
From Simple to Complex Models: Gaits as...
3
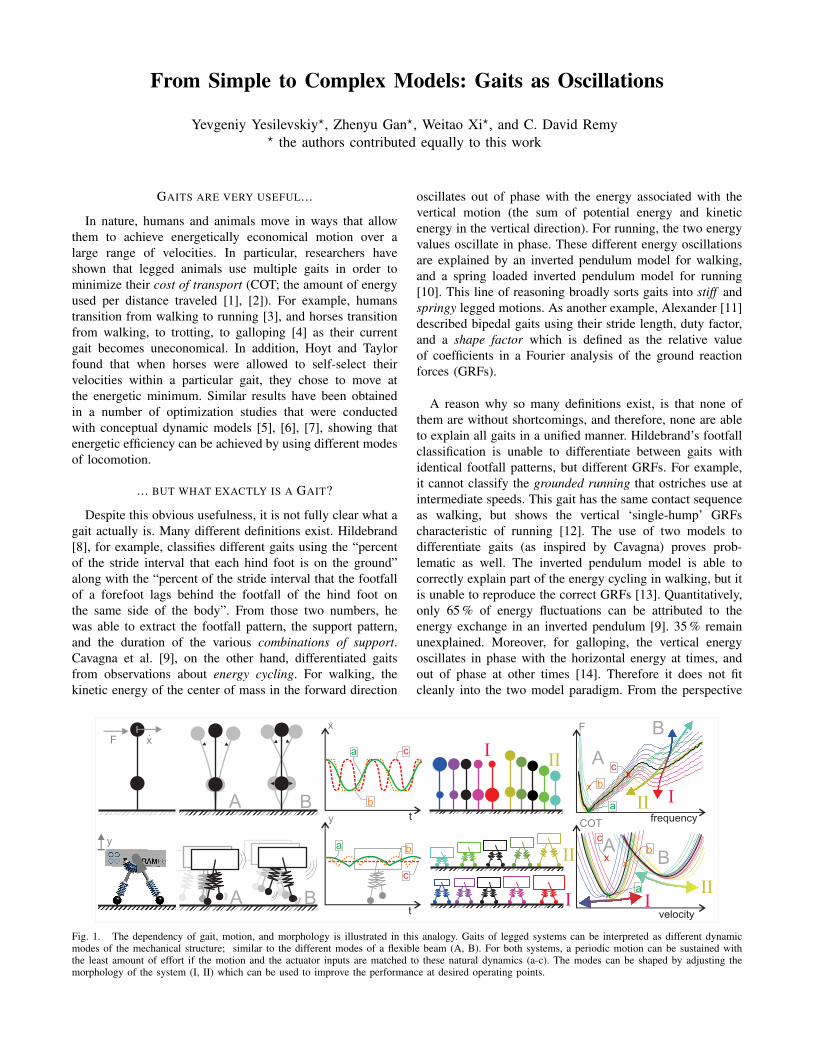
From Simple to Complex Models: Gaits as Oscillations Yevgeniy Yesilevskiy ? , Zhenyu Gan ? , Weitao Xi ? , and C. David Remy ? the authors contributed equally to this work GAITS ARE VERY USEFUL... In nature, humans and animals move in ways that allow them to achieve energetically economical motion over a large range of velocities. In particular, researchers have shown that legged animals use multiple gaits in order to minimize their cost of transport (COT; the amount of energy used per distance traveled [1], [2]). For example, humans transition from walking to running [3], and horses transition from walking, to trotting, to galloping [4] as their current gait becomes uneconomical. In addition, Hoyt and Taylor found that when horses were allowed to self-select their velocities within a particular gait, they chose to move at the energetic minimum. Similar results have been obtained in a number of optimization studies that were conducted with conceptual dynamic models [5], [6], [7], showing that energetic efficiency can be achieved by using different modes of locomotion. ... BUT WHAT EXACTLY IS A GAIT? Despite this obvious usefulness, it is not fully clear what a gait actually is. Many different definitions exist. Hildebrand [8], for example, classifies different gaits using the “percent of the stride interval that each hind foot is on the ground” along with the “percent of the stride interval that the footfall of a forefoot lags behind the footfall of the hind foot on the same side of the body”. From those two numbers, he was able to extract the footfall pattern, the support pattern, and the duration of the various combinations of support. Cavagna et al. [9], on the other hand, differentiated gaits from observations about energy cycling. For walking, the kinetic energy of the center of mass in the forward direction oscillates out of phase with the energy associated with the vertical motion (the sum of potential energy and kinetic energy in the vertical direction). For running, the two energy values oscillate in phase. These different energy oscillations are explained by an inverted pendulum model for walking, and a spring loaded inverted pendulum model for running [10]. This line of reasoning broadly sorts gaits into stiff and springy legged motions. As another example, Alexander [11] described bipedal gaits using their stride length, duty factor, and a shape factor which is defined as the relative value of coefficients in a Fourier analysis of the ground reaction forces (GRFs). A reason why so many definitions exist, is that none of them are without shortcomings, and therefore, none are able to explain all gaits in a unified manner. Hildebrand’s footfall classification is unable to differentiate between gaits with identical footfall patterns, but different GRFs. For example, it cannot classify the grounded running that ostriches use at intermediate speeds. This gait has the same contact sequence as walking, but shows the vertical ‘single-hump’ GRFs characteristic of running [12]. The use of two models to differentiate gaits (as inspired by Cavagna) proves prob- lematic as well. The inverted pendulum model is able to correctly explain part of the energy cycling in walking, but it is unable to reproduce the correct GRFs [13]. Quantitatively, only 65 % of energy fluctuations can be attributed to the energy exchange in an inverted pendulum [9]. 35 % remain unexplained. Moreover, for galloping, the vertical energy oscillates in phase with the horizontal energy at times, and out of phase at other times [14]. Therefore it does not fit cleanly into the two model paradigm. From the perspective t y c a b t x . c a b B A B A y F x . II I II I frequency F B A x x x c a b II I velocity COT II I B A x c x a x b Fig. 1. The dependency of gait, motion, and morphology is illustrated in this analogy. Gaits of legged systems can be interpreted as different dynamic modes of the mechanical structure; similar to the different modes of a flexible beam (A, B). For both systems, a periodic motion can be sustained with the least amount of effort if the motion and the actuator inputs are matched to these natural dynamics (a-c). The modes can be shaped by adjusting the morphology of the system (I, II) which can be used to improve the performance at desired operating points.
-
Upload
nguyenngoc -
Category
Documents
-
view
216 -
download
1
Transcript of From Simple to Complex Models: Gaits as...
From Simple to Complex Models: Gaits as Oscillations
Yevgeniy Yesilevskiy?, Zhenyu Gan?, Weitao Xi?, and C. David Remy? the authors contributed equally to this work
GAITS ARE VERY USEFUL...
In nature, humans and animals move in ways that allowthem to achieve energetically economical motion over alarge range of velocities. In particular, researchers haveshown that legged animals use multiple gaits in order tominimize their cost of transport (COT; the amount of energyused per distance traveled [1], [2]). For example, humanstransition from walking to running [3], and horses transitionfrom walking, to trotting, to galloping [4] as their currentgait becomes uneconomical. In addition, Hoyt and Taylorfound that when horses were allowed to self-select theirvelocities within a particular gait, they chose to move atthe energetic minimum. Similar results have been obtainedin a number of optimization studies that were conductedwith conceptual dynamic models [5], [6], [7], showing thatenergetic efficiency can be achieved by using different modesof locomotion.
... BUT WHAT EXACTLY IS A GAIT?
Despite this obvious usefulness, it is not fully clear what agait actually is. Many different definitions exist. Hildebrand[8], for example, classifies different gaits using the “percentof the stride interval that each hind foot is on the ground”along with the “percent of the stride interval that the footfallof a forefoot lags behind the footfall of the hind foot onthe same side of the body”. From those two numbers, hewas able to extract the footfall pattern, the support pattern,and the duration of the various combinations of support.Cavagna et al. [9], on the other hand, differentiated gaitsfrom observations about energy cycling. For walking, thekinetic energy of the center of mass in the forward direction
oscillates out of phase with the energy associated with thevertical motion (the sum of potential energy and kineticenergy in the vertical direction). For running, the two energyvalues oscillate in phase. These different energy oscillationsare explained by an inverted pendulum model for walking,and a spring loaded inverted pendulum model for running[10]. This line of reasoning broadly sorts gaits into stiff andspringy legged motions. As another example, Alexander [11]described bipedal gaits using their stride length, duty factor,and a shape factor which is defined as the relative valueof coefficients in a Fourier analysis of the ground reactionforces (GRFs).
A reason why so many definitions exist, is that none ofthem are without shortcomings, and therefore, none are ableto explain all gaits in a unified manner. Hildebrand’s footfallclassification is unable to differentiate between gaits withidentical footfall patterns, but different GRFs. For example,it cannot classify the grounded running that ostriches use atintermediate speeds. This gait has the same contact sequenceas walking, but shows the vertical ‘single-hump’ GRFscharacteristic of running [12]. The use of two models todifferentiate gaits (as inspired by Cavagna) proves prob-lematic as well. The inverted pendulum model is able tocorrectly explain part of the energy cycling in walking, but itis unable to reproduce the correct GRFs [13]. Quantitatively,only 65 % of energy fluctuations can be attributed to theenergy exchange in an inverted pendulum [9]. 35 % remainunexplained. Moreover, for galloping, the vertical energyoscillates in phase with the horizontal energy at times, andout of phase at other times [14]. Therefore it does not fitcleanly into the two model paradigm. From the perspective
t
y
c
a b
t
x.
ca
b
BA
BA
y
F x.
III
II
I
frequency
F B
A
x
xx
c
a
b
II I
velocity
COT
III
BAx
c
xa
x
b
Fig. 1. The dependency of gait, motion, and morphology is illustrated in this analogy. Gaits of legged systems can be interpreted as different dynamicmodes of the mechanical structure; similar to the different modes of a flexible beam (A, B). For both systems, a periodic motion can be sustained withthe least amount of effort if the motion and the actuator inputs are matched to these natural dynamics (a-c). The modes can be shaped by adjusting themorphology of the system (I, II) which can be used to improve the performance at desired operating points.

of a roboticist, it is probably most problematic that all ofthese definitions are purely descriptive. While they are ableto classify different characteristics of locomotion, they giveno explanation of where these gaits originate. Alexander’sshape factor is probably the best example for this. HisFourier coefficients can be used in an optimization problemto yield ‘natural looking’ GRFs [11], yet they do not giveany insight into why these different force profiles enableefficient locomotion at different speeds. These definitions arenot useful from a synthetic point of view. For a roboticist,this synthetic perspective still poses an open question: Howcan we build the ability to perform multiple efficient gaitsinto legged robotic systems?
A NEW PERSPECTIVE ON GAIT
Recent work in passive dynamic walking suggests a newinterpretation of gaits. Geyer et al. [15] were able to showthat a bipedal model with two independent, massless legswith springs was able to accurately reproduce the energycycling and GRFs in both walking and running. This dis-covery of multiple gaits within a single model suggests thatgaits are different complex non-linear oscillation modes of asingle oscillatory system. This concept can be understood byits analogy to the different modes of oscillation of a flexiblebeam (Fig. 1, left).
Using similar model approximations as Geyer et al., westudied passive dynamic walking of quadrupeds (Fig. 2a).Our simple model was able to move in a variety of differentmodes while using the same set of model parameters [16].In particular, we discovered passive motions that had thesame footfall patterns as walking, trotting, tolting, pacing,bounding, and galloping of horses. These gaits are merelydifferent dynamic modes of the same mechanical system. Inaddition to replicating a large number of footfall sequences,our models also accurately predicted vertical GRFs of horsesat walk, trot, and tolt (Fig. 3a,b). Analyzing the detailedmotion of these different locomotion modes, we clearlyobserved fundamentally different oscillatory patterns. Walk-ing, for example, had phases of double support alternatingwith phases of triple support. The vertical GRFs exhibited adouble hump with a mid-stance relief. These two force peakscoincided with a double compression of the leg springs.Also, in walking, the main body showed a clear pitchingmotion. The main body oscillated between -1.45 ◦ and 1.23 ◦
during a gait cycle. Tolting has the same footfall sequence aswalking. Yet, in this gait phases of double support alternatewith phases of single support and there are fundamentallydifferent oscillatory patterns. For tolting, the vertical GRFsexhibited only a single hump. This single peak was roughlytwice the magnitude of the peaks in walking. A pitchingmotion is still present, but less pronounced than in walk-ing (the main body oscillates between -0.47 ◦ and 0.48 ◦).Overall, tolting exhibited the least variability in main bodyvertical and horizontal velocity throughout the course of astride. Trotting also showed a single hump in the verticalGRFs - corresponding to a single spring compression. Yet,since trotting is a two-beat gait in which phases of double-
support alternate with an air phase, no noticeable body pitchwas observed. Consequently, the main body exhibited a muchlarger vertical motion than during walking or tolting.
RAMlabm2, j2
m , j3 3
y
x
ταRF
τ lRH
l3
l1
m , j1 1
lR H
u lL HαL H
uαL HuαL H
(b)
y
x αF
l0
αH
kF kH
(a)
la
d lC O M a
m , jo o
k
Fig. 2. Our work explored two different models of legged systems. Passivemodels with massless legs and no damping in the springs (a) and actuatedmodels with distributed mass, series elastic actuators in the legs and hips,and damping in the springs (b). Both models exhibited a number of similaroscillation modes that yielded a variety of gaits.
While these results provide significant evidence towardsinterpreting gaits as different oscillatory modes of a singlesystem, the simplicity of the underlying model limits possibleconclusions with respect to actual robotic systems. The miss-ing actuation, in particular, conceals issues of efficiency andother energetic benefits when exploiting different oscillationsmodes. Such benefits would be expected, given that excitingan oscillatory system in a natural mode would minimize theamount of required excitation power.
To investigate this hypothesis, we additionally conductedoptimal control studies with more complex models of leggedrobots (Fig. 2b). These models have fully distributed masses,which leads to complex swing leg dynamics and collisionlosses at touchdown. Additional losses are created by viscousfriction in the springy legs. The models are not energeticallyconservative and are driven by detailed series elastic actu-ators. In order to not bias the model towards establishedgaits and to reduce the influence of optimization errors, weconducted two studies using different optimization methods.We used direct collocation in an optimization frameworkthat could detect optimal footfall patterns automatically [17],and we used an established multiple shooting framework(MUSCOD [18]) for which we pre-selected a wide range ofdifferent footfall sequences. Both approaches yielded similarresults.
Despite the addition of actuation, collision losses, swing
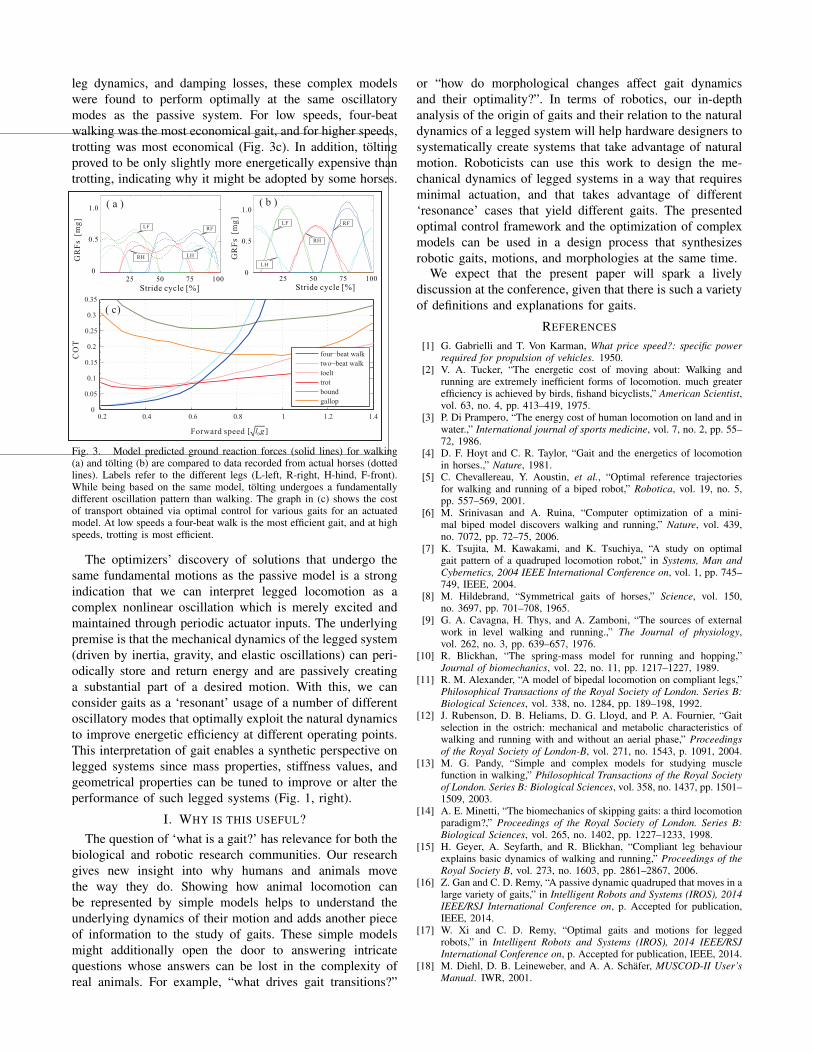
leg dynamics, and damping losses, these complex modelswere found to perform optimally at the same oscillatorymodes as the passive system. For low speeds, four-beatwalking was the most economical gait, and for higher speeds,trotting was most economical (Fig. 3c). In addition, toltingproved to be only slightly more energetically expensive thantrotting, indicating why it might be adopted by some horses.
Fig. 3. Model predicted ground reaction forces (solid lines) for walking(a) and tolting (b) are compared to data recorded from actual horses (dottedlines). Labels refer to the different legs (L-left, R-right, H-hind, F-front).While being based on the same model, tolting undergoes a fundamentallydifferent oscillation pattern than walking. The graph in (c) shows the costof transport obtained via optimal control for various gaits for an actuatedmodel. At low speeds a four-beat walk is the most efficient gait, and at highspeeds, trotting is most efficient.
The optimizers’ discovery of solutions that undergo thesame fundamental motions as the passive model is a strongindication that we can interpret legged locomotion as acomplex nonlinear oscillation which is merely excited andmaintained through periodic actuator inputs. The underlyingpremise is that the mechanical dynamics of the legged system(driven by inertia, gravity, and elastic oscillations) can peri-odically store and return energy and are passively creatinga substantial part of a desired motion. With this, we canconsider gaits as a ‘resonant’ usage of a number of differentoscillatory modes that optimally exploit the natural dynamicsto improve energetic efficiency at different operating points.This interpretation of gait enables a synthetic perspective onlegged systems since mass properties, stiffness values, andgeometrical properties can be tuned to improve or alter theperformance of such legged systems (Fig. 1, right).
I. WHY IS THIS USEFUL?The question of ‘what is a gait?’ has relevance for both the
biological and robotic research communities. Our researchgives new insight into why humans and animals movethe way they do. Showing how animal locomotion canbe represented by simple models helps to understand theunderlying dynamics of their motion and adds another pieceof information to the study of gaits. These simple modelsmight additionally open the door to answering intricatequestions whose answers can be lost in the complexity ofreal animals. For example, “what drives gait transitions?”
or “how do morphological changes affect gait dynamicsand their optimality?”. In terms of robotics, our in-depthanalysis of the origin of gaits and their relation to the naturaldynamics of a legged system will help hardware designers tosystematically create systems that take advantage of naturalmotion. Roboticists can use this work to design the me-chanical dynamics of legged systems in a way that requiresminimal actuation, and that takes advantage of different‘resonance’ cases that yield different gaits. The presentedoptimal control framework and the optimization of complexmodels can be used in a design process that synthesizesrobotic gaits, motions, and morphologies at the same time.
We expect that the present paper will spark a livelydiscussion at the conference, given that there is such a varietyof definitions and explanations for gaits.
REFERENCES
[1] G. Gabrielli and T. Von Karman, What price speed?: specific powerrequired for propulsion of vehicles. 1950.
[2] V. A. Tucker, “The energetic cost of moving about: Walking andrunning are extremely inefficient forms of locomotion. much greaterefficiency is achieved by birds, fishand bicyclists,” American Scientist,vol. 63, no. 4, pp. 413–419, 1975.
[3] P. Di Prampero, “The energy cost of human locomotion on land and inwater.,” International journal of sports medicine, vol. 7, no. 2, pp. 55–72, 1986.
[4] D. F. Hoyt and C. R. Taylor, “Gait and the energetics of locomotionin horses.,” Nature, 1981.
[5] C. Chevallereau, Y. Aoustin, et al., “Optimal reference trajectoriesfor walking and running of a biped robot,” Robotica, vol. 19, no. 5,pp. 557–569, 2001.
[6] M. Srinivasan and A. Ruina, “Computer optimization of a mini-mal biped model discovers walking and running,” Nature, vol. 439,no. 7072, pp. 72–75, 2006.
[7] K. Tsujita, M. Kawakami, and K. Tsuchiya, “A study on optimalgait pattern of a quadruped locomotion robot,” in Systems, Man andCybernetics, 2004 IEEE International Conference on, vol. 1, pp. 745–749, IEEE, 2004.
[8] M. Hildebrand, “Symmetrical gaits of horses,” Science, vol. 150,no. 3697, pp. 701–708, 1965.
[9] G. A. Cavagna, H. Thys, and A. Zamboni, “The sources of externalwork in level walking and running.,” The Journal of physiology,vol. 262, no. 3, pp. 639–657, 1976.
[10] R. Blickhan, “The spring-mass model for running and hopping,”Journal of biomechanics, vol. 22, no. 11, pp. 1217–1227, 1989.
[11] R. M. Alexander, “A model of bipedal locomotion on compliant legs,”Philosophical Transactions of the Royal Society of London. Series B:Biological Sciences, vol. 338, no. 1284, pp. 189–198, 1992.
[12] J. Rubenson, D. B. Heliams, D. G. Lloyd, and P. A. Fournier, “Gaitselection in the ostrich: mechanical and metabolic characteristics ofwalking and running with and without an aerial phase,” Proceedingsof the Royal Society of London-B, vol. 271, no. 1543, p. 1091, 2004.
[13] M. G. Pandy, “Simple and complex models for studying musclefunction in walking,” Philosophical Transactions of the Royal Societyof London. Series B: Biological Sciences, vol. 358, no. 1437, pp. 1501–1509, 2003.
[14] A. E. Minetti, “The biomechanics of skipping gaits: a third locomotionparadigm?,” Proceedings of the Royal Society of London. Series B:Biological Sciences, vol. 265, no. 1402, pp. 1227–1233, 1998.
[15] H. Geyer, A. Seyfarth, and R. Blickhan, “Compliant leg behaviourexplains basic dynamics of walking and running,” Proceedings of theRoyal Society B, vol. 273, no. 1603, pp. 2861–2867, 2006.
[16] Z. Gan and C. D. Remy, “A passive dynamic quadruped that moves in alarge variety of gaits,” in Intelligent Robots and Systems (IROS), 2014IEEE/RSJ International Conference on, p. Accepted for publication,IEEE, 2014.
[17] W. Xi and C. D. Remy, “Optimal gaits and motions for leggedrobots,” in Intelligent Robots and Systems (IROS), 2014 IEEE/RSJInternational Conference on, p. Accepted for publication, IEEE, 2014.
[18] M. Diehl, D. B. Leineweber, and A. A. Schafer, MUSCOD-II User’sManual. IWR, 2001.


















