
High expression of TNF alpha is associated with −308 and −238 TNF alpha polymorphisms in knee...
7
ORIGINAL ARTICLE High expression of TNF alpha is associated with 2308 and 2238 TNF alpha polymorphisms in knee osteoarthritis Jose ´ Francisco Mun ˜ oz-Valle • Edith Orego ´n-Romero • He ´ctor Rangel-Villalobos • Gloria Esther Martı ´nez-Bonilla • Eduardo Castan ˜ eda-Saucedo • Lorenzo Salgado-Goytia • Marco Antonio Leyva-Va ´zquez • Berenice Illades-Aguiar • Luz del Carmen Alarco ´n-Romero • Mo ´nica Espinoza-Rojo • Isela Parra-Rojas Received: 4 April 2012 / Accepted: 13 October 2012 / Published online: 30 October 2012 Ó Springer-Verlag Italia 2012 Abstract Knee osteoarthritis (OA) is a common chronic degenerative disease characterized by the loss of articular cartilage components due to an imbalance between extra- cellular matrix destruction and repair. The proinflammatory cytokines involved in OA, TNFa and IL1b, are considered the major implicated. The aim of this study was to inves- tigate the relationship between TNFa -308 and -238 polymorphisms with messenger RNA (mRNA) and soluble TNFa expression in knee OA patients and healthy subjects (HS). Case–control study involved 50 knee OA patients classified according to 1986 ACR Classification Criteria, as well as 100 HS. The Western Ontario and McMaster Universities Osteoarthritis Index and Lequesne disability index were applied to OA patients. The -308 and -238 polymorphisms were determined by polymerase chain reaction–restriction fragment length polymorphism tech- nique. The TNFa mRNA expression was quantified by real-time PCR using TaqMan method. The sTNFa levels were measured by enzyme-linked immunosorbent assay. The TNFa mRNA expression in knee OA patients was higher than in HS (1.56-fold). In addition, the TNFa mRNA expression was higher in carriers of G allele in the knee OA group for both polymorphisms. The sTNFa levels were increased in G/G versus G/A genotypes in both studied polymorphisms (p \ 0.05). However, the TNFa -308 and -238 genotypes did not show statistical differ- ences between groups. The G allele of TNFa -308 and -238 polymorphisms is associated with high mRNA and soluble expression in knee OA patients. However, it is not a marker of susceptibility in Western Mexico. Further studies are necessary to confirm these findings. Keywords Knee osteoarthritis TNF alpha expression TNF alpha levels TNF alpha polymorphisms Introduction Knee osteoarthritis (OA) is a common chronic degenera- tive disease characterized by the loss of articular cartilage components due to an imbalance between extracellular matrix destruction and repair [1]. Although articular car- tilage breakdown is a major characteristic of OA, other joint tissues such as the synovial membrane and subchon- dral bone actively participate in the progression of the disease [2]. Jose ´ Francisco Mun ˜oz-Valle and Edith Orego ´n-Romero contributed equally to this work. J. F. Mun ˜oz-Valle (&) E. Orego ´n-Romero Grupo de Inmunogene ´tica Funcional, Departamento de Biologı ´a Molecular y Geno ´mica, Centro Universitario de Ciencias de la Salud, Universidad de Guadalajara, Insurgentes 244-1, Colonia Lomas de Atemajac, C.P. 45178 Zapopan, Guadalajara, Jalisco, Mexico e-mail: [email protected] H. Rangel-Villalobos Instituto de Investigacio ´n en Gene ´tica Molecular, Centro Universitario de la Cie ´nega, Universidad de Guadalajara, Ocotla ´n, Jalisco, Mexico G. E. Martı ´nez-Bonilla Servicio de Reumatologı ´a, OPD Hospital Civil de Guadalajara ‘‘Fray Antonio Alcalde’’, Guadalajara, Jalisco, Mexico E. Castan ˜ eda-Saucedo L. Salgado-Goytia M. A. Leyva-Va ´zquez B. Illades-Aguiar L. d. C. Alarco ´n-Romero M. Espinoza-Rojo I. Parra-Rojas Unidad Acade ´mica de Ciencias Quı ´mico Biolo ´gicas, Universidad Auto ´noma de Guerrero, Chilpancingo, Guerrero, Mexico 123 Clin Exp Med (2014) 14:61–67 DOI 10.1007/s10238-012-0216-3
Transcript of High expression of TNF alpha is associated with −308 and −238 TNF alpha polymorphisms in knee...

ORIGINAL ARTICLE
High expression of TNF alpha is associated with 2308 and 2238TNF alpha polymorphisms in knee osteoarthritis
Jose Francisco Munoz-Valle • Edith Oregon-Romero • Hector Rangel-Villalobos •
Gloria Esther Martınez-Bonilla • Eduardo Castaneda-Saucedo • Lorenzo Salgado-Goytia •
Marco Antonio Leyva-Vazquez • Berenice Illades-Aguiar • Luz del Carmen Alarcon-Romero •
Monica Espinoza-Rojo • Isela Parra-Rojas
Received: 4 April 2012 / Accepted: 13 October 2012 / Published online: 30 October 2012
� Springer-Verlag Italia 2012
Abstract Knee osteoarthritis (OA) is a common chronic
degenerative disease characterized by the loss of articular
cartilage components due to an imbalance between extra-
cellular matrix destruction and repair. The proinflammatory
cytokines involved in OA, TNFa and IL1b, are considered
the major implicated. The aim of this study was to inves-
tigate the relationship between TNFa -308 and -238
polymorphisms with messenger RNA (mRNA) and soluble
TNFa expression in knee OA patients and healthy subjects
(HS). Case–control study involved 50 knee OA patients
classified according to 1986 ACR Classification Criteria, as
well as 100 HS. The Western Ontario and McMaster
Universities Osteoarthritis Index and Lequesne disability
index were applied to OA patients. The -308 and -238
polymorphisms were determined by polymerase chain
reaction–restriction fragment length polymorphism tech-
nique. The TNFa mRNA expression was quantified by
real-time PCR using TaqMan method. The sTNFa levels
were measured by enzyme-linked immunosorbent assay.
The TNFa mRNA expression in knee OA patients was
higher than in HS (1.56-fold). In addition, the TNFamRNA expression was higher in carriers of G allele in the
knee OA group for both polymorphisms. The sTNFa levels
were increased in G/G versus G/A genotypes in both
studied polymorphisms (p \ 0.05). However, the TNFa-308 and -238 genotypes did not show statistical differ-
ences between groups. The G allele of TNFa -308 and
-238 polymorphisms is associated with high mRNA and
soluble expression in knee OA patients. However, it is not
a marker of susceptibility in Western Mexico. Further
studies are necessary to confirm these findings.
Keywords Knee osteoarthritis � TNF alpha expression �TNF alpha levels � TNF alpha polymorphisms
Introduction
Knee osteoarthritis (OA) is a common chronic degenera-
tive disease characterized by the loss of articular cartilage
components due to an imbalance between extracellular
matrix destruction and repair [1]. Although articular car-
tilage breakdown is a major characteristic of OA, other
joint tissues such as the synovial membrane and subchon-
dral bone actively participate in the progression of the
disease [2].
Jose Francisco Munoz-Valle and Edith Oregon-Romero contributed
equally to this work.
J. F. Munoz-Valle (&) � E. Oregon-Romero
Grupo de Inmunogenetica Funcional, Departamento de Biologıa
Molecular y Genomica, Centro Universitario de Ciencias de la
Salud, Universidad de Guadalajara, Insurgentes 244-1, Colonia
Lomas de Atemajac, C.P. 45178 Zapopan, Guadalajara, Jalisco,
Mexico
e-mail: [email protected]
H. Rangel-Villalobos
Instituto de Investigacion en Genetica Molecular, Centro
Universitario de la Cienega, Universidad de Guadalajara,
Ocotlan, Jalisco, Mexico
G. E. Martınez-Bonilla
Servicio de Reumatologıa, OPD Hospital Civil de Guadalajara
‘‘Fray Antonio Alcalde’’, Guadalajara, Jalisco, Mexico
E. Castaneda-Saucedo � L. Salgado-Goytia �M. A. Leyva-Vazquez � B. Illades-Aguiar �L. d. C. Alarcon-Romero � M. Espinoza-Rojo � I. Parra-Rojas
Unidad Academica de Ciencias Quımico Biologicas,
Universidad Autonoma de Guerrero, Chilpancingo,
Guerrero, Mexico
123
Clin Exp Med (2014) 14:61–67
DOI 10.1007/s10238-012-0216-3

Among the proinflammatory cytokines involved in OA,
TNFa and IL1b are considered the major implicated. Both
cytokines are produced by chondrocytes, mononuclear
cells, osteoblasts and synovial tissues. In OA patients,
TNFa levels are elevated in the synovial fluid, synovial
membrane, subchondral bone and cartilage [2].
The TNFa gene is located within the human leukocyte
antigen class III region. Previous reports have suggested
that the endogenous production of TNFa may be associated
with TNFa polymorphisms influencing the messenger
RNA (mRNA) and protein expression [3–6]. The TNFa-308 and -238 polymorphisms (G/G genotype) have been
associated with high mRNA expression and soluble TNFalevels in rheumatoid arthritis (RA). Besides, in human
blood leukocytes the G allele of TNFa -308 polymor-
phism was associated with higher mRNA expression
compared to the A allele [7].
The aim of this study was to investigate the association
of the TNFa -308 and -238 polymorphisms with the
mRNA expression and soluble TNFa levels in knee OA
patients.
Materials and methods
Patients and healthy subjects
We selected 50 OA patients attending the Hospital Civil
‘‘Fray Antonio Alcalde,’’ Rheumatology Department. All
patients fulfilled the 1986 classification criteria for knee
OA of the American College of Rheumatology. The
diagnosis of OA was based on clinical evaluations,
excluding metabolic causes. Idiopathic OA patients were
selected. WOMAC and Lequesne disability indexes were
applied to OA patients at the beginning of the study [8, 9].
As a control group, 100 healthy subjects (HS, between the
ages of 22 and 67), residents from Guadalajara, Jalisco,
Mexico, who had the same ethnic and geographical back-
ground than the OA patients, were included. The inclusion
criteria for the control group were as follows: [18 years
old, clinically healthy individuals. All participants were of
the Mexican Mestizo population. According to the National
Institute of Anthropology, the definition of a Mexican
Mestizo states that the individual must be born in the
country, having a Spanish last name, with a family history
of Mexican ancestors and at least back to the third gener-
ation [10].
Ethical considerations
An informed written consent was obtained from all
subjects before enrollment in the study, according to the
ethical guidelines of the 2008 Declaration of Helsinki. The
study was approved by the Ethics Committee of the Hos-
pital Civil ‘‘Fray Antonio Alcalde.’’ Registration number:
CI-25011.
PCR–RFLP of TNFa promoter polymorphism
Genomic DNA was extracted from 3 mL of peripheral
blood leukocyte according to Miller method [11]. Ampli-
fication of TNFa -308 and -238 promoter region was
performed by PCR in a Thermal Cycler (iCyclerTM BIO-
RAD Life Science Research Products) using the following
primers: for the TNFa -308 polymorphism, 50-AGG CAA
TAG GTT TTG AGG GCC AT-30 (Forward) and 50-TCC
TCC CTG CTC CGA TTC CG-30 (Reverse) [12]; and for
the TNFa -238 polymorphism, 50-AGA AGA CCC CCC
TCG GAA CC-30 (Forward) and 50-ATC TGG AGG AAG
CGG TAG TG-30 (Reverse) [13]. The amplification con-
ditions for both TNFa polymorphism were reported pre-
viously [14]. The electrophoretic TNFa genotype
restriction patters were as follows: For TNFa -308 poly-
morphism, digestion fragments of 87 and 20 bp represent
the wild-type genotype (G/G). Fragments of 107, 87 and
20 bp represent the heterozygote genotype (G/A). The
107-bp fragment represents the homozygote genotype (A/
A). For the TNFa -238 polymorphism, fragments of 152
and 133 bp represent the wild genotype (G/G). Fragments
of 152, 133 and 19 bp represent the heterozygote genotype
(G/A). A unique 152-bp fragment represents the homozy-
gote genotype (A/A). The experiments were made in
duplicate, and for each genotype (n = 3), random samples
were sequenced using an ABIPRISM 310 Sequencer
(Applied Biosystems) in order to confirm the above results.
RNA extraction and reverse transcription
Peripheral blood was collected in EDTA blood collection
tubes (BD Vaccutainer, NJ, USA). Immediately, the PBMC
were isolated using dextran reagent (Sigma Chemical Co,
St. Louis MO, USA), and the total RNA was immediately
obtained using Trizol reagent (InvitrogenTM, Carlsbad, CA,
USA) according to Chomiczyki and Sacchi method [15].
The maximum amount of time between drawing blood and
beginning procedures for mRNA extraction from the OA
and control samples was 1 h. RNA concentration was
determined by spectrophotometry, and RNA integrity was
corroborated on 1 % agarose gel. The cDNA synthesis was
performed using oligo(dT)12–18 primer. Briefly, 2 lg of
total RNA was used in a reaction containing RNase-free
water to 9 and 1 lL oligo(dT)12–18 primer. The reaction
was incubated at 75 �C for 15 min followed by 5 min on
ice. After that, 4 lL of 5X first-strand buffer, 4 lL 2.5 mM
of each dNTPs (InvitrogenTM, Carlsbad, CA, USA),
0.25 lL of 10 U/lL RNase inhibitor, 1 lL of 200 U/lL
62 Clin Exp Med (2014) 14:61–67
123

Moloney murine leukemia virus reverse transcriptase and
0.75 lL RNase-free water were added. The reactions were
carried out at 75 �C for 1 h followed by 5 min of incuba-
tion at 95 �C. The cDNA samples were stored at -80 �C
until the real-time PCR assays.
Real-time PCR for TNFa
The TNFa and glyceraldehyde 3-phosphate dehydrogenase
(GAPDH) expression was quantified using ABI Prism 7500
Sequence Detection System (Applied Biosystems), according
to the manufacturer’s protocol. Reaction for the real-time
PCR using TaqMan detection consisted of 10 lL of a 20X
TaqMan buffer (Applied Biosystem, Foster City, CA, USA);
1 lL of a mixture containing 5X TaqMan MGB probes and
18 lM of each primer (Applied Biosystem, Foster City, CA,
USA); 2 lL of cDNA; and 20 lL of water. The reactions
were performed in MicroAmp 96-well plate capped with
MicroAmp optical caps. All samples were incubated at 50 �C
for 2 min and 95 �C for 10 min and then cycled at 95 �C for
15 s and 60 �C for 1 min for 50 cycles. Controls with no
template cDNA were performed in each assay. The mRNA
expression levels were quantified using the critical threshold
value (Ct). Relative gene expression levels were obtained
using the 2�DDCt method (expressed as relative expression
units). Each sample was tested in triplicate.
sTNFa quantification
The sTNFa levels (R&D Systems, Minneapolis, MN, USA)
were measured using serum samples from OA patients and
HS by ELISA. The range of detection was 15.6–1,000 pg/mL,
and the sensitivity of the assay was\4.4 pg/mL. The sTNFaproduction was calculated from a standard curve of the cor-
responding recombinant human TNFa.
Study design and statistics
The study was a case–control type. Differences in genetic
and allelic frequencies between groups were studied using
chi-square test (v2), odds ratio (OR) and 95 % confidence
intervals (95 % CI). If the number in any cell was \5,
Fisher’s exact test was performed. The Student’s t test was
used to compare the two groups. Mann–Whitney U test was
used to evaluate the association between carriership of
TNFa -308, TNFa -238 and sTNFa levels. We used v2
test for comparison of TNFa mRNA expression propor-
tions. Differences in sTNFa levels were evaluated by
Pearson’s correlation (rho). Probability values less than
0.05 were considered statistically significant. The analysis
was performed using SPSS version 10.0, Epi Info version
2002 and GraphPad Prism 5 software.
Results
The mean age of OA patients was 55 years (range 31–86), and
the HS mean age was 40 years (range 22–67). Forty-four OA
patients were treated with nonsteroidal anti-inflammatory
drugs (NSAIDs). The clinical and demographic characteris-
tics of the OA patients in relationship to TNFa -308 and
-238 genotypes are shown in Table 1.
Table 1 Demographic and clinical characteristics according to TNFa genotypes in OA patients
TNFa -308 TNF a -238
All patients
(n = 50)
G/G Genotype
(n = 44)
G/A Genotype
(n = 6)
p G/G Genotype
(n = 47)
G/A Genotype
(n = 3)
p
Demographics
Age, years 55 (31–86) 55 (31–85) 51 (44–72) – 55 (31–85) 55 (45–68) –
Men/women 2/48 2/42 0/6 – 2/45 0/3 –
Disease status
Disease duration, years 5 (0.5–20) 5 (0.5–20) 1 (1–1) NS 3 (0.5–15) 15 (10–20) 0.016
Drug treatment
NSAIDs 42/50 41/44 5/6 – 42/47 2/3 –
Clinical assessment
WOMAC Total score 34.97 (2–72) 33.39 (2–72) 45 (29–64) NS 35.41 (2–72) 29 (23–34) NS
WOMAC-Pain score 7.88 (1–15) 7.52 (1–15) 10.16 (6–13) NS 7.85 (1–15) 8.33 (8–9) NS
WOMAC-Stiffness score 2.90 (0–7) 2.76 (0–7) 3.83 (2–6) NS 3.02 (0–7) 1.33 (1–2) NS
WOMAC-Function score 24.18 (1–52) 23.10 (1–52) 31 (17–48) NS 24.53 (1–52) 19.33 (13–24) NS
Lequesne score 11.88 (3–21) 11.36 (3–18) 15.4 (8–21) 0.028 11.97 (3–21) 10.66 (8–13) NS
Values represent the mean, minimum and maximum scores
NS not significant, NSAIDs nonsteroidal anti-inflammatory drugs, WOMAC Western Ontario and McMaster Universities Osteoarthritis Index
Clin Exp Med (2014) 14:61–67 63
123
Our population was in Hardy–Weinberg equilibrium
(p [ 0.05). Allelic and genotypic frequencies of TNFa-308 and -238 polymorphisms in OA and HS are shown in
Table 2. The allelic frequencies in OA and HS groups were
as follows: 94 versus 97 % for -308G and 6 versus 3 % for
-308A; and 97 versus 94 % for -238G and 3 versus 6 % for
-238A. The TNFa -308 and -238 genotypes did not show
statistical differences between groups.
To investigate the functional impact of the TNFa -308
and -238 variants, the quantitative TNFa mRNA expres-
sion among the different genotypes for both polymor-
phisms (G/G, G/A) was analyzed. The A/A genotype was
not found in the studied population. The TNFa mRNA
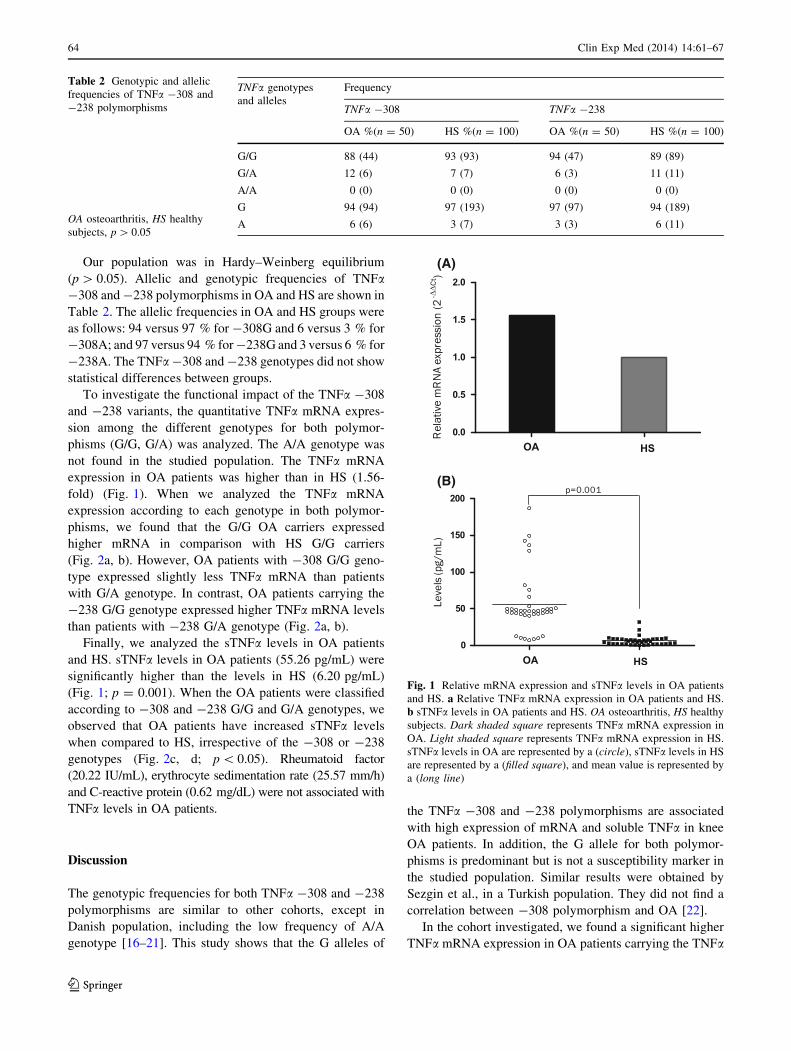
expression in OA patients was higher than in HS (1.56-
fold) (Fig. 1). When we analyzed the TNFa mRNA
expression according to each genotype in both polymor-
phisms, we found that the G/G OA carriers expressed
higher mRNA in comparison with HS G/G carriers
(Fig. 2a, b). However, OA patients with -308 G/G geno-
type expressed slightly less TNFa mRNA than patients
with G/A genotype. In contrast, OA patients carrying the
-238 G/G genotype expressed higher TNFa mRNA levels
than patients with -238 G/A genotype (Fig. 2a, b).
Finally, we analyzed the sTNFa levels in OA patients
and HS. sTNFa levels in OA patients (55.26 pg/mL) were
significantly higher than the levels in HS (6.20 pg/mL)
(Fig. 1; p = 0.001). When the OA patients were classified
according to -308 and -238 G/G and G/A genotypes, we
observed that OA patients have increased sTNFa levels
when compared to HS, irrespective of the -308 or -238
genotypes (Fig. 2c, d; p \ 0.05). Rheumatoid factor
(20.22 IU/mL), erythrocyte sedimentation rate (25.57 mm/h)
and C-reactive protein (0.62 mg/dL) were not associated with
TNFa levels in OA patients.
Discussion
The genotypic frequencies for both TNFa -308 and -238
polymorphisms are similar to other cohorts, except in
Danish population, including the low frequency of A/A
genotype [16–21]. This study shows that the G alleles of
the TNFa -308 and -238 polymorphisms are associated
with high expression of mRNA and soluble TNFa in knee
OA patients. In addition, the G allele for both polymor-
phisms is predominant but is not a susceptibility marker in
the studied population. Similar results were obtained by
Sezgin et al., in a Turkish population. They did not find a
correlation between -308 polymorphism and OA [22].
In the cohort investigated, we found a significant higher
TNFa mRNA expression in OA patients carrying the TNFa
Table 2 Genotypic and allelic
frequencies of TNFa -308 and
-238 polymorphisms
OA osteoarthritis, HS healthy
subjects, p [ 0.05
TNFa genotypes
and alleles
Frequency
TNFa -308 TNFa -238
OA %(n = 50) HS %(n = 100) OA %(n = 50) HS %(n = 100)
G/G 88 (44) 93 (93) 94 (47) 89 (89)
G/A 12 (6) 7 (7) 6 (3) 11 (11)
A/A 0 (0) 0 (0) 0 (0) 0 (0)
G 94 (94) 97 (193) 97 (97) 94 (189)
A 6 (6) 3 (7) 3 (3) 6 (11)
(A)
(B)
Fig. 1 Relative mRNA expression and sTNFa levels in OA patients
and HS. a Relative TNFa mRNA expression in OA patients and HS.
b sTNFa levels in OA patients and HS. OA osteoarthritis, HS healthy
subjects. Dark shaded square represents TNFa mRNA expression in
OA. Light shaded square represents TNFa mRNA expression in HS.
sTNFa levels in OA are represented by a (circle), sTNFa levels in HS
are represented by a (filled square), and mean value is represented by
a (long line)
64 Clin Exp Med (2014) 14:61–67
123
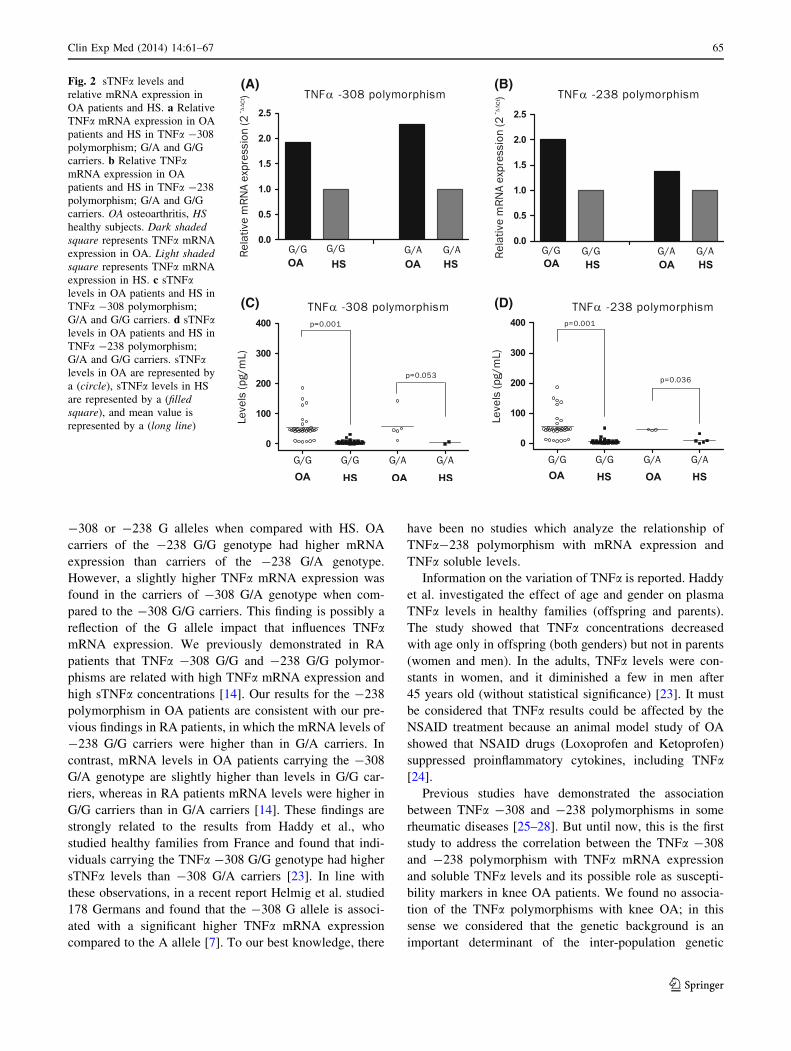
-308 or -238 G alleles when compared with HS. OA
carriers of the -238 G/G genotype had higher mRNA
expression than carriers of the -238 G/A genotype.
However, a slightly higher TNFa mRNA expression was
found in the carriers of -308 G/A genotype when com-
pared to the -308 G/G carriers. This finding is possibly a
reflection of the G allele impact that influences TNFamRNA expression. We previously demonstrated in RA
patients that TNFa -308 G/G and -238 G/G polymor-
phisms are related with high TNFa mRNA expression and
high sTNFa concentrations [14]. Our results for the -238
polymorphism in OA patients are consistent with our pre-
vious findings in RA patients, in which the mRNA levels of
-238 G/G carriers were higher than in G/A carriers. In
contrast, mRNA levels in OA patients carrying the -308
G/A genotype are slightly higher than levels in G/G car-
riers, whereas in RA patients mRNA levels were higher in
G/G carriers than in G/A carriers [14]. These findings are
strongly related to the results from Haddy et al., who
studied healthy families from France and found that indi-
viduals carrying the TNFa -308 G/G genotype had higher
sTNFa levels than -308 G/A carriers [23]. In line with
these observations, in a recent report Helmig et al. studied
178 Germans and found that the -308 G allele is associ-
ated with a significant higher TNFa mRNA expression
compared to the A allele [7]. To our best knowledge, there
have been no studies which analyze the relationship of
TNFa-238 polymorphism with mRNA expression and
TNFa soluble levels.
Information on the variation of TNFa is reported. Haddy
et al. investigated the effect of age and gender on plasma
TNFa levels in healthy families (offspring and parents).
The study showed that TNFa concentrations decreased
with age only in offspring (both genders) but not in parents
(women and men). In the adults, TNFa levels were con-
stants in women, and it diminished a few in men after
45 years old (without statistical significance) [23]. It must
be considered that TNFa results could be affected by the
NSAID treatment because an animal model study of OA
showed that NSAID drugs (Loxoprofen and Ketoprofen)
suppressed proinflammatory cytokines, including TNFa[24].
Previous studies have demonstrated the association
between TNFa -308 and -238 polymorphisms in some
rheumatic diseases [25–28]. But until now, this is the first
study to address the correlation between the TNFa -308
and -238 polymorphism with TNFa mRNA expression
and soluble TNFa levels and its possible role as suscepti-
bility markers in knee OA patients. We found no associa-
tion of the TNFa polymorphisms with knee OA; in this
sense we considered that the genetic background is an
important determinant of the inter-population genetic
(A) (B)
(D)(C)
Fig. 2 sTNFa levels and
relative mRNA expression in
OA patients and HS. a Relative
TNFa mRNA expression in OA
patients and HS in TNFa -308
polymorphism; G/A and G/G
carriers. b Relative TNFamRNA expression in OA
patients and HS in TNFa -238
polymorphism; G/A and G/G
carriers. OA osteoarthritis, HS
healthy subjects. Dark shaded
square represents TNFa mRNA
expression in OA. Light shaded
square represents TNFa mRNA
expression in HS. c sTNFalevels in OA patients and HS in
TNFa -308 polymorphism;
G/A and G/G carriers. d sTNFalevels in OA patients and HS in
TNFa -238 polymorphism;
G/A and G/G carriers. sTNFalevels in OA are represented by
a (circle), sTNFa levels in HS
are represented by a (filled
square), and mean value is
represented by a (long line)
Clin Exp Med (2014) 14:61–67 65
123

variability of the TNFa -308 and TNFa -238 polymor-
phisms and the peculiar characteristics of the Mexican
population. The population of the Western Mexico is
considered Mexican Mestizo and has been estimated that
the paternal ancestry in Western Mexican Mestizos is
mainly European (60–64 %), followed by Amerindian
(25–21 %) and African (15 %) [29, 30].
TNFa, together with IL 1b, is considered one of the most
important players in OA pathophysiology. TNFa is elevated
in synovial fluid, synovial membrane, subchondral bone and
cartilage [31, 32]. TNFa is associated with driving the
inflammatory cascade and stimulating the release of MMP-
1, MMP-3 and MMP-13 which are key regulators of carti-
lage destruction [33–35]. Previous results support the role of
TNFa in knee OA. Satannus et al. suggested that serum level
of TNFa is associated with knee cartilage loss and joint
space narrowing in older people [36]. Likewise, TNFa is
correlated with pain and also is associated with the total
WOMAC score including their subscales (pain, stiffness and
physical function). However, TNFa did not correlate with
the radiographic grading [37]. These findings suggest that
TNFa could be blocked to improve the inflammation pro-
cess, pain and physical function in OA patients.
The importance of our study is based on the relationship
found between the G allele of the polymorphism of TNFa-308 and -238 with high expression of mRNA and sol-
uble TNFa levels in knee OA. However, the limitations of
our study are the low heterozygosity found in the studied
population and that it was not possible to identify any
individual with the homozygous A/A genotype for both
studied polymorphisms.
Conclusions
In conclusion, this study showed that the G allele of TNFa-308 and -238 polymorphisms is associated with high
mRNA and soluble expression in knee OA. However, it is
not a marker of susceptibility in Western Mexico. Further
studies are necessary to confirm these findings.
Acknowledgments This work was supported by Grant No. 69235 to
JFMV of the CONACYT (Fondo Sectorial Secretarıa de Salud-IMSS-
ISSSTE CONACYT, Mexico-Universidad de Guadalajara) and Grant
No. 147778 of the Fondo Mixto CONACYT-Gobierno del Estado de
Guerrero 2010-01.
Conflict of interest None.
References
1. Orita S, Koshi T, Mitsuka T, Miyagi M, Inoue G, Arai G,
Ishikawa T, Hanaoka E, Yamashita K, Yamashita M, Eguchi Y,
Toyone T, Takahashi K, Ohtori S (2011) Associations between
proinflammatory cytokines in the synovial fluid and radiographic
grading and pain-related scores in 47 consecutive patients with
osteoarthritis of the knee. BMC Musculoskelet Disord 12(1):144
2. Kapoor M, Martel-Pelletier J, Lajeunesse D, Pelletier JP, Fahmi
H (2011) Role of proinflammatory cytokines in the pathophysi-
ology of osteoarthritis. Nat Rev Rheumatol 7(1):33–42
3. Pociot F, Briant L, Jongeneel CV, Molvig J, Worsaae H, Abbal M,
Thomsen M, Nerup J, Cambon-Thomsen A (1993) Association of
tumor necrosis factor (TNF) and class II major histocompatibility
complex alleles with the secretion of TNF-alpha and TNF-beta by
human mononuclear cells: a possible link to insulin-dependent
diabetes mellitus. Eur J Immunol 23:224–231
4. Turner DM, Grant SC, Lamb WR, Brenchley PE, Dyer PA,
Sinnott PJ, Hutchinson IV (1995) A genetic marker of high TNF-
alpha production in heart transplant recipients. Transplantation
60:1113–1117
5. Wilson AG, Symons JA, Mcdowell TL, McDevitt HO, Duff GW
(1997) Effects of a polymorphism in the human tumor necrosis
factor alpha promoter on transcriptional activation. Proc Natl
Acad Sci USA 94(7):3195–3199
6. Uglialoro AM, Turbay D, Pesavento PA, Delgado JC, McKenzie
FE, Gribben JG, Hartl D, Yunis EJ, Goldfeld AE (1998) Identi-
fication of three new single nucleotide polymorphisms in the
human tumor necrosis factor-alpha gene promoter. Tissue Anti-
gens 52:359–367
7. Helmig S, Aliahmadi N, Stephan P, Dohrel J, Schneider J (2011)
TNF-a -308 genotypes are associated with TNF-a and TGF-b1
mRNA expression in blood leucocytes of humans. Cytokine
53(3):306–310
8. Bellamy N, Buchanan WW, Goldsmith CH (1988) Validation
study of WOMAC: a health status instrument for measuring
clinically important patient relevant outcomes to antirheumatic
drug therapy in patients with osteoarthritis of the hip or knee.
J Rheumatol 15:1833–1840
9. Lequesne MG, Mery C, Samson M, Gerard P (1987) Indexes of
severity for osteoarthritis of the hip and knee. Validation—value
in comparison with other assessment test. Scand J Rheumatol
65:85–89
10. Gorodezky C, Alaez C, Vazquez-Garcıa MN, de la Rosa G,
Infante E, Balladares S, Toribio R, Perez-Luque E, Munoz L
(2001) The genetic structure of Mexican Mestizos of different
locations: tracking back their origins through MHC genes, blood
groups systems, and microsatellites. Hum Immunol 62:979–991
11. Miller SA, Dykes DD, Polesky HF (1988) A simple salting out
procedure for extracting DNA from human nucleated cells.
Nucleic Acids Res 16:1215
12. Verity DH, Wallace GR, Vaughan RW, Kondeatis E, Madanat W,
Zureikat H, Fayyad F, Marr JE, Kanawati CA, Stanford MR (1999)
HLA and tumour necrosis factor (TNF) polymorphisms in ocular
Behcet’s disease. Tissue Antigens 54:264–272
13. Fargion S, Valenti L, Dongiovanni P, Scaccabarozzi A, Fracan-
zani AL, Taioli E, Mattioli M, Sampietro M, Fiorelli G (2001)
Tumor necrosis factor alpha promoter polymorphisms influence
the phenotypic expression of hereditary hemochromatosis. Blood
97:3707–3712
14. Oregon-Romero E, Vazquez-Del Mercado M, Ruiz-Quezada SL,
Navarro-Hernandez RE, Rangel-Villalobos H, Martınez-Bonilla
G, Bernard-Medina AG, Armendariz-Borunda J, Garcıa-Banue-
los J, Munoz-Valle JF (2008) Tumor necrosis factor a -308 and
-238 polymorphism in rheumatoid arthritis: association with
messenger RNA expression and sTNF-a. J Investig Med 56(7):
937–943
15. Chomiczyki P, Sacchi N (1987) Single step method of RNA
isolation by acid guanidinium thiocyanate-phenol-chloroform
extraction. Annal Biochem 162:156–159
66 Clin Exp Med (2014) 14:61–67
123

16. Moos V, Rudwaleit M, Herzog V, Hohlig K, Sieper J, Muller B
(2000) Association of genotypes affecting the expression of
interleukin-1beta or interleukin-1 receptor antagonist with
osteoarthritis. Arthritis Rheum 43(11):2417–2422
17. de Maat MP, Bladbjerg EM, Hjelmborg JB, Bathum L, Jespersen
J, Christensen K (2004) Genetic influence on inflammation
variables in the elderly. Arterioscler Thromb Vasc Biol 24(11):
2168–2173
18. Rodrıguez-Carreon AA, Zuniga J, Hernandez-Pacheco G, Rod-
rıguez-Perez JM, Perez-Hernandez N, Montes de Oca JV, Cardiel
MH, Granados J, Vargas-Alarcon G (2005) Tumor necrosis fac-
tor-alpha -308 promoter polymorphism contributes indepen-
dently to HLA alleles in the severity of rheumatoid arthritis in
Mexicans. J Autoimmun 24(1):63–68
19. Berdeli A, Tabel Y, Celik HA, Ozyurek R, Dogrusoz B, Aydin
HH (2006) Lack of association between TNFalpha gene poly-
morphism at position -308 and risk of acute rheumatic fever in
Turkish patients. Scand J Rheumatol 35(1):44–47
20. Merza M, Farnia P, Anoosheh S, Varahram M, Kazampour M,
Pajand O, Saeif S, Mirsaeidi M, Masjedi MR, Velayati AA,
Hoffner S (2009) The NRAMPI, VDR and TNF-alpha gene
polymorphisms in Iranian tuberculosis patients: the study on host
susceptibility. Braz J Infect Dis 13(4):252–256
21. Liu C, Wang J, Zhou S, Wang B, Ma X (2010) Association
between -238 but not -308 polymorphism of Tumor necrosis
factor alpha (TNF-alpha)v and unexplained recurrent spontane-
ous abortion (URSA) in Chinese population. Reprod Biol
Endocrinol 8:114
22. Sezgin M, Barlas IO, Ankarali HC, Altintas ZM, Turkmen E,
Gokdogan T, Sahin G, Erdal ME (2008) Tumour necrosis factor
alpha -308G/A gene polymorphism: lack of association with
knee osteoarthritis in a Turkish population. Clin Exp Rheumatol
26(5):763–768
23. Haddy N, Sass C, Maumus S, Marie B, Droesch S, Siest G,
Lambert D, Visvikis S (2005) Biological variations, genetic
polymorphism and familial resemblance of TNF-a and IL-6
concentrations: STANISLAS cohort. Eur J Hum Genet 13:
109–117
24. Orita S, Ishikawa T, Miyagi M, Ochiai N, Inoue G, Eguchi Y,
Kamoda H, Arai G, Suzuki M, Sakuma Y, Oikawa Y, Toyone T,
Aoki Y, Takahashi K, Ohtori S (2012) Percutaneously absorbed
NSAIDs attenuate local production of proinflammatory cytokines
and suppress the expression of c-Fos in the spinal cord of a rodent
model of knee osteoarthritis. J Orthop Sci 17(1):77–86
25. Lee YH, Ji JD, Bae SC, Song GG (2010) Associations between
tumor necrosis factor-alpha (TNF-alpha) -308 and -238 G/A
polymorphisms and shared epitope status and responsiveness to
TNF-alpha blockers in rheumatoid arthritis: a metaanalysis
update. J Rheumatol 37(4):740–746
26. Ozen S, Alikasifoglu M, Bakkaloglu A, Duzova A, Jarosova K,
Nemcova D, Besbas N, Vencovsky J, Tuncbilek E (2002)
Tumour necrosis factor alpha G ? A -238 and G ? A -308
polymorphisms in juvenile idiopathic arthritis. Rheumatology
(Oxford) 41(2):223–227
27. Sousa E, Caetano-Lopes J, Pint P, Pimentel F, Teles J, Canhao H,
Rodrigues A, Resende C, Mourao AF, Ribeiro C, Pinto TL, Rosa
CM, da Silva JA, Branco J, Ventura F, Queiroz MV, Fonseca JE
(2009) Ankylosing spondylitis susceptibility and severity–con-
tribution of TNF gene promoter polymorphisms at positions
-238 and -308. Ann N Y Acad Sci 1173:581–588
28. Pan HF, Leng RX, Wang C, Qin WZ, Chen LL, Zha ZQ, Tao JH,
Ye DQ (2011) Association of TNF-a promoter -308 A/G poly-
morphism with susceptibility to systemic lupus erythematosus: a
meta-analysis. Rheumatol Int. doi:10.1007/s00296-011-1924-9
29. Lisker R, Ramırez E, Gonzalez-Villalpando C, Stern MP (1995)
Racial admixture in a mestizo population from Mexico City. Am
J Hum Biol 7(2):213–216
30. Rangel-Villalobos H, Munoz-Valle JF, Gonzalez-Martın A,
Gorostiza A, Magana MT, Paez-Riberos LA (2008) Genetic
admixture, relatedness, and structure patterns among Mexican
populations revealed by the Y-chromosome. Am J Phys
Anthropol 135(4):448–461
31. Vignon E, Balblanc JC, Mathieu P, Louisot P, Richard M (1993)
Metalloprotease activity, phospholipase A2 activity and cytokine
concentration in osteoarthritis synovial fluids. Osteoarthr Cartil
1(2):115–120
32. Smith MD, Triantafillou S, Parker A, Youssef PP, Coleman M
(1997) Synovial membrane inflammation and cytokine produc-
tion in patients with early osteoarthritis. J Rheumatol 24(2):
365–371
33. Lefebvre V, Peeters-Joris C, Vaes G (1990) Modulation by
interleukin 1 and tumor necrosis factor alpha of production of
collagenase, tissue inhibitor of metalloproteinases and collagen
types in differentiated and dedifferentiated articular chondro-
cytes. Biochim Biophys Acta 1052(3):366–378
34. Reboul P, Pelletier JP, Tardif G, Cloutier JM, Martel-Pelletier J
(1996) The new collagenase, collagenase-3, is expressed and
synthesized by human chondrocytes but not by synoviocytes. A
role in osteoarthritis. J Clin Invest 97(9):2011–2019
35. Westacott CI, Barakat AF, Wood L, Perry MJ, Neison P, Bisbinas
I, Armstrong L, Millar AB, Elson CJ (2000) Tumor necrosis
factor alpha can contribute to focal loss of cartilage in osteoar-
thritis. Osteoarthr Cartil 8(3):213–221
36. Stannus O, Jones G, Cicuttini F, Parameswaran V, Quinn S,
Burgess J, Ding C (2010) Circulating levels of IL-6 and TNF-aare associated with knee radiographic osteoarthritis and knee
cartilage loss in older adults. Osteoarthr Cartil 18(11):1441–1447
37. Orita S, Koshi T, Mitsuka T, Miyagi M, Inoue G, Arai G,
Ishikawa T, Hanaoka E, Yamashita K, Yamashita M, Eguchi Y,
Toyone T, Takahashi K, Ohtori S (2011) Associations between
proinflammatory cytokines in the synovial fluid and radiographic
grading and pain-related scores in 47 consecutive patients with
osteoarthritis of the knee. BMC Musculskelet Disord 12:144
Clin Exp Med (2014) 14:61–67 67
123









![Association between TNF-alpha polymorphism and the age of … · 2020-01-31 · TNF-alpha gene displayed better cognition functions [28] and decreased TNF-alpha serum levels were](https://static.fdocuments.in/doc/165x107/5f9df0f972c98e2f064624b0/association-between-tnf-alpha-polymorphism-and-the-age-of-2020-01-31-tnf-alpha.jpg)








