
Eurooppaoikeus I EU ja Euroopan neuvosta EU-historiaa EU:n instituutiot EU-oikeuden lähteet.

COSTA
NZO
6a EDIÇÃO
LINDA S. COSTANZO
Fisiologia6 a E D I Ç Ã O
LINDA S. COSTANZO, PhD
Os alunos sempre dizem que o livro de Fisiologia da Dra. Costanzo é um dos seus favoritos. É fácil entender o motivo.
O texto simples, lógico e fácil de acompanhar da renomada docente de fisiologia Dra. Linda Costanzo faz com que Fisiologia, agora na 6ª edição, seja ideal para a disciplina e o preparo para provas.
Figuras e tabelas de excelente qualidade ilustram bem os procedimentos ou equações fisiológicas e as explicações passo a passo esclarecem conceitos desafiadores. Este texto colorido e de bom tamanho discute, de maneira abrangente e consistente, os principais conceitos fisiológicos em nível de sistema orgânico e celular, facilitando a compreensão de complexos princípios.
• As informações são apresentadas de forma objetiva, fácil e clara – a maneira perfeita de obter sucesso em cursos e provas.
• Os resumos dos capítulos e as perguntas da seção “Desafie-se”, ao final de cada capítulo, permitem uma revisão extensa do material e reforçam o entendimento e a retenção do conhecimento.
• As equações e amostras de problemas estão integrados por todo o texto.
• NOVO! Mais Quadros de Fisiologia Clínica relacionam a fisiopatologia com o contexto clínico
www.studentsconsult.com.br
A maneira inteligente de estudarstudentconsult
A MANEIRA INTELIGENTE DE ESTUDAR
Este livro tem conteúdo extra e gratuito no sitewww.evolution.com.br.Registre o código que está no verso da capa, dentro deste livro e conheça uma nova maneira de aprender:
- aprenda de forma dinâmica e objetiva com as animações;- teste seus conhecimentos com perguntas e respostas comentadas;- acesse os vídeos complementares.
A aquisição desta obra habilita o acesso ao sitewww.evolution.com.braté o lançamento da próxima edição em português, ou até que esta edição em português não esteja mais disponível para venda pela Elsevier, o que ocorrer primeiro.
Classificação de Arquivo Recomendada
Fisiologiastudentconsult
www.elsevier.com.br
Fisiologia
T R A D U Ç ÃO DA
6ª EDIÇ ÃO
Fisiologia
Costanzo_CoverMechanical Lu aprovada.indd 1 7/9/18 5:45 PM

Fisiologia
C0070.indd iC0070.indd i 04/07/18 9:06 AM04/07/18 9:06 AM

FisiologiaSEXTA EDIÇÃO
LINDA S. COSTANZO, PHDProfessor of Physiology and Biophysics
Virginia Commonwealth University School of MedicineRichmond, Virginia
C0070.indd iiiC0070.indd iii 04/07/18 9:06 AM04/07/18 9:06 AM

© 2018 Elsevier Editora Ltda. Todos os direitos reservados e protegidos pela Lei 9.610 de 19/02/1998. Nenhuma parte deste livro, sem autorização prévia por escrito da editora, poderá ser reproduzida ou transmitida sejam quais forem os meios empregados: eletrônicos, mecânicos, fotográfi cos, gravação ou quaisquer outros.
ISBN: 978-85-352-9034-9 ISBN versão eletrônica: 978-85-352-9035-6
PHYSIOLOGY, SIXTH EDITION Copyright © 2018 by Elsevier, Inc. All rights reserved.
This translation of Physiology 6 th Edition, by Linda S. Costanzo, was undertaken by Elsevier Editora Ltda. and is published by arrangement with Elsevier Inc.
Esta tradução de Physiology 6 th Edition, de Linda S. Costanzo foi produzida por Elsevier Editora Ltda. e publicada em conjunto com Elsevier Inc.
ISBN: 978-0-323-47881-6
Capa Luciana Mello e Monika Mayer
Editoração Eletrônica Thomson Digital
Elsevier Editora Ltda. Conhecimento sem Fronteiras
Rua da Assembleia, n° 100 – 6° andar – Sala 601 20011-904 – Centro – Rio de Janeiro – RJ
Av. Nações Unidas, nº 12995 – 10º andar 04571-170 – Brooklin – São Paulo – SP
Serviço de Atendimento ao Cliente 0800 026 53 40 [email protected]
Consulte nosso catálogo completo, os últimos lançamentos e os serviços exclusivos no site www.elsevier.com.br
Nota Esta tradução foi produzida por Elsevier Brasil Ltda. sob sua exclusiva responsabilidade. Médicos e pesquisadores devem sempre fundamentar-se em sua experiência e no próprio conhecimento para avaliar e empregar quaisquer informações, métodos, substâncias ou experimentos descritos nesta publicação. Devido ao rápido avanço nas ciências médicas, particularmente, os diagnósticos e a posologia de medicamentos precisam ser verifi cados de maneira independente. Para todos os efeitos legais, a Editora, os autores, os editores ou colaboradores relacionados a esta tradução não assumem responsabilidade por qualquer dano/ou prejuízo causado a pessoas ou propriedades envolvendo responsabilidade pelo produto, negligência ou outros, ou advindos de qualquer uso ou aplicação de quaisquer métodos, produtos, instruções ou ideias contidos no conteúdo aqui publicado.
CIP-BRASIL. CATALOGAÇÃO NA PUBLICAÇÃOSINDICATO NACIONAL DOS EDITORES DE LIVROS, RJ
C873f6. ed.
Costanzo, Linda S. Fisiologia / Linda S. Costanzo ; [tradução Aline Santana da Hora, Renata Scavone de Oliveira]. - 6. ed. - Rio de Janeiro : Elsevier, 2018. : il. ; 28 cm.
Tradução de: Physiology Apêndice Inclui índice ISBN 978-85-352-9034-9
1. Fisiologia humana. I. Oliveira, Aline Santana da Hora. II. Scavone de, Renata. III. Título.
18-50154 CDD: 612 CDU: 612
C0075.indd ivC0075.indd iv 04/07/18 9:14 AM04/07/18 9:14 AM

Para
Heinz Valtin e Arthur C. Guyton,
que tão bem escreveram para estudantes de fi siologia
Richard, Dan, Rebecca, Sheila, Elise e Max,
que fazem tudo valer a pena
C0105.indd vC0105.indd v 04/07/18 5:29 AM04/07/18 5:29 AM

vi
Tradução e Revisão Científi ca
REVISÃO CIENTÍFICA
Everson Araújo Nunes Professor Doutor do Departamento de Ciências Fisiológicas Centro de Ciências Biológicas Universidade Federal de Santa Catarina
Gustavo Jorge dos Santos, PhD Biólogo, Mestre em Biologia Funcional e Molecular e Doutor em Biologia Funcional e Molecular - Fisiologia Professor e Coordenador de Pesquisa do Departamento de Ciências Fisiológicas (CFS) do Centro de Ciências Biológicas (CCB) da Universidade Federal de Santa Catarina (UFSC) Professor do Programa de Pós-Graduação Multicêntrico em Ciências Fisiológicas (PPGMCF)
Mariana Terenzi Professora Associada do Departamento de Ciências Fisiológicas de Santa Catarina (UFSC) Pós-doutorado na University of Bristol, Reino Unido Doutora em Fisiologia pela Cardiff University, Reino Unido Mestre em Farmacologia pela Universidade de São Paulo Graduada em Farmácia pela Universidade de São Paulo
Vander Baptista Professor Doutor do Departamento de Ciências Fisiológicas Centro de Ciências Biológicas Universidade Federal de Santa Catarina
TRADUÇÃO
Aline Santana da Hora Doutoranda em Epidemiologia Experimental Aplicada às Zoonoses pela Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo (FMVZ-USP) Mestre em Clínica Veterinária pela FMVZ-USP Médica Veterinária pela Universidade do Estado de Santa Catarina (CAV-UDESC)
Renata Scavone de Oliveira Médica Veterinária formada pela Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo Doutora em Imunologia pelo Instituto de Ciências Biomédicas da Universidade de São Paulo
C0080.indd viC0080.indd vi 04/07/18 9:21 AM04/07/18 9:21 AM

vii
Prefácio
A fi siologia é o fundamento da prática médica. O conhecimento sólido de seus princípios é essencial para o aluno de medicina e para o médico. Este livro é destinado a alunos de medicina e disciplinas relacionadas que estão estudando fi siologia. Pode ser usado como suporte em palestras e conteúdos programáticos de currículos disciplinares ou como fonte primária em currículos integrados ou práticos. Para alunos avançados, o livro pode ser uma referência em cursos de fi siopatologia e estágios clínicos.
Na sexta edição deste livro, como nas edições anteriores, os conceitos importantes da fi siologia são discutidos em níveis celulares e de sistemas orgânicos. Os Capítulos 1 e 2 apresentam os princípios subjacentes da fi siologia celular e do sistema nervoso autônomo. Os Capítulos 3 a 10 , os principais sistemas orgânicos: neurofi siologia e fi siologia cardiovas-cular, respiratória, renal, acidobásica, gastrintestinal, endócrina e reprodutiva. As relações entre os sistemas orgânicos são enfatizadas para destacar os mecanismos integrativos da homeostase.
Nesta edição, as seguintes características facilitam o estudo da fi siologia:
♦ O texto é fácil de ler e conciso: cabeçalhos claros orientam o aluno quanto à organização e à hierarquia do material. As informações fi siológicas complexas são apresentadas de forma sistemática, lógica e gradual. Quando o processo ocorre em uma sequência específi ca, as etapas são numeradas no texto e, de modo geral, correlacionadas com números mostrados na fi gura correspondente. Marcadores são usados para separar e destacar as características de um processo. Em todo o texto, perguntas retóricas antecipam as dúvidas que os alunos possam ter; ao ver e, então, responder a essas perguntas, o aluno aprende a explicar conceitos difíceis e racionalizar achados inesperados ou paradoxais. Os resumos dos capítulos fazem recapitulações curtas.
♦ Tabelas e ilustrações que podem ser usadas com o texto ou, por serem autoexplicativas, como revisão: as tabelas resumem, organizam e comparam as informações. Exemplos são (1) a tabela que compara os hormônios gastrintestinais quanto à família hormonal, ao local e ao estímulo para a secreção e as ações de cada molécula; (2) a tabela que compara as características fi siopatológicas dos distúrbios da homeostase de Ca 2+ ; e (3) a tabela que compara as características do potencial de ação em diferentes tecidos cardíacos. As ilustrações são identifi cadas de maneira clara e apresentam cabeçalhos principais, diagramas simples, diagramas complexos com etapas numeradas e fl uxo-gramas.
♦ Equações e exemplos de problema integrados ao texto: todos os termos e unidades das equações são defi nidos e cada equação é reescrita em palavras, a fi m de colocá-la em seu contexto fi siológico. Os exemplos de problema são acompanhados por soluções numéricas completas e explicações que orientam os alunos ao longo das etapas adequadas de resolução; seguindo essas etapas, os alunos ganham as habilidades e a confi ança para resolver problemas similares.
♦ Fisiologia clínica , apresentada em quadros: cada quadro traz um paciente fictício com uma doença clássica. Os achados clínicos e o tratamento proposto são explicados conforme os princípios fi siológicos subjacentes. A abordagem ao paciente é integrada, enfatizando as relações entre os sistemas orgânicos. Por exemplo, o caso do diabetes mellitus tipo I discute a doença não apenas do sistema endócrino, mas também dos sistemas renal, acidobásico, respiratório e cardiovascular.
C0085.indd viiC0085.indd vii 04/07/18 9:29 AM04/07/18 9:29 AM

viii • Prefácio
♦ Perguntas práticas nas seções “Desafi e-se” ao fi nal de cada capítulo: as perguntas, feitas de forma a terem respostas curtas (uma palavra, uma frase ou uma solução numérica), desafi am o aluno a aplicar os princípios e conceitos à solução de problemas, e não relembrar fatos isolados. As perguntas, que são de diversos formatos e colocadas em ordem aleatória, serão mais efi cazes se usadas como ferramentas após o estudo de cada capítulo e sem consultar o texto. Dessa forma, o aluno pode confi rmar sua compreensão do material e determinar suas áreas de defi ciência. As respostas são dadas no fi nal do livro.
♦ Vídeos didáticos sobre tópicos selecionados: uma vez que a explicação oral de princípios complexos pode ajudar os alunos, pequenos vídeos didáticos sobre tópicos selecionados complementam o texto escrito.
♦ Abreviações e valores normais apresentados em apêndices: ao consultar e utilizar essas abreviações e valores comuns por todo o livro, os alunos os memorizam com naturalidade.
Este livro incorpora três de minhas crenças sobre o ensino: (1) mesmo informações complexas podem ser transmitidas com clareza se a apresentação for sistemá-tica, lógica e gradual; (2) a apresentação pode ser tão efi caz por escrito quanto pessoalmente; e (3) os alunos que começam a estudar medicina querem materiais de ensino precisos e com didática forte, mas sem os detalhes que interessam principalmente aos especialis-tas. Em essência, um livro pode “ensinar” se a voz do professor estiver presente, o material for cuidadosamente selecionado para inclusão de informações essenciais e se houver muito cuidado quanto à lógica e à sequência. Este texto é uma apresentação realista e profi ssional escrita para alunos.
Espero que os leitores deste livro gostem do estudo da fi siologia. Aqueles que aprendem bem seus princípios serão recompensados ao longo de suas carreiras profi s-sionais!
Linda S. Costanzo
C0085.indd viiiC0085.indd viii 04/07/18 9:29 AM04/07/18 9:29 AM

ix
Agradecimentos
Agradeço muito a Elyse O’Grady, Jennifer Ehlers e Dan Fitzgerald, da Elsevier, pelas con-tribuições no preparo da sexta edição de Fisiologia. Ao artista Matthew Chansky, que revisou as fi guras existentes e criou novas ilustrações que complementam lindamente o texto.
Aos colegas da Virginia Commonwealth University, que responderam prontamente às minhas dúvidas, em especial aos Drs. Clive Baumgarten, Diomedes Logothetis, Roland Pittman e Raphael Witorsch. Também agradeço sinceramente aos alunos de medicina de todo o mundo que, de forma generosa, escreveram para mim sobre suas experiências com as edições anteriores do livro.
A meu marido, Richard; aos nossos fi lhos, Dan e Rebecca; à nossa nora, Sheila; e aos nossos netos, Elise e Max, que me deram o apoio entusiasmado e o amor incondicional que permeiam este livro.
C0090.indd ixC0090.indd ix 04/07/18 9:50 AM04/07/18 9:50 AM

xi
Sumário
1 Fisiologia Celular, 1
2 Sistema Nervoso Autônomo, 47
3 Neurofi siologia, 69
4 Fisiologia Cardiovascular, 117
5 Fisiologia Respiratória, 189
6 Fisiologia Renal, 245
7 Fisiologia Acidobásica, 311
8 Fisiologia Gastrintestinal, 339
9 Fisiologia Endócrina, 395
10 Fisiologia Reprodutiva, 461
Apêndice I Abreviações e Símbolos Comuns, 483
Apêndice II Valores Normais e Constantes, 485
Respostas dos Exercícios (Desafi e-se), 487
Índice, 493
C0095.indd xiC0095.indd xi 04/07/18 9:55 AM04/07/18 9:55 AM

47
CAPÍTULO 2
Sistema Nervoso Autônomo
Organização e Características Gerais do Sistema Nervoso Autônomo, 47
Receptores Autonômicos, 59
Resumo, 66
Desafi e-se, 66
O sistema nervoso motor (eferente) possui dois compo-nentes: o somático e o autônomo. Estes dois sistemas são bastante diferentes, principalmente no que se refere aos tipos de órgãos efetores que inervam e os tipos de função que controlam.
O sistema nervoso somático é um sistema motor que tanto pode ser voluntário , sob controle consciente, como involuntário . Cada uma de suas vias é composta por um único motoneurônio e as fi bras musculares esqueléticas por ele inervadas. O corpo celular do motoneurônio está localizado no sistema nervoso central (SNC), seja no tronco encefálico ou na medula espinal, e seus axônios fazem sinapses diretamente na musculatura esquelética, o órgão efetor. O neurotransmissor acetilcolina é liberado de terminais pré-sinápticos dos motoneurônios e ativa receptores nicotínicos localizados nas placas motoras da musculatura esquelética. Um potencial de ação no motoneurônio provoca um potencial de ação na fi bra muscular, causando a contração do músculo. (Ver a discussão completa sobre o sistema nervoso somático no Capítulo 1 )
O sistema nervoso autônomo é um sistema involuntário que controla e modula as funções principalmente dos órgãos viscerais. Cada via do sistema nervoso autônomo é composta por dois neurônios: um pré-ganglionar e um pós-ganglionar. O corpo celular de cada neurônio pré-ganglionar reside no sistema nervoso central. Os axônios desses neurônios pré-ganglionares fazem sinapses com os corpos celulares dos neurônios pós-ganglionares em um dos diversos gânglios autonômicos localizados fora do sistema nervoso central. Os axônios dos neurônios pós-ganglionares trafegam, então, até seus alvos, onde fazem sinapses com órgãos efetores viscerais, como o coração, os bronquíolos, a musculatura lisa vascular, o trato gastrintestinal, a bexiga e a genitália. Todos os neurônios pré-ganglionares do sistema nervoso autônomo liberam acetilcolina. Os neurônios pós-ganglionares liberam acetilcolina ou noradrenalina ou, ainda, em alguns casos, neuropeptídeos.
ORGANIZAÇÃO E CARACTERÍSTICAS GERAIS DO SISTEMA NERVOSO AUTÔNOMO
O sistema nervoso autônomo apresenta duas divisões principais: a divisão simpática e a divisão parassimpática, que geralmente são complementares na regulação da função do órgão visceral. Uma terceira divisão do sistema autônomo, o sistema nervoso entérico, localiza-se nos plexos do trato gastrintestinal. (O sistema nervoso entérico é discutido no Capítulo 8 )
A organização do sistema nervoso autônomo é descrita na Figura 2.1 e na Tabela 2.1 . As divisões simpática e parassimpática, assim como o sistema nervoso somático, são incluídas para comparação.
C0010.indd 47C0010.indd 47 03/07/18 4:08 PM03/07/18 4:08 PM
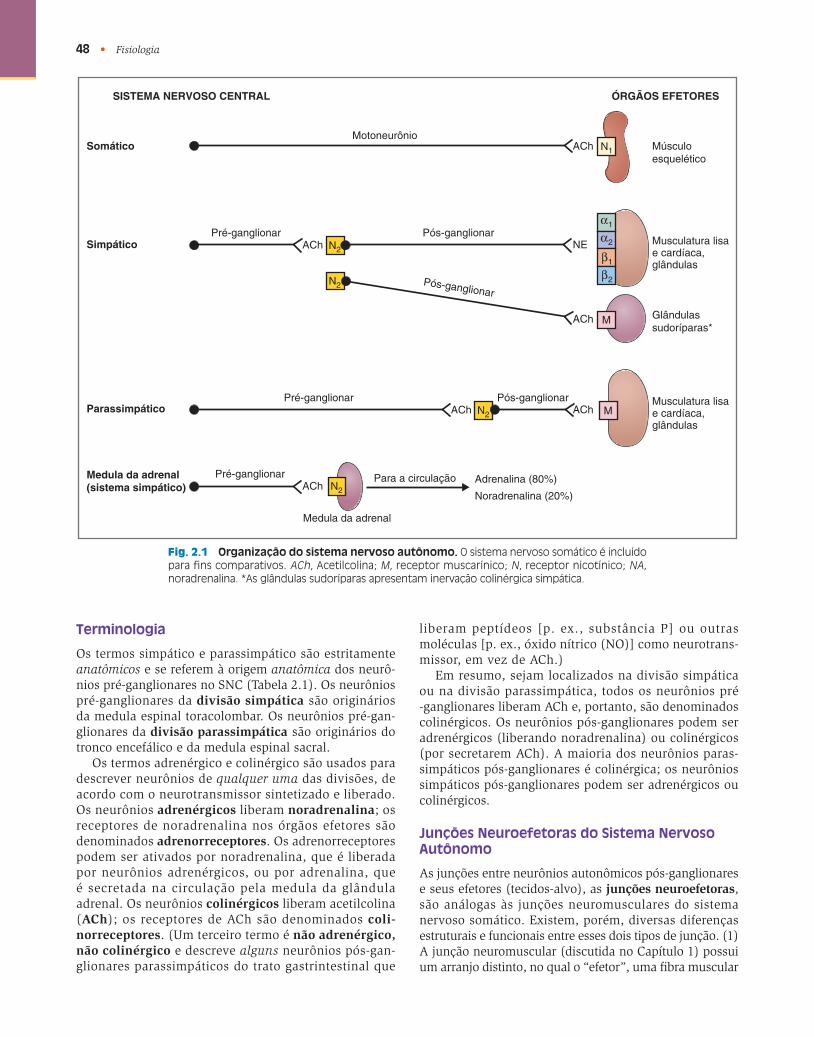
48 • Fisiologia
Terminologia
Os termos simpático e parassimpático são estritamente anatômicos e se referem à origem anatômica dos neurô-nios pré-ganglionares no SNC ( Tabela 2.1 ). Os neurônios pré-ganglionares da divisão simpática são originários da medula espinal toracolombar. Os neurônios pré-gan-glionares da divisão parassimpática são originários do tronco encefálico e da medula espinal sacral.
Os termos adrenérgico e colinérgico são usados para descrever neurônios de qualquer uma das divisões, de acordo com o neurotransmissor sintetizado e liberado. Os neurônios adrenérgicos liberam noradrenalina ; os receptores de noradrenalina nos órgãos efetores são denominados adrenorreceptores . Os adrenorreceptores podem ser ativados por noradrenalina, que é liberada por neurônios adrenérgicos, ou por adrenalina, que é secretada na circulação pela medula da glândula adrenal. Os neurônios colinérgicos liberam acetilcolina ( ACh ); os receptores de ACh são denominados coli-norreceptores . (Um terceiro termo é não adrenérgico, não colinérgico e descreve alguns neurônios pós-gan-glionares parassimpáticos do trato gastrintestinal que
liberam peptídeos [p. ex., substância P] ou outras moléculas [p. ex., óxido nítrico (NO)] como neurotrans-missor, em vez de ACh.)
Em resumo, sejam localizados na divisão simpática ou na divisão parassimpática, todos os neurônios pré-ganglionares liberam ACh e, portanto, são denominados colinérgicos. Os neurônios pós-ganglionares podem ser adrenérgicos (liberando noradrenalina) ou colinérgicos (por secretarem ACh). A maioria dos neurônios paras-simpáticos pós-ganglionares é colinérgica; os neurônios simpáticos pós-ganglionares podem ser adrenérgicos ou colinérgicos.
Junções Neuroefetoras do Sistema Nervoso Autônomo
As junções entre neurônios autonômicos pós-ganglionares e seus efetores (tecidos-alvo), as junções neuroefetoras , são análogas às junções neuromusculares do sistema nervoso somático. Existem, porém, diversas diferenças estruturais e funcionais entre esses dois tipos de junção. (1) A junção neuromuscular (discutida no Capítulo 1 ) possui um arranjo distinto, no qual o “efetor”, uma fi bra muscular
ÓRGÃOS EFETORESSISTEMA NERVOSO CENTRAL
Somático
Simpático
Parassimpático
Medula da adrenal(sistema simpático)
Músculoesquelético
Musculatura lisae cardíaca,glândulas
Glândulassudoríparas*
Musculatura lisae cardíaca,glândulas
Pré-ganglionar
ACh
ACh
Adrenalina (80%)Para a circulação
Noradrenalina (20%)
Medula da adrenal
M
M
NEPós-ganglionar
Pós-ganglionar
AChPré-ganglionar
α1
α2
β1
β2
AChPré-ganglionar Pós-ganglionar
ACh N2
N2
N1
ACh
Motoneurônio
N2
N2
Fig. 2.1 Organização do sistema nervoso autônomo. O sistema nervoso somático é incluído para fi ns comparativos. ACh , Acetilcolina; M , receptor muscarínico; N , receptor nicotínico; NA , noradrenalina. *As glândulas sudoríparas apresentam inervação colinérgica simpática.
C0010.indd 48C0010.indd 48 03/07/18 4:08 PM03/07/18 4:08 PM

2—Sistema Nervoso Autônomo • 49
esquelética, é inervada por um único motoneurônio. Por outro lado, no sistema nervoso autônomo, os neurônios pós-ganglionares que inervam os tecidos-alvo formam redes ramificadas e difusas . Estruturas em formato esférico, ou varicosidades , recobrem esses ramos e são sítios de síntese, armazenamento e liberação de neurotransmissores. As varicosidades são, portanto, análogas aos terminais nervosos pré-sinápticos da junção neuromuscular. (2) Há uma sobreposição entre as redes ramifi cadas de diferentes neurônios pós-ganglionares e, assim, os tecidos-alvo podem ser inervados por várias dessas células. (3) No sis-tema nervoso autônomo, os receptores pós-sinápticos são amplamente distribuídos pelos tecidos-alvo e não há uma região especializada de receptores como a placa motora da fi bra muscular esquelética.
Sistema Nervoso Simpático
A função principal do sistema nervoso simpático é mobilizar o corpo para a atividade . Em caso extremo, quando uma pessoa é exposta a uma situação estres-sante, o sistema nervoso simpático é fortemente ativado, em uma resposta conhecida como “luta ou fuga”, que inclui o aumento da pressão arterial, do fl uxo sanguíneo para os músculos ativados, da taxa metabólica, da glicemia, da atividade mental e do grau de alerta. No entanto, mesmo que essa resposta, por si só, seja raramente empregada, o sistema nervoso
simpático opera continuamente em grau moderado para modular as funções de muitos sistemas de órgãos, como o coração, os vasos sanguíneos, o trato gastrintestinal, os brônquios e as glândulas sudoríparas.
A Figura 2.2 mostra a organização do sistema nervoso simpático em relação à medula espinal, aos gânglios simpáticos e aos órgãos efetores da periferia. Os neurônios simpáticos pré-ganglionares são originários dos núcleos da medula espinal toracolombar, deixam a medula através das raízes motoras ventrais e dos ramos brancos e se projetam para os gânglios paravertebrais da cadeia simpática ou em uma série de gânglios pré-vertebrais. Assim, uma categoria de neurônios pré-ganglionares faz sinapses com neurônios pós-ganglionares nos gân-glios paravertebrais (p. ex., gânglio cervical superior) da cadeia simpática . Essas sinapses podem ocorrer em gânglios do mesmo nível segmentar da cadeia ou as fi bras pré-ganglionares podem cursar em direção cranial ou caudal e inervar gânglios em níveis superiores ou inferiores da cadeia, permitindo, assim, o estabelecimento de sinapses em múltiplos gânglios (o que é consistente com as características difusas das funções simpáticas). A outra categoria de neurônios pré-ganglionares atravessa pela cadeia simpática sem estabelecer sinapses para somente fazê-las nos gânglios pré-vertebrais (celíacos, mesentéricos superiores e mesentéricos inferiores) que suprem os órgãos viscerais, as glândulas e o sistema nervoso entérico do trato gastrintestinal. Nos gânglios,
TABELA 2.1 Organização do Sistema Nervoso Autônomo
Características Divisão Simpática Divisão Parassimpática Sistema Nervoso Somático a
Origem dos neurônios pré-ganglionares
Segmentos T1-L3 da medula espinal (toracolombares)
Núcleos dos NC III, VII, IX e X; segmentos S2-S4 da medula espinal (craniossacrais)
—
Localização dos gânglios autonômicos
Paravertebral e pré-vertebral Em órgãos efetores ou em suas adjacências
—
Comprimento dos axônios pré-ganglionares
Curto Longo —
Comprimento dos axônios pós-ganglionares
Longo Curto —
Órgãos efetores Musculatura lisa; músculo cardíaco; glândulas
Musculatura lisa; músculo cardíaco; glândulas
Musculatura esquelética
Junções neuroefetoras Difusas, ramifi cadas; os receptores não estão concentrados em uma única região
Difusas, ramifi cadas; os receptores não estão concentrados em uma única região
Discretas, organizadas; os receptores de ACh estão localizados na placa motora
Neurotransmissor e tipo de receptor nos gânglios
ACh/receptor nicotínico ACh/receptor nicotínico —
Neurotransmissor em órgãos efetores
Noradrenalina (exceto em glândulas sudoríparas, ACh)
ACh ACh
Tipos de receptores em órgãos efetores
α 1 , α 2 , β 1 , β 2 (exceto em glândulas sudoríparas, muscarínico)
Muscarínicos Nicotínicos
ACh , Acetilcolina; NC , nervo craniano. a O sistema nervoso somático é incluído para comparação.
C0010.indd 49C0010.indd 49 03/07/18 4:08 PM03/07/18 4:08 PM

50 • Fisiologia
Medula espinal
Gânglio cervicalsuperior
Cadeiasimpática
Cadeiasimpática
Par
a os
vas
os s
angu
íneo
s, o
s m
úscu
los
pilo
eret
ores
e a
s gl
ându
las
sudo
rípa
ras
Genitáliamasculina
Plexomesentérico inferior
Intestino grosso
Estômago
Árvore brônquica
Glândula parótida
Glândulassubmandibularese sublinguais
Músculo tarsal
Glândula lacrimal
Músculo radial;dilata a pupila
Intestino delgado
L3
T1
Plexomesentérico superior
Plexo celíaco
Medulada adrenal
Coração
SISTEMA NERVOSO SIMPÁTICO
Fig. 2.2 Inervação do sistema nervoso simpático. Os neurônios pré-ganglionares são originários dos segmentos torácicos e lombares da medula espinal (T1-L3).
C0010.indd 50C0010.indd 50 03/07/18 4:08 PM03/07/18 4:08 PM

2—Sistema Nervoso Autônomo • 51
os neurônios pré-ganglionares fazem sinapses com os neurônios pós-ganglionares, que trafegam para a periferia e inervam os órgãos efetores.
As características do sistema nervoso simpático, dis-cutidas nas próximas seções, são listadas na Tabela 2.1 e ilustradas na Figura 2.2 .
Origem dos Neurônios Pré-ganglionares
Os neurônios pré-ganglionares da divisão simpática são originários de núcleos nos segmentos torácico e lombar da medula espinal, especifi camente do primeiro segmento torácico ao terceiro segmento lombar (T1-L3). Assim, a divisão simpática é chamada toracolombar .
De modo geral, a origem dos neurônios pré-ganglionares na medula espinal é anatomicamente consistente com sua projeção na periferia. Dessa maneira, as vias simpáticas para os órgãos do tórax (p. ex., o coração) possuem neurônios pré-ganglionares originários da medula espinal torácica superior . As vias simpáticas para os órgãos da pelve (p. ex., cólon, genitais) possuem neurônios pré-gan-glionares originários da medula espinal lombar . Os vasos sanguíneos, as glândulas sudoríparas termorreguladoras e os músculos piloeretores da pele apresentam neurônios pré-ganglionares que fazem sinapses com múltiplos neurônios pós-ganglionares acima e abaixo da cadeia simpática, refl etindo sua ampla distribuição pelo corpo.
Localização dos Gânglios Autonômicos
Os gânglios do sistema nervoso simpático estão locali-zados próximos à medula espinal , seja nos gânglios paravertebrais (conhecidos como cadeia simpática) ou nos gânglios pré-vertebrais. Mais uma vez, a anatomia é lógica. O gânglio cervical superior se projeta até os órgãos da cabeça, como os olhos e as glândulas salivares. O gânglio celíaco se projeta até o estômago e o intestino delgado. O gânglio mesentérico superior se projeta até os intestinos delgado e grosso e o gânglio mesentérico inferior atinge a porção inferior do intestino grosso, o ânus, a bexiga e a genitália.
A medula da glândula adrenal é simplesmente um gânglio simpático especializado cujos neurônios pré-ganglionares são originários da medula espinal torácica (T5-T9), passam pela cadeia simpática e pelo gânglio celíaco sem estabelecer sinapses e seguem pelo nervo esplâncnico maior até a adrenal.
Comprimento dos Axônios Pré-ganglionares e Pós-ganglionares
Uma vez que os gânglios simpáticos são próximos à medu-la espinal, os axônios dos neurônios pré-ganglionares são curtos e os axônios dos neurônios pós-ganglionares são longos (para que possam atingir os órgãos efetores periféricos).
Neurotransmissores e Tipos de Receptores
Os neurônios pré-ganglionares da divisão simpática são sempre colinérgicos . Estes neurônios liberam ACh,
que interage com receptores nicotínicos (N 2 ) nos corpos celulares dos neurônios pós-ganglionares. Os neurônios pós-ganglionares da divisão simpática são adrenérgicos em todos os órgãos efetores, exceto nas glândulas sudoríparas termorreguladoras (onde são colinérgicos). Os órgãos efetores inervados por neurônios adrenérgicos simpáticos apresentam um ou mais dos seguintes tipos de receptores: alfa 1 , alfa 2 , beta 1 ou beta 2 ( α 1 , α 2 , β 1 ou β 2 ). As glândulas sudoríparas termorreguladoras são inervadas por neurônios colinérgicos simpáticos que possuem colinorreceptores muscarínicos.
Varicosidades Adrenérgicas Simpáticas
Como anteriormente descrito, os neurônios adrenérgicos pós-ganglionares simpáticos liberam seus neurotrans-missores das varicosidades em tecidos-alvo (p. ex., a musculatura lisa vascular). As varicosidades adrenérgicas simpáticas contêm neurotransmissores clássicos (nora-drenalina) e não clássicos (trifosfato de adenosina [ATP] e neuropeptídeo Y). O neurotransmissor clássico, a noradrenalina , é sintetizada a partir da tirosina nas varicosidades ( Figura 1.18 ) e armazenada em pequenas vesículas de centro denso , prontas para serem liberadas; estas pequenas vesículas de centro denso também contêm dopamina β -hidroxilase, que catalisa a conversão da dopamina em noradrenalina (a etapa fi nal da via sintética) e ATP . O ATP é considerado “colocalizado” à adrenalina. Um grupo separado de grandes vesículas de centro denso contém neuropeptídeo Y .
A estimulação de neurônios adrenérgicos pós-gan-glionares simpáticos leva à liberação de noradrenalina e ATP das pequenas vesículas de centro denso. A noradre-nalina e o ATP atuam como neurotransmissores na junção neuroefetora, onde se ligam a receptores específicos nos tecidos-alvo (p. ex., a musculatura lisa vascular), ativando-os. Na verdade, o ATP age primeiro, ligando-se a receptores purinérgicos no tecido alvo e provocando um efeito fi siológico (p. ex., a contração da musculatura lisa vascular). A ação da noradrenalina ocorre após a ação do ATP; a noradrenalina se liga a seus receptores no tecido alvo (p. ex., receptores α 1 -adrenérgicos na musculatura lisa vascular), causando uma segunda contração, mais prolongada. Por fi m, com a estimulação mais intensa ou de maior frequência, as grandes vesículas de centro denso liberam neuropeptídeo Y, que se liga a seu receptor no tecido alvo e causa uma terceira fase de contração, mais lenta.
Medula da Glândula Adrenal
A medula da adrenal é um gânglio especializado na divisão simpática do sistema nervoso autônomo. Os corpos celula-res de seus neurônios pré-ganglionares estão localizados na medula espinal torácica. Os axônios destes neurônios pré-ganglionares trafegam pelo nervo esplâncnico maior até a medula da adrenal, onde fazem sinapses com as células cromafi ns e liberam ACh, ativando receptores nicotínicos. Quando ativadas, as células cromafi ns da
C0010.indd 51C0010.indd 51 03/07/18 4:08 PM03/07/18 4:08 PM

52 • Fisiologia
medula da adrenal secretam catecolaminas (adrenalina e noradrenalina) na circulação geral. Diferentemente dos neurônios pós-ganglionares simpáticos, que liberam apenas noradrenalina, a medula da adrenal secreta, prin-cipalmente, adrenalina (80%) e uma pequena quantidade de noradrenalina (20%). A razão para essa diferença é a presença de feniletanolamina- N -metiltransferase (PNMT) na medula da adrenal, mas não nos neurônios adrenérgicos pós-ganglionares simpáticos ( Figura 1.18 ). A PNMT catalisa a conversão da noradrenalina em adrenalina, e é interessante notar que essa etapa requer cortisol advindo do córtex adrenal adjacente; o cortisol é disponibilizado para a medula da adrenal graças ao fl uxo venoso do córtex adrenal.
Um tumor na medula da adrenal, ou feocromoci-toma , pode estar localizado nela mesma ou em suas proximidades ou, ainda, ser ectópico (em uma região distante) ( Quadro 2.1 ). Diferentemente da medula da adrenal normal, que secreta principalmente adrenalina,
um feocromocitoma secreta, em especial, noradrenalina , o que é explicado pela grande distância entre o tumor (se ectópico) e o córtex da adrenal, impedindo o recebimento do cortisol exigido pela PNMT.
Resposta de Luta ou Fuga
O corpo responde ao medo, ao estresse extremo e ao exercício intenso com uma ação maciça e coordenada do sistema nervoso simpático, inclusive da medula da adrenal. Essa ativação, a resposta de luta ou fuga, garante que o corpo possa responder adequadamente a uma situação estressante (p. ex., fazer uma prova difícil, correr de uma casa em chamas ou lutar contra um agressor). A resposta aumenta a frequência cardíaca, o débito cardíaco e a pressão sanguínea; redistribui o fl uxo sanguíneo da pele, dos rins e das regiões esplâncnicas para a mus-culatura esquelética; aumenta a ventilação, dilatando as vias aéreas; diminui a motilidade e as secreções do trato gastrintestinal e eleva a glicemia.
QUADRO 2.1 Fisiologia Clínica: Feocromocitoma
DESCRIÇÃO DO CASO. Uma mulher de 48 anos consulta seu médico, queixando-se do que chama de “ataques de pânico”. A paciente relata taquicardia e poder sentir (e até mesmo ver) o coração batendo no peito. Ela também se queixa de cefaleias do tipo pulsátil, mãos e pés frios, calor, distúrbios visuais, náusea e vômito. No consultório médico, sua pressão arterial é bastante alta (230/125). A paciente é internada para avaliação da hipertensão.
O exame de urina de 24 horas revela níveis elevados de metanefrina, normetanefrina e ácido 3-metoxi-4-hidroximandélico (VMA). Depois de descartar outras causas de hipertensão, o médico conclui que a paciente apresenta um tumor na medula da adrenal, denominado feocromocitoma. A tomografia computadorizada do abdômen revela a presença de uma massa de 3,5 cm na medula da adrenal direita. A paciente é tratada com um antagonista α 1 e submetida à cirurgia. A recuperação da paciente é completa; sua pressão arterial volta ao normal e os demais sintomas desaparecem.
EXPLICAÇÃO DO CASO. A mulher apresenta um feo-cromocitoma clássico, um tumor de células cromafi ns da medula da adrenal. O tumor secreta quantidades excessivas de noradrenalina e adrenalina, que produzem todos os sintomas e aumentam a concentração de meta-bólitos das catecolaminas na urina. Diferentemente da medula da adrenal normal, que secreta principalmente adrenalina, os feocromocitomas tendem a liberar nora-drenalina.
Os sintomas da paciente podem ser interpretados por meio do entendimento dos efeitos fisiológicos das catecolaminas. Qualquer tecido que possua adrenorreceptores será ativado pelos níveis elevados
de adrenalina e noradrenalina, que chegam pela circulação. Os principais sintomas apresentados pela paciente são cardiovasculares: palpitações, taquicardia, aumento da frequência cardíaca e da pressão arterial e mãos e pés frios. Estes sintomas podem ser entendidos ao considerarmos a função dos adrenorreceptores no coração e nos vasos sanguíneos. As maiores concen-trações de catecolaminas circulantes ativaram receptores β 1 do coração, aumentando a frequência cardíaca e a contratilidade (batimento cardíaco). A ativação dos receptores α 1 na musculatura lisa vascular da pele produziu vasoconstrição, causando o frio nas mãos e nos pés. A paciente sentiu calor, porém, porque essa vasoconstrição cutânea prejudicou a dissipação do calor. Sua pressão arterial extremamente elevada foi causada pela combinação entre a maior frequência cardíaca, a maior contratilidade e a maior contração (resistência) dos vasos sanguíneos. A cefaleia era secundária ao aumento da pressão.
Os demais sintomas apresentados pela paciente também podem ser explicados pela ativação de adrenorreceptores em outros sistemas de órgãos (isto é, os sintomas gastrintestinais de náusea e vômito e as alterações visuais).
TRATAMENTO. O tratamento da paciente foi compos-to pela localização e excisão do tumor, removendo, assim, a fonte do excesso de catecolaminas. Se o tumor não tivesse sido excisado, a paciente poderia continuar a ser tratada farmacologicamente com uma combinação de antagonistas α 1 (p. ex., fenoxi-benzamina ou prazosina) e antagonistas β 1 (p. ex., propranolol) para impedir as ações das catecolaminas endógenas nos receptores.
C0010.indd 52C0010.indd 52 03/07/18 4:08 PM03/07/18 4:08 PM

2—Sistema Nervoso Autônomo • 53
Sistema Nervoso Parassimpático
A função geral do sistema nervoso parassimpático é restauradora , de conservação de energia . A Figura 2.3 mostra a organização do sistema nervoso parassimpático em relação ao SNC (tronco encefálico e medula espinal), os gânglios parassimpáticos e os órgãos efetores. Os corpos celulares dos neurônios pré-ganglionares da divisão parassimpática estão no tronco encefálico (mesencéfalo, ponte e bulbo) ou na medula espinal sacral. Os axônios pré-ganglionares se projetam para uma série de gânglios localizados nos órgãos efetores ou suas adjacências.
As características do sistema nervoso simpático dis-cutidas nas seções a seguir estão listadas na Tabela 2.1 e ilustradas na Figura 2.2 .
Origem dos Neurônios Pré-ganglionares
Os neurônios pré-ganglionares da divisão parassimpática são originários dos núcleos dos nervos cranianos (NC) III, VII, IX e X ou dos segmentos S2-S4 da medula espinal sacral; assim, a divisão parassimpática é denominada craniossacral . Como na divisão simpática, a origem dos neurônios pré-ganglionares no SNC é consistente com a projeção nos órgãos efetores da periferia. A inervação parassimpática dos músculos dos olhos, por exemplo, é originária do núcleo de Edinger-Westphal no mesencéfalo e trafega até a periferia pelo NC III; a inervação paras-simpática do coração, dos bronquíolos e do trato gas-trintestinal é originária dos núcleos do bulbo e trafega até a periferia pelo NC X (nervo vago); já a inervação parassimpática dos órgãos genitourinários é originária da medula espinal sacral e trafega até a periferia pelos nervos pélvicos.
Localização dos Gânglios Autonômicos
Diferentemente dos gânglios simpáticos, localizados próximos ao SNC, os gânglios do sistema nervoso parassimpático são encontrados no interior dos órgãos efetores, sobre estes órgãos ou nas regiões adjacentes (p. ex., gânglios ciliares, pterigopalatinos, submandibu-lares, óticos).
Comprimento dos Axônios Pré-ganglionares e Pós-ganglionares
O comprimento relativo dos axônios pré-ganglionares e pós-ganglionares da divisão parassimpática é inversa aos comprimentos relativos na divisão simpática. Essa diferença refl ete a localização dos gânglios. Os gânglios parassimpáticos estão localizados nos órgãos efetores ou suas adjacências; os neurônios pré-ganglionares, portanto, possuem axônios longos, enquanto os neurônios pós-gan-glionares apresentam axônios curtos.
Neurotransmissores e Tipos de Receptores
Como na divisão simpática, todos os neurônios pré-ganglionares são colinérgicos e liberam ACh, a qual interage com receptores nicotínicos (N 2 ) nos corpos
celulares dos neurônios pós-ganglionares. A maioria dos neurônios pós-ganglionares da divisão paras-simpática também é colinérgica. Os receptores de ACh nos órgãos efetores são muscarínicos, não nicotínicos. Assim, a ACh liberada por neurônios pré-ganglionares da divisão parassimpática ativa receptores nicotínicos, enquanto a ACh liberada por neurônios pós-gan-glionares da divisão parassimpática ativa receptores muscarínicos. Estes receptores e suas funções são ativados ou inibidos de forma específica por diferentes fármacos ( Tabela 2.2 ).
Varicosidades Colinérgicas Parassimpáticas
Como anteriormente descrito, os neurônios colinérgicos pós-ganglionares parassimpáticos liberam seus neuro-transmissores das varicosidades em tecidos-alvo (p. ex., a musculatura lisa). As varicosidades colinérgicas paras-simpáticas liberam tanto o neurotransmissor clássico (ACh) quanto os neurotransmissores não clássicos (p. ex., peptídeo intestinal vasoativo [VIP], NO). O neurotransmis-sor clássico, ACh , é sintetizado nas varicosidades a partir de colina e acetil coenzima A (acetil CoA) ( Figura 1.17 ) e armazenado em vesículas pequenas e claras . Um grupo separado de grandes vesículas de centro denso contém peptídeos, como o VIP. Por fi m, as varicosidades possuem óxido nítrico sintase e podem sintetizar NO conforme necessário.
TABELA 2.2 Protótipos de Agonistas e Antagonistas de Receptores Autonômicos
Receptor Agonistas Antagonistas
Adrenorreceptores
α 1 Noradrenalina Fenilefrina
Fenoxibenzamina Prazosina
α 2 Clonidina Ioimbina
β 1 Noradrenalina Adrenalina Isoproterenol Dobutamina
Propranolol Metoprolol
β 2 Adrenalina Noradrenalina Isoproterenol Albuterol
Propranolol Butoxamina
Colinorreceptores
Nicotínicos ACh Nicotina
Curare (bloqueia os receptores N 1 neuromusculares)
Hexametônio (bloqueia os receptores N 2 ganglionares)
Muscarínicos ACh Muscarina
Atropina
ACh , Acetilcolina.
C0010.indd 53C0010.indd 53 03/07/18 4:08 PM03/07/18 4:08 PM
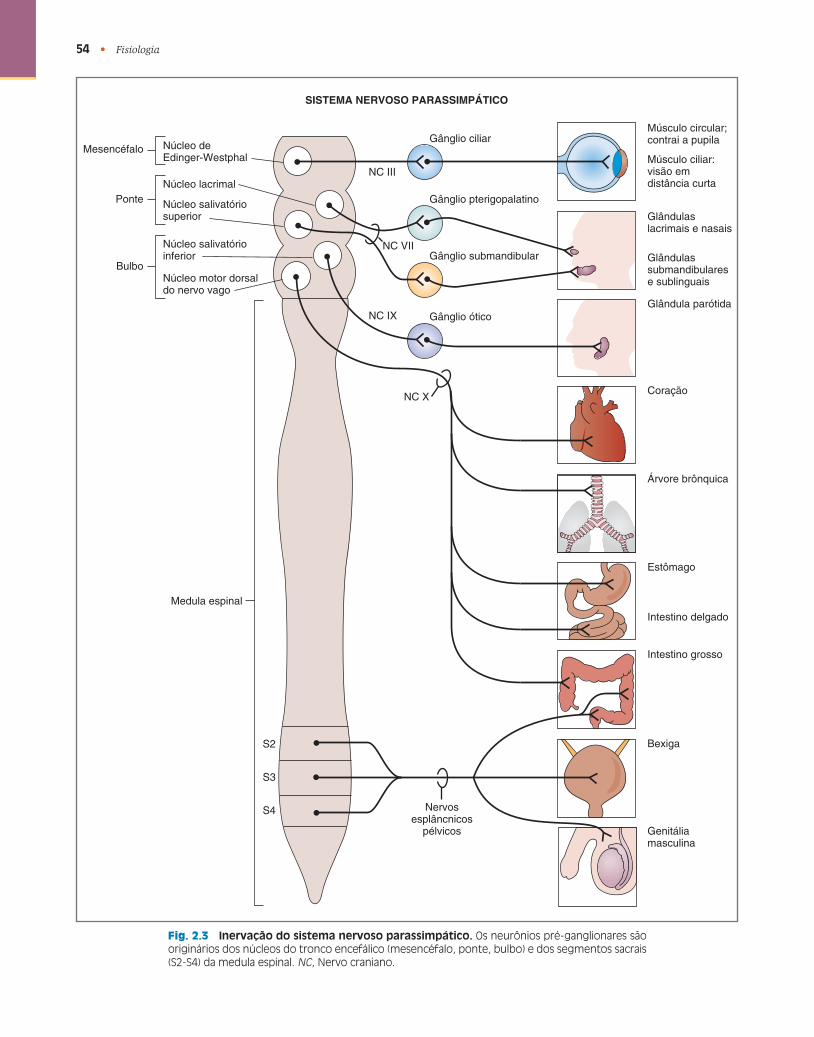
54 • Fisiologia
Coração
Glândula parótida
Glândulassubmandibularese sublinguais
Glândulaslacrimais e nasais
Músculo circular;contrai a pupila
Músculo ciliar:visão emdistância curta
Mesencéfalo
Ponte
Bulbo
Núcleo salivatórioinferior
Núcleo motor dorsaldo nervo vago
Núcleo lacrimal
Núcleo salivatóriosuperior
Núcleo deEdinger-Westphal
SISTEMA NERVOSO PARASSIMPÁTICO
Gânglio óticoNC IX
Medula espinal
Nervosesplâncnicos
pélvicos
S2
S3
S4
Genitáliamasculina
Intestino grosso
Estômago
Árvore brônquica
Intestino delgado
Bexiga
NC X
NC III
NC VIIGânglio submandibular
Gânglio pterigopalatino
Gânglio ciliar
Fig. 2.3 Inervação do sistema nervoso parassimpático. Os neurônios pré-ganglionares são originários dos núcleos do tronco encefálico (mesencéfalo, ponte, bulbo) e dos segmentos sacrais (S2-S4) da medula espinal. NC , Nervo craniano.
C0010.indd 54C0010.indd 54 03/07/18 4:08 PM03/07/18 4:08 PM

2—Sistema Nervoso Autônomo • 55
Quando neurônios colinérgicos pós-ganglionares parassimpáticos são estimulados, a ACh é liberada das varicosidades e se liga a receptores muscarínicos no tecidos-alvo, que determina sua ação fi siológica. Com a estimulação intensa e de alta frequência, as grandes vesículas de centro denso liberam seus peptídeos (p. ex., VIP), que se ligam a receptores nos tecidos-alvo e aumentam as ações da ACh.
Inervação Autônoma dos Sistemas de Órgãos
A Tabela 2.3 serve como uma referência para as informações sobre o controle autônomo da função dos sistemas de órgãos. Esta tabela lista as inervações simpáticas e parassimpáticas dos principais sistemas de órgãos e os tipos de receptores encontrados em seus tecidos. A Tabela 2.3 é ainda mais importante quando a informação mostrada é vista como um conjunto de temas recorrentes, e não como uma lista aleatória de ações e receptores.
Funções Recíprocas – Simpáticas e Parassimpáticas
Muitos órgãos apresentam inervação simpática e parassimpática. Estas inervações operam de forma recíproca ou sinérgica para a produção de respostas coordenadas. O coração, por exemplo, possui inerva-ções simpáticas e parassimpáticas que funcionam de forma recíproca, regulando a frequência cardíaca e a velocidade de condução. As paredes de musculatura lisa do trato gastrintestinal e da bexiga possuem inervação simpática (que produz relaxamento) e parassimpática (que produz contração); os esfíncteres do trato gas-trintestinal e da bexiga também apresentam inervação simpática (que provoca contração) e parassimpática (que causa relaxamento). Os músculos radiais da íris são responsáveis pela dilatação da pupila (midríase) e possuem inervação simpática; o músculo circular da íris é responsável pela contração da pupila (miose) e apresenta inervação parassimpática. Neste exemplo dos músculos oculares, diferentes músculos controlam o tamanho da pupila, mas os efeitos gerais das atividades simpática e parassimpática são recíprocos. Na genitália masculina, a atividade simpática controla a ejaculação, enquanto a atividade parassimpática, ereção; juntas, essas ações são responsáveis pela resposta sexual.
Os três exemplos a seguir ilustram melhor a reciproci-dade e a sinergia das divisões simpática e parassimpática.
NÓ SINOATRIAL
A inervação autônoma do nó sinoatrial (SA) do coração é um excelente exemplo de controle coordenado de função. O nó SA é o marca-passo normal do coração e sua taxa de despolarização controla a frequência cardíaca geral. O nó SA possui inervações simpáticas e parassimpáticas, que funcionam de forma recíproca na modulação da frequência cardíaca. Assim, um aumento da atividade simpática eleva a frequência cardíaca, enquanto um aumento da atividade
parassimpática a diminui. Essas funções recíprocas são ilustradas da seguinte maneira: se a pressão arterial cair, os centros vasomotores do tronco encefálico respondem a essa diminuição e, simultaneamente, aumentam a atividade simpática do nó SA e reduzem a atividade paras-simpática. Cada uma dessas ações, dirigida e coordenada pelo centro vasomotor do tronco encefálico, aumenta a frequência cardíaca. As ações simpática e parassimpática não competem entre si, mas são sinérgicas no aumento da frequência cardíaca (o que ajuda a normalizar a pressão arterial).
BEXIGA
A bexiga é outro exemplo de inervações recíprocas pelas divisões simpática e parassimpática ( Figura 2.4 ). Em adultos, a micção , ou esvaziamento da bexiga, está sob controle voluntário porque o esfíncter externo é compos-to por músculo esquelético. O refl exo da micção em si, porém, é controlado pelo sistema nervoso autônomo. Este refl exo ocorre quando sensores detectam que a bexiga está “cheia”. O músculo detrusor da parede da bexiga e o esfíncter interno do órgão são compostos por músculos lisos, ambos apresentando inervações simpáticas e parassimpáticas. A inervação simpática do músculo detrusor e do esfíncter interno é originária da medula espinal lombar (L1-L3), e a inervação parassimpática tem origem na porção sacral da medula (S2-S4).
Quando a bexiga está sendo enchida por urina, o con-trole simpático é predominante. Esta atividade simpática, por meio de receptores β 2 , relaxa o músculo detrusor e, através de receptores α 1 , contrai o músculo do esfíncter interno. O esfíncter externo é simultaneamente fechado pela ação voluntária treinada. Quando a musculatura da parede está relaxada e os esfíncteres estão fechados, a bexiga pode ser enchida com urina.
Quando a bexiga está cheia , sua distensão é percebida por mecanorreceptores presentes na parede do órgão e os neurônios aferentes transmitem essa informação para a medula espinal e daí para o tronco encefálico. O refl exo de micção é coordenado por centros no mesencéfalo e, agora, o controle parassimpático é predominante. A atividade parassimpática contrai o músculo detrusor (para aumentar a pressão e ejetar a urina) e relaxa o esfíncter interno. Simultaneamente, o esfíncter externo é relaxado pela ação voluntária.
Obviamente, as ações simpáticas e parassimpáticas sobre as estruturas vesicais são opostas, mas coordenadas: as ações simpáticas dominam o enchimento da bexiga e as ações parassimpáticas são responsáveis pelo esvaziamento do órgão.
PUPILA
O tamanho da pupila é reciprocamente controlado por dois músculos da íris: o músculo dilatador (radial) e o músculo constritor (esfíncter) da pupila. O músculo dilatador da pupila é controlado pela inervação simpática,
C0010.indd 55C0010.indd 55 03/07/18 4:08 PM03/07/18 4:08 PM

56 • Fisiologia
TABELA 2.3 Efeitos do Sistema Nervoso Autônomo sobre a Função dos Sistemas de Órgãos
Simpático Parassimpático
Órgão Ação Receptor Ação Receptor
Coração
Nó SA, frequência cardíaca ↑ β 1 ↓ M
Condução do nó AV ↑ β 1 ↓ M
Contratilidade ↑ β 1 ↓ (apenas nos átrios) M
Musculatura Lisa Vascular
Cutânea; esplâncnica Contração α 1
Musculatura esquelética Dilatação β 2
Musculatura esquelética Contração α 1
Endotélio Liberação de EDRF M
Bronquíolos Dilatação β 2 Contração M
Trato Gastrintestinal
Musculatura lisa, paredes Relaxamento α 2 , β 2 Contração M
Musculatura lisa, esfíncteres Contração α 1 Relaxamento M
Secreção de saliva ↑ β 1 ↑ M
Secreção de ácido gástrico ↑ M
Secreção pancreática ↑ M
Bexiga
Parede, músculo detrusor Relaxamento β 2 Contração M
Esfíncter Contração α 1 Relaxamento M
Genitália Masculina Ejaculação α 1 Ereção M
Olho
Músculo radial, íris Dilatação da pupila (midríase) α 1
Músculo circular ou esfíncter, íris
Contração da pupila (miose) M
Músculo ciliar Relaxamento (visão de longe)
β Contração (visão de perto) M
Pele
Glândulas sudoríparas, termorregulação
↑ M a
Glândulas sudoríparas, estresse
↑ α
Músculo piloeretor (arrepios)
Contração α
Glândulas Lacrimais Secreção M
Fígado Gliconeogênese; glicogenólise
α , β 2
Tecido adiposo Lipólise β 1
Rim Secreção de renina β 1
AV , Atrioventricular; EDRF , fator relaxante derivado do endotélio; M , receptor muscarínico; SA , sinoatrial. a Neurônios colinérgicos simpáticos.
C0010.indd 56C0010.indd 56 03/07/18 4:08 PM03/07/18 4:08 PM

2—Sistema Nervoso Autônomo • 57
por meio de receptores α 1 . A ativação desses receptores α 1 causa a contração do músculo radial, provocando a dilatação da pupila, ou midríase. O músculo constritor da pupila é controlado pela inervação parassimpática, por meio de receptores muscarínicos. A ativação desses receptores muscarínicos contrai o músculo do esfíncter, levando à contração da pupila, ou miose.
No refl exo pupilar à luz , por exemplo, a luz atinge a retina e, por meio de uma série de conexões no SNC, ativa neurônios pré-ganglionares parassimpáticos no núcleo de Edinger-Westphal; a ativação dessas fibras parassimpáticas contrai o músculo do esfíncter e a pupila. Na resposta de acomodação , uma imagem retiniana sem nitidez ativa neurônios pré-ganglionares parassimpáticos nos núcleos de Edinger-Westphal, contraindo o músculo do esfíncter e a pupila. Ao mesmo tempo, há contração do músculo ciliar, o que “arredonda” a lente e aumenta seu poder de refração.
Há algumas exceções notáveis na generalização da inervação recíproca. Diversos órgãos apresentam somente inervação simpática : as glândulas sudoríparas, a mus-culatura lisa vascular, os músculos piloeretores da pele, o fígado, o tecido adiposo e o rim.
Coordenação da Função nos Órgãos
A coordenação da função nos sistemas de órgãos, orques-trada pelo sistema nervoso autônomo, é outro tema fi siológico recorrente ( Quadros 2.2 e 2.3 ).
Esse controle é bastante claro, por exemplo, ao con-siderarmos a função da bexiga . Neste órgão, deve haver uma coordenação temporal entre a atividade do músculo detrusor da parede vesical e dos esfíncteres ( Figura 2.4 ). Assim, a atividade simpática é dominante quando a bexiga está se enchendo para relaxar a parede do órgão e, simultaneamente, contrair o esfíncter vesical interno. A bexiga pode ser enchida porque sua parede está relaxada e o esfíncter, fechado. Durante a micção, a atividade paras-simpática é dominante, contraindo a parede da bexiga e, ao mesmo tempo, relaxando o esfíncter.
Uma lógica similar pode ser aplicada ao controle autônomo do trato gastrintestinal : a contração da parede do trato gastrintestinal é acompanhada pelo relaxamento (parassimpático) dos esfíncteres, permitindo que o conteúdo seja propelido para frente. O relaxamento da parede do trato gastrintestinal é acompanhado pela con-tração (simpática) dos esfíncteres; o efeito combinado dessas ações é retardar ou interromper a movimentação de seu conteúdo.
Tipos de Receptores
A Tabela 2.3 traz algumas generalizações sobre os tipos de receptores e seus mecanismos de ação. Essas generalizações são: (1) na divisão parassimpática, os órgãos efetores apresentam receptores muscarínicos; (2) na divisão simpática, existem diversos tipos de receptores em órgãos efetores, incluindo os quatro
β2
α1
M
M
β2
Enchimento da Bexiga Esvaziamento da Bexiga
Músculo
Músculodetrusor
Esfíncterinterno
Esfíncterinterno
Estado
Relaxado
Contraído
Contraído
Mecanismode Controle
Simpático
Simpático
Voluntário
Estado
Contraído
Relaxado
Relaxado
Mecanismode Controle
Parassimpático
Parassimpático
Voluntário
Parassimpático
Simpático
α1
M
M
Medula espinal
L1L2L3
S2S3S4
Fig. 2.4 Controle autonômico do funcionamento da bexiga. Durante o enchimento da bexiga, o controle simpático é predominante e há relaxamento do músculo detrusor e contração do esfíncter interno. Durante a micção, há predominância do controle parassimpático, com contração do músculo detrusor e relaxamento do esfíncter interno. As linhas pontilhadas representam a inervação simpática; as linhas sólidas representam a inervação parassimpática. α 1 , Adrenorreceptor no esfíncter interno; β 2 , adrenorreceptor no músculo detrusor; L1-L3 , segmentos lombares; M, colinorreceptor muscarínico no músculo detrusor e no esfíncter interno; S2.S4 , segmentos sacrais.
C0010.indd 57C0010.indd 57 03/07/18 4:08 PM03/07/18 4:08 PM

58 • Fisiologia
QUADRO 2.2 Fisiologia Clínica: Síndrome de Horner
DESCRIÇÃO DO CASO. Um homem de 66 anos de idade sofreu um derrame do lado direito e apresenta queda da pálpebra direita (ptose), contração da pupila direita (miose) e ausência de sudorese no lado direito da face (anidrose). Seu médico solicita um exame com colírio de cocaína. A solução de cocaína a 10% aplicada no olho esquerdo causa dilatação da pupila (midríase). No olho direito, no entanto, a solução de cocaína não provoca a dilatação da pupila.
EXPLICAÇÃO DO CASO. O homem apresenta um caso clássico de síndrome de Horner secundária ao derrame. Nesta síndrome, há perda da inervação simpática do lado afetado da face. Assim, a perda de inervação simpática para o músculo liso que eleva a pálpebra direita causou ptose desse lado. A perda de inervação simpática para o músculo dilatador da pupila do lado direito provocou a
contração da pupila direita. Por fi m, a perda de inervação simpática para as glândulas sudoríparas do lado direito da face causou anidrose ipsilateral.
Com a instilação do colírio no olho esquerdo (o lado não afetado), a cocaína bloqueou a recaptação de noradrenalina pelos nervos simpáticos que suprem o músculo dilatador da pupila; devido às maiores concen-trações de noradrenalina nessas sinapses adrenérgicas, houve contração do músculo radial da íris, causando a dilatação prolongada da pupila. O uso do colírio de cocaína no olho direito não provocou dilatação da pupila devido à menor quantidade de noradrenalina nessas sinapses.
TRATAMENTO. O tratamento da síndrome de Horner é a resolução da causa subjacente.
QUADRO 2.3 Fisiologia Clínica: Síndrome de Shy-Drager
DESCRIÇÃO DO CASO. Um homem de 58 anos de idade, aparentemente saudável, começa a apresentar sintomas alarmantes. Ocasionalmente impotente, sua impotência progrediu e hoje é contínua. Além disso, ele apresenta enorme urgência de urinar, mas tem difi culdade em produzir um jato de urina. Ele relutou em procurar assistência médica para esses problemas, mas, em uma manhã, desmaiou ao se levantar da cama. Quando marcou uma consulta com o médico, apresentava tontura todas as manhãs e muitos sintomas, inclusive visão dupla, indigestão, diarreia e intolerância ao calor.
O paciente foi encaminhado para um neurologis-ta, que realizou um exame ocular com instilação de metacolina no saco conjuntival; nesse homem, a metacolina provocou miose exagerada (contração da pupila devido à contração do músculo circular da íris). Por causa da natureza global dos sintomas do paciente e dos resultados desse exame ocular, o neurologista o diagnosticou com síndrome de Shy-Drager.
EXPLICAÇÃO DO CASO. A síndrome de Shy-Drager é uma doença rara e progressiva do sistema nervoso autônomo central associada à degeneração dos neurônios pré-ganglionares da coluna celular intermediolateral da medula espinal, dos gânglios autonômicos periféricos e dos centros autonômicos do hipotálamo. Consequen-temente, há grave prejuízo das divisões simpática e parassimpática do sistema nervoso autônomo.
Os sintomas de impotência, difi culdade em urinar e intolerância ao calor são explicados pelos distúrbios simpáticos e parassimpáticos. A resposta sexual masculina é composta por ereção (receptores mus-carínicos parassimpáticos) e ejaculação (receptores α 1 simpáticos). O músculo detrusor da parede da bexiga é composto por musculatura lisa, com inervação simpática (receptores β 2 ) e parassimpática (receptores
muscarínicos); o esfíncter interno da bexiga também é composto por músculo liso, com inervações simpática (receptores α 1 ) e parassimpática (receptores muscaríni-cos). As glândulas sudoríparas termorreguladoras estão sob controle simpático.
A resposta ocular exagerada à metacolina (um agonis-ta muscarínico colinérgico) talvez seja surpreendente, já que o sistema nervoso parassimpático do paciente está prejudicado; no entanto, os resultados do exame fazem sentido, uma vez que a perda da inervação parassimpática do músculo circular da íris provoca a regulação positiva dos receptores colinérgicos e, assim, aumenta a resposta a um agonista colinérgico exógeno (hipersensibilidade por desnervação).
O paciente apresentava hipotensão ortostática, ou seja, diminuição da pressão arterial ao ficar em pé. Ao se levantar, o sangue se acumula nos membros inferiores, o que reduz a pressão arterial. Em pes-soas normais, a queda da pressão arterial evoca o refl exo barorreceptor, com participação dos sistemas nervosos simpático e parassimpático; juntas, essas res-postas autonômicas normalizam a pressão arterial. O mecanismo barorreceptor do paciente foi extremamente prejudicado e sua pressão arterial não pôde ser corrigida por refl exos autonômicos, por isso ele sentia tontura e até desmaiou.
TRATAMENTO. O paciente foi instruído a manter a cabeça elevada durante o sono (para reduzir os efeitos ortostáticos sobre a pressão arterial ao se levantar), usar meias de compressão para impedir o acúmulo de sangue nas pernas e tomar um análogo da aldos-terona para aumentar o volume sanguíneo. Cada uma dessas medidas foi uma tentativa de melhorar a tontura e os desmaios ao se levantar. Os tratamentos foram paliativos; não há cura para essa doença degenerativa e fatal.
C0010.indd 58C0010.indd 58 03/07/18 4:08 PM03/07/18 4:08 PM
2—Sistema Nervoso Autônomo • 59
adrenorreceptores ( α 1 , α 2 , β 1 e β 2 ); nos tecidos com inervação simpática colinérgica, há receptores mus-carínicos; (3) entre os adrenorreceptores simpáticos, o tipo de receptor está relacionado com a função. Os receptores α 1 contraem músculos lisos, como a mus-culatura lisa vascular, os esfíncteres gastrintestinais e vesicais, os músculos piloeretores e o músculo radial da íris. Os receptores β 1 participam de funções metabólicas, como a gliconeogênese, a lipólise e a secreção de renina, assim como de todas as funções cardíacas. Os receptores β 2 relaxam a musculatura lisa dos bronquíolos, da parede da bexiga e da parede do trato gastrintestinal.
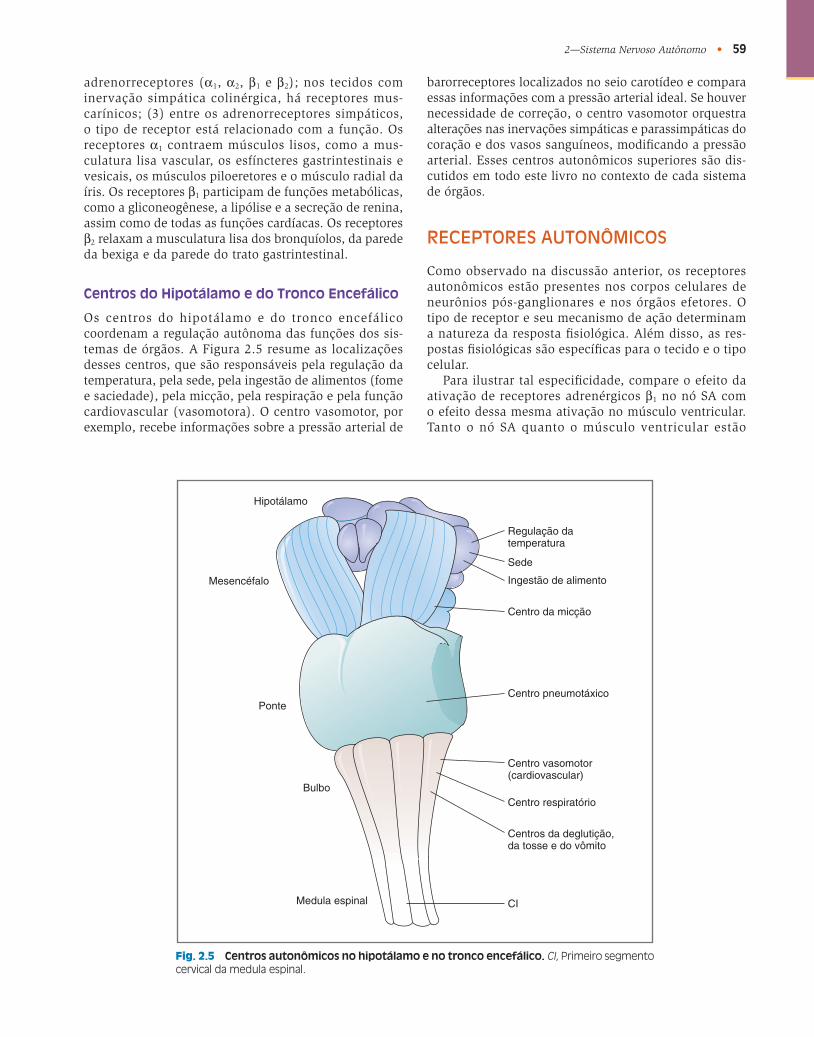
Centros do Hipotálamo e do Tronco Encefálico
Os centros do hipotálamo e do tronco encefálico coordenam a regulação autônoma das funções dos sis-temas de órgãos. A Figura 2.5 resume as localizações desses centros, que são responsáveis pela regulação da temperatura, pela sede, pela ingestão de alimentos (fome e saciedade), pela micção, pela respiração e pela função cardiovascular (vasomotora). O centro vasomotor, por exemplo, recebe informações sobre a pressão arterial de
barorreceptores localizados no seio carotídeo e compara essas informações com a pressão arterial ideal. Se houver necessidade de correção, o centro vasomotor orquestra alterações nas inervações simpáticas e parassimpáticas do coração e dos vasos sanguíneos, modifi cando a pressão arterial. Esses centros autonômicos superiores são dis-cutidos em todo este livro no contexto de cada sistema de órgãos.
RECEPTORES AUTONÔMICOS
Como observado na discussão anterior, os receptores autonômicos estão presentes nos corpos celulares de neurônios pós-ganglionares e nos órgãos efetores. O tipo de receptor e seu mecanismo de ação determinam a natureza da resposta fi siológica. Além disso, as res-postas fi siológicas são específi cas para o tecido e o tipo celular.
Para ilustrar tal especifi cidade, compare o efeito da ativação de receptores adrenérgicos β 1 no nó SA com o efeito dessa mesma ativação no músculo ventricular. Tanto o nó SA quanto o músculo ventricular estão
Centro vasomotor(cardiovascular)
Centro pneumotáxico
Centro da micção
Ingestão de alimento
Sede
Regulação datemperatura
Centro respiratório
Centros da deglutição,da tosse e do vômito
CI
Mesencéfalo
Hipotálamo
Ponte
Bulbo
Medula espinal
Fig. 2.5 Centros autonômicos no hipotálamo e no tronco encefálico. CI , Primeiro segmento cervical da medula espinal.
C0010.indd 59C0010.indd 59 03/07/18 4:08 PM03/07/18 4:08 PM

60 • Fisiologia
localizados no coração, e seus receptores adrenérgicos e mecanismos de ação são os mesmos. As ações fi siológicas resultantes, porém, são completamente diferentes. O receptor β 1 do nó SA está associado a mecanismos que aumentam a frequência espontânea de despolarização e a frequência cardíaca; a ligação de um agonista, como a noradrenalina, a este receptor β 1 aumenta a frequência cardíaca. O receptor β 1 do músculo ventricular está associado a mecanismos que aumentam a concentração intracelular de Ca 2+ e a contratilidade; a ligação de um agonista, como a noradrenalina, a este receptor β 1 aumenta a contratilidade, mas não exerce efeito direto sobre a frequência cardíaca.
O tipo de receptor também indica qual agonista ou antagonista farmacológico o ativa ou bloqueia. Os efeitos desses fármacos podem ser facilmente previstos pela com-preensão das respostas fi siológicas normais . Os fármacos que são agonistas β 1 , por exemplo, devem aumentar a frequência cardíaca e a contratilidade, enquanto os antagonistas β 1 devem diminuir a frequência cardíaca e a contratilidade.
A Tabela 2.4 resume as características dos receptores adrenérgicos e colinérgicos, seus tecidos-alvo e seus meca-nismos de ação. A Tabela 2.2 é disposta de forma similar, por tipo de receptor, e lista os fármacos prototípicos que ativam ( agonistas ) ou bloqueiam ( antagonistas ) os recep-tores. Juntas, essas duas tabelas devem ser usadas como referência durante a discussão sobre os mecanismos de ação. Esses mecanismos, com participação das proteínas ligantes do trifosfato de guanosina (GTP) (proteínas G), da adenilil ciclase e do 1,4,5-trifosfato de inositol (IP 3 ), são também discutidos, no Capítulo 9 , no contexto da ação hormonal.
Proteínas G
Os receptores autonômicos são associados a proteínas ligantes de GTP (proteínas G) e, assim, são denomina-dos receptores ligados à proteína G . Esses receptores, inclusive aqueles do sistema nervoso autônomo, são compostos por uma única cadeia polipeptídica que atravessa a membrana celular sete vezes e, por isso, são conhecidos como receptores proteicos transmem-brânicos de sete passagens. O ligante (p. ex., ACh, noradrenalina) interage com o domínio extracelular de seu receptor ligado à proteína G. O domínio intracelular do receptor se liga (é “associado”) à proteína G.
Essas proteínas G são heterotriméricas . Em outras palavras, apresentam três diferentes subunidades: α , β e γ . A subunidade α se liga ao difosfato de guano-sina (GDP) ou ao GTP. Quando o GDP está ligado, a subunidade α é inativa; quando o GTP está ligado, a subunidade α é ativa. Assim, a atividade da proteína G reside em sua subunidade α e esta proteína alterna entre os estados ativo e inativo de acordo com sua ligação ao GDP ou GTP. Quando a proteína G libera o GDP e se liga ao GTP, por exemplo, passa do estado
inativo para o estado ativo; quando o GTP é novamente convertido em GDP por meio da atividade intrínseca da GTPase da proteína G, há mudança do estado ativo para o inativo.
As proteínas G acoplam receptores autonômicos ligados à proteína G a enzimas que executam ações fisiológicas. Estas enzimas são a adenilil ciclase e a fosfolipase C, que, quando ativadas, dão origem a um segundo mensageiro (monofosfato cíclico de adenosina [AMPc] ou IP 3 , respectivamente). O segundo mensageiro amplifi ca, então, a mensagem e executa a ação fi siológica final. Em alguns casos (p. ex., em certos receptores muscarínicos), a proteína G altera diretamente a função de um canal iônico sem a mediação de um segundo mensageiro.
Adrenorreceptores
Os adrenorreceptores são encontrados em tecidos-alvo do sistema nervoso simpático e são ativados pelas cate-colaminas noradrenalina e adrenalina. A noradrenalina é liberada por neurônios pós-ganglionares do sistema nervoso simpático. A adrenalina é secretada pela medula da adrenal e chega aos tecidos-alvo por meio da circulação. Os adrenorreceptores são divididos em dois tipos, α e β , que ainda se dividem em α 1 , α 2 , β 1 e β 2 . Cada um desses tipos de receptores possui um mecanismo de ação diferente (exceto os receptores β 1 e β 2 , que têm o mesmo mecanismo de ação), provocando efeitos fi siológicos dis-tintos ( Tabelas 2.3 e 2.4 ).
Receptores α 1
Os receptores α 1 são encontrados na musculatura lisa vascular (da pele, nos músculos esqueléticos e na região esplâncnica), nos esfíncteres do trato gastrintes-tinal e da bexiga, além de no músculo radial da íris. A ativação dos receptores α 1 leva à contração de cada um desses tecidos. O mecanismo de ação envolve uma proteína G denominada G q e a ativação da fosfolipase C , ilustrada na Figura 2.6 . Os números circulados na figura correspondem às etapas discutidas, da seguinte maneira:
1. O receptor α 1 está inserido na membrana celular, onde é acoplado, via proteína G q , à fosfolipase C. No estado inativo, a subunidade α q da proteína G q heterotrimérica está ligada ao GDP.
2. Quando um agonista, como a noradrenalina, se liga ao receptor α 1 (etapa 1), a unidade α q da proteína G q sofre uma alteração conformacional. Esta alteração conformacional tem dois efeitos (etapa 2): a liberação do GDP da unidade α q e sua substituição por GTP e a liberação da subunidade α q (ligada ao GTP) do restante da proteína G q .
3. O complexo α q -GTP desloca-se pela membrana celular e se liga à fosfolipase C, ativando-a (etapa 3). A atividade GTPase intrínseca, então, converte o GTP em GDP e
C0010.indd 60C0010.indd 60 03/07/18 4:08 PM03/07/18 4:08 PM

2—Sistema Nervoso Autônomo • 61
a subunidade α q retorna ao estado inativo (não mos-trado).
4. A fosfolipase C ativada catalisa a liberação de diacilglicerol e IP 3 do 4,5-difosfato de fosfatidilinositol (etapa 4). O IP 3 gerado provoca a liberação de Ca 2+ de estoques intracelulares nos retículos endoplasmáticos ou sarcoplasmáticos, aumentando a concentração intracelular de Ca 2+ (etapa 5). Juntos, o Ca 2+ e o diacilglicerol ativam a proteína quinase C (etapa 6), que fosforila proteínas. Estas proteínas fosforiladas executam as ações fi siológicas fi nais (etapa 7), como a contração de músculos lisos.
Receptores α 2
Os receptores α 2 são inibidores, localizados pré e pós-sinapticamente, e são menos comuns do que os recep-tores α 1 . Esses receptores são encontrados em terminais nervosos adrenérgicos e colinérgicos pré-sinápticos e no trato gastrintestinal. Existem duas formas de receptores α 2 , os autorreceptores e os heterorreceptores.
Os receptores α 2 presentes nos terminais nervosos pós-ganglionares simpáticos são denominados autor-receptores . Nessa função, a ativação de receptores α 2 pela noradrenalina liberada por terminais nervosos pré-sinápticos inibe a maior liberação da molécula
pelas mesmas estruturas; esta retroalimentação negativa conserva a noradrenalina em estados de alta estimulação do sistema nervoso simpático. É interessante notar que a medula da adrenal não pos-sui receptores α 2 , portanto não está sujeita à inibição por retroalimentação e, consequentemente, a medula da adrenal pode esgotar as catecolaminas durante períodos de estresse prolongado.
Os receptores α 2 presentes em terminais nervosos pós-ganglionares do trato gastrintestinal são denominados heterorreceptores . A noradrenalina é liberada por fi bras pós-ganglionares simpáticas que fazem sinapse com essas fibras pós-ganglionares parassimpáticas. Quando ativados pela noradrenalina, os receptores α 2 inibem a liberação de ACh pelos terminais nervosos pós-ganglionares parassimpáticos. Dessa forma, o sis-tema nervoso simpático inibe, indiretamente, a função gastrintestinal (ou seja, por meio da inibição da atividade parassimpática).
O mecanismo de ação desses receptores envolve a inibição da adenilil ciclase , descrita pelas seguintes etapas:
1. O agonista (p. ex., noradrenalina) se liga ao receptor α 2 , que está acoplado à adenilil ciclase por uma proteína G inibidora, G i .
TABELA 2.4 Localização e Mecanismo de Ação dos Receptores Autonômicos
Receptor Tecido-Alvo Mecanismo de Ação
Adrenorreceptores
α 1 Musculatura lisa vascular, (cutânea, renal e esplâncnica)
Trato gastrintestinal, esfíncteres Bexiga, esfíncter Músculo radial, íris
IP 3 , ↑ intracelular de [Ca 2+ ]
α 2 Trato gastrintestinal, parede Neurônios adrenérgicos pré-sinápticos
Inibição de adenilil ciclase, ↓ AMPc
β 1 Coração Glândulas salivares Tecido adiposo Rim
Estimulação de adenilil ciclase, ↑ AMPc
β 2 Musculatura lisa vascular de músculos esqueléticos Trato gastrintestinal, parede Bexiga, parede Bronquíolos
Estimulação de adenilil ciclase, ↑ AMPc
Colinorreceptores
Nicotínicos Neurônios pós-ganglionares, SNS e SNP (N 2 ) Medula da adrenal (N 2 )
Abertura dos canais de Na+ e K+ → despolarização
Muscarínicos Todos os órgãos efetores, SNP Glândulas sudoríparas, SNS
IP 3 , ↑ intracelular de [Ca 2+ ] (M 1 , M 3 , M 5 ) ↓ adenilil ciclase, ↓ AMPc (M 2 , M 4 )
AMPc , Monofosfato cíclico de adenosina; IP 3 , 1,4,5-trifosfato de inositol; SNP , sistema nervoso parassimpático; SNS , sistema nervoso simpático.
C0010.indd 61C0010.indd 61 03/07/18 4:08 PM03/07/18 4:08 PM

62 • Fisiologia
2. Com a noradrenalina ligada, a proteína G i libera o GDP e se liga ao GTP, e a subunidade α i se dissocia do complexo da proteína G.
3. A subunidade α i , então, desloca-se pela membrana, liga-se à adenilil ciclase e a inibe. Por causa disso, os níveis de AMPc diminuem, produzindo a resposta fi siológica fi nal.
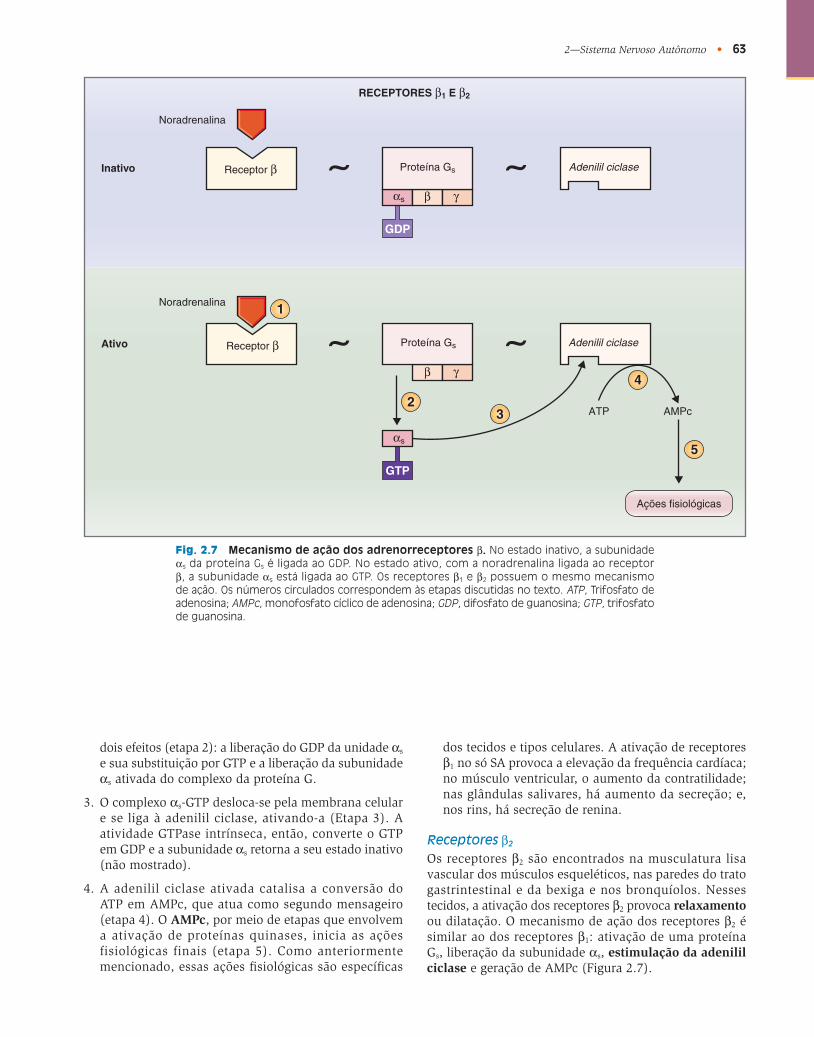
Receptores β 1
Os receptores β 1 são proeminentes no coração, sendo encontrados no nó SA, no nó atrioventricular (AV) e no músculo ventricular. Nesses tecidos, a ativação dos receptores β 1 aumenta a frequência cardíaca no nó SA, a velocidade de condução no nó AV e a contratilidade do músculo ventricular, respectivamente. Os receptores
também estão localizados nas glândulas salivares, no tecido adiposo e nos rins (onde promovem a secreção de renina). O mecanismo de ação dos receptores β 1 envolve uma proteína G s e a ativação da adenilil ciclase . Esta ação é ilustrada na Figura 2.7 e composta pelas seguintes etapas, que correspondem aos números circulados na fi gura:
1. Da mesma forma que outros receptores autonômicos, os receptores β 1 estão inseridos na membrana celular. Esses receptores são acoplados, por uma proteína G s , à adenilil ciclase. No estado inativo, a subunidade α s da proteína G s está ligada ao GDP.
2. Quando um agonista, como a noradrenalina, se liga ao receptor β 1 (etapa 1), a unidade α s sofre uma alteração conformacional. Esta alteração conformacional tem
GDP
GTP
˜1
23
7
6 5
4
Noradrenalina
Inativo
αq β γ˜Receptor α1Proteína Gq ˜
Fosfolipase C
Receptor α1Proteína Gq Fosfolipase C
Noradrenalina
Ativo
αq
β γ˜Diacilglicerol
proteína quinase C
PIP2
Ca2+ liberadodo RE ou RS
RECEPTORES α1
IP3
Ações fisiológicas
Fig. 2.6 Mecanismo de ação dos adrenorreceptores α 1 . No estado inativo, a subunidade α q da proteína G q é ligada ao GDP. No estado ativo, com a noradrenalina ligada ao receptor α 1 , a subunidade α q está ligada ao GTP. α q , β e γ são subunidades da proteína G q . Os números circulados correspondem às etapas discutidas no texto. RE , Retículo endoplasmático; GDP , difosfato de guanosina; G q , proteína G; GTP , trifosfato de guanosina; IP 3 , 1,4,5-trifosfato de inositol; PIP 2 , 4,5-difosfato de fosfatidilinositol; RS ; retículo sarcoplasmático.
C0010.indd 62C0010.indd 62 03/07/18 4:08 PM03/07/18 4:08 PM
2—Sistema Nervoso Autônomo • 63
dois efeitos (etapa 2): a liberação do GDP da unidade α s e sua substituição por GTP e a liberação da subunidade α s ativada do complexo da proteína G.
3. O complexo α s -GTP desloca-se pela membrana celular e se liga à adenilil ciclase, ativando-a (Etapa 3). A atividade GTPase intrínseca, então, converte o GTP em GDP e a subunidade α s retorna a seu estado inativo (não mostrado).
4. A adenilil ciclase ativada catalisa a conversão do ATP em AMPc, que atua como segundo mensageiro (etapa 4). O AMPc , por meio de etapas que envolvem a ativação de proteínas quinases, inicia as ações fisiológicas finais (etapa 5). Como anteriormente mencionado, essas ações fi siológicas são específi cas
dos tecidos e tipos celulares. A ativação de receptores β 1 no só SA provoca a elevação da frequência cardíaca; no músculo ventricular, o aumento da contratilidade; nas glândulas salivares, há aumento da secreção; e, nos rins, há secreção de renina.
Receptores β 2
Os receptores β 2 são encontrados na musculatura lisa vascular dos músculos esqueléticos, nas paredes do trato gastrintestinal e da bexiga e nos bronquíolos. Nesses tecidos, a ativação dos receptores β 2 provoca relaxamento ou dilatação. O mecanismo de ação dos receptores β 2 é similar ao dos receptores β 1 : ativação de uma proteína G s , liberação da subunidade α s , estimulação da adenilil ciclase e geração de AMPc ( Figura 2.7 ).
GDP
GTP
Inativo
1
23
5
4
RECEPTORES β1 E β2
Noradrenalina
αs β γ
Receptor β
Receptor β
Proteína Gs Adenilil ciclase
Noradrenalina
Ativo
αs
β γ
Proteína Gs
AMPcATP
Adenilil ciclase
Ações fisiológicas
˜ ˜
˜ ˜
Fig. 2.7 Mecanismo de ação dos adrenorreceptores β . No estado inativo, a subunidade α s da proteína G s é ligada ao GDP. No estado ativo, com a noradrenalina ligada ao receptor β , a subunidade α s está ligada ao GTP. Os receptores β 1 e β 2 possuem o mesmo mecanismo de ação. Os números circulados correspondem às etapas discutidas no texto. ATP , Trifosfato de adenosina; AMPc , monofosfato cíclico de adenosina; GDP , difosfato de guanosina; GTP , trifosfato de guanosina.
C0010.indd 63C0010.indd 63 03/07/18 4:08 PM03/07/18 4:08 PM

64 • Fisiologia
Respostas dos Adrenorreceptores à Noradrenalina e à Adrenalina
Existem diferenças significativas nas respostas dos adrenorreceptores α 1 , β 1 e β 2 às catecolaminas nora-drenalina e adrenalina. Estas diferenças são explicadas a seguir, lembrando que a noradrenalina é a catecolamina liberada pelas fi bras nervosas adrenérgicas simpáticas pós-ganglionares, enquanto a adrenalina é a principal catecolamina liberada pela medula da adrenal: (1) a noradrenalina e a adrenalina possuem quase a mes-ma potência em receptores α 1 , sendo esta última um pouco mais potente. No entanto, em comparação com os receptores β , os receptores α 1 são relativamente insensíveis às catecolaminas. A ativação de receptores α 1 requer concentrações maiores de catecolaminas do que a ativação de receptores β . Fisiologicamente, essas altas concentrações são localmente atingidas quando a noradrenalina é liberada por fi bras nervosas simpáticas pós-ganglionares, mas não após a liberação de catecolaminas pela medula da adrenal. A quantidade de adrenalina (e noradrenalina) liberada pela medula da adrenal na resposta de luta ou fuga, por exemplo, é insufi ciente para a ativação de receptores α 1 ; (2) a noradrenalina e a adrenalina são equipotentes em receptores β 1 . Como já mencionado, concentrações muito menores de catecolaminas ativam os receptores β 1 em comparação com os receptores α 1 . Assim, a nora-drenalina liberada pelas fi bras nervosas simpáticas ou a adrenalina liberada pela medula da adrenal ativa os receptores β 1 ; (3) os receptores β 2 são preferencialmente ativados pela adrenalina. Assim, a adrenalina liberada pela medula da adrenal deve ativar os receptores β 2 , mas não a noradrenalina liberada pelas terminações nervosas simpáticas.
Colinorreceptores
Existem dois tipos de colinorreceptores: nicotínicos e muscarínicos. Os receptores nicotínicos são encontrados em todos os gânglios autonômicos e nas células cromafi ns da medula da adrenal. Os receptores muscarínicos são encontrados em todos os órgãos efetores da divisão parassimpática e em alguns órgãos efetores da divisão simpática.
Receptores Nicotínicos
Os receptores nicotínicos são encontrados em diversos locais importantes: na placa motora dos músculos esqueléticos, em todos os neurônios pós-ganglionares dos sistemas nervosos simpático e parassimpático e nas células cromafi ns da medula da adrenal. A ACh é seu agonista natural e é liberada pelos motoneurônios e por todos os neurônios pré-ganglionares.
Não se sabe se o receptor nicotínico na placa motora terminal é idêntico ao receptor nicotínico dos gânglios autonômicos. Essa questão pode ser respondida pela análise das ações dos fármacos que atuam como agonis-
tas ou antagonistas nesses receptores. Os receptores nicotínicos dos dois locais certamente são similares: ambos são ativados pelos agonistas ACh, nicotina e carbacol e antagonizados pelo curare ( Tabela 2.2 ). Outro antagonista do receptor nicotínico, no entanto, o hexametônio , bloqueia-o nos gânglios, mas não na placa motora. Assim, pode-se concluir que os receptores desses dois locais são similares, mas não idênticos; os receptores nicotínicos da placa motora da musculatura esquelética são denominados N 1 , e os receptores nicotínicos dos gânglios autonômicos são chamados N 2 . Segundo essa distinção farmacológica, substâncias como o hexame-tônio são agentes bloqueadores ganglionares, mas não bloqueadores neuromusculares.
É possível chegar a uma segunda conclusão sobre os agentes bloqueadores ganglionares , como o hexametônio. Esses agentes devem inibir receptores nicotínicos em gânglios simpáticos e parassimpáticos, e, assim, produzir efeitos disseminados sobre a função autonômica. Para prever as ações de agentes bloqueadores ganglionares em um determinado órgão, porém, é necessário saber se o controle dominante desse órgão é simpático ou parassimpático. A mus-culatura lisa vascular , por exemplo, apresenta apenas inervação simpática, que causa vasoconstrição; dessa forma, os agentes bloqueadores ganglionares relaxam a musculatura lisa vascular e provocam vasodilatação. (Devido a essa propriedade, os agentes bloqueadores ganglionares podem ser usados no tratamento da hipertensão.) Por outro lado, a função sexual mas-culina é dramaticamente prejudicada por agentes bloqueadores ganglionares, já que a resposta sexual do homem apresenta componentes simpáticos (ejaculação) e parassimpáticos (ereção).
O mecanismo de ação dos receptores nicotínicos, seja na placa motora ou nos gânglios, é baseado no fato de que esse receptor de ACh é também um canal para Na + e K + . Quando o receptor nicotínico é ativado pela ACh, o canal se abre, permitindo a passagem simultânea de Na + e K + conforme seus respectivos gradientes ele-troquímicos.
A Figura 2.8 ilustra a função dos receptores/canais nicotínicos em dois estados: fechado e aberto. O receptor nicotínico é uma proteína integral da membrana celular composta por cinco subunidades: duas α , uma β , uma δ e uma γ . Estas cinco subunidades formam um funil ao redor da abertura central. Quando não há ACh ligada , a abertura do canal está fechada. Quando a ACh está ligada a cada uma das duas subunidades α , há uma mudança conformacional em todas as subunidades, abrindo a área central do canal. Com a abertura do centro do canal, o Na + e o K + fl uem de acordo com seus respectivos gradientes eletroquímicos (Na + para dentro da célula e K + para fora da célula) e cada íon tenta levar o potencial de membrana a seu potencial de equilíbrio. O potencial de membrana resultante é a média entre os potenciais de equilíbrio dos dois íons,
C0010.indd 64C0010.indd 64 03/07/18 4:08 PM03/07/18 4:08 PM

2—Sistema Nervoso Autônomo • 65
aproximadamente 0 milivolt, o que corresponde a um estado despolarizado.
Receptores Muscarínicos
Os receptores muscarínicos estão localizados em todos os órgãos efetores do sistema nervoso parassimpático: no coração, no trato gastrintestinal, nos bronquíolos, na bexiga e nos órgãos sexuais masculinos. Esses receptores também são encontrados em certos órgãos efetores do sistema nervoso simpático, especifi camente nas glândulas sudoríparas.
Alguns receptores muscarínicos (p. ex., M 1 , M 3 e M 5 ) apresentam o mesmo mecanismo de ação que os adrenorreceptores α 1 ( Figura 2.6 ). Nesses casos, a ligação do agonista (ACh) ao receptor muscarínico provoca a dissociação da subunidade α da proteína G, a ativação da fosfolipase C e a geração de IP 3 e diacil-glicerol. O IP 3 libera o Ca 2+ armazenado e o aumento da concentração intracelular deste íon associado à presença
de diacilglicerol produz as ações fi siológicas específi cas ao tecido.
Outros receptores muscarínicos (p. ex., M 4 ) inibem a adenilil ciclase e reduzem os níveis intracelulares de AMPc.
Outro tipo de receptores muscarínicos (M 2 ) altera os processos fisiológicos por meio da ação direta da proteína G . Nesses casos, não há participação de segundos mensageiros. Os receptores muscarínicos do nó SA do coração, por exemplo, quando ativados pela ACh, ativam a proteína G i e liberam a subunidade α i , que se liga diretamente aos canais de K + do nó SA. Quando as subunidades α i se ligam aos canais de K + , estes se abrem e diminuem a taxa de despolarização do nó SA e a frequência cardíaca. Nesse mecanismo, não há estimulação ou inibição de adenilil ciclase ou fosfolipase C, nem a participação de quaisquer segundos mensageiros; em vez disso, a proteína G i age diretamente sobre o canal iônico ( Quadro 2.4 )
Líquidoextracelular
ACh ACh
α αγ
βδ
γα α
Canal aberto
K+
Na+
Canal fechadoLíquidointracelular
ACh ACh
RECEPTOR NICOTÍNICO
Fig. 2.8 Mecanismo de ação dos colinorreceptores nicotínicos. O receptor nicotínico para acetilcolina (ACh) é um canal iônico para Na + e K + . O receptor possui cinco subunidades: duas α , uma β , uma δ e uma γ .
C0010.indd 65C0010.indd 65 03/07/18 4:08 PM03/07/18 4:08 PM

66 • Fisiologia
RESUMO
� O sistema nervoso autônomo é composto por duas divisões principais, simpática e parassimpática, que operam de forma coordenada para regular as funções involuntárias. A divisão simpática é toracolombar e originária da medula espinal. A divisão parassimpática é craniossacral, sendo originária do tronco encefálico e da porção sacral da medula espinal.
� As vias eferentes do sistema nervoso autônomo são compostas por um neurônio pré-ganglionar e um neurônio pós-ganglionar, que fazem sinapse em um gânglio autônomo. Os axônios dos neurônios pós-gan-glionares, então, trafegam até a periferia para inervar os órgãos efetores. A medula da adrenal é um gânglio especializado da divisão simpática; quando estimulada, secreta catecolaminas na circulação.
� De modo geral, as inervações simpáticas e parassim-páticas dos órgãos ou sistemas de órgãos têm efeitos recíprocos. Estes efeitos são coordenados por centros autonômicos no tronco encefálico e no hipotálamo. Os centros autonômicos do tronco encefálico controlam a frequência cardíaca, por exemplo, ao modularem a atividade simpática e parassimpática no nó SA.
� Os receptores para neurotransmissores do sistema ner-voso autônomo são adrenérgicos (adrenorreceptores) ou colinérgicos (colinorreceptores). Os adrenorrecep-tores são ativados pelas catecolaminas noradrenalina e adrenalina; os colinorreceptores, por ACh.
� Os receptores autonômicos são acoplados a proteínas G, que podem ser estimuladoras (G s ) ou inibidoras (G i ). As proteínas G, por sua vez, ativam ou inibem enzimas que são responsáveis pelas ações fi siológicas fi nais.
� O mecanismo de ação dos adrenorreceptores pode ser explicado da seguinte maneira: os receptores α 1 atuam mediante a ativação da fosfolipase C e a geração de IP 3 . Os receptores β 1 e β 2 agem por meio da ativação de adenilil ciclase e a geração de AMPc; os receptores α 2 atuam por intermédio da inibição da adenilil ciclase.
� O mecanismo de ação dos colinorreceptores pode ser explicado da seguinte forma: os receptores nicotínicos atuam como canais iônicos para Na + e K + . Muitos receptores muscarínicos apresentam o mesmo mecanis-mo de ação que os receptores α 1 ; em alguns receptores muscarínicos, há ação direta da proteína G no mecanis-mo fi siológico.
QUADRO 2.4 Fisiologia Clínica: Tratamento do Enjoo por Movimento (Cinetose) com um Antagonista de Receptor Muscarínico
DESCRIÇÃO DO CASO. Uma mulher que planeja realizar um cruzeiro de 10 dias pede que seu médico receite algo que previna o enjoo por movimento. O médico prescreve escopolamina, uma substância similar à atropina, e recomenda à paciente que ela tome a medicação durante toda a viagem. Tomando o fármaco, a mulher não tem náuseas ou vomita, como esperado, mas apresenta secura na boca, dilatação das pupilas (midríase), aumento da frequência cardíaca (taquicardia) e difi culdade de micção.
EXPLICAÇÃO DO CASO. A escopolamina, como a atropina, bloqueia os receptores colinérgicos mus-carínicos nos tecidos-alvo. Este fármaco pode, de fato, ser usado com sucesso no tratamento do enjoo por movimento, cuja etiologia envolve receptores muscarínicos do sistema vestibular. As reações adversas que a paciente apresentou durante o tratamento com escopolamina podem ser explicadas pela fisiologia dos receptores muscarínicos nos tecidos-alvo.
A ativação dos receptores muscarínicos aumenta a salivação, contrai as pupilas, diminui a frequência cardíaca (bradicardia) e contrai a parede da bexiga durante a micção ( Tabela 2.2 ), portanto a inibição dos receptores muscarínicos por meio da adminis-tração de escopolamina deve provocar sintomas relacionados com diminuição da salivação (boca seca), dilatação das pupilas (devido à livre infl uência do sistema nervoso simpático sobre o músculo radial), aumento da frequência cardíaca e micção lenta (causada pela perda do tônus contrátil da parede da bexiga).
TRATAMENTO. Interrupção da administração de escopolamina.
Desafi e-se
Responda a cada pergunta com uma palavra, expressão, frase ou solução numérica. Se a questão vier acompanhada por uma lista de possíveis respostas, uma, mais de uma ou nenhuma das escolhas pode estar correta. As respostas corretas são mostradas no fi nal do livro.
1. Qual(is) da(s) seguinte(s) ação(ões) é(são) mediada(s) por receptores β 2 : aumento da frequência cardíaca, contração dos esfíncteres gastrintestinais, contração da musculatura lisa vascular, dilatação das vias aéreas, relaxamento da parede da bexiga?
2. Uma mulher que está tomando atropina para tratamento de uma doença gastrintestinal nota que suas pupilas estão dilatadas. Isto ocorreu porque a atropina bloqueia os receptores _______________ do músculo _________________ da íris.
3. Qual(is) da(s) seguinte(s) é(são) característica(s) do sistema nervoso
C0010.indd 66C0010.indd 66 03/07/18 4:08 PM03/07/18 4:08 PM

2—Sistema Nervoso Autônomo • 67
parassimpático, mas não do sistema nervoso simpático: gânglios em tecidos-alvo ou regiões próximas, receptores nicotínicos em neurônios pós-ganglionares, receptores muscarínicos em alguns tecidos-alvo, receptores β 1 em alguns tecidos-alvo, neurônios pré-ganglionares colinérgicos?
4. O propranolol diminui a frequência cardíaca por ________________ os receptores ________________ no nó sinoatrial do coração.
5. Qual(is) da(s) seguinte(s) ação(ões) é(são) mediada(s) pelo mecanismo da adenilil ciclase: efeito do sistema nervoso parassimpático para aumento da secreção ácida no estômago, efeito da adrenalina para aumento da contratilidade cardíaca, efeito da adrenalina para aumento da frequência cardíaca, efeito da acetilcolina para redução da frequência cardíaca, efeito da acetilcolina para contração das vias aéreas, contração da musculatura lisa vascular nos vasos sanguíneos esplâncnicos?
6. Qual enzima é responsável pela maior síntese de adrenalina do que de noradrenalina pela medula da adrenal?
7. Um homem teve um feocromocitoma que causou uma grave elevação em sua pressão arterial. Antes da cirurgia para remoção do tumor, o paciente foi tratado com o fármaco errado, que aumentou ainda mais sua pressão arterial. Diga duas classes farmacológicas que podem ter sido erroneamente administradas e causado essa elevação.
8. A bexiga de um homem está cheia. Ao urinar, os receptores __________ provocam __________ do músculo detrusor e os receptores __________ provocam __________ do esfíncter interno.
9. Na ação dos receptores α 1 , qual é a ordem correta das etapas: ligação de α q ao GDP, ligação de α q ao GTP, geração de IP 3 , liberação de Ca 2+ dos depósitos intracelulares, ativação da proteína quinase, ativação da fosfolipase C?
10. Quais das seguintes ações são mediadas por receptores muscarínicos: redução da velocidade de condução no nó AV, secreção ácida no estômago, midríase, contração dos esfíncteres gastrintestinais, ereção, secreção de renina, sudorese em um dia quente?
C0010.indd 67C0010.indd 67 03/07/18 4:08 PM03/07/18 4:08 PM

COSTA
NZO
6a EDIÇÃO
LINDA S. COSTANZO
Fisiologia6 a E D I Ç Ã O
LINDA S. COSTANZO, PhD
Os alunos sempre dizem que o livro de Fisiologia da Dra. Costanzo é um dos seus favoritos. É fácil entender o motivo.
O texto simples, lógico e fácil de acompanhar da renomada docente de fisiologia Dra. Linda Costanzo faz com que Fisiologia, agora na 6ª edição, seja ideal para a disciplina e o preparo para provas.
Figuras e tabelas de excelente qualidade ilustram bem os procedimentos ou equações fisiológicas e as explicações passo a passo esclarecem conceitos desafiadores. Este texto colorido e de bom tamanho discute, de maneira abrangente e consistente, os principais conceitos fisiológicos em nível de sistema orgânico e celular, facilitando a compreensão de complexos princípios.
• As informações são apresentadas de forma objetiva, fácil e clara – a maneira perfeita de obter sucesso em cursos e provas.
• Os resumos dos capítulos e as perguntas da seção “Desafie-se”, ao final de cada capítulo, permitem uma revisão extensa do material e reforçam o entendimento e a retenção do conhecimento.
• As equações e amostras de problemas estão integrados por todo o texto.
• NOVO! Mais Quadros de Fisiologia Clínica relacionam a fisiopatologia com o contexto clínico
www.studentsconsult.com.br
A maneira inteligente de estudarstudentconsult
A MANEIRA INTELIGENTE DE ESTUDAR
Este livro tem conteúdo extra e gratuito no sitewww.evolution.com.br.Registre o código que está no verso da capa, dentro deste livro e conheça uma nova maneira de aprender:
- aprenda de forma dinâmica e objetiva com as animações;- teste seus conhecimentos com perguntas e respostas comentadas;- acesse os vídeos complementares.
A aquisição desta obra habilita o acesso ao sitewww.evolution.com.braté o lançamento da próxima edição em português, ou até que esta edição em português não esteja mais disponível para venda pela Elsevier, o que ocorrer primeiro.
Classificação de Arquivo Recomendada
Fisiologia
studentconsult
www.elsevier.com.br
Fisiologia
T R A D U Ç ÃO DA
6ª EDIÇ ÃO
Fisiologia
Costanzo_CoverMechanical Lu aprovada.indd 1 7/9/18 5:45 PM


















