
GFMJongsma Honours Thesis
38
The Impacts of Historical and Ongoing Human Activities on Amphibian Diversity in Bilsa Biological Station, Western Ecuador by Gregor F.M. Jongsma Thesis submitted in partial fulfillment of the requirements for the Degree of Bachelor of Science with Honours in Biology Acadia University 2012 © Copyright by Gregor F.M. Jongsma 2012
-
Upload
gregorius-jongsmaa -
Category
Documents
-
view
38 -
download
2
Transcript of GFMJongsma Honours Thesis

The Impacts of Historical and Ongoing Human Activities on
Amphibian Diversity in Bilsa Biological Station,
Western Ecuador
by
Gregor F.M. Jongsma
Thesis submitted in partial fulfillment of the
requirements for the Degree of
Bachelor of Science with
Honours in Biology
Acadia University
2012
© Copyright by Gregor F.M. Jongsma 2012

ii
This thesis by Gregor F.M. Jongsma is accepted in its present form by the Department of
Biology as satisfying the thesis requirements for the degree of Bachelor of Science with
Honours.
Approved by the Thesis Supervisor
__________________________ ____________________
(Tom Herman) Date
Approved by the Head of the Department
__________________________ ____________________
(Soren Bondrup-Nielsen) Date
Approved by the Honours Committee
__________________________ ____________________
(Pritham Ranjan) Date

iii
I, Gregor F.M. Jongsma, grant permission to the University Librarian at Acadia University to
reproduce, loan or distribute copies of my thesis in microform, paper or electronic formats on a
non-profit basis. I, however, retain the copyright in my thesis.
_________________________________
Signature of Author
_________________________________
Date

iv
Acknowledgements
I am grateful for the assistance provided by staff of the Jatún Sacha Foundation and Bilsa
Biological Station, particularly C. Aulestia and J. Bermingham, and by field researchers D.
Caberera, L. Carrasco, F. Castillo, and J. Olivo. For assistance with species identification many
thanks go to G. Vigle and M. Ortego-Andrade. At Acadia, T. Herman and S. Bondrup-Nielsen
provided valuable input throughout the writing process of my thesis. Research was supported by
the National Science Foundation (OISE-0402137), National Geographic Society, Conservation,
Food and Health Foundation and Disney Wildlife Conservation Fund.

v
Table of Contents
List of Tables............................................................................................................vi
List of Figures ......................................................................................................... vii
Abstract .................................................................................................................. viii
Introduction ............................................................................................................... 1
Materials and Methods ............................................................................................... 5
Results ..................................................................................................................... 11
Discussion ............................................................................................................... 15
Literature Cited ........................................................................................................ 21
Appendix 1 ............................................................................................................................................ 24
Appendix 2..............................................................................................................25
Appendix 3......................................................................................................26
Appendix 4......................................................................................................30

vi
List of Tables
Table 1: Number of individual amphibians recorded across seven habitat categories in Bilsa
Biological Station, Ecuador.
Table 2: Total individuals, diversity (Shannon-Wiener and Simpson‘s respectively), observed
and expected species richness for the seven habitat categories.

vii
List of Figures
Figure 1: Three maps showing where Ecuador is located in South America, where the Mache
Chindul Reserve is located in Ecuador and where Bilsa (where the study was conducted) is
located within the Mache Chindul Reserve.
Figure 2: Expected species accumulation curves for amphibians captured in different habitat
categories in Bilsa Biological Station, according to the Mao Tau index. The vertical hatched line
indicates the minimum abundance level recorded for comparison across habitat types. Curves
were re-scaled to show accumulation of species in relation to number of captured individuals.

viii
Abstract
Human activity in western Ecuador is extensive in the form of logging and slash-and-
burn agriculture, however the impacts of these activities on amphibian diversity is almost
completely unknown. During a nine-month study in the Bilsa Biological Station situated in the
province of Esmeraldas, Ecuador I attempted to evaluate the impacts of these activities.
Amphibian diversity was estimated using visual encounter surveys along transects in pristine
sites (forest and river habitat that has no history of human activities) and historically disturbed
sites (forest and river habitat that was logged between 1989 and 1995) as well as along roads
with varying levels of ongoing human activity. The results of this study indicate that the
influence of disturbance on the diversity and composition of amphibian assemblages in the Bilsa
Biological Station varies with habitat. Amphibian assemblages along rivers were the richest as
well as the least impacted by human activity. Diversity and species richness were lower in
secondary than in primary forest suggesting that amphibian assemblages in interior forest habitat
are more vulnerable to disturbances caused by logging. Road habitat around Bilsa harboured a
very different amphibian assemblage from that of forest and river habitats. The results of this
study show that amphibian assemblages vary markedly in vulnerability to disturbances by human
activity; recognising these differences is critical for developing land management plans that
promote amphibian diversity in north western Ecuador.

1
Introduction
Evidence is mounting that humanity is driving a sixth mass extinction event that is notable
for its unprecedented rate of species loss (Wilson 1992). Amphibians—frogs, salamanders and
caecilians— are the oldest group of terrestrial vertebrates, and are currently facing risk of
extinction world-wide (Stuart 2004; Wake and Vredenburg 2008). Of the 6,800 known
amphibian species approximately one third are listed as threatened and at least 43% are in
decline (Collins et al. 2007). In the past three decades, the rate of amphibian extinction has
exceeded the mean extinction rate of the last 350 million years 200-fold (Roelants et al. 2007).
Amphibians are well suited as indicator species for assessing the health of ecosystems and
because they inhabit most of the world‘s fresh-water and terrestrial biomes, they may serve as
ideal indicators of global biological health and predictors for world-wide species extinctions
(Collins and Halliday 2005).
There are many factors causing amphibians to decline world-wide and the complex
interactions among these factors are poorly understood (Collins et al. 2007). The factors causing
the decline include: environmental contamination, invasive species, disease, climate change, and
deforestation (Stuart 2004). All of these factors result in the alteration of natural habitats and all
are linked to the activities of our ever-growing human population (Wake and Vredenburg 2008).
One of the most prominent forms of habitat alteration, the conversion of forests to agricultural
lands, plantations, and cattle ranches, is occurring at an alarming rate in the tropics. It would be
difficult to find a region in the tropics where this is more evident than in the Chocó
biogeographic region in north-western Ecuador.
Today, conservationists refer to the Chocó region as the Tumbes-Chocó-Magdalena
hotspot—an area covering 274,597 km², extending along the coast of Peru north through coastal

2
Ecuador and Columbia to southern Panama. The region has been coined the ―hottest‖ of
biodiversity hotspots, one demanding immediate international attention as a conservation priority
(Parker and Carr 1992; Mittermeier et al. 1998). Rampant deforestation set this region apart
from other hotspots around the world as less than two percent of the original forest cover remains
(Dodson and Gentry 1991). Even these remnant stands are under siege by a combination of
corporate operations (logging, cattle ranching, African palm and banana plantations) and the
activities of impoverished Ecuadorian farmers (slash and burn agriculture and poaching).
Threats
Logging has played the most pervasive role in transforming the Mache-Chindul— an area
of the Tumbes-Chocó-Magdalena in north coastal Ecuador— both directly and indirectly
(Hamilton et al. Unpublished). Clear-cutting and selective logging transform the landscape by
removing the trees; roads created in the process allow access to land that becomes occupied by
farmers whose activities further transform the area. Cattle pastures and banana and African palm
plantations, as well as small-scale subsistence farms, are prevalent in the region. These activities
inhibit forest regeneration and further deplete the already nutrient-poor soil. As a result, the
Mache-Chindul landscape is now a matrix of pasture-land dotted with patches of primary and
secondary forest— mostly on steep hillsides unfit for grazing cattle or growing crops. The
severe fragmentation of this forest landscape has many implications for amphibians and other
flora and fauna local to the Chocó.
Endemism
An endemic species is one that is restricted to a single locality (Hickman et al. 2008).

3
Coastal Ecuador harbours some of the highest levels of endemism on earth and amphibians in the
region are no exception. Of the 213 recognized amphibian species in the Tumbes-Chocó-
Magdalena, 40 are endemic (Conservation International). Endemic species are inherently at
greater threat of extinction due to their naturally restricted distributions (David 2005).
Most of the world‘s amphibians live in the tropics, where they occupy small geographic
ranges and have specific micro-habitat requirements (Wake and Vredenburg, 2008). These
characteristics leave amphibians vulnerable to rapid extinction. Some species, like the glass frog
Cochranella mache, have only been recorded along a few mountain streams in Western Ecuador
(IUCN). Human encroachment into this small area could very well eliminate the entire species.
Given the intensity of historical and current habitat alteration in the region, many species have
almost certainly perished undocumented and will likely continue to be lost. Habitat destruction
in regions with such high levels of endemism significantly reduces global biodiversity, and
conservation in these regions should be made an international priority.
Conservation
Successful species management and recovery require a number of steps. Amphibian
conservation in the Mache-Chindul is under-developed and a number of initiatives are still
required: (1) current amphibian diversity and population trends need to be accurately assessed,
(2) areas of high conservation priority (i.e. areas with high amphibian diversity or immediately
threatened by human activity) and corridors connecting them need to be identified and protected,
(3) suitable amphibian habitat needs to be allowed to regenerate (Young et al. 2001).
Unfortunately, reserves in Ecuador, as in most of the developing world, are poorly
protected. Illegal logging and slash and burn agriculture in the Mache Chindul are widespread

4
even within protected areas. If the three initiatives above are to be successful Ecuador‘s
government must provide adequate funding for reserves and hold corporations or individuals
who break the law accountable.
Equally important, government and NGOs must provide local residents with sustainable
economic alternatives that promote amphibian diversity, rather than degrade it. Any successful
amphibian conservation initiative will require the collaboration of scientists, government, NGOs,
and, above all, local people.
Objectives
The objectives of this study were two-fold. The first objective was to document the
species diversity of amphibians at Bilsa Biological Station. Over a nine month period amphibian
diversity was estimated using visual encounter surveys along transects in pristine and historically
disturbed forest sites as well as along roads with varying levels of human activity. Because
amphibians in this area are poorly known, these basic surveys contributed to the first complete
inventory of species composition and richness that is critical to any future conservation efforts.
The second objective was to gain insights into the influence of habitat on amphibian
diversity and abundance. To address this question, amphibians were sampled from a range of
habitats including forests, roadsides, and waterways. In this component of the study, I also
addressed how anthropogenic activity may be affecting amphibians by comparing each habitat
type (e.g., forest, road and river) in pristine conditions and in disturbed conditions.

5
Materials and Methods
Study Area
The Mache-Chindul mountain range of north-western Ecuador harbours some of the
greatest biodiversity on earth with a high degree of floral and faunal endemism. Two major
forest categories are found on this mountain range: (1) Humid Pre-Montane forest at elevations
between 500 and 1800 m, and (2) Humid Tropical rainforest below 500 meters. As part of the
Tumbes-Choco-Magdalena (TCM) biodiversity hotspot, international pressure to preserve the
region‘s unique ecosystems resulted in the creation of the Mache-Chindul Ecological Reserve
(MCER) in 1996
(http://www.ambiente.gov.ec/paginas_espanol/4ecuador/docs/areas/mache.htm). Based on the
author‘s personal observations little is being done to curb illegal logging, agriculture or poaching
and much of this 70,000 hectare area remains under threat. As a result, the park today is largely
deforested and consists mainly of small scale farms, cattle ranches, and African-palm
plantations, interspersed with small forest fragments. A smaller reserve inside the MCER, Bilsa
Biological Station, is privately owned and managed by the Jatun Sacha Foundation and is
sufficiently protected.
This study of amphibian diversity was carried out over nine months in 2007 in the Bilsa
Biological Station (79º 45‘ W, 0º 22‘ N, 330-730 meters above sea level) in the province of
Esmeraldas, Ecuador (Figure 1). The Tumbes-Choco-Magdalena region is one of the wettest
places on earth, and Bilsa receives on average more than 300 cm of rain per year. The
temperature is very stable. During the rainy season from January to June the average
temperature ranges between 24°C and 25°C. During the dry season from July to December the
average temperature is between 21°C and 22°C. With elevations ranging from 300m to 730m

6
Bilsa offers protection for both Humid-Pre-Montane forest and Humid Tropical forest. Being
one of the largest patches of remnant forest in North-Western Ecuador, Bilsa protects a mosaic of
primary and secondary forest covering an area of 3300 hectares, 80% of which is primary forest;
the remaining 20% of forest has been allowed to regenerate over the past 12-18 years. This
makes Bilsa one of the most representative areas in terms of existing or former diversity in the
surrounding valleys and rivers of the Mache-Chindul.
Selecting Sites
Rivers: River transect sites were selected based on the type of forest habitat surrounding
them (i.e. primary or secondary). Primary forest (never logged) and secondary forest (logged 12-
18 years previously) were identified with the help of experienced park staff. In addition to forest
habitat type, selection of sites required year-round water-flow and sufficient safety for
investigators to access and sample. Wherever possible, river sites with the most similar bank-
width and water velocity were selected; however, given the high heterogeneity of the area this
was not always possible. In all cases eight common environmental variables, described in detail
below, were recorded for each river transect. In total, 36 sampling sessions were conducted
across 18 river transects in primary forest habitat and 34 sampling sessions were conducted
across 17 transects in secondary forest habitat.
Interior Forest: Forest transect sites were selected in areas that had been characterized by
a team of ornithologists as either primary or secondary; this ensured sampling was conducted in
both forest habitat types equally (Appendix 1). Sixteen transects were set up within 6 primary
forest sites and sixteen transects were set up within 6 secondary forest sites. Transects ran
perpendicular and parallel to walking trails, at random distances within the sites identified by the

7
team of ornithologists. There were a total of 32 sampling sessions in primary forest and 32
sampling sessions in secondary forest.
Road: To investigate the influence of ongoing human disturbances three different road
categories were identified and sampled: (1) roads bordered by secondary forest on both sides (16
sample sessions along 8 transects), (2) roads with secondary forest on one side and pasture land
on the other (14 sample sessions along 7 transects), and (3) roads with pasture land on both sides
(17 sample sessions along 8 transects). The roads were unpaved and inaccessible by vehicles
approximately ten months of the year due to the muddy conditions.
Figure 1: Three maps showing where Ecuador is located in South America, where
the Mache Chindul Reserve is located in Ecuador and where Bilsa (where the study
was conducted) is located within the Mache Chindul Reserve.
Sampling Scheme
Amphibian sampling occurred from March to November 2007. For all habitats (rivers,
interior forest, roads), sampling was conducted using visual encounter surveys (VES) along
transects (Heyer et al. 1994). This involved walking slowly along 50 meter transects, intensively
searching with a stick through all leaf-litter and on all leaves and stems within transects for
amphibians. Transects in interior forest were 2m wide, while river transects were as wide as the

8
river-bank (1-3m) and road transects were as wide as the road-side (1-4m). Each transect was
laid-out at least two weeks prior to sampling to avoid any effects of disturbance. Each transect
was sampled twice: once in the morning and again at night. All amphibians encountered during
morning sampling sessions were captured and only released after the night sampling session was
complete. In addition to capturing individuals encountered during the morning sample session,
the location of each transect was changed each month. Employing this approach is a less
invasive way to avoid re-sampling individuals than toe-clipping, which many studies have shown
to negatively impact frogs (Davis and Ovaska 2001; McCarthy and Parris 2004; Waddle, et al.
2008). Changing the location of transects each month was also important to avoid modifying the
habitat along transects caused by repeated sampling in the same area. This is particularly an
issue in the tropics where the forest floor very readily turns to mud when disturbed.
Four river transects were sampled twice each month (two in secondary forest habitat; two
in primary forest habitat) for a total of 36 sampling sessions along rivers in primary forest and 34
along rivers in secondary forest (sampling along rivers in secondary forest was missed one
month due to health complications). River banks, defined as the area between the water and the
forest edge, were intensively searched for amphibians along both sides of the river for 50 m. All
leaves and branches overhanging the river banks (as high as 3.5 m) were also searched at each
river transect. Similarly, four interior forest transects were sampled twice each month (two in
primary forest; two in secondary) for a total of 18 transects in each habitat. For the forest
transects, two 50 m lengths of string were laid down in parallel rows two meters apart and all
leaf litter, logs, branches and leaves inside the two strings were searched for amphibians. Each
of the three road habitat categories was sampled twice each month.

9
Environmental Variables
For each transect, the weather conditions, number of people searching, and time spent
sampling were recorded. For each frog encountered, the species, age classification (adult,
adolescent, juvenile), and substrate were recorded during the sampling session. Depending on
the habitat, the appropriate environmental variables were recorded.
Environmental variables recorded along river transects included: bank-width, water-
width, water velocity, depth, understory density (1) over the bank and (2) over the water. All of
these variables were measured at 10 m intervals along each transect and then averaged. Water
velocity was measured by dropping a ping-pong ball in the river and timing how long it took to
travel one meter. Relative understory density was obtained by averaging the number of contacts
of vegetation (leaves and branches) with a 2 m tall stick placed vertically every 10 m along a
transect. This was measured at the edge of the forest and the edge of the water. The number of
waterfalls and pools was also recorded for each river transect. Leaf-litter depth and understory
density were measured at each forest transect at 10 m intervals and averaged. The road side
width, road wetness, type of vegetation and average vegetation height were recorded for each
road transect. Vegetation type was categorized as either grass or shrubs. Road wetness was
ranked on a scale of 0-5, with 0 being completely dry and 5 being more water than road.
Species Identification
During the first two months of amphibian sampling, all individuals encountered along
transects were captured and photographed for subsequent identification. Photographs were
shared with herpetologists in the US and Ecuador who had previous experience identifying
amphibians in the tropics. Dr. Greg Vigle has studied herpetofauna in Western Ecuador for the

10
past 25 years and carried out the first herpetological inventory of Bilsa in 2000. Dr. Vigle
provided help identifying amphibians encountered during this study via email. Marcelo Escobar
from the Natural History Museum in Quito has made numerous trips to Bilsa and is working to
create a complete herpetological inventory for the Mache-Chindul. Mr. Escobar offered
invaluable support in identifying the amphibians encountered over the course of this study. So,
although little work had been done on the assemblages of amphibians in altered versus pristine
forests in Bilsa, sufficient inventory work had been done in the park to identify the species
encountered during the course of this study with confidence.
Statistical Analysis
To examine how complete sampling was in different types of habitat and to compare species
richness across habitat types while controlling for differences in sample size, expected species
accumulation curves (also called ‗sample-based rarefaction curves‘, in which a sample is one sampling
occasion for a given station), were computed using the analytical formulas of Colwell et al. (2004) as
implemented in EstimateS v. 8 (Colwell 2006). Virtually any sampling methodology is imperfect in
completely detecting all the species present in a site, and several indices have been developed to estimate
―true‖ species richness based on sampling data (Colwell and Coddington 1994). As an estimation of the
total number of species present in each habitat category, the mean and range of four commonly employed
abundance-based estimators (ACE, Chao1, Jack1 and Boostrap), were calculated using EstimateS v. 8 and
based on 1,000 randomizations of samples without replacement. Two diversity indices were calculated
using EstimateS v. 8: Shannon-Wiener and Simpson`s. The Shannon-Wiener index places more weight on
rare species; conversely, Simpson`s index places less weight on rare species (Krebs 1999).

11
Results
Species Abundance and Diversity
In total 933 individuals were recorded: 613 along river and forest transects, and 320
along road transects (Table 1). Pristimantis achatinus dominated six of the seven habitat
categories and comprised more than half of the total of the individuals encountered during the
study. Hyloxalus awa was the most abundant species encountered along primary rivers and was
the second most abundant along secondary rivers.

12
Table 1: Number of individual amphibians recorded across seven habitat categories in
Bilsa Biological Station, Ecuador.
A sample-based rarefaction curve shows the accumulation curve of each habitat category
(Figure 2). In Bilsa Biological Station river habitat harboured the highest species richness
according to the Mau Tau index. Primary forest habitat harboured the third highest richness
levels, followed by the road habitat category, Pasture-Pasture. The latter two habitat categories
also had the steepest accumulation curves, which suggests that there are still more species to be
Forest River Road
Species Total 1° 2° 1° 2° Pasture 2° Mix
Bolitoglossa Spp. 1 0 0 0 1 0 0 0
Chaunus marinus 1 0 0 0 0 0 1 0
Caecilia leucocephala 1 0 0 1 0 0 0 0
Caecilia nigricans 1 0 0 0 0 1 0 0
Cochranella cf. albomaculata 4 0 0 2 2 0 0 0
C. Mache 2 0 0 2 0 0 0 0
C. Spinosa 1 0 0 1 0 0 0 0
Centrolene prosoblepon 37 0 0 23 14 0 0 0
Craugastor longirostris 8 1 2 2 2 1 0 0
Epipedobates boulengeri 27 1 0 16 10 0 0 0
Hyloxalus awa 114 0 0 79 35 0 0 0
H. sp. nov. 45 3 0 21 21 0 0 0
Hypsiboas pellucens 1 0 0 0 0 1 0 0
H. picturata 48 0 0 18 30 0 0 0
Leptodactylus labrosus 2 0 0 0 0 1 1 0
L. pentadactylus 1 1 0 0 0 0 0 0
Oophaga sylvatica 20 18 0 1 1 0 0 0
Pristimantis achatinus 535 50 97 45 39 91 114 99
P. parvillus 3 1 2 0 0 0 0 0
P.subsigillatus 2 0 0 0 0 1 0 1
P. walkeri 4 0 3 0 0 0 0 1
Rhinella margaritifer 37 0 0 9 28 0 0 0
Rhaebo haematiticus 31 0 0 19 12 0 0 0
Smilisca phaeota 7 0 0 0 0 1 3 3
Total 933 75 104 239 195 97 119 104
Primary habitat is represented by 1°, secondary habitat by 2°.

13
0
2
4
6
8
10
12
14
16
0 50 100 150 200 250 300
No. individuals
No
. s
pe
cie
s
River, Primary
River, Secondary
Forest, Primary
Forest, Secondary
Road, Secondary-Secondary
Road, Secondary-Pasture
Road, Pasture-Pasture
sampled in these habitats, particularly along roads surrounded by pasture. The remaining three
habitats had lower and similar richness levels.
Figure 2: Expected species accumulation curves for amphibians captured in different habitat
categories in Bilsa Biological Station, according to the Mao Tau index. The vertical hatched line
indicates the minimum abundance level recorded for comparison across habitat types. Curves were
re-scaled to show accumulation of species in relation to number of captured individuals.
Secondary river habitat had the highest diversity (Shannon-Wiener and Simpson‘s),
followed by primary river habitat (Table 2). Primary forest had higher amphibian diversity than
secondary forest. Road bordered by pasture had the highest diversity of the three road-categories.
The expected species richness (based on the mean of ACE, Chao1, Jack1, Bootstrap indices) was
most striking for the pasture-road habitat.

14
Table 2: Total individuals, diversity (Shannon-Wiener and Simpson’s respectively),
observed and expected species richness for the seven habitat categories.
Richness Diversity Total Ind.
Obs Expected Shan-W Simpson’s
Forest
Primary 7 13.1 (8.6-19.9) 0.9 2.0 75
Secondary 4 4.4 (4.0-5.0) 0.3 1.2 104
River Primary 14 17.2 (15.5-19.5) 2.0 5.7 239
Secondary 12 14.2 (13.0-15.5) 2.1 7.4 195
Road
Pasture 7 16.4 (9.1-22.0) 0.3 1.1 97
Secondary 4 5.8 (4.8-7.7) 0.2 1.1 119
Mix 4 5.8 (4.8-7.7) 0.2 1.1 104
Number of species observed (Obs), expected species richness (Expected) is the mean of four
common species richness indices (ACE, Chao1, Jack1, Bootstrap), Shannon-Wiener diversity index
(Shan-W), and Simpson‘s diversity index (Simpson‘s).
Environmental Variables
The mean understory density (USD) in primary forest (1.6) was much lower than the mean USD
of secondary forest (2.8) (Two sample t-test, p = 0.005). There was no significant difference
between any environmental variable in primary and secondary forest. Vegetation-height along
roads in pasture land was significantly lower than along roads in secondary forest or roads
bordered by pasture and secondary forest (One-way ANOVA, p = 0.052). No environmental
variable recorded had a significant correlation with diversity. Descriptive statistics (mean,
standard error, standard deviation, skewness, sample size) for each habitat category are included
in Appendix 3.

15
Discussion
Historical Disturbance
Throughout the tropics, alteration of forest habitat by logging and fragmentation
commonly results in reduced amphibian diversity and species richness (Pearman 1997;
Krishnamurthy 2003; Walting and Donnelly 2008). The results from this study show that the
influence of disturbance on the diversity and composition of amphibian assemblages in the
Mache-Chindul varies with habitat. Amphibian assemblages along rivers were the most diverse
as well as the least impacted by human activity in the form of logging 12-18 years prior to this
study. Diversity and species richness were lower in secondary than in primary forest suggesting
that amphibian assemblages in interior forest habitat are more vulnerable to disturbances caused
by logging.
Avian diversity sampled at the same forest sites in Bilsa exhibited the reverse response
and had higher diversity in secondary forest than in primary forest (Karubian unpublished). This
demonstrates how different groups of animals can respond very differently to disturbance and
suggests that amphibians in the Mache-Chindul are more negatively impacted by human
activities, like logging, than birds, in terms of overall diversity and species richness. All forest
sites in this study were part of a mosaic of habitat with direct contact zones between primary and
secondary forest. This type of setting makes it possible for amphibian populations in primary
forest to serve as a source in repopulating secondary forest following a major disturbance like
logging. Despite this, there was still a significant difference in diversity between primary and
secondary forest. What is more alarming about the apparent vulnerability of interior-forest
amphibian assemblages is that one could expect to find even lower amphibian diversity in
isolated secondary forests lacking available source populations.

16
The different responses between forest and river habitats may be explained by regular
disturbances along river-banks caused by rising water-levels during the wet season. This natural
disturbance results in a similar forest structure growing along both primary and secondary rivers.
This was demonstrated by the identical understory densities (USD) in both secondary and
primary river habitat. Conversely, mean understory densities differed greatly between primary
and secondary forest. Amphibian diversity richness is influenced by a number of environmental
variables (canopy cover, soil moisture, distance to nearest stream, invertebrate density) (Urbina-
Cardona et al. 2006). Recording more of these variables during this study may have helped
identify necessary microhabitat characteristics that support greater amphibian diversity or
richness at Bilsa.
Ongoing Disturbance
Road habitat around Bilsa harboured a very different amphibian assemblage from that of
forest and river habitats and of the nine species encountered along roads six were found in no
other habitats. Typically, amphibian assemblages in pasture land have lower diversity and
species richness (Tocher 1996; Pearman 1997, Urbina-Cardona et al. 2006) but roads in pasture
land surrounding Bilsa actually harbour the greatest species richness of the three road-categories.
Road wetness was similar across all road categories; however, vegetation height was
significantly shorter in pasture-road habitat. It is possible that species went undetected in
secondary-road and mixed-road habitats, despite rigorous sampling efforts.
Continued sampling in this habitat is needed to more accurately determine the actual
diversity and species richness. Additional sampling should be done in pasture away from roads
to determine whether the species encountered during this study are restricted to roads or are
widespread throughout pasture lands. This information, in combination with pasture diversity in

17
relation to distance from forest edge, will help gauge the overall amphibian diversity in the
Mache-Chindul as more and more forests are converted to pasture.
Amphibian Detection
Species detectability as defined by Boulinier et al (1998) is the probability of detecting at least
one individual of a given species in a particular sampling effort, given that individuals of that
species are present in the area of interest during the sampling session. This is a crucial factor to
consider for any study involving species richness or diversity estimates. Naturally, not all
species are equally detectable. Additionally, not all species are equally detectable across
different habitats. Fortunately, some species richness estimators take detectability into account.
Abundance-based Coverage Estimator (ACE) for example, separates common and rare species
from a sample and uses the rare subset to predict how many species are likely missing due to
lowered detectability (Chazdon 1998). The issue surrounding detectability is highlighted by
canopy dwelling species in the tropics.
Canopy dwelling species deserve more consideration when assessing amphibian diversity
in the tropics. Utilizing bromeliads as high-rise nurseries, some species may never have to
descend from the canopy and thus would never have been encountered in this study.
Consequently the true amphibian diversity of the forest may be under-estimated. Two species in
Bilsa exemplify the real possibility that amphibian assemblages, particularly frogs, could occupy
the canopy without ever descending.
Cochranella albomaculata, has only been encountered four times in Bilsa and each
encounter was of a mating-pair on a leaf overhanging a river (1-2 m above the water). C.
albomaculata is commonly encountered low on vegetation along rivers in Central America

18
(Savage 2002). Guayasamin (2007) proposed that what is currently identified as C.
albomaculata may actually be several species, as individuals from Columbia and Ecuador
exhibit larger spots on the dorsum than in Central America. These isolated encounters in Bilsa
support the possibility that C. albomaculata represents different species in Ecuador and, in Bilsa
at least, appear to have taken to the canopy, only returning to lower strata of the forest to
procreate. Only during mating was it possible to account for this species and it is likely that
many individuals were not recorded because they were too high in the canopy.
Oophaga sylvatica, is a ground-dwelling frog that uses the water trapped in bromeliads
for rearing its tadpoles. Secondary forest supports far fewer epiphytes than primary forest
(Turner 1997) which may explain why O. sylvatica was never encountered in secondary forest
during this study.
The combination of O. sylvatica‘s use of bromeliads and C. albomaculata‘s canopy-
dwelling life-style demonstrates that amphibian assemblages may exist undetected in the canopy
of primary forest, or in forest where sufficient epiphyte growth has occurred. As such, efforts to
detect canopy-dwelling amphibians should be undertaken more often when assessing diversity in
the tropical forests. This theory is supported by the discovery of Ecnomiohyla phantasmagoria
in Bilsa in 2009 (Patricio M. Olmedo, unpublished. See photograph in Appendix 4). E.
phantasmagoria is a canopy-dwelling species and it is likely that is was never detected during
this study or any previous ones, because of this. New developments in bioacoustics recording
devices may be the best solution to surveying amphibians in the canopy.
Amphibians as Indicator Species
At Jatun Sacha Biological Station in Amazonian Ecuador, Pearman (1997) found that

19
species richness of the genus Pristimantis declined with proximity to pasture and suggested that
Pristimantis may be a useful indicator assemblage for assessing the quality of Tropical Wet
Forest. The Pristimantis spp. at Bilsa Biological Station, however, exhibit the greatest richness
in pasture and secondary forest habitat. All Pristimantis species encountered in and around Bilsa
but one (P. achatinus) were very rare. These findings suggest that Prismantis has little value as
an indicator assemblage for assessing the quality of Pre-Montane or Humid Tropical Forest in
the Mache-Chindul.
Four species (Cochranella mache, C. spinosa, Caecilia leucocephala, and Leptodactylus
pentadactylus) encountered during this study were found exclusively in pristine habitat (primary
forest or river). All of these species were only encountered once or twice over the course of nine
months, thus making them poor indicator species. Oophaga sylvatica, however was encountered
18 times in pristine habitat and never in secondary forest habitat and may be a good candidate for
an indicator of pristine forest habitat in the Mache-Chindul since it (1) was the most abundant
amphibian found solely in pristine forest during this study, (2) is easily identified, (3) vocal
during the day and (4) is wide-spread in the region. All of these traits make its detection easy
and feasible. Surveys for the presence of O. sylvatica, in other pristine (and disturbed) forest
habitat across its range in the Tumbes-Chocó-Magdalena are needed before adopting this species
as a suitable indicator of quality pristine forest habitat.
Conservation Implications
The results of this study in Bilsa Biological Station show that amphibian assemblages
vary markedly in their vulnerability to disturbances by human activity. Recognising these
differences is critical for developing land management plans that promote amphibian diversity in

20
north western Ecuador.
The resilience of riparian amphibian assemblages to historical human activity has strong
implications for the conservation and restoration of amphibian diversity along rivers in Mache-
Chindul and possibly throughout the tropics. Conserving existing forest along rivers and
promoting forest growth along rivers in deforested areas has potential, not only to encourage and
sustain amphibian diversity, but also to provide corridors connecting fragmented interior forest
amphibian populations. The extent of forest surrounding a river necessary to support diversity
levels similar to those in Bilsa as well as to serve as an effective corridor is still unknown and
will require further investigation.
The results of this study show that interior forest amphibian assemblages in Bilsa are
more vulnerable to disturbance than are riparian amphibian assemblages. Pristine, unlogged
forest harboured greater species richness and diversity than secondary forest. In order to
conserve intact interior-forest amphibian assemblages any remaining remnant forest must be
protected. For successful amphibian conservation to be achieved more research is needed into
amphibian responses to habitat size in order to create land-management plans that will sustain
amphibians well into the future.

21
Literature Cited
Bouliner, T., J. D. Nichols, J. R. Sauer, J. E. Hines, K. H. Pollock. 2005 Estimating species
richness : the importance of heterogeneity in species detectability. Ecology 79:1018-1028.
Chazdon, R. L., R. K. Colwell, J. S. Denslow, & M. R. Guariguata. 1998. Statistical methods for
estimating species richness of woody regeneration in primary and secondary rain forests of NE
Costa Rica. Parthenon Publishing, Paris.
Collins, J. P., C. Gascon, J.R. Medelson III. 2007. Amphibian Action Conservation Plan.
Preceedings of the IUCN/SSC Amphibian Conservation Summit in 2005.
Collins, J. P., T. Halliday. 2005. Forecasting changes in amphibian biodiversity: aiming at a
moving target. Philosophical Transactions of the Royal Society of London 360:309-214.
Colwell R.K., AND Coddington J.A. (1994) Estimating terrestrial biodiversity through
extrapolation. Philosophical Transactions of the Royal Society of London 345:101-118.
Colwell R.K., Mao C.X. & Chang J. (2004) Interpolating, extrapolating, and comparing
incidence-based species accumulation curves. Ecology 85:2717-2727.
Colwell, R. K. 2006. EstimateS: Statistical estimation of species richness and shared species
from samples. Version 8. Persistent URL <purl.oclc.org/estimates>
Conservation International. http://web.biodiversityhotspots.org/xp/Hotspots/tumbes_choco/bio
diversity.xml). Access date: 18 January 2009.
Davis, T. M., K. Ovaska. 2001. Individual Recognition of Amphibians: Effects of Toe Clipping and
Fluorescent Tagging on the Salamander Plethodon vehiculum. Journal of Herpetology 35:217-225.
Dodson, C. H., and A.H. Gentry. 1991. Biological extinction in western Ecuador. Annals of the
Missouri Botanical Garden 78:273-295.
Hamilton, P., C. Mouette, A. Almendariz. Initial analysis of coastal Ecuadorian herpetofauna of
dry and moist forests. Reptile and Amphibian Ecology International. Unpublished.
Hickman, C. P., L. S. Roberts, A. Larson, H. I'Anson. 2008. Integrated Principles of Zoology.
Fourteenth edition. McGraw-Hill Publishing Co., New York.
Heyer, W.R, M. A. Donnelly, R. W. McDiarmid, L. C. Hayek, M. S. Foster. 1994. Measuring
and Monitoring Biological Diversity: Standard Methods for Amphibians. First Edition.
Smithsonian.
Krebs, C. J. 1999. Ecological Methodology. Second Edition. Benjamin Cummings, New York.

22
McCarthy, M. A., K. M. Parris. 2004. Clarifying the effect of toe clipping on frogs with
Bayesian statistics. Journal of Applied Ecology 41:780-786.
Mittermeier, R. A., N. Myers, J. B. Thomsen, G. A. B. da Fonseca, S. Olivieri. 1998.
Biodiversity hotspots and major tropical wilderness areas: approaches to setting conservation
priorities. Conservation Biology 12:516-520.
Parker III, T.A., and J. L. Carr. 1992. Status of forest remnants in the Cordillera de la Costa and
adjacent areas of southwestern Ecuador. Conservation International RAP Working Papers 2:172.
Pearman, P.B. 1997. Correlates of amphibian diversity in an altered landscape of Amazonian
Ecuador. Conservation Biology 11:1211-1225.
Roelants, K., S. J. Gower, M. Wilkinson, S. P. Loader, S. D. Biju, K. Guillaume, L. Moriau, F.
Bossuyt. 2007. Global patterns of diversification in the history of modern amphibians.
Proceedings of the National Academy of Science 104:887-892.
Savage, M. J. 2002. The amphibians and reptiles of Costa Rica: a herpetofauna between two
continents, between two seas. University of Chicago Press.
Tocher, D. M. 1996. The effects of deforestation and forest fragmentation on a central
Amazonian frog community. University of Canterbury Press.
Turner, I.A., Y. K. Wong, P. T. Chew, A. Bin Ibrahim. 1997. Tree species richness in primary
and old secondary tropical forest in Singapore. Biodiversity and conservation 6:537-543.
Urbina-Cardona, J. N., M. Olivares-Pe´rez, V. H. Reynoso. 2005. Herpetofauna diversity and
microenvironment correlates across a pasture–edge–interior ecotone in tropical rainforest
fragments in the Los Tuxtlas Biosphere Reserve of Veracruz, Mexico. Biological Conservation
132:61-75.
Waddle, J. H., K. G. Rice, F. J. Mazzotti, H. F. Percival. 2008. Modeling the effect of toe
clipping on treefrog survival: beyond the return rate. Journal of Herpetology 42:467-473.
Wake, D. B., and V. T. Vredenburg. 2008. Are we in the midst of the sixth mass extinction? A
view from the world of amphibians. Proceedings of the National Academy of Science
105:11466-11473.
Watling, J. I., M. A. Donnelly. 2008. Species richness and composition of amphibians and
reptiles in a fragmented forest landscape in northeastern Bolivia. Basic and Applied Ecology
9:523–532.
Wilson, O. E., 1992. The Diversity of Life. W. W. Norton & Company. New York.
Young, B. E., K. R. Lips, J. K. Reaser, R. Ibanez, A. W. Salas, J. R. Cedeno, L. A. Coloma, S.
Ron, E. L. Marca, J.R. Meyer, A. Munoz, F. Bolanos, G. Chaves, D. Romo. 2001. Population

23
declines and priorities for amphibian conservation in Latin America. Conservation Biology
15:1213-1223.

24
Appendix 1
Quoted from Jordan Karubian’s methods (unpublished):
Forest type characterization
Along >15 km of trails that cover about a third of the Bilsa Station area, we establish sampling
points every 200 m (total: 84 points) where we visually classified the forest type around it as
primary (48 points), altered (16 points), or secondary (20 points, Fig. 1). To confirm if our visual
classification reflected actual differences in habitat structure, in each of these points we recorded:
the number of trees with DBH >10 but < 50 cm (within a 10 m-radius circle), number of trees
with DBH ≥ 50 cm (within a 20 m-radius circle), canopy height (in meters), amount of light
reaching the understory (measured with a densiometer, average of four measures), and elevation
(in meters). We then used a Discriminant Analysis (DA) to assess if sites could be confidently
assigned to forest type categories based on these structural variables.

25
Appendix 2
Table 1: Species richness results, calculated using Estimate S program: ACE, Chao 1, Jack 1 and
Bootstrap.
Habitat River Forest Road
Category Primary Secondary Primary Secondary Pasture Forest Mix
Samples 36 34 32 32 17 16 14
Individuals (computed) 239 195 75 104 97 119 104
Sobs (Mao Tau) 14 12 7 4 7 4 4
Sobs 95% CI Lower Bound 10.58 8.49 3.24 3.06 2.76 1.79 1.79
Sobs 95% CI Upper Bound 17.42 15.51 10.76 4.94 11.24 6.21 6.21
Sobs SD (Mao Tau) 1.74 1.79 1.92 0.48 2.16 1.13 1.13
Sobs Mean (runs) 14 12 7 4 7 4 4
ACE Mean 19.51 15.54 19.89 4 22 7.67 7.67
ACE SD (runs) 0 0
0 0
Chao 1 Mean 15.5 13 13 4 22 5 5
Chao 1 95% CI Lower Bound 14.17 12.09 7.96 4 10.32 4.08 4.08
Chao 1 95% CI Upper Bound 26.89 23.06 44.4 4 74.78 17.27 17.27
Chao 1 SD (analytical) 2.29 1.87 7.08 0 13.48 2.17 2.17
Jack 1 Mean 17.89 14.91 10.88 4.97 12.65 5.88 5.86
Jack 1 SD (analytical) 1.86 1.63 1.84 0.97 3.34 1.28 1.26
Bootstrap Mean 15.74 13.24 8.58 4.62 9.14 4.83 4.82
Bootstrap SD (runs) 0
0
0
26
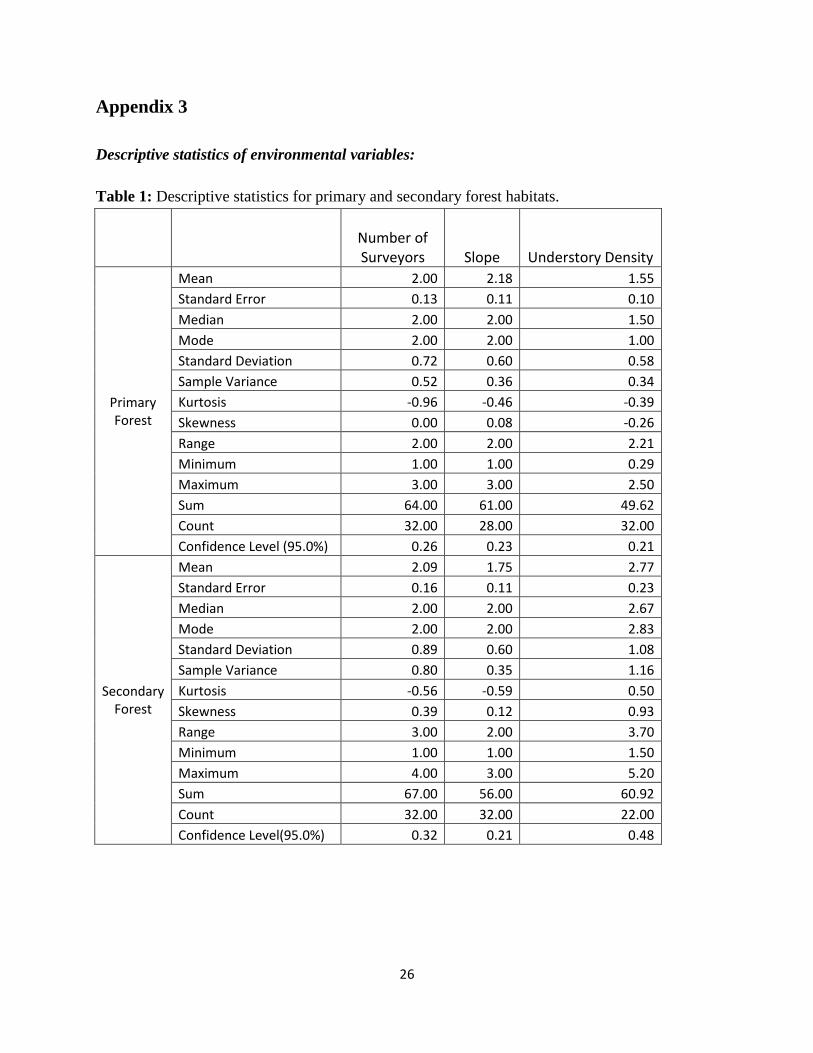
Appendix 3
Descriptive statistics of environmental variables:
Table 1: Descriptive statistics for primary and secondary forest habitats.
Number of Surveyors Slope Understory Density
Primary Forest
Mean 2.00 2.18 1.55
Standard Error 0.13 0.11 0.10
Median 2.00 2.00 1.50
Mode 2.00 2.00 1.00
Standard Deviation 0.72 0.60 0.58
Sample Variance 0.52 0.36 0.34
Kurtosis -0.96 -0.46 -0.39
Skewness 0.00 0.08 -0.26
Range 2.00 2.00 2.21
Minimum 1.00 1.00 0.29
Maximum 3.00 3.00 2.50
Sum 64.00 61.00 49.62
Count 32.00 28.00 32.00
Confidence Level (95.0%) 0.26 0.23 0.21
Secondary Forest
Mean 2.09 1.75 2.77
Standard Error 0.16 0.11 0.23
Median 2.00 2.00 2.67
Mode 2.00 2.00 2.83
Standard Deviation 0.89 0.60 1.08
Sample Variance 0.80 0.35 1.16
Kurtosis -0.56 -0.59 0.50
Skewness 0.39 0.12 0.93
Range 3.00 2.00 3.70
Minimum 1.00 1.00 1.50
Maximum 4.00 3.00 5.20
Sum 67.00 56.00 60.92
Count 32.00 32.00 22.00
Confidence Level(95.0%) 0.32 0.21 0.48

27
Table 2: Descriptive statistics for primary and secondary river habitats.
Surveyors
(#) Slope USD
Forest USD
water Width Bank
Width Water Pool
Waterfalls
Depth Avg (cm)
Velocity
1 River
Mean 2.47 2.72 2.91 1.28 4.63 2.51 2.39 1.39 24.95 0.19
Standard Error 0.22 0.10 0.13 0.13 0.23 0.11 0.35 0.17 2.65 0.02
Standard Deviation 1.34 0.59 0.75 0.78 1.07 0.68 2.09 1.02 15.87 0.12
Sample Variance 1.80 0.35 0.56 0.61 1.15 0.47 4.36 1.04 251.99 0.01
Kurtosis 3.60 0.32 -0.78 0.48 -0.31 0.24 -
1.17 -1.21 -0.93 5.04
Skewness 1.68 0.80 0.27 0.87 -0.90 0.57 0.45 -0.19 0.73 2.24
Range 6.00 2.00 2.62 2.75 3.40 2.62 6.00 3.00 46.20 0.50
Minimum 1.00 2.00 1.58 0.25 2.40 1.54 0.00 0.00 6.80 0.09
Maximum 7.00 4.00 4.20 3.00 5.80 4.16 6.00 3.00 53.00 0.59
Sum 89.00 98.00 104.8
8 46.22 101.9
4 90.26 86.0
0 50.00 898.20 6.92
Count 36.00 36.00 36.00 36.00 22.00 36.00 36.0
0 36.00 36.00 36.00
Confidence Level(95.0%) 0.45 0.20 0.25 0.26 0.47 0.23 0.71 0.35 5.37 0.04
Mean 2.06 2.50 2.92 1.35 4.28 1.65 1.65 0.18 15.72 0.18
2 River
Standard Error 0.20 0.07 0.15 0.13 0.32 0.11 0.33 0.07 1.95 0.01
Standard Deviation 1.15 0.39 0.88 0.77 1.51 0.66 1.91 0.39 11.35 0.06
Sample Variance 1.33 0.15 0.77 0.60 2.27 0.44 3.63 0.15 128.77 0.00
Kurtosis 3.85 1.06 -0.77 -1.16 1.28 -0.16 -
1.20 1.23 -0.66 -0.01
Skewness 1.77 0.82 0.11 0.35 1.13 0.38 0.65 1.78 0.87 0.88
Range 5.00 1.50 3.00 2.34 5.58 2.53 5.00 1.00 34.80 0.22
Minimum 1.00 2.00 1.42 0.33 2.30 0.61 0.00 0.00 3.50 0.09
Maximum 6.00 3.50 4.42 2.67 7.88 3.14 5.00 1.00 38.30 0.31
Sum 70.00 85.00 99.26 45.82 94.08 56.22 56.0
0 6.00 534.34 6.04
Count 34.00 34.00 34.00 34.00 22.00 34.00 34.0
0 34.00 34.00 34.00
Confidence Level(95.0%) 0.40 0.14 0.31 0.27 0.67 0.23 0.66 0.14 3.96 0.02

28
Table 3: Descriptive statistics for pasture, forest and mixed road habitats.
Surveyors (#) Slope Mud
Veg. Height
Road Side
Width
Pasture
Mean 2.59 1.17 2.00 41.62 353.33
Standard Error 0.32 0.17 0.35 14.10 30.26
Median 2.00 1.00 2.00 20.00 450.00
Mode 2.00 1.00 1.50 20.00 450.00
Standard Deviation 1.33 0.41 1.26 50.83 117.21
Sample Variance 1.76 0.17 1.58 2583.26 13738.10
Kurtosis 1.86 6.00 -0.28 2.97 -1.03
Skewness 1.43 2.45 0.07 2.09 -0.71
Range 5.00 1.00 4.00 142.00 300.00
Minimum 1.00 1.00 0.00 13.00 150.00
Maximum 6.00 2.00 4.00 155.00 450.00
Sum 44.00 7.00 26.00 541.00 5300.00
Count 17.00 6.00 13.00 13.00 15.00
Confidence Level(95.0%) 0.68 0.43 0.76 30.71 64.91
Pasture, Forest
Mean 2.14 1.42 2.00 49.40 314.29
Standard Error 0.25 0.14 0.41 12.86 12.21
Median 2.00 1.25 1.75 35.00 350.00
Mode 2.00 1.00 2.00 30.00 350.00
Standard Deviation 0.95 0.47 1.31 40.65 45.69
Sample Variance 0.90 0.22 1.72 1652.71 2087.91
Kurtosis -0.69 -1.93 -0.58 1.09 -1.56
Skewness 0.31 0.38 0.41 1.60 -0.66
Range 3.00 1.00 4.00 108.00 100.00
Minimum 1.00 1.00 0.00 17.00 250.00
Maximum 4.00 2.00 4.00 125.00 350.00
Sum 30.00 17.00 20.00 494.00 4400.00
Count 14.00 12.00 10.00 10.00 14.00
Confidence Level(95.0%) 0.55 0.30 0.94 29.08 26.38
Forest
Mean 2.21 1.22 1.50 55.89 170.77
Standard Error 0.24 0.15 0.22 13.67 29.04
Median 2.00 1.00 2.00 40.00 125.00
Mode 2.00 1.00 2.00 30.00 125.00
Standard Deviation 0.89 0.44 0.71 41.00 104.70
Sample Variance 0.80 0.19 0.50 1680.86 10961.86
Kurtosis 1.00 0.73 0.57 -0.49 2.51
Skewness 1.04 1.62 -1.18 0.76 1.95

29
Range 3.00 1.00 2.00 117.00 300.00
Minimum 1.00 1.00 0.00 3.00 100.00
Maximum 4.00 2.00 2.00 120.00 400.00
Sum 31.00 11.00 15.00 503.00 2220.00
Count 14.00 9.00 10.00 9.00 13.00
Confidence Level(95.0%) 0.52 0.34 0.51 31.51 63.27

30
Appendix 4


















