
Histological survey of Pacific oysters Crassostrea gigas (Thunberg) in Galicia (NW Spain)
8
Histological survey of Pacific oysters Crassostrea gigas (Thunberg) in Galicia (NW Spain) David Iglesias a,⇑ , Luis Rodríguez b , Luis Gómez c , Carlos Azevedo d,e , Jaime Montes a a Centro de Investigacións Mariñas (CIMA), Consellería do Medio Rural e do Mar, 36620 Vilanova de Arousa, Spain b Consellería do Medio Rural e do Mar, Departamento territorial de Vigo, 36201 Vigo, Pontevedra, Spain c Centro Tecnológico del Mar (CETMAR), 36208 Bouzas-Vigo, Pontevedra, Spain d Laboratório de Biologia Celular, Instituto de Ciências Biomédicas de Abel Salazar, Universidade do Porto, 4099-003 Porto, Portugal e College of Science, King Saudi University, 11451 Riyadh, Saudi Arabia article info Article history: Received 13 July 2012 Accepted 31 August 2012 Available online 15 September 2012 Keywords: Crassostrea gigas Histology Prevalence baseline levels Viral gametocytic hypertrophy Haplosporidian infection Disseminated neoplasia abstract In Galicia, there is an increasingly interest among representatives of the oyster industry in the develop- ment of Pacific oyster Crassostrea gigas culture. Nevertheless severe mortalities and emerging health problems in this species have been recently reported in European farming areas. A histological survey was performed from 2004 to 2009 to assess health status in both cultured and wild Galician oysters. Dif- ferent symbiotic organisms and conditions were detected, including viral gametocytic hypertrophy (VGH) which is reported here for first time in Spanish coast. VGH, prokaryote-like colonies and ciliates were observed in oyster tissues without causing host damage. A haplosporidian infection, copepods inducing lesions and a cellular proliferative disorder were detected in some samples causing moderate host damage; their low prevalence suggests these parasites are not a threat for C. gigas in Galicia. None of the parasites detected is OIE (Office International des Epizooties: the World Organization for Animal Health) notifiable. Although the current study did not identify any pathogens or diseases of concern, it provides important prevalence baseline data for future health and epidemiological assessments needed to better understanding the existing and emerging health problems in this species. Ó 2012 Elsevier Inc. All rights reserved. 1. Introduction Oyster industry in Galicia base their activity in a survival strat- egy involving the introduction of submarket-sized flat oysters Os- trea edulis (Linnaeus) from other countries which are on-grown for a short period (less than 1 year) and harvested before bonami- osis caused-mortality decimates the oyster batch (Montes et al., 2003). Nevertheless, the shortage in submarked-sized oyster sup- ply, as well as the high mortality caused by bonamiosis, seriously limit oyster farming profitability. Restoration of natural oyster beds (Laing et al., 2006; Beck et al., 2011) and selective breeding programs for resistance to bonamiosis (da Silva et al., 2005) seems promising strategies to the recovery of native oyster stocks, improving the profitability of oyster farming. Nevertheless, both strategies are very long-term commitments. The development of the culture of Pacific oyster Crassostrea gigas (Thunberg) has been a recurrent interest among representatives of the Galician oyster industry. C. gigas is a fast growing and hardy species native to the Japan/Korea region, relatively inexpensive and easy to produce, that has been introduced to a number of countries worldwide (Miossec et al., 2009). As a result, C. gigas has become the leading species in world shellfish culture, with an estimated production of 4.6 million tons in 2006 (FAO, 2008). First introductions of C. gigas in Galicia took place during the 1980s owing to unintentional immersions of C. gigas seed coming mixed with batches of legally imported O. edulis spat (Molares et al., 1986). In 1991 it was autho- rized by Galician Government the introduction of seeds of C. gigas in 16 rafts for their experimental suspended culture, since there was not natural seed collection or production in Galicia (Xunta de Galicia, 1992). Nowadays, there are a total number of 59 rafts, located in the southern estuaries of the coast of Galicia, which con- tinues to grown C. gigas spat imported from France, still under experimental licences, and achieved a maximum production of 579 tons in 2009 (JACUMAR, 2011). Some C. gigas naturalized pop- ulations are also present in certain Galician northern estuaries, probably as a consequence of spatfalls derived from unintentional immersions of C. gigas spat. The most important naturalized popu- lation, located in Ría de O Barqueiro, has begun to be exploited re- cently by local fishermen, achieving 97 tons of total catches in 2011 (PescadeGalicia, 2012). 0022-2011/$ - see front matter Ó 2012 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.jip.2012.08.015 ⇑ Corresponding author. Address: Centro de Investigacións Mariñas, Pedras de Corón, s/n. P.O. Box 13, 36620, Vilanova de Arousa, Pontevedra, Spain. Fax: +34 886 206372. E-mail address: [email protected] (D. Iglesias). Journal of Invertebrate Pathology 111 (2012) 244–251 Contents lists available at SciVerse ScienceDirect Journal of Invertebrate Pathology journal homepage: www.elsevier.com/locate/jip
-
Upload
david-iglesias -
Category
Documents
-
view
212 -
download
0
Transcript of Histological survey of Pacific oysters Crassostrea gigas (Thunberg) in Galicia (NW Spain)

Journal of Invertebrate Pathology 111 (2012) 244–251
Contents lists available at SciVerse ScienceDirect
Journal of Invertebrate Pathology
journal homepage: www.elsevier .com/ locate / j ip
Histological survey of Pacific oysters Crassostrea gigas (Thunberg) in Galicia(NW Spain)
David Iglesias a,⇑, Luis Rodríguez b, Luis Gómez c, Carlos Azevedo d,e, Jaime Montes a
a Centro de Investigacións Mariñas (CIMA), Consellería do Medio Rural e do Mar, 36620 Vilanova de Arousa, Spainb Consellería do Medio Rural e do Mar, Departamento territorial de Vigo, 36201 Vigo, Pontevedra, Spainc Centro Tecnológico del Mar (CETMAR), 36208 Bouzas-Vigo, Pontevedra, Spaind Laboratório de Biologia Celular, Instituto de Ciências Biomédicas de Abel Salazar, Universidade do Porto, 4099-003 Porto, Portugale College of Science, King Saudi University, 11451 Riyadh, Saudi Arabia
a r t i c l e i n f o
Article history:Received 13 July 2012Accepted 31 August 2012Available online 15 September 2012
Keywords:Crassostrea gigasHistologyPrevalence baseline levelsViral gametocytic hypertrophyHaplosporidian infectionDisseminated neoplasia
0022-2011/$ - see front matter � 2012 Elsevier Inc. Ahttp://dx.doi.org/10.1016/j.jip.2012.08.015
⇑ Corresponding author. Address: Centro de InvesCorón, s/n. P.O. Box 13, 36620, Vilanova de Arousa, Po206372.
E-mail address: [email protected] (D. I
a b s t r a c t
In Galicia, there is an increasingly interest among representatives of the oyster industry in the develop-ment of Pacific oyster Crassostrea gigas culture. Nevertheless severe mortalities and emerging healthproblems in this species have been recently reported in European farming areas. A histological surveywas performed from 2004 to 2009 to assess health status in both cultured and wild Galician oysters. Dif-ferent symbiotic organisms and conditions were detected, including viral gametocytic hypertrophy(VGH) which is reported here for first time in Spanish coast. VGH, prokaryote-like colonies and ciliateswere observed in oyster tissues without causing host damage. A haplosporidian infection, copepodsinducing lesions and a cellular proliferative disorder were detected in some samples causing moderatehost damage; their low prevalence suggests these parasites are not a threat for C. gigas in Galicia. Noneof the parasites detected is OIE (Office International des Epizooties: the World Organization for AnimalHealth) notifiable. Although the current study did not identify any pathogens or diseases of concern, itprovides important prevalence baseline data for future health and epidemiological assessments neededto better understanding the existing and emerging health problems in this species.
� 2012 Elsevier Inc. All rights reserved.
1. Introduction
Oyster industry in Galicia base their activity in a survival strat-egy involving the introduction of submarket-sized flat oysters Os-trea edulis (Linnaeus) from other countries which are on-grownfor a short period (less than 1 year) and harvested before bonami-osis caused-mortality decimates the oyster batch (Montes et al.,2003). Nevertheless, the shortage in submarked-sized oyster sup-ply, as well as the high mortality caused by bonamiosis, seriouslylimit oyster farming profitability. Restoration of natural oysterbeds (Laing et al., 2006; Beck et al., 2011) and selective breedingprograms for resistance to bonamiosis (da Silva et al., 2005) seemspromising strategies to the recovery of native oyster stocks,improving the profitability of oyster farming. Nevertheless, bothstrategies are very long-term commitments. The development ofthe culture of Pacific oyster Crassostrea gigas (Thunberg) has beena recurrent interest among representatives of the Galician oyster
ll rights reserved.
tigacións Mariñas, Pedras dentevedra, Spain. Fax: +34 886
glesias).
industry. C. gigas is a fast growing and hardy species native tothe Japan/Korea region, relatively inexpensive and easy to produce,that has been introduced to a number of countries worldwide(Miossec et al., 2009). As a result, C. gigas has become the leadingspecies in world shellfish culture, with an estimated productionof 4.6 million tons in 2006 (FAO, 2008). First introductions of C.gigas in Galicia took place during the 1980s owing to unintentionalimmersions of C. gigas seed coming mixed with batches of legallyimported O. edulis spat (Molares et al., 1986). In 1991 it was autho-rized by Galician Government the introduction of seeds of C. gigasin 16 rafts for their experimental suspended culture, since therewas not natural seed collection or production in Galicia (Xuntade Galicia, 1992). Nowadays, there are a total number of 59 rafts,located in the southern estuaries of the coast of Galicia, which con-tinues to grown C. gigas spat imported from France, still underexperimental licences, and achieved a maximum production of579 tons in 2009 (JACUMAR, 2011). Some C. gigas naturalized pop-ulations are also present in certain Galician northern estuaries,probably as a consequence of spatfalls derived from unintentionalimmersions of C. gigas spat. The most important naturalized popu-lation, located in Ría de O Barqueiro, has begun to be exploited re-cently by local fishermen, achieving 97 tons of total catches in2011 (PescadeGalicia, 2012).

D. Iglesias et al. / Journal of Invertebrate Pathology 111 (2012) 244–251 245
The development of shellfish-based industries and the concom-itant increase in demand for the introduction and transfer of differ-ent shellfish species and stocks has increased the risks of spreadingtheir parasites and diseases around the world (Bower et al., 1994).High population densities associated to aquaculture conditionsmay also accelerate the pathogenicity of infectious organismsand cause severe mortalities. Proper regulatory guidelines wereimplemented to minimize the risk of introduction of infectious dis-ease agents (ICES, 2005; OIE, 2011a). The OIE (Office Internationaldes Epizooties: the World Organization for Animal Health) list formolluscs some diseases of regulatory significance (OIE, 2011a). Pa-cific oyster is considered a susceptible species for two OIE notifi-able diseases: Perkinsus marinus and P. olseni (OIE, 2011b), butother important no notifiable OIE diseases have been describedaffecting C. gigas (Elston, 1993; Anonymous, 2010a, 2010b; Bower,2010). In summer of 2008 and 2009, severe mortality events in cul-tured Pacific oysters were reported from farming areas along theentire French coastline and some bays of Ireland and the ChannelIslands of United Kingdom (Anonymous, 2010a, 2010b). With re-gard to the etiology of these events, they have been attributed toa combination of adverse environmental factors, together withthe presence of bacteria of the genus Vibrio and Ostreid herpesvi-rus-1 (OsHV-1), including a newly described genotype of that virusnamed OsHV-1 lvar (Anonymous, 2010a, 2010b). No associationhas been found between mortalities and host-related factors suchas origin (wild or hatchery) or ploidy (diploid or triploid), thoughage was found to be a factor where the presence of high viral loadscould explain mortalities in spat and in juveniles, but not in adults(Oden et al., 2011). The presence of the virus in populations notpresenting mortalities has also been shown (Dundon et al.,2011). The European Food Safety Authority (EFSA) Panel on AnimalHealth and Welfare underlined the need of better understandingthe existing and emerging health problems in this species, basedon an assessment of the health status and epidemiological studies(Anonymous, 2010b).
From 2004 to 2009 a multidisciplinary study was performed toassess C. gigas aquaculture’s potential in Galician waters. Thisstudy included an extensive histological survey performed in cul-tured and naturalized Pacific oysters to determine the presenceof symbionts and pathological conditions, their prevalence levelsand geographic distribution among the potential culture sites. Thisinformation could provide important baseline data for future epi-demiological assessment, including comparison studies, in orderto determinate the potential importance of infectious agents on in-creased mortality in Pacific oyster.
2. Materials and methods
2.1. Samples
From 2004 to 2009 C. gigas experimental cultures were con-ducted in seven study stations distributed among four GalicianRías: Vigo, Arousa, O Barqueiro and Ribadeo (Fig. 1), as previouslydescribed (Iglesias et al., 2005). Briefly, oyster seed and juvenileoysters, originating from natural spat in Arcachon bay (France),were placed between February and May 2004, 2005 and 2008 inculture bags deployed onto iron tables in three intertidal stations(Vilaxoán, Moledos and Ribadeo) and on ropes hung from rafts infour stations (Redondela A, Grove A, Grove F and Cambados D).The cultures were terminated when market size (ca. 75 g) had beenexceeded, after a mean reared time of 14 months for seed and9 months for juvenile oysters. A baseline sample of 30 oysters fromevery batch introduced was analyzed using histology to assess thedisease status prior to deployment. Subsequent samples of 20–30individuals at each site were collected at different culture times.
In Ría de O Barqueiro oysters from a local naturalized C. gigas pop-ulation were also routinely sampled and examined by histology.
2.2. Histology
A 4–5 mm transverse section of tissue, posterior to the labialpalps and containing gill, visceral mass, and mantle lobes, was re-moved from every sampled specimen, fixed in Davidson’s solution(OIE, 2011b) and embedded in paraffin. Deparaffinized, 5-lm-thicksections were stained with Harris’ hematoxylin and eosin. Histo-logical sections were examined by light microscopy (100–1000�magnification) for the presence of symbiotic organisms inhabitingthis host and other pathological conditions. Microscopic measuresof some symbionts and conditions were done to characterize themattending to morphometric criteria.
2.3. Transmission electron microscopy
Two samples, one infected by a haplosporidian and anothershowing high intensity of inclusion bodies in gonadal tissue of C.gigas, were selected to perform ultrastructural analysis by trans-mission electron microscopy (TEM). Portions of tissue containingthese conditions were taken out with a scalpel from the paraffinblocks. Paraffin was removed by several rinses in xylene with agi-tation. Tissue was placed in 2.5% glutaraldehyde, postfixed in 2%OsO4, and embedded in Epon. Ultra-thin sections were stainedwith uranyl acetate and lead citrate and examined in a JEOL100CXII TEM transmission electron microscope at 60 kV.
2.4. Statistical and epidemiological analysis
Statistical comparisons of some morphometric values of neo-plastic-like cells vs. normal haemocytes were performed using anindependent samples t-test once verified that data was normallydistributed and variances of the two sets were homogeneous.Minitab 15 statistical software was employed to perform this anal-ysis task.
Prevalence and their 95% confidence interval of each symbiontand condition were calculated for the whole study period at eachsite and also for different oyster groups pooled as: off-bottom cul-tured (OBC), suspended cultured (SC), naturalized population (NP)and French introduced batches (FIB). Prevalence paired compari-sons of these oyster groups were performed using a critical ratio(Z) test on the difference between two independent proportions.The same statistical test was employed to compare viral gametocy-tic hypertrophy weighted prevalence by sex. EpiCalc 2000 softwareVersion 1.02 (Gilman and Myatt, 1998) was used to perform allthese calculations.
3. Results
A total of 2333 oysters were analyzed in this study: 149 corre-sponding to pathological controls prior to deployment of FIB, 1764belonging to experimental culture sites, and 420 picked from thenaturalized C. gigas population. The observation of histologicalslides revealed the presence of various symbionts and pathologicalconditions affecting C. gigas (Tables 1 and 2).
3.1. Virus
Oval to spherical basophilic inclusions, 40.1 ± 11.2 lm(Mean ± SD) of length and 29.7 ± 11.1 lm of width (n = 18), wereobserved in gonoducts (Fig. 3) of both cultured and naturalized Pa-cific oysters from all studied sites, except Cambados D (Table 2).Only one specimen corresponding to a French oyster spat analyzed

Ría de
O Bar
queir
o
RbMo
7°0'W
7°0'W
8°0'W
8°0'W
9°0'W
9°0'W
43°0' N
43°0
'N
42°0' N
42°0
'N
Ría de
Aro
usa
Ría de Vigo
Vi
CD
GFGA
RA
8°45'W
8°45'W
9°0'W
9°0'W
42°30
'N
42°3
0'N
42°15
'N
42°1
5'N
A
B
B
A
30°E
20°E
20°E
10°E
10°E
0°
0°
10°W
10°W20°W
50°N
50°N
40°N
40°N
Fig. 1. Location of the C. gigas study stations in the Galician Rías. Rb: Ribadeo (Ría de Ribadeo); Mo: Moledos (Ría de O Barqueiro); CD: Cambados D, GA: Grove A, GF: Grove F,Vi: Vilaxoán (Ría de Arousa); RA: Redondela A (Ría de Vigo).
246 D. Iglesias et al. / Journal of Invertebrate Pathology 111 (2012) 244–251
prior to their deployment exhibited few similar basophilic inclu-sions (Table 1), being extremely difficult their detection becausethe gonads were in a resting stage. Some inclusion bodies haddense staining margins (Fig. 3) that were identified by TEM asperipherally displaced chromatin (Fig. 4). TEM examination alsorevealed the presence of unenveloped, icosahedral viral particles(38.9 ± 2.7 nm in diameter, n = 23) within the inclusions (Fig. 4),
permitting us to diagnose this condition as a viral gametocytichypertrophy (VGH).
VGH site prevalence ranged from 0% to 1.88% (Tables 1 and 2).Overall prevalences were not statistically different between oystergroups (Fig. 2). In VGH infected samples the prevalence varied be-tween 3.3% and 10.3%. VGH infections were detected in spat andadults, mainly in males (weighted prevalence by sex of 10.13%)
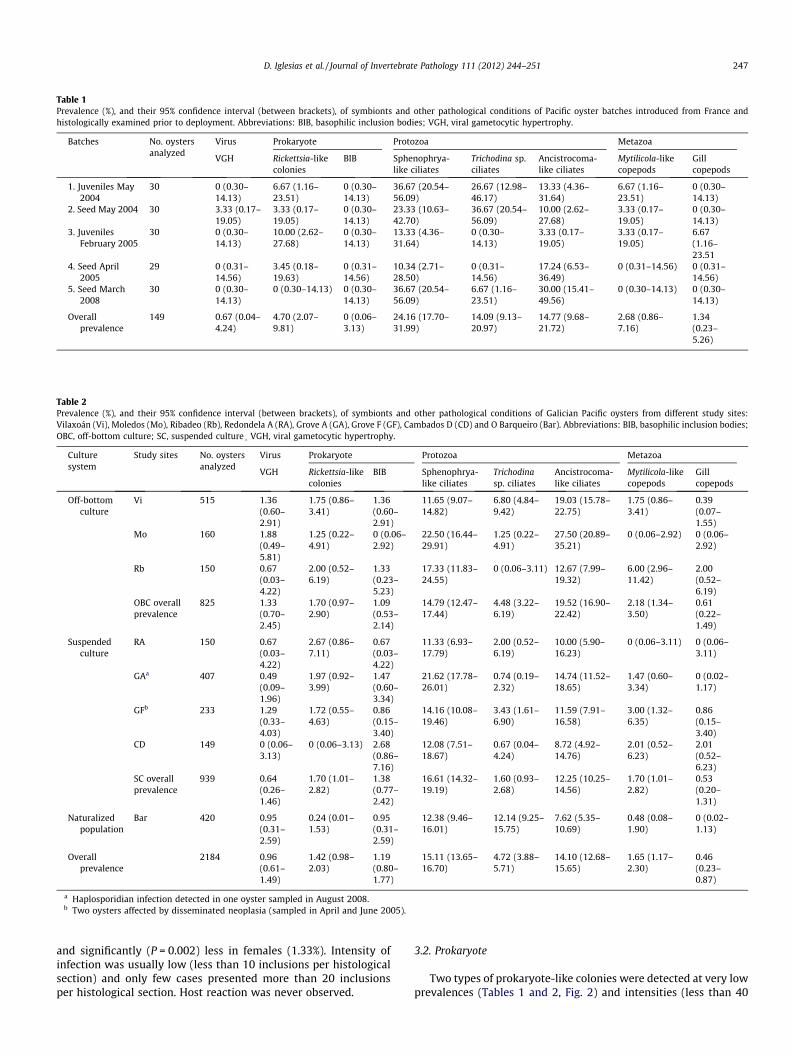
Table 1Prevalence (%), and their 95% confidence interval (between brackets), of symbionts and other pathological conditions of Pacific oyster batches introduced from France andhistologically examined prior to deployment. Abbreviations: BIB, basophilic inclusion bodies; VGH, viral gametocytic hypertrophy.
Batches No. oystersanalyzed
Virus Prokaryote Protozoa Metazoa
VGH Rickettsia-likecolonies
BIB Sphenophrya-like ciliates
Trichodina sp.ciliates
Ancistrocoma-like ciliates
Mytilicola-likecopepods
Gillcopepods
1. Juveniles May2004
30 0 (0.30–14.13)
6.67 (1.16–23.51)
0 (0.30–14.13)
36.67 (20.54–56.09)
26.67 (12.98–46.17)
13.33 (4.36–31.64)
6.67 (1.16–23.51)
0 (0.30–14.13)
2. Seed May 2004 30 3.33 (0.17–19.05)
3.33 (0.17–19.05)
0 (0.30–14.13)
23.33 (10.63–42.70)
36.67 (20.54–56.09)
10.00 (2.62–27.68)
3.33 (0.17–19.05)
0 (0.30–14.13)
3. JuvenilesFebruary 2005
30 0 (0.30–14.13)
10.00 (2.62–27.68)
0 (0.30–14.13)
13.33 (4.36–31.64)
0 (0.30–14.13)
3.33 (0.17–19.05)
3.33 (0.17–19.05)
6.67(1.16–23.51
4. Seed April2005
29 0 (0.31–14.56)
3.45 (0.18–19.63)
0 (0.31–14.56)
10.34 (2.71–28.50)
0 (0.31–14.56)
17.24 (6.53–36.49)
0 (0.31–14.56) 0 (0.31–14.56)
5. Seed March2008
30 0 (0.30–14.13)
0 (0.30–14.13) 0 (0.30–14.13)
36.67 (20.54–56.09)
6.67 (1.16–23.51)
30.00 (15.41–49.56)
0 (0.30–14.13) 0 (0.30–14.13)
Overallprevalence
149 0.67 (0.04–4.24)
4.70 (2.07–9.81)
0 (0.06–3.13)
24.16 (17.70–31.99)
14.09 (9.13–20.97)
14.77 (9.68–21.72)
2.68 (0.86–7.16)
1.34(0.23–5.26)
Table 2Prevalence (%), and their 95% confidence interval (between brackets), of symbionts and other pathological conditions of Galician Pacific oysters from different study sites:Vilaxoán (Vi), Moledos (Mo), Ribadeo (Rb), Redondela A (RA), Grove A (GA), Grove F (GF), Cambados D (CD) and O Barqueiro (Bar). Abbreviations: BIB, basophilic inclusion bodies;OBC, off-bottom culture; SC, suspended culture� VGH, viral gametocytic hypertrophy.
Culturesystem
Study sites No. oystersanalyzed
Virus Prokaryote Protozoa Metazoa
VGH Rickettsia-likecolonies
BIB Sphenophrya-like ciliates
Trichodinasp. ciliates
Ancistrocoma-like ciliates
Mytilicola-likecopepods
Gillcopepods
Off-bottomculture
Vi 515 1.36(0.60–2.91)
1.75 (0.86–3.41)
1.36(0.60–2.91)
11.65 (9.07–14.82)
6.80 (4.84–9.42)
19.03 (15.78–22.75)
1.75 (0.86–3.41)
0.39(0.07–1.55)
Mo 160 1.88(0.49–5.81)
1.25 (0.22–4.91)
0 (0.06–2.92)
22.50 (16.44–29.91)
1.25 (0.22–4.91)
27.50 (20.89–35.21)
0 (0.06–2.92) 0 (0.06–2.92)
Rb 150 0.67(0.03–4.22)
2.00 (0.52–6.19)
1.33(0.23–5.23)
17.33 (11.83–24.55)
0 (0.06–3.11) 12.67 (7.99–19.32)
6.00 (2.96–11.42)
2.00(0.52–6.19)
OBC overallprevalence
825 1.33(0.70–2.45)
1.70 (0.97–2.90)
1.09(0.53–2.14)
14.79 (12.47–17.44)
4.48 (3.22–6.19)
19.52 (16.90–22.42)
2.18 (1.34–3.50)
0.61(0.22–1.49)
Suspendedculture
RA 150 0.67(0.03–4.22)
2.67 (0.86–7.11)
0.67(0.03–4.22)
11.33 (6.93–17.79)
2.00 (0.52–6.19)
10.00 (5.90–16.23)
0 (0.06–3.11) 0 (0.06–3.11)
GAa 407 0.49(0.09–1.96)
1.97 (0.92–3.99)
1.47(0.60–3.34)
21.62 (17.78–26.01)
0.74 (0.19–2.32)
14.74 (11.52–18.65)
1.47 (0.60–3.34)
0 (0.02–1.17)
GFb 233 1.29(0.33–4.03)
1.72 (0.55–4.63)
0.86(0.15–3.40)
14.16 (10.08–19.46)
3.43 (1.61–6.90)
11.59 (7.91–16.58)
3.00 (1.32–6.35)
0.86(0.15–3.40)
CD 149 0 (0.06–3.13)
0 (0.06–3.13) 2.68(0.86–7.16)
12.08 (7.51–18.67)
0.67 (0.04–4.24)
8.72 (4.92–14.76)
2.01 (0.52–6.23)
2.01(0.52–6.23)
SC overallprevalence
939 0.64(0.26–1.46)
1.70 (1.01–2.82)
1.38(0.77–2.42)
16.61 (14.32–19.19)
1.60 (0.93–2.68)
12.25 (10.25–14.56)
1.70 (1.01–2.82)
0.53(0.20–1.31)
Naturalizedpopulation
Bar 420 0.95(0.31–2.59)
0.24 (0.01–1.53)
0.95(0.31–2.59)
12.38 (9.46–16.01)
12.14 (9.25–15.75)
7.62 (5.35–10.69)
0.48 (0.08–1.90)
0 (0.02–1.13)
Overallprevalence
2184 0.96(0.61–1.49)
1.42 (0.98–2.03)
1.19(0.80–1.77)
15.11 (13.65–16.70)
4.72 (3.88–5.71)
14.10 (12.68–15.65)
1.65 (1.17–2.30)
0.46(0.23–0.87)
a Haplosporidian infection detected in one oyster sampled in August 2008.b Two oysters affected by disseminated neoplasia (sampled in April and June 2005).
D. Iglesias et al. / Journal of Invertebrate Pathology 111 (2012) 244–251 247
and significantly (P = 0.002) less in females (1.33%). Intensity ofinfection was usually low (less than 10 inclusions per histologicalsection) and only few cases presented more than 20 inclusionsper histological section. Host reaction was never observed.
3.2. Prokaryote
Two types of prokaryote-like colonies were detected at very lowprevalences (Tables 1 and 2, Fig. 2) and intensities (less than 40

c
Fig. 4. VGH, viral particles (asterisk) detected by transmission electron microscopyinside basophilic inclusions. Arrowhead: edge of inclusion; C: peripheral condensedchromatin. Scale bar: 250 nm.
Fig. 2. Overall prevalences (and 95% confidence interval superior limit) of eachsymbiont/pathological condition for oysters C. gigas pooled as off-bottom cultured(OBC), suspended cultured (SC), naturalized population (NP) and French introducedbatches (FIB). Results of paired comparisons represented using letters (alternatinglowercase and capital letters to improve clarity): different letters for each symbiontindicate significant (P < 0.05) differences. Symbiont abbreviations: VGH, viralgametocytic hypertrophy; R-LC, Rickettsia-like colonies; BIB, basophilic inclusionbodies; S-LC, Sphenophrya-like ciliates; TC, Trichodina sp. Ciliates; A-LC, Ancistroco-ma-like ciliates; M-LC, Mytilicola-like copepods; GC, gill copepods.
248 D. Iglesias et al. / Journal of Invertebrate Pathology 111 (2012) 244–251
colonies per histological section) in sampled oysters from almostevery site studied. Rounded intracellular basophilic bodies,11.4 ± 3.0 lm of length (n = 14), similar to Rickettsia-like colonies,were detected in digestive tubules. Larger (17.2 ± 3.1 lm of lengthand 11.1 ± 2.0 lm of width, n = 18) basophilic inclusion bodies(BIB) of round-elongate shape and surrounded by a halo, were ob-served in epithelial cells of the intestine wall. Associated haemocy-tic reaction was not observed.
3.3. Protozoa
Three types of ciliates were found in the sampled oysters withlow to moderate prevalences (Tables 1 and 2, Fig. 2). Sphe-nophrya-like ciliates, pear-shaped 14 to 42 lm long, were seen at-tached to, or located near, gills and/or labial palps. Intensity waslow and no lesions were observed. Ciliates of the genus Trichodina(43.40 ± 5.51 lm of length, n = 19) were observed free in the palialcavity close to mantle, labial palps or gill surface. Most infections
Fig. 3. Viral gametocytic hypertrophy (VGH). Light micrograph showing basophilicinclusions (stars) among germ cells of male gonad; some of them with chromophilicmargins (arrowheads). Scale bar: 40 lm. H&E stained.
were innocuous with a low intensity but, occasionally, an inflam-matory response of the gill to these ciliates – characterized by des-quamated epithelial cells, haemocytes and tissue debris – wasobserved. A third ciliate type, similar to Ancistrocoma-like ciliates,41–54 lm long, was found in the lumen of the digestive gland tu-bules of oysters from every sampled site. Intensity of infection waslow and pathological effect was negligible.
An advanced haplosporidian infection was detected in one oys-ter cultivated in Grove A and sampled in August 2008. The proto-zoan infection was accompanied by a diffused hemocyticinfiltration into the areas of parasitic infection. Multinucleatedplasmodial stages (2–8 nuclei), 8.2 ± 1.5 lm of length and6.8 ± 1.5 lm of width (n = 18), were seen within the digestivegland and gill connective tissues (Fig. 5). Numerous sporogonic
dte
vct
Fig. 5. Histological section showing haplosporidian plasmodia (arrows) withinvesicular connective tissue (vct) located surrounding digestive tubules. dte,digestive tubule epithelium. Scale bar: 10 lm. H&E stained.

dte
dtldtl
Fig. 6. Haplosporidian spores (arrowheads) within digestive tubule epithelium(dte). dtl, digestive tubule lumen. Scale bar: 10 lm. H&E stained.
n
n n
h
hh
Fig. 8. Presumptive disseminated neoplasia in Crassostrea gigas. Small foci ofabnormal cells (n) in connective tissue surrounding the gut. Arrowheads: mitoticfigures; h, hemocytes. Scale bar: 10 lm. H&E stained.
D. Iglesias et al. / Journal of Invertebrate Pathology 111 (2012) 244–251 249
stages and spores in various stages of development were observedexclusively within the digestive tubules (Fig. 6). Sporocysts,26.0 ± 8.4 lm of length and 16.5 ± 3.4 lm of width (n = 5), contain-ing spores were located within the digestive tubule epithelium.Development of the sporocyst resulted in lysis of digestive epithe-lial cells. Mature, but mainly immature spores, 5.4 ± 0.5 lm ofmaximum diameter (n = 25), were found free within the luminaof affected tubules. Few mature spores could be observed byTEM in the deparaffinized tissues processed for electron micro-scopic examination. These spores presented ellipsoidal morphol-
op
wa
nu
Fig. 7. Electron micrograph of a haplosporidian mature spore showing a layered,striated wall (wa), a hinged operculum (op) and remnants of a nucleus (nu). Scalebar: 1 lm.
ogy with a layered wall and a hinged operculum (Fig. 7).Remnants of the nucleus and haplosporosome formative regionscould be intuited inside the spores. Nevertheless, no spore orna-mentation could be discerned in these samples.
3.4. Metazoa
Unidentified copepods were observed at very low prevalencesin gill and palial cavity of oysters from some study sites (Tables1 and 2). Gill tissue surrounding the copepods exhibited an intenseinflammatory reaction. Mytilicola-like copepods were sometimesdetected in intestinal lumen attached by hooked appendages tothe gut wall and causing focal epithelium disruption.
3.5. Other conditions
Two oysters with a cellular proliferative disorder were found inGrove F (Ría de Arousa) during the study. Focal infiltrations ofabnormal cells were seen in connective tissue surrounding thegut. These transformed cells resembled hypertrophied hemocytesand presented enlarged hyperchromatic rounded nuclei(3.87 ± 0.63 lm vs. 2.67 ± 0.32 lm of length in normal hemocytes,n = 24), sometimes with prominent nucleoli (Fig. 8). As a result,they had a significantly (P = 0.000) higher nucleus to cytoplasm ra-tio (0.70 ± 0.08) than normal hemocytes (0.51 ± 0.09). Mitotic fig-ures were also commonly observed in these foci (Fig. 8).Intensity was low (only very confined small foci of cells). This con-dition was diagnosed as a presumptive disseminated neoplasia.
4. Discussion
This paper presents the first comprehensive histological surveyof C. gigas in Galician estuaries (NW Spain). The study revealed theoccurrence of various symbionts and conditions affecting this spe-cies and established their baseline prevalence levels.
The condition known as VGH or ovacystis denotes massivehypertrophy of gametocytes caused by an associated papilloma-or polyoma-like virus referred to in previous literature as papova-virus (Farley, 1985). This condition was first described in ova ofCrassostrea virginica (Gmelin) from USA (Farley, 1976) and, morerecently, in C. gigas from Korea (Choi et al., 2004), France (Garciaet al., 2006), Germany (Watermann et al., 2008), Ireland (Cheslettet al., 2009) and Alaska, USA (Meyers et al., 2009). According to

250 D. Iglesias et al. / Journal of Invertebrate Pathology 111 (2012) 244–251
our ultrastructural observations, inclusions causing gametocytehypertrophy, previously detected by histology, were associatedwith the presence of viral particles. Although the reduced qualityof the reprocessed TEM material, on the basis of their location, sizeand symmetry, the unenveloped virions observed are in concor-dance with the previous descriptions given by the latter authors.This is the first report of viral gametocytic hypertrophy affectingC. gigas in Spain.
VGH prevalence detected in Galician oysters was similar tothose previously reported for this species. VGH affected predomi-nantly to males in Galicia. In common with previous studies, inten-sity of VGH infection was low and host hemocytic reaction wasnever observed, suggesting limited impact in oyster health.
Rounded intracellular basophilic bodies observed in digestivetubules strongly resembled Rickettsia-like colonies, reported fromdifferent tissues of a variety of bivalve molluscan species withoutobvious significant harm to the host (Bower et al., 1994; Paillardet al., 2004). Larger BIB observed in epithelial cells of the intestinewall were similar to basophilic cytoplasmic inclusion bodies(Meyers, 1981) and prokaryotic inclusion bodies (Gauthier et al.,1990) reported affecting C. virginica from USA. Both prokaryote-like organisms reported in the present study had low levels ofintensity and prevalence, and signs of host response were neverseen, suggesting that they do not constitute a pathologicalproblem.
Ciliates are common inhabitants of bivalve molluscs and theyare considered to be harmless commensals feeding on bacteriarather than parasites (Lauckner, 1983; Bower et al., 1994). How-ever, some controversy surrounds the pathogenic role of Trichodinaciliates. Mortalities attributed to Trichodina sp. have been reportedin cockles and oysters (Lauckner, 1983). Boussaïd et al. (1999)found that C. gigas heavily infected with Trichodina sp. exhibitedan inflammatory response and alterations of the gill epithelium.Similar observations were verified in the present study in someindividuals infected by Trichodina sp., however most ciliate infec-tions were innocuous to the host.
The haplosporidian parasite detected in one Pacific oyster fromGrove A resembles Haplosporidium nelsoni in tissue specificity,morphology, and size of plasmodial and sporogonic stages accord-ing to Bower (2010) and OIE (2011b) reviews. Electron microscopyobservations revealed spores with typical characters of the phylumHaplosporidia because they have walled spores with an orificeexternally covered by a hinged lid (Perkins, 1990). Among thehaplosporidian genera, Haplosporidium and Minchinia have a simi-lar internal organization and the only taxonomic character used todistinguish between these two genera is the ultrastructural organi-zation of the ornaments (Azevedo et al., 2006). It was not observedspore ornamentation in our samples (probably due to the harshtreatment of the tissues during histological processing) and, unfor-tunately, we could not ascertain their taxonomic position. The verylow prevalence of haplosporidian infections detected in the pres-ent study (0.05%) suggests these parasites are not a threat for C.gigas reared in Galicia.
Several species of parasitic copepods have been reported at-tached to gills and intestine wall of oysters and other bivalves(Lauckner, 1983), but the vast majority of species are innocuous(Bower et al., 1994). Limited histological lesions were detected inthe present study caused by parasitic copepods. Nevertheless,overall prevalences were very low and copepods do not seem aparasite of concern.
The cellular proliferative disorder detected in two oysters fromGrove F (Ría de Arousa), was diagnosed as a presumptive low-leveldisseminated neoplasia stage because it consist in the proliferationof circulating cells with relatively large nuclei, a high nucleus-to-cytoplasm ratio, and a relatively high frequency of mitotic figures.A similar abnormal condition, diagnosed as an early stage of dis-
seminated neoplasia, was reported by Villalba (2006) in C. gigascollected from a culture raft located in the same estuary. As faras we know, reports of disseminated neoplasia are practically ab-sent for C. gigas in spite of its now global geographic distributionand the very large numbers of them histologically examinedworldwide (Peters, 1988; Elston et al., 1992; Barber, 2004). To date,one case has been reported in C. gigas from Matsushima Bay, Japan(Farley, 1969), but it has been noted the diagnosis was somewhatequivocal and should not be accepted as a definitive disseminatedneoplasia in C. gigas (Elston et al., 1992). In any case, the low levelintensity and prevalence (two cases in 2184 oysters analyzed) ofthis lesion suggest that is not a threat for the oyster culture inGalicia.
There were not quantitative (in terms of overall prevalence) dif-ferences between oyster groups for the majority of the symbiontsand conditions detected. Overall prevalence higher than 10% wasrecorded only for Sphenophrya and Ancistrocoma-like ciliates,which could be considered the principal species of the symbiontcommunity associated with Pacific oysters. The remaining symbi-onts detected showed overall prevalence lower than 5% and couldbe considered as rare.
In conclusion, none of the parasites detected is OIE notifiable(OIE, 2011a). Despite the variety of histological findings, they donot appear to be a problem for oyster culture in Galicia, eitherdue to low prevalence/intensity levels, or low pathological effects.Moreover, abnormal mortalities (defined as sudden mortalityaffecting more than 15% of a stock within a maximum interval of15 days (Nicolas et al., 2007)) were never observed in the experi-mental batches reared in this study, and cumulative mortalitiesat the end of the culture never exceeded 30% in the worst case(unpublished data). Although the current study did not identifyany pathogens or diseases of concern, it provides important prev-alence baseline data for future health assessments and epidemio-logical studies, needed to better understanding the existing andemerging health problems in this species.
Acknowledgments
We are grateful to Lupe González, María Isabel Meléndez, ElenaPenas, Nieves Rúa, Leticia Romanos, Sonia Balsa, Raquel Alves,Susana Goldar, José A. Fernández, Elena Momboisse, Sara Trabada,Ramón J. Menéndez, Begoña Serra, Guillermo del Río, Ana Alcalde,Aitor Freire and Diego González for technical assistance; to Ansel-mo Bugallo, César García, Jacinto Piñeiro, Andrés Calvo and JoséCarlos Cerqueiro, for performing suspended oyster culture works;and to Javier César Aldariz for producing the map illustrated hereas Fig. 1. We thank Dr. Susana Darriba and Dr. Carmen López forhis critical review of the manuscript. The present study was fundedby Dirección Xeral de Ordenación e Xestión dos Recursos Mariños,Consellería do Mar, Xunta de Galicia.
References
Anonymous, 2010a. COMMISSION REGULATION (EU) No. 175/2010 of 2 March 2010Implementing Council Directive 2006/88/EC as Regards Measures to ControlIncreased Mortality in Oysters of the Species Crassostrea gigas in Connectionwith the Detection of Ostreid Herpesvirus 1 lvar (OsHV-1 lvar).
Anonymous, 2010b. EFSA scientific Opinion on the increased mortality events inPacific oyster (Crassostrea gigas). EFSA J. 8(11), 1894, 1–59.
Azevedo, C., Balseiro, P., Casal, G., Gestal, C., Aranguren, R., Stokes, N.A., Carnegie,R.B., Novoa, B., Burreson, E.M., Figueras, A., 2006. Ultrastructural and molecularcharacterization of Haplosporidium montforti n. sp., parasite of the Europeanabalone Haliotis tuberculata. J. Invertebr. Pathol. 92, 23–32.
Barber, B.J., 2004. Neoplastic diseases of commercially important marine bivalves.Aquat. Living Resour. 17, 449–466.
Beck, M.W., Brumbaugh, R.D., Airoldi, L., Carranza, A., Coen, L.D., Crawford, C., Defeo,O., Edgar, G.J., Hancock, B., Kay, M.C., Lenihan, H.S., Luckenbach, M.W., Toropova,C.L., Zhang, G., Guo, X., 2011. Oyster reefs at risk and recommendations forconservation, restoration, and management. BioScience 61, 107–116.

D. Iglesias et al. / Journal of Invertebrate Pathology 111 (2012) 244–251 251
Boussaïd, B., Grippari, J.L., Renault, T., Tige§, G., Dorange, G., 1999. Trichodina sp.infestation of Crassostrea gigas oyster gills in brittany. Fr. J. Invertebr. Pathol. 73,339–342.
Bower, S.M., McGladdery, S.E., Price, I.M., 1994. Synopsis of infectious diseases andparasites of commercially exploited shellfish. Annu. Rev. Fish Dis. 4, 1–199.
Bower, S.M., 2010. Synopsis of Infectious Diseases and Parasites of CommerciallyExploited Shellfish. <http://www.pac.dfo-mpo.gc.ca/science/species-especes/shellfish-coquillages/diseases-maladies/index-eng.htm>.
Cheslett, D., McKiernan, F., Hickey, C., Collins, E., 2009. Viral gametocytichypertrophy of the Pacific oyster Crassostrea gigas in Ireland. Dis. Aquat. Org.83, 181–185.
Choi, D.L., Lee, N.-S., Choi, H.J., Park, M.-A., McGladdery, S., Park, M.S., 2004. Viralgametocytic hypertrophy caused by a papova-like virus infection in Pacificoyster Crassostrea gigas in Korea. Dis. Aquat. Org. 59, 205–209.
da Silva, P.M., Fuentes, J., Villalba, A., 2005. Growth, mortality and diseasesusceptibility of oyster Ostrea edulis families obtained from brood stocks ofdifferent geographical origins, through on-growing in the Ría de Arousa (Galicia,NW Spain). Mar. Biol. 147, 965–977.
Dundon, W.G., Arzul, I., Omnes, E., Robert, M., Magnabosco, C., Zambon, M., Gennari,L., Toffan, A., Terregino, C., Capua, I., Arcangeli, G., 2011. Detection of type 1Ostreid Herpes variant (OsHV-1 lvar) with no associated mortality in French-origin Pacific cupped oyster Crassostrea gigas farmed in Italy. Aquaculture 314,49–52.
Elston, R.A., Moore, J.D., Brooks, K., 1992. Disseminated neoplasia of bivalvemolluscs. Rev. Aquat. Sci. 6, 405–466.
Elston, R.A., 1993. Infectious diseases of the Pacific oyster, Crassostrea gigas. Annu.Rev. Fish Dis., 259–276.
FAO, 2008. Aquaculture Production: Quantities 1950–2006. Fishstat Plus. <http://www.fao.org/fi/statist/fisoft/FISHPLUS>.
Farley, C.A., 1969. Probable neoplastic disease of the hematopoietic system inoysters, Crassostrea virginica and Crassostrea gigas. Nat. Cancer Inst. Monogr. 31,541–555.
Farley, C.A., 1976. Ultrastructural observations on epizootic neoplasia and lytic virusinfection in bivalve molluscs. Prog. Exp. Tumor Res. 20, 283–294.
Farley, C.A., 1985. Viral gametocytic hypertrophy in oysters. In: Sindermann, C.J.(Ed.), Identification Leaflets for Diseases and Parasites of Fish and Shellfish No.25. ICES, Copenhagen, pp. 1–5.
Garcia, C., Robert, M., Arzul, I., Chollet, B., Joly, J.-P., Miossec, L., Comtet, T., Berthe, F.,2006. Viral gametocytic hypertrophy of Crassostrea gigas in France. fromoccasional records to disease emergence? Dis. Aquat. Org. 70, 193–199.
Gauthier, J.D., Soniat, T.M., Rogers, J.S., 1990. A parasitological survey of oystersalong salinity gradients in coastal Louisiana. J. World Aquacult. Soc. 21, 105–115.
Gilman, J., Myatt, M., 1998. EpiCalc 2000. Version 1.02. Brixton Books. <http://www.brixtonhealth.com/epicalc.html>.
ICES, 2005. Code of Practice on the Introductions and Transfers of MarineOrganisms. ICES, Copenhagen.
Iglesias, D., Rodríguez, L., Montes, J., Conchas, R.F., Pérez, J.L., Fernández, M., Guerra,A., 2005. Estudio de viabilidad del cultivo de ostra rizada Crassostrea gigas(Thunberg, 1793) en diferentes rías gallegas. Primeros resultados biológico-productivos. Bol. Inst. Esp. Oceanogr. 21, 293–309.
JACUMAR, 2011. Official Aquaculture Production from Junta Asesora de CultivosMarinos. Ministerio de Agricultura, Alimentación y Medio Ambiente, Spanish
government. <http://www.magrama.gob.es/app/jacumar/datos_produccion/lista_datos_produccion2.aspx?Id=es> (accessed 31.05.2012.).
Laing, I., Walker, P., Areal, F., 2006. Return of the native – is European oyster (Ostreaedulis) stock restoration in the UK feasible? Aquat. Living Resour. 19, 283–287.
Lauckner, G., 1983. Diseases of mollusca: bivalvia. In: Kinne, O. (Ed.), Diseases ofMarine Animals, Bivalvia to Scaphopoda, vol. II. Biologische Anstalt Helgoland,Hamburg, pp. 477–961.
Meyers, T.R., Burton, T., Evans, W., Starkey, N., 2009. Detection of viruses and virus-like particles in four species of wild and farmed bivalve molluscs in Alaska, USA,from 1987 to 2009. Dis. Aquat. Org. 88, 1–12.
Meyers, T.R., 1981. Endemic diseases of cultured shellfish of Long Island, New York:adult and juvenile American oysters (Crassostrea virginica) and hard clams(Mercenaria mercenaria). Aquaculture 22, 305–330.
Miossec, L., Le Deuff, R.M., Goulletquer, P., 2009. Alien species alert: Crassostreagigas (Pacific oyster). ICES Coop. Res. Rep. 299, pp. 42.
Molares, J., Pascual, C., Quintana, R., 1986. Análisis de los factores determinantes dela producción de la ostra – Crassostrea gigas – cultivada en las rías gallegas.Alimentaria 172, 27–33.
Montes, J., Ferro-Soto, B., Conchas, R.F., Guerra, A., 2003. Determining culturestrategies in populations of the European flat oyster, Ostrea edulis, affected bybonamiosis. Aquaculture 220, 175–182.
Nicolas, J.-L., Renault, T., Gagnaire, B., Garcia, C.M., Garnier, M., Gay, Y., Le Labreuche,F., Roux, L., Miossec, J.F., Saulnier Pepin, D., 2007. Oyster summer mortality risksassociated with pathogens. In: Samain, J.-F., McCombie, H. (Eds.), Mortality ofPacific Oyster Crassostrea Gigas. The Morest Project. Éditions Quae, Versailles,pp. 243–287.
Oden, E., Martenot, C., Berthaux, M., Travaillé, E., Malas, J.-P., Houssin, M., 2011.Quantification of ostreid herpesvirus 1 (OsHV-1) in Crassostrea gigas by real-time PCR: determination of a viral load threshold to prevent summermortalities. Aquaculture 317, 27–31.
OIE, 2011a. Aquatic Animal Health Code. <http://www.oie.int/en/international-standard-setting/aquatic-code/access-online/>.
OIE, 2011b. Manual of Diagnostic Tests for Aquatic Animals. <http://www.oie.int/en/international-standard-setting/aquatic-manual/access-online/>.
Paillard, C., Le Roux, F., Borrego, J.J., 2004. Bacterial disease in marine bivalves, areview of recent studies: trends and evolution. Aquat. Living Resour. 17, 477–498.
Perkins, F.O., 1990. Phylum haplosporidia. In: Margulis, L., Corliss, J.O., Melkonian,M., Chapman, D.J. (Eds.), Handbook of Protoctista. Jones and Bartlett Publishers,Boston, pp. 19–29.
PescadeGalicia, 2012. Fish Landing Statistics of Galician Government. <http://www.pescadegalicia.com/default.htm> (accessed 31.05.2012.).
Peters, E.C., 1988. Recent investigations on the disseminated sarcomas of marinebivalve molluscs. Spec. Publ. Am. Fish. Soc. 18, 74–92.
Villalba, A., 2006. Different types of neoplasia in Crassostrea gigas, Venus verrucosaand Mytilus galloprovincialis from the Spanish coast. J. Shellfish Res. 25, 786–787.
Watermann, B.T., Herlyn, M., Daehne, B., Bergmann, S., Meemken, M., Kolodzey, H.,2008. Pathology and mass mortality of Pacific oysters, Crassostrea gigas(Thunberg), in 2005 at the East Frisian coast. Ger. J. Fish Dis. 31, 621–630.
Xunta de Galicia, 1992. Plan de ordenación dos recursos pesqueiros e marisqueirosde Galicia, pp. 874.


















