
Genotypic and Phenotypic Analysis of HIV Type 1 Primary ...
12
AIDS RESEARCH AND HUMAN RETROVIRUSES Volume 18, Number 1, 2002, pp. 39–48 Mary Ann Liebert, Inc. Genotypic and Phenotypic Analysis of HIV Type 1 Primary Isolates from Western Cameroon DENIS M. TEBIT, 1,2 LÉOPOLD ZEKENG, 3 LAZARE KAPTUÉ, 3 MIKA SALMINEN, 4 HANS-GEORG KRÄUSSLICH, 1,2 and OTTMAR HERCHENRÖDER 1,5 ABSTRACT In this study we report the molecular and biological characteristics of 19 HIV-1 primary isolates obtained in April 1999 from 47 HIV-1-infected individuals living mainly in western Cameroon. Discontinuous portions of gag, pol, and env were amplified by polymerase chain reaction and directly sequenced. Phylogenetic analysis of these sequences showed that all were of HIV-1 group M with the following genotypes: A gag /A pol /A env (n 5 4), A gag /AG pol /AG env (n 5 2), AG gag /A pol /AG env (n 5 1), AG gag /U pol /AG env (n 5 1), AG gag /AG pol /AG env (n 5 6), G gag /G pol /G env (n 5 3), F2 gag /F2 pol /F2 env (n 5 1), and a novel A gag /J pro/rt /A int /U env complex recombinant (n 5 1). This A/J/U recombinant shared the same gag–pol cross-over point with known CRF02.AG viruses and 99CMBD6, an AG recombinant from our panel of isolates. The biological phenotype of most of the isolates correlated with the clinical status of the patient. Six isolates were syncytium inducing (SI) on MT-2 cells whereas 13 isolates were of the non-syncytium-inducing phenotype (NSI). Coreceptor usage by these isolates determined on GHOST cells correlated with their biological phenotype, as all SI isolates used CXCR4 and all NSI isolates used CCR5. Our results show a high predominance of subtype A (mainly CRF02.AG-like viruses) in western Cameroon and fewer HIV-1 subtypes compared with other parts of Cameroon. Genetic variabil- ity was, however, not reflected in the biological characteristics of the isolates. The presence of a novel A/J/U complex recombinant from this region further emphasizes the role of recombination in the global evolution of HIV. 39 INTRODUCTION A MAJOR CHARACTERISTIC of human immunodeficiency viruses (HIV) is their high genetic variability. This led to the classification of HIV-1 into three groups based on nucleo- tide sequence analyses: M (major), O (outlier), and N (non M/non O). 1 Whereas group M viruses are prevalent globally, group O viruses are found mostly in West–Central Africa and group N has so far been detected only in Cameroon. The most recent nomenclature of HIV-1 divides group M into subtypes A–D, F–H, and J–K, with F further subdivided into subsub- types F1 and F2. In addition, there are several groups of circu- lating recombinant forms (CRFs) of HIV-1. 1 CRFs are groups of viruses that are widely distributed and share a common re- combinant lineage. Therefore, different parts of the genomes of CRFs commonly belong to different subtypes, with the cross- over point(s) in each CRF group defined by the recombination event(s). Most of the CRFs described to date contain a portion of subtype A. 1 The high genetic variability of HIV-1 is due to the low fi- delity of the viral reverse transcriptase (RT), which lacks proof- reading activity, and to the high virus turnover in vivo (10 9 viri- ons per day). 2 Retroviruses, including HIV-1, contain two RNA strands of positive polarity, which typically are derived from the same parental provirus. 3 Infected cells can simultaneously harbor two different proviruses, therefore allowing one RNA transcript from each provirus to be encapsidated into a single heterozygous virion. Because RT switches templates during re- verse transcription, such heterozygous virions can give rise to recombinant progeny. 4 In fact, intersubtype recombination has 1 Heinrich-Pette-Institut, D-20251 Hamburg, Germany. 2 Abteilung Virologie, Universität Heidelberg, D-69120 Heidelberg, Germany. 3 University of Yaoundé Teaching Hospital, University of Yaoundé 1, Cameroon. 4 Department of Infectious Disease Epidemiology, National Public Health Institute, Helsinki, Finland. 5 Institut für Virologie, Medizinische Fakultät Carl Gustav Carus, TU Dresden, Germany.
Transcript of Genotypic and Phenotypic Analysis of HIV Type 1 Primary ...

AIDS RESEARCH AND HUMAN RETROVIRUSESVolume 18, Number 1, 2002, pp. 39–48Mary Ann Liebert, Inc.
Genotypic and Phenotypic Analysis of HIV Type 1 PrimaryIsolates from Western Cameroon
DENIS M. TEBIT,1,2 LÉOPOLD ZEKENG,3 LAZARE KAPTUÉ,3 MIKA SALMINEN,4
HANS-GEORG KRÄUSSLICH,1,2 and OTTMAR HERCHENRÖDER1,5
ABSTRACT
In this study we report the molecular and biological characteristics of 19 HIV-1 primary isolates obtained inApril 1999 from 47 HIV-1-infected individuals living mainly in western Cameroon. Discontinuous portions ofgag, pol, and env were amplified by polymerase chain reaction and directly sequenced. Phylogenetic analysisof these sequences showed that all were of HIV-1 group M with the following genotypes: Agag/Apol/Aenv (n 54), Agag/AGpol/AGenv (n 5 2), AGgag/Apol/AGenv (n 5 1), AGgag/Upol/AGenv (n 5 1), AGgag/AGpol/AGenv (n 5 6),Ggag/Gpol/Genv (n 5 3), F2gag/F2pol/F2env (n 5 1), and a novel Agag/Jpro/rt/Aint/Uenv complex recombinant (n 51). This A/J/U recombinant shared the same gag–pol cross-over point with known CRF02.AG viruses and99CMBD6, an AG recombinant from our panel of isolates. The biological phenotype of most of the isolatescorrelated with the clinical status of the patient. Six isolates were syncytium inducing (SI) on MT-2 cellswhereas 13 isolates were of the non-syncytium-inducing phenotype (NSI). Coreceptor usage by these isolatesdetermined on GHOST cells correlated with their biological phenotype, as all SI isolates used CXCR4 and allNSI isolates used CCR5. Our results show a high predominance of subtype A (mainly CRF02.AG-like viruses)in western Cameroon and fewer HIV-1 subtypes compared with other parts of Cameroon. Genetic variabil-ity was, however, not reflected in the biological characteristics of the isolates. The presence of a novel A/J/Ucomplex recombinant from this region further emphasizes the role of recombination in the global evolutionof HIV.
39
INTRODUCTION
AMAJOR CHARACTERISTIC of human immunodeficiencyviruses (HIV) is their high genetic variability. This led to
the classification of HIV-1 into three groups based on nucleo-tide sequence analyses: M (major), O (outlier), and N (nonM/non O).1 Whereas group M viruses are prevalent globally,group O viruses are found mostly in West–Central Africa andgroup N has so far been detected only in Cameroon. The mostrecent nomenclature of HIV-1 divides group M into subtypesA–D, F–H, and J–K, with F further subdivided into subsub-types F1 and F2. In addition, there are several groups of circu-lating recombinant forms (CRFs) of HIV-1.1 CRFs are groupsof viruses that are widely distributed and share a common re-combinant lineage. Therefore, different parts of the genomes of
CRFs commonly belong to different subtypes, with the cross-over point(s) in each CRF group defined by the recombinationevent(s). Most of the CRFs described to date contain a portionof subtype A.1
The high genetic variability of HIV-1 is due to the low fi-delity of the viral reverse transcriptase (RT), which lacks proof-reading activity, and to the high virus turnover in vivo (109 viri-ons per day).2 Retroviruses, including HIV-1, contain two RNAstrands of positive polarity, which typically are derived fromthe same parental provirus.3 Infected cells can simultaneouslyharbor two different proviruses, therefore allowing one RNAtranscript from each provirus to be encapsidated into a singleheterozygous virion. Because RT switches templates during re-verse transcription, such heterozygous virions can give rise torecombinant progeny.4 In fact, intersubtype recombination has
1Heinrich-Pette-Institut, D-20251 Hamburg, Germany.2Abteilung Virologie, Universität Heidelberg, D-69120 Heidelberg, Germany.3University of Yaoundé Teaching Hospital, University of Yaoundé 1, Cameroon.4Department of Infectious Disease Epidemiology, National Public Health Institute, Helsinki, Finland.5Institut für Virologie, Medizinische Fakultät Carl Gustav Carus, TU Dresden, Germany.

been shown to account for more than 10% of all isolates iden-tified and recombinant viruses are increasingly reported in ar-eas harboring more than one HIV-1 subtype.5–7 Furthermore,intergroup recombination between O and M group viruses hasalso been reported.8
The high genetic variability of HIV and its potential for re-combination are clearly important for escape from the immunesystem and also play a major role in the development of resis-tance against antiretroviral drugs. Various drug resistance mu-tations have been described for subtype B isolates of HIV-1,depending on the drug regimen used.9 Some of these mutationshave already been observed in drug-naive individuals as well,but their prevalence and potential for resistance developmentin non-B-type strains of HIV-1 is currently largely unknown.The genetic subtype may influence the biological phenotype ofthe virus. The early phase of HIV-1 infection is dominated bynon-syncytium-inducing (NSI) viruses, which replicate slowly,are macrophage tropic, and use CCR5 as the main coreceptor.10
Later, a switch often occurs to syncytium-inducing (SI) viruses,which replicate rapidly in lymphocytes and T cell lines, and useCXCR4 as the main coreceptor.11 Subtype C viruses appear tobe different from most other subgroups, however, because vir-tually all late C-type isolates have been shown to use predom-inantly CCR5.12
Several previous studies have reported a high genetic vari-ability of subtypes and different recombinant forms of HIV-1from the central and northern parts of Cameroon.6,13,14 The aimof this study was therefore to determine the genetic variability,biological phenotype, and coreceptor usage of primary HIV-1isolates obtained from drug-naive HIV-1-infected individualsmainly in the northwest and southwest provinces of Cameroon.This region lies between central Cameroon, where almost allHIV groups and subtypes have been found, and Nigeria, whereonly a limited number of HIV-1 subtypes circulate.
MATERIALS AND METHODS
Sample collection, serological testing, and virus isolation
Blood was collected from individuals highly suspected of orknown to be infected with HIV in Limbe and Buea (South WestProvince) (n 5 28), Bamenda (North West Province) (n 5 10),and Yaoundé (Center Province) (n 5 9) after obtaining ethicalclearance and informed consent. Blood was screened for HIVantibodies, using an enzyme-linked immunosorbent assay(ELISA; Behring Marburg, Germany), and a larger sample wascollected from HIV-positive individuals. Blood samples werestored at 4°C for a maximum of 7 days (Table 2) and were latershipped at room temperature to Germany. On arrival peripheralblood mononuclear cells (PBMCs) from these samples were pu-rified on Ficoll gradients (Amersham Pharmacia Biotech, Upp-sala, Sweden). After storing an aliquot at 270°C PBMCs werecocultivated with HIV-negative phytohemagglutinin/inter-leukin-2-stimulated PBMCs or with PM-1 cells, a T cell linethat expresses both the CCR5 and CXCR4 coreceptors.15 Bothcultures were maintained by adding growth medium (RPMI1640 with 10–20% fetal calf serum, 2 mM glutamine, and an-tibiotics) twice a week and fresh PBMCs once a week to the
PBMC culture. Cultures were monitored for cytopathic effectsand syncytium formation. Virus growth was analyzed twiceweekly, using a capsid-specific antigen capture ELISA. Posi-tive cultures were harvested and the cell-free supernatants andpellets were stored at 270°C for further analyses.
Biological phenotype and coreceptor usage
Biological phenotypes of all isolates were determined on theHTLV-1-transformed cell line MT-2 as described.16 Briefly,cell-free virus supernatants containing 50 ng of p24 antigenfrom the primary cultures were used to infect 2 3 105 MT-2cells in a 12-well plate and incubated overnight at 37°C. TheMT-2 cells were then washed with phosphate-buffered salineand resuspended in culture medium. MT-2 cultures were testedfor capsid antigen production once a week and monitored forsyncytium formation three times a week.
Chemokine coreceptor usage was determined by infectingGHOST cells stably expressing one of the human chemokinereceptors [CCR1, CCR2b, CCR3b, CXCR4, CCR5, CCR8,BOB, Bonzo, or CX3CR1 (V28)], using cell-free virus super-natants as described.17 Briefly, GHOST cells were plated at 5 3
103 cells/well in 48-well plates and incubated overnight at 37°C.Culture supernatants were removed before addition of cell-freevirus supernatant containing 50 ng of p24 antigen and incubatedovernight at 37°C. Fresh medium was added to a total volumeof 500 ml/well and plates were incubated at 37°C for 3 days.Supernatant was removed and cells were washed, fixed, andimmunostained as described.17
DNA isolation, polymerase chain reaction, andsequence analysis
DNA was extracted from cultured lymphocytes or PM-1 cellswith a Qiagen (Hilden, Germany) DNA extraction kit as rec-ommended by the manufacturer. The following distinct regionsof gag, pol, and env were amplified by nested polymerase chainreaction (PCR): A 778-bp gag region (complete matrix and 59
capsid), a 755-bp pol region (complete protease and 59 reversetranscriptase), and a 678-bp region of env (V3–V5). The primerpairs used for the nested PCR and their hybridization positionsin relation to the standard HxB2r isolate are shown in Table 1.PCR was performed under standard conditions. For long-rangePCR amplification of the complete gag–pol region, we used theExpand high-fidelity PCR system (Boehringer, Mannheim,Germany) as recommended by the manufacturer and primersGag1 (Table 1) and SE5119 (CTTAGCTTTCCTTGAAATAT-ACAT, corresponding to positions 5125–5148 in HxB2r).
The amplimers were purified with a QIAquick gel extractionkit (Qiagen). The long PCR product was cloned into a TA vec-tor (Invitrogen, Groningen, The Netherlands). Nucleotide se-quences were obtained by direct sequencing of both strands ofthe PCR products or clones. Sequencing was performed by thefluorescent dye terminator method, using an ABI model 377Aautomated sequencer (Applied Biosystems, Foster City, CA).
Phylogenetic analysis and drug resistance analysis
Sequence alignment of the gene fragments was done auto-matically by CLUSTAL X with minor manual adjustments tak-ing into consideration protein-coding sequences.18 The new se-
TEBIT ET AL.40

quences were aligned with the compilation of nonrecombinantand circulating recombinant form (CRF) HIV-1 sequences fromthe Los Alamos database. Phylogenetic analyses of these se-quences were performed by the neighbor-joining method of theCLUSTAL X program.18 To prevent the possibility of new se-quences obscuring the genetic relationships (especially withinthe subtype A cluster), unknown sequences were phylogeneti-cally analyzed one at a time. During the final analysis, gaps thatwere introduced in order to create the alignment were stripped.The statistical robustness of the neighbor-joining tree and reli-ability of the branching pattern were confirmed by bootstrap-ping (1000 replicates). SIMPLOT version 2.5 was used to per-form bootscanning and similarity analyses.19
Putative protease and reverse transcriptase drug resistancemutations were determined with the Antiviral Drug ResistanceAnalysis (ADRA) tool from the HIV database.20 Codons 9–99of protease and 1–130 of reverse transcriptase were analyzed.The ADRA tool compares the input sequences with sequencesof HIV-1 B-type isolates shown to confer resistance to anti-HIV drugs.
RESULTS
Virus isolation
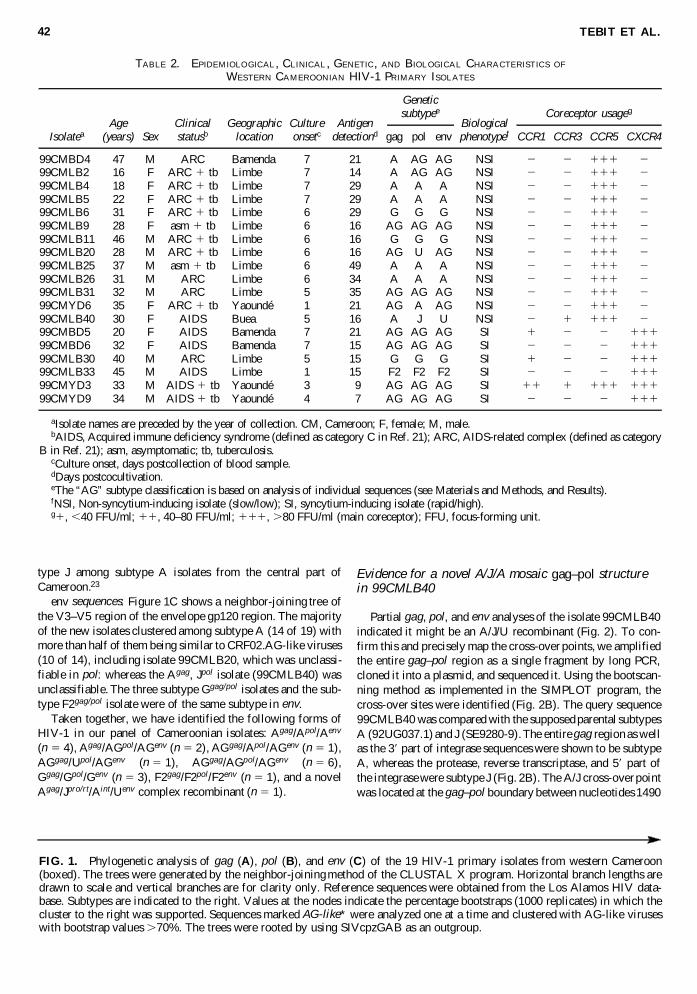
From the 47 blood samples obtained from HIV-infected in-dividuals (most of them tuberculosis patients) from the fourcities we derived 19 primary isolates. Eighteen isolates were ob-tained by coculture with uninfected PBMCs and 1 by coculturewith PM-1 cells. The epidemiological and clinical data are givenin Table 2.21 The cultivation time until first detection of HIV-1capsid antigen was highly dependent on patient clinical statusand isolates from AIDS patients were obtained within a shorter
time (Table 2). Although we could not clearly determine the rea-son for the inability to isolate virus from the other 28 samples,the long period between sample collection and culturing and alsothe shipment conditions (done at room temperature) are the prob-able reasons for this low success in virus isolation.
Phylogenetic analysis
To extend our knowledge of the distribution of various HIV-1 subtypes that are circulating in western Cameroon, we se-quenced discontinuous fragments from the three major struc-tural genes gag, pol, and env of each isolate. Table 2summarizes the genetic characteristics of these isolates.
Gag sequences: Figure 1A shows a gag neighbor-joining treeof the new isolates compared with HIV-1 reference sequences.The new sequences were distributed among subtypes A, G, andF2. Because of the fragment length (, 700 bp) of the variousgenes analyzed, distinction of AG-like viruses from others waspossible only by looking at individual sequences. In the case ofgag, of the 15 isolates that clustered with subtype A, 8 of themsubclustered with the reference CRF02.AG strains (bootstrapvalue, .70%).22 Three isolates clustered among the referenceG strains whereas one was found to be of the newly describedsubsubtype F2.
pol sequences: Sequences from the protease and 59 region ofreverse transcriptase of the 19 isolates were classified into sub-types: A, CRF02.AG, G, F2, and J (Fig. 1B). Five isolates clus-tered with subtype A, eight were CRFAG.02-like viruses, andone isolate (99CMLB20) was unclassifiable. The three isolatesclustering with G had also been found to be subtype G in gagand 99CMLB33 was found to be F2 in gag and pol.
An interesting finding among the pol sequences was the iden-tification of a subtype J sequence in the protease region of the99CMLB40 isolate. Studies have also reported protease sub-
HIV-1 GENOTYPES AND PHENOTYPES IN WESTERN CAMEROON 41
TABLE 1. PRIMERS USED FOR NESTED PCR AND SEQUENCING
Primer Sequence (59 ® 39) Location (HxB2r) Usage/orientation
gag Region
Gag1 TATGGGTGCGAGAGCGTCGGTATTAAG 789–815 Outer/senseGag2 TTTGGTCCTTGTCTTATGTCCAGAATGC 1631–1658 Outer/antisenseGag3 TGGGAAAAAATTCGGTTAAGGCC 836–857 Innera/senseGag4 TACTATTTTATTTAATCCTAGGATTAT 1585–1611 Inner/antisense
pol Region
Pol1 TCAAGGGAAGGCCAGGGAATTT 2111–2132 Outer/sensePol3 TAATATTGCTGGTGATCCTTTCC 3007–3029 Outer/antisensePol2 TCCTTTAACTTCCCTCAAATCACT 2241–2264 Inner/sensePol4 TACATTGTACTGATATCTAATCCCTGGTGT 2964–2993 Inner/antisense
env Region
Env1 TAGCACAGTACAATGTACACATGGAAT 6950–6976 Outer/senseEnv2 GTGCTTCCTGCTCCCAAGAACCCA 7784–7807 Outer/antisenseEnv3 TGTTAAATGGCAGTCTAGCAGAA 7003–7025 Inner/senseEnv4 TTATATAATTCACTTCTCCAATT 7656–7678 Inner/antisense
aInner primers were also used for sequencing PCR products.
type J among subtype A isolates from the central part ofCameroon.23
env sequences: Figure 1C shows a neighbor-joining tree ofthe V3–V5 region of the envelope gp120 region. The majorityof the new isolates clustered among subtype A (14 of 19) withmore than half of them being similar to CRF02.AG-like viruses(10 of 14), including isolate 99CMLB20, which was unclassi-fiable in pol: whereas the Agag, Jpol isolate (99CMLB40) wasunclassifiable. The three subtype Ggag/pol isolates and the sub-type F2gag/pol isolate were of the same subtype in env.
Taken together, we have identified the following forms ofHIV-1 in our panel of Cameroonian isolates: Agag/Apol/Aenv
(n 5 4), Agag/AGpol/AGenv (n 5 2), AGgag/Apol/AGenv (n 5 1),AGgag/Upol/AGenv (n 5 1), AGgag/AGpol/AGenv (n 5 6),Ggag/Gpol/Genv (n 5 3), F2gag/F2pol/F2env (n 5 1), and a novelAgag/Jpro/rt/Aint/Uenv complex recombinant (n 5 1).
Evidence for a novel A/J/A mosaic gag–pol structurein 99CMLB40
Partial gag, pol, and env analyses of the isolate 99CMLB40indicated it might be an A/J/U recombinant (Fig. 2). To con-firm this and precisely map the cross-over points, we amplifiedthe entire gag–pol region as a single fragment by long PCR,cloned it into a plasmid, and sequenced it. Using the bootscan-ning method as implemented in the SIMPLOT program, thecross-over sites were identified (Fig. 2B). The query sequence99CMLB40 was compared with the supposed parental subtypesA (92UG037.1) and J (SE9280-9). The entire gag region as wellas the 39 part of integrase sequences were shown to be subtypeA, whereas the protease, reverse transcriptase, and 59 part ofthe integrase were subtype J (Fig. 2B). The A/J cross-over pointwas located at the gag–pol boundary between nucleotides 1490
TEBIT ET AL.42
TABLE 2. EPIDEMIOLOGICAL, CLINICAL , GENETIC, AND BIOLOGICAL CHARACTERISTICS OF
WESTERN CAMEROONIAN HIV-1 PRIMARY ISOLATES
Age Clinical Geographic Culture Antigen BiologicalIsolatea (years) Sex statusb location onsetc detectiond gag pol env phenotypef CCR1 CCR3 CCR5 CXCR4
99CMBD4 47 M ARC Bamenda 7 21 A AG AG NSI 2 2 111 299CMLB2 16 F ARC 1 tb Limbe 7 14 A AG AG NSI 2 2 111 299CMLB4 18 F ARC 1 tb Limbe 7 29 A A A NSI 2 2 111 299CMLB5 22 F ARC 1 tb Limbe 7 29 A A A NSI 2 2 111 299CMLB6 31 F ARC 1 tb Limbe 6 29 G G G NSI 2 2 111 299CMLB9 28 F asm 1 tb Limbe 6 16 AG AG AG NSI 2 2 111 299CMLB11 46 M ARC 1 tb Limbe 6 16 G G G NSI 2 2 111 299CMLB20 28 M ARC 1 tb Limbe 6 16 AG U AG NSI 2 2 111 299CMLB25 37 M asm 1 tb Limbe 6 49 A A A NSI 2 2 111 299CMLB26 31 M ARC Limbe 6 34 A A A NSI 2 2 111 299CMLB31 32 M ARC Limbe 5 35 AG AG AG NSI 2 2 111 299CMYD6 35 F ARC 1 tb Yaoundé 1 21 AG A AG NSI 2 2 111 299CMLB40 30 F AIDS Buea 5 16 A J U NSI 2 1 111 299CMBD5 20 F AIDS Bamenda 7 21 AG AG AG SI 1 2 2 11199CMBD6 32 F AIDS Bamenda 7 15 AG AG AG SI 2 2 2 11199CMLB30 40 M ARC Limbe 5 15 G G G SI 1 2 2 11199CMLB33 45 M AIDS Limbe 1 15 F2 F2 F2 SI 2 2 2 11199CMYD3 33 M AIDS 1 tb Yaoundé 3 9 AG AG AG SI 11 1 111 11199CMYD9 34 M AIDS 1 tb Yaoundé 4 7 AG AG AG SI 2 2 2 111
aIsolate names are preceded by the year of collection. CM, Cameroon; F, female; M, male.bAIDS, Acquired immune deficiency syndrome (defined as category C in Ref. 21); ARC, AIDS-related complex (defined as category
B in Ref. 21); asm, asymptomatic; tb, tuberculosis.cCulture onset, days postcollection of blood sample.dDays postcocultivation.eThe “AG” subtype classification is based on analysis of individual sequences (see Materials and Methods, and Results).fNSI, Non-syncytium-inducing isolate (slow/low); SI, syncytium-inducing isolate (rapid/high).g1, ,40 FFU/ml; 11, 40–80 FFU/ml; 111, .80 FFU/ml (main coreceptor); FFU, focus-forming unit.
Geneticsubtypee Coreceptor usageg
FIG. 1. Phylogenetic analysis of gag (A), pol (B), and env (C) of the 19 HIV-1 primary isolates from western Cameroon(boxed). The trees were generated by the neighbor-joining method of the CLUSTAL X program. Horizontal branch lengths aredrawn to scale and vertical branches are for clarity only. Reference sequences were obtained from the Los Alamos HIV data-base. Subtypes are indicated to the right. Values at the nodes indicate the percentage bootstraps (1000 replicates) in which thecluster to the right was supported. Sequences marked AG-like* were analyzed one at a time and clustered with AG-like viruseswith bootstrap values .70%. The trees were rooted by using SIVcpzGAB as an outgroup.

HIV-1 GENOTYPES AND PHENOTYPES IN WESTERN CAMEROON 43

TEBIT ET AL.44
FIG. 2. Subtype analysis of the gag–pol region of the novel A/J/U recombinant strain 99CMLB40. (A) Phylogenetic relation-ship of complete gag, protease and reverse transcriptase, and integrase genes of the novel A/J/U complex recombinant strain(boxed). Trees were constructed after determining the cross-over sites, using the bootscanning method of the SIMPLOT program.The reference CRF06.cpx sequences are A/G/J strains from Burkina Faso and Mali. (B) gag–pol bootscanning analysis of99CMLB40. In this analysis a window size of 300 bases with an increment of 20 bases was used. Regions with gaps were notincluded in the alignment. The algorithm used was the Kimura two-parameter method with 100 replicates. Subsubtype F2(95CMMP257C) was used as an outgroup. The y axis represents the bootstrap values while the x axis shows the positions on thegag–pol alignment. The subtype origins of the different genome regions are indicated at the top.

and 1510 (counting from the first nucleotide of gag), similar toour observation when the full-length sequence of isolate99CMBD6 from our panel was analyzed (data not shown) andhad been reported previously for other CRF02.AG viruses.24
The J/A cross-over point lies within the 59 integrase sequenceclose to the boundary with RT between positions 3690 and3710, also similar to the A/G cross-over of 99CMBD6. The dis-tinct A/J/A segments in the gag–pol gene of 99CMLB40 wereanalyzed phylogenetically in relation to other reference se-quences (Fig. 2A). In the subtype A portions in gag and pol,the novel A/J sequence was more closely related to pure sub-type A isolates than to the other recombinant A viruses (Figs2A and 1A). The protease–reverse transcriptase region, on theother hand, clustered with the reference J sequences with a highbootstrap value (100%; Fig. 2A). These findings, which werefurther confirmed after amplification and sequencing of regionsspanning the cross-over points from uncultured PBMCs of thisisolate, indicated that subtype A and J viruses are the parentalstrains of isolate 99CMLB40.
Biological phenotype, coreceptor usage, and V3 loop sequences
The biological phenotype of the primary isolates on MT-2cells could be predicted on the basis of patient clinical status(Table 2). Most SI isolates (four of six) were obtained from pa-tients at late stages of AIDS whereas the NSI isolates were allfrom asymptomatic individuals or patients with AIDS-relatedcomplex, with the exception of 99CMLB40.
The biological phenotypes also correlated with coreceptorusage. All isolates were tested for their ability to infect GHOSTcells expressing one of the following chemokine receptors:CCR1, CCR2, CCR3, CXCR4, CCR5, V28, Bonzo, or BOB(Table 2). Irrespective of their subtype (genotype), all NSI iso-lates used CCR5 as their main coreceptor whereas all SI iso-lates used CXCR4. Three isolates used another coreceptor inaddition to either CCR5 or CXCR4 and one SI isolate was foundto be multitropic, using CCR5, CXCR4, CCR3, and CCR1.None of the 19 isolates used CCR2, V28, Bonzo, or BOB forentry.
The V3 loop of HIV env has been shown to be important forantibody neutralization, phenotypic changes, and coreceptor us-age.25 The V3 loops of 14 new isolates consisted of 35 aminoacids, whereas 2 isolates consisted of either 34 or 37 aminoacids and 1 isolate consisted of 33 amino acids (Fig. 3). Themost common hexapeptide sequences at the apex of the V3 loopwere GPGQAF and GPGQTF (7 and 6 of 19, respectively),whereas 1 isolate each contained the sequence GPGRAV,GAGQTF, GPGRGF, GPGY.F, GRGQTF, and GPGRKY.Apart from the GPGY and GAGQ motifs all others had beenreported from Cameroon.6,13,14 A greater V3 loop sequence het-erogeneity was observed among SI isolates.
The net amino acid charge of the V3 loop was previously foundto be a determinant of biological phenotype.26,27 All our NSI iso-lates except for one had a net charge of less than 16, whereas allSI isolates had a net charge of 16 or more. Xiao et al.27 deter-mined a consensus sequence (S/GXXXGPGXXXXXXXE/D)proposed to be a predictor of CCR5 coreceptor usage. How-ever, only 7 of our 13 NSI isolates contained this consensus se-quence. Furthermore, uncharged and negatively charged amino
acids at positions 11 and 25 have been determined as predic-tors of the NSI phenotype and positively charged amino acidsin these positions have been determined as predictors of the SIphenotype.27,28 Accordingly, the majority of our NSI isolatescontained neutral or negatively charged residues in these posi-tions, whereas five of the six SI isolates contained positivelycharged residues at one of these positions. However, positivecharges at position 11 or 25 were also observed in the case ofthree NSI isolates (Fig. 3).
Determination of putative drug resistance mutations inprotease and reverse transcriptase
Using the same pol sequences as in the phylogenetic analy-sis we determined the occurrence of variants corresponding todrug resistance mutations in B-type isolates, using the AntiviralDrug Resistance Analysis tool (ADRA). Codons 9–99 of pro-tease and 1–130 of reverse transcriptase was analyzed and poly-morphisms were detected in seven positions previously shownto confer drug resistance. Apart from the M36I mutation, whichwas observed in all isolates, the frequency of other mutationsin our panel of isolates was low. Five, six, and eight isolates,respectively, carried three, two, and one alteration in relevantpositions. Only two alterations (V106I and V108I) were ob-served in the reverse transcriptase sequence (data not shown).
DISCUSSION
Genetic variability studies of HIV-1 in Cameroon have sofar been limited to the central urban regions around Yaoundéand Douala, where almost all HIV-1 groups and subtypes havebeen identified.6,13,23 In this study, we determined the molecu-lar and biological characteristics of 19 HIV-1 isolates obtainedmainly from western Cameroon. Our study region included theNorth West and South West Provinces, which lie between cen-tral Cameroon, where all HIV subtypes have been obtained, andNigeria, which shows relatively fewer HIV subtypes. In Westand Central Africa subtype A viruses are the most prevalent,with more than 50% clustering with CRF02AG-like viruses.These viruses cluster differently from subtype A viruses origi-nating from East Africa.29 HIV prevalence studies from someWest African countries indicate less variability in the circulat-ing strains. Peeters et al.30 demonstrated in four main cities ofNigeria the dominance of group M viruses, mainly A, G, andCRF02.AG-like viruses. Our panel of western CameroonianHIV-1 isolates were all HIV-1 group M with fewer subtypeswhen compared with previous studies in the central and north-ern regions of Cameroon.6,13,14 This should, however, be in-terpreted cautiously because of the relatively small number ofstudy isolates and also because this is the first detailed molec-ular study of HIV isolates from this region of Cameroon.
Intersubtype recombination occurs mostly in geographic ar-eas where more than one subtype of HIV prevails. This hasbeen shown to be relatively common in Central Africa.31 Par-tial sequencing of the env region does not allow detection ofrecombinant strains, whereas sequence analysis of regions frommore than one of the structural genes (gag, pol, and env) in-creases the probability of determining possible intersubtype re-combinants.31,32 We therefore analyzed discontinuous portions
HIV-1 GENOTYPES AND PHENOTYPES IN WESTERN CAMEROON 45

from the three major structural genes of HIV, allowing an es-timation of possible nonrecombinant and recombinant strainson the basis of concordant or discordant phylogenies. TheCRF02.AG-like strain was found to be the most prevalent sub-type in our study, accounting for roughly half of the isolates.Earlier studies of HIV-1 strains from Yaoundé found a similarprevalence and CRF02.AG viruses have also been described asthe most prevalent strain circulating in West and CentralAfrica.31
The two complete subtype J sequences currently available inthe HIV database have been obtained from the Democratic Re-public of Congo (Zaire),1 but studies have shown the spread ofJ sequences embedded in subtype A, G, and I sequences fromBurkina Faso and other parts of West Africa and, more recently,also from Yaoundé.23,31 Unlike the complex mosaic structureof an A/J/? strain from Botswana,33 the pol gene of 99CMLB40from this study is almost entirely of subtype J. Generally, the99CMLB40 sequence showed a different pattern of clusteringin the gag–pol region in relation to other J recombinants (A/J/?,A/G/J, and A/G/J/I), suggesting different ancestral recombina-tion events.34
All subtype G viruses that have been completely sequencedso far contain portions that are either unclassified or subtypeA,7 whereas the subtype G isolates obtained in our panel werecomplete G at least in the regions sequenced. Pure subtype Gsequences are required to define the parental forms of the dom-
inant CRF01.AG circulating in West and Central Africa.7 Sub-type F viruses were reclassified into subtype K and subsubtypesF1 and F2.1 These viruses, which have been predominantly re-ported in Cameroon with prevalence rates ranging between 7and 17%, are also present in other parts of Africa, South Amer-ica, and Europe.6,13,30
To date most studies of drug resistance/susceptibility havebeen done with HIV-1 subtype B isolates, which are rarely ob-served in Africa. Currently, few sequence data are available forthe protease and reverse transcriptase genes of nonsubtype Bisolates, but some subtype G and F viruses have been shown tobe less susceptible to respective inhibitors.35 The isolates de-scribed in this study were obtained from drug-naive HIV-in-fected individuals. None of the amino acid alterations observedhas been shown to directly cause drug resistance. The proteaseM36I exchange has been associated with inhibitor resistance inthe presence of additional mutations.9 This M36I mutation,which contributes to nelfinavir and ritonavir resistance, appearsto occur frequently among non-B isolates and Cornelissen etal.36 also found a high prevalence of the M36I substitution(71%) in a panel of subtype A–D isolates.
Several studies have shown a broadening of the coreceptorusage profile of HIV-1 isolates associated with progression toAIDS and about 50% of viruses derived from AIDS patientsshowed usage of multiple coreceptors.37 In a study to determinecoreceptor usage among pregnant women in Cameroon,
TEBIT ET AL.46
FIG. 3. Relationships between V3 loop amino acid sequences, biological phenotype, and coreceptor usage of CameroonianHIV-1 primary isolates. Seq. ID, Sequence identity; dot (.), deletion in the amino acid sequence; dash (–), amino acid identity;Ph, biological phenotype; Co-rec., coreceptor; boldface, amino acid positions determining biological phenotype; boldface italics,apex of V3 loop.

Tscherning-Casper et al.38 described the usage of the orphancoreceptor Bonzo (commonly used by SIV and HIV-239) byfour HIV-1 isolates from pregnant women and one from a new-born baby. These authors suggested that Bonzo may be usedfrequently by env subtype A and CRF02.AG viruses. However,none of the 14 isolates from our panel, which were either sub-type A or of the CRF02.AG lineage in env, was able to useBonzo. Therefore, unlike subtype C viruses, which preferen-tially use CCR5 independent of patient disease status, Bonzousage does not appear to correlate with HIV-1 genotype.12
ACKNOWLEDGMENTS
We are grateful to the authorities of the Cameroon Ministryof Health, especially B. Fogwe, E. Dusi (Limbe), J. Akwa, D.Nota (Buea), and O. Troumba (Bamenda Hospital); the techni-cians and graduate students of the Hematology Laboratory, Uni-versity of Yaounde Teaching Hospital; and A. Schambach forassisting with sampling. We thank the staff of the blood bankat the University Hospital Eppendorf, A. Adam, and J. Stuch,for providing buffy coats and material support, and L. Gürtler,M.T. Dittmar, and K. Wiegers for valuable advice and support.D.M.T. is supported by the Deutscher Akademischer Aus-tauschdienst.
SEQUENCE DATA
Sequences have been deposited in GenBank under the fol-lowing accession numbers:
Isolate gag pol env
99CMBD4 AF355280 AF355299 AF35531899CMBD5 AF355281 AF355300 AF35531999CMBD6 AF355282 AF355301 AF35532099CMLB2 AF355283 AF355302 AF35532199CMLB4 AF355284 AF355303 AF35532299CMLB5 AF355285 AF355304 AF35532399CMLB6 AF355286 AF355305 AF35532499CMLB9 AF355287 AF355306 AF35532599CMLB11 AF355288 AF355307 AF35532699CMLB20 AF355289 AF355308 AF35532799CMLB25 AF355290 AF355309 AF35532899CMLB26 AF355291 AF355310 AF35532999CMLB30 AF355292 AF355311 AF35533099CMLB31 AF355293 AF355312 AF35533199CMLB33 AF355294 AF355313 AF35533299CMLB40 AF355295 AF355314 AF35533399CMYD3 AF355296 AF355315 AF35533499CMYD6 AF355297 AF355316 AF35533599CMYD9 AF355298 AF355317 AF355336
99CMLB40, complete gag-pol AF355337
REFERENCES
1. Robertson DL, Anderson JP, Bradac JA, et al.: HIV-1 nomencla-ture proposal. Science 2000;288:55–65.
2. Mansky LM and Temin HM: Lower in vivo mutation rate of hu-
man immunodeficiency virus type 1 than that predicted from thefidelity of purified reverse transcriptase. J Virol 1995;69:5087–5094.
3. Artenstein AW, VanCott TC, Mascola JR, et al.: Dual infectionwith human immunodeficiency virus type 1 of distinct envelopesubtypes in humans. J Infect Dis 1995;171:805–810.
4. Hu WS and Temin HM: Retroviral recombination and reverse tran-scription. Science 1990;250:1227–1233.
5. Robertson DL, Sharp PM, McCutchan FE, and Hahn BH: Recom-bination in HIV-1. Nature 1995;374:124–126.
6. Takehisa J, Zekeng L, Ido E, et al.: Various types of HIV mixedinfections in Cameroon. Virology 1998;245:1–10.
7. Carr JK, Salminen MO, Albert J, et al.: Full genome sequences ofhuman immunodeficiency virus type 1 subtypes G and A/G inter-subtype recombinants. Virology 1998;247:22–31.
8. Takehisa J, Zekeng L, Ido E, et al.: Human immunodeficiency virustype 1 intergroup (M/O) recombination in Cameroon. J Virol 1999;73:6810–6820.
9. Condra JH, Schleif WA, Blahy OM, et al.: In vivo emergence ofHIV-1 variants resistant to multiple protease inhibitors. Nature1995;374:569–571.
10. Alkhatib G, Combadiere C, Broder CC, et al.: CC CKR5: ARANTES, MIP-a, MIP-1b receptor as a fusion cofactor for mac-rophage-tropic HIV-1. Science 1996;272:1955–1958.
11. Connor RI and Ho DD: Transmission and pathogenesis of humanimmunodeficiency virus type 1. AIDS Res Hum Retroviruses 1994;10:321–323.
12. Tscherning C, Alaeus A, Fredriksson R, et al.: Differences inchemokine coreceptor usage between genetic subtypes of HIV-1.Virology 1998;241:181–188.
13. Nkengasong JN, Janssens W, Heyndrickx L, et al.: Genotypic sub-types of HIV-1 in Cameroon. AIDS 1994;8:1405–1412.
14. Mboudjeka I, Zekeng L, Takehisa J, et al.: HIV type 1 genetic vari-ability in the northern part of Cameroon. AIDS Res Hum Retro-viruses 1999;15:951–956.
15. Lusso P, Cocchi F, Balotta C, et al.: Growth of macrophage-tropicand primary human immunodeficiency virus type 1 (HIV-1) iso-lates in a unique CD41 T-cell clone (PM1): Failure to downreg-ulate CD4 and to interfere with cell-line-tropic HIV-1. J Virol1995;69:3712–3720.
16. Karlsson A, Parsmyr K, Sandstrom E, Fenyö EM, and Albert J:MT-2 cell tropism as prognostic marker for disease progression inhuman immunodeficiency virus type 1 infection. J Clin Microbiol1994;32:364–370.
17. Trkola A, Paxton WA, Monard SP, et al.: Genetic subtype-inde-pendent inhibition of human immunodeficiency virus type 1 repli-cation by CC and CXC chemokines. J Virol 1998;72:396–404.
18. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, and HigginsDG: The CLUSTAL X Windows interface: Flexible strategies formultiple sequence alignment aided by quality analysis tools. Nu-cleic Acids Res 1997;25:4876–4882.
19. Ray SC: Simplot for Windows. http://www.med.lhu.edu/dept/scray/download/simplot (1999).
20. Los Alamos National Laboratory: Antiviral drug resistance analy-sis tool. http://www.hiv.lanl.gov/ADRA/adra.html (2001).
21. Castro KG, Ward JW, Slutsker L, et al.: 1993 revised classifica-tion system for HIV infection and expanded surveillance case def-inition for AIDS among adolescents and adults. Morbid MortalWkly Rep 1992;41:1–19.
22. Howard TM and Rasheed S: Genomic structure and nucleotide se-quence analysis of a new HIV type 1 subtype A strain from Nige-ria. AIDS Res Hum Retroviruses 1996;12:1413–1425.
23. Tscherning-Casper C, Dolcini G, Mauclere P, et al.: Evidence ofthe existence of a new circulating recombinant form of HIV type1 subtype A/J in Cameroon. AIDS Res Hum Retroviruses 2000;16:1313–1318.
HIV-1 GENOTYPES AND PHENOTYPES IN WESTERN CAMEROON 47

24. Cornelissen M, van den Burg R, Zorgdrager F, and Goudsmit J:Spread of distinct human immunodeficiency virus type 1 AG re-combinant lineages in Africa. J Gen Virol 2000;81:515–523.
25. Cocchi F, DeVico AL, Garzino-Demo A, Cara A, Gallo RC, andLusso P: The V3 domain of the HIV-1 gp120 envelope glycopro-tein is critical for chemokine-mediated blockade of infection. NatMed 1996;2:1244–1247.
26. Chesebro B, Wehrly K, Nishio J, and Perryman S: Mapping of in-dependent V3 envelope determinants of human immunodeficiencyvirus type 1 macrophage tropism and syncytium formation in lym-phocytes. J Virol 1996;70:9055–9059.
27. Xiao L, Owen SM, Goldman I, et al.: CCR5 coreceptor usage ofnon-syncytium-inducing primary HIV-1 is independent of phylo-genetically distinct global HIV-1 isolates: Delineation of consen-sus motif in the V3 domain that predicts CCR-5 usage. Virology1998;240:83–92.
28. De Jong JJ, de Ronde A, Keulen W, Tersmette M, and GoudsmitJ: Minimal requirements for the human immunodeficiency virustype 1 V3 domain to support the syncytium-inducing phenotype:Analysis by single amino acid substitution. J Virol 1992;66:6777–6780.
29. Carr JK, Laukkanen T, Salminen MO, et al.: Characterization ofsubtype A HIV-1 from Africa by full genome sequencing. AIDS1999;13:1819–1826.
30. Peeters M, Esu-Williams E, Vergne L, et al.: Predominance of sub-type A and G HIV type 1 in Nigeria, with geographical differencesin their distribution. AIDS Res Hum Retroviruses 2000;16:315–325.
31. Heyndrickx L, Janssens W, Zekeng L, et al.: Simplified strategyfor detection of recombinant human immunodeficiency virus type1 group M isolates by gag/env heteroduplex mobility assay. StudyGroup on Heterogeneity of HIV Epidemics in African Cities. J Vi-rol 2000;74:363–370.
32. Cornelissen M, Kampinga G, Zorgdrager F, and Goudsmit J: Hu-man immunodeficiency virus type 1 subtypes defined by env showhigh frequency of recombinant gag genes. The UNAIDS Networkfor HIV Isolation and Characterization. J Virol 1996;70:8209–8212.
33. Oelrichs RB, Workman C, Laukkanen T, McCutchan FE, and Dea-con NJ: A novel subtype A/G/J recombinant full-length HIV type1 genome from Burkina Faso. AIDS Res Hum Retrovirus 1998;14:1495–1500.
34. Novitsky VA, Gaolekwe S, McLane MF, et al.: HIV type 1 A/Jrecombinant with a pronounced pol gene mosaicism. AIDS ResHum Retroviruses 2000;16:1015–1020.
35. Descamps D, Apetrei C, Collin G, Damond F, Simon F, and Brun-Vezinet F: Naturally occurring decreased susceptibility of HIV-1subtype G to protease inhibitors. AIDS 1998;12:1109–1111.
36. Cornelissen M, van den Burg R, Zorgdrager F, Lukashov V, andGoudsmit J: pol gene diversity of five human immunodeficiencyvirus type 1 subtypes: Evidence for naturally occurring mutationsthat contribute to drug resistance, limited recombination patterns,and common ancestry for subtypes B and D. J Virol 1997;71:6348–6358.
37. Hoffman TL and Doms RW: Chemokines and coreceptors inHIV/SIV–host interactions. AIDS 1998;12(Suppl A):S17–S26.
38. Tscherning-Casper C, Vodros D, Menu E, et al.: Coreceptor usageof HIV-1 isolates representing different genetic subtypes obtainedfrom pregnant Cameroonian women. European Network for InUtero Transmission of HIV-1. J Acquir Immune Defic Syndr 2000;24:1–9.
39. Endres MJ, Clapham PR, Marsh M, et al.: CD4-independent in-fection by HIV-2 is mediated by fusin/CXCR4. Cell 1996;87:745–756.
Address reprint requests to:Ottmar Herchenröder
Institut für Virologie im MTZMedizinische Fakultät Carl Gustav Carus
Technische Universität DresdenFiedlerstr. 42
D-01307 Dresden, Germany
E-mail: [email protected]
TEBIT ET AL.48

This article has been cited by:
1. Denis M. Tebit, Lassana Sangaré, Fabrice Tiba, Yameogo Saydou, Aline Makamtse, Hermann Somlare,Guillaume Bado, Boris G. Kouldiaty, Inoussa Zabsonre, Sibiri L. Yameogo, Kanokporn Sathiandee, JosephY. Drabo, Hans-Georg Kräusslich. 2009. Analysis of the diversity of the HIV-1 pol gene and drugresistance associated changes among drug-naïve patients in Burkina Faso. Journal of Medical Virology 81:10,1691-1701. [CrossRef]
2. Philip A Chan, Rami Kantor. 2009. Transmitted drug resistance in nonsubtype B HIV-1 infection. HIVTherapy 3:5, 447-465. [CrossRef]
3. J.A.M. Brandful , M.E. Coetzer , T. Cilliers , M. Phoswa , M.A. Papathanasopoulos , L. Morris , P.L.Moore . 2007. Phenotypic Characterization of HIV Type 1 Isolates from GhanaPhenotypic Characterizationof HIV Type 1 Isolates from Ghana. AIDS Research and Human Retroviruses 23:1, 144-152. [Abstract][PDF] [PDF Plus]
4. Lucía Pérez-Alvarez, Mercedes Muñoz, Elena Delgado, Celia Miralles, Antonio Ocampo, Valentina García,Michael Thomson, Gerardo Contreras, Rafael Nájera. 2006. Isolation and biological characterization ofHIV-1 BG intersubtype recombinants and other genetic forms circulating in Galicia, Spain. Journal ofMedical Virology 78:12, 1520-1528. [CrossRef]
5. Denis M Tebit, Jean Ganame, Kanokporn Sathiandee, Youssouf Nagabila, Boubacar Coulibaly, Hans-GeorgKrausslich. 2006. Diversity of HIV in Rural Burkina Faso. JAIDS Journal of Acquired Immune DeficiencySyndromes 43:2, 144-152. [CrossRef]
6. Sherri T. Burda , Frank A. J. Konings , Constance A. U. Williams , Christopher Anyangwe , Phillipe N.Nyambi . 2004. HIV-1 CRF09_cpx Circulates in the North West Province of Cameroon Where CRF02_AGInfections Predominate and Recombinant Strains Are CommonHIV-1 CRF09_cpx Circulates in the NorthWest Province of Cameroon Where CRF02_AG Infections Predominate and Recombinant Strains AreCommon. AIDS Research and Human Retroviruses 20:12, 1358-1363. [Abstract] [PDF] [PDF Plus]
7. Nicaise Ndembi, Jun Takehisa, Leopold Zekeng, Eiji Kobayashi, Charlotte Ngansop, Elijah M. Songok,Seiji Kageyama, Taichiro Takemura, Eiji Ido, Masanori Hayami, Lazare Kaptue, Hiroshi Ichimura. 2004.Genetic Diversity of HIV Type 1 in Rural Eastern Cameroon. JAIDS Journal of Acquired Immune DeficiencySyndromes 37:5, 1641-1650. [CrossRef]
8. Avelin Fobang Aghokeng, L??onard Ewane, Bih Awazi, Aubin Nanfack, Eric Delaporte, Martine Peeters,L??opold Zekeng. 2004. Evaluation of Four Simple/Rapid Assays and Two Fourth-Generation ELISAs forthe Identification of HIV Infection on a Serum Panel Representing the HIV-1 Group M Genetic Diversityin Cameroon. JAIDS Journal of Acquired Immune Deficiency Syndromes 37:5, 1632-1640. [CrossRef]
9. Julie Yamaguchi , Pierre Bodelle , Ana S. Vallari , Ruthie Coffey , Carole P. McArthur , Gerald Schochetman, Sushil G. Devare , Catherine A. Brennan . 2004. HIV Infections in Northwestern Cameroon: Identificationof HIV Type 1 Group O and Dual HIV Type 1 Group M and Group O InfectionsHIV Infections inNorthwestern Cameroon: Identification of HIV Type 1 Group O and Dual HIV Type 1 Group M andGroup O Infections. AIDS Research and Human Retroviruses 20:9, 944-957. [Abstract] [PDF] [PDF Plus]
10. Frank A.J. Konings , Phillipe N. Nyambi . 2004. V118I Substitution in the Reverse Transcriptase Geneof HIV Type 1 CRF02_AG Strains Infecting Drug-Naive Individuals in CameroonV118I Substitution inthe Reverse Transcriptase Gene of HIV Type 1 CRF02_AG Strains Infecting Drug-Naive Individuals inCameroon. AIDS Research and Human Retroviruses 20:6, 673-678. [Abstract] [PDF] [PDF Plus]
11. Lucia Fischetti, Ohene Opare-Sem, Daniel Candotti, Francis Sarkodie, Helen Lee, Jean Pierre Allain.2004. Molecular epidemiology of HIV in Ghana: Dominance of CRF02_AG. Journal of Medical Virology73:2, 158-166. [CrossRef]
12. Ping Zhong, Laiyi Kang, Qichao Pan, Frank Konings, Sherri Burda, Liying Ma, Yile Xue, Xiaohong Zheng,Zicheng Jin, Phillipe Nyambi. 2003. Identification and Distribution of HIV Type 1 Genetic Diversity andProtease Inhibitor Resistance???Associated Mutations in Shanghai, P. R. China. JAIDS Journal of AcquiredImmune Deficiency Syndromes 34:1, 91-101. [CrossRef]

13. Rafael Nájera, Elena Delgado, Lucía Pérez-Alvarez, Michael M. Thomson. 2002. Genetic recombinationand its role in the development of the HIV-1 pandemic. AIDS, Supplement 16, S3-S16. [CrossRef]


















