
Heterogeneity of Keratin Expression in Mouse - Cancer Research
10
[CANCER RESEARCH 45, 2760-2768, June 1985] Heterogeneity of Keratin Expression in Mouse Mammary Hyperplastic Alveolar Nodules and Adenocarcinomas1 Harold L. Asch2 and Bonnie B. Asch Department of Experimental Pathology, Roswell Park Memorial Institute, Buffalo, New York 14263 ABSTRACT The keratins and other cytoskeletal proteins expressed by normal, preneoplastic, and malignant mammary tissues in BALB/ c mice and by cells in primary cultures established from these tissues were analyzed and compared. The preneoplastic lesions were hyperplastic alveolar nodules (HAN) derived originally from mice treated by hormonal stimulation (02), exposed to a chemical carcinogen (C4), or spontaneously expressing mouse mammary tumor virus (CV2) and maintained by serial transplantation. All tumors were mammary adenocarcinomas which developed as primary neoplasms from the HAN outgrowth lines. Cytoskeletal extracts were prepared from the tissues and cultured cells and subjected to two-dimensional polyacrylamide gel electrophore- sis. Comparison of the major polypeptides in the normal and abnormal tissue extracts revealed considerable similarities in the cytoskeletal profiles. Three basic and seven acidic polypeptides ranging in molecular weight from 40,000 to 90,000 were regularly identified. However, notable differences were also found. A M, 55,000 keratin (IEF 55) was prominent in one HAN, the D2, and all tumor tissues but not in normal gland. Likewise, a M, 46,000 polypeptide (IEF 46), which has been tentatively identified previ ously as a keratin, was absent in normal epithelium but present in all abnormal tissues except the C4 and CV2 HAN. A M, 58,000 polypeptide (NEPHGE 58) was not detected in normal gland or the C4 lesions but was found in all other abnormal tissues. The overall pattern of polypeptides in cytoskeletal extracts from normal and abnormal mammary cells in primary culture resem bled that of the corresponding tissue but also had important differences. In all cell cultures, IEF 46 and IEF 55 were major species, while the larger and more basic components were markedly reduced. A M, 56,000 polypeptide (NEPHGE 56) was detected only in C4 HAN and C4 and CV2 tumor cells. Trace or small, variable amounts of a M, 57,000 basic keratin (NEPHGE 57) were present in normal and D2 tissues and cultured cells. NEPHGE 57 was dramatically increased in C4 and CV2 tissues and cultured cells and may be related to expression of squamous metaplasia and keratinization which are characteristic of these lesions. Although production of IEF 46 and IEF 55 may be associated with neoplastic progression of mammary epithelium, particularly in vivo, the association is not exclusive since normal cells express these polypeptides when grown in primary culture. In addition, correlations between altered keratin expression and the mode of induction of the mammary lesions were not obvious. INTRODUCTION Immunocytochemical and biochemical analyses have shown 1This work was supported by Grant CA32937 from the National Cancer Institute. 2To whom requests for reprints should be addressed. Received 11/16/84; revised 2/18/85: accepted 2/20/85. that expression of keratins is maintained in epithelial cells during hyperplastic, neoplastic, and metastatic processes (1-12). In fact, the presence or absence of keratins in cells of primary and metastatic tumors can serve as an important indicator of the cell of origin in certain problems of clinical diagnosis (13-17). Fur thermore, the set of keratins expressed by human hepatocellular carcinomas and colonie carcinomas is essentially the same as that of their normal counterparts (7). Although the basic tissue- specific pattern of keratins is usually preserved in tumor cells, epithelia vary considerably in the gain or loss of individual keratins during tumorigenesis (reviewed in Ref. 7). Studies on human and rodent tissues demonstrated that certain keratins were de creased or absent in tumors of epidermal, esophageal, forestom- ach, tongue, vaginal, and anal epithelium as compared to the corresponding normal tissue (7, 9-11). Ductal carcinomas of the human breast both lost and gained expression of keratins relative to those in ductal epithelium of normal breast (6, 7). On the basis of such results, keratins have been proposed as candidate markers of neoplastic progression in epithelial cells (3, 5-9,17). However, not all keratin changes found in tumors may be directly related to the neoplastic state per se. Instead, some may reflect other factors such as increased cell proliferation, differences in the degree of differentiation of the cells, or, in an epithelium with more than one type of cell, dominance of one particular cell that may normally be a minor constituent in a mixed population. Examples of these situations have been found with tumors of the bladder and epidermis (18, 19). Each epithelium must therefore be evaluated individually in order to determine the actual basis for any alterations in keratin expression that may occur in cancer. These considerations are especially pertinent to breast can cers. Three types of epithelial cells comprise the fully developed mammary gland: ductal; alveolar (secretory); and myoepithelial. The roles and fates of the 3 cell types during mammary tumori genesis have been the subject of ongoing controversy for many years (20-26). Carcinomas can develop from either the ductal or lobular compartments of the gland. However, mammary epithelial cells in tumors often lose their distinguishing features as well as their normal architectural organization, and the actual cell of origin of most breast cancers as well as the relative percentages of the 3 epithelial cells in a tumor are unknown (21, 24). Keratins may offer one type of marker for distinguishing among ductal, alveolar, and myoepithelial cells. The use of antikeratin antibodies in immunocytochemistry has suggested that the 3 types of mammary epithelial cells may differ in the subsets of keratins that they express (2,5,27, 28). Recent biochemical and immunoblot data from our laboratory support this idea and indicate that myoepithelial cells in mouse mammary gland pref erentially express M, 50,000 and 40,000 keratins while luminal epithelial cells of ducts and alveoli express little or none of these polypeptides (29). Such differences provide a basis for recogniz- CANCER RESEARCH VOL. 45 JUNE 1985 2760 on April 12, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Transcript of Heterogeneity of Keratin Expression in Mouse - Cancer Research

[CANCER RESEARCH 45, 2760-2768, June 1985]
Heterogeneity of Keratin Expression in Mouse Mammary Hyperplastic AlveolarNodules and Adenocarcinomas1
Harold L. Asch2 and Bonnie B. Asch
Department of Experimental Pathology, Roswell Park Memorial Institute, Buffalo, New York 14263
ABSTRACT
The keratins and other cytoskeletal proteins expressed bynormal, preneoplastic, and malignant mammary tissues in BALB/c mice and by cells in primary cultures established from thesetissues were analyzed and compared. The preneoplastic lesionswere hyperplastic alveolar nodules (HAN) derived originally frommice treated by hormonal stimulation (02), exposed to a chemicalcarcinogen (C4), or spontaneously expressing mouse mammarytumor virus (CV2) and maintained by serial transplantation. Alltumors were mammary adenocarcinomas which developed asprimary neoplasms from the HAN outgrowth lines. Cytoskeletalextracts were prepared from the tissues and cultured cells andsubjected to two-dimensional polyacrylamide gel electrophore-
sis. Comparison of the major polypeptides in the normal andabnormal tissue extracts revealed considerable similarities in thecytoskeletal profiles. Three basic and seven acidic polypeptidesranging in molecular weight from 40,000 to 90,000 were regularlyidentified. However, notable differences were also found. A M,55,000 keratin (IEF 55) was prominent in one HAN, the D2, andall tumor tissues but not in normal gland. Likewise, a M, 46,000polypeptide (IEF 46), which has been tentatively identified previously as a keratin, was absent in normal epithelium but presentin all abnormal tissues except the C4 and CV2 HAN. A M, 58,000polypeptide (NEPHGE 58) was not detected in normal gland orthe C4 lesions but was found in all other abnormal tissues. Theoverall pattern of polypeptides in cytoskeletal extracts fromnormal and abnormal mammary cells in primary culture resembled that of the corresponding tissue but also had importantdifferences. In all cell cultures, IEF 46 and IEF 55 were majorspecies, while the larger and more basic components weremarkedly reduced. A M, 56,000 polypeptide (NEPHGE 56) wasdetected only in C4 HAN and C4 and CV2 tumor cells. Trace orsmall, variable amounts of a M, 57,000 basic keratin (NEPHGE57) were present in normal and D2 tissues and cultured cells.NEPHGE 57 was dramatically increased in C4 and CV2 tissuesand cultured cells and may be related to expression of squamousmetaplasia and keratinization which are characteristic of theselesions. Although production of IEF 46 and IEF 55 may beassociated with neoplastic progression of mammary epithelium,particularly in vivo, the association is not exclusive since normalcells express these polypeptides when grown in primary culture.In addition, correlations between altered keratin expression andthe mode of induction of the mammary lesions were not obvious.
INTRODUCTION
Immunocytochemical and biochemical analyses have shown
1This work was supported by Grant CA32937 from the National Cancer Institute.2To whom requests for reprints should be addressed.
Received 11/16/84; revised 2/18/85: accepted 2/20/85.
that expression of keratins is maintained in epithelial cells duringhyperplastic, neoplastic, and metastatic processes (1-12). In
fact, the presence or absence of keratins in cells of primary andmetastatic tumors can serve as an important indicator of the cellof origin in certain problems of clinical diagnosis (13-17). Fur
thermore, the set of keratins expressed by human hepatocellularcarcinomas and colonie carcinomas is essentially the same asthat of their normal counterparts (7). Although the basic tissue-
specific pattern of keratins is usually preserved in tumor cells,epithelia vary considerably in the gain or loss of individual keratinsduring tumorigenesis (reviewed in Ref. 7). Studies on human androdent tissues demonstrated that certain keratins were decreased or absent in tumors of epidermal, esophageal, forestom-
ach, tongue, vaginal, and anal epithelium as compared to thecorresponding normal tissue (7, 9-11). Ductal carcinomas of the
human breast both lost and gained expression of keratins relativeto those in ductal epithelium of normal breast (6, 7).
On the basis of such results, keratins have been proposed ascandidate markers of neoplastic progression in epithelial cells (3,5-9,17). However, not all keratin changes found in tumors may
be directly related to the neoplastic state per se. Instead, somemay reflect other factors such as increased cell proliferation,differences in the degree of differentiation of the cells, or, in anepithelium with more than one type of cell, dominance of oneparticular cell that may normally be a minor constituent in a mixedpopulation. Examples of these situations have been found withtumors of the bladder and epidermis (18, 19). Each epitheliummust therefore be evaluated individually in order to determinethe actual basis for any alterations in keratin expression thatmay occur in cancer.
These considerations are especially pertinent to breast cancers. Three types of epithelial cells comprise the fully developedmammary gland: ductal; alveolar (secretory); and myoepithelial.The roles and fates of the 3 cell types during mammary tumorigenesis have been the subject of ongoing controversy for manyyears (20-26). Carcinomas can develop from either the ductal or
lobular compartments of the gland. However, mammary epithelialcells in tumors often lose their distinguishing features as well astheir normal architectural organization, and the actual cell oforigin of most breast cancers as well as the relative percentagesof the 3 epithelial cells in a tumor are unknown (21, 24).
Keratins may offer one type of marker for distinguishing amongductal, alveolar, and myoepithelial cells. The use of antikeratinantibodies in immunocytochemistry has suggested that the 3types of mammary epithelial cells may differ in the subsets ofkeratins that they express (2,5,27, 28). Recent biochemical andimmunoblot data from our laboratory support this idea andindicate that myoepithelial cells in mouse mammary gland preferentially express M, 50,000 and 40,000 keratins while luminalepithelial cells of ducts and alveoli express little or none of thesepolypeptides (29). Such differences provide a basis for recogniz-
CANCER RESEARCH VOL. 45 JUNE 1985
2760
on April 12, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

KERATINS IN MOUSE MAMMARY HYPERPLASIAS AND CARCINOMAS
ing the individual cell types. At the same time, these differencescomplicate the interpretation of biochemical analyses of keratinsin breast cancers because an altered keratin profile could be dueto changes in the relative numbers of the 3 epithelial cell typesrather than to expression of cancer.
We have undertaken a series of investigations to assess thepotential of keratins as molecular markers in mammary tumori-
genesis. We recently analyzed the polypeptides in cytoskeletalextracts of normal mouse mammary tissues by gel electropho-
resis combined with immunoblots using antikeratin antibodies(30). The hormonal influences of pregnancy and lactation inducenormal mammary epithelial cells to undergo extensive proliferation, differentiation, and functional activities. Only minor changesin cytoskeletal composition, none of which involved keratins,were found in the epithelium during the various stages of thenormal developmental cycle. Moreover, the data indicated thatductal and alveolar epithelial cells probably express the sameset of keratins. Results of that study provide a foundation forcomparing the cytoskeletal polypeptides, especially keratins,present in normal mouse mammary gland with those expressedin pathological conditions.
In the current investigation, we have asked the followingquestions. Do normal and malignant mouse mammary tissuesdiffer in their cytoskeletal polypeptides, particularly the keratins?If so, does the pattern of cytoskeletal components in preneo-
plastic hyperplasias resemble that of normal or tumor tissue? Ifchanges in keratins are associated with mammary tumorigen-
esis, are they directly related to neoplastia progression or toother factors? Do hyperplastic and malignant lesions of similarhistology but different etiologies have the same profiles of cytoskeletal polypeptides, including keratins? Do cultured epithelialcells derived from such tissues retain the same set of cytoskeletalpolypeptides, including keratins, as that expressed in wVo? Toexamine these questions, we have analyzed and compared thecytoskeletal polypeptides present in normal mouse mammaryepithelium with those found in preneoplastic and neoplasticmammary lesions of hormonal, viral, and chemical etiologies.
MATERIALS AND METHODS
Mammary Tissues. All mammary tissues were obtained from femaleBALB/cCri mice generously provided by Dr. D. Medina. Normal mammaryglands were taken from pregnant mice at 14 to 18 days of gestation.Preneoplastic mammary tissues were derived from 3 HAN3 lines that
were propagated by serial transplantation in cleared mammary fat padsof syngeneic mice as described by Medina (31). All mammary tumorsused in experiments were primary adenocarcinomas that developed inmice bearing transplants of the HAN lines. Pools of tissue from 4 to 5pregnant mice, 12 to 15 animals with HAN, or 1 to 2 mice with primarytumors were used for each respective extraction.
Mammary Cell Cultures. Mammary tissues were dissociated enzy-
matically, and the resulting cells were plated in plastic dishes (ComingGlass Works, Corning, NY) as primary cultures by the method in Ref.32. The cells were grown in Dulbecco's modified Eagle's medium con
taining 13% fetal bovine serum (Grand Island Biological Co., GrandIsland, NY), 18 mw A/-2-hydroxyethylpiperazine-AT-2-ethanesulfonicacid,
and gentamicin sulfate (50 ¿ig/ml;Schering Corp., Kenilworth, NJ). Forserum-free medium, the serum was omitted and fetuin (1 mg/ml; Sigma
3The abbreviations used are: HAN, hyperplastic alveolar nodule; 2-D PAGE, 2-
dimensional polyacrylamide gel electrophoresis; IEF, isoelectric focusing; NEPHGE,nonequilibrium pH gradient gel electrophoresis.
Chemical Co., St. Louis, MO), epidermal growth factor (10 ng/ml; Collaborative Research, Waltham, MA), 50 nw sodium selenite (Aldrich Chemical Co., Milwaukee, Wl), and 5-^g/ml amounts each of transferrin, insulin,
and hydrocortisone (all from Sigma) were included as described in Ref.33. Structural, functional, and in vivo growth properties of mouse mammary cells grown on plastic have been reported (2, 33-35).
Cytoskeletal Extracts. Cytoskeletal fractions enriched in intermediatefilaments were prepared from the mammary tissues and primary mammary cell cultures according to methods modified from those of Frankeef al. (3,36) as reported elsewhere (30). Briefly, samples were extractedby sequential homogenization in buffers containing low and high saltconcentrations and Triton X-100 in the presence of phenylmethylsulfonyl
fluoride, followed by a series of washes to remove the detergent. Forextracts of tissues, precipitation with trichloroacetic acid was necessaryin order to separate fat from proteins. The presence of filaments with adiameter of 9 to 11 nm in the final pellet of extracted material wasconfirmed by electron microscopy. Two or more separate extracts ofeach type of tissue and cultured cells were analyzed.
Electrophoresis. For 2-D PAGE, samples were solubilized in urea and
dithiothreitol, and polypeptides were separated using IEF (pH range, 4to 7.5) or NEPHGE (pH range, 4 to 8.5) as the first dimension accordingto the procedures of O'Farrell (37, 38) as reported previously (30).
Proteins were visualized with Coomassie Brilliant Blue.
RESULTS
Characteristics of Tissues. We have shown previously thatthe pattern of cytoskeletal polypeptides including keratinschanges very little during the normal developmental cycle of theadult mouse mammary gland (30). For convenience, we thereforeused mammary tissue from mice in mid- to late pregnancy as
our example of normal epithelium. Three HAN transplant linesserved as sources of preneoplastic tissue. These lines were: theD2, which originally developed in a mouse following hormonalstimulation (39); the C4, which arose in an animal fed 7,12-
dimethylbenzanthracene (40); and the CV2, which originated ina mouse infected with mouse mammary tumor virus (41). Theprecancerous nature of these lines is well documented (39-41 ),
and the mammary carcinomas emerged spontaneously from theHAN.
Sections of tissue stained with hematoxylin and eosin wereused to examine and compare the histology of the normal andabnormal mammary epithelium (Fig. 1). By the third week ofgestation, the mammary gland of a normal mouse containsabundant alveoli which project from the ductal tree (Fig. 1a). TheD2 HAN consists of well-differentiated alveoli that are almost
indistinguishable from those found in normal tissue (Fig. 1b). TheC4 and CV2 HAN are similarly composed of numerous alveoli,but these lesions also have areas of squamous metaplasia withaccompanying keratinization (Fig. 1, c and d). The D2 tumorswhich arise from the D2 HAN are well-differentiated adenocarci
nomas (Fig. 1e). The C4 and CV2 neoplasms are also adenocarcinomas but, like their respective HAN precursors, they exhibitsquamous metaplasia and keratinization (Fig. 1, fand g). Tumorswith the latter characteristics have also been designated asadenoacanthomas (40).
Electrophoretic Analysis of Cytoskeletal Fractions fromTissues. To determine the polypeptide composition of cytoskeletal fractions isolated from normal, HAN, and tumor tissues,detergent-high salt extracts were analyzed by 2-D PAGE. The
polypeptide pattern obtained from cytoskeletal extracts of normal mammary tissue is shown in Fig. 2, a and b. For the purpose
CANCER RESEARCH VOL. 45 JUNE 1985
2761
on April 12, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

KERATINS IN MOUSE MAMMARY HYPERPLASIAS AND CARCINOMAS
of this study, only those spots stained most heavily by Coomas-
sie blue will be considered. As reported previously (30), 3 majorcomponents of neutral to basic charge were detected only byNEPHGE: M, 90,000 (NEPHGE 90); M, 70,000 (NEPHGE 70);and Mr 52,000 (NEPHGE 52) (Fig. 2a). Seven major acidic spotswere most clearly resolved when IEF was used as the firstdimension (M,, pi); 70,000, 5.1 (IEF 70); 67,000, 5.6 (IEF 67);59,000, 5.6 (IEF 59); a closely clustered set of 2 polypeptides,47,000 and 50,000, 5.1 and 5.3 (IEF 47-50); 43,000, 5.4 (IEF
43); and 40,000, 5.4 (IEF 40) (Fig. 2u). The latter componentwas difficult to differentiate from actin (IEF 43) in Coomassieblue-stained gels. However, IEF 40 was easily distinguished by
immunoblotting with antikeratin antibodies (30). Of these 10polypeptides, immunoblot analysis using antikeratin antibodiesdemonstrated that IEF 47-50 and IEF 40 were keratins (30).
Two minor constituents, M, 57,000 (NEPHGE 57) and 55,000(IEF 55), were also identified as keratins (30). IEF 59 appears tobe a mitochondrial-associated protein. When this componentwas cut from 2-dimensional gels and used to immunize a rabbit,
the resulting antiserum specifically stained mitochondria in cultured cells and reacted only with IEF 59 in immunoblot experiments.4 (For additional discussion about this polypeptide, see
Ref. 30.) The identities of the other major proteins, includingpossible keratins, have not yet been determined.
Analysis of extracts from HAN and tumor tissues revealedpolypeptide profiles similar to that of normal mammary gland butwith notable differences (Figs. 2 and 3). Samples of the D2 HANtissue contained 3 additions (Fig. 2, c and d). A M, 46,000component (IEF 46), isoelectric with actin, was prominent in IEFgels. This polypeptide is probably a keratin because its electro-
phoretic coordinates correspond very closely to those of keratin20 in Schiller's numbering system for mouse keratins (42). How
ever, while antikeratin antibodies reacted strongly with the moreacidic isoelectric variants of IEF 46, they only outlined the majorspot of this series (30). Thus, its confirmation as a keratin is stilllacking. We are currently preparing an antiserum against IEF 46to use in further characterization studies. IEF 46 was not detected in normal tissue from any stage of the developmentalcycle (30). In both IEF and NEPHGE gels, a M, 55,000 polypeptide (IEF 55) was present. As mentioned earlier, a similar moleculewas detectable in normal tissue preparations with the immunoblot technique using antikeratin antibodies (30) but was barelyvisible by Coomassie blue staining (Fig. 2o). A third (M, 58,000)component was best resolved by NEPHGE (Fig. 2c). The patternof polypeptides in cytoskeletal fractions from D2 tumors closelyresembled that of the D2 HAN, including the presence of IEF 46and 55 and NEPHGE 58 (Fig. 2, e and f).
The C4 HAN was also very similar to normal tissue (Fig. 3, aand b). Unlike the D2 HAN, however, IEF 46 was not visible byCoomassie blue staining, and IEF 55 was decreased in abundance. The most striking change seen in C4 HAN tissue wasdemonstrated on NEPHGE gels, wherein NEPHGE 52 appearedto be reduced, while a M, 57,000 component (NEPHGE 57) wasgreatly increased relative to its minor intensity in normal tissue(compare Figs. 2a and 3a). The latter polypeptide has beenidentified previously as a keratin (30). Little or no NEPHGE 58was seen (Fig. 3a). C4 tumors retained the same pattern ofcytoskeletal polypeptides as did the C4 HAN, except that IEF 46
4H. L. Asch and B. B. Asch, unpublished results.
was present, and IEF 55 was more conspicuous in the tumorpreparations (Fig. 3, c and ci). Moreover, NEPHGE 57 wasslightly less pronounced than in the C4 HAN.
The CV2 HAN tissue consistently yielded relatively smallamounts of cytoskeletal material, despite the use of initial quantities of tissue comparable to those used for all other samples.Nonetheless, the pattern resolved sufficiently to determine thatthe CV2 HAN contained a complement of cytoskeletal polypeptides essentially the same as those of the C4 HAN (Fig. 3, e andf), including an increased amount of NEPHGE 57. The corresponding CV2 tumors displayed a similar set of polypeptidesplus IEF 46 and a slight increase in NEPHGE 58 (Fig. 3, g andh).
Vimentin in the cytoskeletal fractions of the various tissuesranged from a very minor component in samples of normal tissueto a major component in some tumor preparations (compareFigs. 2o and 3d). Since immunocytochemistry studies haveshown that vimentin is not detectable in any mammary epithelialcells (2, 27), these differences most probably reflect the variablenumbers of fibroblasts and other stromal cell types present inthe tissues. Studies using cell cultures further support this contention (see below).
Electrophoretic Analysis of Cytoskeletal Fractions fromPrimary Cell Cultures. To determine the effects of growth invitro on expression of cytoskeletal polypeptides by the normaland abnormal mammary epithelium, cells obtained from thevarious tissues were grown in primary culture for 2 to 4 daysand then extracted by the detergent-high salt method. Theresultant 2-D PAGE patterns of cytoskeletal polypeptides were
similar to those of tissues, with certain interesting differences.As we have shown previously (30), cultured cells derived fromnormal mammary tissues expressed substantial amounts of IEF46 and IEF 55 but had marked reductions of the larger (M, >59,000) and more basic polypeptides (Fig. 4, a and b). Also, IEF47-50, which appeared as a broad spot in tissue preparations,
resolved into IEF 47 and IEF 50 in extracts of cultures, and IEF40 was easily recognized beneath actin (Fig. 4b). Other thansome variability in the appearance of IEF 40, the same generalpattern was also seen when cells of preneoplastic and malignantmammary tissues were grown in primary culture (Figs. 4 and 5).One important exception was expression of the high-molecular-
weight basic protein, NEPHGE 57. Although normal and D2 HANand tumor cells had very little of this component (Fig. 4, a, c,and e), it was quite prominent in C4 and CV2 HAN and tumorcells (Fig. 5, a, c, e, and g). Another exception was NEPHGE 56which was not present in tissues but was observed in cellcultures of C4 and CV2 tumors and possibly C4 HAN but notCV2 HAN (Fig. 5, a, c, e, and g). Although NEPHGE 56 migratedin the same vicinity of NEPHGE gels as did IEF 55, the formerpolypeptide was not seen on IEF gels whereas the latter was.
As we have shown recently, the probability that the vimentinobserved in cells cultured from normal tissues originated fromfibroblastic cells was indicated by the selective decrease of thatpolypeptide and its characteristic degradation fragments whenthe cultures were grown in serum-free medium (30). The latter
condition has been shown to reduce fibroblasts in mammarycultures to less than 1% of the cell population (39). Likewise,propagation of HAN and tumor cells in serum-free medium
produced gel patterns in which this intermediate filament proteinwas greatly diminished, whereas none of the other components,
CANCER RESEARCH VOL. 45 JUNE 1985
2762
on April 12, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

KERATINS IN MOUSE MAMMARY HYPERPLASIAS AND CARCINOMAS
including the keratins, were affected (e.g., cf. Fig. 5, f and g).Expression of the various cytoskeletal polypeptides in normal
and abnormal mouse mammary tissues and cultured cells issummarized in Table 1.
DISCUSSION
In the present study, we have compared the polypeptides incytoskeletal fractions of mouse mammary alveolar hyperplasiasand carcinomas with those of normal mammary epithelium. Ourresults demonstrated that, while the polypeptide patterns ofhyperplastic and malignant tissues and cultured cells were verysimilar to that of normal cells and tissues, certain changes inkeratin expression occurred in both of the abnormal states.However, none of the changes appeared to be exclusively relatedto neoplastic progression, and no obvious correlation was foundbetween the altered keratin expression and the mode of induction of the mammary lesions. Thus, although the D2 HAN and alltumors contained IEF 46, a presumptive keratin not found innormal mammary tissue, significant quantities of this componentwere produced by normal mammary cells in primary culture.Moreover, in contrast to the D2 HAN, the C4 and CV2 HANtissues had no detectable amounts of IEF 46, thereby showingthat expression of this polypeptide is not an obligatory accompaniment of the hyperplastic state. These findings also indicatethat the presence of IEF 46 does not correlate with abnormalhistological organization such as that found in the tumors, sincethe histology of the D2 HAN closely resembles the normalmammary gland in late pregnancy. It is also unlikely that IEF 46is linked to increased cell proliferation. Our previous work (30)demonstrated that the latter component is not found in normalmammary tissue during early pregnancy, wherein the proportionof dividing cells is as high as 30%. In addition, Adamson ef al.(43) have shown that HAN which have filled the mammary fatpad (the stage used in our studies) have about the same per
centage of dividing cells as mammary tissue in late pregnancy.Since all of the HAN were maintained by transplantation, the
question arises as to whether any of the keratin changes foundin these hyperplasias and the tumors developing from them weredue to the effects of serial passage in vivo. To examine thispoint, we analyzed cytoskeletal extracts from spontaneous primary mammary carcinomas of BALB/cfC3H mice. The latterneoplasms are derived from primary rather than transplantedHAN (reveiwed in Ref. 31 ). The polypeptide profile obtained wasessentially identical to that of D2 tumors (data not shown),thereby indicating that the altered cytoskeletal patterns of D2,C4, and CV2 HAN and tumors were not a result of the repetitivetransplantation of the HAN. Mammary tumors of BALB/cfC3Hmice also produce mouse mammary tumor virus, which D2tumors do not (reviewed in Ref. 31). The set of cytoskeletalpolypeptides expressed by the mammary cancers therefore appears to be unrelated to the presence or absence of the virus.
An increase in NEPHGE 57 was found in the C4 and CV2 HANand tumors, all of which exhibited areas of squamous metaplasiaand keratinization. Because neither the D2 HAN and tumors nornormal tissue showed evidence of such abnormalities, thechange in NEPHGE 57 may be related to the aberrant differentiation present in the C4 and CV2 lesions. The fact that cellscultured from the C4 and CV2 tissues maintained expression ofNEPHGE 57 in vitro is especially interesting since they areindistinguishable from cells cultured from normal and D2 tissuesboth morphologicaly and in the distribution and organization oftheir keratin filament system as detected by immunocytochem-
istry with polyclonal antikeratin antibodies (data not shown).Squamous metaplasia with keratinization not only occurs in vivoin precancerous and malignant mammary epithelium of mice butalso can be induced in normal mammary glands from mice andhumans during organ culture by exposure to a combination ofA/6,02'-dibutyryl cyclic AMP and prostaglandins (44, 45). Experi
ments to determine if an increase in NEPHGE 57 in the mouse
Table 1Major cytoskeletal polypeptides in mouse mammary tissues and primary cultures
Major polypeptides as defined by Coomassie blue staining, pi (IEF)or resident pH (NEPHGE)valuesare: 5.4 (40); 5.4 (actin, 43); 5.45 (46); 5.1 (47); 5.1 to 5.2 (47 to50); 5.15 (50); 5.6 to 6.3 (55); 5.2 (vimentin,57); 6.4 (58); 5.6 (59); 5.6 (67); 5.1 (70); 7.2 (52);7.1 (57); 7.6 (70); and 7.2 (90).
Acidic Neutral/basic40a Actin 46* 47 47-50*c 50a Vimentin0 59 67 70 52 55a'" 56* 57a 58e 70 90
TissuesNormal (+) ++ - ++ +++++++ (+) - (+) + ++D2HAN + ++ ++ +++++++++- (+) ++ + ++
C4 HAN ++ ++ ++ + (+) (+) + + -++_+ (+)
CV2HAN" ++ ++ + + (+) (+) ++ + - ++ (+) + +CV2 tumor - ++ ++ ++ ++ ++ (+) + ++++-+++ + +
PrimaryculturesNormal ++ ++ ++ ++ •+•++ — — — — ++ — — — — —D2 HAN (+)+++++ + ++ -D2 tumor +++++++ +++--- _++_____C4HAN (+) ++ ++ ++ ++++-- - ++ (+) ++C4 tumor -++++++ ++ - - - - -++ + ++---CV2HAN -+++++ ++ + ___ _++_ + ___CV2 tumor ++++++++ ++ - -++ + ++-" These polypeptides have been shown to be keratins by immunoblotanalysis (9).6 IEF 46 is a presumptive keratin. See text for details.c IEF 47-50 was a single spot in most tissues but resolved into IEF 47 and IEF 50 components in cultures.
The variability in the abundance of vimentin in cell cultures was directly related to the proportion of fibroblasts.8M, 55,000 (55) and M, 58,000 (58) (tissues)and M, 56,000 (56) (cultures)polypeptides appeared in NEPHGEbut not in IEF gels.' -, trace amounts or none detectable; (+), variable,usually low amounts; +, abundant; ++, very abundant.a CV2 HAN tissues were availablein limited amounts; thus, the faint gel profiles were difficult to interpret.
CANCER RESEARCH VOL. 45 JUNE 1985
2763
on April 12, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

KERATINS IN MOUSE MAMMARY HYPERPLASIAS AND CARCINOMAS
tissues or its equivalent in the human tissues and/or any otherkeratin occurs in these situations are in progress.
The lack of NEPHGE 57 in the normal and D2 HAN and tumorcultures does not consitute an absence of basic keratins in thesecells. IEF 55, a major component in all mammary cultures,corresponds to keratin 11 in Schiller's numbering scheme for
mouse keratins (42) and is classified as a member of the basicsubfamily of keratins despite its resolution in a slightly acidic toneutral range of IEF gels. The same is true for the analogouskeratin of the human and bovine species (42). Moreover, ourprevious studies have shown that IEF 55 is recognized by AE3(30), a monoclonal antikeratin antibody which is specific for basickeratins (46). Our results are therefore consistent with the concept of "keratin pairs" recently proposed by Eichner ef al. (46),
which states that epithelial cells must express at least onemember from both the acidic and basic subfamilies of keratinsin order to have filament formation.
Preservation of the overall mammary-specific pattern of keratin
expression in the mouse tumors agrees well with studies onductal carcinomas of the human breast (6,7). However, the latterneoplasms in some cases lost and in other cases gained expression of certain keratins (6, 7). In our study, no loss of keratinwas detected; rather, all of the mouse mammary tumors, whicharose from alveolar hyperplasias, gained expression of a presumptive keratin, IEF 46. Moreover, the 3 types of tumors alsohad increases in another keratin, IEF 55. We have not yetexamined carcinomas developing from mouse mammary ductalhyperplasias, which might present a different profile of keratins.
Results of our study are similar to those of Weiss et al. (19),who analyzed keratin expression in different human epidermaldiseases and found that 2 keratins, M, 48,000 and 56,000, weredetected only in hyperprolif erati ve diseases, including cancers,in vivo. The same 2 components were also found in normalepidermal cells grown in culture. These keratins were thereforeproposed as molecular markers of hyperproliferation in epidermalepithelium (19). In the mouse mammary gland, normal epitheliumundergoes massive proliferation during early pregnancy. However, as mentioned before, we have detected no evidence of IEF46 in the gland during this period (30). Thus, IEF 46 may beassociated with some abnormality in addition to, or instead of,hyperproliferation. The similarities in size and charge of thehuman epidermal keratins, M, 48,000 and 56,000, and the mousemammary proteins, M, 46,000 and 55,000, all of which are linkedto abnormal cellular processes, suggest that their significancemay be more than an interesting coincidence.
The set of keratins expressed in the HAN and tumor tissuesreflects the presence of both epithelial and myoepithelial cells.IEF 50 and IEF 40, keratins expressed preferentially by myoepithelial cells in the mouse mammary gland (29), are representedto a similar degree in cytoskeletal preparations from the normaland abnormal tissues. These results support earlier immunoflu-
orescence studies with an antikeratin antiserum that selectivelyrecognized myoepithelial cells in mouse mammary tissues anddemonstrated that this cell type comprises a minor but significantpopulation in HAN and adenocarcinomas (2). The relative proportions of ductal and alveolar cells in the latter tissues cannotbe assessed on the basis of the present data. Our recent workon keratins in normal mouse mammary gland has indicated thatductal and alveolar cells may express the same set of keratinsor that alveolar cells have a subset of those found in ductal cells
(30). However, since histologically the transplanted HAN arecomposed primarily of alveoli, with few if any ducts, alveolar cellsare presumably the predominant cell type in the HAN and theircorresponding tumors.
While numerous studies have examined keratin patterns ofvarious established epithelial cell lines (3, 4, 7, 11, 12, 18), lessinformation is available regarding cells in primary culture, withthe exception of epidermal cells. Cell selection, adaptation, andother events may occur during the course of prolonged culturein vitro and may be accompanied by changes in keratin composition. Indeed, while long-term cell lines usually retain similarities
in keratin profile to their tissue of origin, they can also exhibit avariety of differences (reviewed in Ref. 7). Our data indicate that,with mouse mammary epithelium, HAN and tumor cells in primaryculture have minimal alterations of their in vivo keratin patterns,but normal cells acquire a pattern more like that of their abnormalcounterparts. In fact, normal and D2 tumor cells had essentiallyidentical sets of cytoskeletal polypeptides. Because normalmouse mammary cells grown in primary culture produce normalepithelium when transplanted into syngeneic hosts (34), thechange in their set of polypeptides in culture is not due toacquisition of neoplastia properties. This information provides abasis for analyzing the effects of serial passages in culture onexpression of keratins in cell lines derived from normal andabnormal mammary tissues. Such studies are in progress.
In summary, although changes in the profile of keratins expressed by mouse mammary epithelium are associated withneoplastic progression, none appears to be related solely tohyperplasia and/or cancer. However, such changes may still beuseful in distinguishing abnormal mammary epithelium, especiallyin vivo and when used in combination with other markers. Inaddition, other types of changes in keratin expression may occurduring mammary tumorigenesis which might not be detected bythe methods used in the present study.
ACKNOWLEDGMENTS
We are especially grateful to Dr. Daniel Medina for donating all of the mice usedin this study. Leon Hall III provided excellent technical assistance.
REFERENCES
1. Altmannsberger, M., Osbom, M., Holscher, A., Schauer, A., and Weber, K.The distribution of keratin type intermediate filaments in human breast cancer:an immunohistochemical study. Virchows Arch. Zellpathol., 37: 277-284,1981.
2. Asch, B. B., Burstein, N. A., Vidrien, A., and Sun, T.-T. Identification of mousemammary epithelial cells by immunofluorescence with rabbit and guinea pigantikeratin antisera. Proc. Nati. Acad. Sci. USA, 78: 5643-5647,1981.
3. Franke, W. W., Mayer, D., Schmid, E., Denk, H., and Borenfreund, E. Differences of expression of cytoskeletal proteins in cultured rat hepatocytes andhepatoma cells. Exp. Cell Res., 734: 345-365,1981.
4. Franke, W. W„Schiller, D. J., Moll, R., Winter, S., Schmid, E„Engelbrecht, l.,Denk, H., Krepier, R., and Platzer, B. Diversity of cytokeratins: differentiationspecific expression of cytokeratin polypeptides in epithelial cells and tissues.J. Mol. Biol., 753: 933-959,1981.
5. Krepier, R., Denk, H., Weinen, E., Schmid, E., and Franke, W. W. Keratin-likeproteins in normal and neoplastic cells in human and rat mammary gland asrevealed by ¡mmunofluorescence microscopy. Differentiation, 20: 242-252,1981.
6. Moll, R., Krepier, R., and Franke, W. W. Complex cytokeratin polypeptidepatterns observed in certain human carcinomas. Differentiation, 23: 256-269,
1983.7. Moll, R. W., Franke, W. W., Schiller, D.. L., Geiger, B., and Krepier, R. The
catalog of human cytokeratins: patterns of expression in normal epithelia,tumors, and cultured cells. Cell. 37: 11-24,1982.
8. Nelson, K. G., and Slaga, T. J. Keratin modifications in epidermis, papillomas,
CANCER RESEARCH VOL. 45 JUNE 1985
2764
on April 12, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

KERATINS IN MOUSE MAMMARY HYPERPLASIAS AND CARCINOMAS
and carcinomas during two-stage carcinogenesis in the SENCAR mouse.Cancer Res., 42: 4176-4181, 1982.
9. Winter, H., and Schweizer, J. Carcinoma-specifickeratin polypeptide patternsin keratinizingepitheliaof rodents: independenceof species-andtissue-specificvariations. Carcinogenesis(Lond.), 2: 613-621,1981.
10. Winter, H., Schweizer, J., and Goerttler, K. Keratinsas markers of malignancyin mouse epidermal tumors. Carcinogenesis(Lond.), 1: 391-398,1980.
11. Winter, S., Jarasch, E.-D.,Schmid, E., Franke,W. W., and Denk,H. Differencesin polypeptide composition of cytokeratin filaments, including tonofilaments.from different epithelial tissues and cells. Eur. J. Cell Bid., 22: 371, 1980.
12. Wu, Y. J., and Rheinwald, J. G. A new small (40Kd) keratin filament proteinmade by some cultured human squamous cell carcinomas.Cell, 25:627-635,1981.
13. Bannasch, P., Zerban, H., Schmid, E., and Franke, W. W. Liver tumorsdistinguished by immunofluorescencemicroscopy with antibodies to proteinsof intermediate-sizedfilaments. Proc. Nati. Acad. Sci. USA, 77: 4948-4952,1980.
14. Battifora, H., Sun, T.-T., Bahu, R. M., and Rao, S. The use of antikeratinantiserum as a diagnostic tool: thymoma versus lymphoma. Hum. Pathol., 11:635-641,1980.
15. Gabbiani,G., Kapanci, Y., Barrazone, P., and Franke, W. W. Immunochemicalidentification of intermediate-sizedfilaments in human neoplasticcells: a diagnostic aid for the surgical pathologist. Am. J. Pathol., 104: 206-216,1981.
16. Ramaekers,F. C. S., Puts, J. J. G., Kant, A., Moesker, 0., Jap, P. H. K., andVooijs, G. P. Useof antibodies to intermediatefilaments in the characterizationof human tumors. Cold Spring Harbor Symp. Quant. Biol., 46:331-339,1982.
17. Schlegel, R., Banks-Schlegel, S., McLeod, J. A., and Pinkus, G. S. Immuno-peroxidase localization of keratin in human neoplasms. Am. J. Pathol., 770:41-49, 1980.
18. Summerhayes, I. C., and Chen, L. B. Localization of a M, 52,000 keratin inbasalepithelialcells of the mouse bladder and expression throughout neoplastic progression. Cancer Res., 42: 4098-4109,1982.
19. Weiss, R. A., Eichner, R., and Sun, T.-T. Monoclonal antibody analysis ofkeratin expression in epidermal diseases: a 48- and a 56-kdalton keratin asmolecularmarkers for hyperproliferativekeratinocytes. J. Cell Biol., 98:1397-1406, 1984.
20. Ahmed, A. The myoepithelium in human breast cancer. J. Pathol., 773: 129-135, 1973.
21. Ahmed, A. In: Atlas of the Ultrastructure of Human Breast Diseases, p. 1.Edinburgh:Churchill Livingstone, 1978.
22. Gabbiani,G., Csank-Brassert, J., Schneedberger,J. C., Kapanci,Y., Trenchev,P., and Holborow, E. J. Contractile proteins in human cancer cells. Am. J.Pathol., 83: 456-467, 1976.
23. Hamperl, H. The myothelia (myoepithelialcells). Curr. Top. Pathol., 53: 161-220,1970.
24. Pitelka,D. R. Evaluationof morphologicalmarkers of mammaryepithelialcells.In: C. M. McGrath, M. J. Brennan,and M. A. Rich (eds.),Cell Biology of BreastCancer, pp. 1-15. New York: Academic Press, Inc., 1980.
25. Slemmer, G. L. Interactions of separate types of cells during normal andneoplastic mammary gland growth. J. Invest. Dermatol., 63: 27-47,1974.
26. Slemmer,G. L. Interactions of mammary cells during progression and metastasis of breast neoplasia. In: C. M. McGrath, M. J. Brennan, and M. A. Rich(eds.),Cell Biology of Breast Cancer, pp. 93-143. New York: AcademicPress,Inc., 1980.
27. Franke, W. W., Schmid, E., Freudenstein, C., Appelhans, B., Osbom, M.,Weber, K., and Keenan,T. Intermediatesized filaments of the prekeratin type
in myoepithelialcells. J. Cell Biol., 84: 633-653,1980.28. Debus, E., Weber, K., and Osbom, M. Monoclonalcytokeratin antibodies that
distinguish simple from stratified squamous epithelia: characterization on human tissues. EMBO J., 7:1641-1647,1982.
29. Asch, H. L., and Asch, B. B. Expression of a 50K keratin is characteristic ofmouse mammary myoepithelialcells. Ann. NY Acad. Sci., in press, 1985.
30. Asch, H. L., and Asch, B. B. Expression of keratins and other cytoskeletalproteins in mousemammaryepitheliumduring the normaldevelopmentalcycleand primary culture. Dev. Biol., 707:470-482,1985.
31. Medina, 0. Preneoplasticlesions in mouse mammarytumorigenesis. MethodsCancer Res., 7: 3-53,1973.
32. Asch, B. B., and Medina, D. ConcanavalinA-induced agglutinabilityof normal,preneoplastic,and neoplasticmouse mammarycells. J. Nati. Cancer Inst, 67:1423-1430,1978.
33. Medina,D., and Obom, C. J. Growth of preneoplasticmammaryepithelialcellsin serum-freemedium.Cancer Res., 40: 3982-3987,1980.
34. Asch, B. B., Medina, D., and Brinkley, B. R. Microtubules and actin-containingfilaments of normal, preneoplastic,and neoplastic mouse mammary epithelialcells. Cancer Res., 39: 893-907,1979.
35. Supowit, S. C., Asch, B. B., and Rosen, J. M. Casein gene expression innomai and neoplastic mammary tissue. In: C. M. McGrath, M. J. Brennan,and M. A. Rich (eds.),Cell Biology of Breast Cancer, pp. 247-263. New York:Academic Press, Inc., 1980.
36. Franke, W. W., Denk, H., Holt, R., and Schmid, E. Biochemicaland immuno-logical identificationof cytokeratin proteins in hepatocytes of mammalianlivertissue. Exp. Cell Res., 737: 299-318, 1981.
37. O'Farrell, P. H. High resolution two-dimensionalelectrophoresisof proteins. J.Biol. Chem.,250: 4007-4021,1975.
38. O'Farrell, P. Z., Goodman, H. M., and O'Farrell, P. H. High resolution two-
dimensionalelectrophoresisof basic as well as acidic proteins. Cell, 72:1133-1142,1977.
39. Medina, D., and DeOme, K. B. Effects of various oncogenic agents on tumor-producing capabilitiesof series D BALB/c mammary nodule outgrowth lines.J. Nati. Cancer Inst., 45:353-363,1970.
40. Medina,D. Preneoplasticlesions in murinemammarycancer. CancerRes.,36:2589-2595,1976.
41. Slagte, B. L., Lanford, R. E., Medina, D., and Butel, J. S. Expression ofmammarytumor virus proteins in preneoplasticoutgrowth linesand mammarytumors of BALB/cV mice. Cancer Res., 44: 2155-2162,1984.
42. Schiller, D. L., Franke, W. W., and Geiger, B. A subfamily of relatively largeand basic cytokeratin polypeptides as defined by peptide mapping is represented by one or several polypeptides in epithelial cells. EMBO J., 7: 761-769, 1982.
43. Adamson, R., Banerjee, M. R., and Medina, D. Susceptibility of mammarytumor-virus free BALB/c mouse mammary nodule outgrowth cells in DNAsynthesis to 3-methylcholanthrenetumorigenesis. J. Nati. Cancer Inst., 46:899-907,1971.
44. Schaefer,F.V., Custer, R. P., and Sorof, S. Inductionof abnormaldevelopmentand differentiation in cultured mammary glands by cyclic adenine nucleotideand prostaglandin.Nature (Lond.),286: 807-810,1980.
45. Schaefer, F. V., Custer, R. P., and Sorof, S. Squamous metaplasia in humanbreast culture: induction by cyclic adeninenucleotide and prostaglandins,andinfluenceof menstrualcycle. Cancer Res., 43: 279-286,1983.
46. Eichner, R., Bonitz, P., and Sun, T.-T. Classification of epidermal keratinsaccording to their immunoreactivity,isoelectricpoint, and mode of expression.J. Cell Biol., 98: 1388-1396,1984.
CANCER RESEARCH VOL 45 JUNE 1985
2765
on April 12, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

KERATINS IN MOUSE MAMMARY HYPERPLASIAS AND CARCINOMAS
'«j&'^Bl OSÜÕ
KÃ MiIL
Fig. 1. Sections of BALB/c mouse mammary tissues, a, a large duct (D, ductal lumen) and several alveoli (arrowheads) in the normal gland of a mouse in latepregnancy. £>to d, D2, C4, and CV2 HAN, respectively. Note the well-differentiated appearance of the alveoli in b. e to g, D2, C4, and CV2 adenocarcinomas, respectively.Squamous metaplasia has not been observed in normal or D2 HAN or tumor tissues, but the C4 and CV2 tissues contain multiple areas of squamous metaplasia withaccompanying keratinization (arrows). H & E, x 365.
CANCER RESEARCH VOL. 45 JUNE 1985
2766
on April 12, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
KERATINS IN MOUSE MAMMARY HYPERPLASIAS AND CARCINOMAS
..•.;>••'-'>•
-N E P H G E
. 90O 70en
57-52—•
70
."759
4>- 50
Q¡f.
-N E P H GE
90
70
S
I E F-
r^
59..V
17-50
A«*
.906/
90
46»'47~-50
.7052-
6770
59
46—»
58-55X"C46
V 47~-50\4/0—90-70
6?47-!70
52
67,70
47-50
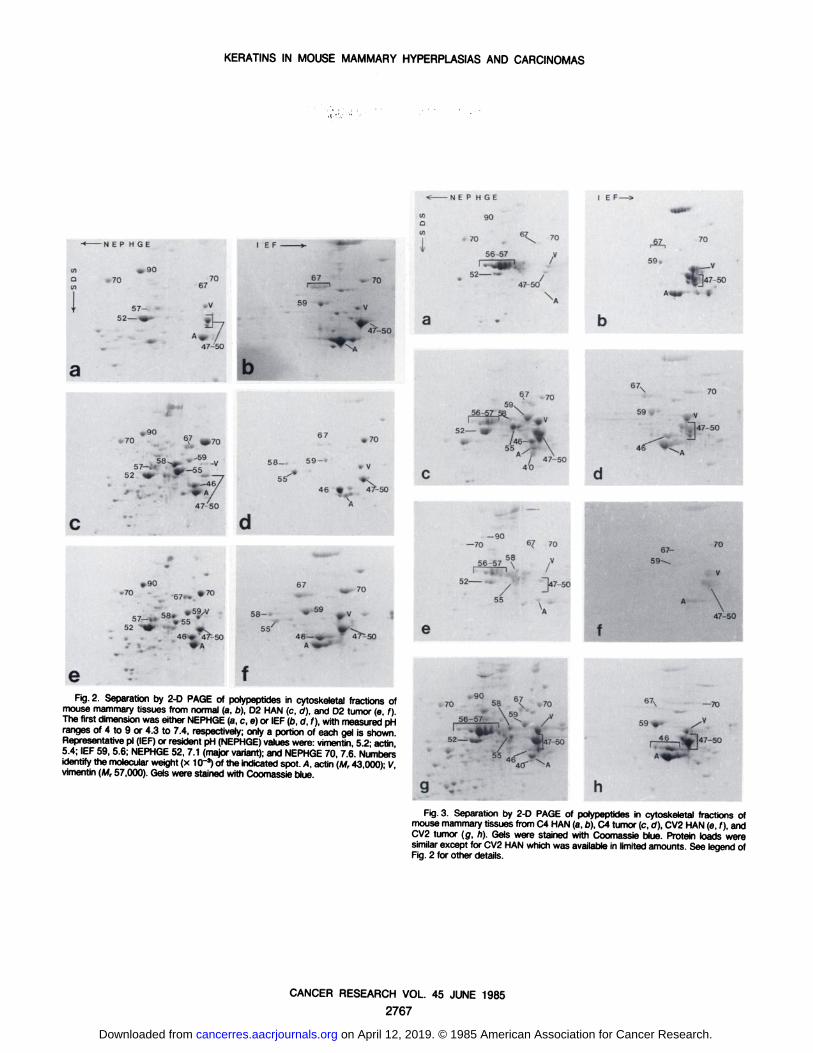
eFig. 2. Separation by 2-D PAGE of polypeptides in cytoskeletal fractions of
mouse mammary tissues from normal (a, b), D2 HAN (c, d), and D2 tumor (e, f).The first dimensionwas either NEPHGE(a, c, e) or IEF(b, d, f), with measuredpHranges of 4 to 9 or 4.3 to 7.4, respectively; only a portion of each gel is shownRepresentativepi (IEF)or resident pH (NEPHGE)valueswere: vimentin, 5.2; actin5.4; IEF 59, 5.6; NEPHGE52, 7.1 (major variant);and NEPHGE70, 7.6. Numbersidentify the molecularweight (x 10'3) of the indicated spot. A, actin (M, 43 000)- Vvimentin (M, 57,000). Gels were stained with Coomassieblue.
59
—70
147-50
Fig.3. Separation by 2-D PAGE of polypeptides in cytoskeletal fractions ofmouse mammarytissues from C4 HAN (a,o), C4 tumor (c, ö),CV2 HAN(e, f), andCV2 tumor (g, h). Gels were stained with Coomassie blue. Protein loads weresimilarexcept for CV2 HAN which was availablein limited amounts. See legend ofFig. 2 for other details.
CANCER RESEARCH VOL. 45 JUNE 1985
2767
on April 12, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

KERATINS IN MOUSE MAMMARY HYPERPLASIAS AND CARCINOMAS
-N E P H G E I EF
-N E P H GE
55 -/"
•
a
b47
56 uL-^l^.-
56-57 «^ y 5
-50—47
46
/
•\47
40
50
5 . TÄ-47
55
-50-47
55
46 e
e fFig. 4. Separation by 2-D PAGE of polypeptides in cytoskeletal fractions of
primary cultures established from mouse mammary tissues. Cells were extracted2 to 5 days after plating, a, b, normal (pregnant) mammary gland cells; c, d, D2HAN cells; e, f, 02 tumor cells. Gels were stained with Coomassie blue. The proteinload was doubled in b in order to demonstrate the absence of any additional majorspots. Arrows, characteristic proteolytic fragments of vimentin. The cells extractedfor c and d were grown in serum-free medium. Note the difference in vimentin (v)in c and d and in a, b, e, and f ; all other components are unchanged. Cells usedfor the latter 4 extracts were grown in medium with serum. Other details are as inFig. 2.
56-57
f gFig. 5. Separation by 2-D PAGE of polypeptides in cytoskeletal fractions of
primary cultures established from mouse mammary epithelium. Cells were harvested 2 to 5 days after plating, a, b, C4 HAN cells; c, d, C4 tumor cells; e, CV2HAN; f, g, CV2 tumor cells. Gels were stained with Coomassie blue. The cellsextracted for d and g were grown in serum-free medium. All other cells were grownin medium with serum. See the legend of Fig 2 for other details.
CANCER RESEARCH VOL. 45 JUNE 1985
2768
on April 12, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1985;45:2760-2768. Cancer Res Harold L. Asch and Bonnie B. Asch Hyperplastic Alveolar Nodules and AdenocarcinomasHeterogeneity of Keratin Expression in Mouse Mammary
Updated version
http://cancerres.aacrjournals.org/content/45/6/2760
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/45/6/2760To request permission to re-use all or part of this article, use this link
on April 12, 2019. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from


















