Fisiología Animal
74
. \ Fisiología Animal o .!:: Ü tU E tU o tU "- tU ¿ cñ--= tU (l.) .--' e O .- a:b5- (l.) tU O "- J tU é O 'tU 6 E-o N ro ::::::¡ .--' .º (/) -o >. tU-o ü tU "- - «0 Biologla
Transcript of Fisiología Animal

. \ ~/ :
Fisiología Animal
o .!:: Ü tU E tU o tU
~ "-tU ¿ cñ--= tU (l.) .--' e O .-a:b5-(l.) tU O "
J tU
é O 'tU 6 E-o N ro ::::::¡ .--'
0~ .º (/) -o >. tU-o ü tU "- -«0
Biologla

UNIVERSIDAD NACIONAL DE COLOMBIA FACULTAD DE CIENCIAS
DEPARTAMENTO DE BIOLOGIA
FISIOLOGIA ANIMAL CODIGO: 11582
GUÍAS DE LABORATORIO

FISIOLOGÍA ANIMAL
© UNIVERSIDAD NACIONAL DE COLOMBIA
Facultad de Ciencias Departamento de Biología
© Autores: Arcadio Guzmán, profesor del Departamento de Ciencias Fisiológicas, Facultad de Medicina Joel Rojas, profesor del Departamento de Ciencias Fisiológicas, Facultad de Medicina y del Departamento de Biología, Facultad de Ciencias Marcela Camacho, Gladys Fajardo y Clara Spinel, profesoras del Departamento Biología, Facultad de Ciencias
Decano: Juan Manuel Tejeiro Vicedecana Académíca: Natalía Ruiz Director de Publicaciones: Gustavo Rubiano
Primera edición, 2003
Impreso por: Universidad Nacional de Colombia UNIBIBLOS
Correo electrónico: [email protected] Bogotá, Colombía

UNIVERSIDAD NACIONAL DE COLOMBIA FACULTAD DE CIENCIAS
DEPARTAMENTO DE BIOLOGIA
Programa Asignatura: Fisiología Animal. semestre 2002.
Introducción.
Código: 11582. Primer
El curso debe brindarle al estudiante de manera clara, conceptos básicos de la Fisiología animal para su apropiación. Esto le permitirá aplicarlos en el análisis de los principios y mecanismos, así como de las estrategias que se han desarrollado en los animales, dentro de los límites químicos y las posibilidades físicas, para su adaptación al medio ambiente. A través del método científico se le orienta y facilitan herramientas para plantear y resolver algunos problemas biológicos.
Tipo de curso: Teórico - práctico.
Intensidad Horaria: Seis horas por semana presenciales, (2 teoría, 4 de laboratorio) .
3

Teoría y Laboratorios.
Intrducción: Introducción al curso, presentación de docentes, generalidades sobre Fisiología animal
La Membrana Celular. Conceptos de modelos de membrana celular, composición de la membrana, propiedades físicas, su capacidad de almacenar carga. Potencial electroquímico. Potencial de membrana, transporte activo, por ejemplo bomba de sodio, transportes activos secundarios.
Potencial de Acción: Del potencial de membrana al potencial de acción. De la ecuación de Nerst y de Golman a los canales iónicos. Canales voltaje dependientes. La conducción del potencial de acción, neuronas mielínicas y amielínicas.
La Sinapsis: estructura sináptica, Sinápsis eléctricas y smapsis qmmlcas. Eventos eléctricos en sinápsis, El potencial sináptico. Sinápsis excitatorias e inhibitorias. Neurotransmisores y liberación de neurotransmisor.
La Recepción Sensorial: Propiedades de las células. Mecanismos de transducción y de salida sensorial. La codificación de estímulos e intensidades sensoriales. Control de la sensibilidad sensorial. Organos de los sentidos.
La Contracción Muscular: Bases estructurales de la contracción muscular esquelética. El acople excitación - contracción. Músculos sincrónicos y asincromcos. Control neuronal de la contracción muscular. Metabolismo energético aeróbico y anaeróbico y su relación con las fibras musculares aeróbicas y anaeróbicas.
Comunicación Hormonal. Generalidades. Mecanismo molecular de la acción hormonal. Hormonas de glándulas periféricas de vertebrados (epífisis,
4

tiroides, paratiroides, glándulas suprarrenales, páncreas, gónadas). Sistema hipotálamo-hipofisiario de vertebrados. Hormonas de los crustáceos. Hormonas de insectos. Ferormonas.
Intercambio De Gases Respiratorios y Sanguíneos. La Regulación Ácido Básica. Generalidades sobre respiración. Bases físicas de la respiración. Respiración cutánea. Respiración branquial: invertebrados, peces. Respiración pulmonar: invertebrados y vertebrados. Control de la respiración. Respiración traqueal. Cociente respiratorio.
El Sistema Circulatorio: Sistemas circulatorios abiertos y cerrados. El corazón y el sistema vascular, aspectos físicos en el control y función circulatorio. La actividad eléctrica en el corazón. Sistema arterial y sistema venoso. El sistema linfático. Respuesta cardiovascular a condiciones extremas, el ejercicio y el buceo.
Balance y Control Iónico y Osmótico. Problemas de la osmoregulación en ambientes terrestres y acuáticos. Su control hormonal.
5

Bibliografía Básica
[1] Eckert, Animal Physiology, Mechanisms and adaptations, Radall, Burggren and French, 4 edición, 1997. Freeman and Company.
[2] Ramón Latorre, José López-Barneo, Francisco Bezanilla, Rodolfo Llinás, Biofísica y Fisiología Celular, Universidad de Sevilla, 1996.
[3] Bertil Hille, Sinauer Associated INC. Ion Channels of Excitable Membranes, Publishers Sunderland, Massachusetts 1992.
[4] Bruce Alberts, Dennis Bray, Julian Lewis, Martin Raff, Keith Roberts, James D. Watson, Molecular Biology of the Cell, Garland Publishing, Inc New York, 1996.
Internet: http://www.ncbi.nlm.nih.gov/PubMed/ http:j /www.geocities.com/unaciona12002
6

Descripción general de un sistema de edición computarizado
Profesor: Arcadio Guzmán, Departamento de Ciencias Fisiológicas, Facultad de Medicina
Una configuración típica para un sistema de medición consiste en: un acondicinador de señales, un convertidor analógico a digital (conversor AjD), el computador y un programa de "software" que convierte el computador en un registrador digital. Este sistema de medición se muestra en la figura.
El Acondicionamiento de la señal: El propósito del acondicionamiento de la señal es el de suministrar la amplificación necesaria y la eliminación de los componentes no deseados de la señal por medio de filtros, antes de pasar la señal al convertidor AjD. Las señales de entrada del acondicionador tienen que ser eléctricas, de tal forma que, muchas señales no eléctricas que proceden de un organismo vivo, tienen que pasar por un transductor de señales antes de ir al acondicionador.
La adquisición de señales: Una vez que la señal ha sido acondicionada de una manera adecuada, necesita convertirse a forma digital para ser mostrada en un monitor o para ser almacenada en un disco en el computador. La señal acondicionada, entonces, se encamina al convertidor AjD, el cual toma muestras de la señal de voltaje a una velocidad determinada por el usuario. Las muestras entonces son convertidas en valores digitales y transferidas a la memoria del computador. La tarjeta digitalizadora que realiza la conversión AjD tiene también la capacidad de generar señales analógicas a partir de señales digitales, es decir, puede efectuar conversiones D j A y además, también puede generar señales digitales.
El análasis de la señal: Los programas de "software" son fundamentales en los sistemas modernos de adquisición de datos y control. Estos programas dirigen la adquisición de datos, el control del experimento y el análisis. Con el software se puede, por ejemplo, colocar la ganancia y determinar los filtros del acondicionador, la velocidad de muestreo del convertidor AjD, disparar el inicio de la estimulación y almacenar los datos en el disco. Después del
7

experimento, el software puede efectuar reducción de datos, análisis de los datos, elaborar archivos con los datos analizados y efectuar las gráficas de los resultados del análisis.
1,'~0. .~/-' -- .'.,' ,,, o.......~",
'~~!:/:~:~~;:t \. . \ 1\, ~:<.,::>;:::---~ "V '¡'llII'IIJIJI . '
\~:'~~~~:~:~~~(~ ~ .. ·="~G ..
~:~,2t~~~f~J.) Figura 1: Configuración típica de un sistema de medición computarizado. Al acondicionador de las señales se conectan los transductores, electrodos y preamplificadores necesarios: la salida del acondicionador se conecta éL
la caja de conexiones de la tarjeta digitalizadora (INTERFACE) que está colocada e n el interior del computador. Por medio de un cable plano (FLAT CABLE) se conecta la caja de conexiones a la tarjeta digitalizadora. Por medio del cableRS-232 se conecta el acondicionador de la señal al puerto serial del computador, para que a través del "software" se pueda manejar el acondicionador .
Anexo No. 1 La conversión de señales análogas a digitales
Una señal es cualquier variable física cuya magnitud o variación con el tiempo contiene información. Desde el punto de vista de cómo una señal lleva la información, esta puede ser analógica o digital. En la señal analógica, la información está contenida en la variación continua con el tiempo. En la señal digital, la información está contenida en el estado (alto, bajo; si, no; 1,0) en que se encuentre la señal.
8

El proceso de conversión de una señal analógica en digital consiste en :
l. Dividir la amplitud de la señal analógica en un número determinado de partes y
2. Asignar a cada parte un código.
Un código se construye con señales digitales. Cada señal digital es una variable binaria. Con tres variables binarias A, B, y C podemos construir el siguiente código:
A B C :-:-:','-11,':
O O O O O 1 O 1 O ._----.-
!
O 1 1 =J--i 1 O O - - - I
1 O 1 ~!J
1 1 O 1 1 1
Con este código se puede dividir una señal con una amplitud determinada en 8 partes iguales. Considerando que cada variable binaria puede solamente presentar dos valores, O ó 1; la asociación de dos variables binarias, establecen un código con 4 valores diferentes. La asociación de tres variables binarias establecen un código con 8 valores diferentes. En general, un conjunto de m variables binarias, puede representar hasta 2m valores distintos, así:
2 variables = 22 = 4 valores
4 variables = 24 = 8 valores
8 variables = 28 = 256 valores
12 variables = 212 = 4096 valores
Observamos que entre más variables binarias utilicemos, el proceso de convertir la señal analógica en digital es más perfecto porque efectivamente dividimos la señal a digitalizar en más partes, es decir resulta más exactamente
9

codificada. Desde el punto de vista tecnológico, lo menos costoso y lo más eficiente son 12 las variables binarias utilizadas, es decir, las señales analógicas se dividen en 4096 valores
La targeta digitalizadora: Las especificaciones básicas de esta tarjeta para las señales analógicas de entrada son:
1. N úmero de canales
2. Velocidad de muestreo
3. Resolución
4. Intervalo (rango) del voltaje de entrada
N úmero de canales: El número de canales de conversión analógica/digital puede ser de 8, 16, 32 o 64 canales. La tarjeta para digitalizar varias señales (varios canales) con una única tarjeta digitalizadora dse denomina "multiplexing". Esta técnica, selecciona y encamina la señal de un canal a la tarjeta digitalizadora y automáticamente, desconecta este canal y conecta el que sigue, hasta que termina con todos los canales, para luego iniciar de nuevo con el primero.
Frecuencia de muestreo (Sampling Rate): Este parámetro especifica que tan rápido suceden las conversiones análogo/digital. Una frecuencia de muestreo alta, adquiere más puntos de la señal en un tiempo dado, suministrando una mejor representación de la señal original. Obviamente, si la señal esta cambiando más rápidamente de lo que la tarjeta digitalizadora puede convertir aparecen errores en los datos medidos, esta distorsión se conoce como "aliasing".
!, .. "\ ,~ j., ,~ r' :\ t\ ,t-¡ 1\ ' I ; I ¡,I, tI , I , \ ' , I I
, ¡ , I
, ¡ , I ~ , I
I I 1 ! I I , I I I
I I , , I I ,
I I I I 1, ; I , , J ' ¡ , ,
~ " ',) " 1: '1)'
'¡.} \ I 'r' o) ,. y' •
10

1 1
, 1 , 1
1 1
" 1 " I
,1'..,
I T 11
/ 1, ,
I l' I
1 J 1 I l' .'
\ I I I
\11 :'/ I " , " U .... /
.1 " l'
,
1 ,
',.)
M uestreo correcto
/ .... , /\ 1:1
/'\, ,K, , ' .' " (, (\
, ' '1 "
\ 1 "
, , , 1 ,
1 , 1 ',' ,
I I , , 1 I 1 1 ,
1 , I
, , , , ~ 1 , II I 1
'1 I 1 ~' I
~ 1 I I ,
I I , 1 I " , ,
1 , " , 1
I , , 1 , , , , 1
1 i , , I 1 ,
'1 1, , , , , I , 1
'\ ; , , , i
\1 1 1
'1, , I ~ ,1/ r 'v 'J "
I j ..... )
M uestreo incorrecto
De acuerdo con el teorema de Nyquist, se debe muestrear al menos, con una frecuencia dos veces mayor que el componente de frecuencia máximo de la señaL Por ejemplo, las señales de audio convertidas en señales eléctricas por medio de un micrófono, tienen componentes de frecuencia de hasta 20 KHz, Se requiere para estas señales, un frecuencia de muestreo por lo menos superior a los 40 KHz,
Resolución: La resolución es el número de variables binarias que utiliza la tarjeta digitalizadora para representar la señal analógica, La resolución más utilizada es de 12 variables binarias, también se le denomina 12 bits de resolución,
!intervalo máximo del voltaje de entrada y ganancia: Este intervalo se refiere a la diferencia entre el máximo y mínimo valor de los niveles de voltaje que la tarjeta digitalizadora puede manejar. Por lo general, las tarjetas digitalizadoras ofrecen varias posibilidades de intervalos de voltaje, Un valor típico de intervalo máximo de voltaje es O a 10 voltios o de -10,24 a +10,24 voltios, Lo aconsejable para aprovechar la capacidad de entrada de la tarjeta digitalizadora, es presentar a su entrada señales analógicas aproximadamente con este valor de entrada, si no es así se desperdicia parte de la resolución, Para ayudar a solucionar el problema con las señales de baja amplitud, la tarjeta digitalizadora posee una etapa de amplificación anterior a la conversión digital teniendo valores, la amplificación, de 1, 2, 4 y 8 programables,
Ancho de código y ganancia de voltaje: El ancho de código es el cambio detectable más pequeño de voltaje de la señal analógica, es decir el mínimo cambio de la señal analógica que queda codificado, También se dice que este
11

cambio en voltaje representa el valor digital del bit menos significativo. Se calcula por medio de la siguiente expresión:
Ancho de código = (Intervalo de voltaje de entrada)/(ganancia)*(2resoluciól1)
Por ejemplo, un intervalo de voltaje de 20 voltios, una resolución de 12 bits y una ganancia de 8 permite obtener una anchura de código de 0,61mV. Entonces la resolución teórica de un bit en el valor digitalizado es 0,61 m V.
12

Anexo No. 2Descripción general del programa "AxoScope 1"
El AxoScope controla la tarjeta digitalizadora permitiendo reemplazar a los antiguos registradores a papel, con cinta magnética y los osciloscopios por un PC. Existen cuatro tipos de ventanas que pueden ser abiertas en AxoScope:
Ventana de registro (Scope Window): allí son mostrados los datos recién adquiridos y que pudieron haber sido grabados o no.
Ventana de análisis (Analisis Window): los archivos de los datos almacenados pueden ser cargados a esta ventana, medidos y editados.
Ventana con los datos medios (Cursor Window): los datos medidos en la ventana de análisis utilizando los cursores, son colocados en una tabla en la presente ventana.
Ventana de notas (Lab Book Window): allí se describen los detalles de la actividad del programa mientras se adquieren los datos y se analiza.
Descripción general de la adquisición: Para adquirir datos utilizando el AxoScope 1, se deben seguir los siguientes pasos:
1. Colocar el nombre a un archivo para salvar los datos. (Set Data File Names)
2. Escoger los nombres de las señales y las unidades para cada canal de datos que desea adquirir. (Signal Names and Units)
3. Editar el protocolo que va a ser usado para el registro de los datos. (Edit Protocol)
4. Salvar el protocolo en un archivo. (Save Protocol). Ud. puede entonces usar después "Open Protocol" para cargar cualquier protocolo ya salvado y comenzar a registrar inmediatamente.
13

5. Abrir una o más ventanas de registro y ajuste sus propiedades de acuerdo a cómo se quiere que sus datos sean registrados.
6. Comenzar la adquisición de los datos de su experimento.
Edición del Protocolo (Edit Protocol): Existen cuatro secciones principales en el protocolo de registro. Estas son:
1. Modo de adquisición y muestreo. (Adquisition Mode and Sampling)
2. Descripción de los canales que se van a utilizar en la adquisición de datos.
3. Disparo (Triggering)
4. Comentarios (File comment)
Modo de adquisición: Se van a utilizar en la realización de nuestros experimentos de los modos de adquisición: modo continuo (Gap-free) y modo osciloscopio. El modo continuo (Gap-free) permite que los datos sean presentados en la ventana de registro continuamente, siendo el usuario el que define exactamente la longitud y la ocurrencia de la adquisición. Si en este modo se registra en el disco, todos los datos son salvados al disco, no hay intervalos ("gaps") en el archivo de datos resultante, porque cada dato se escribe en el disco, hasta el límite de su capacidad. El modo osciloscopio permite que todos los datos de un barrido sean presentados en la ventana de registro o sean registrados en el disco si ellos cruzan el umbral. La longitud de cada segmento de datos es constante y depende de la longitud de barrido. En este modo el disparo (trigger) es indispensable para efectuar la adquisición.
Intervalo de muestra por canal: En el modo continuo, es el tiempo (en microsegundos) entre dos muestras adyacentes del mismo canal de entrada. En el modo de osciloscopio se habla de longitud de barrido por canal, correspondiendo éste al número de muestras de cada canal que serán adquiridas para formar cada barrido.
14

Canales para adquisición (Analog in Channels): Existen 16 canales analógicos de entrada para efectuar adquisiciones. Cualquier número de estos canales puede ser seleccionado. A cada canal se le asocia el nombre de la señal. Los nombres de las señales se dan utilizando el comando "Signal N ames and U nits" .
Información sobre el disparo: La fuente de disparo (Trigger Source) puede ser la misma señal que está adquiriendo, o un pulso de 5 voltios de amplitud, o una acción sobre la barra espaciadora del teclado. Umbral de disparo. Esta opción es aplicable únicamente cuando la misma señal es usada como fuente de disparo (trigger source). Corresponde al valor umbral que la señal debe sobrepasar para disparar una adquisición.
Comentario (File Comment): Ud. puede entrar un comentario para que sea almacenado como encabezamiento del archivo de adquisición que acaba de efectuarse.
Descripción general del "Cyber Amp 380" y del programa "CyberControl": El acondicionador "CyberAmp 380" es un conjunto de 8 amplificadores programables independientes, los cuales pueden amplificar, filtrar, quitar niveles D.C. y acoplar señales A.C. No posee controles manuales, siendo entonces manejado por el programa "CyberControl". Además de los 8 canales de acondicionamiento de las señales, posee un monitor de audio, que no es controlado por el computador.
Cada uno de los 8 canales acondicionadores posee las siguientes características:
1. Posee entradas diferenciales. La señal amplificada es la diferencia entre las señales en las entradas no inversora e inversora.
2. Posee acople de entrada AC (Filtros pasa altos).
3. Posee filtros pasabajos (Bessel, cuarto orden).
4. La ganancia por canal puede tener valores discretos variando entre xl y x2000
15

5. Posee un filtro banda-rechazada (Notch filter) para eliminar 60 Hz de la frecuencia de línea de potencia
6. Se puede introducir o quitar nivel DC en la señal (DC offset y autozero).
7. Existen dos tipos de entrada para cada canal. UN conector BNC y un conector de 15 pines: El contacto interno del conector BNC está conectado a la entrada no inversor del amplificador.
8. El conector de salida para canal es un conector BNC.
Por medio del programa CyberControl, podemos variar las principales características de cada uno de los canales de acondicionamiento de la señal. Podemos entonces variar los filtros pasaaltos desde una frecuencia de corte 0,1 Hz pasando por 1 Hz, 10 Hz, 30 Hz, 100 Hz y 300 HZ. La anterior variación puede ser efectuada en cada una de las entradas del amplificador diferencial. También se puede variar la ganancia en pasos: la ganancia inicial (prefiltro) puede ser xl, x10, x100. Y la ganancia posfiltro xl, x2, x5, x10, x20, x50, x200. Los filtros pasabajo poseen frecuencias de corte que se pueden cambiar desde 2 Hz a 30 KHz y eliminar todos los filtros pasabajo con la opción "Bypass". Con la opción "DC offset" podemos colocar un intervalo de voltaje DC máximo comprendido entre +3 y -3 voltios. Y por medio del "Autozero" podemos colocar en cero el contenido DC de la señal.
16

Preparación de Soluciones para Medición.
Autores: Rojas, MSc, Marcela Camacho, MD., PhD, Departamento de Biología, Facultad de Ciencias.
La adquisición de datos de modelos celulares y animales requiere de la utilización de soluciones y medios de cultivo que garanticen la viabilidad de los mismos y la obtención de información. Para llevar a cabo las prácticas experimentales programadas para este curso el estudiante debe preparar dichas soluciones.
Objetivo: Preparar soluciones fisiológicas para registros de datos.
Soluciones
Solución de Ringer NaCl 114,9 mM; KC12,5 mM; NaH2P04 0,85 mM pH 7,1.
Ringer de anfibio: NaCl112 Mm, KCl1,9 mM, CaC12 1,1 mM, NaH2P04 0,7 mM, pH = 7,2
Soluciones de registro electro fisiológico
KCL 150 Mm, pH: 7.4
NaCl 150 mM, pH 7.4
Solución externa (Hanks Na) NaCl145 mM KCl5 mM CaC12, 1 mM MgC12, 2 mM HEPES-Na, 10 mM Glucosa 5 mM pH 7.34, 300 mOsm
Informe. Presentarlo en forma de guía. Introducción. Soluciones precisas para mantenimiento celular. Principios sobre pH y osmolaridad. Principios de los equipos que permiten la medición de pH y de osmolaridad. Describir el protocolo (receta) seguido para la preparación. Cálculo.
17

81
"sopu'qns81 lUltSOW
"PUPPUloUlsO 8p U9P~p8W "Hd 8p u9P1p8W
"8[US8d

Laboratorio demostrativo canales iónicos.
Lectura complementaria: Laboratory set up, Axon Instruments manual
Lugar: Laboratorio de Biofísica-Centro Interntextbfacho: Departamento de Biología, Facultad de Ciencias Objetivos: Visitar un laboratorio de investigación básica en el cual se llevan a cabo registros de canales ióniclectrofisiólogicos y de microfluorometría
Visita:Laboratorio de Electrofisiología Montaje de patch clamp- un electrodo Montaje de voltage clamp-dos electrodos Mediciones de calcio con microfluorometría Montaje de microinyección
Las demostraciones programadas se harán dependiendo de la disponibilidad de material
Demostraciones:
Patch clamp en configuración de célula entera.
• Simulación de sello con célula modelo
• Registros de corrientes totales de macrófagos murinos
• Demostración de un registro de corrientes totales de salida - farmacología
• Demostración de un registro de corrientes totales de entrada - farmacología
Patch clamp en configuración de célula adherida.
• Registros de canales únicos en macrófago control
• Registro de canales únicos en macrófagos sometidos a APT
Mediciones de calcio en neuronas utilizando la sonda Fura-2
• Demo de Axonimage
19

• Macrófagos cargados con fura-AM
Votage clamp
• Registros de ovocitos de Xenopus laevis
• Registro de Kv 1.1, cambios en la activación
• Microinyección de ovo citos
Práctica experimental Elaboración de pipetas de registro. Adquisición de datos. Comparación de potenciales medidos con diferentes soluciones.
20

Transporte activo de sodio (Piel de sapo)
Profesores: Arcadio Guzmán y Joel Rojas, Departamento de Ciencias Fisiológicas, Facultad de Medicina y Departamento de Biología, Facultad de Ciencias
Objetivo. Medir la corriente de cortocircuito a través de la piel de sapo.
Teoría. La bomba Na-K. La viabilidad de las células nerviosas es mantenida por el transporte constante de Na y K a través de la membrana celular en contra de sus gradientes electroquímicos. Esta tarea permanente es llevada a cabo por la bomba N a - K mediante un transporte activo y la energía para la bomba es obtenida de la hidrólisis de ATP. Un promedio de 3 iones de sodio y 2 iones de potasio son transportados por cada molécula de ATP hidrolizado. El requerimiento de sodio es extremadamente específico. Es el único substrato aceptado para el transporte neto hacia afuera y es el único catión monovalente no aceptado para un transporte hacia adentro. Entonces, el litio, el amonio, el rubidio, el cesio y el talio pueden substituir al potasio en la solución externa, pero no al sodio en la solución interna. El requerimiento para el potasio externo no es absoluto, en su ausencia la bomba podrá sacar sodio a aproximadamente un 10 de su capacidad en un modo "no-acoplado". El sistema de transporte es bloqueado específicamente por glucósidos digitálicos, particularmente la ouabaina. La bomba de N a - K ha sido extensamente estudiada en una gran variedad de preparaciones, incluyendo axones gigantes de calamar, vejiga y piel de sapo.
Transporte epitelial Los tejidos epiteliales están especializados en actuar como barreras entre varios compartimientos corporales y en formar las capas superficiales que se enfrentan al ambiente. Cada órgano o compartimiento dentro de un animal tiene una cobertura de capas superficiales. Algunas de estas hojas sólo sirven como barreras pasivas entre compartimientos, sin ocuparse de un transporte preferente de solutos yagua. En otros casos, están implicadas en el trans-
21

porte activo realizando funciones reguladoras. Los epitelios tienen varias características en común. En primer lugar, se presentan en las superficies que separan los espacios internos del organismo respecto del ambiente. En segundo lugar, las células que forman la capa más externa del epitelio suelen estar selladas entre ellas mediante uniones estrechas lo que, en grado diverso según los epitelios, ocluye las vias paracelulares entre los lados serosal (interno) y mucosal (externo) del epitalio. Las sustancias que son transportadas activamente a través de un epitelio deben seguir las vías transcelulares, en las que participa la membrana celular.
Transporte salino activo El transporte de iones activo entre los dos lados de un epitelio ha sido demostrado en la piel y vejiga urinaria de los anfibios, las branquias de los peces, intestino de vertebrados e invertebrados, el túbulo renal y vesícula biliar de los vertebrados. La mayor parte del trabajo inicial sobre transporte activo epitelial se realizó con piel de rana. En los anfibios la piel actúa como el principal órgano osmoregulador. La sal es transportada activamente desde el lado mucosal (es decir, desde el lado que mira el agua de la charca) al lado serosal de la piel, para compensar las pérdidas salinas desde la piel hacia el agua dulce que rodea la rana.
La preparación experimental de la piel de rana para el estudio del transporte epitelial fue desarrollada por el fisiólogo alemán Ernst Huf y el fisiólogo danés Hans Ussing en la década de los años 30 y 40. Se obtiene un trozo de piel abdominal de rana, de varios centímetros cuadrados de área y se coloca entre las dos mitades de una cámara de Ussing (figura No. 1). La piel se fija entre las dos hemicámaras, se introduce la solución de prueba (por ejemplo Ringer de anfibio: NaCl 112 Mm, KCl 1,9 mM, CaC12 1,1 mM, NaH2P04 0,7 mM, pH = 7,2) con la piel actuando como una separación entre los dos compartimientos.
Using, en 1947 comunicó los primeros experimentos en los que se usaron dos isótopos del mismo ión para medir los flujos bidireccionales. Se preparo la solución de Ringer del compartimiento externo con el isótopo Na 22, mientras que el Ringer del compartimiento externo se preparó con Na 24. Se siguió la aparición de cada uno de los dos isótopos en el lado opuesto de la piel a lo
22

largo del tiempo. En todos los experimentos se observó que el Na muestra un movimiento neto a través de la piel, desde el compartimiento externo hacia el interno. Se concluyó que el movimiento neto de iones de sodio es el resultado del transporte activo, basandose en las siguientes razones:
1. Tiene lugar sin gradiente de concentración alguno, incluso en contra de un gradiente electroquímico.
2. Es bloqueado por inhibidores metabólicos generales, tales como el cianuro y el ácido yodo acético y por inhibidores específicos de transporte, como la ouabaína.
3. Muestra una fuerte dependencia de la temperatura.
4. Presenta una cinética de saturación.
5. Manifiesta especificidad química. Por ejemplo, el Na es transportado, mientras que el ión litio, muy relacionado con el Na, no es transportado.
e
+ L= ______ ~~ Amperlmetro -s: ~----------~~
Electrodo para corriente
Figura 2: Cámara de Ussing. La piel de rana separa las dos hemicámaras, cada una está llena con solución de Ringer u otra solución de prueba. Se puede ajustar la fuente de corriente hasta que la diferencia de potencial a través de la piel es cero. Bajo estas condiciones, la corriente que fluye por el circuito externo, es equivalente a la corriente iónica debida al transporte activo a través de la piel.
23

Potencial eléctrico transpitelial El potencial eléctrico a través de la piel es generado por el transporte activo de sodio, estos iones son transferidos del exterior al interior. La bomba separa los iones de sodio positivos, de los iones de cloro negativos y esta separación de carga es medida como un potencial eléctrico transepitelial, con valores comprendidos entre 10 y 100 m V. El gradiente eléctrico creado por la bomba de sodio (hace la piel negativa fuera y positiva dentro) atrae los iones de cloro hacia adentro a través de la piel, pero dado que la bomba activa de sodio siempre adelanta el movimiento de sodio al de cloro, la separación de carga siempre es mantenida.
Corriente de cortocircuito Las correspondencias eléctricas del transporte activo de sodio fueron publicadas por Ussing y Zehran en 1951. Ellos razonaron que si sólo el sodio es transportado activamente a través del epitelio, debe haber una concordancia cuantitativa entre el número de iones sodio transportados por segundo a través de un área unitaria de piel y la intensidad de corriente resultante. Ussing y Zehran impidieron la formación de una diferencia de potencial, llevándose las cargas negativas excedentes en el lado externo conforme iban quedando separadas del ión sodio que estaba pasando al interior, por medio de una red eléctrica externa (figura No. 1). El ajuste a cero de la diferencia de potencial, haciendo circular una corriente a través de un circuito externo, permite comparar esta corriente con la cantidad neta de iones sodio transportados a través de la piel, en la unidad de tiempo. Experimentalmente se encontró una gran concordancia entre la corriente a través de la piel y el flujo de sodio medido isotópicamente. Tanto la corriente como el transporte de sodio son reducidos o eliminados por la ouabaÍna.
Modelo para la célula epitelial
Las células adyacentes de un epitelio de transporte están Íntimamente unidas por las uniones estrechas. Ésta situación forzaría a todas las sustancias a atravesar la membrana de la célula epitelial dos veces, primero al cruzar la membrana del lado mucoso y luego al dejarla pasar del lado seroso. El transporte activo por esta ruta requiere que la membrana externa de cada célula epitelial sea diferenciada, de manera que la membrana celular que mira
24

FUERA ..
Na+N~};' r. ~:',
I~.;, ,
Membrana celular
.: ,:.: ,. .;.
Célula
'1\' t 1"+ K
Figura 3: Modelo de la célula epitelial que transporta sodio en la piel de sapo, La célula epitelial es asimétrica. La cara externa de la célula se considera permeable al sodio, la cara interna permeable al potasio. La bomba P en la cara interna de la membrana, transporta el sodio que se ha difundido al interior celular, hacia el lado seroso. La bomba también transporta potasio de nuevo al interior celular, para mantener el balance eléctrico y osmótico.
hacia el lado seroso del epitelio difiere en las propiedades funcionales de la porción de la membrana que mira hacia el lado mucoso. Los experimentos con piel de rana han proporcionado distintas líneas de evidencia que apoyan esta hipótesis. Por ejemplo: 7
1. La ouabaína, que bloquea la bomba Na-K, inhibe el transporte transepitelial de sodio, sólo cuando es aplicada en el lado interno (lado seroso) del epitelio. Es inefectiva sobre el lado mucoso. Por el contrario, el fármaco amilorida, un potente inhibidor del transporte facilitado pasivo, bloquea los movimientos de sodio a través de la piel sólo cuando se aplica en el lado externo de la misma.
25

2. Para que telnga lugar el transporte activo de sodio de una manera máxima, el potasio debe estar presente en la solución del lado interno, pero no se necesita en el lado externo.
3. El transporte de sodio muestra una cinética de saturación en función de la concentración de sodio en el lado externo; no se ve afectado por la concentración de sodio en el lado interno.
Tales datos han conducido al modelo de transporte que se muestra en la figura No. 2.
Materiales y métodos
Complejo instrumental El complejo instrumental está conformado por un sistema de adquisición de datos, un estimulador de corriente, los electrodos, la cámara para la piel y la preparación biológica (piel de sapo Bufo). Los electrodos que se utilizan para medir el voltaje se llaman electrodos de calomel. Este electrodo está formado por un tubo de vidrio en forma de U, la parte baja del tubo se llena de mercurio. Por un lado, un hilo de plata que se sumerge en mercurio por el otro, el mercurio está en contacto con una solución de cloruro mercurioso (calomel) la cual está en contacto con una solución de cloruro de potasio. En la figura No. 3 se muestra el montaje experimental.
Resultados
El voltaje transepitelial es de 37, 8mV aproximadamente, positivo en el interior (lado seroso) con respecto al exterior. Se aplica un pulso de corriente de O, 175mA de intensidad y 700ms de duración a través de la piel de sapo. Como se observa, el voltaje transepitelial cae a Om V, denominándose entonces la corriente O, 175mA como "la corriente de cortocircuito". Una vez termina la aplicación de esta corriente, el voltaje transepitelial se restablece a su valor anterior. Es posible calcular el número de iones positivos que atraviesan la piel en 1 segundo, considerando que son iones de sodio y que la corriente de sodio es exactamente igual a la corriente de cortocircuito, medida en el circuito externo. Se sabe por otros estudios que una bomba mueve alrededor de 300 iones cada segundo. El diámetro del sector circular de la piel utilizada
26

J>
,....---,Ag
! ~ __ .-__ j~ __ ~_A_- ¡
Estimulador de corriente
Hg
Figura 4:
0..
G.04
0.0.
0.01-
0.01
o-
-o..t1
100 1000
Figura 5: Medición de la corriente de cortocircuito.
f 2IlOO
8w1/1
en la cámara de 2, 2cm. Por consiguiente Ud. puede obtener un valor aproximado del número de bombas que existen en un área de 1 cm. cuadrado de
27

piel de sapo Bufo.
Referencias
Eckert, R., Randall, D., Augustine, G. Fisiología animal. InteramericanaMcGraw-Hill. 1990. Oackley, B. and Schafer, R. Experimental neurobiology. The University of Michigan Press. Ann Arbor. 1978.
28

Taller percepción sensorial
Profesor: Marcela Camacho: Departamento de Biología, Facultad de CienCIas
Lecturas
• Honeybee navigation: nature and calibration of the odometer. Srinivasan M.V., Zhang S., Altwein M. y Tautz J. (2000) Science, vol 287, p 851-853
• Ant odometry in the third dimension, Wehigemuth, S., Ronacher, B. y Wehner, R. (2001) Nature, vol 411, p 795-797
• Honeybee dances communicate distances measured by optic fiow, Esch, H. E, Zhang, S., Srinivasan, M.V. y Tautz, J. (2001), vol 411, p 581-583
• Perception of three-dimentional shape infiuences colour perception through mutual illumination. Bloj M.G., Kersten D. y Hurlbert A.e. (1999) Nature, vol 402 p 877-879
• Motion Integration and postdiction in visual awareness, Eagleman D.M. y Sejnowski T.T (2000) Science, vol 287, p 2036-2038.
• Pattern recognition and active vision in chickens. Dawkins M.S. y Woodington A. (2000) Nature vol 403, p 652-655
• Motion direction, speed and orientation in binocular matching, van -Ee R. y Andersen B.L. (2001) Nature vol 410, p 691-693
29

Excitación y respuesta del nervio ciático de sapo
Profesor: Arcadio Guzmán, Departamento de Ciencias Fisiológicas, Facultad de Medicina
Objetivo. Describir las principales características eslectrofisiológicas del nervio ciático de sapo Bufo.
Teoría Potencial de acción compuesto Se puede generar un potencial de acción simultáneamente en cada uno de los miles de axones que componen el nervio periférico, por medio de un único estímulo eléctrico. La respuesta colectiva se denomina potencial de acción compuesto (PAC). El registro del PAC es una técnica útil y bastante reproducible. Con el nervio ciático de sapo es posible explorar varios aspectos básicos de la conducción neuronal, como el fenómeno del umbral, la velocidad de conducción, el periodo refractario relativo, las máximas frecuencias de disparo, la propagación diferencial de grupos de fibras nerviosas y nos enseña como varía la forma del PAC dependiendo del modo de registro utilizado.
Los nervios periféricos de vertebrados contienen típicamente miles de axones, pertenecientes a neuronas motoras y sensoriales. Cada axón conduce potenciales de acción de una forma todo - o - nada, de una manera autopropagada, sin embargo, la propiedad todo - o - nada desaparece en el nervio periférico.
El nervio ciático de sapo era la preparación clásica para el estudio del potencial de acción, hasta que fue reemplazada por el axón gigante de calamar. El potencial de acción a que nos referimos se llama compuesto, porque se puede observar la respuesta de grupos poblacionales de las fibras neuronales que componen el nervio ciático. En la figura No. 1 se muestra la propagación de las ondas alfa y beta del PAC de la rana toro, correspondientes a dos poblaciones de fibras neuronales.
Cuando la distancia entre los electrodos de estimulación y de registro es
30

L+/3
ümm l'
21mm I'.e'w l'
1 /3 41 mm ~ )(
1\
II
\ I
81 mm-
143mm
'ir TIME OF
S,IMULUS 10 msec
Figura 6: Los cuatro registros (W-Z) fueron obtenidos en cuatro sitios distintos, con el electrodo de registro colocado a las distancia de 21, 41, 81 Y 143mm desde el sitio de estimulación (Omm). A medida que el potencial de acción se mueve a lo largo del nervio, los componentes alfa y beta se separan cada vez más debido a que la velocidad de propagación de estos componentes es diferente lo cual se aprecia mejor entre mayores sean las distancias entre el sitio de estimulación y el de registro. Los registros han sido acomodados verticalmente en proporción a la distancia de los respectivos electrodos de registro al punto de estimulación (Omm). La línea sólida a la izquierda ha sido dibujada sobre el artefacto del estímulo. Las dos línea intermitentes cruzan por los puntos de iniciación de los componentes alfa y beta, tal como se observa en el registro Z y se han indicado por puntos en cada registro. El hecho de que las líneas intermitentes sean rectas indica que la velocidad de propagación de los componentes alfa y beta a lo largo del nervio es constante pero distinta. (Adaptado de Erlanger y Gasser, 1968).
grande, el PAC consiste de varias ondas (alfa, beta, gamma), tal como se observó en la figura No. 1; lo cual es debido a que el tronco nervioso contiene
31

fibras con diferentes velocidades de conducción. Erlanger y Gasser (1937) clasificaron los grupos de fibras nerviosas, de acuerdo con sus velocidades de conducción, tal como aparece en la tabla No. 1
Fibre group Conduction velocity (mi sec) a 41
A b 22 g 15
B 4
C 0,7
Tabla No. 1. Grupos de fibras en el nervio ciático de la rana Toro, con valores típicos para sus velocidades de conducción a la temperatura del laboratorio. (Basado en Erlanger and Gasser, 1937)
Diferencias entre el registro intra y extracelular de un axon. Cuando se efectúa el registro intracelular, es necesario utilizar un microelectrodo de vidrio que pueda atravesar la membrana celular de tal forma que el interior de él quede conectado eléctricamente con el interior celular. Se mide de esta manera la diferencia de voltaje entre el interior y el exterior de la célula. El interior de la neurona posee un voltaje de reposo aproximadamente de -60mV con respecto al exterior; durante el potencial de acción el interior se vuelve positivo, aproximadamente a un valor de +40m V con respecto al exterior, lo cual implica que la variación de voltaje durante el potencial de acción, medido intracelularmente es de 100mV. Las posibilidades técnicas de efectuar un registro intracelular de un axón son escasas a no ser que sea un axón gigante de calamar.
Para el registro extracelular tenemos varias maneras de hacerlo, sin embargo todas tienen en común el hecho de realizarse en el exterior celular. Además los valores medidos extracelularmente durante el potencial de acción, tienen valores máximos alrededor de los 15mV. En la figura No. 2 se muestra un esquema de las diferencias.
32

Los potenciales de aCClOn medidos intra o extracelularmente en un axón, presentan la propiedad de todo - o - nada, es decir, su amplitud no depende del tamaño del estímulo. Sin embargo, en un nervio, sólo tiene sentido hablar de un registro extracelular y además los potenciales de acción no presentan la característica del todo - o - nada, sino que su amplitud depende del tamaño del estímulo. La razón para esto, es que el potencial de acción registrado es el resultado de la actividad simultánea en un gran número de axones (cada uno de los cuales obedece a la ley del todo - o - nada) que poseen diferentes umbrales.
+++ +++
( ___ ++_+++--,-,0
Intracellular recordln,
Extracetlutar recordin, :v-
Figura 7: Diagrama para mostrar la diferencia en signo de los potenciales de acción registrados por electrodos intra y extracelulares. Los registros extracelulares son a menudo mostrados con los potenciales negativos hacia arriba.
33

El u -~ e
s ql q2
b a 1 1 s ., qz
e 2 3 """'sec:
Figura 8: Esquema para el registro de los potenciales monofásicos y bifásicos. S corresponde a la aplicación del estímulo y el - e2 son los electrodos de registro.
Tipos de registro extracelular sobre el nervio. Se pueden obtener potenciales bifásicos y monofásicos cuando se registran los potenciales con electrodos colocados directamente sobre el nervio y también se pueden obtener potenciales trifásicos si los electrodos están colocados a cierta distancia del nervio. Los registros bifásico y monofásico se obtienen con dos electrodos activos colocados sobre el nervio, en el primer caso ambos detectan el potencial de acción, mientras que en el segundo, solo uno detecta el potencial, como se muestra en la figura No. 3. La técnica utilizada para obtener el registro monofásico consiste en evitar que el potencial de acción llegue al último electrodo activo, lo cual se logra dañando el nervio o aplicándole un anestésico.
El potencial de acción bifásico. La forma de este potencial se explica por el paso de la zona despolarizada bajo los dos electrodos. Los electrodos están conectados al amplificador diferencial y con el objeto de mostrar un potencial de acción bifásico, inicialmente con polaridad positiva, el primer electrodo se conecta a la entrada inversora y el segundo a la no inversora del amplificador diferencial. En la figura No. 4 se muestra que cuando el potencial de acción está lejos de los electrodos, estos están al mismo potencial y el amplificador diferencial tiene una salida igual a cero. Debemos tener en cuenta que la zona despolarizada es una zona de negatividad que se mueve con una velocidad y que tiene una lon-
34

gitud determinada. Cuando esta zona está colocada únicamente por debajo del electrodo que conecta a la entrada inversora, aparece un voltaje positivo de salida del amplificador y cuando la zona despolarizada está colocada únicamente por debajo del electrodo que conecta a la entrada no inversora del amplificador diferencial, aparece un voltaje negativo en su salida. Finalmente, cuando los dos electrodos, detectan el mismo valor de negatividad la salida es igual a cero.
Velocidad de conducción del potencial de acción compuesto Como se muestra en la figura No. 1, la forma del potencial de acción compuesto de nervio se modifica en función de la distancia recorrida. Se tiene la impresión que se trata de ondas distintas, pertenecientes a fibras dotadas de propiedades fisiológicas diferentes, confundidas en el origen, estas ondas se separan progresivamente a medida que avanza sobre el nervio en razón de sus diferentes velocidades. La velocidad de conducción del potencial en las fibras amielínicas es com,tante y está directamente relacionada con la raíz cuadrada del diámetro del axón, mientras que la velocidad de conducción del potencial de acción en las fibras mielinizadas está linealmente relacionada con el diámetro del axón. La velocidad de conducción se puede medir fácilmente en un nervio, desplazando una distancia conocida el primer electrodo de registro del potencial de acción compuesto y midiendo la diferencia temporal en el registro obtenido.
35

e, _-=:::!::=:~==
:2==~ ____ .. ========~
t 3 •
'4 ======::m~-==
t s •
Figura 9: La onda mono fásica de la despolarización de las fibras, recogida por dos electrodos activos, es registrada como una onda bifásica porque ella pasa cuando se propaga bajo cada uno de los electrodos. El primer electrodo está conectado a la entrada inversora del amplificador diferencial. (Instantes de t1 a t5).
Materiales y métodos
Complejo instrumental El complejo instrumental está conformado por un sistema de adquisición de datos, un sistema de acondicionamiento de la señal, un estimulador de corriente eléctrica, la cámara de la preparación, la solución de Ringer y la preparación biológica. La solución de Ringer tiene la siguiente composición: NaCl114,9mM; KCl2,5mM; Nah2po40,85mM y pH7, 1. El sistema de adquisición de datos está conformado por un computador PC compatible IBM, una tarjeta digitalizadora Digidata 1200 y su caja de conexiones y el programa AxoScope 1.1 de Axon Instruments. El sistema de acondicionamiento de la señal está conformado por el acondicionador CyberAmp 380 de Axon y un amplificador diferencial. El estimulador de corriente puede generar pulsos de duración y amplitud variable y controlada, además produce un pulso de sincronismo (trigger) para disparar el sistema de adquisición de
36

datos. La cámara para la preparación está constituida por un conjunto de electrodos horizontales donde se coloca el nervio ciático. En la figura No. 5 se muestra el esquema del montaje experimental.
~ CRO
IN y '---- %
STIMULATOR
+
OUT
I -~
TRIGGER 1 ~
"'" 1 PAEAMPLlFIEA SHIELDED
CABLE
f
\- t-
H -I ~
¡ z
z O O ¡:::: - <: t-u.C!) üo<: w c.. g; O o a:
c..
N AVI~ E ENERVE CHAMBER
Figura 10: Esquema del montaje experimental.
Procedimiento Inmediatamente después de la disección del nervio ciático del sapo, se procede a colocarlo sobre los electrodos y a adicionarle gotas de solución de Ringer en los sitios de unión de los electrodos con el nervio. Los electrodos de estimulación se conectan al estimulador teniendo en cuenta que el electrodo catódico despolariza el nervio, por consiguiente el electrodo anódico lo hiperpolariza y debe estar colocado hacia la extremidad del nervio, con el objeto de evitar que interfiera con el potencial de acción generado en el cátodo. Luego se conectan tres electrodos al amplificador diferencial, primero el común, luego el electrodo inversor y finalmente el electrodo no mversor.
37

Resultados
~~
10 t ,.--r-·T·-····.--~ • • • "-CIIIOI
, ID
.. 111
Figura 11: Potencial de acción compuesto registrado a una escala de tiempo de 50 ms.
~>
~~
.0._
, u
~( ... ) , 7.1
, 10 12
"MI
~f\--_._._. __ .. _ ...... _-_._. __ ._. -¡ r---r i r~·t·-"'_··""·-----r---r--r·~~--~
U S tl W ~ Tlnool_l "1/1
Figura 12: Registro bifásico y monofásico del potencial de acción compuesto.
Este potencial de acción se obtuvo con un pulso de estimulación de 0.07 mA y una duración de 0.1 ms. La amplitud alcanzada por el potencial de acción es aproximadamente de 18 m V. Se observa el artefacto del estímulo, el
38

cual se ha inducido electromagnéticamente a través de los electrodos y cables de conexión del amplificador. Este potencial muestra un after-potential positivo claramente definido después del potencial de acción y posee una duración aproximada de 10 ms y una amplitud porcentual del 8 con relación a la amplitud de la espiga del potencial de acción. Los potenciales posteriores (after-potential) son importantes porque son un indicativo de los mecanismos íntimos de ciertos procesos nerviosos. Un aumento del metabolismo del nervio aumenta los potenciales posteriores, pero tiene poco efecto sobre la espiga y en la asfixia los potenciales posteriores son abolidos, mientras que la espiga sigue presente. Podemos concluir que los procesos representados por los potenciales posteriores son necesarios para el restablecimiento de las condiciones que hacen posible la conducción sucesiva de impulsos. Detlev, W. et al. Bioelectric Studies of the Excitation an response of nerve. Annual Review of Physiology. Vol. 1, 1993.
El registro bifásico se obtuvo con dos electrodot> activot>, ambot> detectaron el potencial de acción. El registro monofásico se obtuvo también con dos electrodos activos pero se impidió que al último electrodo activo llegase el potencial de acción, utilizando un anestésico local (Xilocaína al 2 por ciento). La duración del componente alfa del potencial de acción compuesto es de 2 ms aproximadamente, tal como t>e obtiene en el registro monofásico. En el registro bifásico se obtiene una duración aproximada de 4ms.
0.11.
~~~~~~s~~~~~~-~~
..... (mo) ""'11
Figura 13: Variación de la amplitud del potencial de acción compuesto con la amplitud del estímulo.
39

0.02'
j; 0.1
____ 'O_·01-"~,-~-~r_~·-~;___r~~1 I I 2.1 ti 7.5 10 12
~( ... ) 8w112
Figura 14: Medición de la velocidad de conducción del potencial de acción compuesto.
A medida que aumentamos la amplitud del estímulo de corriente de duración constante 0,1 ms desde 0,025mA hasta 0,060mA en pasos de 0,005mA se incrementa la amplitud del potencial de acción compuesto. Con el valor de 0,025mA no se obtiene respuesta. La primera respuesta se obtiene con el estímulo de O, 030mA y una duración de O, 1ms.
Entonces, la respuesta umbral está comprendida entre 0,025 y 0,030 mA. Con amplitudes de estímulo superiores a 0,060 mA, la amplitud del potencial de acción se acerca a un valor máximo, indicando que todas las fibras están despolarizadas y además en la gráfica se observa que a medida que aumenta la amplitud del estímulo, el valor pico de cada potencial de acción se alcanza un poco más temprano. La variación de la amplitud del potencial de acción con la amplitud del estímulo nos indica que la respuesta del nervio ante un estímulo no es todo - o - nada. Cada fibra nerviosa que conforma el nervio sí responde ante un estímulo como todo - o - nada, es decir, al llegar al estímulo umbral la respuesta siempre es máxima y no varía con la amplitud del estímulo.
En la gráfica encontramos dos potenciales de acción, ambos fueron producidos con el mismo estímulo: 0,0050 mA de amplitud y 0,1 ms de duración. Cuando los dos electrodos de registro activos estaban separados 35 mm se
40

obtuvo el potencial de acción de mayor tamaño, después de acercar el primer electrodo de registro al segundo a una distancia de 20 mm, se obtuvo el potencial de acción de menor tamaño. Entonces, la amplitud del potencial de acción bifásico registrado extracelularmente, depende de la separación de los electrodos de registro. La medición de la separación temporal de los dos valores máximos de las espigas de los potenciales de acción es de 0,075 ms. El cálculo de la velocidad se obtiene utilizando la relación: Velocidad=(espacio)/(tiempo), es decir, V = (20 mm)/ (0,75 ms) = 26,7 mis a la temperatura ambiente. La velocidad del potencial de acción compuesto es constante para una temperatura determinada.
f\
""'--í ~'-~-~~'~·w-··r.-r ~"""._'-"'~',.
11MI(_) • ... , ..
Figura 15: Medición del periodo refractario relativo.
Para obtener los potenciales de acción que se muestran en la figura, se han aplicado pares de pulsos de estimulación. En todos los casos la amplitud del estímulo fue constante e igual a 0,050 mA y la duración de 0,1 ms. El primer par de estímulos se aplico con una separación de 15 ms, en este caso podemos considerar que las amplitudes de los potenciales de acción son prácticamente iguales. El segundo par de estímulos se aplicó con una separación de 12 ms, la amplitud del segundo potencial del par, tiene una amplitud un poco menor que el primero. El tercer par de estímulos se aplicó con una separación de 9 ms, el cuarto par de estímulos con una separación de 6 ms y finalmente el quinto par de estímulos se aplicó con una separación de 3 ms y no se obtuvo una segunda respuesta. La amplitud de los segundos potenciales de acción del par, se encuentra disminuida, porque las fibras nerviosas no se han alcanzado a recuperar de los cambios ocurridos por la generación del primer
41

potencial de acción. Podemos pensar que para que los potenciales de acción posean todos la misma amplitud, se requiere como mínimo una separación de 15 ms, lo cual corresponde a una frecuencia aproximada de 67 Hz.
Discusión de resultados Como se puede apreciar de una manera general de los resultados obtenidos, la preparación del nervio periférico es muy útil para enseñar los principios básicos de la conducción nerviosa. Además el montaje experimental utilizado es muy sencillo. Actualmente, existen muchas investigaciones con el nervio ciático de sapo, sobre todo en las áreas de la farmacología y toxicología. Los siguientes artículos son dos ejemplos:
• Asciouglu, M. and Ozesmi, C. Effects of grayanotoxin
• 1 on threshold intensity and compound action potential of frog sciatic nerve. J. Physiol. Pharmacol. 1966. Jun 47: 2, 341 - 9.
• Bainton, C. R. Strichartz, G.R. Concentration dependence of lidocaineinduced irreversible conduction loss in frog nerve. Anesthesiology. 1994. Sep 81: 3, 657 -67.
Bibliografía
[lJ Oakley, B. and Schafer, R. Experimental Neurobiology: A laboratory manual. The University Michigan Press. Ann Arbor 1984.
[2J Kayser, Charles. Physiologie. Editions Medicales Flammarion. 1963.
[3JAidley, D. J. The Physiology of excitable cells. Cambridge at the University Press. 1971.
42

Conformismo de la frecuencia de descarga del órgano eléctrico con la temperatura en el pez cuchillo fantasma negro (Apteronotus Albifrons)
Profesor: Arcadio Guzmán, Departamento de Ciencias Fisiológicas, Facultad de Medicina
Introducción
Regulación y conformismo Cuando un animal se enfrenta a cambios en su medio ambiente (p. ej., cambios en la temperatura del medio ambiente), puede mostrar una de las dos grandes categorías de respuesta: CONFORMISMO o REGULACIÓN. Los animales denominados conformistas son incapaces de mantener la constancia de parámetros internos con cambios en el medio externo, mientras que los organismos reguladores, utilizan mecanismos fisiológicos para mantener parámetros internos constantes, sin importar las variaciones externas, dentro de rangos apreciablemente amplios. Cuando se considera la regulación de temperatura, la clasificación más útil es la de POIQUILOTERMO y HOMEOTERMO. La mayoría de los animales acuáticos son poiquilotermos, mantienen una temperatura corporal que es igual a la temperatura del agua que los rodea. Aunque los procesos metabólicos de estos animales generan calor, por lo general se disipa rápidamente, aún en los animales grandes. Los primeros animales verdaderamente terrestres, los reptiles (serpientes, lagartos y tortugas), al igual que los peces son poiquilotermos. Los homeotermos son animales que mantienen una temperatura corporal constante a pesar de las fluctuaciones en su ambiente y la mayoría mantiene una temperatura por encima de la de sus alrededores. Entre los animales actuales sólo las aves y los mamiferos son verdaderos homeotermos.
43

Peces que generan señales eléctricas débiles
700-- ~ SOOJsec
IL~ j ,A/I ~ 1\
IvVVV\ V/V TOf\!E OlSCHA.RGE
,- BLACK GI-'OSI K~J¡FEFIS~i # (APTERONOTUS ALBIr:ROf:S)
Los peces que generan señales electricas débiles las utilizan para comunicarse y percibir el medio ambiente (electrolocalización). Dos grupos de peces de agua dulce han evolucionado independientemente con órganos que descargan electricidad: los Gymnotideos de Sudamérica y los Mormíridos en Africa. Los órganos eléctricos de los peces presentan dos orígenes. La mayoría consiste de largas columnas de electro citos, que se concentran hacia la región de la cola del pez. Los electro citos se han derivado de tejido muscular y como el músculo esquelético son controlados por neuronas motoras colinérgicas. Los órganos eléctricos derivados de tejido nervioso sólo se encuentran en una familia de los Gimnotideos: la familia Apteronotidae, estos órganos eléctricos consisten de grandes racimos de axones mielinizados que corren a lo largo del pez hasta la cola. Los peces que emiten señales eléctricas débiles detectan su propio campo eléctrico y el generado por otros peces. La electrorecepción, o la detección de corrientes eléctricas por el pez está realizada por órganos receptores especializados: la ampolla, detecta bajas frecuencias (menores a 50 Hz) y los receptores tuberosos los cuales son estimulados por altas frecuencias.
rt?': NfflWNMNVV\
44
,e ~-
,;.

Los peces que generan señales eléctricas débiles tienen dos modos de generación de señal: especies pulsátiles, producen pulsos y especies tonales, que emiten señales continuas parecidas a tonos eléctricos sinusoidales.
Frecuencia de la descarga del órgano eléctrico
Frequency (Hz) Fantasma Negro O 1000 2000 3{)OO 4000 5000
¡¡; Wo Vr..-v' ,,-, /"\ r-' I /;vi'. ¡ ¡j.!//" ..... • ~ ~1 ¡ ~'.,.W" ¡i.iJ Ir'''~ I :¡ 188 I ,....... \ ¡!f'"' I 1. ,1 r ·180 ~ """ ""
0003
1000 2000 3000 4000 5000 10SIi~! Frequency(Hz) 26grados
6000
'000
1300
1200
Ñ ~ 1100
~ t.)
ffi 1000
hl s: 900
20 22 " 26 ,. 30
TEMPERATURA (e)
La frecuencia de descarga del órgano eléctrico está controlado por un núcleo impar, colocado en la línea media del tallo cerebral, el cual en la mayoría de las especies es denominado: núcleo marcapaso. El grupo de neuronas localizado en el núcleo marcapaso descarga sincrónicamente y excita localmente neuronas de relevo que se proyectan hacia la médula espinal para efectuar sinapsis con neuronas electromotoras. Las neuronas electromotoras comandan directamente el órgano eléctrico, ya sea vía sinápsis con electrocitos o por ellas mismas constituyendo un órgano eléctrico en la forma de procesos axonales especializados.
Objetivo
Obtener la variación de la frecuencia de la descarga del órgano eléctrico del pez cuchillo fantasma negro (Apteronotus Albifrons) con la temperatura.
Materiales
45

1. Acuario
2. Circuito de control de temperatura
3. Pez cuchillo fantasma negro
4. Electrodos
5. Bomba para colocar en movimiento y filtrar el agua del acuario
6. Amplificador diferencial
7. Sistema de adquisición digital de datos
8. Programa Origin
Procedimiento
1. Se baja la temperatura del acuario y el pez, lentamente hasta llegar alrededor de los 20 grados centígrados.
2. Se efectúa la adquisición de la señal del pez durante un segundo.
3. Se ajusta el control de la temperatura a una temperatura ligeramente superior y se espera durante 15 minutos. El agua del acuario permanece circulando con la bomba.
4. Se toma otra adquisición durante otro segundo y se procede como en el paso anterior.
5. Después de haber tomado varias adquisiciones se procede a pasarla al programa Origino
"1 ...¡ IAq'~llq~III~~!'\II,1
~ 1I " l' 1, 11 iI~' l' 1\ /1 " 1\ " 1' " ¡I 1, ,1
R~ J'\\I\I\II\I'\I\/\ \1 '\I\I\/\//\¡/\I\I\I\/\I _¡Pi\U ¡Jjlu H¡~~
~I J
l' . --
lE Mf1J - -~._._:.;,..-:.: .. _-'--- .. -..--- ...... -.
46

6. Para cada una de las adquisiciones se procede a evaluar la transformada rápida de Fourier y a obtener la frecuencia fundamental del conjunto de datos evaluados.
7. Finalmente, se realiza una gráfica con las distintas frecuencias fundamentales obtenidas para distintas temperaturas entre 20 y 30 grados centígrados.
Resultados
_(Hz! o 1000 .... 311CO _ 5000 ....,
t!~;~;~~'~'~'~;
i .. =
lllO1l 10lI0 II!XI ~1lOIl liOOO _
,,-,s ~(Hz!GHOETI7o_
47
I~~_~~ ~ d~ l.,lOi -v- ~-r
0.010: j ji" i
OOOill
I :~ j
0.0G:!; ,
~+Q~~I~~~~~~~a~~.~~,~-·~,~
""'-(kzl

Contracción Isométrica del Músculo Gastronemio de Sapo
Profesor: Arcadio Guzmán, Departamento de Ciencias Fisiológicas, Facultad de Medicina
Objetivos
• Definir las características fundamentales de la sacudida y tétano isométrico.
• Obtener la relación entre las fuerzas pasiva y activa con la longitud muscular.
• Mostrar cualitativamente el fenómeno de la fatiga muscular.
Marco teórico El registro de la sacudida muscular nos permite establecer la existencia de una estimulación umbral y establecer que con el aumento de la amplitud del estímulo se incrementa la fuerza de la sacudida hasta un punto determinado por el reclutamiento total de las fibras. El desarrollo del tétano isométrico nos muestra el efecto de la suma mecánica, de la fuerza de las sacudidas musculares y nos permite establecer su relación con la frecuencia de estimulación. Al estirar el músculo se va incrementando la fuerza pasiva que este realiza. La razón entre la fuerza pasiva y estiramiento, definen la característica del componente elástico en paralelo. Si para algunos valores de fuerza pasiva del músculo, activamos éste para obtener una sacudida muscular, la amplitud de las sacudidas será una función del nivel de fuerza pasiva, es decir de la longitud muscular. Al representar estos valores de fuerza de sacudidas contra longitud, obtenemos la variación de la fuerza activa con la longitud muscular.
La fatiga muscular es un fenómeno reversible, que ocurre al agotar alguna etapa del acoplamiento excitación-contracción. Se ha demostrado que una fibra muscular fatigada, puede producir una contractura por cafeína, mediante la salida de calcio del retículo sarcoplásmico. La fatiga muscular que se observa exclusivamente en el músculo no es la misma fatiga que se observa
48
en el animal completo, debido a que la unión neuromuscular se fatiga antes que el propio músculo y además podría existir fatiga psicológica.
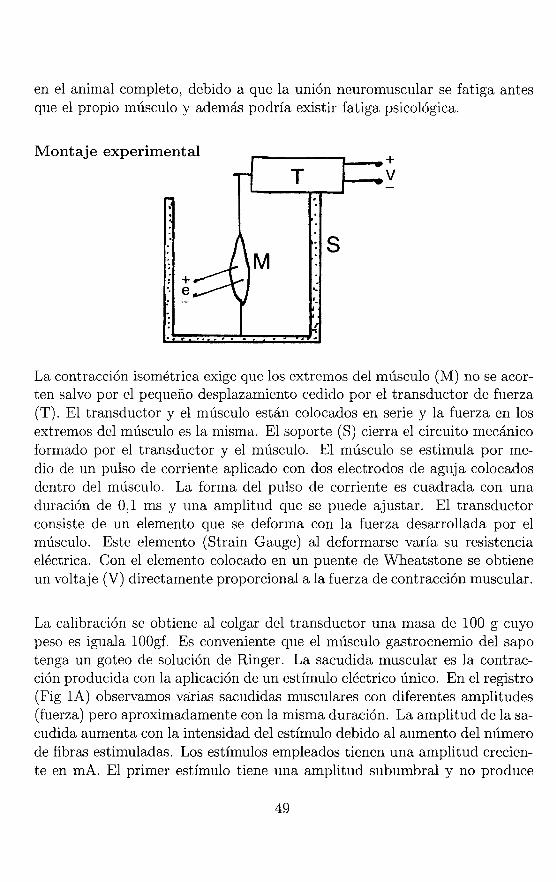
Montaje experimental
+ e
T
M
+ V
. .
S
... • .
La contracción isométrica exige que los extremos del músculo (M) no se acorten salvo por el pequeño desplazamiento cedido por el transductor de fuerza (T). El transductor y el músculo están colocados en serie y la fuerza en los extremos del músculo es la misma. El soporte (8) cierra el circuito mecánico formado por el transductor y el músculo. El músculo se estimula por medio de un pulso de corriente aplicado con dos electrodos de aguja colocados dentro del músculo. La forma del pulso de corriente es cuadrada con una duración de 0,1 ms y una amplitud que se puede ajustar. El transductor consiste de un elemento que se deforma con la fuerza desarrollada por el músculo. Este elemento (8train Gauge) al deformarse varía su resistencia eléctrica. Con el elemento colocado en un puente de Wheatstone se obtiene un voltaje (V) directamente proporcional a la fuerza de contracción muscular.
La calibración se obtiene al colgar del transductor una masa de 100 g cuyo peso es iguala 100gf. Es conveniente que el músculo gastrocnemio del sapo tenga un goteo de solución de Ringer. La sacudida muscular es la contracción producida con la aplicación de un estímulo eléctrico único. En el registro (Fig lA) observamos varias sacudidas musculares con diferentes amplitudes (fuerza) pero aproximadamente con la misma duración. La amplitud de la sacudida aumenta con la intensidad del estímulo debido al aumento del número de fibras estimuladas. Los estímulos empleados tienen una amplitud creciente en mA. El primer estímulo tiene una amplitud subumbral y no produce
49

respuesta, el resto de estímulos tienen amplitudes supraumbrales. Todos los estímulos aplicados tienen la misma duración: 0,1 ms. La duración de la sacudida depende del tipo de músculo (rojo, blanco o intermedio) y de la temperatura.
A
-;
..
'-'\ ,'\ \ \ \\
"~""", .. _--,~ "_1_) '.. té ... 'M'
B
~~~\ 1/ _w4-\ ;" V<'~~*'"''"'~~
I ,,~,Jff" ,l"\\ Ji ,.
i ! ~ 10 ;', v o "-V''ó ..... , .. ',,.,/',, ........ '''v,...,.. ... '\
~"y,..i;~:-,~,,'.,,"~vv,.~ , \~ o '::-..(' ...... ~~"\I::::i.,).,~.<:::~,,\.......:\~,_~..:l' .. ' .... ____ _
-,.., .. ..... 1"
Figura 16: Registros de contracciones musculares. A: Sacudidas musculares con diferentes amplitudes de estimulación. B: Contracciones tetánicas isométricas.
La contracción tetánica isométrica se produce cuando se aplican muchos estímulos eléctricos consecutivos. En el registro (Fig lB) observamos varias contracciones isométricas tetánicas no fusionadas. La amplitud de la fuerza en la contracción tetánica depende de la frecuencia de estimulación. La contracción tetánica que desarrolla más fuerza dentro del grupo de las mostradas en el registro es fusionada y presenta el fenómeno de "fatiga". En la cuarta contracción isométrica no fusionada se observa una falla en el circuito del estimulador que omita la estimulación.
Cuando el músculo no estimulado, sufre un estiramientp, desarrolla una fuerza denominada pasiva, porque no corresponde a una activación del componente contráctil, sino al estiramiento del material de sostén estructural de la
50

máquina contráctil. Cuando un músculo es estimulado a diferentes longitudes musculares, se encuentra que la fuerza desarrollada, denominada fuerza activa, varía con la longitud, presentando un valor máximo a la longitud óptima.
i J
lOO)
J~ 1
:1
Figura 17: Variación de fuerza pasiva y activa con la longitud.
En el registro (Fig 2) se observan dos fuerzas desarrolladas por el músculo, una pasiva que varía con el estiramiento de una forma continua y creciente y las fuerzas de las sacudidas musculares con el mismo estímulo aplicado al músculo a diferentes longitudes. En el registro de la fuerza no se pueden separar las dos fuerzas, si se estira el músculo siempre va a existir una fuerza pasiva, sobre la cual se superpone la fuerza activa de la contracción.
En el registro de la fuerza pasiva se observa que existe una fuerza que desaparece una vez se ha estirado el músculo, esta fuerza es debida a la velocidad de estiramiento pasivo del músculo. La variación de la fuerza activa con la longitud muscular es explicado por la Teoría de los Filamentos Deslizantes. En el registro observamos dos contracciones isométricas tetánicas fusionadas. La primera desarrolla fuerza pero no la sostiene, sino que ésta desciende inicialmente rápido y luego lentamente, luego se termina la estimulación y la
51

fuerza desciende abruptamente al valor inicial de reposo. Después de alrededor de 25 segundos se produce la segunda contracción tetánica fusionada y se alcanza una fuerza intermedia que una vez más no se sostiene y que desciende más rápidamente que la primera.
En los dos registros (Fig 3) observamos el fenómeno de la fatiga muscular. En un músculo existen distintos tipos de fibras musculares. En los dos extremos encontramos fibras resistentes a la fatiga y otras fácilmente fatigables, por esta razón la fuerza disminuye rápidamente al comienzo de la fatiga, para luego permanecer disminuyendo muy lentamente.
~.-.
~ ,aoj ,
ji : 1
, I
~ l' ,., 'n~"too" .• • y ~,~ -r ,~....--, , '50 TI_f.' •
",,'
'75 , I I
ji 10'1
''''1 1
, .. 11
li 7'1!
r'" 25 50 '" ,o.
TIme(.,
,-, 121 150
""M
.~ \..
, .. hin
Figura 18: Fatiga muscular. Fatiga y refatiga.
52

Periodo de Latencia de la Contracción Muscular esquelética
Profesor: Arcadio Guzmán, Departamento de Ciencias Fisiológicas, Facultad de Medicina
Objetivo Mostrar la relación temporal entre el electromiograma y la contracción muscular.
Teoría
Período de Latencia
El periodo de latencia es el tiempo que existe entre el inicio del electro miograma y el inicio de la contracción. Durante el periodo de latencia ocurre el acople electromecánico del músculo. La llegada del potencial de acción a los terminales de los axones motores causa la liberación de acetilcolina. En la placa motora el neurotransmisor abre los canales que dejan pasar cationes, lo que produce una corriente despolarizante, la cual desencadena un potencial de acción todo - nada en el sarcolema. El potencial de acción se propaga desde la placa motora en ambas direcciones. Luego se pone en marcha una secuencia de acontecimientos que conducen a la contracción. El mecanismo contráctil responde con una latencia de varios milisegundos.
El electromiograma El electromiograma es el registro de la actividad eléctrica que produce la contracción muscular. En un músculo al cual se le hace desarrollar fuerza voluntariamente, genera una actividad eléctrica continua, porque el nervio motor descarga ráfagas de potenciales de acción sobre las fibras musculares que inerva ( unidad motora). El potencial eléctrico que se desarrolla en una contracción única de todas las fibras de la unidad motora, se prolonga hasta unos 12 ms. La mayoría de los potenciales de la unidad motora tienen una amplitud alrededor de 0,5 m V. Cuando se registran estos potenciales, aparece una espiga aguda que la mayoría de las veces es trifásica, pero que también puede tener una forma más compleja.
53

Sacudida muscular isométrica La contracción iso métrica se efectúa con los extremos del músculo fijos para impedir su acortamiento. El nombre de la contracción isométrica generada con un sólo estímulo recibe el nombre de sacudida (Twitch). La amplitud de la sacudida depende de la amplitud del estímulo y su duración depende del tipo de músculo (rojo o blanco) y de la temperatura.
Suma mecánica
Si antes de que termina una sacudida muscular, se ha iniciado otra, la fuerza contráctil generada por la primera se suma con la fuerza generada por la segunda sacudida. Si la frecuencia de aplicación de los estímulos es alta, se puede lograr una contracción conocida como tetánica isométrica fusionada, porque la subida de la fuerza se desarrolla de una manera perfectamente continua hasta un valor de fuerza varias veces superior a la de la sacudida.
Materiales y métodos
Complejo instrumental
El complejo instrumental consta de un sistema de adquisición de datos, el transductor de fuerza, el amplificador diferencial, el estimulador de corriente, los electrodos para estimulación y registro y la preparación biológica (un sapo Bufo). En la figura No. 1 se muestra el montaje experimental.
Resultados
Cada sacudida muscular está precedida por el respectivo electromiograma. El electromiograma tiene una duración aproximada de 50 ms y una amplitud pico a pico aproximada de 4 m V, el cual corresponde a la activación simultánea de todas las unidades motoras que conforman el músculo. El estímulo que se aplicó al nervio ciático fue de 0,1 mA y 0,1 ms de duración y correspondió a
54

Sistema de adquisición de datos
Señal Fuerza
Sincronismo
Estimulador
Figura 19: Esquema del montaje experimental.
un estímulo supramáximo. La sacudida muscular del músculo gastrocnemio de sapo tiene una duración aproximada de 287 ms y una amplitud de 0,08 kgf.
. --ay o ....
. ...... ;~\¡',- ..•........... __ ... .
...... O."
D·
.<l. -~_. -,.--~.--. r , r-- -,_·_,.---~~~-----r--1---.------~
250 soo 750 T1fM (1M) 8w 1/1
Figura 20: Las sacudidas musculares y los respectivos electromiogramas.
La marca vertical indica el momento en que se estimuló el nervio ciático con un pulso de corriente de amplitud de 0,1 mA y una duración de 0,1 ms.
55

Entre el estímulo al nervio ciático y el comienzo del electromiograma existe un intervalo de 3,11 ms los cuales corresponden al tiempo de conducción del potencial de acción en el nervio, el retardo sináptico y el inicio de la despolarización de las fibras musculares. Entre el estímulo al nervio ciático y la iniciación de la actividad muscular existe un intervalo de 12,2 ms. El periodo de latencia entre el comienzo del electromiograma y el comienzo de la actividad muscular tiene una duración de 9,09 ms. Durante el periodo de latencia ocurre el acople excitación-contracción.
0.1
~> o
..0.004
:;:1
..0.01 ~~r-~-'--
Ó -----r-~--T~ ~~'~r-'-'--,--~ ..... -'¡-"-'~ .,.---..........,...--.,.............~---r-·--
B ~ n TIme (_1 8w 111
Figura 21: Periodo de latencia.
Se han aplicado tres pulsos de corriente al nervio ciático separados arbitrariamente. Cada nueva sacudida está precedida por el electromiograma correspondiente a la activación de las unidades motoras. Antes de que cada sacudida muscular termine, se ha aplicado un nuevo estímulo, lo cual hace que se inicie una nueva sacudida, cuya fuerza se suma con la anterior. Este registro de fuerza corresponde a una contracción tetánica fusionada.
56

o ...
.0.08
o
.y
~~~------
2110 IlOO TIme (nas)
Figura 22: Suma tetánica.
57
710 Sw1/1

Seguimiento del Ciclo Estral en Ratones
Profesores: Gladys Fajardo, PhD y Clara Spinel, PhD, Departamento de Biología, Facultad de Ciencias.
Introducción Los caracteres sexuales primarios son las gónadas, testículos en los machos y ovarios en las hembras. Los órganos sexuales secundarios o accesorios son las glándulas y los sistemas de conductos que intervienen en la transmisión de los gametos o de los cigotos en el desarrollo. Las gónadas no sólo permiten proliferar los gametos sino que también secretan hormonas que controlan el estado funcional de los órganos sexuales secundarios y en cierto grado influyen sobre los fenómenos psicobiológicos que intervienen en las reacciones del apareamiento. Los caracteres sexuales secundarios son especializaciones más o menos externas que no son esenciales para la proliferación ni para el movimiento de las células germinales, sino que tienen relación con el apareamiento, el nacimiento y la nutrición de los críos.
Ciclo estral: En la mayoría de los vertebrados, con excepción de los primates, la receptividad sexual de las hembras se restringe a períodos llamados estro o celo. Durante este tiempo la hembra está fisiológicamente dispuesta a recibir al macho y se presentan cambios estructurales en los órganos sexuales de la hembra. Los llamados animales monoestrales completan anualmente sólo un ciclo de estro y un largo período de anestro separa las épocas de celo. Las formas poliestrales completan anualmente dos o más ciclos si no son interrumpidos por una gestación (o preñez). El fenómeno dominante en el ciclo estral es la ovulación.
La ovulación depende de la liberación de gonadotropinas que parten de la hipófisis anterior, cuya liberación está controlada por el hipotálamo. Posiblemente en todas las formas, el sistema nervioso central controla el proceso ovulatorio. El ciclo menstrual característico de los primates es fundamentalmente comparable al ciclo estral, pero existen entre éstos dos diferencias importantes: en las hembras de los primates no existe apogeo ineqUÍVoco de aumento de la actividad receptora, la hembra puede aceptar al macho en cualquier período y la ruptura del endometrio al final del ciclo va acom-
58

pañada de pérdida de sangre.
Los frotis vaginales son muy útiles para la estimación del estadio del ciclo estral en mamiferos o menstrual en la mujer, excepto en vacas. En la mujer, el frotis vaginal se realiza periódicamente para detectar cambios anormales en la morfología celular y de este modo prevenir a tiempo patologías como cáncer uterino.
Las etapas del ciclo estral de acuerdo a la morfología celular característica de la mitad de cada etapa, son las siguientes:
Diestro (Anestrus), se observa gran cantidad de leucocitos, puede encontrarse pocas células epiteliales con núcleo y no cornificadas. Puede durar dos días o algo más. Proestro, se encuentra gran número de células pequeñas, redondas y con núcleo picnótico (visible) provenientes del epitelio vaginal, y muchos glóbulos rojos, muy pocos leucocitos y algunos detritos. Dura aproximadamente un día. Crecimiento del folículo y una gran secreción de estrógenos.
Estro, fuerte descamación de células epiteliales, apareciendo como células cornificadas más superiores y sin núcleo, muchos glóbulos rojos y no leucocitos (si aparecen se está al final de esta etapa). Es el período de predisposición (ovulación), dura casi 12 horas. Se finaliza con la ovulación y por ende disminución fuerte de estrógenos, se inicia el cuerpo lúteo.
Metaestro, también de casi 12 horas, puede verse una mezcla de células correspondientes a las de las etapas a, b y c, muchísimos leucocitos, y predominan las células no cornificadas y pocas cornificadas.
Objetivos Aprender a realizar cirugías y seguimiento pos-operatorios en animales de laboratorio. Observar las modificaciones del epitelio vaginal controlado por la hormonas sexuales. Determinar el efecto de la ausencia del ciclo estral.
59

Materiales
• 10 ratones hembras,
• algodón, anestésico (Ketamina y Xilazina),
• tablas de disección,
• jeringas de 1 mI, suero fisiológico (ringer) para mamífero,
• penicilina en polvo o antibióticos de laboratorio,
• tintura de yodo o isodine,
• láminas porta- y cubreobjetos,
• capilares sellados para frotis vaginal,
• Material de disección fina (tijeras de disección pequeñas punta aguda, otra punta roma, pinzas de disección punta fina y otra con punta roma),
• hilo y agujas para suturar,
• colorante azul de metileno,
• mechero,
• Cinta de enmascarar,
• marcadores indelebles,
• frascos goteros,
• Gelatina sin sabor
• Fenol (jugo de limón).
• Lámpara de luz fría.
60

Método
Como son animales de laboratorio que se van a tener durante dos semana, es indispensable que se organice con todos los estudiantes la vigilancia diaria de los animales, se recomienda que el suministro de alimento y del agua sea ad limitum, Adicionalmente, establecer un programa para la limpieza de las jaulas, la toma de muestras y preparación de los frotis vaginales. Durante el fin de semana se pueden mantener los ratones y realizar los frotis en sus casas.
Grupo 1. Ovariectomizadas
1. Se toman tres ratones hembras y se procede a su castración u ovariectomía, de la manera descrita a continuación:
2. Dormir la ratona con Ketamina 90mg/kg y Xilazina 15 mg/kg, para un peso de 30 mg tomar 0,6 mI de ketamina con la jeringa de 1ml y en la misma jeringa 0,3 mI de xilazina. Se inyecta intraperitoneal y se espera a que la ratona muestre los signos de insensibilidad a la presión caudal.
3. Rociar con alcohol al 75 % y afeitar la piel en la parte dorso-lumbar a un lado de la línea media. Se practica en la piel un corte recto de aproximadamente 1 cm de largo. Luego se abre y levanta la piel, lográndose observar en el último tercio del tronco una mancha más clara sobre los riñones que corresponde al tejido graso, el cual sirve de sostén a los ovarios.
4. Realizar a este nivel un corte con el bisturí y extraer delicadamente la masa grasa con una pinza fuera del cuerpo de la rata. Así saldrán en el espesor de la grasa el ovario y el oviducto.
5. Con ayuda de una pinza fina pasar el hilo de sutura debajo de la unión ovario-oviducto y amarrar con firmeza para interrumpir la circulación entre la gónada y el conducto.
6. Retirar el ovario con la ayuda de una tijera fina. De manera cuidadosa, con la pinza volver a colocar el tejido en su sitio. Si la operación ha sido bien realizada no debe fluir sangre de la superficie seccionada.
61

7. Al final, añadir antibióticos en el sitio de la disección y suturar el músculo y la piel. Aplicar tintura de yodo o isodine y dejar el animal en observación para que se recupere. Es conveniente esperar a que se despierten y ponerles una lámpara para dar calor y favorecer la recuperación postoperatoria de las ratonas
8. Pasada una semana de la operación, realizar frotis vaginales de igual manera que con los otros dos grupos experimentales (grupo control y grupo gestante).
Grupo 11. Gestantes En dos cajas de ratones grandes, colocar por caja dos hembras y un macho a las 8 pm. Al día siguiente separar el macho de las hembras, y revisar las hembras muy temprano en la mañana, mejor antes de las 7 am, para observar el tapón vitelina, una estructura blanquecina que tapona la entrada de la vagina, esta estructura es espacio temporal. Sí no se logra observar anotarlo en los resultados. Aquí en el laboratorio, luego de este seguimiento, realizar la toma de células a la salida de la vagina en forma muy delicada y realizar el frotis. Este grupo se inicia el jueves en la noche para que la toma de células vaginales se realice al día siguiente y se comience la parte experimental a la par que los otros dos grupos.
Grupo 111. Control Tres ratas normales. Durante siete días (de viernes a viernes), realizar diariamente en los tres grupos de las ratonas un frotis vaginal, a fin de observar los cambios citológicos de la vagina. Comparar los cambios en la forma celular debidos a la falta de la estrógenos en las ovariectomizadas y exceso de secreción de estrógenos en las gestantes con las del grupo de secreción de estrógeno s control que presenta el ciclo estral normal.
¿Cómo se efectúa el frotis vaginal? Tomar la rata por ambas orejas para inmovilizarla y voltearla de manera que exponga la parte ventral, sujetando la cola del animal entre los dedos meñique y anular. Introducir un aza o copita finos y hacerlos girar en el interior de la vagina. En una gota de suero fisiológico colada sobre una lámina portaobjetos, apoyar el asa o copita para extender la muestra tomada. Proceder
62

a realizar una tinción con azul de metileno, dejar secar, añadir la gelatina sobre el frotis y cubrir con una laminilla. Marcar la lámina y posteriormente montarla al microscopio con el fin de observar el tipo de células que se encuentren y con base en ello identificar: en el grupo III control, la etapa del ciclo a la que corresponda la muestra de cada día. Comparar los resultados de los grupos 1 y II con los del control. Es decir, si se analizan las muestras de los frotis correspondientes a los siete días de duración del experimento, se podrá apreciar los cambios provocados por acción o ausencia hormonal en el epitelio de la vagina.
Consignar los resultados de las observaciones según las condiciones del animal manipulado, esto es:
Rata castrada Rata gestante Rata control Metaestro Estro Proestro Diestro
Bibliografía [1] Guraya 8.8. 1985 Biology of ovarian follicles in mammals. Ed. 8pringer-Verlag, Heidelberg.
[2] Hanke W. 1982. Biologie der hormona. Quelle & Meyer, Heidelberg.
[3] Turner D Endocrinología General. Ed. Interamericana. Colombia
63

Variantes cíclicas de la temperatura corporal
Profesor: Gladys Fajardo, Departamento de Biología, Facultad de Ciencias
El experimento dura un mes entero.
Los objetivos que se persiguen son:
• Mostrar la variación de temperaturas basales en las mujeres, tomando como testigos los varones del grupo.
• Mostrar también el ciclo diario de temperatura, obteniendo bastantes datos para tratarlos estadísticamente y comprender mejor el significado de lo que es "normal" en biología.
Cada estudiante debe tener un termómetro: Cada vez que estos se usen se lavaran con jabón yagua y se enjuagarán. Pueden esterilizarse dejándolos varias horas en una solución germicida y enjuagándolos luego con agua. Nunca deben lavarse con agua caliente ni hervirse. Antes de cada medición de temperatura el termómetro debe sacudirse hasta que la columna de mercurio se encuentre debajo de la señal de 34. 4°C. Para medir la temperatura bucal se coloca el bulbo del termómetro debajo de la lengua, se cierran los labios sobre el termómetro y se mantiene la boca cerrada. No debe hablarse. El sujeto se sienta o se acuesta tranquilamente y respira normalmente durante cinco minutos cuando menos. Luego se saca el termómetro y se lee.
Variaciones mensuales de temperatura basal. El termómetro, listo para ser usado, se coloca sobre la mesa de noche para poder tomar la temperatura antes de levantarse. Cada mañana, durante un mes, después de despertarse, pero antes de levantarse, tome su temperatura bucal. Anote sobre la hoja de resultados la fecha, la hora, la temperatura obtenida, las horas de sueño y las horas transcurridas desde la última ingestión de alimentos. Se construye una gráfica colocando en las abscisas la fecha y en las ordenadas la temperatura basal. Al finalizar el mes se examinarán todas las gráficas construidas por los integrantes del grupo, buscando fluctuaciones de la temperatura basal, para establecer si existe o no en las mujeres una variación de
64

temperatura basal que no sea al azar.
Variaciones de Temperaturas Basales de una Población "Normal": El profesor señalará un día, al azar y concentrará todas las temperaturas basales obtenidas ese día. Se hará el análisis estadístico elemental para establecer la media, la mediana, los límites y la desviación estándar con las temperaturas basales del grupo entero. Se analizará el significado de estos factores.
Ciclo diurno de temperatura. Esta parte del experimento exige colaboración de un compañero de dormitorio o de alguna persona de la familia. Se procede así: se toma una temperatura basal poco tiempo después de despertar, pero antes de levantarse. Se anota la hora y la temperatura, así como cualquier comentario acerca de actividad, excitación, emoción e ingestión de alimentos. También debe ser anotada la temperatura ambiental. Se continúa tomando temperaturas bucales durante todo el día, a intervalos de media hora, anotando estos datos sobre la hoja de resultados. En la noche debe continuarse tomando la temperatura a intervalos de dos horas: el estudiante debe pedir a su "ayudante" que le tome la temperatura mientras duerme y que anote la hora y la lectura hecha. A la mañana siguiente se toma una lectura final de la temperatura basal. Reuniendo estos resultados se construye una curva con la hora del día en las abscisas y la temperatura en las ordenadas. A partir de las observaciones anotadas, señale en la gráfica condiciones como la comida, el ejercicio u otros factores suceptibles de modificar los resultados.
Hoja de resultados
Nombre ___________ _ Fecha ___________ _ Edad ___ _ Sexo ____ _ Peso ____ _ Estatura ___ _
Mediciones diarias de temperatura basal
Fecha Hora Temperatura (oC) Horas de sueño Horas comida Observaciones
65

Variaciones de temperatura en el día
Fecha ______ _
Hora I Temperatura bucal I Temperatura ambiente I Observaciones
Variaciones de temperatura basal en una población normal
Resultados del Análisis
Número de individuos ______________________ _ Temperatura basal media ___________________ _ Desviación estándar ______________________ _ Temperatura basal mediana ___________________ _ Límites de temperatura basales __________________ _
Grafique
1. Temperatura (oC) para las mediciones basales diarias contra día del mes.
2. B)Temperatura ( OC) contra hora del día (A.M, P.M.)
66

Temperatura para las mediciones basales diarias 37.7
37.2
37.0
36.5
36.0 Día del mes
67
Variaciones Cíclicas de la Temperatura Corporal
Nombre ________ _ Grupo. ________ ___ Fech.aB ________ __
Hoja de resultados
Sujeto ____ _ Edad ___ Sexo___ Peso___ EstaturaB ___ _
Mediciones diarias de temperatura basal
I Fecha I Hora I T ('C) I Horas sueño I Horas desde la comida I Observariones
Variaciones de temperatura basal en una población normal Resultados del análisis
Número de individuos ______ _ Temperatura basal media ____ _ Desviación estándar _______ _ Temperatura basal mediana ___ _ Límites de temperatura basales __ _
Variaciones de la temperatura en el día
Fecha _____ _
Hora T bucal TA Observaciones
.-

temperatura basal que no sea al azar.
Variaciones de Temperaturas Basales de una Población "Normal": El profesor señalará un día, al azar y concentrará todas las temperaturas basales obtenidas ese día. Se hará el análisis estadístico elemental para establecer la media, la mediana, los límites y la desviación estándar con las temperaturas basales del grupo entero. Se analizará el significado de estos factores.
Ciclo diurno de temperatura. Esta parte del experimento exige colaboración de un compañero de dormitorio o de alguna persona de la familia. Se procede así: se toma una temperatura basal poco tiempo después de despertar, pero antes de levantarse. Se anota la hora y la temperatura, así como cualquier comentario acerca de actividad, excitación, emoción e ingestión de alimentos. También debe ser anotada la temperatura ambiental. Se continúa tomando temperaturas bucales durante todo el día, a intervalos de media hora, anotando estos datos sobre la hoja de resultados. En la noche debe continuarse tomando la temperatura a intervalos de dos horas: el estudiante debe pedir a su "ayudante" que le tome la temperatura mientras duerme y que anote la hora y la lectura hecha. A la mañana siguiente se toma una lectura final de la temperatura basal. Reuniendo estos resultados se construye una curva con la hora del día en las abscisas y la temperatura en las ordenadas. A partir de las observaciones anotadas, señale en la gráfica condiciones como la comida, el ejercicio u otros factores suceptibles de modificar los resultados.
Hoja de resultados
Nombre ____________ _ Fecha ___________ _
Edad ___ _ Sexo ___ _ Peso ____ _ Estatura ___ _
Mediciones diarias de temperatura basal
Fecha Hora Temperatura (oC) Horas de sueño Horas comida Observaciones
65

Variaciones de temperatura en el día
Fecha ______ _
Hora I Temperatura bucal I Temperatura ambiente I Observaciones
Variaciones de temperatura basal en una población normal
Resultados del Análisis
Número de individuos _____________________ _ Temperatura basal media ___________________ _ Desviación estándar _____________________ _ Temperatura basal mediana __________________ _ Límites de temperatura basales _________________ _
Grafique
1. Temperatura (OC) para las mediciones basales diarias contra día del mes.
2. B)Temperatura ( OC) contra hora del día (A.M, P.M.)
66

Temperatura para las mediciones basales diarias 37.7
37.2
37.0
36.5
36.0 ,
Dla del mes
67

Variaciones Cíclicas de la Temperatura Corporal
Nombre: ________ _ GrupoJ _______ ___ Fechaa ______________ _
Hoja de resultados
Sujeto ____ _ Edad ___ Sexo___ Peso___ Estaturtta~ __ _
Mediciones diarias de temperatura basal
Fecha Hora T ('C) Horas sueño Horas desde la comida Observaciones
----
Variaciones de temperatura basal en una población normal Resultados del análisis
Número de individuos ______ _ Temperatura basal media ____ _ Desviación estándar _______ _ Temperatura basal mediana ___ _ Límites de temperatura basales __ _
Variaciones de la temperatura en el día
Fecha _____ _
Hora I T bucal I TA 1 Observaciones

La (olección Notas de Clase de la Facultad de Ciencias, es un espacio abierto a los profesore5 para publicar su experiencia docente,
recop lada en el tiempo a través de notas escritas. Se espera que las ¡'lterrelaciones autor- colegas y profesores-estudiantes sean la mejor Manera para que estas notas se depuren, a fin de que en un futuro
mediato adquieran carátula de texto.
Comité Editorial
Facultad de Ciencias