
Host usage and seasonal activity patterns of Ixodes kingi...
13
Journal of Vector Ecology June 2006 168 Host usage and seasonal activity patterns of Ixodes kingi and I. sculptus (Acari: Ixodidae) nymphs in a Colorado prairie landscape, with a summary of published North American host records for all life stages Daniel J. Salkeld 1,2 , Rebecca J. Eisen 3 , Michael F. Antolin 4 , Paul Stapp 1 , and Lars Eisen 5 1 Department of Biological Science, California State University, Fullerton, CA 92834, U.S.A. 2 The World Conservation Union, Washington, D.C. 20009, U.S.A. 3 Division of Vector-Borne Infectious Diseases, National Center for Infectious Diseases, Centers for Disease Control and Prevention, Fort Collins, CO 80522, U.S.A. 4 Department of Biology, Colorado State University, Fort Collins, CO 80523, U.S.A. 5 Department of Microbiology, Immunology, and Pathology, Colorado State University, Fort Collins, CO 80523, U.S.A. Received 11 January 2006; Accepted 8 March 2006 ABSTRACT: We examined host usage and seasonal activity patterns of the nymphal stage of the ticks Ixodes kingi and I. sculptus within a prairie rodent community in north-central Colorado. Ixodes kingi was commonly encountered on both northern grasshopper mice (Onychomys leucogaster) and thirteen-lined ground squirrels (Spermophilus tridecemlineatus), whereas I. sculptus frequently infested S. tridecemlineatus but was absent from O. leucogaster. Low numbers of ticks of both species were collected from deer mice (Peromyscus maniculatus) and Ord’s kangaroo rats (Dipodomys ordii). Nymphal loads of I. kingi and I. sculptus increased dramatically on commonly infested rodent species from spring (May-June) to summer (July-August). Further, rodents trapped on prairie-dog towns tended to experience increased nymphal loads of I. kingi (O. leucogaster, S. tridecemlineatus) but decreased loads of I. sculptus (S. tridecemlineatus) following plague epizootics among prairie dog populations. A summary of published North American host records revealed that I. kingi has been recorded from humans, domestic animals (cat, dog), 17 species of carnivores, 40 species of rodents, and four species of lagomorphs, and that I. sculptus has been recorded from humans, domestic animals (cat, dog, goat), 13 species of carnivores, 34 species of rodents, and three species of lagomorphs. In accordance with our observations from Colorado, I. kingi commonly has been found to infest heteromyid and murid rodents (such as grasshopper mice), whereas I. sculptus most frequently has been collected from ground-dwelling sciurid rodents, especially Spermophilus ground squirrels. The potential roles of I. kingi and I. sculptus as enzootic vectors of human pathogens, particularly the agents of tularemia (Francisella tularensis), Q fever (Coxiella burnetii), and Colorado tick fever (CTF virus), are discussed. Journal of Vector Ecology 31 (1): 168-180. 2006. Keyword Index: Ixodes kingi, Ixodes sculptus, host records, rodents, seasonality. INTRODUCTION The ixodid ticks Ixodes kingi and I. sculptus occur throughout western North America and are occasionally encountered also in the east (Bishopp and Trembley 1945, Cooley and Kohls 1945, Gregson 1956, Durden and Keirans 1996, Allan 2001). As these ticks only rarely infest humans (Bishopp and Trembley 1945, Furman and Loomis 1984, Walker et al. 1998, Merten and Durden 2000) and neither species has been clearly implicated as an enzootic vector of a human disease agent, they have received limited attention in recent decades. The causative agents of tularemia, Colorado tick fever, and Q fever have been recorded from I. kingi or I. sculptus (Sidwell et al. 1964, Thorpe et al. 1965, CDC 1976), but their vector competency for these pathogens remains unclear. Ixodes kingi and I. sculptus are commonly found in prairie habitats (Gregson 1956) but both species also have been recorded from shrubland and forested habitats (Allred et al. 1960, Beck et al. 1963, Carey et al. 1980). Both ticks commonly infest rodents as well as carnivores (see summary of published North American host records in Table 2). Several previous studies have provided quantitative data on tick infestation of rodents (Holdenried et al. 1951, Emmons 1966, Johnson 1966, Allred 1968, Hilton and Mahrt 1971, Lane et al. 1981, Spicka 1981, Kietzmann 1987, Kietzmann and Kietzmann 1987, Brillhart et al. 1994) but studies comparing host usage of I. kingi and I. sculptus within a single rodent community have been lacking. Earlier data on the seasonal activity patterns of these ticks have, with a few exceptions (Holdenried et al. 1951, Beck et al. 1963), been qualitative (Bishopp and Trembley 1945, Linsdale 1946, Allred et al. 1960, Egoscue 1962, Johnson 1966, Allred 1968, Wittrock and Wilson 1974, Furman and Loomis 1984). The primary aims of this study were to compare the host usage of I. kingi and I. sculptus within a prairie rodent community, to describe their seasonal activity patterns, and to summarize the published North American host records for these ticks.
Transcript of Host usage and seasonal activity patterns of Ixodes kingi...

Journal of Vector Ecology June 2006168
Host usage and seasonal activity patterns of Ixodes kingi and I. sculptus (Acari: Ixodidae) nymphs in a Colorado prairie landscape, with a summary of published
North American host records for all life stages
Daniel J. Salkeld1,2, Rebecca J. Eisen3, Michael F. Antolin4, Paul Stapp1, and Lars Eisen5
1Department of Biological Science, California State University, Fullerton, CA 92834, U.S.A.2The World Conservation Union, Washington, D.C. 20009, U.S.A.
3Division of Vector-Borne Infectious Diseases, National Center for Infectious Diseases, Centers for Disease Control and Prevention, Fort Collins, CO 80522, U.S.A.
4Department of Biology, Colorado State University, Fort Collins, CO 80523, U.S.A.5Department of Microbiology, Immunology, and Pathology, Colorado State University, Fort Collins, CO 80523, U.S.A.
Received 11 January 2006; Accepted 8 March 2006
ABSTRACT: We examined host usage and seasonal activity patterns of the nymphal stage of the ticks Ixodes kingi and I. sculptus within a prairie rodent community in north-central Colorado. Ixodes kingi was commonly encountered on both northern grasshopper mice (Onychomys leucogaster) and thirteen-lined ground squirrels (Spermophilus tridecemlineatus), whereas I. sculptus frequently infested S. tridecemlineatus but was absent from O. leucogaster. Low numbers of ticks of both species were collected from deer mice (Peromyscus maniculatus) and Ord’s kangaroo rats (Dipodomys ordii). Nymphal loads of I. kingi and I. sculptus increased dramatically on commonly infested rodent species from spring (May-June) to summer (July-August). Further, rodents trapped on prairie-dog towns tended to experience increased nymphal loads of I. kingi (O. leucogaster, S. tridecemlineatus) but decreased loads of I. sculptus (S. tridecemlineatus) following plague epizootics among prairie dog populations. A summary of published North American host records revealed that I. kingi has been recorded from humans, domestic animals (cat, dog), 17 species of carnivores, 40 species of rodents, and four species of lagomorphs, and that I. sculptus has been recorded from humans, domestic animals (cat, dog, goat), 13 species of carnivores, 34 species of rodents, and three species of lagomorphs. In accordance with our observations from Colorado, I. kingi commonly has been found to infest heteromyid and murid rodents (such as grasshopper mice), whereas I. sculptus most frequently has been collected from ground-dwelling sciurid rodents, especially Spermophilus ground squirrels. The potential roles of I. kingi and I. sculptus as enzootic vectors of human pathogens, particularly the agents of tularemia (Francisella tularensis), Q fever (Coxiella burnetii), and Colorado tick fever (CTF virus), are discussed. Journal of Vector Ecology 31 (1): 168-180. 2006.
Keyword Index: Ixodes kingi, Ixodes sculptus, host records, rodents, seasonality.
INTRODUCTION
The ixodid ticks Ixodes kingi and I. sculptus occur throughout western North America and are occasionally encountered also in the east (Bishopp and Trembley 1945, Cooley and Kohls 1945, Gregson 1956, Durden and Keirans 1996, Allan 2001). As these ticks only rarely infest humans (Bishopp and Trembley 1945, Furman and Loomis 1984, Walker et al. 1998, Merten and Durden 2000) and neither species has been clearly implicated as an enzootic vector of a human disease agent, they have received limited attention in recent decades. The causative agents of tularemia, Colorado tick fever, and Q fever have been recorded from I. kingi or I. sculptus (Sidwell et al. 1964, Thorpe et al. 1965, CDC 1976), but their vector competency for these pathogens remains unclear. Ixodes kingi and I. sculptus are commonly found in prairie habitats (Gregson 1956) but both species also have been recorded from shrubland and forested habitats (Allred et al. 1960, Beck et al. 1963, Carey et al. 1980). Both ticks
commonly infest rodents as well as carnivores (see summary of published North American host records in Table 2). Several previous studies have provided quantitative data on tick infestation of rodents (Holdenried et al. 1951, Emmons 1966, Johnson 1966, Allred 1968, Hilton and Mahrt 1971, Lane et al. 1981, Spicka 1981, Kietzmann 1987, Kietzmann and Kietzmann 1987, Brillhart et al. 1994) but studies comparing host usage of I. kingi and I. sculptus within a single rodent community have been lacking. Earlier data on the seasonal activity patterns of these ticks have, with a few exceptions (Holdenried et al. 1951, Beck et al. 1963), been qualitative (Bishopp and Trembley 1945, Linsdale 1946, Allred et al. 1960, Egoscue 1962, Johnson 1966, Allred 1968, Wittrock and Wilson 1974, Furman and Loomis 1984). The primary aims of this study were to compare the host usage of I. kingi and I. sculptus within a prairie rodent community, to describe their seasonal activity patterns, and to summarize the published North American host records for these ticks.

Journal of Vector Ecology 169Vol. 31, no. 1
MATERIALS AND METHODS
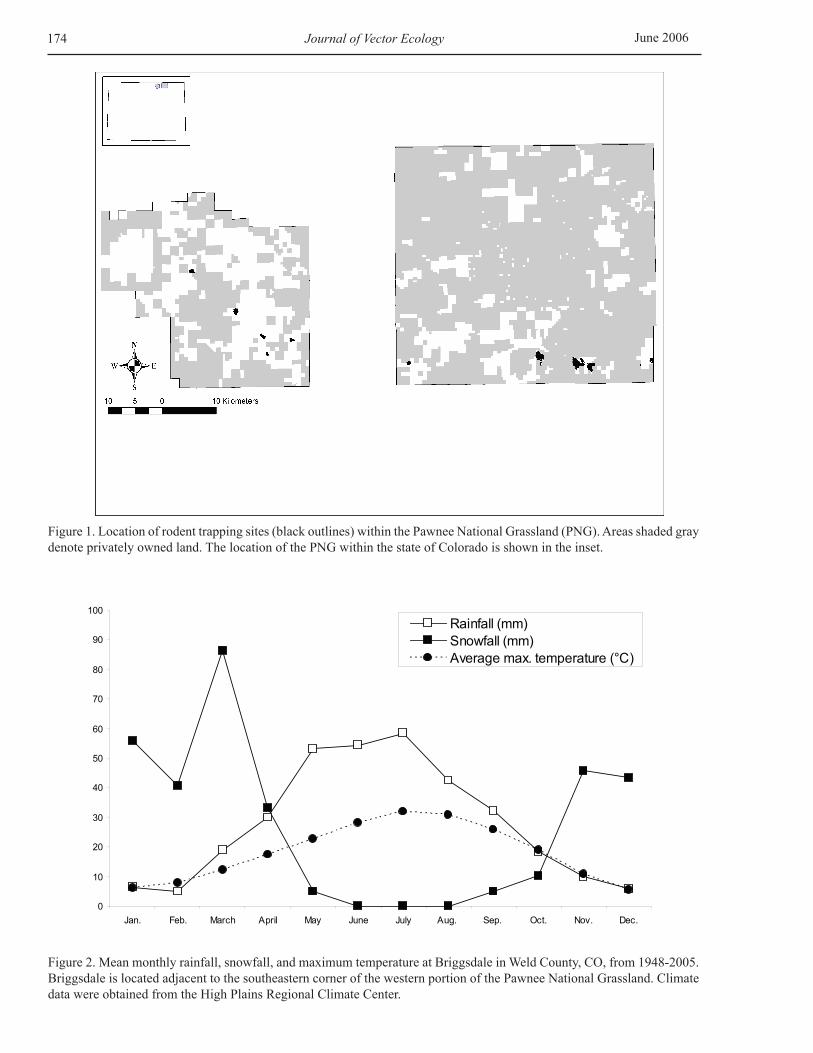
General description of study area The study was conducted within the Pawnee National Grassland (PNG), which is located in Weld County, north-central Colorado (Figure 1). The 80,000 ha PNG ranges in elevation from 1,280-1,950 m and is characterized by a checkerboard of publicly and privately owned land (Figure 1). The climate is semi-arid, with the majority of the annual rainfall (≈320 mm) occurring during the April-September growing season (Figure 2). Short-grass steppe, with intermittent streams, is the predominant habitat type of the PNG. Native vegetation includes blue grama (Bouteloua gracilis), buffalo grass (Buchloe dactyloides), and prickly pear cactus (Opuntia polyacantha), with occasional stands of shrubs on hillsides (Yucca glauca, Chrysothamnus nauseosus, Gutierrezia sarothrae) and floodplains (Atriplex canescens), and with plains cottonwoods (Populus sargentii) lining the few permanent streams.
Rodent trapping, tick collection, and tick identification Small mammals were live-trapped during May-August 2005 within multiple sites on the PNG (Figure 1). Trapping was conducted at sites without presence of black-tailed prairie dog (Cynomys ludovicianus) towns (six sites, total of six trapping grids), sites with uninhabited prairie dog towns because of plague epizootics during the previous year (four sites, total of eight grids), sites with active prairie dog towns free of plague during the year of our trapping (four sites, total of 12 grids), and sites with active prairie dog towns experiencing plague epizootics during the year of our trapping (three sites, total of seven grids). Black-tailed prairie dogs are highly susceptible to infection with the plague bacterium Yersinia pestis, with afflicted prairie dog towns typically suffering near 100% mortality (Cully and Williams 2001, Antolin et al. 2002). All sites were trapped on two separate occasions between May and August, with the exception that the three sites where plague epizootics occurred during 2005 were sampled only from July onwards. Each 2.25 ha trapping grid (150 x 150 m area with traps spaced 15 m apart) held 100 Sherman live-traps (H.B. Sherman Traps Inc., Tallahassee, FL) baited with a mixture of peanut butter and oats. Traps were operated from dusk to mid-morning over four consecutive nights per trapping occasion; animals were recovered from the traps at first light (nocturnal animals) and during mid-morning (diurnally active animals). Trapped animals were anesthetized, weighed and measured, identified to species, sex, age and reproductive status, ear-tagged (National Band and Tag Co., Newport, KY), and examined for ectoparasites (fleas and ticks). Examination of animals for ticks included both careful inspection of the head area and brushing of the body to uncover ticks hidden in the fur. All collected ticks were preserved in 95% ethanol for identification. Ticks were identified to species using published keys (Furman and Loomis 1984, Durden and Keirans 1996) and a tick reference collection made available by M.C. Dolan and J.F. Piesman of the Centers for Disease Control and Prevention, Fort Collins, CO. Some nymphal ticks (n =
10) were too damaged to be identified with certainty; these ticks were not included in the data presented. A surprisingly low number of larval ticks was recovered (n = 20). Of these, six were too damaged to be identified to species. Further, we cannot rule out the possibility that the miniscule larval tick stage was overlooked on the rodents because of the time constraint imposed on the tick detection effort by additional rodent processing procedures (flea detection, blood sampling). Based on these considerations, we have restricted the presentation of larval data to a summary sentence. All statistical analyses were carried out using version 5.1 of JMP® (Sall et al. 2005) and results were considered significant when P < 0.05.
Summary of published North American host records for I. kingi and I. sculptus Published North American host records for I. kingi and I. sculptus were compiled using a combination of computerized searches (OVID Medline, Biological Abstracts) for records from the mid-1960s to the present time and an extensive manual literature search to locate older works with host records of either I. kingi or I. sculptus. This included all studies mentioning these ticks in the 1974 Index Catalogue of Medical and Veterinary Zoology (Doss et al. 1974). Because many references to I. kingi or I. sculptus are given merely as side notes in studies focusing on the ecology of other ectoparasites, mammals, or disease agents, it should be noted that a few published host records may have escaped us. The data presented in Table 2 are broken down by host species, tick species, and tick life stage (larva, nymph, adult, or unknown life stage). Ixodes kingi includes records for the synonym I. pratti. Taxonomic structure and scientific names for North American mammals follow the system used by Wilson and Ruff (1999). We have included only host records for which it was reasonably clear how the common or scientific name used for that particular host corresponds to current nomenclature. Reports that only identified host genus are included only when the data provide new records for a tick species or life stage not evident from the mammalian species records within that particular genus. Data presented for I. sculptus by Chamberlin (1937) were disregarded as they clearly were confused with I. scapularis.
Animal health Animal capture and handling procedures followed guidelines provided by the American Society of Mammalogists (1998) and were approved by the Institutional Animal Care Committee of California State University, Fullerton, and the Colorado Division of Wildlife.
RESULTS
Host usage by I. kingi and I. sculptus in a northern Colorado prairie rodent community During May-August 2005, we recovered a total of 83 I. kingi and 34 I. sculptus nymphs from four rodent species: Ord’s kangaroo rat (Dipodomys ordii), the northern grasshopper mouse (Onychomys leucogaster), the deer

Journal of Vector Ecology June 2006170
mouse (Peromyscus maniculatus), and the thirteen-lined ground squirrel (Spermophilus tridecemlineatus) (Table 1). No ticks were recovered from two rodent species with ≤10 animals examined (Chaetodipus hispidus, Reithrodontomys megalotis). Of 14 collected larval ticks that could be identified to species, 12 were I. kingi (nine larvae recovered from eight northern grasshopper mice and three larvae from a single Ord’s kangaroo rat) and the remaining two were I. sculptus (recovered from one northern grasshopper mouse and one thirteen-lined ground squirrel). To the best of our knowledge, we present the first records of infestation of D. ordii and O. leucogaster by I. sculptus and the first record of I. kingi from S. tridecemlineatus. Incidental collection of ticks from two black-tailed prairie dogs (Cynomys ludovicianus) yielded an additional five I. sculptus nymphs. The thirteen-lined ground squirrel accounted for 66% of all I. kingi nymphs and 94% of I. sculptus nymphs collected (Table 1). We found no significant site-related differences in nymphal infestation rates or nymphal loads for either I. kingi or I. sculptus on this rodent during the July-August peak tick infestation period (data not shown, P > 0.30 in all four cases). Because no significant site-related differences in tick infestation rate or tick load were found for the most commonly infested rodent on the PNG, tick infestation data for a particular rodent species were pooled for all PNG sampling sites in the host usage analyses described below. Statistical analyses are restricted to data from the peak July-August tick infestation period (see Figure 3). The peak infestation rate by I. kingi during July-August was highest for thirteen-lined ground squirrels (6.7% of examined hosts infested) and northern grasshopper mice (6.5%), followed by deer mice (1.8%) and Ord’s kangaroo rats (0%) (Table 1). In contrast, the peak infestation rate by I. sculptus was highest for thirteen-lined ground squirrels (6.0%), followed by deer mice (1.8%) and Ord’s kangaroo rats and northern grasshopper mice (0% in both cases) (Table 1). Comparisons between the two most commonly captured rodent species, the thirteen-lined ground squirrel and the northern grasshopper mouse, showed that I. sculptus was encountered more frequently on ground squirrels than on grasshopper mice (6.0 versus 0%; Fisher’s exact 2-tailed test, P = 0.005) whereas I. kingi was equally likely to infest both rodents (6.7 versus 6.5%; P = 1.00). Further, northern grasshopper mice were more likely to be infested with I. kingi than I. sculptus (6.5 versus 0%; P = 0.007) whereas no significant difference was seen for thirteen-lined ground squirrels (6.7 versus 6.0%; P = 1.00). Potential differences in infestation rates between male and female or adult and juvenile rodents were not explored because of the small number of nymphal ticks collected. No individual animal was infested by more than eight I. kingi nymphs or 11 I. sculptus nymphs and the mean loads of I. kingi or I. sculptus nymphs during May-August were low for all rodent species examined (means of ≤0.21 per animal in all cases) (Table 1). Comparisons of median nymphal loads during the peak July-August infestation period for the two most commonly captured rodents, the thirteen-lined ground squirrel and the northern grasshopper mouse, revealed that
R
oden
t spe
cies
Num
ber o
f rod
ents
exa
min
ed /
Dip
odom
ys
Ony
chom
ys
Pero
mys
cus
Sper
mop
hilu
sIn
fest
atio
n by
nym
phal
tick
s or
dii
leuc
ogas
ter
man
icul
atus
tr
idec
emlin
eatu
sTo
tal n
o. in
divi
dual
rode
nts e
xam
ined
71
26
7 17
7 25
9To
tal n
o. I.
kin
gi /
I. sc
ulpt
us re
cove
red
3 / 1
24
/ 0
1 / 1
55
/ 32
Perc
enta
ge o
f tot
al n
o. I.
kin
gi /
I. sc
ulpt
us re
cove
red
4% /
3%
29%
/ 0%
1%
/ 3%
66
% /
94%
Infe
stat
ion
rate
by
I. ki
ngi /
I. sc
ulpt
us, M
ay-A
ug.a
2.8%
/ 1.
4%
4.9%
/ 0%
0.
6% /
0.6%
4.
3% /
3.5%
In
fest
atio
n ra
te b
y I.
king
i / I.
scul
ptus
, Jul
y-A
ug.b
0% /
0%
6.5%
/ 0%
1.
8% /
1.8%
6.
7% /
6.0%
M
ean
no. I
. kin
gi /
I. sc
ulpt
us p
er a
nim
al, M
ay-A
ug.a
0.03
/ 0.
01
0.06
/ 0
0.01
/ 0.
01
0.14
/ 0.
08M
ean
no. I
. kin
gi /
I. sc
ulpt
us p
er a
nim
al, J
uly-
Aug
.b 0
/ 0
0.08
/ 0
0.02
/ 0.
02
0.21
/ 0.
15M
ean
no. I
. kin
gi /
I. sc
ulpt
us p
er in
fest
ed a
nim
ala
1.0
/ 1.0
1.
3 / 0
1.
0 / 1
.0
3.3
/ 2.4
Med
ian
no. I
. kin
gi /
I. sc
ulpt
us p
er a
nim
ala
0 / 0
0
/ 0
0 / 0
0
/ 0R
ange
of I
. kin
gi /
I. sc
ulpt
us p
er a
nim
ala
0-1
/ 0-1
0-
3 / 0
0-
1 / 0
-1
0-8
/ 0-1
1a I
nclu
ding
tick
s col
lect
ed o
n th
e fir
st c
aptu
re o
f an
indi
vidu
al a
nim
al d
urin
g M
ay-A
ugus
t.b I
nclu
ding
tick
s col
lect
ed o
n th
e fir
st c
aptu
re o
f an
indi
vidu
al a
nim
al d
urin
g th
e pe
ak Ju
ly-A
ugus
t tic
k in
fest
atio
n pe
riod.
Tabl
e 1.
Infe
stat
ion
of ro
dent
s by
nym
phal
Ixod
es k
ingi
and
I. sc
ulpt
us in
a n
orth
ern
Col
orad
o pr
airie
land
scap
e.
Journal of Vector Ecology 171Vol. 31, no. 1
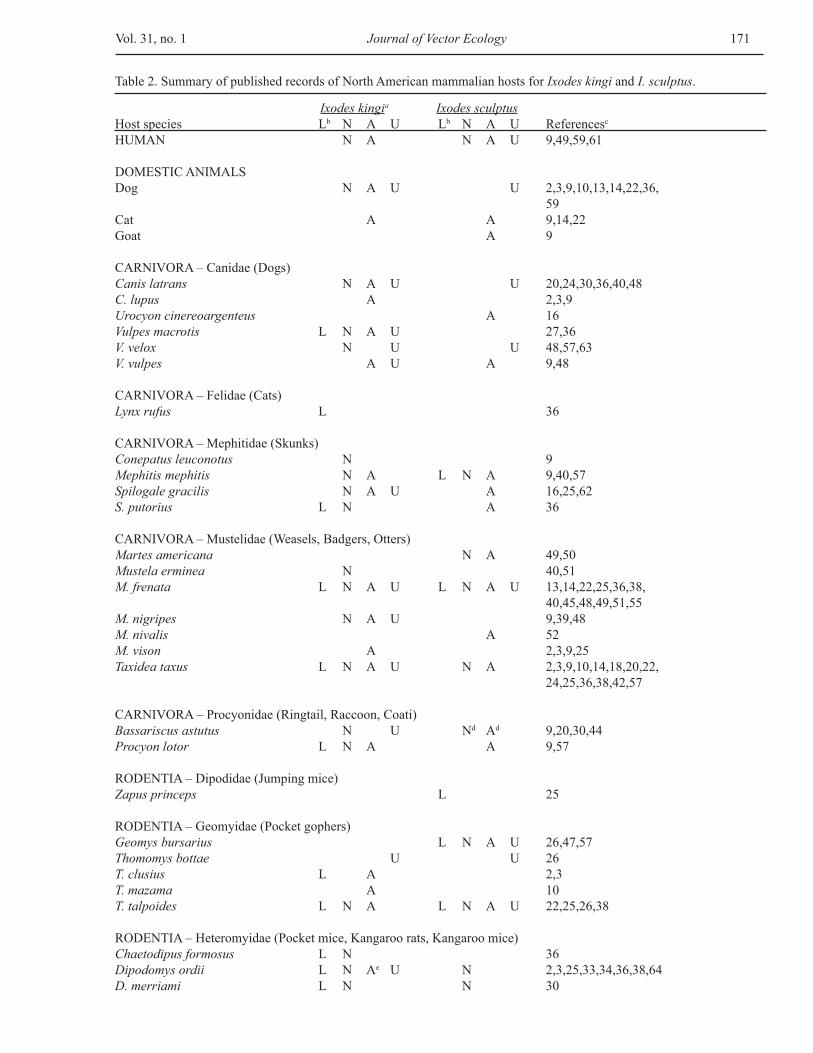
Ixodes kingia Ixodes sculptusHost species Lb N A U Lb N A U Referencesc
HUMAN N A N A U 9,49,59,61
DOMESTIC ANIMALSDog N A U U 2,3,9,10,13,14,22,36, 59Cat A A 9,14,22Goat A 9
CARNIVORA – Canidae (Dogs)Canis latrans N A U U 20,24,30,36,40,48C. lupus A 2,3,9Urocyon cinereoargenteus A 16Vulpes macrotis L N A U 27,36V. velox N U U 48,57,63V. vulpes A U A 9,48
CARNIVORA – Felidae (Cats)Lynx rufus L 36
CARNIVORA – Mephitidae (Skunks)Conepatus leuconotus N 9Mephitis mephitis N A L N A 9,40,57Spilogale gracilis N A U A 16,25,62S. putorius L N A 36
CARNIVORA – Mustelidae (Weasels, Badgers, Otters)Martes americana N A 49,50Mustela erminea N 40,51M. frenata L N A U L N A U 13,14,22,25,36,38, 40,45,48,49,51,55M. nigripes N A U 9,39,48M. nivalis A 52M. vison A 2,3,9,25Taxidea taxus L N A U N A 2,3,9,10,14,18,20,22, 24,25,36,38,42,57
CARNIVORA – Procyonidae (Ringtail, Raccoon, Coati)Bassariscus astutus N U Nd Ad 9,20,30,44Procyon lotor L N A A 9,57
RODENTIA – Dipodidae (Jumping mice)Zapus princeps L 25
RODENTIA – Geomyidae (Pocket gophers)Geomys bursarius L N A U 26,47,57Thomomys bottae U U 26T. clusius L A 2,3T. mazama A 10T. talpoides L N A L N A U 22,25,26,38
RODENTIA – Heteromyidae (Pocket mice, Kangaroo rats, Kangaroo mice)Chaetodipus formosus L N 36Dipodomys ordii L N Ae U N 2,3,25,33,34,36,38,64D. merriami L N N 30
Table 2. Summary of published records of North American mammalian hosts for Ixodes kingi and I. sculptus.

Journal of Vector Ecology June 2006172
RODENTIA – Heteromyidae (Pocket mice, Kangaroo rats, Kangaroo mice) continuedD. microps L N A U 25,29,30,36Microdipodops megacephalus L N 30,36Perognathus longimembris L N N 25,36P. parvus L N A U 6,25,30,36,38
RODENTIA – Muridae (Rats, Mice, Voles, Lemmings)Lemmiscus curtatus N 22Microtus longicaudus L N 25M. montanus N 25,38M. ochrogaster A 12Neotoma cinerea L N 36,38N. fuscipes L A 10,49N. lepida L N U 25,29,30,36Onychomys leucogaster L N A L 25,32,36,38,64O. torridus L N 25,30Peromyscus crinitus L N U 25,29,31,36P. eremicus L N 25P. leucopus N 13,56P. maniculatus L N A U N U 13,25,29,33,34,36, 38,45,48,51,64P. truei L N 25,36Phenacomys intermedius U 37Reithrodontomys megalotis L N 25,36
RODENTIA – Sciuridae (Squirrels, Chipmunks, Marmots, Prairie dogs)Cynomys gunnisoni N A U A U 4,10,17,25,28C. leucurus L N N A 10,25C. ludovicianus N A U 10,48,53,64C. parvidens N L 25Marmota flaviventris L N A L N 2,3,25M. monax N A N A 9,10Ammospermophilus leucurus L N A L N A 30,36Spermophilus armatus L L N A U 10,15,25S. beecheyi L N A 10,11,16,46,49,55,60S. beldingi N A 9,10,25,49S. brunneus U 58S. columbianus L N N A U 3,5,9,10,14,22S. elegans A N A 10S. franklinii L N A U 9,10,41,52S. lateralis N L N A 10,23,25,35,43,49,55S. richardsonii L N A U L N A U 8,10,14,19,22,25, 41,43S. spilosoma A 20S. townsendii L N 38S. tridecemlineatus N L N A U 5,7,9,10,13,48,52,54, 64S. variegatus L Af U 1,5,10,36Tamiasciurus hudsonicus L N 3Tamias dorsalis U 29T. merriami N A 10,49T. minimus L N A 38
Ixodes kingia Ixodes sculptusHost species Lb N A U Lb N A U Referencesc
Table 2. Continued.

Journal of Vector Ecology 173Vol. 31, no. 1
LAGOMORPHA – Ochotonidae (Pikas)Ochotona princeps L N N 3,25
LAGOMORPHA – Leporidae (Rabbits and Hares)Lepus californicus L 38 Lepus spp N A 49 Sylvilagus audubonii L 30,36S. floridanus U 21S. nuttallii L 25,36Sylvilagus spp. A L A 9,10,22
Including records where the host species could be determined with reasonable certainty.
Ixodes kingia Ixodes sculptusHost species Lb N A U Lb N A U Referencesc
Table 2. Continued.
Information based on host genus included only when this added data for a tick species or life stage not evident from the species records within that genus.aIncludes records for the synonym I. pratti.bL, larva; N, nymph; A, adult; U, unknown life stage.c1, Banks 1908; 2, Bishopp 1911; 3, Hooker et al. 1912; 4, Burnett and McCampbell 1926; 5, Hixson 1932; 6, Chamberlin, 1937; 7, Eddy and Joyce 1942; 8, Brown 1944; 9, Bishopp and Trembley 1945; 10, Cooley and Kohls 1945; 11, Linsdale 1946; 12, Jameson 1947; 13, Knipping et al. 1950; 14, Brown and Kohls 1950; 15, Edmunds 1951; 16, Holdenried et al. 1951; 17, Eads et al. 1952; 18, Irons et al. 1952; 19, Burgess 1955; 20, Eads et al. 1956; 21, Ecke and Yeatter 1956; 22, Gregson 1956; 23, Eklund et al. 1958; 24, Gier 1959; 25, Allred et al. 1960; 26, Miller and Ward 1960; 27, Egoscue 1962; 28, Johnson 1962; 29, Marchette et al. 1962; 30, Beck et al. 1963; 31, Egoscue 1964; 32, Hansen 1964; 33, Sidwell et al. 1964; 34, Thorpe et al. 1965; 35, Emmons 1966; 36, Johnson 1966; 37, Kinsella and Pattie 1967; 38, Allred 1968; 39, Boddicker 1968; 40, Wilkinson 1970; 41, Hilton and Mahrt 1971; 42, Wittrock and Wilson 1974; 43, CDC 1975; 44, Toweill and Price 1976; 45, Carey et al. 1980; 46, Lane et al. 1981; 47, Spicka 1981; 48, Easton 1983; 49, Furman and Loomis 1984; 50, Lane 1984; 51, Wilkinson 1984; 52, Demaree 1986; 53, Kietzmann Jr. 1987; 54, Kietzmann and Kietzmann 1987; 55, Webb et al. 1990; 56, Cleven et al. 1992; 57, Brillhart et al. 1994; 58, Yensen et al. 1996; 59, Walker et al. 1998; 60, Lang 1999; 61, Merten and Durden 2000; 62, Crooks et al. 2004; 63, Pence et al. 2004; 64, Current study.dReferred to as civet cat for records of I. sculptus (Bishopp and Trembley 1945). This could possibly refer to Spilogale skunks but the authors specifically use the nomenclature skunk in other instances. eReferred to as Perodipus richardsoni by Bishopp (1911) and Hooker et al. (1912). fReferred to as Citellus grammurus by Cooley and Kohls (1945).
Journal of Vector Ecology June 2006174
Figure 2. Mean monthly rainfall, snowfall, and maximum temperature at Briggsdale in Weld County, CO, from 1948-2005. Briggsdale is located adjacent to the southeastern corner of the western portion of the Pawnee National Grassland. Climate data were obtained from the High Plains Regional Climate Center.
Figure 1. Location of rodent trapping sites (black outlines) within the Pawnee National Grassland (PNG). Areas shaded gray denote privately owned land. The location of the PNG within the state of Colorado is shown in the inset.
0
10
20
30
40
50
60
70
80
90
100
Jan. Feb. March April May June July Aug. Sep. Oct. Nov. Dec.
Rainfall (mm)Snowfall (mm)Average max. temperature (°C)

Journal of Vector Ecology 175Vol. 31, no. 1
Seasonal activity patterns of I. kingi and I. sculptus nymphs Based on the low overall numbers of ticks encountered, nymphal infestation data for a particular rodent species were pooled for all PNG sampling sites in the seasonality analyses. This was justified by the previously mentioned lack of significant site-related differences in nymphal infestation rate or nymphal load for the most commonly infested rodent on the PNG. The seasonal activity patterns of nymphal I. kingi (on northern grasshopper mice and thirteen-lined ground squirrels) and I. sculptus (on thirteen-lined ground squirrels) in our northern Colorado study area during May-August are shown in Figures 3A and 3B. All time points included in the figures are based on at least nine animals examined for presence of ticks (range, 9-71 animals for mice and 11-77 animals for ground squirrels). Ixodes kingi was first recorded from mice in early May and from ground squirrels in late June, whereas I. sculptus was not recorded from ground squirrels until early July (Figures 3A and 3B). Peak nymphal infestation rates and nymphal loads of both ticks occurred during July-August (Figures 3A and 3B), which coincides with the warmest period of the year and the latter part of the May-August peak period of rainfall on the PNG (Figure 2).
DISCUSSION
Host usage patterns of I. kingi and I. sculptus We found I. kingi nymphs commonly infesting both a murid rodent (the northern grasshopper mouse) and a sciurid rodent (the thirteen-lined ground squirrel), whereas I. sculptus nymphs were recorded only from the sciurid rodent. This finding is in accordance with the summary of North American tick host records presented in Table 2. Although both ticks infest a variety of mammals (including rodents, lagomorphs, and carnivores), I. kingi appears to be more likely to parasitize heteromyid and murid rodents (e.g., grasshopper mice) than I. sculptus. Our finding that plague epizootics among prairie dog populations result in increased loads of I. kingi on both grasshopper mice and ground squirrels but decreased loads of I. sculptus on ground squirrels demonstrates that subtle differences in host usage patterns by these ticks can have dramatic effects on tick population dynamics. We speculate that a plague-induced die-off of prairie dogs and other susceptible sciurid rodents merely results in I. kingi switching to infesting other less plague-susceptible rodent hosts, whereas the more narrowly sciurid-specializing I. sculptus is less capable of switching hosts and, consequently, suffers population decreases due to diminished host-finding success. Our observation of heavy usage of the thirteen-lined ground squirrel by I. sculptus is in accordance with the results of previous studies including this rodent (see Table 2; Hixson 1932, Eddy and Joyce 1942, Cooley and Kohls 1945, Easton 1983, Demaree 1986, Kietzmann and Kietzmann 1987). Although our record of I. kingi was the first for the thirteen-lined ground squirrel, this tick has previously been collected from several other species of Spermophilus ground squirrels (Table 2). Our finding that northern grasshopper mice were
ground squirrels carried heavier loads of I. sculptus nymphs than grasshopper mice (Table 1; Wilcoxon ranked sums test: χ2 = 7.71, df = 1, P = 0.006) whereas no difference was seen for these rodents in the case of I. kingi nymphs (P = 0.87).
Host usage by I. kingi and I. sculptus following plague outbreaks in prairie dog populations We also examined whether nymphal infestation rates or nymphal loads for I. kingi on grasshopper mice or ground squirrels or for I. sculptus on ground squirrels differed during the peak July-August infestation period between prairie dog town-associated sites with vs. without prairie dog plague epizootics occurring during the year preceding our trapping (2004), the year of the trapping (2005), or during either year (2004-2005). The only statistically significant result of these analyses was a heavier nymphal load of I. kingi on grasshopper mice in sites where plague epizootics had occurred in the prairie dog populations during either 2004 or 2005 than in plague-free towns (means of 0.11 versus 0.02 nymphs; medians of 0 in both cases; χ2 = 3.89, df = 1, P = 0.049). A similar trend towards higher nymphal loads in sites experiencing prairie dog plague epizootics during either 2004 or 2005, relative to plague-free sites, was seen for I. kingi also on ground squirrels (means of 0.37 versus 0.12 nymphs per animal). In the case of I. sculptus on ground squirrels, there was a reverse trend towards higher nymphal loads in plague-free sites than in plague sites (means of 0.22 vs. 0.02 nymphs per animal).
Summary of published North American host records for I. kingi and I. sculptus Both tick species commonly infest rodents as well as carnivores. Ixodes kingi has been reported from humans, two species of domestic animals (cat, dog), 17 species of carnivores, 40 species of rodents, and four species of lagomorphs (Table 2). Ixodes sculptus has been reported from humans, three species of domestic animals (cat, dog, goat), 13 species of carnivores, 34 species of rodents, and three species of lagomorphs (Table 2). In addition to the host records for mammals, a single larval I. kingi has been collected from the striped whipsnake (Masticophis taeniatus) and a single adult I. sculptus from a quail (Lophortyx sp.) (Johnson 1966, Furman and Loomis 1984). To the best of our knowledge, these are the only documented cases of infestation by either I. kingi or I. sculptus for non-mammalian hosts. The clearest differences in host usage between I. kingi and I. sculptus are that I. kingi predominates on heteromyid and murid rodents, whereas I. sculptus is more frequently collected than I. kingi from ground-dwelling sciurid rodents, especially Spermophilus spp. ground squirrels (Table 2). This is in accordance with our finding that I. sculptus was found almost exclusively on thirteen-lined ground squirrels, whereas I. kingi commonly was recorded from both the sciurid thirteen-lined ground squirrel and the murid northern grasshopper mouse (Table 1).

Journal of Vector Ecology June 2006176
Figure 3. Seasonal patterns of nymphal infestation (A) and nymphal loads (B) of Ixodes kingi on northern grasshopper mice and thirteen-lined ground squirrels and of I. sculptus on thirteen-lined ground squirrels in a north-central Colorado prairie landscape. Based on pooled data from all sampling sites on the Pawnee National Grassland.
commonly infested with I. kingi but did not harbor I. sculptus also agrees with previous studies (see Table 2; Allred et al. 1960, Hansen 1964, Johnson 1966, Allred 1968). On the other hand, the low number of I. kingi ticks recorded from Ord’s kangaroo rats and deer mice was surprising because both rodents can be commonly parasitized by this tick in other areas (Allred et al. 1960, Johnson 1966). We speculate that the dearth of I. kingi nymphs on these rodents on the PNG may be related to rodent community structure. Specifically, I. kingi may be less likely to infest kangaroo rats and deer mice in areas where these rodents co-exist with colonial sciurid rodents such as prairie dogs and ground squirrels.
Additional studies on host usage patterns are needed to clarify the ecological mechanisms underlying the observed patterns of host usage by I. kingi and I. sculptus.Infestation of rodents by ixodid ticks in north-central Colorado In addition to our records of I. kingi and I. sculptus from the PNG in Weld County, I. sculptus has been recovered from deer mice, ground squirrels, and chipmunks in the Rocky Mountain National Park in Larimer County, approximately 80 km to the southwest of the PNG (CDC 1975). However, neither I. sculptus nor I. kingi were collected from various murid rodents collected during multiple studies in foothills

Journal of Vector Ecology 177Vol. 31, no. 1
shrub habitats located just west of Fort Collins in central Larmier County, approximately 40 km west of the PNG, or from various shrub or pine habitats along the eastern slope of the Rocky Mountains in Colorado (Maupin et al. 1994; Gage et al. 1997; Burkot et al. 2000, 2001; Zeidner et al. 2000; DeNatale et al. 2002). These studies yielded either only I. spinipalpis (Burkot et al. 2000, 2001; Zeidner et al. 2000) or predominantly I. spinipalpis and low numbers of Dermacentor andersoni (Gage et al. 1997, Maupin et al. 1994, DeNatale et al. 2002). Additional studies are needed to gain an understanding of the overall geographical distribution patterns, and the ecological mechanisms underlying these patterns, of I. kingi and I. sculptus in the western U.S.
Seasonal activity patterns of nymphal I. kingi and I. sculptus The seasonal activity patterns of I. kingi and I. sculptus are, in large part, dependant on local climatic conditions. We observed distinct increases in nymphal abundance from spring (May-June) to summer (July-August) for both tick species. Although it was not possible to determine nymphal abundance during the fall months, we conclude that the nymphal stages of both I. kingi and I. sculptus most likely exhibit a unimodal seasonal activity pattern in northern Colorado, with the peak activity occurring during the summer months. The occurrence of peak abundances of nymphal I. kingi and I. sculptus during July-August on the PNG is not surprising as this period coincides with the warmest time of the year and the latter part of the May-August peak period of rainfall (Figure 2). Although we did not conduct rodent sampling from fall to early spring, the occurrence of a second peak tick activity period during this part of the year is highly unlikely because of the low winter temperatures characteristic of north-central Colorado (Figure 2). The seasonal activity patterns observed on the PNG are in stark contrast to those observed for I. kingi and I. sculptus in other parts of the U.S. For example, the nymphal stage of I. kingi exhibits a bimodal seasonal activity pattern, with peaks in spring and winter, on rodents in hot, dry desert shrubland/piñon-juniper woodland habitats in Nevada (Beck et al. 1963). The same study reported a unimodal seasonal activity pattern for I. sculptus nymphs on white-tailed antelope squirrels (Ammospermophilus leucurus), with peak tick abundance occurring during November-March. On the other hand, the seasonal activity pattern observed for I. sculptus nymphs on thirteen-lined ground squirrels on the PNG is similar to that reported previously for California ground squirrels (S. beecheyi) in the more temperate climate of north coastal California (Holdenried et al. 1951). These findings clearly demonstrate that the seasonal activity patterns of I. kingi and I. sculptus are strongly related to climatic conditions and that the peak tick activity period differs dramatically within the geographical ranges of the ticks. As shown in Figure 3, I. kingi was encountered earlier in the season (early May for grasshopper mice) than I. sculptus (early July for ground squirrels). However, this pattern may have been related, in part, to host usage. Curiously, I. kingi was collected from grasshopper mice throughout May and
June but not encountered on ground squirrels until late June. We speculate that this phenomenon is associated with changes in some aspect of ground squirrel behavior, such as microhabitat usage, during the late spring period when ground squirrel populations increase due to the emergence of juvenile animals.
Potential roles of I. kingi and I. sculptus as enzootic vectors of human pathogens The vector competency of I. kingi or I. sculptus has not been examined for any human pathogen. Thus, we cannot rule out the possibility that these non-human biting ticks are involved in the enzootic maintenance of one or several human pathogens. For example, there is a report of an I. kingi tick being infected with the tularemia agent Francisella tularensis (Thorpe et al. 1965). As shown in Table 2, both I. kingi and I. sculptus commonly infest carnivores as well as rodents. This raises some intriguing questions regarding the possibility of these ticks serving as bridging vectors of F. tularensis from carnivores, which are considered to tolerate infection with the tularemia agent relatively well (Stagg et al. 1956, Lundgren et al. 1957, McKeever et al. 1958, Marchette et al. 1961, Zarnke et al. 2004), to highly F. tularensis-susceptible rodents (Stagg et al. 1956, Marchette et al. 1961). Although most tick species implicated as vectors of F. tularensis belong to the genus Dermacentor (Jellison 1974), at least one member of the genus Ixodes (I. scapularis) is capable of transmitting this pathogen (Hopla 1962). Infection with Coxiella burnetii, the causative agent of Q-fever, also has been reported for I. kingi (Sidwell et al. 1964). Known experimental tick vectors of this pathogen include, among others, D. andersoni and the Australian tick I. holocyclus (Parker and Davis 1938, Smith 1942, Maurin and Raoult 1999). Finally, there is a report of infection of I. sculptus with the Colorado tick fever virus (CTFV) (CDC 1976). This finding is especially interesting since I. sculptus commonly infests Spermophilus spp. ground squirrels (Table 2) and these rodents are considered key enzootic reservoirs of CTFV (Burgdorfer and Eklund 1959, Emmons 1966, Carey et al. 1980, Bowen et al. 1981). The only currently proven experimental vector of CTFV is D. andersoni (Florio et al. 1944, Emmons 1988, Bowen 1989). The above-mentioned findings underscore the possibility that I. kingi and I. sculptus may contribute to enzootic transmission cycles of human pathogens driven primarily by other ticks, especially D. andersoni and I. spinipalpis, in foothills shrub or montane forest habitat in Colorado. Further, since I. kingi and I. sculptus occur in prairie landscapes of Colorado where human-biting ticks (D. andersoni, D. variabilis) are rare or absent, we need to consider the possibility that these ticks are involved in “silent” transmission cycles of other human pathogens. As a case in point, the tick I. spinipalpis maintains “silent” transmission cycles of at least three potential human pathogens (Anaplasma phagocytophilum, Borrelia bissettii, Babesia microti) in foothills shrub habitats in central Colorado lacking a human-biting bridging vector (Maupin et al. 1994; Gage et al. 1997; Burkot et al. 2000, 2001; Zeidner et al. 2000). We conclude

Journal of Vector Ecology June 2006178
that experimental vector competency studies targeting I. kingi and I. sculptus are needed to clarify the potential roles of these ticks as enzootic vectors of selected human pathogens.
Acknowledgments
We thank M. Lindquist of the Shortgrass Steppe Long-term Ecological Research station (SGS-LTER) for logistical support, D.W. Tripp of Colorado State University for logistical support and for providing ticks from prairie dogs, A. Benson, H. Houghton, and C. Knox for field assistance, and M.C. Dolan and J.F. Piesman of the Centers for Disease Control and Prevention for use of their tick reference collection. This study was funded, in part, by grants from the National Science Foundation to the SGS-LTER (DEB 9632852, DEB 0217631) and to M.F. Antolin and P. Stapp (DEB 0327052).
REFERENCES CITED
Allan, S.A. 2001. Ticks (Class Arachnida: Order Acarina), In: W.M. Samuel, M.J. Pybus, and A.A. Kocan (eds.), Parasitic Diseases of Wild Mammals. pp. 72-106. Iowa State University Press, Ames, IA.
Allred, D.M. 1968. Ticks of the national reactor testing station. Brigham Young Univ. Sci. Bull. Biol. Ser. 10: 1-29.
Allred, D.M., D.E. Beck, and L.D. White. 1960. Ticks of the genus Ixodes in Utah. Brigham Young Univ. Sci. Bull. Biol. Ser. 1: 1-42.
American Society of Mammalogists. 1998. Guidelines for the capture, handling, and care of mammals as approved by the American Society of Mammalogists. J. Mammal. 79: 1416-1431.
Antolin, M.F., P. Gober, B. Luce, D.E. Biggins, W.E. Van Pelt, D.B. Seery, M. Lockhart, and M. Ball. 2002. The influence of sylvatic plague on North American wildlife at the landscape level, with special emphasis on black-footed ferret and prairie dog conservation. Trans. N. Am. Wildl. Nat. Res. Conf. 67: 104-127.
Banks, N. 1908. A revision of the Ixodoidea, or ticks, of the United States. U.S. Dept. Agric. Bur. Entomol. Tech. Ser. 15: 1-61.
Beck, D.E., D.A. Allred, and E.P. Brinton. 1963. Ticks of the Nevada test site. Brigham Young Univ. Sci. Bull. Biol. Ser. 4: 1-11.
Bishopp, F.C. 1911. Some new North American Ixodidae with notes on other species. Proc. Biol. Soc. Wash. 24: 197-208.
Bishopp, F.C. and H.L. Trembley. 1945. Distribution and hosts of certain North American ticks. J. Parasitol. 31: 1-54.
Boddicker, M.L. 1968. Parasites of the black-footed ferret. Proc. South Dakota Acad. Sci. 47: 141-148.
Bowen, G.S. 1989. Colorado tick fever virus, In: T. P. Monath (ed.), The Arboviruses: Epidemiology and Ecology. Vol. II. pp. 159-176. CRC Press, Boca Raton, FL.
Bowen, G.S., R.G. McLean, R.B. Shriner, D.B. Francy, K.S. Pokorny, J.M. Trimble, R.A. Bolin, A.M. Barnes, C.H. Calisher, and D.J. Muth. 1981. The ecology of Colorado tick fever in Rocky Mountain National Park in 1974. II.
Infection in small mammals. Am. J. Trop. Med. Hyg. 30: 490-496.
Brillhart, D.B., L.B. Fox, and S.J. Upton. 1994. Ticks (Acari: Ixodidae) collected from small and medium-sized Kansas mammals. J. Med. Entomol. 31: 500-504.
Brown, J.H. 1944. The spotted fever and other Albertan ticks. Can. J. Res. 22: 36-51.
Brown, J.H. and G.M. Kohls. 1950. The ticks of Alberta with special reference to distribution. Can. J. Res. 28: 197-205.
Burgdorfer, W. and C.M. Eklund. 1959. Studies on the ecology of Colorado tick fever virus in western Montana. Am. J. Hyg. 69: 127-137.
Burgess, G.D. 1955. Arthropod ectoparasites of Richardson’s ground squirrel. J. Parasitol. 41: 639-640.
Burkot, T.R., B.S. Schneider, N.J. Pieniazek, C.M. Happ, J.S. Rutherford, S.B. Slemenda, E. Hoffmeister, G.O. Maupin, and N.S. Zeidner. 2000. Babesia microti and Borrelia bissettii transmission by Ixodes spinipalpis ticks among prairie voles, Microtus ochrogaster, in Colorado. Parasitology 121: 595-599.
Burkot, T.R., G.O. Maupin, B.S. Schneider, C. DeNatale, C.M. Happ, J.S. Rutherford, and N.S. Zeidner. 2001. Use of a sentinel host system to study the questing behavior of Ixodes spinipalpis and its role in the transmission of Borrelia bissettii, human granulocytic ehrlichiosis, and Babesia microti. Am. J. Trop. Med. Hyg. 65: 293-299.
Burnett, W.L. and S.C. McCampbell. 1926. The Zuni prairie dog in Montezuma County, Colorado. Colorado Agric. Coll. Circular 49: 1-15.
Carey, A.B., R.G. McLean, and G.O. Maupin. 1980. The structure of a Colorado tick fever ecosystem. Ecol. Monographs 50: 131-152.
CDC. 1975. Studies on Colorado tick fever (CTF) in Colorado, In: Vector-Borne Diseases Division 1975 Report. pp. 116-138. Centers for Disease Control and Prevention, Vector-Borne Diseases Division, Fort Collins, CO.
CDC. 1976. Colorado tick fever (CTF) studies, In: Vector-Borne Diseases Division 1976 Report. Part I: Arbovirus Reference & Research. pp. 47-72. Centers for Disease Control and Prevention, Vector-Borne Diseases Division, Fort Collins, CO.
Chamberlin, W.J. 1937. The ticks of Oregon. Oregon Agric. Exp. Stn. Bull. 349: 1-34.
Cleven, T.D., E.C. Burgess, R.W. Howe, and A.I. Goldsby. 1992. Absence of Ixodes dammini (deer ticks) on Peromyscus leucopus (white-footed mice) in Brown and Door Counties, Wisconsin. Bull. Soc. Vector Ecol. 17: 70-74.
Cooley, R.A. and G.M. Kohls. 1945. The genus Ixodes in North America. U.S. Publ. Health Serv. Nat. Inst. Health Bull. 184: 1-243.
Crooks, K.R., D.K. Garcelon, C.A. Scott, J.E. Depue, J.T. Wilcox, R.B. Kimsey, and D.H. van Vuren. 2004. Ectoparasites of a threatened insular endemic mammalian carnivore: The island spotted skunk. Am. Midl. Nat. 151: 35-41.
Cully, J.E. and E.S. Williams 2001. Interspecific comparisons

Journal of Vector Ecology 179Vol. 31, no. 1
of sylvatic plague in prairie dogs. J. Mammal. 82: 894-905.
Demaree, H. 1986. Ticks of Indiana. Indiana Dept. Nat. Resources Pittman Robertson Bull. 16: 1-178.
DeNatale, C.E., T.R. Burkot, B.S. Schneider, and N.S. Zeidner. 2002. Novel potential reservoirs for Borrelia sp. and the agent of human granulocytic ehrlichiosis in Colorado. J. Wildl. Dis. 38: 478-482.
Doss, M.A., M.M. Farr, K.F. Roach, and G. Anastos. 1974. Index-catalogue of medical and veterinary zoology. U.S. Dept. Agric. Special Publ. 3 - Ticks and tick-borne diseases. I. Genera and species of ticks. Part 2. Genera H-N: 1- 593.
Durden, L.A. and J.E. Keirans. 1996. Nymphs of the genus Ixodes (Acari: Ixodidae) of the United States: Taxonomy, identification key, distribution, hosts, and medical/veterinary importance. Entomological Society of America, Lanham, MD.
Eads, R.B., G.C. Menzies, and V.I. Miles. 1952. Acarina taken during west Texas plague studies. Proc. Ent. Soc. Wash. 54: 250-253.
Eads, R.B., G.C. Menzies, and B.G. Hightower. 1956. The ticks of Texas, with notes on their medical significance. Texas J. Sci. 8: 7-24.
Easton, E.R. 1983. The ticks of South Dakota: An annotated checklist (Acari: Ixodoidea). Entomol. News 94: 191-195.
Ecke, D.H. and R.E. Yeatter. 1956. Notes on the parasites of cottontail rabbits in Illinois. Illinois Acad. Sci. Trans. 48: 208-214.
Eddy, G.W. and C.R. Joyce. 1942. Ticks collected on the Tama (Iowa) Indian Reservation with notes on other species. Iowa State Coll. J. Sci. 16: 539-543.
Edmunds, L.R. 1951. A check list of the ticks of Utah. Pan-Pacific Entomologist 27: 23-26.
Egoscue, H.J. 1962. Ecology and life history of the kit fox in Tooele County, Utah. Ecology 43: 481-497.
Egoscue, H.J. 1964. Ecological notes and laboratory life history of the canyon mouse. J. Mammal. 45: 387-396.
Eklund, C.M., G.M. Kohls, and W.L. Jellison. 1958. Isolation of Colorado tick fever virus from rodents in Colorado. Science 128: 413.
Emmons, R.W. 1966. Colorado tick fever: Prolonged viremia in hibernating Citellus lateralis. Am. J. Trop. Med. Hyg. 15: 428-433.
Emmons, R.W. 1988. Ecology of Colorado tick fever. Ann Rev. Microbiol 42: 49-64.
Florio, L., M.D. Stewart, and E.R. Mugrage. 1944. The experimental transmission of Colorado tick fever. J. Exp. Med. 80: 165-188.
Furman, D.P. and E.C. Loomis. 1984. The ticks of California (Acari: Ixodida). Bull. Calif. Insect Survey 25: 1-239.
Gage, K.L., G.O. Maupin, J. Montenieri, J. Piesman, M. Dolan, and N.A. Panella. 1997. Flea (Siphonaptera: Ceratophyllidae, Hystrichopsyllidae) and tick (Acarina: Ixodidae) control on wood rats using host-targeted liquid permethrin in bait tubes. J. Med. Entomol. 34: 46-51.
Gier, H.T. 1959. Parasites and diseases of Kansas coyotes.
Kansas State Univ. Agric. Exp. Stn. Tech. Bull. 91: 1-34.
Gregson, J.D. 1956. The Ixodoidea of Canada. Canada Dept. Agric. Sci. Serv. Publ. 930: 1-92.
Hansen, C.G. 1964. Ectoparasites of mammals from Oregon. Great Basin Nat. 24: 75-81.
Hilton, D.F. J. and J.L. Mahrt. 1971. Ectoparasites from three species of Spermophilus (Rodentia: Sciuridae) in Alberta. Can. J. Zool. 49: 1501-1504.
Hixson, H. 1932. The life history and habits of Ixodes sculptus Neumann (Ixodidae). Iowa State Coll. J. Sci. 7: 35-42.
Holdenried, R., F.C. Evans, and D.S. Longanecker. 1951. Host-parasite-disease relationships in a mammalian community in the central coast range of California. Ecol. Monographs 21: 1-18.
Hooker, W.A., F.C. Bishopp, and H.P. Wood. 1912. The life history and bionomics of some North American ticks. U.S. Dept. Agric. Bur. Entomol. Bull. 106: 1-239.
Hopla, C.E. 1962. Ixodes scapularis as a vector of tularemia organisms in the southern United States, pp. 95-97, 11th International Congress of Entomology, Vienna, August 17-25, 1960.
Irons, J.V., R.B. Eads, C.W. Johnson, O.L. Walker, and M.A. Norris. 1952. Southwest Texas Q fever studies. J. Parasitol. 38: 1-5.
Jameson, E.W., Jr. 1947. Natural history of the prairie vole (mammalian genus Microtus). Univ. Kansas Museum Nat. Hist. Publ. 1: 125-151.
Jellison, W.L. 1974. Tularemia in North America, 1930-1974. University of Montana. Missoula, MT.
Johnson, D.E. 1962. General lists of insects of Curecanti Reservoir Basins, 1961. Univ. Utah Anthropol. Papers 59: 168-177.
Johnson, D.E. 1966. Ticks of Dugway Proving Ground and vicinity and their host associations. Proc. Utah Acad. Sci. Arts Lett. 43: 49-66.
Kietzmann, G.E., Jr. 1987. Ectoparasites of black-tailed prairie dogs (Cynomys ludovicianus) from South Dakota. J. Wildl. Dis. 23: 331-333.
Kietzmann, G.E., Jr. and E.A. Kietzmann. 1987. Parasites associated with thirteen-lined ground squirrels (Spermophilus tridecemlineatus Mitchill) from Ames, Iowa. Proc. Iowa Acad. Sci. 94: 22-23.
Kinsella, J.M. and D.L. Pattie. 1967. Ectoparasites of small mammals of the alpine Beartooth Plateau, Wyoming. Can. J. Zool. 45: 233-235.
Knipping, P.A., B.B. Morgan, and R.J. Dicke. 1950. Notes on the distribution of Wisconsin ticks. Trans. Wisconsin Acad. Sci. Arts Lett. 40: 185-197.
Lane, R.S. 1984. New host records of ticks (Acari: Argasidae and Ixodidae) parasitizing wildlife in California and a case of tick paralysis in a deer. Calif. Fish Game 70: 11-17.
Lane, R.S., R.W. Emmons, D.V. Dondero, and B.C. Nelson. 1981. Ecology of tick-borne agents in California. I. Spotted fever group rickettsiae. Am. J. Trop. Med. Hyg. 30: 239-252.
Lang, J.D. 1999. Ixodid ticks (Acari, Ixodidae) found in San

Journal of Vector Ecology June 2006180
Diego County, California. J. Vector Ecol. 24: 61-69.Linsdale, J.M. 1946. The California Ground Squirrel.
University of California Press, Berkeley, CA.Lundgren, D.L., W.J. Marchette, and K.L. Smart. 1957.
Tularemia in the coyote, Canis latrans lestes Merriam. J. Infect. Dis. 101: 154-157.
Marchette, W.J., D.L. Lundgren, P.S. Nicholes, and E.D. Vest. 1961. Studies on infectious diseases in wild animals in Utah. I. Susceptibility of wild animals to experimental tularemia. Zoonoses Res. 1: 49-73.
Marchette, N.J., J.B. Bushman, D.D. Parker, and E.E. Johnson. 1962. Studies on infectious diseases in wild animals in Utah. IV. A wild rodent (Peromyscus spp.) plague focus in Utah. Zoonoses Res. 1: 341-361.
Maupin, G.O., K.L. Gage, J. Piesman, J. Montenieri, S.L. Sviat, L. Vanderzanden, C.M. Happ, M. Dolan, and B.J.B. Johnson. 1994. Discovery of an enzootic cycle of Borrelia burgdorferi in Neotoma mexicana and Ixodes spinipalpis from northern Colorado, an area where Lyme disease is nonendemic. J. Infect. Dis. 170: 636-643.
Maurin, M. and D. Raoult. 1999. Q fever. Clin. Microbiol. Rev. 12: 518-553.
McKeever, S., J.A. Schubert, M.D. Moody, G.W. Gorman, and J.F. Chapman. 1958. Natural occurrence of tularemia in marsupials, carnivores, lagomorphs, and large rodents in southwestern Georgia and northwestern Florida. J. Infect. Dis. 103: 120-126.
Merten, H.A. and L.A. Durden. 2000. A state-by-state survey of ticks recorded from humans in the United States. J. Vector Ecol. 25: 102-113.
Miller, R.S. and R.A. Ward. 1960. Ectoparasites of pocket gophers from Colorado. Am. Midl. Nat. 64: 382-391.
Parker, R.R. and G.E. Davis. 1938. A filter-passing infectious agent isolated from ticks. II. Transmission by Dermacentor andersoni. Publ. Health Rep. 53: 2267-2269.
Pence, D.B., J.F. Kamler, and W.B. Ballard. 2004. Ectoparasites of the swift fox in northwestern Texas. J. Wildl. Dis. 40: 543-547.
Sall, J., L. Creighton, and A. Lehman. 2005. JMP Start Statistics. Third Edition. Brooks/Cole, Belmont, CA.
Sidwell, R.W., D.L. Lundgren, J.B. Bushman, and B.D. Thorpe. 1964. The occurrence of a possible epizootic of Q fever in fauna of the Great Salt Lake desert of Utah. Am. J. Trop. Med. Hyg.13: 754-762.
Smith, D.J.W. 1942. Studies in the epidemiology of Q fever. X. The transmission of Q fever by the tick Ixodes holocyclus (with notes on the paralysis in bandicoots). Aust. J. Exp. Bio. Med. Sci. 20: 213-217.
Spicka, E.J. 1981. Ectoparasites and other arthropod associates on two subspecies of plains pocket gophers: Geomys bursarius illinoensis and Geomys bursarius missouriensis. Can. J. Zool. 59: 1903-1908.
Stagg, G., W.S. Tanner, and J. Lavender. 1956. Experimental infections of native animals with P. tularensis. J. Infect. Dis. 99: 34-37.
Thorpe, B.D., R.W. Sidwell, D.E. Johnson, K.L. Smart, and D.D. Parker. 1965. Tularemia in the wildlife and livestock of the Great Salt Lake desert region, 1951 through 1964. Am. J. Trop. Med. Hyg. 14: 622-637.
Toweill, D.E. and M.A. Price. 1976. Ectoparasites of ringtails collected from Kerr County, Texas. Southwestern Entomol. 1: 20.
Walker, E.D., M.G. Stobierski, M.L. Poplar, T.W. Smith, A.J. Murphy, P.C. Smith, S.M. Schmitt, T.M. Cooley, and C.M. Kramer. 1998. Geographic distribution of ticks (Acari: Ixodidae) in Michigan, with emphasis on Ixodes scapularis and Borrelia burgdorferi. J. Med. Entomol. 35: 872-882.
Webb, J.P., Jr., S.G. Bennett, and G.L. Challet. 1990. The larval ticks of the genus Ixodes Latreille (Acari: Ixodidae) of California. Bull. Soc. Vector Ecol. 15: 73-124.
Wilkinson, P.R. 1970. Dermacentor ticks on wildlife and new records of paralysis. J. Entomol. Soc. Brit. Columbia 67: 24-29.
Wilkinson, P.R. 1984. Hosts and distribution of Rocky Mountain wood ticks (Dermacentor andersoni) at a tick focus in British Columbia rangeland. J. Entomol. Soc. Brit. Columbia 81: 57-71.
Wilson, D.E. and S. Ruff. 1999. The Smithsonian Book of North American Mammals. Smithsonian Institution Press, Washington, D.C.
Wittrock, D.D. and N. Wilson. 1974. Ectoparasites of the badger, Taxidea taxus (Schreber, 1778), in northwestern Iowa with a list of species recorded from North America. Iowa State J. Res. 49: 9-15.
Yensen, E., C.R. Baird, and P.W. Sherman. 1996. Larger ectoparasites of the Idaho ground squirrel (Spermophilus brunneus). Great Basin Nat. 56: 237-246.
Zarnke, R.L., J.M. Ver Hoef, and R.A. DeLong. 2004. Serologic survey for selected disease agents in wolves (Canis lupus) from Alaska and the Yukon territory, 1984-2000. J. Wildl. Dis. 40: 632-638.
Zeidner, N.S., T.R. Burkot, R. Massung, W.L. Nicholson, M.C. Dolan, J.S. Rutherford, B.J. Biggerstaff, and G.O. Maupin. 2000. Transmission of the agent of human granulocytic ehrlichiosis by Ixodes spinipalpis ticks: Evidence of an enzootic cycle of dual infection with Borrelia burgdorferi in northern Colorado. J. Infect. Dis. 182: 616-619.


















