
Growth of Streptococcusfaecalis var. liquefaciens · Recovery of Streptococcus faecalis, S. lactis,...
4
Growth of Streptococcus faecalis var. liquefaciens on Plants' J. ORVIN MUNDT, J. H. COGGIN, JR.,2 AND L. F. JOHNSON Departments of Bacteriology and Food Technology, and Plant Pathology, University of Tennessee, Knoxville, Tennessee Received for publication Jutne 4, 1962 ABSTRACT MUNDT, J. ORVIN (University of Tennessee, Knoxville), J. H. COGGIN, JR., AND L. F. JOHNSON. Growth of Strep- tococcus faecalis var. liquefaciens on plants. Appl. Micro- biol. 10:552-555. 1962-The proliferation of Streptococcus faecalis var. liquefaciens on two varieties of beans, and on corn, rye, and cabbage was investigated. Comparisons were made with growth patterns on these same plants ex- hibited by S. lactis and Lactobacillus plantarum. The ability of each of the bacteria to multiply and to spread to new plant parts as they developed from seed was studied under several environmental conditions. Plants were grown aseptically in glass culture and in sterilized and non- sterilized soil in the greenhouse. Quantitative estimations of increase in bacterial numbers were made. S. faecalis established commensal growth on each of five plants, al- though selectivity was noted for some plant parts. The organism increased in numbers on the plants equally as well as did the control bacteria, both alone, and in com- petition with the control bacteria and the microflora of the soil. Opposing views have been expressed to account for the widespread occurrence of enterococci on plants. Ostrolenk and Hunter (1946) considered their presence more an indi- cation of survival than of reproduction; Sherman (1937), however, considered as evidence against accidental occur- rence and survival the fact that, of the many cultures iso- lated from plants, none of the hemolytic types of en- terococci had been obtained. Streptococcus faecalis has an appreciable survival in nature (Mallmann and Litsky, 1951; Bartley and Slanetz, 1960). There is, however, increasing evidence for the adaptation of this organism and its variants to growth on plants. It is present in silage (Mieth, 1962) and prominent in the early stages of some silage fermentations (Langston and Bouma, 1960). Widespread occurrence on plants with- out concomitant coliform organisms and within the young seeds of corn and beans (Mundt, Johnson, and Khatchi- kian, 1958), in seeds of corn and peas (Fitzgerald, 1947), and in substantial numbers on corn flowers (Mundt, 1961) ash been reported. Occurrence in seed or soil, movement from the seed to 1 Published with the approval of the Director, Tennessee Arigcultural Experiment Station. 2 Present address: Southern Research Institute, Birmingham, Ala. the emergent parts, and ability to reproduce on the growing plant are criteria of bacterial plant epiphytes (Burri, 1903; Dueggeli, 1904; Vosnyakovskaya and Khudyakov, 1960; for early review see Thayssen and Bunker, 1930). The present studies were designed to supplement the present fragmentary knowledge of the enterococcal-plant associ- ation and to determine whether S. faecalis is a potential epiphyte on plants. MATERIALS AND METHODS Three experimental approaches were used to study the ability of S. faecalis to proliferate on plants. The first was designed to determine the behavior of the test organism and the control bacteria on germinating seeds, and the abilitv of the bacteria to spread with the newly emerged plant parts. The second involved experiments with plants grown from surface-sterilized, reinoculated seeds in steri- lized soil within sterile glass containers. The third involved growing plants from sterilized, reinoculated seeds in steri- lized and in nonsterile soil in the greenhouse. The test organism was a culture of S. faecalis, which pro- duced a soft curd in milk and which was originally iso- lated from the feces of a black bear. It was cultivated routinely in Azide Dextrose (AD) Broth (Difco) and on the KF medium of Kenner, Clark, and Kabler (1961). Two microorganisms, generally considered to be epi- phytic on plants, were used as controls. A local isolate of S. lactis was cultivated routinely in milk, and recovery and quantitative studies were made with Tryptone Glucose Extract (TGE) Agar (Difco) on which the organism pro- duced typically white, opaque, smooth colonies. Lacto- bacillus plantarum (ATCC 8962, strain 250Y) was culti- vated in AD Broth, and recovery and quantitative studies were made on fresh Tomato Juice (TJ) Agar (Difco) on which the organism produced smooth, button-type colonies. This organism is widely distributed in nature on a variety of garden crops (Pederson, 1936) and on orange blossoms and fruits (Hays, 1951). In preliminary screening of common garden seeds, five kinds were selected which would survive the procedure of surface sterilization (according to the method of Rovira, 1956) and which would germinate and grow under the en- vironmental conditions of the experiments. These were white and brown bean, corn, rye, and cabbage. Inoculation of seeds. Sterilized seeds were immersed in 24-hr-old liquid cultures of each organism, singly or in a mixtuire of the three cultures, for several minutes. The 552 on November 13, 2018 by guest http://aem.asm.org/ Downloaded from
Transcript of Growth of Streptococcusfaecalis var. liquefaciens · Recovery of Streptococcus faecalis, S. lactis,...

Growth of Streptococcus faecalis var. liquefaciens on Plants'J. ORVIN MUNDT, J. H. COGGIN, JR.,2 AND L. F. JOHNSON
Departments of Bacteriology and Food Technology, and Plant Pathology, University of Tennessee, Knoxville, Tennessee
Received for publication Jutne 4, 1962
ABSTRACT
MUNDT, J. ORVIN (University of Tennessee, Knoxville),J. H. COGGIN, JR., AND L. F. JOHNSON. Growth of Strep-tococcus faecalis var. liquefaciens on plants. Appl. Micro-biol. 10:552-555. 1962-The proliferation of Streptococcusfaecalis var. liquefaciens on two varieties of beans, and oncorn, rye, and cabbage was investigated. Comparisonswere made with growth patterns on these same plants ex-hibited by S. lactis and Lactobacillus plantarum. Theability of each of the bacteria to multiply and to spread tonew plant parts as they developed from seed was studiedunder several environmental conditions. Plants weregrown aseptically in glass culture and in sterilized and non-sterilized soil in the greenhouse. Quantitative estimationsof increase in bacterial numbers were made. S. faecalisestablished commensal growth on each of five plants, al-though selectivity was noted for some plant parts. Theorganism increased in numbers on the plants equally aswell as did the control bacteria, both alone, and in com-petition with the control bacteria and the microflora of thesoil.
Opposing views have been expressed to account for thewidespread occurrence of enterococci on plants. Ostrolenkand Hunter (1946) considered their presence more an indi-cation of survival than of reproduction; Sherman (1937),however, considered as evidence against accidental occur-rence and survival the fact that, of the many cultures iso-lated from plants, none of the hemolytic types of en-terococci had been obtained.
Streptococcus faecalis has an appreciable survival innature (Mallmann and Litsky, 1951; Bartley and Slanetz,1960). There is, however, increasing evidence for theadaptation of this organism and its variants to growth onplants. It is present in silage (Mieth, 1962) and prominentin the early stages of some silage fermentations (Langstonand Bouma, 1960). Widespread occurrence on plants with-out concomitant coliform organisms and within the youngseeds of corn and beans (Mundt, Johnson, and Khatchi-kian, 1958), in seeds of corn and peas (Fitzgerald, 1947),and in substantial numbers on corn flowers (Mundt, 1961)ash been reported.
Occurrence in seed or soil, movement from the seed to1 Published with the approval of the Director, Tennessee
Arigcultural Experiment Station.2 Present address: Southern Research Institute, Birmingham,
Ala.
the emergent parts, and ability to reproduce on the growingplant are criteria of bacterial plant epiphytes (Burri,1903; Dueggeli, 1904; Vosnyakovskaya and Khudyakov,1960; for early review see Thayssen and Bunker, 1930). Thepresent studies were designed to supplement the presentfragmentary knowledge of the enterococcal-plant associ-ation and to determine whether S. faecalis is a potentialepiphyte on plants.
MATERIALS AND METHODS
Three experimental approaches were used to study theability of S. faecalis to proliferate on plants. The first wasdesigned to determine the behavior of the test organismand the control bacteria on germinating seeds, and theabilitv of the bacteria to spread with the newly emergedplant parts. The second involved experiments with plantsgrown from surface-sterilized, reinoculated seeds in steri-lized soil within sterile glass containers. The third involvedgrowing plants from sterilized, reinoculated seeds in steri-lized and in nonsterile soil in the greenhouse.The test organism was a culture of S. faecalis, which pro-
duced a soft curd in milk and which was originally iso-lated from the feces of a black bear. It was cultivatedroutinely in Azide Dextrose (AD) Broth (Difco) and on theKF medium of Kenner, Clark, and Kabler (1961).Two microorganisms, generally considered to be epi-
phytic on plants, were used as controls. A local isolate ofS. lactis was cultivated routinely in milk, and recovery andquantitative studies were made with Tryptone GlucoseExtract (TGE) Agar (Difco) on which the organism pro-duced typically white, opaque, smooth colonies. Lacto-bacillus plantarum (ATCC 8962, strain 250Y) was culti-vated in AD Broth, and recovery and quantitative studieswere made on fresh Tomato Juice (TJ) Agar (Difco) onwhich the organism produced smooth, button-typecolonies. This organism is widely distributed in nature on avariety of garden crops (Pederson, 1936) and on orangeblossoms and fruits (Hays, 1951).
In preliminary screening of common garden seeds, fivekinds were selected which would survive the procedure ofsurface sterilization (according to the method of Rovira,1956) and which would germinate and grow under the en-vironmental conditions of the experiments. These werewhite and brown bean, corn, rye, and cabbage.
Inoculation of seeds. Sterilized seeds were immersed in24-hr-old liquid cultures of each organism, singly or in amixtuire of the three cultures, for several minutes. The
552
on Novem
ber 13, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

GROWTH OF S. FAECALIS VAR. LIQUEFACIENS
cultures were diluted to contain 1.8 X 106 viable cells perml. The average quantity of inoculum adsorbed per seedwas estimated to be 0.004 ml for cabbage seed, containing400 to 600 cells; 0.005 ml for rye seed, containing 300 to500 cells; 0.04 ml for white bean seed, containing 3,700 to4,500 cells; 0.05 ml for corn seed, containing 4,500 to 5,700cells; and 0.055 ml for brown bean seed, containing 4,600 to4,800 cells per seed. Uninoculated seeds used as controlswere immersed for a comparable period of time in distilledwater.
Behavior of bacteria on germinating seeds. Sterilized,inoculated, and uninoculated seeds were placed on 2.5 %nonnutrient (NN) agar in petri plates. Dihydrostreptomy-cin (2 mg) and penicillin (1,000 units per ml) were added tothe agar to suppress the microflora of the unsterilized seeds.When 90 % of the control seeds, which were incubated inmoist chambers at 17 to 20 C, had germinated, tips of thehypocotyls and primary roots were excised aseptically, and,together with the seed coats, were cultured in appropriatemedia.
Growth of plants in a gnotobiotic environment. Seedsinoculated with single cultures and with mixtures of thebacteria, and sterilized uninoculated seeds, were placed insterilized greenhouse soil (one-third each of peat moss,field soil, and sand) in sterilized glass jars closed withcotton-gauze pads. After incubation for 96 hr in darkness,the containers were removed to a constant-temperatureroom equipped with "day-brite" fluorescent lamps 3 ftabove the containers. Temperature at the soil level was25 C under light and 19.5 C in darkness at approximately85 % relative humidity.
Growth in the greenhouse. Seeds sterilized and inoculatedas described above were planted in sterilized and in non-sterile soil in pots embedded in sand. None of the threebacteria was recovered from the nonsterile soil or the sandprior to planting the seeds
Recovery of bacteria from plants. Excised primary andsecondary roots, arils, cotyledons, and primary leaves ofall plants, and secondary leaves of beans, were placed intoKF and AD Broths and into Litmus Milk for enrichment.From the liquid cultures, streaks were made on KF Agarfor the recovery and identification of S. faecalis, on TJAgar for recovery and identification of L. plantarum, andon TGE Agar for the recovery and identification of S.lactis. Quantitative estimates of populations were made bymechanical disintegration of whole plants in phosphatebuffer, followed by surface plating of serial dilutions onKF, TJ, and TGE Agars.
RESULTS AND DIscussIoN
Recovery of bacteria from germinating seeds. Bacteriaadded to the seed prior to germination were recovered fromthe aril, primary root, and hypocotyl of each plant, exceptrye (Table 1). S. faecalis and L. plantarum failed to movewith the hypocotyl of the white bean, and S. lactis not onlyfailed to move with the primary root and the hypocotyl of
cabbage but died in a number of instances on the seed coatof this plant. El Shammaa (1962) isolated from rye seedsand germ-free bran a water-soluble, crystallizable, heat-stable antibiotic substance, which is active against manyTABLE 1. Recovery of bacteria from emiergent plant parts of sterilized,
inoculated, and germinated seeds
Per cent recovery (avg)*Plant Inoculumstructure ~~White Brown Cab-
bean bean Rye Corn bage
Seed coat S. faecalis 100a 93a 08 97ab 65aS. lactis 89b 93a 0. 87b 17bL. plantarum 1008a l0a Oa lOOa 75b
Primary S. faecalis 98a 64a O 57b 15broot S. lactis 86b 2b O 68b 9h
L. plantarum 1008 6b Oa 94a 77a
Hypocotyl S. faecalis lb 89ab Oa 87a 50bS. lactis 838 72b 0a 69b OeL. plantarumtt 1B 93a 08 90a 763
* Values for each plant anid structure were analyzed statisti-cally by Duncan's multiple-range test. In each case, the threcvalues being compared are not significantly different from eachother at the 5% level if followed by the same superscript letter.
SEC. LEAr
PRI. LEAF
COTYLEDON
STEM
ARI L
PRI. ROOT
SEC. ROOT
WHITE BROWN RYEBEAN BEAN
S. FAECALS I S. LAGTIS
SEC LEAF
PRI LEAF
COTYLEDON
STEM
ARI L
PRI. ROOT
SEC. ROOT
SOI LL
WHITEBE AN
- S. FAECALIS
I*
......- :
*.........
...........
.R.....
>......
..................
:....:
..:::
..-.........Fi...................... E,S
CABBAGE
X :1 L. PLANTARUM
BROWN R E CORN CABBAGE
BE AN
El-S LACTIS LJ PLANTARUM
FIG. 1. Recovery of Streptococcus faecalis, S. lactis, and Lacto-bacillus plantarum from plant structures and rhizosphere soil ofeach of five plants, grown from surface-sterilized seeds inoculatedwith single cultures (upper) and with a mixture of the bacteria(lower) in sterilized soil in sterilized glass containers. White spacesindicate no recovery. Asterisks indicate structures were not cultured.
19621 553
L---j
on Novem
ber 13, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
MUNDT, COGGIN, AND JOHNSON
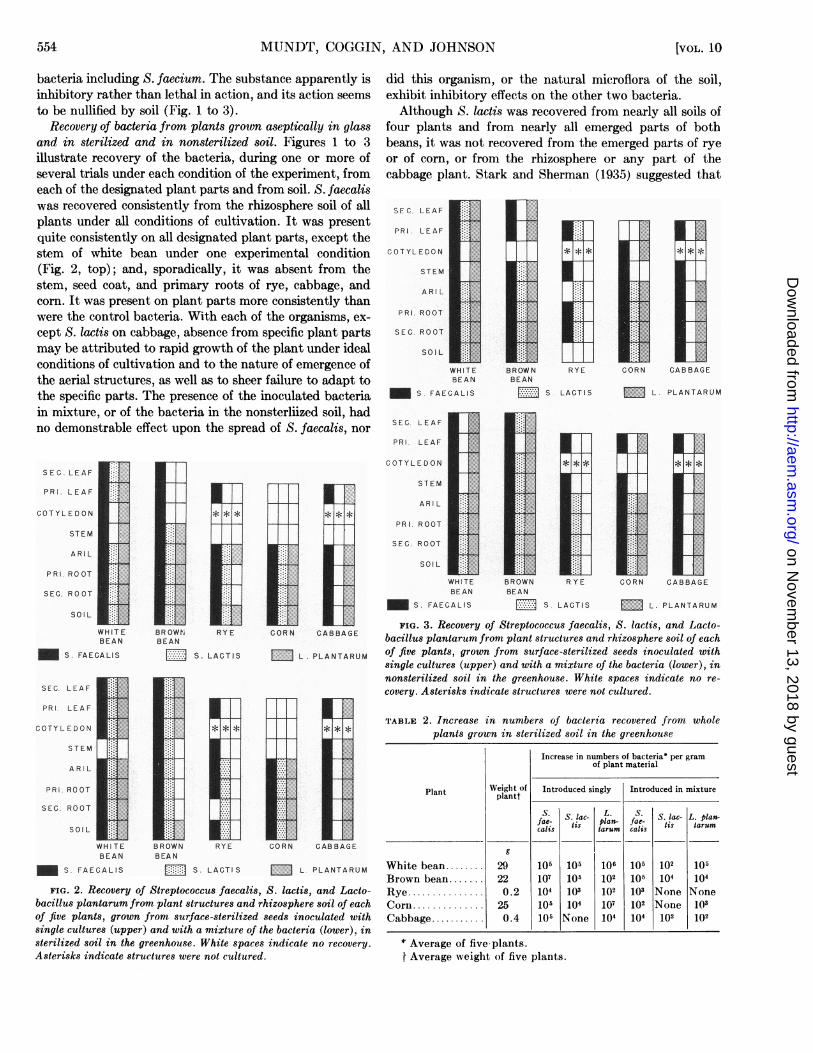
bacteria including S. faecium. The substance apparently isinhibitory rather than lethal in action, and its action seemsto be nullified by soil (Fig. 1 to 3).
Recovery of bacteria from plants groum aseptically in glassand in sterilized and in nonsterilized soil. Figures 1 to 3illustrate recovery of the bacteria, during one or more ofseveral trials under each condition of the experiment, fromeach of the designated plant parts and from soil. S. faecaliswas recovered consistently from the rhizosphere soil of allplants under all conditions of cultivation. It was presentquite consistently on all designated plant parts, except thestem of white bean under one experimental condition(Fig. 2, top); and, sporadically, it was absent from thestem, seed coat, and primary roots of rye, cabbage, andcorn. It was present on plant parts more consistently thanwere the control bacteria. With each of the organisms, ex-
cept S. lactis on cabbage, absence from specific plant partsmay be attributed to rapid growth of the plant under idealconditions of cultivation and to the nature of emergence ofthe aerial structures, as well as to sheer failure to adapt tothe specific parts. The presence of the inoculated bacteriain mixture, or of the bacteria in the nonsterliized soil, hadno demonstrable effect upon the spread of S. faecalis, nor
SEC LEAF
PRI. LEAF
COTYLEDON
STEM
AR.Rorl. - LPRI. ROOT
SEC. ROOT
SOI
WHITE BROWPh RYE CORN CABBAGEBEAN BEAN
S FAECALIS S. LACTIS L.PLANTARUM
SEC LEAF
PR L E A F
COTYLEDON
STEM E
PRI. ROOT
SEC ROOT
WHITE BROWN RYE CORN CABBAGEBEAN BEAN
S FAECALIS m S LACTI S L PLANTARUM
FIG. 2. Recovery of Streptococcus faecalis, S. lactis, and Lacto-bacillus plantarum from plant structures and rhizosphere soil of eachof five plants, grown from surface-sterilized seeds inoculated withsingle cultures (upper) and with a mixture of the bacteria (lower), insterilized soil in the greenhouse. White spaces indicate no recovery.
Asterisks indicate structures were not cultured.
did this organism, or the natural microflora of the soil,exliibit inhibitory effects on the other two bacteria.Although S. lactis was recovered from nearly all soils of
four plants and from nearly all emerged parts of bothbeans, it was not recovered from the emerged parts of rye
or of corn, or from the rhizosphere or any part of thecabbage plant. Stark and Sherman (1935) suggested that
SEC LEAF
PRI LEAFF
COT YL E DON
STEM
A RI L
PRI ROOT
SEC ROOT
SOIL
WHITE BROWNBEAN BEAN
S FAEGALIS S
RYE
LACT IS
*
1e
Mi................
....................
1'''
j
CORN CABBAGE
m L PLANTARUM
SEC LEAF
COTYLEDON
STEM [
RI
PR ROOT
SEC ROOT
SOIL [[
WHITE BROWN RYE CORN CABBAGEBEAN BEAN
S. FAEGALIS S LACTIS C L PLANTARUM
FIG. 3. Recovery of Streptococcus faecalis, S. lactis, and Lacto-bacillus plantarum from plant structures and rhizosphere soil of eachof five plants, grown from surface-sterilized seeds inoculated withsingle cultures (upper) and with a mixture of the bacteria (lower), innonsterilized soil in the greenhouse. White spaces indicate no re-covery. Asterisks indicate structures were not cultured.
TABLE 2. Increase in numbers of bacteria recovered from wholeplants grown in sterilized soil in the greenhouse
Increase in numbers of bacteria* per gramof plant material
Plant Weight of Introduced singly Introduced in mixtureplantt______________________S. S. lac- L. S. S. lac- L. plan-fae- lis plan- fae- ais farumcalis tarum calis
White bean........ 29 105 105 106 105 102 106Brown bean....... 22 107 105 102 105 104 104Rye ............. 00.2 104 103 102 103 None NoneCorin ............. 25 105 104 107 102 None 103Cabbage ........... 0 .4 10- None 104 104 102 102
* Average of five-plants.Average weight of five plants.
554 [VOL. 10
on Novem
ber 13, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

GROWTH OF S. FAECALIS VAR. LIQUEFACIENS
cabbage, among other plants, may be a natural habitatfor this organism. Although L. plantarum was isolated fromthe rhizosphere and from the parts of four plants withnearly the same frequency as was S. faecalis, it was notrecovered from either the roots or stems of rye cultivatedin the greenhouse.The numbers of the three organisms per gram of plant
material, of plants grown from seeds inoculated singly,and with mixtures of the bacteria in sterilized soil in thegreenhouse are shown in Table 2. Data obtained withplants grown in sterilized glass containers and in nonsterilegreenhouse soil are nearly identical with those of the tableand are not included here.
Increase in the number of S. faecalis was substantialwhen compared with the number of cells estimated to havebeen added on the seed. It produced higher populations pergram of plant material when introduced to the seed alonethan when introduced with a mixture of the bacteria. Withbut two exceptions (L. plantarum on white bean and oncorn), its population on plating was at least as great as, andfrequently higher than, the populations of either S. lactisor L. plantarum. In several instances, no increases in cellcounts of the organisms used for comparison were obtainedwhen those obtained from the plant were compared withthose introduced to the seed (counts of S. lactis and L.plantarum on cabbage, rye, and corn plants).When considered with the results of previous studies,
the findings in this study strongly suggest that S. faecalisis a potential epiphyte on plants. It was found consistentlyon most plant parts; it established a cycle on plants inwhich transmission is via the seed; and it reproduced onthe growing plant. Although its population density wasnot excessive when compared with the millions of bacteriaper gram of plant, as reported by Burri (1903) and Dueggeli(1904) and, more recently, by Mundt (1956), it did pro-duce populations comparable with those of the acceptedplant epiphytes, S. lactis and L. plantarum.
Previous studies (Eaves and Mundt, 1960) and concur-rent unpublished work suggest that S. faecalis and itsvariants may be spread with relative ease from animalwastes to plants by insects and possibly by other agents.This report indicates that these bacteria also are capableof adaptation to an environment substantially differentfrom that of the intestinal tract in quantity of nutrients,temperature, and moisture. The fact that they do adaptpresents an interesting problem, namely, the distinctionbetween those bacteria introduced to plants from animalwastes, those which may grow temporarily following suchintroduction, and those which grow in a truly epiphyticrelationship. It is unfortuinate that isolates from plants
bear no presently known marker which will positivelydistinguish these streptococci from those of animal origin.
LITERATURE CITED
BARTLEY, C. H., AND L. W. SLANETZ. 1960. Types and sanitarysignificance of fecal streptococci isolated from feces, sewage,and water. Am. J. Public Health 50:1545-1552.
BURRI, R. 1903. Die Bakterienvegetation auf der Oberfliache nor-mal entwickelter Pflanzen. Zentr. Bakteriol. Parasitenk.,Abt. II 10:756-763.
DUEGGELI, M. 1904. Die Bakterienflora gesunder Samen und darausgezogener Keimpflanzchen. Zentr. Bakteriol. Parasitenk.,Abt. II 13:56-63.
EAVES, G. N., AND J. 0. MUNDT. 1960. Distribution and characteri-zation of streptococci from insects. J. Insect Pathol. 2:289-298.
EL SHAMMAA, Z. A. 1962. Versuche zur Isolierung eines bakterien-hemmenden Faktors aus Roggenkleie. Zentr. Bakteriol.Parasitenk., Abt. I, Orig. 183:527-539.
FITZGERALD, G. A. 1947. Are frozen foods a public health problem?Am. J. Public Health 37:695-701.
HAYS, G. L. 1951. The isolation, cultivation, and identification oforganisms which have caused spoilage in frozen concentratedorange juice. Proc. Florida State Hort. Soc. 64:135-137.
KENNER, B. A., H. F. CLARK, AND P. W. KABLER. Fecal strep-tococci. I. Cultivation and enumeration of streptococci insurface waters. Appl. Microbiol. 9:15-20.
LANGSTON, C. W., AND C. BOUMA. 1960. Types and sequence changeof bacteria in orchardgrass and alfalfa silages. J. Dairy Sci.43:1575-1584.
MALLMANN, W. L., AND W. LITSKY. 1951. Survival of selectedenteric organisms in various types of soil. Am. J. PublicHealth 41:38-44.
MIETH, H. 1962. Untersuchungen ueber das Vorkommen vonEnterokokken bei Tieren und Menschen. III. Die Enterokok-kenflora in den Faeces von Rindern. Zentr. Bakteriol. Para-sitenk., Abt. I, Orig. 185:47-52.
MUNDT, J. 0. 1956. The direct microscopic enumeration of bacteriain the washings of green beans. Food Research 21 :21-26.
MUNDT, J. 0. 1961. Occurrence of enterococci: bud, blossom andsoil studies. Appl. Microbiol. 9:541-544.
MUNDT, J. O., A. H. JOHNSON, AND R. KHATCHIKIAN. 1958. Inci-dence and nature of enterococci on plant materials. Food Re-search 23:186-193.
OSTROLENK, M., AND A. C. HUNTER. 1946. The distribution ofenteric streptococci. J. Bacteriol. 51:735-741.
PEDERSON, C. S. 1936. A study of the species Lactobacillus plan-tarum (Orla-Jensen) Bergey et al. J. Bacteriol. 31:217-224.
RovIRA, A. D. 1956. Plant root excretions in relation to therhizosphere effect. Plant and Soil 7:178-217.
SHERMAN, J. M. 1937. The streptococci. Bacteriol. Rev. 1:3-97.STARK, P., AND J. M. SHERMAN. 1935. Concerning the habitat of
Streptococcus lactis. J. Bacteriol. 30:639-646.THAYSSEN, A. C., AND L. D. BUNKER. 1930. The microbiology of
starch and sugars. Oxford University Press, London.VOZNYAKOVSKAYA, Y. M., AND Y. P. KHUDYAKOV. 1960. Species
composition of the epiphytic microflora. Mikrobiologiya 29:73-76. (English translation)
1962] i--555O
on Novem
ber 13, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from








![Doc1 - CDC...126 Gr. A Streptococci erythromycin 2001 2003 [24] 53% faecalis erythromycin chloram- phenicol ciprofloxacin gentamicin avoparcin ' E. faecalis vancomycin](https://static.fdocuments.in/doc/165x107/6118145c1932226e937f5e05/doc1-cdc-126-gr-a-streptococci-erythromycin-2001-2003-24-53-faecalis-erythromycin.jpg)









