
FoxO regulates expression of decidual protein induced by progesterone (DEPP) in human endothelial...
5
FoxO regulates expression of decidual protein induced by progesterone (DEPP) in human endothelial cells Simin Chen, Junfang Gai, Yeyu Wang, Hao Li ⇑ Key Laboratory of Cardiovascular Disease and Molecular Intervention, Department of Pathophysiology, Nanjing Medical University, Nanjing 210029, China article info Article history: Received 12 December 2010 Revised 30 March 2011 Accepted 11 April 2011 Available online 15 April 2011 Edited by Robert Barouki Keywords: DEPP Forkhead box protein Endothelial cell Transcription factor abstract DEPP was initially cloned from the human endometrial stromal cell cDNA library, but the transcrip- tional regulation of DEPP remains largely unknown. We demonstrate here that expression of DEPP is FoxO-dependent in human endothelial cells. Two functional FoxO-responsive elements are iden- tified in the DEPP promoter. Hypoxia stimulates DEPP expression in the endothelial cell line EA.hy926. Hypoxia-induced upregulation of DEPP is dependent on FoxO expression. We conclude that DEPP is regulated at the level of transcription by FoxO in human vascular endothelial cells. Ó 2011 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. 1. Introduction Decidual protein induced by progesterone (DEPP) was initially cloned from the human endometrial stromal cells cDNA library enriched for progesterone-induced genes [1]. Fasting Induced Gene (FIG) was isolated independently from a human adipose tissue library. When the sequences of both genes are available to the pub- lic, it is evident that these two genes are the same (UniGene Cluster Hs.93675 as Chromosome 10 open reading frame 10 (C10orf10)). Human DEPP was highly expressed in various tissues including placenta, ovary, kidney, white adipose and liver [1–3]. DEPP was also detected in endothelial cells of peripheral tissues [3]. Although the tissue distribution of DEPP is well known, its transcriptional regulation remains to be elucidated. It was reported that DEPP was regulated by FoxO1 by binding to IRS element in its promoter in a renal adenocarcinoma cell line, but the specific binding site remains unknown [4]. Increased expression of DEPP was observed in several conditions. Progesterone or androgen induces DEPP mRNA in endometrial stromal cells [1]. DEPP was induced in a malignant glioma cell line in response to hypoxic stress [5]. Recently, DEPP was reported to be upregulated by feeding in insu- lin-sensitive tissues including white adipose tissue and liver [2]. However, the physiological significance of DEPP remains largely unknown. Forkhead box O (FoxO) transcription factor family consists of FoxO1, FoxO3a, and FoxO4 (also known as FKHR, FKHRL1, and AFX, respectively). FoxOs regulate hormonal, nutrient, and stress responses [6]. Activation of the PI3K pathway blocks the function of FoxO factors by Akt-dependent phosphorylation of three con- served residues, which leads to inhibition of DNA binding, nuclear exclusion, and subsequent sequestration in the cytoplasm. Dephos- phorylation of FoxOs in turn stimulates nuclear entry, leading to the activation or repression of target genes [7,8]. In a microarray analysis of expression profile in human endo- thelial cell line transfected with FoxO plasmids, we found that DEPP mRNA increased with FoxO overexpression. The present study was designed to investigate molecular mechanisms by which FoxO regulates DEPP transcription in vascular endothelial cells. 2. Materials and methods Materials and methods are shown as Supplementary material. 3. Results 3.1. Expression of DEPP is FoxO-dependent in human endothelial cells EA.hy926 cells or HUVECs were transfected with FoxO expres- sion plasmids and DEPP mRNA was quantified using real-time 0014-5793/$36.00 Ó 2011 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.febslet.2011.04.024 Abbreviations: HUVEC, human umbilical vein endothelial cells; EMSA, electro- phoretic mobility shift assay; DMOG, dimethyloxalyl glycine; ChIP, chromatin immunoprecipitation ⇑ Corresponding author. Fax: +86 25 86862888. E-mail address: [email protected] (H. Li). FEBS Letters 585 (2011) 1796–1800 journal homepage: www.FEBSLetters.org
-
Upload
simin-chen -
Category
Documents
-
view
214 -
download
1
Transcript of FoxO regulates expression of decidual protein induced by progesterone (DEPP) in human endothelial...

FEBS Letters 585 (2011) 1796–1800
journal homepage: www.FEBSLetters .org
FoxO regulates expression of decidual protein induced by progesterone (DEPP)in human endothelial cells
Simin Chen, Junfang Gai, Yeyu Wang, Hao Li ⇑Key Laboratory of Cardiovascular Disease and Molecular Intervention, Department of Pathophysiology, Nanjing Medical University, Nanjing 210029, China
a r t i c l e i n f o a b s t r a c t
Article history:Received 12 December 2010Revised 30 March 2011Accepted 11 April 2011Available online 15 April 2011
Edited by Robert Barouki
Keywords:DEPPForkhead box proteinEndothelial cellTranscription factor
0014-5793/$36.00 � 2011 Federation of European Biodoi:10.1016/j.febslet.2011.04.024
Abbreviations: HUVEC, human umbilical vein endphoretic mobility shift assay; DMOG, dimethyloxaimmunoprecipitation⇑ Corresponding author. Fax: +86 25 86862888.
E-mail address: [email protected] (H. Li).
DEPP was initially cloned from the human endometrial stromal cell cDNA library, but the transcrip-tional regulation of DEPP remains largely unknown. We demonstrate here that expression of DEPPis FoxO-dependent in human endothelial cells. Two functional FoxO-responsive elements are iden-tified in the DEPP promoter. Hypoxia stimulates DEPP expression in the endothelial cell lineEA.hy926. Hypoxia-induced upregulation of DEPP is dependent on FoxO expression. We concludethat DEPP is regulated at the level of transcription by FoxO in human vascular endothelial cells.� 2011 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
1. Introduction
Decidual protein induced by progesterone (DEPP) was initiallycloned from the human endometrial stromal cells cDNA libraryenriched for progesterone-induced genes [1]. Fasting Induced Gene(FIG) was isolated independently from a human adipose tissuelibrary. When the sequences of both genes are available to the pub-lic, it is evident that these two genes are the same (UniGene ClusterHs.93675 as Chromosome 10 open reading frame 10 (C10orf10)).Human DEPP was highly expressed in various tissues includingplacenta, ovary, kidney, white adipose and liver [1–3]. DEPP wasalso detected in endothelial cells of peripheral tissues [3]. Althoughthe tissue distribution of DEPP is well known, its transcriptionalregulation remains to be elucidated. It was reported that DEPPwas regulated by FoxO1 by binding to IRS element in its promoterin a renal adenocarcinoma cell line, but the specific binding siteremains unknown [4]. Increased expression of DEPP was observedin several conditions. Progesterone or androgen induces DEPPmRNA in endometrial stromal cells [1]. DEPP was induced in amalignant glioma cell line in response to hypoxic stress [5].Recently, DEPP was reported to be upregulated by feeding in insu-lin-sensitive tissues including white adipose tissue and liver [2].
chemical Societies. Published by E
othelial cells; EMSA, electro-lyl glycine; ChIP, chromatin
However, the physiological significance of DEPP remains largelyunknown.
Forkhead box O (FoxO) transcription factor family consists ofFoxO1, FoxO3a, and FoxO4 (also known as FKHR, FKHRL1, andAFX, respectively). FoxOs regulate hormonal, nutrient, and stressresponses [6]. Activation of the PI3K pathway blocks the functionof FoxO factors by Akt-dependent phosphorylation of three con-served residues, which leads to inhibition of DNA binding, nuclearexclusion, and subsequent sequestration in the cytoplasm. Dephos-phorylation of FoxOs in turn stimulates nuclear entry, leading tothe activation or repression of target genes [7,8].
In a microarray analysis of expression profile in human endo-thelial cell line transfected with FoxO plasmids, we found thatDEPP mRNA increased with FoxO overexpression. The presentstudy was designed to investigate molecular mechanisms by whichFoxO regulates DEPP transcription in vascular endothelial cells.
2. Materials and methods
Materials and methods are shown as Supplementary material.
3. Results
3.1. Expression of DEPP is FoxO-dependent in human endothelial cells
EA.hy926 cells or HUVECs were transfected with FoxO expres-sion plasmids and DEPP mRNA was quantified using real-time
lsevier B.V. All rights reserved.

S. Chen et al. / FEBS Letters 585 (2011) 1796–1800 1797
PCR. Transfection with constitutively active FoxO1TM or Fox-O3aTM, the two predominant FoxO isoforms in human endothelialcells [9,10] led to about 4-fold increase of DEPP mRNA in EA.hy926cells (Fig. 1A) and about 2-fold increase in HUVECs (Fig. 1B). Wenext depleted FoxO1 or FoxO3a with specific siRNA respectively.Depletion of either FoxO1 or FoxO3a was associated with a 40–50% reduction of DEPP mRNA in EA.hy926 (Fig. 1D), while deple-tion of both FoxOs resulted in a 70% reduction. Dramatic decreaseof DEPP expression was also observed in FoxO1- or FoxO3a-si-lenced HUVEC (Fig. 1E), consistent with a previous microarray re-port [9]. Thus, expression of DEPP is dependent on FoxO factorsin human endothelial cells.
3.2. FoxO activates DEPP promoter in human endothelial cells
We scanned the DEPP promoter for putative FoxO binding sitesusing rVista2.0 and found three conserved optimal Forkhead-bind-ing sites. The first located from �537 to �530 (S1), the second lo-cated from �179 to �172 (S2), and the third located from �151 to�143 (S3), relative to the start of cDNA (Fig. 2A). To test whetherFoxO-induced expression of DEPP is mediated by its putative bind-ing sites, the DEPP promoter was cloned upstream of luciferase inthe pGL3-Basic plasmid. Then, we co-transfected EA.hy926 cellswith this construct and either a control or a FoxO-expressing plas-mid and measured luciferase activity 24 h later. We observed dra-matic increase of DEPP promoter activity with FoxO co-expression,
Fig. 1. FoxO-dependent expression of DEPP in endothelial cells. (A, B) EA.hy926cells or HUVECs were transfected as indicated and DEPP mRNA was quantified usingreal-time PCR. Data are mean ± SD from three experiments. ⁄P < 0.05 versus control.(C) EA.hy926 cells were transfected as indicated and harvested for western blotafter 24 h. (D, E) DEPP mRNA was quantified after transfection with siRNA. Data aremean ± SD from three experiments. ⁄P < 0.05 versus control.
the highest of which was FoxO1TM, leading to a 6-fold increaseversus control (Fig. 2B, left panel). In HEK293 cells, the highest acti-vation of DEPP promoter was observed with FoxO3aTM overex-pression while the effect of FoxO1TM was mild (Fig. 2B, rightpanel). We further generated a series of deletion constructs. Dele-tion of the first binding site (DEPP-D2-luc) had no significant effect,whereas deletion of the last two binding sites (DEPP-D1-luc)caused a prominent fall of the responsiveness of the DEPP pro-moter to FoxO in HEK293 cells (Fig. 2C). Consistent with this,mutation of the first binding site (DEPP-mut1-luc) almost had noeffect on the luciferase activity in EA.hy926 cells, while mutationof the second (DEPP-mut2-luc) or third binding site (DEPP-mut3-luc) resulted in an obvious decrease of the luciferase activativity,and mutation of the last two binding sites (DEPP-mut2/3-luc) com-pletely abolished the activition of the DEPP promoter to FoxO(Fig. 2D). In HUVECs, mutation of either one of the last two bindingsites abolished DEPP promoter response to FoxO (Fig. S1). Thesefindings indicate that the two forkhead binding sequences locatedwithin 200 bp upstream of the start of cDNA are the major sitesresponsible for the effects of FoxO on the DEPP promoter.
3.3. FoxO binds its cognate sequences within the DEPP promoter
To determine whether FoxO binds to DEPP promoter at the sitesshown above, we first performed EMSA. We found that a 25 bpdouble-stranded oligonucleotide probe encompassing FoxO bind-ing site S2 in the DEPP promoter (underlined in Fig. 3A) exhibitedan electromobility shift (Fig. 3C) when incubated with a nuclearextract prepared from cells transfected with FoxO expression plas-mids. The binding was blocked by unlabeled competitor DNA. Pre-incubation with antibodies against FoxO caused a supershift. Whenthe putative binding site was mutated, the electromobility shiftwas almost abolished. Similar results were obtained with the thirdputative binding site S3 (Fig. 3B). These data demonstrate thatFoxO binds its cognate sequences within the DEPP promoter invitro.
To confirm that FoxO also binds to the DEPP promoter in vivo,we performed CHIP. First, we transfected HEK293 cells with aMyc-FoxO1TM expressing plasmid and performed PCR amplifica-tion of the 305 bp DNA fragment encompassing the double FoxObinding sites of DEPP promoter. As expected, in the presence ofantibody, the DNA fragment could be amplified by PCR (Fig. 4A)whereas the amplification in the absence of the Myc antibodywas much weaker. We also tested whether endogenous FoxO1binds to DEPP promoter in EA.hy926 cells and obtained similar re-sults (Fig. 4B). Binding of FoxO1 to the promoter region of DEPPwas specific, as no chromatin fragment in the coding region ofDEPP was immunoprecipitated in both experiments (data notshown). Taken together, these results confirm that FoxO binds tothe DEPP promoter.
3.4. Upregulation of DEPP by hypoxia is FoxO-dependent in endothelialcell line EA.hy926
Prominent upregulation of DEPP mRNA was detected by quan-titative PCR in EA.hy926 cells after 8 or 24 h culture under hypoxicconditions (Fig. 5A). Upregulation of DEPP was also detected withhypoxia mimetic DMOG treatment for 4 or 6 h (Fig. 5B). Increasedexpression of DEPP was confirmed by western blot (Fig. 5C). In con-trast, no alteration of DEPP mRNA was detected in HUVECs stimu-lated with DMOG (data not shown) and DEPP protein evendecreased by hypoxia (Fig. 5D). No detectable effect of hypoxiaon FoxO binding to the double FoxO binding sites of DEPP pro-moter was observed in HUVECs (Fig. S2). It was reported that Fox-O3a protein and transcripts were increased in hypoxia in an HIF1-dependent way in mouse embryonic fibroblasts [11] and in micro-

Fig. 2. Effect of FoxO on DEPP promoter activity in human endothelial cells. (A) A scan of DEPP gene promoter revealed three putative binding sites (bold in the sequence) forFoxO. (B) Cells were co-transfected with reporter plasmid containing the putative FoxO binding sites and FoxOTM plasmids as indicated. Luciferase activity in cell lysate wasanalyzed 24 h after transfection. Data are given as means ± SD of three independent experiments. ⁄P < 0.05 versus control. (C) 293 cells were transfected with indicateddeletion reporter plasmids. Data are given as means ± SD of three independent experiments. The results for DEPP-D1-luc were shown in insert. (D) EA.hy926 cells weretransfected with mutated reporter plasmids. Data are given as means ± SD of three independent experiments.
1798 S. Chen et al. / FEBS Letters 585 (2011) 1796–1800
vascular endothelial cell line glEND.2 [12]. In HUVEC and EA.hy926cells, no detectable increase of FoxO3a or its cellular distributionwas observed with hypoxia, though HIF1 target genes includingVEGF and NIX transcripts upregulated as expected (Fig. 6A and B,Fig. S3). Expression of FoxO1 was not significantly altered by hy-poxia either (data not shown). Despite of the unchanging amountof FoxOs, interference with either FoxO1 or FoxO3a using siRNAinhibited hypoxia- as well as DMOG-induced DEPP significantly,and knockdown of both FoxOs resulted in maximal effect of inhibi-tion (Fig. 6C and D), suggesting hypoxia-stimulated upregulation ofDEPP was dependent on FoxO.
4. Discussion
In the present study, we provide evidence that transcription fac-tor FoxO regulates expression of DEPP in human endothelial cells.FoxO-regulated transcription may be divided to several classesaccording to whether an insulin response sequence (IRS) is re-quired [4]. We demonstrate that binding to IRS element is essentialin FoxO-activated DEPP expression, which is consistent with previ-ous report [4]. We further demonstrate that FoxO binds to the
DEPP promoter at �179 to �172 and �151 to �143 relative tothe start of cDNA. These new findings support that DEPP is a noveltarget gene of FoxO in human endothelial cells.
Recently, DEPP was reported to be upregulated by feeding ininsulin-sensitive tissues [2]. Treatment of cultured adipocytesand HepG2 hepatoma cells with insulin significantly decreasedDEPP mRNA levels in dose- and time-dependent manners [2].Our findings help to explain the above observations. Since insulin-and Akt-dependent phosphorylation results in inactivation andtranslocation of FoxO into the cytoplasm, thereby DEPP expressionis inhibited.
The physiological significance of DEPP largely remains un-known. Our findings indicate that the function of DEPP is connectedto FoxO. In the current study, we observed a prominent increase ofDEPP expression in endothelial cells line EA.hy926 when stimulatedby hypoxia. Similar results were reported previously in a malignantglioma cell line [5]. Therefore, we speculate that DEPP may partic-ipate in cellular stress response. Interestingly, the induction of DEPPby hypoxia was not observed in HUVEC, indicating the response tohypoxia is different between transformed and primary endothelialcells. Whether DEPP is upregulated by hypoxic stress in HUVEC ex-

Fig. 3. Identification of FoxO-responsive sites in the DEPP promoter region. (A)Sequence of probes used in EMSA, mutations of FoxO binding sites are indicated asbold in sequence. (B, C) Binding of FoxO to DEPP promoter in vitro. RepresentativeEMSA demonstrated an electromobility shift when a probe containing the indicatedsequence of the DEPP promoter incubated with nuclear extracts from HEK293 cellstransfected with FoxO plasmids.
Fig. 4. FoxO binds to DEPP promoter in vivo. (A) CHIP assays were performed inHEK293 cells transfected with control or Myc-FoxO1TM plasmids. FoxO1 wasimmunoprecipitated with anti-Myc antibody and PCR amplification was performedusing the primer pair encompassing the last two FoxO binding sites. (B) CHIP assaywith endogenous FoxO1 was performed in EA.hy926 cells with anti-FoxO1antibodies and amplified using the same primer pair.
S. Chen et al. / FEBS Letters 585 (2011) 1796–1800 1799
posed to laminar flow conditions or in vivo remains to be eluci-dated. Besides, several studies have revealed that FoxO factors play
Fig. 5. DEPP expression was stimulated by hypoxia. EA.hy926 cells were placed in a hypanalyzed by real-time PCR. Data are mean ± SD from three experiments. ⁄P < 0.01 versustimulated with 1 mM DMOG for 6 h and analyzed by western blot. (D) HUVECs were e
an essential role in endothelial cell morphogenesis and vascularhomeostasis [13–15]. It was reported that DEPP was upregulatedin subsets of endothelial cells in settings of adult neo-vasculariza-tion including tumor angiogenesis [3]. Since hypoxia is a drivingforce in neo-vascularization in physiology and disease, whether hy-poxia-induced DEPP upregulation is involved in vascularizationneeds further analysis.
Although hypoxia has been shown to stimulate expression ofFoxO3a in mouse embryonic fibroblasts 11 and in glEND.2 cells[12], no increase of FoxO3a protein or mRNA was detected in HU-VEC or EA.hy926. As other HIF1 target genes were upregulated asexpected in our studies, we speculate that the different responseof FoxO to hypoxia was due to the distinct cells studied, sincethe function of FoxO proteins has been shown to be highly con-text-specific and may differ even in endothelial cells obtained fromdifferent vascular beds [15]. As there is no detectable alteration ofFoxO1 and FoxO3a at protein levels or their subcellular distribu-tion, whether the induction of DEPP mRNA by hypoxia in endothe-lial cell line EA.hy926 is mediated by increased FoxO binding to theDEPP promoter or via other molecules remains to be investigated.
oxia incubator chamber (A) or treated with 1 mM DMOG (B) for indicated time ands control. (C) EA.hy926 cells were exposed to hypoxia (pO2 = 40 mmHg) for 8 h orxposed to hypoxia for 8 h and analyzed by western blot.
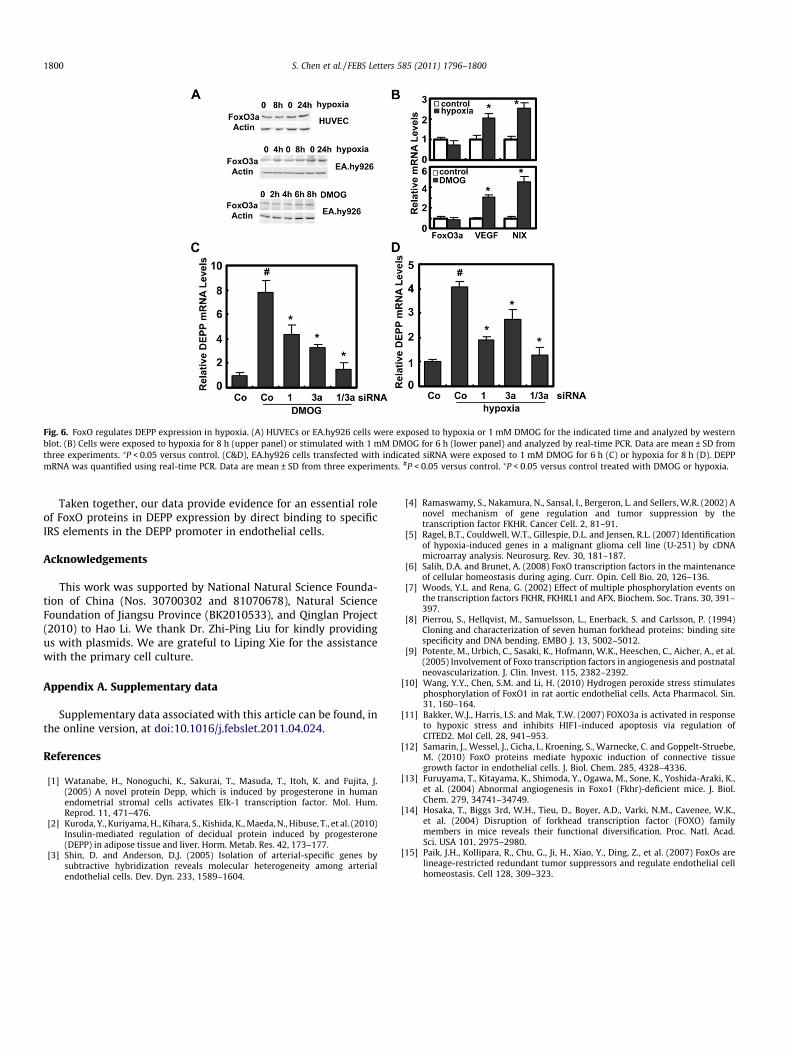
Fig. 6. FoxO regulates DEPP expression in hypoxia. (A) HUVECs or EA.hy926 cells were exposed to hypoxia or 1 mM DMOG for the indicated time and analyzed by westernblot. (B) Cells were exposed to hypoxia for 8 h (upper panel) or stimulated with 1 mM DMOG for 6 h (lower panel) and analyzed by real-time PCR. Data are mean ± SD fromthree experiments. ⁄P < 0.05 versus control. (C&D), EA.hy926 cells transfected with indicated siRNA were exposed to 1 mM DMOG for 6 h (C) or hypoxia for 8 h (D). DEPPmRNA was quantified using real-time PCR. Data are mean ± SD from three experiments. #P < 0.05 versus control. ⁄P < 0.05 versus control treated with DMOG or hypoxia.
1800 S. Chen et al. / FEBS Letters 585 (2011) 1796–1800
Taken together, our data provide evidence for an essential roleof FoxO proteins in DEPP expression by direct binding to specificIRS elements in the DEPP promoter in endothelial cells.
Acknowledgements
This work was supported by National Natural Science Founda-tion of China (Nos. 30700302 and 81070678), Natural ScienceFoundation of Jiangsu Province (BK2010533), and Qinglan Project(2010) to Hao Li. We thank Dr. Zhi-Ping Liu for kindly providingus with plasmids. We are grateful to Liping Xie for the assistancewith the primary cell culture.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.febslet.2011.04.024.
References
[1] Watanabe, H., Nonoguchi, K., Sakurai, T., Masuda, T., Itoh, K. and Fujita, J.(2005) A novel protein Depp, which is induced by progesterone in humanendometrial stromal cells activates Elk-1 transcription factor. Mol. Hum.Reprod. 11, 471–476.
[2] Kuroda, Y., Kuriyama, H., Kihara, S., Kishida, K., Maeda, N., Hibuse, T., et al. (2010)Insulin-mediated regulation of decidual protein induced by progesterone(DEPP) in adipose tissue and liver. Horm. Metab. Res. 42, 173–177.
[3] Shin, D. and Anderson, D.J. (2005) Isolation of arterial-specific genes bysubtractive hybridization reveals molecular heterogeneity among arterialendothelial cells. Dev. Dyn. 233, 1589–1604.
[4] Ramaswamy, S., Nakamura, N., Sansal, I., Bergeron, L. and Sellers, W.R. (2002) Anovel mechanism of gene regulation and tumor suppression by thetranscription factor FKHR. Cancer Cell. 2, 81–91.
[5] Ragel, B.T., Couldwell, W.T., Gillespie, D.L. and Jensen, R.L. (2007) Identificationof hypoxia-induced genes in a malignant glioma cell line (U-251) by cDNAmicroarray analysis. Neurosurg. Rev. 30, 181–187.
[6] Salih, D.A. and Brunet, A. (2008) FoxO transcription factors in the maintenanceof cellular homeostasis during aging. Curr. Opin. Cell Bio. 20, 126–136.
[7] Woods, Y.L. and Rena, G. (2002) Effect of multiple phosphorylation events onthe transcription factors FKHR, FKHRL1 and AFX. Biochem. Soc. Trans. 30, 391–397.
[8] Pierrou, S., Hellqvist, M., Samuelsson, L., Enerback, S. and Carlsson, P. (1994)Cloning and characterization of seven human forkhead proteins: binding sitespecificity and DNA bending. EMBO J. 13, 5002–5012.
[9] Potente, M., Urbich, C., Sasaki, K., Hofmann, W.K., Heeschen, C., Aicher, A., et al.(2005) Involvement of Foxo transcription factors in angiogenesis and postnatalneovascularization. J. Clin. Invest. 115, 2382–2392.
[10] Wang, Y.Y., Chen, S.M. and Li, H. (2010) Hydrogen peroxide stress stimulatesphosphorylation of FoxO1 in rat aortic endothelial cells. Acta Pharmacol. Sin.31, 160–164.
[11] Bakker, W.J., Harris, I.S. and Mak, T.W. (2007) FOXO3a is activated in responseto hypoxic stress and inhibits HIF1-induced apoptosis via regulation ofCITED2. Mol Cell. 28, 941–953.
[12] Samarin, J., Wessel, J., Cicha, I., Kroening, S., Warnecke, C. and Goppelt-Struebe,M. (2010) FoxO proteins mediate hypoxic induction of connective tissuegrowth factor in endothelial cells. J. Biol. Chem. 285, 4328–4336.
[13] Furuyama, T., Kitayama, K., Shimoda, Y., Ogawa, M., Sone, K., Yoshida-Araki, K.,et al. (2004) Abnormal angiogenesis in Foxo1 (Fkhr)-deficient mice. J. Biol.Chem. 279, 34741–34749.
[14] Hosaka, T., Biggs 3rd, W.H., Tieu, D., Boyer, A.D., Varki, N.M., Cavenee, W.K.,et al. (2004) Disruption of forkhead transcription factor (FOXO) familymembers in mice reveals their functional diversification. Proc. Natl. Acad.Sci. USA 101, 2975–2980.
[15] Paik, J.H., Kollipara, R., Chu, G., Ji, H., Xiao, Y., Ding, Z., et al. (2007) FoxOs arelineage-restricted redundant tumor suppressors and regulate endothelial cellhomeostasis. Cell 128, 309–323.


















