
Firing relations of medial entorhinal neurons to the hippocampal theta rhythm in urethane...
8
Exp Brain Res (1992) 90:21-28 Experimental BrainResearch Springer-Verlag 1992 Firing relations of medial entorhinal neurons to the hippocampal theta rhythm in urethane anesthetized and walking rats M. Stewart, G.J. Quirk, M. Barry, and S.E. Fox Department of Physiology, State University of New York Health Science Center, 450 Clarkson Avenue, Brooklyn, NY 11203, USA Received December 2, 1991 / Accepted March 17, 1992 Summary. The firing of neurons from layers II and III of medial entorhinal cortex (MEC) was examined in rela- tion to the hippocampal theta rhythm in urethane anes- thetized and walking rats. 1) MEC neurons showed a significant phase relation to the hippocampal theta rhythm in both walking and urethane anesthetized rats, suggesting that this region contributes to the generation of both atropine-resistant and atropine-sensitive theta rhythm components. 2) The proportion of phase-locked cells was three times greater in walking rats (22/23 cells) as compared to anesthetized rats (8/23 cells), indicating that MEC cells made a greater contribution during walk- ing theta rhythm. This difference was also manifest in the greater mean vector length for the group of phase-!ocked MEC cells during walking" 0.39 4- 0.13 versus 0.21 4- 0,08. Firing rate differences between walking and urethane conditions were not significant. 3) In walking.rats, MEC cells fired on the positive peak of the dentate theta rhythm (group mean phase = 5~ 0 ~= positive peak at the hippocampal fissure). This is close to the reported phases for dentate granule and hippocampal pyramidal cells. The distribution of MEC cell phases in urethane anes- thetized rats was broader (group mean phase = 90~ con- sistent with the phase data reported for hippocampal projection cells. These findings suggest that medial entorhinal neurons are the principal determinant of theta-related firing of hippocampal neurons and that their robust rhythmicity in walking as compared to urethane anesthesia accounts for EEG differences across the two conditions. Key words: Rhythmical slow activity - Entorhinal cortex - EEG - Rats Correspondence to. M. Stewart, Dept. Pharmacology, Box 29, SUNY Health Sci. Ctr., Brooklyn NY 11203, USA Introduction The hippocampal theta rhythm, one of the best studied EEG phenomena, is a large amplitude, sinusoidal field potential that can be recorded throughout the hippocam- pal formation and the overlying neocortex. Many experi- ments, using a variety of methods, have been performed, first to determine which structures actually generate the theta rhythm, and then to identify the specific synaptic circuitry involved. The most powerful method for determining if a corti- cal region acts as a generator for the theta rhythm is to record the EEG at multiple depths within the region. The voltage records as a function of depth can be used to estimate the appearance (at sources) or disappearance (at sinks) of extracellular currents from the recording site (current source density analysis; CSD). Active sources and sinks are associated with membrane events such as post- synaptic potentials and action potentials. Currents ap- pearing in or disappearing from the extracellular space at one location on a cell must be associated with passive currents of the opposite direction at some other location on the cell. A pair of active and passive currents form a "dipole" that identifies the region as a generator of the field potential. Coupled with anatomical data, current source density profiles can be used to suggest the iden- tities of afferent systems generating the EEG. CSD data, however, do not distinguish between active and passive currents: an extracellular current sink in the apical den- dritic region might be active, due to excitatory synapses at that location, or passive, due to inhibitory synapses at another depth. Other experimental methods must be used to distin- guish between active and passive transmembrane cur- rents. Intracellular recordings, for example, permit the identification of subthreshold events as excitatory or inhibitory. Such recordings have even been used to con- firm the location of synaptic inputs, since electrotonic distances of particular conductance changes from the stimulating/recording microelectrode can be calculated from membrane impedance changes (Fox 1985, 1989).
Transcript of Firing relations of medial entorhinal neurons to the hippocampal theta rhythm in urethane...

Exp Brain Res (1992) 90:21-28
Experimental Brain Research �9 Springer-Verlag 1992
Firing relations of medial entorhinal neurons to the hippocampal theta rhythm in urethane anesthetized and walking rats
M. Stewart, G.J. Quirk, M. Barry, and S.E. Fox
Department of Physiology, State University of New York Health Science Center, 450 Clarkson Avenue, Brooklyn, NY 11203, USA
Received December 2, 1991 / Accepted March 17, 1992
Summary. The firing of neurons from layers II and III of medial entorhinal cortex (MEC) was examined in rela- tion to the hippocampal theta rhythm in urethane anes- thetized and walking rats. 1) MEC neurons showed a significant phase relation to the hippocampal theta rhythm in both walking and urethane anesthetized rats, suggesting that this region contributes to the generation of both atropine-resistant and atropine-sensitive theta rhythm components. 2) The proportion of phase-locked cells was three times greater in walking rats (22/23 cells) as compared to anesthetized rats (8/23 cells), indicating that MEC cells made a greater contribution during walk- ing theta rhythm. This difference was also manifest in the greater mean vector length for the group of phase-!ocked MEC cells during walking" 0.39 4- 0.13 versus 0.21 4- 0,08. Firing rate differences between walking and urethane conditions were not significant. 3) In walking.rats, MEC cells fired on the positive peak of the dentate theta rhythm (group mean phase = 5 ~ 0 ~ = positive peak at the hippocampal fissure). This is close to the reported phases for dentate granule and hippocampal pyramidal cells. The distribution of MEC cell phases in urethane anes- thetized rats was broader (group mean phase = 90~ con- sistent with the phase data reported for hippocampal projection cells.
These findings suggest that medial entorhinal neurons are the principal determinant of theta-related firing of hippocampal neurons and that their robust rhythmicity in walking as compared to urethane anesthesia accounts for EEG differences across the two conditions.
Key words: Rhythmical slow activity - Entorhinal cortex - EEG - Rats
Correspondence to. M. Stewart, Dept. Pharmacology, Box 29, SUNY Health Sci. Ctr., Brooklyn NY 11203, USA
Introduction
The hippocampal theta rhythm, one of the best studied EEG phenomena, is a large amplitude, sinusoidal field potential that can be recorded throughout the hippocam- pal formation and the overlying neocortex. Many experi- ments, using a variety of methods, have been performed, first to determine which structures actually generate the theta rhythm, and then to identify the specific synaptic circuitry involved.
The most powerful method for determining if a corti- cal region acts as a generator for the theta rhythm is to record the EEG at multiple depths within the region. The voltage records as a function of depth can be used to estimate the appearance (at sources) or disappearance (at sinks) of extracellular currents from the recording site (current source density analysis; CSD). Active sources and sinks are associated with membrane events such as post- synaptic potentials and action potentials. Currents ap- pearing in or disappearing from the extracellular space at one location on a cell must be associated with passive currents of the opposite direction at some other location on the cell. A pair of active and passive currents form a "dipole" that identifies the region as a generator of the field potential. Coupled with anatomical data, current source density profiles can be used to suggest the iden- tities of afferent systems generating the EEG. CSD data, however, do not distinguish between active and passive currents: an extracellular current sink in the apical den- dritic region might be active, due to excitatory synapses at that location, or passive, due to inhibitory synapses at another depth.
Other experimental methods must be used to distin- guish between active and passive transmembrane cur- rents. Intracellular recordings, for example, permit the identification of subthreshold events as excitatory or inhibitory. Such recordings have even been used to con- firm the location of synaptic inputs, since electrotonic distances of particular conductance changes from the stimulating/recording microelectrode can be calculated from membrane impedance changes (Fox 1985, 1989).

22
Stable intracellular recordings can be difficult to ob- tain in vivo and in some instances, like freely moving animals, they are impossible to obtain. For practical rea- sons, extracellular single unit recordings are more com- monly used to study neuronal circuit organization in intact animals. Extracellular unit recordings can be used to distinguish between active and passive t ransmembrane currents identified in depth profiles. To accomplish this, recordings f rom neurons afferent to the generator region must be combined with anatomical knowledge of their laminae of termination, and electrophysiological data on the excitatory or inhibitory nature of the input.
On the basis of much anatomical (Hjorth-Simonsen 1972; Hjorth-Simonsen and Jeune 1972; Steward 1976; Wyss 1981) and physiological data (Andersen et al. 1966a, b; Dudek et al. 1976; White et al. 1977; Doller and Weight 1982; Vanderwolfand Leung 1983; Tomasu- lo et al. 1991), the entorhinal cortices are suspected of generating a massive phasic extracellular current sink in the apical dendrites of the dentate gyrus and CA1 in both walking (Buzsaki et al. 1986; Brankack and Fox 1987) and urethane anesthetized rats (Buzsaki et al. 1986; Fox and Stewart 1986). The contribution of entorhinal cortex to the hippocampal E E G was suggested by early unit studies (Ranck 1973; Mitchell and Ranck 1980) and confirmed when bilateral lesions of the entorhinal cor- tices abolished the atropine-resistant component of the hippocampal theta rhythm (Vanderwolf and Leung 1983). Finding entorhinal units phase-locked to the theta rhythm in walking and ether anesthetized rats (Quirk and Stewart 1988) indicated that the entorhinal cortex con- tributed to the generation of both the atropine-resistant and the atropine-sensitive components of the hippocam- pal theta rhythm (Kramis et al. 1975; Buzsaki et al. 1983; Vanderwolf 1988).
As described above, an examination of the firing rela- tions of single entorhinal units to the hippocampal theta rhythm in walking and urethane anesthetized rats is essential for interpreting the hippocampal CSD data available f rom these two preparations. In addition, since the entorhinal cortices are the principal excitatory input to the dentate gyrus (Andersen et al. 1966a; Dudek et al. 1976; White et al. 1977), the firing relations of dentate neurons, as well as CA3 and CA1 pyramidal cells, may be controlled by the firing of entorhinal projection cells. This control can be assessed, at least indirectly, by deter- mining the firing relations of entorhinal inputs to the hippocampus. Studies of entorhinal control of hippo- campal activity are central for understanding the flow of information (specifically, spatial information) through the hippocampal format ion (e.g. Quirk et al. 1992).
Portions of the data in this manuscript have appeared elsewhere in abstract form (Quirk and Ranck 1986; Ste- wart et al. 1991). A similar study, by Alonso and Garcia- Austt (1987b), reported phase data for entorhinal units in curarized rats to complement their depth profiles through entorhinal cortex in those animals (Alonso and Garcia-Austt 1987a). Their unit data, however, cannot be readily applied to the interpretation of walking or urethane CSD data, since depth profiles of the hippo- campal theta rhythm vary among all of these conditions.
Materials and methods
Acute experiments
Male Sprague-Dawley albino rats (310-450 g) were anesthetized with urethane (1.0 g/kg i.v. or 1.5 g/kg i.p.) and prepared with a tracheal cannula for connection to a source of moisturized air. When animals received urethane intravenously, they were fitted with the tracheal tube and jugular cannula under ether anesthesia. The ether was gradually replaced with the calculated dose of ure- thane. Small supplemental doses of urethane were given, as needed, during the experiments.
Animals were held in a Kopf stereotaxic frame and maintained at 37 ~ C with an isothermal heating pad (Braintree Scientific, Brain- tree, Mass., USA) and/or a shielded heating lamp. In the 16 rats used for this part of the study, small holes were drilled through the skull and enlarged with a rongeur. The dura at the base of a hole was carefully removed and the exposed brain was covered with vaseline.
All implants were made using stereotaxic coordinates (Paxinos and Watson 1986). Unit recording electrodes (1-2 M~ teflon- insulated stainless steel, Microprobe Inc., Clarksburg, Md., USA) were lowered through the entorhinal cortex (AP 1.5 mm anterior to interaural line, ML 5.5 mm lateral to midline) at an angle of 0-15 ~ off the vertical in the coronal plane. Monopolar recordings of the hippocampal EEG were taken from each of a pair of stainless steel wires (125 g diameter) that were varnish insulated and cut square. The tips were vertically separated by about 1 mm and referred to a screw over cerebellum. The EEG electrode pair was lowered vertically (AP 4.7, ML 3.0) until the theta rhythm recorded on the deeper electrode was phase reversed with respect to that on the superficial electrode, and was maximal in amplitude. The deeper electrode tip was then near the hippocampal fissure (referred to here as "dentate" EEG) and the superficial electrode was dorsal to the pyramidal cell layer of CA1 (referred to here as "CAI" EEG) (Green and Rawlins 1979).
Unit activity and EEG recordings were amplified, filtered (single units: bandpass=500 Hz to 10 kHz; -24 dB/octave high pass, - 6 dB/octave low pass. EEG: bandpass=0.1 Hz to 10 kHz; - 6 dB/octave rolloffs), and stored on tape for off-line analysis.
Chronic experiments
A total of 10 female Long Evans hooded rats (240-310 g pre-train- ing weight) were used for this portion of the study. Rats were food deprived to approximately 80% of their normal body weight, and maintained that way for the remainder of the experiments. Rats were handled daily. After two days, each rat was introduced into a confining space (76 cm diam. grey cylinder) in which small food pellets were delivered at random intervals (Muller et al. 1987). Rats walked about the space eating the pellets. After several days of training, long periods of walking behavior could be induced. After a training period of about two weeks (15-20 exposures to the testing chamber), rats were surgically implanted with chronic electrode assemblies.
Under nembutal anesthesia (50 mg/kg i.p.), small holes were drilled in the skull to pass two separate electrode assemblies. The first was a pair of fine wires (78 p in diam.), offset vertically by about 1 mm. The pair was inserted above the hippocampus for EEG recordings (skull coordinates: 3.5 mm posterior to bregma, 2.0 mm lateral to midline, 2.5 mm ventral to surface of brain). The second electrode assembly was a modified version of that described by Kubie (1984). It consisted of a moveable bundle of ten Formvar- insulated microwires (each 25 g in diam.) for recording single units in entorhinal cortex. Just prior to implantation, the electrode tips were coated with earbowax to prevent bending of the tips as they were driven down to near the level of entorhinal cortex. Skull coordinates used to reach MEC were as follows: 4.0 mm posterior to bregma and 5.2 mm lateral to midline. The electrode assembly

23
was angled 30 ~ off the vertical in the sagittal plane and driven initially to a depth of 4.5 mm below the surface of the brain before the end of surgery. Stainless steel screws were placed over cerebel- lum and anterior to bregma. Both electrode assemblies were an- chored to the skull with Grip Cement (Dentsply, Maumee, Ohio). A topical antibiotic was applied around the wound at the end of surgery. Rats were allowed to recover for approximately one week before testing for unit activity.
The unit recording and data collection procedures have been described previously (Muller et al. 1987; Quirk et al. 1992). Briefly, each rat was connected to a cable that permitted free movement within the cylindrical chamber. Electrode contacts on the rat's head were connected to field-effect transistors in source-follower con- figuration located at the end of the input cable. Signals were am- plified, filtered (single units: bandpass=300 Hz to 10 kHz; - 6 dB/octave. EEG: bandpass = 0.1 Hz to 10 kHz; - 6 dB/octave rolloffs), and stored on video tape for off-line analysis. To aid discrimination of single units, active high pass filters with sharper rolloffs ( - 2 4 dB/octave) were sometimes used. In addition, three window discriminators (Bak, time-amplitude, model DIS-l) con- nected in series were used to isolate waveforms. Acceptance pulses from the last discriminator were also stored on tape. The driveable unit electrode was advanced in 50 g steps between unit recordings.
Histology of electrode placements
Electrode locations for the last unit on each pass were marked at the ends of the experiments by passing positive current through the recording electrodes (2-5 gA, 10-30 s). Rats were overdosed with urethane (acute experiments) or Nembutal (chronic experiments) and perfused through the heart. Brains were kept in 10% buffered formalin and exposed to potassium ferrocyanide (about 2%) prior to sectioning. Lesions were identified in Nissl-stained 40 g frozen sections with the Prussian blue reaction. Units not at the lesion site were located by measuring back from a lesion along the electrode track.
All lesions were found to be in layer II of medial entorhinal cortex. We identified three groups on the basis of histology (layers II, II/III, and III), but cells in layers II and III were electrophysio- logically indistinguishable. For this reason, no distinctions are made between layers II and III in the Results section. Occasional units located away from the two projection layers, II and III (Hjorth- Simonsen 1972; Steward 1976; Steward and Scoville 1976; Germ- roth et al. 1989), were readily identified and are not included in this report.
Data analysis
Phase histograms were constructed using either a DEC PDP 11/45 or a Hewlett Packard 9836U computer system (Corvallis, Oregon, USA). Detailed descriptions of the construction of phase histo- grams have been published (Fox et al. 1986; Stewart and Fox 1989c). Briefly, estimates of rhythmic quality, based on autocorrela- tions of 1 s epochs of EEG (Stewart and Fox 1989a, c), were used to identify periods of rhythmic theta frequency activity. Peaks of identified theta cycles were detected, and acceptable theta cycles were those whose periods fell within pre-set time windows (typically 176-272 ms for urethane data and 111-166 ms for walking data). Cycles were averaged by normalizing slow wave voltage data and spike data collected during each theta cycle, redistributing them into 32 histogram bins. Final scaling yielded the slow wave voltage at each 11.25 ~ segment of the average theta cycle. A mean phase angle was calculated for each unit by representing each phase bin as a vector whose length and phase are given by the height and phase angle of the bin. The mean phase for the unit is given by the angle of the resultant vector when all 32 phase bin vectors are plotted in polar coordinates. The length of the resultant vector is taken as the mean vector length for the unit and provides an estimate of modula- tion of the unit's firing. For example, if all action potentials from the unit fell into a single phase bin, the mean vector length would
be 1. Likewise, if the unit's action potentials were distributed evenly amongst all phase bins, the length of the mean vector would be zero.
Significant phase-locking of the unit to the theta rhythm was determined by the Runs (Siegel 1956; Fox et al. 1986) and Rayleigh (Batschelet 1981) tests. The mean phase for the dentate theta rhythm was used as the reference for summarizing the mean phases of the spike histograms for cells recorded in urethane anesthetized rats. CA1 theta rhythm was used as the reference for the walking data because it was not possible to ensure good placement of an electrode near the hippocampal fissure on the basis of stereotaxic coordinates. For comparison with the urethane data, summaries of the mean phases of spike histograms for maits collected in chroni- cally prepared animals are presented as if the CA1 and dentate signals were 180 ~ apart (Fox et al. 1986; Stewart and Fox 1989c).
Results
Acute experiments
A to ta l o f 23 cells were r eco rded f rom 11 ra ts anes- thet ized wi th u re thane . Al l cells were loca ted in layers I I and I I I o f med ia l en to rh ina l cor tex. On ly 8/23 h is to logi - cal ly identif ied cells f rom layers I I / I I I o f media l en torh ina l cor tex ( M E C ) were s ignif icant ly p h a s e - l o c k e d to the h i p p o c a m p a l the ta r h y t h m with a m e a n phase for the g roup o f 90 ~ The posi t ive p e a k o f the " d e n t a t e " the ta ( recorded at the h i p p o c a m p a l fissure) is def ined as 0 ~ Phase d a t a for the ind iv idua l cells are given in Tab le 1. R a w d a t a f rom one cell and a phase h i s t o g r a m for a second cell are shown in Fig. 1. As descr ibed in the M e t h o d s section, the m e a n phase for an ind iv idua l cell is der ived, in par t , f r om a phase h i s t o g r a m such as those i l lus t ra ted in Figs. 1 and 2. The m e a n phases for the g roup o f 8 pha se - l oc ke d M E C neurons in u r e thane are s u m m a r i z e d in Fig. 3 ( top). M e a n vec to r lengths, which p rov ide an es t imate o f the m o d u l a t i o n o f cell firing, were 0.21 _+ 0.08 and 0.08 _+ 0.05 for the pha se - l oc ke d cells and n o n p h a s e - l o c k e d cells, respect ively.
In several ra ts , a t t e m p t s were m a d e to a n t i d r o m i c a l l y ac t iva te M E C cells by app ly ing s t imuli to the den ta t e E E G record ing e lec t rode ( loca ted near the h i p p o c a m p a l fissure). One p h a s e - l o c k e d cell ou t o f the four cells tes ted cou ld be dr iven by s t imuli de l ivered t h r o u g h the den ta t e E E G elec t rode , ips i la te ra l to the e n to rh ina l uni t elec- t rode. The cell 's l oca t i on ( m a r k e d by a lesion) was in M E C layer II . A l t h o u g h it fo l lowed pa i rs o f s t imuli a t 1 kHz , col l i s ion o f the p r e s u m e d a n t i d r o m i c a l l y dr iven spikes, however , cou ld n o t be de mons t r a t e d .
F i r i ng ra tes for M E C cells du r ing h i p p o c a m p a l the ta r h y t h m (bo th phas ic and non -pha s i c cells) ave raged 8 . 6 + 7 . 7 spikes/s. To faci l i ta te c o m p a r i s o n across p r e p a r a t i o n s wi th different the ta r h y t h m frequencies , this ra te conver t s to an average o f 1.9 :t: 1.7 spikes/cycle. The m e d i a n ra te was 5.9 spikes/s ( range : 0 .1-29.3 spikes/s) . There were no d is t inc t g roups o f slow a n d fast f ir ing M E C cells. The d i s t r i bu t ion o f firing ra tes was u n i m o d a l and skewed to the r ight . In add i t ion , none o f ou r layer I I / I I I cells were seen to fire complex-sp ikes . Since we have li t t le d a t a f rom cells in the deeper layers , we c a n n o t dismiss the existence o f complex - sp ike cells in the en tor -
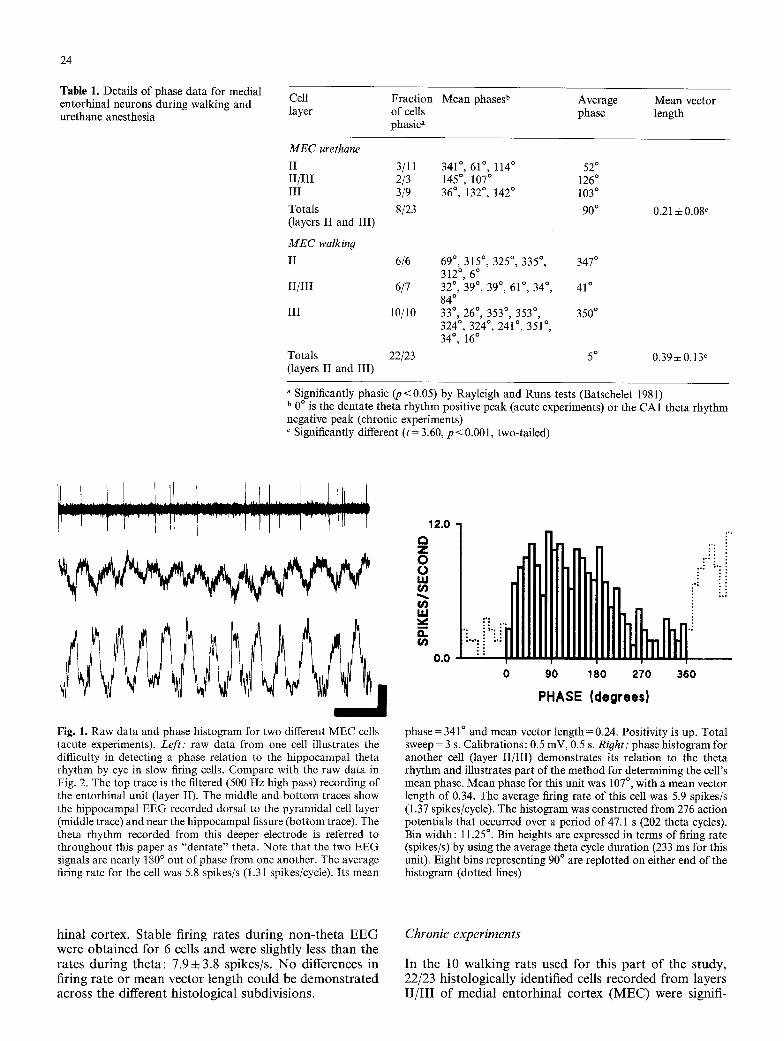
24
Table 1. Details of phase data for medial entorhinal neurons during walking and urethane anesthesia
Cell layer
Fraction Mean phases b Average of cells phase phasic"
Mean vector length
MEC urethane II II/III III
Totals (layers II and III)
MEC walking II
II/III
III
3/11 341 ~ , 61 ~ , 114 ~ 52 ~ 2/3 145 ~ , 107 ~ 126 ~ 3/9 36 ~ , 132 ~ , 142 ~ 103 ~
8/23 90 ~
6/6 69 ~ , 315 ~ , 325 ~ , 335 ~ , 347 ~ 312 ~ 6 ~
6/7 32 ~ , 39 ~ , 39 ~ , 61 ~ , 34 ~ , 41 ~ 84 ~
10/10 33 ~ 26 ~ 353 ~ 353 ~ 350 ~ 324 ~ , 324 ~ , 241 ~ , 351 ~ , 34 ~ ' 16 ~
0.21 • 0.08 ~
Totals 22/23 5 ~ 0.39 + 0.13 c (layers II and III)
" Significantly phasic (p < 0.05) by Rayleigh and Runs tests (Batschelet 1981) b 0 o is the dentate theta rhythm positive peak (acute experiments) or the CA1 theta rhythm negative peak (chronic experiments) c Significantly different ( t= 3.60, p < 0.001, two-tailed)
Fig. 1. Raw data and phase histogram for two different MEC cells (acute experiments). Left: raw data from one cell illustrates the difficulty in detecting a phase relation to the hippocampal theta rhythm by eye in slow firing cells. Compare with the raw data in Fig. 2. The top trace is the filtered (500 Hz high pass) recording of the entorhinal unit (layer II). The middle and bot tom traces show the hippocampal EEG recorded dorsal to the pyramidal cell layer (middle trace) and near the hippocampal fissure (bottom trace). The theta rhythm recorded from this deeper electrode is referred to throughout this paper as "dentate" theta. Note that the two EEG signals are nearly 180 ~ out of phase from one another. The average firing rate for the cell was 5.8 spikes/s (1.31 spikes/cycle). Its mean
12.0 C~ Z 0 0 UJ
t~ 1.1.1
a .
0 , 0 i i i
i
0
i" ii " i i i i
90 180 270 360
PHASE {degrees}
phase= 341 ~ and mean vector length=0.24. Positivity is up. Total sweep= 3 s. Calibrations: 0.5 mV, 0.5 s. Right: phase histogram for another cell (layer II/III) demonstrates its relation to the theta rhythm and illustrates part of the method for determining the cell's mean phase. Mean phase for this unit was 107 ~ with a mean vector length of 0.34. The average firing rate of this cell was 5.9 spikes/s (1.37 spikes/cycle). The histogram was constructed from 276 action potentials that occurred over a period of 47.1 s (202 theta cycles). Bin width: 11.25 ~ Bin heights are expressed in terms of firing rate (spikes/s) by using the average theta cycle duration (233 ms for this unit). Eight bins representing 90 ~ are replotted on either end of the histogram (dotted lines)
h i n a l co r t ex . S t a b l e f i r i ng r a t e s d u r i n g n o n - t h e t a E E G w e r e o b t a i n e d f o r 6 cel ls a n d w e r e s l i gh t ly less t h a n t h e r a t e s d u r i n g t h e t a : 7.9 + 3.8 sp ike s / s . N o d i f f e r ences in f i r ing r a t e o r m e a n v e c t o r l e n g t h c o u l d b e d e m o n s t r a t e d a c r o s s t h e d i f f e r e n t h i s t o l o g i c a l s u b d i v i s i o n s .
Chronic experiments
I n t h e 10 w a l k i n g r a t s u s e d f o r t h i s p a r t o f t h e s t u d y , 22 /23 h i s t o l o g i c a l l y i d e n t i f i e d cells r e c o r d e d f r o m l a y e r s I I / I I I o f m e d i a l e n t o r h i n a l c o r t e x ( M E C ) w e r e s ignif i -

25
' , :i :"i " ~' ! ' "
Fig. 2. Raw data and phase histogram for two different MEC cells (chronic experiments). Left: raw data for one MEC cell (top trace) and the simultaneously recorded CA1 EEG (bottom trace). The phase relation of this cell (layer III) to the hippocampal theta rhythm is readily apparent in the raw data. Note: negativity is up for this figure. Mean phase = 353~ mean vector length= 0.50. The average firing rate of this cell (30 spikes/s) is much higher than the cell shown in Fig. 1 because the theta rhythm frequency is higher
74.0 a Z 0 L) IJJ r
r g,I
a . r
0.0
' L i ,~!. ; , t , , ,
~.;
i 0
ii i 90 180 270 360
PHASE (degrees} and the cell fires more spikes per cycle (4.1 spikes/cycle). Calibra- tions: 0.5 mV, 0.2 s. Total sweep= 2 s. Right: phase histogram for a different cell (layer II) showing its pronounced phase relation to the theta rhythm. Mean phase = 315~ mean vector length= 0.47. The firing rate of the cell was 34 spikes/s (4.63 spikes/cycle). Histo- gram constructed from 1751 spikes occurring over 51.5 s (378 theta cycles). Average cycle duration= 136 ms. Bin width= 11.25 ~
r . - I . - I UJ 0
d z
MEC urethane
i i i - 1 8 0 - 1 3 5 - 9 0 - 4 5 0 45
PHASE [degrees]
90 135 180
12
- J --J LM
d z
10 MEC walking
- 1 8 0 - 1 3 5 - 9 0 - 4 5 0 45
PHASE (degrees)
i i
90 135 180
Fig. 3. Summary phase h is tograms for acute and chronic experi- ments. Top: phase dis t r ibut ion for 8 phase- locked medial entorhi- nal neurons recorded in ure thane anesthet ized rats. The denta te theta rhy thm posit ive peak is at 0 ~ The mean phase for the group was 90 ~ . Bottom: same as above but for cells recorded during walking behaviors. The mean phase for the group was 5 ~ . Fo r addi t ional details, see Table 1
cantly phase-locked to the h ippocampal theta rhythm. Raw data f rom one identified M E C neuron are shown in Fig. 2 together with a phase his togram f rom a second cell. The difference in rhythmic "quali ty" for M E C cells in walking and anesthetized rats is dramatic (compare Figs. 1 and 2). The mean phase for the group of 22 layer I I / I I I cells was 5 ~ (0 ~ dentate theta rhythm positive peak), summarized in Fig. 3 (bottom). Mean vector lengths were 0.39 + 0.13 for the 22 phasic cells and 0.10 for the single non-phasic cell.
The average firing rate for the entire group of cells during theta rhythm was 11.8 4- 11.8 spikes/s (1.6 + 1.6 spikes/ cycle), with a median rate of 6.6 spikes/s. The range of firing rates was 0.8-34.8 spikes/s. Like the data f rom M E C cells in urethane, the distribution of firing rates was unimodal and skewed to the right. Although medial entorhinal cells did seem to fire faster in walking rats (if only because of the higher theta frequency) no differences could be demonstrated in the firing rates of M E C cells between walking and urethane anesthesia. The spikes/ cycle values (urethane: 1.9_+ 1.7; walk: 1.6_+ 1.6) were virtually identical.
The propor t ion of phase-locked cells in walking rats did, however, differ significantly f rom the propor t ion obtained f rom urethane anesthetized rats (z=4.38, p<0.0001, two-tailed; McNemar 1969). The average mean vector lengths for the groups of phasic cells differed as well: 0.394-0.13 for walking rats compared to 0.214-0.08 for urethane anesthetized rats ( t=3.60, p < 0.01, two-tailed). These last two measures confirm the superior rhythmic quality of the M E C output in walking rats that was visible in the raw data of Figs. 1 and 2.
Discussion
E• recordings taken f rom single medial entor- hinal neurons (layers I I / I I I ) in walking and urethane

26
anesthetized rats have revealed the following: 1) there exist phase-locked entorhinal units in both of our experi- mental conditions, confirming an entorhinal contribu- tion to theta rhythm generation in walking and urethane anesthetized rats; 2) the distribution of preferred firing phases for MEC cells in walking and urethane were similar to the firing phases for dentate neurons, suggest- ing that medial entorhinal neurons are the principal de- terminant of theta-related firing of hippocampal neurons and that they generate much of the theta-related current sink in the mid-molecular layer of the dentate; and 3) more cells were phase-locked in walking rats and their rhythmic firing was more striking. These differences in rhythmic "quality" may account for EEG differences across the two conditions, such as the prominence of an atropine-resistant component of the walking theta rhythm.
Entorhinal control of hippocampal unit activity
As reviewed in the Introduction, the principal cortical input to the dentate gyrus are the entorhinal cortices. Surgical ablation of this input (Reeves and Steward 1988) or entorhinal injections of tetrodotoxin (Tomasulo et al. 1991) suppress dentate cell firing. These data, taken with the more proximal zone of termination on granule cell dendrites of the medial entorhinal inputs, lead one to expect MEC inputs to be more influential than LEC inputs in determining the firing of dentate neurons (see e.g. Wyss 1981).
Medial entorhinal cortical cells were found to fire near the positive peak of the dentate theta rhythm in both walking and urethane anesthetized rats. The distribution of preferred firing phases for MEC cells during walking behaviors was very clear: 18/22 cells fired within 45 ~ of the dentate theta rhythm positive peak. The positive peak is also the preferred phase for hippocampal granule and pyramidal cell firing during walking (Buzsaki et al. 1983, 1986; Fox et al. 1986). For walking rats, there is crude agreement across laboratories on the firing phases of hippocampal neurons. Small differences in the reported average firing phases for cells in the different hippocam- pal areas depend in part on their being averages for many cells, and partly on differences in techniques between labs for determining the preferred phase for individual cells. For such reasons, it is probably unreasonable to expect to see timing differences (e.g. MEC cells leading dentate cells, dentate cells leading CA3 cells, etc.) that would be on the order of a fraction of the theta cycle.
In urethane anesthetized rats, the phase-locked firing of MEC cells was much less robust, but averaged to 90 ~ for the group. Consistent with these data is the finding that the firing phases of individual dentate and CA3 neurons in urethane anesthetized rats are also more wide- ly scattered when compared to the walking data (Fox et al. 1986). Some dentate cells have been shown to fire closer to the negative peak of the dentate theta rhythm (Buzsaki et al. 1983 ; Fox et al. 1986). From the summary figure (Fig. 3, top) it can be seen that some of our MEC cells fire closer to the dentate negative peak (180~ This phase is also coincident with the firing of phase-locked lateral entorhinal neurons (Stewart et al. 1991). For rea- sons that remain unclear, control of dentate cell firing by
medial entorhinal cortex is decidedly weaker in urethane anesthetized animals. Urethane itself may be largely re- sponsible, perhaps through an action on glutamate activ- ity (Moroni et al. 1981). Urethane has been found to abolish the perforant path to dentate evoked response in vitro (M Stewart and IS Kass, unpublished observations) and Buzsaki et al. (1986) showed a dramatic decrease in the overall firing rate of dentate neurons when chroni- cally implanted rats were given urethane.
Entorhinal contributions to hippocampal field potentials
Buzsaki et al. (1986) described the current sources and sinks associated with the hippocampal theta rhythm in walking rats. A large extracellular current sink was found in the distal apical dendrites of dentate granule cells coincident with the peak negativity of the dentate theta rhythm (see also Branka~k and Fox 1987). Deep to this region, they demonstrated a sink at the level of the EPSP evoked by perforant path stimulation. Projections from the medial entorhinal cortex appear to make up the largest portion of the perforant path input (Hjorth- Simonsen 1972; Hjorth-Simonsen and Jeune 1976; Ste- ward and Scoville 1976). During the hippocampal theta rhythm, a sink appeared at this level 30-60 ms earlier than the maximal sink in the distal apical dendrites (Buzsaki et al. 1986). This time turncd out to be very near the positive peak of the dentate theta rhythm. Given a mean phase of 5 ~ for the group of medial entorhinal neurons during walking theta rhythm, this sink is likely the result of medial entorhinal inputs to the dentate.
Unlike the current source density values calculated from AC-coupled depth profiles, DC-coupled recordings do not constrain the CSD values to alternate between sink and source at a particular lamina. CSD data from our lab using DC-coupled recordings (Fox and Stewart 1986; Branka~k and Fox 1987) show the peak of the extracellular current sink located in the midmolecular layer of the dentate gyrus to occur at 300 ~ almost a quarter cycle ahead of the positive peak (0 ~ dentate theta positive peak). This phasic current sink rides atop a substantial DC component that may result from some firing of MEC cells throughout the cycle. Other con- tributors to the CSD at this level remain unclear.
A limited preliminary sample of lateral entorhinal neurons (Stewart et al. 1991) suggests that the firing of LEC cells may account for the large current sink occur- ring near the hippocampal fissure at the negative peak of the dentate theta rhythm (Buzsaki et al. 1986; Fox and Stewart 1986; Branka6k and Fox 1987). Data from lat- eral entorhinal neurons in freely moving rats are essential for confirming any contributions to theta generation or assessing their relative contributions (compared with medial entorhinal neurons) to hippocampal unit activity.
Entorhinal cortex and the atropine-resistant theta rhytm
The discovery of atropine-resistant and atropine- sensitive components of the hippocampal theta rhythm (Kramis et al. 1975) begged for the elucidation of the cellular bases of these two components (Buzsaki et al. 1983; Vanderwolf 1988). Over the past 30 years, the role of the medial septal nuclei in the production of the hippo-

27
campal theta rhythm has become clearer (reviewed in Stewart and Fox 1990). It appears that rhythmically bursting medial septal cells act as "pacemakers" for at least the atropine-sensitive (cholinergic) theta com- ponent. Vanderwolf and others have suggested that a separate, non-septal system mediates the atropine-resis- tant theta component. Lesion studies showed that bilat- eral destruction of the entorhinal cortices (Vanderwolf and Leung 1983) or lesions of entorhinal afferents (Van- derwolf et al. 1985) eliminated the atropine-resistant component of the hippocampal theta rhythm. There was no apparent effect of these lesions on the atropine- sensitive theta component. On the basis of these data and manipulations of whole-brain serotonergic activity, a brainstem-hippocampal system, passing through the en- torhinal cortex, was argued to mediate the atropine-resis- tant theta rhythm (reviewed in Vanderwolf 1988).
Our finding of rhythmic MEC units in walking and urethane anesthetized rats reemphasizes the role of en- torhinal cortex in the generation of theta rhythm in both walking and urethane anesthetized animals. The most striking result of our experiments, however, is the dif- ference in rhythmic quality of MEC neurons in walking as compared to urethane. This difference supports a distinction between atropine-resistant and atropine- sensitive theta components based largely on the rhyth- micity of entorhinal cortical cells: pronounced entorhi- nal activity contributes to atropine-resistant theta rhythm and diminished entorhinal activity leaves the atropine-sensitive theta component to dominate. More- over, the differences in entorhinal rhythmicity between walking and urethane anesthetized rats obviates, but cannot exclude, a role for direct non-septal inputs to the hippocampus for the atropine-resistant theta rhythm.
Two broad categories of possibilities exist to account for the enhanced rhythmicity of MEC neurons in walk- ing rats. First, there may be a fundamental difference in the phasic firing of medial entorhinal neurons in the two conditions. The notion of separate pacemaker systems (described above) is in this category. While a non-septal (brainstem) "pacemaker" system is possible, we feel it is unlikely, primarily because clear "pacemaker" activity has not yet been demonstrated in any of the brainstem afferents. Alternatively, the firing rate changes that have been reported for certain brainstem areas (Sheu et al. 1974; Vertes 1979) may gate on and off the intrinsic membrane oscillations that were seen in layer II cells in undrugged entorhinal slices (Alonso and Llinfis 1989). The gating of intrinsic oscillatory activity could operate in complete isolation from rhythmic medial septal af- ferent activity. In walking animals, concurrent activity in scptal afferents (Alonso and K6hler 1984) may serve only to entrain the rhythmic entorhinal activity.
The second category supposes that the differences between urethane and walking MEC firing patterns are quantitative rather than qualitative. We have argued that the activity of the medial septum is unchanged from walking to urethane anesthetized rats (Stewart and Fox 1990; see also Givens and Breese 1990). It is entirely possible that depression of glutamatergic activity by urethane (Moroni et al. 1981) impairs the septum's abil- ity to organize the activity of entorhinal (and hippocam- pal) neurons, hence the weaker rhythmic entorhinal (and
hippocampal) output. The septum could act as the "pacemaker" for both atropine-resistant and atropine- sensitive theta components: the atropine-sensitive com- ponent resulting from a direct cholinergic activation of hippocampal theta cells (interneurons), and the atropine- resistant component arising from rhythmic septal inhibi- tion of baseline activity in hippocampus and entorhinal cortex. Loss of this activity baseline, either by elimina- tion of entorhinal excitatory afferents or the attenuation of glutamate synapses by urethane, prevents a prominent atropine-resistant theta component from being detected in the hippocampus. Averaging of the hippocampal EEG triggered on the rhythmic activity of atropine-resistant septal cells permitted the detection of an atropine-resis- tant theta component of the EEG in urethane anes- thetized rats (Stewart and Fox 1989b). More recently, we have found that medial septal cells remain phase-locked to the hippocampal theta rhythm in walking rats even after massive doses of atropine (C Jia and SE Fox, un- published observations). The relation of medial septal neurons to both theta components leads us to interpret the differences seen in entorhinal activity between walk- ing and urethane anesthetized animals as a quantitative effect of urethane on the cortical targets of a single septo-hippocampal "pacemaker" and not a qualitative action of urethane on a separate non-septal "pacemaker" system.
Identification of the specific contributions to the hippocampal theta rhythm made by septal and brainstem systems is hindered by their considerable overlap. Many brainstem affcrcnts asccnd through the septal nuclei, and septal and brainstem "pacemaker" systems appear to act at both the hippocampus and the entorhinal cortex. Be- cause we have found septal units locked to atropine-resis- tant theta components in walking and urethane anes- thetized rats, it is our belief that septum "paces" all theta rhythm components. We cannot, however, rule out the possibility that the non-cholinergic septal output acts together with some non-septal system to cause rhythmic firing in entorhinal neurons. Until additional data be- come available, including the actions of serotonin on the two types of rhythmically bursting medial septal cells (Stewart and Fox 1989a) and layer II/III entorhinal neu- rons, the precise origin of the atropine-resistant theta rhythm remains uncertain.
Acknowledgements. This work was supported by National Institutes of Health grants: NS17095, NS07117, NS20686, and NS14497.
References
Alonso A, Garcia-Austt E (1987a) Neuronal sources of theta rhythm in the entorhinal cortex of the rat. I. Laminar distribu- tion of theta field potentials. Exp Brain Res 67:493-501
Alonso A, Garcia-Austt E (1987b) Neuronal sources of theta rhythm in the entorhinal cortex of the rat. II. Phase relations between unit discharges and theta field potentials. Exp Brain Res 67: 502-509
Alonso A, K6hler C (1984) A study of the reciprocal connections between the septum and the entorhinal area using anterograde and retrograde axonal transport methods in the rat brain. J Comp Neurol 225:327-343
Alonso A, Lllnas RR (1989) Subthreshold Na§ theta- like rhythmicity in stellate cells of entorhinal cortex layer II. Nature 342:175-177

28
Andersen P, Holmqvist B, Voorhoeve PE (1966a) Entorhinal ac- tivation of dentate granule cells. Acta Physiol Scand 66:448-460
Andersen P, Holmqvist B, Voorhoeve PE (1966b) Excitatory synap- ses on hippocampal apical dendrites activated by entorhinal stimulation. Acta Physiol Scand 66:461-472
Batschelet E (1981) Circular statistics in biology. Academic, Lon- don
Branka~k J, Fox SE (1987) Current sources for the alternating and sustained potentials of the hippocampal theta rhythms of the rat. Soc Neurosci Abstr 13:1331
Buzsaki G, Czopf J, Kondakor I, Kellenyi L (1986) Laminar distri- bution of hippocampal rhythmic slow activity (RSA) in the behaving rat: current-source density analysis, effects of urethane and atropine. Brain Res 365:125-137
Buzsaki G, Leung LS, Vanderwolf CH (1983) Cellular bases of hippocampal EEG in the behaving rat. Brain Res Rev 6:13%171
Doller HJ, Weight FF (1982) Perforant pathway activation of hippocampal CA1 stratum pyramidale neurons: electrophys- iological evidence for a direct pathway. Brain Res 237:1-13
Dudek FE, Deadwyler SA, Cotman CW, Lynch G (1976) Intracel- lular responses from granule cell layer in slices of rat hippocam- pus: perforant path synapse. J Neurophysiol 39:384-393
Fox, SE (1985) Location of membrane conductance changes by analysis of the input impedance of neurons. I. Theory. J Neu- rophysiol 54:1578-1593
Fox, SE (1989) Membrane potential and impedance changes in hippocampal pyramidal cells during theta rhythm. Exp Brain Res 77: 283-294
Fox SE, Stewart M (1986) Analysis of hippocampal theta rhythm shows that correct location of current sources and sinks generat- ing EEG patterns can require DC recording. Soc Neurosci Abstr 12:1527
Fox SE, Wolfson S, Ranck JB Jr (1986) Hippocampal theta rhythm and the firing of neurons in walking and urethane anestherized rats. Exp Brain Res 62:495-508
Germroth P, Schwerdtfeger WK, Buhl EH (1989) Morphology of identified entorhinal neurons projecting to the hippocampus. A light microscopical study combining retrograde tracing and intracellular injection. Neuroscience 30:683-691
Givens BS, Breese GR (1990) Electrophysiological evidence that ethanol alters function of medial septal area without affecting lateral septal function. J Pharm Exp Ther 253:95-103
Green KF, Rawlins JNP (1979) Hippocampal theta in rats under urethane: generators and phase relations. EEG Clin Neuro- physiol 47: 420-429
Hjorth-Simonsen A (1972) Projection of the lateral part of the entorhinal area to the hippocampus and fascia dentata. J Comp Neurol 146: 219-232
Hjorth-Simonsen A, Jeune B (1972) Origin and termination of the hippocampal perforant path in the rat studied by silver im- pregnation. J Comp Neurol 144:215 232
Kramis R, Vanderwolf CH, Bland BH (1975) Two types of hippo- campal rhythmical slow activity in both the rabbit and the rat: relations to behavior and effects of atropine, diethyl ether, urethane, and pentobarbital. Exp Neurol 49:58-85
Kubie JL (1984) A driveable bundle of microwires for collecting single unit data from freely moving rats. Physiol Behav 32:115-118
McNemar Q (1969) Psychological statistics, fourth edition. Wiley, New York, p 62
Mitchell SJ, Ranck JB Jr (1980) Generation of theta rhythm in medial entorhinal cortex of freely moving rats. Brain Res 189: 49-66
Mitchell SJ, Rawlins JNP, Steward O, Olton DS (1982) Medial septal area lesions disrupt theta rhythm and cholinergic staining in medial entorhinal cortex and produce impaired radial arm maze behavior in rats. J Neurosci 2: 292-302
Moroni F, Corradetti R, Casamenti F, Moneti G, Pepeu G (1981) The release of endogenous GABA and glutamate from the cerebral cortex in the rat. Naunyn-Schmiedebergs Arch Pharmacol 316:235-239
Muller RU, Kubie JL, Ranck JB Jr (1987) Spatial firing patterns of hippocampal complex-spike cells in a fixed environment. J Neurosci 7:1935-1950
Quirk G J, Muller RU, Kubie JL, Ranck JB Jr (1992) The positional firing properties of medial entorhinal neurons: description and comparison with hippocampal place cells. J Neurosci 12:1945-1963
Quirk GJ, Ranck JB Jr (1986) Firing of single cells in entorhinal cortex is location specific and phase-locked to the hippocampal theta rhythm. Soc Neurosci Abstr 12:1524
Quirk GJ, Stewart M (1988) Neurons in entorhinat cortex of etherized rats exhibit rhythmic bursting correlated with the hippocampal EEG. Ann NY Acad Sci 529:310-313
Paxinos G, Watson C (1986) The rat brain in stereotaxic coordina- tes, 2nd. edn. Academic, Sydney
Ranck JB Jr (1973) Studies on single neurons in dorsal hippocampal formation and septum in unrestrained rats. I. Behavioral cor- relates and firing repertoires. Exp Neurol 41:461 531
Reeves TM, Steward O (1988) Changes in the firing properties of neu- rons in the dentate gyrus with denervation and reinnervation: Implications for behavioral recovery. Exp Neurol 102:37-49
Sheu Y, Nelson JP, Bloom FE (1974) Discharge patterns of cat raphe neurons during sleep and waking. Brain Res 73 : 263-276
Siegel S (1956) Nonparametric statistics for the behavioral sciences. McGraw Hill, New York pp 52-58
Steward O (1976) Topographic organization of the projections from the entorhinal area to the hippocampal formation of the rat. J Comp Neurol 167:285-314
Steward O, Scoville SA (1976) Cells of origin of entorhinal cortical afferents to the hippocampus and fascia dentata of the rat. J Comp Neurol 169:347-370
Stewart M, Fox SE (1989a) Two populations of rhythmically burst- ing neurons in the rat medial septum are revealed by atropine. J Neurophysiol 61:982-993
Stewart M, Fox SE (1989b) Detection of an atropine-resistant component of the hippocampal theta rhythm in urethane anes- thetized rats. Brain Res 500:55-60
Stewart M, Fox SE (1989c) Firing relations of medial septal neu- rons to the hippocampal theta rhythm in urethane anesthetized rats. Exp Brain Res 77:507-516
Stewart M, Fox SE (1990) Do septal neurons pace the hippocampal theta rhythm? Trends Neurosci 13:163-168
Stewart M, Barry M, Quirk GJ, Fox SE (1991) Firing relations of entorhinal neurons to the hippocampal theta rhythm in walking and urethane anesthetized rats. Soc Neurosci Abstr 17:1036
Tomasulo RA, Burger B, Steward O (1991) Injection of tet- rodotoxin into the entorhinal cortex suppresses cell firing in the dentate gyrus. Exp Neurol 111 : 340-348
Vanderwolf CH (1988) Cerebral activity and behavior: control by central cholinergic and serotonergic systems. Int Rev Neurobiol 30: 225-340
Vanderwolf CH, Baker GB (1986) Evidence that serotonin mediates non-cholinergic neocortical low voltage fast activity, non-cho- linergic hippocampal rhytmical slow activity and contributes to intelligent behavior. Brain Res 374:342-356
Vanderwolf CH, Leung LS (1983) Hippocampal rhythmical slow activity: a brief history and the effects of entorhinal lesions and phencyclidine. In: Seifert W (ed) The neurobiology of the hippo- campus. Academic, London, pp 275-302
Vanderwolf CH, Leung, LS, Cooley RK (1985) Pathways through cingulate, neo- and entorhinal cortices mediate atropine-resis- tant hippocampal rhythmical slow activity. Brain Res 347 : 58-73
Vertes RP (1979) Brain stem gigantocellular neurons: patteins of activity during behavior and sleep in the freely moving rat. J Neurophysiol 42:214-228
White WF, Nadler JV, Hamberger A, Cotman CW, Cummins JT (1977) Glutamate as transmitter of hippocampal perforant path. Nature 270:356-357
Wyss JM (1981) An autoradiographic study of the efferent connec- tions of the entorhinal cortex in the rat. J Comp Neurol 199:495-512


















