
Fish Rhabdovirus Replication Non-Piscine Cell Culture ... · Fish Rhabdovirus Replication in...
10
INFECTION ANI) IMMUNITY, JUIV 1974/ p. 180-188 Copyright © 1974 American Society for Micro biology \'ol. 10. No. 1 Printed in U.S.A. Fish Rhabdovirus Replication in Non-Piscine Cell Culture: New System for the Study of Rhabdovirus-Cell Interaction in Which the Virus and Cell Have Different Temperature Optima H FRED CLARK AND ELVIRA Z. SORIANO The Wistar Institute of AnatomY and Biology. Philadelphia, PennsYlvania 19104 Received for publication 25 Februarv 1974 The replication of three rhabdoviruses associated with diseases of f'ish has been demonstrated in cells of' continuously cultivated non-piscine cell lines. Spring viremia of carp (SVC) virus and the salmonid f'ish viruses, Egtved and infectious hematopoietic necrosis virus, all replicated in mammalian WI-38 (human diploid cell strain) and BHK/21 cells and in cells of one or more reptilian cell lines at the temperatures commonly used to propagate these viruses in fish cells. The infections were cytopathic: SVC virus plaque assays may be performed in several types of' mammalian cell culture. "Autointerf'erence" apparently mediated by abortive "T" particle formation was observed during serial nondiluted passages of SVC virus in BHK/21 and THI cells, but not in RTG-2 or WI-38 cells. Optimal temperatures for replication of SVC and Egtved viruses in BHK/21 cells were identical to those determined in poikilothermic vertebrate cell cultures. However, these viruses replicated relatively more efficiently at suboptimal temperatures in "cold-blooded" vertebrate cells than in the hamster cells. Studies of' [3H]uridine incorporation into uninfected BHK/21 cells incubated at different temperatures revealed that [3H Juridine uptake is sharply reduced at temperatures below 24.5 C. Growth curve studies of SVC virus in BHK/21 cells incubated at 23 C revealed that a clear-cut large excess of' virus-induced [3H ]uridine incorporation could be demonstrated in the absence of' actinomycin D. Actinomycin D treatment (1 ,ug/ml) led to efficient inhibition of control cell [3H]uridine uptake, but also markedly reduced the total counts per minute of' virus-induced [3H]uridine uptake, without depressing the yield of released infectious virus. Actinomycin D added to SVC virus-infected BHK/21 cell cultures at concentra- tions as low as 0.01 Ag/ml caused a significant decrease in the level of' virus-induced [3H]uridine uptake, despite the fact that this concentration is insufficient to efficiently suppress "background" cellular [3H ]uridine incorpora- tion. Four different rhabdoviruses have been iso- lated from fish; each has been associated with economically important disease in its natural host. Egtved (22, 29) and infectious hematopoi- etic necrosis (IHN) (1) viruses have been iso- lated from salmonid fish, from Europe, and from the United States, respectively. Spring viremia of carp (SVC) virus has been found associated with disease (infectious dropsy dis- ease complex) (16) that is widespread in Euro- pean carp (17), and a fourth rhabdovirus has been recovered from Dutch pike afflicted with "red disease" (13). These four viruses may be distinguished from one another on the basis of their host range in vivo or in cell culture, plaque morphology in piscine cell culture (28), temper- ature limits for growth in cell culture, or anti- genic composition (23). In vitro propagation of each of' these fish rhabdoviruses has been reported only in f'ish cell culture. We have also investigated the cultiva- bility of SVC, IHN, and Egtved viruses in several types of continuously cultivated cells of mammalian or reptilian origin. The finding that many non-fish cell types do support f'ish rhab- dovirus growth has been exploited in studies of' the influence of diverse types of' host cells on the temperature requirements for virus replication. The demonstration of virus-induced ribonucleic acid (RNA) in amounts greatly in excess of' control cell RNA concentrations is described in a system comprised of hamster cells infected 180 on February 11, 2019 by guest http://iai.asm.org/ Downloaded from on February 11, 2019 by guest http://iai.asm.org/ Downloaded from on February 11, 2019 by guest http://iai.asm.org/ Downloaded from
Transcript of Fish Rhabdovirus Replication Non-Piscine Cell Culture ... · Fish Rhabdovirus Replication in...

INFECTION ANI) IMMUNITY, JUIV 1974/ p. 180-188Copyright © 1974 American Society for Microbiology
\'ol. 10. No. 1Printed in U.S.A.
Fish Rhabdovirus Replication in Non-Piscine Cell Culture:New System for the Study of Rhabdovirus-Cell Interaction inWhich the Virus and Cell Have Different Temperature Optima
H FRED CLARK AND ELVIRA Z. SORIANOThe Wistar Institute of AnatomY and Biology. Philadelphia, PennsYlvania 19104
Received for publication 25 Februarv 1974
The replication of three rhabdoviruses associated with diseases of f'ish has beendemonstrated in cells of' continuously cultivated non-piscine cell lines. Springviremia of carp (SVC) virus and the salmonid f'ish viruses, Egtved and infectioushematopoietic necrosis virus, all replicated in mammalian WI-38 (human diploidcell strain) and BHK/21 cells and in cells of one or more reptilian cell lines at thetemperatures commonly used to propagate these viruses in fish cells. Theinfections were cytopathic: SVC virus plaque assays may be performed in severaltypes of' mammalian cell culture. "Autointerf'erence" apparently mediated byabortive "T" particle formation was observed during serial nondiluted passagesof SVC virus in BHK/21 and THI cells, but not in RTG-2 or WI-38 cells. Optimaltemperatures for replication of SVC and Egtved viruses in BHK/21 cells wereidentical to those determined in poikilothermic vertebrate cell cultures. However,these viruses replicated relatively more efficiently at suboptimal temperatures in"cold-blooded" vertebrate cells than in the hamster cells. Studies of' [3H]uridineincorporation into uninfected BHK/21 cells incubated at different temperaturesrevealed that [3H Juridine uptake is sharply reduced at temperatures below24.5 C. Growth curve studies of SVC virus in BHK/21 cells incubated at 23 Crevealed that a clear-cut large excess of' virus-induced [3H ]uridine incorporationcould be demonstrated in the absence of' actinomycin D. Actinomycin Dtreatment (1 ,ug/ml) led to efficient inhibition of control cell [3H]uridine uptake,but also markedly reduced the total counts per minute of' virus-induced[3H]uridine uptake, without depressing the yield of released infectious virus.Actinomycin D added to SVC virus-infected BHK/21 cell cultures at concentra-tions as low as 0.01 Ag/ml caused a significant decrease in the level of'virus-induced [3H]uridine uptake, despite the fact that this concentration isinsufficient to efficiently suppress "background" cellular [3H ]uridine incorpora-tion.
Four different rhabdoviruses have been iso-lated from fish; each has been associated witheconomically important disease in its naturalhost. Egtved (22, 29) and infectious hematopoi-etic necrosis (IHN) (1) viruses have been iso-lated from salmonid fish, from Europe, andfrom the United States, respectively. Springviremia of carp (SVC) virus has been foundassociated with disease (infectious dropsy dis-ease complex) (16) that is widespread in Euro-pean carp (17), and a fourth rhabdovirus hasbeen recovered from Dutch pike afflicted with"red disease" (13). These four viruses may bedistinguished from one another on the basis oftheir host range in vivo or in cell culture, plaquemorphology in piscine cell culture (28), temper-
ature limits for growth in cell culture, or anti-genic composition (23).
In vitro propagation of each of' these fishrhabdoviruses has been reported only in f'ish cellculture. We have also investigated the cultiva-bility of SVC, IHN, and Egtved viruses inseveral types of continuously cultivated cells ofmammalian or reptilian origin. The finding thatmany non-fish cell types do support f'ish rhab-dovirus growth has been exploited in studies of'the influence of diverse types of' host cells on thetemperature requirements for virus replication.The demonstration of virus-induced ribonucleicacid (RNA) in amounts greatly in excess of'control cell RNA concentrations is describedin a system comprised of hamster cells infected
180
on February 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
on February 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
on February 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

FISH RHABDOVIRUS IN NON-PISCINE CELLS
with SVC virus and incubated at room tempera-ture without actinomycin D (AD).
MATERIALS AND METHODS
Cell cultures. The methods used for cultivation offish and reptilian cell lines have been described indetail elsewhere (3, 27). The growth medium forreptilian cells was Eagle basal medium (14) contain-ing 10%C fetal calf serum. Fish cell lines, humandiploid cell strain WI-38 and green monkey cell lineVero, and hamster cell line BHK/21 were grown inEagle minimal essential medium (MEM) (15) con-
taining 10% fetal calf serum. Rainbow trout cells(RT-G-2) (26) were propagated at 18 C, fathead min-now (FHM) (20), Terrapene heart (TH1) (9), andGekko lung (GL1) (6) cell lines were propagated at23 C, and the viper cell lines VSW and VH2 (7) were
propagated at 30 C.Virus infection experiments. Monolayer cell cul-
tures were washed once with phosphate-bufferedsaline (PBS) (Dulbecco), infected with virus adsorbedat the normal incubation temperature of the virus for30 min washed two times with PBS, and refed withgrowth (see above) or maintenance medium (MEMwith 0.1% bovine serum albumin or BHK medium[24] with 2% fetal calf serum). When plaque assayswere performed, cell cultures in 30-ml plastic flaskswere infected with serial dilutions of virus, the cellcultures were not washed after adsorption, and an
overlay consisting of 0.5%. agarose in BHK mediumwas added. The staining overlay consisted of 0.01%neutral red in the same medium.
Radioisotope incorporation studies. ['HJuridineincorporation was measured in cells grown on dupli-cate glass cover slips in 35-mm diameter plastic petridishes. Isotope concentrations and labeling times are
given in Results. The incorporation of radioactiveuridine by infected cells was measured by scintillationspectrometry of duplicate cover slips washed in PBS,fixed in 10% trichloroacetic acid-methanol (3:1) for30 min and 5% trichloroacetic acid for 10 min, andfinally rinsed in methanol.
RESULTSSpectrum of host cell susceptibility to fish
rhabdoviruses. The released virus yields ob-tained during the course of three serial non-
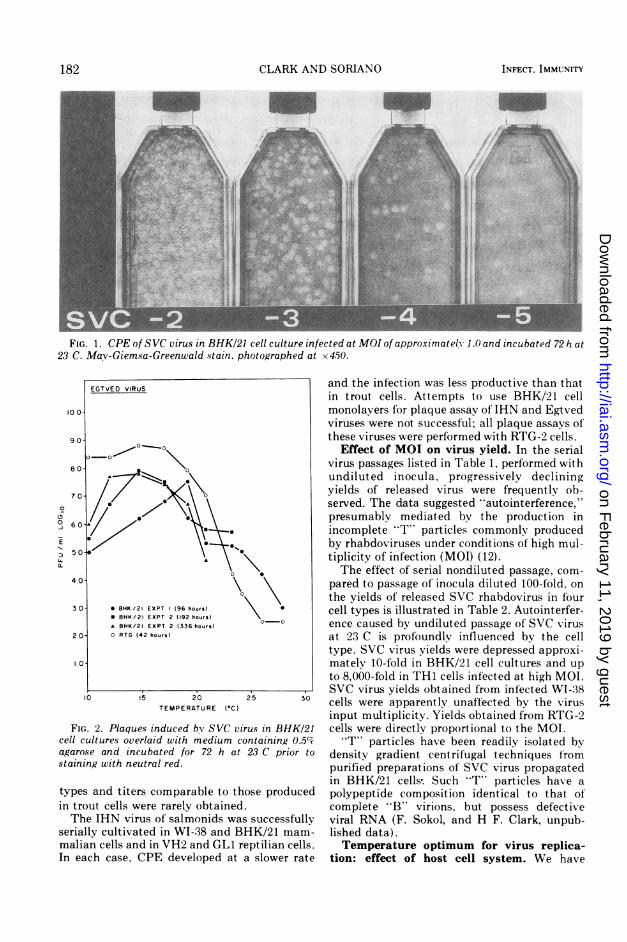
diluted virus passages in several cell lines incu-bated at temperatures appropriate for eachvirus are presented in Table 1. SVC virusreplicated efficiently in each of' the cell typestested, including three from mammals, fourfrom reptiles, and two from fish. The infectionwas cytopathic in each system (see Fig. 1).Cytopathic effect (CPE) appeared in mamma-lian cells and in turtle cell line TH1 at a ratesimilar to that observed in fish cells; the devel-opment of CPE was delayed for several days inthe other types of reptilian cells. Nevertheless,released virus yields at the time of near max-
imum CPE were similar in all cell types tested.BHK/21 cells were tested as a substrate for
plaque assay of SVC virus. Clear lytic plaqueswere induced after incubation of SVC virus-in-fected BHK/21 cell monolayers for 3 to 4 daysat room temperature (Fig. 2). Comparativetitrations indicated that plaque-forming unit(PFU) titers obtained in BHK/21 cell cultureswere three to fourfold lower than those obtainedin FHM cell culture. However, because of con-
venience, this cell system was adopted for allsubsequent assays of SVC virus infectivity.SVC virus was also found to induce clear pla-ques in human WI-38 cell monolayers and inBHK clone S13 cells, both after a similar incu-bation period.Egtved virus was serially propagated at 18 C
without difficulty in mammalian WI-38 andBHK/21 cells and in reptilian THI and GLIcells, but replication was not demonstrable inVero cells or in the viper cell lines. Egtved CPEwas invariably delayed in the non-salmonid cell
TABLE 1. Replication of piscine rhabdouiruses in cell cultures of mammlian. reptilian, or piscine origin'
Viruses
Cell lines SVC Egtved IHN
Passage I Passage II Passage III Passage I Passage II Passage III Passage I Passage II Passage III
WI-38 5.5 107 2.5 106r 1.3 106 4.0) 11( 6.0 1(6 5.0) 1()3 1. 5 10' 5.0 1( 1. 1(4BHK2I 2.8 1(' 3.5 10' 3.5 10' 1.( 107 9.5@) 10' )5.* 10() > 2 1)0 2.5 I 0 2.5 10(Vero 3.:3 107 2.9 1(7 5.0 106 4.5 106 4.0 1(3 <5.0 10( _ h
Vsw 9.5 107 4.05 10' 1.05- 108 6.0 104 1 .( 103 l.5 10' > 105 - _VH2 1.6 107 3.0 106 7.0 1()6 1.0 10' 5.0- 1()2 5.0 10( :I) 1(4 2.0 104 1.2 1(4GL1 1.5 10( 2.5 X 107 1 .5 107 3.0 10' :I3.0 106 9.5 1( 1.( *1(6 - 5.)* 1()3TH1 >5.0 X 107 2.6 x 10' 2.0 - 106 1.2 * 10' 1.0 1(04 5. 1(03 > )) 10( 6 _RTG 5.0 106 5.0 x 10' 1.5, 10' 9.() 107 1.0 107 4.0 1)5 4. 106 4.0 104 4.5 10FHM 3.0 106 6.5 k 105 - 5.0 105 :3.0 1()4
a Yields in PFU per milliliter of released virus. Cell cultures were originally inftected at a Xirus multiplicity ot from 1..0 to 10.0:released virus was harvested when > 50' of cells were involved with CPE. Serial passage was performed with nondilutedsupernatant fluids. SVC-infected cultures were incubated at 2:3 C. Trout viruses *vere propagated at 18 C. SVC virus p)laquetitrations were performed in BHK/21 cells: Egtved and IHN viruses were assaved in RTG-2 cells.
'Dash indicates infection not performed.
VOL. 10, 1974 181
on February 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
CLARK AND SORIANO
FIG. 1. CPE of SVC virus in BHK/21 cell culture infected at MOI of approximatelY 1.0 and incubated 72 h at23 C. Mav-Giemsa-Greenwald stain, photographed at x450.
10 0
0.
-i
10 15 20 25 30TEMPERATURE (°C)
FIG. 2. Plaques induced by SVC virus in BHK/21cell cultures overlaid with medium containing 0.5%cagarose and incubated for 72 h at 23 C prior tostaining with neutral red.
types and titers comparable to those producedin trout cells were rarely obtained.The IHN virus of' salmonids was successfully
serially cultivated in WI-38 and BHK/21 mam-malian cells and in VH2 and GL1 reptilian cells.In each case, CPE developed at a slower rate
and the inf'ection was less productive than thatin trout cells. Attempts to use BHK/21 cellmonolavers for plaque assay of' IHN and Egtvedviruses were not successf'ul; all plaque assays of'these viruses were performed with RTG-2 cells.
Effect of MOI on virus yield. In the serialvirus passages listed in Table 1, performed withundiluted inocula. progressively decliningyields of released virus were frequently ob-served. The data suggested "autointerference,"presumably mediated by the production inincomplete 'T" particles commonly producedby rhabdoviruses under conditions of high mul-tiplicity of infection (MOI) (12).The effect of serial nondiluted passage, com-
pared to passage of inocula diluted 100-f'old, onthe yields of' released SVC rhabdovirus in fourcell types is illustrated in Table 2. Autointerfer-ence caused by undiluted passage of' SVC virusat 23 C is profoundly influenced by the celltype. SVC virus yields were depressed approxi-mately 10-fold in BHK/21 cell cultures and upto 8,000-fold in THI cells infected at high MOI.SVC virus yields obtained from inf'ected WI-38cells were apparently unaffected by the virusinput multiplicity. Yields obtained from RTG-2cells were directly proportional to the MOI."T" particles have been readily isolated by
density gradient centrif'ugal techniques frompurif'ied preparations of' SVC virus propagatedin BHK/21 cells. Such "T" particles have apolypeptide composition identical to that of'complete "B"' virions, but possess defectiveviral RNA (F. Sokol, and H F. Clark, unpub-lished data).Temperature optimum for virus replica-
tion: effect of host cell system. We have
182 INFECT. IMMUNITY
on February 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
FISH RHABDOVIRUS IN NON-PISCINE CELLS
TABLE 2. Production of SVC virus during serialpassage, nondiluted, or diluted (10-2-fold). in diverse
cell culture svstemsa
Virus vield
Pas- Diluted Nondiluted passageCell type sage passage
no.Fraction of
PFt per ml PFU per ml diluted pas-sage yield
BHK/21 1 8.0 x 108 6.5 x 107 0.0812 1.0 x 108 8.5 x 106 0.0853 4.5 x 10' 4.0 x 106 0.089
WI-38 1 1.8 x 106 8.5 x 106 4.72 1.4x 106 1.8 x 106 1.33 2.0 x 106 3.0 x 106 1.5
RTG-2 1 3.2 x 105 9.0 X 105 2.82 3.5 x 106 >2.5 x 107 >7.13 4.0 x 104 4.0 x 105 10.0
TH 1 1 6.5 x 105 4.0 x 105 0.622 1.6 x 106 7.5 x 104 0.0473 6.5 x 108 8.0 x 104 0.00012
a First passage cell cultures were infected at a MOIof 10.0 (nondiluted series) or 0.10 (diluted series) andincubated at 23 C. Supernatant fluids were harvestedwhen 50 to 75% of cells were involved in CPE and usedfor subsequent passages, nondiluted or diluted 10-2,as indicated.
previously suggested (10) that systems com-prised of cells infected with viruses havingtemperature optima at which cell metabolism isretarded may provide propitious opportunitiesfor the study of preponderantly viral metabolicprocesses in cells not treated with selectivemetabolic poisons. We have similarly suggestedthat the ability of a virus to replicate attemperatures nonoptimal for the host cell maypossibly shed light on the dependence (or in-dependence) of virus replication upon host cellmetabolism.The results of comparative determinations of
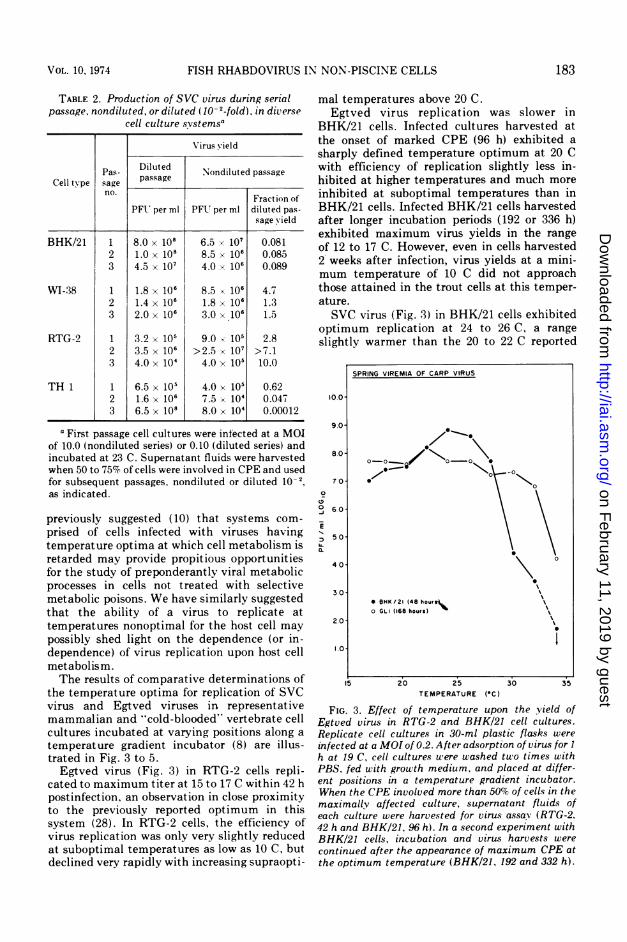
the temperature optima for replication of SVCvirus and Egtved viruses in representativemammalian and "cold-blooded" vertebrate cellcultures incubated at varying positions along atemperature gradient incubator (8) are illus-trated in Fig. 3 to 5.Egtved virus (Fig. 3) in RTG-2 cells repli-
cated to maximum titer at 15 to 17 C within 42 hpostinfection, an observation in close proximityto the previously reported optimum in thissystem (28). In RTG-2 cells, the efficiency ofvirus replication was only very slightly reducedat suboptimal temperatures as low as 10 C, butdeclined very rapidly with increasing supraopti-
mal temperatures above 20 C.Egtved virus replication was slower in
BHK/21 cells. Infected cultures harvested atthe onset of marked CPE (96 h) exhibited asharply defined temperature optimum at 20 Cwith efficiency of replication slightly less in-hibited at higher temperatures and much moreinhibited at suboptimal temperatures than inBHK/21 cells. Infected BHK/21 cells harvestedafter longer incubation periods (192 or 336 h)exhibited maximum virus yields in the rangeof 12 to 17 C. However, even in cells harvested2 weeks after infection, virus yields at a mini-mum temperature of 10 C did not approachthose attained in the trout cells at this temper-ature.SVC virus (Fig. 3) in BHK/21 cells exhibited
optimum replication at 24 to 26 C, a rangeslightly warmer than the 20 to 22 C reported
10.0-
9.0-
8.0-
70O
2o 6.0
, 50-
40O
30-
2.0
I.0*
15 20 25 30 35TEMPERATURE tIC)
FIG. 3. Effect of temperature upon the yield ofEgtved virus in RTG-2 and BHK/21 cell cultures.Replicate cell cultures in 30-ml plastic flasks were
infected at a MOI of 0.2. After adsorption of virus for I
h at 19 C, cell cultures uere washed two times withPBS. fed with growth medium, and placed at differ-ent positions in a temperature gradient incubator.When the CPE involved more than 50% of cells in themaximally affected culture, supernatant fluids ofeach culture were harvested for virus assay (RTG-2.42 h and BHK/21, 96 h). In a second experiment withBHK/21 cells, incubation and virus harvests were
continued after the appearance of maximum CPE atthe optimum temperature (BHK/21, 192 and 332 h).
SPRING VIREMIA OF CARP VIRUS
* 88K /21 (48 hours4,%O GLI (168 hours)
VOL. 10, 1974 183
on February 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
184
10.0
90
8 0
7 0
60-
5.0-
4.0-
3.0-
20-
1.0-
10 15 20 25TEMPERATURE (-C)
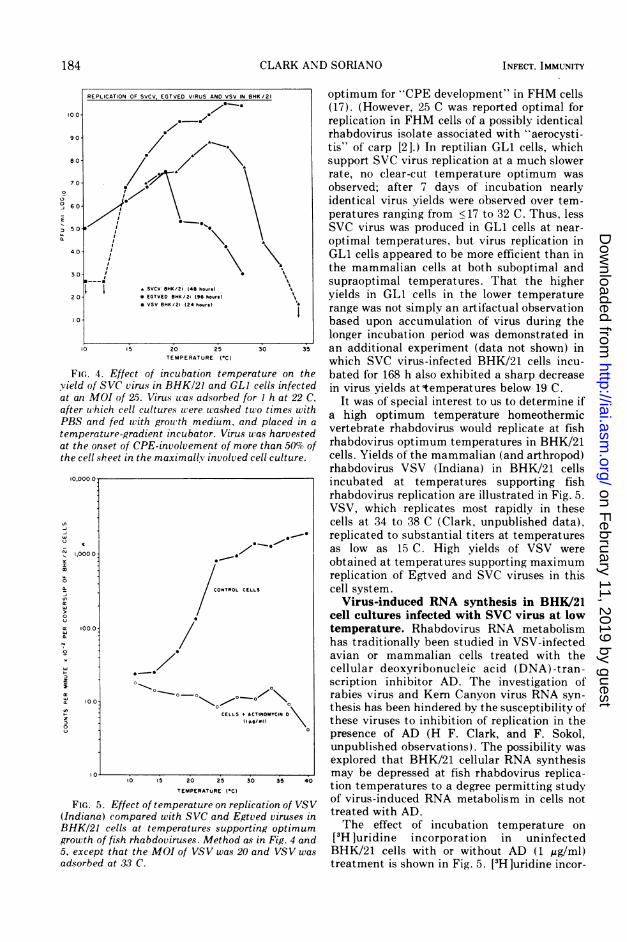
FiG.. 4. Effect of incubation tempercyield of SVC virus in BHK/21 and GLIat an MOI of 25. Virus was adsorbed forafter which cell cultures were washed tuPBS and fed with growth medium. anctemperature-gradient incu bator. Virus uat the onset of CPE-involvement of mor(the cell sheet in the maximally involved c
1,0000
10.0
10 15 20 25 30
TEMPERATURE -C)
FIG. 5. Effect of temperature on replic(Indiana) compared with SVC and EgtvBHK/21 cells at temperatures supportigrowth of fish rhabdoviruses. Method as
5, except that the MOI of VSV was 20 cadsorbed at 33 C.
CLARK AND SORIANO INFECT. IMMUNITY
HK /21 optimum for "CPE development" in FHM cells(17). (However, 25 C was reported optimal forreplication in FHM cells of a possibly identicalrhabdovirus isolate associated with "aerocysti-tis" of' carp [2].) In reptilian GL1 cells, whichsupport SVC virus replication at a much slowerrate, no clear-cut temperature optimum wasobserved; after 7 days of incubation nearlyidentical virus yields were observed over tem-peratures ranging from <17 to 32 C. Thus, lessSVC virus was produced in GLI cells at near-optimal temperatures, but virus replication inGLI cells appeared to be more efficient than inthe mammalian cells at both suboptimal andsupraoptimal temperatures. That the higheryields in GLI cells in the lower temperaturerange was not simply an artifactual observationbased upon accumulation of virus during thelonger incubation period was demonstrated in
30 3 an additional experiment (data not shown) inwhich SVC virus-infected BHK/21 cells incu-
lture on the bated for 168 h also exhibited a sharp decreasecells infected in virus yields attemperatures below 19 C.r I h at 22 C. It was of' special interest to us to determine if'o times with a high optimum temperature homeothermicI placed in a vertebrate rhabdovirus would replicate at fish'7as harvestede than 50% of rhabdovirus optimum temperatures in BHK/21ell culture. cells. Yields of' the mammalian (and arthropod)
rhabdovirus VSV (Indiana) in BHK/21 cellsincubated at temperatures supporting fishrhabdovirus replication are illustrated in Fig. 5.VSV, which replicates most rapidly in thesecells at 34 to 38 C (Clark, unpublished data),
*.e@ replicated to substantial titers at temperaturesas low as 15 C. High yields of VSV wereobtained at temperatures supporting maximumreplication of Egtved and SVC viruses in this
LS cell system.Virus-induced RNA synthesis in BHK/21
cell cultures infected with SVC virus at lowtemperature. Rhabdovirus RNA metabolismhas traditionally been studied in VSV-infectedavian or mammalian cells treated with thecellular deoxyribonucleic acid (DNA)-tran-scription inhibitor AD. The investigation of
0\ rabies virus and Kem Canyon virus RNA syn-0\ thesis has been hindered by the susceptibility of
:TINOMYCIN DII-fl \ these viruses to inhibition of replication in the0 presence of AD (H F. Clark, and F. Sokol,
unpublished observations). The possibility wasexplored that BHK/21 cellular RNA synthesis
)iS 4'0 may be depressed at fish rhabdovirus replica-tion temperatures to a degree permitting study
ationof VSV of virus-induced RNA metabolism in cells notViV treated with AD.
eng optimum The effect of incubation temperature onin Fig. 4 and [3H luridine incorporation in uninfected2nd VSV was BHK/21 cells with or without AD (1 ,ug/ml)
treatment is shown in Fig. 5. [3H ]uridine incor-
REPLICATION OF SVCV, EGTVED VIRUS AND VSV IN RI
l//
SVCV 66MK/21 146 hours)* EGTVEO 86K/21 (96 hours)a VSV 86RK/21 124 hours)
jONTROL CELL
CELLS + ACO iLC
20
on February 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
FISH RHABDOVIRUS IN NON-PISCINE CELLS
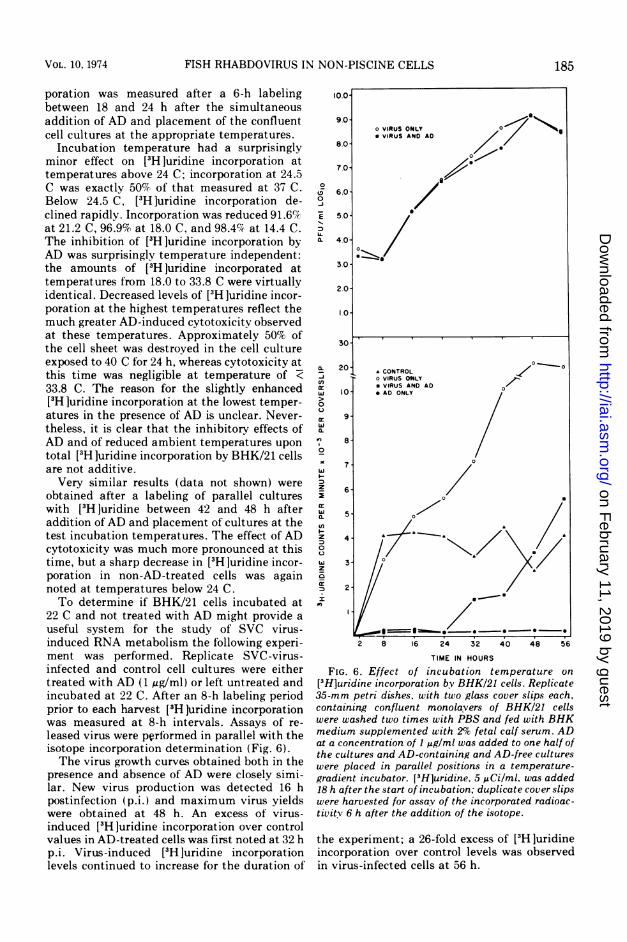
poration was measured after a 6-h labelingbetween 18 and 24 h after the simultaneousaddition of AD and placement of the confluentcell cultures at the appropriate temperatures.
Incubation temperature had a surprisinglyminor effect on [3H]uridine incorporation attemperatures above 24 C; incorporation at 24.5C was exactly 50% of that measured at 37 C.Below 24.5 C, [3H]uridine incorporation de-clined rapidly. Incorporation was reduced 91.6%at 21.2 C, 96.9% at 18.0 C, and 98.4% at 14.4 C.The inhibition of [H ]uridine incorporation byAD was surprisingly temperature independent:the amounts of [3H luridine incorporated attemperatures from 18.0 to 33.8 C were virtuallyidentical. Decreased levels of [3H ]uridine incor-poration at the highest temperatures reflect themuch greater AD-induced cytotoxicity observedat these temperatures. Approximately 50% ofthe cell sheet was destroyed in the cell cultureexposed to 40 C for 24 h, whereas cytotoxicity atthis time was negligible at temperature of <33.8 C. The reason for the slightly enhanced[3H ]uridine incorporation at the lowest temper-atures in the presence of AD is unclear. Never-theless, it is clear that the inhibitory effects ofAD and of reduced ambient temperatures upon
total [3H ]uridine incorporation by BHK/21 cellsare not additive.Very similar results (data not shown) were
obtained after a labeling of parallel cultureswith [3H ]uridine between 42 and 48 h afteraddition of AD and placement of cultures at thetest incubation temperatures. The effect of ADcytotoxicity was much more pronounced at thistime, but a sharp decrease in [3H ]uridine incor-poration in non-AD-treated cells was againnoted at temperatures below 24 C.To determine if BHK/21 cells incubated at
22 C and not treated with AD might provide auseful system for the study of SVC virus-induced RNA metabolism the following experi-ment was performed. Replicate SVC-virus-infected and control cell cultures were eithertreated with AD (1 gg/ml) or left untreated andincubated at 22 C. After an 8-h labeling periodprior to each harvest [3H Juridine incorporationwas measured at 8-h intervals. Assays of re-
leased virus were performed in parallel with theisotope incorporation determination (Fig. 6).The virus growth curves obtained both in the
presence and absence of AD were closely simi-lar. New virus production was detected 16 hpostinfection (p.i.) and maximum virus yieldswere obtained at 48 h. An excess of virus-induced [3H ]uridine incorporation over controlvalues in AD-treated cells was first noted at 32 hp.i. Virus-induced [3H ]uridine incorporationlevels continued to increase for the duration of
10.0o
0
0
-J
i
0.
a.-i
Cf)
>crw
0~CL
tr
w
z
Ionw
z0.
(-)
z
CX
0
TIME IN HOURS
FIG. 6. Effect of incubation temperature on
[3H]uridine incorporation by BHK/21 cells. Replicate35-mm petri dishes, with two glass cover slips each,containing confluent monolavers of BHK/21 cellswere washed two times with PBS and fed with BHKmedium supplemented with 2% fetal calf serum. ADat a concentration of I pg/ml was added to one half ofthe cultures and AD-containing and AD-free cultureswere placed in parallel positions in a temperature-gradient incubator. [3H]uridine. 5 uCi/ml. was added18 h after the start of incubation; duplicate cover slipswere harvested for assay of the incorporated radioac-tivitv 6 h after the addition of the isotope.
the experiment; a 26-fold excess of [3H]uridineincorporation over control levels was observedin virus-infected cells at 56 h.
185VOL. 10, 1974
on February 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

CLARK AND SORIANO
The level of' [3H luridine incorporation innon-drug-treated uninfected cell cultures re-
mained constant throughout the experiment. Aclear excess of [3H ]uridine incorporation inuntreated infected cells was first noted 24 h p.i.The virus-induced excess isotope incorporationincreased steadily to reach peak levels at 48 and56 h p.i. which were 8.4- and 5.0-f'old greaterthan control cell levels, respectively. The totalvirus-induced excess of' [3H ]uridine incorpora-tion in non-AD-treated cells was in all cases
much greater than in cells treated with AD, theexcess varying in magnitude from 3.0- to 7.1-fold. The observations indicate that AD mark-edly inhibits SVC virus-induced RNA synthesisunder conditions causing no inhibition of infec-tious virus production.A further experiment was performed to deter-
mine whether concentrations of' AD less than 1
mg/ml might suppress cellular uridine incorpo-ration without affecting the amount of' virus-induced uridine incorporation. The results oftreatment of SVC-infected BHK/21 cells with0.1 and 0.01 Ag/ml, respectively, of' AD duringincubation at 23 C are presented in Table 3.AD used at concentrations of' 0.1 and 0.01
Ag/ml caused 85 to 90% and 50 to 75%; reduction,respectively, in control cell levels of [3H ]uridineincorporation at 24 or 48 h after initiation of'incubation at 23 C. Clear-cut virus-induced[3H]uridine incorporation was evident only inthe 48 h p.i. samples. At this time, in additionto the suppression of' cellular [3H ]uridine incor-poration, the excess virus-induced incorpora-tion of' [3H]uridine into infected cells was re-
duced by 60.5 and 28.4c, respectively, in thepresence of AD concentrations of' 0.10 and 0.01,ug/ml. The results suggest that any concentra-tion of' AD capable of reducing cellular uridine
corporation may also affect virus-induced meta-bolic processes.
DISCUSSIONWe have previously suggested (10) that vi-
ruses of' cold-blooded vertebrates cultivated inmammalian or avian cell culture might provideuseful systems for distinguishing viral and hostcell metabolic processes. Such potential existswith polyhedral cytoplasmic frog viruses (5, 19),but the propagation of other viruses isolatedfrom fish, reptiles, or amphibians representing a
wide variety of virus groups has been limited tocell culture systems from species closely relatedto the natural poikilothermic host of the virus(frog viruses, reviewed by Granoff [18]; fishviruses, listed by Wolf and Quimby [28]; iguanavirus [11]). We herein report that three rhab-doviruses isolated from fish, each of which haspreviously been studied only in fish cell culture,may be readilv propagated in a variety ofreptilian and mammalian cell cultures incu-
bated at room temperature or lower tempera-t ures.The efficiency of' fish rhabdovirus replication
in BHK/21 cells suggests especially interestingcomparative studies of nucleic acid and proteinmetabolism and virion morphogenesis in thiscell type most often used for the study of themore thoroughly characterized mammalianrhabdoviruses of' the vesicular stomatitis andrabies serogroups. Such studies are in progress.
A unique opportunity provided by the fishrhabdovirus BHK/21 cell system may be in theidentification of' host cell-derived versus virusgenome-coded virion enzymes (kinase, adeno-sine 5'-triphosphatase, RNA polymerase) in lowtemperature optimum fish viruses comparedwith mammalian rhabdoviruses cultivated at
TABLE 3. Effect of treatment with lou concentrations of AD upon SVC virus-induced [3H]uridineincorporation in infected BHK/21 cells incubated at 23 Ca
tEfftect
24 h p.i. 48 h 1).I.
AD Intected cells Excess Infected (ells l3HEurxdineconcn ControlHcellin(ig/mi) 1Control H luridine H luridinie Control cells incorplo-Hiuridine inorporation 3H luiin Hluridinerainn
incorporated ratned irus titer in intected incorporat ed inrd virus titer intected(counts/min) (counts/(PFUr/ml cellIsi (counts/min) (counts/ (PFU/ml ( cells
(counts/ (cPFntm/mi mill)counts!mini(counts/mm) mini ~~~~~~~~~mini0.10 915 1,700 >2.5 x 106 785 1,850 6,060 1.9 x 108 4,2100.01 4,850 3,830 1.5 x 106 3,400 10,400 > 2.5 x 108 7,.000None 10,000 11,000 >2.5 x 106 1,000 13,250 22,800 >2.5 x 108 9,550
a Infection of cover slips with SVC, treatment with AD, and 8-h pulse-labeling were performed as describedfor Fig. 6.
I NFECT. I MMU,NITYl186
on February 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

FISH RHABDOVIRUS IN NON-PISCINE CELLS
identical temperatures in the same cell culture(BHK/21) substrate. Observation of' differingtemperature responses of enzymes of warm- andcold-temperature optimum viruses grown underthese identical conditions would strongly sug-gest that the enzymes are virus-genome-coded.The converse situation would also apply.An analogy with mammalian rhabdovirus
replication patterns is suggested by the observa-tion of presumably incomplete "T" particle-mediated autointerference after serial non-diluted passage of fish rhabdovirus SVC virus inmammalian or reptile-origin cell culture. Aswith VSV (21, 25) the degree of autointerferenceobserved was strongly host-cell influenced,being pronounced in BHK/21 and one reptiliancell line, and negligible in human WI-38 cellsand in trout cells. Particularly efficient expres-sion of "autointerference" accompanied by "T"particle formation in reptile cell cultures in-fected with VSV at high MOI has also beenobserved (P. D. Lunger and H F. Clark, manu-script in preparation).
Previous studies (8, 10) have suggested thatthe temperature optimum of the host cell hasonly minimal effects on temperature effects onthe replication of infecting viruses. Preliminarystudies reported here suggest that the host cellhas a pronounced effect on only the lowertemperature level limits of piscine rhabdovirusreplication. Both SVC and Egtved viruses repli-cated more efficiently at suboptimal tempera-tures in cells of cold-blooded vertebrate originthan in BHK/21 cells, suggesting a requisitehost cell metabolic contribution to virus replica-tion that is temperature limited.An appealing theoretical question is that of'
whether fish viruses may be especially rapidlyattenuated in mammalian cells. It has fre-quently been assumed that the role of mamma-lian virus attenuation varies directly with thephylogenetic distance of' the test host cell fromthat of the natural host. Attenuation of mam-malian rabies rhabdovirus by serial propagationin reptilian cells has been demonstrated (4).The possibility that the converse situation maybe similarly eff'ective is now being investigated.
Piscine rhabdoviruses propagated in BHK/21cells appear to provide a particularly promisingsystem for the study of rhabdovirus RNA me-tabolism. RNA-type virus-induced RNA me-tabolism has traditionally been investigatedalmost exclusively in mammalian or avian cellstreated with AD. The artifacts that may beinduced by AD treatment of cells are almostentirely unknown.We have demonstrated that, although cellu-
lar levels of [3H luridine incorporation are much
higher in untreated BHK/21 cells incubated at23 C than in AD-treated cells, such cells in-fected with SVC virus exhibit a clear-cut excessof virus-induced [3H]uridine incorporation overcontrol cell levels. The total virus-induced uri-dine incorporation in untreated infected cells ismuch greater than in AD-treated infected cellsand the onset of virus-induced excess uridineincorporation is delayed by AD treatment. Theresults indicate that untreated cells produce agreat excess of virus-associated RNA over theamounts needed for maximal production ofinf'ectious virions. The concentration of' thisvirus-induced RNA is reduced by treatmentwith AD even at levels too low to efficientlydepress cellular [3H ]uridine incorporation.Whether the excess virus-induced [3H ]uridineincorporation represents exclusively virus-specific RNA or consists in part of virus-induced cellular RNA, and whether AD selec-tively inhibits particular species of virus-induced RNA are questions that remain to beanswered by further experimentation. We sug-gest that BHK/21 cells inf'ected with SVC virusmay provide a particularly usef'ul source of largeamounts of rhabdovirus-specific RNA and mayalso provide insights into aspects of virus-induced RNA metabolism that may be madeobscure by cell treatment with AD.
ACKNOWLEDGMENTS
This investigation was supported in part by Public HealthService research grants AI-09706 from the National Instituteof Allergy and Infectious Diseases and RR 05540 trom theDivision of Research Resources.
LITERATURE CITED
1. Amend. D. F.. and V. C. Chambers. 1970(. MIorphology ofcertain viruses of' salmonid f'ishes. I. In Lvitro studies of'some viruses causitng hematopoietic necrosis. ,J. Fish.Res. Rd. Can. 27:1285-129:8.
2. Bachmalnn. P. A., and W. Ahne. 197,:3. Isolation andcharacterization of'agent causinig swim bladder inflam-mation in carp. Nature (London) 244:235-237.
3. Clark. H F. 1972a. Cultivation of cells from poikilother-mic vertebrates. p. 2873-:325. In G. H. Rothblat and V.J. Cristot'alo (ed.), Growth, nutrition, and metabo-lism of' cells in culture. vol. 2. Academic Press Inc.New York.
4. Clark. H F. 1972b. Grow-th and attenuation oftrabies virusin cell cultures of' reptilian origin. Proc. Soc. Exp. Biol.Med. 139:1317-1325.
.. Clark. H F.. J. C. Brennan. R. F. Zeigel. and D. T.Karzon. 1968. Isolation and characterization of virusesfrom the kidneys of Rana pipienms with renal adenocar-cinoma bet'ore and after passage in the red ef't ( Triturusviridescens). J. Virol. 2:629-64(0.
6. Clark, H F.. M1. N1. Cohen. and D. T. Karzon. 1970.Characterizationi of' reptilian cell lines established atincubation temperatures of' 2:3 to :360. (:34622). Proc.Soc. Exp. Biol. Med. 133:10:39-1t)47.
7. Clark, H F., M. M. Cohen, and P. D. Lunger. 197:3.Comparative characterizatioin of a C-type virus-pro-ducing cell line (VSW) f'rom a virus-free cell line (VH2)
VOL. 10, 19714 187
on February 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

188 CLARK AN
from Vipera russelli. J. Nat. Cancer Inst. 51:645-657.8. Clark, H F., F. Kaminski, and D. T. Karzon. 1970.
Thermoelectrically cooled temperature-gradient appa-ratus for comparative cell and virus temperature stud-ies. Appl. Microbiol. 19:848-854.
9. Clark, H F., and D. T. Karzon. 1967b. Terrapene Heart(TH-1). A continuous cell line from the heart of the BoxTurtle (Terrapene carolina). Exp. Cell Res. 48:263-268.
10. Clark, H F., and D. T. Karzon. 1968. Temperatureoptima of mammalian and amphibian viruses in cellcultures of homeothermic and poikilothermic origin.Arch. Gesamte Virusforsch. 23:270-279.
11. Clark, H F., and D. T. Karzon. 1972. Iguana virus, aherpes-like virus isolated from cultured cells of lizard,Iguana iguana. Infect. Immunity 5:559-569.
12. Cooper, P. D., and A. J. D. Bellett. 1959. A transmissibleinterfering component of vesicular stomatitis viruspreparations. J. Gen. Microbiol. 21:485-597.
13. De Kinkelin, P., B. Galimard, and R. Bootsma. 1973.Isolation and identification of' the causative agent of"Red Disease" of pike (Esox lucius L. 1766). Nature(London) 241:465-467.
14. Eagle, H. 1955. Nutrition needs of' mammalian cells intissue culture. Science 122:501-504.
15. Eagle, H. 1959. Amino acid metabolism in mammaliancell cultures. Science 130:432-437.
16. Fijan, N. 1972. Infectious dropsy in carp-A diseasecomplex. Symp. Zool. Soc. Lond. 30:39-51.
17. Fijan, N., Z. Petrinec, D. Sulimanovic. and L. 0.Zwillenberg. 1971. Isolation of the viral causative agentfrom acute form of infectious dropsy of carp. Vet. Arch.41:125-138.
18. Granoff, A. 1969. Viruses of amphibia, p. 107-1:37. InW. Arber et al. (ed.), Current topics in microbiologyand immunology. vol. 50. Springer-Verlag, New York.
ID SOUR1ANO INFECT. IMMUNITY
19. Granoff. A., P. E. Came, and D. C. Breeze. 1966. Virusesand renal carcinoma of' Rana pipien.s. I. The isolationand properties of virus f'rom normal and tumor tissue.Virology 29:133-148.
20. Gravell, M., and R. G. Malsberger. 1965. A permanentcell line from the fathead minnow (Pimephalespromelas). Ann. N.Y. Acad. Sci. 126:555-565.
21. Huang, A. S., and D. Baltimore. 197)). Defective viralparticles and viral disease processes. Nature (London)226:325-327.
22. Jensen, M. H. 196:3. Preparation of fish tissue cultures forvirus research. Bull. Off'. Intern. Epizool. 59:131-134.
23. J0rgensen, P. E. V. 1972. Egtved virus: antigenic varia-tion in 76 virus isolates examined in neutralizationtests and by means of the fluorescent antibody tech-nique. Symp. Zool. Soc. Lond. 30:333-340.
24. MacPherson, I., and M. Stoker. 1962. Polvoma transf'or-mation of' hamster cell clones-an investigation ofgenetic factors affecting cell competence. Virology16:147-151.
25. Perrault, J., and J. Holland. 1972. Variability of vesicularstomatitis virus autointerference with dif'ferent hostcells and virus serotypes. Virology 50:148-158.
26. Wolf, K., and M. C. Quimby. 1962. Established euryther-mic line of' f'ish cells in vitro. Science 135:1065-1066.
27. Wolf, K., and M. C. Quimby. 1969. Fish cell and tissueculture. Fish Phvsiol. 3:253-30)5.
28. Wolf, K., and M. C. Quimby. 1973. Fish viruses: buffersand methods for plaquing eight agents under normalatmosphere. Appl. Microbiol. 25:659-664.
29. Zwillenberg, L. O., M. H. Jensen. and H. H. L. Zwillen-berg. 1965. Electron microscopy of' the virus of' viralhemorrhagic septicaemia of rainbow trout (Egtvedvirus). Arch. Gesamte Virusforsch. 17:1-19.
T- -I.11 . T -
on February 11, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

ERRATUM
Fish Rhabdovirus Replication in Non-Piscine Cell Culture:New System for the Study of Rhabdovirus-Cell Interaction
in Which the Virus and Cell Have DifferentTemperature Optima
H FRED CLARK AND ELVIRA Z. SORIANOThe Wistar Institute ofAnatomy and Biology, Philadelphia, Pennsylvania 19104
Volume 10, no. 1, p. 180. The erratum previously published in regard to this article (Infect.Immun. 10:970) was printed with an error in the second author's name. We regret the error.
213


















