
HIV Copyright © 2019 AAV-delivered eCD4-Ig protects rhesus ...
13
Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019 SCIENCE TRANSLATIONAL MEDICINE | RESEARCH ARTICLE 1 of 12 HIV AAV-delivered eCD4-Ig protects rhesus macaques from high-dose SIVmac239 challenges Matthew R. Gardner 1 * † , Christoph H. Fellinger 1 *, Lisa M. Kattenhorn 2‡ , Meredith E. Davis-Gardner 1 , Jesse A. Weber 1 , Barnett Alfant 1 , Amber S. Zhou 1 , Neha R. Prasad 1 , Hema R. Kondur 1 , Wendy A. Newton 3 , Kimberly L. Weisgrau 3 , Eva G. Rakasz 3 , Jeffrey D. Lifson 4 , Guangping Gao 5,6 , Nancy Schultz-Darken 3 , Michael Farzan 1 A number of simian and simian human immunodeficiency viruses (SIV and SHIV, respectively) have been used to assess the efficacy of HIV-1 vaccine strategies. Among these, SIVmac239 is considered among the most stringent because, unlike SHIV models, its full genome has coevolved in its macaque host and its tier 3 envelope glycoprotein (Env) is exceptionally hard to neutralize. Here, we investigated the ability of eCD4-Ig, an antibody-like entry inhibitor that emulates the HIV-1 and SIV receptor and coreceptor, to prevent SIVmac239 infection. We show that rh-eCD4-Ig I39N expressed by recombinant adeno-associated virus (AAV) vectors afforded four rhesus macaques complete protection from high-dose SIVmac239 challenges that infected all eight control macaques. However, rh-eCD4-Ig I39N –expressing macaques eventually succumbed to serial escalating challenge doses that were 2, 8, 16, and 32 times the chal- lenge doses that infected the control animals. Despite receiving greater challenge doses, these macaques had significantly lower peak and postpeak viral loads than the control group. Virus isolated from three of four ma- caques showed evidence of strong immune pressure from rh-eCD4-Ig I39N , with mutations located in the CD4-binding site, which, in one case, exploited a point-mutation difference between rh-eCD4-Ig I39N and rhesus CD4. Other escape pathways associated with clear fitness costs to the virus. Our data report effective protection of rhesus macaques from SIVmac239. INTRODUCTION Conventional HIV-1 vaccine designs have not been successful due in part to their inability to elicit neutralizing antibodies. However, over the past decade, broadly neutralizing antibodies (bNAbs) that emerge naturally several years after infection in some HIV-1–positive individuals have been identified and characterized (1). Whereas vaccine immunogen design is currently focused on eliciting such bNAbs, passive administration of bNAbs has successfully prevented and treated SHIV infection in rhesus macaques (2–9). In addition, clinical trials have shown bNAbs to be safe in uninfected individuals and both safe and effective in HIV-1–infected individuals (10–17). A limitation to passive administration of bNAbs is the need for repeated infusions to maintain concentrations at protective and therapeutic levels. Adeno-associated virus (AAV) vectors can bypass this difficulty by expressing a secreted protein for years after a single inoculation (18). These vectors can deliver a transgene payload to the nuclei of transduced tissue including skeletal muscle and liver. The transgenes persist as an unintegrated episome that can express a protein of interest. AAV-expressed bNAbs or immunoadhesins can also prevent HIV-1 or SIVmac316 transmission in animal models (19–21). However, more recent macaque studies show that AAV- delivered bNAbs, either of rhesus origin or rhesusized in their constant regions, are frequently targeted by host immune responses, limiting their expression and efficacy (22–24). The macaques often develop severe anti-drug antibody (ADA) responses that have been correlated to the degree of somatic hypermutation of the expressed bNAb (22). In addition, a recent study reported 20% of macaques developing ADA against an anti–simian immunodeficiency virus (SIV) antibody with lower amounts of somatic hypermutation compared to HIV-1 bNAbs (25). We have previously described an HIV-1 entry inhibitor, desig- nated eCD4-Ig, that is more self-like than a bNAb (26–28). eCD4- Ig is an Fc-fusion protein composed of domains 1 and 2 of human CD4 fused to an immunoglobulin G (IgG) Fc with a sulfated CCR5-mimetic peptide attached to the Fc C terminus. This inhibi- tor functions like an antibody, in that it binds to the CD4- and coreceptor-binding sites on the HIV-1 or SIV envelope glycoprotein (Env) to prevent infection. eCD4-Ig shows greater neutralization breadth than any bNAb described to date, because it potently neu- tralizes all HIV-1, HIV-2, and SIV isolates assayed (28). We have previously demonstrated that AAV-delivered rhesus eCD4-Ig (rh-eCD4-Ig I39N ) could protect rhesus macaques from SHIV-AD8 challenges that infected all control macaques (28). rh-eCD4-Ig I39N uses rhesus CD4 domains 1 and 2 modified with an asparagine at residue 39 to better emulate the potency of human eCD4-Ig. As pre- viously reported, this human CD4-derived residue improves binding to both HIV-1 and SIV Envs (29, 30). In this study, we examined the efficacy of AAV-expressed eCD4-Ig to protect rhesus macaques from SIVmac239 challenges. Together with protection observed against SHIV-AD8 challenges, this study highlights the utility of AAV-expressed eCD4-Ig as an alternative to a conventional HIV-1 vaccine. 1 Department of Immunology and Microbiology, The Scripps Research Institute, Jupiter, FL 33458, USA. 2 Department of Microbiology and Immunobiology, Harvard Medical School, New England Primate Research Center, Southborough, MA 01772, USA. 3 Wisconsin National Primate Research Center, University of Wisconsin, Madison, WI 53715 USA. 4 AIDS and Cancer Virus Program, Frederick National Laboratory for Cancer Research, Frederick, MD 21702, USA. 5 Horae Gene Therapy Center, University of Massachusetts Medical School, Worcester, MA 01605, USA. 6 Department of Microbiology and Physiological Systems, University of Massachusetts Medical School, Worcester, MA 01655, USA. *These authors contributed equally to this work. †Corresponding author. Email: [email protected] ‡Present address: Novartis Institutes for Biomedical Research, Cambridge, MA 02139, USA. Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works at UNIV OF CALIFORNIA - SAN DIEGO on August 9, 2019 http://stm.sciencemag.org/ Downloaded from
Transcript of HIV Copyright © 2019 AAV-delivered eCD4-Ig protects rhesus ...

Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
1 of 12
H I V
AAV-delivered eCD4-Ig protects rhesus macaques from high-dose SIVmac239 challengesMatthew R. Gardner1*†, Christoph H. Fellinger1*, Lisa M. Kattenhorn2‡, Meredith E. Davis-Gardner1, Jesse A. Weber1, Barnett Alfant1, Amber S. Zhou1, Neha R. Prasad1, Hema R. Kondur1, Wendy A. Newton3, Kimberly L. Weisgrau3, Eva G. Rakasz3, Jeffrey D. Lifson4, Guangping Gao5,6, Nancy Schultz-Darken3, Michael Farzan1
A number of simian and simian human immunodeficiency viruses (SIV and SHIV, respectively) have been used to assess the efficacy of HIV-1 vaccine strategies. Among these, SIVmac239 is considered among the most stringent because, unlike SHIV models, its full genome has coevolved in its macaque host and its tier 3 envelope glycoprotein (Env) is exceptionally hard to neutralize. Here, we investigated the ability of eCD4-Ig, an antibody-like entry inhibitor that emulates the HIV-1 and SIV receptor and coreceptor, to prevent SIVmac239 infection. We show that rh-eCD4-IgI39N expressed by recombinant adeno-associated virus (AAV) vectors afforded four rhesus macaques complete protection from high-dose SIVmac239 challenges that infected all eight control macaques. However, rh-eCD4-IgI39N–expressing macaques eventually succumbed to serial escalating challenge doses that were 2, 8, 16, and 32 times the chal-lenge doses that infected the control animals. Despite receiving greater challenge doses, these macaques had significantly lower peak and postpeak viral loads than the control group. Virus isolated from three of four ma-caques showed evidence of strong immune pressure from rh-eCD4-IgI39N, with mutations located in the CD4-binding site, which, in one case, exploited a point-mutation difference between rh-eCD4-IgI39N and rhesus CD4. Other escape pathways associated with clear fitness costs to the virus. Our data report effective protection of rhesus macaques from SIVmac239.
INTRODUCTIONConventional HIV-1 vaccine designs have not been successful due in part to their inability to elicit neutralizing antibodies. However, over the past decade, broadly neutralizing antibodies (bNAbs) that emerge naturally several years after infection in some HIV-1–positive individuals have been identified and characterized (1). Whereas vaccine immunogen design is currently focused on eliciting such bNAbs, passive administration of bNAbs has successfully prevented and treated SHIV infection in rhesus macaques (2–9). In addition, clinical trials have shown bNAbs to be safe in uninfected individuals and both safe and effective in HIV-1–infected individuals (10–17).
A limitation to passive administration of bNAbs is the need for repeated infusions to maintain concentrations at protective and therapeutic levels. Adeno-associated virus (AAV) vectors can bypass this difficulty by expressing a secreted protein for years after a single inoculation (18). These vectors can deliver a transgene payload to the nuclei of transduced tissue including skeletal muscle and liver. The transgenes persist as an unintegrated episome that can express a protein of interest. AAV-expressed bNAbs or immunoadhesins can also prevent HIV-1 or SIVmac316 transmission in animal models
(19–21). However, more recent macaque studies show that AAV- delivered bNAbs, either of rhesus origin or rhesusized in their constant regions, are frequently targeted by host immune responses, limiting their expression and efficacy (22–24). The macaques often develop severe anti-drug antibody (ADA) responses that have been correlated to the degree of somatic hypermutation of the expressed bNAb (22). In addition, a recent study reported 20% of macaques developing ADA against an anti–simian immunodeficiency virus (SIV) antibody with lower amounts of somatic hypermutation compared to HIV-1 bNAbs (25).
We have previously described an HIV-1 entry inhibitor, desig-nated eCD4-Ig, that is more self-like than a bNAb (26–28). eCD4-Ig is an Fc-fusion protein composed of domains 1 and 2 of human CD4 fused to an immunoglobulin G (IgG) Fc with a sulfated CCR5-mimetic peptide attached to the Fc C terminus. This inhibi-tor functions like an antibody, in that it binds to the CD4- and coreceptor-binding sites on the HIV-1 or SIV envelope glycoprotein (Env) to prevent infection. eCD4-Ig shows greater neutralization breadth than any bNAb described to date, because it potently neu-tralizes all HIV-1, HIV-2, and SIV isolates assayed (28). We have previously demonstrated that AAV-delivered rhesus eCD4-Ig (rh-eCD4-IgI39N) could protect rhesus macaques from SHIV-AD8 challenges that infected all control macaques (28). rh-eCD4-IgI39N uses rhesus CD4 domains 1 and 2 modified with an asparagine at residue 39 to better emulate the potency of human eCD4-Ig. As pre-viously reported, this human CD4-derived residue improves binding to both HIV-1 and SIV Envs (29, 30). In this study, we examined the efficacy of AAV- expressed eCD4-Ig to protect rhesus macaques from SIVmac239 challenges. Together with protection observed against SHIV-AD8 challenges, this study highlights the utility of AAV-expressed eCD4-Ig as an alternative to a conventional HIV-1 vaccine.
1Department of Immunology and Microbiology, The Scripps Research Institute, Jupiter, FL 33458, USA. 2Department of Microbiology and Immunobiology, Harvard Medical School, New England Primate Research Center, Southborough, MA 01772, USA. 3Wisconsin National Primate Research Center, University of Wisconsin, Madison, WI 53715 USA. 4AIDS and Cancer Virus Program, Frederick National Laboratory for Cancer Research, Frederick, MD 21702, USA. 5Horae Gene Therapy Center, University of Massachusetts Medical School, Worcester, MA 01605, USA. 6Department of Microbiology and Physiological Systems, University of Massachusetts Medical School, Worcester, MA 01655, USA.*These authors contributed equally to this work.†Corresponding author. Email: [email protected]‡Present address: Novartis Institutes for Biomedical Research, Cambridge, MA 02139, USA.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from

Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
2 of 12
RESULTSComplete, long-term eCD4-Ig–mediated protection from SHIV-AD8We have reported that AAV1-delivered rh-eCD4-IgI39N protected four of four rhesus macaques from four infectious challenges with SHIV-AD8, with a final challenge at 34 weeks after AAV1 inoculation (28). We continued this study through week 60, when rh-eCD4-IgI39N concentrations for the four macaques were 23, 30, 34, and 52 g/ml (fig. S1A). As previously shown, modest ADA responses were ob-served in two inoculated macaques but largely disappeared by week 37 and remained below background through week 60 (fig. S1B). All four inoculated animals were challenged again intravenously at weeks 52 and 56 with doses of SHIV-AD8 that were two and four times greater (1600 and 3200 pg of p27), respectively, than the highest previous dose (fig. S1C). These challenges were 8 and 16 times the 50% animal infectious dose of this stock of SHIV-AD8, but no macaque became infected. In contrast, as we previously reported, AAV1 vectors encoding HIV-1 bNAbs (3BNC117, NIH45-46, 10-1074, or PGT121) (31) did not afford significant protection (P = 0.23, Mantel-Cox test) from SHIV-AD8 challenge doses that were 16 and 32 times lower than the final challenge dose of the rh-eCD4-IgI39N group (fig. S1D). We conclude that, following a single inoculation, AAV1-rh-eCD4-IgI39N can completely protect macaques from high-dose SHIV-AD8 challenges for more than 1 year.
Robust protection from SIVmac239To further assess the in vivo breadth of AAV1-rh-eCD4-IgI39N, we subsequently investigated whether rh-eCD4-IgI39N could protect from the highly pathogenic tier 3 virus SIVmac239. We first confirmed that rh-eCD4-IgI39N neutralized SIVmac239 produced in human embryonic kidney (HEK) 293T cells and rhesus macaque peripheral blood mononuclear cells (PBMCs) with nearly identical efficiencies, with 50% inhibitory concentrations (IC50) of 0.03 and 0.036 g/ml, respectively (fig. S2). We then inoculated four macaques with 2.0 × 1013 genome copies (GCs) AAV1 vectors encoding rh-eCD4-IgI39N together with AAV1 vectors encoding rhesus tyrosine protein sulfotranserase 2 (TPST2) (0.5 × 1013 GCs), an enzyme that ensures efficient sulfation of tyrosines present at the rh-eCD4-IgI39N C terminus. Serum rh-eCD4-
IgI39N concentrations peaked at 13 to 44 g/ml, plateauing to between 3 and 18 g/ml (Fig. 1A).
Compared with our previous SHIV-AD8 study (fig. S1B), we observed higher ADA against rh-eCD4-IgI39N in all four macaques beginning at week 4 after inoculation (Fig. 1B). However, ADA in rh-eCD4-IgI39N–expressing macaques reverted to background by 14 weeks in three of four animals, whereas in one animal with the lowest rh-eCD4-IgI39N concentration (rh2448), ADA persisted for 30 weeks. Increased ADA was observed in all four animals after infection, as shown for rh2448. ADA for this group of macaques primarily recognized the rh-eCD4-IgI39N sulfopeptide (Fig. 2, A and B), with little or no recognition of the CD4 domains of rh-eCD4-IgI39N, or the ectodomains of human or rhesus CD4 (Fig. 2, C and D).
Beginning at 22 weeks after AAV1 inoculation, rh-CD4-IgI39N–expressing macaques were challenged with SIVmac239. SIVmac239 was selected because its Env is highly divergent from that of SHIV-AD8, used in fig. S1, and because humoral prophylaxis against this tier 3 virus is considered especially challenging. None of the rh-eCD4-IgI39N–expressing animals became infected after intravenous SIVmac239 challenges that infected all eight untreated control animals (Fig. 3A). Macaque rh2448, with the highest ADA and lowest rh-eCD4-IgI39N concentrations (3 g/ml), became infected 34 weeks after inoculation at twice the challenge dose that infected all control animals. With repeated escalating challenge inoculum doses, the remaining macaques eventually succumbed to infection at 8, 16, and 32 times the dose that infected all control animals, at 42, 46, and 50 weeks after inoculation, respectively. Although the macaque expressing the lowest amounts of rh-eCD4-IgI39N became infected first, we did not observe a significant correlation (P = 0.33, Pearson correlation analysis) between number of challenges needed to infect the macaques and the amount of expressed rh-eCD4-IgI39N at the time of SIVmac239 acquisition (fig. S3). Despite the absence of a detectable correlation, this protection was derived from rh-eCD4-IgI39N rather than from a nonspecific consequence of AAV1 inoculation. For example, unlike serum from rh-eCD4-IgI39N–inoculated macaques, serum from animals inoculated with AAV1-expressing antibodies did not neutralize SIVmac239 (fig. S4A) or mediate antibody-dependent cellular cyto-toxicity (ADCC) against this virus (fig. S4B). We then compared viral
Fig. 1. rh-eCD4-IgI39N concentrations and anti–rh-eCD4-IgI39N responses in macaques inoculated with AAV1-rh-eCD4-IgI39N. (A) Serum concentrations of rh-eCD4-IgI39N in four rhesus macaques inoculated intramuscularly with 2.0 × 1013 genomic copies (GCs) of AAV1 vectors encoding rh-eCD4-IgI39N and 5 × 1012 GCs of AAV1 vectors encoding rhesus TPST2. (B) Anti–rh-eCD4-IgI39N IgG antibodies in the sera of macaques expressing rh-eCD4-IgI39N, as determined by enzyme-linked immunosorbent assay (ELISA) with plates coated with rh-eCD4-IgI39N. Note that macaque rh2448 (red) became infected at week 36. Error bars represent the range of measured duplicates per experiment. Data are representative measurements of at least two experiments.
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from

Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
3 of 12
loads in infected rh-eCD4-IgI39N–expressing macaques (Fig. 3B). Peak and postpeak (weeks 6 through 12 after infection) viral loads were significantly lower (P <0.05, unpaired t test) in rh-eCD4-IgI39N–expressing macaques compared to controls (Fig. 3, C and D). This difference is especially noteworthy because rh-eCD4-IgI39N–expressing macaques were challenged with doses of SIVmac239 that were con-siderably higher than what control animals received. These latter data suggest that rh-eCD4-IgI39N exerts continuing pressure on the rep-licating virus, although it may also suggest differences in initial seeding of infected cells. Collectively, our data show that AAV- delivered rh-eCD4-IgI39N affords substantial and robust protection against acquisition of both SHIV-AD8 and SIVmac239 and suggest a role in partial control of SIVmac239 replication after acquisition.
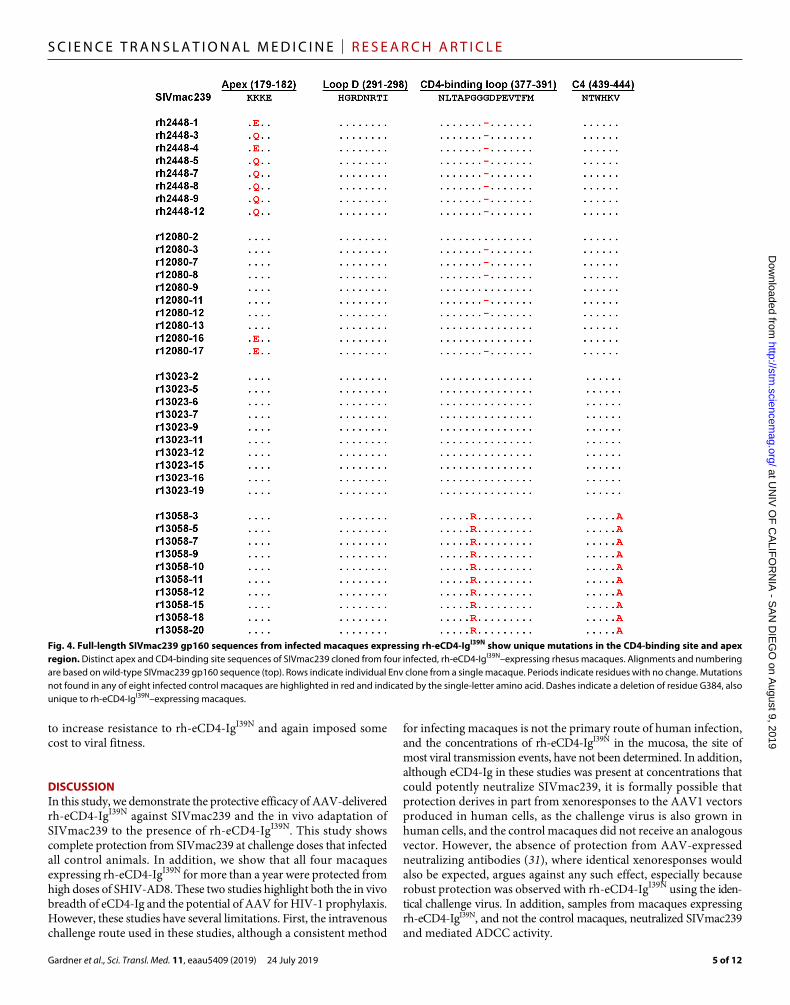
Evolution of SIVmac239 Env in the presence of rhesus eCD4-IgTo examine whether AAV-expressed rh-eCD4-IgI39N exerted pressure on the replicating virus in macaques that became infected with SIVmac239, we sequenced full-length env genes from all four rh-eCD4-IgI39N–expressing macaques and all eight controls. Samples were obtained from macaques expressing rh-eCD4-IgI39N at the time of necropsy and from control animals 10 to 12 weeks after infection, after which they were placed on combination antiretroviral therapy for another study. We observed multiple amino acid changes in Env sequences in the rh-eCD4-IgI39N group not found in the control group (fig. S5, A and B). Some of these mutations may reflect ongoing escape from the rhesus humoral response, whereas others may confer resistance to rh-eCD4-IgI39N. We therefore characterized those that emerged multiple times in one or more rh-eCD4-IgI39N–expressing macaque. env genes from two such macaques, rh2448 and r12080, encoded a deletion of G384 in the CD4-binding loop, analogous to residue 367 of HIV-1 (Fig. 4). Sequences obtained from r13058 encoded mutations at residue 382 (glycine to arginine, G382R) and
residue 444 (valine to alanine, V444A) in the CD4-binding loop and C4 region, respectively. No changes in the Loop D region of the CD4-binding site were observed in the rh-eCD4-IgI39N group. In addition, no mutations in the CD4-binding site were observed in r13023. Furthermore, we did not observe any mutations in the known residues that comprise the coreceptor-binding site on SIVmac239 (fig. S5B). Sequences from rh2448 and r12080 also encoded mutations at residue 180 (lysine to glutamine or lysine to glutamic acid, K180Q or K180E) (Fig. 4). This result is interesting because K180 is in the predicted apex region of SIVmac239, a region of HIV-1 Env shown to bind sulfated tyrosines of apex antibodies such as PGT145, PGDM1400, CAP256-VRC26.25, and PG9 (32, 33).
We then sought to determine whether the observed Env mutations altered susceptibility to rh-eCD4-IgI39N. Specifically, we generated SIVmac239 gp160 expression plasmids encoding the G382R and V444A mutations (G382R/V444A), the K180Q and G384 deletion mutations (K180Q/G384), the G382R mutation, or the G384 deletion mutation (G384). When tested in neutralization assays, the K180Q/G384 variant mediated a 48-fold increase in IC50 by rh-eCD4-IgI39N, and the G384 variant alone was 10-fold more resistant than wild-type SIVmac239 (Fig. 5A). A similar decrease in potency against the K180Q/G384 and the G384 variants was also observed for rh-CD4-IgI39N and rh-CD4-Ig, suggesting that the G384 mutation resulted in a loss of rhesus CD4 binding. In contrast, the G382R/V444A variant was not more resistant to rh-eCD4-IgI39N or rh-CD4-IgI39N. However, this variant was threefold less sensitive to rh-CD4-Ig, lacking the I39N mutation in CD4 domain 1. All three inhibitors, rh-eCD4-IgI39N, rh-CD4-IgI39N, and rh-CD4-Ig, showed an increase in potency against the G382R single-mutation variant suggesting that the V444A was a compensatory mutation. The neutralization results correlated with the affinity of the gp120 variants on rh-CD4-Ig and rh-CD4-IgI39N as determined by surface plasmon resonance (SPR) (Fig. 5B and fig. S6). The G382R gp120 variant had greater
Fig. 2. Host ADA against expressed rh-eCD4-IgI39N primarily targets the sulfated CCR5-mimetic peptide. (A to D) ELISA plates were coated with rh-CD4-IgI39N (A), CCR5mim2-Ig (B), the human CD4 ectodomain (C), or the rhesus CD4 ectodomain (D). rh-CD4-IgI39N and CCR5mim2-Ig included human rather than rhesus Fc domains. The CCR5mim2 sulfopeptide of CCR5mim2-Ig is placed at the Fc C terminus. Values indicate absorbance at 450 nm. Error bars represent the range of duplicates per experiment. Data are representative measurements of at least two experiments.
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from

Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
4 of 12
affinity for rh-CD4-Ig, consistent with previous reports (29). Both the K180Q/G384 and the G384 variants exhibited decreased affinity for rh-CD4-Ig and rh-CD4-IgI39N. The G382R/V444A variant bound rh-CD4-Ig, but not rh-CD4-IgI39N, less efficiently, consistent with the decrease in potency observed in the neutralization assay. Last, we investigated SIVmac239 Env variants from macaque r13023. No Env variants from this macaque included CD4-binding site changes, but a combination of three gp41 mutations (Y690C/R774S/A833T) resulted in roughly a 13-fold loss of neutralization by rh-eCD4-IgI39N (Fig. 5C). This Env variant was also similarly more resistant to rh-CD4-IgI39N (Fig. 5D), suggesting that these gp41 changes resulted in less efficient binding to CD4. Two other variants, A131T/I152M and A612T/K643R, did not increase resistance to rh-eCD4-IgI39N.
Resistance mutations to rh-eCD4-IgI39N come at a fitness costWe next determined whether the resistance mutations reported in Fig. 5 had an effect on infection of cells expressing rhesus macaque CD4. We observed that the G382R variant was markedly more efficient at infecting Cf2Th cells expressing rhesus CD4 and human CCR5 than was wild-type SIVmac239 (Fig. 6A). This result is consistent with our previous studies showing that G382R, together with H442Y, facilitated binding to and utilization of rhesus CD4 by adapting to rhesus CD4 isoleucine 39 (29). We also observed a modest loss of infection with the G382R/V444A variant and a more pronounced de-crease for both the K180Q/G384 and the G384 variants, consistent with their respective abilities to bind rh-CD4-Ig. Infection observed with the gp41 mutations Y690C/R774S/A833T was near background,
suggesting a high fitness cost associated with these changes. As shown in Fig 6A, none of these changes conferred CD4 independence to these Envs. To better understand the relationship between the G382R and G382R/V444A mutations in macaque r13058, we analyzed Env sequences from this macaque longitudinally (Fig. 6B). Although the G382R single-mutation variant predominated in weeks 18 through 22 after infection, the double G382R/V444A variant outcompeted this G382R by week 28, despite the greater infectivity of G382R in vitro. Last, we sought to confirm these results on rhesus PBMCs (Fig. 6C). As in Cf2Th-rhCD4/CCR5 cells, the G382R variant replicated more efficiently than wild-type SIVmac239 on these cells, whereas the G382R/V444A replicated less efficiently. Thus, the late emergence of V444A appears to complement the G382R variant by making it more resistant to rh-eCD4-IgI39N, but this alteration associates with a fitness cost.
These data of Figs. 5 and 6 indicate that SIVmac239 can resist rh-eCD4-IgI39N by at least two mechanisms. First, mutations in Env can reduce affinity for the CD4 domain of rh-eCD4-Ig I39N, thereby increasing the IC50 of rh-eCD4-IgI39N. This is the pathway taken by variants with the CD4-binding site mutation G384 and apparently those with gp41 mutations Y690C/R774S/A833T. This pathway associates with a high fitness cost. Second, Env can exploit a difference between rh-eCD4-IgI39N and rhesus CD4 at CD4 residue 39 to increase the efficiency with which it binds and uses rhesus CD4, as observed with the G382R Env variant. This mutation not only increased infection of cells expressing rhesus CD4 but also increases sensitivity to rh-eCD4-IgI39N. A compensatory mutation, V444A, emerged later
Fig. 3. AAV1-delivered rh-eCD4-IgI39N protects rhesus macaques from high-dose challenges with SIVmac239. (A) Kaplan-Meier curves comparing infection of four rh-eCD4-IgI39N–expressing (blue) macaques with eight controls (gray). Challenge doses are indicated on the horizontal axis. **P = 0.002, Mantel-Cox test. (B) Viral loads of macaques expressing rh-eCD4-IgI39N (blue) aligned at the time of infection. Gray lines show viral loads of control macaques that were placed on antiviral therapy on week 12. (C to D) Comparison of viral loads peak and postpeak (mean observed after 5 weeks) between rh-eCD4-IgI39N–expressing macaques (blue) and their corre-sponding controls (gray). Bars indicate mean and SD. *P < 0.05 based on unpaired t test. Viral loads are the result of individual experiments per time point.
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from
Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
5 of 12
to increase resistance to rh-eCD4-IgI39N and again imposed some cost to viral fitness.
DISCUSSIONIn this study, we demonstrate the protective efficacy of AAV-delivered rh-eCD4-IgI39N against SIVmac239 and the in vivo adaptation of SIVmac239 to the presence of rh-eCD4-IgI39N. This study shows complete protection from SIVmac239 at challenge doses that infected all control animals. In addition, we show that all four macaques expressing rh-eCD4-IgI39N for more than a year were protected from high doses of SHIV-AD8. These two studies highlight both the in vivo breadth of eCD4-Ig and the potential of AAV for HIV-1 prophylaxis. However, these studies have several limitations. First, the intravenous challenge route used in these studies, although a consistent method
for infecting macaques is not the primary route of human infection, and the concentrations of rh-eCD4-IgI39N in the mucosa, the site of most viral transmission events, have not been determined. In addition, although eCD4-Ig in these studies was present at concentrations that could potently neutralize SIVmac239, it is formally possible that protection derives in part from xenoresponses to the AAV1 vectors produced in human cells, as the challenge virus is also grown in human cells, and the control macaques did not receive an analogous vector. However, the absence of protection from AAV-expressed neutralizing antibodies (31), where identical xenoresponses would also be expected, argues against any such effect, especially because robust protection was observed with rh-eCD4-IgI39N using the iden-tical challenge virus. In addition, samples from macaques expressing rh-eCD4-IgI39N, and not the control macaques, neutralized SIVmac239 and mediated ADCC activity.
Fig. 4. Full-length SIVmac239 gp160 sequences from infected macaques expressing rh-eCD4-IgI39N show unique mutations in the CD4-binding site and apex region. Distinct apex and CD4-binding site sequences of SIVmac239 cloned from four infected, rh-eCD4-IgI39N–expressing rhesus macaques. Alignments and numbering are based on wild-type SIVmac239 gp160 sequence (top). Rows indicate individual Env clone from a single macaque. Periods indicate residues with no change. Mutations not found in any of eight infected control macaques are highlighted in red and indicated by the single-letter amino acid. Dashes indicate a deletion of residue G384, also unique to rh-eCD4-IgI39N–expressing macaques.
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from

Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
6 of 12
Although rh-eCD4-IgI39N–expressing macaques resisted doses of SIVmac239 that infected all control animals, these animals were eventually infected at higher doses. We observed that, once infected, these macaques had lower peak and postpeak viral loads, despite having received higher challenge doses. These data suggest that rh-eCD4-IgI39N exerts pressure on SIVmac239 over at least 10 to 12 weeks after infection (after which all untreated control animals were placed on combination antiretroviral therapy). Consistent with these observations, we observed in three of four macaques a deletion in a critical region of the CD4-binding site of Env. This G384 muta-tion frequently paired with a mutation in the Env apex region recog-nized by sulfotyrosines of tyrosine-sulfated bNAbs (32, 33). This observation raises the interesting possibility that one of the rh-eCD4-IgI39N sulfopeptides could engage the apex region, either transiently or more stably, before a CD4-induced conformational change disrupts the
apex epitope. Resistance to the CD4-domain of rh-eCD4-IgI39N was also observed because of changes in the cytoplasmic tail of gp41, con-sistent with previous reports that show that perturbation of the cyto-plasmic tail of SIV can increase its neutralization resistance (34, 35).
We further observed a second, unexpected pathway for resistance. Specifically, a CD4-binding site mutation that we have previously shown to accommodate isoleucine 39 of rhesus CD4 (G382R) emerged under pressure from rh-eCD4-IgI39N. The G382R mutation increased sensitivity to rh-eCD4-IgI39N, whereas the G382R/V444A remained as resistant as wild-type SIVmac239. The G382R mutation markedly increased the efficiency with which the SIVmac239 Env used cell- surface rhesus CD4, accommodating isoleucine 39 present on rhesus CD4 but modified to asparagine in rh-eCD4-IgI39N. Of course, this pathway would not be available to HIV-1 in humans presuming a complete identity in the CD4 domains of human eCD4-Ig and
Fig. 5. Two pathways for resistance to rh-eCD4-IgI39N occur in the CD4-binding site or gp41 region. (A) Neutralization assays performed with wild-type SIVmac239 Env and Env variants using the indicated concentration of rh-eCD4-IgI39N (left), rh-CD4-IgI39N (middle), or rh-CD4-Ig (right). Neutralization is normalized to SIVmac239 infection alone. (B) SPR analyses showing the association rate constant (Kon), dissociation rate constant (Koff), and equilibrium dissociation constant (Kd) of SIVmac239 gp120 or gp120 variants to rh-CD4-Ig (left) or rh-CD4-IgI39N (right). (C to D) Neutralization assays evaluating the indicated mutations from three SIVmac239 Env sequences against rh-eCD4-IgI39N (C) or Y690C/R774S/A883T against rh-CD4-IgI39N (D). Error bars in (A), (C), and (D) indicate SEM. Neutralization data are representative of at least three experiments.
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from

Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
7 of 12
human CD4. We have yet to identify any Env that fully escapes any human or rhesus eCD4-Ig variant, either in cell culture studies (36) or, as we show here, in vivo.
The ability of AAV-rh-eCD4-IgI39N to protect from SIVmac239 is strongly suggestive that the same approach would be similarly protective in humans, but also identifies some obstacles with this approach. For example, although AAV-expressed rh-eCD4-IgI39N is regularly less immunogenic than AAV-expressed rhesusized bNAbs, the ADA activity against eCD4-IgI39N appeared to attenuate protection in animal rh2448, due to recognition of the sulfopeptide. Modified sulfopeptide variants may be less immunogenic. In addi-tion, most adult humans are 10 to 12 times larger than macaques, and production of AAV vectors remains expensive, so more efficient eCD4-IgI39N expression and lower production costs will be neces-sary for broad application of this approach. Last, the consequences of long-term expression of eCD4-Ig have not been evaluated in humans, and a means of halting transgene expression may be important to its use in AAV-mediated prophylaxis.
That said, AAV-eCD4-Ig is one of the most effective vaccine or vaccine alternatives thus far tested in primates. Its ability to protect against both SHIV-AD8 and SIVmac239 is unique but wholly consistent with its 100% breadth in cell culture studies. Unlike current con-ventional HIV-1 vaccines, this approach only requires a one-time inoculation, and protection is clearly durable, lasting more than a year. Some of these properties may also extend to combinations of AAV-expressed bNAbs, where available data suggest that the challenges of ADA (22, 31) and population-level escape will nonetheless
be greater. In addition, all AAV-based vaccine approaches require further study of their safety. Nonetheless, our data make clear that AAV-eCD4-Ig remains a viable approach to providing sustained, effective, and universal protection from HIV-1.
MATERIALS AND METHODSStudy designThe purpose of this study was to determine whether rhesus macaques expressing rh-eCD4-IgI39N from AAV1 vectors were protected from SIVmac239 challenges. SIVmac239 is a hard-to-neutralize tier 3 isolate and considered more difficult for vaccine studies than SHIV challenge viruses. In addition, we sought to characterize Env escape mutations that arose in macaques expressing rh-eCD4-IgI39N and infected with SIVmac239. Twelve female macaques were age- and weight-matched and randomly assigned except that the group receiving AAV1-rh-eCD4-IgI39N were consistently AAV1-seronegative before inoculation, and Mamu A*01– and Mamu A*02–positive animals were distributed equally among inoculated and control groups. Mamu A*01 and Mamu A*02 are two rhesus macaque major histocompat-ibility complex alleles that have been described to slow SIV-related disease progression. Each group included at least four macaques, a number that provided statistical significance in our previously pub-lished study (28). Viral load analyses were blinded. All other animal procedures and experiments were not blinded. The number of experimental replicates is indicated in the figure legends. Primary data are available in data file S1.
Fig. 6. Fitness costs associated with resistance to rh-eCD4-IgI39N. (A) Infection assays of Cf2Th cells stably expressing rh-CD4 and CCR5 (left) or CCR5 alone (right) using the indicated p24 amounts of luciferase-expressing HIV-1 NL4.3 pseudotyped with the indicated SIVmac239 gp160. (B) Percentages of observed Env mutations over the course of infection from macaque r13058 in which the G382R variant was isolated. (C) p27 values of samples harvested from rhesus macaque PBMCs infected ex vivo with indicated SIVmac239 variant. Biological triplicates of rhesus macaque PBMCs were infected with 10 ng of replication-competent SIVmac239 or its variants (G382R or G382R/V444A). Error bars in (A) and (C) indicate range of independent results (n = 3). Data are representative of at least two experiments.
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from

Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
8 of 12
Animals, inoculations, and challengesThe 20 animals described in this study were Indian-origin rhesus macaques (Macaca mulatta), between 2 and 5 years of age at the time of AAV1 inoculations. Eight macaques were housed at the New England Primate Research Center (NEPRC) in accordance with standards set forth by the American Association for Accreditation of Laboratory Animal Care. Twelve macaques were housed at the Wisconsin National Primate Research Center (WNPRC) and were similarly cared for with standards set forth by the American Association for Accreditation of Laboratory Animal Care and following the guidelines of the Weatherall Report under a protocol approved by the University of Wisconsin Graduate School Animal Care and Use Committee. All studies were performed with the approval of the appropriate Institutional Animal Care and Use Committees, protocol numbers 04888 (NEPRC) and G005045 (WNPRC). All macaques were AAV1- and SIV-negative at the beginning of the study. All macaques were negative for Mamu B*08 and B*17 alleles. Macaques that were positive for the Mamu A*01 or A*02 alleles were divided evenly among groups when possible. Macaque weights at the time of AAV inoculation ranged from 2.5 to 6.38 kg. Four macaques were inoculated with 2 ml of saline containing 2.0 × 1013 AAV1 GCs encoding the rh-eCD4-IgI39N transgene and 5 × 1012 AAV1 GCs encoding the rhesus macaque TPST2 transgene into both quadriceps (two 0.5 ml per injection per quadriceps muscle). One milliliter of sera was obtained every 1 to 2 weeks after AAV1 inoculation beginning at week 4. Animals were initially challenged intravenously with 20 pg of p27 SIVmac239. Uninfected animals were rechallenged every 4 weeks with twice the previous dose of challenge virus. Animals in the SHIV-AD8 study were challenged with 1600 and 3200 pg of p27 SHIV-AD8. Plasma viral loads were quantified by reverse transcription polymerase chain reaction (RT-PCR) as previously described (37).
Cell lines and plasmidsHEK293T cells were obtained from American Type Culture Collection and grown in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine serum (FBS) at 37°C. TZM-bl cells were obtained through the National Institutes of Health (NIH) AIDS Reagent Program, Division of AIDS, National Institute of Allergy and Infectious Diseases (NIAID), NIH from J. C. Kappes, X. Wu, and Tranzyme Inc. and grown in DMEM with 10% FBS at 37°C (38–42). Cf2Th-rhCD4/R5 and Cf2Th-R5 cells were cultured in DMEM with 10% FBS, G418 (500 g/ml), and puromycin (3 g/ml) at 37°C as previously described (29, 43). ADCC target and effector cells have been described previously (44) and were a gift from M. Alpert and D. Evans (University of Wisconsin, Madison). Briefly, CEM.NKR-CCR5-sLTR-Luc ADCC target cells, harboring an SIV Tat-inducible luciferase, were derived from CEM.NKR-CCR5 CD4+ T cells obtained from the NIH AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH, contributed by A. Trkola, and have been previously described (44–48). Target cells were grown in R10 media, specifically RPMI 1640 supplemented with 10% FBS, 25 mM Hepes, 2 mM l-glutamine, and primocin (0.1 mg/ml; InvivoGen). The KHYG-1–derived natural killer cell (NK cell) line expressing rhesus macaque CD16a (FCGR3A allele) have been previously described (44, 45). KHYG-1–derived NK cells were grown in R10 media additionally supplemented with cyclosporine (1 g/ml) and interleukin-2 (IL-2). The SIVmac239 SpX plasmid was obtained through the NIH AIDS Reagent Program, Division of AIDS, NIAID, and NIH from R. C. Desrosiers (49). Plasmids encoding rh-eCD4-Ig, rh-CD4-IgI39N, rh-CD4-Ig, rh-TPST2, CCR5mim2-Ig, and
SHIV-AD8- EO have been previously described (28). SIVmac239 gp160, NL4.3Env, and NL4.3Env-luc expression plasmids were previously described (29). SIVmac239 gp160 variants were cloned using synthesized gBlocks (Integrated DNA Technologies) con-taining the mutations into the SIVmac239 gp160 expression plasmid. SIVmac239 gp120 and variants were cloned by adding in a stop codon at position 526 of the gp160 expression plasmids by round-the-horn site-directed mutagenesis.
Protein production and purificationProduction of rh-eCD4-IgI39N and rh-CD4-Ig variants was performed as previously described (43). Briefly, HEK293T cells in 175-cm2 flasks were transfected with 80 g of total DNA per flask at 50% con-fluency with a calcium phosphate transfection kit (Takara). Plasmids encoding sulfated proteins were cotransfected with a plasmid-encoding human TPST2 at a 4:1 ratio. At 12 to 16 hours after transfection, 10% FBS-DMEM media was replaced with serum-free 293 Free-style media (Invitrogen). Media was collected after 48 hours, debris was cleared by centrifugation for 10 min at 1500g, and the super-natant was filtered using 0.45-m filter flasks (Millipore). Proteins were isolated with HiTrap columns (GE Healthcare) and eluted with IgG Elution Buffer (Thermo Fisher Scientific) into 1 M tris-HCl Buffer (pH 9.0) (G-Biosciences). Buffer was exchanged with phosphate-buffered saline (PBS) and protein-concentrated to 1 mg/ml with Amicon Ultra Centrifugal Filters (Millipore). Proteins were stored at 4°C.
SIVmac239 gp120s were produced as previously described (50). Briefly, Expi293F cells were transfected SIVmac239 gp120 or variant expressor plasmids using FectoPro transfection reagent (Polyplus Transfection) per the manufacturer’s instructions. Supernatants were collected at 5 days after transfection, filtered with a 0.45-m filter flask, and mixed with 1 ml of Capto Lentil Lectin beads (GE Healthcare) per 200 ml of supernatant overnight at 4°C. Supernatants were then run through a column, and beads were collected. The beads were washed with 30 ml of 0.65 M NaCl in PBS and then with 20 ml of PBS. A total of 6 ml of 1 M methyl--d mannopyranoside (in PBS) was added to the beads and incubated at 4°C for 1 to 2 hours. The eluate was collected and subjected to buffer exchange in PBS using Amicon Ultra Centrifugal Filters (Millipore) to a concentration of 1 mg/ml. gp120 proteins were stored at 4°C until use.
TZM-bl neutralization assayTZM-bl neutralization assays were performed as previously described (51). Briefly, SIVmac239 was preincubated with titrated amounts of inhibitor or serum in DMEM (10% FBS) for 1 hour at 37°C. TZM-bl cells were detached by trypsinization, diluted in DMEM (10% FBS) to 100,000 cells/ml, and added to the virus/inhibitor mixture. Cells were then incubated for 44 hours at 37°C. Viral entry was analyzed using Britelite Plus (PerkinElmer), and luciferase was measured using a Victor X3 plate reader (PerkinElmer).
Luciferase-based antibody-dependent cell cytotoxicity assayADCC assays were performed as previously described (27). Briefly, CEM.NKR-CCR5-sLTR-Luc target cells were infected by spinoculation with SIVmac239SpX (200 ng of p27) 4 days before assay. On the day of the assay, infected target cells were mixed with NK effector cells at a 10:1 ratio in the presence of heat-inactivated serum. Cells were mixed with heat-inactivated serum samples or known concentrations of rh-eCD4-IgI39N and incubated for 8 hours at 37°C. ADCC activity
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from

Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
9 of 12
was measured by luciferase using BriteLite Plus (PerkinElmer) and measured using a Victor X3 plate reader (PerkinElmer).
AAV production and purificationProduction of recombinant AAV, at the University of Massachusetts Medical School Vector Core, has been previously described (52). In short, HEK293T cells were transfected with an AAV vector plasmid, a plasmid encoding AAV2 rep and AAV1 cap, and helper plasmid encoding adenovirus genes. After harvesting lysates of transfected cells, AAV was purified through three sequential cesium chloride centrifugation steps. The vector genome copy number (in GC/ml) was assessed by RT-PCR. The integrity of AAV particles was verified by electron microscopy, and the purity of the AAV preparations was verified by silver-stained SDS–polyacrylamide gel electrophoresis.
AAV neutralization assayRhesus macaques were screened for preexisting AAV1-neutralizing antibodies using an AAV1.LacZ-neutralizing assay. Specifically, Huh7 cells were added to 96-well plates at 5 × 104 cells per well and incubated overnight at 37°C. The next day, 5 × 106 wild-type adeno-virus diluted in unsupplemented DMEM was added to each well. Serum samples from macaques were diluted 1:5, 1:10, and 1:20 in DMEM supplemented with 10% FBS. Serum dilutions were then mixed with AAV1.LacZ vector for 1 hour at 37°C. The mixture of AAV1.LacZ vector and macaque serum was added to the plates of Huh7 cells and incubated for 1 hour at 37°C. A total of 100 l of medium was removed from the 96-well plates and replaced with DMEM supplemented with 20% FBS. Plates were incubated overnight at 37°C. One day after infection, 40 l of medium was removed from the 96-well plates, and cells were washed with 200 l PBS. All supernatant was removed, and 40 l of lysis buffer (Thermo Fisher Scientific) was added to the cells. Plates were incubated for 10 min, and 100 l of substrate and reaction buffer (Thermo Fisher Scientific) was added. Plates were incubated for 1 hour, and LacZ production was measured on a luminometer (Clarity, Bio-Tek). Infection was calculated by dividing the signal observed in wells from the serum samples by the amount of signal observed in wells without serum.
Challenge virus and pseudovirus productionSIVmac239 for challenge studies was made in HEK293T cells plated in 175-cm2 flasks and transfected with 80 g of SIVmac239 SpX DNA per flask by calcium phosphate technique. Pseudoviruses for neutraliza-tion studies were produced in HEK293T cells plated in 175-cm2 flasks and transfected with 40 g of NL4.3Env-luc expression plasmid and 40 g of SIVmac239 gp160 expression plasmid DNA per flask using the jetPRIME transfection reagent (Polyplus Transfection). At 12 hours after transfection, medium was replaced with fresh DMEM (10% FBS). Medium was harvested at 48 hours after transfection, and debris was cleared by centrifugation for 10 min at 1500g and filtered using 0.45-m filter flasks (Millipore). Virus stocks were aliquoted and frozen at −80°C. Virus titers were determined by an SIV p27 enzyme- linked immunosorbent assay (ELISA) kit Advanced Bioscience Labo-ratories (ABL) for challenge studies and infections studies of rhesus PBMCs. Viral titers were determined by an HIV p24 ELISA kit (ABL) for pseudovirus infection studies.
gp120 and anti-transgene ELISAsHalf-area 96-well Costar assay plates (Corning) were coated with gp120 (5 g/ml; Immune Tech). Plates were washed with PBS-T
(PBS + 0.05% Tween-20) twice and blocked with 5% bovine serum albumin (BSA) in PBS for 1 hour at 37°C. Serum samples were serially diluted in 5% BSA in PBS and were added to the plate in duplicate. Standard curves were generated with a starting concentration of standard protein (4 g/ml) in 5% BSA then serially diluted and added to the plate in duplicate. Plates were incubated for 1 hour at 37°C. Samples were washed five times with PBS-T, and a horseradish peroxidase secondary antibody was added. To determine rh-eCD4-IgI39N con-centrations, a secondary antibody recognizing human IgG1 (Jackson ImmunoResearch) was used. Plates were incubated for 1 hour at 37°C and then washed 10 times with PBS-T. 3,3′,5,5′-Tetramethylbenzidine (TMB) solution (Thermo Fisher Scientific) was added, and plates were incubated at room temperature until their standard curves developed (typically 5 to 10 min). TMB Stop Solution (SouthernBiotech) was then added, and absorbance was read at 450 nm by a Victor X3 plate reader (PerkinElmer). Standard curves were analyzed using GraphPad Prism 6.0 software and used to determine protein titers from sera samples.
Anti–rh-eCD4-IgI39N antibodies were measured in the same manner except that ELISA plates were coated with various constructs (5 g/ml). Serum samples from macaques expressing rh-eCD4-IgI39N were assayed against rh-eCD4-IgI39N, rh-CD4-IgI39N, C-terminal CCR5mim2-Ig (human IgG1 Fc and hinge without CD4 domains), and the ecto-domains of human and rhesus macaque CD4 (Immune Tech). Serum samples were diluted 10-fold and blocked in 5% BSA in PBS. Anti-transgene antibodies were detected using anti- and anti- secondary antibodies (SouthernBiotech). Plates were incubated for 1 hour at 37°C. TMB solution was added for 15 min at room temperature. Stop solution was then added, and absorbance at 450 nm was measured as described above.
Full-length env sequencingSIVmac239 gp160 sequences were obtained from infected plasma samples as previously described (3, 53). Viral RNA was isolated from plasma samples using a QIAamp viral RNA kit (Qiagen). Isolated RNA was reverse-transcribed to cDNA using SuperScript III reverse transciptase (Invitrogen). cDNA synthesis was performed using 1× First-Strand buffer (Invitrogen), 0.5 mM dNTP (New England Biolabs), 5 mM dithiothreitol (DTT), 40 units (U) of RNaseOUT (Invitrogen), 200 U of SuperScript III reverse transcriptase, and 0.25 mM SIVEnvR1 antisense primer (5′-TGTAATAAATCCCTTCCAGTC-CCCCC-3′). RNA, SIVEnvR1, and dNTPs were heated at 65°C for 5 min followed by 1 min at 4°C. First-Strand buffer, dNTP, DTT, and RNaseOUT were then added, and the reaction mixture was incu-bated for 1 hour at 50°C followed by 55°C for 1 hour. The reaction mixture was heat-inactivated for 15 min at 70°C. A total of 1 l of RNase H (New England Biolabs) was added, and the reaction mix-ture was incubated for 20 min at 37°C. SIVmac239 gp160 sequences were PCR-amplified using the Platinum Taq (Invitrogen) with the following mixture conditions: 1× buffer, 2 mM MgCl2, 0.2 mM dNTP, 0.2 M of sense and antisense primer, and 0.025 U of Platinum Taq polymerase. For the first round of PCR, the primers used were SIVEnvF1 (5′-CCTCCCCCTCCAGGACTAGC-3′) and SIVEnvR1. The primers used for the second round were SIVEnvF2 (5′-GTTTCTTTATAATAGACATGGAGA CACCCTTGAGGGAGC-3′) and SIVEnvR2 (5′-GTTTCTTATGAGACATGTCTATTGCCAATTTG-TA-3′). Cycle conditions were 94°C for 2 min, then 35 cycles of 94°C for 15 s, 55°C for 30 s, and 68°C for 4 min, and a final extension of 68°C for 10 min. A total of 2 l of the first PCR was used as the
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from

Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
10 of 12
template for the second PCR that used the same conditions but were run for a total of 45 cycles. After the second PCR, 2.5 U of Platinum Taq polymerase was added, and the reaction mixture was incubated at 72°C for 15 min. The PCR product was gel-extracted and eluted with 20 l of elution buffer (Qiagen). The gel-purified PCR product was then cloned into the pcDNA3.1/V5-His vector by Topo TA cloning per the manufacturer’s instructions. Cloned vectors were then trans-formed using Top10 competent cells. Sequencing of the SIVmac239 env was performed using the following primers: SIVEnvF3 (5′-GGTAAT-CATATCTATAATAGACATGGAGACAC-3′), mac239EnvSeqF1 (5′-GGGGAACAACTCAGTGCCTACC-3′), mac239EnvSeqF3 (5′-GCACCTCCAGGTTATGCTTTGC-3′), mac239EnvSeqF5 (5′-GCA-GAGGAGAGTTCCTCTAC-3′), mac239EnvSeqF7 (5′-GCAACAGCT-GTTGGACGTGG-3′), and mac239EnvSeqF9 (5′-CCAGCAGAC-CCATATCCAACAGG-3′). Sequences were aligned to wild-type SIVmac239 gp160 to identify mutations using the Los Alamos Highlighter tool (www.hiv.lanl.gov/content/sequence/HIGHLIGHT/highlighter_top.html).
Surface plasmon resonanceSPR analysis was performed using a Biacore X100, Biacore software, and Biacore reagents (GE Healthcare). A mouse antihuman IgG antibody was immobilized on a CM5 censor based on the manu-facturer’s instructions (GE Healthcare). rh-CD4-Ig (2 g/ml) and rh-CD4-IgI39N proteins were captured by the antihuman IgG anti-body. For affinity analysis, SIVmac239 gp120s were injected at 400, 200, 100, 50, and 25 nM concentrations. Each analysis included an empty flow cell to deduct background signal. Sensor chips were regenerated after each concentration of gp120 using 3 M MgCl2. Association (kon) and dissociation (koff) rate constants were calculated using a 1:1 Langmuir binding model via the Biacore software. The equilibrium dissociation constant (Kd) was calculated from koff/kon.
Infection assaysHIV-1 pseudovirus bearing SIVmac239 variant Envs and encoding firefly luciferase were diluted to 30 ng of p24 and serially diluted threefold in DMEM (10% FBS). CD4-negative and rhesus CD4-positive Cf2Th-CCR5 cells were harvested and diluted in DMEM (10% FBS) to 100,000 cells/ml and added to the pseudovirus/inhibitor mixture. Cells were then incubated for 48 hours at 37°C. Viral entry was analyzed using Britelite Plus (PerkinElmer), and luciferase was measured by a Victor X3 plate reader (PerkinElmer).
Rhesus macaque PBMCs from a single donor (iQ Biosciences) isolated by Ficoll gradient centrifugation were infected with replication- competent SIVmac239 or variants with the G382R or G382R/V444A mutations as previously described (28). Briefly, rhesus PBMCs were thawed and grown for 3 days in RPMI 1640 supplemented with 20% FBS, penicillin/streptomycin (100 U/ml), 10 mM Hepes, and phytohemagglutinin-L (PHA-L) (5 g/ml). Rhesus PBMCs were then split into a 12-well plate at 1 × 106 cells/ml in RPMI 1640 supplemented with 20% FBS, penicillin/streptomycin (100 U/ml), 10 mM Hepes, and human IL-2 (40 U/ml; growth medium). Cells were infected with 10 ng of SIVmac239 or one of the variants in biological triplicates for 3 hours at 37°C. Cells were pelleted and washed with unsupplemented RPMI 1640, and medium was replaced with fresh growth medium. At days 1, 3, 6, 9, and 12 after infection, supernatants were collected, and cells were pelleted and replaced with fresh growth medium. Supernatants were analyzed for p27 amounts using an SIV p27 ELISA kit (ABL).
Statistical analysesAll statistical analyses were performed using GraphPad Prism 6.07 software (GraphPad Software). Comparisons of groups were performed using Student’s t test (two-tailed) or Mantel-Cox test as reported in the figure legends. A P < 0.05 was considered significant.
SUPPLEMENTARY MATERIALSstm.sciencemag.org/cgi/content/full/11/502/eaau5409/DC1Fig. S1. AAV1-delivered rh-eCD4-IgI39N affords complete protection from high-dose challenges with SHIV-AD8.Fig. S2. Functional characterization of rh-eCD4-IgI39N expressed from an AAV gene cassette.Fig. S3. In vivo rh-eCD4-IgI39N concentrations plotted against number of SIVmac239 challenges until infection.Fig. S4. Protection from SIVmac239 is not due to a xenogenic response to the AAV1 vector.Fig. S5. Alignment of full-length SIVmac239 gp160 sequences cloned from infected macaques.Fig. S6. Sensograms of SIVmac239 gp120 variants binding rh-CD4-Ig and rh-CD4-IgI39N.Data file S1. Primary data.
REFERENCES AND NOTES 1. P. D. Kwong, J. R. Mascola, HIV-1 vaccines based on antibody identification, B cell
ontogeny, and epitope structure. Immunity 48, 855–871 (2018). 2. R. Gautam, Y. Nishimura, N. Gaughan, A. Gazumyan, T. Schoofs, A. Buckler-White,
M. S. Seaman, B. J. Swihart, D. A. Follmann, M. C. Nussenzweig, M. A. Martin, A single injection of crystallizable fragment domain–modified antibodies elicits durable protection from SHIV infection. Nat. Med. 24, 610–616 (2018).
3. B. Julg, L. J. Tartaglia, B. F. Keele, K. Wagh, A. Pegu, D. Sok, P. Abbink, S. D. Schmidt, K. Wang, X. Chen, M. G. Joyce, I. S. Georgiev, M. Choe, P. D. Kwong, N. A. Doria-Rose, K. Le, M. K. Louder, R. T. Bailer, P. L. Moore, B. Korber, M. S. Seaman, S. S. Abdool Karim, L. Morris, R. A. Koup, J. R. Mascola, D. R. Burton, D. H. Barouch, Broadly neutralizing antibodies targeting the HIV-1 envelope V2 apex confer protection against a clade C SHIV challenge. Sci. Transl. Med. 9, eaal1321 (2017).
4. B. Julg, A. Pegu, P. Abbink, J. Liu, A. Brinkman, K. Molloy, S. Mojta, A. Chandrashekar, K. Callow, K. Wang, X. Chen, S. D. Schmidt, J. Huang, R. A. Koup, M. S. Seaman, B. F. Keele, J. R. Mascola, M. Connors, D. H. Barouch, Virological control by the CD4-binding site antibody N6 in simian-human immunodeficiency virus-infected rhesus monkeys. J. Virol. 91, e00498-17 (2017).
5. B. Julg, P.-T. Liu, K. Wagh, W. M. Fischer, P. Abbink, N. B. Mercado, J. B. Whitney, J. P. Nkolola, K. McMahan, L. J. Tartaglia, E. N. Borducchi, S. Khatiwada, M. Kamath, J. A. LeSuer, M. S. Seaman, S. D. Schmidt, J. R. Mascola, D. R. Burton, B. T. Korber, D. H. Barouch, Protection against a mixed SHIV challenge by a broadly neutralizing antibody cocktail. Sci. Transl. Med. 9, eaao4235 (2017).
6. J. Liu, K. Ghneim, D. Sok, W. J. Bosche, Y. Li, E. Chipriano, B. Berkemeier, K. Oswald, E. Borducchi, C. Cabral, L. Peter, A. Brinkman, M. Shetty, J. Jimenez, J. Mondesir, B. Lee, P. Giglio, A. Chandrashekar, P. Abbink, A. Colantonio, C. Gittens, C. Baker, W. Wagner, M. G. Lewis, W. Li, R.-P. Sekaly, J. D. Lifson, D. R. Burton, D. H. Barouch, Antibody-mediated protection against SHIV challenge includes systemic clearance of distal virus. Science 353, 1045–1049 (2016).
7. R. Gautam, Y. Nishimura, A. Pegu, M. C. Nason, F. Klein, A. Gazumyan, J. Golijanin, A. Buckler-White, R. Sadjadpour, K. Wang, Z. Mankoff, S. D. Schmidt, J. D. Lifson, J. R. Mascola, M. C. Nussenzweig, M. A. Martin, A single injection of anti-HIV-1 antibodies protects against repeated SHIV challenges. Nature 533, 105–109 (2016).
8. M. Shingai, Y. Nishimura, F. Klein, H. Mouquet, O. K. Donau, R. Plishka, A. Buckler-White, M. Seaman, M. Piatak, J. D. Lifson, D. Dimitrov, M. C. Nussenzweig, M. A. Martin, Antibody-mediated immunotherapy of macaques chronically infected with SHIV suppresses viraemia. Nature 503, 277–280 (2013).
9. D. H. Barouch, J. B. Whitney, B. Moldt, F. Klein, T. Y. Oliveira, J. Liu, K. E. Stephenson, H.-W. Chang, K. Shekhar, S. Gupta, J. P. Nkolola, M. S. Seaman, K. M. Smith, E. N. Borducchi, C. Cabral, J. Y. Smith, S. Blackmore, S. Sanisetty, J. R. Perry, M. Beck, M. G. Lewis, W. Rinaldi, A. K. Chakraborty, P. Poignard, M. C. Nussenzweig, D. R. Burton, Therapeutic efficacy of potent neutralizing HIV-1-specific monoclonal antibodies in SHIV-infected rhesus monkeys. Nature 503, 224–228 (2013).
10. M. R. Gaudinski, E. E. Coates, K. V. Houser, G. L. Chen, G. Yamshchikov, J. G. Saunders, L. A. Holman, I. Gordon, S. Plummer, C. S. Hendel, M. Conan-Cibotti, M. G. Lorenzo, S. Sitar, K. Carlton, C. Laurencot, R. T. Bailer, S. Narpala, A. B. McDermott, A. M. Namboodiri, J. P. Pandey, R. M. Schwartz, Z. Hu, R. A. Koup, E. Capparelli, B. S. Graham, J. R. Mascola, J. E. Ledgerwood; VRC 606 Study Team, Safety and pharmacokinetics of the Fc-modified HIV-1 human monoclonal antibody VRC01LS: A phase 1 open-label clinical trial in healthy adults. PLOS Med. 15, e1002493 (2018).
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from

Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
11 of 12
11. K. H. Mayer, K. E. Seaton, Y. Huang, N. Grunenberg, A. Isaacs, M. Allen, J. E. Ledgerwood, I. Frank, M. E. Sobieszczyk, L. R. Baden, B. Rodriguez, H. Van Tieu, G. D. Tomaras, A. Deal, D. Goodman, R. T. Bailer, G. Ferrari, R. Jensen, J. Hural, B. S. Graham, J. R. Mascola, L. Corey, D. C. Montefiori; HVTN 104 Protocol Team; NIAID HIV Vaccine Trials Network, Safety, pharmacokinetics, and immunological activities of multiple intravenous or subcutaneous doses of an anti-HIV monoclonal antibody, VRC01, administered to HIV-uninfected adults: Results of a phase 1 randomized trial. PLOS Med. 14, e1002435 (2017).
12. M. Caskey, T. Schoofs, H. Gruell, A. Settler, T. Karagounis, E. F. Kreider, B. Murrell, N. Pfeifer, L. Nogueira, T. Y. Oliveira, G. H. Learn, Y. Z. Cohen, C. Lehmann, D. Gillor, I. Shimeliovich, C. Unson-O’Brien, D. Weiland, A. Robles, T. Kummerle, C. Wyen, R. Levin, M. Witmer-Pack, K. Eren, C. Ignacio, S. Kiss, A. P. West Jr., H. Mouquet, B. S. Zingman, R. M. Gulick, T. Keler, P. J. Bjorkman, M. S. Seaman, B. H. Hahn, G. Fätkenheuer, S. J. Schlesinger, M. C. Nussenzweig, F. Klein, Antibody 10-1074 suppresses viremia in HIV-1-infected individuals. Nat. Med. 23, 185–191 (2017).
13. J. F. Scheid, J. A. Horwitz, Y. Bar-On, E. F. Kreider, C.-L. Lu, J. C. C. Lorenzi, A. Feldmann, M. Braunschweig, L. Nogueira, T. Oliveira, I. Shimeliovich, R. Patel, L. Burke, Y. Z. Cohen, S. Hadrigan, A. Settler, M. Witmer-Pack, A. P. West Jr., B. Juelg, T. Keler, T. Hawthorne, B. Zingman, R. M. Gulick, N. Pfeifer, G. H. Learn, M. S. Seaman, P. J. Bjorkman, F. Klein, S. J. Schlesinger, B. D. Walker, B. H. Hahn, M. C. Nussenzweig, M. Caskey, HIV-1 antibody 3BNC117 suppresses viral rebound in humans during treatment interruption. Nature 535, 556–560 (2016).
14. K. J. Bar, M. C. Sneller, L. J. Harrison, J. S. Justement, E. T. Overton, M. E. Petrone, D. B. Salantes, C. A. Seamon, B. Scheinfeld, R. W. Kwan, G. H. Learn, M. A. Proschan, E. F. Kreider, J. Blazkova, M. Bardsley, E. W. Refsland, M. Messer, K. E. Clarridge, N. B. Tustin, P. J. Madden, K. Oden, S. J. O’Dell, B. Jarocki, A. R. Shiakolas, R. L. Tressler, N. A. Doria-Rose, R. T. Bailer, J. E. Ledgerwood, E. V. Capparelli, R. M. Lynch, B. S. Graham, S. Moir, R. A. Koup, J. R. Mascola, J. A. Hoxie, A. S. Fauci, P. Tebas, T.-W. Chun, Effect of HIV antibody VRC01 on viral rebound after treatment interruption. N. Engl. J. Med. 375, 2037–2050 (2016).
15. R. M. Lynch, E. Boritz, E. E. Coates, A. DeZure, P. Madden, P. Costner, M. E. Enama, S. Plummer, L. Holman, C. S. Hendel, I. Gordon, J. Casazza, M. Conan-Cibotti, S. A. Migueles, R. Tressler, R. T. Bailer, A. McDermott, S. Narpala, S. O’Dell, G. Wolf, J. D. Lifson, B. A. Freemire, R. J. Gorelick, J. P. Pandey, S. Mohan, N. Chomont, R. Fromentin, T.-W. Chun, A. S. Fauci, R. M. Schwartz, R. A. Koup, D. C. Douek, Z. Hu, E. Capparelli, B. S. Graham, J. R. Mascola, J. E. Ledgerwood; VRC 601 Study Team, Virologic effects of broadly neutralizing antibody VRC01 administration during chronic HIV-1 infection. Sci. Transl. Med. 7, 319ra206 (2015).
16. J. E. Ledgerwood, E. E. Coates, G. Yamshchikov, J. G. Saunders, L. Holman, M. E. Enama, A. DeZure, R. M. Lynch, I. Gordon, S. Plummer, C. S. Hendel, A. Pegu, M. Conan-Cibotti, S. Sitar, R. T. Bailer, S. Narpala, A. McDermott, M. Louder, S. O’Dell, S. Mohan, J. P. Pandey, R. M. Schwartz, Z. Hu, R. A. Koup, E. Capparelli, J. R. Mascola, B. S. Graham; VRC 602 Study Team, Safety, pharmacokinetics and neutralization of the broadly neutralizing HIV-1 human monoclonal antibody VRC01 in healthy adults. Clin. Exp. Immunol. 182, 289–301 (2015).
17. M. Caskey, F. Klein, J. C. C. Lorenzi, M. S. Seaman, A. P. West Jr., N. Buckley, G. Kremer, L. Nogueira, M. Braunschweig, J. F. Scheid, J. A. Horwitz, I. Shimeliovich, S. Ben-Avraham, M. Witmer-Pack, M. Platten, C. Lehmann, L. A. Burke, T. Hawthorne, R. J. Gorelick, B. D. Walker, T. Keler, R. M. Gulick, G. Fatkenheuer, S. J. Schlesinger, M. C. Nussenzweig, Viraemia suppressed in HIV-1-infected humans by broadly neutralizing antibody 3BNC117. Nature 522, 487–491 (2015).
18. G. Buchlis, G. M. Podsakoff, A. Radu, S. M. Hawk, A. W. Flake, F. Mingozzi, K. A. High, Factor IX expression in skeletal muscle of a severe hemophilia B patient 10 years after AAV-mediated gene transfer. Blood 119, 3038–3041 (2012).
19. A. B. Balazs, Y. Ouyang, C. M. Hong, J. Chen, S. M. Nguyen, D. S. Rao, D. S. An, D. Baltimore, Vectored immunoprophylaxis protects humanized mice from mucosal HIV transmission. Nat. Med. 20, 296–300 (2014).
20. A. B. Balazs, J. Chen, C. M. Hong, D. S. Rao, L. Yang, D. Baltimore, Antibody-based protection against HIV infection by vectored immunoprophylaxis. Nature 481, 81–84 (2012).
21. P. R. Johnson, B. C. Schnepp, J. Zhang, M. J. Connell, S. M. Greene, E. Yuste, R. C. Desrosiers, K. Reed Clark, Vector-mediated gene transfer engenders long-lived neutralizing activity and protection against SIV infection in monkeys. Nat. Med. 15, 901–906 (2009).
22. J. M. Martinez-Navio, S. P. Fuchs, S. Pedreño-López, E. G. Rakasz, G. Gao, R. C. Desrosiers, Host anti-antibody responses following adeno-associated virus-mediated delivery of antibodies against HIV and SIV in rhesus monkeys. Mol. Ther. 24, 76–86 (2016).
23. K. O. Saunders, L. Wang, M. G. Joyce, Z.-Y. Yang, A. B. Balazs, C. Cheng, S.-Y. Ko, W.-P. Kong, R. S. Rudicell, I. S. Georgiev, L. Duan, K. E. Foulds, M. Donaldson, L. Xu, S. D. Schmidt, J.-P. Todd, D. Baltimore, M. Roederer, A. T. Haase, P. D. Kwong, S. S. Rao, J. R. Mascola, G. J. Nabel, Broadly neutralizing human immunodeficiency virus type 1 antibody gene transfer protects nonhuman primates from mucosal simian-human immunodeficiency virus infection. J. Virol. 89, 8334–8345 (2015).
24. S. P. Fuchs, J. M. Martinez-Navio, M. Piatak Jr., J. D. Lifson, G. Gao, R. C. Desrosiers, AAV-delivered antibody mediates significant protective effects against SIVmac239 challenge in the absence of neutralizing activity. PLOS Pathog. 11, e1005090 (2015).
25. H. C. Welles, M. F. Jennewein, R. D. Mason, S. Narpala, L. Wang, C. Cheng, Y. Zhang, J.-P. Todd, J. D. Lifson, A. B. Balazs, G. Alter, A. B. McDermott, J. R. Mascola, M. Roederer, Vectored delivery of anti-SIV envelope targeting mAb via AAV8 protects rhesus macaques from repeated limiting dose intrarectal swarm SIVsmE660 challenge. PLOS Pathog. 14, e1007395 (2018).
26. I. Fetzer, M. R. Gardner, M. E. Davis-Gardner, N. R. Prasad, B. Alfant, J. A. Weber, M. Farzan, eCD4-Ig variants that more potently neutralize HIV-1. J. Virol. 92, e02011-17 (2018).
27. M. E. Davis-Gardner, M. R. Gardner, B. Alfant, M. Farzan, eCD4-Ig promotes ADCC activity of sera from HIV-1-infected patients. PLOS Pathog. 13, e1006786 (2017).
28. M. R. Gardner, L. M. Kattenhorn, H. R. Kondur, M. von Schaewen, T. Dorfman, J. J. Chiang, K. G. Haworth, J. M. Decker, M. D. Alpert, C. C. Bailey, E. S. Neale, C. H. Fellinger, V. R. Joshi, S. P. Fuchs, J. M. Martinez-Navio, B. D. Quinlan, A. Y. Yao, H. Mouquet, J. Gorman, B. Zhang, P. Poignard, M. C. Nussenzweig, D. R. Burton, P. D. Kwong, M. Piatak, J. D. Lifson, G. Gao, R. C. Desrosiers, D. T. Evans, B. H. Hahn, A. Ploss, P. M. Cannon, M. S. Seaman, M. Farzan, AAV-expressed eCD4-Ig provides durable protection from multiple SHIV challenges. Nature 519, 87–91 (2015).
29. C. H. Fellinger, M. R. Gardner, C. C. Bailey, M. Farzan, Simian immunodeficiency virus SIVmac239, but not SIVmac316, binds and utilizes human CD4 more efficiently than rhesus CD4. J. Virol. 91, e00847-17 (2017).
30. D. Humes, S. Emery, E. Laws, J. Overbaugh, A species-specific amino acid difference in the macaque CD4 receptor restricts replication by global circulating HIV-1 variants representing viruses from recent infection. J. Virol. 86, 12472–12483 (2012).
31. M. R. Gardner, I. Fetzer, L. M. Kattenhorn, M. E. Davis-Gardner, A. S. Zhou, B. Alfant, J. A. Weber, H. R. Kondur, J. M. Martinez-Navio, S. P. Fuchs, R. C. Desrosiers, G. Gao, J. D. Lifson, M. Farzan, Anti-drug antibody responses impair prophylaxis mediated by AAV-delivered HIV-1 broadly neutralizing antibodies. Mol. Ther. 27, 650–660 (2019).
32. J. H. Lee, R. Andrabi, C.-Y. Su, A. Yasmeen, J.-P. Julien, L. Kong, N. C. Wu, R. McBride, D. Sok, M. Pauthner, C. A. Cottrell, T. Nieusma, C. Blattner, J. C. Paulson, P. J. Klasse, I. A. Wilson, D. R. Burton, A. B. Ward, A broadly neutralizing antibody targets the dynamic HIV envelope trimer apex via a long, rigidified, and anionic -hairpin structure. Immunity 46, 690–702 (2017).
33. D. Sok, M. J. van Gils, M. Pauthner, J.-P. Julien, K. L. Saye-Francisco, J. Hsueh, B. Briney, J. H. Lee, K. M. Le, P. S. Lee, Y. Hua, M. S. Seaman, J. P. Moore, A. B. Ward, I. A. Wilson, R. W. Sanders, D. R. Burton, Recombinant HIV envelope trimer selects for quaternary-dependent antibodies targeting the trimer apex. Proc. Natl. Acad. Sci. U.S.A. 111, 17624–17629 (2014).
34. E. White, F. Wu, E. Chertova, J. Bess, J. D. Roser, J. D. Lifson, V. M. Hirsch, Truncating the gp41 cytoplasmic tail of simian immunodeficiency virus decreases sensitivity to neutralizing antibodies without increasing the envelope content of virions. J. Virol. 92, e01688-17 (2018).
35. E. Yuste, W. Johnson, G. N. Pavlakis, R. C. Desrosiers, Virion envelope content, infectivity, and neutralization sensitivity of simian immunodeficiency virus. J. Virol. 79, 12455–12463 (2005).
36. C. H. Fellinger, M. R. Gardner, J. A. Weber, B. Alfant, A. S. Zhou, M. Farzan, eCD4-Ig limits HIV-1 escape more effectively than CD4-Ig or a broadly neutralizing antibody. J. Virol. 93, e00443-19 (2019).
37. A. N. Cline, J. W. Bess, M. Piatak Jr., J. D. Lifson, Highly sensitive SIV plasma viral load assay: Practical considerations, realistic performance expectations, and application to reverse engineering of vaccines for AIDS. J. Med. Primatol. 34, 303–312 (2005).
38. E. J. Platt, M. Bilska, S. L. Kozak, D. Kabat, D. C. Montefiori, Evidence that ecotropic murine leukemia virus contamination in TZM-bl cells does not affect the outcome of neutralizing antibody assays with human immunodeficiency virus type 1. J. Virol. 83, 8289–8292 (2009).
39. Y. Takeuchi, M. O. McClure, M. Pizzato, Identification of gammaretroviruses constitutively released from cell lines used for human immunodeficiency virus Research. J. Virol. 82, 12585–12588 (2008).
40. X. Wei, J. M. Decker, H. Liu, Z. Zhang, R. B. Arani, J. M. Kilby, M. S. Saag, X. Wu, G. M. Shaw, J. C. Kappes, Emergence of resistant human immunodeficiency virus type 1 in patients receiving fusion inhibitor (T-20) monotherapy. Antimicrob. Agents Chemother. 46, 1896–1905 (2002).
41. C. A. Derdeyn, J. M. Decker, J. N. Sfakianos, X. Wu, W. A. O’Brien, L. Ratner, J. C. Kappes, G. M. Shaw, E. Hunter, Sensitivity of human immunodeficiency virus type 1 to the fusion inhibitor T-20 is modulated by coreceptor specificity defined by the V3 loop of gp120. J. Virol. 74, 8358–8367 (2000).
42. E. J. Platt, K. Wehrly, S. E. Kuhmann, B. Chesebro, D. Kabat, Effects of CCR5 and CD4 cell surface concentrations on infections by macrophagetropic isolates of human immunodeficiency virus type 1. J. Virol. 72, 2855–2864 (1998).
43. M. R. Gardner, C. H. Fellinger, N. R. Prasad, A. S. Zhou, H. R. Kondur, V. R. Joshi, B. D. Quinlan, M. Farzan, CD4-induced antibodies promote association of the HIV-1 envelope glycoprotein with CD4-binding site antibodies. J. Virol. 90, 7822–7832 (2016).
44. M. D. Alpert, L. N. Heyer, D. E. J. Williams, J. D. Harvey, T. Greenough, M. Allhorn, D. T. Evans, A novel assay for antibody-dependent cell-mediated cytotoxicity against
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from

Gardner et al., Sci. Transl. Med. 11, eaau5409 (2019) 24 July 2019
S C I E N C E T R A N S L A T I O N A L M E D I C I N E | R E S E A R C H A R T I C L E
12 of 12
HIV-1- or SIV-infected cells reveals incomplete overlap with antibodies measured by neutralization and binding assays. J. Virol. 86, 12039–12052 (2012).
45. M. D. Alpert, J. D. Harvey, W. A. Lauer, R. K. Reeves, M. Piatak Jr., A. Carville, K. G. Mansfield, J. D. Lifson, W. Li, R. C. Desrosiers, R. P. Johnson, D. T. Evans, ADCC develops over time during persistent infection with live-attenuated SIV and is associated with complete protection against SIVmac251 challenge. PLOS Pathog. 8, e1002890 (2012).
46. A. Trkola, J. Matthews, C. Gordon, T. Ketas, J. P. Moore, A Cell line-based neutralization assay for primary human immunodeficiency virus type 1 isolates that use either the CCR5 or the CXCR4 coreceptor. J. Virol. 73, 8966–8974 (1999).
47. H. K. Lyerly, D. L. Reed, T. J. Matthews, A. J. Langlois, P. A. Ahearne, S. R. Petteway Jr., K. J. Weinhold, Anti-GP 120 antibodies from HIV seropositive individuals mediate broadly reactive anti-HIV ADCC. AIDS Res. Hum. Retroviruses 3, 409–422 (1987).
48. D. N. Howell, P. E. Andreotti, J. R. Dawson, P. Cresswell, Natural killing target antigens as inducers of interferon: Studies with an immunoselected, natural killing-resistant human T lymphoblastoid cell line. J. Immunol. 134, 971–976 (1985).
49. D. A. Regier, R. C. Desrosiers, The complete nucleotide sequence of a pathogenic molecular clone of simian immunodeficiency virus. AIDS Res. Hum. Retroviruses 6, 1221–1231 (1990).
50. K. Honarmand Ebrahimi, G. M. West, R. Flefil, Mass spectrometry approach and ELISA reveal the effect of codon optimization on N-linked glycosylation of HIV-1 gp120. J. Proteome Res. 13, 5801–5811 (2014).
51. M. Li, F. Gao, J. R. Mascola, L. Stamatatos, V. R. Polonis, M. Koutsoukos, G. Voss, P. Goepfert, P. Gilbert, K. M. Greene, M. Bilska, D. L. Kothe, J. F. Salazar-Gonzalez, X. Wei, J. M. Decker, B. H. Hahn, D. C. Montefiori, Human immunodeficiency virus type 1 env clones from acute and early subtype B infections for standardized assessments of vaccine-elicited neutralizing antibodies. J. Virol. 79, 10108–10125 (2005).
52. C. Mueller, D. Ratner, L. Zhong, M. Esteves-Sena, G. Gao, Production and discovery of novel recombinant adeno-associated viral vectors. Curr. Protoc. Microbiol. 26, 14D.1.1–14D.1.21 (2012).
53. K. A. Buckley, P.-L. Li, A. H. Khimani, R. Hofmann-Lehmann, V. Liska, D. C. Anderson, H. M. McClure, R. M. Ruprecht, Convergent evolution of SIV env after independent inoculation of rhesus macaques with infectious proviral DNA. Virology 312, 470–480 (2003).
Acknowledgments: We would like to thank the staff at the NEPRC and WNPRC for their efforts in handling and performing the required procedures on the animals used in this study and the staff of the Quantitative Molecular Diagnostics Core of the AIDS and Cancer Virus Program, Frederick National Laboratory for Cancer Research for expert technical assistance with viral quantitation
assays. We would also like to thank H. Peng and R. Goydel for technical assistance with the Biacore X100. Funding: This project was supported by NIH grants R01 AI080324 (to M.F.), UM1 AI126623 (to M.F., defeatHIV Delaney Cure Collaboratory), P01 AI100263 (to G.G. and M.F.), and F32AI122980 (to M.R.G.) and by the Bill and Melinda Gates Foundation OPP1132169 (to M.F.). M.R.G. is a recipient of the NIH Loan Repayment Program award. J.D.L. was supported in part with federal funds from the National Cancer Institute, NIH, under contract HHSN261200800001E. Research reported in this publication was supported in part by the Office of the Director, NIH under award number P51OD011106 to the WNPRC, University of Wisconsin-Madison. This research was conducted in part at a facility constructed with support from Research Facilities Improvement Program grant numbers RR15459-01 and RR020141-01. The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services or the NIH nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. government. Author contributions: M.R.G. and M.F. conceived the study. M.R.G., C.H.F., L.M.K., and M.F. designed the experiments. M.R.G., C.H.F., L.M.K., M.E.D.-G., J.A.W., B.A., A.S.Z., N.R.P., and H.R.K. performed the experiments. L.M.K. conducted the nonhuman primate studies at NEPRC. W.A.N. and N.S.-D. conducted the nonhuman primate studies at WNPRC. E.G.R. and K.L.W. prepared reagents and processed animal samples for the nonhuman primate study at WNPRC. G.G. oversaw production of AAV1 vectors and screening for AAV1 seropositivity. J.D.L. oversaw viral load quantification experiments. M.R.G., J.D.F., and M.F. analyzed the data. M.R.G. and M.F. wrote the manuscript with input and approval from the other co-authors. Competing interests: M.R.G. and M.F. are cofounders of Emmune Inc., a company that is developing technologies for eCD4-Ig and AAV-eCD4-Ig. M.R.G. and M.F. are inventors on patent application no. 15/140,919 submitted on 28 April 2016 by The Scripps Research Institute that covers the development of AAV-delivered eCD4-Ig. M.R.G. and M.F. are inventors on patent application no. 62/645,903 submitted on 21 March 2019 by the Scripps Research Institute that covers improvements to eCD4-Ig proteins. Data and materials availability: All data associated with this study are present in the paper or the Supplementary Materials.
Submitted 20 June 2018Resubmitted 9 February 2019Accepted 31 May 2019Published 24 July 201910.1126/scitranslmed.aau5409
Citation: M. R. Gardner, C. H. Fellinger, L. M. Kattenhorn, M. E. Davis-Gardner, J. A. Weber, B. Alfant, A. S. Zhou, N. R. Prasad, H. R. Kondur, W. A. Newton, K. L. Weisgrau, E. G. Rakasz, J. D. Lifson, G. Gao, N. Schultz-Darken, M. Farzan, AAV-delivered eCD4-Ig protects rhesus macaques from high-dose SIVmac239 challenges. Sci. Transl. Med. 11, eaau5409 (2019).
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from

AAV-delivered eCD4-Ig protects rhesus macaques from high-dose SIVmac239 challenges
Jeffrey D. Lifson, Guangping Gao, Nancy Schultz-Darken and Michael FarzanAlfant, Amber S. Zhou, Neha R. Prasad, Hema R. Kondur, Wendy A. Newton, Kimberly L. Weisgrau, Eva G. Rakasz, Matthew R. Gardner, Christoph H. Fellinger, Lisa M. Kattenhorn, Meredith E. Davis-Gardner, Jesse A. Weber, Barnett
DOI: 10.1126/scitranslmed.aau5409, eaau5409.11Sci Transl Med
could protect against HIV infection. puts selective pressure on SIV. Their results show that gene therapy with CD4-based proteinsI39Nrh-eCD4-Ig
treated animals. Subsequent analysis revealed that viral loads in the treated group were lower and that−I39Nchallenges. All control animals quickly became infected, but much higher doses were needed to infect rh-eCD4-Igdelivered to nonhuman primates with an adeno-associated virus vector before repeated escalated SIVmac239
, wasI39Nability of a modified CD4 protein to protect against a stringent SIV challenge. The protein, rh-eCD4-Ig tested theet al.therapeutics have been designed based on CD4 to prevent viral binding and cell entry. Gardner
HIV and the related simian immunodeficiency virus (SIV) enter cells through the CD4 receptor, soStepping up protection against SIV
ARTICLE TOOLS http://stm.sciencemag.org/content/11/502/eaau5409
MATERIALSSUPPLEMENTARY http://stm.sciencemag.org/content/suppl/2019/07/22/11.502.eaau5409.DC1
CONTENTRELATED
http://science.sciencemag.org/content/sci/365/6453/530.fullhttp://stm.sciencemag.org/content/scitransmed/11/504/eaav5685.fullhttp://stm.sciencemag.org/content/scitransmed/11/493/eaav7325.fullhttp://stm.sciencemag.org/content/scitransmed/9/408/eaao4235.fullhttp://stm.sciencemag.org/content/scitransmed/9/402/eaaf1483.fullhttp://stm.sciencemag.org/content/scitransmed/11/501/eaaw2607.full
REFERENCES
http://stm.sciencemag.org/content/11/502/eaau5409#BIBLThis article cites 53 articles, 24 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAAS.Science Translational Medicinetitle licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. TheScience, 1200 New York Avenue NW, Washington, DC 20005. 2017 © The Authors, some rights reserved; exclusive
(ISSN 1946-6242) is published by the American Association for the Advancement ofScience Translational Medicine
at UN
IV O
F C
ALIF
OR
NIA
- SA
N D
IEG
O on A
ugust 9, 2019http://stm
.sciencemag.org/
Dow
nloaded from












![Winners list - Motor Car [AAV]excise-punjab.gov.pk/system/files/AAV...pdf · 22 Motor Car AAV 518 2,000 3,000 AAV-3520232060***-518 tariq shahzad 23 Motor Car AAV 132 2,000 3,000](https://static.fdocuments.in/doc/165x107/6035859f3caf8564033319d5/winners-list-motor-car-aavexcise-22-motor-car-aav-518-2000-3000-aav-3520232060-518.jpg)





