
How are the dynamics of woodland dominants influenced by ...
135
How are the dynamics of woodland dominants influenced by climate and disturbances in south eastern Australia? Janet S. Cohn Bachelor of Natural Resources (Honours) Thesis submitted to Charles Sturt University for the degree of Doctor of Philosophy August 2010 Faculty of Science School of Environmental Sciences
Transcript of How are the dynamics of woodland dominants influenced by ...

How are the dynamics of woodland dominants
influenced by climate and disturbances in south
eastern Australia?
Janet S. Cohn
Bachelor of Natural Resources (Honours)
Thesis submitted to Charles Sturt University for the degree of Doctor of
Philosophy
August 2010
Faculty of Science
School of Environmental Sciences

i
Table of Contents
Acknowledgements iii
Substantial Contributions to the thesis iv
Publications v
Abstract vi
Chapter 1. General introduction and thesis overview 1
Chapter 2. Demographic patterns of Callitris glaucophylla are associated
with rainfall and disturbances along rainfall gradients 21
Abstract and Introduction 22
Methods 25
Results 29
Discussion 37
Chapter 3. Competitive feedback between Eucalyptus species and
Callitris glaucophylla trees and C. glaucophylla regeneration influences
stand dynamics along a rainfall gradient 43
Abstract and Introduction 44
Methods 47
Results 52
Discussion 62
Chapter 4. Feedback between patches of Callitris glaucophylla and fire
intensity influences Callitris survival in flammable Eucalyptus woodlands
in different fire-weather conditions 67
Abstract and Introduction 68
Methods 71
Results 76
Discussion 85
Chapter 5. Synthesis 89
Chapters 2-4 90
The dynamics of woodland dominants in geographic zones 96
Predicted dynamics of woodland dominants in geographic zones under
climate change 98
Further work 100
References 102

ii
Certificate of Authorship

iii
Acknowledgements
Firstly, I would like to thank my supervisors Ian Lunt, Ross Bradstock and Karen Ross.
They were all of immense help during the production of this thesis. Their different
backgrounds and interests complimented one another very well. I am particularly
grateful to Ian, who was always generous with his time, enthusiasm and extraordinary
sense of logic. We had many creative and exciting discussions which helped to shape
the research and I look forward to many more.
Jack Baker and Klaus Koop agreed to my leave from DECCW NSW to pursue this
research. John Whittall, also from DECCW NSW smoothed the way for the
flammability work. Thanks to Stephen Campbell and Alan Smith from State Forests
NSW for their assistance and permission to take tree samples. Statisticians Simon
McDonald (CSU) and Terry Koen (DECCW NSW) were wonderfully generous with
their time and patience.
Finally, I would like to thank my dear family and friends. Andrew, Katrina and Bernie
Cohn, Grace Bradstock, Livia Tigwell, John Warren and Kate Hammill helped me in
the field and in many other ways, for which I am grateful.

iv
Substantial Contributions to the thesis
The research was funded by the Australian Research Council (ARC) Linkage Grant
between Charles Sturt University (CSU) and NSW Department of Environment and
Climate Change (NSW DECC; Project number LP066829) and Australian Institute for
Nuclear Science and Energy (AINSE; AINGRA09075).
Discussions with supervisors Ian Lunt, Ross Bradstock and Karen Ross variously
assisted with the formulation of objectives, methods and data interpretation. They also
edited drafts of the thesis. I carried out the fieldwork with the help of assistants and I
wrote the thesis. Statisticians Simon McDonald and Terry Koen helped to advise me on
the appropriate analyses for difficult datasets and they assisted me in carrying out some
analyses.

v
Publications
Cohn, J. S., Lunt I.D., Ross K. A. & Bradstock R. A. (in review) How do slow-growing,
fire-sensitive conifers survive in flammable eucalypt woodlands? Journal of Vegetation
Science.
The relative contributions of the authors to this manuscript are similar to those outlined
in Substantial Contributions to the Thesis (page iv).

vi
Abstract
Woodlands in south eastern Australia are dominated by trees of contrasting functional
types. Callitris glaucophylla is a slow growing, obligate-seeding conifer and Eucalyptus
species are fast growing, resprouting angiosperms. The success of these functional types
is likely to vary with climate and different levels and rates of disturbances along
climatic gradients. I examined how climate and disturbances influenced the dynamics of
these woodland dominants, by using surveys of varying scales.
Regeneration failure of Callitris has led to ageing populations (mid - late 1800s) below
405 mm annual rainfall in the winter rainfall zone. Failure was associated with farms,
which graze livestock and where rabbits are common. In contrast, Callitris populations
are younger (since 1950s) and expanding on tenures other than farms (roadside,
travelling stock routes, State Forests), where livestock grazing levels are low-moderate,
and rabbits and fires are less frequent above 405 mm mean annual rainfall.
Competition from canopy trees had little effect on the success of Callitris regeneration.
Although regeneration under canopy trees had lower densities, were smaller and less
likely to be reproducing than those in gaps, they have persisted 50 years after their
establishment. In contrast, competition from regeneration had a significant effect on
canopy tree resilience during drought. Callitris canopy trees surrounded by regeneration
had higher mortality and Eucalyptus canopy trees had increased stress levels at lower
rainfall.
In large woodland remnants, where unplanned fires occur (e.g. National Parks), dense
Callitris regeneration can reduce incoming fire intensity from the flammable Eucalyptus
matrix in low-moderate fire weather conditions. This increases the survival probability
of larger Callitris, allowing coexistence with Eucalyptus.
These results highlight how the dynamics of woodland dominants are associated with
disturbance regimes and climate along climatic gradients. Disturbance regimes were
associated with rainfall through different land use types. As climate changes, rainfall,
land use and disturbance regimes will shift along climatic gradients, influencing the
distributions of these woodland dominants.

vii

Chapter 1. General introduction and thesis overview
1
Chapter 1
General introduction and thesis overview
Callitris glaucophylla and Eucalyptus species woodland, Merrimerriwa Range, central western New South Wales (Source: K. Ross)

Chapter 1. General introduction and thesis overview
2
Chapter 1
General introduction and thesis overview
Introduction
In recent centuries woodlands worldwide have undergone dramatic changes (Belsky &
Blumenthal 1997; Wright et al. 2004; Noss et al. 2006) and those in south eastern
Australia are no exception (Clarke 2000; Lunt et al. 2006). Their occurrence in
agricultural zones has led to landscape-scale changes in disturbance regimes (Sivertsen
& Clarke 2000). Introduced grazers, tree removal, and changes in fire regimes and soils
have resulted in a mosaic of woodland compositional and structural states (Austin &
Williams 1988; Westoby et al. 1989; Clarke 2000; Lunt et al. 2006). Interactions among
these processes are often complex and poorly understood (Clarke 2000; Burrows 2002;
Graz 2008).
Woodlands are widespread and extend from the temperate to the arid zone in south
eastern Australia (Fig. 1.1; Specht & Specht 1999; Keith 2004). Within this climatic
range, different processes are likely to control the dynamics of woody plants. Climate
can directly affect species distributions through physiological limitations, imposed by
rainfall and temperature, or indirectly by influencing disturbance regimes (Gworek et al.
2007). For example, in a study along an elevational gradient, the dynamics of Pinus
jeffreyi were variously associated with interactions between rainfall (drought),
parasitism by mistletoe and seed predation by insects (Gworek et al. 2007). Disturbance
regimes may also be influenced by predominant human land uses that vary with climate.
For example, as rainfall increases from the arid to the temperate zone, a shift from
grazing to a range of land uses becomes possible at higher rainfall (e.g. forestry and
cropping; Dale 1997; Bureau of Rural Sciences 2009). Changes in regional land use can
further alter disturbances (e.g. fires and grazing) within patches of native vegetation.
Trees of contrasting functional types dominate woodlands in south eastern Australia.
Callitris glaucophylla, a slow growing, fire-sensitive obligate-seeder often coexists with
a range of Eucalyptus species, which are fast growing, fire-tolerant resprouters (Lacey
1972, 1973; Clayton-Greene 1981; FCNSW 1988; Florence 1996; Specht & Specht
1999). Differences in growth rates and methods of regeneration between these
evergreen trees are likely to result in different responses to climatic conditions and

Chapter 1. General introduction and thesis overview
3
disturbances, leading to variations in age-structure and abundances across their ranges.
At extremes, Callitris can occur as dense regeneration or as widely spaced mature trees
(Lacey 1972, 1973; Noble 1997). The regeneration success of Eucalyptus is also
spatially variable (Reid & Landsberg 2000). Despite their prominence, there are gaps in
our understanding of how the comparative dynamics of Callitris and Eucalyptus vary in
relation to disturbances and climate across their ranges (Clayton-Greene 1981; Bowman
et al. 1988; Bowman & Wilson 1988; Clayton-Greene & Ashton 1990).
Figure 1.1 The distribution of woodland types in Australia. The study area is circled in
blue (Source: Lindenmayer 2005).
Interactions among functional types, productivity and rainfall are hypothesised as
determinants of a species’ success (Bellingham & Sparrow 2000; Huston 2003). Few
studies have examined these interactions using woody plants (Loehle 2000; Bond et al.
2001a; Zavala & Zea 2004; Sankaran et al. 2005; Pausas & Bradstock 2007). Of those
that have, most have concentrated on relationships between shrubs or between grasses
and trees (Scholes & Archer 1997; Bond et al. 2001; Sankaran et al. 2005), rather than
between tree species (Loehle 2000; Zavala & Zea 2004). A useful first step in predicting
patterns of Callitris and Eucalyptus success in inland south eastern Australia is to use
theoretical models to examine the relative success of plant functional types along
productivity and disturbance gradients (Bellingham & Sparrow 2000; Huston 2003).

Chapter 1. General introduction and thesis overview
4
The aim of this thesis is to examine how the dynamics of woodland dominants of
contrasting functional types, Callitris glaucophylla and Eucalyptus species, are
influenced by climate and disturbance regimes in south eastern Australia.
In this first chapter, I give an account of the current knowledge of the dynamics of
woodland dominants, Callitris glaucophylla and Eucalyptus species. The chapter begins
with the climatic setting of study region, a description of disturbance regimes,
physiological and life history characteristics of functional types (Callitris and
Eucalyptus) and our knowledge of how functional types respond to varying rainfall and
disturbances. Then I predict functional type success (Callitris and Eucalyptus) along
productivity and disturbance gradients, by examining a range of theoretical models.
Finally, I give an outline of the thesis.
Callitris and Eucalyptus woodlands
Climate and habitat
The study area lies within woodlands in south eastern Australia (Fig. 1.1). It extends
from the temperate zone in the east to the arid zone in the west (Specht & Specht 1999),
a distance of approximately 600 kms. Mean annual rainfall ranges from 600 to 200 mm.
From north to south, there is a change in seasonal rainfall from summer to winter
dominance (Colls & Whittaker 2001; BOM 2010a). Under climate change, much of the
study area is predicted to have increases in drought severity and frequency and a switch
to summer dominant rainfall (Nicholls 2004; CSIRO & BOM 2007; Hennessey et al.
2008; DECCW NSW 2009). Only in the north east is the water balance expected to
remain relatively unchanged (DECCW NSW 2009).
Woodlands decline in height and become more open from the temperate to the arid zone
(Specht & Specht 1999). They are dominated by Callitris glaucophylla and range of
Eucalyptus species (Fig. 1.2). Whilst both genera can coexist at higher rainfall (Lindsay
1967; Keith 2004), in the drier parts of their range, Eucalyptus become confined to
intermittent water courses or rock outcrops (Brooker & Kleinig 1990). Callitris
glaucophylla most commonly occurs on sandy to loamy textured soils, whilst
Eucalyptus species can occur on a wider range of soils (Lacey 1973; Wardell-Johnson et
al. 1997).

Chapter 1. General introduction and thesis overview
5
Figure 1.2 A mixed stand of Callitris glaucophylla and Eucalyptus species (left
foreground) in central New South Wales. Callitris which established in the 1950s is on
the right beside a 2m tall standard and on the left is a Callitris which established in the
late 1800s (Source: J. Cohn).
Disturbance regimes
Woodlands in south eastern Australia have undergone major changes in disturbance
regimes since European management in the 1800s (Austin & Williams 1988; Westoby
et al. 1989; Clarke 2000; Lunt et al. 2006). Landscape-scale manipulation of tree
densities, changes in fire regimes and introduced herbivores have led to a mosaic of
disturbance regimes and resultant stand types (Noble 1997; Lunney 2001; Lunt &
Spooner 2005).
Extensive areas of woodlands have been cleared for grazing, cropping and building
materials (Sivertsen & Clarke 2000; Cox et al. 2001). Selective ringbarking of
Eucalyptus to improve growth rates of the commercially valuable Callitris (Allen 1995),
has resulted in a switch from Eucalyptus to Callitris dominance in State Forests (Lunt et
al. 2006).

Chapter 1. General introduction and thesis overview
6
Little information is available on past and present fire regimes across the study area.
However, it is generally agreed that fire extent and frequencies have declined since
European management, largely as a result of habitat fragmentation, a reduction in
biomass caused by livestock grazing, as well as active fire suppression and exclusion
(Wilson 1990; Pulsford et al. 1992; Noble 1997, 2001). Unplanned fires still occur
regularly in large woodland remnants, such as conservation reserves (Brookhouse
1999).
Sheep and cattle grazing were introduced during the 1800s (Noble 1997). As a result,
farms have historically had higher grazing levels than more fragmented tenures like
roadside verges, travelling stock routes, State Forests and National Parks (Lunt 1995;
Fensham 1998a). Rabbits were introduced to the region in the late 1800s and spread
rapidly reaching ‘plague proportions’ over much of south eastern Australia until the
1950s (Austin & Williams 1988; Myers et al. 1994), when their numbers declined
dramatically after the release of myxomatosis, a mosquito borne virus (Noble 1997).
Since then, rabbit populations have been maintained at lower levels (Norris et al. 1991;
Tiver & Andrew 1997) with the help of rabbit haemorrhagic disease virus since the
1990s (Scanlan et al. 2006).
Callitris and Eucalyptus functional types and responses to disturbances
Physiological and life history attributes
Callitris glaucophylla and Eucalyptus species are long-lived evergreen trees, capable of
reaching a maximum height of approximately 30m (Cunningham et al. 1992; Fig. 1.2).
The two genera represent contrasting functional types (Table 1.1). Callitris
glaucophylla is a slow-growing, shade tolerant, obligate-seeding conifer, whilst
Eucalyptus species are faster-growing, shade-intolerant, resprouting angiosperms
(Lacey 1972, 1973; Clayton-Greene 1981; FCNSW 1988; Florence 1996). These life-
history differences are likely to influence their respective responses to productivity and
disturbance regimes (Bellingham & Sparrow 2000).
A major difference between Callitris and Eucalyptus is in their responses to fire.
Following 100% canopy scorch, Callitris die, whilst coexisting Eucalyptus resprout
(Gill & Bradstock 1992). Following death, persistence of Callitris is dependent on
seeds. Given these responses, Callitris is termed an obligate-seeder and Eucalyptus a
resprouter (Gill & Bradstock 1992).
Chapter 1. General introduction and thesis overview
7
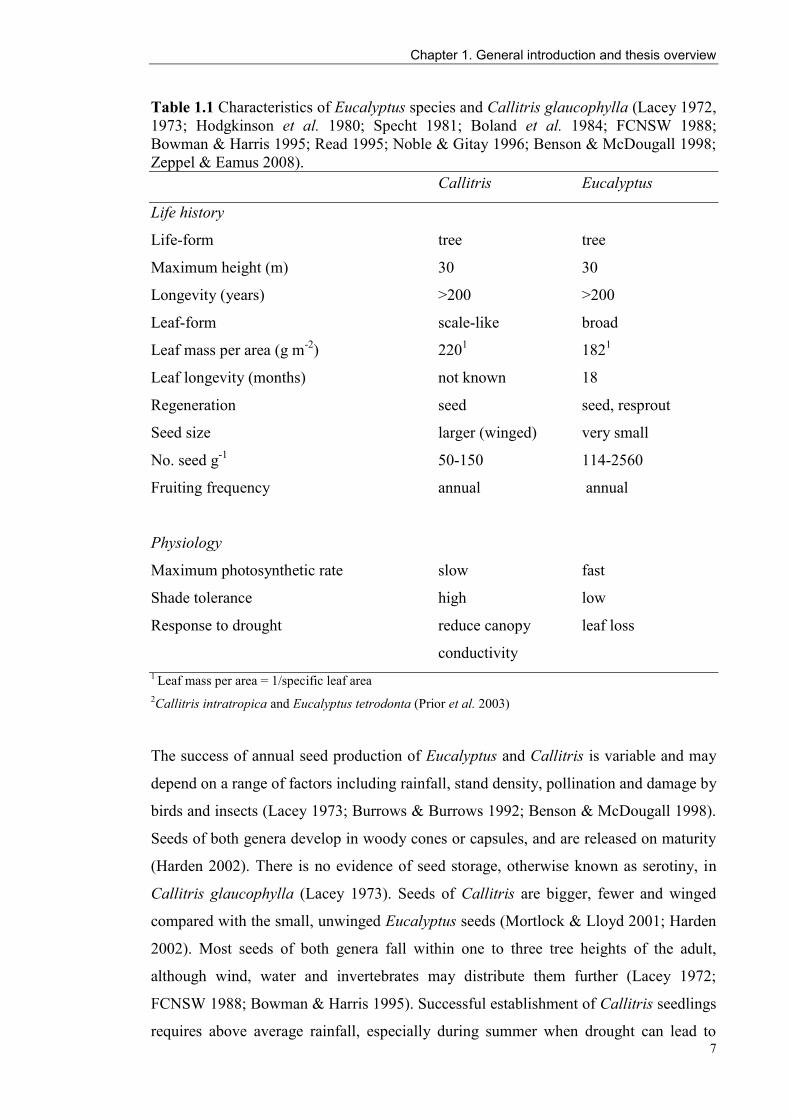
Table 1.1 Characteristics of Eucalyptus species and Callitris glaucophylla (Lacey 1972,
1973; Hodgkinson et al. 1980; Specht 1981; Boland et al. 1984; FCNSW 1988;
Bowman & Harris 1995; Read 1995; Noble & Gitay 1996; Benson & McDougall 1998;
Zeppel & Eamus 2008).
Callitris Eucalyptus
Life history
Life-form tree tree
Maximum height (m) 30 30
Longevity (years) >200 >200
Leaf-form scale-like broad
Leaf mass per area (g m-2
) 2201 182
1
Leaf longevity (months) not known 18
Regeneration seed seed, resprout
Seed size larger (winged) very small
No. seed g-1
50-150 114-2560
Fruiting frequency annual annual
Physiology
Maximum photosynthetic rate slow fast
Shade tolerance high low
Response to drought reduce canopy leaf loss
conductivity
1 Leaf mass per area = 1/specific leaf area
2Callitris intratropica and Eucalyptus tetrodonta (Prior et al. 2003)
The success of annual seed production of Eucalyptus and Callitris is variable and may
depend on a range of factors including rainfall, stand density, pollination and damage by
birds and insects (Lacey 1973; Burrows & Burrows 1992; Benson & McDougall 1998).
Seeds of both genera develop in woody cones or capsules, and are released on maturity
(Harden 2002). There is no evidence of seed storage, otherwise known as serotiny, in
Callitris glaucophylla (Lacey 1973). Seeds of Callitris are bigger, fewer and winged
compared with the small, unwinged Eucalyptus seeds (Mortlock & Lloyd 2001; Harden
2002). Most seeds of both genera fall within one to three tree heights of the adult,
although wind, water and invertebrates may distribute them further (Lacey 1972;
FCNSW 1988; Bowman & Harris 1995). Successful establishment of Callitris seedlings
requires above average rainfall, especially during summer when drought can lead to

Chapter 1. General introduction and thesis overview
8
desiccation (FCNSW 1988). Eucalyptus regeneration occurs more commonly following
fires and above average rainfall (FCNSW 1988; Bell & Williams 1997; Gill 1997). Fire
contributes to Eucalyptus establishment by stimulating mass seed release, reducing
competition and favourably altering the microenvironment (Bell & Williams 1997).
Callitris grows slowly compared with coexisting Eucalyptus, especially in the earlier
stages of development (Clayton-Greene 1981). Self-thinning of dense Callitris saplings
occurs at slow rates, as they reduce growth rates to compensate for intraspecific
competition (Lacey 1972, 1973; FCNSW 1988). In comparison Eucalyptus self-thin at
faster rates (Clayton-Greene 1981; FCNSW 1988). Slower growth rates of Callitris
increase their time to reproductive maturity, thereby increasing the period of
vulnerability to biotic and abiotic factors (Bond 1989). Without intense competition
individual Callitris may reach reproductive maturity in approximately six years (Lacey
1972). Following germination Eucalyptus quickly develop a lignotuber, from which
they can resprout (Bell & Williams 1997). Callitris and Eucalyptus are similarly long-
lived, reaching ages of at least 200 years (Bowman & Harris 1995; Williams & Brooker
1997).
Callitris have small scale-like leaves, which form a denser canopy than the pendulous,
broader leaves of Eucalyptus. Both leaf types are considered sclerophyllous (Berry &
Roderick 2002), which is interpreted as an adaptation to seasonal drought in
Mediterranean type systems (Cunningham et al. 1999; Ninemets 2001; Wright et al.
2001; Lamont et al. 2002). Studies during droughts have found that Callitris and
Eucalyptus similarly reduce water usage, but by different means. Callitris reduce
canopy conductance or gas exchange and Eucalyptus shed leaves (Attiwill & Clayton-
Greene 1984; Zeppel & Eamus 2008). The recovery of Eucalyptus is dependent on their
ability to resprout (Pook et al. 1966; Attiwill & Clayton-Greene 1984), which may be
tested during periods of prolonged drought.
Differences in leaf size influence the flammability of leaf litter through differential
spatial packing (Schwilk & Ackerly 2001; Scarff & Westoby 2006). Larger leaves like
those of Eucalyptus provide better ventilated ground fuels which increases fire intensity
compared with the smaller leaves of Callitris which pack more tightly (Scarff &
Westoby 2006). The formation of tightly packed litter advantages slow growing,

Chapter 1. General introduction and thesis overview
9
obligate-seeders like Callitris, by increasing their chances of surviving fire and
increasing their probability of reaching reproductive maturity (Scarff & Westoby 2006).
Rainfall
The success of Callitris regeneration is strongly influenced by rainfall. Widespread
establishment of Callitris was observed in the late 1800s and the 1950s following above
average rainfall during La Nina events (Austin & Williams 1988; Noble 1997).
Regeneration during these periods contributed to the formation of dense thickets of
Callitris regeneration, giving it ‘woody weed’ status in parts of the semi-arid zone
(Noble 1997; Fig. 1.3). Conversely, during periods of drought, desiccation of Callitris
seedlings has contributed to Callitris mortality in arid areas (Johnson 1969; Read 1995).
Regeneration of Eucalyptus similarly coincides with above average rainfall (Fensham et
al. 2009).
Survival of Callitris and Eucalyptus trees is negatively affected by drought (Lacey
1972, 1973; Allen 1998; Fensham 1998b; Fensham & Holman 1999; Fensham et al.
2009). During drought higher mortality of Eucalyptus and Callitris trees has been
observed or recorded at higher tree densities (Lacey 1972, 1973; Fensham & Holman
1999). There is anecdotal information that Eucalyptus and Callitris trees surrounded by
dense Callitris sapling regeneration are selectively killed (Lacey 1972, 1973; Allen
1998). Foresters have also observed higher mortality of Callitris trees in lower rainfall
regions (Lacey 1972).
Fire regimes
Given its sensitivity to fire, fire regimes are likely to have a major influence on the
population dynamics of Callitris. A regime of frequent high intensity fire is believed
responsible for contracting populations of Callitris intratropica in northern Australia
(Bowman et al. 2001). Callitris intratropica is a slow-growing obligate-seeder like
Callitris glaucophylla (Bowman & Harris 1995). Patchy burning has been observed in
monospecific stands of Callitris compared with continuous burning in adjacent
Eucalyptus stands (Clayton-Greene 1981; Bradstock & Cohn 2002). Minimal damage to
dense patches of Callitris regeneration has been observed in ‘severe fires’ (Lacey 1972).
Low biomass of grasses, shrubs and lower leaf litter flammability are likely to
contribute to lower flammability associated with dense stands of Callitris (Clayton-
Greene 1981; Bowman & Wilson 1988; Scarff & Westoby 2006).

Chapter 1. General introduction and thesis overview
10
Figure 1.3 Dense regeneration of Callitris glaucophylla in central New South Wales.
Callitris saplings that established in the 1950s surround an adult Callitris that
established in the late 1800s. The standard is 4m tall (Source: J. Cohn).
Interactions between rainfall, grazing and fire regimes are hypothesised as determinants
of woody species encroachment in semi-arid woodlands (Westoby et al. 1989; Walker
1993; Noble 1997; Burrows 2002). Regular fires following above average rainfall and
herbaceous fuel production are believed to have reduced opportunities for woody plant
establishment prior to European management. However, under high levels of introduced
grazers, reductions of herbaceous fuels have contributed to declines in fire frequency,
allowing the establishment of woody species, like Callitris (Noble 1997).
Grazing regimes
Grazing by sheep and cattle has reduced or prevented the regeneration success of both
Callitris (FCNSW 1988; Lacey 1973; Allcock & Hik 2004) and Eucalyptus (Parker &
Lunt 2000; Allcock & Hik 2004). Widespread grazing by large numbers of rabbits
prevented the regeneration of Callitris from 1890 to 1950 (Austin & Williams 1988;
Noble 1997), and is still affecting regeneration in some locations (Allcock & Hik 2004).

Chapter 1. General introduction and thesis overview
11
Summary
Whilst this literature review provides an indication of how rainfall, and disturbance
regimes (logging, fire, grazing) influence Callitris and Eucalyptus dynamics, it also
highlights the relative incompleteness of this knowledge. It reveals that: above average
rainfall stimulates establishment and drought results in mortality of Callitris and
Eucalyptus; high frequencies and intensities of fires do not favour the obligate-seeding
Callitris; regeneration of Callitris and Eucalyptus are negatively affected by grazing.
However, we know little about Callitris survival during low-moderate intensity fires
and how this may vary with site productivity. We do not know how different levels of
grazing may affect Callitris and Eucalyptus success and how this may vary with site
productivity. We also know little about the comparative effects of drought on Callitris
and Eucalyptus, which is especially important as droughts increase in frequency and
severity under global warming (Nicholls 2004; CSIRO & BOM 2007; Hennessey et al.
2008; DECCW NSW 2009). By examining theoretical models of functional type
responses, we can predict the relative success of Callitris and Eucalyptus at varying
levels of productivity and disturbances.
Models of functional type success along productivity and disturbance
gradients
Gymnosperms dominated global vegetation up until the late Cretaceous (Krassilov
1978). The fossil record in Australia provides evidence of a dramatic reduction in
conifer diversity and distribution during the Tertiary (Wells & Hill 1989; Hill 1995).
Possible explanations include competition with angiosperms, which were spreading and
diversifying at that time (Bond 1989), increasing aridity (Brodribb & Hill 1998) and an
increase in disturbance frequency (Hill & Brodribb 1999).
Competition, drought and disturbance regimes are worthy of exploration in explaining
contemporary dominance shifts in Eucalyptus species and C. glaucophylla across their
ranges. By examining a range of theories that use physiological and life history traits to
predict dominance along gradients of productivity and disturbance (Specht 1981;
Keeley 1986; Bond 1989; Westoby 1999; Bellingham & Sparrow 2000; Huston 2003), I
aim to generate hypotheses on the likely mechanisms for the relative success of
Eucalyptus and Callitris in south eastern Australia. Models are ordered from those
which predict patterns along (a) productivity gradients, (b) disturbance gradients, and
(c) productivity and disturbance combinations.

Chapter 1. General introduction and thesis overview
12
Physiological traits and productivity
It is widely accepted that competition for resources, especially light determines
dominance at higher productivity sites (Gurevitch et al. 2006). However, there is debate
on whether competition for water and nutrients (Tilman 1988) is more important than
stress tolerance (Grime 1979) in determining dominance at lower productivity sites. In
the ‘slow seedling’ hypothesis, Bond (1989) proposed that the competitive ability of
conifers in the regeneration phase is limited by physiological characteristics. Because of
intense competition from faster growing angiosperms on higher productivity sites,
conifers are left more vulnerable to disturbances, restricting them to lower productivity
sites.
Leaf traits reflect resource uptake and use-efficiency strategies in plants, so are good
indicators of whole-plant function (Reich et al. 1999). Comparative studies suggest
significant differences in leaf traits between conifers and angiosperms (Ackerly & Reich
1999; Lusk et al. 2003; Prior et al. 2003). Conifers have smaller leaves with longer life
span, lower leaf specific area, lower photosynthetic capacity, stomatal conductance, and
higher area based nitrogen concentration. The implications of these traits are discussed.
The higher photosynthetic capacity per unit leaf mass of evergreen angiosperms is likely
to give angiosperms a competitive advantage over conifers in productive habitats (Lusk
& Matus 2000; Lusk et al. 2003). The difference in photosynthetic performance
between evergreen conifers and angiosperms is partly attributable to variation in
stomatal conductance, which is higher on an area basis in angiosperms (Lusk et al.
2003). Lower stomatal conductivity in conifers is consistent with the theory that
conifers may still be disadvantaged by stomatal traits evolved under higher atmospheric
levels of CO2 (Beerling & Woodward 1996).
Conifers may be advantaged at lower productivity sites by their lower photosynthetic
capacity placing fewer demands on resources (water, nutrients; Grime 1979). Wright et
al. (2001) argued that leaves with higher area based nitrogen concentration, as were
found on conifers (Ackerly & Reich 1999), make greater investments in photosynthetic
‘machinery’ allowing a given photosynthetic rate to be achieved at lower stomatal
conductivity. This encourages water-use efficiency (Schulze et al. 1998; Lamont et al.
2002; Llorens et al. 2004). The longer-lived leaves of conifers allows nutrients to be
accrued over a longer residence time (Roderick et al. 2000; Lusk et al. 2003; Wright et

Chapter 1. General introduction and thesis overview
13
al. 2004), which helps persistence in lower productivity sites. These characteristics
suggest that conifers may be best described as stress-tolerators according to Grime’s
(1979) Competitor-Stress-tolerator-Ruderal scheme. Keeley & Zedler (1998) believe
that this scheme is useful in distinguishing the qualities that separate Pinus species from
woody angiosperms in their ability to tolerate abiotically stressful environments.
Leaf specific area is the ratio of projected leaf area to leaf dry mass (Roderick et al.
2000). Many studies have shown that leaf specific area increases with soil water
availability along aridity gradients (e.g. Pierce et al. 1994; Schulze et al. 1998; Reich et
al. 1999). This adds further weight to the prediction that conifers and angiosperms may
dominate at the lower and higher ends of productivity gradients, respectively.
The gap availability hypothesis relates recruitment opportunities of seeders (non-
resprouters) to the availability of gaps along resource gradients (Specht 1981; Bond &
van Wilgen 1996). In humid environments, regeneration of resprouters predominates
because competition from dense stands of resprouters reduces the successful
establishment of seeders (Specht 1981). In contrast, seeder establishment is more
successful in drier environments, because lower densities of resprouters result in more
open vegetation. Thus the proportional representation of resprouters and seeders shifts
along gradients of water availability. In large-scale gradient studies comparable with
Specht’s (1981), resprouter success increased with increasing moisture availability
(Pausas & Bradstock 2007) and increasing summer rainfall dominance (Ojeda 1998).
However, this trend may be confounded by fire frequency, which Pausas & Bradstock
(2007) found to increase with increasing rainfall. Disentangling the effects of water
availability from fire regimes is important if we are to understand the drivers of the
relative success of resprouters and seeders (Pausas & Bradstock 2007).
At a local scale, seeders are often more abundant on drier sites than resprouters in a
range of Mediterranean ecosystems (Keeley 1986; Keith 1991; Pausas et al. 1999;
Meentemeyer et al. 2001; Clarke & Knox 2002). Since resprouters tend to be deeper
rooted than seeders (Keeley 1986; Bell 2001) and water potential for resprouters is
usually higher during summer (Davis et al. 1999; Ackerly 2004), the above ground
organs of seeders must have physiological, chemical and/or structural compensations
(Paula & Pausas 2006). Comparative studies on physiological tolerance of seeders and
resprouters to drought vary. Some studies have found that seeders are more drought
tolerant than resprouters because they have greater resistance to embolisms caused by

Chapter 1. General introduction and thesis overview
14
water stress and later stomatal closure than resprouters (Davis et al. 1999), or higher
potential for structural resistance to drought and higher water use efficiency (Ackerly
2003, 2004; Paula and Pausas 2006). In other studies resprouters have shown higher
carbon assimilation at low water potentials and later stomatal closure indicating greater
drought tolerance than seeders (Calamassi et al. 2001; Vilagrosa et al. 2003). Although
no pattern in the relative drought tolerance of seeders and resprouters has also been
identified (Smith et al. 1992), Knox & Clarke (2005) found that they respond differently
to the availability of water and nutrients. Seedlings of resprouters may be advantaged at
more fertile sites because of allocation to the formation of roots and starch reserves,
giving them a competitive advantage over seeders after fires (Knox & Clarke 2005).
The studies on leaf traits of conifers and angiosperms support the slow ‘seedling
hypothesis’ (Bond 1989). Faster growth rates may contribute to competitive advantage
of angiosperms over slower growing conifers at higher productivity sites. This restricts
conifers to lower productivity sites where slower growth rates and longer-lived leaves
contribute to their tolerance of stress. There is some support for the gap availability
hypothesis (Specht 1981), which predicts that seeders and resprouters will be more
successful at lower and higher productivity sites, respectively. Seeders may be more
drought tolerant than resprouters allowing better survival in lower productivity sites and
resprouters may have a competitive advantage over seeders in higher productivity sites.
Life history and disturbances
Bellingham & Sparrow (2000) predict that woody seeders can only survive where
disturbances are less frequent, mild and predictable enough for maturity to be reached
before they are killed. Conversely, woody resprouters should increase with increasing
frequency of disturbance (Bellingham & Sparrow 2000; Bond & Midgley 2001), and
decline at the highest frequencies where it is very difficult to grow and maintain storage
levels. Here, herbaceous seeders are more likely to be successful (Bellingham &
Sparrow 2000; Sparrow & Bellingham 2001). In fire-prone Mediterranean systems the
success of resprouters and seeders in relation to fire frequency is variable. In some
ecosystems non-respouters are more successful at low to intermediate frequencies of
fire (Keeley & Zedler 1978; Hilbert 1987; Burgman & Lamont 1992; Enright et al.
1998; Clarke 2002; Clarke & Knox 2002). Conversely, resprouters are more successful
at the lowest and higher frequencies, where seeders senesce and do not reach
reproductive maturity, respectively (Keeley & Zedler 1978; Kruger & Bigalke 1984;

Chapter 1. General introduction and thesis overview
15
Keeley 1986; Enright et al. 1998; Pausas 1999). In other ecosystems, such as Australian
heathlands, the relative success of seeders and resprouters with fire frequency can not be
generalised (Pausas et al. 2004). Traits such as time to maturity, shade tolerance,
serotiny and plant longevity may also need consideration when predicting species’
responses to fire regimes (Pausas 2001).
The fire frequency hypothesis assumes that space limits the regeneration of seeders
(Keeley & Zedler 1978; Specht 1981; Keeley 1986). As fire interval lengthens, more
gaps should occur as mortality rates of resprouters increase due to senescence and death
from higher intensity fires (Keeley & Zedler 1978; Keeley 1986). Consequently, there
will be more opportunities for seeders to establish, while sprouters would be more
common at shorter fire intervals. Whilst a number of field studies support this
hypothesis (Keeley & Zedler 1978; Kruger & Bigalke 1984; Traubaud 1991), Kruger
(1983) found that resprouters were prominent in fynbos shrublands, irrespective of fire
regime.
Westoby (1999) predicts that certain plant traits are more likely to be favoured under
different grazing regimes. Moderate intensity grazing is likely to select for species with
higher specific leaf area (SLA; e.g. Eucalyptus), which are considered more palatable,
because of lower structural strength than leaves with lower specific leaf area (e.g.
Callitris; Reich et al. 1997). Under this grazing intensity, species with lower specific
leaf area are likely to dominate. Under higher grazing intensity where selectivity is low,
species with higher specific leaf area may be more successful, as they can grow more
quickly, especially during early life (Reich et al. 1997; Ninemets 2001; Wright et al.
2001). Although this theory is sound, the comparatively small differences between
specific leaf areas of Callitris and Eucalyptus may not be sufficient to distinguish
different responses to grazing regimes.
Empirical studies indicate that the ability to predict grazing responses using plants traits
may vary with location (Vesk et al. 2004). In the productive grasslands of Argentina
and Israel, grazing resistance of herbaceous species was associated with a range of traits
like higher specific leaf area, short stature and smaller leaves (Diaz et al. 2001), but no
such associations were found in shrubs of the less productive arid and semi-arid
woodlands of Australia (Vesk et al. 2004). Vesk et al. (2004) believe that the different
outcomes of these studies may be related to structural differences in vegetation. In the

Chapter 1. General introduction and thesis overview
16
tropical grasslands where swards are simple and continuous, taller plants are selected, as
there is a ‘top down’ approach to grazing. However in drier woodlands, where the
vegetation is more open, allowing easier movement of grazers and there is a higher
diversity of growth forms (Vesk et al. 2004), simple traits are unlikely to be good
predictors of grazing response (Landsberg et al. 1999). These trends highlight the need
for further empirical studies on plant traits and grazing responses, especially in relation
to site productivity (Vesk et al. 2004).
Gaps created by grazing are most likely to favour plants which produce the highest
quantity of seed per unit biomass (Westoby 1999). Since seed mass is inversely related
to seed output per m2 of canopy (Henery & Westoby 2001), smaller seeded species (e.g.
Eucalyptus) are likely to be more successful in colonising than larger seeded species
(e.g. Callitris) under grazing (Westoby 1999). The applicability of this prediction may
be restricted to higher productivity sites, where the vegetation is denser, rather than
more open lower productivity sites, such as semi-arid woodlands where successful
establishment is dependent on above average rainfall, rather than gap availability (Vesk
et al. 2004).
The theories on the relative success of seeders and resprouters along disturbance
gradients are relatively consistent in their predictions. Seeders (e.g. Callitris) are likely
to succeed at lower intensities and frequencies of disturbances because, firstly they are
usually killed by higher intensity disturbances, and secondly they need sufficient time
between disturbances to reach reproductive maturity. In contrast, resprouters (e.g.
Eucalyptus) are more likely to succeed at higher frequencies and intensities, because of
faster growth rates and their ability to resprout, respectively.
Life history, productivity and disturbances
Relationships between site productivity and disturbances are likely to influence the
relative success of plants with different life histories (Bellingham & Sparrow 2000;
Huston 2003). Site productivity determines growth rates, which determine the time to
reproductive maturity. Since the time required to reach reproductive maturity is longer
at lower productivity sites, resprouters are likely to be more successful than seeders at
lower than higher productivity sites, given a similar disturbance regime (Bellingham &
Sparrow 2000).

Chapter 1. General introduction and thesis overview
17
Productivity and disturbances may also influence life history success through
establishment opportunities. The storage effect (Warner and Chesson 1985) has been
suggested as an important selective pressure for resprouting in drought affected fire-
prone systems. Here seedlings are likely to face a high probability of drought death, so
resprouters are more likely to be successful (Enright & Lamont 1992). Resprouters have
an advantage over seeders in being able to ‘store’ their reproductive potential via their
resprouting ability (Warner and Chesson 1985).
Feedback between vegetation and disturbance regimes can influence the success of life
history types. Vegetation can influence disturbance regimes, since higher biomass
production has the potential to support higher frequencies and intensities of disturbances
(Huston 2003). For example, in fire-prone systems of south eastern Australia, fires are
driven by herbaceous fuels in drier regions and leaf litter in wetter regions (Bradstock
2010). As the climate becomes drier (Nicholls 2004; CSIRO & BOM 2007; Hennessey
et al. 2008; DECCW NSW 2009), fire potential is predicted to decline in drier regions,
as herbaceous biomass declines (Bradstock 2010). In contrast, in wetter regions where
moist fuels may dry, fire potential may increase (Bradstock 2010). Differences in fire
regimes are likely to contribute to varying proportions of life history types along
productivity gradients.
Studies on the success of life history types in relation to productivity and disturbances
are rare. However in one comprehensive study, the evolution of life history types in the
genus Pinus was determined by the interaction of site productivity and fire frequency
(Keeley & Zedler 1998). Resprouting was a common trait where fires were predictable
and stand replacing. In contrast, bark thickness and self-pruning were common traits
where predictable ground fires predominated. Bark thickness and self-pruning were also
common traits at productive sites where less predictable stand replacing fires occurred.
In latter circumstances, the fire return interval may exceed the life span of some Pinus
species, so the ability to resprout was not an advantage.
The theories predict that resprouters are likely to be more successful than seeders at
lower productivity sites when disturbances are present, because of slower growth rates
and limited establishment opportunities for seeders.

Chapter 1. General introduction and thesis overview
18
Summary of predictions
The models are relatively consistent in their predictions of Eucalyptus and Callitris
success with varying productivities and disturbances. Eucalyptus is more likely to
succeed in sites of higher productivity because faster growth rates give Eucalyptus a
competitive advantage over Callitris. Callitris can dominate at lower productivity sites
because of higher water use efficiency. Eucalyptus are predicted to succeed at higher
frequencies and intensities of disturbances, irrespective of productivity, because faster
growth rates allow quicker recovery than Callitris, which is restricted to sites with lower
frequencies and intensities of disturbances.

Chapter 1. General introduction and thesis overview
19
Thesis aims
The aim of this thesis is to examine how the dynamics of woodland dominants of
contrasting functional types, Callitris glaucophylla and Eucalyptus species, are
influenced by climate and disturbance regimes in south eastern Australia.
Specific aims are addressed in chapters 2-4, and the final chapter summarises and
integrates the findings.
Chapter 2. Examines how demographic patterns of Callitris glaucophylla are
associated with rainfall and disturbances along rainfall gradients. This study extended
across the entire study area (Fig. 1.4).
Chapter 3. Examines how competitive feedback between Eucalyptus species and
Callitris glaucophylla trees and C. glaucophylla regeneration influences stand dynamics
along a rainfall gradient. This study occurred along a rainfall gradient in the south east
of the study area.
Chapter 4. Examines how feedback between patches of Callitris glaucophylla and fire
intensity influences Callitris survival in flammable Eucalyptus woodlands in different
fire-weather conditions. This study took place in a large fire-prone reserve in the north
east of the study area.
Chapter 5. Summarises and then integrates the findings using a spatial approach to
illustrate and explain, firstly the current dynamics and distributions of Callitris and
Eucalyptus, and secondly future dynamics under predicted climate changes.
2
3
4
Figure 1.4 A satellite image of Australia showing the scale of each study, numbered by
chapter (Source: Geosciences, Australia)

Chapter 1. General introduction and thesis overview
20

Chapter 2. Demographic patterns of Callitris
21
Chapter 2
Demographic patterns of Callitris glaucophylla are associated with rainfall
and disturbances along rainfall gradients.
Callitris glaucophylla of varying ages in north eastern New South Wales (Source: J. Cohn)

Chapter 2. Demographic patterns of Callitris
22
Chapter 2
Demographic patterns of Callitris glaucophylla are associated with rainfall
and disturbances along rainfall gradients.
Abstract
Predicting species distributions with changing climate has often relied on climatic
variables, but increasingly there is recognition that disturbance regimes should also be
included in distribution models. I examined how changes in rainfall and disturbances
along climatic gradients determined demographic patterns in a widespread and long-
lived tree species, Callitris glaucophylla in SE Australia. I examined recruitment since
1950 in relation to annual (200-600 mm) and seasonal (summer, uniform, winter)
rainfall gradients, edaphic factors (topography) and disturbance regimes (vertebrate
grazing (tenure and species), fire). A switch from recruitment success to failure
occurred at 405 mm mean annual rainfall, coincident with a change in grazing regime.
Recruitment was lowest on farms with rabbits below 405 mm rainfall (mean = 0-0.89
cohorts) and highest on less disturbed tenures with no rabbits, above 405 mm rainfall
(mean = 3.25 cohorts). Moderate levels of recruitment occurred where farms had no
rabbits or less disturbed tenures had rabbits, above and below 405 mm rainfall (mean =
1.77-1.71 cohorts). These results show that low annual rainfall and high levels of
introduced grazing has led to ageing, contracting populations, whilst higher annual
rainfall with low levels of grazing has led to younger, expanding populations. This
study demonstrates how demographic patterns vary with rainfall and spatial variations
in disturbances, which are linked in complex ways to climatic gradients. Predicting
changes in tree distribution with climate change requires knowledge of how rainfall and
key disturbances (tenure, vertebrate grazing) will shift along climatic gradients.

Chapter 2. Demographic patterns of Callitris
23
Introduction
A current challenge in ecology is to determine the processes influencing species
distributions, so shifts in distributions under climate change can be predicted. Many
models identify empirical relationships between current distributions of species and
climatic variables that are used to estimate future distributions of species under climate
scenarios (Thomas et al. 2004; Araujo & New 2007). Since they do not consider
population dynamics, biotic factors and disturbances, it is uncertain whether species will
track predicted shifts in suitable climate space (Pearson & Dawson 2003; Guisan &
Thuiller 2005; Keith et al. 2008). In the few studies where these factors have been
measured, biotic interactions and disturbance regimes were just as important as climate
in determining distribution and thus potential range shifts (Miller & Halpern 1998;
Garcia et al. 1999; Gworek et al. 2007; Boulant et al. 2009). This highlights the need to
include biotic factors and disturbances along with climate in studies identifying drivers
of species distributions.
For widespread species, different processes are likely to control demographic patterns in
different parts of their distributions (Gworek et al. 2007). Climate can directly affect
species distributions by limiting rainfall and temperature (Gworek et al. 2007). For
example, establishment of woody species in low rainfall systems is more dependent on
the infrequent periods of above average rainfall (Denham & Auld 2004; Castro et al.
2005; Holmgren et al. 2006a; Squeo et al. 2007), resulting in fewer opportunities than
in high rainfall systems. Climate can indirectly affect species distributions through
disturbance regimes (Gworek et al. 2007), which may be determined by land use, the
distribution of which is determined by climate (Dale 1997; Luck 2008). For example,
high levels of livestock grazing characteristic of arid systems (Walker 1993; Bureau of
Rural Sciences 2009) have contributed to ageing populations of woody species (Garcia
et al. 1999; Meyer & Pendleton 2005; Auld & Keith 2009). By comparison, in higher
rainfall systems, where a range of land uses are possible (Dale 1997), disturbance
regimes may similarly vary. Livestock grazing levels are likely to be higher on farms
than on other tenures (e.g. roadsides; Lunt 1995; Fensham 1998a), which may act as
refuges, especially for palatable woody species. Whilst fire frequencies have declined in
some agricultural systems, leading to an expansion of fire sensitive woody species
(Belsky & Blumenthal 1997; Noble 1997; Noss et al. 2006), large patches of woodland
may maintain a regime of regular fires (Brookhouse et al. 1999). Identifying where and

Chapter 2. Demographic patterns of Callitris
24
how processes are associated with demographic patterns along climatic gradients, will
allow more precise predictions of distributions under climate change.
Large-scale surveys of the demographic patterns of widespread and long-lived species
can be used to identify the climatic factors and disturbance regimes associated with their
distribution (Peters et al. 2007). Species with widespread distributions offer
comparatively more information on the factors influencing distributions than those with
restricted distributions, since their ranges will encompass greater variability in abiotic
factors (e.g. rainfall), environmental factors (e.g. topography, soil type) and disturbance
regimes (e.g. land use, grazing, fire; Pearson & Dawson 2003). The distribution of trees
is especially valuable because their longevity tells us about past influences (Dale et al.
2001). Seedlings of trees often act as demographic ‘bottle necks’ (Grubb 1977;
Richardson & Bond 1991), because their recruitment is more sensitive to changes in
climate and disturbances than the mortality of adults (Lloyd 1997; Hanson & Weltzin
2000). Changes in the distribution of climatic and disturbance regimes associated with
the demographic patterns of a species will influence its future distribution (Thuiller et
al. 2008).
My aim was to use a long-lived and widespread tree species to examine how rainfall
and disturbances are associated with demographic patterns along climatic gradients.
Callitris glaucophylla provides an ideal model to address this issue. It is a widespread,
long-lived canopy dominant of woodlands ranging from arid to temperate regions of
Australia (Bowman & Harris 1995). It is a slow growing obligate-seeding conifer,
which is sensitive to disturbances, since it does not resprout (Bowman & Harris 1995).
A number of small scale studies have identified varying demographic patterns, which
have been variously linked to climatic and disturbance variables. Recruitment success
has been associated with above average rainfall and recruitment failure with introduced
grazing and high intensity fires (Austin & Williams 1988; Bowman & Latz 1993; Noble
1997). However, it is unclear how these factors vary across climatic gradients and our
ability to predict future changes in distribution is dependent on how these factors
interact in space and time. I predict that: (1) disturbance regimes will vary along rainfall
gradients; and in the absence of inter- and intra-specific competition (2) fewer cohorts
are more likely where rainfall is low, especially where disturbances are frequent; and (3)

Chapter 2. Demographic patterns of Callitris
25
more cohorts are expected where productivity is higher, especially where disturbances
are less frequent.
Methods
Stands of Callitris glaucophylla Thompson & Johnson were sampled at 56 sites along
three transects of declining mean annual rainfall and varying seasonal rainfall
dominance in New South Wales, Australia (Colls & Whitaker 2001; BOM 2010a; Fig.
2.1). Transects extended from the arid zone in the west (200 mm annual rainfall) to the
temperate zone in the east (600 mm annual rainfall; Specht & Specht 1999; Bureau of
Meteorology 2006). Rainfall varies seasonally from north to south with summer
dominant rainfall in the north (60% excess in summer cf winter, Lacey 1973), uniform
rainfall in the middle and winter dominant rainfall in the south (40% excess rainfall in
winter cf summer). Hereafter, transects will be referred to as the summer (149.76oS, -
29.36oE to 141.44
oS, -29.03
oE
), uniform (148.17
oS, -33.15
oE to 141.63
oS, -32.77
oE)
and winter rainfall zones (147.20oS, -34.80
oE to 141.05
oS, -34.01
oE). Each transect was
approximately 600 kms in length. Fieldwork was undertaken from May to August 2008.
S1
S2
S3
S4
S5
S6
0 200
Kilometres
Figure 2.1 The location of transects used to survey Callitris recruitment along gradients
with mean annual rainfall (200 - 600 mm) from east to west in seasonal rainfall zones
from summer in the north, uniform in the middle to winter in the south in New South
Wales. Sites where wood samples were taken for radiocarbon dating are marked from
S1 to S6.

Chapter 2. Demographic patterns of Callitris
26
Sites were chosen to sample the maximum number of cohorts at intervals along each
transect. In the higher rainfall area to the east, sites were on average 30 km apart,
however in the lower rainfall areas in the west, sites were less frequent, and previous
vegetation surveys were used to identify sites within 50 kms north or south of each
transect (Fox 1991; Scott 1992; Porteners 1993, Porteners et al. 1997; Keith 2004,
Benson et al. 2006). A total of 56 sites were sampled including 17 in the winter, 23 in
the uniform and 16 in the summer rainfall zone.
Prerequisites for site inclusion were the presence of mature C. glaucophylla trees (circa
1890 recruitment; Whipp 2009) as a seed source, gaps between canopies and at least
10% bare ground to provide space for potential recruitment, flat to undulating
topography, no evidence of earthworks, logging or fires since the 1950s and minimal
stock grazing. Where possible, fenced roadsides and state forests were sampled, since
these were generally less intensely grazed than travelling stock routes and paddocks
(Fensham 1998a; Lunt 2005). However, the availability of less disturbed sites declined
in the lower rainfall areas in the west, where roadsides were often unfenced from
adjacent farms, and state forests were less common.
At each site a range of variables were measured within a 0.1 ha quadrat. The number of
cohorts which had established since the 1950s was counted. Cohorts were distinguished
by height differences, canopy shape, presence of bark lichens and lower stems and local
knowledge of land managers (Read 1995; Whipp 2009). Cohorts were allocated to a
relative age category from A to D, with A being the oldest and D the youngest at each
site. Where there was uncertainty in differentiating between two cohorts I lumped them
rather than split them, resulting in a higher probability of underestimating than
overestimating cohort numbers. The height and diameter of the two tallest individuals
from each cohort were measured.
An attempt to age Callitris samples using ring counts was not pursued, because of its
imprecision. Instead radiocarbon dating, based on the signature of the bomb-spike was
used (Hua 2009). Radiocarbon dating (14
C) of representative basal disks of Callitris
from two distant sites along each transect (where possible) was used to confirm
recruitment dates (Hua 2009; Fig. 2.1). This involved ageing one replicate from each
cohort at four sites (winter and uniform transects) and two replicates from each cohort
to examine within site reliability of cohort assessment at two sites (summer transect).

Chapter 2. Demographic patterns of Callitris
27
Disks from the younger trees were removed near ground level, but disks from the oldest
trees were removed 20 cm from the ground, reducing the precision of dating the year of
recruitment. Radiocarbon dating used the three innermost rings of each sample and was
confirmed by counting the total number of rings (Hua 2009). Since the precision of
radiocarbon dating is low for the most recent recruits (> 2003), they were aged by ring
counting only.
Topographic position and topsoil texture were recorded in each quadrat (McDonald et
al. 1990). Recent grazing was assessed by recording the presence (presence-absence
data) of native macropods and introduced vertebrates including cattle (Bos taurus
Linnaeus, 1758), sheep (Ovis aries Linnaeus, 1758), European rabbits (Oryctolagus
cuniculus Linnaeus, 1758), and goats (Capra hircus Linnaeus, 1758). Herbivores were
either sighted directly or indirectly by scats, warrens or diggings. Evidence of fire since
the 1950s relied on charred stems or stags of trees that established after 1950. Land
tenure was used as a surrogate for longer term grazing intensity, measured as low to
moderate and high: low to moderate = non-farms, including roadsides fenced from
adjacent farms, travelling stock routes, state forests and national parks; high = farms and
adjacent unfenced roadsides.
At a number of sites in the winter rainfall zone, C. glaucophylla and C. gracilis subsp.
murrayensis J. Garden coexisted. Despite their taxonomic distinction (Hill 1998; Pye et
al. 2003), it was apparent from diagnostic characteristics (cones and pedicels) that they
were producing hybrids (M. Pye pers. comm.). These sites were excluded from
sampling.
Statistical analyses
I used regression tree analyses to examine the suite of variables associated with the
number of cohorts at each site (Table 2.1). Regression trees are one of the few methods
that can model interactions without a priori choice or using stepwise procedures as with
generalised linear modelling (GLM, Breiman et al. 1993). They also accept mixes of
categorical and continuous variables, which is not possible with generalised additive
models (GAM, Thuiller et al. 2003) and they do not assume relations are linear across
the dataset (Breiman et al. 1993). Overfitting was avoided by pruning the tree to
minimise the cross-validated error (Venables & Ripley 2002). Categorical variables
which occurred in fewer than 30% of sites were excluded from the analyses.

Chapter 2. Demographic patterns of Callitris
28
Spearman’s rank coefficient was used to examine collinearity between remaining pairs.
In the first regression tree analysis all explanatory variables with r2 < 0.5 were included
(Tabachnick & Fidell 2007), i.e. annual rainfall (mm), latitude, topographic position
(flat, dune), land use (non-farm, farm), and the presence of rabbits, sheep, cattle, goats
and kangaroos. In the second regression tree analysis, annual rainfall was removed, as a
number of explanatory variables were significantly correlated with annual rainfall
including land use (r2=-0.461, P < 0.001), rabbits (r
2=-0.48, P < 0.0001), goats (r
2=-
0.34, P < 0.011) and topographic position (r2=-0.34, P < 0.011). No other explanatory
variables were significantly correlated. All statistical analyses were undertaken in
SPLUS software version 8 for windows (Insightful Corp. 2007).
Based on the results from the regression tree analysis, I examined trends in the number
of cohorts with annual rainfall in each seasonal rainfall zone, by fitting linear, loess and
sigmoidal relationships. The curve of best fit was chosen using the residual sum of
squares (RSS) and Akaike Information Criterion (AIC). I used RSS and AIC to
discriminate between the models, with lower values of either indicating a better fit
(Quinn & Keough 2002).
I used an analysis of covariance (ANCOVA) to examine the effects of mean annual
rainfall (covariate) and seasonal rainfall (summer, uniform, winter) on an index of
potential opportunities for Callitris regeneration during summer, since 1950. This index
was derived at each site by assuming that Callitris regeneration is dependent on high
rainfall in summer, a well documented phenomenon (FCNSW 1988). Calculation of the
index involved a number of steps. Firstly, within the estimated time of establishment for
each radiocarbon dated sample (Fig. 2.2), I chose the highest summer rainfall
(December-February). Secondly, I derived the minimum summer rainfall required for
regeneration by selecting the lowest rainfall figure from this dataset of high summer
rainfall. Thirdly, I counted the frequency of this minimum value for each site from 1950
to 2008, and from this was able to construct an index of potential opportunities for
Callitris regeneration along each seasonal rainfall transect. Lastly an analysis of
variance (ANOVA) was used to examine the effects of seasonal rainfall (summer,
winter, uniform) and annual average rainfall (< 405 mm, >405 mm) on the index of
potential opportunities for Callitris regeneration. In both ANCOVA and ANOVA, data

Chapter 2. Demographic patterns of Callitris
29
were square root transformed to satisfy homogeneity of variances. Post hoc pairwise
comparisons used Boniferoni correction.
I used non-parametric proportion tests to examine the proportional occurrence of
disturbances, namely farms, rabbits, sheep, cattle, goats, and kangaroos with annual
rainfall (< 405 mm, > 405 mm) and seasonal rainfall zone (winter, uniform, summer).
Fires were too infrequent to analyse. Post hoc tests used the Bonferroni correction.
Table 2.1 A description of the variables used in the regression tree analyses
Variable Values Assessed
Annual rainfall (mm yr-1
) 200-600 BOM (2006)
Seasonal rainfall winter, uniform, summer BOM (2010a)
Landuse fenced roadside, travelling tenure maps and
stock route, state forest observation
national park, farm and
unfenced roadside
Herbivores (rabbits, sheep, present / absent direct (sighting)
cattle, goats) indirect (scats, warrens,
diggings)
Fire since 1950 present / absent charred stems or stags of
Callitris that established
after 1950
Results
Radiocarbon dating and ring counting were used to estimate periods of establishment
for representative Callitris samples. The results indicate that whilst field recognition of
discrete cohorts within sites was reliable, recognition of the same age class between
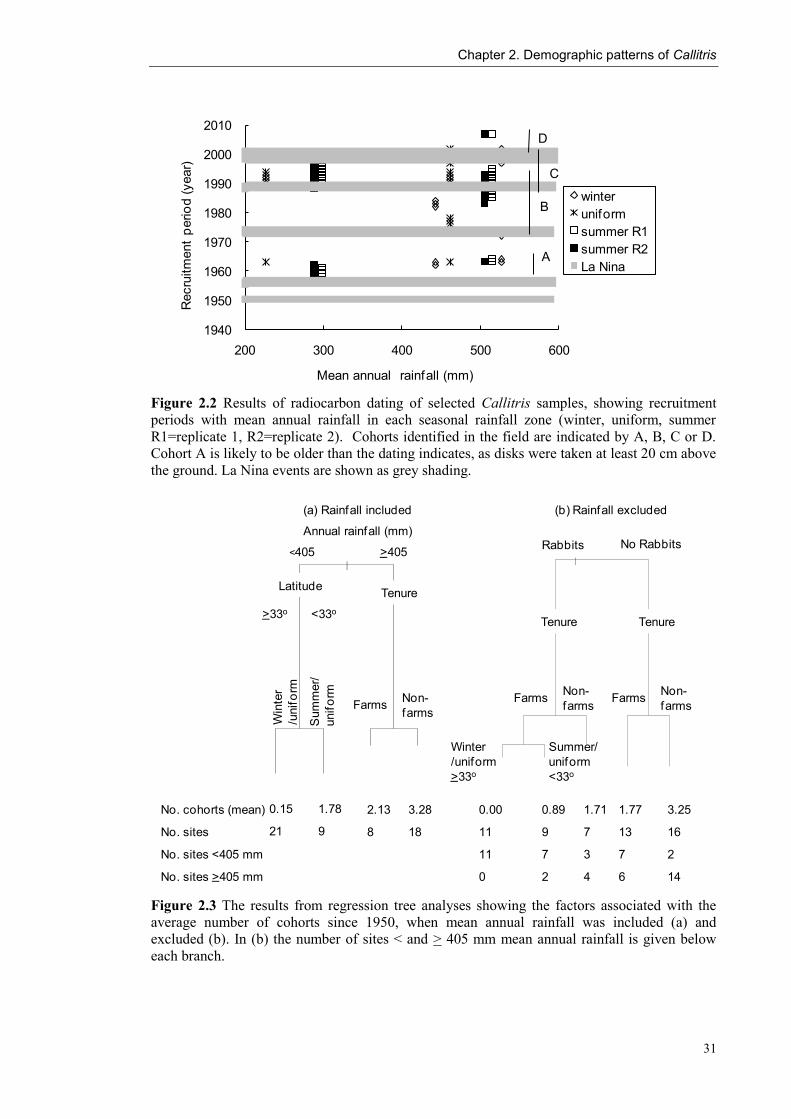
sites was not always reliable. Cohort A was consistently dated from 1959-1964,
although it is likely that these established in the 1950s, since the samples were cut 20
cm above the ground (Fig. 2.2). Cohort B was considered older than C in the field,
however dating showed overlap between sites. Cohort B dated from 1972 to 1996 and
cohort C from 1988 to 2002. Ring counts supported these trends and indicated that
cohort D, which established from 2003 to 2006 was the youngest. In five of the six
cases where replicate samples were dated within a site, both replicates were from the

Chapter 2. Demographic patterns of Callitris
30
same recruitment year. On the one occasion where this did not occur, there was only
one year difference between the two replicates.
The first regression tree analysis indicated that cohort number was associated with
annual rainfall, seasonal rainfall (i.e. latitude) and land tenure (adjusted R2
= 0.65, Fig.
2.3a). The root or primary division in the regression tree occurred at 405 mm annual
rainfall. There were fewer cohorts below 405 mm annual rainfall, especially in the
winter rainfall zone compared with the summer rainfall zone (0.15 cf 1.78). Above 405
mm annual rainfall, farms had fewer cohorts than less disturbed tenures, such as
roadsides, travelling stock routes, state forests and national parks (2.13 cf 3.28).
The second regression tree analysis, which excluded mean annual rainfall, indicated that
cohort number was associated with combinations of rabbit occurrence and land tenure
(adjusted R2=0.65, Fig. 2.3b). The number of cohorts was lowest on farms with rabbits
in the winter and uniform rainfall zones (0.00) and marginally higher in the summer and
uniform rainfall zones (0.89). Most of these sites occurred below 405 mm annual
rainfall. The highest number of cohorts occurred where there were no rabbits on less
disturbed tenures, namely roadsides, travelling stock routes, state forests and national
parks (3.25). Most of these sites occurred above 405 mm annual rainfall. Moderate
numbers of cohorts were on farms without rabbits (1.77) and less disturbed tenures with
rabbits (1.71). These sites were distributed equally above and below 405 mm annual
rainfall.
Curve fitting identified linear and non-linear trends in the number of cohorts with
annual rainfall, depending on the seasonal rainfall zone (Fig. 2.4). Abrupt declines in
the number of cohorts at approximately 400 mm annual rainfall in the winter and
uniform rainfall zones were described by a sigmoid and loess curve, respectively, based
on low values for the residual sum of squares and AIC (Table 2.2). In the summer
rainfall zone, either a linear or a loess curve best describes the relatively unchanging
relationship between cohort number and annual rainfall, depending on whether AIC or
residual sum of squares is used (Table 2.2).
Chapter 2. Demographic patterns of Callitris
31
1940
1950
1960
1970
1980
1990
2000
2010
200 300 400 500 600
winter
uniform
summer R1
summer R2
La Nina
Mean annual rainfall (mm)
Recru
itm
ent
perio
d (year)
A
B
C
D
Figure 2.2 Results of radiocarbon dating of selected Callitris samples, showing recruitment
periods with mean annual rainfall in each seasonal rainfall zone (winter, uniform, summer
R1=replicate 1, R2=replicate 2). Cohorts identified in the field are indicated by A, B, C or D.
Cohort A is likely to be older than the dating indicates, as disks were taken at least 20 cm above
the ground. La Nina events are shown as grey shading.
|
Annual rainfall (mm)
>405
FarmsNon-
farms
1.78
9
3.28
18
0.15
21
<405
TenureLatitude
2.13
8
>33o <33o
Win
ter
/unif
orm
Sum
mer/
unif
orm
|
FarmsNon-
farms
0.89
9
7
2
3.25
16
2
14
0.00
11
11
0
Rabbits No Rabbits
FarmsNon-
farms
1.77
13
7
6
1.71
7
3
4
TenureTenure
Winter
/uniform
>33o
Summer/
uniform
<33o
(a) Rainfall included (b) Rainfall excluded
No. cohorts (mean)
No. sites
No. sites <405 mm
No. sites >405 mm
Figure 2.3 The results from regression tree analyses showing the factors associated with the
average number of cohorts since 1950, when mean annual rainfall was included (a) and
excluded (b). In (b) the number of sites < and > 405 mm mean annual rainfall is given below
each branch.

Chapter 2. Demographic patterns of Callitris
32
Mean annual rainfall (mm)
Num
ber
of
co
ho
rts
(a) Summer rainfall
(b) Uniform rainfall
(c) Winter rainfall
Figure 2.4 Curves of best fit for the number of cohorts since 1950 with mean average rainfall in
each seasonal rainfall zone.
An index of potential opportunities for Callitris regeneration (square root transformed)
during summer, since 1950, was significantly and positively associated with mean
annual rainfall (Table 2.3, Fig. 2.5) and significantly associated with seasonal rainfall
(summer, uniform, winter). Post-hoc pairwise tests indicated that the index was
significantly greater in summer than either the uniform or winter rainfall zones (mean =
5.18, sd = 1.19, mean = 3.53, sd = 0.99, mean=3.50, sd = 0.83, respectively), but not
significantly different between the winter and uniform rainfall zones.

Chapter 2. Demographic patterns of Callitris
33
Table 2.2 Results of fitting curves to the number of cohorts with mean annual rainfall in
each seasonal rainfall zone. Models are ordered from best to worst according to AIC.
Rainfall Curve Residual AIC
zone sum squares
Summer linear 32.96 15.56
loess 26.96 20.40
Uniform loess 5.42 -21.43
linear 29.53 9.75
sigmoid 7.40 -18.08
Winter sigmoid 1.83 -29.86
loess 2.20 -22.87
linear 9.24 -6.36
Table 2.3 Results of ANCOVA used to examine the effects of mean annual rainfall
(covariate) and seasonal rainfall (summer, uniform, winter) on an index of
potential opportunities for Callitris regeneration during summer, since 1950. The index
was square root transformed. Adjusted R2 = 0.925.
Source SS df MS F P
Corrected model 79.33 3 26.44 227.96 0.000
Intercept 0.17 1 0.17 1.42 0.238
Annual rainfall (mm yr-1
) 47.88 1 47.88 412.74 0.000
Seasonal rainfall (s,u,w) 12.79 2 6.39 55.11 0.000
Error 6.03 52 0.116
Total 978.00 56
Corrected total 85.37 55

Chapter 2. Demographic patterns of Callitris
34
0
2
4
6
8
200 250 300 350 400 450 500 550 600 650
winter
uniform
summer
Mean annual rainfall (mm yr-1)
Index o
f re
genera
tion p
ote
ntia
l (s
quare
root tr
ansf
orm
ed)
Figure 2.5 Linear trends in an index of potential opportunities for Callitris regeneration during
summer, since 1950 in different seasonal rainfall zones (summer, uniform, winter) with mean
annual rainfall.
An index of potential opportunities for Callitris regeneration in summer since 1950
(square root transformed), was significantly associated with mean annual rainfall (< 405
mm, > 405 mm) and seasonal rainfall (summer, uniform, winter), but not their
interaction (Table 2.4). There were fewer opportunities for regeneration below 405 mm
than above 405 mm mean annual rainfall (mean = 3.26, se = 0.34 vs mean = 4.43, se =
0.06 ). Opportunities were greater in the summer rainfall zone than in the uniform or
winter (mean = 4.93, se = 0.156; mean = 3.70, se = 0.13; mean = 3.62, se =0.15,
respectively). There was no significant difference in opportunities between winter and
uniform rainfall zones.
Proportion tests were used to examine the occurrence of disturbances with annual
rainfall and with seasonal rainfall (Table 2.5, Fig. 2.6). A significantly greater
proportion of sample sites contained farms, rabbits and sheep below 405 mm annual
rainfall than above 405 mm annual rainfall. The proportion of sites with farms, sheep,
cattle and kangaroos were not significantly different between the seasonal rainfall
zones. The proportion of sites with rabbits was significantly greater in the winter than
the summer rainfall zone and for goats was significantly greater in the uniform than the
winter rainfall zone.

Chapter 2. Demographic patterns of Callitris
35
Table 2.4 Results of ANOVA which examined the effects of seasonal timing of rainfall
(summer, uniform, winter) and annual average rainfall (< 405 mm, >405 mm) on the
index of potential opportunities for Callitris regeneration, since 1950. The index was
square root transformed.
Source SS df MS F P
Corrected model 67.38 5 13.48 37.47 0.000
Intercept 867.43 1 867.43 2411.90 0.000
Seasonal rainfall (s,u,w) 17.23 2 8.61 23.95 0.000
Annual rainfall (< or > 405) 35.04 1 35.04 97.42 0.000
Seasonal*Annual rainfall 0.78 2 0.39 1.08 0.348
Error 17.98 50 0.36
Total 978.00 56
Corrected total 85.37 55
Table 2.5 Results of proportions tests examining the occurrence of disturbances with
mean annual rainfall (below 405 mm, above 405 mm) and seasonal rainfall (winter W,
uniform U, summer S).
Disturbance Mean annual rainfall (mm) Seasonal rainfall zone
X2 df P X
2 df P
Farms 13.80 1 0.002 below>above 3.37 2 0.190
Rabbits 10.20 1 0.001 below>above 6.86 2 0.030
W>S
Sheep 3.91 1 0.048 below>above 0.41 2 0.810
Cattle 1.31 1 0.250 1.49 2 0.480
Goats 1.31 1 0.290 9.61 2 0.008 U>W
Kangaroos 0.04 1 0.840 0.11 2 0.950

Chapter 2. Demographic patterns of Callitris
36
Pro
po
rtio
n o
f sites
Mean annual rainfall (mm)
0.0
0.2
0.4
0.6
0.8
1.0Farms
0.0
0.2
0.4
0.6
0.8
1.0Rabbits
0.0
0.2
0.4
0.6
0.8
1.0
<405 >405
Sheep
Cattle
winter
uniform
summer
Kangaroos
0.0
0.2
0.4
0.6
0.8
1.0 Dunes
Goats
<405 >405
Fire
Figure 2.6 Proportion of sample sites containing farms, grazing species and dunes in
annual (< and > 405 mm) and seasonal rainfall zones (winter, uniform, summer). Means
and standard errors are given.

Chapter 2. Demographic patterns of Callitris
37
Discussion
These results indicate that demographic patterns of this widespread and long-lived tree
species varied greatly along rainfall and disturbance gradients. Ageing Callitris
populations were associated with low annual rainfall and high grazing levels, whilst
younger, expanding populations were associated with higher annual rainfall and low to
moderate grazing levels. Although these processes cannot be easily disentangled at the
survey scale, the results indicate the inextricable linking of climate and disturbance
regimes.
Annual rainfall
Demographic patterns varied widely across the species distribution. Recruitment failure
has led to ageing Callitris populations below 405 mm annual rainfall in the uniform and
winter rainfall zones. Today, these populations are largely represented by trees which
established in the 1850s or 1890s (Austin & Williams 1990). This is despite a number
of opportunities for recruitment since 1950, provided by favourable high rainfall during
summer, necessary for seedling survival (Fig. 2.5). Studies in this region (Auld & Keith
2009) and in other low rainfall systems worldwide have similarly reported ageing
populations of long-lived species (Garcia et al. 1999; Meyer & Pendleton 2005).
Elsewhere in the region where annual rainfall was higher, Callitris populations are
younger, similar to other semi-arid systems (Kraaij & Ward 2006; Graz 2008; Sankaran
et al. 2008; Boulant et al. 2009). These populations are consistently represented by trees
which established in the 1950s and then at various times through to 2007. A maximum
of four cohorts was recorded at any one site, but this is likely to be an underestimate,
especially where structural complexity was high. These trends are consistent with the
predictions that cohort numbers were more likely to increase with rainfall or
productivity.
Seasonal rainfall
Recruitment success was greater in the summer than either the winter or uniform
rainfall zones. Summer drought limits recruitment, especially in low rainfall systems, so
significant rainfall during this period can increase the recruitment success, by increasing
growth rates and reducing the probability of desiccation and subsequent damage by
grazers (Denham & Auld 2004; Castro et al. 2005; Holmgren et al. 2006b; Squeo et al.
2007). However, an index of opportunities for seedling recruitment during summer was

Chapter 2. Demographic patterns of Callitris
38
comparable between all seasonal rainfall zones below 405 mm mean annual rainfall.
This suggests that disturbances were responsible for the absence of Callitris recruitment
in the winter and uniform rainfall zones compared with some success in the summer
rainfall zone, below this threshold in annual rainfall. This is consistent with the
hypothesis that fewer cohorts were more likely at lower rainfall, especially where
disturbances were more frequent.Since long-lived species need few recruitment events
to maintain their populations (Watson et al. 1997), one recruitment event in 120 - 160
years may be the difference between local extinction or survival, given that the
longevity of Callitris is approximately 250 years (Bowman & Harris 1995).
Herbivory and fire
As hypothesised the distribution of disturbance regimes varied inextricably with rainfall
gradients. Below 405 mm annual rainfall, most of the sites were on farms which grazed
livestock, a common form of agriculture in lower rainfall systems (Walker 1993; Kraaij
& Ward 2006; Bureau of Rural Sciences 2009). Above 405 mm annual rainfall, a
broader range of land uses reflected greater agricultural flexibility with higher rainfall.
Since the study’s intention was to examine a site’s maximum potential for recruitment, I
was able to select tenures with lower levels of disturbance above 405 mm annual
rainfall. These land uses often included fenced roadsides, travelling stock routes, state
forests and national parks, all of which have historically lower levels of grazing than
farms, thus acting as refuges for native plants (Lunt 1995; Fensham 1998a). The
difference in grazing levels either side of 405 mm annual rainfall was reflected in the
higher occurrence of sheep and rabbits below 405 mm. Fires were almost absent above
405 mm annual rainfall, occurring in only 1 of 30 sites compared with 7 of 26 sites
below 405 mm annual rainfall. Noble (1997) believes there has been a decline in fire
frequency in this fragmented agricultural landscape since European management in the
1850s, suggesting that pre European fire regimes may have made establishment more
difficult for Callitris. These overall trends suggest higher levels of disturbances and or
fewer refuges from disturbances below 405 mm annual rainfall, since European
management.
The distribution of disturbances also varied with seasonal rainfall. Smaller herbivores,
like rabbits and sheep, prefer C3 grasses (Andrzejewska & Gyllenberg 1980; Van Dyne
et al. 1980; Heckathorn et al. 1999), which are more abundant in the winter rainfall

Chapter 2. Demographic patterns of Callitris
39
zone of southern Australia (Hattersley 1983; Murphy & Bowman 2007), whereas larger
herbivores, like cattle, prefer C4 grasses, which are more abundant in the summer
rainfall zone of northern Australia. Although I did not detect a pattern in the distribution
of cattle and sheep with seasonal rainfall, rabbits, whose successful breeding from
autumn to early summer is dependent on forage produced from winter rainfall, were
more common in the south than the north (Cooke 1977; Parer 1987). Goats were most
common in the uniform rainfall zone. Their generalist browsing diet (Landsberg & Stol
1996), may give them a greater tolerance to drought than sheep. Kangaroos were
ubiquitous. These trends suggest that at least rabbits are distributed in relation to food
availability, which is largely determined by seasonal rainfall.
Relationships between annual rainfall and grazing regimes were closely associated with
trends in recruitment success. In particular, the distributions of land tenure and rabbits
were associated with recruitment success across the species range. Callitris recruitment
was lowest below 405 mm annual rainfall, where rabbits occurred on farms which
grazed mainly sheep, but also goats and cattle. In other studies of low rainfall systems,
recruitment failure of woody species is related to a combination of drought and high
levels of grazing (Holmgren et al. 2006b; Gutierrez et al. 2007; Auld & Keith 2009).
While I cannot easily disentangle grazing from climatic effects in the survey, numerous
studies have illustrated the negative effects of grazing on recruitment in low rainfall
systems (see Holmgren et al. 2006b; Auld & Keith 2009). In particular, introduced
rabbits and livestock have reduced or prevented the regeneration of a range of palatable
woody species, in the arid zones of Australia (see Auld & Keith 2009) and elsewhere in
the world (Holmgren et al. 2006b; Gutierrez et al. 2007). From 1890 to 1950, Callitris
recruitment was prevented by high numbers of rabbits in SE Australia (Austin &
Williams 1988; Noble 1997). Widespread recruitment resumed in the 1950s following
widespread above average rainfall (La Nina) and the release of a virus which reduced
rabbit numbers (Austin & Williams 1988; Noble 1997). Trends in recruitment in my
study agree with this. However, the infrequency of recruitment below 405 mm annual
rainfall in the winter and uniform rainfall zones, suggests, that at least in some
locations, rabbit numbers did not decline to levels allowing recruitment. Indeed, the
lowest recruitment occurred in the winter rainfall zone where rabbits and Callitris often
coexisted on dunes. Warrens, which are necessary for the reproductive success of

Chapter 2. Demographic patterns of Callitris
40
rabbits in arid zones, are easily constructed in the sandy soils of dunes (Parer 1987).
Higher recruitment in the uniform and summer rainfall zones where rabbits were absent,
suggests patchily distributed livestock grazing (Holmgren et al. 2006a, 2006b; Gutierrez
et al. 2007), either from chance, or management or a lag in return to pre-drought levels,
since wet conditions usually follow drought (Meserve et al. 2003; Letnic et al. 2005).
These trends suggest that a combination of high levels of grazing, typical of rangelands
and rainfall outside the growing season has led to recruitment failure since the mid to
late 1800s. Exclusion of grazers is likely to increase the probability of recruitment
success (Holmgren et al. 2006b; Auld & Keith 2009).
Expanding populations were associated with lower levels of grazing. The highest
recruitment success was in less disturbed tenures with no rabbits. These tenures
included roadsides, travelling stock routes, state forests and national parks. Callitris is
sensitive to grazing, especially by rabbits and livestock (Lacey 1973; Allcock & Hik
2004). The recruitment successes of other woody species have been similarly associated
with lower rather than higher levels of grazing (Lunt 2005; Boulant et al. 2009), but not
in all cases (Richardson & Bond 1991; Brown & Archer 1999). Callitris is also fire
sensitive, since it is often killed by fire and must regenerate from seed. A decline in fire
frequency since European management (Noble 1997) has no doubt contributed to
recruitment success of Callitris, as it has with fire sensitive species in other systems
(Cooper 1960; Belsky & Blumenthal 1997). This contrasts markedly with ageing and
contracting populations of the fire sensitive Callitris intratropica in northern Australia,
where fires have increased in frequency and intensity since European management
(Bowman & Panton 1993). The relative absence of disturbances in certain tenures above
405 mm annual rainfall, is undoubtedly advantageous to the recruitment of the slow
growing and shade tolerant Callitris.
Climate change
Climate change is likely to have both direct and indirect effects on recruitment and thus
the future distributions of Callitris. The predicted increases in frequency, areal extent
and severity of droughts in SE Australia (Nicholls 2004; CSIRO & BOM 2007;
Hennessy et al. 2008), is likely to most severely affect arid and semi-arid regions
(Holmgren et al. 2006a). These conditions may further reduce opportunities for
recruitment below or immediately above 405 mm annual rainfall, if there is an easterly

Chapter 2. Demographic patterns of Callitris
41
shift in this isohyet, by increasing the chance of desiccation, identified by other studies
as contributing to recruitment failure in Callitris (Johnson 1969; Read 1995). It may
also accelerate senescence of ageing Callitris, removing the only remaining seed
sources in the south west (Foden et al. 2007; Jump et al. 2009). Indirectly, changes or
shifts in key disturbances, such as land tenure and grazing species, might also influence
the distribution of Callitris, as in other woody species (Loehle & Le Blanc 1996;
Turner, et al. 1998; Dale et al. 2001). If farms which graze livestock expanded east to
displace unprofitable crop lands (Dale 1997; Garnaut 2010), recruitment failure may
shift in the same direction, especially if refuges like travelling stock routes disappear.
Predicted declines in the populations of rabbits and livestock as pasture biomass
declines with higher temperatures (Scanlan et al. 2006; Gunasekera et al. 2008) may not
be enough to compensate the direct effects of drought on Callitris seedling recruitment
in an expanding low rainfall zone.
Summary
Demographic patterns in Callitris, a widespread and long-lived tree species were closely
associated with the spatial arrangement of disturbance regimes and their relationship
with rainfall along rainfall gradients. Disturbance regimes were associated with rainfall
through different land use types. As climate changes, rainfall, land use and disturbance
regimes will shift along climatic gradients, influencing species distributions. To predict
changes in the distribution of Callitris and other species, we need to predict shifts in
both disturbance regimes and climate, and disentangle their relative importance along
climatic gradients.
Acknowledgments
Australian Research Council (ARC) Linkage Grant between Charles Sturt University
(CSU) and NSW Department of Environment and Climate Change (DECC) provided
the funding for this research (Project number LP066829). A grant from Australian
Institute for Nuclear Science and Energy (AINSE) provided funding for the radiocarbon
dating of Callitris wood samples (AINGRA09075). I would like to thank Stephen
Campbell and Alan Smith from State Forests NSW for their assistance and permission
to take samples. I would also like to thank John Warren and Kate Hammill for
assistance in the field and Simon McDonald for statistical advice.

Chapter 2. Demographic patterns of Callitris
42

Chapter 3. Competitive feedback between canopy trees and regeneration
43
Chapter 3
Competitive feedback between Eucalyptus species and Callitris
glaucophylla trees and C. glaucophylla regeneration influences stand
dynamics along a rainfall gradient
Eucalyptus species canopy tree (left background) surrounded by dense Callitris glaucophylla regeneration in central western New South Wales (Source: K. Ross ).

Chapter 3. Competitive feedback between canopy trees and regeneration
44
Chapter 3
Competitive feedback between Eucalyptus species and Callitris
glaucophylla trees and C. glaucophylla regeneration influences stand
dynamics along a rainfall gradient
Abstract
Little attention has been given to the influence of canopy tree competition on the
encroachment of woody species in semi-arid woodlands of SE Australia. I examined
how canopies of Eucalyptus species and Callitris glaucophylla influenced the survival,
growth and reproduction of C. glaucophylla saplings and how these in turn influenced
the survival of the canopy trees along a rainfall gradient during severe drought
conditions. I sampled in six forests along a rainfall gradient (400 - 600 mm). In each
forest I sampled saplings under canopies of Eucalyptus and Callitris and in adjacent
gaps (4-6 replicates per canopy species). Canopy tree survival was assessed along two
transects in each forest, one with and one without regeneration. Saplings under canopies
had lower densities, were smaller and were less likely to be reproducing than those in
gaps. Sapling densities under canopies decreased from 1.5 to 1.3 m-2
as annual rainfall
increased from 400 to 600 mm. Mortality was low for canopy trees not surrounded by
Callitris regeneration (0 – 10%), but Callitris and Eucalyptus canopy trees surrounded
by Callitris regeneration had higher mortality and stress levels, respectively. Callitris
canopy tree mortality was relatively uniform across the rainfall gradient (0 – 25%) and
the proportion of Eucalyptus canopy trees with higher stress levels varied from 0.95 to
0.30 as annual rainfall increased from 400 to 600 mm. These results suggest that
canopies do not exclude but rather moderate Callitris sapling densities, which in turn
increase the mortality and stress levels of Callitris and Eucalyptus canopy trees during
drought. As drought severity increases under climate change, and ageing Eucalyptus
canopy trees die and are not replaced, the system may shift further from Eucalyptus and
Callitris co-dominance, typical of woodlands prior to European management to a
simplified Callitris dominated system.

Chapter 3. Competitive feedback between canopy trees and regeneration
45
Introduction
Woody plant encroachment is a major issue in semi-arid regions throughout the world
(Scholes & Archer 1997; Asner et al. 2004). Mechanisms responsible for its occurrence
are still poorly understood, but have variously been attributed to increased CO2, and
changes in climate, soils, grazing and fire regimes (Bond & Midgley 2000; Sankaran et
al. 2005; Fensham et al. 2005). The most commonly cited mechanism is a decline in fire
frequency and intensity, resulting from reductions in grass fuels by introduced grazers
(Noble 1997; Scholes & Archer 1997; Asner et al. 2004). In particular, this has
contributed to the encroachment of fire-sensitive woody plants (Belsky & Blumenthal
1997; Burrows 2005).
The effects of existing tree canopies on woody plant encroachment are less easily
predicted from the ecological literature. Patterns vary from establishment beneath
canopies or in gaps or continuous filling from gap to canopy (Harrington et al. 1981;
Haase et al. 1996; Belsky & Blumenthal 1997; Barnes & Archer 1999). The success of
woody species establishment under canopies is partly dependent on their requirements
for water, nutrients and light and how these are modified by canopy tree species along
rainfall gradients (Veblen 1992; Bertness & Callaway 1994). Most evidence for positive
effects of canopies on woody establishment is at lower rainfall, where canopies improve
micro-climatic conditions (e.g. temperature and water), thereby facilitating
establishment (Holmgren et al. 1997).
The influence of woody encroachment on canopy plants is generally negative (Callaway
& Walker 1997; Belsky & Blumenthal 1997; Barnes & Archer 1999). The increased
competition can accelerate mortality of canopy plants by reducing their vigour and
increasing their vulnerability to insect attack and pathogens, especially during periods of
water deficit (Belsky & Blumenthal 1997; Barnes & Archer 1999). Increased
competition can also reduce opportunities for establishment of canopy plants by
reducing their seed production or making conditions unsuitable for their establishment
by pre-empting resources (McPherson et al. 1988; Barnes & Archer 1999) or changing
fire regimes (Covington & Moore 1994). So, strong inter-specific competition has the
potential to lead to major changes in species composition (Belsky & Blumenthal 1997,
Barnes & Archer 1999).

Chapter 3. Competitive feedback between canopy trees and regeneration
46
Climate, especially rainfall influences the demography of woody plants in semi-arid
systems (Schwinning et al. 2004; Fensham et al. 2005; Holmgren et al. 2006a;
Shinneman & Baker 2009). Woody plants establish during periods of above average
rainfall (Schwinning & Sala 2004; Fensham et al. 2005; Kraaij & Ward 2006) and have
increased rates of mortality during droughts (Holmgren et al. 2006a; Fensham et al.
2009; Shinneman & Baker 2009). As droughts are predicted to increase in severity in
many semi-arid systems (IPCC 2007), there may be fewer opportunities for woody
species establishment (Garcia et al. 1999; Castro et al. 2005) and accelerated rates of
thinning (Fensham et al. 2009). Given that droughts preferentially thin at higher levels
of competition (Belsky & Blumenthal 1997; Guarin & Taylor 2005; Fensham et al.
2009), dense woody regeneration and ageing canopy trees that overtop them are likely
to suffer higher levels of mortality than those at lower levels of competition (Belsky &
Blumenthal 1997).
Understanding how climate influences competitive interactions between canopy trees
and regeneration are vital for predicting future changes in semi-arid systems, especially
under climate change. Within this system, competitive interactions may vary along a
rainfall gradient. For example, canopies more commonly facilitate seedling
establishment at lower than higher rainfall (Holmgren et al. 1997). Canopy tree
densities are likely to decline with declining rainfall (Specht & Specht 1999) or may be
influenced by disturbances, like logging (Lunt et al. 2006). Drought may also have a
greater negative effect on plant survival at lower than higher rainfall (Lacey 1972). So,
the combined effects of competition and drought may variably drive plant dynamics
along a rainfall gradient.
Semi-arid SE Australia underwent dense encroachment by a range of woody species,
including Callitris glaucophylla, after European settlement in the 1800s (Hodgkinson &
Harrington 1985; Noble 1997). This encroachment coincided with widespread changes
in fire and grazing regimes and canopy tree removal (Noble 1997). In many places
woodland dominants, C. glaucophylla and a range of Eucalyptus species were both
removed or ringbarked at different rates, shifting dominance from Eucalyptus species to
C. glaucophylla (Lunt et al. 2006). Although the dynamics of monospecific stands of C.
glaucophylla have been well researched at the stand scale (Lacey 1972, 1973; FCNSW
1988), there is comparatively little research on dynamics within mixed stands of

Chapter 3. Competitive feedback between canopy trees and regeneration
47
Eucalyptus species and C. glaucophylla (Clayton-Greene 1981; Clayton-Greene &
Ashton 1990).
Eucalyptus species are fast growing, resprouting angiosperms with large open canopies,
whilst C. glaucophylla is a slow growing, obligate-seeding gymnosperm with a
relatively small dense canopy (Lacey 1973; FCNSW 1988; Bowman & Harris 1995;
Williams & Woinarski 1997). Both genera are considered drought tolerant, and achieve
low water usage by different means (Zeppel & Eamus 2008). Callitris glaucophylla
reduce canopy conductance, whereas Eucalyptus species reduce leaf area by shedding
leaves before recovery by resprouting (Pook et al. 1966; Attiwill & Clayton-Greene
1984; Clayton-Greene & Ashton 1990). During drought, mortality of a range of
Eucalyptus species and C. glaucophylla has been greatest at higher levels of
competition and in drier areas (Lacey 1972, 1973; Allen 1998; Fensham & Holman
1999; Fensham et al. 2009). These patterns have only rarely been quantified (e.g.
Fensham & Holman 1999; Fensham et al. 2005; Fensham et al. 2009). Quantification
will allow better predictions of woody dynamics as climate changes in these semi-arid
systems.
My aim was to examine how canopy trees of Eucalyptus species and C. glaucophylla
influenced the survival, growth and reproduction of C. glaucophylla saplings and how
these in turn influenced the survival of the canopy trees along a rainfall gradient during
drought conditions. The following hypotheses were proposed: (1) densities of saplings
will be greater in gaps than under canopies, and as rainfall increases, densities will
increase in gaps and decrease under canopies; (2) growth rates and the probability of
reproducing will be greater for saplings without canopy or sapling competition and will
increase in areas of higher rainfall; (3) sapling densities, growth rates and the
probability of reproducing will be greater under Eucalyptus than Callitris canopies; (4)
denser saplings will experience greater mortality during a drought; (5) recent drought
mortality of Eucalyptus and Callitris canopy trees will be greater at lower rainfall when
dense sapling regeneration is present.
Methods
The study was restricted to the zone of coexistence of Callitris glaucophylla Thompson
& Johnson and a range of Eucalyptus species in semi-arid central New South Wales,
south-eastern Australia. The study area encompassed a 250 km long gradient of mean

Chapter 3. Competitive feedback between canopy trees and regeneration
48
annual rainfall, from 600 mm in the east (148.21oS, -33.34
oE) to 400 mm in the west
(145.57oS, -32.74
oE). Sampling occurred within six State Forests distributed along this
rainfall gradient. Fieldwork was undertaken from October to December 2008. A
drought which started in 2001 resulted in extremely dry years in 2002 and 2006 (CSIRO
2008; Fig. 3.1). Mean annual rainfall at each site from 2001 to 2008 was lower than the
long-term mean annual rainfall at each site: from west to east sites experienced 0.88,
0.76, 0.78, 0.88, 0.72 and 0.79 of their long-term mean annual rainfall (Fig. 3.1; Bureau
of Meteorology 2010).
Callitris saplings
Within each of the six sites, I randomly selected canopy trees of Callitris and
Eucalyptus that were surrounded by dense Callitris saplings (Fig. 3.2a). I defined two
zones, the canopy zone under the canopy and the gap zone which extended beyond the
edge of the canopy and was free from any overhead foliage. At each selected canopy
tree, I sampled Callitris saplings within a quadrat under the canopy and another in the
adjacent gap. Quadrats were 8 m2 and there were 4-6 replicates of each canopy tree
species at each site. I sampled the dominant cohorts, namely Callitris and Eucalyptus
canopy trees which established in the 1890s and Callitris saplings from the 1950s
(Lacey 1972; Allen 1998; Whipp 2009). Comparative sampling of Eucalyptus saplings
was not undertaken because of their infrequency in the region.
The density of live and recently dead Callitris saplings was recorded in each quadrat.
Recently dead Callitris saplings were identified by the presence of brown leaves or fine
twigs. To compare the intensity of competition between canopies and gaps, the height,
diameter at breast height (DBH) and the presence of fruiting cones on a range of live
Callitris sapling types were measured: closest to the trunk of canopy trees; random in
gaps (randomly throwing a stone); dominant (tallest) in gaps; and isolated from other
saplings or tree canopies by at least 1m in gaps. Isolated saplings were measured outside
the gap quadrat. Saplings closest to the trunk of the canopy tree approximated maximum
canopy competition and minimal sapling competition and are hereafter referred to as
saplings under canopies. I measured one of each sapling type at each canopy tree and
also recorded the height and canopy area of the latter.
Chapter 3. Competitive feedback between canopy trees and regeneration
49
0
200
400
600
800
1000
1200 (a) Yathong NR
0
200
400
600
800
1000
1200 (b) Yelkin SF
0
200
400
600
800
1000
1200 (c) Weelah SF
0
200
400
600
800
1000
1200 (d) East Cookeys Plains SF
0
200
400
600
800
1000
1200 (e) Weddin SF
0
200
400
600
800
1000
1200
1950
1953
1956
1959
1962
1965
1968
1971
1974
1977
1980
1983
1986
1989
1992
1995
1998
2001
2004
2007
(f ) Backyamma SF
Year
Rain
fall
(mm
yr-1
)
Figure 3.1 Annual rainfall at the six sites from west to east (top to bottom) along the
rainfall gradient from 1950 to 2008 (BOM 2010). The period of recent drought from
2001 to 2008 is shaded grey. Long-term mean annual rainfall is represented by the
horizontal lines.
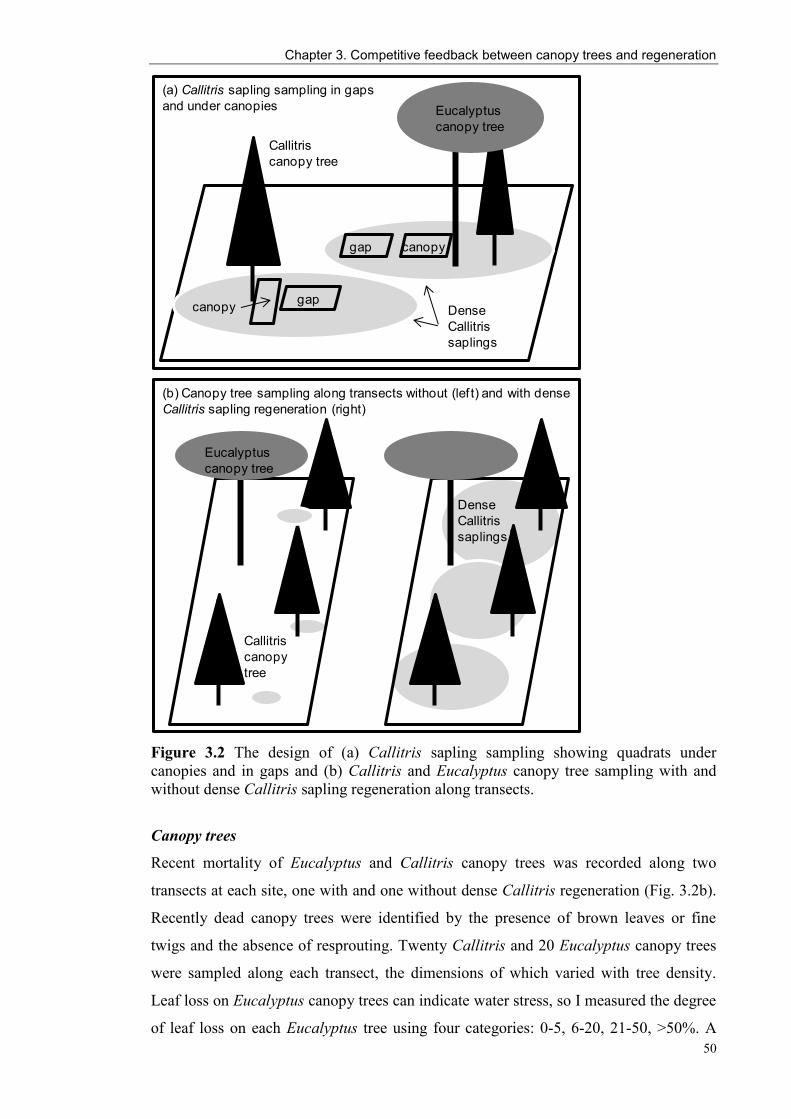
Chapter 3. Competitive feedback between canopy trees and regeneration
50
Dense
Callitris
saplings
Callitris
canopy tree
Eucalyptus
canopy tree
(a) Callitris sapling sampling in gaps
and under canopies
gap
gap canopy
canopy
Dense
Callitris
saplings
Callitris
canopy
tree
Eucalyptus
canopy tree
(b) Canopy tree sampling along transects without (lef t) and with dense
Callitris sapling regeneration (right)
Figure 3.2 The design of (a) Callitris sapling sampling showing quadrats under
canopies and in gaps and (b) Callitris and Eucalyptus canopy tree sampling with and
without dense Callitris sapling regeneration along transects.
Canopy trees
Recent mortality of Eucalyptus and Callitris canopy trees was recorded along two
transects at each site, one with and one without dense Callitris regeneration (Fig. 3.2b).
Recently dead canopy trees were identified by the presence of brown leaves or fine
twigs and the absence of resprouting. Twenty Callitris and 20 Eucalyptus canopy trees
were sampled along each transect, the dimensions of which varied with tree density.
Leaf loss on Eucalyptus canopy trees can indicate water stress, so I measured the degree
of leaf loss on each Eucalyptus tree using four categories: 0-5, 6-20, 21-50, >50%. A

Chapter 3. Competitive feedback between canopy trees and regeneration
51
full canopy was considered to be one where twigs and smaller branches had the
potential to have leaves, so gaps in this potential were measured as a percentage loss.
Stem damage progresses from the apex to the base, so lower resprouting positions
indicate higher stress levels (Vesk & Westoby 2003). I recorded the lowest reprouting
position on each Eucalyptus canopy tree using four categories: base, trunk, branch,
none.
Statistical analyses
I examined how a range of variables influenced sapling density, height and DBH in
three separate analyses. Since measurements of saplings under each canopy and in the
adjacent gap were not independent, they required an analysis that could accommodate a
split design and also continuous (rainfall) and categorical variables. So, I used nested
analyses of variance and I partitioned the five degrees of freedom for site into linear,
quadratic and cubic effects to examine regressions between rainfall and the response
variables (Steel & Torrie 1960, Quinn & Keough 2002). Testing for linear, quadratic
and cubic effects allowed me to examine the shape of the relationship between rainfall
and the response variables. More specifically, I examined the effects of site (1-6), mean
annual rainfall, canopy species (Eucalyptus, Callitris) and canopy position (gap,
canopy) on the density of live Callitris saplings. I also examined the effects of site,
rainfall, canopy species and sapling type (under canopy, random in gap, dominant in
gap, isolated in gap) on the height and DBH of live Callitris saplings. Post hoc Fishers
LSD tests were used for paired comparisons. Diagnostic checks of these and subsequent
models included examination of the residuals for outliers and goodness of fit. Analyses
were undertaken in GenStat (GenStat 2009).
Linear regressions were used to examine the recent mortality of Callitris saplings in
gaps and under canopies. In gaps, I examined the effects of the total density of saplings
(live and recently dead) and mean annual rainfall on the density of recently dead
saplings. Under canopies, I examined the effects of the same variables and canopy tree
species (Callitris, Eucalyptus) on the density of recently dead Callitris saplings. The
density of recently dead saplings was log transformed to satisfy Levene’s test of
homogeneity and to improve normality. These tests and the following were undertaken
in SPLUS software v8 for Windows (Insightful Corp. 2007).

Chapter 3. Competitive feedback between canopy trees and regeneration
52
Wilcoxon signed-rank tests were used to examine differences in canopy area, height and
DBH between Callitris and Eucalyptus canopy trees. Data within sites were paired, to
take into account between site differences. Non-parametric testing was used because
data on canopy area and DBH could not be transformed to satisfy the assumption of
normal distribution, required for parametric testing (Quinn& Keough 2002).
Wilcoxon signed-rank tests (paired) were used to examine whether the proportion of (a)
Callitris that died, (b) Eucalyptus with leaf stress (>5% leaf loss), and (c) Eucalyptus
resprouting from the trunk, were influenced by the presence of dense Callitris sapling
regeneration. I then used Spearman’s rank correlation tests to examine relationships
between the proportion of (a-c) where Callitris sapling regeneration was present, and a
range of factors. These factors included mean annual rainfall, density of Callitris or
Eucalyptus canopy trees, and the mean density of Callitris saplings (live and recently
dead) under the respective canopies of Callitris and Eucalyptus at each site. There were
no analyses of Eucalyptus mortality, because only two deaths were recorded.
Results
Canopy effects on Callitris sapling density along the rainfall gradient
There was a significant linear relationship between Callitris sapling density and canopy
position. As predicted sapling densities were greater in gaps than under canopies and as
mean annual rainfall increased, the density of Callitris saplings increased in gaps and
decreased under canopies (Table 3.1, Fig. 3.3). A significant cubic relationship between
the Callitris sapling density and mean annual rainfall indicated site effects, when
saplings in gaps and under canopies were not differentiated. These regression results
agreed with the significant effects of site, canopy position and their interactions on
Callitris sapling density in the ANOVAs (Table 3.1). Sapling densities varied with site,
with canopy position and their interaction. Contrary to predictions, densities under
Callitris canopies were not significantly different from those under Eucalyptus
canopies.
Canopy effects on Callitris sapling growth and reproduction along the rainfall
gradient
The height and DBH of saplings was significantly associated with site, canopy species,
sapling type and their interactions, but not mean annual rainfall (Table 3.1). As
predicted, Callitris saplings in gaps had higher growth rates than those under canopies,

Chapter 3. Competitive feedback between canopy trees and regeneration
53
but contrary to predictions growth rates were not greater at higher rainfall. Callitris
saplings under canopies (Eucalyptus and Callitris combined) were similar in height to
random saplings in gaps, both being shorter than dominant saplings in gaps, which were
shorter than isolated saplings in gaps (Fig. 3.4a). Similarly, Callitris saplings under
canopies (Eucalyptus and Callitris combined) had bigger DBHs than random saplings in
gaps but smaller than dominant saplings in gaps which were smaller than isolated in
gaps (Fig. 3.4b). As predicted, Callitris saplings under Eucalyptus canopies were taller
and had larger DBHs than those under Callitris canopies (Figs 3.4a, 3.4b). Significant
cubic and quadratic relationships between mean annual rainfall and Callitris sapling
height and DBH indicated site effects, as did the significant cubic relationship between
height or DBH of sapling types and rainfall.
The proportion of Callitris saplings with fruiting cones varied with sapling type along
the rainfall gradient (Fig. 3.4c). As predicted Callitris saplings in gaps were more likely
to reproduce than those under canopies, but contrary to predictions this probability did
not appear to increase with rainfall. Whilst Callitris saplings under canopies and
random saplings in gaps rarely produced cones, dominant saplings in gaps were more
likely to produce cones at low rainfall. Most isolated Callitris saplings in gaps produced
cones except at two sites around 500 mm mean annual rainfall.
Callitris sapling mortality
As predicted the density of recently dead Callitris saplings was significantly associated
with the combined densities of live and recently dead Callitris saplings in both gaps and
under canopies (Table 3.2). The density of recently dead saplings increased as the
combined density increased (Figs 3.5a, 3.5b). The density of recently dead Callitris
saplings in gaps and under canopies was not significantly associated with annual
rainfall. This implies that the combined densities of saplings were not linearly related to
annual rainfall, but are thinning to densities which are (Fig. 3.3). The density of recently
dead Callitris saplings in gaps and under canopies was not significantly associated with
canopy species.

Chapter 3. Competitive feedback between canopy trees and regeneration
54
Mean annual rainfall (mm yr-1)
y = 0.0028x + 0.6475R² = 0.1362
y = -0.0018x + 2.2541R² = 0.0974
0
1
2
3
4
400 450 500 550 600
gap
canopy
Density (no
. m
-2)
Figure 3.3 Linear regressions of live Callitris sapling densities in gaps (solid lines) and
under canopies (dotted lines) with mean annual rainfall (95% CI). Each data point
shows mean sapling density at each site.

Chapter 3. Competitive feedback between canopy trees and regeneration
55
Table 3.1 Results of the partly nested analyses of variance with orthogonal
polynomials, used to examine the effects of a range of variables on (a) the density of
saplings, (b) sapling height and (c) sapling diameter at breast height (sapling location: c
closest to trunk under canopy; r random in gap; d dominant in gap; i isolated in gap).
Source of variation d.f. SS MS F P
(a) Sapling density
Tree stratum
Site 5 23.50 4.70 3.67 0.006
Linear 1 0.22 0.22 0.18 0.680
Quadratic 1 3.40 3.40 2.65 0.110
Cubic 1 6.48 6.48 5.07 0.030
Deviations 2 13.40 6.70 5.24 0.008
Species (Eucalypt, Callitris) 1 2.03 2.03 1.59 0.210
Site.Species 5 12.85 2.57 2.01 0.090
Residual 52 66.50 1.28 2.50
Tree.Position stratum
Position (gap, canopy) 1 19.16 19.16 37.48 <0.001
Position.Site 5 6.90 1.38 2.70 0.030
Position.Linear 1 3.45 3.45 6.74 0.012
Position.Quadratic 1 0.09 0.09 0.18 0.676
Position.Cubic 1 0.11 0.11 0.21 0.649
Deviations 2 3.26 1.63 3.19 0.050
Position.Species 1 0.05 0.05 0.10 0.756
Position.Site.Species 5 0.92 0.18 0.36 0.872
Residual 52 26.58 0.51
Total 127 158.50
(b) Sapling height
Tree stratum
Site 5 29.72 5.95 4.62 0.001
Linear 1 2.53 2.53 1.96 0.167
Quadratic 1 8.39 8.39 6.52 0.014
Cubic 1 7.36 7.36 5.72 0.020
Deviations 2 11.45 5.73 4.45 0.016
Species (Eucalypt, Callitris) 1 8.52 8.52 6.62 0.013
Site.Species 5 6.29 1.26 0.98 0.440
Residual 52 66.94 1.28 1.57

Chapter 3. Competitive feedback between canopy trees and regeneration
56
Table 3.1 cont.
Source of variation d.f. SS MS F P
(b) Sapling height cont.
Tree.Position stratum
Sapling location (c,r,d,i) 3 314.32 104.77 127.94 <0.001
Sapling location.Site 15 45.41 3.03 3.70 <0.001
Sapling location.Linear 3 2.00 0.67 0.81 0.490
Sapling location.Quad ratic 3 1.89 0.63 0.77 0.510
Sapling location.Cubic 3 24.14 8.05 9.83 <0.001
Deviations 6 17.37 2.89 3.54 0.003
Sapling location.Species 3 7.47 2.49 3.04 0.031
Sapling location.Site.Species 15 21.99 1.47 1.79 0.041
Residual 149 122.02 0.82
Total 248 604.31
(c) Sapling diameter at breast height
Tree stratum
Site 5 5.81 1.16 6.40 <0.001
Linear 1 0.12 0.12 0.67 0.416
Quadratic 1 2.46 2.46 13.58 <0.001
Cubic 1 2.63 2.63 14.51 <0.001
Deviations 2 0.59 0.30 1.63 0.206
Species (Eucalypt, Callitris) 1 0.68 0.68 3.77 0.058
Site.Species 5 1.02 0.20 1.12 0.361
Residual 52 9.43 0.18 1.83
Tree.Position stratum
Sapling location (c,r,d,i) 3 60.67 20.22 203.99 <0.001
Sapling location.Site 15 5.37 0.36 3.610 <0.001
Sapling location.Linear 3 0.65 0.22 2.18 0.093
Sapling location.Quad ratic 3 0.32 0.11 1.09 0.357
Sapling location.Cubic 3 1.36 0.46 4.59 0.004
Deviations 6 3.04 0.51 5.10 <0.001
Sapling location.Site.Species 15 1.53 0.10 1.03 0.430
Residual 149 14.77 0.10
Total 248 97.178

Chapter 3. Competitive feedback between canopy trees and regeneration
57
Pro
po
rtio
n w
ith c
ones
Mean annual rainfall (mm)
0.0
0.2
0.4
0.6
0.8
1.0
400 450 500 550 600
(c) Saplings with cones
Isolated
Dominant
Canopy (both spp.)
Random
0
1
2
3
4
5
6
7(a) Sapling height
0
2
4
6
8
10
12
14
Random Callitris canopy
Eucalyptus canopy
Dominant Isolated
(b) Sapling dbh
Sapling type
Heig
ht (m
)D
BH
(cm
)
Figure 3.4 (a) Height, (b) diameter at breast height (DBH) and (c) fruiting success of
Callitris sapling types: under Callitris or Eucalyptus canopies or random, dominant and
isolated in gaps (mean and standard error).

Chapter 3. Competitive feedback between canopy trees and regeneration
58
Table 3.2 Results of regression analyses of the density of recently dead Callitris
saplings and the combined density of saplings (alive and recently dead) in gaps and
under canopies.
Variable Value SE t P R2
Gap
Constant 0.10 0.07 1.36 0.178 0.22
Sapling density 0.10 0.02 4.15 0.0001
Canopy
Constant 0.01 0.04 0.16 0.87 0.41
Sapling density 0.14 0.02 5.52 <0.001
0.0
0.2
0.4
0.6
0.8
1.0
1.2 (a) Gap
Ln
(density+
1)
of
recently
dead
(no
.m-2
)
Density of live and recently dead (no. m-2)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
0 1 2 3 4 5 6 7
(b) Canopy
Figure 3.5 Linear regressions of densities of recently dead Callitris saplings with
combined sapling density (alive and recently dead) in (a) gaps and under (b) canopies
(95% CI).

Chapter 3. Competitive feedback between canopy trees and regeneration
59
Effects of sapling regeneration on canopy trees along the rainfall gradient
Eucalyptus canopy trees had 3-4 times larger canopy areas, were taller and had larger
DBHs than coexisting Callitris canopy trees (Z = -4.69, P < 0.001; Z = -4.00; P <
0.0001; Z = -4.69, P < 0.0001; Fig. 3.6).
Contrary to predictions recent mortality of Callitris canopy trees was relatively
consistent along the rainfall gradient, but was significantly and negatively associated
with the presence of Callitris sapling regeneration, which was predicted (Z = 2.02, P <
0.04; Fig. 3.7a). For Callitris canopy trees surrounded by regeneration, mortality was
not significantly correlated with mean annual rainfall (Z = -0.27, P < 0.79, r = -0.09),
density of Callitris or Eucalyptus canopy trees (Z = -0.54, P < 0.59, r = -0.21; Z = -1.36,
P < 0.17, r = 0.64), or the mean density of live and recently dead Callitris sapling
regeneration (Z = 1.09, P < 0.28, r = 0.52).
Eucalyptus canopy tree mortality was low and did not respond negatively to the
presence of Callitris sapling regeneration or positively to increasing rainfall, as
predicted by the hypotheses. Only two of the sampled 240 Eucalyptus canopy trees died
during the recent drought and these were not surrounded by Callitris sapling
regeneration. However, Eucalyptus canopy trees suffered leaf stress, defined as > 5%
leaf loss, which mostly ranged from 6 – 20%. The proportion of Eucalyptus canopy
trees with leaf stress was not significantly associated with the presence of Callitris
sapling regeneration (Z = 1.83, P < 0.07). This result is close to significance at P < 0.05
level, indicating some ambiguity. At the lower end of the rainfall gradient, Eucalyptus
surrounded by regeneration are more likely to be stressed than those that are not (Fig.
3.7b). For Eucalyptus canopy trees surrounded by regeneration the proportion of trees
with leaf stress was not significantly correlated with the densities of Callitris or
Eucalyptus canopy trees, the mean density of sapling regeneration or rainfall (Z = 1.23,
P < 0.22, r = 0.58; Z = -0.84, P < 0.40, r = -0.35; Z = -0.78, P < 0.43; r = -0.32; Z = -
1.49, P < 0.14; r = -0.64). These results indicate that at lower mean annual rainfall,
higher leaf stress in Eucalyptus canopy trees may be associated with the presence of
Callitris sapling regeneration.
The proportion of Eucalyptus canopy trees resprouting from the trunk was not
significantly affected by the presence of Callitris sapling regeneration (Z = 0.11, P <
0.91; Fig. 3.7c). Among Eucalyptus canopy trees surrounded by regeneration, the

Chapter 3. Competitive feedback between canopy trees and regeneration
60
proportion resprouting from trunk was significantly and positively correlated with
Callitris canopy tree density (Z = 2.17, P < 0.03, r = 1.00; Fig. 3.7d), but not
Eucalyptus canopy tree density, the mean density of sapling regeneration or rainfall (Z
= 1.02, P < 0.31, r = 0.49, Z = -0.51, P < 0.61, r = 0.26, Z = -0.89, P < 0.37, r = -0.37).
0
50
100
150 (a)
Heig
ht (m
)
0
5
10
15
20
25 (b)
Cano
py a
rea (
m2)
0
20
40
60
80
100
Callitris Eucalyptus
(c)
DB
H (cm
)
Figure 3.6 (a) Canopy area, (b) height and (c) diameter at breast height (DBH) of
Callitris and Eucalyptus canopy trees.

Chapter 3. Competitive feedback between canopy trees and regeneration
61
0
50
100
150
200
250
400 450 500 550 600
(d) Density of Callitris & Eucalyptus
Callitris (no regeneration)
Callitris (regeneration)
Eucalyptus (regeneration)
Eucalyptus (no regeneration)
0.0
0.2
0.4
0.6
0.8
1.0 (c) Eucalyptus resprouting f rom trunk
0.0
0.2
0.4
0.6
0.8
1.0(b) Eucalyptus with leaf stress
Eucalyptus (regeneration)
Eucalyptus (no regeneration)
Mean annual rainfall (mm)
0.0
0.2
0.4
0.6
0.8
1.0(a) Callitris
Callitris (regeneration)
Callitris (no regeneration)P
rop
ort
ion d
ead
Pro
po
rtio
nD
en
sity
(no
. ha
-1)
Figure 3.7 Trends in mortality and stress levels of canopy trees with and without
Callitris sapling regeneration along the rainfall gradient: (a) Callitris mortality; (b)
Eucalyptus with leaf stress (>5% leaf loss); (c) Eucalyptus resprouting from the trunk;
and (d) the density of Callitris and Eucalyptus canopy trees.

Chapter 3. Competitive feedback between canopy trees and regeneration
62
Discussion
The results indicate feedback between Callitris saplings and Eucalyptus and Callitris
canopy tree dynamics, which varied along the rainfall gradient. Saplings were less
dense, smaller and less likely to be reproducing under canopies than in the gaps, with
canopies reducing sapling density more at the higher end of the rainfall gradient.
Drought preferentially thinned saplings at higher density. In turn, the combined effects
of drought and sapling regeneration reduced the resilience of canopy trees. Callitris
canopy trees surrounded by dense sapling regeneration suffered higher mortality than
similarly placed Eucalyptus canopy trees, which suffered greater levels of stress at the
lower end of the rainfall gradient. These trends highlight how interactions among
climate, canopies and regeneration influence stand dynamics in this semi-arid system.
Canopy effects on Callitris sapling density along the rainfall gradient
As hypothesised, densities of Callitris saplings were lower under canopies than in gaps,
but the magnitude of this difference was relatively small and changed along the rainfall
gradient. Difference in sapling densities in gaps and under canopies were greater at
higher than lower rainfall. This implies increasing canopy competition as rainfall
increases. At high rainfall, competition for resources under canopies may have lowered
sapling survival more than in adjacent gaps (Fuentes et al. 1984; Belsky 1994). By
comparison, at the low end of the rainfall gradient, competitive effects of trees may
have been reduced by positive effects of canopies, such as increased humidity and
nutrients or reduced temperatures, wind and herbivory (Barnes & Archer 1999;
Pugnaire & Luque 2001; Bruno et al. 2003; Arrieta & Suarrez 2005). If canopies did
improve conditions for Callitris saplings at low rainfall, it was not enough to sustain
higher densities than in adjacent gaps. Thus the net effect of canopy trees on
regeneration was competitive rather than facultative.
Canopy effects on Callitris sapling growth and reproduction along the rainfall
gradient
As predicted growth rates of Callitris saplings were slower under canopies than in the
gaps, where there was also a greater probability of fruiting . Saplings under canopies
were short and without fruits, as were the majority of saplings surrounded by dense
sapling regeneration in gaps. Slow growth rates and negligible seed production of
similarly aged Callitris saplings in dense stands, have previously been recorded (Lacey

Chapter 3. Competitive feedback between canopy trees and regeneration
63
1972; Hawkins 1966; FCNSW 1988). By comparison, growth rates and reproductive
success were significantly greater for the less frequent but taller or isolated saplings in
gaps. These growth rate differences suggest a longer period of vulnerability for Callitris
saplings under canopies than in gaps. Slow development, leaves non-resprouters, like
Callitris, particularly vulnerable to disturbances like fire, since their survival is
dependent on seeds or thick bark which increases with plant size (Bowman et al. 1988;
Bond 1989; Bowman & Harris 1995). If fire penetrates the stand, the higher
flammability of litter under Eucalyptus compared with Callitris canopy trees (Scarff &
Westoby 2006) heightens this vulnerability (Clayton-Greene 1981).
Density, growth and reproduction of Callitris saplings under Callitris and Eucalyptus
canopies
Contrary to predictions, densities of Callitris saplings were similar beneath Eucalyptus
and Callitris canopies. This trend suggests comparable levels of competition by the
different canopy species on Callitris saplings. Indeed other studies have found that
during drought Eucalyptus and Callitris trees use similar amounts of water (Zeppel &
Eamus 2004).
In contrast and as hypothesised, growth rates of Callitris saplings under Eucalyptus
canopy trees were faster than those under Callitris canopy trees. This may be related to
higher nutrient litter under Eucalyptus because of higher leaf turnover or greater light
infiltration through the more open Eucalyptus canopies (Clayton-Greene 1981; Bowman
& Wilson 1988). Other studies have shown that nutrient levels and light availability can
influence the performance plants in semi-arid systems (Holmgren et al. 1997; Armas &
Pugnaire 2005). Further research is needed to determine the mechanisms underlying
trends in the density and growth of Callitris saplings under Eucalyptus and Callitris
canopy trees. Reproductive success of Callitris saplings under canopy trees was so low,
that comparison between Eucalyptus and Callitris canopy trees was not made.
Callitris sapling mortality
Mortality of Callitris saplings was related to sapling density during the drought. As
hypothesised, the density of recently dead Callitris saplings increased as the density of
saplings increased, resulting in 10 to 30% mortality. Density dependent mortality of
woody species, including Callitris has previously been observed during droughts (Lacey
1973; FCNSW 1988; Belsky & Blumenthal 1997). Drought is especially important as a

Chapter 3. Competitive feedback between canopy trees and regeneration
64
‘thinning agent’ for species with slow rates of self-thinning, like Callitris (Lacey 1972,
1973; FCNSW 1988), when other disturbances, like fire are absent (Lacey 1973;
Bowman et al. 1988; Fule & Covington 1998).
Effects of sapling regeneration on canopy trees along the rainfall gradient
Despite their ability to similarly reduce water usage during periods of water deficit
(Zeppel & Eamus 2005), Callitris canopy trees suffered higher mortality than
Eucalyptus canopy trees during the drought. As hypothesised, the mortality of Callitris
canopy trees not surrounded by sapling regeneration was lower (0 - 10%), than those
surrounded by sapling regeneration (0 - 25%). Dense Callitris regeneration has
promoted the death of both Callitris and Eucalyptus canopy trees in previous droughts,
but has not been quantified (Lacey 1973; FCNSW 1988; Allen 1998). By comparison,
the mortality of Eucalyptus canopy trees was low (<2%), irrespective of sapling
regeneration presence. In previous droughts, higher levels of mortality of a range
Eucalyptus species have been recorded in northern Australian savannas (Fensham
1998b; Fensham & Holman 1999; Fensham et al. 2005; Fensham et al. 2009).
Although Callitris canopy tree mortality was not greater at lower rainfall, as
hypothesised, a greater proportion of Eucalyptus trees showed signs of stress at lower
rainfall. These stressed Eucalyptus trees had greater leaf loss than and were resprouting
from the trunk. These stress indicators were associated with the presence of dense
sapling regeneration and higher Callitris canopy density, respectively. These Eucalyptus
canopy trees may be predisposed to mortality during future droughts, if their resprouting
ability becomes compromised (Attiwill & Clayton-Greene 1984). Indeed, it is possible
that this ability contributed to the higher survival of Eucalyptus than Callitris canopy
trees during this latest drought.
The effects of predicted changes in climate
Whilst Callitris sapling regeneration was common, Eucalyptus sapling regeneration was
infrequent in this semi-arid system. Eucalyptus species most commonly regenerate after
fires (Gill 1997) and since European management, fire frequency and intensity have
declined, often to the extent of exclusion (Noble 1997). Once dense patches of Callitris
become established, their low flammability can maintain this fire regime in low-
moderate, but not in extreme fire weather conditions (see Chapter 4). Despite predicted
increases in extreme fire-weather days (Lucas et al. 2007), fire frequency is unlikely to

Chapter 3. Competitive feedback between canopy trees and regeneration
65
increase because of predicted declines in herbaceous fuels, which limit fires in this
region, together with high levels of landscape fragmentation (Bradstock 2010).
In contrast, the predicted drought regime (Nicholls 2004; CSIRO & BOM 2007;
Hennessy et al. 2008), is likely to accentuate Callitris dominance, if the current fire
regime is maintained. As droughts increase in frequency and intensity, mortality rates of
Callitris sapling regeneration may increase. As gaps appear they are more likely to be
filled by Callitris than Eucalyptus seedlings, since Eucalyptus seedlings do not compete
well with dense Callitris regeneration (FCNSW 1988). Thus Callitris and Eucalyptus
sapling regeneration may be maintained at their current levels of dominance and
infrequency, respectively. As ageing canopy trees die, Eucalyptus are unlikely to be
replaced by conspecifics because of their infrequency. Consequently, the system may
shift further toward Callitris dominance, especially in the west where Eucalyptus
canopy trees were most stressed during this latest drought.
Conclusion
Interactions among climate, sapling regeneration and canopy trees influenced dynamics
in this semi-arid system. Feedback between sapling regeneration and canopy trees was
largely negative. Canopy trees moderated rather than excluded dense sapling
regeneration, allowing Callitris to persist at relatively high densities, 50 years after their
establishment. In turn, dense Callitris sapling regeneration increased mortality rates or
stress levels in canopy trees during drought. As drought severity increases and fires
continue to be excluded from these semi-arid systems, the fire-sensitive Callitris is
likely to maintain and perhaps increase its dominance.
Acknowledgments
An Australian Research Council (ARC) Linkage Grant between Charles Sturt
University (CSU) and NSW Department of Environment and Climate Change and
Water (DECCW NSW) provided the funding for this research (Project number
LP066829). I would like to thank Stephen Campbell and Alan Smith from State Forests
NSW for their assistance and Livia Tigwell, Andrew Cohn and John Warren for
assistance in the field. Terry Koen (DECCW NSW) and Simon McDonald (CSU)
assisted with statistical advice.

Chapter 3. Competitive feedback between canopy trees and regeneration
66

Chapter 4. Feedback between Callitris patches and fire intensity
67
Chapter 4
Feedback between patches of Callitris glaucophylla and fire intensity
influences Callitris survival in flammable Eucalyptus woodlands in
different fire-weather conditions
Resprouting Eucalyptus species following fire in the Eucalyptus matrix in extreme fire-weather conditions (top). A Callitris patch following fire in low to moderate fire-weather conditions (bottom). in Pilliga Conservation Area, north eastern NSW (Source: K. Ross, J.Cohn)

Chapter 4. Feedback between Callitris patches and fire intensity
68
Chapter 4
Feedback between patches of Callitris glaucophylla and fire intensity
influences Callitris survival in flammable Eucalyptus woodlands in
different fire-weather conditions
Abstract
I examined how interactions between weather and vegetation influence spatial
heterogeneity in fire intensity, permitting the survival of fire-sensitive Callitris
glaucophylla within a matrix of highly flammable Eucalyptus species. The study was
undertaken in Pilliga State Forest, south eastern Australia. In areas burned under
extreme and low-moderate fire-weather conditions, fire intensity and survival of
Callitris were measured in the flammable Eucalyptus matrix and in Callitris dominated
patches. Pre-fire fuel characteristics were measured in an adjacent unburnt area.
Flammable fuel levels were lower in Callitris patches than in the matrix. In extreme
fire-weather conditions, Callitris patches did not reduce incoming fire intensity from the
matrix, since weather controlled fire intensity. However in low-moderate, fire-weather
conditions, Callitris patches reduced incoming fire intensity from the matrix, the
magnitude of which increased with increasing Callitris basal area. Survival of Callitris
was 4% and 54% in extreme and low-moderate fire-weather conditions, respectively.
Callitris survival was negatively related to char height and positively related to trunk
diameter. The relative importance of fuels and weather in determining fire intensity
varied, influencing Callitris survival. In low-moderate, fire-weather conditions, Callitris
survival was higher in patches dominated by Callitris, where flammable fuels and fire
intensities were lower than in the matrix. In extreme fire-weather conditions,
flammability differences were less important than weather, resulting in high fire
intensities in Callitris patches and very low Callitris survival. Thus, under low-
moderate, fire-weather conditions, patch structure created by the less flammable
obligate-seeding Callitris reduced fire intensity, enabling it to persist in a flammable
Eucalyptus matrix.

Chapter 4. Feedback between Callitris patches and fire intensity
69
Introduction
In many plant communities, disturbance prevents competitive exclusion and promotes
species coexistence, as described by non-equilibrium theories of diversity (Bond & van
Wilgen 1996; Huston 2003). Coexistence of species of varied resilience and competitive
status may depend upon heterogeneity in disturbances, such as fire regimes (frequency,
intensity and season), across differing spatial and temporal scales (Cowling 1987;
Whelan 1995; Williams et al. 1994).
A fundamental question in fire ecology is – how do long-lived, obligate-seeders persist
in flammable landscapes (Bond & van Wilgen 1996)? Adaptations, such as, serotinous
cones and thick bark are major determinants of persistence (Agee 1998; Keeley &
Zedler 1998). Bark thickness is important for species that are weakly serotinous and
varies among species and increases with tree age (Bond & van Wilgen 1996; Keeley &
Zedler 1998). Interactions with fire intensity determine survival, with thicker barked
individuals surviving higher temperatures (Peterson & Arbaugh 1986).
Spatial variation in fire intensity is influenced by interactions among topography,
weather and fuels (Catchpole 2002; Boer et al. 2008). The relative importance of
weather and fuels has been debated but appears to vary greatly among ecosystems
(Bessie & Johnson 1995; Hely et al. 2001; Turner et al. 2003). In some ecosystems,
variations in fuel levels influence fire intensity under moderate weather conditions, but
fuels have little influence on fire intensity under extreme weather conditions (i.e. high
temperature and wind speed, and low humidity and soil moisture; Bessie & Johnson
1995; Turner et al. 2003). By contrast, in other ecosystems, variations in fuel levels
influence fire intensity across the entire range of weather conditions, including extreme
conditions (Hely et al. 2001).
Flammability varies greatly among vegetation types due to trait differences among
component species (Williamson & Black 1981; van Wilgen et al. 1990; Schwilk &
Ackerly 2001). Flammability is determined by a range of characteristics including fuel
type, biomass and spatial continuity. Low flammability may result from shedding of
lower branches and understorey suppression, both of which reduce the vertical
continuity of fuels, resulting in less damage to the canopy (Schwilk 2003; Knapp &
Keeley 2006). Leaf litter also affects stand flammability. Litter with a high surface-area
to volume ratio (e.g. needles) can pack more tightly, reducing aeration and increasing

Chapter 4. Feedback between Callitris patches and fire intensity
70
moisture levels, thereby reducing flammability (Schwilk & Ackerly 2001; Scarff &
Westoby 2006). Patches of species with less flammable traits may therefore promote
their own survival and coexistence among competing flammable species by influencing
the spatial heterogeneity of fire intensity (Cowling 1987; van Wilgen et al. 1990;
Bowman 1998).
Whilst studies have examined how fire intensity affects the survival of long-lived, non-
serotinous, obligate-seeders (e.g. Rodrigo et al. 2004 2007), few have attempted to
examine survival in relation to the spatial variation of fuels in varying fire-weather
conditions (Bowman & Wilson 1988; Bowman & Panton 1993). No study appears to
have quantified interactions amongst flammability, fire intensity and obligate-seeder
survival at the patch scale under varying fire-weather conditions.
In Australian woodlands, flammable and resprouting Eucalyptus species often coexist
with less flammable, non-serotinous, obligate-seeding conifers of the genus Callitris
(Bowman & Harris 1995). Mixed forests dominated by these two genera provide an
ideal model system to examine two key questions that underlie the survival of long-
lived, obligate-seeders in flammable landscapes: (1) how do weather and vegetation
(fuel) patterns interact to influence spatial heterogeneity in fire intensity; and (2) to what
extent do these interactions permit the survival of patches of fire-sensitive species
(obligate-seeders) within a matrix dominated by highly flammable, fire-tolerant species
(resprouters)?
In this study I used this model system to investigate these two issues, by examining the
following hypotheses: (1) under extreme fire-weather conditions, patches of Callitris
will have little influence on the incoming fire intensity from the Eucalyptus matrix; (2)
under low-moderate fire-weather conditions, patches of Callitris of high basal area will
reduce the incoming fire intensity from the Eucalyptus matrix; (3) survival of Callitris
will be negatively related to fire intensity and positively related to tree size; and (4)
ground fuels will be of lower flammability in patches of Callitris than in the Eucalyptus
matrix, thereby reducing fire intensity in patches of Callitris.

Chapter 4. Feedback between Callitris patches and fire intensity
71
Methods
Study area
The study was undertaken in the Pilliga Conservation Reserve and State Forest in north-
eastern New South Wales, south-eastern Australia. ‘The Pilliga’ holds the most
extensive stands of Callitris glaucophylla in Australia (270 000 ha; Thompson &
Eldridge 2005). The area experiences a quasi Mediterranean climate of warm, dry
summers and mild, moist winters (Gill & Moore 1990). The dominant trees include
non-serotinous, obligate-seeding C. glaucophylla J. Thomps. & L. Johnson and C.
endlicheri (Parl.) Bailey, and resprouting Eucalyptus crebra F. Muell., E. fibrosa
F.Muell., E. nubila Maiden & Blakely, and E. trachyphloia (F. Muell.) K.D. Hill &
L.A.S. Johnson (Binns & Beckers 2001). Dense patches of C. glaucophylla and C.
endlicheri, typically 0.01-0.25 km2 in size (hereafter ‘Callitris patches’) are scattered
throughout a more open matrix dominated by Eucalyptus with sparse Callitris (hereafter
the ‘Eucalyptus matrix’). Topography is flat to gently undulating.
On 29th
November 2006, during a lengthy drought (Australian Bureau of Meteorology
unpubl.), lightning ignited a number of fires in the Pilliga, one of which is the focus of
this study. Approximately 70 000 ha burnt in the first 24 hours in extreme fire-weather
conditions, and over the following 2 weeks, a further 27 000 ha burnt in low-moderate
fire-weather conditions (DECC NSW unpubl.). Fire-weather conditions are defined by
the Forest Fire Danger Index (FFDI), which is based on a drought factor and ambient air
temperature, wind speed and relative humidity (Noble et al. 1980). During extreme
conditions, the maximum temperature and wind speed reached 37oC and 57 km hr
-1, and
the minimum humidity was 12%. In the low-moderate fire-weather conditions
temperatures and wind speeds were generally lower and relative humidity higher and all
were more variable (Australian Bureau of Meteorology unpubl.).
Sampling was undertaken within the burnt area and data on pre-fire fuel characteristics
were collected from an adjacent unburnt area. Within the burnt area there were two fire-
weather zones, extreme and low-moderate. Sampling locations were determined from
1:50 000 scale aerial photos taken in 2003. Callitris patches were at least 1 ha and
dominated by C. glaucophylla and/or C. endlicheri, which have comparable architecture
and leaf characteristics inferring similar flammabilities (Harden 2000; Scarff &
Westoby 2006). Sampling occurred between September and November 2007 and was
restricted to areas of < 5o slope.

Chapter 4. Feedback between Callitris patches and fire intensity
72
Fire intensity indicators and Callitris survival
Paired quadrats were placed in the direction of the fire, so that the incoming quadrat was
burnt before the outgoing quadrat (Fig. 4.1). Quadrat pairs were located across four
vegetation transitions: in the Eucalyptus matrix (both quadrats placed in the matrix);
across the Eucalyptus matrix – Callitris patch boundary (incoming quadrat in matrix
and outgoing in Callitris patch); in the centre of Callitris patches (both in Callitris
patch); and across the Callitris patch – Eucalyptus matrix boundary (incoming in
Callitris patch and outgoing in adjacent Eucalyptus matrix). Each transition pair was
replicated ten times in each fire-weather zone, giving 40 pairs in the extreme fire-
weather zone and 40 pairs in the low-moderate fire-weather zone. To minimise spatial
auto correlative effects, all paired locations were separated by at least 100 m.
Fire
E/E Ee/Ce
C/C
> 100 m
Eucalyptus matrix
Ce/Ee
Callitris patchin out
in outin out
in out
Fig. 4.1 Diagrammatic representation of the sampling design within the burned area.
Data were collected in quadrat pairs located across four vegetation transitions: in
Eucalyptus matrix (E/E); across Eucalyptus matrix – Callitris patch boundary (Ee/Ce;
‘e’ indicates an edge quadrat); in Callitris patches (C/C); and across the Callitris patch –
Eucalyptus matrix boundary (Ce/Ee). Paired quadrats were placed in the direction of the
fire, so that the incoming (in) quadrat was burnt before the outgoing (out) quadrat and
all pairs were separated by at least 100 m.
Fire direction was determined by a combination of means: (1) reports on prevailing
wind direction; (2) char heights were lower on the windward side of trees than the more
protected leeward side; and (3) shrubs often bent and remained fixed in the direction of
the fire.
Char height and post-fire stem tip diameter were measured as indicators of fire intensity
(Cain 1984; Moreno & Oechel 1989). Char height was measured as the maximum
height of charring on the bole (Sah et al. 2006), which provides a relative measure of

Chapter 4. Feedback between Callitris patches and fire intensity
73
fire intensity (Cain 1984). Leaf scorch height, a direct measure of fire intensity
(Williams et al. 2003), was not used because the substantial distance to the first leaves
on many Eucalyptus would have resulted in imprecise measures of intensity in low-
moderate weather conditions. The ability of some bark types (e.g. stringybarks) to carry
flames more easily than others (McCarthy et al. 1999), and their interaction with wind
speed (Cheney 1981) was addressed by sampling Eucalyptus species with a range of
bark types at each site. Stem tip diameter is a measure of the minimum diameter of
branches remaining after the fire, with more intense fires leaving larger diameter
branches (Moreno & Oechel 1989). It is correlated with maximum surface temperatures
during a fire. Canopy scorch and survival were recorded as measures of fire severity
(Bond & van Wilgen 1996; Turner et al. 1999).
Char height, percent canopy scorch, survival and sizes of individuals (height and
diameter at breast height at 1.3 m, DBH) were measured on targeted size classes of
unconsumed Callitris and the Eucalyptus in the centre of each 10 m x 10 m quadrat
(two trees of each): small Callitris (3-15 cm DBH), large Callitris and Eucalyptus (> 15
cm DBH). Small Callitris established after 1952, and large Callitris originated from
1890 to 1911 (Austin & Williams 1988; Norris et al. 1991; Whipp 2009). Targeted size
classes were chosen to provide a consistent index of fire intensity. Stem tip diameters
were measured on Callitris with DBHs of 1-4 cm. Stem tip diameters were measured on
branches of at least 1 cm length and approximately 1.3 m from the ground. Five
branches were sampled on each of five Callitris in each quadrat. To determine basal
area and density of Callitris and Eucalyptus in each quadrat, the number of all trees in
each DBH size class was counted (0-5, 5.1-10, 10.1-20, 20.1-30, 30.1-40, 40.1-50, 50.1-
60, 60.1-70 cm). Sparsely distributed larger trees (> 20 cm DBH) were measured in a
larger quadrat (15 m x 15 m) centred on the main inner quadrat (10 m x 10 m).
Fuel characteristics
Ground fuel characteristics were sampled in an unburnt zone adjacent to the burnt zone,
with four quadrats one in each vegetation type (Eucalyptus matrix, Eucalyptus matrix
edge, Callitris patch edge, Callitris patch). To achieve comparable fuel characteristics,
unburned vegetation types were chosen in the same manner as the burned vegetation
types and fire history records for the unburned and the burned zones were similar.
Sampling occurred in November 2007 to approximate grass and herb conditions at the
time of the fire.

Chapter 4. Feedback between Callitris patches and fire intensity
74
The following fuel characteristics were measured in four 1 m x 1 m quadrats, positioned
2 m inside each corner of a 10 m x 10 m quadrat: percent cover of bare ground, leaf
litter of Callitris and Eucalyptus, grass, herbs and shrubs; the number of all shrubs and
shrubs > 1 m tall; and the average height of shrubs, grasses and herbs. Litter depths
were measured in each quadrat and at six other fixed locations within each 10 m x 10 m
quadrat. Tree density was measured in DBH size classes (see above) to estimate
Callitris and Eucalyptus basal area and density. Ground fuel was collected in a 0.25 m x
0.25 m plot in the centre of each 1m x 1m quadrat. Fuel was sorted into total fine fuel
(litter, twigs < 6 mm diameter, bark), grasses, herbs, and twigs 6-20 mm diameter.
Samples were dried for 24 hrs at 70oC and weighed.
Statistical analyses
Basal areas of Callitris and Eucalyptus
Two factor Analyses of Variance (ANOVA) were used to examine whether Callitris
and Eucalyptus basal areas varied between the four vegetation types (Eucalyptus matrix,
Eucalyptus matrix edge, Callitris patch edge, Callitris patch) and two fire-weather
zones (low-moderate, extreme). Only data from the incoming quadrats were used, since
data from the outgoing quadrats were not independent. Basal area data were
transformed to correct for skewing and heterogeneity of variance. Tukey tests were used
for post-hoc pairwise comparisons (Quinn & Keough 2002).
To assess whether Callitris and Eucalyptus basal areas in each vegetation type in the
unburnt area were representative of the burnt area, Kruskal-Wallis tests and associated
post-hoc tests were used.
Fire intensity indicators
Linear regressions were used to examine the relationship between fire intensity
indicators in the incoming quadrat (‘Intensity Indicator In’) and the abundance of
Callitris and Eucalyptus in the outgoing quadrat, and fire intensity indicators in the
outgoing quadrat (‘Intensity Indicator Out’) within each fire-weather zone (low-
moderate, extreme) for all paired quadrats:
Intensity Indicator Out ~ Intensity Indicator In – Callitris Abundance Out + Eucalyptus
Abundance Out

Chapter 4. Feedback between Callitris patches and fire intensity
75
Intensity indicators included Eucalyptus char height in the low-moderate fire-weather
zone and Callitris stem tip diameter in the extreme fire-weather zone. In both fire-
weather zones, measures of Callitris and Eucalyptus abundances included basal area
and density. Pearson’s correlation coefficient was used to check for collinearity between
predictor variables. Akaike Information Criterion (AIC) was used to compare models,
the best of which had the smallest AIC (Quinn & Keough 2002).
A Kruskal-Wallis test followed by post-hoc tests were used to compare the change in
magnitude of fire intensity indicators from the incoming to the outgoing quadrat for
each transition pair within each fire-weather zone.
Callitris survival
Logistic regressions (logit-link) were used to examine the survival of Callitris in
relation to char height on Callitris and Callitris size (DBH and height) in the low-
moderate fire-weather zone only (incoming quadrats), since most Callitris died in the
extreme fire-weather zone:
Callitris Survival ~ Char Height on Callitris * Callitris Size
Correlation tests were used to determine significant collinearity between predictor
variables, which precluded them from testing within the same model. A pseudo R2 was
calculated for each model (R2
= 1- (residual deviance/null deviance)) and AIC was
used to compare models (Veall & Zimmerman 1996). Kruskal-Wallis tests followed by
post-hoc tests were used to compare the proportion of Callitris which survived amongst
vegetation types. Only data from the incoming quadrats were used, since data from the
outgoing quadrats were not independent.
Fuel characteristics
Kruskal-Wallis tests followed by post-hoc tests were used to examine the effects of
vegetation type on ground fuel characteristics, including shrub cover, shrub density,
Eucalyptus and Callitris leaf litter cover, litter depth, bare ground cover, shrub height,
fine fuel weight and total fuel weight (fine fuel weight, grass, herbs, twigs 6-20 mm
diameter). Given their sparseness, data on grass and herb cover were not analysed.

Chapter 4. Feedback between Callitris patches and fire intensity
76
Results
Basal areas of Callitris and Eucalyptus
Callitris and Eucalyptus basal areas varied significantly between vegetation types, but
not between fire-weather zones. In the burnt area, Callitris basal area was significantly
greater in the Callitris patches and Callitris patch edges than in the Eucalyptus matrix
and Eucalyptus matrix edges, indicating a sharp boundary between the Eucalyptus
matrix and Callitris patches (F3, 71 = 25.33, P < 0.001; Fig. 4.2). Eucalyptus basal area
was significantly greater in the Eucalyptus matrix and Eucalyptus matrix edge than the
Callitris patch edge but not the Callitris patches (F3, 71 = 5.71, P = 0.0015), indicating a
relatively even distribution of Eucalyptus basal area. Callitris had similar mean basal
area in Callitris patches as did Eucalyptus in the Eucalyptus matrix (0.11-0.14 m2 100
m-2
).
Basal areas of Callitris and Eucalyptus were generally comparable between the burnt
and unburnt areas (where fuels were sampled). Only the basal area of Callitris
(incoming quadrats) was significantly greater in the unburnt than the burnt area in
Eucalyptus matrix edges and Callitris patches (X2
1 = 7.32, P = 0.0068, X2
1 = 5.0, P =
0.019; Fig. 4.2).
0
10
20
30
40
E b
E u
b
Ee b
Ee u
b
Ce b
Ce u
b
C b
C u
b
Eucalyptus
Callitris
Vegetation type (burned , unburned)
Basal a
rea (
m2
ha
-1)
Fig. 4.2 Basal areas of Callitris and Eucalyptus in vegetation types in the burned area
(b) and in the unburned area (ub) where fuel data were collected (E = Eucalyptus
matrix, Ee = Eucalyptus matrix edge, Ce = Callitris patch edge, C = Callitris patch).
Means and standard errors are given.

Chapter 4. Feedback between Callitris patches and fire intensity
77
Fire intensity indicators
Char heights on Eucalyptus in the low-moderate fire-weather zone (0-8.5 m; Fig. 4.3a)
corresponded with Cheney’s (1981) low-moderate fire intensities (1.5-6 m, < 500-3000
kW m-1
). In the extreme fire-weather zone, most char heights (7-16 m; Fig. 4.3b)
corresponded with Cheney’s (1981) moderate to very high fire intensities (6-16 m,
3000-70000 kW m-1
). The ‘moderate’ values were an underestimation, since char
heights were limited by Eucalyptus heights. For this reason stem tip diameters were
used as indicators of fire intensity for analyses within the extreme fire-weather zone,
while char heights were used for analyses within the low-moderate fire-weather zone.
In the low-moderate fire-weather zone, average char heights on Callitris were lower
than those on adjacent Eucalyptus, especially below 3 m char heights on the Eucalyptus
(Fig. 4.3a). Stem tip diameters on Callitris mostly ranged from 2-4.5 mm in the extreme
fire-weather zone (Fig. 4.3b) compared to 0.5-2 mm in the low-moderate fire-weather
zone.

Chapter 4. Feedback between Callitris patches and fire intensity
78
0
2
4
6
0 5 10 15 20
Char height on Eucalyptus (m)
Char
heig
ht
on C
alli
tris
(m
)
0
2
4
6
0 5 10 15 20
Ste
m tip
dia
mete
r o
n C
alli
tris
(m
m)
(a) Low-moderate f ire weather zone
(b) Extreme f ire weather zone
Fig. 4.3 Comparisons of fire intensity indicators: (a) maximum char heights on the
trunks of Callitris (> 15 cm DBH) versus maximum char heights on the trunks of
Eucalyptus (> 15 cm DBH) in the low-moderate fire-weather zone and; (b) stem tip
diameter on Callitris (1-4 cm DBH) versus char heights on Eucalyptus (> 15 cm DBH)
in the extreme fire-weather zone. Data are means for each quadrat.

Chapter 4. Feedback between Callitris patches and fire intensity
79
There was only one significant correlation between a fire intensity indicator and
Callitris and Eucalyptus abundance measures used in the analyses of intensity and
Callitris survival. In the low-moderate fire-weather zone, the incoming char height was
negatively correlated with Callitris basal area in the outgoing quadrat (r = -0.34, t34 = -
2.12, P = 0.04). Since the correlation strength was relatively low, the variables were not
excluded from the analyses.
In the extreme fire-weather zone, linear regressions indicated that stem tip diameter in
the outgoing quadrat was linearly and positively related to stem tip diameter in the
incoming quadrat (Table 4.1, Fig. 4.4), indicating that variation in fuels had no
significant effect on fire intensity. Outgoing stem tip diameter was not significantly
affected by Callitris or Eucalyptus basal area or density in the outgoing quadrat.
Table 4.1 Models of fire intensity indicators in the outgoing quadrat as a function of
intensity indicators in the incoming quadrat and abundances of Callitris and Eucalyptus
in the outgoing quadrat for each fire-weather zone. Intensity Indicator Out ~ B0 + B1
Intensity Indicator In - B2 Callitris Abundance Out + B3 Eucalyptus Abundance Out.
Models are ordered from best to worst according to AIC.
Intensity Abundance Coefficients R2
AIC
indicator B0 B1 B2 B3
(a) Extreme fire-weather zone
Stem tip diameter Basal area 0.76 0.78 0.47 48
Stem tip diameter Density 0.76 0.78 0.47 48
(b) Low-moderate fire-weather zone
Char height Basal area 2.02 0.69 11.00 0.64 131
Char height Density 0.97 0.76 0.59 135
By contrast, in the low-moderate fire-weather zone, the better model indicated by AIC,
showed that char height in the outgoing quadrat was positively related to char height in
the incoming quadrat and negatively related to Callitris basal area in the outgoing
quadrat (Table 4.1). Thus char height decreased as Callitris basal area increased (Fig.
4.5). For example, an incoming char height of 5 m at the lowest Callitris basal area (0
m2 100 m
-2) was reduced to 3 m in the outgoing quadrat at the highest Callitris basal
area (0.2 m2
100 m-2
). In the second model, outgoing char height was positively related
to incoming char height but not significantly related to Callitris density. In neither
model did Eucalyptus abundance in the outgoing quadrat have a significant effect on the
outgoing char height.

Chapter 4. Feedback between Callitris patches and fire intensity
80
0
1
2
3
4
5
0 1 2 3 4 5
Stem tip diameter In (mm)
Ste
m tip
dia
mete
r O
ut
(mm
)
Fig. 4.4 Stem tip diameter in the outgoing quadrat as a function of stem tip diameter in
the incoming quadrat in the extreme fire-weather zone. Stem Tip Diameter Out ~ 0.76 +
0.78 Stem Tip Diameter In (95% confidence intervals).
Char
heig
ht
Out
(mm
)
0 2
4
6
8 1
0
Fig. 4.5. Char height in the outgoing quadrat as a function of char height in the
incoming quadrat and Callitris basal area (BA) in the outgoing quadrat in the low-
moderate fire-weather zone. Char height was measured on the Eucalyptus. Char Height
Out ~ 2.02 + 0.69 Char Height In – 11.0 Callitris Basal Area Out.

Chapter 4. Feedback between Callitris patches and fire intensity
81
The Kruskal-Wallis test and post hoc tests indicated a significant decline in char height
from the Eucalyptus matrix edge – Callitris patch edge transition (Table 4.2), coinciding
with an increase in Callitris basal area (Fig. 4.2). There were no other significant
changes in char height between vegetation transitions, despite a significant decline in
Callitris basal area across the Callitris patch edge – Eucalyptus matrix edge transition
(Fig. 4.2). In the extreme fire-weather zone there was no significant change in stem tip
diameter across any vegetation transition (Table 4.2).
Table 4.2 Results of the Kruskal-Wallis tests showing the median change in intensity
indicators from the incoming to the outgoing quadrats across four vegetation transitions
in two fire-weather zones: in Eucalyptus matrix (E/E); across Eucalyptus matrix –
Callitris patch boundary (Ee/Ce); in Callitris patches (C/C); and across the Callitris
patch – Eucalyptus matrix boundary (Ce/Ee). The median (Med.) change and mean
intensity indicator (standard error) for each incoming and the outgoing quadrat are also
given.
Vegetation Low-moderate fire-weather Extreme fire-weather
transition Char height (m) Stem tip diameter (mm)
Med. Mean (se) Med. Mean (se)
change change
E/E 0.15 5.3 (0.8)/5.6 (0.6) 0.23 3.2 (0.3)/3.0 (0.4)
Ee/Ce -1.65 4.4 (0.5)/2.5 (0.4) 0.18 3.0 (0.3)/3.1 (0.2)
C/C 0.40 2.0 (0.8)/2.8 (0.8) 0.11 2.5 (0.3)/2.6 (0.4)
Ce/Ee 0.75 3.8 (1.0)/5.0 (1.0) 0.15 3.0 (0.2)/3.1 (0.3)
X2 15.3 0.9
P 0.002 0.9
Callitris survival
In the extreme fire-weather zone, 4% (8/202) of Callitris individuals survived, all of
which were within one Callitris patch, near the edge of the extreme fire-weather zone.
In the low-moderate fire-weather zone, 54% (133/234) of Callitris individuals survived.
Here, Callitris survival was significantly greater in Callitris patches and Callitris patch
edges than in the Eucalyptus matrix and Eucalyptus matrix edges (X2
3 = 25.4, P < 0.001;
Fig. 4.6).
Callitris survival was best related to percent canopy scorch. Since 99% (239/241) of
Callitris individuals were killed by 100% canopy scorch, modelling was unnecessary.
This threshold in Callitris survival was reached in the low-moderate fire-weather zone.

Chapter 4. Feedback between Callitris patches and fire intensity
82
In this zone, logistic regression modelling indicated that Callitris survival was
negatively related to char height and positively related to Callitris DBH and height
(Table 4.3, Fig. 4.7). Callitris DBH was a slightly better predictor of Callitris survival
than height, as indicated by AIC values and by the higher proportion of correctly
classified cases when fitted values were compared to actual values (Table 4.4). Most
Callitris died when they experienced char heights exceeding 1 m, with survival
restricted to trees > 25 cm DBH
0.0
0.2
0.4
0.6
0.8
1.0
E Ee Ce C
Vegetation type
Surv
ival (
pro
po
rtio
n)
Fig. 4.6. Proportional survival of the Callitris in each vegetation type in the low-
moderate fire-weather zone (E = Eucalyptus matrix, Ee = Eucalyptus matrix edge, Ce =
Callitris patch edge, C = Callitris patch). Means and standard errors are given.
Table 4.3 Models of Callitris survival in relation to char height on Callitris and
Callitris size in the low-moderate fire-weather zone. Models are ordered from best to
worst according to AIC. Survival ~ B0 + B1 Char Height + B2 Callitris Size + B3 Char
Height * Callitris Size.
Factors Coefficients R2 AIC
Intensity
Indicator Size B0 B1 B2 B3
Char height DBH -0.6 -3.5 0.3 0.58 147
Char height Height -2.5 -3.3 0.6 0.54 154

Chapter 4. Feedback between Callitris patches and fire intensity
83
Surv
ival p
rob
ab
ility
00.5
1
.0
Fig. 4.7. Probability of Callitris survival with char height and diameter at breast height
(DBH) in the low-moderate fire-weather zone.
Table 4.4 Classification table comparing the actual and the fitted values for alive and
dead Callitris for the survival models.
Model Fitted
Survival~ Dead Alive Correct (%)
Char height + DBH Actual dead 83 15 85
Actual alive 7 128 95
Average 91
Char height + Height Actual dead 78 19 80
Actual alive 8 127 94
Average 87

Chapter 4. Feedback between Callitris patches and fire intensity
84
Fuel characteristics
Five of the nine fuel characteristics differed significantly among vegetation types (Table
4.5). Shrub cover was greater in the Eucalyptus matrix than in all other vegetation types,
and shrub density was lower in Callitris patches than in all other vegetation types.
Eucalyptus leaf litter cover was greater in the Eucalyptus matrix and Eucalyptus matrix
edges than in Callitris patches and Callitris patch edges; the converse was true for
Callitris leaf litter cover and as well the Eucalyptus matrix had lower cover than
Eucalyptus matrix edge. Litter depth was greater in the Eucalyptus matrix than in all
other vegetation types. There were no significant effects of vegetation type on bare
ground cover, shrub height, fine fuel weight (litter, twigs < 6 mm diameter, bark) and
total ground fuel weight (fine fuel, grass, herbs, twigs 6-20 mm diameter).
Table 4.5 Results of Kruskal-Wallis tests on the effects of vegetation type (Eucalyptus
matrix (E), Eucalyptus matrix edge (Ee), Callitris patch edge (Ce), Callitris patch (C)),
on ground fuel characteristics. Means of significant tests are given in ascending order
and underlining indicates no significant difference (ns = not significant).
Factor Means for vegetation types X2
P
E Ee Ce C
Shrub cover (%) 24.1 6.0 5.3 2.8 14.02 0.003
Shrub density (number m-2
) 5.5 2.3 4.7 1.0 8.41 0.040
Eucalypt leaf litter cover (%) 52.1 45.9 23.2 23.9 12.01 0.005
Callitris leaf litter cover (%) 0.7 14.6 58.6 67.2 44.07 <0.001
Litter depth (mm) 20.4 15.9 14.6 16.2 8.61 0.040
Bare ground cover (%) 15.63 6.84 ns
Shrub height (m) 0.52 2.60 ns
Fine fuel wt (kg ha-1
) 7786 1.95 ns
Total ground fuel wt (kg ha-1
) 17581 3.76 ns

Chapter 4. Feedback between Callitris patches and fire intensity
85
Discussion
These results show that fire intensity was influenced by interactions between fire-
weather and vegetation structure, which in turn influenced Callitris survival. Under
extreme fire-weather, Callitris patches had very little effect on the incoming fire
intensity from the surrounding Eucalyptus matrix, resulting in a very low probability of
Callitris survival. By contrast, under low-moderate fire-weather, Callitris patches
reduced incoming fire intensity from the Eucalyptus matrix, resulting in a higher
probability of Callitris survival. Fewer shrubs and the greater cover of Callitris leaf
litter may have contributed to lower flammability in the Callitris patches. Thus patch
structure contributes to spatial heterogeneity of fire intensities in low-moderate fire-
weather, which enhances Callitris survival and coexistence with its more flammable
Eucalyptus competitor. This is especially important for the persistence of non-
serotinous Callitris, which are less likely to have seeds available post-fire than
serotinous Callitris.
Fire intensity indicators
In flat landscapes, fire intensity is influenced by interactions between weather and fuels
(Catchpole 2002; Boer et al. 2008), with their relative importance varying among
ecosystems (Bessie & Johnson 1995; Hely et al. 2001; Turner et al. 2003). In this study,
as hypothesised, fuels had less influence than weather on inferred fire intensity in the
extreme fire-weather zone. Fire intensity in outgoing quadrats, as measured by the
remaining stem tip diameter, was positively and linearly related to intensity in incoming
quadrats. Most Callitris patches experienced similar fire intensities to the Eucalyptus
matrix, despite lower flammability fuels. This result is consistent with a number of
studies, which found fuels to be less important in extreme fire-weather conditions
(Bessie & Johnson 1995; Turner et al. 2003). In at least two studies, the occurrence of
small patches of surviving obligate-seeders after crown fires, suggests that fuels have
some influence (Greene & Johnson 2000; Rodrigo et al. 2007).
As predicted in the low-moderate fire-weather zone, vegetation influenced fire intensity,
as measured by char height. This trend has been found in a range of other ecosystems
(Bessie & Johnson 1995; Turner et al. 1999; Knapp & Keeley 2006). In this study,
Callitris patches of more than 1 ha reduced the incoming fire intensity from the
Eucalyptus matrix within 10 m of patch edges. The magnitude of this abrupt decline in
intensity at the patch edge increased with increasing Callitris basal area, with a

Chapter 4. Feedback between Callitris patches and fire intensity
86
maximum of 20 m2 ha
-1 having the greatest effect. In other studies, lower intensity fires
have been observed in clumps of non-serotinous, obligate-seeding Callitris intratropica
compared with the surrounding more flammable forest (Bowman et al. 1988; Bowman
& Panton 1993).
Since trees with larger diameters contribute significantly more to basal area, larger
diameter Callitris were more likely to lower flammability than smaller diameter
Callitris in low-moderate fire-weather conditions. Indeed, larger diameter trees are
usually taller and have fewer lower branches, reducing the vertical continuity of fuels
(Alexander et al. 2006; Thompson & Spies 2009). The largest Callitris in this study,
established from 1890 to 1911 and in the early 1950s (Austin & Williams 1988; Norris
et al. 1991; Whipp 2009). Whilst those from 1890 - 1911 period were characterised by
an absence of lower branches, those from the 1950s had no foliage on the lower
branches. The older of these cohorts (1890 – 1911) survived a fire in the early 1950s,
when they were approximately 50 years old, a similar age to the Callitris in this study,
which established in the early 1950s. This suggests that a fire free interval of at least 50
years allows Callitris patches to grow old and dense enough to reduce fire intensity in
low-moderate fire-weather conditions and survive. This is consistent with the
interpretation that weak serotiny, which characterises Callitris glaucophylla, is an
adaptation to facilitate inter-fire recruitment (Enright et al. 1998; Whelan et al. 1998).
Callitris in this study successfully recruit in the absence of fire, following above
average rainfall (Austin & Williams 1988).
Callitris survival
Differences in fire intensity have consequences for the survival of Callitris. Only 4% of
Callitris survived fire in the extreme fire-weather zone, since most of their foliage was
either scorched or burnt, and like most conifers, Callitris does not resprout following
this level of damage (Gill 1981; Bond & Midgley 2001; Russell-Smith 2006). Given
this susceptibility, regular high intensity fires have led to declines in non-serotinous
seeder persistence and shifts in favour of resprouters or serotinous seeders in a range of
ecosystems (e.g. Prior et al. 2007; Pausas et al. 2008).
By comparison, Callitris survival was relatively high, reaching 54% in the low-
moderate fire-weather zone. Callitris survival increased as char height, a surrogate for
fire intensity declined and Callitris diameter, a surrogate for bark thickness and tree

Chapter 4. Feedback between Callitris patches and fire intensity
87
height increased. These trends are confirmation of the hypotheses. The survival of other
non-serotinous, obligate-seeder species are similarly affected by these interactions
(Bowman et al. 1988; Rodrigo et al. 2004 2007; Russell-Smith 2006). Callitris survival
was higher in Callitris patches, since they experienced lower fire intensities than in the
adjacent Eucalyptus matrix. When char heights on Callitris exceeded 1 m, most
Callitris suffered mortality, with survival restricted to trees > 25 cm DBH. This
preferential mortality or thinning of smaller size-classes is especially important for
obligate-seeder species not capable of self-thinning (Fule & Covington 1998; Russell-
Smith 2006), since it increases growth rates and shortens the time to maturity for the
survivors (Lacey 1972).
Fuel characteristics
How do we account for the differences in fire intensity between the Eucalyptus matrix
and the Callitris patches in the low-moderate fire-weather zone? Other studies have
found greater fuel biomass beneath Eucalyptus species, largely due to contributions
from woody material and also grasses (Clayton-Greene 1981; Bowman & Wilson
1988). It was unlikely that grasses played a significant role in the drought coincident
fire in this study, since grasses were sparse when I sampled one year after the drought
had broken. I also found similar levels of ground fuels in the Eucalyptus matrix and in
the Callitris patches, which may be explained by the ubiquity of Eucalyptus across the
patch transition (Fig. 2) and differences in the specific leaf area, which influence leaf
longevity (Lusk et al. 2003; Prior et al. 2003). Leaves of higher specific area,
characterising Eucalyptus, have a shorter leaf life span on the tree, so are more likely to
make a larger contribution to leaf litter than the leaves of lower specific area
characterising Callitris (Lusk et al. 2003; Prior et al. 2003).
This study did however find differences in shrub cover and leaf litter characteristics
between the Eucalyptus matrix and the Callitris patches. Greater shrub cover in the
Eucalyptus matrix is likely to have increased intensity by increasing elevated fuel levels
(Cheney 1981; Alexander et al. 2006; Thompson & Spies 2009). Higher cover of
Callitris and Eucalyptus leaf litters, in their respective locations may have influenced
flammability through differential spatial packing (Schwilk & Ackerly 2001; Scarff &
Westoby 2006). Despite comparable fine fuel biomass, the needle-like leaves of the
Callitris resulted in tightly packed, shallower leaf litter in the Callitris patches than in
the Eucalyptus matrix, which in experiments by Scarff & Westoby (2006) resulted in

Chapter 4. Feedback between Callitris patches and fire intensity
88
lower flammability. Correspondingly, at lower intensities, char heights were
consistently lower on Callitris than on Eucalyptus, although the influence of higher
shrub cover in the Eucalyptus matrix cannot be discounted. These trends are consistent
with the hypotheses on flammability differences between Callitris patches and
Eucalyptus matrix and agree with results from other studies on the relative litter
flammabilities of these coexisting functional types (Clayton-Greene 1981; Bowman &
Wilson 1988).
Summary
In a flat landscape, patch scale interactions between fuels and weather during fires
determine the fire intensity and thus the survival probability of non-serotinous, obligate-
seeding Callitris within a matrix of more flammable resprouting Eucalyptus. If Callitris
can establish patches of critical size and basal area during long fire free intervals,
vegetation–fire intensity feedbacks increase their chances of survival during low-
moderate fire-weather conditions. Thus, patch structure can influence spatial
heterogeneity of fire intensities and may be the key to the persistence of less flammable
species in a matrix of flammable species. In contrast, in extreme weather conditions,
where there is no such feedback, since weather has greater influence than fuel, the
survival of less flammable Callitris is very low. Predictions of an increase in the
frequency of extreme fire-weather days under global warming (Lucas et al. 2007),
suggest that fuel differences may be overridden by fire-weather in the future,
encouraging Eucalyptus to profit over Callitris.
Acknowledgements.
Australian Research Council (ARC) Linkage Grant between Charles Sturt University
(CSU) and NSW Department of Environment and Climate Change and Water (DECCW
NSW) provided the funding for this research (Project number LP066829). I would like
to thank NSW DECC staff at the Baradine office and Andrew and Katrina Cohn for
assistance in the field. Simon McDonald (CSU) assisted with statistical advice.

Chapter 5. Synthesis
89
Chapter 5.
Synthesis
Callitris glaucophylla and Eucalyptus species woodland, Merrimerriwa Range, central western New South Wales (Source: K. Ross)

Chapter 5. Synthesis
90
Chapter 5.
Synthesis
This research contributes to a better understanding of the dynamics of dominant
woodland trees in relation to climate and disturbance regimes in south eastern Australia.
The focus was on Callitris glaucophylla and a range of Eucalyptus species, which vary
in abundance across their range of coexistence and beyond (Sivertsen & Clarke 2000;
Harris & Lamb 2001). Functional differences between Callitris and Eucalyptus
contribute to these trends. Callitris glaucophylla is a slow-growing, obligate-seeding
gymnosperm and Eucalyptus species are faster growing, resprouting angiosperms
(Clayton-Greene 1981; Bowman & Harris 1995; Bell & Williams 1997; Gill 1997). The
relative success of Callitris and Eucalyptus was expected to vary with different rates
and intensities of disturbances, which varied along rainfall gradients (Bellingham &
Sparrow 2000).
I examined the following:
(1) How demographic patterns of Callitris glaucophylla are associated with rainfall
and disturbances along rainfall gradients;
(2) How competitive feedback between Eucalyptus species and Callitris
glaucophylla trees and C. glaucophylla regeneration influences stand dynamics
along a rainfall gradient;
(3) How feedback between patches of Callitris glaucophylla and fire intensity
influences Callitris survival in flammable Eucalyptus woodlands in different fire-
weather conditions.
The study area was in south eastern Australia, extending from the temperate region in
the east to the arid region in the west (600 - 200 mm). The thesis was written in three
chapters, with each chapter addressing one of the above questions. Surveys of varying
scales were used to address each question (Fig. 5.1). Since the first question examined
climatic effects, this study encompassed the entire rainfall gradient. The second
question related to comparative competitive effects of Callitris and Eucalyptus, and

Chapter 5. Synthesis
91
extended from the temperate to the semi-arid zone, where they coexist. The third
question was examined within a large area of woodland in the temperate zone, where
fires still occur regularly.
In this chapter I summarize the findings of the three chapters. I then integrate the
findings by defining geographic zones, within which demographic patterns of Callitris
and Eucalyptus are associated with particular suites of disturbance regimes (grazing,
fire, tree removal). I then make predictions of future dynamics and distributions of these
woodland dominants, as disturbance regimes change and shift with climate change.
Finally I outline directions for future research.
2
3
4
Figure 5.1 A satellite image of Australia showing the scale of each study, numbered by chapter
(Source: Geosciences, Australia)

Chapter 5. Synthesis
92
Chapter 2
Demographic patterns of Callitris glaucophylla are associated with rainfall and
disturbances along rainfall gradients
An important and topical ecological question is - how will climate change influence
species distributions? To predict changes in species distributions, it is important to
determine the factors responsible for current demographic patterns (Pearson & Dawson
2003). A first step in this process is to identify associations between demographic
patterns and environmental factors using broad-scale surveys along climatic gradients
(Peters et al. 2007). Species that are widespread, long-lived and can be aged are ideal
and can indicate how other species with similar traits may behave under climate change.
Callitris is widespread, ranging from the arid to the temperate zone. It lives for
approximately 200 - 250 years, and can be aged, so is ideal for this challenge (Bowman
& Harris 1995).
In this chapter I examined how demographic patterns of Callitris glaucophylla were
associated with rainfall and disturbances along rainfall gradients.
The number of Callitris cohorts since 1950 was recorded in relation to mean annual
(200 - 600 mm) and seasonal (summer, uniform, winter) rainfall, edaphic factors
(topography) and disturbance regimes (tenure, vertebrate grazing, fire). Data were
collected along three transects, each within a seasonal rainfall zone spanning the annual
rainfall gradient.
A switch from recruitment success to failure occurred at 405 mm annual rainfall in the
south-west. Below 405 mm annual rainfall where winter or uniform rainfall
predominate, Callitris has failed to recruit since the mid to late 1800s. This has resulted
in ageing Callitris, which live for approximately 200 - 250 years (Bowman & Harris
1995), so the older trees are approaching senescence. Recruitment failure was
associated with farms which graze livestock and the presence of rabbits. Sheep and
rabbits are renowned for depressing or preventing the establishment of Callitris and a
range of other palatable woody species in Australia (Auld & Keith 2009). By
comparison, recruitment has occurred more often since the 1950s, both above 405 mm
annual rainfall and also below 405 mm annual rainfall where summer rainfall is
predominant. In these regions recruitment was most successful where Callitris occurred

Chapter 5. Synthesis
93
on tenures other than farms, like roadsides and travelling stock routes, which have
historically lower grazing levels and where rabbits were less abundant. The absence of
fires may have also contributed to Callitris expansion, as it has with other fire sensitive
woody species (Belsky & Blumenthal 1997).
The study demonstrates how demographic patterns in Callitris were closely associated
with the spatial arrangement of disturbance regimes, dictated by regional land use, and
their relationship with rainfall along rainfall gradients. Low annual rainfall and high
levels of introduced grazing has led to ageing, contracting populations, whilst higher
annual rainfall with low levels of grazing and infrequent fire has led to younger,
expanding populations. As climate changes, rainfall, land use and disturbance regimes
will shift along climatic gradients, influencing the demography and distribution of
Callitris.
Chapter 3
Competitive feedback between Eucalyptus species and Callitris glaucophylla trees
and C. glaucophylla regeneration influences stand dynamics along a rainfall
gradient
Woody plant encroachment is a major issue in semi-arid regions throughout the world
(Scholes & Archer 1997; Asner et al. 2004), and the mechanisms triggering it are still
actively debated (Bond & Midgley 2000; Fensham et al. 2005; Sankaran et al. 2005).
Little attention has been given to the influence of canopy trees on woody encroachment
and how encroachment may in turn influence the survival of canopy trees.
In this chapter I examined how competitive feedback between Eucalyptus species and
Callitris glaucophylla trees and C. glaucophylla regeneration influences stand dynamics
along a rainfall gradient.
I sampled in six forests along a rainfall gradient during a drought. In each forest I
sampled saplings under canopies of Eucalyptus and Callitris and in adjacent gaps.
Canopy tree survival was assessed along two transects in each forest, one with and one
without regeneration.

Chapter 5. Synthesis
94
The results indicate feedback between Callitris saplings and Eucalyptus and Callitris
canopy tree dynamics, which varied along the rainfall gradient. Under canopies,
saplings were less dense, smaller and less likely to be reproducing than those in gaps.
Thus canopies increased the vulnerability of Callitris saplings, which are dependent on
seeds or thick bark, to survive disturbances like fire (Bowman et al. 1988; Bowman &
Harris 1995). There were higher densities of Callitris saplings under canopies at the
lower than higher end of the rainfall gradient, but the net effect of canopy trees on
sapling regeneration along the rainfall gradient was competitive rather than facultative.
Callitris canopy trees suffered higher mortality than Eucalyptus canopy trees during the
latest drought, despite similar abilities to reduce water usage during periods of water
deficit (Zeppel & Eamus 2005). The mortality of Callitris canopy trees not surrounded
by Callitris sapling regeneration ranged from 0 to 10%, compared with a consistent
10% for those surrounded by Callitris sapling regeneration. By comparison, the
mortality of Eucalyptus canopy trees was low (< 2%), irrespective of Callitris sapling
regeneration presence. Despite high survival, Eucalyptus canopy trees showed signs of
stress at low rainfall, coincident with the presence of Callitris sapling regeneration and
higher densities of Callitris canopy trees.
This study shows that interactions among climate, sapling regeneration and canopy trees
influenced dynamics in this semi-arid system. Feedback between sapling regeneration
and canopy trees was negative, especially during drought. Canopy trees moderated but
did not exclude sapling regeneration, which in turn increased mortality rates of Callitris
or stress levels in Eucalyptus canopy trees during drought. As droughts increase in
frequency and severity, saplings and canopy trees may continue to thin (Nicholls 2004;
CSIRO & BOM 2007; Hennessy et al. 2008).
Chapter 4
Feedback between patches of Callitris glaucophylla and fire intensity influences
Callitris survival in flammable Eucalyptus woodlands in different fire-weather
conditions
A fundamental question in fire ecology is – how do long-lived, obligate-seeders persist
in flammable landscapes (Bond & van Wilgen 1996)? Spatial heterogeneity in fire

Chapter 5. Synthesis
95
intensity, driven by interactions between weather and fuels or flammability may
contribute to species persistence (Whelan 1995; Boer et al. 2008).
In this chapter I examined how feedback between patches of Callitris glaucophylla and
fire intensity influenced Callitris survival in flammable Eucalyptus woodlands in
different fire-weather conditions.
The study was undertaken in Pilliga State Forest. In areas burned under extreme and
low-moderate fire-weather conditions, fire intensity and survival of Callitris were
measured in the flammable Eucalyptus matrix and in Callitris dominated patches. Pre-
fire fuel characteristics were measured in an adjacent unburnt area.
The results showed that fire intensity was influenced by interactions between fire-
weather and vegetation structure, which in turn influenced the survival of the obligate-
seeding, Callitris. Under extreme fire-weather conditions, Callitris patches had very
little effect on the incoming fire intensity from the surrounding Eucalyptus matrix,
resulting in a very low probability of Callitris survival (4%). By contrast, under low-
moderate fire-weather conditions, Callitris patches reduced incoming fire intensity from
the Eucalyptus matrix, resulting in a higher probability of Callitris survival (54%).
Callitris survival increased as char height declined and Callitris diameter increased.
This selective mortality or thinning of smaller size-classes is especially important for
obligate-seeders like Callitris, which are not capable of self-thinning (Fule & Covington
1998; Russell-Smith 2006).
Differences in shrub cover and leaf litter characteristics between the Eucalyptus matrix
and Callitris patches may explain lower fire intensity in the latter. Greater shrub cover
in the Eucalyptus matrix is likely to have increased fire intensity by increasing elevated
fuel levels (Cheney 1981; Alexander et al. 2006; Thompson & Spies 2009). Higher
covers of Callitris and Eucalyptus leaf litters in their respective locations, may have
influenced flammability, through differences in spatial packing (Schwilk & Ackerly
2001; Scarff & Westoby 2006). Despite comparable fine fuel biomass, the needle-like
leaves of Callitris result in tightly packed, shallower leaf litter in Callitris patches than
in the Eucalyptus matrix, which in experiments by Scarff & Westoby (2006) resulted in
lower flammability.

Chapter 5. Synthesis
96
In this flat landscape, patch scale interactions between fuels and weather during fires
determined the fire intensity and thus the survival probability of less flammable Callitris
within a matrix of more flammable resprouting Eucalyptus. If Callitris can establish in
dense patches during long fire free intervals, vegetation–fire intensity feedbacks
increase their chances of survival during low-moderate fire-weather conditions. Thus,
patch structure can influence spatial heterogeneity of fire intensities and may be the key
to the persistence of less flammable species in a matrix of flammable species. In
contrast, in extreme fire-weather conditions, where there is no such feedback, since
weather has greater influence than fuel, the survival of less flammable Callitris is very
low.
The dynamics of woodland dominants in geographic regions
The aim of this section is to integrate the results from the three studies. The studies
indicated that the dynamics of Callitris and Eucalyptus were associated with climate
and disturbance regimes along climatic gradients. Disturbance regimes were broadly
determined by land use (e.g. farms, roadsides, state forest, national parks), the nature of
which varied along rainfall gradients. A useful integrative approach is to define
geographic regions, within which different patterns of Callitris and Eucalyptus
dynamics are associated with unique suites of disturbance regimes (grazing, fire, tree
removal). This spatial approach is used to illustrate and explain, firstly the current
dynamics and distributions of Callitris and Eucalyptus, and secondly to speculate on
those of the future, under predicted trends in climate in south eastern Australia.
Callitris has contracted in the region (Region 1) below 405 mm mean annual rainfall,
where rain falls predominantly in winter or uniformly throughout the year (Fig. 5.2a).
Callitris populations in this region are ageing, with the majority dating from the mid to
late 1800s. Regeneration failure of Callitris was associated with low annual rainfall,
farms with high levels of livestock grazing and the presence of rabbits.
In a region of expansion (Region 2), Callitris has recruited a number of times since the
1950s (Fig. 5.2a). Above 405 mm mean annual rainfall, irrespective of seasonal
rainfall, regeneration of Callitris was most successful on tenures with low to moderate
levels of livestock grazing (e.g. roadsides, travelling stock routes, State Forests and
National Parks) and where rabbits and fire were absent. Less successful Callitris
regeneration occurred on farms and/or where rabbits were present. A combination of

Chapter 5. Synthesis
97
Sydney
CobarBroken Hill
200 600
Un
ifo
rm
Annual rainfall (mm)
0 200
Kilometres
N
400
Se
aso
na
l ra
infa
ll
Region 1. Callitris
contraction (winter &
uniform rainfall, farms,
rabbits)
Region 2. Callitris
expansion &
dominance (winter &
uniform rainfall,
roadsides , no rabbits, no fires)
Region 2. Callitris expansion
(summer rainfall, farms (west),
roadsides (east), few rabbits) Region 3. Callitris
& Eucalyptus
coexist (national
park, regular fires ,
few rabbits)
(a)
Sydney
CobarBroken Hill
200 600
Annual rainfall (mm)
0 200
Kilometres
N
400
Se
aso
na
l ra
infa
ll
Region 2.
Callitris expand
(slower) &
increase
dominance as Eucalyptus die
(drier)
Region 3.Callitris &
Eucalyptus coexist if
similar fire regime or
Eucalyptus dominate
if fires increase(similar climate to
now )
3.
Region 1.
Callitris extinct
(drier)
(b)
Region 2. Callitris expand slower in west (drier)
than in the east (similar climate to now)
?
?
?
Figure 5.2 Satellite images of New South Wales showing the dynamics of woodland
dominants in different geographic regions: (a) since 1950s; and (b) predicted under
climate change (Source: Geosciences, Australia).

Chapter 5. Synthesis
98
successful Callitris regeneration, ringbarking of Eucalyptus canopy trees (Allen 1995),
and low levels of Eucalyptus regeneration, has led to a switch from Eucalyptus to
Callitris domination in State Forests in the south east (Lunt et al. 2006; see Chapter 3).
Below 405 mm mean annual rainfall in the summer rainfall region of the north west,
Callitris regeneration has been successful on farms, where rabbits were infrequent. This
suggests that rabbit infrequency, rainfall during the growing season (summer) or their
interactions have contributed to establishment success here, when it failed further south
in winter rainfall zone.
Callitris and Eucalyptus coexist, where low levels of introduced grazers and regular
fires occur in the Pilliga forests of the north east (Fig. 5.2a; Region 3). If the fire-
sensitive Callitris can form dense patches of regeneration between fires (i.e. ~ 50 years,
see Chapter 4) that are capable of reducing fire intensity from the incoming Eucalyptus
matrix, then larger Callitris can survive fires under low-moderate fire weather
conditions. Large changes in the fire regime are likely to affect dominance in this
region.
Predicted dynamics of woodland dominants in geographic zones under climate
change
Drier conditions are expected over most of south eastern Australia (Nicholls 2004;
CSIRO & BOM 2007; Hennessy et al. 2008; DECCW 2009). Droughts are expected to
increase in intensity and severity. A shift to summer dominant rainfall is not expected to
compensate for a decline in rainfall, especially in the south west. Only in the north east
might a predicted increase in summer rainfall compensate for increasing temperatures
(DECCW 2009). These climatic changes may influence Callitris establishment and
survival both directly through processes like desiccation, and indirectly by changing
disturbance regimes, like grazing and fire (Gworek et al. 2007; Boulant et al. 2009). In
this section I predict changes in the dynamics of Callitris and Eucalyptus, under
predicted climate change in each geographic region.
The local extinction of Callitris is possible in the south west, as opportunities for
regeneration decline and ageing Callitris trees senesce (Region 1; Fig. 5.2b). Predicted
declines in the populations of rabbits and livestock as pasture biomass declines with
higher temperatures (Scanlan et al. 2006; Gunasekera et al. 2008) may not be enough to
compensate the direct effects of drought on Callitris seedling recruitment. This south

Chapter 5. Synthesis
99
west region may also expand further east if grazing by livestock and other introduced
herbivores displace unprofitable crop lands (Dale 1997; Dunlop & Brown 2008;
Garnaut 2010). Refuges from grazing (e.g. fenced roadsides, travelling stock routes)
will become increasingly important for the regeneration of Callitris, as the combined
effects of drought stress and grazing challenge survival thresholds in this region
(Dunlop & Brown 2008).
Callitris dominance is predicted to increase in the south east (Region 2; Fig. 5.2b), as
ageing Eucalyptus trees senesce and are replaced by Callitris, since Eucalyptus saplings
are infrequent. Their infrequency may be related to competition from dense Callitris
regeneration (FCNSW 1988), and also fire exclusion, which reduces establishment
opportunities (Bell & Williams 1997; Gill 1997). Fire frequency is unlikely to increase
because of predicted declines in herbaceous fuels, which limit fires in this region
together with high levels of landscape fragmentation (Bradstock 2010). Drier
conditions, fire exclusion and low-moderate grazing levels are likely to shift the system
further toward Callitris dominance, especially in the west of this region where
Eucalyptus canopy trees were most stressed during the latest drought (see Chapter 3).
In the north, changes in Callitris regeneration is predicted to vary from east to west. In
the north east, Callitris regeneration may continue at similar rates, if climate and
disturbance regimes remain relatively unchanged (Region 2; Fig. 5.2b). In the north
west, where the climate is predicted to be similar to the south west, opportunities for
regeneration may similarly decline (Region 2). But unlike the south west, this region
has younger Callitris cohorts to replace ageing Callitris trees, at least in the short term.
As temperatures increase, there is potential for Callitris to extend its range in the east,
where low temperatures are currently believed to limit seed germination (FCNSW 1988;
Fig. 5.3b). This expansion will also depend on disturbance regimes. If the potential for
fire increases in the more easterly ecosystems, as wet fuels dry (Bradstock 2010), the
expansion of Callitris will depend on the altered fire regime.
Dominance of Callitris will most likely to be determined by future fire regimes in the
Pilliga forests in the north east (Region 3; Fig. 5.2b). If predicted increases in
temperatures are matched by increases in summer rainfall (DECCW 2009) and the
current fire regime persists, coexistence of Callitris and Eucalyptus may continue.

Chapter 5. Synthesis
100
However, if rising temperatures reduce fuel moisture increasing fire potential
(Bradstock 2010), the system may shift toward Eucalyptus dominance. Intervals shorter
than 50 years may not be sufficient for Callitris to form the critically dense patches
capable of reducing incoming fire intensity from a Eucalyptus matrix during low-
moderate fire weather conditions (see Chapter 4).
Further work
Determination of causal relationships between Callitris dynamics and the associated
variables identified in this research requires smaller scale experimental studies. An area
of obvious interest is below 405 mm mean annual rainfall, where recruitment is
associated with interactions between seasonal rainfall (summer, winter) and grazing
(sheep, rabbits). Disentangling and thus determining the relative importance of these
variables on recruitment success will increase our ability to predict Callitris dynamics
and distributions, as drivers change and shift with climate.
Eucalyptus vary in abundance across their range in south eastern Australia. In this study
low levels of Eucalyptus regeneration were common. Determining the extent of
regeneration success and failure and the variables associated with these patterns will
increase our understanding of Eucalyptus dynamics and advise targeted smaller scale
research. This will increase our ability to predict Eucalyptus dynamics and distribution,
as drivers change and shift with climate. Because Eucalyptus are functionally different
from Callitris, drivers of demographic patterns may also differ.
Persistence of obligate-seeders in flammable systems is dependent on survival during
fire and post-fire regeneration. Little is known of post-fire seedling dynamics of
Callitris at sites which have experienced different fire intensities and how competition
with Eucalyptus seedlings determines the subsequent patterns of dominance. For
example, are fruiting cones (and seeds) on Callitris consumed during high intensity
fires, so that post-fire colonisation can only occur from areas, such as edges, which have
burned at lower intensity? If so, it may be difficult for Callitris to recolonise this area
before the next fire, resulting in Eucalyptus domination. This research would enhance
our ability to predict the likelihood of obligate-seeder persistence in flat landscapes,
where fires of mixed intensities occur regularly.

Chapter 5. Synthesis
101
Adaptive management is one approach that may be taken to address further work on
Callitris and Eucalyptus. Adaptive management is defined as ‘the integration of science,
management and learning within a collaborative framework’ (Jacobson et al. 2006). In
practice, adaptive management actions are experiments designed to gain information
(Holling 1978), and management decisions are reviewed regularly rather than following
a rigid experimental protocol (Lee 1999). By addressing the previous hypotheses on
Callitris and Eucalyptus regeneration success across the landscape, setting up
appropriate experiments to address these hypotheses and monitoring the results is likely
to contribute to disentangling the effects of climate and disturbances. For example,
results from this research suggest that recruitment failure of Callitris below 405 mm
mean annual rainfall in the winter rainfall zone is related to grazing by herbivores,
particularly rabbits. Seasonal timing of rainfall may also have some influence.
Establishing exclosures (exclude rabbits, exclude sheep/cattle/kangaroos, exclude all,
exclude none) to monitor Callitris recruitment success at sites below and above 405 mm
mean annual rainfall in the winter and summer rainfall zones, will help us understand
the relative contribution of grazing and seasonal rainfall to recruitment success. The
monitoring results would be fed interactively into the adaptive management approach to
inform further research and management (Schreiber et al. 2004).
Summary
It is clear that the relative success of Callitris and Eucalyptus was associated with
climate and disturbance regimes along climatic gradients. Prescribing different
disturbance regimes along the climatic gradient will contribute to the persistence of
these functional types. At the higher end of the rainfall gradient, a regime of regular
mixed intensity fires and low levels of grazing by introduced herbivores is likely to
contribute to the coexistence of Eucalyptus and Callitris. At the lower end of the rainfall
gradient where Callitris is in decline, reducing the levels of introduced grazers is likely
to contribute its persistence in this region, especially if droughts increase in intensity
and frequency and opportunities for establishment decline.

References
102
References
Ackerly, D. (2004). Functional strategies of chaparral shrubs in relation to seasonal
water deficit and disturbance. Ecological Monographs, 74(1), 25-44.
Ackerly, D. D. (2003). Community assembly, niche conservatism, and adaptive
evolution in changing environments. International Journal of Plant Sciences,
164(3), S165-S184.
Ackerly, D. D., & Reich, P. B. (1999). Convergence and correlations among leaf size
and function in seed plants:a comparative test using independent contrasts.
American Journal of Botany, 86(9), 1272-1281.
Agee, J. K. (1998). Fire and pine systems. In D. M. Richardson (Ed.), Ecology and
biogeography of Pinus (pp. 193-218). Cambridge: Cambridge University Press.
Alexander, J. D., Seavy, N. E., Ralph, C. J., & Hogoboom, B. (2006). Vegetation and
topographical correlates of fire severity from two fires in the Klamath-Siskiyou
region of Oregon and California. International Journal of Wildland Fire, 15(2),
237-245.
Allcock, K. G., & Hik, D. S. (2004). Survival, growth and escape from herbivory are
determined by habitat and herbivore species for three Australian woodland
plants. Oecologia, 138, 231-241.
Allen, M. (1995). The landscape archaeology of two western New South Wales forests.
Unpublished Master of Letters, University of New England, Armidale.
Allen, M. R. (1998). Forest history projects for State Forests of New South Wales: case
studies of three cypress pine forests in the Lachlan and Bogan River
Catchments, Forbes Forestry Distict on Back Yamma, Euglo South and Strahorn
State Forests. Sydney: State Forests of New South Wales.
Andrzejewska, L., & Gyllenberg, G. (1980). Small herbivore subsystem. In A. I.
Breymeyer & G. M. van Dyne (Eds.), Grasslands, Systems analysis and Man
(ist ed., pp. 201-268). Cambridge: Cambridge University Press.
Araújo, M. B., & New, M. (2007). Ensemble forecasting of species distributions.
Trends in Ecology & Evolution, 22(1), 42-47.
Armas, C., & Pugnaire, F. I. (2005). Plant interactions govern population dynamics in a
semi-arid plant community. Journal of Ecology, 93(5), 978-989.
Arno, S. F., & Gruell, G. E. (1986). Douglas-fir encroachment into mountain grasslands
in southwestern Montana. Journal of Range Management, 39(3), 272-276.

References
103
Arrieta, S., & Suárez, F. (2005). Spatial patterns of seedling emergence and survival as
a critical phase in holly (Ilex aquifolium L.) woodland recruitment in Central
Spain. Forest Ecology and Management, 205(1-3), 267-282.
Asner, G. P., Elmore, A. J., Olander, L. P., Martin, R. E., & Harris, T. (2004). Grazing
sysytems, ecosystem responses, and global change. Annual Review of
Environment & Resources, 29(1), 261-C-264.
Attiwill, P. M., & Clayton-Greene, K. A. (1984). Studies in gas exchange and
development in a subhumid woodland. Journal of Ecology, 72, 285-294.
Auld, T. D., & Keith, D. A. (2009). Dealing with threats: integrating science and
management. Ecological Management and Restoration, 10, s79-s87.
Austin, M. P., & Williams, O. B. (1988). Influence of climate and community
composition on the population demography of pasture species in semi-arid
Australia. Vegetatio, 77, 43-49.
Barnes, P. W., & Archer, S. (1999). Tree-shrub interactions in a subtropical savanna
parkland: competition or facilitation? Journal of Vegetation Science, 10(4), 525-
536.
Beerling, D. J., & Woodward, F. I. (1996). Palaeo-ecophysiological perspectives on
plant responses to global change. Trends in Ecology & Evolution, 11(1), 20-23.
Bell, D., T. (2001). Ecological response syndromes in the flora of southwestern Western
Australia:fire resprouters versus reseeders. The Botanical Review, 67(4), 417-
440.
Bell, D. T., & Williams, J. E. (1997). Eucalypt ecophysiology. In J. Williams & J.
Woinarski (Eds.), Eucalypt Ecology:Individuals to Ecosystems (pp. 168-196).
Cambridge: Cambridge University Press.
Bellingham, P. J., & Sparrow, A. D. (2000). Resprouting as a life history strategy in
woody plant communities. Oikos, 89, 409-416.
Belsky, A. J. (1994). Influences of Trees on Savanna Productivity: Tests of Shade,
Nutrients, and Tree-Grass Competition. Ecology, 75(4), 922-932.
Belsky, A. J., & Blumenthal, D. M. (1997). Effects of Livestock Grazing on Stand
Dynamics and Soils in Upland Forests of the Interior West. Conservation
Biology, 11(2), 315-327.
Benson, D., & McDougall, L. (1998). Ecology of Sydney plant species: Part 6.
Dicotyledon family Myrtaceae. Cunninghamia, 5, 808-987.

References
104
Benson, J. S., Allen, C. B., Togher, C., & Lemmon, J. (2006). New South Wales
vegetation classification and assessment:Part 1 plant communities of NSW
western plains. Cunninghamia, 9(3), 383-450.
Bertness, M. D., & Callaway, R. M. (1994). Positive interactions in communities.
Trends in Ecology and Evolution, 9, 191-193.
Bessie, W. C., & Johnson, E. A. (1995). The relative importance of fuels and weather
on fire behaviour in subalpine forests. Ecology, 76(3), 747-762.
Binns, D., & Beckers, D. (2001). Floristic patterns in the Pilliga. In J. Dargavel (Ed.),
Perfumed Pineries:Environmental History of Australia's Callitris Forests (pp.
104-110). Canberra: Australian National University.
Boer, M., Sadler, R., Bradstock, R., Gill, A., & Grierson, P. (2008). Spatial scale
invariance of southern Australian forest fires mirrors the scaling behaviour of
fire-driving weather events. Landscape Ecology, 23(8), 899-913.
Boland, D. J., Brooker, M. I. H., Chippendale, G. M., Hall, N., Hyland, B. P. M.,
Johnston, R. D., et al. (2006). Forest Trees of Australia (5th ed.). Collingwood:
CSIRO Publishing.
Bond, W., J. , & Midgley, G., F. . (2000). A proposed CO2 controlled mechanism of
woody plant invasion in grasslands and savannas. Global Change Biology, 6(8),
865-869.
Bond, W. J. (1989). The tortoise and the hare:ecology of angiosperm dominance and
gymnosperm persistence. Biological Journal of the Linnean Society, 36, 227-
249.
Bond, W. J., & Midgley, J. J. (2001). Ecology of sprouting in woody plants:persistence
niche. Trends in Ecology and Evolution, 16(1), 45-51.
Bond, W. J., Smythe, K.-A., & Balfour, D. A. (2001a). Acacia species turnover in space
and time in an African savanna. Journal of Biogeography, 28, 117-128.
Bond, W. J., & van Wilgen, B. W. (1996). Fire and Plants. London: Chapman & Hall.
Boulant, N., Garnier, A., Curt, T., & Lepart, J. (2009). Disentangling the effects of land
use, shrub cover and climate on the invasion speed of native and introduced
pines in grasslands. Diversity and Distributions, 15, 1047-1059.
Boulant, N., Kunstler, G., Rambal, S., & Lepart, J. (2008). Seed supply, drought, and
grazing determine spatio-temporal patterns of recruitment for native and
introduced invasive pines in grasslands. Diversity and Distributions, 14(5), 862-
874.

References
105
Bowman, D. M. J. S. (1998). Tansley Review No. 101. The impact of aboriginal
landscape burning on the Australian biota. New Phytologist, 140(3), 385-410.
Bowman, D. M. J. S., & Harris, S. (1995). Conifers of Australia's dry forests and open
woodlands. In N. J. Enright & S. Hill Robert (Eds.), Ecology of the Southern
Conifers (1 ed., pp. 252-270). Melbourne: Melbourne University Press.
Bowman, D. M. J. S., & Latz, P. K. (1993). Ecology of Callitris glaucophylla
(Cupressaceae) on the MacDonnell Ranges, Central Australia. Australian
Journal of Botany, 41, 217-225.
Bowman, D. M. J. S., Price O,, Whitehead, P. J., & Walsh, A. (2001). The 'wilderness
effect' and the decline of Callitris intratropica on the Arnhem Land Plateau,
northern Australia. Australian Journal of Botany, 49, 665-672.
Bowman, D. M. J. S., & Panton, W. J. (1993). Decline of Callitris intratropica
R.T.Baker & H.G. Smith in the Northern Territory: implications for pre- and
post-European colonization fire regimes. Journal of Biogeography, 20, 373-381.
Bowman, D. M. J. S., & Wilson, B. A. (1988). Fuel characteristics of coastal monsoon
forests, Northern Territory. Journal of Biogeography, 15, 807-817.
Bowman, D. M. J. S., Wilson, B. A., & Davis, G. W. (1988). Response of Callitris
intratropica R.T.Baker & H.G. Smith to fire protection, Murgenella, Northern
Australia. Australian Journal of Ecology, 13, 147-159.
Bradstock, R. A. (2010). A biogeographic model of fire regimes in Australia: current
and future implications. Global Ecology and Biogeography, 19(2), 145-158.
Bradstock, R. A., & Cohn, J. C. (2002). Fire regimes and biodiversity in semi-arid
mallee ecosystems. In R. A. Bradstock, J. E. Williams & A. M. Gill (Eds.),
Flammable Australia:the fire regimes and biodiversity of a continent (pp. 238-
258). Cambridge: Cambridge University Press.
Breiman, L., Freidman, J. H., Olshen, R. A., & Stone, C. J. (1993). Classification and
regression trees. New York: Chapman & Hall Inc.
Brodribb, T., & Hill, R. S. (1998). The photosynthetic drought physiology of a diverse
group of southern hemisphere conifer species is correlated with minimum
seasonal rainfall. Functional Ecology, 12(3), 465-471.
Brooker, M. I. H., & Kleinig, D. A. (1990). Field Guide to Eucalypts. Southwestern and
Southern Australia (Vol. 2). Sydney: Inkata Press.
Brookhouse, P., Nicholson, D., Green, D., & Lord, B. (1999). Large Pilliga fires and the
development of fire management and fire suppression strategies. In I. Lunt (Ed.),
Proceedings of the Australian Bushfire Conference.

References
106
Brown, J. R., & Archer, S. (1999). Shrub invasion of grassland: recruitment is
continuous and not regulated by herbaceous biomass or density. Ecology, 80(7),
2385-2396.
Bruno, J. F., Stachowicz, J. J., & Bertness, M. D. (2003). Inclusion of facilitation into
ecological theory. Trends in Ecology & Evolution, 18(3), 119-125.
Bureau of Meteorology. (2006). Special climate statement 9:An exceptionally dry
decade in parts of southern and eastern Australia:October 1996-September
2006. Melbourne: Bureau of Meteorology.
Bureau of Meteorology. (2010a). Major seasonal rainfall zones of Australia Retrieved
1/7/2010, 2010
Bureau of Meteorology. (2010b). Maps of rainfall percentages. Melbourne: Bureau of
Meteorology.
Bureau of Rural Sciences. (2009). 2001/02 Land uses of Australia, V.3. 2010
Burgman, M. A., & Lamont, B., B. . (1992). A stochastic model for the viability of
Banksia cuneata populations: environmental, demographic and genetic effects.
Journal of Applied Ecology, 29(3), 719-727.
Burrows, D. M., & Burrows, W. H. (1992). Seed production and litter fall in some
eucalypt communities in central Queensland. Australian Journal of Botany,
40(3), 389-403.
Burrows, W. H. (2002). Seeing the wood(land) for the trees-an individual perspective of
Queensland woodland studies (1965-2005). Tropical Grasslands, 36, 202-217.
Calamassi, R., Rocca, G. D., Falusi, M., Paoletti, E., & Strati, S. (2001). Resistance to
water stress in seedlings of eight European provenances of Pinus halapensis
Mill. Annals of Forest Science, 58, 663-672.
Callaway, R. M., & Walker, L. R. (1997). Competition and facilitation: a synthetic
approach to interactions in plant communities. Ecology, 78(7), 1958-1965.
Camarero, J. J., & Gutierrez, E. (2007). Response of Pinus uncinata recruitment to
climate warming and changes in grazing pressure in an isolated population of the
Iberian system (NE Spain). Arctic, Antarctic and Alpine Research, 39, 210-217.
Castro, J., Zamora, R., Hodar, J. A., & Gomez, J. M. (2005). Alleviation of summer
drought boosts establishment success of Pinus sylvestris in a mediterranean
mountain: an experimental approach. Plant Ecology, 181(2), 191-202.
Catchpole, W. (2002). Fire properties and burn patterns in heterogeneous landscapes. In
R. A. Bradstock, J. E. Williams & A. M. Gill (Eds.), Flammable Australia: the

References
107
fire regimes and biodiversity of a continent (pp. 49-75). Cambridge: Cambridge
University Press.
Cheney, N. P. (1981). Fire behaviour. In A. M. Gill, R. H. Groves & I. R. Noble (Eds.),
Fire and the Australian Biota (pp. 151-175). Canberra: Australian Academy of
Science.
Chesterfield, C. J., & Parsons, R. F. (1985). Regeneration of three tree species in arid
south-eastern Australia. Australian Journal of Botany, 33, 715-732.
Clarke, P. J. (2000). Plant population processes in temperate woodlands in eastern
Australia. In R. J. Hobbs & J. Yates Colin (Eds.), Temperate Eucalypt
Woodlands in Australia:Biology,Conservation, Management and Restoration
(pp. 248-270). Sydney: Surrey Beatty and Sons.
Clarke, P. J. (2002). Habitat insularity and fire response traits:evidence from a
sclerophyll archipelago. Oecologia, 132, 582-591.
Clarke, P. J., & Knox, J. E. (2002). Post-fire response of shrubs in the tablelands of
eastern Australia:do existing models explain habitat differences? Australian
Journal of Botany, 50, 53-62.
Clayton-Greene, K. A. (1981). The autecology of Callitris columellaris F.Muell. and
associated Eucalyptus spp. in south-eastern Australia. Unpublished PhD thesis,
University of Melbourne, Melbourne.
Clayton-Greene, K. A., & Ashton, D. H. (1990). The dynamics of Callitris
collumellaris/Eucalyptus albens communities along the Snowy River and its
tributaries in south-eastern Australia. Australian Journal of Botany, 38, 403-432.
Colls, K., & Whitaker, R. (2001). The Australian Weather Book. Sydney: Reed New
Holland Pty Ltd.
Cooke, B. D. (1977). Factors limiting the distribution of wild rabbit in Australia. Paper
presented at the Exotic species in Australia-their establishment success,
Adelaide.
Covington, W. W., & Moore, M. M. (1994). Southwestern ponderosa forest structure-
changes since Euro-American settlement. Journal of forestry (USA), 92, 39-47.
Cowling, R. M. (1987). Fire and its role in coexistence and speciation in Gondwanan
shrublands. South African Journal of Science, 83, 106-112.
Cox, S. J., Sivertsen, D. P., & Bedward, M. (2001). Clearing of native woody vegetation
in the New South Wales northern wheatbelt:extent,rate of loss and implications
for biodiversity. Cunninghamia, 7(1), 101-155.

References
108
CSIRO. (2008). Special climate statement 16:Long term rainfall deficiencies continue
in southern Australia while wet conditions dominate the north. Canberra
[www.bom.gov.au/climate/current/statements/scs16.pdf]: CSIRO.
CSIRO & BOM. (2007). Climate change in Australia: technical report 2007. Canberra:
CSIRO, Bureau of Meterology and Australian Greenhouse Office in partnership
with the Australian Climate Change Program.
Cunningham, G. M., Mulham, W. E., Milthorpe, P. L., & Leigh, J. H. (1992). Plants of
Western New South Wales. Sydney: Inkata Press.
Curtin, F. K. (1987). Notes on the establishment of cypress pine regeneration: Forestry
Commission of NSW.
Dale, V. H. (1997). The relationship between land-use change and climate change.
Ecological Applications, 7(3), 753-769.
Dale, V. H., Joyce, L. A., McNulty, S., Neilson, R. P., Ayres, M. P., Flannigan, M. D.,
et al. (2001). Climate Change and Forest Disturbances. (Cover story).
Bioscience, 51(9), 723.
Davis, S. D., Ewers, F. W., Wood, J., Reeves, J. J., & Kolb, K. J. (1999). Differential
susceptibility to xylem cavitation among three pairs of Ceanothus species in the
Transverse Mountain Ranges of southern California. Ecoscience, 6, 180-186.
DECCW NSW. (2009). NSW State of the Environment 2009. Sydney: NSW Dept of
Environment, Climate Change and Water.
Denham, A. J., & Auld, T. D. (2004). Survival and recruitment of seedlings and suckers
of trees and shrubs of the Australian arid zone following habitat management
and the outbreak of Rabbit Calicivirus Disease (RDC). Austral Ecology, 29(5),
585-599.
Denny, M. (1992). Historical and ecological study of the effects of european settlement
on inland NSW. Sydney: Nature Conservation Council of New South Wales.
Diaz, S., Noy-Meir, I., & Cabido, M. (2001). Can grazing response of herbaceous plants
be predicted from simple vegetative traits? Journal of Applied Ecology, 38(3),
497-508.
Dunlop, M., & Brown, P. R. (2008). Implications or climate change for Australia's
national reserve system: a preliminary assessment. Canberra: Report to
Department of Climate Change.
Eldridge, D. J., & Greene, R. S. B. (1994). Microbiotic soil crusts - a review of their
roles in soil and ecological processes in the rangelands of Australia. Australian
Journal of Soil Research, 32(3), 389-415.

References
109
Enright, N. J., & Lamont, B. B. (1992). Recruitment variability in the resprouting shrub
Banksia attenuata and non-sprouting congeners in the northern sandplain heaths
of southwestern Australia. Acta Oecologica, 13(6), 727-741.
Enright, N. J., Marsula, R., Lamont, B. B., & Wissel, C. (1998). The ecological
significance of canopy seed storage in fire-prone environments: a model for non-
sprouting shrubs. Journal of Ecology, 86(6), 946-959.
Facelli, J. M., & Springbett, H. (2009). Why do some species in arid lands increase
under grazing? Mechanisms that favour increased abundance of Maireana
pyramidata in overgrazed chenopod shrublands of South Australia. Austral
Ecology, 34(5), 588-597.
FCNSW. (1988). Notes on the Silviculture of Major NSW Forest Types.10. Cypress
Pine Types. Sydney: Forestry Commission of NSW.
Fensham, R. J. (1998a). The grassy vegetation of the Darling Downs, south-eastern
Queensland, Australia. Floristics and grazing effects. Biological Conservation,
84(3), 301-310.
Fensham, R. J. (1998b). The influence of cattle grazing on tree mortality after drought
in savanna woodland in north Queensland. Australian Journal of Ecology, 23(4),
405-407.
Fensham, R. J. (2008). Leichhardts maps: 100 years of change in vegetation structure in
inland Queensland. Journal of Biogeography, 35(1), 141-156.
Fensham, R. J., Fairfax, R. J., & Ward, D. P. (2009). Drought-induced tree death in
savanna. Global Change Biology, 15(2), 380-387.
Fensham, R. J., & Holman, J. E. (1999). Temporal and spatial patterns in drought-
related tree dieback in Australian savanna. Journal of Applied Ecology, 36(1),
1035-1050.
Florence, R. G. (1996). Ecology and Siviculture of Eucalypt Forests. Melbourne:
CSIRO.
Foden, W., Midgley, G. F., Hughes, G., Bond, W. J., Thuiller, W., Hoffman, M. T., et
al. (2007). A changing climate is eroding the geographical range of the Namib
Desert tree Aloe through population declines and dispersal lags. Diversity &
Distributions, 13(5), 645-653.
Foran, B. D. (1986). The impact of rabbits and cattle on an arid calcareous shrubby
grassland in central Australia. Vegetatio, 66, 49-59.
Fox, M. D. (1991). The natural vegetation of the Ana Branch-Mildura 1:250000 map
sheets. Cunninghamia, 2(3), 443-493.

References
110
Fuentes, E. R., Otaiza, R. D., Alliende, M. C., Hoffmann, A., & Poiani, A. (1984).
Shrub clumps of the Chilean matorral vegetation: structure and possible
maintenance mechanisms. Oecologia, 62(3), 405-411.
Fulé, P. Z., & Covington, W. W. (1998). Spatial patterns of Mexican pine-oak forests
under different recent fire regimes. Plant Ecology, 134(2), 197-209.
García, D., Zamora, R., Hódar, J. A., & Gómez, J. M. (1999). Age structure of
Juniperus communis L. in the Iberian peninsula: Conservation of remnant
populations in Mediterranean mountains. Biological Conservation, 87(2), 215-
220.
Garnaut, R. (2010). Climate change and the Australian agricultural and resource
industries. Australian Journal of Agricultural and Resource Economics, 54(1),
9-25.
GenStat. (2009). The guide to GenStat Release 12. Part 2: Statistics. Waterhouse Street,
Hemel Hempstead, Hertfordshire HP1 1ES, UK: VSN International
Gill, A. M. (1981). Adaptive responses of Australian vascular plant species to fires. In
A. M. Gill, R. H. Groves & I. R. Noble (Eds.), Fire and the Australian Biota.
Canberra: The Australian Academy of Science.
Gill, A. M., & Bradstock, R. A. (1992). A national register for the fire response of plant
species. Cunninghamia, 2, 653-660.
Gill, A. M., & Moore, P. H. R. (1990). Fire intensities in eucalyptus forests of
southeastern Australia. Paper presented at the International Conference on
Forest Fire Research, Coimbra, Portugal.
Gill, M. (1997). Eucalypts and fires:interdependent or independent? In J. Williams & J.
Woinarski (Eds.), Eucalypt Ecology:Individuals to Ecosystems (pp. 151-167).
Cambridge: Cambridge University Press.
Graz, F. P. (2008). The woody weed encroachment puzzle: gathering pieces.
Ecohydrology, 1(4), 340-348.
Greene, D. F., & Johnson, E. A. (2000). Tree recruitment from burn edges. Canadian
Journal of Forest Research, 30(8), 1264-1274.
Grime, J. P. (1979). Plant Strategies and Vegetation Processes. Chichester: John Wiley
and Sons.
Grime, J. P. (2001). Plant Strategies, vegetation processes and ecosystem properties.
Chichester: John Wiley and Sons.
Grubb, P. J. (1977). The maintenance of species-richness in plant communities:the
importance of the regeneration niche. Biological Review, 52, 107-145.

References
111
Guarín, A., & Taylor, A. H. (2005). Drought triggered tree mortality in mixed conifer
forests in Yosemite National Park, California, USA. Forest Ecology and
Management, 218(1-3), 229-244.
Guisan, A., & Thuiller, W. (2005). Predicting species distribution: offering more than
simple habitat models. Ecology Letters, 8(9), 993-1009.
Gunasekera, D., Tulloh, C., Ford, M., & Heyhoe, E. (2008, 13 June 2008). Climate
change:opportunities and challenges in Australian agriculture. Paper presented
at the Facing climate change:reserach on adaptation og agrosystems, University
of Sydney.
Gurevitch, J., Scheiner, S. M., & Fox, G. A. (2006). The Ecology of Plants. Sunderland,
MA: Sinauer Associates, Inc.
Gutiérrez, J. R., Holmgren, M., Manrique, R., & Squeo, F. A. (2007). Reduced
herbivore pressure under rainy ENSO conditions could facilitate dryland
reforestation. Journal of Arid Environments, 68(2), 322-330.
Gworek, J. R., Vander Wall, S. B., & Brussard, P. F. (2007). Changes in biotic
interactions and climate determine recruitment of Jeffrey pine along an elevation
gradient. Forest Ecology and Management, 239(1-3), 57-68.
Haase, P., Pugnaire, F. I., Clark, S. C., & Incoll, L. D. (1996). Spatial patterns in a two-
tiered semi-arid shrubland in southeastern Spain. Journal of Vegetation Science,
7(4), 527-534.
Hanson, P. J., & Weltzin, J. F. (2000). Drought disturbance from climate change:
response of United States forests. The Science of The Total Environment, 262(3),
205-220.
Harden, G. J. (2000). Flora of New South Wales (Vol. 1). Sydney: New South Wales
University Press.
Harden, G. J. (2002). Flora of New South Wales (Vol. 2). Sydney: New South Wales
University Press.
Harrington, G. N. (1991). Effects of soil moisture on shrub seedling survival in a semi-
arid grassland. 1338-1149.
Harrington, G. N., Dawes, G. T., & Ludwig, J. A. (1981). An analysis of the vegetation
pattern in a semi-arid Eucalyptus populnea woodland in north-west New South
Wales. Australian Journal of Ecology, 6, 279-287.
Harris, M., & Lamb, D. (2001). Recent disturbances to Callitris forests in Southern
Queensland. In J. Dargavel (Ed.), Perfumed Pineries:Environmental History of

References
112
Australia's Callitris Forests (pp. 125-135). Canberra: Australian National
University.
Hattersley, P. W. (1983). The distribution of C3 and C4 grasses in Australia in relation to
climate. Oecologia, 57(1/2), 113-128.
Hawkins, P. J. (1966). Seed production and litterfall studies of Callitris columellaris.
Australian Forest Research, 2, 3-16.
Heckathorn, S. A., McNaughton, S. J., & Coleman, J. S. (1999). C4 plants and
herbivory. In R. F. Sage & R. K. Monson (Eds.), C4 Plant Biology (pp. 285-
312). London: Academic Press.
Hely, C., Flannigan, M., Bergeron, Y., & McRae, D. (2001). Role of vegetation and
weather on fire behavior in the Canadian mixedwood boreal forest using two fire
behavior prediction systems. Canadian Journal of Forest Research, 31(3), 430-
441.
Henery, M. L., & Westoby, M. (2001). Seed mass and seed nutrient content as
predictors of seed output variation between species. Oikos, 92(3), 479-490.
Hennessy, K., Fawcett, R., Kirono, D., Mpelasoka, F., Jones, D., Bathols, J., et al.
(2008). An assessment of the impact of climate change on the nature and
frequency of exceptional climatic circumstances: Commonwealth of Australia.
Hennessy, K., Lucas, C., Nicholls, N., Bathols, J., Suppiah, R., & Ricketts, J. (2005).
Climate change impacts on fire-weather in south-east Australia: CSIRO
Atmospheric Research
Australian Government Bureau of Meterology.
Hilbert, D. W. (1987). A model of life history strategies of chapparal shrubs in relation
to fire frequency. In J. D. Tenhunen, F. M. Catarino, O. L. Lange & W. C.
Oechel (Eds.), Plant Response to Stress:Functional Analysis in Mediterranean
Ecosystems. (pp. 597-606). Berlin: Springer-Verlag.
Hill, K. D. (1998). Pinophyta. In Flora of Australia, vol. 48: Ferns, Gymnosperms and
Allied Groups (pp. 545-596). Melbourne: CSIRO, Australia.
Hill, R., S. (1995). Conifer origin, evolution and diversification in the southern
hemisphere. In N. J. Enright & S. Hill Robert (Eds.), Ecology of the Southern
Conifers (1 ed., pp. 10-29). Melbourne: Melbourne University Press.
Hill, R. S., & Brodribb, T. J. (1999). Turner Review No. 2. Southern conifers in time
and space. Australian Journal of Botany, 47, 639-696.

References
113
Hodgkinson, K., Harrington, G. N., & Miles, G. E. (1980). Composition, spatial and
temporal variability of the soil seed pool in a Eucalyptus populnea shrub
woodland in central New South Wales. Australian Journal of Ecology, 5, 23-29.
Hodgkinson, K. C. (1979). The shrubs of poplar box (Eucalyptus populnea) lands and
their biology. The Rangeland Journal, 1(4), 280-293.
Hodgkinson, K. C. (1991). Shrub recruitment response to intensity and season of fire in
a semi-arid woodland. Journal of Applied Ecology, 28, 60-70.
Hodgkinson, K. C., & Harrington, G. N. (1985). The case for prescribed burning to
control shrubs in eastern semi-arid woodlands. Australian Rangeland Journal,
64-74.
Holling, C.S. 1978. Adaptive environmental assessment and management. John Wiley,
London.
Holmgren, M., Lopez, B. C., Gutierrez, J. R., & Squeo, F. A. (2006b). Herbivory and
plant growth rate determine the success of El Nino Southern Oscillation-driven
tree establishment in semiarid South America. Global Change Biology, 12(12),
2263-2271.
Holmgren, M., & Scheffer, M. (2001). El Niño as a window of opportunity for the
restoration of degraded arid ecosystems. Ecosystems, 4(2), 151-159.
Holmgren, M., Scheffer, M., & Huston, M. A. (1997). The interplay of facilitation and
competition in plant communities. Ecology, 78(7), 1966-1975.
Holmgren, M., Stapp, P., Dickman, C. R., Gracia, C., Graham, S., Gutierrez, J. R., et al.
(2006a). Extreme climatic events shape arid and semiarid ecosystems. Frontiers
in Ecology and the Environment, 4(2), 87-95.
Hua, Q. (2009). Radiocarbon:a chronological tool for the recent past. Quaternary
Geochronology, 4, 378-390.
Huntly, N. (1991). Herbivores and the dynamics of communities and ecosystems.
Annual Review of Ecology & Systematics, 22, 477-503.
Huston, M. (1979). A general hypothesis of species diversity. The American Naturalist,
81-101.
Huston, M. (2003). Understanding the effects of fire and other mortality causing
disturbances on species diversity. In I. Abott & N. Burrows (Eds.), Fire in
Ecosystems of south-west Western Australia: Impacts and Management (pp. 37-
70). Leiden, The Netherlands: Backhuys Publishers.
Insightful Corp. (2007). S-Plus 8 Guide to Statistics. Seattle, Washington: Insightful
Corporation.

References
114
IPCC. (2007). Climate change 2007: The physical basis. Contributions of working
group 1 to the fourth assessment report of the intergovernmental panel on
climate change. New York.
Jacobson, S.K., Morris, J.K., Sanders, J.S., Wiley, E.N., Brooks, M., Bennetts,
R.E., Percival, H.F. & Marynowski, S. 2006. Understanding Barriers to
Implementation of an Adaptive Land Management Program.
Conservation Biology 20: 1516-1527.
Johnston, T. N. (1969). The effect of sheep and rabbit grazing on regeneration of white
cypress pine. Australian Forest Research, 4, 3-12.
Jump, A. S., Mátyás, C., & Peñuelas, J. (2009). The altitude-for-latitude disparity in the
range retractions of woody species. Trends in Ecology & Evolution, 24(12), 694-
701.
Keeley, J., E. , & Zedler, P. H. (1978). Reproduction of chaparral shrubs after fire: a
comparison of sprouting and Seeding strategies. American Midland Naturalist,
99(1), 142-161.
Keeley, J. E., Hopkins, A. J. M., & Lamont, B. B. (1986). Resilience of mediterranean
shrub communities to fire. In B. Dell (Ed.), Resilience in Mediterranean type
ecosystems (pp. 95-112). Dordrecht, The Netherlands: W.Junk Publishers.
Keeley, J. E., & Zedler, P. H. (1998). Evolution of life histories in Pinus. In D. M.
Richardson (Ed.), Ecology and biogeography of Pinus (1 ed., pp. 219-249).
Cambridge: Cambridge University Press.
Keith, D. (2004). Ocean Shores to Desert Dunes:the Native Vegetation of New South
Wales and the ACT. Sydney: Department of Conservation (NSW).
Keith, D. A. (1991). Coexistence and species diversity in upland swamp vegetation:the
roles of an environmental gradient and recurring fires. Unpublished PhD,
University of Sydney, Sydney, Australia.
Keith, D. A., Akcakaya, H. R., Thuiller, W., Midgely, G. F., Pearson, R. G., Phillips, S.
J., et al. (2008). Predicting extinction risks under climate change: coupling
stochastic population models with dynamic bioclimatic habitat models. Biology
Letters, 4, 560-563.
Knapp, E. E., & Keeley, J. E. (2006). Heterogeneity in fire severity within early season
and late season prescribed burns in a mixed-conifer forest. International Journal
of Wildland Fire, 15(1), 37-45.

References
115
Knox, K. J. E., & Clarke, P. J. (2005). Nutrient availability induces contrasting
allocation and starch in resprouting and obligate seeding shrubs. Functional
Ecology, 19, 690-698.
Kraaij, T., & Ward, D. (2006). Effects of rain, nitrogen, fire and grazing on tree
recruitment and early survival in bush-encroached savanna, South Africa. Plant
Ecology, 186(2), 235-246.
Krassilov, V. A. (1978). Late Cretaceous gymnosperms from Sakhalin and the terminal
Cretaceous event. Palaeontology, 21, 893-905.
Kruger, F. J. (1983). Plant community diversity and dynamics in relation to fire. In F. J.
Kruger, D. Mitchell & J. Jarvis (Eds.), Mediterranean-type ecosystems: the role
of nutrients (pp. 446-472). Berlin: Springer-Verlag.
Kruger, F. J., & Bigalke, R. C. (1984). Fire in fynbos. In P. V. Booysen & N. M.
Tainton (Eds.), Ecological effects of fire in South African ecosystems
(pp. 67-114). New York: Springer-Verlag.
Lacey, C. J. (1972). Factors influencing occurrence of cypress pine regeneration in
New South Wales. Sydney: Forestry Commission of NSW.
Lacey, C. J. (1973). Silvicultural characteristics of white cypress pine. Sydney: Forestry
Commission of NSW.
Lamont, B. B., Groom, P. K., & Cowling, R. M. (2002). High leaf area mass per area of
related species assemblages may reflect low rainfall and carbon isotope
discrimination rather than phosphorous and nitrogen concentrations. Functional
Ecology, 16, 403-412.
Landsberg, J., Lavorel, S., & Stol, J. (1999). Grazing response groups among
understorey plants in arid rangelands. Journal of Vegetation Science, 10, 683-
696.
Landsberg, J., & Stol, J. (1996). Spatial distribution of sheep, feral goats and kangaroos
in woody rangeland paddocks. Rangeland Journal, 18(2), 270-291.
League, K., & Veblen, T. (2006). Climatic variability and episodic Pinus ponderosa
establishment along forest-grassland ecotones of Colorado. Forest Ecology and
Management, 228, 98-107.
Lee, K.N. 1993. Compass and gyroscope: integrating science and politics for the
environment. Island Press, Washington D.C.
Letnic, M., Tamayo, B., Dickman, C. R., & Kelt, D. A. (2005). The responses of
mammals to La Nina (EL Nino Southern Oscillation)-associated rainfall,

References
116
predation and wildfire in central Australia Journal of Mammalogy, 86(4), 689-
703.
Lindsay, A. D. (1967). Forest types of the New South Wales Cypress Pine Zone (No.
Technical report No. 8). Sydney: Forestry Commission of NSW.
Llorens, L., Peñuelas, J., Beier, C., Emmett, B., Estiarte, M., & Tietema, A. (2004).
Effects of an experimental increase of temperature and drought on the
photosynthetic performance of two Ericaceous shrub species along a north–
south European gradient. Ecosystems, 7(6), 613-624.
Lloyd, A. H. (1997). Response of tree-line populations of foxtail pine (Pinus
balfouriana) to climate variation over the last 1000 years. Canadian Journal of
Forest Research, 27, 936-942.
Loehle, C. (2000). Strategy space and the disturbance spectrum:a life-history model for
tree species coexistence. The American Naturalist, 156(1), 14-33.
Loehle, C., & LeBlanc, D. (1996). Model-based assessments of climate change effects
on forests: a critical review. Ecological Modelling, 90(1), 1-31.
López, B. C., Holmgren, M., Sabaté, S., & Gracia, C. A. (2008). Estimating annual
rainfall threshold for establishment of tree species in water-limited ecosystems
using tree-ring data. Journal of Arid Environments, 72(5), 602-611.
Lucas, C., Hennessy, K., Mills, G., & Bathols, J. (2007). Bushfire weather in
southeastern Australia: recent trends and projected climate change impacts.
Sydney: The Climate Institute of Australia.
Luck, G. W. (2007). The relationships between net primary productivity, human
population density and species conservation. Journal of Biogeography, 34(2),
201-212.
Ludwig, J., Tongway, D. J., Freudenberger, D., & Noble, J. (Eds.). (1997). Landscape
Ecology Function and Management: Principles from Australia's Rangelands.
Collingwood: CSIRO, Australia.
Lunney, D. (2001). Causes of the extinction of native mammals of the Western Division
of New South Wales: an ecological interpretation of the nineteenth century
historical record. The Rangeland Journal, 23(1), 44-70.
Lunt, I., & Spooner, P. G. (2005). Using historical ecology to understand patterns of
biodiversity in fragmented agricultural landscapes. Journal of Biogeography, 32,
1859-1873.
Lunt, I. D. (1995). European management of remnant grassy forests and woodlands in
south-eastern Australia. Victorian Naturalist, 112, 239-249.

References
117
Lunt, I. D. (2005). Effects of Stock Grazing on Biodiversity Values in Temperate Native
Grasslands and Grassy Woodlands in SE Australia:A Literature Review.
Canberra: Environment ACT.
Lunt, I. D. (2005). Technical Report 18. Effects of Stock Grazing on Biodiversity Values
in Temperate Native Grasslands and Grassy Woodlands in SE Australia:A
Literature Review. Canberra: Environment ACT.
Lunt, I. D., Jones, N., Spooner, P. G., & Petrow, M. (2006). Effects of European
colonization on indigenous ecosystems:post-settlement changes in tree stand
structures in Eucalyptus-Callitris woodlands in central New South Wales,
Australia. Journal of Biogeography, 33, 1102-1115.
Lusk, C. H., & Matus, F. (2000). Juvenile tree growth rates and species sorting on fine-
scale soil fertility gradients in a Chilean temperate rain forest. Journal of
Biogeography, 27(4), 1011-1020.
Lusk, C. H., Wright, I., & Reich, P. B. (2003). Photosynthetic differences contribute to
competitive advantage of evergreen angiosperm trees over evergreen conifers in
productive habitats. New Phytologist, 160, 329-336.
McCarthy, G.J., Tolhurst, K.G. & Chatto, K. (1999). Overall Fuel Hazard Guide. In:
Research Report No. 47, Department of Natural Resources and Environment,
Melbourne.
McDonald, R. C., Isbell, R. F., Speight, J. G., Walker, J., & Hopkins, M. S. (1990).
Australian Soil and Land Survey:Field Handbook (2nd ed.). Melbourne: Inkata
Press.
McKinney, M. L., & Lockwood, J. L. (1999). Biotic homogenization: a few winners
replacing many losers in the next mass extinction. Trends in Ecology &
Evolution, 14(11), 450-453.
McPherson, G. R., Wright, H. A., & Wester, D. B. (1988). Patterns of shrub invasion in
semiarid texas grasslands. American Midland Naturalist, 120(2), 391-397.
Meentemeyer, R. K., Moody, A., & Franklin, J. (2001). Landscape-scale patterns of
shrub-species abundance in California chaparral – The role of topographically
mediated resource gradients. Plant Ecology, 156(1), 19-41.
Meserve, P. L., Kelt, D. A., Milstead, W. B., & Gutierrez, J. R. (2003). Thirteen years
of shifting top-down and bottom-up control. Bioscience, 53(7), 633-646.
Meyer, S. E., & Pendleton, B., K. (2005). Factors affecting seed germination and
seedling establishment of a long-lived desert shrub (Coleogyne ramosissima:
Rosaceae). Plant Ecology, 178(2), 171-187.

References
118
Miller, E. A., & Halpern, C. B. (1998). Effects of environment and grazing disturbance
on tree establishment in meadows of the central Cascade Range, Oregon, USA.
Journal of Vegetation Science, 9(2), 265-282.
Moreno, J. M., & Oechel, W. C. (1989). A simple method for estimating fire intensity
after a burn in Californian chapparral. Acta Oecologica, 10(1), 57-68.
Mortlock, W., & Lloyd, M. (2001). Floradata-A Guide to Collection, Storage and
Propagation of Australian Native Plant Seed. Searchable Database February
2001. Canberra: Australian Centre for Mining Environmental Research
(Brisbane), Australian National Botanic Gardens, CSIRO Forestry and Forest
Products, Greening Australia Limited.
Murphy, B. P., & Bowman, D. M. J. S. (2007). Seasonal water availability predicts the
relative abundance of C3 and C4 grasses in Australia. Global Ecology &
Biogeography, 16(2), 160-169.
Myers, K., Parer, I., Wood, D., & Cooke, B. D. (1994). The rabbit in Australia. In H. V.
Thompson & C. M. King (Eds.), The European Rabbit (pp. 108-157). Oxford:
Oxford University Press.
Nicholls, N. (1992). Historical El Nino/Southern Oscillation variability in the
Australasian region. In H. F. Diaz & V. Markgraf (Eds.), El Nino: Historical
and paleo aspects of the Southern Oscillation (pp. 151-172). Cambridge:
Cambridge University Press.
Nicholls, N. (2004). The changing nature of Australian droughts. Climatic Change,
63(3), 323-336.
Niinemets, U. (2001). Global-scale climatic controls of leaf dry mass per area, density
and thickness in trees and shrubs. Ecology, 82(3), 453-469.
Noble, I. R., Bary, G. A. V., & Gill, A. M. (1980). McArthur's fire-danger meters
expressed as equations. Australian Journal of Ecology, 5, 201-203.
Noble, I. R., & Gitay, H. (1996). A functional classification for predicting the dynamics
of landscapes. Journal of Vegetation Science, 7, 329-336.
Noble, J. C. (1997). The delicate and noxious scrub. Canberra: CSIRO.
Noble, J. C. (2001). Regulating Callitris populations: tale of two pineries. In J. Dargavel
(Ed.), Perfumed Pineries:Environmental History of Australia's Callitris Forests
(pp. 173-183). Canberra: Australian National University.
Noble, J. C., & Vines, R. G. (1993). Fire studies in mallee (Eucalyptus spp.)
communities of western New South Wales: grass fuel dynamics and associated
weather patterns. The Rangeland Journal, 15(2), 270-297.

References
119
Norris, E. H., Mitchell, P. B., & Hart, D. M. (1991). Vegetation changes in the Pilliga
forests: a preliminary evaluation of the evidence. Vegetatio, 91, 209-218.
Noss, R. F., Franklin, J. F., Baker, W. L., Schoennagel, T., & Moyle, P. B. (2006).
Managing fire-prone forests in western United States. Frontiers in Ecology and
the Environment, 4(9), 481-487.
Noy-Meir, I. (1973). Desert ecosystems: environment and procedures. Annual Review of
Ecology & Systematics, 4, 25-51.
Ojeda, F. (1998). Biogeography of seeder and resprouter Erica species in the Gape
Floristic Region-Where are the resprouters? Biological Journal of the Linnean
Society, 63(3), 331-347.
Ojeda, F., Brun, F., G. , & Vergara, J., J. . (2005). Fire, rain and the selection of seeder
and resprouter life-histories in fire-recruiting, woody plants. New Phytologist,
168(1), 155-165.
Parer, I. (1987). Factors influencing the distrubtion and abundance of rabbits
(Oryctolagus cuniculus) in Queensland. Proceedings of the Royal Society of
Queensland, 98, 73-82.
Parker, D., & Lunt, I. D. (2000). Stand structure and recruitment patterns in Callitris-
Eucalyptus woodlands in Terrick Terrick National Park, Victoria. Victorian
Naturalist, 117, 207-213.
Paula, S., & Pausas, J. G. (2006). Leaf traits and resprouting ability in the
Mediterranean basin. Functional Ecology, 1-7.
Pausas, J. G., & Bradstock, R. A. (2007). Fire persistence traits of plants along a
productivity and disturbance gradient in mediterranean shrublands of south-east
Australia. Global Ecology & Biogeography, 16(3), 330-340.
Pausas, J. G., Bradstock, R. A., Keith, D. A., Keeley, J. E., & GCTE. (2004). Plant
functional traits in relation to fire in crown-fire ecosystems. Ecology, 85(4),
1085-1100.
Pausas, J. G., Carbó, E., Caturla, R. N., Gil, J. M., & Vallejo, R. (1999). Post-fire
regeneration patterns in the eastern Iberian Peninsula. Acta Oecologica, 20(5),
499-508.
Pausas, J. G., Llovet, J., Rodrigo, A., & Vallejo, R. (2008). Are wildfires a disaster in
the Mediterranean basin? A review. International Journal of Wildland Fire,
17(6), 713-723.

References
120
Pearson, R., G. , & Dawson, T., P. . (2003). Predicting the impacts of climate change on
the distribution of species: are bioclimate envelope models useful? Global
Ecology & Biogeography, 12(5), 361-371.
Peters, D. P. C., Bestelmeyer, B. T., Herrick, J. E., Fredrickson, E. L., Monger, H. C., &
Havstad, K. M. (2006). Disentangling complex landscapes: new insights into
arid and semiarid system dynamics. Bioscience, 56(6), 491-501.
Peterson, D. L., & Arbaugh, M. J. (1986). Postfire survival in Douglas-fir and lodgepole
pine:comparing the effects of crown and bole damage. Canadian Journal of
Forest Research, 16, 1175-1179.
Pierce, L. L., Running, S. W., & Walker, J. (1994). Regional-scale relationships of leaf
area index to specific leaf area and leaf nitrogen content. Ecological
Applications, 4(2), 313-321.
Platt, W. J., Evans, G. W., & Rathbun, S. L. (1988). The population dynamics of a long-
lived conifer (Pinus palustris). The American Naturalist, 131(4), 491-525.
Pook, E. W. (1985). Canopy dynamics of Eucalyptus maculata Hook. III Effects of
drought. Australian Journal of Botany, 33(1), 65-79.
Pook, E. W., Costin, A. B., & Moore, C. W. E. (1966). Water stress in native vegetation
during the drought of 1965. Australian Journal of Botany, 14, 257-267.
Porteners, M. F. (1993). The natural vegetation of the Hay Plain:Booligal-Hay and
Deniliquin-Bendigo 1:250 000 maps. Cunninghamia, 3(1), 1-122.
Porteners, M. F., Ashby, E. M., & Benson, J. S. (1997). The natural vegetation of
Pooncarie 1:250 000 map. Cunninghamia, 5(1), 139-231.
Prior, L. D., Bowman, D. M. J. S., & Brook, B. W. (2007). Growth and survival of two
north Australian relictual tree species, Allosyncarpia ternata (Myrtaceae) and
Callitris intratropica (Cupressaceae). Ecological Research, 22(2), 228-236.
Prior, L. D., Eamus, D., & Bowman, D. M. J. S. (2003). Leaf attributes in the seasonally
dry tropics: a comparison of four habitats in northern Australia. Functional
Ecology, 17, 504-515.
Pugnaire, F. I., & Luque, M. T. (2001). Changes in plant interactions along a gradient of
environmental stress. Oikos, 93(1), 42-49.
Pulsford, I. F., Banks, J. C. G., Hodges, L., & Good, R. B. (1992). Land use history of
the white cypress pine forests in the snowy valley, Kosciusko National Park. In
P. Grenier (Ed.), Les Alpes Australiennse Review De Geographie Alpine (Vol.
Tome LXXX, pp. 86-104). Grenoble: Institut de Geographie Alpine.

References
121
Pye, M. G., Gadek, P. A., & Edwards, K. J. (2003). Divergence, diversity and species of
the Australasian Callitris (Cupressaceae) and allied genera: evidence from ITS
sequence data. Australian Systematic Botany, 16(4), 505-514.
Quinn, G. P., & Keough, M. J. (2002). Experimental design and data analysis for
biologists (1st ed.). Cambridge: Cambridge University Press.
Read, J. (1995). Recruitment characteristics of the white cypress pine (Callitris
glaucophylla) in arid South Australia. The Rangeland Journal, 17(2), 228-240.
Rebertus, A. J., Williamson, G. B., & Moser, E. B. (1989). Longleaf pine pyrogenicity
and turkey oak mortality in Florida xeric sandhills. Ecology, 70(1), 60-70.
Reich, P. B., Ellsworth, D. S., Walters, M. B., Vose, J. M., Gresham, C., Volin, J. C., et
al. (1999). Generality of leaf trait relationships:a test across six biomes. Ecology,
80(6), 1955-1969.
Reich, P. B., Walters, M. B., & Ellsworth, D. S. (1997). From tropics to tundra:global
convergence in plant functioning. Proceedings of National Academy of Sciences,
USA, 94, 13730-13734.
Reid, N., & Landsberg, J. (2000). Tree decline in agricultural landscapes:what we stand
to lose. In R. J. Hobbs & J. Yates Colin (Eds.), Temperate Eucalypt Woodlands
in Australia:Biology,Conservation, Management and Restoration (pp. 127-166).
Sydney: Surrey Beatty and Sons.
Richardson, D. M., & Bond, W. J. (1991). Determinants of plant distribution: evidence
from pine invasions. The American Naturalist, 137(5), 639-668.
Roderick, M. L., Berry, S. L., & Noble, I. R. (2000). A framework for understanding the
relationship between environment and vegetation on the surface area to volume
ratio of leaves. Functional Ecology, 14, 423-437.
Rodrigo, A., Quintana, V., & Retana, J. (2007). Fire reduces Pinus pinea distribution in
the northeastern Iberian peninsular. Ecoscience, 14(1), 23-30.
Rodrigo, A., Retana, J., & Pico, F. X. (2004). Direct regenerationis not the only
response of mediterranean forests to large fires. Ecology, 85(3), 716-729.
Russell-Smith, J. (2006). Recruitment dynamics of the long-lived obligate seeders
Callitris intratropica (Cupressaceae) and Petraeomyrtus punicea (Myrtaceae).
Australian Journal of Botany, 54(5), 479-485.
Sankaran, M., Hanan, N. P., Scholes, R. J., Ratnam, J., Augustine, D. J., Cade, B. S., et
al. (2005). Determinants of woody cover in African savannas. Nature,
438(7069), 846-849.

References
122
Sankaran, M., Ratnam, J., & Hanan, N. (2008). Woody cover in African savannas: the
role of resources, fire and herbivory. Global Ecology and Biogeography, 17(2),
236-245.
Scanlan, J. C., Berman, D. M., & Grant, W. E. (2006). Population dynamics of the
European rabbit (Oryctolagus cuniculus) in north eastern Australia: Simulated
responses to control. Ecological Modelling, 196(1-2), 221-236.
Scarff, F. R., & Westoby, M. (2006). Leaf litter flammability in some semi-arid
Australian woodlands. Functional Ecology, 20, 745-752.
Scheffer, M., Carpenter, S., Foley, J. A., Folke, C., & Walker, B. (2001). Catastrophic
shifts in ecosystems. Nature, 413(6856), 591-596.
Scholes, R. J., & Archer, S. R. (1997). Tree-grass interactions in savannas. Annual
Review of Ecology and Systematics, 28(1), 517-544.
Schreiber, E.S.G., Bearlin, A.R., Nicol, S.J. & Todd, C.R. 2004. Adaptive
management: a synthesis of current understanding and effective
application. Ecological Management & Restoration 5: 177-182.
Schulze, E., Kelliher, F. M., Korner, C., Lloyd, J., & Leuning, R. (1994). Relationships
among maximum stomatal conductance, ecosystem surface conductance, carbon
assimilation rate, and plant nitrogen nutrition: a global ecology scaling exercise.
Annual Review of Ecology and Systematics, 25(1), 629-662.
Schulze, E. D., Williams, R. J., Farquhar, G. D., Schulze, W., Langridge, J., Miller, J.
M., et al. (1998). Carbon and nitrogen isotope discrimination and nitrogen
nutrition of trees along a rainfall gradient in northern Australia. Functional Plant
Biology, 25(4), 413-425.
Schwilk, D. W. (2003). Flammability is a niche construction trait: canopy architecture
affects fire intensity. The American Naturalist, 162(6), 725-733.
Schwilk, D. W., & Ackerly, D. D. (2001). Flammability and serotiny as
strategies:correlated evolution in pines. Oikos, 94, 326-336.
Schwinning, S., & Sala, O. E. (2004). Hierarchy of responses to resource pulses in arid
and semi-arid ecosystems. Oecologia, 141(2), 211-220.
Schwinning, S., Sala, O. E., Loik, M. E., & Ehleringer, J. R. (2004). Thresholds,
memory, and seasonality: understanding pulse dynamics in arid/semi-arid
ecosystems. Oecologia, 141(2), 191-193.
Scott, J. A. (1992). The natural vegetation of the Balranald-Swan Hill area.
Cunninghamia, 2(4), 597-652.

References
123
Shinneman, D. J., & Baker, W. L. (2009). Historical fire and multidecadal drought as
context for pinon juniper woodland restoration in western Colorado. Ecological
Applications, 19(5), 1231-1245.
Sivertsen, D., & Clarke, P. J. (2000). Temperate woodlands in New South Wales:a brief
overview of distribution, composition and conservation. In R. J. Hobbs & J.
Yates Colin (Eds.), Temperate Eucalypt Woodlands in
Australia:Biology,Conservation, Management and Restoration (pp. 6-16).
Sydney: Surrey Beatty and Sons.
Smith, R. E., van Wilgen Brian, W., Forsyth, G. G., & Richardson, D. M. (1992).
Coexistence of seeders and resprouters in a fire-prone environment:the role of
ecophysiology and soil moisture. In W. van Wilgen Brian, D. M. Richardson, F.
J. Kruger & H. J. van Hensbergen (Eds.), Fire in South Africa Mountain Fynbos
(pp. 108-122). Berlin: Springer-Verlag.
Sparrow, A. D., & Bellingham, P. J. (2001). More to resprouting than fire. Oikos, 94(1),
195-197.
Specht, R. L. (1981). Responses to fires of heathlands and related shrublands. In A. M.
Gill, R. H. Groves & I. R. Noble (Eds.), Fire and the Australian Biota.
Canberra: Australian Academy of Science.
Specht, R. L., & Specht, A. (1999). Australian Plant Communities:dynamics of
structure, growth and biodiversity. Melbourne: Oxford University Press.
Squeo, F. A., Holmgren, M., Jimenez, M., Alban, L., Reyes, J., & Gutierrez, J. R.
(2007). Tree establishment along an ENSO experimental gradient in the
Atacama desert. Journal of Vegetation Science, 18(2), 195-202.
Steel, G. D., & Torrie, J. H. (1960). Principles and Procedures of Statistics with Special
Reference to Biologivcal Sciences. London: McGraw-Hill Book Company, Inc.
Suding, K. N., & Hobbs, R. J. (2009). Threshold models in restoration and
conservation: a developing framework. Trends in Ecology & Evolution, 24(5),
271-279.
Tabachnick, B. G., & Fidell, L. S. (2007). Using Multivariate Statistics (5th ed.).
Boston: Pearson Education Inc.
Thomas, C. D., Cameron, A., Green, R. E., Bakkenes, M., Beaumont, L. J., Collingham,
Y. C., et al. (2004). Extinction risk from climate change. Nature, 427(6970),
145-148.

References
124
Thompson, J. R., & Spies, T. A. (2009). Vegetation and weather explain variation in
crown damage within a large mixed-severity wildfire. Forest Ecology and
Management, 258(7), 1684-1694.
Thompson, W. A., & Eldridge, D. A. (2005). White cypress pine (Callitris
glaucophylla):a review of its roles in landscape and ecological processes in
eastern Australia. Australian Journal of Botany, 53, 555-570.
Thuiller, W., Araujo, M. B., & Lavorel, S. (2003). Generalized models vs. classification
tree analysis: predicting spatial distributions of plant species at different scales.
Journal of Vegetation Science, 14(5), 669-680.
Tilman, D. (1988). Plant Strategies:Dynamics and Structure of Plant Communities.
Princeton: Princeton University Press.
Tiver, F., & Andrew, M. H. (1997). Relative effects of herbivory by sheep, rabbits,
goats and kangaroos on recruitment and regeneration of shrubs and trees in
eastern South Australia. Journal of Applied Ecology, 34(4), 903-914.
Trabaud, L. (1991). Fire regimes and phytomass growth dynamics in a Quercus
coccifera garrigue. Journal of Vegetation Science, 2(3), 307-314.
Turner, M. G., Baker, W. L., Peterson, C. J., & Peet, R. K. (1998). Factors Influencing
Succession: Lessons from Large, Infrequent Natural Disturbances. Ecosystems,
1(6), 511-523.
Turner, M. G., Romme, W. H., & Gardner, R. H. (1999). Prefire heterogeneity, fire
severity, and early postfire plant reestablishment in subalpine forests of
Yellowstone National Park, Wyoming. International Journal of Wildland Fire,
9(1), 21-36.
Turner, M. G., Romme, W. H., & Tinker, D. B. (2003). Surprises and lessons from the
1988 Yellowstone fires. Frontiers in Ecology and Environment, 1(7), 351-358.
van de Koppel, J., Rietkerk, M., & Weissing, F. J. (1997). Catastrophic vegetation shifts
and soil degradation in terrestrial grazing systems. Trends in Ecology &
Evolution, 12(9), 352-356.
van Dyne, G. M., Brokington, N. R., Sczocs, Z., Duek, J., & Ribic, C. A. (1980). Large
herbivore subsystem. In A. I. Breymeyer & G. M. van Dyne (Eds.), Grasslands,
Systems analysis and Man (ist ed., pp. 270-516). Cambridge: Cambridge
University Press.
Van Wagner, C. E. (1973). Height of crown scorch in forest fires. Canadian Journal of
Forest Research, 3, 373-378.

References
125
van Wilgen, B. W., Higgins, K. B., & Bellstedt, D. U. (1990). The role of vegetation
structure and fuel chemistry in excluding fire from forest patches in the fire-
prone fynbos shrublands of South Africa. Journal of Ecology, 78, 210-222.
Veall, M. R., & Zimmermann, K. F. (1996). Pseudo-R2 measures for some limited
dependent variable models Journal of Economic Surveys, 10, 241-259.
Veblen, T. T. (1992). Regeneration dynamics. In D. C. Glenn-Lewin, R. K. Peet & H.
Veblen Thomas (Eds.), Plant Succession: Theory and Prediction (ist ed., pp.
152-187). London: Chapman & Hall.
Venables, W. N., & Ripley, B. D. (2002). Statistics and Computing (4th ed.). New
York: Springer-Verlag.
Vesk, P., A. , Leishman, M., R. , & Westoby, M. (2004). Simple traits do not predict
grazing response in Australian dry shrublands and woodlands. Journal of
Applied Ecology, 41(1), 22-31.
Vesk, P. A., & Westoby, M. (2003). Drought damage and recovery-a conceptual model.
New Phytologist, 160, 7-14.
Vilagrosa, A., Cortina, J., Gil-PelegrÃn, E., & Bellot, J. (2003). Suitability of drought-
Preconditioning techniques in mediterranean climate. Restoration Ecology,
11(2), 208-216.
Walker, B. H. (1993). Rangeland ecology: understanding and managing change. Ambio,
22(2/3), 80-87.
Wardell-Johnson Grant, W., Williams, J. E., Hill, K. D., Cumming, R., & Woinarski, J.
(1997). Evolutionary biogeography and contemporary distribution of eucalypts.
In J. Williams (Ed.), Eucalypt Ecology:Individuals to Ecosystems (pp. 92-128).
Cambridge: Cambridge University Press.
Warner, R. R., & Chesson, P. L. (1985). Coexistence mediated by recruitment
fluctuations:a field guide to the storage effect. The American Naturalist, 125(6),
769-787.
Watson, I. W., Westoby, M., & Holm, A. M. (1997). Continuous and episodic
components of demographic change in arid zone shrubs:models of two
Eremophila species from Western Australia compared with published data on
other species. Journal of Ecology, 85, 833-846.
Wells, P. M., & Hill, R. S. (1989). Fossil imbricate-leaved Podocarpaceae from Tertiary
sediments in Tasmania. Australian Systematic Botany, 2, 387-423.
Westoby, M. (1999). The LHS scheme in relation to grazing and fire. Proceedings of
6th International Rangeland Congress, Townsville, 2, 893-896.

References
126
Westoby, M., Walker, B., & Noy-Meir, I. (1989). Opportunistic management for
rangelands not at equilibrium. Journal of Range Management, 42(4), 266-274.
Whelan, R. J. (1995). The ecology of fire. Cambridge: Cambridge University Press.
Whelan, R. J., De Jong, N. H., & Von Der Burg, S. (1998). Variation in bradyspory and
seedling recruitment without fire among populations of Banksia serrata
(Proteaceae). Australian Journal of Botany, 23, 121-128.
Whipp, R. K. (2009). Historical vegetation change in relation to forest management in
the Pilliga State Forests of northern NSW, Australia. . Unpublished PhD thesis,
Charles Sturt University, Albury.
Wiegand, K., Jeltsch, F., & Ward, D. (2004). Minimum recruitment frequency in plants
with episodic recruitment. Oecologia, 141(2), 363-372.
Williams, J., & Woinarski, J. (1997). Eucalypt Ecology:Individuals to Ecosystems.
Cambridge: Cambridge University Press.
Williams, J. E., & Brooker, I. H. (1997). Eucalypts:an introduction. In J. Williams & J.
Woinarski (Eds.), Eucalypt Ecology:Individuals to Ecosystems (pp. 1-15).
Cambridge: Cambridge University Press.
Williams, J. E., Whelan, R. J., & Gill, A. M. (1994). Fire and environmental
heterogeneity in southern temperate forest ecosystems: implications for
management. Australian Journal of Botany, 42(2), 125-137.
Williamson, G. B., & Black, E. M. (1981). High temperature of forest fires under pines
as a selective advantage over oaks. Nature, 293(5834), 643-644.
Wilson, A. D. (1990). The effect of grazing on Australian ecosystems. Proceedings of
Ecological Society of Australia, 16, 235-244.
Wright, C. S., & Agee, J. K. (2004). Fire and vegetation history in the Eastern Cascade
Mountains, Washington. Ecological Applications, 14(2), 443-459.
Wright, I. J., Reich, P. B., & Westoby, M. (2001). Strategy shifts in leaf physiology,
structure and nutrient content between species of high- and low-rainfall and
high- and low-nutrient habitats. Functional Ecology, 15, 423-434.
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al.
(2004). The worldwide leaf economics spectrum. Nature, 428, 821-827.
Zavala, M. A., & Zea, E. (2004). Mechanisms maintaining biodiversity in
Mediterranean pine-oak forests: insights from a spatial simulation model. Plant
Ecology, 171, 197-207.

References
127
Zeppel, M., & Eamus, D. (2008). Coordination of leaf area, sapwood area and canopy
conductance leads to species convergence of tree water use in a remnant
evergreen woodland. Australian Journal of Botany, 56(2), 97-108.


















