
Growth and accumulation of N, K+, Ca2+ and Mg2+ in barley exposed to various nutrient regimes and...
8
PHYSIOL, PLANT, 67: 159^165. Copenhagoi 1986 Growth and accumulation of N, K^, Ca^^ and Mg^"^ in barley exposed to various nutrient regimes and root/shoot temperatures Paul Jensen and Harald Perby Jensen, P. and Perby, H. 1986. Growth and accumulation of N, K*, Ca-* and Mg^* in bariey exposed to various nutrient regimes and root/shoot temperatures. - Physioi. Piant. 67: 159-165, Six cuitivars of spring barley (Hordeum vulgare L. cvs Saive, Nurnberg ii, Bomi. Rise 1508, Mona and Sv 73 608) were grown in water culture for tiiree weeks with various comhinations of minerai suppiy and differentiai root/shoot temperatures dur- ing tiie growth period. Most important for growtli and accumulation of N, K*, Ca'* and Mg^* was the minerai suppiy, foiiowed by the root temperature and the ciioice of cuitivar. Treatments with iow mineral supply or iow root temperature induced a uni- form reduction in growth and accumulation of the ions studied. The effects of iow minerai supply and low root temperature on growth and N accumuiation was addi- tive, which indicates that these factors exert their infiuence independently of each other. Roots grown at 10°C were smaiier and Rb*(*'Rb) influx was higher than in roots grown at 20°C. It is suggested that the controi of Rb*(''Rb) influx is affected by the root temperature and the age of the plants. The higher *'Rb* ("Rb) influx into the iow temperature roots couid not compensate for the smaiier root size. However, tiie iower totai mineral accumulation made up for the needs of tiie smaiier plants and can- not explain tiie reduction in growtii. Additional key words - Age, aliosteric regulation, cultivar, Rb* influx. P. Jensin artd H. Perby (reprint requests), Dept of Plant Physiology, Univ. of Lund, P.O. Box 7007, S-220 07 Lund, Sweden. Introduction In the laboratory, temperate cereals often encounter considerably higher temperatures than in the field. For convenience, shoots and roots of such plants are aiso kept at the same temperature. Ciarkson (1976) ques- tioned whether resuits obtained with piants cultivated at a high and uniform temperature refiect the physioiogy of fieid-grown plants. Temperate piants adapt to iow soii temperatures, but shoot and root growth decrease (Power et ai. 1967, Mack and Finn 1970, Moraghan and Porter 1975, Cumbus and Nye 1982, Smakman and Hof- stra 3982) and roots become thicker with fewer laterals (Nye and Tinkei 1977, Clarkson and Warner 1979, Cumbus and Nye 1982, Abbas-Ai-Ani and Hay 1983). More realistic growth conditions can be obtained with systems which mantain different shoot and root tem- Received 23 September, 1985; revised 24 January, 1986 Physiol, Planl. 67. 1986 peratures (Cooper et al. 1960, Mack and Barber 1960, Willis et al. 1963, Kemp 1972, Stone and Taylor 1982). Ion uptake by roots adapted to temperatures in the range of 18 to 28°C decreased after transfer to iower temperatures (Carey and Berry 1978, Schimansky 1981). However, ion uptake is usualiy higher in roots adapted to about 8 to ]5°C, compared with roots kept at temperatures around 20°C (Ciarkson et ai. !974, Clark- son 1976, Deane-Drummond and Glass 1983, Siddiqi et ai. 1984). The higher ion uptake is thought to be due to an increased number of carrier entities for active trans- port. V^,, for K+(*'Rb)-uptake in piants grown at 20°C increased after adaptation to 10°C, whiie K^ was about the same or increased slowiy. In roots initially kept at 20°C, elements were fed to the xylem at a higher rate af- ter adaptation to 12°C. With regard to the relationships discussed, it becomes 159
-
Upload
paul-jensen -
Category
Documents
-
view
216 -
download
2
Transcript of Growth and accumulation of N, K+, Ca2+ and Mg2+ in barley exposed to various nutrient regimes and...

PHYSIOL, PLANT, 67: 159^165. Copenhagoi 1986
Growth and accumulation of N, K^, Ca^^ and Mg " in barleyexposed to various nutrient regimes and root/shoot temperatures
Paul Jensen and Harald Perby
Jensen, P. and Perby, H. 1986. Growth and accumulation of N, K*, Ca-* and Mg * inbariey exposed to various nutrient regimes and root/shoot temperatures. - Physioi.Piant. 67: 159-165,
Six cuitivars of spring barley (Hordeum vulgare L. cvs Saive, Nurnberg ii, Bomi.Rise 1508, Mona and Sv 73 608) were grown in water culture for tiiree weeks withvarious comhinations of minerai suppiy and differentiai root/shoot temperatures dur-ing tiie growth period. Most important for growtli and accumulation of N, K*, Ca'*and Mg * was the minerai suppiy, foiiowed by the root temperature and the ciioice ofcuitivar. Treatments with iow mineral supply or iow root temperature induced a uni-form reduction in growth and accumulation of the ions studied. The effects of iowminerai supply and low root temperature on growth and N accumuiation was addi-tive, which indicates that these factors exert their infiuence independently of eachother.Roots grown at 10°C were smaiier and Rb*(*'Rb) influx was higher than in rootsgrown at 20°C. It is suggested that the controi of Rb*(''Rb) influx is affected by theroot temperature and the age of the plants. The higher *'Rb* ("Rb) influx into theiow temperature roots couid not compensate for the smaiier root size. However, tiieiower totai mineral accumulation made up for the needs of tiie smaiier plants and can-not explain tiie reduction in growtii.
Additional key words - Age, aliosteric regulation, cultivar, Rb* influx.
P. Jensin artd H. Perby (reprint requests), Dept of Plant Physiology, Univ. of Lund,P.O. Box 7007, S-220 07 Lund, Sweden.
Introduction
In the laboratory, temperate cereals often encounterconsiderably higher temperatures than in the field. Forconvenience, shoots and roots of such plants are aisokept at the same temperature. Ciarkson (1976) ques-tioned whether resuits obtained with piants cultivated ata high and uniform temperature refiect the physioiogyof fieid-grown plants. Temperate piants adapt to iowsoii temperatures, but shoot and root growth decrease(Power et ai. 1967, Mack and Finn 1970, Moraghan andPorter 1975, Cumbus and Nye 1982, Smakman and Hof-stra 3982) and roots become thicker with fewer laterals(Nye and Tinkei 1977, Clarkson and Warner 1979,Cumbus and Nye 1982, Abbas-Ai-Ani and Hay 1983).More realistic growth conditions can be obtained withsystems which mantain different shoot and root tem-
Received 23 September, 1985; revised 24 January, 1986
Physiol, Planl. 67. 1986
peratures (Cooper et al. 1960, Mack and Barber 1960,Willis et al. 1963, Kemp 1972, Stone and Taylor 1982).
Ion uptake by roots adapted to temperatures in therange of 18 to 28°C decreased after transfer to iowertemperatures (Carey and Berry 1978, Schimansky1981). However, ion uptake is usualiy higher in rootsadapted to about 8 to ]5°C, compared with roots kept attemperatures around 20°C (Ciarkson et ai. !974, Clark-son 1976, Deane-Drummond and Glass 1983, Siddiqi etai. 1984). The higher ion uptake is thought to be due toan increased number of carrier entities for active trans-port. V^,, for K+(*'Rb)-uptake in piants grown at 20°Cincreased after adaptation to 10°C, whiie K^ was aboutthe same or increased slowiy. In roots initially kept at20°C, elements were fed to the xylem at a higher rate af-ter adaptation to 12°C.
With regard to the relationships discussed, it becomes
159

of interest for applied piant science to analyze the re-actions of different cuitivars under conditions that arereasonably close to what occurs in practicai agriculture.In the present investigation the combined effects ofvariations in root/shoot temperature and minerai suppiyon growth, accumuiation of N, K*, Ca-* and Mg * andinfiux of Rb*('*Rb) into the roots are compared forone-, two- and three-week-oid barley piants. Since datafor six cuitivars are pooied in this paper, the influence ofcuitivar differences on variation in growth and minerainutrition wiii be compared with the effects of the growthtemperature and minerai suppiy.
Abbreviations — DNP. dinitrophenoi.
Materials and methods
The piant material inciuded six cuitivars of spring bariey{Hordeum vulgare L ) . Mona, Saive and Bomi havebeen cuitivated in Scandinavia. Sv 73 608 and Ris0 1508are high-lysine iines derived from Mona and Bomi, re-spectively. Niirnberg II originates from a 19th centuryGerman land race.
Germination took piace in darkness at ca 20°C in Pe-tri dishes on fiiter papers moistened with distiiied water.
Fig. i. Arrangement in climate chamber for cultivation ofplants with shoots and roots at different temperatures in the in-terval 10-20°C. A, Cooling unit (Thermofrig), alcohol batiiand temperature regulator; B. polytiiene vessel, 40 L with cir-cuiating distilled water; C. biack-painted glass beaker, 2 1, with1.8 1 nutrient solution (six beakers per vessel); D. outlet; E, in-let; F. connection between vessels.
After three days, the seediings were transferred to blackpiastic discs, 35 mm in diameter, each containing 10piants. Four such pJant groups were grown together in2 i black-painted glass beakers containing 1.8 i nutrientmedium.
Z
-4IA
.0.2
1 0.1
0"o
.- E 20
c « ' 10o «
10
2020
12020
2020
Trftatment
1010
III1510
20
101010
IV1510
2010
* •
14 2 1 14 21 7Age, days
14 21 14 21
Fig. 2. Design of experiment A (top) and fresh weights and total contents of K*, Ca * and Mg'* in shoots (circles) and roots (trian-gles) of bariey (with one plant as a basis) exposed to four different nutrient aad temperature treatments (I-IV) during threeweeks. Temperature regime (T): upper figure represents shoot temperature and lower figure root temperature. Nutrient supply(NS): Double-iined area (treatments! to HI) represents cultivation in fuli strength medium, and singie-iined area (treatment IV)represents cultivation in the same medium diluted ten times. Fresh weights and ion contents are shown as mean values for six culti-vars ±SD (vertical hars). The value for each cultivar is the mean of eight groups with ten plants in each.
160 Physiol. PUnl. 67.

The plants were grown in a complete nutrient me-dium and in 10 and 100 times dilutions of it. The fullstrength medium was composed of 2.0 mM KNO,, 1.0mM Ca(NO,),, 1.0 mM MgSO,, 1.0 mM KH,PO4, 0.5miWNa,HPO4,0.1 mAf Fe-EDTA, 1.5 nA/MnSOj, 0.25|iM ZnSOj, 0.25 fiM CuCI,, 0.04 \xM NajMoO.,, 10 \iMH,BO,; pH was initially 6.1, The nutrient solutions werechanged three times a week and always on the day be-fore harvest. They were continuously aerated. Waterlosses from the beakers were compensated for by dailyadditions of distiiied water.
The plants were piaced in ciimate chambers with con-tinuous iight provided by General Electric F 48 power
groove 17-CVX ilO W fiuorescent iamps, giving an ir-radiance of ca 25 W m - on the shoots. Air temperaturewas kept at 10, 15 or 20°C. For some treatments, thetemperature around the roots (10°C) was regulated bythe equipment shown in Fig. 1. The designs of experi-ments A and B are included in Figs 2 and 3.
In experiment A (Fig. 2), uptake experiments wereperformed 7, 14 and 21 days after the transfer of seed-lings to the nutrient media. For both shoots and rootsthe temperature regime was the same as during the pre-ceeding week. Two plant groups were placed in each 2 Iblack-painted glass beaker containing 1.8 i '*Rb*-ia-beiled nutrient medium with the same composition as
NS
0,2
O 0.1
0 . 6
0 .4
o
E 0 . 2
z E
Treatment
2020
2Q20
2020
1010
VI1510
2010
2020
VII
2 02020
V I I I
JJL J5.10 10 10
14 21 14 21 /
Age, dayB
14 21 14 21
Fig. 3, Design of experiment B (top) and fresh and dry weights, totai contents and concentrations of N in barley exposed to fourdifferent nutrient and temperature treatments (V-VIII) during three weeks. White area represents cultivation in a fuii strengthmedium diluted 100 times. The vaiue for each cuitivar is the mean of four groups with ten plants in each. Otherwise as for Fig, 2.
Physiol, Plant 67. 1986 161

, 0 . 2LL
E " •
'•r- 3 0
i'l~f 20
"oE
M" 10
5^ 10
Treatment (See top of Fig. 2)III IV
I-i'
-I-
Z 0 14 21 1,4 21 7Age, days
14 21 14 21
Fig. 4. Levels of K*, Ca" * and Mg * in bariey exposed to four different nutrient and temperature regimes during three weeks.Otherwise as for Fig. 2.
the fuli strength medium except that the 2 mM KNO,was replaced by 1 mM RbNO,. The specific activity wasca 0.75 MBq i ' for ""Rb*. After 60 min the roots wererinsed for 10 min in 1.8 1 of uniabeiied medium with thesame composition as the radioiabeiled medium. Theroots were blotted between filter papers, and roots andshoots were separated and weighed for fresh weights.Contents of K*, Ca * and Mg" were determined byatomic absorption spectrophotometry after digestion ofthe plant parts in a mixture of concentrated HNO, andHCiOj(2.1 v/v). Radioactivity was determined by liquidscintiliation counting.
In experiment B (Fig. 3), the piants were harvestedafter 7, 14 and 21 days and dried for two days at 70°Cbefore determination of dry weights. N-contents weredetermined titrimetricaUy by an automatic Kjeidahltechnique (Tecator Kjeitec 1003; Hoganas, Sweden),modified for efficient reduction of NOj (98-99%; datanot shown). The digestion procedure can be dividedinto four main steps:
a. Sampies (0.10 to 0.25 g) are soaked and digested in13 ml of a mixture of concentrated H^SO, (1000 ml),concentrated HjPO, (50 ml) and salicylic acid (50 g).
b. Reduction with Zn before digestion.
c. The digestion temperature is raised to 35O°C for 75min, whereafter Hg-catalyst is added.
d. A final digestion at 420°C for 40 min.
Results
General trends
Pooled data for up to three-week-old plants of the sixbarley cultivars are presented. For each combination oftemperature/nutrient treatment, a general pattern wasfound for changes in fresh and dry weight and contentsof K*, Ca", Mg" and N (Figs 2 and 3).
Seedlings grown at 10°C the first week produced dur-ing this period less fresh and dry weight than seedlingsgrown at 20°C (Figs 2 and 3; treatment II and III vs I; VIvs V; Vni vs VII). When both root and shoot tempera-ture increased stepwise on days 7 and 14 (iO-15-2O°C),this effect was later compensated for (II vs I). Whenonly the shoot temperature increased stepwise (10-15-20°C) and the root was kept at KfC, growth and accu-muiation of minerais were restricted (III vs I; VI vs V;VIII vs VII). Similarly, a limited mineral suppiy duringthe whole cultivation period, 21 days, or during part ofit, decreased growth and mineral contents of the plants
162 Physiol. Plam. 67, 1<>86
I I
E Og •- 0,4
Treatmant (See lop of Fig. Z)HI IV
14 21 14 21 7Age, day*
14 21 14 21
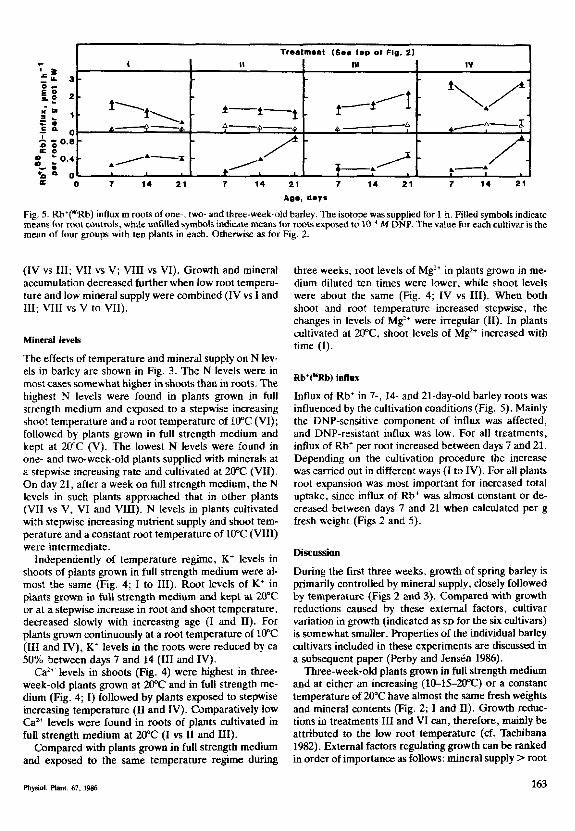
Fig. 5. Rb*(*'Rb) influx in roots of one-, two- and three-week-o!d bariey. The isotope was suppiied for 1 ii. Filled symbois indicatemeans for root controis, whiie unfiiied symbois indicate means for roots exposed to 10" M DNP. The vaiue for each cultivar is themean of four groups with ten piants in each. Otherwise as for Fig. 2.
(IV vs III; VII vs V; VIII vs VI). Growth and mineraiaccumulation decreased further when iow root tempera-ture and iow minerai supply were combined (IV vs I andIII; v m vs V to VII).
Minerai leveis
The effects of temperature and mineral suppiy on N lev-eis in bariey are shown in Fig. 3. The N levels were inmost cases somewhat higher in shoots than in roots. Thehighest N leveis were found in piants grown in ftiiistrength medium and exposed to a stepwise increasingshoot temperature and a root temperature of ICI°C (VI);foiiowed by piants grown in fuii strength medium andkept at 20°C (V). The iowest N ieveis were found inone- and two-week-oid piants suppiied with minerais ata stepwise increasing rate and cuitivated at 20°C (VII).On day 2i, after a week on fuii strength medium, the Nlevels in such piants approached that in other piants(VII vs V, VI and VIII). N leveis in piants cultivatedwith stepwise increasing ntitrient suppiy and shoot tem-perature and a constant root temperature of WC (VIII)were intermediate.
Independently of temperature regime, K* leveis inshoots of piants grown in fuii strength medium were ai-most the same (Fig. 4; I to III). Root ievels of K* inplants grown in fuli strength medium and kept at 20°Cor at a stepwise increase in root and shoot temperature,decreased siowiy with increasing age (I and II). Forpiants grown continuously at a root temperature of 10°C(in and IV), K* levels in the roots were reduced by ca50% between days 7 and 14 (III and IV).
Ca" levels in shoots (Fig. 4) were highest in three-week-old piants grown at 20°C and in fuii strength me-dium (Fig. 4; I) foiiowed by plants exposed to stepwiseincreasing temperature (II and IV). Comparatively lowCa" leveis were found in roots of plants cultivated infuii strength medium at 20°C (I vs II and III).
Compared with plants grown in full strength mediumand exposed to the same temperature regime during
three weeks, root leveis of Mg" in plants grown in me-dium diluted ten times were lower, whiie shoot ievelswere about the same (Fig. 4; IV vs III). When biothshoot and root temperature increased stepwise, thechanges in levels of Mg * were irregular (II). In piantscultivated at 20°C, shoot leveis of Mg * increased withtime (I).
Rb CRb) influx
Infiux of Rb* in 7-, 14- and 21-day-oid bariey roots wasinfiuenced by the cuitivation conditions (Fig. 5). Mainiythe DNP-sensitive component of infiux was affected,and DNP-resistant infiux was iow. For aii treatments,infiux of Rb* per root increased between days 7 and 21.Depending on the cuitivation procedure the increasewas carried out in different ways (I to IV). For ali piantsroot expansion was most important for increased totaiuptake, since influx of Rb* was almost constant or de-creased between days 7 and 21 when calculated per gfresh weight (Figs 2 and 5).
Discussion
During the first three weeks, growth of spring barley isprimarily controlled by minerai supply, closely foiiowedby temperature (Figs 2 and 3). Compared with growthreductions caused by these extemai factors, cuitivarvariation in growth (indicated as SD for the six cultivars)is somewhat smaller. Properties of the individual barleycuitivars included in these experiments are discussed ina subsequent paf)er (Perby and Jensen 1986).
Three-week-old plants grown in fuli strength mediumand at either an increasing (1CI-15-2O°C) or a constanttemperature of 20°C have aimost the same fi-esh weightsand mineral contents (Fig. 2; I and II). Growth reduc-tions in treatments III and VI can, therefore, mainly beattributed to the low root temperature (cf. Tachibana1982). Extemai factors regulating growth can be rankedin order of importance as follows; mineral supply > root
Phyaol. Plant. 67. 1986 163

temperature (10 and 20°C) > cuitivar (Figs 2 and 3).This verifies the resuits of Mack and Finn (1970), whoexposed ciones of timothy to varying fertiiizer, moistureand soii temperature conditions. The totai yield of herb-age was infiuenced iti the order fertiiizer > soii tem-perature (10 and 20°C) > soii moisture > cionai stock.
Increase in dry weight (dry weight at harvest minusdry weight day 0; dry weight on day 0 was ca 35 mg perseedling) for treatments V to VIII can be used to testthe interaction between a moderate root temperaturestress and a restricted mineral supply on growth and ni-trogen accumulation (Fig. 3). Related to treatment V(high temperature and minerai suppiy), values for dryweight accumuiation on day 21 in VI (low root tempera-ture) and VII (initial mineral stress) were 29 ± 1% and23 ± 2%. respectively. If there is no synergistic actionof temperature and mineral stress, the expected vaiuefor their combined effect on dry weight accumulation intreatment VIII will be 7 ± 1%. The real value was 5 ±2%. The difference is insignificant (P>0.1). A similarcaicuJation for N accumuiation gave the same resuit.This indicates that moderate root temperature stressand restricted minerai supply independently depress dryweight production and N accumulation in young barleypiants. Apart from this, iow temperature during the firstweek of cuitivation partly inhibits the development ofstressing N ieveis in one- and two-week-oid seedlings bydepressing growth. Consequentiy the demand for nitro-gen decreases (Fig. 3; VIII vs VII).
Low root temperatures decrease growth of roots andshoots of bariey and other chiiling-toieraat piants (Figs2 and 3; treatment III vs I, VI vs V; Moraghan and Por-ter 1975, Clarkson and Warner 1979, Cumbus and Nye1982, Smakman and Hofstra 1982). Abbas Al-Ani andHay (1983) studied root expansion in four cereal speciescultivated at root temperatures of 5, 15 and 25°C.Among bariey roots of equai iength those growti at theiowest temperature were somewhat thicker than theothers. However, the increase in diameter was smaiiand the authors assumed this to he of minor importancefor minerai nutrition. Tlie temperature also affects themorphoiogy of the root (Nye and Tinker 1977).
Barley roots adapt the fluidity of their membranes tothe ambient temperature, changing the composition ofphosphoiipids (Clarkson et al. 1980). Some ion carriersystems adapt to the root temperature (Clarkson et al.1974, Ciarkson 1976, Deane-Drummond and Giass1983). In our experiments influx of Rb* per root inthree-week-old plants grown in full strength medium ishigher in roots grown at 10°C than in roots grown athigher temperatures (Fig. 5; III vs I and II). Since thereare minor differences in K* levels among the roots ofthese plants (Fig. 4; I to III), differences in aliostericregulation as weii as in number of ion carriers might beinvoived (Ciarkson 1976, Giass 1976, Jensen and Pet-tersson 1978, Siddiqi et al. 1984). Similariy, in perenniairyegrass with roots kept at 10 and 20°C, uptake of NH;,
but not of NO,, adapts to the root temperature (Clar-kson and Warner 1979).
ln the long run, uptake of minerals is depressed if theroots are continuousiy kept at iow temperatures (Figs 2and 3; III vs I, VI vs V). Although the roots adapted to10°C absorb ions at high rates per unit root fresh weight,they are not able to compensate for the difference insize (Figs 2, 3 and 5). The long-term growth depressionis caused by the moderate root temperature stress andnot by lack of minerals. The background for decreasedgrowth may be explained by the growth regulator bal-ance which presumabiy is affected by the root tempera-ture (Ciarkson et al. 1974). In our experiments leveis ofN in one- and two-week-old piants exposed to low roottemperatures were usualiy higher than in piants kept athigh root temperature (Fig. 3; VI vs V, VIII vs VII). Inthe case of Ca'* and Mg", leveis of the eiements variedgreatiy between different temperature treatments(Fig. 4), but in no case was severe stress of Ca'* andMg" induced at low root temperature. K+ ievels, es-pecially in the roots, were low in two- to three-week-oldpiants with roots kept at lO'C (Fig. 4; III and IV). How-ever, they were not low enough to fuily explain growthreduction caused by a moderate temperature stress.This view supports Tachibana (1982) who, when com-paring growth of cucumber varieties at different roottemperatures, claimed that suppression of minerai up-take is not the primary cause of growth reduction at iowroot temperatures. Watts (1972) found that low roottemperature decreased the expansion of corn ieaves byeffects on the ieaf meristematic region.
Jensen (1978) described how DNP-insensitive uptakeof K*('*Rb) and ["Sj-sulphate increased in importanceas wheat grown in water culture aged. This is aiso shownfor one- to three-week-oid piants of bariey grown in fuiistrength medium and kept at 20°C (Fig. 5; I). It meansthat while leveis of K* in the roots are aimost constant,they affect the aiiosteric reguiation system differently atdifferent ages. On the contrary, DNP-sensitive Rb* in-flux is of great importance in three-week-old plantsgrown in full strength medium and exposed to iow roottemperature (Fig. 5; III). Maybe physioiogicai ageingproceeds differentiy as an effect of the cuitivation tech-nique, keeping the active uptake systems intact for aionger time in roots adapted to iow temperatures.
Acknowledgements - The authors thank Mrs Lena Lundii andMiss Kajsa Hedlund for skiifui technicai assistance and DrJanet Bomman for correcting the ianguage. Seeds were kindlyprovided by Svaiof AB, Svaiov, Sweden. The work was sup-ported by grants from the Swedisii Councii for Forestry andAgricuiturai Research and O.E. and Edia Johanssons Founda-tion.
ReferencesAbbas Ai-Ani, M. K. & Hay, R. K. M. 1983. The infiuence of
growing temperature on the growth and morpiioiogy of ce-reai seediing root systems. - J. Exp. Bot. 34: 1720-1730.
164 PhysioL Plant 67. 1986

Carey, R. W. & Berry, J. A, i978. Effects of iow temperatureon respiration and uptake of rubidium ions by excised bar-iey and corn roots. - Piant Physioi. 6i: 858-860.
Ciarkson, D. T. i976. The infiuence of temperature on the ex-udation of xyiem sap from detached root systems of rye (Se-cale cereale) and bariey (Hordeum vulgare). - Planta 132:297-3(M.
- & Warner. A. J, 1979. Relationships between root tem-perature and the transport of ammonium and nitrate ionsby italian and perennial ryegrass (Lolium multiflorum andLolium perenne). - Plant Physiol. 64: 557-56i.
- . Siione. M. G. T. & Wood, A. V. 1974. The effect of pre-treatment temperature on the exudation of xylem sap bydetached barley root systems. - Planta i2i: 8i-92.
- . Hall, K. C. & Roberts. J. K, M. 1980. Phosphoiipid com-position and fatty acid desaturation in the roots of rye dur-ing acciimatization of iow temperature. Positionai anaiysisof fatty acids. - Pianta 149: 464-47 i.
Cooper, D. J,, Neiisen, K. F. Wiiite. J. W. & Kaibfleisch, W.1960. Note on an apparatus for controlling soil tempera-tures. - Can. J. Soil Sci. 40: 105-i07.
Cumbus, 1. P. & Nye, P. H. 1982. Root zone temperature ef-fects on growth and nitrate ahsorbtion in rape {Brassica na-pus cv. Emerald). - J. Exp, Bot. 33: 1138-1146.
Deane-Drummond, C. E. & Giass. A. D, M. 1983. Compen-satory changes in ion fluxes into barley {Hordeum vulgareL. cv, Betzes) seediings in response to differentiai root/shoot growth temperature. - J. Exp. Bot. 34: 17ii-i7i9.
Glass, A. D. M. i976. Reguiation of potassium absorption inbariey roots: An aiiosteric modei. - Piant Piiysioi. 58: 33-37.
Jensen. P. i978. Changes in ion transport in spring wheat dur-ing ontogenesis. - Physioi. Piant. 43: i29-135.
- & Pettersson, S. i978. Aiiosteric regulation of potassiumuptake in plant roots, - Physiol. Plant. 42: 207-213.
Kemp. G, A, 1972. Water bath for germination and root devel-opment studies under various temperature combinations ofsoil and ambient air. - Can. J. Plant Sci. 52: 677-679.
Mack, A, R. & Barber. S. A. i960, A hath for soil tempera-ture control in pot culture work. - Agron. J. 52: 299.
- & Finn, B. J. 1970. Differential response of timothy cionailines and cuitivars to soii temperature, moisture and fertii-ity. - Can. J. Piant Sci. 50: 295-305,
Moraghan. J. T. & Porter, O. A. 1975. Maize growth as af-fected by root temperature and form of nitrogen. - PiantSoii 43: 479-487.
Nye, P H. & Tinker, P. B. 1977. Solute Movement in the Soil-Root System. - Blackwell Scientific Publications, Oxford,pp. 208-209. iSBN 0-632-09730-2.
Perby, H. & Jensen, P. 1986. Variation in growth and accumu-iation of N, K*, Ca-* and Mg-* among barley cuitivars ex-posed to various nutrient regimes and root/siioot tempera-tures. - Physioi. Plant. 67: 166-172.
Power, J. F. Willis, W. O , Grunes, D. L. & Reichman, G. A.1967. Effect of soil temperature, phosphorus, and plant ageon growth analysis of ijarley. - Agroa, J 59: 231-234.
Schimansky, C. i981. Die Aufname von ™Mg, "Rb und =Cadurcii Gerstenpfianzen bei unterschiedlichen Wurzeltem-peraturen. - Z. Pfianzenernaiir. Bodenk, 144: 356-365.
Siddiqi, M. Y., Memon, A. R. & Glass, A. D. M. 1984, Regu-iation of K* infiux in bariey. Effects of low temperature, -Piant Piiysioi. 74: 730-734.
Smakman, G. & Hofstra, J. J. 1982. Energy metaiwiism ofPlantago lanceoloata, as affected by change in root tem-perature, - Physiol. Plant. 56: 33-37.
Stone. J. A. & Taylor. H. M, 1982. A water bath system to ob-serve temperature effects on taproot and lateral root devei-opment of piants. - Soil Sci. Soc. Am. J. 46: i343-1345.
Tachibana, S. 1982. Comparison of effects of root temperatureon tile growth and minerai nutrition of cucumber cultivarsand figleaf gourd, - J. Jpn. Soc. Hortic, Sci. 51: 299-308.
Watts. W. R. 1972. L af extension in Zea mays. II. Leaf exten-sion in response to independent variation of the tempera-ture of tiie apical meristem. ofthe air around the ieaves andof tiie root-zone. - J. Exp. Bot, 23: 713-721,
Wiiiis. W. O., Power. J, F , Reichman, G. A. & Grunes, D. L.1963, Constant temperature water baths for plant growthexperiments. - Agron. J. 55: 200.
Edited by A. Kylin
Physiol. PUnt. 67. 1986 165



















