://hmg.oxfordjournals.org/cgi/content/full/12/17/2229 The full text of this article, along with updated information and services is available online at References http://hmg.oxfordjournals.org/cgi/content/full/12/17/2229#BIBL This article cites 44 references, 18 of which can be accessed free at Cited by http://hmg.oxfordjournals.org/cgi/content/full/12/17/2229#otherarticles This article has been cited by 2 articles at 5 June 2008 . View these citations at Reprints http://www.oxfordjournals.org/corporate_services/reprints.html Reprints of this article can be ordered at Email and RSS alerting Sign up for email alerts, and subscribe to this journal’s RSS feeds at http://hmg.oxfordjournals.org image downloads PowerPoint® Images from this journal can be downloaded with one click as a PowerPoint slide. Journal information at http://hmg.oxfordjournals.org Additional information about Human Molecular Genetics, including how to subscribe can be found Published on behalf of http://www.oxfordjournals.org Oxford University Press at Emory University Libraries on 5 June 2008 http://hmg.oxfordjournals.org Downloaded from
Transcript of Human Molecular Genetics - Emory University · PDF fileHuman Molecular Genetics ... Anne-Marie...

Human Molecular Genetics
doi:10.1093/hmg/ddg220 12:2229-2239, 2003. First published 8 Jul 2003; Hum. Mol. Genet.
Anne-Marie Laurent, Meizhang Li, Stephanie Sherman, Gérard Roizès and Jérôme Buard Recombination across the centromere of disjoined and non-disjoined chromosome 21
http://hmg.oxfordjournals.org/cgi/content/full/12/17/2229The full text of this article, along with updated information and services is available online at
References http://hmg.oxfordjournals.org/cgi/content/full/12/17/2229#BIBL
This article cites 44 references, 18 of which can be accessed free at
Cited by http://hmg.oxfordjournals.org/cgi/content/full/12/17/2229#otherarticles
This article has been cited by 2 articles at 5 June 2008 . View these citations at
Reprints http://www.oxfordjournals.org/corporate_services/reprints.html
Reprints of this article can be ordered at
Email and RSS alerting Sign up for email alerts, and subscribe to this journal’s RSS feeds at http://hmg.oxfordjournals.org
image downloadsPowerPoint® Images from this journal can be downloaded with one click as a PowerPoint slide.
Journal informationat http://hmg.oxfordjournals.org Additional information about Human Molecular Genetics, including how to subscribe can be found
Published on behalf ofhttp://www.oxfordjournals.org Oxford University Press
at Emory University Libraries on 5 June 2008 http://hmg.oxfordjournals.orgDownloaded from

Recombination across the centromere ofdisjoined and non-disjoined chromosome 21
Anne-Marie Laurent1, Meizhang Li1, Stephanie Sherman2, Gerard Roizes1 and
Jerome Buard1,*
1Institut de Genetique Humaine, CNRS UPR 1142, Montpellier, France and 2Department of Genetics,
Emory University School of Medicine, Atlanta, USA
Received May 21, 2003; Revised and Accepted June 25, 2003 DDBJ/EMBL/GenBank accession no. AF254982
Meiotic recombination is generally suppressed across the centromere of eukaryotic chromosomes. Inhuman, megabase-long satellite sequences and contiguous segmental duplications hamper both physicaland fine scale genetic mapping in regions flanking centromeric DNA. We have developed polymorphicmicrosatellite markers embedded within the duplicated most proximal sequences of the long arm and of theshort arm of chromosome 21 by using paralogous specific bases as anchor points for their specificdetection. Segregation analysis in CEPH reference pedigrees shows that recombination is repressedsignificantly across the centromere of chromosome 21 both in male and in female but not in the mostproximal 21q region in female. Extreme size variations of the alpha-satellite I blocks transmitted in thesefamilies and deduced from quantitative FISH analysis are not correlated with the inter-individual variations ofrecombination activity observed in the peri-centromeric region. Finally, none of 28 families with a trisomy 21child previously associated with a nullitransitional meiosis I non-disjunction event presents a recombinationexchange across the centromere. This confirms that, for this group of errors, the lack of recombination is theprimary susceptibility factor, not abnormal recombination in the centromeric region.
INTRODUCTION
Meiotic exchange is not distributed randomly along eukaryoticchromosomes. The most pronounced departure from unifor-mity is observed near centromeres and telomeres in a widevariety of plant and animal genomes. The centromere exerts anegative effect on recombination activity both within itself andin proximal regions (1).
In human, specific physical and genetic mapping efforts haveshown that sex-average recombination rate per physical unit isreduced by at least one order of magnitude across thecentromere of chromosome 10 (2), chromosome X (3,4),chromosome 5 (5) and in most proximal regions of chromo-some 19 (6). This reduction is more intense in male withrepression extending more than 5 Mb away from the threeautosomal centromeres. Chromosome X analysis demonstratesthat the centromere effect exists in female as well, with a 8-foldsuppresssion across the centromere compared with the averagerate of recombination along the whole chromosome (4).
Several kinds of redundant sequences, including megabase-long satellite DNA and contiguous segmental duplications,hamper the characterization of unique polymorphic markers
within centromeric and juxtacentromeric regions respectively.Indeed, a typical juxta-centromeric region consists of apatchwork of contiguous genomic sequences duplicated onseveral non-homologous chromosomes with high levels ofnucleotide identity between copies (7–9). This complexityaccounts for the lack of reliable physical map and ofchromosome-specific polymorphic markers for most juxta-centromeric regions.
In the absence of such tools, only genome-wide recombina-tion maps and female-to-male genetic distance ratio could sheda light on the centromere effect in human. Metacentricchromosomes present a strong positive correlation betweensex average recombination rates and the physical distance fromcentromere (10–12). This correlation is mainly due to thecentromeric repression in male that is 2–8-fold higher than infemale. In contrast, no significant gender difference has beenobserved in the most proximal regions for acrocentricchromosomes except for chromosome 21, which behaves likea metacentric chromosome in this respect (10). A meta-analysisof recombination along 21q indicates that the femalerecombination rate is higher in the most proximal interval(4 cM/Mb) than along the rest of the chromosome (13) and
*To whom correspondence should be addressed at: Institut de Genetique Humaine, 141 rue de la Cardonille, 34396 Montpellier cedex 5, France.Tel: þ33 499619977; Fax: þ33 499619901; Email: [email protected]
Human Molecular Genetics, 2003, Vol. 12, No. 17 2229–2239DOI: 10.1093/hmg/ddg220
Human Molecular Genetics, Vol. 12, No. 17 # Oxford University Press 2003; all rights reserved

strengthens previous reports showing no suppression ofrecombination within a 2.4 Mb 21q11.1 interval, 400 kb awayfrom the centromeric alphoid tandem repeats (14). However,the proximal boundaries of these maps consist of microsatelliteelements located within the duplication-rich juxtacentromericregion. Whether the microsatellite typed for each CEPH parentis actually located in 21q11.1 thus remains uncertain.Characterization of chromosome-specific marker in this regioncould confirm this unexpectedly high female recombination ratenear the centromere. Also, the level of recombination activityacross the centromere of chromosome 21 is unknown.
Exploring centromeric female recombination activity forchromosome 21 relative to other acrocentric chromosomesmight also be relevant for better understanding of the molecularbasis of segregation errors resulting in trisomy 21. Indeed, thebest documented molecular correlate of meiotic chromosomalnon-disjunction (NDJ) is an abnormal frequency and distribu-tion of recombination events between the two non-disjoinedchromosomes (reviewed in 15,16). Half of NDJ events with thetwo 21q homologues transmitted during the first meioticdivision present no detectable exchange between the twochromosomes. In contrast, those involving two 21q sisterchromatids—so called MII events—are associated with a largeexcess of juxtacentromeric recombination events (17,18). Thelevel of confidence in determining whether an NDJ eventoccurred in MI or MII depends on the genetic distance betweenthe centromere and the most proximal genetic marker used(19). In most recent studies, the proximal microsatellite markerused is located 1.3 Mb away from the alphoid centromericblock (17). Consequently, the relative importance of absence ofrecombination versus centromeric recombination in non-disjunction could be overestimated. However, alphoid poly-morphism analysis of non-disjoined chromosome capturedindividually in hybrid cells failed to reveal any 21q proximalexchange among 10 informative trisomy events (20).Therefore, if they exist, the undetected recombination eventsmust occur within the very proximal 21q region, within thecentromeric region itself or along the short arm of thechromosome.
A second molecular correlate with meiotic non-disjunction ofchromosome 21 is the size of the alpha-satellite repeat array.Alpha-satellite, the major type of human centromeric tandemrepeat, is present at all human centromeres and is variable insize between homologs for most chromosome pairs (21–24).An association between small alphoid size of chromosome 21and maternal non-disjunction has been reported in twoindependent studies, with different methods for size estimationof the chromosome 21 alpha-satellite I blocks (25,26). Onanother hand, satellite sequences constitute an important part ofcentromeric heterochromatin and heterochromatic regions havelong been recognized as a poor substrate for recombinationregardless of their chromosomal location (27). As suggested bygenetic mapping analysis of Drosophila X chromosomes withpartial deletions of centromeric heterochromatin, the largeblocks of satellite DNA may influence the centromere effect(28). Whether small alphoid size and altered recombinationdistribution are correlated with non-disjunction of humanchromosome 21 in dependent mechanistic pathways—with thealphoid block influencing recombination distribution—remainsto be investigated.
We report here the characterization of juxta-centromericmarkers for chromosome 21, including a short arm micro-satellite marker, and the first estimates of recombinationactivity in male and in female across and near the centromerefor both correctly and incorrectly disjoined chromosome 21.We have also investigated whether the size of the alpha-satellitecentromeric DNA influences juxta-centromeric recombination.
RESULTS
Extension of the 21p contig sequence usingparalogous duplications
Homology search in the BAC end sequence database using the281 kb 21p contig sequence reported previously (AL163201)(29) identified BAC insert 2503J9 (AF254982.1), the T7 end ofwhich presents 100% identity with the telomeric end of the 21pcontig. Sequence analysis of this BAC insert revealed that threecontigs (nos 5, 14 and 16) out of its 16 unordered contigs (30April 2000 version) contain unique, non-tandemly repeatedDNA, the remaining contigs consisting of satellite 3 and alpha-satellite sequences. Homology search in the human sequencedatabase using ends of each of these three ‘non-satellite’ contigsidentified two BAC sequences (AC027723 and AL139386)assigned to chromosome 10 and chromosome 13 respectively,each of them overlapping with contigs 5 and 14 and with contigs14 and 16 respectively, with more than 98% nucleotide identity(Fig. 1). This suggested both an order for these three contigs anda size for each sequence gap between them. PCR amplificationof BAC 2503J9 DNA across the two predicted gaps followedby sequencing of the PCR products were then performed andconfirmed these predictions. Finally, a 56 026 bp contig wasassembled that includes 2.7 kb of sequence overlapping with thedistal end of the previously reported 21p contig (99.7%nucleotide identity; see Materials and Methods) followed by45.6 kb consisting of non-tandemly repeated DNA plus, towardsthe telomere, 7.6 kb of the satellite 3 block. An independenteffort produced by others at the Institute of Molecular Biologyof Iena, and aimed at completing the AF254982 sequence, veryrecently confirmed the order between contig 5 and contig 14(Dr Platzer, personal communication).
Microsatellite markers in chromosome 21juxta-centromeric sequences
Fifteen microsatellite elements were analysed, including 11short tandem repeats (STRs) retrieved from the most proximalsequences of 21q (250 kb scanned) and 21p (326.6 kb), twomicrosatellites detected within a 41 kb 21p BAC insert notmapped precisely yet (BAC B1L1C6; A. DeSario, personalcommunication) and two proximal 21q microsatellite markersdescribed elsewhere (30,31). In silico homology search usingthe BLAST tool (32) in human sequence databases togetherwith PCR amplification data using DNAs of a monochromo-somal hybrid cell line panel showed that none of these 15elements is found exclusively as a unique copy within thejuxtacentromeric region of chromosome 21 (Table 1).
Paralogous copies of D21S215 (30) and of D21S369 (31)have been found in databases within sequences assigned to
2230 Human Molecular Genetics, 2003, Vol. 12, No. 17

chromosomes 13, 18 and 10 (Table 1). Experimental data confirmthat they are present on chromosome 10, 18 and 21 but not onchromosome 13, strongly suggesting incorrect chromosomeassignation for this sequence in the database. D21S215 istherefore contained in an interchromosomal duplication butalso in an intrachromosomal tandem duplication as it is presentin several copies on each of chromosome 21 (two copies),chromosome 18 (two copies) and chromosome 10 (four copies).For the remaining 13 other STRs,�600 bp of sequence flanking,on one side, paralogous copies of each STR were obtained bysequencing the PCR products amplified from monochromosomalDNAs. Multiple alignment between sequences produced experi-mentally plus those available in the sequence databases revealeda high level of nucleotide identity between paralogous copies,often close to allelic levels (less than 1/500 base substitutionaldifference between pairs of sequences). For the six most proximal21p microsatellites plus one unassigned 21p microsatellite(B1L1C6.CA) there was no flanking base present only on chro-mosome 21. Visual inspection of electropherograms of PCRproducts amplified from chromosome 21, 13 and 15 revealedseveral double peaks scattered along the sequence, indicating thatat least two distinct but highly similar copies were present on eachof these chromosomes for the seven microsatellites. Furthermore,the same nucleotidic differences found between these intra-chromosomal copies were observed for the three different acro-centric chromosomes and fewer sequence variations were foundbetween inter-chromosomal copies. This suggests that duplica-tion occurred first within a chromosome short arm, before therecent (or ongoing) inter-chromosomal spreading of each copy.
Only in the six remaining cases was a base at a variant site inthe flanking sequence specific for the 21 juxta-centromericcopy. These were used to detect the microsatellite either bydesigning PCR primers specific for the variant nucleotide at
their 30 end or by restriction analysis following PCR, the variantbase creating a restriction polymorphism between paralogouscopies (Fig. 2). In three cases out of six, the unique STRdetected was polymorphic between unrelated individuals. Ahighly polymorphic CA dinucleotide repeat (2503J9.TG), with80% heterozygosity and 12 alleles among CEPH parents, waslocated 800 bp proximal from the satellite 3 block in 21p10(BAC insert 2503J9). An imperfect CYTT tetranucleotiderepeat (f92b11.tetra) and a TA dinucleotide repeat (f92b11.TA)located 550 bp away from each other, in 21q11.1, at 214 kb fromthe centromeric alphoid block (within clone f92b11) (29)presented respectively two and three alleles among CEPHparents. Analysis of inheritance of alleles of f92b11.TA and of2503J9.TG microsatellite showed that one of the two alleles wasnot detected for two and four individuals, respectively, among57 unrelated grand-parents. Moreover, the paralogous specificbase used for the detection of 2503J9.TG, that had beendetected by sequencing monochromosomal hybrid cell DNA ofchromosome 21, was not present in the sequence available indatabase and assigned to chromosome 21 (AF254982; Fig. 2).These two observations suggest that the nucleotidic variationwhich allows discrimination of chromosome 21 juxtacentro-meric microsatellite from other paralogous copies in the twocases was not present on all chromosomes 21, being an allelicvariant base as well as a paralogous variant base.
Recombination rates across and near the centromereof chromosome 21
Estimates of recombination rates per physical unit across andnear the centromere (Fig. 3) were obtained from CRIMAPanalyses of segregation data in 40 CEPH reference families ofthe two juxtacentromeric markers f92B11.TA and 2503J9.TG
Figure 1. Juxtacentromeric sequences of human chromosome 21. Proximal 21p contig sequences AL163201 and 21q contig sequences AL163202 are shown oneach side of the centromeric alpha-satellite I block (stripped box interrupted, CEN). Thirteen microsatellite elements among which 11 have a known location (ver-tical ticks) have been chosen all along the region for developing polymorphic markers. Sequence AF254982 of BAC insert 2503J9 extends the 21p contig towardsthe telomere, up to the satellite 3 block (dotted box) and the alpha-satellite II blocks (stripped boxes). Its proximal end shows 99.7% identity over a 2.7 kb overlapwith the distal end of contig AL163201. The upper box magnifies the proximal end of sequence AF254982. This end comprises three contigs which have beenassembled in silico using highly homologous paralogous sequences AC027723 and AL139386. This organization was confirmed experimentally by PCR ampli-fication across the putative gaps (arrows) and sequencing of the resulting products.
Human Molecular Genetics, 2003, Vol. 12, No. 17 2231
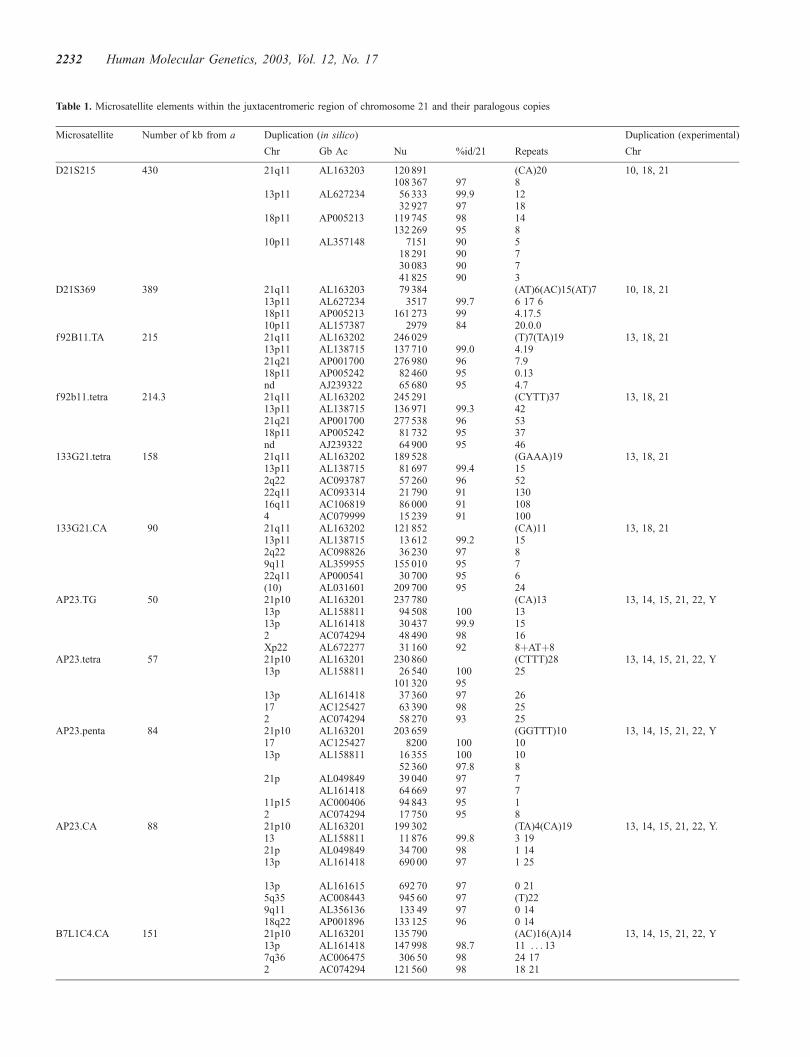
Table 1. Microsatellite elements within the juxtacentromeric region of chromosome 21 and their paralogous copies
Microsatellite Number of kb from a Duplication (in silico) Duplication (experimental)
Chr Gb Ac Nu %id/21 Repeats Chr
D21S215 430 21q11 AL163203 120 891 (CA)20 10, 18, 21108 367 97 8
13p11 AL627234 56 333 99.9 1232 927 97 18
18p11 AP005213 119 745 98 14132 269 95 8
10p11 AL357148 7151 90 518 291 90 730 083 90 741 825 90 3
D21S369 389 21q11 AL163203 79 384 (AT)6(AC)15(AT)7 10, 18, 2113p11 AL627234 3517 99.7 6 17 618p11 AP005213 161 273 99 4.17.510p11 AL157387 2979 84 20.0.0
f92B11.TA 215 21q11 AL163202 246 029 (T)7(TA)19 13, 18, 2113p11 AL138715 137 710 99.0 4.1921q21 AP001700 276 980 96 7.918p11 AP005242 82 460 95 0.13nd AJ239322 65 680 95 4.7
f92b11.tetra 214.3 21q11 AL163202 245 291 (CYTT)37 13, 18, 2113p11 AL138715 136 971 99.3 4221q21 AP001700 277 538 96 5318p11 AP005242 81 732 95 37nd AJ239322 64 900 95 46
133G21.tetra 158 21q11 AL163202 189 528 (GAAA)19 13, 18, 2113p11 AL138715 81 697 99.4 152q22 AC093787 57 260 96 5222q11 AC093314 21 790 91 13016q11 AC106819 86 000 91 1084 AC079999 15 239 91 100
133G21.CA 90 21q11 AL163202 121 852 (CA)11 13, 18, 2113p11 AL138715 13 612 99.2 152q22 AC098826 36 230 97 89q11 AL359955 155 010 95 722q11 AP000541 30 700 95 6(10) AL031601 209 700 95 24
AP23.TG 50 21p10 AL163201 237 780 (CA)13 13, 14, 15, 21, 22, Y13p AL158811 94 508 100 1313p AL161418 30 437 99.9 152 AC074294 48 490 98 16Xp22 AL672277 31 160 92 8þATþ8
AP23.tetra 57 21p10 AL163201 230 860 (CTTT)28 13, 14, 15, 21, 22, Y13p AL158811 26 540 100 25
101 320 9513p AL161418 37 360 97 2617 AC125427 63 390 98 252 AC074294 58 270 93 25
AP23.penta 84 21p10 AL163201 203 659 (GGTTT)10 13, 14, 15, 21, 22, Y17 AC125427 8200 100 1013p AL158811 16 355 100 10
52 360 97.8 821p AL049849 39 040 97 7
AL161418 64 669 97 711p15 AC000406 94 843 95 12 AC074294 17 750 95 8
AP23.CA 88 21p10 AL163201 199 302 (TA)4(CA)19 13, 14, 15, 21, 22, Y.13 AL158811 11 876 99.8 3 1921p AL049849 34 700 98 1 1413p AL161418 690 00 97 1 25
13p AL161615 692 70 97 0 215q35 AC008443 945 60 97 (T)229q11 AL356136 133 49 97 0 1418q22 AP001896 133 125 96 0 14
B7L1C4.CA 151 21p10 AL163201 135 790 (AC)16(A)14 13, 14, 15, 21, 22, Y13p AL161418 147 998 98.7 11 . . . 137q36 AC006475 306 50 98 24 172 AC074294 121 560 98 18 21
2232 Human Molecular Genetics, 2003, Vol. 12, No. 17

plus 22 other 21q markers, up to 6.6 Mb (6597 kb) away from thealphoid block (D21S1905). The location of these markers alongthe sequence map of chromosome 21 (29) was preciselydetermined from BLAST homology analyses. Using theCHROMPIC function of CRIMAP, 71 female and 10 malerecombination events could be mapped in this region, withinintervals as small as 44 kb (D21S409–D21S364). As norecombination event was detected between the three mostproximal 21q11 loci (f92b11-TA, D21S369 and D21S215),these were considered together as a megalocus. As none of theuninformative families for these three markers was informativefor f92b11-tetra, this marker was not used for segregationanalyses. In the males, no recombination event among 146double informative meioses was found between this 21q11megalocus and the 21p10 locus (2503J9.TG), an intervalspanning 2.3 Mb on average (see below for size estimation ofalphoid blocks). Two crossovers among 298 informative malemeioses were found in the proximal 2 Mb region, betweenthis most proximal megalocus and a set of four loci clusteredwithin 59 kb (D21S192, D21S1231, D21S415, D21S1234; onlyD21S192 is presented in Fig. 3 out of these four). Eightcrossovers among 236 male informative events were found in thedistal 3.7 Mb interval (D21S1234–D21S11). These figuresindicate a strong repression of recombination across and also atdistance from the centromere in male. In the females, onerecombination event was found among 153 double informativemeioses within the alphoid-centromeric interval delineated by
the 21p10 marker and the 21q11 most proximal megalocus.Twenty-six crossovers among 273 informative meioses could bemapped unambiguously within the most proximal 2 Mb region,including nine crossovers within the region proximal to D21S408,and 41 crossovers among 255 informative meioses were foundwithin the distal 3.8 Mb interval (D21S1231–D21S1905). Themarkers considered arbitrarily for delineating these two intervalswere not informative in the three remaining cases. Therefore,the repressive effect upon recombination across the centromerewas also observed in the females, with an �10-fold reduction ofrecombination activity (0.23 cM/Mb) compared with chromo-some average (12,13). However, in contrast to the male, thisrepressive effect was restricted to the centromeric interval in thefemales, the most proximal interval of 21q11, at 389 kb awayfrom the alphoid centromeric block, showing a 4.3 cM/Mbrecombination rate. A female-specific recombinationally hyper-active region lay within a 359 kb interval (D21S1256–D21S406),at 5 Mb away from the centromeric alphoid block. Withinthis interval, a maximum recombination rate occurred at over12 cM/Mb within the 44 kb interval D21S409–D21S364[reported in Lynn et al. (13) as 1.2 cM/44 kb].
Inter-individual variations in recombination eventsdistribution and in size of alpha-satellite block
Some mothers among the seven largest CEPH families presentmore recombination events within the proximal 6.6 Mb 21q
Table 1. (Continued)
Microsatellite Number of kb from a Duplication (in silico) Duplication (experimental)
Chr Gb Ac Nu %id/21 Repeats Chr
7q36 AC104692 184 25 90 10 8B15L0C0.TG 273 21p10 AL163201 133 10 (AC)14 13, 15, 21, Y
13 AL139386 482 10 99.9 1810 AC027723 118 830 99.6 2013q11 AL590076 195 70 97 19Yq11 AC019099 360 30 95 1213q12 AL359538 616 80 95 2013q12 AL354798 126 785 94 2013q32 AL137058 927 60 89 2213q14 AL158066 634 50 89 25
2503J9.TG 340 21p10 AF254982 832 30 (TG)18 13, 15, 21, Y10 AC027723 870 90 99.5 13nd AC084096 394 25 99.2 13nd AC127381 198 000 97 1314 NG001019 654 400 96 1814q32 AC013778 995 20 90 8
B1L1C6.CA nd 21p AL049849 115 40 (CA)20 7, 13, 15, 21, 22, Y13 AL161418 914 30 99.6 217q36 AC005631 126 050 95 21
B1L1C6.tetra nd 21p AL049849 175 45 (TAAA)7 7, 13, 15, 21, 22, Y13 AL161418 862 65 99.4 77q36 AC005631 120 040 97 6
For each of 15 short tandem repeats (named after its D number or clone name), the first row presents information concerning the chromosome 21 juxtacentromericcopy of the microsatellite. This includes the distance from the centromeric alpha-satellite block (number of kb from a), the chromosome band assignation (Chr), thenucleotide coordinate of the element (Nu) within the chromosome 21 contig sequence (GenBank accession number; Gb Ac), the number and type of repeats of themicrosatellite and the chromosomes on which paralogous copies have been detected experimentally by PCR screening of a monochromosomal hybrid cell panel.The following rows give the same information for paralogous copies of the microsatellite found within sequence databases together with the percentage ofnucleotide identity with the chromosome 21 juxtacentromeric microsatellite locus comprising 1000 bp of flanking sequence on each side of the tandem array (%id/21). Chromosome assignation of paralogous sequences available in databases can be erroneous and then conflict with experimental data. Conversely, paralogouscopies available in sequence databases, especially most diverged ones, could be undetected by PCR amplification from monochromosomal hybrid cell DNA.
Human Molecular Genetics, 2003, Vol. 12, No. 17 2233

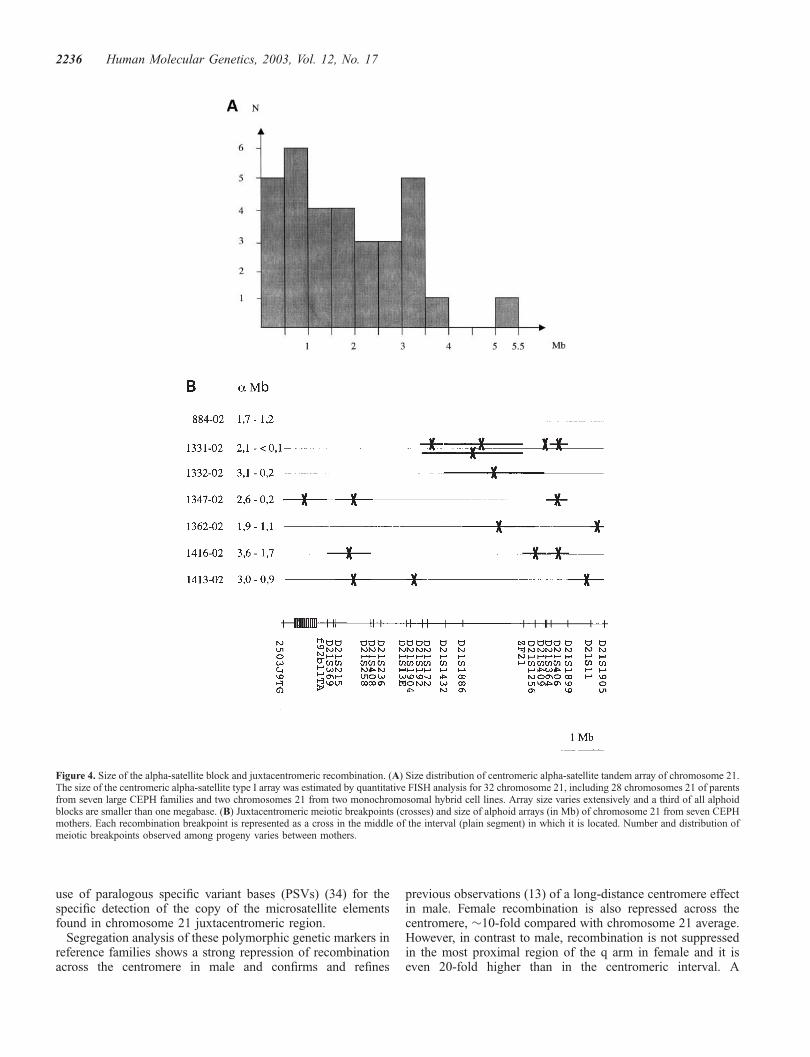
region than others, extremes being family 884 with no femalerecombination event in the region among five children havinginherited a recombinant chromosome and family 1331 showingfive maternal recombination events in this region among sevenrecombinant children. This observation is reminiscent of inter-individual variations observed in the overall frequency ofrecombination in female (10,12) and it also suggests that thedistribution of juxtacentromeric recombination varies betweenwomen. We hypothesized that the size of the alphoid DNAcould influence in cis the extent of repression of recombinationnear 21cen. The size of the alpha-satellite blocks of chromo-some 21 was therefore estimated by quantitative FISH(Q-FISH) using both alpha-satellite I probe aRI-680 and anon-repeated YAC DNA probe, located in 21q22, as aninternal standard (25). Relationship between the intensity of aFISH signal and the actual physical size of the correspondingalpha-satellite block was deduced from pilot Q-FISH experi-ments with four chromosome 21 (one individual plus two
monochromosomal hybrids) for which alphoid size had beenestimated previously by Pulse Field Gel Electrophoresis andSouthern blot analyses (23 and data not shown). Alpha-satellitetype I block of chromosome 21 varied considerably in sizebetween individuals, from less than 100 kb to 5.3 Mb (1.8 Mbon average) among 32 chromosome 21 (Fig. 4A), includingthose of the parents of the seven largest CEPH families.However, we found no strong relationship between distributionof recombination events in the pericentromeric region and sizeor combined size of the alphoid block of the two recombiningchromosome 21 for seven CEPH mothers (Fig. 4B).
Juxtacentromeric recombination and non-disjunction
To further examine the characteristics of recombination onchromosome 21 non-disjunction, 67 father–mother–trisomicchild trios for which no recombination event had been detectedpreviously along the two non-disjoined maternal 21q
Figure 2. Alignments between flanking sequences of microsatellite elements found in chromosome 21 juxta-centromeric regions and their paralogous copies.Chromosomal assignment of each sequence is given with its GenBank accession number when available. Other sequences were obtained by direct sequencingof PCR products amplified from DNA of monochromosomal hybrid cell lines (NA prefix). Only sequences of primer binding sites and of restriction sites (under-lined) used for the specific detection of the chromosome 21 juxta-centromeric copy of each microsatellite are shown with their nucleotidic coordinate in the 21juxta-centromeric GenBank sequence. Only nucleotide variants relative to the chromosome 21 target sequence are shown for paralogous copies. (A) A tetranucleo-tide microsatellite repeat present in 21q11 (f92b11.Tet) is detected after BclI restriction digest of the PCR product amplified with primer f92b11.Tetra F (left site)and primer f92b11.Tetra E (right site). (B) The TA dinucleotide repeat f92b11.TA is discriminated from paralogous copies by RsaI cleavage of PCR productamplified with primer f92b11.TAnD (left site) and primer f92b11.TAnE (right site). (C) The 21p10 copy of dinucleotide repeat 2503J9.TG is specifically amplifiedwith the copy-specific primer 2503J9.TGX (left site) and primer 2503J9.TG5 (right site).
2234 Human Molecular Genetics, 2003, Vol. 12, No. 17

homologues (17) were typed with the juxtacentromeric markers.The hypothesis that we explored was that these errors were notrelated to the lack of recombination, but instead to a centromericrecombinant event that had not been identified using onlypericentromeric markers on 21q.
The mother was heterozygous for the 21p10 marker in eachof 26 trios (Table 2). In three cases, the mother, the father andthe child shared the same two bands and therefore they wereuninformative. Overall, 23 trios were thus informative for the21p10 marker. In two of the 23 informative cases, the fathershowed one band, the mother was heterozygous and her childshowed only the two maternal alleles that were of the sameintensity. Given the results from the CEPH families presentedabove, the likely explanation for these two cases is that thefather transmitted a null allele to the trisomic child.
Ten trios were informative for the 21q11 proximal marker,f92b11.TA, among which five were already informative withthe 21p10 maker. All 10 cases, including seven cases solved bydosage analysis and three cases for which the three alleles of
the trisomic child were distinct, showed transmission of the twomaternal alleles.
Therefore, none of the non-disjoined chromosomes that wereinformative for at least one of the two juxtacentromeric markersshow a recombination event within the newly exploredcentromeric region (23 cases) or within the most proximaljuxtacentromeric interval (five additional cases). These resultssupport the original hypothesis that the non-disjunctional errorin this subset of cases was associated with the lack ofrecombination, not undetected centromeric recombination.
DISCUSSION
We have characterised unique microsatellite markers embeddedwithin the highly duplicated sequences found on both sides ofthe alpha-satellite centromeric block of chromosome 21. Toachieve this, we extended 21p sequence contig and developedan approach derived from ARMS-PCR (33) and based on the
Figure 3. Recombination activity in the juxtacentromeric region of chromosome 21 in CEPH reference families. Recombination rate per physical unit (cM/Mb) infemale (black dots, solid line) and in male (white dots, dashed line) have been deduced from segregation data of 24 markers delineating intervals of known physicalsize, up to 6.6 Mb away from the alphoid centromeric block (hatched box).
Human Molecular Genetics, 2003, Vol. 12, No. 17 2235
use of paralogous specific variant bases (PSVs) (34) for thespecific detection of the copy of the microsatellite elementsfound in chromosome 21 juxtacentromeric region.
Segregation analysis of these polymorphic genetic markers inreference families shows a strong repression of recombinationacross the centromere in male and confirms and refines
previous observations (13) of a long-distance centromere effectin male. Female recombination is also repressed across thecentromere, �10-fold compared with chromosome 21 average.However, in contrast to male, recombination is not suppressedin the most proximal region of the q arm in female and it iseven 20-fold higher than in the centromeric interval. A
Figure 4. Size of the alpha-satellite block and juxtacentromeric recombination. (A) Size distribution of centromeric alpha-satellite tandem array of chromosome 21.The size of the centromeric alpha-satellite type I array was estimated by quantitative FISH analysis for 32 chromosome 21, including 28 chromosomes 21 of parentsfrom seven large CEPH families and two chromosomes 21 from two monochromosomal hybrid cell lines. Array size varies extensively and a third of all alphoidblocks are smaller than one megabase. (B) Juxtacentromeric meiotic breakpoints (crosses) and size of alphoid arrays (in Mb) of chromosome 21 from seven CEPHmothers. Each recombination breakpoint is represented as a cross in the middle of the interval (plain segment) in which it is located. Number and distribution ofmeiotic breakpoints observed among progeny varies between mothers.
2236 Human Molecular Genetics, 2003, Vol. 12, No. 17

recombinationally hyperactive female-specific region has alsobeen localized at 5 Mb away from the centromere betweenD21S409 and D21S364. This high level of recombinationactivity was already noticeable in a previous study (13), butadding segregation data of other nearby markers confirmed thisobservation and refined the localization of this hot region.Further refinement of the genetic map in this short intervalmight help fine-scale characterization of the first femalerecombination hot-spot(s) in human.
The mechanistic basis of non-disjunction of chromosome 21,leading to Down syndrome, is poorly understood. The twoknown molecular correlates with segregation errors ofchromosome 21 are altered recombination and size of thecentromeric alphoid block. Two independent studies havesuggested that prevalence of small alphoid blocks is higheramong mothers with a trisomic child than among correctlydisjoined chromosomes 21 (25,26). Our study, which repre-sents the first attempt to examine whether these two differentobservations share a common mechanism, suggests that thetwo effects are independent of each other. Size of the alpha-satellite block does not have a major effect on frequency ofpericentromeric recombination. Segregation errors involvingalphoid size could be more closely related to the centromerefunction itself than to the repression of recombination thatthe centromere exerts in its vicinity. Such an hypothesis hasbeen tested previously and no relationship could be foundbetween centromeric alphoid size of human chromosome Yandfrequency of disomy in sperm (35). However, as it seems thatnon-disjunction results from the sum of various phenomena,each having a minor influence in the final outcome, onlystudies involving a much larger number of families (withoutand with trisomy 21 children) could rule out subtle relation-ships between these phenomena.
We show here that none of 28 informative non-disjoinedchromosome 21 previously typed as nullitransitional homo-logs (i.e. without detectable crossover event) present a
recombination event across the centromeric region or in mostproximal sequences. This strengthens previous analysis of such21q nullitransitional MI non-disjoined chromosomes capturedindividually in hybrid cells that indicated that proximalrecombination in 21q is infrequent in trisomy-generatingmeioses (20). However, the discrepancies observed betweenmeiotic stage determination based on proximal 21q markersand those based on 21p chromosome heteromorphisms (36,37)must still be considered. These could be attributed torecombination along short arm sequences located distally fromthe 21p10 marker described here, at least 340 kb distal from thealphoid centromeric block. Recent analysis of crossoverdistribution along chromosome 21 in Down syndrome patientspresenting a (14q21q) robertsonian translocation suggests thatthis illegitimate exchange between short arms interferes withthe location of crossover between long arms (38). In turn and asshown for free trisomy 21 events (17), these distal exchangesbetween long arms would increase the probability of asegregation error. Legitimate recombination activity along theshort arm of chromosome 21 therefore remains to beinvestigated. In addition, the relationship between alphoidsequences and non-disjunction associated with detectablepericentromeric recombination described previously (17,18)needs to be further investigated.
The approach for characterizing chromosome-specific poly-morphic markers embedded within duplication-rich regionsshould open the route to recombination analysis across mosthuman centromeres and also within short arms of acrocentricchromosomes. However, only three markers were developed inthis study out of 13 microsatellite elements tested because it wasnot possible, for most microsatellites, to find one variant basenot shared with any other paralogous copies and close enough tothe tandem repeat to be useful. This was due both to anextremely high nucleotide identity between copies, often above99%, and also to the high number of inter- and intra-chromosomal paralogous copies. The extension of the contigsequence of chromosome 21p reported here relies on a 2772 bpoverlap with five base substitutions between the distal end of theexisting contig and the new clone sequenced. Although thisfigure is consistent with estimates of nucleotide diversity forchromosome 21 (39,40), the extremely high level of identitybetween non-homologous chromosomes in these regions makesit still possible that this clone originates from anotherchromosome. It could also originate from a paralogousduplication on chromosome 21p, on the distal side of thesatellite 3 block. This would reconcile the discrepancy betweenthe distribution of satellite blocks on the short arm suggested bythe present analysis (cen> satellite 3> alpha II> tel) and theone reported previously (41). In this latter case, the micro-satellite typed here and embedded in this clone would be locatedless than 400 kb further distal from the centromeric alphoidblock, a figure that would not substantially change the overallpicture of repressed recombination activity across the region.A better physical characterization of the short arm of allacrocentric chromosomes could respond to this uncertainityas the high probability of physical mapping errors in theseregions originates in part from the fact that very few of theparalogous copies are properly mapped. Further, in-depthanalysis of the level of nucleotide identity between andalong short arm sequences of acrocentric chromosomes
Table 2. Summary of the genotype results of the two most proximal cen-tromeric markers among the trios in which there was a non-disjunctional errorof maternal origin
Mating type(father�mother)
Trisomicchild
2503J9.TG21p10
f92b11.TA21q11
ab� cd acd 15 3ab� (cd?)a acd 1 —ab� ac aac 5 7a(—)b� bc (—)bc 2 —ab� ab abbc 3 —Total informative 23 5d
aOne case for which the maternal genotype could not be obtained, the trisomicchild presents three bands, two of them were different from the fathers alleleand were therefore deduced to be the maternal alleles.bTwo fathers present only one band that has not been transmitted to the trisomicchild. This is interpreted as segregation of a null allele, as evidenced in CEPHfamilies.cThe parental origin of the child’s alleles can not be determined in case bothparents show the same heterozygous genotype.dFive cases that were not informative with the 21p10 marker were informativewith the 21q11 most proximal marker, adding up to 28 families that wereinformative with one or the other juxtacentromeric marker.
Human Molecular Genetics, 2003, Vol. 12, No. 17 2237

should allow the characterization of new specific polymorphicmarkers and should also provide hints concerning themechanisms underlying exchanges between non-homologouschromosomes.
MATERIALS AND METHODS
Extension of the 21p contig sequence
The TIGR BAC end sequence database (www.tigr.org/tdb/humgen/bac_end_search.html) was screened with sequenceAL163201. The T7 end of BAC insert 2503J9 was detected,showing 100% identity over 553 overlapping bases with thetelomeric end of the contig. The BAC insert was sequenced inthe genome sequencing center of the Institute of MolecularBiotechnology in Jena (Germany). Size estimates of the wholeBAC insert and of the satellite blocks that it contains werededuced from pulse-field gel electrophoresis and Southern blotanalyses and were consistent with the figures given bysequence analysis (data not shown).
Using the BLAST tool, the T7 end of the sequenced BAC2503J9 (AF254982.1) showed 16 mismatches over 2772overlapping bases with the 21p contig sequence AL163201,including three mismatches within polyA tracts. Elevenmismatches including these latter three plus eight basesubstitutions were found grouped within a 421 base-longregion. Re-sequencing of this short region amplified fromDNA of BAC 2503J9 using PCR primers 2503J9-olpB8.F(50 GACATGAATGAAAGCCCAGT 30) and primers 2503J9-olpB8.R (50 GGTCTGCACACTTGCAATGC 30) showed thatfour mismatches out of these eight base substitutions weresequencing errors and were, instead, identical to the corres-ponding AL163201 bases. Recent re-sequencing of the wholeregion in the Genome Sequencing laboratory of Iena confirmedthat these four mismatches plus four others outside the 421 bpregion were errors. The level of nucleotide identity betweenAL163201 and AF254982 over the overlap is therefore 99.8%(five base substitutions/2772 bp), excluding short tandemrepeat polymorphisms.
Detection of juxtacentromeric microsatellite elements
Microsatellite elements were sought in silico in the 250 kb mostproximal sequences of 21q, in the 326.6 kb chromosome 21psequence and in 21p sequence AL049849 (BAC B1L1C6)with the Tandem Repeat Finder program (42) (http://tandem.biomath.mssm.edu/trf.html). Mononucleotide repeats andmicrosatellite elements containing less than 10 perfect repeatsor flanked with dispersed repeats on each side were notconsidered further for analysis. The BLAST tool (32) was usedto screen non-redundant (nr) and unfinished (htgs) humangenomic sequence databases with the flanking sequences of theselected microsatellite elements in order to detect sequence ofparalogous copies. The ClustalW multialignment program (43)was used to detect variant bases between flanking sequences ofthese paralogous copies.
For dinucleotide repeat 2503J9.TG, the 50 end of primer2503J9.TG5 (50 GGATCAATGCCCACTCCATGTC 30) wasradioactively labelled with P33 using polynucleotide kinase.
The microsatellite was then amplified from �100 ng genomicDNA, using 0.4 mM of this labelled primer, 0.4 mM of thecopy-specific primer 2503J9.TGX (50 AACAGCTTGATGAA-CGGGT 30) and one unit of Taq DNA polymerase (Promega)in a buffer described previously (44) and in 20 ml volume.Cycling conditions in a Hybaid PCR express thermocyclerwere: 94�C, 1 min followed by 30 cycles at 94�C, 20 s,66�C, 20 s and 72�C, 1 min. Dinucleotide repeat f92b11.TAwas amplified using the labelled primer f92b11.TanE (50
GACCCTGACTTCCTGATTTG 30) and primer f92b11.TanD(50 TTCTGTCCACCCTTCACAAG 30) in the same conditionsexcept for annealing temperature (58�C). Following amplifica-tion, f92b11.TA PCR product was ethanol-precipitated, washed,resuspended in 5 ml water and subjected to 2 h RsaI digest(1 unit). In the two cases, the products were subsequentlysubjected to electrophoresis in a 35 cm long denaturing (urea7 M) 5% polyacrylamide gel (acrylamide : bisacrylamide 39 : 1)and the gel was dried and exposed to an autoradiography film.
Tetranucleotide repeat f92b11.Tet was similarly amplifiedwith unlabelled primers f92b11.TetE (50 GCAAAGTATTTG-CTTATTGACTG 30) and f92b11.TetF (50
GAAGCTTCCAAA-CATGGCG 30) at 58�C annealing tem-perature followed by BclI restriction of the PCR product.Restriction digest products were then subjected to electrophor-esis in a 4.5% non-denaturing 20 cm long polyacrylamide gelwhich was subsequently stained in ethidium bromide solution.
ACKNOWLEDGEMENTS
We thank C. de Toma and H. Cann for the cell lines of CEPHparents, Y. Usson for valuable help on quantitative FISH imageanalysis, A. Rosenthal’s laboratory for initial sequencing of BAC2503J9, Matthias Platzer for help on merging its recent update ofthis sequence with ours and colleagues for useful discussion.This work was funded by the Fondation pour la RechercheMedicale, by the Association Francaise pour la Recherche sur laTrisomie 21 and by NIH R01 HD38979 (S.L.S.).
REFERENCES
1. Choo, K.H.A. (1998) Why is the centromere so cold? Genome Res., 8,81–82.
2. Jackson, M.S., Gee See, C., Mulligan, L.M. and Lauffart, B.F. (1996) A9.75-Mb map across the centromere of human chromosome 10. Genomics,33, 258–270.
3. Nagaraja, R., MacMillan, S., Kere, J., Jones, C., Griffin, S., Schmatz, M.,Terrell, J., Shomaker, M., Jermak, C., Hott, C. et al. (1997) X chromosomemap at 75-kb STS resolution, revealing extremes of recombination and GCcontent. Genome Res., 7, 210–222.
4. Mahtani, M.M. and Willard, H.F. (1998) Physical and genetic mapping ofthe human X chromosome centromere: repression of recombination.Genome Res., 8, 100–110.
5. Puechberty, J., Laurent, A.-M., Gimenez, S., Billault, A., Brun-Laurent, M.-E., Calenda, A., Marcais, B., Prades, C., Ioannou, P., Yurov, Y.et al. (1999) Genetic and physical analyses of the pericentromeric regionsof human chromosomes 5 and 19. Recombination across 5cen. Genomics,56, 274–287.
6. Mohrenweiser, H.W., Tsujimoto, S., Gordon, L. and Olsen, A.S. (1998)Regions of sex-specific hypo- and hyper-recombination identified throughintegration of 180 genetic markers into the metric physical map of humanchromosome 19. Genomics, 47, 153–162.
2238 Human Molecular Genetics, 2003, Vol. 12, No. 17

7. Horvath, J.E., Bailey, J.A., Locke, D.P. and Eichler, E.E. (2001)Lessons from the human genome: transitions between euchromatin andheterochromatin. Hum. Mol. Genet., 10, 2215–2223.
8. Bailey, J.A., Yavor, A.M., Massa, H.F., Trask, B.J. and Eichler, E.E. (2001)Segmental duplications: organization and impact within the current humangenome project assembly. Genome Res., 11, 1005–1017.
9. Bailey, J.A., Gu, Z., Clark, R.A., Reinert, K., Samonte, R.V., Schwartz, S.,Adams, M.D., Myers, E.W., Li, P.W. and Eichler, E.E. (2002)Recent segmental duplications in the human genome. Science, 297,1003–1007.
10. Broman, K.W., Murray, J.C., Sheffield, V.C., White, R.L. and Weber, J.L.(1998) Comprehensive human genetic maps: individual and sex-specificvariation in recombination. Am. J. Hum. Genet., 63, 861–869.
11. Yu, A., Zhao, C., Fan, Y., Jang, W., Mungall, A.J., Deloukas, P., Olsen, A.,Doggett, N.A., Ghebranious, N., Broman, K.W. et al. (2001) Comparisonof human genetic and sequence-based physical maps. Nature, 409,951–953.
12. Kong, A., Gudbjartsson, D.F., Sainz, J., Jonsdottir, G.M., Gudjonsson, S.A.,Richardsson, B., Sigurdardottir, S., Barnard, J., Hallbeck, B., Masson, G.et al. (2002) A high-resolution recombination map of the human genome.Nat. Genet., 31, 241–247.
13. Lynn, A., Kashuk, C., Petersen, M.B., Bailey, J.A., Cox, D.R.,Antonarakis, S.E. and Chakravarti, A. (2000) Patterns of meioticrecombination on the long arm of human chromosome 21. Genome Res.,10, 1319–1332.
14. van Hul, W., van Camp, G., Stuyver, L., Delabar, J.M., McInnis, M.G.,Warren, A.C., Antonarakis, S.E. and van Broeckhoven, C. (1993) Acontiguous physical map of the pericentromeric region of chromosome 21qbetween D21Z1 and D21S13E. Genomics, 15, 626–630.
15. Hassold, T., Sherman, S. and Hunt, P. (2000) Counting cross-overs:characterizing meiotic recombination in mammals. Hum. Mol. Genet., 9,2409–2419.
16. Hassold, T. and Hunt, P. (2001) To err (meiotically) is human: the genesisof human aneuploidy. Nat. Rev. Genet., 2, 280–291.
17. Lamb, N.E., Feingold, E., Savage, A., Avramopoulos, D., Freeman, S.,Gu, Y., Hallberg, A., Hersey, J., Karadima, G., Pettay, D. et al. (1997)Characterization of susceptible chiasma configurations that increase therisk for maternal nondisjunction of chromosome 21. Hum. Mol. Genet.,6, 1391–1399.
18. Lamb, N.E., Freeman, S.B., Savageaustin, A., Pettay, D., Taft, L.,Hersey, J., Gu, Y.C., Shen, J., Saker, D., May, K.M. et al. (1996)Susceptible chiasmate configurations of chromosome 21 predispose tonondisjunction in both maternal meiosis-I and meiosis-II. Nat. Genet.,14, 400–405.
19. Antonarakis, S.E., Petersen, M.B., McInnis, M.G., Adelsberger, P.A.,Schinzel, A.A., Binkert, F., Pangalos, C., Raoul, O., Slaugenhaupt, S.A.,Hafez, M. et al. (1992) The meiotic stage of nondisjunction in trisomy 21:determination by using DNA polymorphisms. Am. J. Hum. Genet., 50,544–550.
20. Shen, J.J., Sherman, S.L. and Hassold, T.J. (1998) Centromeric genotypingand direct analysis of nondisjunction in humans: Down syndrome.Chromosoma, 107, 166–172.
21. Wevrick, R. and Willard, H.F. (1989) Long-range organization of tandemarrays of alpha satellite DNA at the centromeres of human chromosomes:high-frequency array-length polymorphism and meiotic stability. Proc. NatlAcad. Sci. USA, 86, 9394–9398.
22. Mahtani, M.M. and Willard, H.F. (1990) Pulse-field gel analysis of alpha-satellite DNA at the human X chromosome centromere: high-frequencypolymorphisms and array size estimate. Genomics, 7, 607–613.
23. Marcais, B., Bellis, M., Gerard, A., Pages, M., Boublik, Y. and Roizes, G.(1991) Structural organization and polymorphism of the alpha-satelliteDNA sequences of chromosomes 13 and 21 as revealed by pulse field gelelectrophoresis. Hum. Genet., 86, 311–316.
24. Jabs, E.W., Warren, A.C., Taylor, E.W., Colyer, C.R., Meyers, D.A. andAntonarakis, S.E. (1991) Alphoid DNA polymorphisms for chromosome21 can be distinguished from those of chromosome 13 using probeshomologous to both. Genomics, 9, 141–146.
25. Lo, A.W., Liao, G.C., Rocchi, M. and Choo, K.H. (1999) Extremereduction of chromosome-specific alpha-satellite array is unusuallycommon in human chromosome 21. Genome Res., 9, 895–908.
26. Maratou, K., Siddique, Y., Kessling, A.M. and Davies, G.E. (2000)Variation in alphoid DNA size and trisomy 21: a possible cause ofnondisjunction. Hum. Genet., 106, 525–530.
27. Baker, W.K. (1958) Crossing over in heterochromatin. Am. Nat., 92, 59–60.28. Yamamoto, M. and Miklos, G.L.G. (1978) Genetic studies on hetero-
chromatin in Drosophila melanogaster and their implications for thefunctions of satellite DNA. Chromosoma, 66, 71–98.
29. Hattori, M., Fujiyama, A., Taylor, T.D., Watanabe, H., Yada, T., Park, H.S.,Toyoda, A., Ishii, K., Totoki, Y., Choi, D.K. et al. (2000) The DNAsequence of human chromosome 21. Nature, 405, 311–319.
30. Warren, A.C., Petersen, M.B., van Hul, W., McInnis, M.G.,van Broeckhoven, C., Cox, T.K., Chakravarti, A. and Antonarakis, S.E.(1992) D21S215 is a (GT)n polymorphic marker close to centromericalphoid sequences on chromosome 21. Genomics, 13, 1365–1367.
31. Bosch, A., Nunes, V., Patterson, D. and Estivill, X. (1993) Isolation andcharacterization of 14 CA-repeat microsatellites from human chromosome21. Genomics, 18, 151–155.
32. Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W.and Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: a newgeneration of protein database search programs. Nucl. Acids Res., 25,3389–3402.
33. Newton, C.R., Graham, A., Heptinstall, L.E., Powell, S.J., Summers, C.,Kalsheker, N., Smith, J.C. and Markham, A.F. (1989) Analysis of any pointmutation in DNA. The amplification refractory mutation system (ARMS).Nucl. Acids Res., 17, 2503–2516.
34. Horvath, J.E., Schwartz, S. and Eichler, E.E. (2000) The mosaic structure ofhuman pericentromeric DNA: a strategy for characterizing complex regionsof the human genome. Genome Res., 10, 839–852.
35. Abruzzo, M.A., Griffin, D.K., Millie, E.A., Sheean, L.A. and Hassold, T.J.(1996) The effect of y-chromosome alpha-satellite array length on therate of sex-chromosome disomy in human sperm. Hum. Genet., 97,819–823.
36. Lorber, B.J., Grantham, M., Peters, J., Willard, H.F. and Hassold, T.J. (1992)Nondisjunction of chromosome 21: comparisons of cytogenetic andmolecular studies of the meiotic stage and parent of origin. Am. J. Hum.Genet., 51, 1265–1276.
37. Petersen, M.B., Frantzen, M., Antonarakis, S.E., Warren, A.C.,van Broeckhoven, C., Chakravarti, A., Cox, T.K., Lund, C., Olsen, B.,Poulsen, H. et al. (1992) Comparative study of microsatellite andcytogenetic markers for detecting the origin of the nondisjoinedchromosome 21 in Down syndrome. Am. J. Hum. Genet., 51, 516–525.
38. Berend, S.A., Page, S.L., Atkinson, W., McCaskill, C., Lamb, N.E.,Sherman, S.L. and Shaffer, L.G. (2003) Obligate short-arm exchange inde novo Robertsonian translocation formation influences placement ofcrossovers in chromosome 21 nondisjunction. Am. J. Hum. Genet., 72,488–495.
39. Patil, N., Berno, A.J., Hinds, D.A., Barrett, W.A., Doshi, J.M.,Hacker, C.R., Kautzer, C.R., Lee, D.H., Marjoribanks, C.,McDonough, D.P. et al. (2001) Blocks of limited haplotype diversityrevealed by high-resolution scanning of human chromosome 21.Science, 294, 1719–1723.
40. Sachidanandam, R., Weissman, D., Schmidt, S.C., Kakol, J.M.,Stein, L.D., Marth, G., Sherry, S., Mullikin, J.C., Mortimore, B.J.,Willey, D.L. et al. (2001) A map of human genome sequence variationcontaining 1.42 million single nucleotide polymorphisms. Nature, 409,928–933.
41. Trowell, H.E., Nagy, A., Vissel, B. and Choo, K.H.A. (1993) Long-rangeanalyses of the centromeric regions of human chromosomes 13, 14 and 21:identification of a narrow domain containing two key centromeric DNAelements. Hum. Mol. Genet., 2, 1639–1649.
42. Benson, G. (1999) Tandem repeats finder: a program to analyze DNAsequences. Nucl. Acids Res., 27, 573–580.
43. Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994) CLUSTAL W:improving the sensitivity of progressive multiple sequence alignmentthrough sequence weighting, position-specific gap penalties and weightmatrix choice. Nucl. Acids Res., 22, 4673–4680.
44. Jeffreys, A.J., Neumann, R. and Wilson, V. (1990) Repeat unit sequencevariation in minisatellites: a novel source of DNA polymorphism forstudying variation and mutation by single molecule analysis. Cell, 60,473–485.
Human Molecular Genetics, 2003, Vol. 12, No. 17 2239














