
Gurdon Lecture Slides
115
The egg and the nucleus: a battle for supremacy J. B. Gurdon Cambridge, England. Stockholm: 7 December, 2012.
-
Upload
le-minh-duc -
Category
Documents
-
view
25 -
download
0
Transcript of Gurdon Lecture Slides

The egg and the nucleus: a battle for supremacy
J. B. Gurdon
Cambridge, England.
Stockholm: 7 December, 2012.

Attack by the egg
Background
Content
Prospects
Defense by the nucleus

The original question Do all cells in the body have the same sets of genes?
Background


Briggs and King, 1952. Proc. Nat. Acd. Sci., USA


Michail Fischberg

1-nucleolus 2-nucleolus


Intestinal Tract of Feeding Tadpole
(GFP-marked))

Partial blastula

Nuclear Transplant Embryo Graft
GFP

Muscle derived from intesti by nuclear transfer and grafts
GFP-muscle derived from intestine nuclei

Efficiency of nuclear reprogramming by nuclear transfer to eggs
Switch between cell-types: e.g. intestinal epithelium to muscle and nerve.
First nuclear transfers……………..……..…15%
Serial nuclear transfers…………..………..….7%
Grafts from nuclear transfer embryos……….8%
30% Total:

Wilmut, Campbell et al 1996 and 1997.

40
30
20
10
% of total nuclear transfers
Stage of donor nuclei
Mammalian nuclear transfers
Xenopus nuclear transplants reaching feeding tadpole stage
Nuclear transfer success decreases as donor cells differentiate
reaching birth
Blastocystt Blastula Blastula Adult Differentiation

Derivation of functional heart from adult monkey skin (Byrne et al., 2008)
Donor of skin
Embryo
Embryonic stem cells
Egg
+
Add factors
Skin cell nucleus
Cloning
Donor of eggs
Stem cell creation
Increase cell number
Differentiation
Beating heart muscle


Epigenetic memory

muscle
Donor tadpole
Muscle cell
Nuclear Blastula
Neurect -oderm
Endoderm
Embryos derived from muscle nuclei remember their origin even in their nerve and endoderm cells
No transcription
High expression
of
56%
52%
genes
Transcription
transfer
Nature Cell Biol. 2006

muscle
Donor tadpole
Muscle cell
Nuclear Blastula
Neurect -oderm
Endoderm
H3.3 is required for epigenetic memory.
No transcription
High expression
of
3%
5%
genes
Transcription
transfer
Elimination by H3.3 mutated from K4 to E4.
H3.3-E4 mutant mRNA
K4, methylatable lysine.
E4, gutamine

EPIGENETIC MEMORY
Can be explained if:
1. H3.3 promotes continuing transcription of active genes, and if
2. Egg cytoplasm reverses gene transcription with a 50% efficiency.

First meiotic prophase oocytes
to analyse the mechanism of nuclear reprogramming

UV
DNA replication
transcription only
Blastula SSomatic cell nucleus
New gene expression New cell types
Frog
Replication errors in Transplanted somatic nuclei
Postzygotic
Single nuclear transfer to eggs in second meiotic metaphase

Incomplete DNA replication
damages somatic nuclei
transplanted to eggs

Days culture O 4
Intense transcription
(no DNA replication)
New gene expression
Germinal vesicle
[Egg progenitor]
Multiple nuclei transferred to growing oocytes in first meiotic prophase
Oocyte
No new cell type

100 days
Oocyte
Egg
Blastula
Oocyte formation prepares the egg for develpment
Germ cell
Transcription G
Lineage selection
Chromatin decondensation Intense gene transcription DNA demethylation

Xenopus oocyte and germinal vesicle
GV
1 mm

.
.
Oocyte Egg Embryo
First meiotic prophase
Second meiotic metaphase
Mid blastula
The oocyte germinal vesicle contents
Matur -ation
contribute to post-fertilization development
Germinal vesicle

Mammalian stem cell genes are rapidly activated in mammalian nuclei transplanted to Xenopus oocytes
1 2 3 4
nuclei
Mouse/human somatic cell
Days
High
Low
thymus Mouse
HeLa Oct4 Nanog
Nuclei of differentiated cells are reprogrammed slowly.
Sox2

Oocyte transcription assay

Living oocyte transcription assay 1. Multiple somatic nuclei in one oocyte.
2. Linear accumulation of new transcripts.
3. Multiple initiations of transcription per gene per day.
4. Oocyte injections show resistance

Transcriptional activation:
attack by egg cytoplasm

Male
Female pronucleus
pronucleus
HFertilized mouse egg Mouse sperm

Mammalian cultured cell nuclei
Just after injection
:

1-2 days after nuclear transfer
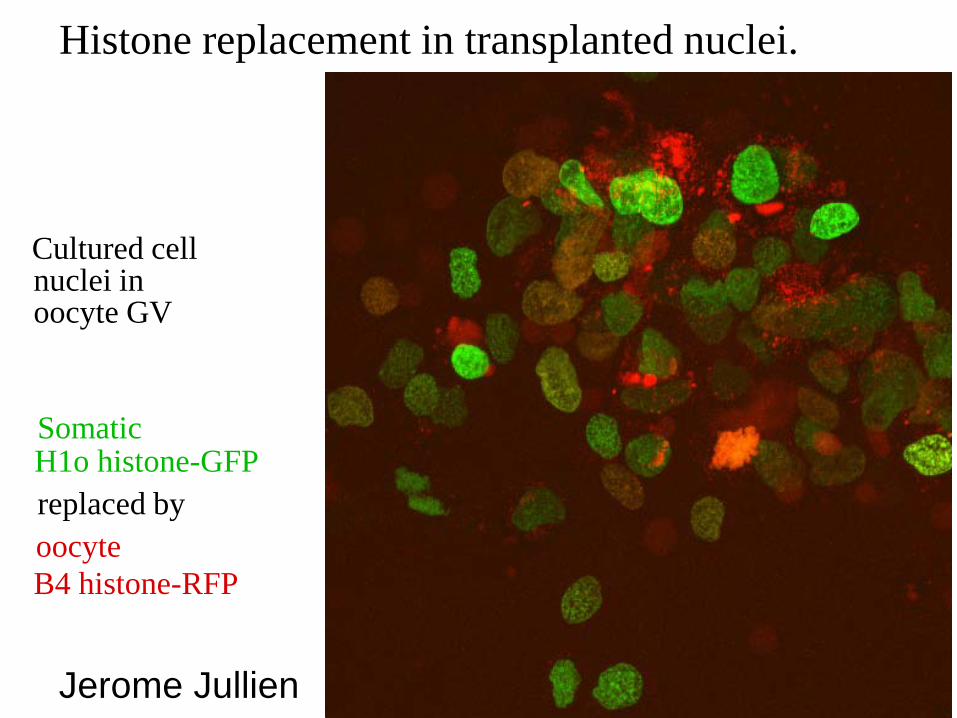
Histone replacement in transplanted nuclei.
Cultured cell nuclei in oocyte GV
H1o histone-GFP replaced by
Somatic
oocyte B4 histone-RFP
Jerome Jullien



Donor H3.3 Oocyte H3.3 Merge
0%
82%
99%
5
10
15
. Histone H3.3 is incorporated into translanted nuclei

Uptake of linker histone and pol II correlates with reprogramming
Linker histone B4 (oocyte origin)
1 hour 24 hours 48 hours
Polymerase II (unphosphorylated)
Polymerase II (serine 5 phosph.)
DAPI

H2B-cherry
YFP-RPB1 (RNA polymerase)
(core histone)
GFP-TBP (TATA-binding protein
of somatic cells)
TBP2 -cherry (TATA-binding protein
of oocytes)
Uptake and loss of chromosomal proteins 0 hour 24 hours
Loss
Loss
Loss
Gain
Gain

Time sequence of polymerase II components binding to genes
Hours since nuclear injection 24 48
% o
f nuc
lei
boun
d
100
50
75
25
0
Histone B4 Pol II ser5P Pol II ser2P
Component bound

2
4
6
Abso
lute
tran
scrip
t lev
el
norm
alize
d to
24
hr sa
mpl
e
0 24 48 Hours after nuclear injection
,
, , with a-amanitin
, normal
Transcriptional reprogramming depends on polymerase II of oocyte origin
inject a-am resistant donor nuclei
(which kills ooc pol II)

Linker histone B4
Polymerase II Ser2
Transcription
Globin TSS Sox 2 TSS High
Low High
Low
High
Low
Reprogramming is selective at the level of polymerase II

Hours after nuclear transfer at 17 C o 48 36 24 12
High
Low
Tran
scrip
tion
3 6
Time course of transcriptional activation of somatic cell nuclei by oocytes
40

Resistance to reprogramming:
defence by the nucleus

Repressed Xi in female mammals

MEF Xi
3
Xi
0
EPI
Xi
0
Xi
3
EPI
100%
50%
Oct
4 tran
script
s rel
ativ
e to
active
X o
n da
y 3
Days
Epiblast-Xi, but not MEF-Xi, genes are strongly reactivated in injected oocytes
MEF

MacroH2A helps to explain resistance to reprogramming
MacroH2A is knocked down by inhibitory RNA,
and induces Oct4 and Sox2 in MEF-Xi cells.

Conclusion
macroH2A marks embryonic
differentiation and acts as an
epigenetic resistance to nuclear
reprogramming
Presenter
Presentation Notes
required to maintain the high stability of the differentiated state Xenopus NT + iPS as tools

Selective gene transcription 48 hours after nuclear transfer to Xenopus oocytes
High
Low
Tra
nscr
ipt
accu
mul
atio
n
Hours since injection 0 48
3368 (21%)
1176 (7%)
3805 (23%)
7890 (49%)
Numbers (%) of genes

Resistance is gene and cell-type specific

Chromatin modification

Nil U16
H3K27me3 H3K9me2/3
Histone H3 Histone H3 Histone H3
Histone modifications in nuclei can be changed after transfer to oocytes
H2AUb
Inject mRNs on day 1. Transplant nuclei on day 2. Reisolate transplanted nuclei on day 3 for Western analysis
Nil K6b Nil K4D
K6b, H3K27me3, H3K27 demethylase.
K4D, H3K9 demethylase.
U116, H2A deubiquitinase.
mRNAs:

DAPI H3K9Me3
No overexpression
Histone modifiers overexpressed in the oocyte efficiently modify transplanted nuclei chromatin
Anti-HA
Overexpressed HA-histone demethylase
methylation
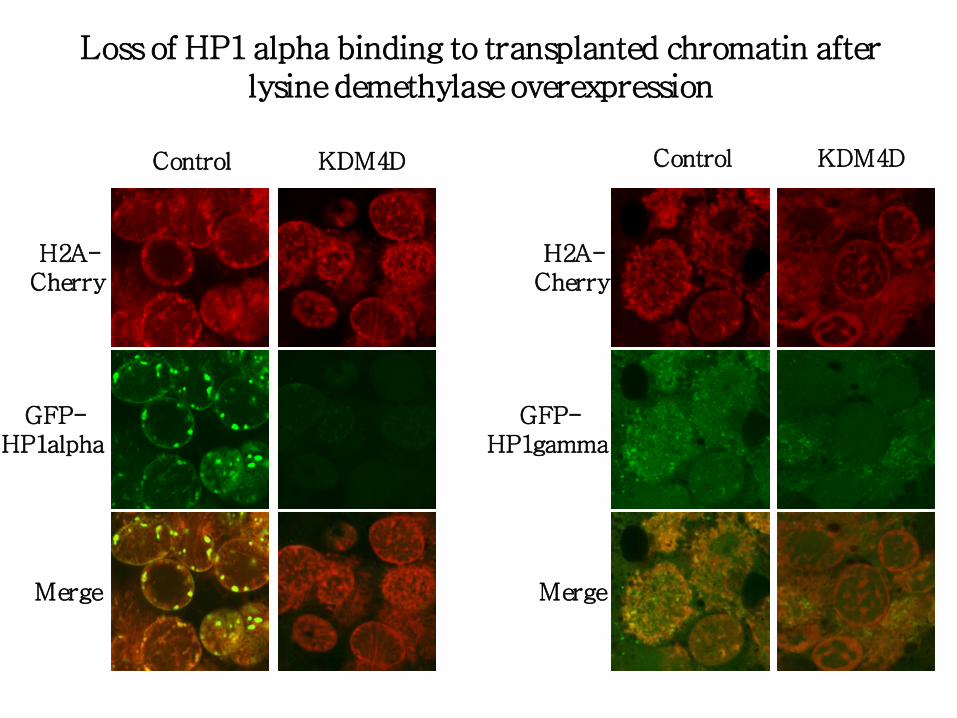
KDM4D Control
H2A- Cherry
GFP- HP1alpha
Merge
Loss of HP1 alpha binding to transplanted chromatin after lysine demethylase overexpression
H2A- Cherry
GFP- HP1gamma
Merge
KDM4D Control

0
2
4
6
8
10
12
14
16
T0 RFP T48 KDM4D T48 USPs T48
Prok 2
Overexpression of histone 2A deubiquitinase removes resistance
Tra
scri
pt le
vel
mRNAs Injected

Chromatin depletion

10
4
5
6
7
8
9
10
10
10
10
10
Log
tra
nscr
ipt co
nten
t
, Oct4 mRNA , luciferase mRNA
.
RNA is removed, replaced, and quantitated by RT-PCR
RNA +RNase +RNase +RNase inhib. +More RNA
Oct
4 an
d lu
cife
rase
43
RNA RNA

1 2 3 4 1 2 3 4 Days
Tra
nscr
ipts
per
gen
e
Normal RNA _
Oct4. Pluripotency. Thymus nuclei
560
1 2 3 4 1 2 3 4 Days
Normal RNA _
Thymus nuclei
1000
Resistance of nuclei transplanted to oocytes RNA depletion from donor nuclei does not affect rate or extent of reprogramming
Sox2. Pluripotency.

1
2
3
4
5
6
7
8
9
10 400 mM NaCl
75 mM NaCl
, Oct4
, c-Jun
R. Halley-Stott
Protein depletion in somatic nuclei removes memory and enhances transcription

P Protein removal from nuclei by salt and Triton

Nil 75mM 200mM 400mM NaCl
S
ox2
tran
scrip
ts
C2C12 ES
HMG 10 10 3 1 EED 10 10 8 0 BMi1 10 10 5 0
HP! 10 10 9 0 Brg1 10 10 6 1
H2A 10 10 10 10
chromosomal proteins
Resistance to reprogramming is maintained at high salt concentrations
Levels of
Pol II 10 10 9 0 after salt treatment

The battle for supremacy
The egg The nucleus
Designed to transform sperm to an embryo
active nucleus
Designed to maintain the same pattern of gene expression
Tries to do the same for somatic nuclei
Tries to resist any change

Prospects
To defeat resistance and win
efficient cell replacement

Gurdon lab
Charles Bradshaw (bioinformatics)
Funding:
Acknowledgements
Jerome Jullien
Rick Halley-Stott
Vincent Pasque
Kei
Miyamoto
Nigel Garrett
Patrick Narbonne

Past colleagues
Ronald Laskey Donald Brown
Laurence Korn Eduardo De Robertis
Marvin Wickens Alan Colman
Ann Clarke
Valerie Moar
Christpher Graham John Knowland
Present colleagues
James Byrne
Jerome Jullien Kei Miyamoto
Rick Halley-Stott
Vincent Pasque
Marta Teperek
Eva Hoermanseder
Stan Wang
Celia Delahay
ACKNOWLEDGEMENTS
MEDICAL RESEARCH COUNCIL WELLCOME TRUST
CANCER RESEARCH CAMPAIGN
Stina Simonsson Carolina Astrand
Doug Melton

END

Sperm cell Embryonic cell Specialised cell
Images from Dr Kei Miyamoto Marta Teperek
99% 35% 1%
% of normal development after nuclear transfer (to a feeding tadpole)
A sperm nucleus is specially designed to yield normal development

Conclusions
2. 2. Stable comitment can be reversed by nuclear transfer to eggs.
3. Nuclei from diferentiated cells show a strong resistance to reprogramming.
4. Resistance is strongly cell-type and gene specific.
5. Resistance depends on histone modifications and on other stable chromosomal components.
.
1. Some cells (endoderm) undergo a very early stable commitment to their lineage pathway.

Nuclear reprogramming
Vincent Pasque (Xi) Richard Halley-Stott (Trn)
Kei Miyamoto Polym. Actin)
Acknowledgements
Jerome Jullien (B4)
39
Kazutaka Murata (Histone mods) Marta Teperek (Sperm)
Other Laboratories G. Crabtree (Stanford)
G. Almouzni (Paris) K. Ohsumi (Nagoya)
K.Shinkai (Kyoto)
Welcome Trust
Welcome Trust
Medical Research Council
Medical Reseacrh Councill

.
Single nuclear transfer to unfertilized eggs
Skin cells
nucleus in egg
Cloned animal
Somatic
Skin
Skin

Oocyte
Multiple somatic nuclei Germinal vesicle FRAP
GV isolation
Fluorescence recovery after photobleaching
Germinal vesicle with
injected nuclei
To determine the exchange rate of a defined protein in transplanted nuclei


1 hour 6 hours 24 hours 48 hours
Time after nuclear transfer
Histone B4
Pol II total
Pol II Ser 2
DAPI
Increase in polymerase II after nuclear transfer

0 24 48 72 Hrs
0 24 48 72 Hrs
0 24 48 72 Hrs
0 24 48 72 Hrs
0 24 48 72 Hrs
2
4 6
2
4 6
2
4 6
2
4 6
2
4 6
H3K4Me2
H3K4Me3 ,
,
B-globin promoter
Oct4 coding region
Sall4 promoter
Sox2RR2 reg. region
Sox2 promoter
Histones in gene control regions are methylated – Chip analysis
Nuclei from retinoic acid treated ES cells
Epigenetics and chromatin, 2010.

MacroH2A helps to explain resistance to reprogramming
MacroH2A is high on MEF-X:i resists reprogramming. but absent from EPI-Xi: is reprogrammed.
MacroH2A is knocked down by inhibitory RNA, and induces Oct4 and Sox2 in MEF-Xi cells

muscle
Donor tadpole
Muscle cell
Nuclear Blastula
Neurect -oderm
Endoderm
Epigenetic memory
No transcription
High expression
of
56%
52%
genes
Transcription
transfer
Nature Cell Biol. 2006

The resistance of MEF Xi nuclei to reprogramming
by oocytes is not explained by
DNA methylation or by
histone H3K27 me

Nature Cell Biol.2007

Nature Cell Biol.2007

WAVE-1 is required for zygotic genome activation and embryonic development
Oocyte
Egg
Embryo development
+ Antisense Wave-1 _
Nucleus
Nucleus
Arrest as gastrulae
Meiotic maturation
(Wiskott-Aldrich syndrome)
(no antisense)
(with antisense)

Histone modifiers overexpression in the oocyte:
- H3K9 demethylase KDM4D efficiently removes H3K9Me2/3 from transplanted nuclei and leads to loss of HP1 alpha
-H2A deubiquitinases (USP16&21) reduce ubiquitinated H2A
level in transplanted nuclei

10
4
5
6
7
8
9
10
10
10
10
10
Log
RN
A c
onte
nt
, endogenous G3PDH
, endogenous Xist
RNA can be depleted from donor nuclei by RNase.
Permeabilized cells, containing RNA, are treated with RNase,
+RNase
of tre
ated
cel
ls
43
then assayed for residual RNA.
No RNase
Rick Halley Stott

Past colleagues
Ronald Laskey
Donald Brown
Laurence Korn
Eduardo De Robertis
Marvin Wickens
Alan Colman
Ann Jewkes
Valerie Speight
Christpher Graham
John Knowland
Present colleagues
James Byrne Jerome Jullien
Kei Miyamoto
Rick Halley-Stott
Vincent Pasque
Marta Teperek
Eva Hoermanseder
Stan Wang
Celia
ACKNOWLEDGEMENTS
MEDICAL RESEARCH COUNCIL WELLCOME TRUST
CANCER RESEARCH CAMPAIGN


Nil 75mM 200mM 400mM NaCl
S
ox2
tran
script
s
C2C12 ES
HMG 10 10 3 1
EED 10 10 8 0
BMi1 10 10 5 0
HP! 10 10 9 0
Brg1 10 10 6 1
H2A 10 10 10 10
chromosomal
proteins
Resistance to reprogramming is maintained at high salt concentrations
Levels of
Pol II 10 10 9 0 after salt
treatment

Newt chromosome II.
Amphibian lampbrush chromosomes

Oogenesis and development in the mouse
Meiotic
Mouse 17
days days 10
Germ cell
divisions
Mature oocyte Zygote
Muscular response
Oogenesis Development

Major events in nuclear reprogramming
Chromatin decondensation
New (pluripotency) gene expression
DNA demethylation
DNA replication, cell proliferation
Repression of unwanted genes
(lineage selection)
Amphibia
Oocyte in meiotic prophase
Eggs and embryos
DNA demethylation

Stem cell genes are rapidly activated in mammalian nuclei transplanted to Xenopus oocytes
1 2 3 4
nuclei
Mouse/human somatic cell
Days
High
Low
Thymus
Mouse
ES nuclei
Oct4 Nanog
Nuclei of most differentiated cells resist reprogramming.
Sox2
nuclei Differentiated
(resistant)

Resistance to reprogramming is pronounced when comparing different donor cell-types. [by up to 50X]
Time following nuclear transplantation (hrs)
480 400 320 240
160 80
0
35
28
24
12
4
3 24 48 72 96 3 24 48 72 96
Sox2 Oct4
C2C12 MEF C2C12
MEF

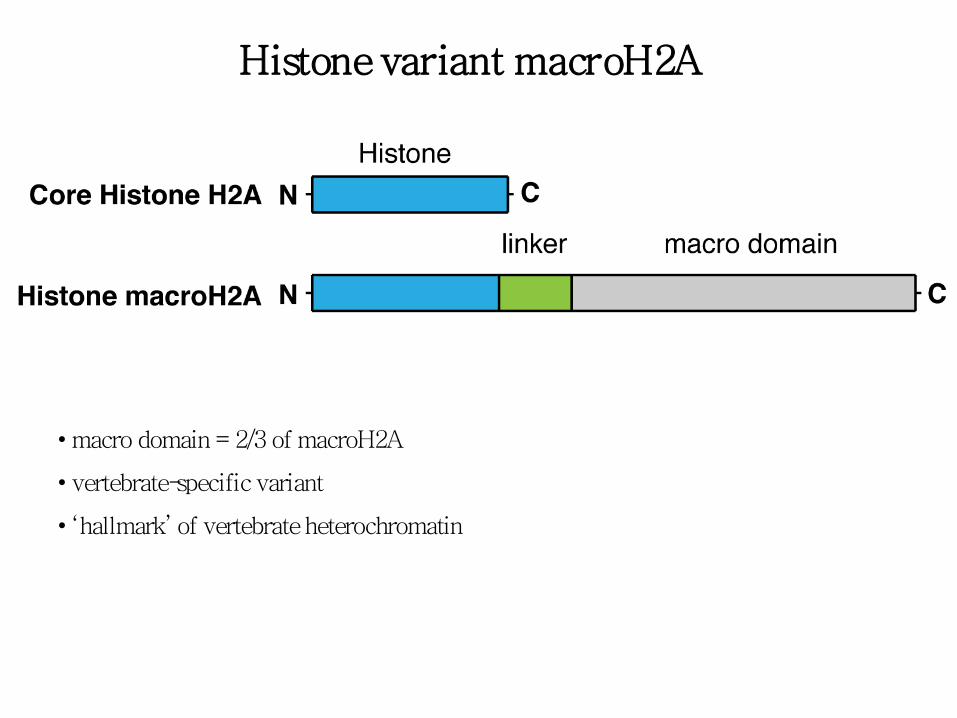
Histone variant macroH2A
• macro domain = 2/3 of macroH2A
• vertebrate-specific variant
• ‘hallmark’ of vertebrate heterochromatin

0
25
50
ES 48h MEF 48h
Prok2 (RIP)
0
1
2
3
4
ES 48h MEF 48h
Ooep (RIP)
0
50
100
150
ES 48h MEF 48h
Gadd45 a (RIP)
0 0,5
1 1,5
2 2,5
ES 48h MEF 48h
GAPDH (RIP)
Examples of genes with restricted expression in MEF nuclei after transplantation to Xenopus oocytes

0 24 48 72 Hrs
0 24 48 72 Hrs
0 24 48 72 Hrs
0 24 48 72 Hrs
0 24 48 72 Hrs
2
4 6
2
4 6
2
4 6
2
4 6
2
4 6
H3K4Me2
H3K4Me3 ,
,
B-globin promoter
Oct4 coding region
Sall4 promoter
Sox2RR2 reg. region
Sox2 promoter
Histones in gene control regions are methylated – Chip analysis
Nuclei from retinoic acid treated ES cells
Epigenetics and chromatin, 2010.

Gene activation in somatic nuclei transplanted to oocytes is selective
Expresssed in MEFs, But NOT in transplanted MEF nuclei,
Number %
NOT expressed in MEFs, BUT in transplanted MEF nuclei
Expressed in MEFs and in transplanted MEF nuclei ,
Repressed 7113 41
Activated 1176 9
No change 3308 29
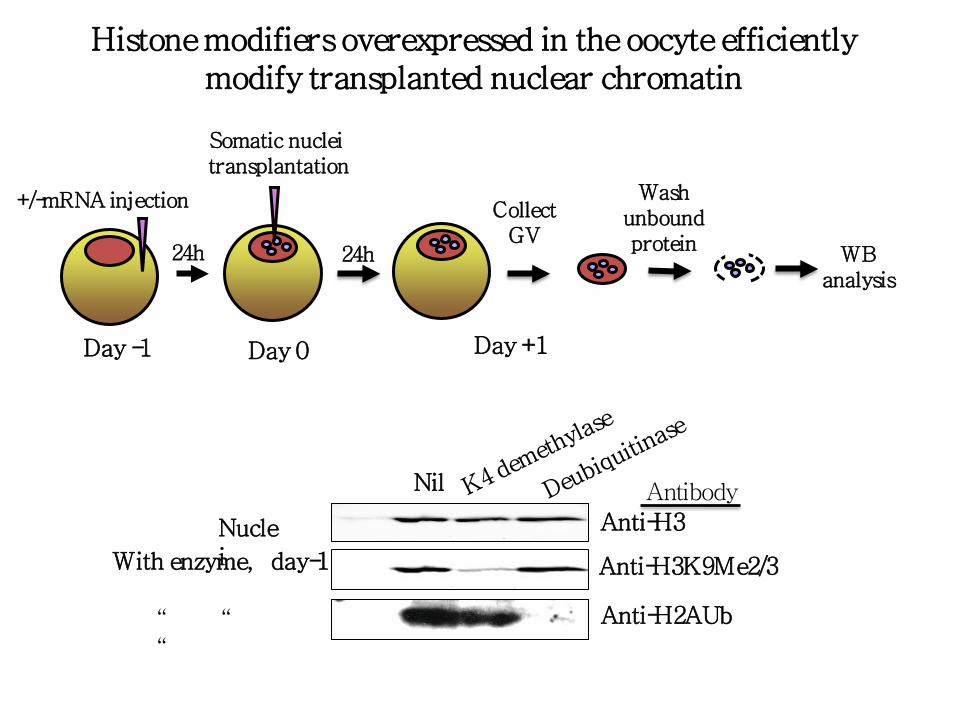
+/-mRNA injection
Anti-H3K9Me2/3
Anti-H3
Anti-H2AUb
Histone modifiers overexpressed in the oocyte efficiently modify transplanted nuclear chromatin
Somatic nuclei transplantation
24h 24h
Collect GV
Wash unbound protein
WB analysis
Day -1 Day 0 Day +1
Nil
With enzyme, day-1
Nuclei
“ “ “
Antibody

Chromatin modifiers that alter the epigenetic state of transplanted nuclei

Acknowledgements
39
Welcome Trust
Welcome Trust Medical Research Council
Vincent Pasque (Xi) Belgium Richard Halley-Stott (Resistance) S. Africa
Kei Miyamoto (Polymerized actin) Japan
Jerome Jullien (Histone B4, H3.3) France
Kazutaka Murata (Histone mods) Japan
Marta Teperek (Sperm progenitors) Poland
Stan Wang USA
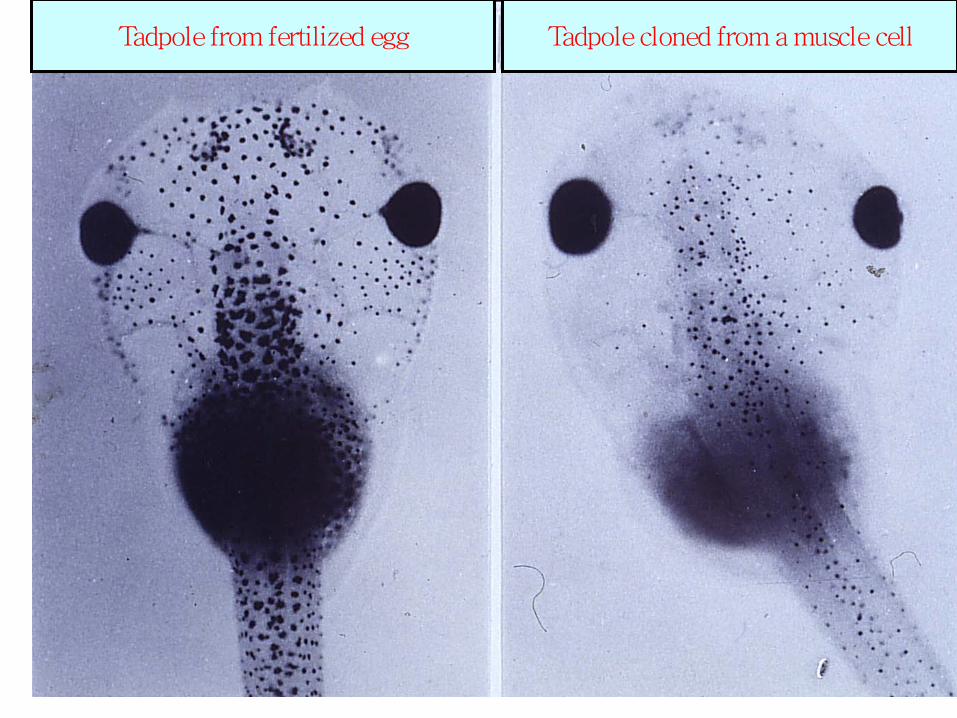
Tadpole cloned from a muscle cell Tadpole from fertilized egg

derived from transplanted muscle cell nucleus
Normal eye
Lens
Retinal layers
Pigmented iris
Normal eye
by cloning from a muscle cell


DNA replication is retarded in Amphibian somatic cell nuclear transfers
30 60 90 120 150
Mins after fertilization or nuclear transfer
replic-
DNA
ation
comp-
lete
100%
0%
Nuclear transfers
with chromosomal damage
Fertilized
eggs
oooo
Mitosis
DNA DNA

Sox
2 tr
ansc
ription
2
1
rela
tive
to
contr
ol a
t 24
hou
rs
B4 histone is required for gene activation in oocytes
, Sox2 transcripts , c-jun transcripts
B4 Dom. Neg. 24 hrs
B4 antibody
24 hrs Control
24 hrs 0 hrs
Sox Jun Sox Jun Sox Jun Sox Jun
Jerome Jullien

5
4
3
2
1 Indu
ced
tran
scrip
t lev
el
Transcription is enhanced by actin polymerization
K. Miyamoto. Gen.Devel.2011.
Oct
4/GA
PDH

A model of nuclear actin function

Transcriptional activation is much enhanced by WAVE-1
Toca-1
WAVE-1
Rac1
N-WASP Enhances reprogramming seen by pluripotency gene transcription in oocytes
No effect on Reprogramming
Actin polymerization
(Wiskott-Aldrich syndrome)

WAVE-1 is required for zygotic genome activation and embryonic development
Oocyte
Egg
Embryo development
+ Antisense Wave-1 _
Nucleus
Nucleus
Arrest as gastrulae
Meiotic maturation
(Wiskott-Aldrich syndrome)
(no antisense)
(with antisense)

Genes with restricted expression in MEF or ES nuclei after transplantation to Xenopus oocytes
(41%) 2123 2864 2048 NOT
expressed in ES after NT.
NOT expressed in MEF nuclei after NT.
Expressed in ES nuclei after NT.
Expressed in MEF nuclei after NT.
Expressed in MEF and ES
(40%) (31%)
(29%)

Histone modifications in nuclei can be changed after transfer to oocytes
Nil K6b Nil U16
H3K27me3 H3K9me3/2 H2AUb
Histone H3 Histone H3 Histone H3
Nil K4D Overexpressed mRNAs
0 hour: mRNA injections. 24 hours: nuclear injections. 48 hours:reisolation of injected nuclei and Western analysis.
K6b, H3K27 demethylase. K4D, H3K9 demethylase. U16, H2A deubiquitinase. Western blots to show loss of histone modifications 48 hours after mRNA injection.

Overexpression of H2A deubiquitinases removes restriction
0
5
10
15
T0 RFP T48 KDM4D T48 USPs T48
prok2
0
0,5
1
1,5
T0 RFP T48 KDM4D T48 USPs T48
ooep
0
0,2
0,4
0,6
0,8
1
1,2
1,4
T0 RFP T48 KDM4D T48 USPs T48
sox2
0
0,5
1
1,5
2
T0 RFP T48 KDM4D T48 USPs T48
Jun
0
5
10
15
T0 RFP T48 KDM4D T48 USPs T48
prok2

Transcriptional reprogramming

Eggs and oocytes have a very high content of histone H3.3.
Histone H3.3 prolongs transcription of somatic nuclei in oocytes.
CONCLUSIONS


















