hypomorphic lethal mutations and their implications for the ...

ARTICLE
Generation of Novel Conditional and Hypomorphic Allelesof the Smad2 GeneYe Liu,1 Maria H. Festing,2 Mark Hester,2 John C. Thompson,2 and Michael Weinstein1,2*1Molecular, Cellular, and Developmental Biology Program, The Ohio State University, Columbus, Ohio2Department of Molecular Genetics, The Ohio State University, Columbus, Ohio
Received 15 January 2004; Accepted 7 July 2004
Summary: Smad2 is an intracellular mediator of thetransforming growth factor beta signaling (TGF�) path-way. It has been previously shown that, in the mouse,ablation of functional Smad2 results in embryonic lethal-ity due to gastrulation defects. To circumvent the earlylethality and study the spatially and temporally specificfunctions of Smad2, we utilized the Cre-loxP system togenerate a Smad2 conditional allele. Here we show thata conditional allele, Smad2flox, was generated. In thisallele, exons 9 and 10 are flanked by loxP sites and thegene is functionally wildtype. Cre-mediated recombina-tion results in a deletion allele which phenocopies ourpreviously reported Smad2�C null mutation. To generatethis conditional allele, we first made a targeted mutationwhich introduced a floxed neo cassette into intron 10.This allele (Smad23loxP) functions hypomorphically whenplaced opposite a null allele, and unlike the other pub-lished Smad2 hypomorphic allele, can be maintained inthe homozygous state. genesis 40:118–123, 2004.© 2004 Wiley-Liss, Inc.
Key words: Cre; loxP; hypomorph; null; conditional; Smad2;mouse
INTRODUCTION
Smad2 and Smad3 are intracellular transmitters forTGF�, nodal, and activin signaling ligands. Upon ligandbinding, the receptors are activated, and in turn activateSmad2 and Smad3 via phosphorylation of the C-terminalserine residues in the MH2 domain. These activatedmediators then bind with Smad4, the common Smad,and subsequently translocate to the nucleus where,through interactions with other transcription factors,regulate the expression of responsive genes (Attisanoand Wrana, 2000; Derynck and Zhang, 2003; Massague,1998, 2000). To elucidate the function of Smad2 duringmammalian development, different loss of function al-leles have been generated. Each of these Smad2 allelesresults in embryonic lethal phenotypes (Hamamoto etal., 2002; Heyer et al., 1999; Nomura and Li, 1998;Tremblay et al., 2000; Waldrip et al., 1998; Weinstein etal., 1998). In an effort to circumvent this early lethality,the Cre-loxP system (Sauer, 1998) has been adopted to
create Smad2 conditional alleles. A previously publishedSmad2 conditional allele (Vincent et al., 2003) was de-signed to remove the first coding exon of the protein.However, a truncated Smad2 protein has been found incells homozygous for a similar N-terminal Smad2 muta-tion (Heyer et al., 1999) that can be phosphorylated andtranslocated to the nucleus (Randall et al., 2004). There-fore, we designed a novel conditional allele, recombina-tion of which results in a C-terminal truncation thatdeletes the activation sites due to a frameshift in exon11. Thus, if a protein is created it will lack the majorityof the MH2 domain, including the C-terminal phosphor-ylation sites which are needed for function. Here we alsoreport the generation of this Smad2 conditional allele(Smad2flox), which can be excised by Cre-mediated re-combination. The excision allele (Smad2�E9,10) pheno-copies the previously reported Smad2�C mutation in thehomozygous state (Weinstein et al., 1998). In addition,we report that Smad23loxP, the targeted mutation usedto create the Smad2flox conditional allele, is itself hypo-morphic. Smad23loxP can be maintained in the homozy-gous state, but causes lethality when placed opposite aSmad2 null mutation.
A targeting construct used to create the Smad23loxP
allele was generated by placing a loxP site into intron 8of the Smad2 genomic sequence, and a floxed PGKneocassette (Yang et al., 1998) into intron 10 (Fig. 1a). Theconstruct was electroporated into TC1 ES cells (Deng etal., 1996) and after G418 and gancyclovir double selec-tion, 10 homologous recombinants were identified bySouthern blot analysis using a 5� external probe(Fig. 1a,b). Injection of targeted ES cells into C57BL/6blastocysts generated chimeric mice, and when crossedto Black Swiss mice, resulted in germline transmission ofthe Smad23loxP allele. Mice heterozygous for the
Y. Liu and M.H. Festing contributed equally to this work.* Correspondence to: M. Weinstein, Molecular, Cellular, and Develop-
mental Biology Program, The Ohio State University, Columbus, OH 43210.E-mail: [email protected] online inWiley InterScience (www.interscience.wiley.com)DOI: 10.1002/gene.20072
© 2004 Wiley-Liss, Inc. genesis 40:118–123 (2004)
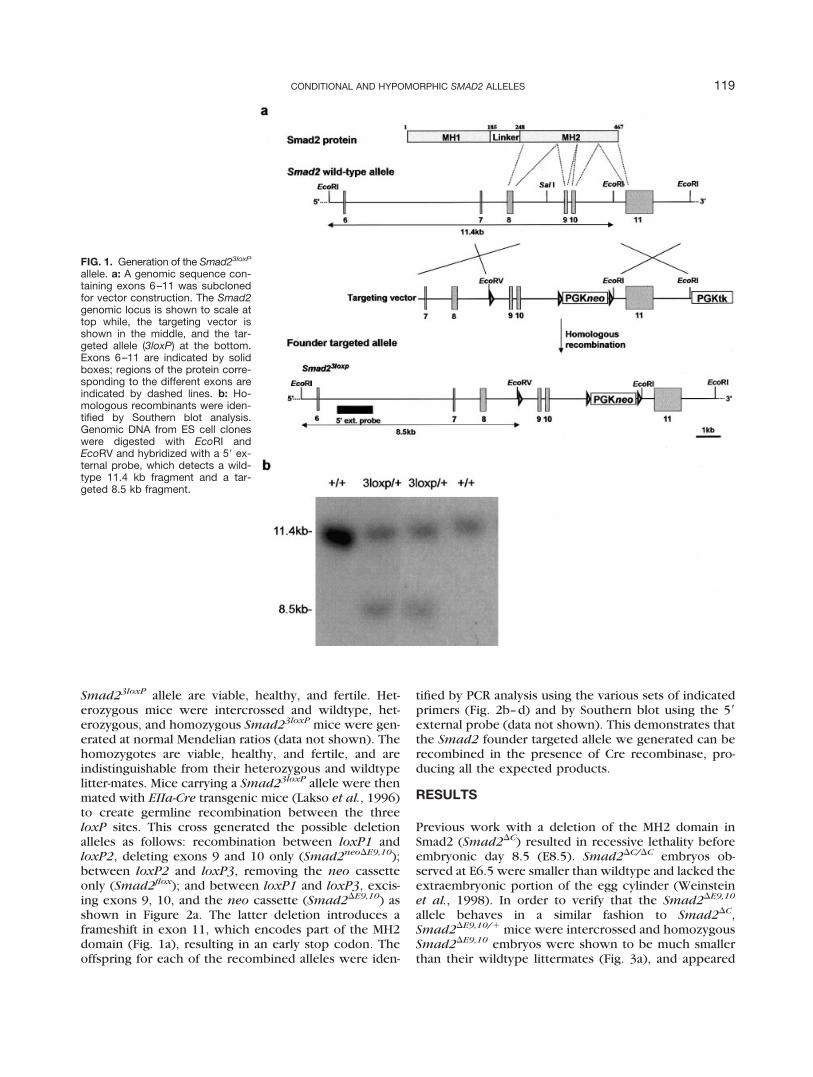
Smad23loxP allele are viable, healthy, and fertile. Het-erozygous mice were intercrossed and wildtype, het-erozygous, and homozygous Smad23loxP mice were gen-erated at normal Mendelian ratios (data not shown). Thehomozygotes are viable, healthy, and fertile, and areindistinguishable from their heterozygous and wildtypelitter-mates. Mice carrying a Smad23loxP allele were thenmated with EIIa-Cre transgenic mice (Lakso et al., 1996)to create germline recombination between the threeloxP sites. This cross generated the possible deletionalleles as follows: recombination between loxP1 andloxP2, deleting exons 9 and 10 only (Smad2neo�E9,10);between loxP2 and loxP3, removing the neo cassetteonly (Smad2flox); and between loxP1 and loxP3, excis-ing exons 9, 10, and the neo cassette (Smad2�E9,10) asshown in Figure 2a. The latter deletion introduces aframeshift in exon 11, which encodes part of the MH2domain (Fig. 1a), resulting in an early stop codon. Theoffspring for each of the recombined alleles were iden-
tified by PCR analysis using the various sets of indicatedprimers (Fig. 2b–d) and by Southern blot using the 5�external probe (data not shown). This demonstrates thatthe Smad2 founder targeted allele we generated can berecombined in the presence of Cre recombinase, pro-ducing all the expected products.
RESULTS
Previous work with a deletion of the MH2 domain inSmad2 (Smad2�C) resulted in recessive lethality beforeembryonic day 8.5 (E8.5). Smad2�C/�C embryos ob-served at E6.5 were smaller than wildtype and lacked theextraembryonic portion of the egg cylinder (Weinsteinet al., 1998). In order to verify that the Smad2�E9,10
allele behaves in a similar fashion to Smad2�C,Smad2�E9,10/� mice were intercrossed and homozygousSmad2�E9,10 embryos were shown to be much smallerthan their wildtype littermates (Fig. 3a), and appeared
FIG. 1. Generation of the Smad23loxP
allele. a: A genomic sequence con-taining exons 6–11 was subclonedfor vector construction. The Smad2genomic locus is shown to scale attop while, the targeting vector isshown in the middle, and the tar-geted allele (3loxP) at the bottom.Exons 6–11 are indicated by solidboxes; regions of the protein corre-sponding to the different exons areindicated by dashed lines. b: Ho-mologous recombinants were iden-tified by Southern blot analysis.Genomic DNA from ES cell cloneswere digested with EcoRI andEcoRV and hybridized with a 5� ex-ternal probe, which detects a wild-type 11.4 kb fragment and a tar-geted 8.5 kb fragment.
119CONDITIONAL AND HYPOMORPHIC SMAD2 ALLELES

indistinguishable from Smad2�C/�C embryos (Weinsteinet al., 1998). We and others have occasionally observedheterozygous Smad2 mutant embryos that are externalto the yolk sac (Nomura and Li, 1998; Liu et al., 2004),which were also seen in about 10% of the Smad2�E9,10/�
embryos (Fig. 3a).There has been controversy surrounding the various
Smad2 deletion alleles, which may be at least partiallydue to the deletion of different regions of the Smad2protein. N-terminal mutations of Smad2 lead to mild-er homozygous phenotypes than Smad2�C andSmad2�E9,10 (Heyer et al., 1999; Vincent et al., 2003;Waldrip et al., 1998; Weinstein et al., 1998). It ispossible that the Smad2�E9,10 mutation creates aSmad2 protein with antimorphic effects on TGF� sig-nal transduction (Hoodless et al., 1999). In or-
der to address this possibility, a Smad2 cDNA corre-sponding to the transcript of the Smad2�E9,10 allele(Smad2�E9-11) was tested for dominant-negative activ-ity in a Xenopus laevis embryo injection assay. The 32uninjected and 19 LacZ injected embryos developednormally (Fig. 3b), with a DAI score of 5 (Kao andElinson, 1988), as compared to injection of the dom-inant-negative Smad23SfA (n � 15), which exhibitedanterior truncation (Fig. 3c) and a DAI score of 4.67 �0.7. However, no effects were seen on the develop-ment of the Xenopus embryos injected withSmad2�E9-11 mRNA (Fig. 3d), as all 54 embryos in-jected exhibited a DAI score of 5, despite the presenceof a truncated Smad2 protein (Fig. 3e). Therefore,deletion of exons 9 and 10 is likely to create a nullallele of Smad2.
FIG. 2. Deletion alleles fromCre-mediated recombination ofSmad23loxp and the PCR strat-egy used to identify them. a:Founder mice heterozygous forthe 3loxP allele were bred withEIIa-Cre mice to obtain off-spring, which were mosaic forthe different recombination al-leles. Black arrowheads indi-cate the location and directionof the primer sets used for iden-tification of specific alleles.b–d: Expected lengths of thePCR products used in genotyp-ing. The PCR reactions for de-tecting the neo cassette (b),loxP1 (c), and the full deletion(d) alleles.
120 LIU ET AL.

DISCUSSION
As stated previously, deletion of the Smad2 N-terminusresults in the production of a truncated protein (Randallet al., 2004). In addition, a hypomorphic allele of Smad2has been reported, which phenocopies N-terminalSmad2 mutations (Vivian et al., 2002). Therefore, thepreviously published Smad2 conditional allele (Vincentet al., 2003) could result in a hypomorphic mutationafter recombination. In contrast, the Smad2 conditionalallele described here will be recombined into a null. Thiswill provide a powerful tool to analyze Smad2 functionsunder a variety of physiological and pathological condi-tions.
It has been reported that the neo cassette contains acryptic splice donor site and, therefore, insertion of aneo cassette intronically can result in a hypomorphicallele (Meyers et al., 1998). However, homozygousSmad23loxP mice are obtained at Mendelian ratios andshow no observable abnormality. In order to more com-pletely characterize the Smad23loxP allele, Smad23loxP
mutant mice were bred with Smad2�C/� mice. Fromthese crosses, no Smad23loxP/�C offspring were found in71 offspring, suggesting that this allele combination islethal. The Smad2�E9,10 mutant allele was also crossedwith Smad23loxP, resulting in the same apparent lethal-ity, with no Smad23loxP/�E9,10 animals being found in138 offspring.
The timing of lethality was determined throughcrosses of Smad2�E9,10/� with Smad23loxP/3loxP animals.All of the Smad23loxP/�E9,10 embryos exhibited lethalitybefore E11.5. Between E8.5 and E10.5 they displayed arange of phenotypes (Fig. 3f–i), which can be dividedinto three classes. Class I mutants (Fig. 3f) consist of anempty yolk sac, phenocopying Smad2Robm1/Robm1,Smad2m1Mag/m1Mag, and Smad2m1Mag/Robm1 embryos(Vivian et al., 2002; Waldrip et al., 1998). Class II mutantembryos (Fig. 3g) have a mass of tissue attachedto the distal tip of the yolk sac and, in some cases, thistissue is large enough to display obvious anterior/poste-rior polarity. Class III (Fig. 3h) consists of embryos withsevere midline defects as compared to normal siblings(Fig. 3i). The phenotypic variability observed in theSmad23loxP/�E9,10 embryos may be due to the outbrednature of the genetic background. To determinewhether this hypomorphic effect is due to the presenceof the intronic neo cassette, but not the insertedloxP sites, crosses were established to produceSmad2flox/�E9,10 mice. These were found at normal Men-delian ratios, and are indistinguishable from littermates(data not shown). The embryonic lethality and the de-velopmental abnormalities observed in Smad23loxP/�C
and Smad23loxP/�E9,10 mutants supports the conclusionthat the Smad23loxP allele is a novel hypomorph. It isable to be maintained in the homozygous state, andexhibits no observable anomaly until it is placed oppo-site a null allele. This is a milder hypomorph than thepreviously reported Smad2m1Mag allele (Vivian et al.,
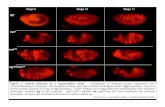
FIG. 3. Analysis of Smad2�E9,10 and Smad23loxP alleles. a: Embryosfrom Smad2�E9,10/� intercrosses were observed at E7.5.Smad2�E9,10/�E9,10 embryos (right, top, and bottom) are muchsmaller than their wildtype littermate (left). The middle embryo isSmad2�E9,10/�, which has embryonic tissue external to the yolk sac.This defect is observed in less than 10% of the heterozygous usingthe Smad2�E9,10/� or Smad2�C/� (Liu et al., 2004). b–d: Xenopusinjections were used to compare the function of Smad2 mutantalleles. b: Uninjected embryo control. c: Injection of the dominant-negative Smad2 allele, Smad23SfA. d: Injection of Smad2�E9-11. e:Western blot of injected Xenopus embryos showing the presence ofSmad2: flag fusion proteins. The lower panel shows a nonspecificband used for a loading control. f–h: Smad23loxP/�E9,10 embryosdisplay a range of phenotypes at E9.5. f: Class I. g: Class II. h: ClassIII. i: Normal Smad23loxP/� littermate. Scale bar in i � 303 �m for a;1.6 mm for f,g; 565 �m for h; and 943 �m for i.
121CONDITIONAL AND HYPOMORPHIC SMAD2 ALLELES

2002), since Smad23loxP/3loxP mice are viable, whileSmad2m1Mag/m1Mag are not. This new hypomorphic al-lele should prove to be extremely useful for determiningnovel functions of Smad2 in different developmentalprocesses.
MATERIALS AND METHODS
Construction of the Targeting Vector andGeneration of Germline ChimerasA 9-kb Smad2 genomic fragment containing exons 6–11was subcloned into pBluescript KS (Stratagene, La Jolla,CA) from a BAC clone (Osoegawa et al., 2000). A loxPsite with an upstream EcoRV site was inserted into theSalI site in intron 8. A 3.1-kb EcoRI/EcoRI fragment wasplaced downstream of the floxed neo cassette of pLox-pneo (Yang et al., 1998). A 5.9-EcoRI/NotI fragment wasinserted upstream of the floxed neo. The NotI site wasnot genomic, but was carried over from the polylinker ofpBluescript KS. These manipulations flank the genomicsequence of exons 9 and 10, as well as the neo cassettewith loxP sites. This Smad23loxp vector was electropo-rated into 129/SvEv TC1 ES cells (Deng et al., 1996) andselected with G418 and gancyclovir; 192 resistant colo-nies were selected and further analyzed by Southernblot. Correctly targeted clones were microinjected intoC57BL/6 blastocysts and implanted into foster mothersto obtain chimeras by standard procedures. The chi-meric founders were crossed with Black Swiss mice toobtain Smad23loxp/� offspring.
Xenopus Embryo Manipulations
Eggs were fertilized in vitro and raised in 0.1� modi-fied Barth’s saline (MBS). Microinjections were per-formed in 1� MBS with 4% ficoll. Embryos were injectedinto the subequatorial dorsal marginal zone at the 4-cellstage with 1.5 ng of synthetic mRNA encoding either thedominant-negative Smad23SfA or Smad2�E9-11, alongwith LacZ. The Smad23SfA has the C-terminal phos-phorylatable serines replaced by alanines, while theSmad2�E9-11 allele truncates after exon 8, thus removinga majority of the MH2 domain with the phosphorylationsites. Lineage tracing with �-galactosidase (with eitherRed-Gal or X-Gal) confirmed injection into dorsal areas ofthe embryos. Western blots on neurula stage embryoswere performed using standard procedures and probedfor the flag epitope attached to the N-terminus of Smad2(Hoodless et al., 1999) using an M2 anti-flag antibodyfrom Sigma (St. Louis, MO).
Genotyping Analysis
Mice were genotyped either by Southern blot or PCR.For PCR analysis, the following primers were used todetect both the wildtype and conditional alleles. Fordetecting the loxP1, SM2-16 (5�-GAG CTG CGC AGACCT TGT TAC-3�) and R2 (5�-TGC CTG ACA AAC AGTCCT GG-3�) amplify a 370-bp product from the wt alleleand a 400-bp product from the conditional allele(Fig. 2a,c). The neo cassette is detected with 3JNEO
(5�-GTT CTA ATT CCA TCA GAA GCT GAC TCA AG-3�)and WTR (5�-GAA GGG GAT CCC ATC TGA GT-3�)(Fig. 2a,b). The Smad2�E9,10 allele was amplified usingSM2-16 and WTR, which generates a 950-bp productfrom the mutant allele (Fig. 2a,d). The Smad2neo�E9,10
allele was amplified using SM2-16 and RINA (5�-CCAGAC TGC CTT GGG AAA AGC-3�), which generates a400-bp product (Fig. 2a, and data not shown). TheSmad2flox allele can also be detected using LEFT (5�-TACTTG GGG CAA TCT TTT CG -3�) and SM2AA (5�-GTCACT CCC TGA ACC TGA AG -3�), which generates a470-bp wt product and a 500-bp mutant product (Fig. 2a,and data not shown).
ACKNOWLEDGMENTS
We thank Heithem El-Hodiri for expertise with Xenopusinjection experiments, and Nicole Spahich for assistancewith Western analysis. We also thank Dr. NatarajanMuthusamy for performing the blastocyst microinjec-tions to make chimeric Smad23loxP mice, and membersof the Weinstein laboratory for critical advice and sug-gestions.
LITERATURE CITED
Attisano L, Wrana JL. 2000. Smads as transcriptional co-modulators.Curr Opin Cell Biol 12:235–243.
Deng C, Wynshaw-Boris A, Zhou F, Kuo A, Leder P. 1996. Fibroblastgrowth factor receptor 3 is a negative regulator of bone growth.Cell 84:911–921.
Derynck R, Zhang YE. 2003. Smad-dependent and Smad-independentpathways in TGF-beta family signalling. Nature 425:577–584.
Hamamoto T, Beppu H, Okada H, Kawabata M, Kitamura T, MiyazonoK, Kato M. 2002. Compound disruption of smad2 acceleratesmalignant progression of intestinal tumors in apc knockout mice.Cancer Res 62:5955–5961.
Heyer J, Escalante-Alcalde D, Lia M, Boettinger E, Edelmann W, StewartCL, Kucherlapati R. 1999. Postgastrulation Smad2-deficient em-bryos show defects in embryo turning and anterior morphogene-sis. Proc Natl Acad Sci U S A 96:12595–12600.
Hoodless PA, Tsukazaki T, Nishimatsu S, Attisano L, Wrana JL, Thom-sen GH. 1999. Dominant-negative Smad2 mutants inhibit activin/Vg1 signaling and disrupt axis formation in Xenopus. Dev Biol207:364–379.
Kao KR, Elinson RP. 1988. The entire mesodermal mantle behaves asSpemann’s organizer in dorsoanterior enhanced Xenopus laevisembryos. Dev Biol 127:64–77.
Lakso M, Pichel JG, Gorman JR, Sauer B, Okamoto Y, Lee E, Alt FW,Westphal H. 1996. Efficient in vivo manipulation of mousegenomic sequences at the zygote stage. Proc Natl Acad Sci U S A93:5860–5865.
Liu Y, Festing M, Thompson JC, Hester M, Rankin S, El-Hodiri HM, ZornAM, Weinstein M. 2004. Smad2 and Smad3 coordinately regulatecraniofacial and endodermal development. Dev Biol 270:411–426.
Massague J. 1998. TGF-beta signal transduction. Annu Rev Biochem67:753–791.
Massague J. 2000. How cells read TGF-beta signals. Nat Rev Mol CellBiol 1:169–178.
Meyers EN, Lewandoski M, Martin GR. 1998. An Fgf8 mutant allelicseries generated by Cre- and Flp-mediated recombination. NatGenet 18:136–141.
Nomura M, Li E. 1998. Smad2 role in mesoderm formation, left-rightpatterning and craniofacial development. Nature 393:786–790.
Randall RA, Howell M, Page CS, Daly A, Bates PA, Hill CS. 2004.
122 LIU ET AL.

Recognition of phosphorylated-Smad2-containing complexes by anovel Smad interaction motif. Mol Cell Biol 24:1106–1121.
Sauer B. 1998. Inducible gene targeting in mice using the Cre/loxsystem. Methods 14:381–392.
Tremblay KD, Hoodless PA, Bikoff EK, Robertson EJ. 2000. Formationof the definitive endoderm in mouse is a Smad2-dependent pro-cess. Development 127:3079–3090.
Vincent SD, Dunn NR, Hayashi S, Norris DP, Robertson EJ. 2003. Cellfate decisions within the mouse organizer are governed by gradedNodal signals. Genes Dev 17:1646–1662.
Vivian JL, Chen Y, Yee D, Schneider E, Magnuson T. 2002. An allelicseries of mutations in Smad2 and Smad4 identified in a genotype-
based screen of N-ethyl-N-nitrosourea-mutagenized mouse embry-onic stem cells. Proc Natl Acad Sci U S A 99:15542–15547.
Waldrip WR, Bikoff EK, Hoodless PA, Wrana JL, Robertson EJ. 1998.Smad2 signaling in extraembryonic tissues determines anterior-posterior polarity of the early mouse embryo. Cell 92:797–808.
Weinstein M, Yang X, Li C, Xu X, Gotay J, Deng CX. 1998. Failure ofegg cylinder elongation and mesoderm induction in mouse em-bryos lacking the tumor suppressor smad2. Proc Natl Acad SciU S A 95:9378–9383.
Yang X, Li C, Xu X, Deng C. 1998. The tumor suppressor SMAD4/DPC4is essential for epiblast proliferation and mesoderm induction inmice. Proc Natl Acad Sci U S A 95:3667–3672.
123CONDITIONAL AND HYPOMORPHIC SMAD2 ALLELES