
Self-Referential Thinking and Equilibrium as States of Mind in Games: FMRI Evidence

Chapter
FMRI Evidence for ACV98 Connectionist Model for Reading of Mono and Multi-Syllabic Words and
Pseudo-Words
Author: Monica Baciu1 Co-authors: Bernard Ans, Serge Carbonnel, Sylviane Valdois, Alexandra Juphard
Author and co-authors affiliation: Laboratoire de Psychologie et Neurocognition, UMR CNRS 5105
Université Pierre Mendès-France, BP 47, 38040 Grenoble Cedex 09
France
Abstract According to predictions made by ACV98 connectionist model of reading, behavioral experiments have shown that syllabic length affects naming latencies for pseudo-words but not for words. ACV98 postulates the existence of two successive reading procedures. According to it, any orthographical object (word or pseudo-word) is first submitted to a global processing which aims to find a “lexical familiarity” computed from previously experienced words. If it is found, the corresponding phonological form of the stimulus is activated and reading is performed. If it is not found, a subsequent analytical processing is performed within the input stimulus using the same route, in order to extract familiar orthographic components (typically, syllables) as well as their corresponding phonological forms. The syllabic phonological forms are successively and temporarily maintained in a phonological buffer, before being assembled into a whole phonological form. Overall, this model predicts that words are read by using the global procedure, while the pseudo-words are read by using the analytical procedure. The present event-related fMRI study aimed to assess the effect of syllabic length on cerebral activity during reading, in order to obtain additional
1 Corresponding author :Monica Baciu, MD, PhD,Laboratoire de Psychologie et Neurocognition, UMR CNRS
5105,Université Pierre Mendès-France,BP 47, 38040 Grenoble Cedex 09, France, Phone : 00 33 4 76 82 78 07,Fax : 00 33 4 76 82 78 34,Email : [email protected]

Monica Baciu 2
anatomo-functional information as support to this model as well as to behavioral results. Based on ACV98 predictions and in terms of cerebral network, we hypothesized (1) a lexicality effect: there should be no differences between words and pseudo-words, they are processed within a common network of cerebral regions, and (2) a syllable length effect: the length influences cerebral activity only during pseudo-words reading because only long pseudo-words are processed following an analytical procedure involving supplementary visual analysis, visuo-spatial attention and working memory processes. Eight right-handed volunteers performed a silent reading task on French printed words and pseudo-words. A pseudo-randomized event-related paradigm with 6 types (words=W and pseudo-words=PW composed of 1, 2 and 3 syllables) of stimuli was used. Data processing was performed using SPM’99. Our results have shown that (1) Words and pseudo-words involved common mechanisms such as visuo-orthographic, phonological, attentional and motor processes. No process was significantly more involved for one or the other of two types of stimuli (word or pseudo-word). This result is a priori more in agreement with “single way models” and particularly with ACV’98 than with dual-route models. Nonetheless the dual-route model cannot be excluded; (2) A length effect on cerebral activity was obtained only during pseudo-word reading suggesting an analytical procedure involvement during reading of long pseudo-words, as ACV’98 predicts.
Introduction Understanding cerebral mechanisms of reading means understanding how the
orthographic form of visual linguistic stimuli is translated into their phonological form. Several theoretical models were developed and tried to answer this question. The classical cognitive “dual-route” model (Coltheart et al., 2001) for reading postulates the existence of two completely isolated routes that work in parallel and rely on radically different computational principles: one “lexical-semantic” used for the processing of words and another “phonological” used mainly for the sequential processing of pseudo-words (grapheme-to-phoneme rules). According to dual-route model, two different networks of cerebral regions will be expected to be activated, one for words and another one for pseudo-words, independently on the item length. These differences are explained in terms of sub-lexical phonological processing required by pseudo-word reading with respect to word (even for monosyllabic stimuli). Although the psychological models of reading do not directly predict that length has an effect on cerebral activation, we can nonetheless formulate hypotheses from effects expected at the behavioural level and from cerebral imaging data on reading. Therefore, within the framework of dual-route models (Coltheart et al., 2001), a length effect could be predicted during pseudo-word reading only. Indeed, the phonological form of pseudo-words can only be obtained from the sequential processing using the non lexical route (grapheme-to-phoneme translation). Furthermore, reading words would mainly be performed by using the lexical route. As this route allows to access directly the global phonological form of the word, no length effect is predicted for words. Based on this functional architecture of the dual-route model, we can assume that the reading of words and pseudo-words would involve different cerebral regions underlying each of the two routes. Cerebral regions allowing accessing orthographical and phonological, as well as semantic representations, such as occipito-temporal (Dehaene et al., 2002; Fiebach et al., 2002; Jobard et al., 2003; Price et al., 1994), temporo-parietal and inferior frontal cortex (Fiez and Petersen, 1998, St George et al., 1999; Xu et al., 2002) would mainly be related to the lexical route. As for

FMRI Evidence for ACV8 Connectionist Model for Reading … 3
the non lexical route, it may implicate regions related to grapheme-phoneme transformation, such as the left superior temporal region (Price et al., 1994; Jobard et al., 2003). The non-lexical route would additionally require the working memory processes in order to store, maintain and manipulate extracted phonological segments. The left supramarginal gyrus and the dorsal portion of the inferior frontal cortex were shown as being related to these processes (Paulesu et al, 1993; Gold and Buckner, 2002; Jobard et al., 2003; Thierry et al., 1999). Although the dual-route model did not make predictions on syllabic length, one could suppose that increasing the pseudo-words length should more intensively activate regions inherent to the non lexical route, whereas increasing the words length would have no influence on the degree of activation of regions underlying the lexical route.
Several connectionist models of reading were subsequently developed and they all proposed a unitary theory for reading. These models postulate that all orthographic items are read in the same way, i.e. the pronunciation is always computed from knowledge about previously experienced exemplars (familiarity). For instance, within PDP (Parallel Distributed Processing) models (Plaut, et al., 1996; Seidenberg and McClelland, 1989), the reading of words and of pseudo-words is performed via a unique global processing way, without involving any serial process In consequence, a common network of cerebral regions involved in orthographic (occipito-temporal cortex), phonological (posterior superior temporal and supramarginal cortex, as well as the dorsal part of the left inferior prefrontal gyrus) and semantic (middle and inferior temporal cortex and angular gyrus, as well as the anterior inferior part of the inferior prefrontal gyrus) processes, should be involved, whatever the lexical nature of the items or their length is. Depending on orthographic and phonological characteristics of the items, a length effect could emerge. It would then consist in a more significant activation of this cerebral network for the longest items in comparison with the shortest items. However, the increased activation of this cerebral network, due to the increase in length, should not differ between words and pseudo-words, the latter being processed by a common mechanism.
Among the unitary conception of reading, the model developed by Ans et al. (Ans, et al., 1998) (thereafter ACV98), is the only one that can simulate reading of multi-syllabic items, the others being restricted to a monosyllabic processing. It postulates the existence of two sequential reading procedures. According to it, any orthographical object (word or pseudo-word) is first submitted to a global processing which aims to find a “lexical familiarity” computed from previously experienced words. If it is found, the corresponding phonological form of the stimulus is activated and reading is performed. If it is not found, a subsequent analytical processing is performed within the input stimulus using the same route, in order to extract familiar orthographic components (typically, syllables) as well as their corresponding phonological forms. These syllabic phonological forms are successively and temporarily maintained in a phonological buffer, before being assembled into a whole phonological form. This model predicts that words are read by using the global procedure, while pseudo-words are read by using the analytical procedure. The simulations done with the ACV98 model have shown that monosyllabic pseudo-words also required a short analytical processing but the differences between monosyllabic words and monosyllabic pseudo-words have not been detected in behavioural experiments, in terms of time responses. This can be explained by the weak analytical procedure required by monosyllabic pseudo-words (only one syllable to be re-analyzed after the prior unsuccessfully global analysis) with respect to multi-syllabic pseudo-words. In terms of cerebral neuroanatomy of language and based on ACV98

Monica Baciu 4
predictions, the reading of the two types of items should generate a common network activation of cerebral regions related to the global procedure, mainly related to orthographic (occipito-temporal cortex) and phonological (posterior superior temporal and supramarginal cortex, as well as the dorsal part of the left inferior prefrontal gyrus) processes. Although not predicted by the model, semantic processing (middle and inferior temporal cortex and angular gyrus, as well as the anterior inferior part of the inferior prefrontal gyrus) could be also involved. Furthermore, as the model predicts, the analytical procedure is activated when long pseudo-words are presented. It implies additional processes such as a supplementary visuo-orthographic analysis (striate and extrastriate occipital areas) and visuo-spatial attention (superior parietal lobule). Those two types of processes would be indicative respectively of the reduction of the size of the visuo-attentional window and its shift to the different segments of the input item (typically syllables). This analytical procedure would also involve the verbal working memory (left inferior frontal cortex, Broca's area, BA 44 and temporo-parietal cortex, Wernicke's area, BA 22 ; supramarginal gyrus, BA 40), more particularly related to the subvocal articulation, phonological process and storage.
In this chapter, we present an fMRI study which aims to assess the cerebral regions that underlie silent reading of words and of pseudo-words of different syllabic lengths. According to our theoretical frame of reference, the ACV98 multi-trace model, the syllabic length effect should impact on cerebral activation only during pseudo-word reading.
More specifically, according to the ACV98 model and in terms of cerebral activation, we formulated the following hypotheses:
Firstly, independently of the syllabic length, as words and pseudo-words are processed within a unique system, we expect to observe the activation of a common network of cerebral regions for both types of items. More precisely, this network should include the occipito-temporal cortex (related to orthographic processing) as well as the temporo-parietal cortex and the anterior part of the inferior frontal cortex, Broca's area (related to phonological processing). The semantic processing is not predicted by ACV’98 model as being essential for reading, but regions classically related to this process, as well as to other auxiliary processes (such as motor processes related to the preparation and the control of the motor act of speech, even if speech was inner in this study), could be observed and included within the common network. We will refer at this first assumption, as the lexicality effect.
Secondly, we expect to observe a length effect on the cerebral activation for pseudo-words only. Increasing syllabic length of pseudo-words, implies the analytical procedure, meaning, as described previously, the reduction of size and the shift of visuo-attentional window, sequentially from one segment (typically the syllable) of the visual input, to the next one, till the whole visual item is processed. Verbal working memory will be involved and responsible for generating phonological form of each syllable and maintaining them within the phonological buffer in order to constitute the whole phonological form of the visual input. Consequently, the length effect during pseudo-word reading, should be illustrated by stronger involvement of visual associative (~ BA 18,19), of superior parietal (~ BA 7), supramarginal (~ BA 40) and Broca's (~ BA 44, 45 left) regions for long pseudo-words when compared to short pseudo-words and long words. We will refer at this second assumption as the length effect.

FMRI Evidence for ACV8 Connectionist Model for Reading … 5
Material and Methods
Subjects
Eight healthy volunteers (5 women, average age 23.6 years) gave their informed
consent to participate at this experiment that was approved by the local ethics committee. All the subjects of this experience were right-handed as assessed by the Edinburgh Handedness inventory (Oldfield, 1971) and they were all French native speakers and had no visual or reading problems.
Stimuli and Procedure
A pseudo-randomized ER-fMRI paradigm with six types of stimuli (words composed
of 1, 2 and 3 syllables and pseudo-words composed of 1, 2 and 3 syllables), was used. Overall, 144 words and pseudo-words composed of one (3-5 letters), two (6-8 letters) and three (9-11 letters) syllables were presented during a single fMRI session. The words were mid-frequency regular words while the pseudo-words have been obtained by manipulating words. In addition to words and pseudo-words, 34 null-events (ten of them at the end of the session) composed of a blank screen and a fixation cross on the center of the screen were also included. A fixation cross was presented between stimuli on the center of the screen. The presentation time of each stimulus was 200 ms and the average interval between the stimuli of the same type was 14 seconds. The stimuli were written in black lower-case letters on a white screen (« Courier new 45 points » bold). The stimuli were generated by means of Psyscope V.1.1 (Carnegie Mellon Department of Psychology) on a Macintosh computer (Power Macintosh 9600). They were transmitted into the magnet by means of a video projector (Eiki LC 6000), a projection screen situated behind the magnet and a mirror centered above the patient’s eyes.
The subjects were instructed to read each item silently, without articulating and vocalizing.
MR acquisition
Functional MR imaging was performed on a 1.5 Tesla MR imager (Philips NT) with
echo-planar (EPI) acquisition. Twenty-three adjacent, axial slices (thickness 5 mm each) were imaged. The imaging volume was oriented parallel to the bicommissural plane. Positioning of the image planes was performed on scout images acquired in the sagittal plane. An EPI MR pulse sequence was used. The major MR acquisition parameters of the EPI sequence were: TR = 2000 ms, TE = 45 ms, flip angle = 90°, field-of-view = 256x256 mm2, imaging matrix = 64x64, reconstruction matrix = 128x128. Subsequent to the functional scan, a high resolution 3D anatomical MR scan was obtained from the volume previously examined.
Data Processing

Monica Baciu 6
Data analysis was performed based on the general linear model (Friston, et al., 1995) for event-related designs, implemented in SPM’99 software (Wellcome Department of Imaging Neuroscience, London, UK) running on a Unix workstation under the MATLAB environment (Mathworks, Sherbon, USA).
A. Spatial Pre-Processing
MR images were processed using the following steps. First, during the slice timing step, the functional volumes were corrected for sampling bias effects caused by the different time acquisition of each slice composing the functional volume, relative to the haemodynamic response. In the second step, the realignment, motion correction was applied by using rotations and translations in order to realign each functional volume to the first acquired one. In a third step, the anatomical volume was spatially normalized into the Talairach and Tournoux reference space (Talairach and Tournoux, 1988) using as template a representative brain from the Montreal Neurological Institute. The anatomical normalization parameters were subsequently applied to functional volumes. Finally, in conformity with the assumption that data are normally distributed, the functional images were spatially smoothed by using a Gaussian filter (8 mm width).
B. Statistics
For each type of individual events, regressors of interest were created by convolving a delta function at each event onset with a canonical haemodynamic response function. A fixed effect group analysis was performed. Clusters of activated voxels were then identified, based on the intensity of the individual responses (p<0.05 corrected and p<0.001 uncorrected for multiple comparisons) and the spatial extent (clusters composed of at least 5 voxels).
B.1. Evaluation of cerebral substrates for word and pseudo-word
reading (lexicality effect)
In order to assess the neural substrates involved during word and pseudo-word reading, we calculated the following contrasts: W vs. fixation (regions related to word reading, PW vs. fixation (regions related to pseudo-word reading), W vs. PW (regions significantly more activated during word than during pseudo-word reading) and PW vs. W (regions significantly more activated during pseudo-word than during word reading).
B.2. Evaluation of Cerebral Substrates for Mono and Three-Syllabic
(Length Effect) Word and Pseudo-Word Reading
We focused only on the extreme stimuli, meaning words and pseudo-words composed of one and three syllables. In order to assess the neural substrate presenting a length effect, we calculated the following contrasts: W1 vs. W3 (regions significantly more activated for short word reading with respect to long words), W3 vs. W1 (regions significantly more activated for long word reading with respect to short word), PW1 vs. PW3 (regions significantly more activated for short pseudo-word reading with respect to long pseudo-word reading) and PW3 vs. PW1 (regions significantly more activated for long pseudo-word reading with respect to short pseudo-word reading). By taking into

FMRI Evidence for ACV8 Connectionist Model for Reading … 7
account the lexical type (word or pseudo-word), the contrast W3 vs. PW3 was performed in order to assess regions significantly more activated for long word than long pseudo-word reading, while the contrast PW3 vs. W3 has highlighted regions significantly more activated for long pseudo-word than long word reading. We also calculated the following contrasts (interactions): (PW3 vs. PW1) vs. (W3 vs. W1) in order to assess if a possible length effect for pseudo-words is significantly greater than a possible length effect for words, and (W3 vs. W1) vs. (PW3 vs. PW1) in order to assess if a possible length effect for words is significantly greater than a possible length effect for pseudo-words.
Results All activations reported in this sub-section were obtained based on a fixed effect
group analysis.
A. Lexicality Effect
As shown in Figure 1, independently of the item length, the same cerebral regions were activated during word (W vs. fixation) and pseudo-word (PW vs. fixation) reading. Overall, these regions mentioned in detail in Table 1, were activated bilaterally: the infero-medial occipito-temporal gyrus, the superior parietal lobule, the left inferior parietal lobule (supramarginal and angular gyrus), the motor and premotor cortices, as well as the right cerebellum. The direct contrast between words and pseudo-words W vs. PW and PW vs. W did not show any cerebral region activated significantly more by one type of item with respect to the other one.
Figure 1 Lexicality effect: The images (lateral views of the left hemisphere on the right and of the right hemisphere on the left) represent in yellow the commonly activated cerebral regions during word and
pseudo-word reading, independently of their length. More precisely, the contrast W vs. Fixation should induce activation represented in « green », while the contrast PW vs. Fixation should induce activation
represented in « red ». All activations represented on these images are « yellow » suggesting that all regions were commonly activated by words and pseudo-words. The activated regions are shown in Table
1. LH = left hemisphere, RH = right hemisphere.

Monica Baciu 8
B. Syllabic Length Effect
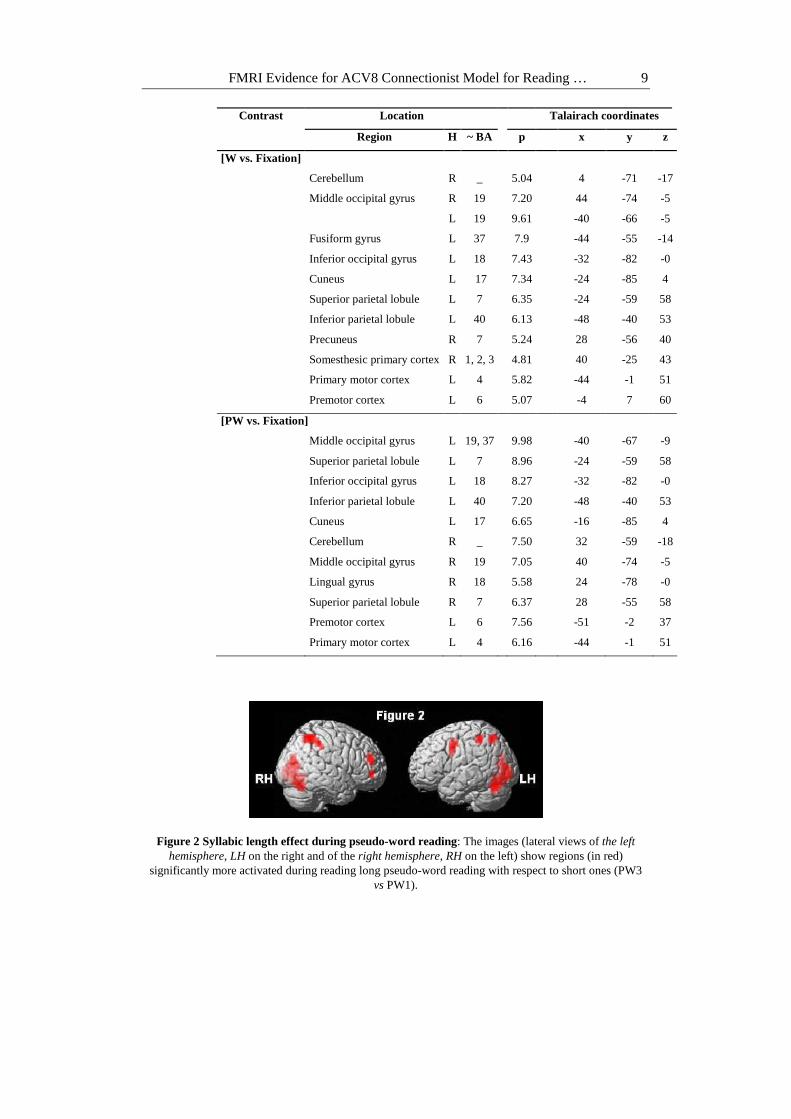
During reading, three-syllabic words (W3) did not induce significantly more activation when contrasted to mono-syllabic words (W1). Three-syllabic pseudo-words (PW3) induced significantly greater cerebral activity when contrasted to monosyllabic pseudo-words (PW1) bilaterally within middle and inferior occipital gyrus, inferior and superior parietal lobule and cerebellum. The premotor cortex was activated to the left whereas the middle and inferior frontal gyrus were activated to the right (Figure 2, Figure 3 and Table 2).
Table 1. Significantly ctivated regions obtained during word and pseudo-word reading (W vs. fixation and PW vs. fixation). H corresponds to the hemisphere with L for activation lateralized within the left and R for activation lateralized within the right hemisphere; BA represents the approximate Brodmann Area corresponding to the activated region; p=significance value; x, y and z indicate for each activated region, the Talairach coordinates.
Table 1
FMRI Evidence for ACV8 Connectionist Model for Reading … 9
Contrast Location Talairach coordinates
Region H ~ BA p x y z
[W vs. Fixation]
Cerebellum R _ 5.04 4 -71 -17
Middle occipital gyrus R 19 7.20 44 -74 -5
L 19 9.61 -40 -66 -5
Fusiform gyrus L 37 7.9 -44 -55 -14
Inferior occipital gyrus L 18 7.43 -32 -82 -0
Cuneus L 17 7.34 -24 -85 4
Superior parietal lobule L 7 6.35 -24 -59 58
Inferior parietal lobule L 40 6.13 -48 -40 53
Precuneus R 7 5.24 28 -56 40
Somesthesic primary cortex R 1, 2, 3 4.81 40 -25 43
Primary motor cortex L 4 5.82 -44 -1 51
Premotor cortex L 6 5.07 -4 7 60
[PW vs. Fixation]
Middle occipital gyrus L 19, 37 9.98 -40 -67 -9
Superior parietal lobule L 7 8.96 -24 -59 58
Inferior occipital gyrus L 18 8.27 -32 -82 -0
Inferior parietal lobule L 40 7.20 -48 -40 53
Cuneus L 17 6.65 -16 -85 4
Cerebellum R _ 7.50 32 -59 -18
Middle occipital gyrus R 19 7.05 40 -74 -5
Lingual gyrus R 18 5.58 24 -78 -0
Superior parietal lobule R 7 6.37 28 -55 58
Premotor cortex L 6 7.56 -51 -2 37
Primary motor cortex L 4 6.16 -44 -1 51
Figure 2 Syllabic length effect during pseudo-word reading: The images (lateral views of the left hemisphere, LH on the right and of the right hemisphere, RH on the left) show regions (in red)
significantly more activated during reading long pseudo-word reading with respect to short ones (PW3 vs PW1).
Monica Baciu 10
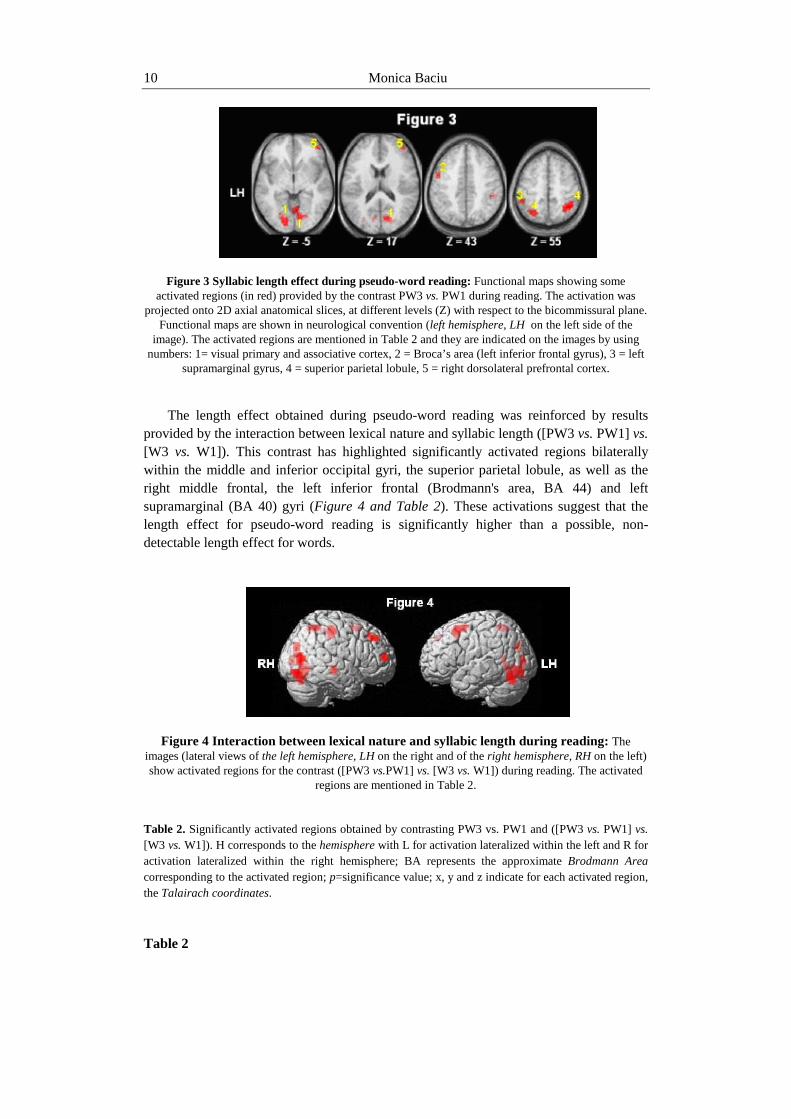
Figure 3 Syllabic length effect during pseudo-word reading: Functional maps showing some activated regions (in red) provided by the contrast PW3 vs. PW1 during reading. The activation was
projected onto 2D axial anatomical slices, at different levels (Z) with respect to the bicommissural plane. Functional maps are shown in neurological convention (left hemisphere, LH on the left side of the
image). The activated regions are mentioned in Table 2 and they are indicated on the images by using numbers: 1= visual primary and associative cortex, 2 = Broca’s area (left inferior frontal gyrus), 3 = left
supramarginal gyrus, 4 = superior parietal lobule, 5 = right dorsolateral prefrontal cortex.
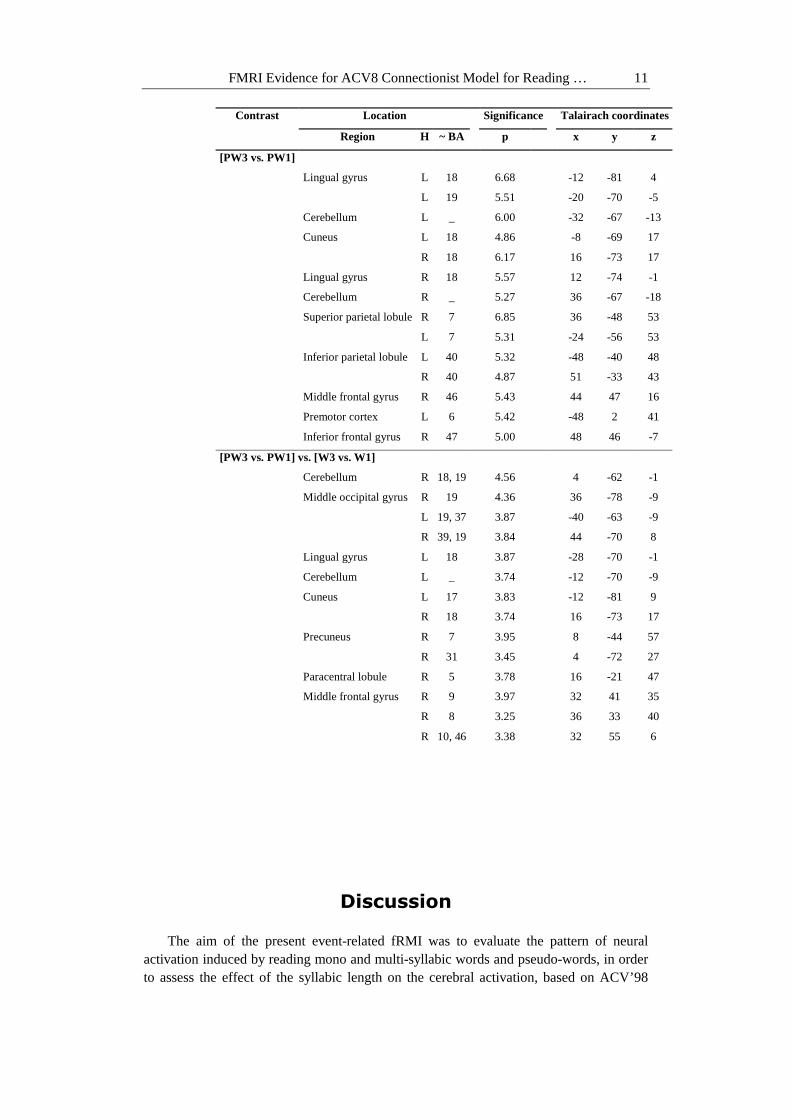
The length effect obtained during pseudo-word reading was reinforced by results
provided by the interaction between lexical nature and syllabic length ([PW3 vs. PW1] vs. [W3 vs. W1]). This contrast has highlighted significantly activated regions bilaterally within the middle and inferior occipital gyri, the superior parietal lobule, as well as the right middle frontal, the left inferior frontal (Brodmann's area, BA 44) and left supramarginal (BA 40) gyri (Figure 4 and Table 2). These activations suggest that the length effect for pseudo-word reading is significantly higher than a possible, non-detectable length effect for words.
Figure 4 Interaction between lexical nature and syllabic length during reading: The images (lateral views of the left hemisphere, LH on the right and of the right hemisphere, RH on the left) show activated regions for the contrast ([PW3 vs.PW1] vs. [W3 vs. W1]) during reading. The activated
regions are mentioned in Table 2.
Table 2. Significantly activated regions obtained by contrasting PW3 vs. PW1 and ([PW3 vs. PW1] vs. [W3 vs. W1]). H corresponds to the hemisphere with L for activation lateralized within the left and R for activation lateralized within the right hemisphere; BA represents the approximate Brodmann Area corresponding to the activated region; p=significance value; x, y and z indicate for each activated region, the Talairach coordinates.
Table 2
FMRI Evidence for ACV8 Connectionist Model for Reading … 11
Contrast Location Significance Talairach coordinates
Region H ~ BA p x y z
[PW3 vs. PW1]
Lingual gyrus L 18 6.68 -12 -81 4
L 19 5.51 -20 -70 -5
Cerebellum L _ 6.00 -32 -67 -13
Cuneus L 18 4.86 -8 -69 17
R 18 6.17 16 -73 17
Lingual gyrus R 18 5.57 12 -74 -1
Cerebellum R _ 5.27 36 -67 -18
Superior parietal lobule R 7 6.85 36 -48 53
L 7 5.31 -24 -56 53
Inferior parietal lobule L 40 5.32 -48 -40 48
R 40 4.87 51 -33 43
Middle frontal gyrus R 46 5.43 44 47 16
Premotor cortex L 6 5.42 -48 2 41
Inferior frontal gyrus R 47 5.00 48 46 -7
[PW3 vs. PW1] vs. [W3 vs. W1]
Cerebellum R 18, 19 4.56 4 -62 -1
Middle occipital gyrus R 19 4.36 36 -78 -9
L 19, 37 3.87 -40 -63 -9
R 39, 19 3.84 44 -70 8
Lingual gyrus L 18 3.87 -28 -70 -1
Cerebellum L _ 3.74 -12 -70 -9
Cuneus L 17 3.83 -12 -81 9
R 18 3.74 16 -73 17
Precuneus R 7 3.95 8 -44 57
R 31 3.45 4 -72 27
Paracentral lobule R 5 3.78 16 -21 47
Middle frontal gyrus R 9 3.97 32 41 35
R 8 3.25 36 33 40
R 10, 46 3.38 32 55 6
Discussion The aim of the present event-related fRMI was to evaluate the pattern of neural
activation induced by reading mono and multi-syllabic words and pseudo-words, in order to assess the effect of the syllabic length on the cerebral activation, based on ACV’98

Monica Baciu 12
predictions. According to ACV98 model predictions, we evaluated the lexicality effect (we expected to obtain the activation of a common network of cerebral regions during both word and pseudo-word reading), as well as the syllabic length effect (we expected that the syllabic length influences only pseudo-word reading, related to the supplementary analytical procedure required for reading long pseudo-words).
Lexicality Effect
Overall, words and pseudo-words activated a common network of cerebral regions.
This network included the left primary visual area (BA 17), the inferior and middle temporo-occipital gyrus (BA 18, 19, 37) as well as the inferior (SMG, BA 40) and superior (precuneus, BA 7) parietal lobule, bilaterally. As stimuli were visually presented, the primary visual area, responsible for visual perceptive processing, was activated (Cornelissen et al., 2003; Petersen et al., 1988, 1990, Tarkainien et al., 1999; Wise et al., 1991). The activation of the middle and inferior temporo-occipital gyrus could be related to orthographic processing (Fiez and Petersen, 1998; Petersen et al., 1988, 1990; Rumsey et al, 1997), whereas the inferior parietal lobule (supramarginal gyrus) would reflect the phonological processing (Devlin et al., 2003; Mc Dermott et al., 2003). The activation of the superior parietal lobule could be related to visuo-spatial attention processes (Chen et al., 2002; Corbetta et al., 1993; ; Corbetta et al., 1998; Simon et al., 2002) while the right cerebellum and the left premotor cortex (BA 6) activation could be related to motor aspects, such as the coordination and preparation of articulation in relation to the inner speech (Fiez, 1997; Fiez and Petersen, 1998; Haggort et al., 1999; Wise et al., 1991). Moreover, it was hypothesized that the cerebellum could be responsible for the control of attention (Ravizza and Ivry, 2001). Overall, the activation of the cerebellum obtained here could be attributed quite as much to the motor control of linguistic production (notably its medial part) as to the cognitive processes (notably its lateral part), such as attention and phonology (Ravizza and Ivry, 2001; Fiez, et al., 1996).
The activation of a common network of cerebral regions during the reading of words and of pseudo-words is in line with previous studies showing that similar regions are activated during the processing of those two types of stimuli (Chen, et al., 2002; Rumsey et al., 1997; Tagamets, et al., 2000). In return, no cortical region was significantly more implicated in the processing of one of the two types of item. This result is in disagreement with the idea suggesting that words and pseudo-words activate different regions (Chen et al., 2002; Fiez et al., 1997; Fulbright et al., 1999; Hagoort et al., 1999; Herbster et al., 1997; Rumsey et al., 1997; Xu et al., 2001). However, we can not completely exclude other reasons which could, even partially, have explained the absence of differential activation between words and pseudo-words. For instance, the very conservative statistical threshold of significance (p corrected < .05) used for this comparison, combined with the poor sampling of subjects having participated to this experiment, could explain this result. Moreover, even if the event-related paradigm, used in the present study, offers the advantage of allowing a randomized presentation of different types of stimuli, it is less sensitive in detecting activation than the « block » paradigm. Also, the limited number of items of each type could have also played a role in the absence of activation between words and pseudo-words.
The common network of regions activated for words and pseudo-words suggest common mechanisms to process the two types of stimuli. With respect to various models

FMRI Evidence for ACV8 Connectionist Model for Reading … 13
of reading, the intervention of common processes for word and pseudo-word reading, is rather in agreement with the “one way models” for reading, such as PMSP96 (Plaut, et al., 1996) and ACV98 (Ans, et al., 1998), that postulates the existence of a single system to process both, words and pseudo-words. However, it does not invalidate the dualistic approach for reading, underlying dual-route models, according to which words and pseudo-words would be processed within distinct systems. In fact, the fMRI spatial resolution, although relatively satisfying by comparing with other techniques such as the MEG or the ERP, techniques is not sufficient to guarantee the absence of intervention of different neuronal populations within this common cerebral network, specific to each types of item.
Syllabic Length Effect
Our main result is that the comparison of the longest items (3 syllables) with the
shortest ones (1 syllable) showed an influence of the increase of the syllabic length on the cerebral activity only during the reading of pseudo-words. This result is in agreement with the ACV98 reading model. It is also in agreement with the dual-route model (Coltheart et al., 2001), insofar as the number of letters increased with the number of syllables. On the other hand, within the framework of the PMSP96 model (Plaut et al., 1996), as words and pseudo-words are processed by a common procedure within a unique way, if a length effect appears, it should affect words as much as pseudo-words.
Compared to short pseudo-words, long pseudo-words induced an increased cerebral activity in a certain number of regions that are classically attributed to phonological, visuo-orthographic and visuo-spatial attention processing. More precisely, the bilateral inferior parietal lobule (supramarginal gyrus, BA 40) was more intensively activated during the reading of long than of short pseudo-words. This region, especially to the left, is typically associated with phonological processing, more particularly involved in phonological storage (Démonet et al., 1992; Fiez and Petersen, 1998; Fiez et al., 1999; McDermott et al., 2003; Gold and Buckner, 2002; Paulesu et al., 1993; Xu et al., 2002). The increased activation of the bilateral inferior parietal lobule suggests that phonological analysis was more intensively involved while processing long pseudo-words than short pseudo-words. Such an increase in activation, predominantly within the left hemisphere, could reflect a stronger involvement of phonological storage component of the verbal working memory during long pseudo-words reading (Gold and Buckner, 2002; Paulesu et al., 1993; Thierry et al., 1999). This activation is therefore in line with the ACV98 model (Ans et al., 1998) that postulates that the different syllables successively generated during the analytical procedure of long pseudo-words are maintained within the working memory in order to be assembled into a whole phonological form. According to this model, the working memory is not much involved during reading of short pseudo-words and of short and long words. The activation of the supramarginal gyrus is also in agreement with the dual-route model. In fact, this model postulates the short term maintenance of different phonemes extracted following the grapheme-to-phoneme rules, during the non lexical processing required to read pseudo-words.
In addition, long pseudo-words induced significantly greater activation, than short pseudo-words, within extrastriate visual cortex (BA 18, 19). The bilateral activation of the extrastriate visual cortex is typically associated with visual processing of orthographic sequences (Fiez and Petersen, 1998). Indefrey et al. (1997) have explicitly studied the

Monica Baciu 14
length effect of pseudo-words and of false characters of the same length, on the activation pattern of this region. Activation of the medial extrastriate cortex was found for each type of stimuli with respect to the baseline. On the other hand, those activations disappeared when pseudo-words were compared to sequences of false characters of same length. The authors concluded that the length difference in itself was sufficient to account for the extrastriate activation, suggesting that this region was mainly sensitive to physical characteristics of visual input. However, in the present study, the increased activation of extrastriate regions was obtained only by contrasting long pseudo-words with short pseudo-words. Nevertheless, it was not observed when we compared the long words with short words. This result therefore contradicts the hypothesis according to which the medial extrastriate activations are generated by the differences in physical length between items. In fact, in agreement with this hypothesis, increased activations of those regions should have been obtained for all the comparisons between the longest and the shortest items. According to other studies, the increased activation of extrastriate region could rather be related to greater visual complexity of the input stimuli (Hagoort et al., 1999; Tagamets et al., 2000; Tarkainien et al., 2002), rather than merely their physical characteristics. Such an interpretation has proven more consistent with the ACV98 model insofar as it postulates a visual processing, the degree of complexity of which increases with the increase in the pseudo-word length. Actually, according to this model, long pseudo-words would be processed syllable by syllable following the analytical procedure. The processing of each new syllable would require a new visuo-attentional capture. On the other hand, a strong implication of extrastriate regions during reading of long pseudo-words is not directly predicted by the DVC2001 model. This latter hypothesizes that visual analysis component (i.e., the layers of visual features and of letters) is common to lexical and non-lexical reading procedures. In consequence, a similar visual processing is supposed to apply to both words and pseudo-words. However, the non lexical route involves a segmentation mechanism that fractionates the internal representation of the orthographic input sequence into its different letters before the application of the grapheme-to-phoneme rules. If we hypothesize that those areas are activated during the manipulation of internal orthographic representations, such an orthographic segmentation could probably then result in an increased activation of the visual extrastriate areas. The increased activation of the extrastriate cortex could also result from the increase in visual attention (Clark et al., 1997; Corbetta et al., 1993). In fact, the increase in concomitant activation of the bilateral superior parietal lobule (BA 7) for long pseudo-words compared with short pseudo-words suggests a more extensive involvement of visuo-spatial attention processes during the analytic procedure using for long pseudo-words. The superior parietal area would be involved in the visuo-spatial analysis and attention (Behrmann, et al., 2004; Corbetta et al., 1998; Corbetta, et al., 1993; Posner and Petersen, 1990) and would be sensitive to the attention level allocated to a particular task (Fiez et al., 1995). Tagamets et al. (2000) have obtained parietal activations for sequences of false characters and of illegal letters but not for words and for pseudo-words composed of four letters. They argued that the activation of the superior parietal cortex (BA 7) could reflect the spatial processing necessary for organizing symbols in the unfamiliar sequences and/or an additional cognitive cost to process the new stimuli. The increased activation of this region, during three-syllabic pseudo-word reading, could therefore be related to the task properties that require a spatial processing allowing serial shifts of attention from one syllable to the other, within a sequence of letters. The activation of the superior parietal lobule could also be explained by the relative increase in eye movements in order to

FMRI Evidence for ACV8 Connectionist Model for Reading … 15
process long and unfamiliar sequences (Chen et al., 2002). However, since eye movements have not been measured, it is impossible for us to know if the activation of parietal regions reflects the direction of attention or the eye movements. In both cases, the activation of the superior parietal lobule for long pseudo-words reading in comparison with short pseudo-words is in line with the ACV98 model. The latter assumed that the analytical reading of polysyllabic pseudo-words implies a serial shift of the visuo-attentional window from the first to the last syllable of the input. In consequence, the analytical procedure would involve a redirection of both the attention and the eye movements.
Activations of the superior parietal lobule (BA 7) have also been reported in studies related to working memory, in particular when the task requires visual processing (Cabeza and Nyberg, 2000). The activation of the superior parietal lobule and of the extrastriate regions could then reflect an increase in visual attention within the context of short term memory processes. This latter alternative is in line with the additional activations of the inferior parietal lobule (BA 40) typically involved in phonological storage (Gold and Buckner, 2002; Paulesu et al., 1993 ; Thierry et al., 1999).
The left premotor cortex (BA 6), probably associated with the programming of articulation was activated. Its activation could also be interpreted as related to the articulatory loop (together with Broca’s area) within the framework of verbal working memory. The activation of the right middle (BA 46) and inferior frontal (BA 47) gyri is more difficult to interpret. This region could be related to “central administrator” within the framework of working memory but this region was generally activated to the left during verbal working memory. Furthermore, the ACV98 model predicts a “comparison” between external visual input and an internal visual input generated after recognizing the visual item within the lexicon. This comparison has a major role in triggering the analytical procedure: when external and internal visual input is the same, the analytical procedure is not activated, when they are different, this procedure is activated. The “comparison” process should depend on one or several cerebral regions. It is well known that the frontal/prefrontal cortex is very much involved in control and strategy processes, that is why this region could be involved, based on ACV’98 predictions, as “comparator” for triggering the analytical procedure. The increase in item’s length should solicit more intensively the “comparator”, especially when long items are pseudo-words.
Conclusion The main results of the present fMRI study were:
a) Word and pseudo-word reading involves common processes such as visuo-orthographic, phonological, attentional and motor processes. No process was significantly more involved for one or the other of two types of stimuli (word or pseudo-word). This result is a priori more in agreement with “single way models” than with “two distinct processing ways models”. Nonetheless the dual-route model cannot be excluded, insofar as we can consider that lexical and non lexical procedures involve different neural populations within the commonly activated regions obtained in this study.
b) We have obtained a syllabic length effect on cerebral activity only during pseudo-word reading and it represents the main

Monica Baciu 16
result of the present study. Within the framework of the ACV98 model, the activation of such regions can easily be attributed to analytical procedure involvement, which is predominantly activated while reading pseudo-word.
We will evaluate, in a subsequent study, a larger number of subjects in order to carry
out randomized effect analysis. In addition, the amplitude, the form and the time of the hemodynamic response will be evaluated within the different activated cerebral regions, so as to detect more subtle differences, with respect to both, the lexical nature and the syllabic length.
References
Ans, B., Carbonnel, S. and Valdois, S. (1998). A connectionist multiple-trace memory model for polysyllabic word reading. Psychological Review 105(4), 678-723.
Behrmann, M., Geng, J. J. and Shomstein, S. (2004). Parietal cortex and attention.
Current Opinion in Neurobiology, 14(2), 212-217. Cabeza, R. and Nyberg, L. (2000). Imaging cognition II: An empirical review of 275 PET
and fMRI studies. Journal of Cognitive Neuroscience, 12, 1-47. Chen, Y., Fu, S., Iversen, S. D. et al. (2002). Testing for dual brain processing routes in
reading: A direct contrast of Chinese character and pinyin reading using fMRI. Journal of Cognitive Neuroscience, 14(7), 1088-1098.
Clark, V.P., Parasuranam, R., Keil, K. et al. (1997). Selective attention to face identity
and color studied with fMRI. Human Brain Mapping, 5, 293-297. Coltheart, M., Rastle, K., Perry, C. et al. (2001). DRC: A Dual Route Cascaded model of
visual recognition and reading aloud. Psychological Review, 108(1), 204-256 Corbetta, M., Akbudak, E., Conturo, T. et al. (1998). A common network of functional
areas for attention and eye movements. Neuron, 21,761-773. Corbetta, M., Miezin, F.M., Shulman, G.L. and Petersen S.E. (1993). A PET study of
visuospatial attention. Journal of Neuroscience, 13(3), 1202-1226. Cornelissen, P., Tarkiainen, A., Helenius, P. and Salmelin, R. (2003). Cortical effects of
shifting letter position in letter strings of varying length. Journal of Cognitive Neuroscience, 15(5), 731-746.
Dehaene, S., Le Clec'H, G., Poline, J.-B. et al.(2002). The visual word form area: A
prelexical representation of visual words in the fusiform gyrus. NeuroReport, 13(3), 321-325. Démonet, J.-F., Chollet, F., Ramsay, S. et al. (1992).The anatomy of phonological and
semantic processing in normal subjects. Brain, 115(6), 1753 1768. Devlin, J. T., Matthews, P. M. and Rushworth, M. F. S. (2003). Semantic processing in
the left inferior prefrontal cortex: A combined functional magnetic resonance imaging and transcranial magnetic stimulation study. Journal of Cognitive Neuroscience, 15(1), 71-84.

FMRI Evidence for ACV8 Connectionist Model for Reading … 17
Fiebach, C. J., Friederici, A. D., Müller, K. and von Cramon, D. Y. (2002). fMRI
evidence for dual routes to the mental lexicon in visual word recognition. Journal of Cognitive Neuroscience, 14(1), 11-23.
Fiez, J. A. (1996). Cerebellar contributions to cognition. Neuron, 16, 13-15. Fiez, J. A. (1997). Phonology, semantics, and the role of the left inferior prefrontal cortex.
Human Brain Mapping, 5, 79-83. Fiez, J. A., Balota, D. A., Raichle, M. E. and Petersen, S. E. (1999). Effects of lexicality,
frequency, and spelling-to-sound consistency on the functional anatomy of reading. Neuron,24, 205-218.
Fiez, J. A. and Petersen, S. E. (1998). Neuroimaging studies of word reading. Proceeding
National Academic Science, 95, 914-921. Fiez, J. A., Raichle, M. E., Balota, D. A. et al. (1996). PET activation of posterior
temporal regions during auditory word presentation and verb generation. Cerebral Cortex, 6, 1-10.
Friston, K. J., Holmes, A. P., Worsley, K. J. et al. (1995). Statistical parametric maps in
functional imaging: A general linear approach. Human Brain Mapping, 3, 189-210. Fulbright, R. K., Jenner, A. R., Mencl, W. E. et al. (1999). The cerebellum's role in
reading: A functional MR imaging study. American Journal of Neuroradiology, 20, 1925-1930.
Gold, B. T. and Buckner, R. L. (2002). Common prefrontal regions coactivate with
dissociable posterior regions during controlled semantic and phonological tasks. Neuron, 35, 803-812.
Hagoort, P., Indefrey, P., Brown, C. et al. (1999). The neural circuitry involved in the
reading of German words and pseudowords: A PET study. Journal of Cognitive Neuroscience, 11(4), 383-398.
Herbster, A. N., Mintun, M. A., Nebes, R. D. and Becker, J. T. (1997). Regional cerebral
blood flow during word and non-word reading. Human Brain Mapping, 5, 84-92. Indefrey, P., Kleinschmidt, A., Merboldt, K.-D. et al. (1997). Equivalent responses to
lexical and nonlexical visual stimuli in occipital cortex: A functional magnetic resonance imaging study. NeuroImage, 5(1), 78-81.
Jobard, G., Crivello, F., and Tzourio-Mazoyer, N. (2003). Evaluation of the dual route
theory of reading: A metanalysis of 35 neuroimaging studies. NeuroImage, 20(2), 693-712. McDermott, K. B., Petersen, S. E., Watson, J. M. and Ojemann, J. G. (2003). A procedure
for identifying regions preferentially activated by attention to semantic and phonological relations using functional magnetic resonance imaging. Neuropsychologia, 41, 293-303.
Oldfield, R. C. (1971). The assessment and analysis of handness: The Edinburgh
inventory. Neuropsychologia, 9, 97-113. Paulesu, E., Frith, C. D. and Frackowiak, R. S. J. (1993). The neural correlates of the
verbal component of working memory. Nature, 362, 342-345.

Monica Baciu 18
Petersen, S. E., Fox, P. T., Posner, M. I. et al. (1988). Positron emission tomograpihc
studies of the cortical anatomy of single-word processing. Nature, 331, 585-589. Petersen, S. E., Fox, P. T., Snyder, A. Z. and Raichle, M. E. (1990). Activation of
extrastriate and frontal cortical areas by visual words and word-like stimuli. Science, 24, 1041-1043.
Plaut, D. C. (1997). Structure and function in the lexical system: Insights from distributed
models of word reading and lexical decision. Language and Cognitive Processes, 12(5/6), 765-805.
Plaut, D. C., McClelland, J. L., Seidenberg, M. S. and Patterson, K. (1996).
Understanding normal and impaired word reading: Computational principles in quasi-regular domains. Psychological Review, 103, 56-115.
Posner, M. I. and Petersen, S. E. (1990). The attention system of the human brain. Annual
Reviews of Neuroscience, 13, 25-42. Price, C. J., Wise, R., Watson, J. et al. (1994). Brain activity during reading: The effect of
task and exposure duration. Brain, 117, 1255-1269. Ravizza, S.M. and R.B. Ivry. (2001). Comparison of the Basal Ganglia and Cerebellum in
Shifting Attention. Journal of Cognitive Neuroscience, 13(3), 285-297.
Rumsey, J. M., Horwitz, B., Donohue, B. C. et al. (1997). Phonological and orthographic components of word recognition. A PET-rCBF study. Brain, 120, 739-759.
Seidenberg, M. S. and McClelland, J. L. (1989). A distributed, developmental model of
word recognition and naming. Psychological Review, 96, 523-568. Simon, O., Mangin, J. F., Cohen, L., Le Bihan, D. and Dehaene, S. (2002). Topographical
layout of hand, eye, calculation, and language-related areas in the human parietal lobe. Neuron, 33(3), 475-487.
St George, M., Kutas, M., Martinez, A. and Sereno, M. I. (1999). Semantic integration in
reading: Engagement of the right hemisphere during discourse processing. Brain, 122(7), 1317-1325.
Tagamets, M.-A., Novick, J. M., Chalmers, M. L. and Friedman, R. B. (2000). A
parametric approach to orthographic processing in the brain: An fMRI study. Journal of Cognitive Neuroscience, 12, 281-297.
Talairach, J. and Tournoux, P. (1988). Co-planar stereotaxic atlas of the human brain. 3-
Dimensional proportional system: An approach to cerebral imaging. New York: Thieme Medical.
Tarkiainen, A., Cornelissen, P. L. and Salmelin, R. (2002). Dynamics of visual feature
analysis and object-level processing in face versus letter-string perception. Brain, 125, 1125-1136.
Tarkiainen, A., Helenius, P., Hansen, P. C. et al. (1999). Dynamics of letter string
perception in the human occipitotemporal cortex. Brain, 122, 2119-2131.

FMRI Evidence for ACV8 Connectionist Model for Reading … 19
Thierry, G., Boulanouar, K., Kherif, F. et al. (1999). Temporal sorting of neural components underlying phonological processing. NeuroReport, 10, 2599-2603.
Wise, R., Hadar, U., Howard, D. and Patterson, K. (1991). Language activation studies
with positron emission tomography. Ciba Foundation Symposia, 163:218-28; discussion 228-34. Review.
Xu, B., Grafman, J., Gaillard, W. D. et al. (2001). Conjoint and extended neural networks
for the computation of speech codes: The neural basis of selective impairment in reading words and pseudowords. Cerebral Cortex, 11(3), 267-277.
Xu, B., Grafman, J., Gaillard, W. D. et al. (2002). Neuroimaging reveals automatic speech
coding during perception of written word meaning. NeuroImage, 17, 859-870.




















