

Functional characterization of three specific acyl...
22
Functional characterization of three specific acyl-coenzyme A synthetases 1 involved in anaerobic cholesterol degradation in Sterolibacterium denitrificans 2 Chol1S 3 4 Markus Warnke a , Tobias Jung a , Christian Jacoby a , Michael Agne a,b , Franziska Maria Feller c , Bodo Philipp c , 5 Wolfgang Seiche d , Bernhard Breit d , and Matthias Boll a,# 6 7 8 a Faculty of Biology – Microbiology, Albert-Ludwigs-University Freiburg, Freiburg, Germany 9 b Spemann Graduate School of Biology and Medicine (SGBM), Albert-Ludwigs-University Freiburg 10 c Institute of Molecular Microbiology and Biotechnology, University of Münster, Münster, Germany 11 d Institute of Organic Chemistry, Albert-Ludwigs-University Freiburg, Freiburg, Germany 12 13 14 15 16 17 18 19 20 21 22 23 24 Running title: Acyl-CoA synthetases in anaerobic steroid degradation 25 26 27 28 # Correspondent footnote: 29 M Boll, Institute for Biology II, Faculty of Biology, University of Freiburg, 30 Schänzlestr. 1, 79104 Freiburg, Germany 31 E-mail: [email protected] 32 33 AEM Accepted Manuscript Posted Online 26 January 2018 Appl. Environ. Microbiol. doi:10.1128/AEM.02721-17 Copyright © 2018 American Society for Microbiology. All Rights Reserved. on August 11, 2019 by guest http://aem.asm.org/ Downloaded from
Transcript of Functional characterization of three specific acyl...

Functional characterization of three specific acyl-coenzyme A synthetases 1
involved in anaerobic cholesterol degradation in Sterolibacterium denitrificans 2
Chol1S 3
4
Markus Warnkea, Tobias Junga, Christian Jacobya, Michael Agnea,b, Franziska Maria Fellerc, Bodo Philippc, 5
Wolfgang Seiched, Bernhard Breitd, and Matthias Bolla,# 6
7
8
aFaculty of Biology – Microbiology, Albert-Ludwigs-University Freiburg, Freiburg, Germany 9
bSpemann Graduate School of Biology and Medicine (SGBM), Albert-Ludwigs-University Freiburg 10
cInstitute of Molecular Microbiology and Biotechnology, University of Münster, Münster, Germany 11
dInstitute of Organic Chemistry, Albert-Ludwigs-University Freiburg, Freiburg, Germany 12
13
14
15
16
17
18
19
20
21
22
23
24
Running title: Acyl-CoA synthetases in anaerobic steroid degradation 25
26
27
28
#Correspondent footnote: 29
M Boll, Institute for Biology II, Faculty of Biology, University of Freiburg, 30
Schänzlestr. 1, 79104 Freiburg, Germany 31
E-mail: [email protected] 32
33
AEM Accepted Manuscript Posted Online 26 January 2018Appl. Environ. Microbiol. doi:10.1128/AEM.02721-17Copyright © 2018 American Society for Microbiology. All Rights Reserved.
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

Abstract 34
35
The denitrifying -proteobacterium Sterolibacterium denitrificans Chol1S catabolizes steroids such as 36
cholesterol via an oxygen-independent pathway. It involves enzyme reaction sequences described for 37
aerobic cholesterol and bile acid degradation as well as enzymes uniquely found in anaerobic steroid-38
degrading bacteria. Recent studies provided evidence that in Stl. denitrificans the cholest-4-en-3-one 39
intermediate is oxygen-independently oxidized to 4-dafachronic acid (C26-oic acid), which is 40
subsequently activated by a substrate-specific acyl-coenzyme A (CoA) synthetase (ACS). Further 41
degradation was suggested to proceed via unconventional -oxidation where aldolases, aldehyde 42
dehydrogenases and additional ACS substitute for classical -hydroxyacyl-CoA dehydrogenases and 43
thiolases. Here, we heterologously expressed three cholesterol-induced genes that putatively code for 44
AMP-forming ACS, and characterized two of the products as specific 3-hydroxy-5-cholenoyl-CoA (C24-45
oic acid) and pregn-4-en-3-one-22-oyl-CoA (C22-oic acid) forming ACS, respectively. A third 46
heterologously produced ATP-dependent ACS was inactive with 26-, 24-, or 22-oic-acids but activated 47
3a-H-4-(3'propanoate)-7a-methylhexahydro-1,5-indanedione (HIP) to HIP-CoA, a rather late 48
intermediate of aerobic cholesterol degradation that still contains the CD-rings of the sterane skeleton. 49
This work provides experimental evidence that anaerobic steroid degradation proceeds via numerous 50
alternate CoA-ester-dependent or -independent enzymatic reaction sequences as a result of aldolytic 51
side-chain and hydrolytic sterane ring C–C-bond cleavages. The aldolytic side-chain degradation pathway 52
comprising highly exergonic ACS and aldehyde dehydrogenases is considered to be essential for driving 53
the unfavorable oxygen-independent C26 hydroxylation forward. 54
55
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

Importance 56
57
The biological degradation of ubiquitously abundant steroids is hampered by their low solubility and the 58
presence of two quaternary carbon atoms. The degradation of cholesterol by aerobic Actinobacteria has 59
been studied in detail since more than thirty years and involves a number of oxygenase-dependent 60
reactions. In contrast, much less is known about oxygen-independent degradation of steroids in 61
denitrifying bacteria. In the cholesterol-degrading anaerobic model organism Sterolibacterium 62
denitrificans Chol1S initial evidence has been obtained that steroid degradation proceeds via numerous 63
alternate CoA-ester dependent/independent reaction sequences. Here we describe the heterologous 64
expression of three highly specific and characteristic acyl-CoA synthetases, two of which play a key role 65
in the degradation of the side-chain whereas a third one is specifically involved in the B-ring 66
degradation. The results obtained shed light into oxygen-independent steroid degradation comprising 67
more than 40 enzymatic reactions. 68
69
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

Introduction 70
71
Steroids constitute a highly abundant class of natural compounds with a common sterane ring system. 72
They play essential roles as components of biological membranes and signaling molecules (1). Steroids 73
serve as growth substrate for certain Actinobacteria and Proteobacteria that use them as carbon and 74
together with an electron acceptor as energy source. Bacterial degradation represents the major means 75
of eliminating steroids from the environment (2, 3, 4). Research on microbial steroid degradation has 76
also a biotechnological impact as enzymes transforming steroids may be used for the synthesis of value-77
added products, e.g. the specific conversion of cholecalciferol to 25-hydroxyvitamin D3 by 78
monooxygenases (5) or oxygen- and electron donor-independent hydroxylases (6). Finally, steroid 79
degradation is medically relevant, because Mycobacterium tuberculosis is known to use cholesterol from 80
macrophages as growth substrate during intracellular survival (7, 8). 81
82
Aerobic steroid degradation has mainly been studied in several cholesterol degrading Actinobacteria 83
such as the model organisms Rhodococcus jostii RHA1 and Mycobacterium tuberculosis H37Rv (9), while 84
aerobic degradation of steroidal core ring has mainly been studied in Commamonas testosterone (10). 85
Bile salts degradation has originally been studied in Pseudomonas species but recently also in R. jostii 86
(11, 12, 13, 14). 87
88
During both, aerobic and anaerobic cholesterol degradation, catabolism is initiated by the oxidation of 89
ring A by 3-hydroxysteroid dehydrogenases or cholesterol oxidases and 3-ketosteroid-∆1-90
dehydrogenases yielding cholest-4-en-3-one and cholesta-1,4-diene-3-one, respectively (15, 16, 17). The 91
further degradation involves the oxidation of C26 primary carbon atom to 4-dafachronic acid (26-oic 92
acid; Fig. 1). The enzymes involved in this reaction sequence fundamentally differ in aerobic and 93
anaerobic bacteria. In aerobic Actinobacteria, the cytochrome p450 oxygenases Cyp125 or Cyp142 94
catalyze C26 hydroxylation and further oxidation to the C26-oic acid (18, 19). In contrast the denitrifying 95
Sterolibacterium denitrificans first hydroxylates the tertiary C25 with water by a Mo-dependent steroid 96
C25 dehydrogenase (C25DH) (20) followed by an apparent hydroxyl shift from tertiary C25 to primary 97
C26 by an unknown enzyme (21). The subsequent oxidation of the primary alcohol to the C26-oic acid is 98
then achieved by the action of putative cholesterol-induced alcohol and aldehyde dehydrogenases (22). 99
In both, aerobic and anaerobic cholesterol-degrading bacteria, an ATP-dependent acyl-coenzmye A 100
(CoA) synthetase (ACS) specific for C26-oic acid was identified and characterized (22, 23), (Fig. 1). 101
102
In all cholesterol degrading bacteria -oxidation of the C26-oyl-CoA intermediate yields androsta-1,4-103
diene-3,17-dione (ADD), two propionyl-CoA and one acetyl-CoA (Fig. 1). In Actinobacteria classical acyl-104
CoA dehydrogenases, enoyl-CoA hydratases, 3-hydroxyacyl-CoA dehydrogenases and thiolases are 105
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

involved (8). The only exception is the last C–C bond cleavage at a tertiary carbon atom, which is 106
catalyzed by an aldolase at the level of a -hydroxyacyl-CoA intermediate (24). In contrast, a recent 107
integrated multi omics study suggested that cholesterol side-chain degradation in Stl. denitrificans 108
proceeds via aldolytic cleavage of all 3-hydroxacyl-CoA intermediates to propionyl-CoA/acetyl-CoA and 109
the corresponding C24-/C22-aldehydes. The latter were proposed to be oxidized and activated to the 110
corresponding C24-/C22-oyl-CoAs (22). Consequently, the conventional 3-hydroxyacyl-CoA 111
dehydrogenase and thiolases would be replaced by aldolases, aldehyde dehydrogenases and ACS (Fig. 112
1). However, besides the C26-oic acid activating ACS, none of the proposed enzymes involved in anoxic 113
cholesterol side-chain degradation has been isolated and characterized, yet. Notably, the aldolytic 114
pathway proposed for cholesterol degradation in Stl. denitrificans for steroid side-chain degradation 115
resembles to that reported for aerobic cholate degradation (4, 14), demonstrating the composite 116
character of the anaerobic cholesterol degradation pathway. 117
118
After side-chain removal, degradation of the ADD formed again greatly differs in aerobic and anaerobic 119
steroid degrading bacteria. In the aerobic pathway, a series of oxygenase-dependent reactions results in 120
ring A and B cleavage via a name giving 9,10-seco intermediate, finally yielding the CD-rings containing 121
3a-H-4(3’propanoate)-7a-methylhexahydro-1,5-indanedione (HIP) and its CoA ester HIP-CoA (8). In 122
contrast, denitrifying bacteria degrade ring A by hydrolysis to 17-hydroxy-1-oxo-2,3-seco-androstan-3-123
oic acid via the 2,3-seco pathway (25). Nothing is known about ring B cleavage in anaerobic cholesterol 124
degrading denitrifying bacteria. But the identification of high-resolution masses fitting to HIP-CoA 125
suggested that this compound results from ring B cleavage in Stl. denitrificans and represents an 126
intermediate in both, aerobic and anaerobic cholesterol degradation (22) (Fig. 1). Very recently, 127
degradation of HIP-CoA to central intermediates was revealed in Actinobacteria, which involves two 128
distinct ring-cleaving hydrolases (26). Genomic analyses suggested that HIP-CoA is similarly degraded in 129
Stl. denitrificans and possibly other denitrifying bacteria (22). 130
131
In this work, we heterologously produced and characterized three ACS from Stl. denitrificans and 132
demonstrate that two of them are involved in side-chain degradation (specific for C24-, C22-oic acids 133
respectively), whereas the third ACS is involved in B-ring degradation (specific for HIP). The work 134
provides enzymatic evidence for the omics-based proposal for side-chain degradation and CD-rings 135
cleavage in an anaerobic, cholesterol-degrading model organism; it sheds light into the in large parts still 136
obscure anaerobic steroid degradation pathway. 137
138
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

Results 139
140
ACS genes involved in anaerobic cholesterol degradation. Recent MS-based analyses of metabolites in 141
Stl. denitrificans cells grown with cholesterol suggested a -oxidation-like side-chain degradation 142
pathway (21, 22, 27). With the cholesterol-induced gene product SDENChol_v1_11189, that specifically 143
catalyzed the ATP- and CoA-dependent formation of C26-oyl-CoA, the first enzyme of the proposed -144
oxidation sequence was isolated and characterized (22). Analyses of further metabolites suggested 145
aldolytic cleavage of C–C bonds during side-chain degradation, which would involve two additional C24- 146
and C22-oic acid converting ACS. Finally, a CoA-ester fitting to authentic HIP-CoA standard was identified 147
in whole cells during UPLC-coupled HRMS; its formation may be catalyzed by a further ACS (22). 148
149
Previous proteome analyses identified next to C26-oyl-CoA forming SDEN_v1_11189 three cholesterol-150
induced, ACS-like gene products (Table S1). Two of them, SDENChol_v1_10299 and SDENChol_v1_10305 151
were induced 1.45 and 2.35 (log2 ratios) in cells grown with cholesterol than with testosterone 152
respectively, which contains the sterane skeleton but which lacks the side-chain (22). This finding 153
suggests that the two ACS are involved in side-chain degradation. In contrast, a third ACS candidate 154
(SDENChol_v1_10766) was higher abundant in cells grown with cholesterol vs propionate but was not 155
induced in cells grown with cholesterol vs testosterone. Consequently, the latter is assumed to be 156
involved in sterane ring system degradation. 157
158
Heterologous production, purification and characterization of three ACS involved in anaerobic 159
cholesterol degradation. The genes encoding three putative ACS involved in anaerobic cholesterol 160
degradation, SDENChol_v1_10299, SDENChol_v1_10305, and SDENChol_v1_10766, were heterologously 161
expressed in a soluble form in E. coli BL21 with a N-terminal Strep-tag. The products were purified via 162
StrepTactin® affinity chromatography (see Material and Methods). SDS-PAGE analysis of the three highly 163
enriched enzymes revealed three soluble proteins at molecular masses of 65, 70 and 55 kDa fitting to 164
the theoretical masses of 65.1, 71.3 and 58.6 kDa with enrichment factors of 43.4, 18.4 and 33, 165
respectively (Fig. S2). 166
167
Kinetic properties of SDENChol_v1_10299, SDENChol_v1_10305, and SDENChol_v1_10766. The 168
heterologously produced ACS were tested for their substrate preference using numerous carboxylic 169
acids including 4-dafachronic acid (C26), 3-hydroxy-5-cholenic acid (C24), cholic acid and its 170
analogues desoxy- and lithocholic acid (C24 each), pregn-4-en-3-one-22-oic acid (C22), HIP (C13), 171
palmitic acid (C18), decanoic acid (C10), hexanoic acid (C6) and propionate (C3) as potential substrates. 172
Concentrations of all carboxylic acids tested were 0.3 mM in the presence of 7.8% (w/v) 2-173
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

hydroxypropyl--cyclodextrin as solubilizing agent. Addition of the latter largely increased the solubility 174
of the carboxylic acid substrates and in turn the activity of the individual ACS. The time-dependent 175
formation of CoA ester products was followed by UPLC-analysis in the presence of MgATP, CoA and the 176
individual, heterologously produced ACS. The identity of the reaction products were confirmed by co-177
elution during UPLC-analyses with standards, characteristic UV/vis spectra and LC-ESI/MS analyses (Fig. 178
S1, Fig. S3 and Table S2). No conversion was observed in any case when MgATP was omitted from the 179
assay. 180
181
SDENChol_v1_10299 showed the highest activity with the 3-hydroxy-5-cholenic acid (C24-oic acid), 182
whereas virtually no conversion of the C26-, C22-oic acids and shorter carboxylic acids was observed 183
(Table 1). These results strongly indicate that cholesterol-induced SDENChol_v1_10299 is specifically 184
involved in the activation of the side-chain degradation product 3-hydroxy-5-cholenic acid to its CoA 185
ester. It also activated the C24 bile acids litocholic acid, cholic acid and deoxycholic acids to their 186
respective CoA thioester at lower rates, suggesting that the chain-length rather than modifications at 187
the sterane ring system governs the substrate preference. In contrast SDENChol_v1_10305 only 188
converted pregn-4-en-3-one-22-oic acid to its CoA ester, whereas it was apparently inactive (<0.1%) 189
with all other carboxylic acids tested (Table 1). In conclusion, the gene product is now assigned to as a 190
specific pregn-4-en-3-one-22-oyl-CoA synthetase. Notably, the 4-dafachronic acid (C26-oic acid), the 191
preferred substrate of the recently characterized SDENChol_v1_11189 was not converted by 192
SDENChol_v1_10299 or SDENChol_v1_10305. These results suggest that all three ACS are involved in 193
side-chain degradation with a strong preference for the chain-length of the individual substrates. 194
SDENChol_v1_10766 neither converted any of the C26-/C24- /C22-oic acid substrates nor any of the 195
aliphatic fatty acids. Instead, it showed a high activity with HIP, indicating that it plays a role in the 196
activation of the CD rings containing carboxylic acid (Table 1). 197
198
Using the coupled spectrophotometric assay, the initial rates of all three ACS strongly depended on the 199
substrate concentration. A fit to Michaelis-Menten curves revealed Km-values of SDENChol_v1_10299, 200
SDENChol_v1_10305, and SDENChol_v1_10766 for their individual preferred carboxylic acid substrates 201
(74-156 µM) as well as kcat-values (1.8-10.8 s-1); the results are summarized in Table 2. 202
203
ACS activities in extracts from cells grown with different substrates. The substrate preference of the 204
three ACS for side-chain-containing C24- and C22-oic acids or for HIP suggested a specific role in steroid 205
degradation, whereas they should not be required during growth with propionate. This finding is in full 206
agreement with the differential induction of the individual genes during growth on different carbon 207
sources (22). To further analyze the differential induction of ACS activities, we tested extracts grown 208
with cholesterol and propionate using different carboxylic acid substrates. The activities with the 209
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

individual carboxylic substrates of all three ACS were induced by a factor of 6 to 9 in cells grown with 210
cholesterol vs propionate strongly confirming their specific role in anaerobic cholesterol degradation 211
(Table 3). 212
213
Phylogenetic analyses of ACS involved in steroid degradation. Using BLAST a total of 14 putative genes 214
encoding ACS were identified in the genome of Stl. denitrificans (Expect value threshold = e–14). A 215
multiple sequence alignment with the clustalW program revealed that the experimentally verified ACS 216
from Stl. denitrificans specific for C26-oic acid and HIP cluster with the corresponding FadD19 and FadD3 217
enzymes involved in cholesterol degradation in M. tuberculosis H37v, R. jostii RHA1, respectively (Fig. 2). 218
In contrast, the closest related enzymes to the Stl. denitrificans ACS specific for C24-, C22-oic acids are 219
those involved in bile acid degradation in R. jostii RHA1 and Pseudomonas sp. DOC21. According to the 220
relationship to corresponding enzymes from E. coli, two further putative ACS of Stl. denitrificans most 221
likely function as acetate and propionate activating enzymes, whereas the function of the remaining five 222
putative ACS remains elusive. 223
224
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

Discussion 225
226
The three heterologously produced and characterized ACS from Stl. denitrificans Chol1S in this work, 227
together with a recently identified C26-oyl-CoA forming ACS show all remarkable specificities for their 228
individual C26-, C24-, C22-oic acid and HIP substrates, respectively. Two of which show similarities to 229
enzymes involved in aerobic cholesterol degradation (C26-oyl-CoA and HIP-CoA forming), whereas the 230
other two are rather related to enzymes involved in bile acid degradation and are absent in aerobic 231
cholesterol degradation (C24-, and C22-oyl-CoA forming). This finding corroborates the composite 232
anaerobic steroid degradation pathway involving numerous CoA-ester dependent/independent reaction 233
sequences known from aerobic cholesterol or bile acid degradation. 234
235
The three ACS involved in side-chain degradation strictly discriminate between the structurally related 236
C26-, C24-, C22-oic acids that strongly supports their assignment to individual carboxylic acid substrates 237
of cholesterol side-chain degradation. The assignment of SDENChol_v1_10766 to a highly specific HIP-238
CoA synthetase appears to be less clear as no structurally related substrate analogues that could serve 239
as potential intermediates in anaerobic steroid degradation were available. However, the kcat of the 240
enzyme from Stl. denitrificans is in a similar range (3.7 s-1) as the one reported for HIP-CoA synthetase 241
from R. jostii RHA1 (kcat 7.1 s-1) (28). Thus, the steady-state kinetic properties of SDENChol_v1_10766 242
support that HIP or a structurally closely related analogue act as an intermediate of anaerobic 243
cholesterol degradation. Moreover, previous studies identified clustered, cholesterol-induced genes in 244
Stl. denitrificans (22), that are highly similar to those involved in -oxidation of HIP-CoA to 4-methyl-5-245
oxooctanedioyl-CoA in other organisms (26). In conclusion, both findings together suggest that CD-ring 246
degradation proceeds via identical or highly similar pathways in aerobic and anaerobic cholesterol 247
degrading bacteria. 248
249
The identification of ACS specific for C24-oic and C22-oic acids in Stl. denitrificans is in full agreement 250
with the recently proposed aldolytic C–C-bond cleavage during isoprenoid side-chain -oxidation (22). 251
The classical thiolytic C–C cleavage of -oxidation yields two CoA-esters from a -ketoacyl-CoA 252
intermediate, whereas the aldolytic C–C cleaving mechanism yields an aldehyde plus acetyl-CoA or 253
propionyl-CoA. As a result the aldehyde has to be oxidized to a carboxylic acid, followed by an ACS 254
(AMP-forming) dependent thioesterification to initiate a new round of -oxidation. On the first view, 255
this strategy appears to be energetically less efficient than thiolytic cleavage as two additional 256
phosphoanhydride bonds are hydrolyzed for each -oxidation reaction sequence, assuming that the 257
pyrophosphate formed by AMP-forming ACS is readily hydrolyzed by a pyrophosphatase. Consequently, 258
anaerobic cholesterol degradation via two aldolytic side-chain cleavages yields four ATP less than via the 259
thiolytic pathway. However, the insights into the anaerobic cholesterol degradation pathway obtained 260
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

so far suggest that in total around ten acetyl-CoA/propionyl-CoA units plus one succinyl-CoA are formed 261
from cholesterol, that are fully oxidized in the tricarboxylic acid cycle. Taken into account that NADH 262
oxidation coupled to denitrification yields more than one ATP per NADH oxidized (29), the four 263
additional ATP consumed during aldolytic side-chain degradation will only slightly effect the overall ATP 264
yield. Thus, energetic considerations appear to be rather marginal when comparing the employment of 265
the aldolytic vs the thiolytic side-chain degradation pathway. A possible much more important point is 266
that the aldolytic pathway comprises with the NAD+-dependent aldehyde oxidation to a carboxylic acid 267
(G°’ ≈ –45 kJ mol-1) (30) and the CoA-ester formation by AMP-forming ACS/pyrophosphatase (G°’ ≈ –268
20 kJ mol-1) two highly exergonic steps that are missing in the thiolytic pathway. In contrast, the latter 269
involves an unfavorable reaction that is missing in the aldolytic pathway: the oxidation of -hydroxyacyl-270
CoA to -ketoacyl-CoA with NAD+ as electron acceptor (G°’ ≈ +25 kJ mol-1), that almost compensates 271
for the subsequent exergonic -ketothiolase reaction G = –25.1 kJ mol-1 (31). In conclusion, the 272
equilibrium of the aldolytic pathway is clearly more shifted in the oxidative direction in comparison to 273
the thiolytic one, which will strongly promote the unidirectionality of the overall catabolic pathway. 274
275
The question rises why denitrifying steroid degrading bacteria employ an aldolytic pathway? The most 276
probably rate limiting step in anaerobic cholesterol degradation is the formation of the C26-hydroxy 277
species via a tertiary C25-hydroxy intermediate. This conversion of a tertiary to a primary alcohol is 278
unprecedented in biology and probably highly unfavorable; it might be only possible if highly exergonic 279
downstream -oxidation reactions pull the reaction forward. This assumption is in accordance with 25-280
OH-cholest-4-en-3-one (and its ring A 1,4-diene analogue) representing the highest abundant 281
metabolites in MS-based analyses (22), indicating that its conversion is rate-limiting in the overall 282
degradation pathway. Thus, the rational of using the energetically less efficient aldolytic instead of the 283
thiolytic pathway may be to provide a virtually irreversible reaction sequence to drive an unfavorable, 284
rate-limiting reaction forward. 285
286
It is obvious, that in bacteria specialized for using fatty acids and steroids as growth substrates the 287
number of ACS is usually high (>>10). Indeed, phylogenetic analyses with Stl. denitrificans identified next 288
to the experimentally verified four ACS, ten further genes encoding putative ACS. Among them, the 289
SDENCholv1_20262 and SDENCholv1_20218 most possibly code for propionyl-CoA and acetyl-CoA 290
synthetases, respectively, according to their similarities to experimentally verified enzymes (Fig. 2). The 291
function of the remaining seven putative ACS cannot be easily predicted. Likely functions are the 292
activation of aliphatic fatty acids with chain-length longer than C3, e.g. palmitic acid, a known growth 293
substrate of Stl. denitrificans (32), or the activation of C26-oic acid analogues during growth with -294
sitosterol, stigmasterol or ergosterol (6). Finally, hydrolytic cleavage of ring A yields in denitrifying 295
steroid degraders 1,17-dioxo-2,3-seco-androstan-3-oic acid (DSAO) (25), that needs to be activated to a 296
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

CoA thioester to initiate a new round of -oxidation (22). Taken together, Stl. denitrificans contains a 297
highly versatile inventory of ACS as a result of its specialization for steroid and fatty acid growth 298
substrates. 299
300
Material and Methods 301
302
Materials. Cholest-4-en-3-one-26-oic acid (analytical standard), pregn-4-en-3-one-22-oic acid, 3β-303
hydroxycholenic acid, lithocholic acid, cholic acid, deoxycholic acid, palmitic acid, decanoic acid, 304
hexanoic acid and propionic acid and CoA were purchased from Sigma-Aldrich (Darmstadt, Germany), 305
Cayman Chemicals (Ann Arbor, Michigan, USA) or Santa Cruz Biotechnology (Heidelberg, Germany). In 306
addition to the HIP synthesized by the authors (see below), it was kindly provided by Prof. L. Eltis (Dept. 307
Microbiology & Immunology, University of British Columbia, Vancouver, Canada). Other chemicals and 308
reagents were of analytical or HPLC grade. Sterolibacterium denitrificans Chol1S (DSM = 13999) was 309
obtained from the Deutsche Sammlung für Mikroorganismen und Zellkulturen (DSMZ; Braunschweig, 310
Germany). 311
312
Cloning of acs genes. Cholesterol-grown cells were used as template for extraction of genomic DNA 313
using the Illustra bacteria genomicPrep (GE Healthcare). The genes encoding putative ACS were 314
amplified with specific primers (Table 4); a N-terminal Strep-tag was fused to each subunit. After 315
restriction and ligation into the vector pASK-IBA15plus (IBA Lifesciences) the construct was transformed 316
into Escherichia coli BL21(DE3) (New England Biolabs). 317
318
Heterologous expression of genes and protein purification. E. coli cells expressing the individual genes 319
encoding ACS were grown in LB-media at 30 °C. At an OD600 of 0.6, gene expression was induced with 320
250 µM isopropyl β-D-1-thiogalactopyranoside. Growth temperature was subsequently set to 16°C and 321
cells were harvested after 18 h. 5 g cells were suspended in buffer A (50 mM HEPES/HCl pH 7.5, 150 mM 322
potassium chloride) with 0.1 mg ml-1 DNAse I and lysed using a French pressure cell. The cell lysate was 323
ultracentrifuged and applied to a Strep-Tactin® affinity column (GE Healthcare) using an FPLC System 324
(ÄKTA purifier, GE Healthcare). Removal of non-specifically bound proteins and elution of the 325
heterologously expressed proteins was conducted using buffer A and buffer A containing 5 mM 326
desthiobiothin, respectively. The eluted protein was concentrated using a centrifugal concentrator 327
(Sartorius). Protein concentration was determined with the Bradford method (33) and protein purity 328
was determined by SDS-PAGE. 329
330
Spectrophotometric enzyme assays. For determination of specific ACS activities in cell extracts the 331
time-dependent formation of CoA esters was followed using a discontinuous, ultra performance liquid 332
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

chromatography (UPLC®)-based assay (see below). Specific activities of heterologously produced ACS 333
were conducted using a spectrophotometric assay that coupled AMP formation to NADH oxidation as 334
described (28) with slight modifications. Reactions were performed in a total volume of 0.1 mL with 0.1 335
M HEPES buffer (pH 8), containing 1 mM CoA, 1.5 mM ATP, 1 mM PEP, 4 units pyruvate kinase, 4 units 336
adenylate kinase, 4 units lactate dehydrogenase, 500 µM of NADH, 2.5 mM MgCl2, 2 mM DTE, 7.8% 337
(w/v) 2-hydroxypropyl--cyclodextrin and 787 nM SDENchol_10299, 328 nM SDENchol_10305 or 820 338
nM SDENchol_10766. The mixture was incubated 5 min at 30°C and the reaction was initiated by adding 339
300 µM of the respective carboxylic acid substrate (from 5-10 mM stock, dissolved in 70% isopropanol). 340
All measurements were recorded on a Varian Cary 100 BIO UV/Vis spectrophotometer with the standard 341
software CaryWinUV Kinetics (Version 3.00). 342
343
Synthesis of HIP. A mixture of 5α-OH-HIP and 7β-OH-HIP was purified from cultures of P. stutzeri Chol1 344
Δscd3A with 12 mM succinate and 1 mM deoxycholate as described (34). After complete transformation 345
of deoxycholate to HIPs, the supernatant was acidified to pH 2-3 with HCl and extracted with 346
ethylacetate. HIPs were resolved in MilliQ-H2O. Complete transformation of deoxycholate and purity of 347
HIPs was determined by LC-MS analysis as described for other steroid compounds (35). 348
349
Ultra performance liquid chromatography (UPLC®) analysis of CoA esters. The products formed by 350
purified ACS or cell extracts were analyzed by an UPLC-based assay using a Waters Acquity H-class UPLC 351
system equipped with diode array detector (Waters, Eschborn, Germany) and a Knauer Bluespher 100-2 352
C18 column (2 mm x 100 mm, 2 µm particle size, Knauer, Berlin, Germany). Samples (50 µL) were 353
precipitated by addition of 25 µL 50% (v/v) 1 M HCl or 100 µL MeOH and the supernatant was applied 354
onto the column. If necessary (low concentrations) CoA-esters were enriched by solid phase extraction 355
as described elsewhere (36). For UPLC analyses, a gradient of 15-90 % acetonitrile in 10 mM ammonium 356
acetate buffer and a flow rate of 0.3 mL min-1 were used for separation. Identification of the individual 357
CoA esters was accomplished by their retention times, their UV/vis spectra and by high resolution mass 358
spectrometry (HRMS, see below), (Table S2). 359
360
Liquid chromatography-mass spectrometry (LC-MS). CoA-esters were analyzed using a Waters Acquity 361
I-class UPLC system with a Knauer Bluespher 100-2 C18 column (2 mm x 100 mm, 2 µm particle size, 362
Knauer, Berlin, Germany) coupled to a Waters Synapt G2-Si HDMS electrospray ionization 363
(ESI)/quadrupole time-of-flight (Q-TOF) system (Waters, Eschborn, Germany). A gradient of 15-90 % 364
acetonitrile in 10 mM ammonium acetate buffer and a flow rate of 0.3 mL/min were used for 365
separation. Any compounds were measured in MS positive mode with a capillary voltage of 3 kV, 150 °C 366
source temperature, 450°C desolvation temperature, 1000 L min–1 N2 desolvation gas flow and 100 L 367
min–1 N2 cone gas flow. Collision induced dissociation of precursor ions was performed using a collision 368
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

energy of 20 V. LC/UV-vis analyses were conducted on a Waters Acquity H-class UPLC with a Knauer 369
Eurospher C18 column and the same gradient as described above. Evaluation of LC/MS metabolite data 370
was performed using MassLynx (Waters); for evaluation of LC/UV-vis data MassLynx or Empower 371
(Waters) was used. CoA esters were verified by detection of their characteristic fragment ion at m/z = 372
428.0367 (35). 373
374
Phylogenetic analyses. The protein sequences of ACS detected in the genome of Stl. denitrificans were 375
aligned with a set of published sequences (identification numbers are given in brackets). Phylogenetic 376
analyses were done at the server of www.phylogeny.fr (37) by using the MUSCLE algorithm (default 377
parameters), Gblocks was used for data curation (default parameters) and PhyML for phylogenetic 378
tree construction by maximum-likelihood method, built with 3000 bootstrap replicates. The resulting 379
tree was exported as newick format and visualized in MEGA 7 (38). 380
381
382
ACKNOWLEDGEMENTS 383
This work was funded by the German research council (DFG, BO 1565, 10-2, 14-1). We thank Mario 384
Mergelsberg, University of Freiburg for the help with MS measurements and data handling. We thank 385
Dr. Lindsay D. Eltis, University of British Columbia for providing substrates for enzymatic assays. 386
387 on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

References 388
1. Nes WD. 2011. Biosynthesis of cholesterol and other sterols. Chem Rev 111:6423-51. 389
2. Khanal SK, Xie B, Thompson ML, Sung S, Ong SK, Van Leeuwent J. 2006. Fate, transport, and 390
biodegradation of natural estrogens in the environment and engineered systems. Environ Sci 391
Technol 40:6537-46. 392
3. Barbosa MO, Moreira NF, Ribeiro AR, Pereira MF, Silva AM. 2016. Occurrence and removal of 393
organic micropollutants: An overview of the watch list of EU Decision 2015/495. Water Res 394
94:257-79. 395
4. Holert J, Kulić Ž, Yücel O, Suvekbala V, Suter MJ, Möller HM, Philipp B. 2013. Degradation of the 396
acyl side chain of the steroid compound cholate in Pseudomonas sp. strain Chol1 proceeds via 397
an aldehyde intermediate. J Bacteriol 195:585-95. 398
5. Wikvall K. 2001. Cytochrome P450 enzymes in the bioactivation of vitamin D to its hormonal 399
form (review). Int J Mol Med 7:201-9. 400
6. Warnke M, Jung T, DermerJ, Hipp K, Jehmlich N, Bergen M, Ferlaino S, Fries S, Müller M, Boll M. 401
2016. 25-Hydroxyvitamin D3 Synthesis by Enzymatic Steroid Side-Chain Hydroxylation with 402
Water. Angewandte Chemie (International ed. in English) 55 (5):1881–1884. 403
7. García JL, Uhía I, Galán B. 2012. Catabolism and biotechnological applications of cholesterol 404
degrading bacteria. Microb Biotechnol 5:679-99. 405
8. Wipperman MF, Sampson NS, Thomas ST. 2014. Pathogen roid rage: cholesterol utilization by 406
Mycobacterium tuberculosis. Crit Rev Biochem Mol Biol 49:269-93. 407
9. Van der Geize R, Yam K, Heuser T, Wilbrink MH, Hara H, Anderton MC, Sim E, Dijkhuizen L, 408
Davies JE, Mohn WW, Eltis LD. 2007. A gene cluster encoding cholesterol catabolism in a soil 409
actinomycete provides insight into Mycobacterium tuberculosis survival in macrophages. Proc 410
Natl Acad Sci USA 104:1947-52. 411
10. Horinouchi M, Hayashi T, Kudo T. 2012: Steroid degradation in Comamonas testosteroni. J 412
Steroid Biochem Mol Biol 129:4–14. 413
11. Mohn WW, Wilbrink MH, Casabon I, Stewart GR, Liu J, van der Geize R, Eltis LD. 2012. Gene 414
cluster encoding cholate catabolism in Rhodococcus spp. J Bacteriol 194:6712-9. 415
12. Holert J, Alam I, Larsen M, Antunes A, Bajic VB, Stingl U, Philipp B. 2013. Genome Sequence of 416
Pseudomonas sp. Strain Chol1, a Model Organism for the Degradation of Bile Salts and Other 417
Steroid Compounds. Genome Announc 1(1):e00014-12. 418
13. Holert J, Jagmann N, Philipp B. 2013. The essential function of genes for a hydratase and an 419
aldehyde dehydrogenase for growth of Pseudomonas sp. strain Chol1 with the steroid 420
compound cholate indicates an aldolytic reaction step for deacetylation of the side chain. J 421
Bacteriol 195:3371-80. 422
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

14. Barrientos Á, Merino E, Casabon I, Rodríguez J, Crowe AM, Holert J, Philipp B, Eltis LD, Olivera 423
ER, Luengo JM. 2015. Functional analyses of three acyl-CoA synthetases involved in bile acid 424
degradation in Pseudomonas putida DOC21. Environ Microbiol 17:47-63. 425
15. Simard J, Ricketts ML, Gingras S, Soucy P, Feltus FA, Melner MH. 2005. Molecular biology of the 426
3beta-hydroxysteroid dehydrogenase/delta5-delta4 isomerase gene family. Endocr Rev 26:525-427
82. 428
16. Vrielink A, Ghisla S. 2009. Cholesterol oxidase: biochemistry and structural features. FEBS J 429
276:6826-43. 430
17. Chiang YR, Ismail W, Heintz D, Schaeffer C, Van Dorsselaer A, Fuchs G. 2008. Study of anoxic and 431
oxic cholesterol metabolism by Sterolibacterium denitrificans. J Bacteriol 190:905-14. 432
18. Rosłoniec KZ, Wilbrink MH, Capyk JK, Mohn WW, Ostendorf M, van der Geize R, Dijkhuizen L, 433
Eltis LD. 2009. Cytochrome P450 125 (CYP125) catalyses C26-hydroxylation to initiate sterol 434
side-chain degradation in Rhodococcus jostii RHA1. Mol Microbiol 74:1031-43. 435
19. Driscoll MD, McLean KJ, Levy C, Mast N, Pikuleva IA, Lafite P, Rigby SE, Leys D, Munro AW. 2010. 436
Structural and biochemical characterization of Mycobacterium tuberculosis CYP142: evidence 437
for multiple cholesterol 27-hydroxylase activities in a human pathogen. J Biol Chem 285:38270-438
82. 439
20. Dermer J, Fuchs G. 2012. Molybdoenzyme that catalyzes the anaerobic hydroxylation of a 440
tertiary carbon atom in the side chain of cholesterol. J Biol Chem 287:36905-16. 441
21. Wang PH, Lee TH, Ismail W, Tsai CY, Lin CW, Tsai YW, Chiang YR. 2013. An oxygenase-442
independent cholesterol catabolic pathway operates under oxic conditions. PLoS One 8:e66675. 443
22. Warnke M, Jacoby C, Jung T, Agne M, Mergelsberg M, Starke R, Jehmlich N, von Bergen M, 444
Richnow HH, Brüls T, Boll M. 2017. A patchwork pathway for oxygenase-independent 445
degradation of side chain containing steroids. Environ Microbiol 19: 4668-4699. 446
23. Casabon I, Swain K, Crowe AM, Eltis LD, Mohn WW. 2014. Actinobacterial acyl coenzyme A 447
synthetases involved in steroid side-chain catabolism. J Bacteriol 196:579-87. 448
24. Gilbert S, Hood L, Seah SYK. 2017. Characterization of an aldolase involved in cholesterol side 449
chain degradation in Mycobacterium tuberculosis. J Bacteriol. doi:10.1128/JB.00512-17. 450
25. Wang PH, Leu YL, Ismail W, Tang SL, Tsai CY, Chen HJ. 2013. Anaerobic and aerobic cleavage of 451
the steroid core ring structure by Steroidobacter denitrificans. J Lipid Res 54: 452
1493-504. 453
26. Crowe AM, Casabon I, Brown KL, Liu J, Lian J, Rogalski JC, Hurst TE, Snieckus V, Foster LJ, Eltis LD. 454
2017. Catabolism of the last two steroid rings in Mycobacterium tuberculosis and other bacteria. 455
MBio 8(2): e00321-17. 456
27. Lin CW, Wang PH, Ismail W, Tsai YW, El Nayal A, Yang CY, Yang FC, Wang CH, Chiang YR. 2015. 457
Substrate uptake and subcellular compartmentation of anoxic cholesterol catabolism in 458
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

Sterolibacterium denitrificans. J Biol Chem 290:1155-69. 459
28. Casabon I, Crowe AM, Liu J, Eltis LD. 2013. FadD3 is an acyl-CoA synthetase that initiates 460
catabolism of cholesterol rings C and D in actinobacteria. Mol Microbiol 87:269-83. 461
29. Strohm TO, Griffin B, Zumft WG, Schink B. 2007. Growth yields in bacterial denitrification and 462
nitrate ammonification. Appl Environ Microbiol 73:1420-4. 463
30. Simon H, White H, Lebertz H, Thanos I. 1987. Reduktion von 2-Enoaten und Alkanoaten mit 464
Kohlenmonoxid oder Formiat, Viologenen und Clostridium thermoaceticum zu gesättigten 465
Säuren und ungesättigten bzw. gesättigten Alkoholen. Angewandte Chemie 99:785-787. 466
31. Thauer RK, Jungermann K, Decker K. 1977. Energy conservation in chemotrophic anaerobic 467
bacteria. Bacteriol Rev 41:100-80. 468
32. Tarlera S, Denner EB. 2003. Sterolibacterium denitrificans gen. nov., sp. nov., a novel 469
cholesterol-oxidizing, denitrifying member of the beta-Proteobacteria. Int J Syst Evol Microbiol 470
53:1085-91. 471
33. Bradford MM. 1976. A rapid and sensitive method for the quantitation of microgram quantities 472
of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248-54. 473
34. Holert J, Yücel O, Jagmann N, Prestel A, Möller HM, Philipp B. 2016. Identification of bypass 474
reactions leading to the formation of one central steroid degradation intermediate in 475
metabolism of different bile salts in Pseudomonas sp. strain Chol1. Environ Microbiol 18: 3373–476
89. 477
35. Yücel O, Drees S, Jagmann N, Patschkowski T, Philipp B. 2016. An unexplored pathway for 478
degradation of cholate requires a 7α-hydroxysteroid dehydratase and contributes to a broad 479
metabolic repertoire for the utilization of bile salts in Novosphingobium sp. strain Chol11. 480
Environ Microbiol 18: 5187–203. 481
36. Gan-Schreier H, Okun JG, Kohlmueller D, Langhans CD, Peters V, Ten Brink HJ, Verhoeven NM, 482
Jakobs C, Voelkl A, Hoffmann GF. 2005. Measurement of bile acid CoA esters by high-483
performance liquid chromatography-electrospray ionisation tandem mass spectrometry (HPLC-484
ESI-MS/MS). J Mass Spectrom 40:882-9. 485
37. Dereeper A, Guignon V, Blanc G, Audic S, Buffet S, Chevenet F, Dufayard JF, Guindon S, Lefort V, 486
Lescot M, Claverie JM, Gascuel O. 2008. Phylogeny.fr: robust phylogenetic analysis for the non-487
specialist. Nucleic Acids Res 36:W465-9. 488
38. Kumar S, Stecher G, Tamura K. 2016. MEGA7: Molecular Evolutionary Genetics Analysis Version 489
7.0 for Bigger Datasets. Mol Biol Evol 33:1870-4. 490
491
492
493
494
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from
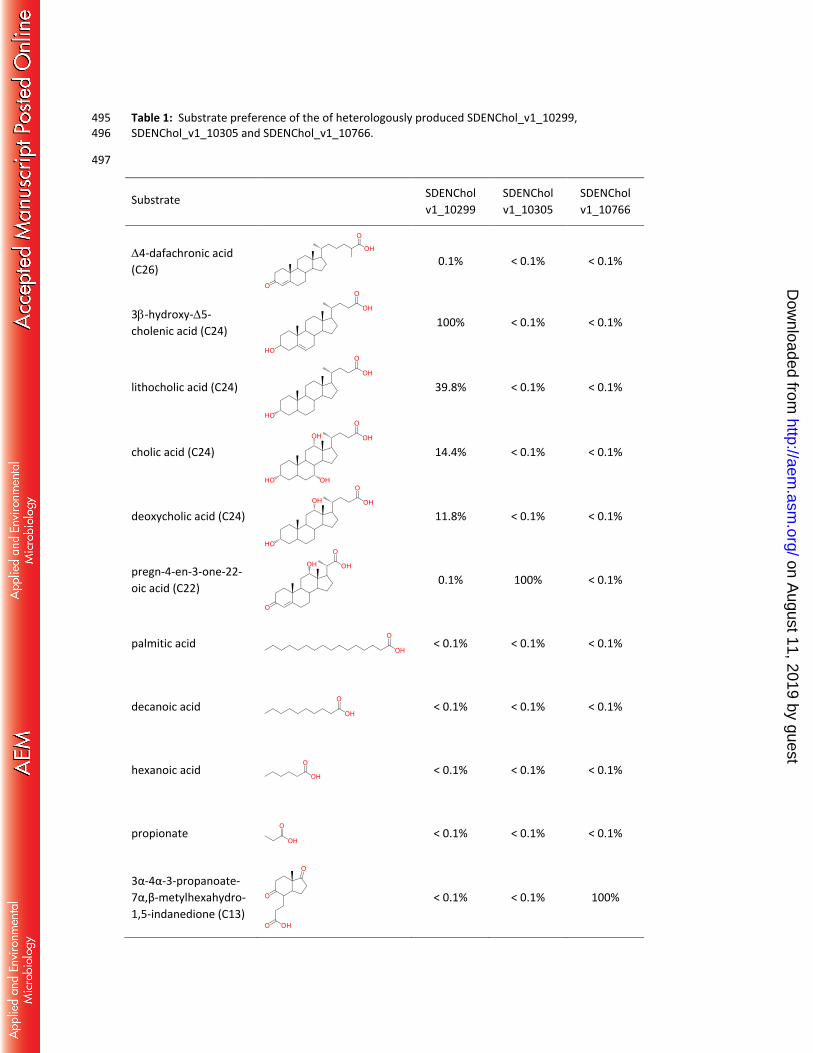
Table 1: Substrate preference of the of heterologously produced SDENChol_v1_10299, 495 SDENChol_v1_10305 and SDENChol_v1_10766. 496
497
Substrate SDENChol
v1_10299
SDENChol
v1_10305
SDENChol
v1_10766
4-dafachronic acid
(C26)
0.1% < 0.1% < 0.1%
3-hydroxy-5-
cholenic acid (C24)
100% < 0.1% < 0.1%
lithocholic acid (C24)
39.8% < 0.1% < 0.1%
cholic acid (C24)
14.4% < 0.1% < 0.1%
deoxycholic acid (C24)
11.8% < 0.1% < 0.1%
pregn-4-en-3-one-22-
oic acid (C22)
0.1% 100% < 0.1%
palmitic acid
< 0.1% < 0.1% < 0.1%
decanoic acid
< 0.1% < 0.1% < 0.1%
hexanoic acid < 0.1% < 0.1% < 0.1%
propionate
< 0.1% < 0.1% < 0.1%
3α-4α-3-propanoate-
7α,β-metylhexahydro-
1,5-indanedione (C13)
< 0.1% < 0.1% 100%
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from
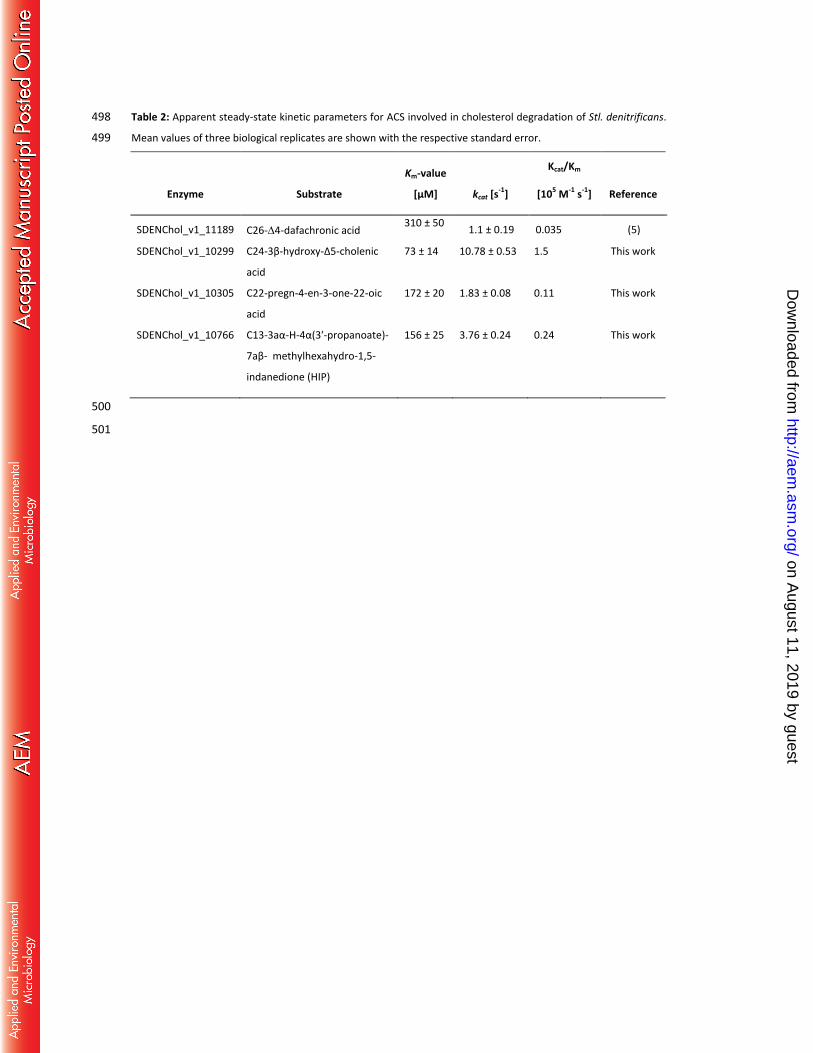
Table 2: Apparent steady-state kinetic parameters for ACS involved in cholesterol degradation of Stl. denitrificans. 498
Mean values of three biological replicates are shown with the respective standard error. 499
Enzyme Substrate
Km-value
[µM] kcat [s-1]
Kcat/Km
[105 M-1 s-1] Reference
SDENChol_v1_11189 C26-4-dafachronic acid 310 ± 50
1.1 ± 0.19 0.035 (5)
SDENChol_v1_10299 C24-3β-hydroxy-Δ5-cholenic
acid
73 ± 14 10.78 ± 0.53 1.5 This work
SDENChol_v1_10305 C22-pregn-4-en-3-one-22-oic
acid
172 ± 20 1.83 ± 0.08 0.11 This work
SDENChol_v1_10766 C13-3aα-H-4α(3′-propanoate)-
7aβ- methylhexahydro-1,5-
indanedione (HIP)
156 ± 25 3.76 ± 0.24 0.24 This work
500
501
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

Table 3: Kinetic parameters of ACS activities in extracts from cells grown with cholesterol and propionate. The 502
mean values of three biological replicates with respective standard deviations are shown. 503
504
ACS substrates
Cholesterol
Vmax [nmol min-1 mg-1]
Propionate
Vmax [nmol min-1 mg-1]
C24-3β-hydroxy-Δ5-cholenic acid 244 ± 11.2 31 ± 3.8
C22-Pregn-4-en-3-one-22-oic acid 98 ± 6.5 11 ± 2.1
C13-3aα-H-4α(3′-propanoate)-7aβ-
methylhexahydro-1,5-indanedione (HIP) 112 ± 6.9 12 ± 1.4
505
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from

Table 4. Primers used for heterologous expression of ACS genes. 506
507
508
509
510
511
512
513
514
515
516
517
518
519
520
521
522
523
Figure legends 524
525
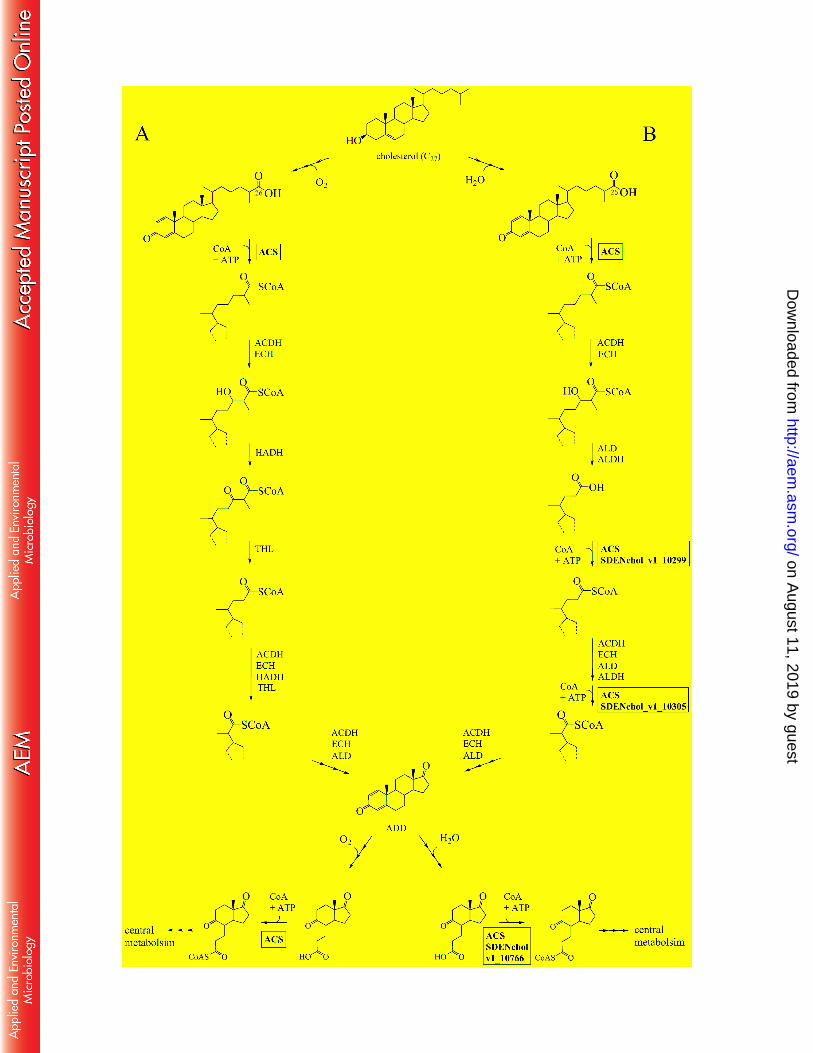
Figure 1. Aerobic (A) and anaerobic (B) cholesterol degradation pathway. The involvement of ATP-526
dependent acyl-CoA synthetases (ACS) is indicated; in the anaerobic pathway only the first ACS acting on 527
C26-oic acid has been isolated and characterized at the beginning of this work. ACDH, acyl-CoA 528
dehydrogenase; ECH, enoyl-CoA hydratase; HADH, 3-hydroxyacyl-CoA dehydrogenase; THL, thioloase; 529
ALD, aldolase; ALDH, aldehyde dehydrogenase. 530
531
Figure 2. Phylogenetic analysis of ACS from Stl. denitrificans (bold) and other steroid-degrading 532
organisms. The clustering of ACS specific for individual substrates is shown with the four experimentally 533
verified ACS being highlighted in red: C26ACS, 4-dafachronic acid; C24ACS, 3-hydroxy-5-cholenic 534
acid; C22ACS, pregn-4-en-3-one-22-oic acid; HIPACS, HIP. The accession numbers are given. The scale 535
bar represents the number of substitutions per site. 536
Primer Sequence (5´-3´) Ta (°C) Restriction site
SDENv1_10299 forw (C24-ACS)
GGAGGCGGTACCACGGAAGCGCTGAAAC
56 KpnI
SDENv1_10299 rev (C24-ACS)
GGTGGAAAGCTTCTCAGTCGCGTTCAAAATC
55 HindIII
SDENv1_10305 forw (C22-ACS)
GGAGGCGGTACCGCAATTCCCCGCGAAG
56 KpnI
SDENv1_10305 rev (C22-ACS)
GGTGGAAAGCTTATCGGTAGTTTAGACGCC
54 HindIII
SDENv1_10766 forw (HIP-ACS)
GGAGGCGAATTCTCGCCGTTGCCGCAAAC
60 EcoRI
SDENv1_10766 rev (HIP-ACS)
GCCTCCAAGCTTGGAGTACGAATCAGCGCAACTGATAC
61 HindIII
on August 11, 2019 by guest
http://aem.asm
.org/D
ownloaded from














![Ubiquitin Ein regulatorisches Wunderwerk Quellen · • E2= Ub-konjugierendes Enzym • E3= Ubiquitin-Protein-Ligase [4] (Berg, Stryer & Tymoczko, 2013) Letzter Schritt der katalytischen](https://static.fdocuments.in/doc/165x107/5e2091384123e612f72c7c31/ubiquitin-ein-regulatorisches-wunderwerk-quellen-a-e2-ub-konjugierendes-enzym.jpg)


