
Gall structure and specificity in Bostrychia culture ... · 411 Rue des Moguerou, 29680 Roscoff...
10
Algae 2013, 28(1): 83-92 http://dx.doi.org/10.4490/algae.2013.28.1.083 Open Access Research Article Copyright © The Korean Society of Phycology 83 http://e-algae.kr pISSN: 1226-2617 eISSN: 2093-0860 Gall structure and specificity in Bostrychia culture isolates (Rhodomelaceae, Rhodophyta) John A. West 1, * , Curt M. Pueschel 2 , Tatyana A. Klochkova 3 , Gwang Hoon Kim 3 , Susan de Goër 4 and Giuseppe C. Zuccarello 5 1 School of Botany, University of Melbourne, Parkville VIC 3010, Australia 2 Department of Biological Sciences, State University of New York at Binghamton, Binghamton, NY 13902-6000, USA 3 Department of Biology, Kongju National University, Kongju 314-701, Korea 4 11 Rue des Moguerou, 29680 Roscoff, France 5 School of Biological Sciences, Victoria University of Wellington, PO Box 600, Wellington, 6140, New Zealand The descriptions of galls, or tumors, in red algae have been sparse. Kützing (1865) observed possible galls of Bostrychia but only presented a drawing. Intensive culture observations of hundreds of specimens of the genus Bostrychia over many years have revealed that galls appeared in only a small subset of our unialgal cultures of B. kelanensis, Bostrychia moritziana / radicans, B. radicosa, B. simpliciuscula, and B. tenella and continued to be produced intermittently or con- tinuously over many years in some cultures but were never seen in field specimens. Galls appeared as unorganized tissue found primarily on males and bisexuals, but occasionally on females and tetrasporophytes. The gall cells usually were less pigmented than neighboring tissue, but contained cells with fluorescent plastids and nuclei. The galls were not trans- ferable to other potential hosts. Galls could be produced from gall-free tissue of cultures that originally had galls even after transfer to new culture dishes. Electon microscopy of galls on one isolate (3895) showed that virus-like particles are observed in some gall cells. It is possible that a virus is the causative agent of these galls. Key Words: Bostrychia; galls; Rhodomelaceae; Rhodophyta; unialgal culture; virus-like particles INTRODUCTION Galls, or tumors, are unorganized tissue on otherwise normal plants. Galls are usually associated with abnor- mal cell division patterns and / or cell enlargement (Apt 1988, Scheffer 1997). Galls have not been reported ex- tensively in red algae possibly because they are rare and not considered important, but the deformation of tissue could have fitness consequences for the host. While the cause of all red algal galls is not known several causative agents have been shown or suggested. Cyanobacteria were reported to cause galls in Mazzaella laminarioides (Bory de Saint-Vincent) Fredericq (as Iridaea laminarioi- des Bory de Saint-Vincent) (Correa et al. 1993). Bacteria were seen in the galls of the red algae Chondracanthus teedei (Mertens ex Roth) Kützing (as Gigartina teedii [Roth] Lamouroux) (Tsekos 1982), Grateloupia ameri- cana Kawaguchi et Wang (as Prionitis lanceolata [Harvey] Harvey), and Polyneuropsis stolonifera M. J. Wynne, D. L. McBride & J. A. West (McBride et al. 1974). Subsequently bacteria were proven to be the cause of galls on G. ameri- cana (Apt and Gibor 1989, Ashen and Goff 1996, 1998, Received November 19, 2012, Accepted February 1, 2013 *Corresponding Author E-mail: [email protected] Tel: +61-3-8344-8080 This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://cre- ativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Transcript of Gall structure and specificity in Bostrychia culture ... · 411 Rue des Moguerou, 29680 Roscoff...

Algae 2013, 28(1): 83-92http://dx.doi.org/10.4490/algae.2013.28.1.083
Open Access
Research Article
Copyright © The Korean Society of Phycology 83 http://e-algae.kr pISSN: 1226-2617 eISSN: 2093-0860
Gall structure and specificity in Bostrychia culture isolates (Rhodomelaceae, Rhodophyta)
John A. West1,*, Curt M. Pueschel2, Tatyana A. Klochkova3, Gwang Hoon Kim3, Susan de Goër4 and Giuseppe C. Zuccarello5 1School of Botany, University of Melbourne, Parkville VIC 3010, Australia2Department of Biological Sciences, State University of New York at Binghamton, Binghamton, NY 13902-6000, USA 3Department of Biology, Kongju National University, Kongju 314-701, Korea411 Rue des Moguerou, 29680 Roscoff, France5School of Biological Sciences, Victoria University of Wellington, PO Box 600, Wellington, 6140, New Zealand
The descriptions of galls, or tumors, in red algae have been sparse. Kützing (1865) observed possible galls of Bostrychia
but only presented a drawing. Intensive culture observations of hundreds of specimens of the genus Bostrychia over
many years have revealed that galls appeared in only a small subset of our unialgal cultures of B. kelanensis, Bostrychia
moritziana / radicans, B. radicosa, B. simpliciuscula, and B. tenella and continued to be produced intermittently or con-
tinuously over many years in some cultures but were never seen in field specimens. Galls appeared as unorganized tissue
found primarily on males and bisexuals, but occasionally on females and tetrasporophytes. The gall cells usually were
less pigmented than neighboring tissue, but contained cells with fluorescent plastids and nuclei. The galls were not trans-
ferable to other potential hosts. Galls could be produced from gall-free tissue of cultures that originally had galls even
after transfer to new culture dishes. Electon microscopy of galls on one isolate (3895) showed that virus-like particles are
observed in some gall cells. It is possible that a virus is the causative agent of these galls.
Key Words: Bostrychia; galls; Rhodomelaceae; Rhodophyta; unialgal culture; virus-like particles
IntroductIon
Galls, or tumors, are unorganized tissue on otherwise
normal plants. Galls are usually associated with abnor-
mal cell division patterns and / or cell enlargement (Apt
1988, Scheffer 1997). Galls have not been reported ex-
tensively in red algae possibly because they are rare and
not considered important, but the deformation of tissue
could have fitness consequences for the host. While the
cause of all red algal galls is not known several causative
agents have been shown or suggested. Cyanobacteria
were reported to cause galls in Mazzaella laminarioides
(Bory de Saint-Vincent) Fredericq (as Iridaea laminarioi-
des Bory de Saint-Vincent) (Correa et al. 1993). Bacteria
were seen in the galls of the red algae Chondracanthus
teedei (Mertens ex Roth) Kützing (as Gigartina teedii
[Roth] Lamouroux) (Tsekos 1982), Grateloupia ameri-
cana Kawaguchi et Wang (as Prionitis lanceolata [Harvey]
Harvey), and Polyneuropsis stolonifera M. J. Wynne, D. L.
McBride & J. A. West (McBride et al. 1974). Subsequently
bacteria were proven to be the cause of galls on G. ameri-
cana (Apt and Gibor 1989, Ashen and Goff 1996, 1998,
Received November 19, 2012, Accepted February 1, 2013
*Corresponding Author
E-mail: [email protected]: +61-3-8344-8080
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://cre-ativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.

Algae 2013, 28(1): 83-92
http://dx.doi.org/10.4490/algae.2013.28.1.083 84
rochrome DAPI (4′,6-diamidino-2-phenylindole, Sigma-
Aldrich, St. Louis, MO, USA) using a heat fixation method.
Algal thalli were dipped in 5 µg mL-1 DAPI solution in
seawater for 5 min, and then the cover slips were slightly
heated over a boiler for a few seconds. After staining, algal
thalli were mounted on slides in the DAPI solution and
were examined under a UV filter.
Micrographs were taken with Olympus DP50 digital
camera affixed to an Olympus BX50 microscope (Olym-
pus, Tokyo, Japan) using Viewfinder Lite and Studio Lite
computer programs (Better Light Inc., Placerville, CA,
USA) or with a Zeiss GFL bright field microscope (Carl
Zeiss AG, Oberkochen, Germany) using a Canon G3 cam-
era (Canon Inc., Tokyo, Japan) and Photoshop CS4 com-
puter program ( http://www.adobe.com/au/).
Transmission electron microscopy (TEM)
Galls were fixed in phosphate buffered saline (PBS) buf-
fer containing 2% glutaraldehyde at 4°C for 2 h. The glu-
taraldehyde was then rinsed out with PBS buffer and the
cells were postfixed with 2% osmium tetroxide at 4°C for
1.5 h. Thereafter, the cells were rinsed out with PBS buffer
and were dehydrated in a graded acetone series, embed-
ded in Spurr’s epoxy resin (Spurr 1969) and polymerized
overnight in a 70°C oven (Polysciences Inc., Warrington,
PA, USA). Sections stained with uranyl acetate and Reyn-
olds’s lead citrate (Reynolds 1963) were viewed and pho-
tographed on a Phillips Bio Twin Transmission Electron
Microscope (Phillips Electron Optics, Eindhoven, Nether-
lands). We were able to carry out TEM studies on only one
B. simpliciuscula isolate (3895).
rESuLtS
Refer to Table 1 for the time periods (years) in which
galls were seen in the various isolates. These galls were
predominantly found in isolates from Australia where
most isolates were obtained. Only a small percentage of
our isolates had galls and they were only observed in B.
simpliciuscula Harvey ex J. Agardh, B. moritziana / radi-
cans complex, B. radicosa (Itono) J. A. West, G. C. Zucca-
rello & M. H. Hommersand, B. kelanensis Grunow, and B.
tenella (Lamouroux) J. Agardh.
Bostrychia simpliciuscula
B. simpliciuscula is a polyphyletic species consisting
of three lineages (Zuccarello et al. 1999, Zuccarello and
2000, Ashen et al. 1999). A fungus possibly causes galls of
Catenella nipae Zanardini (Zuccarello 2008). The galls of
Prionitis and Catenella were originally described as red
algal parasites (Lobocolax deformans Howe and Catenel-
locolax leeuwenii Weber-van Bosse, respectively). Experi-
mental infection and formation of galls was not achieved
with fungi in Catenella nipae (Zuccarello 2008).
Galls have also been associated with viruses. Virus-
like particles (VLPs) were found in the galls of Gracilaria
epihippisora Hoyle (Apt and Gibor 1991). In this species
gall tissue was capable of autonomous growth but only in
an undifferentiated state. Pueschel (1995) also observed
VLPs in the filamentous red alga Acrochaetium savianum
(Meneghini) Nägeli (as Audouinella saviana).
We have studied Bostrychia over many years and it has
become useful in research on speciation, ecophysiology,
evolution and cell biology / video microscopy of repro-
duction (reviewed in Zuccarello and West 2011). A very
large collection of over 1,000 isolates of all the recognized
species of Bostrychia has been established (http://www.
botany.unimelb.edu.au/West). A small subset of these
isolates has produced undifferentiated tissue that persist-
ed in culture. Galls of Bostrychia were observed in 1865,
when Kützing illustrated many species of Bostrychia and
depicted a gall on B. cornifera Montagne (Kützing 1865,
Pl. 24) (Fig. 1A in this paper), later synonymized with B.
moritziana (Sonders ex Kützing) J. Agardh (King and Put-
tock 1989). Our observations on Bostrychia galls are pre-
sented below.
MAtErIALS And MEtHodS
Algal material and laboratory culture
Unialgal culture methods were described in West and
Zuccarello (1999) and West (2005). Culture isolates were
all maintained at 18-23°C, 12 : 12 LD daily photoperiods,
3-5 µmol photons m-2 s-1 cool white fluorescent or LED
lighting, MPM/2 culture medium (30‰ salinity). For fast-
er growth and reproduction cultures were placed in 10-15
µmol photons m-2 s-1 on rotary shaker (70 rpm).
Most isolates used for this research program are now
available at the Korean Marine Plants Collection, Chun-
gnam National University, 220 Gung-dong, Yuseong-gu,
Daejeon, Korea.
Bright field and fluorescence microscopy
Algal nuclei were stained with the DNA-specific fluo-

West et al. Galls of Bostrychia
85 http://e-algae.kr
Fig. 1. Morphology of galls in Bostrychia. (A) Drawing of B. cornifera (currently B. moritziana) with several ‘galls’ developing on a monosiphonous branch (arrows). Reproduced from Kützing (1865). (B-E) B. simpliciuscula microscopic images of galls developing on polysiphonous axis and laterals in different isolates (3895, 3931, and 3910). (D & E) Through-focus images of the same gall (3910). (F) B. simpliciuscula, fluorescent DAPI staining of the gall cells nuclei (blue color, DAPI-stained nuclei; red color, plastid autofluorescence; arrow points to the dead cells inside the gall). (G & H) B. simpliciuscula, transmission electron microscopy images of virus-like particles in gall tissues of isolate 3895. Scale bars represent: B-F, 50 µm; G & H, 200 nm.
A C
D E
B
F
HG
Algae 2013, 28(1): 83-92
http://dx.doi.org/10.4490/algae.2013.28.1.083 86
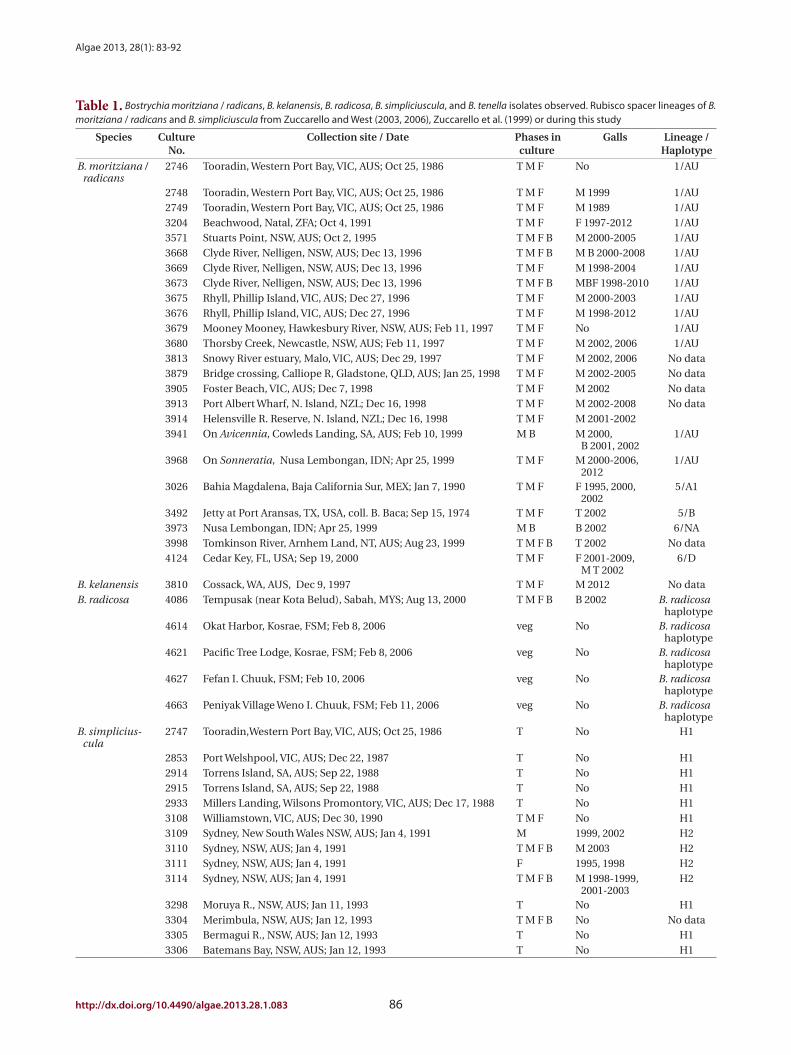
Table 1. Bostrychia moritziana / radicans, B. kelanensis, B. radicosa, B. simpliciuscula, and B. tenella isolates observed. Rubisco spacer lineages of B. moritziana / radicans and B. simpliciuscula from Zuccarello and West (2003, 2006), Zuccarello et al. (1999) or during this study
Species Culture No.
Collection site / Date Phases in culture
Galls Lineage / Haplotype
B. moritziana / radicans
2746 Tooradin, Western Port Bay, VIC, AUS; Oct 25, 1986 T M F No 1/AU
2748 Tooradin, Western Port Bay, VIC, AUS; Oct 25, 1986 T M F M 1999 1/AU2749 Tooradin, Western Port Bay, VIC, AUS; Oct 25, 1986 T M F M 1989 1/AU3204 Beachwood, Natal, ZFA; Oct 4, 1991 T M F F 1997-2012 1/AU3571 Stuarts Point, NSW, AUS; Oct 2, 1995 T M F B M 2000-2005 1/AU3668 Clyde River, Nelligen, NSW, AUS; Dec 13, 1996 T M F B M B 2000-2008 1/AU3669 Clyde River, Nelligen, NSW, AUS; Dec 13, 1996 T M F M 1998-2004 1/AU3673 Clyde River, Nelligen, NSW, AUS; Dec 13, 1996 T M F B MBF 1998-2010 1/AU3675 Rhyll, Phillip Island, VIC, AUS; Dec 27, 1996 T M F M 2000-2003 1/AU3676 Rhyll, Phillip Island, VIC, AUS; Dec 27, 1996 T M F M 1998-2012 1/AU3679 Mooney Mooney, Hawkesbury River, NSW, AUS; Feb 11, 1997 T M F No 1/AU3680 Thorsby Creek, Newcastle, NSW, AUS; Feb 11, 1997 T M F M 2002, 2006 1/AU3813 Snowy River estuary, Malo, VIC, AUS; Dec 29, 1997 T M F M 2002, 2006 No data3879 Bridge crossing, Calliope R, Gladstone, QLD, AUS; Jan 25, 1998 T M F M 2002-2005 No data3905 Foster Beach, VIC, AUS; Dec 7, 1998 T M F M 2002 No data3913 Port Albert Wharf, N. Island, NZL; Dec 16, 1998 T M F M 2002-2008 No data3914 Helensville R. Reserve, N. Island, NZL; Dec 16, 1998 T M F M 2001-20023941 On Avicennia, Cowleds Landing, SA, AUS; Feb 10, 1999 M B M 2000,
B 2001, 20021/AU
3968 On Sonneratia, Nusa Lembongan, IDN; Apr 25, 1999 T M F M 2000-2006, 2012
1/AU
3026 Bahia Magdalena, Baja California Sur, MEX; Jan 7, 1990 T M F F 1995, 2000, 2002
5/A1
3492 Jetty at Port Aransas, TX, USA, coll. B. Baca; Sep 15, 1974 T M F T 2002 5/B3973 Nusa Lembongan, IDN; Apr 25, 1999 M B B 2002 6/NA3998 Tomkinson River, Arnhem Land, NT, AUS; Aug 23, 1999 T M F B T 2002 No data4124 Cedar Key, FL, USA; Sep 19, 2000 T M F F 2001-2009,
M T 20026/D
B. kelanensis 3810 Cossack, WA, AUS, Dec 9, 1997 T M F M 2012 No dataB. radicosa 4086 Tempusak (near Kota Belud), Sabah, MYS; Aug 13, 2000 T M F B B 2002 B. radicosa
haplotype4614 Okat Harbor, Kosrae, FSM; Feb 8, 2006 veg No B. radicosa
haplotype4621 Pacific Tree Lodge, Kosrae, FSM; Feb 8, 2006 veg No B. radicosa
haplotype4627 Fefan I. Chuuk, FSM; Feb 10, 2006 veg No B. radicosa
haplotype4663 Peniyak Village Weno I. Chuuk, FSM; Feb 11, 2006 veg No B. radicosa
haplotypeB. simplicius-
cula2747 Tooradin,Western Port Bay, VIC, AUS; Oct 25, 1986 T No H1
2853 Port Welshpool, VIC, AUS; Dec 22, 1987 T No H12914 Torrens Island, SA, AUS; Sep 22, 1988 T No H12915 Torrens Island, SA, AUS; Sep 22, 1988 T No H12933 Millers Landing, Wilsons Promontory, VIC, AUS; Dec 17, 1988 T No H13108 Williamstown, VIC, AUS; Dec 30, 1990 T M F No H13109 Sydney, New South Wales NSW, AUS; Jan 4, 1991 M 1999, 2002 H23110 Sydney, NSW, AUS; Jan 4, 1991 T M F B M 2003 H23111 Sydney, NSW, AUS; Jan 4, 1991 F 1995, 1998 H23114 Sydney, NSW, AUS; Jan 4, 1991 T M F B M 1998-1999,
2001-2003H2
3298 Moruya R., NSW, AUS; Jan 11, 1993 T No H13304 Merimbula, NSW, AUS; Jan 12, 1993 T M F B No No data3305 Bermagui R., NSW, AUS; Jan 12, 1993 T No H13306 Batemans Bay, NSW, AUS; Jan 12, 1993 T No H1

West et al. Galls of Bostrychia
87 http://e-algae.kr
Species Culture No.
Collection site / Date Phases in culture
Galls Lineage / Haplotype
3308 Broughton Ck., NSW, AUS; Jan 11, 1993 T M F T 1995, M 1998-1999, 2002, F 1996, 1998, 2002
H2
3315 Tuross Lake, NSW, AUS; Jan 12, 1993 T No H1
3317 Narooma, NSW, AUS; Jan 12, 1993 T No H1
3319 Narooma, NSW, AUS; Jan 12, 1993 T M F B B 2002, M 2003 H1
3321 Wapengo Ck., NSW, AUS; Jan 11, 1993 T No H1
3322 Merimbula, NSW, AUS; Jan 12, 1993 F No H1
3330 Garden Island, SA, AUS; Jan 22, 1993 F No H1
3546 Brunswick Heads, NSW, AUS; Oct 23, 1995 T No H3
3562 Forster, NSW, AUS; Oct 23, 1995 T M F Only on M, at 17°C, not 23°C
H2
3576 Woolooware Bay, NSW, AUS; Dec 14, 1995 T No H1
3581 Williamstown, VIC, AUS; Feb 29, 1996 T M F B F 2002 H1
3612 Jawbone Reserve, Williamstown, VIC, AUS; Jun 21, 1996 T M F B B 2004 H1
3657 Sussex Inlet, NSW, AUS; Dec 14, 1996 T M F M 2004-2006, 2009
H1
3658 Sussex Inlet, NSW, AUS; Dec 14, 1996 T M F M 2001-2004, B 2003, 2005, 2009
H1
3663 Bermagui, NSW, AUS; Dec 13, 1996 T M F B B M 1999-2012 H1
3671 Merimbula, NSW, AUS; Dec 14, 1996 T M F B B M 2000-2001, 2003-2006
H1
3672 Sussex Inlet, NSW, AUS; Dec 14, 1996 T No H1
3677 Rhyll, Phillip I., VIC, AUS; Dec 27, 1996 T M F B B 2002 H1
3814 Corringle Beach, VIC, AUS; Jan 1, 1998 T M F No H1
3895 American R., Kangaroo I., SA, AUS; Sep 19, 1998 T M F M 2001-2012 H1
3897 Wynyard Wharf, TAS, AUS; Sep 13, 1998 T M F M 2003-2010 H1
3900 Batman Bridge, S. of George Town, TAS, AUS; Nov 17, 1998 T M F No H1
3910 Mimosa Rock National Park, NSW, AUS; Dec 16, 1998 M 2000-2010 No data
3912 Wapengo Lake, NSW, AUS; Dec 16, 1998 T No No data
3915 Kiama, Minnamura R., NSW, AUS; Dec 19, 1998 T No No data
3916 Kiama, Minnamura R., NSW, AUS; Dec 19, 1998 T No No data
3917 Sussex Inlet, NSW, AUS; Dec 19, 1998 T F No No data
3918 Sussex Inlet, NSW, AUS; Dec 19, 1998 T No No data
3929 Tumby Bay, SA, AUS; Feb 10, 1999 T B B 2001-2006 No data
3930 Tumby Bay, SA, AUS; Feb 10, 1999 M B M B 2000-2012 No data
3931 Tumby Bay, SA, AUS; Feb 10, 1999 T M F M 2003, 2006 No data
3932 Arno Bay, SA, AUS; Feb 10, 1999 T M F B B F 2002-2012 No data
3938 Blanche Harbour, SA, AUS; Feb 10, 1999 T M F No No data
3940 Blanche Harbour, SA, AUS; Feb 10, 1999 T M F B M 2001, B 2002-2012
No data
4042 Price, SA, AUS; Jan 13, 2000 F No No data
4045 Port Clinton, SA, AUS; Jan 10, 2000 T B B 2004-2012 No data
4046 Pt. Arthur, SA, AUS; Jan 10, 2000 T No No data
4203 Woolooware Bay, NSW, AUS; Oct 15, 2001 M No No data
4600 Lehn Mesi R., Pohnpei, FSM; Feb 4, 2006 NR No H3
4629 Fefan I. Chuuk, FSM; Feb 10, 2006 NR No H3
4636 Old Taliafak Bridge, GUM; Feb 12, 2006 NR No H3
B. tenella 2751 Initao, Misamis Oriental, PHI; Oct 25, 1986 T M F F 2003-2012 No data
VIC, Victoria; AUS, Australia; T, tetrasporophyte; M, male; F, female; ZFA, South Africa; NSW, New South Wales; B, bisexual; NZL, New Zealand; SA, South Australia; IDN, Indonesia; MEX, Mexico; NA, not available; WA, Western Australia; MYS, Malaysia; FSM, Micronesia; TAS, Tasmania; NR, no re-production; GUM, Guam; PHI, Philippines.
Table 1. continued

Algae 2013, 28(1): 83-92
http://dx.doi.org/10.4490/algae.2013.28.1.083 88
West 2003, 2006, 2011) and galls were found on isolates
from three lineages (1, 5, and 6). Again the preponder-
ance of galls was found on isolates from Australia.
In 26 years of culture isolate 2748 produced galls only
once (May 1999) on the male spermatangial stichidia and
these were very similar in overall structure to those seen
in B. simpliciuscula. On the female of isolate 2749 galls
were noted only in 1989. By contrast, isolate 2747 from
the same locality never had galls. Isolate 3026 female
formed galls at irregular intervals in 1995, 2000, and 2002
whereas the tetrasporophyte and male showed no gall de-
velopment during 22 years of culture. Isolate 3492 (Texas,
USA) was isolated in 1974 and did not show galls until
July, 2002 and these developed primarily on cladohaptera
of tetrasporophytes.
Isolate 3204 female (South Africa) was collected in 1991
and had numerous galls on procarpic lateral branches
from 1997-2012 (Fig. 2A & B). Males and tetrasporophytes
had no galls. Isolate 4124 female (Florida, USA) was col-
lected in 2000 and developed galls with mostly colorless
living cells on the tips of vegetative laterals. Regeneration
of viable branches frequently occurred from gall tissue
(Fig. 2C & D). Galls were evident for almost 8 years (2001-
2009). Males and tetrasporophytes produced galls briefly
in 2002.
Galls were observed on 16 males, 4 females, 4 bisexu-
als, and 3 tetrasporophytes of all isolates in Table 1.
Bostrychia radicosa
Isolate 4086 (Sabah, Malaysia) was obtained in Au-
gust 2000 and showed gall formation on the nodes and
gametangial sectors of the bisexual gametophytes first in
May, 2002. Isolate 4178 (New Caledonia) males developed
galls in August, 2012 (Fig. 3A & B). Initially the gall cells
enlarged and retained fully pigmented chloroplasts (Fig.
3A) but as the galls matured pale hypertrophied cells with
enlarged vacuoles were evident (Fig. 3B). Other isolates of
B. radicosa from Thailand (4207) and Micronesia (4614,
4621, 4627, and 3662) did not develop galls.
Bostrychia kelanensis
In 31 isolates of B. kelanensis from Australia, Guam, In-
dia, Indonesia, Malaysia, and Micronesia only one male
(3810) from Western Australia developed galls. Grouped
cells divided and enlarged, projecting from tier cells (Fig.
3C) in spermatangial sectors of lateral branches. Eventu-
ally irregularly shaped masses developed numerous short
branch apices (Fig. 3D).
West 2006). Galls were found on 15 of 87 (17%) isolates of
lineages H1 and H2, exclusively with Australian isolates.
Galls were usually associated with viable spermatangial
stichidia, either on unisexual or bisexual plants, although
some were evident on the non-reproductive sectors. Tet-
rasporophytes and female gametophytes occasionally
had galls. This has been true over 26 years of culture for
some isolates. Galls never developed on young gameto-
phytes, only on reproductively mature gametophytes. On
isolate 3562 no galls formed on the tetrasporophytes, fe-
males or males grown at 23°C. However, galls developed
in the vicinity of spermatangial stichidia of males but not
on females or tetrasporophytes when grown at 17°C. Iso-
late 3562, having numerous galls, was placed in a culture
(on a shaker and in brighter light) for 3-6 weeks with the
male of another isolate (3108) having a long history with-
out galls. No galls developed on isolate 3108. This is only a
partial test of Koch’s postulates (http://en.wikipedia.org/
wiki/Koch’s postulates).
Galls varied in appearance and size. The initial stages
appeared as enlarged proliferating cells. Cell divisions
appeared random and very different from the polysipho-
nous tier cell division pattern of the normal host (Fig. 1B-
E). The cells in galls were often smaller than tier cells, of
various shapes and sizes and had enlarged vacuoles. Gall
cells were less pigmented than normal host tissue cells
(Fig. 1B). They appeared to have viable nuclei that often
were larger than tier-cell nuclei (Fig. 1F). In smaller galls
the cells all appeared viable but as the galls expanded
and the cell number increased some dead cells were seen
(arrow in Fig. 1F). In some larger galls new shoots arose
within the cell mass (no photo).
In many isolates galls were present intermittently, how-
ever, galls were continuously present on the male and bi-
sexual phases of 3663 (Fig. 1F) from 1999-2012 and on the
male phase of 3895 from 2001-2012 (Fig. 1B). Isolate 3932
was unusual because galls appeared on the bisexual or fe-
male thalli but not on males from 2002-2012.
Electron microscopic (TEM) observations. While TEM
fixation of cellular structure was difficult our micrographs
did show VLPs in gall cells of isolate 3895 (Fig. 1G & H).
These VLPs are of two distinct morphologies (staining dif-
ferently) approximately 70-75 nm in size and hexagonal
in shape.
Bostrychia moritziana / radicans
From 390 isolates of the B. moritziana / radicans spe-
cies-complex 24 (6%) produced galls. This species com-
plex consists of seven different lineages (Zuccarello and

West et al. Galls of Bostrychia
89 http://e-algae.kr
Fig. 2. Galls on Bostrychia moritziana / radicans isolates 3204 and 4124. (A) Habit image of 3204 female with numerous white galls on branches bearing procarps. (B) Galls at tips of procarp bearing branches. Trichogynes (tr) visible projecting from gall at lower right. (C) 4124 female with gall at tip of vegetative lateral. Two regenerating shoots developed from gall cells. (D) 4124 female gall with mass of colorless cells, a few appear dead with collapsed protoplasts but most appear to be viable living cells. Healthy, branched shoot regenerated from gall tissue. Scale bars represent: A, 1 mm; B & D, 80 µm; C, 60 µm.
A
C
B
D

Algae 2013, 28(1): 83-92
http://dx.doi.org/10.4490/algae.2013.28.1.083 90
Fig. 3. Galls on Bostrychia radicosa, isolate 4178 (A & B); B. kelanensis, isolate 3810 (C & D); B. tenella isolate 2751 (E-G). (A) B. radicosa, elongate spermatangia bearing lateral branches with two galls visible on left. Released spermatia near branch tips also visible, small developing gall with normally pigmented cells visible on far left. (B) B. radicosa, high magnification, enlarged cells lightly pigmented with enlarged vacuoles. (C) B. kelanensis, developing gall with normally pigmented cells projecting from branch. (D) B. kelanensis, older galls with extensive branch formation. (E) B. tenella, normal procarpic branches with abundant trichogynes (tr). (F) B. tenella, numerous galls on procarpic branches, trichogynes (tr). (G) B. tenella, high magnification of gall cells, tier and cortical cells with fully pigmented chloroplasts, trichogynes (tr). Scale bars represent: A & B, 50 µm; C & D, 77 µm; E & F, 100 µm; G, 25 µm.
A C D
B
E
G
F

West et al. Galls of Bostrychia
91 http://e-algae.kr
also indicate that much of the transmission is vertical
within a single host. We did not test gall transmission to
progeny of gall bearing sexual parents.
Although not tested extensively three antibiotics (Peni-
cillin G, Ciprofloxacin, and Rifampin) were routinely add-
ed to various cultures without any effect on gall presence.
During antibiotic treatments various bacteria were also
clearly present on the hosts as well.
The greater susceptibility of males and bisexuals to gall
formation in B. simpliciuscula may be due to the numer-
ous spermatangia formed on male branches and the fre-
quent release of spermatia opening more surface areas to
attachment by viruses and bacteria.
We know very little about causative agents of the galls
seen in Bostrychia, the effects of environmental stressors
on gall formation, the transmission of the causative agent
or the effects of galls on the fitness of their hosts. The
study of potential pathogens of marine red algae should
be pursued more critically.
AcKnoWLEdGEMEntS
The Australian Research Council grants (A19917056
[1999-2001]; SG0935526 [1994]; S198122824 [1998];
S005005 [2000]), Australian Biological Resources Study
program (2002-2005), and Hermon Slade Foundation
(2005-2007) partially supported this work. Many thanks
to Ulf Karsten and Doug McBride for help with collecting
samples in various localities around Australia and to Alan
Critchley (South Africa), Rosario Braga (Brazil), and E. K.
Ganesan (Venezuela).
rEFErEncES
Apt, K. & Gibor, A. 1989. Development and induction of
bacteria-associated galls on Prionitis lanceolata (Rho-
dophyta). Dis. Aquat. Org. 6:151-156.
Apt, K. E. 1988. Galls and tumor-like growths on marine mac-
roalgae. Dis. Aquat. Org. 4:211-217.
Apt, K. E. & Gibor, A. 1991. The ultrastructure of galls on the
red alga Gracilaria epihippisora. J. Phycol. 27:409-413.
Ashen, J. B., Cohen, J. D. & Goff, L. J. 1999. GC-SIM-MS de-
tection and quantification of free indole-3-acetic acid
in bacterial galls on the marine alga Prionitis lanceolata
(Rhodophyta). J. Phycol. 35:493-500.
Ashen, J. B. & Goff, L. J. 1996. Molecular identification of a
bacterium associated with gall formation in the marine
red alga Prionitis lanceolata. J. Phycol. 32:286-297.
Bostrychia tenella
In 86 isolates of B. tenella from many different geo-
graphic regions only isolate 2751 (Philippines) developed
galls and that was only after 17 years in culture. Galls oc-
curred continuously from 2003-2012 on the procarp bear-
ing female, but never on the males or tetrasporophytes.
In normal females the lateral branches were uniform in
shape and heavily corticated, bearing numerous pro-
carps (Fig. 3E). Many gall structures appeared as irregu-
lar club-shaped enlargements around the procarps (Fig.
3F). The variably-shaped cells contained fully pigmented
chloroplasts (Fig. 3G). No branch shoot proliferation oc-
curred from these galls.
dIScuSSIon
While the presence of galls is infrequent in Bostrychia
culture isolates, their similar morphology and the pres-
ence of VLP in gall cells of B. simpliciuscula implicate vi-
ruses are the causative agent. Galls have been observed
previously in other red algal cells with VLP suggesting
them as a causative agent (Apt and Gibor 1991). Tumor-
ous growth is commonly caused by viruses in higher
plants (Francki et al. 1985, Scheffer 1997).Galls were not observed in laboratory culture on any
other Bostrychia species although this could be due to
limited sampling. We have many more isolates of the B.
moritziana / radicans (390), B. simpliciuscula (87), and B.
tenella (89) than of other species but it could be that these
species were more susceptible to the causative agent of
gall formation.
It is noteworthy that almost all species with galls lack
cortication except for well-corticated B. tenella in which
only one female (2751) had galls.
We have never observed galls on any Bostrychia spe-
cies in the field although we have examined thousands
of specimens, however Kützing (1865) observed and illus-
trated possible galls on field specimens.
The ability of galls to form from healthy tissue sepa-
rated from other gall tissue, suggests that the causative
agent (possibly a virus) may be latent in cells of some
Bostrychia isolates. Latent bacteria are known in higher
plants (Francki et al. 1985), and the stimulation of their
effects (e.g., cell proliferation) could be due to stressors
in the cells. This is seen in our experiments in which galls
were induced in low temperature conditions. The inabil-
ity of galls to be transmitted from one isolate to another
in Bostrychia and in Gracilaria (Apt and Gibor 1991) may

Algae 2013, 28(1): 83-92
http://dx.doi.org/10.4490/algae.2013.28.1.083 92
Spurr, A. R. 1969. A low-viscosity epoxy resin embedding me-
dium for electron microscopy. J. Ultrastruct. Res. 26:31-
43.
Tsekos, I. 1982. Tumour-like growths induced by bacteria in
the thallus of a red alga, Gigartina teedii (Roth) Lamour.
Ann. Bot. 49:123-126.
West, J. A. 2005. Long term macroalgal culture maintenance.
In Andersen, R. A. (Ed.) Algal Culturing Techniques. Aca-
demic Press, New York, pp. 157-163.
West, J. A. & Zuccarello, G. C. 1999. Biogeography of sexual
and asexual populations in Bostrychia moritziana (Rho-
domelaceae, Rhodophyta). Phycol. Res. 47:115-123.
Zuccarello, G. C. 2008. A fungal gall of Catenella nipae (Caul-
acanthaceae, Rhodophyta) and a review of Catenelloco-
lax leeuwenii. Bot. Mar. 51:436-440.
Zuccarello, G. C. & West, J. A. 2003. Multiple cryptic spe-
cies: molecular diversity and reproductive isolation in
the Bostrychia radicans/B. moritziana complex (Rho-
domelaceae, Rhodophyta) with focus on North Ameri-
can isolates. J. Phycol. 39:948-959.
Zuccarello, G. C. & West, J. A. 2006. Molecular phylogeny of
the subfamily Bostrychioideae (Ceramiales, Rhodophy-
ta): subsuming Stictosiphonia and highlighting poly-
phyly in species of Bostrychia. Phycologia 45:24-36.
Zuccarello, G. C. & West, J. A. 2011. Insights into evolution
and speciation in the red alga Bostrychia: 15 years of re-
search. Algae 26:21-32.
Zuccarello, G. C., West, J. A., Karsten, U. & King, R. J. 1999.
Molecular relationships within Bostrychia tenuissima
(Rhodomelaceae, Rhodophyta). Phycol. Res. 47:81-85.
Ashen, J. B. & Goff, L. J. 1998. Galls on the marine red alga
Prionitis lanceolata (Halymeniaceae): specific induction
and subsequent development of an algal-bacterial sym-
biosis. Am. J. Bot. 85:1710-1721.
Ashen, J. B. & Goff, L. J. 2000. Molecular and ecological evi-
dence for species specificity and coevolution in a group
of marine algal-bacterial symbioses. Appl. Environ. Mi-
crobiol. 66:3024-3030.
Correa, J. A., Flores, V. & Sánchez, P. 1993. Deformative dis-
ease in Iridaea laminarioides (Rhodophyta): gall develo-
ment associated with an endophytic cyanobacterium. J.
Phycol. 29:853-860.
Francki, R. I. B., Milne, R. G. & Hatta, T. 1985. Atlas of plant
viruses. CRC Press, Boca Raton, FL, Vol. 1. 222 pp, Vol.
2. 284 pp.
King, R. J. & Puttock, C. F. 1989. Morphology and taxonomy
of Bostrychia and Stictosiphonia (Rhodomelaceae/Rho-
dophyta). Aust. Syst. Bot. 2:1-73.
Kützing, F. T. 1865. Tabulae phycologicae. Abbildungen der
Tange. Vol. 15. W. Koehne, Nordhausen, 36 pp.
McBride, D. L., Kugrens, P. & West, J. A. 1974. Light and elec-
tron microscopic observations on red algal galls. Proto-
plasma 79:249-264.
Pueschel, C. M. 1995. Rod-shaped virus-like particles in the
endoplasmic reticulum of Audouinella saviana (Acro-
chaetiales, Rhodophyta). Can. J. Bot. 73:1974-1980.
Reynolds, E. S. 1963. The use of lead citrate at high pH as an
electron-opaque stain in electron microscopy. J. Cell
Biol. 17:208-212.
Scheffer, R. P. 1997. The nature of disease in plants. Cam-
bridge University Press, Cambridge, 325 pp.




![ICS Summer School, Roscoff Introduction to Scientific Visualization · 2017. 5. 9. · Introduction [Scientific] visualization Chart junk (E. Tufte) Which of these visualizations](https://static.fdocuments.in/doc/165x107/60b7ce2d62e5f7154217588a/ics-summer-school-roscoff-introduction-to-scientiic-visualization-2017-5-9.jpg)













