
Glycocalyx of Bodies Versus Tails Schistosoma … of Bodies Versus Tails of Schistosoma mansoni...
7
THE JOURNAL OF BIOLOGICAL CHEMISTRY !c> 1991 by The American Society for Biochemistry and Molecular Biology, Inc. Val. 266, No. 2, Issue of January 15. pp. 1341-1347, 1991 Printed in U.S.A. Glycocalyx of Bodies Versus Tails of Schistosoma mansoni Cercariae LECTIN-BINDING, SIZE, CHARGE, AND ELECTRON MICROSCOPIC CHARACTERIZATION* (Received for publication, March 26, 1990) Jayasri Nanduri, James E. Dennis$, Terrone L. Rosenberrys, Adel A. F. Mahmoud, and Alan M. Tartakoffn From the Division of Geographic Medicine, Department of Medicine and the Departments of $Biology and §Pharmacology and Ylnstitute of Pathology, Case Western Reserve University and University Hospitals, Cleveland, Ohio 44106 Infection with Schistosoma mansoni is initiated by penetration of the intact skin of the mammalian host with the head but not the tail of the parasite cercariae. The surface of cercariae is covered by a 1-2-micron thick carbohydrate-rich glycocalyx (gx). Furthermore, the transformation of cercariae to schistosomula (the next parasitic stage in the mammalian host) is associ- ated with loss of gx from the bodies. To understand the role of gx in the host-parasite relationship, we have characterized the gx of bothbodies and tails of S. mansoni cercariae. A fluorescent fucose-specific lec- tin-stained bodies and not tails of the organism. More- over, when an enriched preparation of gx obtained by extractionwith 40% aqueousphenoland exclusion from Sepharose CL-GB was subjected to affinity chro- matography on insolubilized Lotus lectin, which binds fucose-containing glycans, only body gx was retained. Body gx is smaller and less negatively charged than tail gx. Electron microscopy showed that gx from bod- ies and tails is composed of 25-40-nm particles and fibrillar material. Carbohydrate composition of gx of bodies and tails indicate that fucose and glucose are major components, respectively. &Elimination exper- iments indicate that the linkage sugar is N-acetylga- lactosamine in both cases. Upon treatment with alka- line borohydride, nearly 90% of gx of both bodies and tails was recovered as two glycan chains: I and I1 (M, -10,500 and 5,600, respectively). Glycan I was in both cases more negatively charged than glycan 11. Fucose is the predominant sugar in glycan I of the bodies while glycan I of tails is mainly composed of glucose. The gx was resistant to several proteases. This resistance and the abundance of carbohydrate in gx may be of biolog- ical importance for survival of cercariae. The substan- tial differences observed between the gx of bodies and tails may provide the basis for understanding the mechanism of selective release of body gx during trans- formation. The existence of a conspicuous carbohydrate-rich glycoca- * This work was supported by the United Nations Development Program/World Bank/World Health Organization Special Program for Research and Training in Tropical Diseases, The McArthur Foundation, and National Institutes of Health Grant AI 15351 and Biomedical Resource Center Program Grant l-P41-RR05351-01, The costs of publication of thisarticle were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisernent” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. lyx (gx)’ at the external surface of many animalcells has been known for more than 20 years (1-3). In parasitic infections, the glycocalyx of the invader may be of importance, since it covers the parasite surfacewhere critical biochemical and physiological interactionswiththehostaretaking place. These interactions often lead to adaptive modificationsin both parasite and host and, more importantly, determine the outcome of successful infection or resistance. Nevertheless, the origin, composition, and functional complexity of the gx of parasites have been incompletely characterized. In certain cestodes and trematodes, light and electron microscopic stud- ies have shown that the gx contains “mucosubstances” and glycoproteins and is anionic (4-7). The present study concerns Schistosoma mansoni, a human trematode parasite,whose surface undergoes radical morpho- logical, physiological, and biochemical changes, as it adapts to fresh water, snail, and mammalian hosts (8,9). The surface of the infective stage to mammalian hosts, cercariae is limited by a lipid bilayer covered with a 1-2-micron thick carbohy- drate-rich gx. This structure is formed late in cercarial devel- opment within the snail, the intermediate host (10-12). Dur- ing the process of infection of mammalian hosts only the bodies of cercariae penetrate the intact skin; the tails are shaken off and do not participate in further parasite-host interactions. Subsequent transformation of cercariae into schistosomulaisassociated with gx release only from the bodies of the organism. The loss of gx exhibits a time course similar to the decrease in the susceptibility of the organism to host effector mechanisms (13). Histochemical staining of intact cercariae of S. mansoni shows that gx contains carbohydrate and polyanions ( l l ) , but reacts in situ with few lectins other than those which are specific for fucose (14). Both scanning and transmission elec- tron microscope studies have shown that the gx is composed of fibrils and particles (15). Limited biochemical information has been obtained by studying gx extracted from intact orga- nisms (15). These studies indicate that the major component has an apparent molecular weight >5 x lo6 under denaturing conditions, an isoelectric point of 5.0, and is 18% amino acid and 82% carbohydrate, with an unusual sugar composition (51% fucose, 30% galactose with small amounts of glucosa- mine, galactosamine, and mannose). Thus little is known about the molecular structure of the gx. Such knowledge is essential for understanding the mechanism of gx release dur- ing transformation and for identifying possible epitopes in- volved in immune recognition. Moreover, the immense molec- ular size of the gx raises questions pertaining to how such ’ The abbreviations used are: gx, glycocalyx; SDS, sodium dodecyl sulfate; FITC, fluorescein isothiocyanate; Me&, tetramethylsilyl; HPLC, high performance liquid chromatography; GC/MS, gas chro- matography/mass spectrometry. 1341
Transcript of Glycocalyx of Bodies Versus Tails Schistosoma … of Bodies Versus Tails of Schistosoma mansoni...

THE J O U R N A L OF BIOLOGICAL CHEMISTRY !c> 1991 by The American Society for Biochemistry and Molecular Biology, Inc.
Val. 266, No. 2, Issue of January 15. pp. 1341-1347, 1991 Printed in U.S.A.
Glycocalyx of Bodies Versus Tails of Schistosoma mansoni Cercariae LECTIN-BINDING, SIZE, CHARGE, AND ELECTRON MICROSCOPIC CHARACTERIZATION*
(Received for publication, March 26, 1990)
Jayasri Nanduri, James E. Dennis$, Terrone L. Rosenberrys, Adel A. F. Mahmoud, and Alan M. Tartakoffn From the Division of Geographic Medicine, Department of Medicine and the Departments of $Biology and §Pharmacology and Ylnstitute of Pathology, Case Western Reserve University and University Hospitals, Cleveland, Ohio 44106
Infection with Schistosoma mansoni is initiated by penetration of the intact skin of the mammalian host with the head but not the tail of the parasite cercariae. The surface of cercariae is covered by a 1-2-micron thick carbohydrate-rich glycocalyx (gx). Furthermore, the transformation of cercariae to schistosomula (the next parasitic stage in the mammalian host) is associ- ated with loss of gx from the bodies. To understand the role of gx in the host-parasite relationship, we have characterized the gx of both bodies and tails of S. mansoni cercariae. A fluorescent fucose-specific lec- tin-stained bodies and not tails of the organism. More- over, when an enriched preparation of gx obtained by extraction with 40% aqueous phenol and exclusion from Sepharose CL-GB was subjected to affinity chro- matography on insolubilized Lotus lectin, which binds fucose-containing glycans, only body gx was retained. Body gx is smaller and less negatively charged than tail gx. Electron microscopy showed that gx from bod- ies and tails is composed of 25-40-nm particles and fibrillar material. Carbohydrate composition of gx of bodies and tails indicate that fucose and glucose are major components, respectively. &Elimination exper- iments indicate that the linkage sugar is N-acetylga- lactosamine in both cases. Upon treatment with alka- line borohydride, nearly 90% of gx of both bodies and tails was recovered as two glycan chains: I and I1 (M, -10,500 and 5,600, respectively). Glycan I was in both cases more negatively charged than glycan 11. Fucose is the predominant sugar in glycan I of the bodies while glycan I of tails is mainly composed of glucose. The gx was resistant to several proteases. This resistance and the abundance of carbohydrate in gx may be of biolog- ical importance for survival of cercariae. The substan- tial differences observed between the gx of bodies and tails may provide the basis for understanding the mechanism of selective release of body gx during trans- formation.
The existence of a conspicuous carbohydrate-rich glycoca-
* This work was supported by the United Nations Development Program/World Bank/World Health Organization Special Program for Research and Training in Tropical Diseases, The McArthur Foundation, and National Institutes of Health Grant AI 15351 and Biomedical Resource Center Program Grant l-P41-RR05351-01, The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisernent” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
lyx (gx)’ at the external surface of many animal cells has been known for more than 20 years (1-3). In parasitic infections, the glycocalyx of the invader may be of importance, since it covers the parasite surface where critical biochemical and physiological interactions with the host are taking place. These interactions often lead to adaptive modifications in both parasite and host and, more importantly, determine the outcome of successful infection or resistance. Nevertheless, the origin, composition, and functional complexity of the gx of parasites have been incompletely characterized. In certain cestodes and trematodes, light and electron microscopic stud- ies have shown that the gx contains “mucosubstances” and glycoproteins and is anionic (4-7).
The present study concerns Schistosoma mansoni, a human trematode parasite, whose surface undergoes radical morpho- logical, physiological, and biochemical changes, as it adapts to fresh water, snail, and mammalian hosts (8,9). The surface of the infective stage to mammalian hosts, cercariae is limited by a lipid bilayer covered with a 1-2-micron thick carbohy- drate-rich gx. This structure is formed late in cercarial devel- opment within the snail, the intermediate host (10-12). Dur- ing the process of infection of mammalian hosts only the bodies of cercariae penetrate the intact skin; the tails are shaken off and do not participate in further parasite-host interactions. Subsequent transformation of cercariae into schistosomula is associated with gx release only from the bodies of the organism. The loss of gx exhibits a time course similar to the decrease in the susceptibility of the organism to host effector mechanisms (13).
Histochemical staining of intact cercariae of S. mansoni shows that gx contains carbohydrate and polyanions ( l l ) , but reacts in situ with few lectins other than those which are specific for fucose (14). Both scanning and transmission elec- tron microscope studies have shown that the gx is composed of fibrils and particles (15). Limited biochemical information has been obtained by studying gx extracted from intact orga- nisms (15). These studies indicate that the major component has an apparent molecular weight >5 x lo6 under denaturing conditions, an isoelectric point of 5.0, and is 18% amino acid and 82% carbohydrate, with an unusual sugar composition (51% fucose, 30% galactose with small amounts of glucosa- mine, galactosamine, and mannose). Thus little is known about the molecular structure of the gx. Such knowledge is essential for understanding the mechanism of gx release dur- ing transformation and for identifying possible epitopes in- volved in immune recognition. Moreover, the immense molec- ular size of the gx raises questions pertaining to how such
’ The abbreviations used are: gx, glycocalyx; SDS, sodium dodecyl sulfate; FITC, fluorescein isothiocyanate; Me&, tetramethylsilyl; HPLC, high performance liquid chromatography; GC/MS, gas chro- matography/mass spectrometry.
1341

1342 S. mansoni Glycocalyx
large molecules can be packaged and transported. The present investigation is centered on the biochemical
characterization of the gx of cercarial bodies and tails. The objectives are to examine differences between the gx of bodies and tails with respect to size, charge, and lectin affinity, to determine the linkage between carbohydrate and the putative backbone, and to characterize well-defined fragments of the intact glycocalyx. The long-term aim is to develop a molecular model of gx.
EXPERIMENTAL PROCEDURES
Materials Lotus lectin coupled to Sepharose, FITC-conjugated Latus, Anguib
anguilla, and Ulex europaeus Iectins were obtained from EY Labs (San Mateo, CA). Sepharose CL-GB, Sephacryl S-1000, Sephacryl S- 200, and DEAE-Sepharose CL-GB were from Pharmacia LKB Bio- technology Inc. NaB3H4 (5-20 Cilmmol) and NalZ5I (100 mCi/ml) were from Amersham Corp. Eserine sulfate, sodium m-periodate, sodium borohydride, neuraminidase (Clostridium perfringens, type V), bovine trypsin, thermolysin, proteinase K, a-amylase, and DNase I were from Sigma. Neuraminidase (Vibro cholerae and Arthobacter urafaciem) and Pronase were from Calbiochem-Boehringer Chemi- cals.
Parasites A Puerto Rican strain of S. mansoni was maintained in Bwmphal-
aria glabruta snails and CF1 mice. Cercariae were obtained by expos- ing infected snails to light for 2 h (16). Cercariae were then immobi- lized with 1 mM eserine sulfate to block transformation and shedding of gx during the labeling procedure (11). The cercarial suspension was concentrated by sedimentation at 600 X g for 2 min at 4 "C. The pellet was washed with spring water and resuspended in spring water with 1 mM eserine sulfate. Cercariae are sheared into bodies and tails by pushing them through 2 syringes connected through a 3-way stopcock (17). Cercarial bodies were then separated from tails on a 60% (w/v) Percoll gradient centrifuged at 600 X g for 10 min at 4 "C (18). The bodies that were in the pellet and the tails that accumulated at the top of the gradient were washed 3 times with spring water to remove Percoll. Examination by light microscopy showed that effi- ciency of separation of bodies and tails achieved by this procedure was around 95%.
Labeling and Extraction of Gr The gx of the cercarial bodies and tails was labeled separately by
periodate-NaB3H4 reduction, a procedure which should introduce label without fragmenting glycan chains (11). Typically lo6 cercariae suspended in 4.5 ml of spring water with 1 mM eserine sulfate were incubated with 5 mM NaI04 for 10 min at 4 "C, washed, incubated with 1 mCi of NaB3H4 (5-20 Ci/mmol) in 5 ml of spring water for 45 min at room temperature and washed extensively. To stabilize the preparation, the reaction is then terminated with an excess (10 mM) of unlabeled NaBH, for 30 min at room temperature and the orga- nisms washed thoroughly. Alternatively eserine sulfate-treated orga- nisms were iodinated with Iodo-Beads (Pierce Chemical CO.) (19). Briefly, 4-5 Iodo-Beads were incubated with 200 pCi of NaiZ5I (100 mCi/ml) in 500 pl of phosphate-buffered saline (pH 7.4) for 10 min at room temperature. IO6 cercariae were then added to 500 p1 of the Na'? reaction mixture for 15 min at room temperature and washed extensively with spring water. After labeling with either procedure, the organisms were still motile. They were then resuspended in 0.5
80% phenol at 70 "C for 15 min as described (20). The extraction ml of spring water and gx was extracted using an equal volume of
procedure was repeated and the aqueous phases pooled and dialyzed against spring water for 48 h at 4 "C. The resulting soluble prepara- tions are referred to as "3H-gx" or "1251-gx". The 3H labeling procedure incorporated 2-5 X lo' cpm/106 cercariae. The yield of protein and carbohydrate from lo6 cercariae was -50 and -300 pg, respectively. Some experiments also made use of unlabeled gx extracted from the organisms by the above phenol-extraction procedure.
Column Chromatography Lectin Affinity Chromatography-Dialyzed radiolabeled gx was
chromatograpbed on a 2-ml Lotus lectin Sepharose affinity column at room temperature in 20 mM Tris-HC1 buffer (pH 7.6) containing
1 mM MnC12, 1 mM MgC12, 1 mM CaC12, 0.1% Triton X-100, and 0.1 M NaC1. Material bound to the column was eluted with 0.5 M L-fucose in the column buffer. Fractions of 1 ml were collected and aliquots of each fraction were counted for radioactivity.
Gel Filtration Chromatography-Chromatography on Sephacryl S- 1000 was performed at 4 "C on a 1.5 X 100-cm column equilibrated in 0.01 M NH4HCOZ buffer (pH 7.8) containing 0.1% SDS and run at 3 ml/h. The column was calibrated with DNA and cytochrome c to mark the void volume and total volume. Sepharose CL-GB (1 X 50- cm column) and Sephacryl S-200 (1 X 100-cm column) were eluted with 0.01 M NH4HC03 (pH 7.8) containing 0.1% SDS with flow rates of 15 and 3 ml/h, respectively, and the cohmns were calibrated with blue dextran (2 X lo6 kDa) and cytochrome e. Fractionation was followed by counting aliquots of each 1-ml fraction for radioactivity.
Ion Exchange Chromatography-To analyze the charge distribu- tion of gx, dialyzed aqueous extracts of gx were loaded on DEAE- Sepharose under two experimental conditions: A, DEAE-Sepharose CL-GB 5-ml columns equilibrated with 50 mM sodium acetate buffer (pH 5.8), 8.0 M urea (21). The column was eluted with 10 ml of the column buffer. A 50-ml salt gradient from 0.1 to 1.0 M NaCI in the column buffer was then applied and was followed with 2.0 M NaCl in the column buffer. Fractions were assayed for radioactivity or absorb- ance to measure carbohydrate when unlabeled material was used. E, ion exchange chromatography was also performed in 20 mM Tris- HCl buffer (pH 9.0) without any urea (22). The procedure was exactly the same as that described above. The salt gradient was determined by conductivity measurements. Recovery of labeled material from each step of purification was 80-90%.
Fluorescent Staining 3000 cercariae in 200 pl of phosphate-buffered saline containing 1
mM eserine sulfate were incubated with FITC-conjugatedhtus lectin, FITC-conjugated A. anguilla, FITC-conjugated U. europaeus (UEA 1) (50 pg/ml) for 30 min at room temperature (23). Control samples were supplemented with 0.1 M fucose. Such incubations do not induce transformation (24). Organisms were then fixed in 1% p-formalde- hyde in phosphate-buffered saline for 10 min on ice, washed, stored in 90% glycerol, and observed under a Nikon Ultraphot microscope.
Release of Oligosaccharides from 'H-Gx by Mild Alkaline Borohydride Treatment
3H-Gx and unlabeled gx was treated with alkaline NaBH, to characterize the putative carbohydrate peptide linkage (25). Samples were treated with an equal volume of freshly prepared 2.0 M NaBH, in 0.1 M NaOH (pH 11) for 19 h at 45 "C. At the end of the incubation period, the reaction was terminated by neutralizing the mixture with 5 N acetic acid. The samples were then adjusted to 0.1% SDS, and chromatographed on gel filtration and ion exchange columns.
Chemical and Enzymatic Treatments The presence of sialic acid in 3H-gx was examined by treating with
50 mM H2S04 at 80 "C for 1 h followed by neutralization with NaOH and clarification by centrifugation at 400 rpm for 10 min (26). Samples were also treated with neuraminidase (2 units/ml) from V. cholerae, C. perfringens, and A. urafaciens. Incubations were for 24 h at 37 "C in 0.2 M sodium acetate buffer (pH 5.6), containing 2 mM CaC12 (27). To examine the possible presence of glycogen, samples were incubated with 500 units of a-amylase in 0.02 M sodium phos- phate buffer (pH 6.9) containing 0.006 M NaCl at 37 "C for 1 h (28). Treated and control (no acid/enzyme treatment) samples were chro- matographed on Sepharose CL-GB and Sephadex G-25.
To examine the putative peptide backbone of gx, aliquots of 1251- gx were subjected to four different proteases (10 mg/ml): Pronase and trypsin in 50 mM ammonium bicarbonate buffer (pH 8.0) at 37 'c for 19 h, proteinase K and thermolysin in 50 mM ammonium bicarbonate buffer (pH 7.4) containing 0.1% SDS at 65 "C for 6 h. Treated samples were concentrated on a speed Vac concentrator (Savant, Hicksville, NY) and subjected to chromatography on CL-GB column. In addition, samples were also electrophoresed on 10% SDS-polyacrylamide gel electrophoresis gels after dilution with equal amounts of reducing sample buffer. Gels were fixed in methanol/acetic acid/water (5/1/4, by v/v) for 1 h, soaked in Enlighter (Du Pont-New England Nuclear) for 30 min, dried, and exposed to Kodak X-AR film (Eastman Kodak Co., Rochester, NY) at -70 "C for fluorography.
Protein measurements were done using a Bio-Rad assay kit. Car- bohydrate was estimated by phenol sulfuric acid assay (29). Briefly, to 100 p1 of the sample, 100 p1 of 85% phenol was added. TO this

S. mansoni Glycocalyx 1343
reaction mixture 2.0 ml of sulfuric acid was added, samples were cooled on ice, and absorbance read immediately at 490 nm. Glucose (5-100 pg) processed as above was used as standard in the assay.
Linkage Sugar Analysis
In order to determine the linkage sugar, gx from bodies and tails were treated with 50 mM NaOH containing 1 M NaB'H4 at 45 "C for 16 h. Samples were neutralized by dropwise addition of 4 N acetic acid, and deionized with Dowex 50-H+ (Sigma). Residual borate was removed by repeated evaporation of the samples with a solution of 1 M acetic acid in methanol. The released oligosaccharides were hydro- lyzed in 2 N HC1 for 3 h at 100 "C. Following hydrolysis the acid was removed by evaporation under reduced pressure, the sugars were reacetylated in 0.3 ml of H20 with 0.03 ml of 4 N Na2C03 and 0.045 ml of freshly prepared ice-cold 12.5% acetic anhydride solution in H20. After 10 min at room temperature followed by 5 min at 100 "C, the mixture was desalted over a 3-ml column of AG 501-X8 (Bio- Rad) mixed-bed resin. The samples were subjected to descending paper chromatography on Whatman No. 1 paper (previously impreg- nated with 0.57% Na2B207.10H20 containing 0.01 M NaCI) in the solvent system 1-butanol/pyridine/water (6/4/3) for 60 h (22). The chromatogram was cut into 1-cm strips which were counted for radioactivity. Standard sugars N-[3H]acetylgalactosaminitol and N- [~'H]acetylglucosaminitol were prepared by labeling nonradioactive sugars, as described above.
Analysis of Carbohydrate Composition Carbohydrate composition was determined by three different meth-
ods. 1) Alditol Acetate Deriuatiues-Alditol acetates were prepared by
hydrolyzing salt-free samples with 2 M trifluoroacetic acid for 2 h at 121 "C. Glycosyl residues were reduced with NaBD4, and acetylated with acetic anhydride/pyridine, followed by gas chromatography, and combined gas chromatography/mass spectrometry (GC/MS) analysis. For alditol acetates the column was a Supelco 2330 fused silica capillary column. GC/MS analysis was performed using a Hewlett- Packard 5890 GC coupled to a 5970 MSD.
2) Me& Deriuatiues--Me.lSi methylglycosides were prepared by methanolysis in 1 M HCI in methanol, followed by N-acetylation with pyridine and acetic anhydride (for detection of amino sugars). The samples were then treated with Tri-Sil, followed by GC and GC/MS analysis (30). GC analysis of the MelSi methylglycosides was done on a HP 5890 GC using a Supelco DB1-fused silica capillary column.
3) Dioner-Salt-free samples were added to 1-ml vials and dried by lyophilization. 0.1 ml of 2 M trifluoroacetic acid was added, flame sealed, and heated at 100 "C for 4-5 h (31). After hydrolysis, trifluo- roacetic acid was removed by evaporation under reduced pressure, the aqueous phase dried on a Speed Vac concentrator, resuspended in 100 pl of distilled water containing 5 mM NaOH, 10 mM NaOAc. Resuspended samples were analyzed on Dionex HPLC equipped with a pulse amperometric detector. Chromatography was carried out with Carbopac PA 1 column at 1 ml/min using 100 mM NaOH, 1 M NaOAc in distilled water as the gradient system.
Procedures 1 and 2 were conducted by the Complex Carbohydrate Research Center (Athens, Georgia) using standard procedures. Con- trols for sugar degradation were run in parallel.
Rotary Shadowing, Negative Staining, and EM Studies
'H-Gx was digested with DNase I (10 mg/ml) a t 37 "C for 30 min to remove any traces of DNA and chromatographed on Sepharose CL-GB in the presence of 7 M guanidine HCl in 10 mM NH4HCOs (pH 7.8). The excluded volume was recovered, concentrated, and resuspended in 10 mM ammonium bicarbonate buffer (pH 7.8) con- taining 50% glycerol to a final concentration of 20-50 pg/ml. The samples were sprayed onto a freshly cleaved mica substrate a t a pressure of 28 psi at a distance of 30 cm. The dried specimens were then rotary shadowed at a 6" angle, 10-cm distance with 1.0 cm of 0.08-inch thick 100% platinum, evaporated off 0.050-inch tungsten. Shadowed samples were then coated with a thin layer of carbon and floated off on a water surface, and picked up on 400-mesh copper grids (32). For negative staining, samples were adsorbed from 0.01 M NH4HCOs solution onto carbon films coated on a copper grid, ren- dered hydrophilic by glow discharge. Staining was performed with a freshly prepared saturated uranyl acetate solution of pH 4.0 (32). Samples in both cases were observed in a JEOL 100s microscope at 60 Kev using a 30-pm objective aperture.
RESULTS
Characterization of Intact Gx-We have labeled the gx by a reductive procedure employing NaB3H4 which is known from autoradiographic studies to label only the surface of S. man- soni (11). Since 3H-gx extracted from whole cercariae was found to be heterogeneous, we separated the bodies and tails before labeling them. As gx is rich in fucose and since Lotus lectin binds fucose, we used Lotus lectin affinity as a first step in the characterization. 70% of 'H-gx from bodies bound to the column and could be eluted with 0.5 M fucose (Fig. 1). By contrast, only 20% of 'H-gx from the tails bound to the column. Rechromatography showed no change in the chro- matographic properties of 'H-gx of bodies or tails.
A striking fluorescent pattern was obtained using a FITC- conjugated fucose-specific lectin. FITC-conjugated A. anguilla lectin showed intense specific fluorescence on the surface of the bodies (Fig. 2) consistent with the affinity chromatogra- phy data (Fig. 1). Fluorescence was markedly less on tails and was restricted to creases and the forked region of the tails (Fig. 2). No fluorescence was observed in the presence of 0.1 M fucose, the competing sugar. Commercial FITC-conjugated Lotus lectin and Ulex lectin did not consistently stain bodies or tails, possibly because of variations in the preparation or stability of these conjugates.
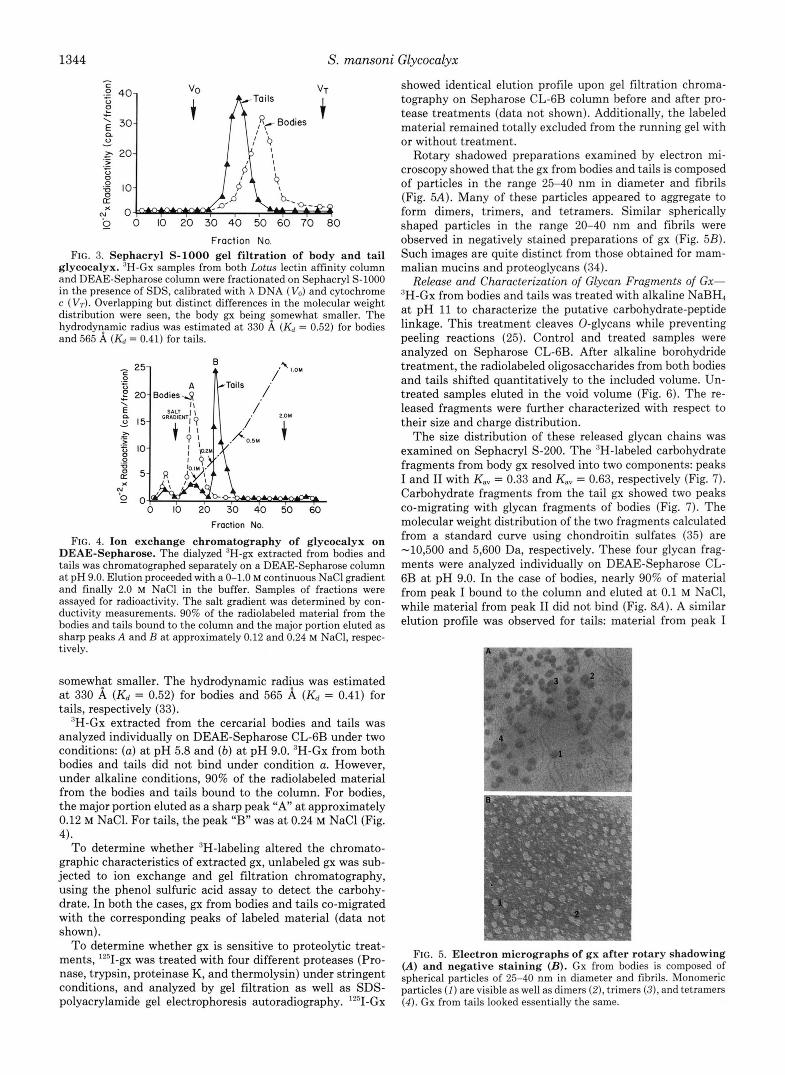
To study the size distribution of 3H-gx, samples of gx were fractionated on Sephacryl S-1000 in the presence of 0.1% SDS. As illustrated in Fig. 3, both bodies and tails exhibited polydispersity. Overlapping but distinct differences in the molecular weight distributions were seen, body 3H-gx being
A
g "1 Y
1 5 1 hToi ls I k B o d i e s E R
A
Fraction No. FIG. 1. Lotus lectin affinity chromatography of 3H-gx. 'H-
Gx from bodies and tails was loaded on the Lotus lectin affinity column and was washed with column buffer. The bound material was eluted with 0.5 M fucose in column buffer. 70% of 3H-gx from bodies bound to the column. By contrast, only 20% of 'H-gx from tails bound to the column. Rechromatography showed no change in the chromat- ographic properties of 'H-gx bodies and tails.
FIG. 2. Fluorescent staining with a fucose-specific lectin. 3000 cercariae in 200 p1 of phosphate-buffered saline containing 1 mM eserine sulfate were incubated with FITC-conjugated A. anguilh lectin (50 pg/ml)for 30 min at room temperature. Organisms were then fixed in 1% formaldehyde in phosphate-buffered saline. Intense specific fluorescence was observed on the surface of bodies (b). Fluo- rescence was markedly less on tails ( t ) and was restricted to creases and the forked region on tails. No fluorescence was observed in the presence of 0.1 M fucose.
1344 S. mansoni Glycocalyx
Q ‘4 IO 0 CC d
qJx 0 Q 0 IO 20 30 40 50 60 70 80
Fraction No. FIG. 3. Sephacryl S-1000 gel filtration of body and tail
glycocalyx. “H-Gx samples from both Lotus lectin affinity column and DEAE-Sepharose column were fractionated on Sephacryl S-1000 in the presence of SDS, calibrated with X DNA (Vo) and cytochrome c (Vr). Overlapping but distinct differences in the molecular weight distribution were seen, the body gx being somewhat smaller. The hydrodyyamic radius was estimated at 330 A (& = 0.52) for bodies and 565 A (& = 0.41) for tails.
0 10 20 30 40 50 60 Fraction No.
FIG. 4. Ion exchange chromatography of glycocalyx on DEAE-Sepharose. The dialyzed ‘H-gx extracted from bodies and tails was chromatographed separately on a DEAE-Sepharose column at pH 9.0. Elution proceeded with a 0-1.0 M continuous NaCl gradient and finally 2.0 M NaCl in the buffer. Samples of fractions were assayed for radioactivity. The salt gradient was determined by con- ductivity measurements. 90% of the radiolabeled material from the bodies and tails bound to the column and the major portion eluted as sharp peaks A and B at approximately 0.12 and 0.24 M NaCI, respec- tively.
somewhFt smaller. The hydrodynamic radius was estimated at 330 A (Kd = 0.52) for bodies and 565 A ( K d = 0.41) for tails, respectively (33).
”H-Gx extracted from the cercarial bodies and tails was analyzed individually on DEAE-Sepharose CL-GB under two conditions: (a) at pH 5.8 and (b) at pH 9.0. ‘H-Gx from both bodies and tails did not bind under condition a. However, under alkaline conditions, 90% of the radiolabeled material from the bodies and tails bound to the column. For bodies, the major portion eluted as a sharp peak “ A at approximately 0.12 M NaCl. For tails, the peak “B” was at 0.24 M NaCl (Fig. 4).
To determine whether “-labeling altered the chromato- graphic characteristics of extracted g x , unlabeled gx was sub- jected to ion exchange and gel filtration chromatography, using the phenol sulfuric acid assay to detect the carbohy- drate. In both the cases, gx from bodies and tails co-migrated with the corresponding peaks of labeled material (data not shown).
To determine whether gx is sensitive to proteolytic treat- ments, I2’I-gx was treated with four different proteases (Pro- nase, trypsin, proteinase K, and thermolysin) under stringent conditions, and analyzed by gel filtration as well as SDS- polyacrylamide gel electrophoresis autoradiography. ‘2sI-Gx
showed identical elution profile upon gel filtration chroma- tography on Sepharose CL-GB column before and after pro- tease treatments (data not shown). Additionally, the labeled material remained totally excluded from the running gel with or without treatment.
Rotary shadowed preparations examined by electron mi- croscopy showed that the gx from bodies and tails is composed of particles in the range 25-40 nm in diameter and fibrils (Fig. 5A). Many of these particles appeared to aggregate to form dimers, trimers, and tetramers. Similar spherically shaped particles in the range 20-40 nm and fibrils were observed in negatively stained preparations of gx (Fig. 5B). Such images are quite distinct from those obtained for mam- malian mucins and proteoglycans (34).
Release and Characterization of Glycan Fragments of Gx- 3H-Gx from bodies and tails was treated with alkaline NaBH4 at pH 11 to characterize the putative carbohydrate-peptide linkage. This treatment cleaves 0-glycans while preventing peeling reactions (25). Control and treated samples were analyzed on Sepharose CL-GB. After alkaline borohydride treatment, the radiolabeled oligosaccharides from both bodies and tails shifted quantitatively to the included volume. Un- treated samples eluted in the void volume (Fig. 6). The re- leased fragments were further characterized with respect to their size and charge distribution.
The size distribution of these released glycan chains was examined on Sephacryl S-200. The ‘H-labeled carbohydrate fragments from body gx resolved into two components: peaks I and I1 with K,, = 0.33 and K,,, = 0.63, respectively (Fig. 7). Carbohydrate fragments from the tail gx showed two peaks co-migrating with glycan fragments of bodies (Fig. 7). The molecular weight distribution of the two fragments calculated from a standard curve using chondroitin sulfates (35) are -10,500 and 5,600 Da, respectively. These four glycan frag- ments were analyzed individually on DEAE-Sepharose CL- 6B at pH 9.0. In the case of bodies, nearly 90% of material from peak I bound to the column and eluted at 0.1 M NaCl, while material from peak I1 did not bind (Fig. 8A). A similar elution profile was observed for tails: material from peak I
!. ,
I
I
FIG. 5. Electron micrographs of gx after rotary shadowing (A) and negative staining ( B ) . Gx from bodies is composed of spherical particles of 25-40 nm in diameter and fibrils. Monomeric particles ( 1 ) are visible as well as dimers (2), trimers (3), and tetramers (4 ) . Gx from tails looked essentially the same.

S. mansoni Glycocalyx 1345
4 vo Bodies
t
CT
IO 20 30 40 50 60 70
0 0 10 20 30 40 50 60 70
Froction No.
FIG. 6. Release of labeled glycan chains by alkaline borhy- dride treatment. 3H-Gx from bodies and tails was treated with alkaline NaBH4 at pH 11 to characterize the putative carbohydrate- peptide linkage. Control and treated samples were analyzed on Seph- arose CL-GB (0.1% SDS in NH4HCO3, pH 7.8) calibrated with blue dextran (Vo) and cytochrome c (VT). The radiolabeled oligosaccha- rides from both bodies ( A ) and tails ( B ) shifted quantitatively to the included volume.
Fraction No.
FIG. 7. Sephacryl 5-200 elution profile of the 3H-glycan chains released from 3H-gx by alkaline borohydride treat- ment. The labeled fragments released from bodies and tails were analyzed for their size distribution on Sephacryl S-200 calibrated with blue dextran (Vo) and cytochrome c (VT). The 3H-labeled car- bohydrate fragments from body gx resolved into two components, fragment I of M, 10,500, fragment I1 of lower M, 5,600. Carbohydrate fragments from the tail gx showed two peaks co-migrating with peaks from bodies, indicating the presence of two different size 0-glycan chains.
bound and eluted at 0.2 M NaCl, while peak I1 material did not bind (Fig. 8B). Glycans released from unlabeled gx showed elution profiles identical to those seen for labeled samples.
To identify the putative linkage sugar(s), both body and tail gx were treated with NaOH in the presence of NaB3H4, hydrolyzed, and analyzed by descending paper chromatogra- phy. The released radioactive sugar co-migrated with standard N-[3H]acetylgalacto~aminitol in both cases (Fig. 9, A and B ) .
Sugar Composition of Intact gx and Carbohydrate Fragments Released by Alkaline Borohydride Treatment-Carbohydrate composition was determined by three different methods to eliminate the possibility of underestimating some sugars due to destruction or resistance to hydrolysis. In addition, optimal reaction conditions were chosen and standard sugars were run simultaneously under identical conditions. Carbohydrate compositional analysis obtained by the three methods agreed well with each other. Table I gives the neutral sugar values
Bodies ,.OM
I.
1 > 0 IO 20 .- c
Toils I OM
9
Fraction No.
FIG. 8. Ion exchange chromatography of 3H-glycan chains. The four different glycan fragments from S-200 column were analyzed individually on DEAE-Sepharose CL-GB at pH 9.0. In case of bodies ( A ) , nearly 90% of material from peak I bound to the column and eluted at 0.1 M NaCI, while material from peak I1 did not bind. A similar elution profile was observed for tails ( B ) , material from peak I bound and eluted at 0.2 M NaCl, while peak I1 material did not bind.
5 0 0 0 0 ~ 10000 i
0 10 20 30 40
cms
750 i i 1
0 10 20 30 40
crns FIG. 9. Identification of linkage sugar. Body and tail gx were
subjected to p-elimination in the presence of NaB3H4. The released saccharides were hydrolyzed in 2 N HC1 for 3 h at 100 “C, re-N- acetylated and analyzed by descending paper chromatography as described under “Experimental Procedures.” The migration position of standards are indicated: 1, N-acetylgalactosaminitol; 2, N-acetyl- glucosaminitol.
determined by the alditol acetate method and the amino sugars determined by the MesSi method. In agreement with the previous findings on total gx (E), we found that fucose

1346 S. mansoni Glycocalyx
TABLE I Carbohydrate composition of cercarial body and tail gx
Values are expressed as the percent of total carbohydrate deter- mined by the alditol acetate method (neutral sugars) and Me3Si method (amino sugars). Sialic acid was not detected by any of the three analytic procedures.
Monosaccharide Body Tail
% Fucose 46.6 5.3 Galactose 15.9 3.2 Galactosamine 10.5 3.1 Glucose" 17.3 84.0 Glucosamine 3.2 1.8 Mannose 2.8 1.0
Glucose values were found to be slightly higher by the MesSi and Dionex-HPLC methods in the case of bodies.
TABLE I1 Carbohydrate composition of glycan chains I and 11
of body and tail gx Body
Monosaccharide Tail
I I1 I I1
%" % Fucose 48.5 15.1 8.6 1.1 Galactose 13.6 30.4 8.2 13.6 Galactosamine 12.8 5.4 5.0 2.8 Glucose" 10.0 31.8 72.0 80.0 Glucosamine 14.3 11.7 3.8 0 Mannose 0.4 5.6 2.4 1.3
See Table I.
was a major component of gx of bodies. In addition, glucose, which has not been previously described in gx, was found to be the most abundant sugar in the tail gx. Analysis of mock column eluants showed that the glucose could not be attrib- uted to contaminants. Galactose, galactosamine, glucosamine, and mannose were present in lesser amounts. Sialic acid was not detected. Analysis of the two glycans released by alkaline borohydride treatment indicate that fucose is the predomi- nant sugar in fraction I of bodies, while glucose is the major sugar contributing to fraction I from tails (Table 11). Both for bodies and tails, the total amount of carbohydrate recovered from fraction I was 5-6 times that obtained from fraction 11.
Analysis of Gx for the Presence of Sialic Acid-Sialic acid is readily labeled by the procedure we have used to produce 3H- gx (26) and is also easily released by mild H2S04 treatment (26). As shown in Fig. 10, A and B, respectively, 3 H - g ~ from both bodies and tails was sensitive to mild acid hydrolysis: all 'H label shifted to the total volume upon gel filtration on Sephadex G-25. Further characterization of acid-released ra- dioactivity by thin layer chromatography on silica gel showed slower migration than [3H]sialic acid released from labeled fetuin. In addition, gel filtration showed that 3H-gx was re- sistant to neuraminidases of C. perfringens, v. cholerae, and A. urafaciens which are known to be active on other glycopro- teins labeled with the procedures we have used (Ref. 26 and data not shown).
DISCUSSION
There have been several anatomic and cytochemical studies of the cell surface glycocalyx of parasitic helminths (2-7); however, little corresponding chemical information is avail- able. s. mansoni is among the better-studied organisms, in that recent chemical and immunochemical studies of its gly- cocalyx indicate that the major component of the surface coat is rich in fucose and has a molecular weight in excess of 5 X lo6 (15). Furthermore, several smaller (15,000-220,000 dal-
Fraction No.
FIG. 10. Analysis of gx for the possible presence of sialic acid. 3H-Gx of bodies and tails was treated with 50 mM H,SO, at 80 "C for 1 h followed by neutralization with NaOH and subjected to chromatography on Sepharose CL-GB. Control samples with no acid were incubated under identical conditions. 3H-Gx from both bodies (A) and tails ( B ) was sensitive to mild acid hydrolysis: all 3H label shifted to the total volume. The released labeled material on Sephadex G-25 eluted close to the total volume.
tons) surface glycoproteins of cercariae have been identified and monoclonal antibodies raised against this stage of the parasite detect epitopes which are shared by several compo- nents of the cercarial surface (14). I t is striking that the binding of at least one such antibody can be inhibited by free fucose (14). The question of possible chemical differences between the glycocalyx of cercarial bodies uersus tails has not been previously addressed. A substantial difference between the two parts of cercariae may account for the observations that the gx of tails is not lost during transformation (11) and that tails, which do not penetrate mammalian hosts, are more sensitive than bodies to lysis by anti-gx antibodies and com- plement (13).
Our observations show that the high molecular weight gx of bodies differs from that of the tails in several regards: 1) only body gx binds to fucose-specific lectins, and therefore contains accessible terminal nonreducing fucose residues; 2) the apparent size of body gx is smaller than that of tail gx; 3) body gx is much richer in fucose than tail gx, which is rich in glucose; 4) although both varieties of gx are anionic, tail gx is more anionic than body gx. The negative charge cannot be accounted for by sulfate (15), but, judging from the acid lability of the tritium label, might reflect the presence of sialic acid. Previous studies have suggested the presence of sialic acid on the surface of adult schistosomes based on neuramin- idase treatment (36) and indirect lectin-binding techniques (37). However, no sialic acid was detected on biochemical analysis of the major 0-linked oligosaccharides in schistoso- mula or adult worm glycoproteins (38, 39). In the present studies, three neuraminidases are ineffective in releasing label from 'H-gx. Furthermore, carbohydrate analysis did not de- tect sialic acid.
The abundance of glucose in tail gx is surprising since tail gx does not bind lectins which interact with terminal nonre- ducing glucose residues (40). The observation that gx of both body and tail is resistant to a-amylase treatment' rules out
J. Nanduri, unpublished results.

S. mansoni Glycocalyx 1347
the possibility that the glucose could be derived from associ- ated glycogen.
Up on treatment with alkaline borohydride, nearly 90% of the carbohydrate of both body and tail gx is converted to a pair of smaller fragments. Thus, a total of at least four types of glycans are present. These observations are consistent with glycan linkages to serine or threonine residues. The major 0- linked sugar in both body and tail gx is N-acetylgalactosa- mine. The apparent sizes of the released glycans and the 0- linkages are different from the 0- and N-linked oligosaccha- rides in other glycoproteins synthesized by schistosomula and adult worms (39).
lZ5I-Gx was resistant to proteolytic treatments. This resist- ance may reflect the abundance of carbohydrate which con- ceals a putative peptide backbone. Resistance may also be of biological importance for protection of cercariae from other organisms in the natural habitat. Apart from the presence of amino acids in total preparations of gx (15), and suggestions that proteases secreted by transforming schistosomula play a role in the release of schistosomular surface molecules (41), there is no evidence that gx includes a peptide backbone. Indeed, one study suggests that a glycolipid moiety may be responsible for anchoring of surface components of cercariae and schistosomula (42).
Electron microscopic studies of gx after rotary shadowing and negative staining demonstrate a globular structure for gx and do not show obvious resemblance to proteoglycan, i.e. a polypeptide backbone with multiple lateral oligosaccharide chains. The possibility that the anatomic polydispersity ob- served could be due to aggregation is unlikely since incubation with various agents known to prevent aggregation such as guanidine (8 M ) and SDS (1.0%) resulted in no change in the size or shape of the molecule. On the other hand, the polydis- persity may be due to different degrees of hydration.
Although the size of labeled and unlabeled gx preparations extracted by phenol is impressive, our electron microscopic observations, as well as those of others ( E ) , make it clear that the dimensions of the isolated molecule are far smaller than those of the glycocalyx in situ which is 1-2 p m thick (7, 10). The magnitude of the size discrepancy in the case of S. mansoni and the degree of polydispersity are so striking that one wonders to what extent there may be linkages within the gx (such as the fibrils which are seen) which are broken by the extraction conditions. Such “breakage” might also account for the water solubility of gx after extraction. It is also striking that although body and tail gx differ significantly in their carbohydrate composition, the glycan chains are of the same molecular weight distribution. Further studies are needed to elucidate the biological mechanism of gx release and to iden- tify the sites of action of any proteases and/or glycosidases which may be involved.
Acknowledgments-We thank Drs. Lloyd Culp, Arnold Caplan, Neil Jentoft, and Martin Snider for helpful suggestions, S. H. Yang for carbohydrate analysis, and Marie Ward for beautifying the text.
REFERENCES
1. Ito, S. (1969) Fed. Proc. 28, 12-25 2. Fawcett, D. W. (1965) J. Histochem. Cvtochem. 13. 75-91
4. Bennett, H. S. (1963) J. Histochem. Cytochem. 11, 14-23 5. Lumsden, R. D. (1975) Exp. Parasitol. 37, 267-339 6. Lumsden, R. D. (1972) J. Parasitol. 58, 229-234 7. Threadgold, L. T. (1976) Exp. Parasitol. 39, 119-134 8. Stirewalt, M. A,, Cousin, C. E., and Dorsey, C. H. (1983) Exp.
9. Stirewalt, M. A. (1975) Adu. Parasitol. 12, 115-181 Parasitol. 56, 358-368
10. Hockley, D., and McLaren, D. (1973) Int. J . Parasitol. 3, 13-25 11. Samuelson, J. C., and Caulfield, J. P. (1985) J. Cell Biol. 100,
12. Caulfield, J. P., Yuan, H. C., Cianci, C. M. L., and Hein, A. (1988)
13. Samuelson, J. C., and Caulfield, J. P. (1986) Infect. Immun. 51,
14. Dalton, J. A,, Levis, S. A., Aronstein, W. S., and Strand, M.
15. Caulfield, J . P., Cianci, C. M. L., McDiarmid, S. S., Suyemitsu,
16. Smithers, S. R., and Terry, R. J. (1965) Parasitology 55,695-699 17. Colley, D. G., and Wikel, S. K. (1974) Exp. Parasitol. 35, 44-51 18. Ramalho-Pinto, F. J., Gazzinelli, G., Howells, R. E., Mota-Santos,
T . A,, Figueredo, E. A., and Pellegrino, J. (1976) Exp. Parasitol. 36,360-372
1423-1434
Exp. Parasitol. 65, 10-19
181-186
(1987) Exp. Parasitol. 63, 215-226
T., and Schmid, K. (1987) J. Parasitol. 73, 514-522
19. Markwell, M. (1972) Anal. Biochem. 25, 464-470 20. Westphahl, O., and Jann, L. (1965) Methods Carbohydr. Chem.
21. Wightman, B., Weltman, E., and Culp, L. (1986) Biochem. J .
22. Cummings, R. D., Merkle, R. K., and Stults, N. L. (1989) Methods
23. Wiest, P., Tartakoff, A., Aikawa, M., and Mahmoud, A. (1988)
24. Wiest, P., Kossman, R. J., and Tartakoff, A. M. (1989) A m . J .
25. Carlson, D. M. (1968) J. Biol. Chem. 243, 616-626 26. Gahmberg, C. G., and Andersson, L. C. (1977) J. Biol. Chem.
252,5888-5894 27. Woodward, H. D., Ringler, N. J., Selvakumar, R., Simet, I. M.,
Bhavandan, V. P., and Davidson, E. A. (1987) Biochemistry
28. Boctor, F. N., Nash, T. E., and Cheever, A. W. (1979) J. Immunol. 122,39-43
29. Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., and Smith, F. (1956) Anal. Chem. 28, 350-356
30. York, W. S., McNeil, A. G., Stevenson, T. T., and Albersheim, P. (1985) Methods Enzymol. 118, 3-40
31. Biermann, C. J. (1988) Adu. Carbohydr. Chem. Biochem. 46,251- 270
32. Paulsson, M., Morgelin, M., Wiedemann, H., Beardmore-Gray, M., Dunham, D., Hardingham, T., Heinegard, D., Timpl, T., and Engel, J. (1987) Biochern. J . 245, 763-772
33. Shogren, R., Jentoft, N., Jamieson, A., Gerkan, P., and Blackwell, J. (1987) Carbohydr. Res. 160, 317-327
34. Constantopoulos, G., Dekaban, A. S., and Carroll, W. R. (1969) Anal. Biochem. 31, 59-70
35. Sheehan, J. K., and Carlstedt, I. (1990) Biochem. J . 265, 169- 178
36. McDiarmid, S. S., and Podesta, R. B. (1984) Mol. Biochem. Parmitol. 10, 33-43
37. Simpson, A. J., Corria-A-Oliveria, R., Smithers, R. S., and Sher, A. (1983) Mol. Biochem. Parasitol. 8, 191-205
38. Nyame, K., Cummings, R. D., and Damian, R. T . (1987) J. Biol. Chem. 262,7990-7995
39. Nyame, K., Cummings, R. D., and Damian, R. T. (1988) J . Parasitol. 74, 562-572
40. Murrell, K., Taylor, D., Vannier, W., and Dean, D. (1978) Exp. Parasitol. 46, 247-255
41. Marikovsky, M., Arnon, R., and Fishelson, Z. (1988) J . Immunol.
5,83-91
235,469-479
Cell Biol. 32, 141-183
Proc. Natl. Acad. sci. U. s. A . 85, 3825
Trop. Med. Hyg. 41, 70-77
26,5315-5322
141,273-278 3. Martinez-Palom’o, A.’(1970) Int. Rev. &ol. 29, 29-74 42. Pearce, E. J., and Sher, A. (1989) J. Immunol. 142, 979-984


















