Gene order and chromosome dynamics coordinate ... · Gene order and chromosome dynamics coordinate...
9
Gene order and chromosome dynamics coordinate spatiotemporal gene expression during the bacterial growth cycle Patrick Sobetzko a , Andrew Travers b,c , and Georgi Muskhelishvili a,1 a School of Engineering and Science, Jacobs University Bremen, D-28759 Bremen, Germany; b Medical Research Council Laboratory of Molecular Biology, Cambridge CB2 0QH, United Kingdom; and c Fondation Pierre-Gilles de Gennes pour la Recherche, Laboratoire de Biologie et Pharmacologie Appliquée, Ecole Normale Supérieure de Cachan, 94235 Cachan, France Edited by Sankar Adhya, National Cancer Institute, National Institutes of Health, Bethesda, MD, and approved November 23, 2011 (received for review May 23, 2011) In Escherichia coli crosstalk between DNA supercoiling, nucleoid-as- sociated proteins and major RNA polymerase σ initiation factors regulates growth phase-dependent gene transcription. We show that the highly conserved spatial ordering of relevant genes along the chromosomal replichores largely corresponds both to their tem- poral expression patterns during growth and to an inferred gradient of DNA superhelical density from the origin to the terminus. Genes implicated in similar functions are related mainly in trans across the chromosomal replichores, whereas DNA-binding transcriptional reg- ulators interact predominantly with targets in cis along the repli- chores. We also demonstrate that macrodomains (the individual structural partitions of the chromosome) are regulated differently. We infer that spatial and temporal variation of DNA superhelicity during the growth cycle coordinates oxygen and nutrient availabil- ity with global chromosome structure, thus providing a mechanistic insight into how the organization of a complete bacterial chromo- some encodes a spatiotemporal program integrating DNA replica- tion and global gene expression. gene order conservation | transcriptional regulatory network | protein gradients I n Escherichia coli cells the physiological transitions induced by the changing growth environment are accompanied by changes in DNA superhelical density (1–3), nucleoid structure (4–6), and the promoter selectivity of the RNA polymerase (RNAP) holo- enzyme (3, 7). During the growth cycle both the relative and absolute concentrations of the abundant nucleoid-associated proteins (NAPs; Table S1) change substantially and correspond- ingly generate bacterial chromatin of variable composition (8, 9). The NAPs stabilize distinct supercoil structures (10–12) selec- tively favoring particular RNAP holoenzymes (13–15). These variable nucleoprotein complexes modulate DNA topology during the growth cycle (Fig. 1A), optimizing the channeling of supercoil energy into appropriate metabolic pathways (16, 17). The expression of the genes determining superhelical density, polymerase selectivity, and nucleoid structure is coordinated by cross-regulation. Thus, factor for inversion stimulation (FIS), a NAP abundant during the early exponential phase (18), regulates expression not only of the superhelicity determinants DNA gyrase subunits A and B (gyrA and gyrB) and topoisomerase I (topA) (19– 21) but also other NAP-encoding genes including hns, α subunit of histone-like protein from E. coli strain U93 (hupA), and DNA binding protein from starved cells (dps) (22–24) and components of the transcription machinery such as σ 38 subunit of RNA poly- merase rpoS (25). Similarly mutations affecting the selectivity of RNAP influence NAP production (26–28). Again, mutations in the genes controlling DNA superhelicity affect the production of both NAPs and the basal transcription machinery (27, 29). This pattern of integrated control constitutes a heterarchical network co- ordinating chromosome structure with cellular metabolism (27, 28, 30). A further pointer to this integrated network is the observed selection of mutations in fis and tRNA dihydrouridine synthase (dusB) (essential for fis expression) and also in topA (31), as well as in rpoC (the β′ subunit of RNAP) under conditions of adaptive evolution (32). Although there is substantial evidence for integrated regulation of NAPs, DNA superhelicity, and RNAP selectivity during the growth cycle, the mechanism by which this regulation is accom- plished remains obscure. We report here that the conserved or- dering of the stage-specific regulatory genes and their targets along the replichores corresponds with their temporal expression pat- terns during the growth cycle. We propose that this ordering reflects a gradient of DNA gyrase-binding sites and hence negative superhelicity from chromosomal origin (OriC) to terminus (Ter) of replication and that the generation of this superhelicity gradient is coupled to energy availability. During the growth cycle changes in local superhelicity drive morphological changes in chromosome structure that facilitate the integration of DNA replication and gene expression. Results Gene Order. We observed that the ordering, relative to OriC, of the genes encoding the NAPs specific to particular stages of the growth cycle [i.e., fis, hupA, β subunit of histone-like protein from E. coli strain U93 (hupB), suppression of td phenotype (stpA), lrp, dps, cbpA, and β subunit of integration host factor (ihfB)] ap- proximately reflects their relative abundance during the growth cycle (8, 9). With one exception, hns, the NAPs associated with the higher overall superhelicity characteristic of exponential growth are closer to the origin. Conversely, those associated with the lower superhelical density characteristic of the stationary phase are closer to the replication terminus (Fig. 1B). The or- dering of the genes for the transcriptional-machinery components exhibits a pattern similar to that of the NAPs, with rpoD, encoding the σ 70 factor for vegetative growth, located closer to OriC than rpoS, encoding the stationary phase σ S factor. Similarly, gyrB (but not gyrA), encoding subunit B of DNA gyrase, is located in close proximity to OriC. Gyrase increases negative superhelicity, es- pecially with the higher ATP/ADP ratios prevailing on nutritional shift-up (33). In contrast, both topA and topoisomerase III (topB), encoding the DNA-relaxing topoisomerases, are closer to the Author contributions: A.T. and G.M. designed research; P.S. performed research; P.S., A.T., and G.M. analyzed data; and P.S., A.T., and G.M. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. Freely available online through the PNAS open access option. 1 To whom correspondence should be addressed. E-mail: g.muskhelishvili@jacobs-university. de. See Author Summary on page 355. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1108229109/-/DCSupplemental. E42–E50 | PNAS | January 10, 2012 | vol. 109 | no. 2 www.pnas.org/cgi/doi/10.1073/pnas.1108229109 Downloaded by guest on August 22, 2020
Transcript of Gene order and chromosome dynamics coordinate ... · Gene order and chromosome dynamics coordinate...

Gene order and chromosome dynamics coordinatespatiotemporal gene expression during thebacterial growth cyclePatrick Sobetzkoa, Andrew Traversb,c, and Georgi Muskhelishvilia,1
aSchool of Engineering and Science, Jacobs University Bremen, D-28759 Bremen, Germany; bMedical Research Council Laboratory of Molecular Biology,Cambridge CB2 0QH, United Kingdom; and cFondation Pierre-Gilles de Gennes pour la Recherche, Laboratoire de Biologie et Pharmacologie Appliquée,Ecole Normale Supérieure de Cachan, 94235 Cachan, France
Edited by Sankar Adhya, National Cancer Institute, National Institutes of Health, Bethesda, MD, and approved November 23, 2011 (received for reviewMay 23, 2011)
In Escherichia coli crosstalk between DNA supercoiling, nucleoid-as-sociated proteins and major RNA polymerase σ initiation factorsregulates growth phase-dependent gene transcription. We showthat the highly conserved spatial ordering of relevant genes alongthe chromosomal replichores largely corresponds both to their tem-poral expression patterns during growth and to an inferred gradientof DNA superhelical density from the origin to the terminus. Genesimplicated in similar functions are related mainly in trans across thechromosomal replichores, whereas DNA-binding transcriptional reg-ulators interact predominantly with targets in cis along the repli-chores. We also demonstrate that macrodomains (the individualstructural partitions of the chromosome) are regulated differently.We infer that spatial and temporal variation of DNA superhelicityduring the growth cycle coordinates oxygen and nutrient availabil-ity with global chromosome structure, thus providing a mechanisticinsight into how the organization of a complete bacterial chromo-some encodes a spatiotemporal program integrating DNA replica-tion and global gene expression.
gene order conservation | transcriptional regulatory network |protein gradients
In Escherichia coli cells the physiological transitions induced bythe changing growth environment are accompanied by changes
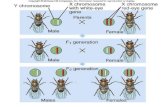
in DNA superhelical density (1–3), nucleoid structure (4–6), andthe promoter selectivity of the RNA polymerase (RNAP) holo-enzyme (3, 7). During the growth cycle both the relative andabsolute concentrations of the abundant nucleoid-associatedproteins (NAPs; Table S1) change substantially and correspond-ingly generate bacterial chromatin of variable composition (8, 9).The NAPs stabilize distinct supercoil structures (10–12) selec-tively favoring particular RNAP holoenzymes (13–15). Thesevariable nucleoprotein complexes modulate DNA topology duringthe growth cycle (Fig. 1A), optimizing the channeling of supercoilenergy into appropriate metabolic pathways (16, 17).The expression of the genes determining superhelical density,
polymerase selectivity, and nucleoid structure is coordinated bycross-regulation. Thus, factor for inversion stimulation (FIS),a NAP abundant during the early exponential phase (18), regulatesexpression not only of the superhelicity determinants DNA gyrasesubunits A and B (gyrA and gyrB) and topoisomerase I (topA) (19–21) but also other NAP-encoding genes including hns, α subunit ofhistone-like protein from E. coli strain U93 (hupA), and DNAbinding protein from starved cells (dps) (22–24) and componentsof the transcription machinery such as σ38 subunit of RNA poly-merase rpoS (25). Similarly mutations affecting the selectivity ofRNAP influence NAP production (26–28). Again, mutations in thegenes controlling DNA superhelicity affect the production of bothNAPs and the basal transcription machinery (27, 29). This patternof integrated control constitutes a heterarchical network co-ordinating chromosome structure with cellular metabolism (27, 28,30). A further pointer to this integrated network is the observed
selection of mutations in fis and tRNA dihydrouridine synthase(dusB) (essential for fis expression) and also in topA (31), as well asin rpoC (the β′ subunit of RNAP) under conditions of adaptiveevolution (32).Although there is substantial evidence for integrated regulation
of NAPs, DNA superhelicity, and RNAP selectivity during thegrowth cycle, the mechanism by which this regulation is accom-plished remains obscure. We report here that the conserved or-dering of the stage-specific regulatory genes and their targets alongthe replichores corresponds with their temporal expression pat-terns during the growth cycle. We propose that this orderingreflects a gradient of DNA gyrase-binding sites and hence negativesuperhelicity from chromosomal origin (OriC) to terminus (Ter)of replication and that the generation of this superhelicity gradientis coupled to energy availability. During the growth cycle changesin local superhelicity drive morphological changes in chromosomestructure that facilitate the integration of DNA replication andgene expression.
ResultsGene Order. We observed that the ordering, relative to OriC, ofthe genes encoding the NAPs specific to particular stages of thegrowth cycle [i.e., fis, hupA, β subunit of histone-like protein fromE. coli strain U93 (hupB), suppression of td phenotype (stpA), lrp,dps, cbpA, and β subunit of integration host factor (ihfB)] ap-proximately reflects their relative abundance during the growthcycle (8, 9). With one exception, hns, the NAPs associated withthe higher overall superhelicity characteristic of exponentialgrowth are closer to the origin. Conversely, those associated withthe lower superhelical density characteristic of the stationaryphase are closer to the replication terminus (Fig. 1B). The or-dering of the genes for the transcriptional-machinery componentsexhibits a pattern similar to that of the NAPs, with rpoD, encodingthe σ70 factor for vegetative growth, located closer to OriC thanrpoS, encoding the stationary phase σS factor. Similarly, gyrB (butnot gyrA), encoding subunit B of DNA gyrase, is located in closeproximity to OriC. Gyrase increases negative superhelicity, es-pecially with the higher ATP/ADP ratios prevailing on nutritionalshift-up (33). In contrast, both topA and topoisomerase III (topB),encoding the DNA-relaxing topoisomerases, are closer to the
Author contributions: A.T. and G.M. designed research; P.S. performed research; P.S., A.T.,and G.M. analyzed data; and P.S., A.T., and G.M. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Freely available online through the PNAS open access option.1To whom correspondence should be addressed. E-mail: [email protected].
See Author Summary on page 355.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1108229109/-/DCSupplemental.
E42–E50 | PNAS | January 10, 2012 | vol. 109 | no. 2 www.pnas.org/cgi/doi/10.1073/pnas.1108229109
Dow
nloa
ded
by g
uest
on
Aug
ust 2
2, 2
020

Ter. Spatial organization of genes modifying the transcriptionmachinery [transcriptional terminator Rho (rho), the N utiliza-tion substance gene (nus) factors, the gre factors, dnaK suppressor
(dksA), curly (crl)] and genes sustaining catabolism and energyproduction under aerobic [ATP synthase (atp) operon], micro-aerobic [two component signal transduction system (arcA/B)], and
NAP abundance
RNA polymerase sigma factors
Plasmid superhelical
density ( )
Growth stage
S
-0.068 -0.043
Shift up Early log Mid/late log Transition Stationary (ppGpp spike)
TerOriC
Ori NS Right Ter
Ori NS Left Ter
Ter
TerOriC
rpoSrpoDTerOriC
rpoBC
rpoA
OriC
Chromosomal macrodomains
Aerobic/anaerobic metabolism,DNA replication,rrn genes, etc
DNA topology
NAPs
RNA polymerase modulators
TerOriCRight
Left
fliArpoE
fecI
rpoH
rpoN
cbpA hns
stpA
fnr
atp
arcA
arcB
hupA hupB
ihfA
ihfBlrp
dps
fiscrp
hfq
seqA
dnaA
rmf
rpoZ ssrS
crl
rsd
dksA
greA
greB
nusA
nusG
rho
gyrB gyrAparCparE
yacG topA
topBsbmC
H
G
E
D
BAC
A
B
Fig. 1. Spatiotemporal organization of chromosomal expression. (A) Temporal changes of NAPs, RNAP composition, and average plasmid DNA superhelicity (σ)during bacterial growth. The phases of growth cycle are correlatedwith preferred expression of particular NAPs, RNAP holoenzymes and the supercoiling temporalgradient are indicated below. A transient increase in guanosine tetraphosphate (ppGpp) levels occurs at the transition between the exponential and stationaryphases. (B) Spatial ordering of regulatory genes on the E. coli chromosome along the OriC–Ter axis. (Top line) Correspondence of macrodomains defined by Valenset al. (40) to linear map. (First bar) Selected genes involved in aerobic/anaerobic metabolism (dark blue), DNA replication (orange), rrn genes (red), and transitionphase (brown). Genes on the clockwise (right) replichore are above the bar, and genes on anti-clockwise (left) replichore are below the bar. The atp operon encodesATP synthase. arcA/arcB encode a two-component system active undermicroaerobic conditions (61, 62). ArcA also represses rpoS (63). fnr has a dominant role undermore strictly anaerobic conditions (61). dnaA, encoding the principal initiator of DNA replication, maps close to OriC, whereas seqA, an inhibitor of replicationinitiation atOriC,maps closer to Ter. rmfdecreases the availability of ribosomes andmaps to amacrodomain immediately adjacent to the Termacrodomain. (Secondbar) Selected genes involved in control of DNA topology. gyrB, a component of DNA gyrase responsible for increasing negative superhelicity, maps close to OriC,whereas the gyrase inhibitor susceptibility to B17microcin, locus C (sbmC), and topA and topB, both responsible for relaxing DNA, map either close to or within theTer macrodomain. DNA gyrase inhibitor (yacG), encoding an inhibitor of GyrB, maps close to the center. Chromosomal partition genes C and E (parC and parE)encode the subunits of topoisomerase IV, responsible for decatenation of newly replicatedDNA in the terminal region (64) and relaxation of negative supercoils (65).(Third bar) Selected genes encoding NAPs. The NAP-encoding gene closest to OriC is hupA, encoding histone-like protein from E. coli strain U93 (HU)α. Its earlyexpression relative to hupB, encodingHuβ (9), could buffer high negative superhelicity generated by DNA gyrase (36). HUα2 and HUαβ, but not HUβ2, constrain highsuperhelical densities in vitro (9). A mutation in hupA both increases growth rate and antagonizes histone-like nucleoid-structuring protein (H-NS) regulation ofcertain transcription units (6). High frequency of recombination (Hfq) is a nucleic acid-binding protein whose major role is that of an RNA chaperone, but it also mayact as a DNA-binding NAP (8). lrp is activated by ppGpp (38). (Fourth bar) Selected genes involved in modulating RNAP activity, including σ factor-utilization reg-ulators, secondary channel-binding proteins, termination/elongation factors, and RNAP subunits. σ factor-utilization regulators (light green): ω subunit of RNApolymerase (rpoZ), mapping close to the origin, encodes the ω subunit of RNAP, which confers a preference for utilization of σ70 (28). Regulator of sigma D (rsd)encodes an anti-σ70 (66),whereas crl confers a strongpreference for σS utilization (67). Note that both rpoZ and crlmap closer to OriC thando the respective σ factorswhose activity they affect. The encoded regulatory pattern thus reflects a shift from predominantly σ70 use close toOriC to σS availability in the central region of thechromosome. Secondary channel-binding proteins (plum): growth regulator A and B, transcription elongation factors (greA and greB) both map in the regioncontaining many genes expressed during rapid growth. GreA has been shown to stimulate initiation and transcription of genes involved in aerobic metabolism,including the atp operon (68, 69) as well as the rrnB P1 promoter in vitro (70, but also see ref. 71). DksA, like the plasmid-encoded quorum sensing regulator (TraR)protein (72), inhibits ribosomal protein promoters and rrn initiation (73, 74) and is more distant fromOriC than is greA. In vivo it would act to reduce the rate of rrninitiation and hence antagonize transcription foci formation. Termination/elongation factors (red): The termination factor Rho is encoded by a gene located veryclose to OriC. This location may compensate for the antagonistic effect of high negative superhelicity on transcription termination, which involves the rewinding ofDNA. RNAP subunits: Themappositions relative toOriC of rpoD and rpoS, respectively encoding σ70 and σS, correspond to their relative order of temporal expression.
Sobetzko et al. PNAS | January 10, 2012 | vol. 109 | no. 2 | E43
GEN
ETICS
PNASPL
US
Dow
nloa
ded
by g
uest
on
Aug
ust 2
2, 2
020

anaerobic (fnr) conditions, as well as those involved in activationand negative modulation of replication [DNA replication initiatorprotein DnaA (dnaA) and sequestration of origin (seqA)] andtranslation [ribosomal RNA (rrn) operons and ribosome matura-tion factor (rmf)], exhibit a similar pattern of chromosomal or-dering (legend of Fig. 1B and Table S2).We asked whether the gene order of regulatory elements as-
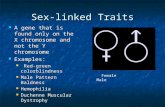
sociated with the temporal pattern of transcription in Escherichiacoli was conserved in other γ-Proteobacteria. Analysis of 131γ-proteobacterial genomes showed that the relative distance of
such genes from the origin and terminus was highly conserved (Fig.2A). Similar analysis of all orthologous genes demonstrated that,although the relative distance was conserved, the specific repli-chore was not conserved to the same extent (Fig. 2B). Simulationssupported this observed bias (Fig. S1). Furthermore, essentiallythe same picture emerged when we analyzed the more distantlyrelated Gram-positive bacteria (Fig. S2 A and B). We concludethat conservation of relative distance along the OriC–Ter axisoverrides the replichore coherence.
Targets. Not only are the genes coordinating the major regulatorypathways ordered; their targets are ordered as well. Analyses ofthe distribution density of binding sites for Eσ70 and EσS hol-oenzymes compiled in RegulonDB (34) show opposite spatialbiases. For the vegetative σ70 factor the highest percentage oftargets is found around the origin, whereas for the stationary phaseσS factor the highest target density is close to the terminus (Fig. 3Aand B), consistent with both the closer location of rpoD to OriCand the temporal division of labor between σ70 and σS during thebacterial growth cycle (7). Similarly, the average density of bindingsites for DNA gyrase (35) diminishes by five- to 10-fold (Fig. 3C)fromOriC to Ter (36). This organization could generate a gradientof superhelical density (Fig. 3D) correlating with that of Eσ70targets and anti-correlating with EσS targets, as expected from theopposite supercoiling preferences of these holoenzymes (3, 28)and in keeping with the requirement of high negative superhelicityfor initiation of OriC replication (37).For the major NAPs, despite distinct chromosomal locations
and abundances during the growth cycle, a high percentage ofthe binding sites compiled in RegulonDB occurs around theorigin (Fig. 3 E–H). Among the NAPs encoded in the Ter-proximal region, only integration host factor (IHF) targets acti-vating binding sites in the vicinity of Ter (Fig. 3F), whereas thestationary-phase regulator leucine responsive protein (LRP) (38)and the global repressor H-NS (39) both preferentially target theOriC-proximal region (Fig. 3 G and H). Additionally HU, themajor supercoil-constraining NAP for which no binding site in-formation is available, has distinct and opposite functionaleffects at the OriC and Ter ends of the chromosome, respectivelyreducing and increasing transcription (Fig. S3).To explore the relevance of this apparent chromosomal ordering
of regulators for chromosomal expression, we used the Gene On-tology (GO) database describing the gene products in terms of as-sociated metabolic processes and investigated the spatial organi-zation of functional groups of genes. Many of these genes would bethe ultimate targets of the regulation. Using scanning windows ofvariable sizes (0.1–0.5 Mb), we mapped the genes in the GO data-base, identifying the significant matches between functionally com-plementing windows on the chromosome (Figs. S4 and S5). Wedetermined the ratio of all significant combinations for cis (matchingwindows located on the same replichore) and trans (matching win-dows on distinct replichores) arrangements. Although the GO treeorganization comprises 13 complexity levels from the most broaddown to specific functions, most significant matches were observedat levels three and four (Figs. S6 and S7). Fig. 4A shows that func-tionally matching groups are localized predominantly on oppositereplichores within the rrn macrodomain (36) comprising OriC andthe nonstructured left (LNS) and right (RNS) macrodomains (40).Importantly, the predominantly trans organization of these groupswas significant only when the chromosomal gene order was alignedalong the OriC–Ter axis. We infer that a majority of the functionallyrelated groups are organized at comparable distances from thecenter of symmetry at OriC and are related in trans across thereplichores. We denote these spatially coordinated matching GOgroups of genes as “maGOGs.”We next analyzed the interactions between the E. coli DNA-
binding transcriptional regulators and their targets compiled inthe RegulonDB in the form of a static transcriptional regulatory
A
B
Fig. 2. Arrangement of important regulatory elements according to their po-sition relative toOriC in the γ-Proteobacteria. (A) Genes located on the right andleft replichores are indicated above and below the chart, respectively, as inFig.1B. Horizontal bars show spatial distributions of orthologs; black color indi-cates the highest density for each individual distribution. The plot shows a strongconservation of chromosomal positions suggesting that origin-focused genepositioning is a major selection criterion in γ-Proteobacteria. (B) Relationshipamong the phylogenetic distance, the correlation of distance to origin, and thereplichore coherence (the conserved replichore identity of orthologs) for allγ-Proteobacteria. The points represent the data on pair-wise species compar-isons computed using the Pearson correlation coefficient of either distances toorigin or replichore identity (right/left) of all orthologous pairs. The points arecolor-coded in the 3D plot. Red indicates a higher correlation of distance toorigin than replichore coherence; blue indicates higher replichore coherence.The predominance of red points indicates the stronger conservation of distanceto origin. The phylogenetic distances in B were derived from the tree of γ-Pro-teobacteria (http://www.cbrg.ethz.ch/research/orthologous/speciestrees).
E44 | www.pnas.org/cgi/doi/10.1073/pnas.1108229109 Sobetzko et al.
Dow
nloa
ded
by g
uest
on
Aug
ust 2
2, 2
020

network (TRN). These analyses again revealed macrodomains inwhich spatially coordinated interactions between the regulatorsand targets were significantly enriched. However, in contrast tothe spatial organization of maGOGs, significant TRN inter-actions occur mainly in cis along the replichores, largely corre-sponding to intermacrodomain communication (Fig. 4B).We also analyzed the spatial organization of couplons, entities
corresponding to intersections of regulons of distinct σ factorsand NAPs and containing functionally related genes (27). Cou-plon analyses again demonstrated preferential communicationsin trans across the replichores (Fig. 4C). We infer that chromo-somal arrangement of functionally related genes and the spatialcommunications between transcriptional regulators and theirtargets are essentially orthogonal with respect to each other.
Expression Profiles. Any valid regulatory network must describemeaningfully the changes in gene expression both during normalgrowth and when expression is perturbed by mutation. The
analyses shown in Figs. 3 and 4 use the static E. coli Regulondatabase, which contains neither temporal nor spatial orderinginformation. To analyze the dynamic organization of communi-cations in the effective transcript profiles (41) obtained underdefined conditions, we integrated the maGOGs, TRN, andcouplon networks in a single combined heterarchical network(HEN). In such a network, as opposed to a hierarchical network,there is no subordination to one single dominant component.The HEN (Fig. 4D) served as a template for mapping the ef-fective transcript profiles. The observed gene ordering predictedthat relative transcription during exponential and stationarygrowth phases would be ordered in a similar spatial mannerrelative to OriC and Ter. The results fully confirmed this pre-diction, clearly demonstrating a division of labor between theOriC and Ter chromosomal ends in temporal expression profiles.In exponentially growing wild-type cells the OriC-proximal regionwas transcriptionally more active than the Ter-proximal regionwith the active regions corresponding largely to the rrn domain
A E
B F
C G
D H
Fig. 3. Organization of binding sites for the major σ initiation factors, DNA, gyrase, and NAPs in the chromosome (RegulonDB). Distributions were calculated byusing a sliding window of 400 kb and normalizing over the total gene number for each window. The replichores are organized from OriC to Ter (left to right).The frequency distributions (ordinate) are plotted above the zero in the ordinate for the right replichore and below the zero for the left replichore. (A) Genomicdistributions for RNAPσ70-regulated promoters. (B) Genomic distributions for σS-regulated promoters. (C) Genomic distributions of gyrase-binding sites. (D)Inferred gradient of negative superhelical density along the OriC–Ter axis in exponentially growing cells. (E) Genomic distribution of FIS-binding sites. (F)Genomic distribution of IHF-binding sites. (G) Genomic distribution of H-NS–binding sites. (H) Genomic distribution of LRP-binding sites. The chromosomalposition for each regulator gene is indicated, and the direction of the effect is rainbow color-coded with blue for repression and red for activation. The Ori, Ter,left, and right macrodomains (green, cyan, blue, and red lines, respectively) are indicated on the chromosomal replichores above and below the distributions.
Sobetzko et al. PNAS | January 10, 2012 | vol. 109 | no. 2 | E45
GEN
ETICS
PNASPL
US
Dow
nloa
ded
by g
uest
on
Aug
ust 2
2, 2
020

comprising the Ori and LNS macrodomains (Fig. 5 A and B).Similar spatiotemporal division of labor was observed in an rpoSmutant lacking σS, but the pattern relative to wild-type cells wasreversed, with the Ter-proximal region being more favored duringthe exponential phase and the OriC-proximal region during thestationary phase (Fig. 5 C and D). Also, in an rpoZ mutant fa-voring the EσS holoenzyme (28), the Ter-proximal region wasfavored during the exponential phase, an effect that could be re-versed by overproduction of σ70 activating the OriC end (Fig. 5 E
and F). Importantly, although the effects of σ factors mainly in-volved trans communications, consistent with the orthogonal or-ganization of regulatory communications in HEN, the mutationsof fis and hns genes also substantially affected the cis communi-cations, which were largely buffered in wild-type cells (Fig. 5 G–
L). Furthermore, some of these spatiotemporal patterns could beclosely imitated by manipulating the composition of NAPs andDNA superhelicity (Fig. S8), validating the usefulness of HEN forgene-expression analyses.
A B
C D
Fig. 4. Organization of functional groups and regulatory communications in the E. coli chromosome. (A) maGOGs. (B) TRN. (C) Couplons. (D) HEN. Thecircular genome is represented as a pair of multicolored parallel lines corresponding to the right (Upper line) and left (Lower line) replichores. On thereplichores the macrodomains (colored as in Fig. 3) and the rrn functional domain (orange dashed line) are indicated. All trans communications occur betweenthe upper and lower lines, whereas cis communications occur along the lines. The order of regulatory genes on the right and left replichores is indicatedabove and below each network, respectively, organized from OriC to Ter.
E46 | www.pnas.org/cgi/doi/10.1073/pnas.1108229109 Sobetzko et al.
Dow
nloa
ded
by g
uest
on
Aug
ust 2
2, 2
020

DiscussionWe have demonstrated that during the E. coli growth cycle inbatch culture the temporal pattern of stage-specific gene ex-pression corresponds largely with the gene order along the tworeplichores. This correspondence is apparent not only for theprincipal regulatory genes but also for their targets. Importantly,this property is a highly conserved characteristic of the repli-chores in both Gram-negative γ-Proteobacteria and Gram-posi-tive bacteria, implying that the ordering is related to DNAreplication. Moreover, the most conserved property for a partic-ular gene appears to be not the precise location but the relativedistance of the gene from OriC and Ter, independent of repli-chore (Fig. 2 and Figs. S1 and S2 A and B). This finding suggeststhat the selective determinant is itself a variable property thatalso is dependent on the relative OriC/Ter distance and is con-sistent with macrodomains sharing many functions (Fig. S9).In fast-growing E. coli the bidirectional replication starting from
OriC creates a gradient of gene dosage fromOriC to Ter, such thatthe resulting origin-proximal relative increase in gene dosage po-tentially could increase gene expression in that region. Indeed, it isprecisely this region that is preferentially active during exponentialgrowth (Fig. 5A). However, variation in gene dosage is likely to beonly one of a number of regulatory influences. Thus, the origin-proximal region contains a five- to 10-fold higher average densityof gyrase-binding sites than the Ter-proximal region (Fig. 3C) andalso is the preferred target region for Eσ70 holoenzyme and for FISprotein (Fig. 3 A and E), implicated in evolutionary modulation ofDNA superhelicity (25). We infer that the gradient of gyrasebinding sites is indicative of potential gradients of negative su-perhelical density from OriC to Ter along the replichores (Fig.3D). The existence of such gradients corresponds well with boththe pattern of holoenzyme and NAP targets and the greater den-sity of supercoiling-sensitive genes (e.g., rpoZ, fis, hupA, and rrn) inthe OriC end (16, 36, 42), as well as with the activation of the rrnmacrodomain under conditions of high negative superhelicityduring exponential growth (Fig. S10 A and B). The gradient ofgyrase-binding sites would provide a simple mechanism for cou-pling energy availability to superhelical density (2, 33) and, be-cause initiation of DNA replication is enhanced by high negativesuperhelicity (37), also to replication itself.Although the spatial order of selected stage-specific genes
corresponds well to the temporal order of expression during thegrowth cycle, the position of certain other genes, although highlyconserved, does not. Genes in this category in E. coli include hnsand gyrA. Although in Gram-positive bacteria gyrA and gyrB mapin close proximity to OriC (Fig. S2C), a rationale for the separateconserved locations of gyrA and gyrB in γ-Proteobacteria is notobvious. On shift-up expression of gyrB increases to a propor-tionally much greater extent than expression of gyrA (19). GyrAand GyrB together form a heterotetramer, and if GyrB werelimiting the distinct locations would favor the use of the gyrase-binding sites in the origin-proximal region. We hope to clarify anyrequirement for separate placement of the gyrA and gyrB genes inE. coli by switching their chromosomal positions.Both FIS and H-NS principally target the origin-proximal re-
gion, potentially delimiting short topological domains (43). How-ever, fis is expressed at maximal levels during the exponentialphase and maps in the origin-proximal region, in contrast to hns,which maps in the Ter macrodomain. In this context MonteroLlopis et al. (44) recently reported that both Escherichia coli andCaulobacter crescentus spatially organize translation so that themRNA product of a gene is translated in close proximity to itsposition in the nucleoid. This observation has profound implica-tions for gene regulation and chromosome structure. If NAPproduction is localized, then diffusion of the resultant DNA-binding proteins within the nucleoid will be anomalous, generatingconcentration gradients. The outcome for nucleoid organization at
A
B
G
H
M
C
D
I
J
E
F
K
L
N
Fig. 5. Spatiotemporal HEN patterns. The circular genome is represented asa pair of thin parallel lines corresponding to the right (Upper line) and left(Lower line) replichores as in Fig. 4.Macrodomains are indicated in color on theright (Upper) and left (Lower) replichores organized from OriC to Ter. (A) Ef-fective HEN (eHEN) pattern of exponentially growing wild-type cells. (B) eHENpattern of transcripts from stationary wild-type cells. Red indicates coherentlyenhanced and blue indicates coherently reduced numbers of expressed genesin matching windows; black indicates an absence of coherence. Note theswitch in the activation of the OriC and Ter ends of the chromosome at thetransition from exponential to stationary growth. (C) eHEN pattern of rpoScells lacking σS obtained during exponential phase. (D) eHEN pattern of rpoScells in stationary phase. Note the spatiotemporal inversion of the activation ofthe OriC and Ter ends of the chromosome with respect to wild-type cells. (E)eHEN pattern of exponentially growing rpoZ cells favoring EσS. (F) As in E, butthe cells were overproducing σ70 from an episome (28). Note the σ70-de-pendent coherent activation and repression of communications in the OriCand Ter ends of the chromosome, respectively. (G and H) eHEN patterns ofwild-type cells grown under conditions of high and low superhelicity, re-spectively. (I and J) Patterns of hns-mutant cells grown under conditions ofhigh and low superhelicity, respectively. Note the supercoiling-dependentcoherent activation and repression of communications in the left and rightreplichores, respectively. (K and L) eHEN patterns of fis-mutant cells grownunder conditions of high and low superhelicity, respectively. Note the coherentactivation of distinct supercoiling-dependent cis and trans communications. InA–D the cells were grown in minimal medium. In E–L the cells were harvestedduring exponential growth in rich double-YT medium. (M and N) Model ofchromosomal morphology changing with DNA superhelicity and NAP gra-dients on transition from exponential (M) to stationary (N) growth. For clarity,only the interactions between the gradients of FIS (pink) and H-NS (blue) areshown. Although FIS levels decline dramatically on transition to stationaryphase, the compaction of the nucleoid along the OriC–Ter axis enables H-NS toestablish repression. The chromosome is depicted as a plectoneme, but themodel is equally consistent with a toroidal scaffold, which would maintain theseparation of the replichores (49, 50). The macrodomains are indicated bycolors, and approximate chromosomal positions of the fis and hns genes areshown. The expression data for mapping onto the HEN connectivity patternswere taken from Dong and Schellhorn (59) in A–D, from Geertz et al. (28) in Eand F, and from Blot et al. (16) in G–L.
Sobetzko et al. PNAS | January 10, 2012 | vol. 109 | no. 2 | E47
GEN
ETICS
PNASPL
US
Dow
nloa
ded
by g
uest
on
Aug
ust 2
2, 2
020

a given locus then depends on the relative local concentrations ofdifferent NAPs, which in turn depend on the distance from the siteof production and the total number of molecules available. In thismodel FIS and H-NS would be expected to be more dominant onnucleoid structure and function in the origin-proximal regionduring the exponential and stationary phases, respectively.The relation of NAP function to negative superhelicity is con-
sistent with the concept that NAPs can act to buffer superhelicaldensity not only at the local level, as previously reported for therrnA P1 promoter (45), but also at the more global level of thechromosome and macrodomains (Fig. 5 G–L). In this respecta correlation between the gradient of gyrase-binding sites and thefunctional effect of HU (36) is particularly striking (Fig. S3). In thecontext of the transcriptional effect of HU, the apparentlyanomalous positions of the rrnG and rrnH genes approximatelyhalfway between OriC and Ter delimit the rrn functional macro-domain (36). Importantly, although all seven rRNA operons havebeen shown to be necessary for rapid adaptation, only five arenecessary to support near-optimal growth (46), and it is not knownhow the rebalancing of rRNA transcription after shift-up is dis-tributed among the rrn operons. Also, in a strain lacking ppGpp(the negative regulator of rRNA transcription), the regions con-taining up-regulated genes whose expression is stimulated by highnegative superhelicity are in close proximity to rrnG and rrnH (Fig.S10C andD). This finding together with the propensity for formingvariable transcription foci (47) raises the possibility that, in growingcells with a full complement of rrn operons, the regulation of in-dividual rrn operons depends on their position in the chromosome.A related issue is how the scattered distribution of tRNA
genes around the chromosome can be reconciled with chromo-somal positioning (Fig. S11). Unlike rRNA genes, the expressionof tRNA genes must be consistent both with growth and withdifferences in the relative amounts of iso-accepting tRNAs andcodon use at different growth rates (ref. 48 and legend of Fig.S11). For both rRNA and iso-accepting tRNA genes, as for othergenes described here, distance from the origin is more conservedthan replichore (Fig. S11B).
Topological Model for Temporal Gene Expression. The deduced logicof spatial communications in the E. coli chromosome suggestsa simple topological model of the circular chromosome folded asa negative supercoil in which close spatial proximity of replichoreseither within the rrn macrodomain comprising the Ori and flank-ing flexible macrodomains or between the rrn and Ter macro-domains supports alternative communication during exponentialand stationary growth. These morphological transitions could oc-cur as a consequence of changes in superhelical density, whichwould affect the configuration of DNA directly and also wouldalter the relative affinities of the different NAPs for DNA. Thealternative configurations, with varying OriC and Ter separation,are consistent with the formation of transcription foci (47), thereorganization and physical extension of nucleoids during expo-nential growth (49–51), and the compaction of nucleoids in thestationary phase (51, 52). This last effect is counteracted by theactivator of rRNA operons FIS, which is abundant in exponentialphase (4, 18), suggesting that the conformational transition in thenucleoid is associated with changing gradients of competing NAPs(Fig. 5 M and N). Although the extent to which gene expressionhas a linear relationship with gene copy number is unknown, it isconceivable that the relative increase in gene dosage resultingfrom the initiation of replication would increase the competitiveadvantage of regulators encoded in the origin-proximal region,whereas a balanced OriC-to-Ter stoichiometry on cessation ofgrowth would abolish such an advantage (Fig. S12).How could such morphological transitions be coupled to en-
ergy availability? A primary response to nutritional shift-up un-der aerobic conditions is an increase in the ATP/ADP ratio (2,33) that activates DNA gyrase, favoring the DnaA–ATP complex
active in replication initiation (53) and coordinating RNAP ac-tivity with energy availability (54). We propose that gyrase ac-tivity creates a gradient of superhelical density corresponding tothe gradient of gyrase-binding sites (Fig. 3 C and D and Fig. S10A and B) such that the selectively increased superhelicity of theorigin-proximal region and the resultant change in chromosomemorphology precede the initiation of replication. Such selectivitysuggests a lower superhelical density in the Ter region, consistentwith the finding that in the prereplicative state in slow-growingcells the Ter region is extended (55). Concomitantly the in-creased spatial separation between origin-proximal H-NS targetgenes and the Ter-proximal location of the hns gene (Fig. 5M)would attenuate the silencing effect H-NS (39, 56, 57) on the rrnmacrodomain. On transition to stationary phase. a dramaticdecline of FIS levels (18) in conjunction with DNA relaxation(Fig. S10 A and B) and compaction of the nucleoid would enableH-NS, as well as the stationary-phase regulator LRP (38), toreestablish efficient repression (Fig. 5N) in the vicinity of repli-cation origin. This model provides a dynamic mechanism for theconcerted impact of the NAPs and DNA superhelicity on chro-mosome structure and cellular physiology (4–6, 9, 10, 16, 17, 19,36, 39, 58).
ConclusionThe conservation of gene order and spatial organization ofregulator–target interactions in chromosomal macrodomainsreported in this paper argue that the gene order specifies thetemporal pattern of gene expression during the bacterial growthcycle. Our work thus illuminates why the timing of the expressionof a gene is linked to its position on the chromosome, providinga view of the bacterial nucleoid as a highly organized dynamicentity optimized for coordinating oxygen and nutrient availabilitywith spatiotemporal gene expression during rapid growth andits cessation.
Materials and MethodsIdentification of Functional Groups in the GO Tree and Modeling of the TFGs. Toaddress the similarity of chromosomal regions, we applied a sliding-windowapproach using a 500-kb window size and a 4-kb window shift creating 1,160windows, each containing about 500 genes. All genes within a window wereassigned to their specific locations on the GO tree (http://www.geneontology.org/). For a fixed level of the GO tree we summed up all genes assigned tothe specific subtree, determining a unique one-dimensional pattern of GO-subtree (cluster) sizes for each window. Subsequently the similarity score oftwo windows (i and j) was determined by the equation:
si;j ¼∑c
k¼1ni;k∗nj;k
∑c
k¼1ni;k∗∑
c
k¼1nj;k
where c is the number of clusters on the current GO-tree level, and n is thenumber of genes in a certain branch of this level, reflecting their covariancewith a mean that equals zero. Hence, high similarity scores (s values) indicatea high correlation of the functional composition. All s values were comparedwith scores of 10,000 random genomes, generated by a random remappingof gene (operon) IDs to gene (operon) positions. For subsequent analyseswindow pairs with a P value < 0.05 were considered significant, and allothers, including scores of overlapping windows [distance(i,j) <500 kb], wereexcluded. However, higher significance values up to a false-discovery rate of0.05 indicated no qualitative change. In practice, the exclusion of over-lapping windows introduces a bias toward trans matches. To address thisissue and to compile a reference data set with the same bias, we shifted theorigin position along the chromosome and compared cis/trans ratios withthe native origin location. Subsequently, the significance of the cis/transratio peaks for a shifted origin position was compared with 10,000 randomgene sets with the same number of best-scoring matches to rule out randomnoise peaks. GO levels three and four turned out to be statistically signifi-cant; lower GO levels lacked a reasonable functional resolution, whereashigher levels represented an extremely detailed functional resolution that
E48 | www.pnas.org/cgi/doi/10.1073/pnas.1108229109 Sobetzko et al.
Dow
nloa
ded
by g
uest
on
Aug
ust 2
2, 2
020

suffered from varying resolutions among distinct functional clusters becauseof a nonuniform depth of the GO tree.
Modeling of the TRN. In correspondence with the GO-tree function analysis,we parsed the TRN communication along the chromosome by counting thenumber of connections between the subnetworks of any two 500-kb win-dows normalized by the total number of connections spreading from bothwindows into the genome. Using procedures analogous to GO similarityanalysis, we determined highly connected regions on the chromosome anddetermined the peaks for the cis/trans ratio by shifting the origin.
Couplon Similarity. As a measure of couplon similarity, we determined foreach couplon (27) whether the number of contained genes for the currentwindow was greater or less than the expected number of genes. Hence, thesimilarity of two windows equals the number of couplons coherently over-or underrepresented in both windows. The determination of the signifi-cance of matches and cis/trans ratio peaks was carried out as for the GO-treesimilarity with random sets of both genes and operons.
Regulator Targets. To determine target site frequency distributions, the rel-evant data derived from the RegulonDB database were analyzed usinga sliding 400-kb window with the map position at the window midpoint.
Expression Patterns. The expression data for mapping onto the HEN con-nectivity patterns were taken from Dong and Schellhorn (59) in Fig. 5 A–D,from Geertz et al. (28) in Fig. 5 E and F, and from Blot et al. (16) in Fig. 5 G–L.
Phylogenetic Analysis. In the phylum of γ-Proteobacteria the majority of reg-ulators is conserved in a plethora of species including human and plantpathogens such as Vibrio cholerae and Pseudomonas syringae. In total we in-vestigated 131 species (using a reciprocal best blast hit approach at the proteinsequence level followed by the determination of orthologous clusters bya modified Girvan–Newman algorithm, resulting in densely connected clusters(node degree ≥ n/2), where n denotes the cluster size. Species informationcontaining protein sequences was derived from the RefSeq sequence databaseand origin positions using the DoriC database (60). The phylogenetic distancesin Fig. 2B were derived from the tree of γ-Proteobacteria (http://www.cbrg.ethz.ch/research/orthologous/speciestrees). The same methodology was ap-plied for analysis of all the annotated genomes of Gram-positive bacteria.
ACKNOWLEDGMENTS. Theauthors thankMalcolmBuckle andSylvie Rimsky forfruitful discussions. This work was supported by grants from the Ecole NormaleSupérieure de Cachan and Deutsche Forschungsgemeinschaft (to G.M.). A.T. re-ceived a Chaire d’Excellence award from l’Agence Nationale de la Recherche.
1. Balke VL, Gralla JD (1987) Changes in the linking number of supercoiled DNAaccompany growth transitions in Escherichia coli. J Bacteriol 169:4499–4506.
2. Hsieh LS, Burger RM, Drlica K (1991) Bacterial DNA supercoiling and [ATP]/[ADP].Changes associated with a transition to anaerobic growth. J Mol Biol 219:443–450.
3. Bordes P, et al. (2003) DNA supercoiling contributes to disconnect sigmaSaccumulation from sigmaS-dependent transcription in Escherichia coli. Mol Microbiol48:561–571.
4. Ohniwa RL, et al. (2006) Dynamic state of DNA topology is essential for genomecondensation in bacteria. EMBO J 25:5591–5602.
5. Rimsky S, Travers A (2011) Pervasive regulation of nucleoid structure and function bynucleoid-associated proteins. Curr Opin Microbiol 14:136–141.
6. Kar S, Edgar R, Adhya S (2005) Nucleoid remodeling by an altered HU protein:Reorganization of the transcription program. Proc Natl Acad Sci USA 102:16397–16402.
7. Ishihama A (2000) Functional modulation of Escherichia coli RNA polymerase. AnnuRev Microbiol 54:499–518.
8. Ali Azam T, Iwata A, Nishimura A, Ueda S, Ishihama A (1999) Growth phase-dependent variation in protein composition of the Escherichia coli nucleoid.J Bacteriol 181:6361–6370.
9. Claret L, Rouvière-Yaniv J (1997) Variation in HU composition during growth ofEscherichia coli: The heterodimer is required for long term survival. J Mol Biol 273:93–104.
10. Dame RT (2005) The role of nucleoid-associated proteins in the organization andcompaction of bacterial chromatin. Mol Microbiol 56:858–870.
11. Guo F, Adhya S (2007) Spiral structure of Escherichia coli HUalphabeta providesfoundation for DNA supercoiling. Proc Natl Acad Sci USA 104:4309–4314.
12. Maurer S, Fritz J, Muskhelishvili G (2009) A systematic in vitro study of nucleoproteincomplexes formed by bacterial nucleoid-associated proteins revealing novel types ofDNA organization. J Mol Biol 387:1261–1276.
13. Colland F, Barth M, Hengge-Aronis R, Kolb A (2000) Sigma factor selectivity ofEscherichia coli RNA polymerase: Role for CRP, IHF and lrp transcription factors. EMBOJ 19:3028–3037.
14. Shin M, et al. (2005) DNA looping-mediated repression by histone-like protein H-NS:Specific requirement of Esigma70 as a cofactor for looping. Genes Dev 19:2388–2398.
15. Maurer S, Fritz J, Muskhelishvili G, Travers A (2006) RNA polymerase and an activatorform discrete subcomplexes in a transcription initiation complex. EMBO J 25:3784–3790.
16. Blot N, Mavathur R, Geertz M, Travers A, Muskhelishvili G (2006) Homeostaticregulation of supercoiling sensitivity coordinates transcription of the bacterialgenome. EMBO Rep 7:710–715.
17. Sonnenschein N, Geertz M, Muskhelishvili G, Hütt MT (2011) Analog regulation ofmetabolic demand. BMC Syst Biol 5:40.
18. Ball CA, Osuna R, Ferguson KC, Johnson RC (1992) Dramatic changes in Fis levels uponnutrient upshift in Escherichia coli. J Bacteriol 174:8043–8056.
19. Schneider R, Travers A, Kutateladze T, Muskhelishvili G (1999) A DNA architecturalprotein couples cellular physiology and DNA topology in Escherichia coli. MolMicrobiol 34:953–964.
20. Keane OM, Dorman CJ (2003) The gyr genes of Salmonella enterica serovarTyphimurium are repressed by the factor for inversion stimulation, Fis. Mol GenetGenomics 270:56–65.
21. Weinstein-Fischer D, Altuvia S (2007) Differential regulation of Escherichia colitopoisomerase I by Fis. Mol Microbiol 63:1131–1144.
22. Claret L, Rouvière-Yaniv J (1996) Regulation of HU α and HU β by CRP and FIS inEscherichia coli. J Mol Biol 263:126–139.
23. Falconi M, Brandi A, La Teana A, Gualerzi CO, Pon CL (1996) Antagonistic involvementof FIS and H-NS proteins in the transcriptional control of hns expression. MolMicrobiol 19:965–975.
24. Grainger DC, Goldberg MD, Lee DJ, Busby SJ (2008) Selective repression by Fis and H-NS at the Escherichia coli dps promoter. Mol Microbiol 68:1366–1377.
25. Hirsch M, Elliott T (2005) Fis regulates transcriptional induction of RpoS in Salmonellaenterica. J Bacteriol 187:1568–1580.
26. Zhou YN, Jin DJ (1998) The rpoB mutants destabilizing initiation complexes atstringently controlled promoters behave like “stringent” RNA polymerases inEscherichia coli. Proc Natl Acad Sci USA 95:2908–2913.
27. Muskhelishvili G, Sobetzko P, Geertz M, Berger M (2010) General organisationalprinciples of the transcriptional regulation system: A tree or a circle? Mol Biosyst 6:662–676.
28. Geertz M, et al. (2011) Structural coupling between RNA polymerase composition andDNA supercoiling in coordinating transcription: A global role for the omega subunit?mBio 2, 4; e00034-11.
29. Peter BJ, et al. (2004) Genomic transcriptional response to loss of chromosomalsupercoiling in Escherichia coli. Genome Biol 5:R87.
30. Travers A, Muskhelishvili G (2005) DNA supercoiling - a global transcriptionalregulator for enterobacterial growth? Nat Rev Microbiol 3:157–169.
31. Crozat E, et al. (2010) Parallel genetic and phenotypic evolution of DNA superhelicityin experimental populations of Escherichia coli. Mol Biol Evol 27:2113–2128.
32. Conrad TM, et al. (2010) RNA polymerase mutants found through adaptive evolutionreprogram Escherichia coli for optimal growth in minimal media. Proc Natl Acad SciUSA 107:20500–20505.
33. van Workum M, et al. (1996) DNA supercoiling depends on the phosphorylationpotential in Escherichia coli. Mol Microbiol 20:351–360.
34. Gama-Castro S, et al. (2008) RegulonDB (version 6.0): Gene regulation model ofEscherichia coli K-12 beyond transcription, active (experimental) annotated promotersand Textpresso navigation. Nucleic Acids Res 36(Database issue):D120–D124.
35. Jeong KS, Ahn J, Khodursky AB (2004) Spatial patterns of transcriptional activity inthe chromosome of Escherichia coli. Genome Biol 5:R86.
36. Berger M, et al. (2010) Coordination of genomic structure and transcription by themain bacterial nucleoid-associated protein HU. EMBO Rep 11:59–64.
37. Fuller RS, Kornberg A (1983) Purified dnaA protein in initiation of replication at theEscherichia coli chromosomal origin of replication. Proc Natl Acad Sci USA 80:5817–5821.
38. Tani TH, Khodursky A, Blumenthal RM, Brown PO, Matthews RG (2002) Adaptation tofamine: A family of stationary-phase genes revealed by microarray analysis. Proc NatlAcad Sci USA 99:13471–13476.
39. Dorman CJ (2004) H-NS: A universal regulator for a dynamic genome. Nat RevMicrobiol 2:391–400.
40. Valens M, Penaud S, Rossignol M, Cornet F, Boccard F (2004) Macrodomainorganization of the Escherichia coli chromosome. EMBO J 23:4330–4341.
41. Marr C, Geertz M, Hütt MT, Muskhelishvili G (2008) Dissecting the logical types ofnetwork control in gene expression profiles. BMC Syst Biol 2:18.
42. Ferrándiz MJ, Martín-Galiano AJ, Schvartzman JB, de la Campa AG (2010) Thegenome of Streptococcus pneumoniae is organized in topology-reacting geneclusters. Nucleic Acids Res 38:3570–3581.
43. Hardy CD, Cozzarelli NR (2005) A genetic selection for supercoiling mutants ofEscherichia coli reveals proteins implicated in chromosome structure. Mol Microbiol57:1636–1652.
44. Montero Llopis P, et al. (2010) Spatial organization of the flow of genetic informationin bacteria. Nature 466:77–81.
45. Rochman M, Aviv M, Glaser G, Muskhelishvili G (2002) Promoter protection bya transcription factor acting as a local topological homeostat. EMBO Rep 3:355–360.
46. Condon C, Liveris D, Squires C, Schwartz I, Squires CL (1995) rRNA operon multiplicityin Escherichia coli and the physiological implications of rrn inactivation. J Bacteriol177:4152–4156.
Sobetzko et al. PNAS | January 10, 2012 | vol. 109 | no. 2 | E49
GEN
ETICS
PNASPL
US
Dow
nloa
ded
by g
uest
on
Aug
ust 2
2, 2
020

47. Cabrera JE, Jin DJ (2003) The distribution of RNA polymerase in Escherichia coli isdynamic and sensitive to environmental cues. Mol Microbiol 50:1493–1505.
48. Dong H, Nilsson L, Kurland CG (1996) Co-variation of tRNA abundance and codonusage in Escherichia coli at different growth rates. J Mol Biol 260:649–663.
49. Lau IF, et al. (2003) Spatial and temporal organization of replicating Escherichia colichromosomes. Mol Microbiol 49:731–743.
50. Adachi S, Fukushima T, Hiraga S (2008) Dynamic events of sister chromosomes in thecell cycle of Escherichia coli. Genes Cells 13:181–197.
51. Kim J, et al. (2004) Fundamental structural units of the Escherichia coli nucleoidrevealed by atomic force microscopy. Nucleic Acids Res 32:1982–1992.
52. Frenkiel-Krispin D, et al. (2004) Nucleoid restructuring in stationary-state bacteria.Mol Microbiol 51:395–405.
53. Olliver A, Saggioro C, Herrick J, Sclavi B (2010) DnaA-ATP acts as a molecular switch tocontrol levels of ribonucleotide reductase expression in Escherichia coli.Mol Microbiol76:1555–1571.
54. Travers AA, Kari C, Mace HAF (1981) Transcriptional regulation by bacterial RNApolymerase. Genetics as a Tool in Microbiology, SGM Symposia, eds Glover S,Hopwood D (Cambridge Univ Press, Cambridge, UK), Vol XXXI, pp 111–130.
55. Wiggins PA, Cheveralls KC, Martin JS, Lintner R, Kondev J (2010) Strong intranucleoidinteractions organize the Escherichia coli chromosome into a nucleoid filament. ProcNatl Acad Sci USA 107:4991–4995.
56. Afflerbach H, Schröder O, Wagner R (1998) Effects of the Escherichia coli DNA-binding protein H-NS on rRNA synthesis in vivo. Mol Microbiol 28:641–653.
57. Spurio R, et al. (1992) Lethal overproduction of the Escherichia coli nucleoid proteinH-NS: Ultramicroscopic and molecular autopsy. Mol Gen Genet 231:201–211.
58. Scolari VF, Bassetti B, Sclavi B, Lagomarsino MC (2011) Gene clusters reflectingmacrodomain structure respond to nucleoid perturbations. Mol Biosyst 7:878–888.
59. Dong T, Schellhorn HE (2009) Control of RpoS in global gene expression of Escherichiacoli in minimal media. Mol Genet Genomics 281:19–33.
60. Gao F, Zhang CT (2007) DoriC: A database of oriC regions in bacterial genomes.Bioinformatics 23:1866–1867.
61. Alexeeva S, Hellingwerf KJ, Teixeira de Mattos MJ (2003) Requirement of ArcA forredox regulation in Escherichia coli under microaerobic but not anaerobic or aerobicconditions. J Bacteriol 185:204–209.
62. Levanon SS, San KY, Bennett GN (2005) Effect of oxygen on the Escherichia coli ArcA
and FNR regulation systems and metabolic responses. Biotechnol Bioeng 89:556–564.63. Mika F, Hengge R (2005) A two-component phosphotransfer network involving ArcB,
ArcA, and RssB coordinates synthesis and proteolysis of sigmaS (RpoS) in E. coli. Genes
Dev 19:2770–2781.64. Zechiedrich EL, Khodursky AB, Cozzarelli NR (1997) Topoisomerase IV, not gyrase,
decatenates products of site-specific recombination in Escherichia coli. Genes Dev 11:
2580–2592.65. Zechiedrich EL, et al. (2000) Roles of topoisomerases in maintaining steady-state DNA
supercoiling in Escherichia coli. J Biol Chem 275:8103–8113.66. Jishage M, Ishihama A (1998) A stationary phase protein in Escherichia coli with
binding activity to the major sigma subunit of RNA polymerase. Proc Natl Acad Sci
USA 95:4953–4958.67. Robbe-Saule V, et al. (2006) Physiological effects of crl in Salmonella are modulated
by SigmaS and promoter activity. J Bacteriol 189:2976–2987.68. Stepanova E, et al. (2007) Analysis of promoter targets for Escherichia coli
transcription elongation factor GreA in vivo and in vitro. J Bacteriol 189:8772–8785.69. Susa M, Kubori T, Shimamoto N (2006) A pathway branching in transcription
initiation in Escherichia coli. Mol Microbiol 59:1807–1817.70. Potrykus K, et al. (2006) Antagonistic regulation of Escherichia coli ribosomal RNA
rrnB P1 promoter activity by GreA and DksA. J Biol Chem 281:15238–15248.71. Rutherford ST, et al. (2007) Effects of DksA, GreA, and GreB on transcription
initiation: Insights into the mechanisms of factors that bind in the secondary channel
of RNA polymerase. J Mol Biol 366:1243–1257.72. Blankschien MD, et al. (2009) TraR, a homolog of a RNAP secondary channel
interactor, modulates transcription. PLoS Genet 5:e1000345.73. Paul BJ, et al. (2004) DksA: A critical component of the transcription initiation
machinery that potentiates the regulation of rRNA promoters by ppGpp and the
initiating NTP. Cell 118:311–322.74. Lemke JJ, et al. (2011) Direct regulation of Escherichia coli ribosomal protein
promoters by the transcription factors ppGpp and DksA. Proc Natl Acad Sci USA 108:
5712–5717.
E50 | www.pnas.org/cgi/doi/10.1073/pnas.1108229109 Sobetzko et al.
Dow
nloa
ded
by g
uest
on
Aug
ust 2
2, 2
020