
FINAL NARRATIVE REPORT GRANT # 05-3044-SCR … NARRATIVE REPORT GRANT # 05-3044-SCR-E-O I. APPLICANT...
14
FINAL NARRATIVE REPORT GRANT # 05-3044-SCR-E-O I. APPLICANT INFORMATION Applicant Name Ron Weiss Check 0' Applicant Career Level: D Postdoctoral Fellow X Young Investigator (five years post completion of formal training) D Established Investigator a D in another field b D in spinal cord injury research Applicant Organization Name Princeton University Check 0' Appropriate Organization Type D Public Institution of Higher Learning X Private Institution of Higher Learning D Non-Profit D For Profit II. PROPOSAL INFORMATION Check 0' Proposal Type X Basic Science D Clinical Science D Alternative Medicine D Psycho/Social Rehabilitation Proposal Title Programmed Tissue Engineering for Spinal Cord Regeneration Hypothesis Precise control over stem cell differentiation for tissue engineering can be achieved by careful orchestration of master switch gene expression using synthetic gene networks KeyWords Tissue engineering, Synthetic gene networks, master switches II. PROPOSAL INFORMATION, Continued Dollar Amount Requested Year 2: $192,643 Check 0' Institutional Review Board/Institutional Animal Care and Use Committee Approval: DYES - Approval Obtained, Copy Attached D NO - Approval Not Obtained Date Anticipated for Review: (Copy must be forwarded to NJCSCR as soon as approval is obtained.) D PENDING - Currently Under Review Date Anticipated: (Copy must be forwarded to NJCSCR as soon as approval is obtained.) X EXEMPT All applicants must conform with the State of New Jersey Department of Health and Senior Services Policy for Investigating and Reporting Instances of Alleged or Apparent Misconduct Involving Scientific Research. This policy can be viewed at: www.state.nj.us/health/spinalcord/. Proposal Number: Date of Submission: _
Transcript of FINAL NARRATIVE REPORT GRANT # 05-3044-SCR … NARRATIVE REPORT GRANT # 05-3044-SCR-E-O I. APPLICANT...

FINAL NARRATIVE REPORT
GRANT # 05-3044-SCR-E-O
I. APPLICANT INFORMATION
Applicant Name
Ron Weiss
Check 0' Applicant Career Level:D Postdoctoral FellowX Young Investigator (five years post completion of
formal training)D Established Investigator
a D in another fieldb D in spinal cord injury research
Applicant Organization Name
Princeton University
Check 0' Appropriate Organization TypeD Public Institution of Higher LearningX Private Institution of Higher LearningD Non-ProfitD For Profit
II. PROPOSAL INFORMATION
Check 0' Proposal TypeX Basic ScienceD Clinical ScienceD Alternative MedicineD Psycho/Social Rehabilitation
Proposal Title
Programmed Tissue Engineering for SpinalCord Regeneration
Hypothesis
Precise control over stem cell differentiationfor tissue engineering can be achieved bycareful orchestration of master switch geneexpression using synthetic gene networks
KeyWords
Tissue engineering, Synthetic genenetworks, master switches
II. PROPOSAL INFORMATION, Continued
Dollar Amount Requested
Year 2: $192,643
Check 0' Institutional Review Board/Institutional Animal Careand Use Committee Approval:
DYES - Approval Obtained, Copy Attached
D NO - Approval Not Obtained
Date Anticipated for Review:
(Copy must be forwarded to NJCSCR as soon asapproval is obtained.)
D PENDING - Currently Under Review
Date Anticipated:
(Copy must be forwarded to NJCSCR as soon asapproval is obtained.)
X EXEMPT
All applicants must conform with the State of NewJersey Department of Health and Senior ServicesPolicy for Investigating and Reporting Instancesof Alleged or Apparent Misconduct InvolvingScientific Research. This policy can be viewed at:www.state.nj.us/health/spinalcord/.
Proposal Number:
Date of Submission: _

· FOR STATE USE ONLY
New Jersey Commission on Spinal Cord Research NJCSCR Number
FINAL NARRATIVE REPORT Spending Plan Number
(Type or print all data.) Funding Authorization Number(s)
1. Name of Principal Investigator 1a. Email AddressRon Weiss [email protected]
2. Name of OrganizationPrinceton University
3. Street Address City County State Zip CodeB312 E·Quad Olden Street Princeton Mercer NJ 08540
4. Name and Title of Fiscal Contact 5. Telephone No.John Floersch, Manager, Sponsored Research Accounting 609·258·3070
6. Street Address City County State Zip Code5 New South Building Princeton Mercer NJ 08540
7. Name of Attorney for Agency 8. Telephone No.
9. Name and Title of Principal Contact 10. Telephone No.Michelle D. Christy, Director, Office of Research and Project Administration 609·258·3090
11. Employer ID No.21·0634501
12. a. Will any member of the Board of DirectorsfTrustees receive any direct or indirect personalor monetary gain from the funding of this grant? DYes XNo
b. Does any member of the Board of DirectorsfTrustees serve on any board, council commission,committee or Task Force which has regulatory or advising influence on the funding program? DYes XNo
(Member) (Board Council Etc.13. Type of Payment Plan Preferred 14. Location Where Payments Should be Sent:
DCost Reimbursement X Advance Payment Office of Research and Project Administration PO Box36, 4New South Buildino. Princeton NJ 08540
15. Type of Agency (check one) 16. Does this Agency meet the following Licensure Requirement?X Private Non-Profit DGovernment DHospital Yes No Pending N/ADPrivate Profit DOther: For Facility X 0 0 0
17. Agency Fiscal 18. Agency Accounting System: For Services X 0 0 0YearEnd DCash Basis DAccrual BasisJune 30 DOther (Specify): For Personnel X 0 0 0
19. Type of Request 20. a. Budget Period (Mo/Day/Yr)X Continuation of Grant No.: 05·3044·SCR·E·O From: 12/15/2005 Through: 12/14/2006
21. If grant is awarded, will funds be used to replace other funds b. Project Period (Mo/Day/Yr)which would be available in absence of award? From: 12/15/2004 Through: 12/14/2006DYes DNo
COST OF PROJECT22a. Total Funds Needed
lb.Funds Requested from NJCSCR c. Funds from Other Sources
392,586 192,643
Principal Investigator/Program Director Assurance: I agree to accept responsibility for the scientific conduct of the project and to providethe required progress reports if a grant is awarded as a result of this application.
23. Signature of Principal Investigator/Program Director (In Ink, "Per" signature not acceptable) I Date
Certification: The applicant certifies that to the best of his/her knowledge and belief, all data supplied in this application and attachmentsare true and correct, the document has been duly authorized by the governing body of the applicant, and further understands and agreesthat any grant received as a result of this application shall be subject to the grant conditions and other policies, regulations and rulesissued by the N. J. Department of Health and Senior Services which include provisions described in grant application instructions.
24. Name and Title of Official Signing for Applicant OrganizationMichelle D. Christy, Director, Office of Research and Project Administration
25. Signature of Official Date

Lay Abstract: Please describe your research project in simple, non-technical language that isunderstandable by a person not trained in science. Include in your discussion: 1) the significance ofyour project to developing effective interventions and cures for paralysis and other consequences ofspinal cord injury and disease as noted in the program guidelines; and 2) any special value it mighthave for the citizens of New Jersey. This abstract is meant to serve as a public description of theproposed research and, should the award be made, it will be used in press releases and variousNJCSCR publications. DO NOT EXCEED THE SPACE PROVIDED.
Abstract of Research Plan: State the application's long-term objectives and specific aims, makingreference to the spinal cord relatedness of the project, and describe concisely the methods for achievingthese goals. Avoid summaries of past accomplishments and the use of the first person. The abstract ismeant to serve as a succinct and accurate description of the proposed work when separated from theapplication. DO NOT EXCEED THE SPACE PROVIDED.
Proposal Narrative: Proposed narrative limited to four pages maximum, not including appendices.DO NOT EXCEED FOUR (4) PAGES.
Narrative information should address the following:• Specific aims, summarized from the original proposal·• Overall summary of progress made during the contract period·• Problems encountered in this report period·• Changes in research plan·• List of publications emerging from this research, including those in preparation

Lay Abstract (Do Not Exceed The Space Provided)Project Title (do not exceed 60 spaces)
Programmed Tissue Engineering for Spinal Cord Regeneration
Please provide a one sentence description of your project appropriate for a lay personThe project aims to program embryonic stem cells (ES cells) to achieve controlled differentiation towards spinal cord regeneration.
Description (Do not exceed space provided. Type in single spaced format.)
Spinal cord injury is a major cause of paralytic disabilities. Currently, there are no effective therapies that result insignificant alleviation of such disabilities. The therapies that are in use are generally limited to preventing more extensiveaccumulation of tissue damage. No effective therapies can reverse spinal damage through the regeneration, or re-growth of new cells. Several experimental studies suggest that transplantation of undamaged cells into the site of injurymay eventually be an effective avenue for therapeutic intervention. While encouraging, such therapies are inherentlylimited by a lack of suitable transplantable material that will fully regenerate the spinal cord without complications suchas tissue rejection. Recently, it has been suggested that stem cell populations, whether from embryonic or adult tissuesources, may represent a source of transplantable material for therapeutic tissue repair. Stem cells are cells that cangenerate many types of mature cell populations for lifelong time periods. The major barrier to realizing the potential ofstem cells is to devise ways to direct these cells toward different pathways of mature tissue or cell production. That is,for spinal cord repair it is necessary to ensure the production of the nerve and associated cells that are normally found inthis tissue. It will also be necessary to ensure that the mature cells are produced in the proper amounts and in anappropriate three-dimensional tissue structure. The mechanisms that control the production of mature cells from stemcells in normal fetal or adult physiological contexts are poorly understood. Nevertheless, a number of molecules thatfunction as genetic "master switches" in stem cells to direct the production of different cell types have been identified.We propose to harness these "master switches" by implementing them in artificially controlled regulatory circuitsconstructed inside stem cells by genetic engineering. In this way it will be possible to trigger the controlled production ofdifferent mature cell populations after transplantation of the genetically modified stem cells into the site of tissue injury.In addition, it will eventually be possible to produce these cells in a correct spacial and temporal tissue organization.Precedents for the successful construction of artificial genetic circuits have been reported. In general, these have beenlimited to simpler cells such as bacteria or yeast; however, we have ample preliminary results that demonstrate thesuccessful construction of such circuits in mammalian cells. Thus, there is no insurmountable barrier to extending ourresults to stem cells. One of the laboratories (Weiss) in this proposal has pioneered the design and implementation ofartificially controlled genetic circuits. The second laboratory (Lemischka) has been a leader in stem cell research formany years. Our joint expertise should facilitate the rapid progress of the studies in this proposal. We will begin bydeveloping a number of "proof-of-principal" genetic circuits in mouse embryonic stem (ES) cells. We will take advantageof the known "master switch" molecules, and ask if we can build circuits where these "master switches" can be artificiallycontrolled by small pharmaceutical agents used to treat the stem cells that have been genetically modified to contain theartificial circuits. We will address how effectively the stem cells can be triggered to produce alternate types of maturecells in response to controlled stimuli. These relatively simple circuits will also produce a wealth of necessary informationregarding the dynamics and overall behavioral properties of artificial circuits in stem cells. Finally, we will take advantageof new results from the Lemischka laboratory that have begun to identify the naturally occurring regulatory components,pathways, and networks that control biological functions of stem cells. We anticipate that the successful completion ofthe studies in this proposal will lay a broad and rigorous framework that will have broad applications in stem cellmediated repair of damaged tissues including the spinal cord.

Technical Abstract (Do not exceed space provided. Type in single spaced format.)
Precise control of stem cell differentiation offers tremendous potential for spinal cord regeneration. The objective ofthis proposal is to engineer synthetic gene networks that would ultimately control temporal and spatial differentiationof stem cells into structures that resemble the spinal cord. Such programmed control will enable treatment of spinalcord injuries that are difficult to repair given the means that are available today in clinical settings. These genenetworks will turn on "master switch" genes in stem cells at precise times and under well-controlled and well-definedconditions based on external stimuli and the internal state of the cell. In the future, genetically engineered stem cellswill be implanted at the site of spinal cord injury and programmed to differentiate appropriately in order to restorepartial or full functionality. For this proposal, two new gene networks for programmed differentiation will beimplemented, a 1:2 multiplexer and a toggle switch. The 1:2 multiplexer is a network with one input (e.g. an inducermolecule) that selects to activate one of two potential gene outputs (master switches). This will form the basis formore sophisticated multiplexer networks that incorporate more input signals and output master switches that cantrigger the differentiation of a homogeneous population of stem cells into many cell types based on the syntheticinputs provided. Importantly, such networks will achieve desired differentiation in vivo despite hostile environments.The toggle switch network is a bistable system with positive feedback that can be set to one of its states withtransient administration of one of its two exogenous inputs. Such a network is advantageous in clinical settingsbecause implanted stem cells will be given a transient cue to force them into a particular differentiation pathwaythereby obviating the need to introduce and sustain the appropriate growth factors in-vivo. The proposed methodincludes initial validation and characterization of master switches relevant to spinal cord regeneration, followed bydesign and implementation of stem cell synthetic gene networks to control and analyze differentiation, commitment,and self-renewal pathways. In the future, these networks will be integrated with cell-cell signaling to engineer artificialdifferentiation patterns on demand. While the proposed work is application oriented, it will also provide an importantplatform for discovering the fundamental mechanisms of stem cell differentiation.

Final Narrative Report (Do Not Exceed 5 Pages)A. Specific Aims summarized from the original proposal
Precise control of stem cell differentiation offers tremendous potential for spinal cord regeneration. Here we proposeto engineer synthetic gene networks that would ultimately control temporal and spatial differentiation of stem cells intostructures that resemble the spinal cord. Such programmed control will enable treatment of spinal cord injuries thatare difficult to repair given the means that are available today in clinical settings. In general, current approachestowards tissue engineering rely on carefully creating environments that induce cells to differentiate into desiredtissues or organs. While these approaches have proven partially effective for certain applications, they are inherentlylimited since they rely on innate cellular response to existing host conditions or exogenous cues. Often naturallyoccurring host conditions are insufficient to trigger the correct differentiation pathways for certain wound healingapplications. In those instances, researchers have attempted to provide the appropriate environmental cues usingscaffolds and exogenous signals. However, it is often difficult, if not impossible, to create and maintain the preciseconditions that are required for tissue regeneration using such means.
We propose a paradigm shift in tissue engineering and spinal cord regeneration: synthetic gene networks will be usedto determine cell fate by precisely controlling gene expression. Synthetic gene networks are at the forefront ofsystems biology and provide a framework for understanding and engineering life. Our proposed networks will becapable of turning on "master switch" genes in stem cells at precise times and under well-controlled and well-definedconditions based on external stimuli and the internal state of the cell. The over-expression of these master regulatorsgenes is sufficient to trigger particular differentiation pathways in embryonic stem cells as well as many adult celltypes. We envision spinal cord treatments where genetically engineered stem cells will be implanted at the site ofinjury and programmed to differentiate appropriately in order to restore partial or full functionality.
Our overall objective is to achieve precise control over stem cell differentiation for spinal cord regeneration viasynthetic genetic networks. The proposed research on this programmed tissue engineering encompasses four keycomponents:
Spinal cord development is a complex process requiring the precise temporal and spatial orchestration of manyfactors. The redundant nature of many systems in vertebrates complicates the identification of master switcheswhich, when over expressed, can force naIve cells to differentiate into various cell types. Nonetheless, a few masterswitches have been identified. For example, over expression of the transcription factor NeuroD3 will cause stem cellsto adopt a neural fate [Ma, Q., et aI., 1996] while over expression of MyoD should cause the same cells to becomeskeletal muscle tissue [Davis, R.L., et aI., 1987]. Other master switches relevant to spinal cord regeneration includeSox9 which regulates differentiation of chondrocytes [Nakashima, K., et aI., 2001], and Osterix which leads to theformation of osteocytes [Nakashima, K., et aI., 2002]. It is still not known which adult stem cell lines can be induced todifferentiate along these particular pathways. We will investigate this issue by regulating expression of these knownmaster switches in a variety of cell lines using our synthetic gene networks. Using these inducible constructs, we willquantify the expression levels and durations required for each master switch to induce differentiation. We will alsoexplore the self-renewal and commitment of stem cells by controlling expression of Nanog and Oct4 [Chambers, I., etal.,2003].
As we assemble our library of master switches, we will design and construct multiplexer and toggle switch genenetworks in mouse ES cells. These networks will express a specific gene out of a possible set of genes based on thepresence or absence of extracellular ligands. As reported in Section 3, we recently constructed synthetic genenetworks in adult mouse muscle stem cells that implement the initial versions of a 1:2 multiplexer and a toggle switch.With the multiplexer network, we are able to control whether cells express EGFP or DsRedExpress based on theaddition of a single input (Doxycycline). With the toggle switch, we can control the sustained expression of EGFP orDsRedExpress using transient administration of Isopropyl-beta-D-thiogalactoside (IPTG) or anhydrotetracycline (aTe)inducer molecules. We will fine-tune and optimize the kinetic rates of the genetic elements comprising these circuitsto achieve desired dynamic behavior (e.g. response delays, expression levels, dosage responses). We will integratecircuit design based on computer models with the experimental implementation. Specifically, we will use the dataobtained from the experiments to build quantitative models of the circuits, and also use the models to forward-engineer circuit behavior.

The multiplexer and toggle switch networks afford a unique ability to control and analyze stem cell differentiation. Wewill first integrate master switches relevant to spinal cord regeneration (NeuroD3, MyoD, Sox9, Osterix, Nanog, Oct4)into these networks. We will induce differentiation of homogeneous populations of stem cells (ES and adult) into twodistinct cell types as dictated by our choice of external Iigands. We will analyze the commitment and self-renewalcapabilities of the differentiated cells by inducing and alternating the expression of the master switches. Themultiplexer and toggle switch networks allow us to perform a variety of experiments with various temporal sequencingof master switches. For example, NeuroD3 can be expressed initially to specify neuronal precursor cell fate, followedby expression of Pax? to further differentiate these precursor cells into dorsal spinal cord neurons. In addition, we willalso explore the effects of expressing master switches that promote differentiation along unrelated pathways, byexpressing these genes both simultaneously and sequentially. These experiments can provide some initial insightinto the relationships between the different pathways.
As discussed above, a number of transcriptional regulators have been implicated in ES self-renewal anddifferentiation into defined cell types. However, very little is known about the targets of these regulators, and aboutthe architecture and dynamics of the regulatory pathways and networks. An integral requirement to an eventualprecise regulation of stem cell fates is an understanding of such pathways and networks. Therefore an importantaspect of the proposed studies is to identify global regulatory pathways and networks in ES cells as these cells areinduced to self-renew or differentiate under controlled conditions. Extracting pathway or network architecture formglobal gene expression, proteomic, or other highthroughput analyses is a new and exciting area of research, andthere are as yet no "written-in-stone" methodologies to accomplish such goals. However, a number of laboratorieshave successfully implemented perturbation approaches where individual gene products are up or down-regulated,and the resulting effects on global transcriptional profiles have been measured using microarray technologies.Statistical and computational strategies have been used to extract pathway and network architectures from such datasets. The Lemischka laboratory has been developing systematic methodologies to perform similar perturbationanalyses in ES cells. Key to their studies is the ability to utilize inhibitory RNA (RNAi) to downregulate the expressionlevels of transcriptional regulators in ES cells. The exact strategy utilizes the lentiviral vector delivery system.Importantly, there is an iterative aspect to our overall project. Any regulatory pathways and networks that areidentified in the Lemischka laboratory will provide additional components to incorporate into the artificially controlledswitch circuits. It can be anticipated that we will identify numerous novel master switches that are found up ordownstream of the ones that are the focus of our current proposal.
In order to validate the cell fate regulators, we built lentivirus vectors that allow us to induce expression of MyoD,Ppary, NeuroD3, and Nanog. Each of the vectors was constructed from a newly acquired lentivirus backbone, pLV-TRE-IRES2-EGFP (Lemischka Lab). This backbone has a CMV promoter driving the expression of the viral mRNA,beginning in the 5' LTR and ending at the deltaU3-3' LTR [Miyoshi, H., et aI., 1998]. In order to limit viral replication toa single round, separate viral protein constructs are used to trans-complement the overexpression vector. The gag,pol and env genes are removed from the virus and expressed on two separate packaging plasmids usingheterologous promoters. The packaging and vector plasmids can be co-transfected into cells [Buchschacher, 2003].The viral proteins will package vector DNA, and vector virus that is released from these transfected cells can beharvested for transduction of target cells. Because the helper packaging plasm ids do not contain the cis-actingsequences necessary for propagation, they will not be packaged or transferred to the target cells; therefore, there willbe only a single round of vector replication and the foreign genes can be introduced into the target cells in a relativelypredictable and efficient manner. The backbone also contains elements necessary for bacterial cloning.
For induction of the cell fate regulators, a minimal CMV promoter with seven tet operators (Tet Response Element;TRE) is used to transcribe the gene of interest. The Internal Ribosome Entry Site (IRES2) allows for co-expression ofEGFP that serves as a reporter. We acquired an ES cell line (TetON-ES) that encodes the reverse Tetracycline-controlled transactivator (rtTA). When the growth media of these cells is supplemented with DOX, it results intranscriptional activation from the TRE promoter. We have validated that TetON-ES cells transfected with the circuitsin Figure 1 express EGFP when induced with Doxycycline (DOX), and result in differentiation resulting in a cell fatecorresponding to the overexpressed master regulator as shown in Figure 1.

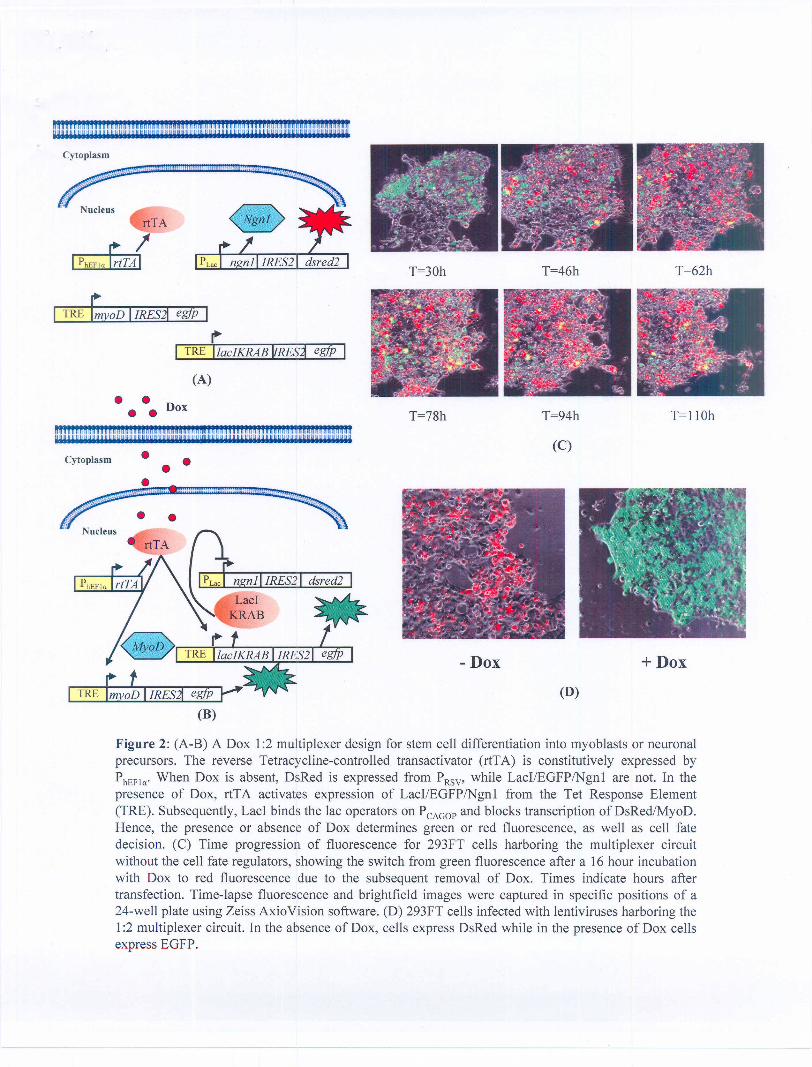
We built and testing a 1:2 multiplexer with a single inducer input and two fluorescent protein outputs as shown inFigure 2. The 1:2 multiplexer uses the membrane-diffusible compound (DOX) as the input, and expression of theEnhanced Green Fluorescent Protein (EGFP) and red fluorescent protein (DsRed2) as its two possible outputs.Figure 2 shows the plasm ids constructed for the multiplexer. pLV-CAGOP-IRES2-DsRed2 was constructed frompLV-Hef1a-IRES2-DsRed2 (Lemischka Lab) by inserting the the chicken .a-actin promoter with a CMV enhancer(CAG) with Lac01 and LacOid sites at position +3 of the transcription start site of the human EF1a promoter. Thedesign and characterization of the pLV-CAGOP-IRES2-DsRed2 plasmid is depicted in Figure 3. Also, pLV-TRE-Lacl-IRES2-EGFP was constructed from pCMVlacl (Stratagene) and pLV-TRE-IRES2-EGFP. The reverseTetracycline-controlled transactivator (rtTA) is constitutively expressed by the PCMV promoter. When DOX is absent,DsRed2 is expressed from PCAGOP, while Lacl and EGFP are not. In the presence of DOX, rtTA forms an activatorcomplex that binds the Tet Response Element (TRE) and activates expression of EGFP and Lacl. Subsequently, Laclbinds the Lac operator sites on PCMV and blocks the transcription of DsRed2. Hence, the presence or absence ofDOX determines green or red fluorescence output accordingly.
Subsequently it was determined that the addition of a KRAB domain to repressors (Figure 4) was necessary forefficient and tight repression of the promoter. pCMVLaclKRAB was constructed from pLV-tTRKRAB-Red (Trono Lab,ETH, Zurich) and pCMVlacl (Stratagene). The LaclKRAB was inserted in pLV-TRE-IRES2-EGFP to obtain thelentiviral vector pLV-TRE-LacIKRAB-IRES2-EGFP.
Figure 4 shows the plasm ids constructed during the contract period towards achieving the toggle switch network.The final design of the toggle switch was as follows: The pLV-Hef1a-tet02-IRES2-EGFP precursor plasmid wasconstructed by insertion of 2X tetO operator sites in the plasmid pLV-Hef1a-IRES2-EGFP. The laclKRAB gene frompCMVlaclKRAB was inserted before IRES2 to construct pLV-Hef1a-tet02-lacIKRAB-IRES2-EGFP. The TeTRKRABgene from pLV-tTRKRAB-Red was inserted before IRES2 from pLV-CAGOP-IRES2-DsRed2 to construct pLV-CAGOP-tetRKRAB-IRES2-DsRed2. The two components comprise the full toggle switch network. The mainelements of the toggle switch network are two transcription factors, LaclKRAB and TetRKRAB, that cross-represseach other. The network design is based on our earlier computer models [Weiss et. ai, 1999] and a recentimplementation in E. coli [Gardner, 2000]. The double negative feedback of the repressors translates into a positivefeedback that sustains the current state despite the absence of inputs. Under normal conditions, either LaclKRAB orTetRKRAB are expressed, but not both. EGFP and DsRed2 are used to approximate cytoplasmic levels of LaclKRABand TetRKRAB, respectively. Assume that initially both LaclKRAB and TetRKRAB exist in low concentrations. WhenIPTG is added to the media, it freely diffuses into the nucleus, binds LaclKRAB, and prevents LaclKRAB fromrepressing TetRKRAB expression. After some time, LaclKRAB decays and only TetRKRAB remains. Even after IPTGis removed from the media, the system maintains this steady state and continues to express TetRKRAB. To switchthe state of the system, aTc is added to the media. aTc freely diffuses into the nucleus, binds TetRKRAB, andprevents repression of LacIKRAB. As a result, LaclKRAB concentrations rise and subsequently TetRKRAB decays.Again, if aTc is removed from the media, the system maintains its other steady state where Lacl is expressed at highlevels. Thus it is possible to select the permanent expression of circuit output with transient administration of theappropriate inducer. Some problems were encountered packaging the pLV-CAGOP-IRES2-DsRed2 and efforts arecurrently underway to overcome this.
Over the duration of this project significant progress was made towards identification of potential cell fate regulatorgenes implicated in the myogenic and neuronal differentiation pathways. As a model cell fate decision to investigatewe have chosen self-renewal versus differentiation. We have utilized short hairpin inhibitory RNA (shRNA)methodologies to down-regulate the expression levels of numerous gene-products in ES cells. We have chosen todown-regulate molecules that are candidate regulators of self-renewal. These were chosen from a set of gene-products that are turned-off with rapid kinetics following induced differentiation of ES cells. We have developedefficient procedures to identify exact mRNA sequences to target with shRNAs. We constructed lentiviral vectorsexpressing individual shRNAs, and introduced these into ES cells. In a number of cases the individual shRNAscompromised the ability of the ES cells to undergo self-renewal. Our efforts so far have therefore uncovered anumber of molecules that are required of self-renewal in the ES cell system. Most of these are transcription factors,and therefore may control numerous downstream genes that control ES cell fate decisions. We have begun toidentify such downstream target genes using shRNA down-regulation together with genome-wide microarraytechniques. Our results suggest the existence of numerous "master regulatory" gene-products that control

differentiation along distinct lineages and pathways. These gene-products are excellent candidates for inclusion inartificially controllable regulatory modules.
In order to identify gene-products that may be critical in promoting ES cells to differentiate into neuronal populations,we have obtained an ES cell line engineered to express Green Fluorescent Protein from the regulatory regions of theSox-1 gene. This is a marker for neuronal differentiation. Identifying such regulators should facilitate the controlleddifferentiation of ES cells into neuronal lineages. In the future, similar strategies can be envisioned to identify "masterregulators" for other cell types necessary in spinal cord function.
We initially encountered problems packaging some of the genetic circuits into lentiviruses with the requisite highefficiency. Specifically, we had difficulties packaging constructs that have more than one promoter in the same virus.Also, constructs that coded for dsRed-Express, one of the reporters originally used for determining the cell state inboth the inverter and toggle switch plasmids were packaged inefficiently with low viral titers (105 particles / ml). Wealso encountered difficulties with the construction of 'tight' promoters that had low levels of leaky expression in theabsence of the inducer molecules. Specifically, construction of the lac promoter proved to be the biggest challengeand we had to go through several variants, battling lentiviral packaging problems en route before landing on ourcurrent design of CAGOP which shows promise. Efforts are underway to construct and test another lac promoterHef1 a-Lac01 Oid resulting in the plasmid pLV-Hef1 a-Lac01 Oid-IRES2-DsRed2 from pLV-Hef1 a-IRES2-DsRed2(Lemischka Lab) by inserting the Lac01 and LacOid sites at position +3 of the transcription start site of the humanEF1 a promoter.
The Weiss lab built and tested artificial pathways that orchestrate gene expression in mammalian cells. These effortsrepresent the first steps to building more complex circuits that will ultimately allow us to direct the cells to followparticular differentiation pathways in a programmed manner. Specifically, considerable progress was madeconstructing a 1:2 multiplexer and a toggle switch as described above. The collaboration with the Lemischka lab gavebirth to a unique environment with interdisciplinary expertise in both life sciences and engineering. Members of ourlabs freely and routinely interacted on a daily basis, and often work together in one of the individual labs. The overallenvironment that we have created fostered the rapid transition from ideas to implementation. The research donetowards accomplishing the aims of this proposal led to the establishment of the basic groundwork necessary for theapplication of synthetic biology and bio-engineering to stem cell research and has resulted in research towards moreambitious proposals such as to create a 3-D tissue pattern of differentiated tissue cells from an autonomouspopulation of mouse embryonic stem cells based on engineered cell-cell communication, quorum sensing coupled tomaster cell fate regulator genes. One such proposal titled "Artificial signaling pathways for tissue regeneration" basedon an adaptation of the Lux quorum sensing pathway to mammalian cells was submitted to NJSCR in December2006.
E. List of publications emerging from this research, including those inpreparation
S. Subramanian, P. Purnick, I. Lemishcka, R Weiss, "Experimental analysis of a synthetic toggle switch forcell fate determination," in preparation.
S. Subramanian, P. Purnick, I. Lemishcka, R Weiss, "Multiplexed cell fate determination based on artificialsender-receiver communication in mouse embryonic stem cells," in preparation.
Chambers, I., et aI., (2003) Cell 113: 643-655.Davis, RL., et aI., (1987) Cell 51: 987-1000.Gardner, T., et aI., (2000) Nature 403: 339.Ma, Q., et aI., (1996) Cell 87: 43-52.Miyoshi, H., et al. (1998) Journal of Virology 72: 8150-8157.

Nakashima, K., et al. (2001) Curr Opin Cell Bioi 13(6):721-7.Nakashima, K., et al. (2002) Cell 108(1 ):17-29.Weiss, R., et aI., (1999) Proceedings of the Dimacs Workshop on Evolution as Computation.

• •••••!!!!I!!!!!!I!!!!I!!!!!!!U!UMiIftI!,i!I!!!!!!I!III!I!!!!mUIImlIl!U!UII" II ~ U H II U II II uU!lUUlluHHuYSfUY~n!!f!l!U III UU II H II ~ U II II U~~U~~yyu;t1nYHUnUY
•• •~ 101
s::Q)()
~ 10°I-o~u: 10-1
10-3 10-2 10-1 100 101
Dox concentration (ug/ml)
Figure 1: (A) mES cells expressing cell fate regulators (the CFR is either MyoD, Nanog or Ngn 1) from the1:2 multiplexer construct. rtTA is expressed constitutively in the Ainv15 mES cell line and activatestranscription from the TRE promoter in the presence of Dox. (B) A dosage response curve of the TREpromoter depicting relative levels of EGFP expressed from the promoter as a function of Dox concentration.The mean fluorescence values were measured using FACS. (C-E) Ainv15 mES cells that constitutivelyexpress rtTA were infected with a virus encoding either Ngnl/EGFP, MyoDIEGFP, or NanoglEGFP underthe control of the TRE promoter. Cells were grown in the presence of LIF with and without Dox 24 hourspost infection. In the absence of Dox, cells maintained self renewal (not shown), while the presence of Doxresulted in differentiation into (C) cells with a neuronal morphology when Ngnl was overexpressed afterthree days; (D) cells that resemble muscle precursors when MyoD was overexpressed after seven days; and(E) cells that maintained stem cell morphology when Nanog was overexpressed for four days. The Ngnlimage is zoomed in by a factor of approximately four relative to the MyoD and Nanog images.
•••ftftlll!!•••ft••••!!·ft •••ftft••••ft1·1ft••! II·U •••ft••••lI!·a•••ftftlllu•••n••••lftllln"ln"'ln·n••nhllllllft·II "'1111"'ft,.,ft"'UlllU•••ft••••ftI·ft"'ftft"'ftft"'ft"nftlllhft"'ft"'I'"',ft"'ftI"'ftI"'ift""Um I I I I I II I Ii Ii I I I I mmY9uummy" I I I I I I I I" mm I I I I KHiypHRHUpumy
~~laclKRAB~
(A)
• • Dox• •ftftmft!ftftftftftftftftmmmmmmmnftftftlftlft!ftftftftftftftftmmftmmmm~ V V H Y U u u u U U H U Y U U H" H H UYHHBB g H H HYMN NY WY MY M H H V Y Y Y H Y H Y Y Y MY II K VYHHPHBHHH HRNSUY
• •••C'.""".."""'''''.'''.''.'''..'''.'~..... .. '-.. Nucleus
Figure 2: (A-B) A Dox 1:2 multiplexer design for stem cell differentiation into myoblasts or neuronalprecursors. The reverse Tetracycline-controlled transactivator (rtTA) is constitutively expressed byPhEFla'When Dox is absent, DsRed is expressed from PRSV' while LacI/EGFP/Ngnl are not. In thepresence of Dox, rtTA activates expression of LacI/EGFP/Ngnl from the Tet Response Element(TRE). Subsequently, LacI binds the lac operators on PCAGOPand blocks transcription of DsRed/MyoD.Hence, the presence or absence of Dox determines green or red fluorescence, as well as cell fatedecision. (C) Time progression of fluorescence for 293FT cells harboring the multiplexer circuitwithout the cell fate regulators, showing the switch from green fluorescence after a 16 hour incubationwith Dox to red fluorescence due to the subsequent removal of Dox. Times indicate hours aftertransfection. Time-lapse fluorescence and brightfield images were captured in specific positions of a24-well plate using Zeiss AxioVision software. (D) 293FT cells infected with lentiviruses harboring the1:2 multiplexer circuit. In the absence of Dox, cells express DsRed while in the presence of Dox cellsexpress EGFP.

r""'"'''''''''' ,.
~~~laclKRAB ~
• ••
• •~' ~""''''''''''''
• • •
-::::>::S 50Q)(.)
cQ)(.)
Cf)
~ 25o::::l
LL
Figure 3: Characterization of the lac promoter. (A) Circuit used to characterize the PCAGOP promoter.In the absence of IPTG, LacIKRAB which is expressed constitutively respressed the production ofDsRed from the PCAGOP promoter. When IPTG is added to the system, it binds LacIKRAB andprevents the binding of LacIKRAB to the PCAGOP promoter, hence DsRed is expressed. (B)Transfection of the circuit shown in (A) into 293FT cells. Brightfield and DsRed channels in theabsence (top) or presence (bottom) ofIPTG. (C) FACS analysis of (B).

- i"'an"'h"'h"'h"'h"'h"'U"'ijU"'ii Ilft"'hi"'iiii"'iU"'Ih"'H6"'U"'U'"W"'h"'h"'ii"'''''U''U'''i U"'U"',iif,"',HH"'ftri"'U"'H"U'"'U-RftH.IHb~Dij;;iUiji Utl" IbRMIlRHU'IHSFBBHF22HWnU;bl"UU! p UiSiY"HYYYUarBIlC2lJtitIY•Cytoplasm
~!1..2L[ lac1 ~'-- ...•..._ ....•..••"'--
~~ lac1 ~ myoD-egflJ
~ ..
.". .
I DBlank. - aTe. + aTe I
-:-120=><C 100- 80
604020
o
~~ tetR ~I ngnl
ad yoD
~! !~lacl ~-m-y-o-D---eg-fj-lJ- +aTc
(E) Effect of KRAB domain
Figure 4: Toggle switch network in stem cells. The lac promoter PCAGOP-ni (PLac in the figure) wasconstructed by placing the lacDl and lac Did operators 74.5 bp apart at the +3 transcription site of achicken ~-actin promoter with a CMV enhancer. The non-coding intron region of the promoter wasremoved to enhance packaging of the lentivirus. The tet promoter was constructed by placing twocopies of the bacterial tet operator downstream of the TATA box ofa human EFI-a promoter (hEFla).(A) Initially both Lacl and TetR exist in low concentrations. IPTG is added and freely diffuses into thenucleus, binds Lacl, and prevents Lad from repressing TetR. After some time, Lad decays and onlyTetR and Ngnl remain. (B) Even after IPTG is removed from the media, the system maintains thissteady state and continues to express Ngn 1. (C) To change the state of the system, aTe is added andbinds TetR, preventing repression of Lad. As a result, Lad and MyoD/EGFP concentrations rise.Subsequently, TetR and Ngnl decay. (D) If aTe is then removed from the media, the system maintainsits other steady state condition where Lacl and MyoD/EGFP are expressed at high levels. (E) Theeffect of KRAB domain fusions on the repression levels: Fusion of a KRAB domain to TetR results inalmost complete repression of the fluorescence from the PhEFlaJTet02 promoter. Brightfield andfluorescence images of 293FT cells co-infected with lentiviruses encoding PhEFla-TetRKRAB-IRES2-Puro and PhEFlaJTet02-Lacl-IRES2-EGFP48 hours post induction with (top) or without (bottom) 100ng/ml aTe.










![[HE-3044] Groupe Adèle - adeleplus.com€¦ · [HE-3044] Groupe Adèle Brief to PSD2HTML May 1st, 2014 Document completed by Vanessa Moreau Nadine Lessard Groupe Adèle Project-](https://static.fdocuments.in/doc/165x107/5f117192862b5e59e42e1dd8/he-3044-groupe-adle-he-3044-groupe-adle-brief-to-psd2html-may-1st-2014.jpg)







