
Fine structure, distribution and frequency of plasmodesmata and pits in the cortex ofLaminaria...
8
Planta (1982)154:385-392 Planta Springer-Verlag 1982 Fine structure, distribution and frequency of plasmodesmata and pits in the cortex of Laminaria hyperborea and L. saccharina Klaus Schmitz and Rainer Kfihn Botanisches Institut der Universit~it, Gyrhofstrasse 15, D-5000 K61n 41, Federal Republic of Germany Abstract. Prior to a long-distance transport of pho- toassimilate in the sieve elements of Laminaria, a pa- renchyma transport across the cortex must occur. It is suggested that this transport is a symplastic one. The structural basis for this statement, continuous cytoplasmic interconnections of cells along the trans- port pathway, is demonstrated here for L. hyperborea and L. saccharina. The distribution, size, and fre- quency of pit fields in cell walls of all planes were determined. The data suggest that the conductivity for assimilate transport in the cortex is highest in the long axis of the thallus, not radially across the cortex. The fine structure, arrangement and number of plasmodesmata in pit fields were studied. The esti- mated flux rates and the anatomical findings clearly point to a symplastic parenchyma transport of pho- toassimilate in the cortex of Laminaria. Key words: Laminaria - Parenchyma transport - Phaeophyta - Pit fields - Plasmodesmata - Symplastic transport. Introduction Long-distance transport of photoassimilate and inor- ganic ions is a common feature of the Laminariales (Parker 1965; Nicholson and Briggs 1972; Floc'h and Penot 1972; Schmitz and Lobban 1976). Transloca- tion of ~4C-labeled photosynthate was repeatedly demonstrated in the genus Laminaria (Lfining et al. 1972; Schmitz et al. 1972; Ltining et al. 1973; Willen- brink et al. 1975; Schmitz and Lobban 1976), and it was shown by histoautoradiography that long-dis- tance transport occurs in the sieve element system in the medulla (Steinbil3 and Schmitz 1973). The photosynthetic capacity of a Laminaria thal- lus is nearly exclusively confined to the meristoderm and the outermost layer of cortex cells (Hellebust and Haug 1972) separated from the conducting tissue in the medulla by several layers of parenchyma cortex cells. Prior to a long-distance transport of photosyn- thate in the sieve elements, a medium-distance trans- port between the phaeoplast-rich meristoderm and the sieve elements in the medulla must occur. A time- dependent advancing front of 14C was previously re- corded by autoradiography after radiocarbon had been applied to the thallus surface (Steinbig and Schmitz 1973). An identification of the medium-dis- tance transport pathway however was not achieved by this technique. There are basically two pathways for this paren- chyma transport: the symplast and/or the apoplast. A prerequisite for symplastic transport is the continu- ous cytoplasmic interconnection of cells along the transport pathway. It was the aim of this investigation to trace an uninterrupted cytoplasmic continuum between the meristoderm and the long-distance transport system in the medulla and to investigate the fine structure, arrangement, and frequency of plasmodesmata. On the basis of the data obtained, the mode of cortex parenchyma transport of photoassimilate is discussed. Material and methods Laminaria saccharina (L.) Lamour. and L. hyperborea (Gunn.) Fosl. (Phaeophyta, Laminariales) were taken from a natural habitat of the subtidal zone near Helgoland, North Sea in February, April, and July 1977. Plants were selected for uniformity in length, mor- phology, and cleanliness. Small pieces of stipe, intercalary growing region, and mature phylloids were fixed in glutaraldehyde/osmiumtetroxideand either embedded in epon-araldite for transmission electron microscopy, as described previously (Schmitz and Srivastava 1974), or dehy- drated via aceton and amylacetate followed by critical point drying with CO> Gold-coated specimensof dried samples were examined with a scanning electron microscope (ETEC-Biosem). Moreover, rapidly frozen tissue samples (melting nitrogen) were lyophilized and gold coated for scanning electron microscopy, or the frozen tissue was sectioned with a cryotome for light microscopy. The 0032-0935/82/0154/0385/$01.60
-
Upload
klaus-schmitz -
Category
Documents
-
view
214 -
download
2
Transcript of Fine structure, distribution and frequency of plasmodesmata and pits in the cortex ofLaminaria...

Planta (1982)154:385-392 Planta �9 Springer-Verlag 1982
Fine structure, distribution and frequency of plasmodesmata and pits in the cortex of Laminaria hyperborea and L. saccharina
Klaus Schmitz and Rainer Kfihn Botanisches Institut der Universit~it, Gyrhofstrasse 15, D-5000 K61n 41, Federal Republic of Germany
Abstract. Prior to a long-distance transport of pho- toassimilate in the sieve elements of Laminaria, a pa- renchyma transport across the cortex must occur. It is suggested that this t ransport is a symplastic one. The structural basis for this statement, continuous cytoplasmic interconnections of cells along the trans- port pathway, is demonstrated here for L. hyperborea
and L. saccharina. The distribution, size, and fre- quency of pit fields in cell walls of all planes were determined. The data suggest that the conductivity for assimilate t ransport in the cortex is highest in the long axis of the thallus, not radially across the cortex. The fine structure, arrangement and number of plasmodesmata in pit fields were studied. The esti- mated flux rates and the anatomical findings clearly point to a symplastic parenchyma transport of pho- toassimilate in the cortex of Laminaria.
Key words: Laminaria - Parenchyma transport - Phaeophyta - Pit fields - Plasmodesmata - Symplastic transport.
Introduction
Long-distance transport of photoassimilate and inor- ganic ions is a common feature of the Laminariales (Parker 1965; Nicholson and Briggs 1972; Floc 'h and Penot 1972; Schmitz and Lobban 1976). Transloca- tion of ~4C-labeled photosynthate was repeatedly demonstrated in the genus Laminaria (Lfining et al. 1972; Schmitz et al. 1972; Ltining et al. 1973; Willen- brink et al. 1975; Schmitz and Lobban 1976), and it was shown by histoautoradiography that long-dis- tance transport occurs in the sieve element system in the medulla (Steinbil3 and Schmitz 1973).
The photosynthetic capacity of a Laminaria thal- lus is nearly exclusively confined to the meristoderm and the outermost layer of cortex cells (Hellebust
and Haug 1972) separated from the conducting tissue in the medulla by several layers of parenchyma cortex cells. Prior to a long-distance t ransport of photosyn- thate in the sieve elements, a medium-distance trans- port between the phaeoplast-rich meristoderm and the sieve elements in the medulla must occur. A time- dependent advancing front of 14C was previously re- corded by autoradiography after radiocarbon had been applied to the thallus surface (Steinbig and Schmitz 1973). An identification of the medium-dis- tance transport pathway however was not achieved by this technique.
There are basically two pathways for this paren- chyma transport: the symplast and/or the apoplast. A prerequisite for symplastic t ransport is the continu- ous cytoplasmic interconnection of cells along the transport pathway.
It was the aim of this investigation to trace an uninterrupted cytoplasmic continuum between the meristoderm and the long-distance transport system in the medulla and to investigate the fine structure, arrangement, and frequency of plasmodesmata. On the basis of the data obtained, the mode of cortex parenchyma transport of photoassimilate is discussed.
Material and methods
Laminaria saccharina (L.) Lamour. and L. hyperborea (Gunn.) Fosl. (Phaeophyta, Laminariales) were taken from a natural habitat of the subtidal zone near Helgoland, North Sea in February, April, and July 1977. Plants were selected for uniformity in length, mor- phology, and cleanliness.
Small pieces of stipe, intercalary growing region, and mature phylloids were fixed in glutaraldehyde/osmiumtetroxide and either embedded in epon-araldite for transmission electron microscopy, as described previously (Schmitz and Srivastava 1974), or dehy- drated via aceton and amylacetate followed by critical point drying with CO> Gold-coated specimens of dried samples were examined with a scanning electron microscope (ETEC-Biosem). Moreover, rapidly frozen tissue samples (melting nitrogen) were lyophilized and gold coated for scanning electron microscopy, or the frozen tissue was sectioned with a cryotome for light microscopy. The
0032-0935/82/0154/0385/$01.60

386 K. Schmitz and R. Ktihn: Symplast in the cortex of Laminaria
thickness of cryosections was 20 40 ~tm, but could be reduced to < 10 gm. Cell walls were stained with toluidine blue (Ito and Win- chester 1963) or ruthenium red in order to increase the contrast of cell walls, for better recognition of pits.
The distribution, frequency, and diameter of pits were studied fromadequately stained cryosections. Anticlinal cross sections, para- dermal and radial longitudinal sections provided surface views of correspondingly oriented cell walls. For each dimension, 50 surface views of cell walls from the mature blade, the growing region and the stipe of both species were examined, the pits were counted, their diameter determined, and the surface area of each cell wall calculated. The determination of wall areas was difficult due to their variable and irregular forms. Anticlinal cross walls of cortex cells appeared polygonal to round, their area therefore was consid- ered to be a circular one. Longitudinal cell walls of mature phyl- loids and growing regions are square, hexagonal, Or slightly rectan-
gular, with their long axis up to 1.5 times their width. The area of such cell walls was uniformly considered to be circular and calculated on the basis of the measured average diameter. The data therefore are sIightly underestimated. Longitudinal cortex ceil walls of s t ipes are more or less rectangular and up to 5 times longer than they are wide. The area therefore had to be calculated for each wall, assuming it was an ideal rectangle.
Results
The internal organization of the thallus. The tissue topography of a Laminaria thallus is illustrated in Fig. 1 a, b. It is basically the same in the blade and in the stipe. Cross sections of stipes reveale a radial
Fig. 1 a. Scanning electron micrograph of a radial longitudinal section through a blade of Laminaria hyperborea. Tissue arrangement is lateraly symmetric. M meristoderm; Md medulla; SD slime duct. Magnification b a r = 200 gm
Fig. I b. Cross section of a stipe of L. hyperborea. Thissue topography is radially symmentric here. The lense shaped medulla (Md) is surrounded by a multilayered cortex (C); the densely stained periphery indicated by the white arrow is composed of the meristoderm (NO and the outermost cortex cells which contain numerous phaeoplasts. Magnification bar = 1 m m
K. Schmitz and R. Kfihn: Symplast in the cortex of Laminaria 387
symmetry, the medulla often appears lense shaped (Fig. 1 b). The phaeoplast-containing cell layers (indi- cated by a white arrow) are separated from the medul- la by a wide parenchymatic cortex. A radial longitudi- nal section of a mature blade (Fig. 1 a) reveals a lateral symmetry and demonstrates that the small cells of the meristoderm and the layer of outermost cortex cells, which are characterized by a dense protoplast with numerous plastids, are separated from the sieve cell system in the medulla by only 6-8 layers of cortex parenchyma cell 400-600 gm thick. Embedded in the inner cortex is the slime duct system (c.f. Steinbil3 and Schmitz 1974). The medulla is composed mainly of long streched cells which were previously described as sieve elements and hyphal cells (Ziegler and Ruck !967; Schmitz and Srivastava 1974; Sideman and Scheirer 1977).
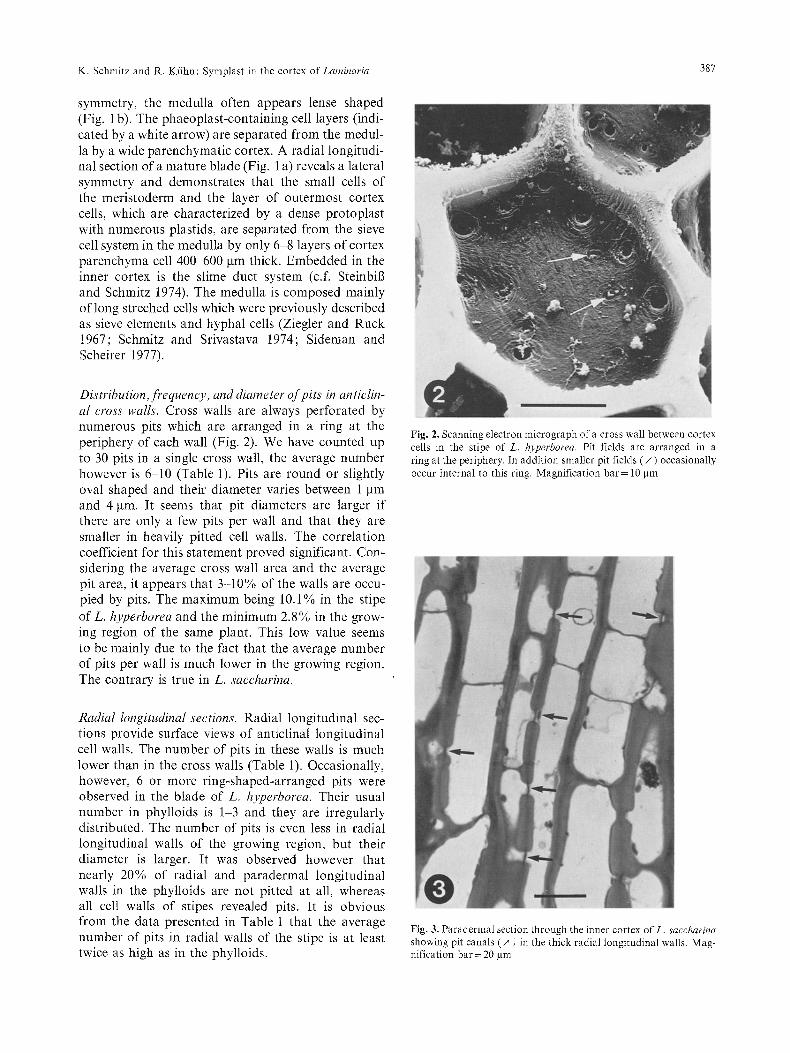
Distribution,frequency, and diameter of pits in anticlin- al cross walls. Cross walls are always perforated by numerous pits which are arranged in a ring at the periphery of each wall (Fig. 2). We have counted up to 30 pits in a single cross wall, the average number however is 6-10 (Table 1). Pits are round or slightly oval shaped and their diameter varies between 1 gm and 4 gin. It seems that pit diameters are larger if there are only a few pits per wall and that they are smaller in heavily pitted cell walls. The correlation coefficient for this statement proved significant. Con- sidering the average cross wall area and the average pit area, it appears that 3-10% of the walls are occu- pied by pits. The maximum being 10.1% in the stipe of L. hyperborea and the minimum 2.8% in the grow- ing region of the same plant. This low value seems to be mainly due to the fact that the average number of pits per wall is much lower in the growing region. The contrary is true in L. saccharina.
Radial longitudinal sections. Radial longitudinal sec- tions provide surface views of anticlinal longitudinal cell walls. The number of pits in these walls is much lower than in the cross walls (Table 1). Occasionally, however, 6 or more ring-shaped-arranged pits were observed in the blade of L. hyperborea. Their usual number in phylloids is 1-3 and they are irregularly distributed. The number of pits is even less in radial longitudinal walls of the growing region, but their diameter is larger. It was observed however that nearly 20% of radial and paradermal longitudinal walls in the phylloids are not pitted at all, whereas all cell walls of stipes revealed pits. It is obvious from the data presented in Table 1 that the average number of pits in radial walls of the stipe is at least twice as high as in the phylloids.
Fig. 2. Scanning electron micrograph of a cross wall between cortex cells in the stipe of L. hyperborea. Pit fields are arranged in a ring at the periphery. In addition smaller pit fields (/~ ) occasionally occur internal to this ring. Magnification bar = 10 Itm
Fig. 3. Paradermal section through the inner cortex o fL . saccharina showing pit canals ( ,~ ) in the thick radial longitudinal walls. Mag- nification bar = 20 l, tm

388 K. Schmitz and R. Ktihn : Symplast in the cortex of Laminaria
Table 1. Measurements and calculations of cell wall area, number of pit fields per wall and the diameter of pit fields in different plant organs and section planes of Laminaria hyperborea and L. saccharina based on light microscopy of cryosections. The mean values (X) and standard deviations (SD) relate to N=50 each. The average area of plasmodesmata per cell wall is based on an average frequency of 153 gm -2 of pit field area and an Inner diameter of the plasmalemma tubule of 20 nm
Orientation of cell walls Wall area and plant organs
Number of pits Diameter of pits Area of pits Area of plasmodesmata
2 gm 2 SD )7 SD )7 gm SD gm 2 % of gm 2 % of cell wall cell wall
Laminaria hyperborea
Anticlinal cross walls Mature lamina Growing region Stipe
Anticlinal longitudinal walls Mature lamina Growing region Stipe
Periclinal longitudinal walls Mature lamina 0-200 gm
200-400 gm Growing region 0-200 rtm
200-400 ~tm 400-600 gm
Stipe
Laminaria saccharina
Anticlinal cross walls Mature lamina Growing region Stipe
Anticlinal longitudinal walls Mature lamina Growing region Stipe
Periclinal longitudinal walls Mature lamina 0-200 lam
200 400 gm Growing region 0 200 gm
200-400 gm 400-600 gm
Stipe
871 364.0 9.8 5.3 2.6 0.4 52.0 6.0 2.5 0.29 677 231.2 2.9 2.4 2.9 0.7 19.2 2.8 0.9 0.13 615 248.0 7.7 3.5 3.2 0.6 61.9 10.1 3.0 0.49
992 431.7 3.0 4.3 2.6 0.5 15.9 1.6 0.8 0.08 1.132 366.2 2.2 0.9 3.2 0.6 17.7 1.6 0.9 0.08 1.428 287.6 5.9 2.6 3.1 0.6 44.5 3.1 2.1 0.15
300 152.4 1.9 0.7 2.9 0.6 12.5 4.2 0.6 0.20 1.897 472.4 1.5 0.6 3.3 0.6 12.8 0.7 0.6 0.03
335 145.7 2.2 1.2 2.7 0.5 12.6 3.8 0.6 0.18 1.099 389.5 1.6 0.8 3.3 0.8 13.7 1.2 0.7 0.06 1.910 442.3 1.6 0.9 3.5 0.7 15.4 0.8 0.7 0.04 1.262 381.7 10.6 3.8 3.0 0.6 74.9 5.9 3.6 0.29
1.354 655.3 15.6 7.5 1.7 0.3 89.3 6.6 4.3 0.32 1.293 555.7 7.1 4.6 2.7 0.7 40.7 3.1 2.0 0.15
612 187.9 5.5 2.7 3.1 0.5 41.5 6.8 2.0 0.33
1.675 605.0 1.6 0.8 2.4 0.7 7.2 0.4 0.3 0.02 946 368.6 1.3 0.6 3.6 0.9 13.2 1.4 0.6 0.06
1.126 487.4 5.4 2.5 2.9 0.5 35.7 3.2 1.7 0.15
705 301.6 1.3 0.6 2.9 0.8 8.6 1.2 0.4 0.06 1.303 534.8 1.4 0.6 3.4 0.5 12.7 1.0 0.6 0.05
545 235.9 2.0 0.7 3.8 0.6 22.7 4.2 1.1 0.20 884 435.5 1.6 0.6 3.6 0.7 16.3 1.8 0.8 0.09
2.787 731.3 1.7 0.8 3.9 2.3 20.3 0.7 1.0 0.04 562 248.7 10.1 2.8 2.4 0.3 45.7 8.1 2.2 0.39
Paradermal longitudinal walls. I f the p h o t o s y n t h a t e
f r o m the p i g m e n t e d p e r i p h e r a l cell layers m o v e s ra-
d ia l ly s t r a igh t t o w a r d the medu l l a , one s h o u l d expec t
t ha t the p a r a d e r m a l cell wal l s a re p r e f e r a b l y pi t ted .
T o t race the d i s t r i b u t i o n a n d f r e q u e n c y o f pi ts in
these walls , success ive p a r a d e r m a l sec t ions o f e q u a l
th ickness (40 ~tm) were cu t f r o m t issue samples o f
g r o w i n g r eg ions and m a t u r e blades. The d a t a pre- sen ted in T a b l e 1 are de r i ved f r o m a series o f sec t ions , i n v a r i a b l y c o v e r i n g 200 g m c o r t e x th ickness . T h e
n u m b e r o f pits in these cell wal l s was re la t ive ly l ow
and cons t an t . T h e ca l cu l a t ed ave rage was 1,3 to 2,2.
The re were no s ign i f i can t d i f fe rences in the n u m b e r
o f pits pe r wal l b e t w e e n the a n a l y z e d g r o w i n g r eg ions and the m a t u r e b lades o f b o t h species. T h e ave rage
d i a m e t e r o f the pits seems to inc rease f r o m the m e r -
i s t o d e r m to the i nne r cor tex . The s a m e is o b v i o u s
fo r the wal l area. T h e p e r c e n t a g e o f wa l l a r ea occu-
p ied by pits, h o w e v e r , s ign i f ican t ly decreases f r o m
the m e r i s t o d e r m to the m e d u l l a . In t a n g e n t i a l cell wal l s o f st ipes, 10 o r m o r e i r r egu la r ly d i s t r i b u t e d pits pe r
wa l l were c o u n t e d wh ich o c c u p y 6 to 8 % o f the co r re -
s p o n d i n g wal l a r ea (Tab le 1).
Fine structure o f pits and plasmodesmata. Pits a p p e a r u n d e r the l ight m i c r o s c o p e a n d the s c a n n i n g e l e c t r o n
m i c r o s c o p e as r o u n d o r s l ight ly o v a l - s h a p e d wal l per-
f o r a t i o n s (Fig. 2). T h e pit m e m b r a n e is a lways t h i n n e r
t h a n the s u r r o u n d i n g cell wa l l and this f ea tu re m a y
be such tha t real pi t cana l s a re f o r m e d (Fig. 3). T r a n s m i s s i o n e l ec t ron m i c r o g r a p h s revea l t ha t a pit
is a loca l a c c u m u l a t i o n o f p l a s m o d e s m a t a p e r f o r a t i n g

K. Schmitz and R. Kfihn : Symplast in the cortex of Laminaria 389
Fig. 4. Tangential section of a pit field between two cortex cells of L. saccharina. Plasmodesmal canals are lined by open p lasmalemma tubules. The tripartite b iomembrane of the p lasmalemma is clearly shown in the inset. Note the light outer rim between the p lasmalemma tubule and the cell wall. Magnification ba r=0 .2 ~trn, inset 0.1 ~tm
the pit membrane. Their distribution within the pit field - which may be the apropriate term here - is even, but not regular (Fig. 4). Transverse sections re- veal that each of the numerous plasmodesm canals is lined by a tripartite biomembrane (Fig. 4). The plasmalemma tubules appear round in transverse sec- tions, the membrane thickness is approximately 8 nm, and the outer diameter of the tubules varies between 30 and 40 nm. The lumen of plasmalemma tubules is electron light and approximately 20 nm wide, a desmotubule was never observed. There is usually a narrow space between the plasmalemma membrane and the inner rim of the wall perforation. This seems to be an artifact due to the fixation and preparation procedure. The diameter of the wall perforations (plasmodesmal canals) was determined from 50 trans-
verse and 50 longitudinal sections, their width ranged between 30 and 60 nm. In slightly oblique transverse sections, plasmodesmata appeared more or less oval shaped; here, only the smallest diameter of the wall perforation was measured. The average diameter cal- culated from cross sections is 45 nm but 40 nm was the average determined from longitudinal sections. This difference is due to the fact that longitudinal sections only partially reveale the median section plane of plasmodesmata, most were cut tangentially.
Considering a section thickness of 50 nm or more, it is obvious that a clear electron micrograph of these structures in longitudinal sections can hardly be achieved. Nevertheless, Fig. 5 tentatively shows the continuity of plasmalemma through the plasmodesm canals (cf. arrows), thus providing a cytoplasmic con-

390 K. Schmitz and R. Kfihn: Symplast in the cortex of Laminar&
Fig. 5. Micrograph of plasmodesmata cut longitudinally. The plasmalemma is continuous from cell to cell establishing a cytoplasmic continuity between cortex cells. Numerous vesicles (*) are always closely associated with pit fields. Magnification bar= 0.2 gm
tinuity from cell to cell. Very frequently small vesicles and tubular endoplasmic reticulum are closely asso- ciated with plasmodesmata, a continuity of tubular endoplasmic reticulum through plasmodesmata was not observed.
It may be seen from Fig. 4 that the frequency of plasmodesmata in a pit field is very high. From numerous sections of all organs and the section planes of both species investigated, an average density of 153 plasmodesmata per gm 2 was calculated. The vari- ability was high, the absolute measurements ranged from 100 to 200 lain -2. Taken an average diameter of 45 nm for the plasmodesmal canal and a frequency of 153 ~tm -2, the area occupied by plasmodesmata amounts to 24.3% of the pit field area, but it is reduced to 4.8% when only the lumen of the plasma- lemma tubules is considered.
Discussion
Photosynthate from the meristoderm and the outer- most cortex cells moves through the cortex toward the medulla where it is loaded into the sieve element system for further long-distance transport (Hellebust and Haug 1972; Schmitz et al. 1972; Steinbil3 and Schmitz 1973). Both the symplast and the apoplast could act as a pathway for this parenchyma transport, though it is most probable that aquatic plants use the symplast rather than the apoplast. Cell mem- branes of brown seaweeds seem rather impermeable for organic carbon compounds, the uptake of organic carbon from aqueous media is very limited and does not play any significant role in the carbon balance of these plants (Bidwell and Ghosh 1963; Drew 1969; Kiippers 1978) and the release of photoassimilate under normal conditions seems very minor (Moebus and Johnson 1974; Harlin and Craigie 1975; Bry-
linsky 1977; Munda and Kremer 1977), even though contradictory opinions have been expressed (Khailov and Burlakova 1969; Sieburth 1969).
If the apoplast were involved in parenchyma trans- port, the plants would be covered or sealed off by a more or less impermeable barrier to carbon organic compounds, or the apoplast would be compartmenta- lized in order to avoid uncontrolled leaching and a loss of nutrients to the surrounding medium. Such barriers have not been demonstrated in Laminariales.
Still another observation pointing to symplastic transport is the lack of transport selectivity (Parker 1966; Schmitz etal . 1972; Schmitz and Srivastava 1975, 1979). We therefore suggest that the parenchy- ma transport in Laminaria is a symplastic transport.
The present study presents evidence for a structur- al basis of symplastic transport across the cortex; it demonstrates that all cells of the meristoderm and cortex of Laminaria hyperborea and L. saccharina are interconnected by the plasmodesmatal establishment of a symplast. It was recognized much earlier that plasmodesmata in Laminariales are mainly confined to pit fields which may be arranged in a ring at the periphery of anticlinal cross walls (Smith 1939; Parker and Fu 1965; Bisalputra 1966). The number and width of pit fields is greatly variable. Pits as small as 1 gm and as wide as 6 ~tm were measured, the pit frequency varied between 0.06 and 1.8. 100 gm -2 of the cell wall area. It is apparent that the conductivi- ty in a longitudinal direction is higher than in a radial or tangential direction. This was stated before for stipes of several other Laminariales also (Smith 1939; Parker and Fu 1965).
The data presented in Table 1 also show that the pit frequency and therefore the conductive area across the paradermal longitudinal cell walls is minor and does not match preconceived expectations, a re- sult which contradicts statements of Wille (1885), who

K. Schmitz and R. Kiihn : Symplast in the cortex of Laminaria 391
claimed that the paradermal longitudinal walls in the stipes of Laminaria were the most densely pitted walls. The measurements presented here reveale that the wall area occupied by pits is 4.2% in the outermost 200 gm of the meristoderm-cortex region of L. hyper- borea and only 0.7% in the innermost 200 gm of the cortex. The figures for L. saccharina are 1.2% and 1.0%, respectively. The higher values for the mer- istoderm-cortex region are probably due to the fact that the inner tangential walls of the meristoderm are relatively small but densely pitted (Bisalputra 1966).
The fine structure of single plasmodesmata, as described here for Laminaria, is in accordance with observations reported earlier for Egregia menziesii and Fucus evanescens (Bisalputra 1966). The average frequency of plasmodesmata in pit fields of both spe- cies of Laminaria is rather high, but corresponds well with those calculated from published micrographs of other brown algae. For Himanthalia lorea, 168 gm-2 were calculated (Plate VII, Fig. b in Berkaloff 1963), for Egregia menziesii 132 and 172 gm -2 were deter- mined (Figs. 2 and 4 in Bisalputra 1966). Comparable frequencies of plasmodesmata of up to 140 gm- 2 are reported for young walls of Dryopteris filix-mas though in older cell walls of the same plant the fre- quency was only 10-20 gm -2 (Burgess 1971).
The area of a cell wall occupied by plasmodesmata varies between 0.3 and 4.3 gm 2, i.e., 0.02-0.5% of the corresponding wall area (Table 1). These data re- late to the lumina of the plasmalemma tubules which seem to represent the conducting area rather than the whole sectional area of the plasmodesmal canals. The data presented here are in the range of those measured on higher plant cell walls (c.f. Robards 1976).
Photosynthetic CO2-fixation rates of L. hyperbo- rea and L. saccharina were determined by different methods and range from 15 to 133 gmol CO 2 dm 2 h - t (Ltining 1971 ; Willenbrink et al. 1975; Johnston et al. 1977). An average rate of 65 gmol CO 2 dm -2 h ~ will yield the same rate of [ C H 2 0 ] . All these measurements relate to the simple thallus area al- though the dense layer of phaeoplasts on both sides of a lamina (meristoderm and outermost cortex cells) equally contribute to photosynthesis. Considering the parenchyma transport in question, only half the amount would have to move from the pigmented layer of one side toward the medulla. Assuming that all photosynthate is translocated and that the amount of fixed carbon produced by 1 cm 2 of thallus surface will move radially through the same area of plasma- lemma, a flux density of 32.5 gmol [CHzO] cm -2 h -~ would result which equals 0.9.10 6tool m -2 s- 1. This is much higher than the upper flux capacity
of higher plant cell membranes of 10-s-10 -7 mol m -2 s -1, as determined by Mac Robbie (1971), and suggests a transport through a much larger membrane area or a transport through plasmodesmata. If we assume an average pit density of 1% in the cortex of both plants, and this may even be less because the calculated wall area is certainly underestimated (cf. materials and methods) and because up to 20% of the walls is not pitted at all, and, further, if the pit density is 153 gm -2 of a pit field area and the fixed carbon actually moves through these plasmodes- mata, the flux density would at least be 0.6.10 is tool plasmodesma-1 s-1. Gunning and Robards (1976) suggested that the limiting flux rate for plasmo- desmata might be in the range of between 2.10-19 10-~8 tool plasmodesma-1 s 1. Hence, the flux rate estimated here seems close to saturation, and this might also force the substances to move in a longitudi- nal direction through the plasmodesmata in the anti- clinal cross walls. It is obvious from Table 1 that these walls are preferably pitted, providing the largest conductive area and probably a much lower resistance for a symplastic transport of organic compounds. In conclusion, a deviation from a straight radial trans- port should result and the translocate should move oblique toward the sieve elements not-merging oppo- site to the site of synthesis, but diagonally sloping toward the sink of long distance transport.
Pit connections in the longitudinal cell walls of sieve elements connecting them with neighboring cor- tex parenchyma cells have not yet been observed. This is in accordance with observations reported ear- lier by Nicholson (1976) for the stipes of Nereocystis. A symplast connecting the phaeoplast-rich meristo- derm and the cortex parenchyma cells with the sieve elements may exist after all, though the connections may not be straight radially. This however remains to be proven and to be adequately documented.
This work was supported by Deutsche Forschungsgemeinschaft. We thank Mrs. Poelmann for technical assistance.
References
Berkaloff, C. (1963) Les cellules m6rist6matiques d'Himanthalia lorea (L.) S.F. Gray. Etude au microscope 61ectronique. J. Microsc. (Paris) 2, 213 228
BidwelI, R.G., Ghosh, N.R. (1963) Photosynthesis and metabolism of marine algae V. Respiration and metabolism of C~4-1abelled glucose and organic acids supplied to Fucus vesiculosus. Can. J. Bot. 41, 155 163
Bisalputra, T. (1966) Electron microscopic study of the protoplas- mic continuity in certain brown algae. Can. J. Bot. 44, 89-93
Brylinsky, M. (1977) Release of dissolved organic matter by some marine macrophytes. Mar. Biol. 39, 213-220
Burgess, J. (1971) Observations on structure and differentiation in plasmodesmata. Protoplasma 73, 8345

392 K. Schmitz and R. Kfihn: Symplast in the cortex of Laminar&
Drew, E.A. (1969) Uptake and metabolism of exogenously supplied sugars by brown algae. New Phytol. 68, 35-43
Floc'h, J.Y., Penot, M. (1972) Transport du 32p et du 86Rb chez quelques algues brunes: orientation des migrations et voles de conduction. Physiol. V6g. 10, 677 686
Gunning, B.E.S., Robards, A.W. (1976) Plasmodesmata: Current knowledge and outstanding problems. In : Intercellular commu- nication in plants: Studies on plasmodesmata, pp. 297 311, Gunning, B.E.S., Robards, A.W., eds. Springer, Berlin Heidel- berg New York
Harlin, M.M., Craigie, J.S. (1975) The distribution of photosyn- thate in Ascophyllum nodosum as it relates to epiphytic Polysi- phonia lanosa. J. Phycol. 11, 109-113
Hellebust, J.A., Haug, A. (1972) Photosynthesis, translocation, and alginic acid synthesis in Laminaria digitata and Laminaria hyper- borea. Can. J. Bot. 50, 169-176
Ito, S., Winchester, R.S. (1963) The fine structure of the gastric mucosa in the rat. J. Cell Biol. 16, 541-577
Johnston, C.S., Jones, R.G., Hunt, R.T. (1977) A seasonal carbon budget for a laminarian population in Scottish sealoch. Helgo- lfinder Wiss. Meeresuuters. 30, 527-545
Khailov, K.M., Burlakova, Z.P. (1969) Release of dissolved organic matter by marine seaweeds and distribution of their total organ- ic production to inshore communities. Limnol. Oceanogr. 14, 521-527
Ktippers, U. (1978) Enzymologie der CO2-Fixierung bei Laminaria hyperborea: Jahresperiodische, umweltbezogene Ver/inderungen von Enzymaktivitgten. Thesis, K61n
Liining, K. (1971) Seasonal growth of Laminaria hyperborea under recorded underwater light conditions near Helgoland. In: Proc. IVth European Marine Biology Symposium, pp. 347 361, Crisp, D.J., ed. Cambridge University Press, Cambridge
Lfining, K., Schmitz, K., Willenbrink, J. (1972) Translocation of 1*C-labeled assimilates in two Laminaria species. Proc. VIIth Int. Seaweed Syrup., pp. 420-425
Lfining, K., Schmitz, K., Willenbrink, J. (1973) CO2 Fixation and translocation in benthic marine algae. III. Rates and ecological significance of translocation in Laminaria hyperborea and L. saccharina. Mar. Biol. 23, 275~81
MacRobbie, E.A.C. (1971) Phloem translocation. Facts and mech- anisms: a comparative survey. Biol. Rev. 46, 429-481
Moebus, K., Johnson, K.M. (1974) Exudation of dissolved organic carbon by brown algae. Mar. Biol. 26, 117-125
Munda, J.M., Kremer, B.P. (1977) Chemical composition and physiological properties of fucoids under conditions of reduced salinity. Mar. Biol. 42, 9-16
Nicholson, N.L. (1976) Anatomy of the medulla of Nereocystis. Bot. Marina 19, 23-31
Nicholson, N.L., Briggs, W.R. (1972) Translocation of photosyn- thate in the brown alga Nereocystis. Am. J. Bot. 59, 97 106
Parker, B.C. (1965) Translocation in the giant kelp Macrocystis.
I. Rates, direction, quantity of Cl~-labeled products and fluo- rescein. J. Phycol. 1, 41-46
Parker, B.C. (1966) Translocation in Macrocystis. III. Composition of sieve tube exudate and identification of the major C14-1abeled products. J. Phycol. 2, 38 41
Parker, B.C., Fu, M. (1965) The internal structure of the elk kelp (Pelagophycus species). Can. J. Bot. 43, 1293-1305
Robards, A.W. (1976) Plasmodesmata in higher plants. In: Inter- cellular communication in plants: Studies on plasmodesmata, pp, 15 57, Gunning, B.E.S., Robards, A.W., eds. Springer, Berlin Heidelberg New York
Schmitz, K., Lobban, C.S. (1976) A survey of translocation in Laminariales (Phaeophyceae). Mar. Biol. 36, 207-216
Schmitz, K., Lfining, K., Willenbrink, J. (1972) CO2-Fixierung und Stofftransport in benthischen marinen Algen. II. Zum Fern- transport 14C-markierter Assimilate bei Laminaria hyperborea und Laminaria saccharina. Z. Pflanzenphysiol. 67, 418-429
Schmitz, K., Srivastava, L.M. (1974) Fine structure and develop- ment of sieve tubes in Laminaria groenlandica Rosenv. Cytobiol. 10, 66-87
Schmitz, K., Srivastava, L.M. (1975) On the fine structure of sieve tubes and the physiology of assimilate transport in Alaria mar- ginata. Can. J. Bot. 53, 861-876
Schmitz, K., Srivastava, L.M. (1979) Long distance transport in Macrocystis integrifolia. I. Translocation of 14C-labeled assimi- lates. Plant Physiol. 63, 995-1002
Sideman, E.J., Scheirer, D.C. (1977) Some fine structural observa- tions on developing and mature sieve elements in the brown alga Laminaria saccharina. Am. J. Bot. 64, 649~557
Sieburth, J.McN. (1969) Studies on algal substances in the sea. III. The production of extracellular organic matter by littoral marine algae. J. Exp. Mar. Ecol. 3, 290-309
Smith, A.J. (1939) The comparative histology of some of the La- minariales. Am.J. Bot. 26, 571 585
Steinbil3, H.H., Schmitz, K. (1973) COz-Fixierung und Stofftrans- port in benthischen marinen Algen. V. Zur autoradiogra- phischen Lokalisation der Assimilattransportbahnen im Thallus von Laminaria hyperborea. Planta 112, 253-263
Steinbil3, H.H., Schmitz, K. (1974) Zur Entwicklung und funktio- nellen Anatomie des Phylloids von Laminaria hyperborea. Hel- gol/inder Wiss. Meeresunters. 26, 134 152
Wille, N. (1885) Siebhypen bei den Algen. Ber. Dtsch. Bot. Ges. 3, 29-31
Willenbrink, J., Rangoni-Kfibbeler, M., Tersky, B. (1975) Frond development and CO2-fixation in Laminaria hyperborea. Planta 125, 161-170
Ziegler, H., Ruck, I. (1967) Untersuchungen fiber die Feinstruktur des Phloems. III. Mitteilung: Die "Trompetenzellen" von La- minaria-Arten. Planta, 73, 62-73
Received 7 November 1980; accepted 2 February 198l







![Selective Targeting of Mobile mRNAs to Plasmodesmata for ... · Selective Targeting of Mobile mRNAs to Plasmodesmata for Cell-to-Cell Movement1[OPEN] Kai-Ren Luo, Nien-Chen Huang,](https://static.fdocuments.in/doc/165x107/5f13a2fca34f6100383e7928/selective-targeting-of-mobile-mrnas-to-plasmodesmata-for-selective-targeting.jpg)










