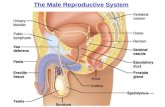
Reproductive Hormones; Male Reproductive Endocrinology Chapters 16, 17.

1993Volume 4 No.
ORNITOLOGIA NEOTROPICAL 4: 1-41, 1993@ The Neotropical Ornithological Society
FIELD STUDY OF ALLOPATRY, SYMPATRY,PARAPATRY, AND REPRODUCTIVE ISOLATION
IN STEPPE BIRDS OF PATAGONIA
Fran~ois Vuilleumier
Department of Ornithology. American Museum of Natural History,Central Park West at 79th Street, New York, N.Y. 10024, U.S.A.
Resumen. La vegetación estépica o subdesértica de Patagonia (América del Sur desde 36 oS hasta 56 OS) representae170% de la superficie total de la región (1.140.000 kilómetros cuadrados). Esta zona, con una población humanaescasa sirve sin embargo para una industria ovejera extensiva. De las aproximadamente 90 especies de aves terrestresque anidan en estepas patagónicas, unas 70 apartenecen a un gremio ecológico de especies que buscan su alimen-tación en el suelo o cerca del suelo en la vegetación baja, herbácea o arbustiva. Aunque la fauna esta empoberecida,se encuentran varios casos de distribución que sugieren patrones de especiación local, incluyendo ejemplos deespecies alopátricas o parcialmente simpátricas, y ejemplos de aislamiento reproductivo post-especiacional quemerecen investigación detallada. Durante una transecta desde el Oceano Atlántico hasta la wna pre-Andina en laProvincia del Chubut, Argentina, en Noviembre de 1991, se estudiaron de manera preliminar problemas de alopa-tria, simpatria, y aislamiento reproductivo en una muestra de 26 especies (en 14 generos: Eudromia, Tinamotis,Thinocorus, Geositta, Upucerthia, Eremobius, Asthenes, Leptasthenura, Agriornis, Muscisaxicola, Mimus, Anthus, Si-calis, y Phrygilus), representando e130% de las especies terrestres anidando en estepas de Patagonia. Las interaccio-nes entre varias especies congenéricas (o afines) estan discutidas en términos de diferencias o semejanzas al nivelecológico al nivel etológico, y en términos de territorialidad interespecífica. Investigaciones futuras deberán averi-guar si los patrones descritos en este artículo pueden estar verificados por medio de trabajos a largo plaw. Parececlaro que fenómenos eco-evolutivos acerca de problemas especiacionales y distribucionales en el extremo sur delcontinente Sudamericano merecen más amplia investigación.
Abstract. The steppe or semi-desert vegetation (shrubsteppe) of Patagonia (southern South America between 36°Sand 56°S) covers about 70% of the total area of this region (1,140,000 square kilometers). Although ~his wne hasa sparse human population, it is used extensively to raise sheep. Of the 90-0dd species of landbirds that breed inPatagonian steppes, about 70 belong to the ground or near-ground foraging guild. In spite of the low species diversi-ty in the Patagonian steppe avifauna, speciation appears to have been active in that region in the past, as revealedby the occurrence of severa! distribution patterns among congeneric or closely related species, which suggests thatthese patterns are the results of local speciation. These cases include instances of alIopatry, partial sympatry, andreproductive isolation. During a transect from the Atlantic coast westward to the foothills of the Andes in ChubutProvince, Argentina, in November 1991, preliminary field studies were carried out on a sample of 26 species (in14 gene1"a: Eudromia, Tinamotis, Thinocorus, Geositta, Upucerthia, Eremobius, Asthenes, Leptasthenura, Agriornis,Muscisaxicola, Mimus, Anthus, Sicalis, and Phrygilus), representing about 30% of the species breeding in Patagoniansteppes. Interspecific interactions (including differences or similarities in habitat preferences and behaviora! traits)were studied in order to better understand patterns of eco-geographic overlap versus non-overlap. Preliminaryconclusions about the modalities of reproductive isolation suggest a number of questions for future, long-termresearch on the details of the speciational history of Patagonian birds. Accepted 3 july 1992.
Key words: Steppes, Patagonia, Chubut, Argentina, landbirds, allopatry, sympatry, reproductive isolation, bio-
geography.

VUILLEUMIER
In spite of their relative structural and botani-cal uniformity the steppes of Patagonia have avery interesting avifauna. First, although it isdepauperate and includes only about 90 speciesof breeding landbirds, it is taxonomically varied.Especially remarkable is the guild ( cf. Root 1967)of species foraging on the ground or in lowshrubs near the ground and eating seeds and/orsmall invertebrates. This guild has about 70 spe-cies in eleven families: Rheidae (rheas), Tinami-dae (tinamous), Charadriidae(plovers), Thinoco-ridae (seedsnipe), Furnariidae (ovenbirds), Rhi-
nocryptidae (tapaculos), Tyrannidae (flycatch-ers), Mimidae (mockingbirds), Motacillidae (pi-pits), Icteridae (meadowlarks), and Emberizidae(finch-Iike birds). Secondly, members of thisguild are not only speciose, but they are alsonumerically dominant at many sites (especial-ly Furnariidae, Tyrannidae, and Emberizidae).Thirdly, in several avian groups, geographical andecological overlaps between similar speciessuggest that the physiognomically simple steppehabitats of Patag2nia can sustain a rather com-
INTRODUCTION
The vegetation of In()st of Patagonia (southernSouth America; defined in Vuilleumier 1985,1991a) consists of dry shrubsteppes on vast pla-teaus, which often stretch, gray-green in color,from one end of the horizon to the other (Fig.1). Perqaps as much as 800,000 km2 of Patago-nia's 1,140,000 km2, or about 70%, is coveredwith steppes. A very sparse human populationuses these steppes to raise sheep. Steppe habitatsextend for hundreds of kilometers, from theshores of the Atlantic Ocean in the east all theway to beech (Nothofagus) forests at the foothillsof the Andes in the west (map in Hueck & Sei-bert 1972). Here and there, however, the relativemonotony of this arid landscape is broken byspectacular cliffs of colorful rocks or huge screes(Fig. 2), by shallow lagoons, extensive salt flats,and green and shady manmade oases (chacras)where willows (Salix) and poplars (Populus) growalong some river valleys (Fig. 3) and where vari-ous crops are cultivated on the irrigated valleyfloor.
FIG. 1. Very open, dry shrubsteppe on soft, sandy substrate near Piedra Parada, Chubut, looking westward towardthe snow-capped peaks of the Andes. Vegetation is "Patagonian Steppes and Semideserts, Subandean and WesternSector," No.64 in Hueck & Seibert (1972). Photo F. Vuilleumier, November 1991.

STEPPE BIRDS OF PATAGONIA
FIG. 2. Top: cliffs in the Chubut river valley a few km west of Paso del Sapo, Chubut; valley floor in foreground,Chubut River behind photographer. Bottom: scree of large rocks and boulders, Chubut river valley a few km westof Piedra Parada, Chubut; Chubut river behind photographer. Photos F. Vuilleumier, November 1991.
3

'UILLEUMIER
plex ecological assembly of birds. Fourth, in afew of these groups, t~e species concerned are ei-ther congeners or members of closely related ge-nera. Finally, several cases of sympatry orparapatry occur between species ;hat appear tobe reproductively isolated and to have speciatedeither in Patagonia or in neighboring regions
(Vuilleumier 1991a, 1991b).To the biogeographer the steppe avifauna of
Patagonia poses an ecological challenge in termsof niche segregation and interspecific competi-tion (proximate factors of community struc-turing), and an evolutionary challenge in termsof the spatio-temporal origins of the parapatricor sympatric situations observed today (ultimatefactors of speciation). Community structure hasusually been studied by ecologists who haveanalyzed the factors ( climate, vegetation, inter-specific competition) thought to be responsiblefor overlaps between or among species of birds ingiven yegetation types. For example, Wiens &Rotenberry (1980) and Rotenberry & Wiens(1980) described avian communities in the grass-lands and shrubsteppes of North America, envi-ronments that are equivalent to some of thesteppes of Patagonia.
Speciation has usually been studied by evolu-tionists interested in patterns of differentiationdetectable between sister species. Rarely have pat-terns of allopatry/sympatry between or amongspecies been studied in an entire avifauna, in anattempt to combine an analysis of proximate(ecological) and of ultimate (evolutionary) fac-tors.
In this paper, I examine the ecological ques-tion of overlaps versus non-overlaps and assessthe evolutionary problem of reproductive isola-tion in several congeneric or closely related pairsof species occurring in north-central Patagonia,in an attempt to document patterns of overlapsin a substantial portion of the Patagonian land-bird fauna. The 26 species discussed below con-stitute about 30% of the breeding landbird faunaof Patagonian steppes (total about 90 species).
In two recently published surveys of specia.ition phenomena in Patagonian landbirds, I con-cluded that this region showed many instances ofvarious stages in the speciation process (Vuilleu-mier 1991a, 1991b). I suggested that an analysis()f the nature of secondary overlaps (includingparapatry) was necessary before significant pro-
gress could be made in our understanding ofavian evolution in that region. Among specificquestions that need answers I included (Vuilleu-mier 1991a: 25): (1) "How do the species behavetoward each other in areas of secondary overlap?",(2) "Is interspecific territoriality common?", and(3) "What is the nature of reproductive isolationin parapatric zones?"
Gochfeld (1978) studied habitat selection be-tween two species of Mimus in northeastern Pata-gonia and interspecific territoriality between twospecies of Stumella ( 1979) at the northern edge ofthe Patagonian region. These two papers appearto be the only ones focusing on the problem in-vestigated here, although Maclean (1969) men-tioned habitat differences between species ofThinocorus and Short (1968) studied sympatry inStumella north of Patagonia. Gochfeld (pers.comm.) also studied Phrygilus and Anthus, butunfortunately did not publish his results. Paperssuch as those by Durnford (1877, 1878), Peters(1923), Wetmore (1926a, 1926b), and Fjeldsa(1988), and the book by Fjeldsa & Krabbe (1990)all give valuable information on the distributionof many Patagonian bird species, but are of littleuse in elucidating patterns of overlap and repro-ductive isolation, because these authors were notworking on these problems. Cody (1970) di-scussed a series of patterns of overlaps amongcongeneric species of Chilean birds, includingground birds like Muscisaxicola. His paper ex-plored some of the questions of interest here, butbecause he worked west of the Andes and in theAndes themselves, and not in Patagonian steppeseast of these mountains, his results may not bedirectly applicable. Thus, fresh field work isnecessary. Evolutionary questions posed by al-lopatry or sympatry can be approached by fieldwork carried out at two out of severallevels ofbiogeographic perception (Blondel & Choisy1983): local (biotope in Blondel & Choisy 1983)and regional.
On a local geographical scale, my ongoingfield work in Chilean Patagonia and northwest-ern Tierra del Fuego on the genera Phrygilus andGeositta (Vuilleumier 1991a: 14-18,21-22, andunpublished) has been directed at one instance ofparapatry (Phrygilus) and one ofsecondary over-lap (Geositta). In order to study problems ofoverlaps on a regional scale, I carried out atransect in November 1991 in Chubut, across the
4

STEPPE BIRDS OF PATAGONIA
FIG. 3. Top: abrupt transition between shrubsteppe (foreground) and dense riverine vegetation of willows (Salix)and poplars (Populus) along Arroyo Telsen, a few km west of Telsen, Chubut. Bottom: open, overgrazed, grassyriverine vegetation with groves of willows (Salix) and poplars(Populus) along the Río Chubut, a few km west ofPaso del Sapo, Chubut. Photos F. Vuilleumier, November 1991.
5

steppes of north-central Patagonia from the At-lantic Ocean to the Ándean foothills. Duringthis trip I focused my attention on several groupsof ground or bush inhabiting birds presentingevolutionary problems and gathered informa-tion on distribution, habitat selection, relativeabundance, and interspecific interactions of thedifferent species involved. In this paper I reportfield observations concerning species in the gen-era Eudromia and Tinamotis (Tinamidae); 7hino-corus (Thinocoridae); Geositta, Upucerthia, Ere-mobius, Asthenes, and Leptasthenura (Furnarii-dae); Agriornis and Muscisaxicola (Tyrannidae);Mimus (Mimidae); Anthus (Motacillidae); and Si-calis and Phrygilus (Emberizidae). A few obser-vations I made in 1965 near Bariloche (RíoNegro Province) are also cited. This field work ispart of a long-term research program on the bio-geography, ecology, and evolution of the avifau-na of Patagonia (Vuilleumier 1967 a, 1967b, 1972,
1985, 1991a, 1991b).
(map of entire territory of Chubut Province),and 1:400000 (insets for Península Valdés/Ma-dryn/Trelew/Rawson area, and for El Bolsón/Esquel area). Two other maps that are widelyused and available in Argentina have either in-complete road localizations, or incomplete local-ity names, or both, at least in the transect area.They are the Esso/Exxon road map ("Mapa car-retero República Argentina," 1986, Esso S.A. Pe-trolera Argentina, scale 1:4000000) and theundatedmap, "República Argentina: Red Cami-nera Principal," published by the ACA, scale1:4000000. Of the two, the ACA map is thebetter one as far as roads and place names go.Note that the Times Atlas, Eight Comprehen-sive Edition, 1990, includes several of the local-ities mentioned in this paper on Plate 121, scalel:s000000.
Field observationsThe target taxa of birds selected for analysis ofallopatry versus sympatry and reproductive iso-lation were observed at about 20 study siteschosen to represent the range of steppe habitatsas well as other vegetation and landscape types,as described below. At each site, a period of from1-8 hours (average about 3 hours) was spent ob-serving birds and noting the following: habitatpreference, relative abundance, territorial behav-ior, nesting behavior, foraging behavior, vocalbehavior, and interspecific behavioral interac-tions, if any. No birds could be collected. Duringthe study period the weather was generally good,with sunny skies, little or no cloud cover, goodto excellent visibility, mild temperatures reachingabout 25 °C by midday or early afternoon, andlittle windj rain and thunderstorms were en-countered only on the high basaltic plateaus be-tween Telsen and Gan-Gan. Locally (near PuertoMadryn and Sierra Chataj and near the intersec-tion of routes 3 and 30 south of Uzcudún) ashclouds from Cerro Hudson in Chile at about46 OS impeded the visibility slightly. VolcanoHudson (identified in Fig. 4) erupted in August1991 and produced vast volumes of fine ash,much of which was deposited not only near theChilean border but also all the way to theAtlantic Ocean as far west as the ports of PuertoDeseado and San Julián (see N.C. Nash, NewYork Times, Monday, October 21, 1991, pp. Al
and A6).
MATERIAL AND METHODS
TransectField observations were made in the australspring, between 5 November and 18 November1991 in Chubut Province, Argentina, along atransect from the Atlantic Ocean at PenínsulaValdés and Cabo Dos Bahías westward to Esquelat the foot of the Andes, between about 42 oS and45 oS and 64 OW and 71 oW. Fig. 4 shows the loca-lization of the transect (dotted line) and indicatesthe routes and the main localities along them.From Trelew to Puerto Madryn I followed route3. From Puerto Madryn to Península Valdés Ifollowed routes 2, 47, and 52. From Puerto Ma-dryn to Esquel I followed route 4 to Telsen, Gan-Gan, Gastre, and El Molle,then route 13 to Pasodel Sapo, and route 12 to Gualjaina and Esquel.From Esquel, I went backto the Atlantic coastfollowing routes 40 and 25 to Las Chapas, thenroute 31 to Uzcudún and routes 3 and 30 to theCamarones/Cabo Dos Bahías/Puerto Melo area.I ret1;!rned to Trelew via routes 30, 3, and 9through Gaimán.
Locality names on Fig. 4 are taken from the
undated map "Provincia del Chubut," publishedin Buenos Aires by the Automóvil Club Argen-tino (ACA). This map has a scale of l:l000000

;TEPPE BIRDS OF PATAGONIA
I L E ~~ A R G E N \T I N A VaJChe~\
I /-" , Lago R¡O NEGRONahuel
~ ) ~ ~~api Ingenjero Mequjncheo ó9 I~ : ~ JeccbeOO O ..,. ~
Puerto{¡o--' Beriloche O ~e"","""\Montt OHuanuluan SO
El \\'00",Bo '-" c .-., ~n J!"\ -..c .-.
.-., Gastra I Puerto
ElMolleCf""4.[j] ::;; ~..~5an Madryn~ Piedrel ...~...o :~ ~Gualjajna Paraca. Gan-Gan
I !!l 'ia ~ ~ ..p ..p ~ del Sapo F, ~n." .~.~ ~
, <).
La5... ~ 'Salinas O o
,.ckaO Colan.\ C,Conhué ..
b..Pempe de Agnja
San Antonio Oeste ""-0~
~Golfo San Matías
).../
} San Golfo I -42"
J~Sa~~
si.,go"alau((
, !TH
J4.TLANTIC
,OCEAt:J- 44°
Laslas " 'I
~
1 " , I
l..-,",
.-,,)-',
'...I
-del -, ,Indios Altaras
CHUBUT\
Ainegh¡;';'r
'40.-f-I c ~
, ~I' ~
.t! \ UI -
II.
U
~~ -
~..44° \w ¡¡(t::.L -40"
f)'nes
~ i# L
Colh~' ~'
DosPuerto BahíasMelo
GO/~mD \San Jo:-j-
~ CHIL{,' I : .
.
' \46" -50'
s.rmle~to ~ ,
r--~. I I
CRUZ k;lometers\ ! 5'0 100 1 ~o 200 2~0 300
..;;..;.;Aq¡; ~
50'
46°-
"tFIG. 4. Schematic map of Chubut Province, Argentina, showing the transect route (dotted line) followed inNovember 1991. The numbers along the route are the road numbers (see text). The main localities mentionedin the text are indicated.
sand dunes near the ocean (Fig. 5). Areas ofgrassy steppes were seen near Punta Delgada onPenínsula Valdés (Fig. 6) and near Colan Conhué(no illustration). At scattered localities, substan-tial man-made oases with relatively dense grovesof willows (Salix humboldtiana) and poplars(Populus sp.), and with locally extensive riverinemarshy vegetation were encountered, notably onPenínsula Valdés, in the Telsen area (ArroyoTelsen, Fig. 3, top), near Paso del Sapo (RíoChubut Valley, Fig. 3, bottom), near Gualjaina,along the Río Chubut below the F. AmeghinoDam, and in the Gaimán area (Río Chubut). Avery narrow, often discontinuous, band of wil-lows and other riverine vegetation lies along thebanks of the Río Chubut between Piedra Paradaand Paso del Sapo, between Paso de Indios andLas Chapas, and in the Gaimán::rrelew area. Inmost of these areas, the green riverine oasesextend just a few meters away from the Río orArroyo and abruptly make way to arid shrub-
steppe (Fig. 3, top).Figs. 7 and 8 illustrate several types of steppe
vegetation along the transect from the base ofPenínsula Valdés westward to the Gastre area
VegetationShrubsteppes composed of low, spaced outbushes (many of them thorny) are the dominantvegetation throughout the transect. In mostareas, the ground is bare and made up of rela-tively fine material, often including wind- andsand-polished pebbles. In the central part of thetransect, especially between Telsen and Gastrt;outcrops of basaltic rocks are prominent. Ac-cording to Hueck and Seibert (1972: 43, 51-53),the main vegetation formations from east to westalong the transect are: (1) Monte-Shrubsteppe("Monte-Strauchsteppe" or "Estepa arbustiva deMonte;" no. 51), including Larrea, Prosopis, Cas-sia, and Chuquiraga, and (2) Patagonian Steppesand Semideserts ("Patagonische Steppen undHalbwüsten" or "Estepas y semidesiertos patag6-nicos;" central sector no.66, San Jorge sector no.67, and subandean and western sector no.64).Physiognomically important plants in nos. 64,66, and 67 include Berberis, Senecio, Chuquiraga,Verbena, and Mulinum spinosum (the last espe-cially abundant in the western sector).
In the Península Valdés area and near PuertoMadryn, the steppe vegetation grows locally on

-\, VUILLEUMIER
FIG. 5. Top: low, open shrubsteppe growing on sand dunes at the top of coastal cliffs, Puerto Pirámides, Chubut.Bottom: low, open shrubsteppe growing on coastal dunes and on top of low cliffs, a few km north of Puerto
Madryn, Chubut. Photos F. Vuilleumier, November 1991.
8

STEPPE BIRDS OF PATAGONIA
FIG. 6. Top: very open steppe of low grnsses interspersed with tiny shrubs, Punta Delgada, Chubut. Bottom: low,grassy steppe at the top of coastal cliffs, Punta Delgada, Chubut. Photos F. Vuilleumier, November 1991.
9

JILLEUMIER
FIG. 7. Top: relatively dense shrubsteppe at the base of Penfnsula Valdés, a few km west of Puerto Pirámides,Chubut; note absence of grass cover. Bottom: very open, low steppe on soft pebbly soil with grazed, hard grasstussocks in flat area of foreground and low shrubs on ridges of background, about 30 km east of Gan-Gan,Chubut. Photos F. Vuilleumier, November 1991.
10

"I
(;
STEPPE BIRDS OF PATAGONIA
11

VUILLEUMIER
(and Fig. 1 shows the vegetation toward Esquel).West of Esquel toward the Chilean border,Andean valleys receive more and more rainfall,and have, first relatively dry and open Austroce-drus woodlands and then mesic to very humidbeech (Nothofagus) forests. The avifauna of Pata-gonian beech forests has been treated elsewhere(Vuilleumier 1967a, 1967b, 1972, 1985, Vuilleu-mier & Kikkawa 1991) and was not studied dur-i.ng this transect.
butions, in Patagonia at least, appear to be depen-dent on their interactions.
Eudromia elegans was seen (isolated, or up tothree birds together) and heard commonly insteppes from Península Valdés to the Telsen area,and after an absence in the Gan-Gan area, heardagain near Gastre at about 1200 m, near Paso delSapo, and near Gualjaina. It was not seen orheard from there on to Esquel, or along the roadfrom Esquel to Los Altares in the Chubut valley,but was noted again in steppes between theF. Ameghino Dam and Uzcudún, and was seencommonly in the steppes of the Camarones/Puerto Melo/Cabo Dos Bahías coastal area (seeFig. 9). In spite of its relative abundance, E.elegans was not as common as Wetmore (1926b:32) reported from western Neuquén, where, hestates, "it was not rare to see 30 or 40, or even100, all adults, banded together" in 1920. Thespecies may be rarer today than 70 years ago (seeAppendix 1 about Nothura). Fig. 9 shows local-ity records of E. elegans in Río Negro accordingto Peters (1923), Wetmore (1926a, b), and Betti-nelli & Chebez (1986). Fig. 10 illustrates thecharacteristic habitats of E. elegans.
RESULTS
In this section, observations on overlaps, non-overlaps, and reproductive isolation are presentedfor each pair (or triplet) of species studied in thefield. For convenience, the order and nomencla-ture of birds follow Meyer de Schauensee (1982).
Eudromia and Tinamotis (Tinamidae)Eudromia elegans and Tinamotis ingoufi are theonly tinamous that I observed along the transect.Even though they are not congeneric and didnoL originate from the same speciation event, Iinclude a discussion of their distribution here be-cause of their similarity in size and color, as wellas in habitat preference. Their respective distri-
FIG. 9. Distribution of Eudromia elegans (black dots) and Tinamotis ingoufi (black triangles) along the transect
route; additionallocalities in R¡o Negro from the literature (see text).
12

STEPPE BIRDS OF PATAGONIA
FIG. 10. Two views of characteristic shrubsteppe habi-tat of Eudromia elegans. Top: a few km west of PuertoPirámides, Chubut. Bottom: a few km west of PuntaDelgada, Chubut. Photos F. Vuilleumier, November1991.
FIG. 11. Two views of shrubsteppe habitat of Tina-motis ingoufi; note very open nature of vegetation.Top: about 30 km east of Gan-Gan, altitude about1100 m, Chubut. Bottom: about 10 km west of Gan-Gan, altitude about 1050 m, Chubut. Photos F. Vuil-leumier, November 1991.
Tinamotis ingoufi was noted at only threelocalities (Fig. 9): (1) along route 4 about 30 kmE of Gan-Gan at 1100 m, (2) along route 4 about10 km W of Gan-Gan at 1050 m, and (3) about25 km W of Gan-Gan at 1050 m along a privateranch road off route 4. The three sites are similarto each other in that they have open steppe withlow and spaced out shrubs growing on sandy orpebbly soil, with hard cushion plants and sparsetufts of coarse, sheep-grazed tussock grass inter.spersed here and there (Figs. 11 and 12). Thesesites differ from one another in that the first oneis located on a vast basaltic plateau, whereas the
other two are somewhat different geologically,the soil being composed of pebbles and rocks ofsedimentary, rather than volcanic, material. Pe-ters (1923: 287) reported a specimen of T ingoufifrom Huanuluan, where E. elegans Boccurred spar-ingly in a few localities" (p. 286).
Thus Eudromia and Tinamotis were bothfound in steppes, but Tinamotis occurred only athigher elevations in the central part of the tran-sect, whereas Eudromia was found all the wayfrom sea level to a high elevation near 1200 m.Only one species was recorded (seen or heard) atany one site (Fig. 9). Although actual habitat
13

VUILLEUMIER
FIG. 12. rwo close-up views of the habitat of Tina-motis ingoufi about 25 km west of Gan-Gan, altitudeabout 1050 m, Chubut. Top: sparse shrubs and a hardcushion plant (center, diameter about 25 cm) onpebbly ground. Bottom: sparse woody shrubs onpebbly ground. Photos F. Vuilleumier, November1991.
and more melancholy kiewla of Tinamotis ingou-fi. Wetmore (1926b: 31) described the call of E.elegans as "a low mournful whistle given slowly"and compared it to the call of Rhynchotus rufes-cens. One slight behavioral difference betweenEudromia and Tinamotis is that, when alarmed,Eudromia keeps its raised neck slightly curved,whereas Tinamotis maintains its neck quitestraight, with the head held at a right angle, pe-riscope-like. When startled, Eudromia flies off ea-sily, but Tinamotis runs and disappears, simplymelting away in the low vegetation.
My observations of the habitat of Tinamotisingoufi do not seem to match those published byFjeldsa & Krabbe (1990: 64), who wrote that itoccurs in "grassland steppes, in sheltered valleyswith patches of dense, low brush (&rberis, Perne-tyia [sic], Verbena)," and "mainly at 200-800 min sheltered valleys between the barren andwindy plateaus of Arg. Patagonia from w RíoNegro to Sta. Cruz. .." Fjeldsa (1988: 87) hadwritten earlier: "Southern Patagonia is irihabitedby the Elegant Crested Tinamou (Eudromias [sic]elegans) and the Patagonian Tinamou (Tinamotisingoufi), which are both restricted to brush andshrub in sheltered valleys, and avoid the wind-
swept plateaus."My observations suggest that Tinamotis in-
goufi occurs on these plateaus and that it is re-placed by Eudromia elegans in lower areas orsheltered valleys. In fact, the distribution map inFjeldsa & Krabbe (1990: 63) shows a gap in thedistribution of Eudromia elegans in Río Negro -
Chubut, precisely where high elevation basalticplateaus are located and where Tinamotis ingoufioccurs.
Clearly the geographical and ecological distri-bution of these tinamous in Patagonia requiresmore field work before it can be understood. Anattempt should be made to locate a site whereEudromia elegans and Tinamotis ingoufi occurnear each other and where their habitat prefe-rences, foraging habits (including food items),and direct or indirect interactions (includingcompetition) could be quantified. The variousmechanisms that keep them separate in areas ofcontact or overlap need to be studied.
overlap should be expected (see Peters 1923, citedabove, and range description in Olrog 1979), Idid not observe it. These two species are verysimilar and may exclude each other mutually.The crest of Eudromia is lacking in Tinamotis,but otherwise both species are large tinamouswith similar head and neck patterns and withlong necks which they raise vertically in similarways to observe their surroundings. Both speciesare fond of foraging on dirt roads. The flute-like,two or three syllabic, whistled calls of Eudromiaelegans remind me of the calls of certain AndeanGrallaria spp., but also of the louder, whistled,
Thinocorus (ThiJ1ocoridae )Notwithstanding some differences in size and
color, the two species of the genus Thinocorus are
14

very similar to each other and are likely to haveevolved from a common ancestor through asingle vicariant event: The apparent extent ofoverlap between these species makes it difficultto infer the nature and localization of that vicari-ant event, however. Descriptions or maps of thegeographic distribution of L rumicivorus and Lorbignyianus (e.g., Meyer de Schauensee 1982,Fjeldsa & Krabbe 1990) suggest a very extensivewne of overlap between them. Actually, the twospecies of 7hinocorus seem to have only partiallyoverlapping distributions and may in fact be
largely allopatric.Maclean (1969: 35) thus wrote of L rumici-
vorus that "in Patagonia ...it occurs far inlandon the open steppe, " and of L orbignyianus that
it "seldom descends below 700 meters except inthe extreme southern part of its range" (i.e., Pata-gonia). Elsewhere, Maclean (1969: 37) stated that"just as the Least Seedsnipe [L rumicivorus] isessentially a bird of the lowlands, the Gray-brea-sted S~dsnipe [L orbignyianus] is a bird of themountains." And further: "in the southernmostpart of its range the Gray-breasted is still more a
bird of higher elevations than the Least (Craws-hay 1907) although the inhabitants of ArgentinePatagonia assured me that in winter the Gray-breasted is common on the pampa" (p. 37).
In my experience 7hinocorus orbignyianuslives in mountain valleys and slopes and is usu-ally relatively scarce wherever it occurs, whereas7:' rumicivorus lives in open plains where it canbe extremely abundant. Thus, in a transect Icarried out through the steppes of Chilean Pata-gonia at about 52 oS, from Morro Chico west-ward to Gallegos Chico, O'Higgins, Kimiri Aike,and Punta Dungeness on February 27 and 28,1987 and on March 1, 1987, I saw only one 7hi-nocorus orbignyianus but thousands of 7:' rumici-vorus.
During the present transect in Chubut Pro-vince, 7hinocorus spp. were encountered at 12sites: 1 pair rumicivorus at Caleta Valdés, 1 pairrumicivorus 30 km E of Gan-Gan, 1 O' rumicivo-rus 35 km E of Gan-Gan, 1 pair rumicivorusabout 25 km W of Gan-Gan, song orbignyianusabout 40 km E of Gastre, song orbignyianus nearGastre, 1 pair orbignyianus with 3 downy chicks
FIG. 13. Distribution of Thinocorus rumicivorus (black dots) and T orbignyianus (black triangles) along transectroute; additional localities in Río Negro for these two species from the literature and personal observations
(see text).

VUILLEUMIER
FIG. 14. Top: habitat of Thinocorus rumii:ivorus about25 km west of Gan-Gan, altitude about 1050 m,Chubut. Bottom: close-up view of habitat of r rumii:i-vorus at same locality; low shrub (Verbena sp.) in centeris about 30 cm in diameter and 15 cm tall. Photos F.Vuilleumier, November 1991.
whereas L rumicivorus occurs mostly in thelower eastern part of the study area (habitat illu-strated in Fig. 14). The distribution pattern of7binocorus spp. is somewhat similar to that ofthe tinamous Eudromia elegans and Tinamotis in-
goufi.Durnford (1877, 1878) mentioned only
7binocorus rumicivorus, which he called "com-mon" (1877) or "abundant" (1878). In his 1877paper, he wrote that L rumicivorus was "seenmost frequently on the higher stony plateaux,but occasionally in the valley." Peters (1923: 292)found L orbignyianus to be "a characteristic resi-dent of the western portion of the Plain of Pata-gonia." He observed it "only in the vicinityof Huanuluan, almost invariably up an1ong thehigher gullies and ravines which cut back intothe table-lands or head far up on El Escorial." OfL rumicivorus, Peters (1923: 293) wrote that itwas "a common resident in the western part ofRío Negro" but that "unlike its larger relative, Lorbignyianus (sic), it does not frequent the rockygullies and ravines, but is found on the gravellyplains and sandy valleys." Peters (1923: 293) re-ported an adult female "taken seven miles east ofBariloche on February 13." I have seen L rumici-vorus in the Pampa de Nahuel Huapí near Bari-loche on 11 February 1965. Wetmore (1926a) re-ported specimens of L rumicivorus from ArroyoSeco (Río Negro, near Valcheta, Fig. 13), and ofL orbignyianus from Huanuluan and ArroyoAnecon Grande (both in Río Negro, the twolocalities being about 20 miles from each otherjFig. 13). In his 1926b paper, Wetmore stated thathe "encountered the small seed snipe on itsbreeding grounds on the closely grazed slopes ofan open valley in which there was a tiny streamand occasional little seeps or spring holes" at
Zapala, Neuquén.In the transect area, 7binocorus rumicivorus
would therefore appear to occur from the coastwestward to the Andean foothills, in suitablehabitats at relatively low elevations, whereas Lorbignyianus would seem to be restricted to thehigher altitude plateaus of the central area. Theapparent broad geographical overlap one sees onpublished maps (e.g., Fjeldsa & Krabbe 1990)does not seem to be accompanied by an equiva-lent ecological overlap, and the two species mayin fact not breed side by side. In an earlier paper(Vuilleumier 1991b: 329) I classified the situation
about 30 km W of Gualjaina, 1 pair rumicivorusnear Puerto Melo, 1 pair rumicivorus near CaboDos Bahías, 1 + 2 + 4 birds at 3 sites along route30, between Camarones and the intersection ofroute 30 with route 3.
These records, mapped on Fig. 13, suggest,first that Thinocorus is patchily distributed, andsecond that the two species do not overlap geo-graphically. Thinocorus orbignyianus seems con-fined to the central area of plateaus and highermountains (habitat similar to that in Fig. 17),
16

STEPPE BIRDS OF PATAGONIA
in Thinocorus as a zone of parapatry with anarrow but long overlap zone along the Andeanfoothills. There is appa~ntly no published infor-mation on the interactions of these closely re-lated species in areas of overlap or parapatry, andno statement based on actual field data can bemade at this time about reproductive isolatingmechanisms. One may speculate, however, thatthe color differences in male plumage, especiallythe presence (T rumicivorus) or absence (T orbig-nyianus) of a black bar between the throat andthe breast, combined with overall size differencesand differences in vocalizations, act, togetherwith the habitat differences mentioned above, asisolating mechanisms. A field study of thesemechanisms would be very rewarding.
discuss the distribution of these two speciesalong the transect to illustrate differences in habi-tat preference and patchiness.
Geositta rufipennis was observed only once, 2birds (paired?) in a scree of huge boulders with al-most no vegetation along route 12 at 530 m inthe Chubut River Valley near Piedra Paradaabout 75 km W of Paso del Sapo (Fig. 15, Fig.16, top). Peters (1923: 312) collected the speciesin rocky habitats near Huanuluan and Maquin-chao (Rio Negro). Wetmore (1926a: 438) re-ported G. rufipennis from Rio Negro (ArroyoCumallo) and Chubut (Maitén). In his 1926bpaper, Wetmore saw G. rufipennis "among lowbrush on rocky slopes" near Mendoza. Olrog(1979: 166) stated that G. rufipennis occurred"possibly" in Chubut and Santa Cruz.
Geositta cunicularia was noted on 9 occa-sions, 6 of them in Península Valdés, where thespecies was locally abundant, especially in thePunta Delgada area, in very open, low grassy andscrubby steppes on sandy and dune-like substrates(Fig. 16, bottom). I did not see G. cuniculariaat the base of the Península in denser and tallershrubsteppe with no or very little grass. In spite
Geositta (Furnariidae)The two species of Geositta (rufipennis and cuni-cularia) encountered along the transect are notvery closely related to each other (Vuilleumier1967, Vaurie 1980). G. rufipennis seems to be iso-lated within the genus and G. cunicularia seemsto be related to high Andean G. punensis andsouthern Patagonian G. antarctica. I nevertheless
FIG. 15. DistributioQ of Geositta cunicularia (black dots) and G. rufipennis (black triangles) along the transectroute; additionallocalities in Río Negro for these two species from the literature and unpublished personal obser-vations (see text).
17

VUILLEUMIER
FIG. 17. Habitat of Geositta cunicularia ("wittu" vocal-ization type) about 30 km east of Gastre, altitude about1300 m, Chubut; the birds occurred in the very opengrassy area at the edge of the smalllagoon in the middleground. Photo F. Vuilleumier, November 1991.
FIG. 16. Top: area of sympatry between Geosittacunicularia (habitat: open riverside vegetation in leftmiddle ground) and G. rufipennis (habitat: rockyscree in foreground) along the Chubut river near PiedraParada, Chubut. Bottom: shrubsteppe with low grass,habitat of G. cunicularia a few km west of Punta Del-gada, Chubut. Photos F. Vuilleumier, November1991.
of active search, I did not encounter G. cunicula-ria along the rest of the transect, with three ex-ceptions: (1) 2 birds at about 1300 m, about 30km E of Gastre, in an open, sandy valley withlow bushy steppe on hills and grassy slopes (Fig.17); these birds emitted the "wittu-wittu-wittu"vocalization characteristically heard in Tierra delFuego; (2) 1 bird in a damp, grassy area near asmall pond and open and dry steppe near PiedraParada at 530 m (bird not heard); (3) 1-2 birds("wittu" call type) in a damp meadow along theedge of an artificial pond about 20 km SE ofColan Conhué at about 800 m.
My observations strongly suggest that G.cunicularia is patchily distributed and is absentfrom large areas of north-central Patagonia cov-ered with pure shrubsteppe, whether at relativelylow altitudes near the coast or higher up on thebasaltic plateaus, but that it occurs only in areaswith very open, grazed, grassy steppe on softsandy soil (perhaps only in areas with Ctenomyscolonies, as in Tierra del Fuego). Fig. 15 illustra-tes this distribution. Furthermore, it seems thattwo populations, with two distinguishable songtypes, occur in the transect area: the trill (fol-lowed by repeated notes) in the Península Valdés,and the "wittu" vocali7,ation locally inland.
It is of interest to point out that Durnford(1878) did not find Geositta cunicularia to becommon in the Chubut Valley, but that Peters(1923: 312) found that species "very common" inRío Negro. (Unfortunately, Peters did not giveany locality data for G. cunicularia.) Of theseveral specimens cited by Wetmore (1926a: 438),only the one from Guaguel Niyeu, Río Negro,November 14, 1911 is mapped on Fig. 15. Theother birds are from late summer or winter andcould be migrants from elsewhere. Wetmore(1926b: 244) stated that "near Zapala [Neuquén]the miner frequented sandy areas along theslopes of little valleys."
Habitat co-occupancy was not noted betweenthe two species of Geositta, which live in very
18

STEPPE BIRDS OF PATAGONIA
different environments (see also Peters 1923: 312,who stated that G. .rufipennis "is invariablyfound in rocky situations, whereas G. c. cunicula-ria frequents the dry, sandy plains"). Records ofG. rufipennis and G. cunicularia from the Barilo-che area on Fig. 15 represent my own field obser-vations made in 1965. I observed rufipennis inrocky areas and cunicularia in open steppes.
In southern Patagonia and Tierra del Fuegowhere Geositta cunicularia and G. antarcticaoverlap and even breed side by side locally, I haveobtained no evidence of interbreeding. G. cuni-cularia and G. antarctica are very similar to eachother morphologically but differ in voice. Onecan thus suppose that reproductive isolation isprimarily achieved through differences in vocal-izations. Because of substantial differences inmorphology, reproductive isolation betweenGeositta cunicularia and G. rufipennis is morelikely to be due to their external appearance thantheir vocalizations, if one reasons by analogywith the situation between similar-looking G.cunicularia and G. antarctica, although thevoices of cunicularia and rufipennis are quite
different. Overlaps between species of the genusGeositta, irrespective of the taxonomic rela-tionship of these species, have not been studiedin detail in the field and are worth investigating.The two Geositta species encountered along thetransect are largely segregated by habitat, whereasthe two species in Tierra qel Fuego coexist in thesame habitat. These two pairs of species repre-sent the extremes observed in the genus: mostother species pairs found together or near eachother exhibit at least some difference in habitat
preference.
Upucerthia and Eremobius (Furnariidae)Three species of earthcreepers were observedalong the transect, Upucerthia ruficauda, U du-metaria, and Eremobius phoenicurus. The twospecies of Upucerthia are not each others' closestrelatives (Vaurie 1980). The monotypic Eremobi-us, although very similar morphologically tosome species of Upucerthia, like ruficauda andandaecola, does differ from them in its nest siteand nest structure. As in the case of Geosittaabove, overlaps in this group of furnariids are dis-
FIG. 18. Distribution of Eremobius phoenicurus (black dots), Upucerthia dumetaria (black triangles), and U rufi.cauda (star) along the transect route; additionallocalities in Río Negro for E. phoenicurus and U dumetaria fromthe literature and unpublished personal observations (see text).
19

STEPPE BIRDS OF PATAGONIA
(I.
FIG. 20. Nesting hole of Upucerthia dumetaria a fewkm west of Punta Delgada, Chubut (see Fig. 19, bot-tom for site location). Photo F. Vuilleumier, November1991.
FIG. 19. Two views of the habitat of Upucerthia dume-taria. Top: open steppe on rocky (basaltic) ground afew km west of Telsen, Chubut; birds were displayingfrom the top of the shrubs in the left background.Bottom: unused roadside gravel pit a few km west ofPunta Delgada, Chubut; a pair was breeding in a hole(arrow) near the top of the cut just below the shruboff the center of the picture (see Fig. 20)- Photos F.Vuilleumier, November 1991.
the bill, then the bird flew to the top of a rockto finish the kill and start eating his prize, beforedisappearing out of sight. Neither Peters (1923)nor Wetmore (1926a, 1926b) mentioned Upucer-thia ruficauda from northern Patagonia. Olrog(1979: 169) gave its distribution and habitat asfollows: ..Arenales pedregrosos entre 3500 y 4000m de altura en los cerros de Jujuy, Salta, Cata-marca y Tucumán y después por el oeste sucesiva-mente más bajo, hasta el sur de Chubut."
Of the two other species, Upucerthia dumeta-ria was observed regularly from the PenínsulaValdés (where two different pairs were feedingyoung in nests in holes in road-side ditches on 8and 9 November; Fig. 19, bottom, and Fig. 20)all the way to the steppes west of Gualjaina. Itoccurred in a variety of habitats, including scrub-by, open steppes on basaltic plateaus (Fig. 19,top), fairly dense shrubsteppe on level, pebblyareas, low grass-scrub steppe in sandy areas, anddenser shrubsteppe on coastal plateaus. Peters(1923: 312-313) cited the species from San Anto-nio (..in the bushes growing close to the edge ofthe saltmarsh"), from west-central Río Negro(..on the plains, up the ravines and gullies, but al-ways among the bushes"), and from the LakeNahuel Huapí area (one record from near thebeach of the lake). Wetmore (1926a: 439) listedonly one breeding season specimen from west-central Río Negro (juvenile male). Wetmore(1926b: 249-250) found the species near General
cussed here because interspecific interactionsmight influence their distribution patterns.
Upucerthia ruficauda was seen only once,about 30 km W of Paso del Sapo in the ChubutRiver Valley at an elevation of 480 m (Fig. 18).The single bird was actively foraging for food atthe foot of a vertical, 50 m high cliff, at the edgeof the valley (Fig. 21, bottom). It searched forlarvae in the semi-soft ground, at the base ofsmall stones. One large prey item (probably abeetle larva) was killed with repeated blows of
20

STEPPE BIRDS OF PATAGONIA
FIG 21. Top: large stick nest of Eremobius phoenicurusin a low shrub, open shrubsteppe about 10 km west ofGan-Gan, Chubut (arrow points to 9 cm long Swissknife on top of nest). Bottom: habitat of Upucerthiaruficauda a few km west of Paso del Sapo, Chubut; thebird was foraging on the rocky ground at the bottomof the cliff among the sparse shrubs in the left fore-ground. Photos F. Vuilleumier, November 1991.
Roca (Rfo Negro) "among the heaviest growthsof Atriplex and other sbrubs in the lowland f1oodplain of the Rio Negro" and near Zapala (Neu-quén) "in heavy tracts of thorny brush in an ar-royo leading toward the lowlands." Wetmore(1926b: 250) aptly mentioned the "thrasherlike[Toxostorl1a, Mimidae] appearance of u: dumeta-ria in bill shape and habits.
Eremobius was observed in the same areas andhabitats as Upucerthia dumetaria, from PenfnsulaValdés to near Gastre (5 observations), from sealevel (bilt not in the open steppes on dunes andsandy soil near Punta Delgada) to about 1300 m(Fig. 18). Several nests were seen (Fig. 21, top),but those I opened up turned out to be inactive.Peters (1923: 314) called Eremobius phoenicurus"a common resident in arid northwestern Patago-nia" (Huanuluan/Maquinchao area). Wetmore(1926a: 440) mentioned two specimens from SanAntonio and Paja Alta (near Valcheta), both ineasternRfo Negro. Wetmore (1926b: 253) foundEremobius near Zapala {Neuquén) "amid patchesof low thorny brush that grew on the slopes ofrolling hills, where the soil was composed ofsand and stones." He compared the species to"long-tailed wrens" but found them more terres-trial.
My observations suggest that habitat sharingis common between two of the three earth-creepers (Upucerthia dumetaria and Eremobiusphoenicurus). Along the transect, these two spe-cies were characteristically found in similarshrubsteppes on open stony ground with little orno grass tussocks. Durnford (1878) called Upu-certhia dumetaria and Eremobius phoenicurus"common ...throughout our journey".
Sympatry and habitat co-occupancy amongthese two earthcreepers could be achieved be-cause of differences in size and in bill shape.Eremobius phoenicurus is relatively small andhas a long, thin, and slightly decurved bill. Upu-certhia dumetaria is larger and has a longer,thicker, and more decurved bill. Their nests alsodiffer. Eremobius makes large stick nests inshrubs (Fig. 21, top). On the other hand, Upu-certhia dumetaria makes its nest at the bottom ofholes in small cliffs or banks (Fig. 19, bottom;
Fig. 20).In the field, Eremobius phoenicurus and Upu-
certhia ruficauda appear quite similar in color,pattern, bill size and shapt; and in the way they
cock their tail up at an angle. In this case, repro-ductive isolation between these birds would beachieved chiefly through their substantial diffe-rences in habitat preference. Eremobius phoeni-curus and Upucerthia dumetaria, on the otherhand, are quite different from each other in bothmorphology and vocalization.
Leptasthenura (Furnariidae )The two species of Leptasthenura (L. platensis andL. aegithaloides) observed along the transect areprobably sister species (Wetmore 1926b: 256;
21

VUILLEUMIER
Fjeldsa 1991: 349) that have evolved from a com-mon ancestor, althou.gh the vicariance eventresponsible for this speciation event is unknown.Although largely allopatric, these two near-sib-ling species seem to overlap in steppes of north-eastern Patagonia and in parts of the Monte andEspinal yegetation types (Hueck & Seibert,1972) of eastern and central Argentina.
Leptasthenura platensis was identified on 7November at Riacho San José, base of PenínsulaValdés, on 8 November in Península Valdésabout 5 km W of Puerto Piramides, 11 Novem-ber in the Telsen area, and 14 November about5 km W ofPaso del Sapo (Figs. 22 and 23). L. pla-tensis was not recorded from Chubut by Durn-ford (1877), or from Río Negro by Peters (1923),Wetmore (1926a), or Bettinelli & Chebez (1986),or from Neuquén by Wetmore (1926b). Myobservations seem to extend the range of L.platensis to parts of Chubut, although Meyerde Schauensee (1982: 207) states "Argentinato Chubut," and Olrog(1963: 214 and 1979: 175)indicates "hasta el norte de Chubut." It is unfor-tunate that Fjeldsa & Krabbe (1990) did not
include L. platensis in their book. This species,which is very similar to L. aegithaloides, doesoccur in part of the area they cover. Users of thebook working in Patagonia will not be aware ofthe potential occurrence of L. platensis there.The distribution of Leptasthenura spp. may notbe as well known as previously believed. Forexample, I discovered a hitherto unknown breed-ing population of L. aegithaloides in ChileanTierra del Fuego (specimens collected in 1985and 1987).
L. aegithaloides was observed at two localitiesin Península Valdés ( about 20 km W of P. Pirámi-des; near Caleta Valdés), near Puerto Madryn,near Gan-Gan, a few kilometers west of Paso delSapo (nesting; parents were actively feeding nest-lings at a nest in a hole in a cliff, 14 November,see Fig. 24; a few km west of area with L. platen.sis), between Gualjaina and Esquel, betweenColan Conhué and Pampa de Agnia, and atCabo Dos Bahías. Peters (1923: 315, 316) foundL. aegithaloides a "common resident" and col-lected it at San Antonio, Huanuluan/Maquin-chao and Bariloche. Wetmore (1926a: 441) re-
22

"I STEPPE BIRDS OF PATAGONIA
FIG. 23. Dense shrubsteppe habitat of Leptasthenuraplatensis a few km east of Puerto Pirámides, Chubut.Photo F. Vuilleumier, November 1991.
ported t,?!O winter specimens from near Valcheta,Río Negro. In 1965, I found this species commonin a variety of brushy habitats near Bariloche
{Fig. 22).Wetmore {1926b: 256) encountered L. platen-
sis in La Pampa Province {Argentina) and Uru-guay "in trees of the densest foliage ...wherethey clambered like titmice in a leisurely mannerthrough the dense groWth of limbs." Of L.aegithaloides, Wetmore {1926b: 257) stated that itwas "distinguished from L. platensis by some-what more bushy crest, darker coloration [Chi-lean population], and grayish white on the innerwebs of the retrices." Patagonian populations,however, are paler and look more like L. platensisin color. In Río Negro, Wetmore {1926b: 257)found L. aegithaloides "in the tops of thickbushes in a region where the atriplex and othergroWth typical of alkaline flats was tall anddense." Wetmore {1926b: 256-257) gave thevoice of L. platensis as "a faint tsee-ee-ee" and thatof L. aegithaloides as "a low buzzing trill."
Fig. 22 shows that, according to my obser-vations, these two species of Leptasthenura arepotentially sympatric. Even though observednear each other in two areas {Península Valdésand near Paso del Sapo) I did not actually seethem side by side. Thus, I have no first-hand in-formation on interspecific interactions in sympa-try. In the future, specimens should be collectedat localities where these two species either occur
FIG. 24. Top: habitat of Leptasthenura aegithaloides afew km west of Paso del Sapo in the Río ChubutValley; birds were foraging among the shrubs in themiddle distance as well as at the foot of the tall cliff inthe left of the photograph. Bottom: breeding habitat ofL. aegithaloides a few km west of Paso del Sapo in theRío Chubut Valley; arrow indicates nest location inhole in cliff; the two parents were actively feeding nest-lings (nest was too high to check age and number ofnestlings). Photos F. Vuilleumier, November 1991.
together or near each other in order to fully doc-ument their occurrence.
If L. platensis and L. aegithaloides live to-gether in the same shrubsteppe habitat, study oftheir reproductive isolation should be extremelyinteresting because they are very similar to eachother morphologically and perhaps also vocally.Four specimens of L. aegithaloides (AMNH
23

to two different species groups (Vuilleumier1967), and are thus probably not very closelyrelated to each other (see also Vaurie 1980). In thefield, however, they look very similar and oftenbehave in similar fashion. It is thus reasonable toexpect that, if found together, they may interactand hence influence their respective distribu-tions.
Asthenes patagonica was seen at the base ofPenínsula Valdés (adult flushed from a nest thatI did not open, 7 November), Península Valdés(near Puerto Piramides), Sierra Chata, W ofSierra Chata (nest building), about 10 km W ofTelsen (inactive nest), and about 30 km W ofPaso del Sapo (Fig. 25). Peters (1923: 316) statedthat he "found [this species] only at San Antoniowhere a male and a female were taken on August18." He did not describe its habitat. Wetmore(1926a: 442) similarly mentioned two specimensfrom San Antonio without specifying their habi-tat. In his 1926b paper, Wetmore stated (p. 271)that A. patagonica "was found in the semiaridregion that bordered the Rio Negro, where it fre-quented the denser, taller stands of Atriplex andother shrubs that grew in the river bottom or
818454, 817073, 826140, 818455) that I collectedin Tierra del Fuego ¡n 1985 and 1987 weighrespectively 9.0,9.1,9.3, and 9.5 grams. Two spe-cimens of L. platensis (AMNH 779562, 779584)collected in Entre Rios weigh respectively 9.8and 10.4 grams. L. platensis may therefore be alittle heavier than L. aegithaloides. Plumage diffe-rences between these two speciesinclude the pres-ence of a crest in L. platensis (absent in aegithal-oides), less heavily streaked crown in platensis(heavily marked with buffy in aegithaloides),paler and buffier tail feathers in platensis (darkerand grayer in aegithaloides), and streaked throatin platensis (in aegithaloides the throat is eitherunstreaked or less streaked). These differences arefairly well-marked on study skins but much lessso when observing birds in the field. If vocaliza-tions are similar, then such morphological diffe-rences as presence!absence of a crest, poorlymarked versus well marked crown, and streakedversus unstreaked throat might be characters usedas isolating mechanisms.
Asthenes (Furnariidae)The two species of Asthenes observed along thetransect, patagonica and pyrrholeuca, may belong
24

STEPPE BIRDS OF PATAGONIA
,i
FIG. 26. Top: habitat of Asthenes pyrrholeuca in ariverine grove of willows a few km west of Paso delSapo along the Chubut River, Chubut. Bottom:habitat of Asthenes patagonica in dense shrubsteppe afew km east of Puerto Pirámides, Chubut; note largestick nest in shrub in center of photograph. Photos F.
Vuilleumier, November 1991.
the two species differ. The one nest of A. pyrrho-leuca I saw was in a 70 cm tall shrub; it was spher-ical, about 25 cm in diameter, with a side open-ing. For a description of a nest of A. pyrrholeucasee Wetmore (1926b: 270-271). I found severalnests of Asthenes patagonica, usually very visibleand voluminous, placed near the top of thornybushes (Fig. 26, bottom). They were elliptical, atleast about 45 cm long and 25 cm wide in thecenter with an entrance tunnel sometimes aslong as 15 to 20 cm or more. Wetmore (1926b:
occurred more sparingly in the smaller, morescattered growth that clothed the gravel hillsabove the flood plaiñ." Fig. 26 (bottom) illus-trates the habitat of A. patagonica.
Asthenes pyrrholeuca was observed at RiachoSan José (base of Península Valdés, nest withthree eggs, 7 November), about 60 km E of Tel-sen, near"Telsen, at two localities in the Paso delSapo area, and one locality between Gualjainaand Esquel (Fig. 25). Peters (1923: 317) wrotethat "this was the common spiney-tail of the
Huanul~an-Maquinchao region, always found inthe thicker and denser groWths of bushes." Wet-more (1926a: 442) listed two specimens fromwestern Chubut, "both in barred juvenal plum-agt; " from 4 February and 3 March. He (1926b:
270) stated that "in places, as near the coast atBahia Blanca, these birds were abundant andformed the dominant element among passerinespecies." A. pyrrholeuca is listed from Chipau-quil, Meseta de Somuncurá, by Bettinelli & Che-bez (1986).
Actual sympatry and habitat co-occupancywas detected between the two species of Asthenesat only one locality (about 30 km west ofPasodel Sapo). At this site, the vegetation ranged fromriverine willows along the Río Chubut to over-grazed meadows with scattered Berberis bushesand to open scrub at the foot of a cliff. Asthenespatagonica and A. pyrrholeuca both occurred inthe drier vegetation near the base of the cliff, butA. pyrrholeuca was also found in the Berberisbushes and the riverine willows (Fig. 26, top). Atthat sitt; A. pyrrholeuca thus occurred in abroader range of habitats, and may have pre-ferred denser vegetation types near the river.This difference may help explain why I did notobserve A. patagonica and A. pyrrholeuca to-gether more often. Olrog (1979: 179-180) statedthat A. pyrrholeuca occurred generally near watercourses, and that A. patagonica was found in aridscrub.
Morphologically A. patagonica is shortertailed and appears slightly plumper than A.pyrrholeuca. A. patagonica appears at times to bepale or more sandy in color, whereas A. pyrrho-leuca is more mouse-brown. In some individualsof A. pyrrholeuca the tail is very long. Both spe-cies emit trills. A. pyrrholeuca has characteristiccall notes that can be transcribed as" huit" or"twit," either isolated or repeated. The nests of
25

open grazed grassland and shrubbery near ba-saltic rocks (Fig. 28). The Bariloche records inFig. 27 are from observations I made in 1965.Wetmore (1926a: 446) reported birds from nearCholila (Chubut) and one 60 miles W of Val-cheta (Río Negro). Wetmore (1926b: 297) collect-ed A. montana near Mendoza "on the ground oron low bushes near streams." He also stated(1926b: 297) that A. montana "did not differ inactions from 5triata [ = microptera] or livida, but
[was] readily distinguished by the white in thetail."
Agriomi5 microptera was seen at the samelocality as A. montana, foraging on the groundin a patch of grassy steppe (Fig. 28), as well as attwo localities in Península Valdés in shrubbysteppe on sandy soil between Punta Delgada andCaleta Valdés (Fig. 29, top), near Puerto Madrynin shrubsteppe (foraging on the ground and on agravel road), and near Puerto Melo in scrubsimilar to that near Puerto Madryn. Displayflights were seen near Puerto Madryn and PuntaDelgada (Península Valdés). The displaying birdflies up to about 100 m above the ground, thenalternates between upward. flights with active
272) described a similarly-shaped nest of A. pata-
gomca.The differences between these two species of
Asthenes (morphology, voice, nest structure)could easily explain their reproductive isolation.In this pair of species, as well as in other sym-patric pajrs of Asthenes elsewhere, no study ofinterspecific interactions exists. Because thesespecies are common in northern Patagonia, andtheir nests are very easy to find, it would berelatively easy to investigate such interactionsthrough detailed field study.
Agriornis (Tyrannidae)Speciation in flycatchers of the genus Agriorniswas discussed previously (Vuilleumier 1971), butproblems of interspecific interactions or repro-ductive isolation were not treated in that paper,and sympatry was discussed in one paragraphonly. Three of the five species of Agriornis (mon-tana, microptera, and murina) were seen alongthe transect (Fig. 27).
Agriornis montana was observed only onceon 13 November at about 1300 m near Gastre ina valley with mixed vegetation including very
FIG. 27. Distribution of Agriornis microptera (black dots), A. murina (black triangles) and A. montana (stars)along the transect route; additionallocalities for A. murina and A. montana in R¡o Negro and Chubut from theliterature and unpublished personal observations (see text).
26

1
STEPPE BIRDS OF PATAGONIA
FIG. 28. Locality where Agriornis microptera and A.montana occurred side by side a few km west of Gastreat about 1300 m, Chubut; A. microptera fornged on theground in the low grassy steppe at the edge of the drypool in the background, whereas A. montana foragedfrom the dense shrubbery and perched on the basalticrocks in the foreground. Photos F. Vuilleumier, No-vember 1991.
FIG. 29. Top: shrubsteppe habitat of Agriornis microp-tera on sandy soil near Punta Delgada, Chubut. Bot-torn: shrubsteppe habitat of A. murina at the top ofcoastal cliffs near Caleta Valdés, Chubut. Note structu-ral sirnilarity of habitats of two species. Photos F. Vuil-leurnier, Novernber 1991.
microptera and A. murina together in PenínsulaValdés. A. microptera was seen in more openhabitat than A. murina. Near Gastre, A. microp.tera and A. montana were observed within 200 mof each other. A. microptera was foraging on theground in a grazed meadow, whereasA. montanaforaged in an area of boulders surrounded bythorny shrub at the edge of the meadow, and didnot f1y toward the meadow (Fig. 28). Study ofinteractions among Agriornis spp. would be veryinteresting but are likely to be difficult to carryout because of the relative shyness and scarcity ofthese tyrants.
wing flaps and downglides with closed wings, inrelatively rapid sequence. The performance ap-peared to be silent. Wetmore (1926a: 446) reportsthree specimens from near Valcheta, R¡o Negro,where A. murina was also collected. In his 1926bpaper (p. 296), Wetmore reported finding A. mi-croptera in R¡o Negro "in a region of arid gravelhills covered with an open growth of low brush."
Agriornis murina was seen only twice, onebird near Caleta Valdés in relatively dense coastalshrubsteppe on 9 November, and one bird atCabo Dos Bah¡as in similar, but more opencoastal steppe on 17 November (Fig. 27). In bothcases the birds disappeared after a short period ofobservation and could not be found again. Peters(1923: 320) collected one A. murina in coastalRío Negro on 18 August, but did not find it else-where; Wetmore (1926a: 447) reported two A.murina from coastal Río Negro, and one fromnear Valcheta, further inland in R¡o Negro, and(1926b: 301) other specimens from near GeneralRoca, R¡o Negro (Fig. 27). The first nest of thatspecies was found by Nores and Salvador (1990)in Catamarca, a locality extending the breedingrange by about 1200 km northward.
Although seen within a short distance ofeach other (a few km) I did not observe A.
27

VUILLEUMIE
tance (like the calls of Motacilla jlava) ["pe-chee"in Peters 1923: 323], but closer by, the vocaliza-tion is more complex in structure: "tsit-tsit-tsit-tsitsitsipirisih. " I found M. maculirostris in several
habitats, including low open shrubsteppe on flatstony ground, shrubsteppe of the high basalticplateaus (Fig. 34 bottom), in rocky screes at theedge of escarpments, in arid plains with verysparse vegetation, and in low scrub near thecoast. Wetmore (1926b: 306) stated that M. macu-lirostris "seen on their breeding grounds werefound among low bushes on sandy or gravellyhillsides." Fjeldsa & Krabbe (1990: 508) wrotethat M. maculirostris "has a broad habitat prefer-ence, but is always close to vertical rocks, banksor walls." However, in several areas along thetransect, I found M. maculirostris to be abundantin flat, uniform shrubsteppe with no rocks,banks or walls (e.g., between Telsen and Gan-Gan, see Fig. 32). After Zonotrichia capensis andLessonia rufa, M. maculirostris was probably thethird most common and widespread passerinespecies along the transect. Interestingly, Durn-ford (1878: 395) found M. maculirostris to be un-common, but Peters (1923: 323) qualified it as "afairly common resident in western Río Negro"
(Huanuluan area).
Muscisaxicola (Tyrannidae)Speciation patterns il\ the genus Muscisaxicolawere described in a previous publication (Vuil-leumier 1971). The two species observed alongthe transect, M capistrata and M. maculirostris,were placed in two different species groups inthat pap~r. Muscisaxicola capistrata was observedon three occasions: one pair near Gastre at about1300 m, an isolated bird between Paso del Sapoand Piedra Parada, and an isolated bird about 30km NE of Esquel (Fig. 30, and Fig. 31, top).Peters (1923: 323) found that species "ratheruncommon" in the Huanuluan area, but did notindicate its breeding habitat. Wetmore (1926a:450) cited a September specimen from the Val-cheta area, but did not mention the species in hisother paper (1926b).
By contrast, M. maculirostris was abundantalong the transect, from near Telsen all the wayto the Gualjaina area. Along the coast I did notobserve it on Península Valdés but found it nearPuerto Melo (Fig. 30). Locally it was common,and at any one locality, up to 3-6 birds could beeither seen or heard simultaneously during theirflight songs (well described by Peters 1925: 323).The flight song sounds like "pssip" from a dis-
FIG. 30. Distribution of Muscisaxicola maculirostris (black dots) and M. capistrata (black triangles) along thetransect route; additionallocalities in Río Negro from the literature (see text).

"I STEPPE BIRDS OF PATAGONIA
FIG. 31. Left: habitat of Muscisaxicola capistrata a few km west of Gastre at about 1300 m, Chubut; the birdsforaged in the open steppe area on sandy soil of the foreground. Right: dense shrubsteppe habitat of M. maculiro.stris a few km west of Telsen, Chubut. Photos F. Vuilleumier, November 1991.
FIG. 32. Two views of flat shrubsteppe where Muscisaxicola maculirostris occurred at high densities, betweenTelsen and Gan-Gan, Chubut. Photos F. Vuilleumier, November 1991.
29

VillLLEUMIER
the Telsen area, but not between Telsen and Gan-Gan. It was seen again W of Gan-Gan to theEsquel region, and in coastal areas near Camaro-nes. Habitats where I found M. patagonicusvaried from arid steppe with sparse and lowthorny shrubs growing on bare soil to moist,man-made oases with fairly dense groves of wil-lows growing along arroyos and the Río Chubut.
At the two localities where both M. triurusand M. patagonicus were observed together (Fig.34), individuals of the two species were seen afew meters from each other, singing from the topof adjacent shrubs. In one case, I even saw onebird of each species singing from the top of thesame shrub. I never saw individuals of M. triuruspay any attention to individuals of M. patagoni-CUS, no matter how close they were to each other.
Seen next to each other, the two species of Mi-mus look different. M. patagonicus appears some-what larger and coarser-plumaged, with a longerbill, whereas M. triurus appears more slender,with a smaller and thinner bill and a longer tail.Furthermore, the large areas of white in thewings and tail of M. triurus are quite conspic-uous, especially in flight, and also when the bird
In the Meseta de Somuncurá, Bettinelli &Chebez (1986) report&d M. capistrata at LagunaBlanca and M. maculirostris at Cerro CoronaChico. Because their observations are from thesecond half of February, it is possible that thesebirds were already migrants, rather than breed-
mg.
Mimus (Mimidae)Two species of mockingbirds occur in the tran-sect area, Mimus patagonicus and M. triurus. M.patagonicus is closely related to M. satuminus(map in Short 1975: 290), and M. triurus to M.dorsalis (Short 1975: 290-291). Mimus triuruswas observed at only two localities, one or twobirds about 15 km west of Puerto Madryn alongroute 4 on 5 and 10 November, and one individ-ual about 65 km west of Puerto Madryn alongroute 4 on 10 November (Fig. 33). Both observa-tions were made in shrubsteppe growing onrather flat ground. By contrast, Mimus patagoni-cus was observed at many localities along thetransect, from coastal areas in the Península Val-dés region (including: family with one fledglingon 7 November at Riacho San José; family withone adult and two fledglings on 9 November) to
FIG. 33. Distribution of Mimus patagonicus (black dots) and two localities of overlap between M. patagonicus andM. triurus (stars) along the transect route (see text).
30

STEPPE BIRDS OF PATAGONIA
is perched. The songs of these two species arequite similar, but that of M. triurus may be moremelodious and more.varied, with rich liquidtrills and gurgling sounds. At times, the song ofM. triurus reminded me of phrases of Alauda ar-vensis or of Acrocephalus schoenobaenus; at othertimes, it seemed that the bird imitated some ofthe trill~ emitted by Geositta cunicularia (in-terestingly, since there were none nearby). Wet-more (1926b: 351) indicated that M. triurusfrequently mimicked the Vermilion Flycatcher
(Pyrocephalus rubinus).Durnford (1877: 31) wrote that Mimus pata-
gonicus was "not uncommon," but did notmention M. triurus. Peters (1923: 328) did notmention M. triurus from Río Negro. He foundM. patagonicus "abundant" along the coast but"rather uncommon" in the Huanuluan-Maquin-chao area. Wetmore (1926a: 455) reported twospecimens of M. patagonicus from eastern RíoNegro, but did not list M. triurus. Wetmore(1926b:350-353) cited M. triurus "in fair num-bers" from General Roca (Río Negro) and M.patagonicus from General Roca (Río Negro),Challaco (Neuquén), but not Zapala (Neuquén)(Fig. 33). According to Wetmore (1926b: 351), M.triurus's song "strongly resembles that of Mimuspolyglottos, and is accompanied frequently byaerial gyrations, in which the birds spring intothe air and support themselves with slow beatsof the widely open wings, that, with the spreadtail, display their contrasted markings to the ut-most." Wetmore (1926b: 353) foundM. patagoni-cus "common in growths of atriplex (Atriplexlampa and A. crenatifolia), creosote bush, andgreasewoods on the flood plain of the Rio Negro,near General Roca, and proved to be a true desertform since it spread out through the arid, gravelhills north of the railroad, where water waswholly lacking." He described the song as "simi-lar to that of M. triurus, though the performerswere less flamboyant in actions during delivery."Boswall and Prytherch (1972) mention only M.patagonicus from Punta Tombo, coastal Chubut.
Gochfeld (1978) described the sympatry be-tween M. triurus and M. patagonicus, which hestudied in northeastern Chubut, eastern RíoNegro, and southern Buenos Aires Provinces.Gochfeld (1978: 64-65) concluded as follows:"At the Bahia Blanca study area there was no evi-dence that the territories of the two species over-
FIG. 34. Habitat where Mimus patagonicus and M.triurus were found sympatricly west of Sierra Chata,Chubut; the other locality where these two specieswere found together, near Puerto Madryn has verysimilar vegetation. Photo F. Vuilleumier, November1991.
lapped, but there was also no evidence of inter-specific aggression." And further: "The presentstudy has shown that where they are sympatricthe Patagonian and White-banded Mockingbirdshave different habitat preferences or tolerances.The former was found in both desert scrub andmonte, while the latter was essentially absentfrom desert scrub areas."
It thus appears that the two species of Mimuscoexist with overlapping territories in a rathernarrow zone of geographical overlap in north-central and northern Patagonia. Gochfeld's(1978) pioneering work deserves to be pursuedfurther.
Anthus (Motacillidae)Three species of pipits (Anthus furcatus, A. hell-mayri, and A. correndera) occur in the transectarea. Durnford (1877: 32) mentioned only A. cor-rendera, stating that it was "common throughoutthe valley [of the Río Chubut] and on the hillswhere there was any grass." Wetmore (1926a:456) only reported A. correndera, from easternRío Negro, but Bettinelli & Chebez (1986) citedA. hellmayri (but no other species) from theMeseta de Somuncurá. Wetmore (1926b: 360-361) did not cite A. furcatus from Río Negro, butfoundA. correndera at Zapala (Neuquén). He didnot cite A. hellmayri. Peters (1923: 329) stated:"Two species of pipits very closely resemblingeach other in the field, occur in Argentina;
31

VUILLEUMIER
that in Buenos Aires Province, A. furcatus was"common over rolling, open country coveredwith low tufts of grass."
Near Las Salinas, several pairs of pipits wereseen in grassy meadows and very low scrub. Allof them were Anthus correndera. I heard no flightsongs; calls I heard can be transcribed as "tziht,""twit," or "tziht."
My observations thus add little informationabout the distribution and relationships of pipitsin north-central Patagonia, other than to suggestthat these birds may be patchily distributed andlocalized, and that they may overlap only little.Wetmore (1926b: 361), however, stated thatAnthus furcatus "was locally common in some ofthe areas visited, and though closely similar to A.c. correndera, with which it was often associated,was readily told by its grayer, less distinctlystreaked dorsal surface, and by the fact that inwalking it did not tilt the tail." In Uruguay,Wetmore (1926b: 361) found A. furcatus andA. correndera breeding side by side (specimenscollected). The latest published color plates ofpipits (by Fjeldsa in Fjeldsa & Krabbe 1990; andby Tudor in Ridgely & Tudor 1989) are unfortu-
nearly all field-observers confuse the two and Imust plead guilty to ti4e same error." He collectedonly 2 Anthus furcatus but "over twenty" A. cor.rendera at Huanuluan. Peters (1923: 330) alsostated that "the possibility of the occurrence ofAnthus hellmayri dabbenei Hellmayr (Hornero,1921,2, p. 191. Río Traful, Gob. de Neuquén) inextreme western Río Negro should also be consi-dered."
I saw pipits at only two localities along thetransect (Fig. 35) at Punta Delgada near sea levelon Península Valdés on 8 and 9 November, andnear Las Salinas, about 90 km east of Esquelalong route 25, at 650 m on 15 November. Noother suitable-looking habitats yielded pipits,
very surprisingly.At Punta Delgada, pipits were found in
grassy steppes (Fig. 36) and were abundant in flatareas with a very sparse growth of short grass(Fig. 36, bottom). All the birds that I saw wellwere A. furcatus. Many birds emitted flight songsthat can be transcribed by" tzi-ti-tititiriri " and
sharp calls by "whitt" or "pitt," or else "dzipp"or "dzitt" (somewhat reminiscent of calls ofMotacilla Jlava). Wetmore (1926b: 361) found
"i).
FIG. 35. Distribution of Anthus furcatus (black dot) and A. carrendera (black triangle) along the transect route;additionallocalities in Río Negro from the literature (see text).
32

STEPPE BIRDS OF PATAGONIA
FIG. 36. Two views of habitat of Anthus furcatus nearPunta Delgada, Chubut. Top: dense grassland. Bottom:very open steppe with low and sparse grass cover. Notethat habitat of A. conmdera near Las Salinas (notillustrated) was very similar to the bottom photograph.Photos F. Vuilleumier, November 1991.
Phrygilus (Emberizidae)Wetmore (1926b: 405) has discussed the affinitieswithin Phrygilus. Phrygilus fruticeti and P. carbo-narius are probably closely related to each other.Both were seen along the transect (Fig. 39). Ofthe two, P. fruticeti was the more common andthe more widespread. In some areas it was abun-dant (between Puerto Madryn and Telsen alongroute 4, for instance). I did not find it in Penfn-sula Valdés. P. fruticeti was found in open steppeson pebbly or rocky ground, with sparse to rela-tively dense thorny shrubs. More densely vege-tated areas had more birds, reaching a density ofabout one singer per hectare. Phrygilus carbonari-
nately not sufficient for accurate field identifica-tion of these very difficult birds that look andsound very similar. R1rthermore, the texts inthese two books are not very informative either,insofar as field identification goes. Extremecaution is therefore advised for observers whowant to identify pipits in the field in Patagonia.What is needed to clarify problems of identifica-tion and hence distribution and speciation is aresearch program including the collection of ade-quate series of specimens, tape recordings ofvocalizations and playback experiments.
Sicalis (Emberizidae)In an earlier paper (Vuilleumier 1991b: 329), Isuggested that the allopatric distribution of S.lebruni (Patagonia) and S. olivascens (high Andesfurther north) could be due to competition withS. auriventris, which occurs in the interveningarea. I was not able to verify this statement dur-ing the transect in Chubut because I did not seeauriventris. However, I observed S. lebruni and S.luteola and found them to occupy very differenthabitats.
Sicalis lebruni was seen in the Punta Delgada-Caleta Valdés area of Península Valdés, where itis abundant and breeds in holes along road cuts(Fig. 37). S. luteola was observed in man-made oa-ses (Telsen, along Arroyo Telsen; and various 10-calities in the Chubut Valley, especially near l.osAltares), where it is abundant and occurs in tallgrass and dense riverine vegetation of weedyplants (Fig. 37). Fig. 38 illustrates the habitats ofS. lebruni and S. luteola along the transect. Wet-more (1926b: 403) described the habitat and be-havior of S. luteola, but not those of S. lebruni.
Peters (1923: 331-332) found 3 species ofSicalis in the Huanuluan-Maquinchao area: S.luteola, S. auriventris, and S. lebruni, althoughonly the last one was "fairly common". Wetmore(1926a: 458) cited specimens of S. luteola fromBariloche and of S. lebruni from near Lake N a-huel Huapí (western Río Negro), records whichsuggest some geographic, if not ecological over-lap, between these two species (Fig. 38).
The distribution of Sicalis lebruni and S.luteola in northern Patagonia appears to bepatchy, each species preferring a different habitat.In places where these habitats occur side by side,one should expect parapatry. It would be worth-while investigating such parapatric contacts inthe future.
33

VUILLEUMIER
FIG. 37. Distribution of Sicalis lebruni (black dots) and S. luteola (black triangles) along the transect route;additionallocalities in Río Negro from the literature (see text).
us was seen at only five localities: near PuertoMadryn, near Sierra Chata, near Telsen, near LasChapas, and near Uzcudún, always in habitatcooccupancy with P. fruticeti.
At two of these five localities I was able toobserve the interactions between P. fruticeti andP. carbonarius. At the first locality (near SierraChata, Fig. 40, top) I thought that they hadinterspecific territoriality, but at the second (nearTelsen, Fig. 40, bottom), I noted that these twospecies had overlapping territories, and that di-splaying males did not pay any attention to eachother. At these two localities, where shrubs wererelatively high and abundant, P. fruticeti out-numbered P. carbonarius about 75 to 1.
Although both P. fruticeti and P. carbonariushave similar display flights, they differ in somerespects. First, their vocalizations are distinct.The song of P. carbonarius is pipit-like, a buzzyseries of notes that can be transcribed as "wheezy-wheezy-wheezy-wheezy-wheezy," each sound aclearly demarcated two-syllable "whee-zee," re-peated quickly about 5-9 times during the para-chuting sequence, which lasts no more than 2-3seconds. There is much individual variation,however, and some birds have a more trill-like
FIG. 38. Top: shrubsteppe on dunes, habitat of Sicalislebruni at Punta Delgada, Chubut. Bottom: dense riv-erine weedy vegetation, habitat of S. luteola at ArroyoTelsen, a few km west of Telsen, Chubut. Photos F.Vuilleumier, November 1991.
34

STEPPE BIRDS OF PATAGONIA
FIG. 39. Distribution of Phrygilus fruticeti (black dots) and five localities of overlap between E fruticeti and Ecarbonarius (stars) along the transect route (see text).
song, "pzii-pzee-pzee-pzee-pzee." By contrast, P.fruticeti has a shorter, more nasal song, with ablackbird-quality to it. Wetmore (1926b: 406)wrote very aptly that "males were singing a songthat sounded like the effort of some icterid." Thesinger of P. carbonarius can fly up from a perchright within the vegetation, and return to a perchout of sight of the observer, whereas many P. fru-ticeti remain well in sight. The down flight in P.carbonarius is parachute-like, tail spread out,banking from side to side. P. fruticeti 's down-flight is more butterfly-like and less a simpleparachuting descent, as in P. carbonarius.
Phrygilus carbonarius was not mentioned byDurnford (1877, 1878), Peters (1923), or Betti-nelli & Chebez (1986). Wetmore (1926a: 460)cited only an immature specimen from NahuelNiyeu, eastern Río Negro (5 July 1911, a winterdate). Wetmore (1926b: 406-407) recorded P.fru-ticeti at Zapala (Neuquén) and General Roca(Río Negro), and P. carbonarius near BahíaBlanca (Buenos Aires) and Victorica (La Pampa).Hence the literature is of no help in providingdata on the details of the overlap between thesetwo species in Patagonia. Because these two spe-cies are easy to observe at the localities where I
35
FIG. 40. Two views oí shrubsteppes where Phrygilusfruticeti and R carbonarius were sympatric. Top: nearSierra Chata, Chubut. Bottom: west oí Telsen, Chu-but. Photos F. Vuilleumier, November 1991.

JILLEUMIER
saw them, it should be possible to quantify theirhabitat preferences, fe.eding habits, breeding be-havior, and interspecific interactions. The visibi-lity of males during their display flights shouldenable one to map their territories precisely andhence to ascertain whether their territories doindeed o.verlap, as my observations near Telsen
seem to suggest.
altitudinal dimension present in the Andes far-ther west, species replacements and/or overlapstake place largely latitudinally, rather than altitu-dinally. Hence Cody's (1970) conclusions maynot apply to the area I visited.
The data presented in this report show twoclasses of distribution patterns. In some cases thespecies pairs or triplets studied were largely allo-patric, whereas in other cases I found evidence ofsympatry or parapatry. Superimposed on thispattern of geographic distribution of non-over-lap versus overlap are patterns of similarities ordifferences of habitat preferences.
Allopatric situations. Allopatric is used here asdefined by Futuyma (1986: 550): "Of a popula-tion or species, occupying a geographic regiondifferent from that of another population or spe-cies." Well marked differences in habitat preferen-ces between pairs of allopatric congeners werenoted in Sicalis. S. lebruni was found in opengrassy or scrubby coastal steppes on sandy soil.S. luteola occurred inland in dense, varied vegeta-tion of grasses and weeds along riverbanks. Thetwo habitats are geographically separated, henceit is not surprising that these two species wouldbe disjunct (see Figs. 37 and 38). However, Pe-ters' (1923) collection of one S. luteola and ofseveral S. lebruni at Huanuluan (see Fig. 37)suggests sympatry. It would be interesting toknow in what habitats Peters (1923) found thesebirds. In situations where these two species occurrelatively near each other, then the habitat diffe-rences they show should minimize interactionsbetween them. Furthermore reproductive isola-tion could be achieved through a combination ofother factors including size, color, nest locationand vocalizations (see Table 1).
DISCUSSION
The data presented in this paper are preliminaryin nature, since they are based on a survey of alarge area of north-central Patagonia. Never-theless, these observations allow me to makeseveral comments about patterns of habitat over-lap or non-overlap in situations of allopatry,sympatry, and parapatry, and make some specu-lations about factors of reproductive isolation inabout 30% of the breeding landbird fauna of
Patagonia.Before discussing these patterns, however, I
wish to point out that several other groups ofcongeners that I expected to study in the fieldwere not encountered, thus suggesting the possi-bility of either changes in distribution patternssince visits by earlier observers, or man-inducedmodifications of the habitats in the last 50-100years. I discuss these (apparent) absences in Ap-
pendix 1.Interactions of various sympatric congeners
have been discussed for the Chilean avifauna byCody (1970), who analyzed some of the groupsof ground or bush inhabiting birds reportedupon in this paper, including GeQsitta, Muscisaxi-cola, Sicalis, and Phrygilus. Cody (1970) studiedespecially altitudinal replacements in the Andes.In Patagonian steppes, however, which lack the
TABLE 1. Differences and similarities between allopatric and sympatric species pairs or triplets in Chubut.
black lineon breast
no line onbreast
open steppe,often sandy plains
open steppe,often valley slopes
scrnpe in
groundscrnpe m
ground
36

STEPPE BIRDS OF PATAGONIA
larger thancunicularia
smaller than
rufipennis
in hole orcrevice in rocks
in hole inbanks
Geositta grassy steppe
Upucertbia dumetaria in holein banks
in hole orcrevice inrocks
stick nestin shrubs
trills shrubsteppes
Upucerthia trills shrubsteppesruficauda
E",mobius phoenicurus
la~er than scallopedruficauda& breastEremobius
smaller than streakeddumetari4, same breastas E",mobius
smaller than streakeddumetari4, same breastas ruficauda
trills shrubsteppes
Leptasthenura platensis similar toaegitbaloides
slightlypaler than
aegithaloides
slightly morestreaked than
platensis
in tree holesor abandonednests of otherfurnariids
often in holein cliff orroad cut
trills monte and
shrubsteppes
Leptasthenura aegithaloides similar to
platensis
trills shrubsteppes andsemideserts
very similarto murina
very similarto microptera
Agriornis often inshrubs
in shrubs
mi=ptera
Agriornis mur¡na
Muscisaxicola capistrata la~er thanmaculirostris
Muscisaxicola maculirostris smaller than
capistrata
dark facial
pattern
very plain
often in rodentburrows
under rocks,in crevices
jruticetiPhrygilus
Phrygilus carbonarius smaller than
fruticeti
very similar in shrubsto carbonarius,wing barswell marked
very similar to in shrubs
fruticeti, wingbars less marked
wheezy trill shrubsteppe
37

VUILLEUMIER
Three kinds of situations were detected alongthe transect in Chubut: (1) sympatry and habitatoverlap, (2) sympatry with little or non-overlapof habitats, and (3) parapatry with similar habi-tat preferences for the two species in the pair.
(1) Truly sympatric species pairs, where indi-viduals of each species were observed in closeproximity of each other in the same habitat,were found in Upucerthia-Eremobius, Asthenes,Mimus, and Phrygilus. In these five genera I wasable to study instances of habitat overlap, withsinging or displaying birds of each species onlya few meters from each other.
Upucerthia dumetaria and Eremobius phoeni-curus were observed side by side in three areasalong the transect (Fig. 18). In each they oc-curred in similar kind of shrubsteppes (Fig. 19,top; Fig. 21, top). Differences between these twospecies that could minimize their reproductiveinteractions are given in Table 1.
Sympatry or near sympatry was observed inAsthenes in three regions along the transect (Fig.25). A. patagonica and A. pyrrholeuca differ inseveral characters including morphology andnest structure (Table 1). Only in the Paso delSapo area, however, were the two species seenside by side. They were found in dry thornyshrubs on the floor of the Chubut river valley.In Asthenes, color and nest structure (Table 1)may be factors of reproductive isolation. In Mi-mus and Phrygilus singing and displaying indivi-duals of each species in the pair were observed atlocalities with open shrubsteppe (Figs. 34 and40). Differences in color, size, and vocalizationspresumably keep the two species of each pair
separate (Table 1).(2) Sympatry with little or non-overlap of
habitat was observed in Geositta and Agriornis.In Geositta the two species differ in size andcolor (Table 1). Although the vocalizations ofboth species include trills (Table 1), the soundsemitted are quite different. I found an area nearPiedra Parada (see Fig. 16) where both specieswere neighbors and lived within about 500 m ofeach other but in very different habitats. G. cuni-cularia occurred in open, flat, grassy steppes withor without shrub cover along the bank of theR¡o Chubut. G. rufipennis foraged in rockyscrees with sparse shrubs above the valley floor.
In Agriornis I observed A. microptera and A.montana at the same site near Gastre (Figs. 28
More subtle differences in habitat preferenceswere noted in four other allopatric species pairs.In the tinamous Eudromia and Tinamotis, nooverlap was detected. Eudromia seemed to preferdenser shrubsteppes at lower elevations thanTinamotis. Table 1 lists other differences be-tween these two species which could reducereproductive interactions, should these birdsoccur. in contact elsewhere. In 7hinocorus andMuscisaxicola one species (I' orbignyianus andM. capistrata, respectively) was found in higherelevatio:n steppes of valley slopes. The othermember of the species pair (I' rumicivorus andM. maculirostris, respectively) occurred in flat,lower elevation steppes. In Anthus both specieswere observed in grassy and scrubby steppes.One of them, A. furcatus, was found in coastalareas, whereas A. correndera was found in theinterior. Although coastal and interior grassysteppes looked very similar to me, further studyof these habitats is likely to reveal differencesthat might help explain better the distribution ofthese pipits.
Note that, even though I did not observeparapatry or sympatry between species of tina-mous, 7hinocorus, Muscisaxicola, and Anthus,such contacts can be expected, especially perhapsin areas other than the transect in Chubut. Theinterspecific interactions that might occur insuch cases of contact are unknown. One mightspeculate that differences in color and crest intinamous, and in size and male breast pattern(Table 1) between 7hinocorus orbignyianus andI' rumicivorus, could serve as isolating mecha-nisms. Size and color pattern differences andslight differences in display flights (Table 1)could isolate Muscisaxicola capistrata and M. ma-culirostris. Differences in morphology and be-havior between Anthus furcatus and A. corren-dera seem poorly marked (Table 1), and it wouldbe of great interest to study an area of actual con-tact between them.
Sympatric or parapatric situations. Sympatric orparapatric situations were observed in severalgenera. Sympatric is used here as defined byFutuyma (1986: 556): "Of two species or popula-tions, occupying the same geographic locality sothat the opportunity to interbreed is presented."Futuyma (1986: 554) defines parapatric as "Popu-lations that haye contiguous but non-over-lap-ping geographic distributions."

STEPPE BIRDS OF PATAGONIA
and 29). A. microptera foraged by running on theground in grassy steppe, whereas A. montanaforaged by flying down from perches on thornyshrubs and basaltic rocks to catch invertebrateprey on the ground. Further differences betweenthese two species include tail pattern and nest 10-cation (Table 1).
In both Geositta and Agriomis a combinationof habitat and other characters (listed in Table 1)presumably prevent reproductive interactionsbetween the two species in each pair.
(3) Parapatry with little habitat differentia-tion was found in Leptasthenura and Agriomis.Leptasthenura platensis and L. aegithaloides wereobserved at sites only a few km from each otherin the Península Valdés area and near Paso delSapo (Figs. 23 and 24). Both species were seen insimilarly structured shrubsteppe so that contactbetween them could be expected. Similarities anddifferences that could minimize reproductive in-teractions are listed in Table 1.
In Agriomis, parapatry with little habitatdifferentiation between A. microptera and A.murina was noted in two coastal areas (Figs. 27and 29). The main difference that probably mini-mizes reproductive interactions between thesetwo species is size (Table 1). I observed displayflights of only A. microptera and hence do notknow what behavioral difference, if any, couldkeep these two species apart.
(2) Is there spatial segregation and exclusion,or is there habitat overlap, between Patagonianspecies of Anthus ? How different are flight songsand vocalizations among species of Anthus ?What are their exact breeding habitat require-ments? Do they have overlapping territories?
(3) Is there habitat overlap between Leptasthen-ura aegithaloides and L. platensis, or is therespatial exclusion? What specific habitat preferen-ces do these two very similar sister species have?Do they have any interspecific interactions? Arethere differences in vocalizations?
(4) What behavioral or other mechanismskeep Mimus triurus and M. patagonicus, and Phry-gilus fruticeti and P. carbonarius apart in theirareas of habitat overlap? Do they always haveoverlapping territories or do they show occa-sional interspecific territoriality?
ACKNOWLEDGEMENTS
I thank the Leonard C. Sanford Fund for contin-uing financial support of my research on the evo-lution of Patagonian birds. I am grateful to Os-valdo A. Reig for advice about localities to visitin Chubut, to Paul R. Sweet for assistance in thepreparation of this trip and the preparation ofthis paper, and to Helen Kwon and Elizabeth DeJesus for typing the manuscript. I thank MichaelGochfeld for information on some of the birdsdiscussed in this paper. The illustrations wereprepared by Joel A. Pollick. I thank Dennis Fin-nin, Jackie Beckett and Craig Chesek for help inpreparing the photographs.
Prospects for further workClearly the ecological and evolutionary study ofPatagonian birds in allopatric, sympatric andparapatric situations is a field that is wide openfor further exploration. Two major questionsare: (1) How can so many similar-looking conge-ners or closely related species of birds coexist insuch a structurally simple and uniformly distri-buted landscape type as the shrubsteppes of Pata-gonia? and (2) How did these distribution pat-terns originate? In order to approach these gene-ral problems, answers to more specific questionsare needed. A sample of such questions includes:
(1) Is there spatial segregation and exclusionbetween Patagonian Eudromia elegans and Tinam-otis ingoufi, or between Thinocorus rumicivorusand L orbignyianus ? To what extent are thesetwo species pairs mutually exclusive in theirdistribution?
REFERENCES
Bettinelli, M. D., & j. C. Chebez. 1986. Notas sobreaves de la Meseta de Sornuncurá, Rio Negro, Argen-tina. Hornero 12: 230-234.
Blondel, j., & j.-P. Choisy. 1983. Biogéographie despeuplernents d'oiseaux a différentes échelles deperception: de la théorie a la pratique. Acta Oecolo-gica, Oecol. Gener. 4: 89-110.
Boswall, j., & R.j. Prytherch. 1972. Sorne notes on thebirds of Point Tornbo, Argentina. Bull. BritishOrnith. Club 92: 118-129.
Cody, M.L. 1970. Chilean bird distribution. Ecology51: 455-464.
Durnford, H. 1877. Notes on sorne birds observed inthe Chuput Valley, Patagonia, and in the neighbor-ing district. Ibis, fourth series, vol. I: 27-46.
Durnford, H. 1878. Notes on the birds of central Pata-gonia. Ibis, fourth series, vol. II: 389-406.
39

VUILLEUMIE
Vaurie, C. 1980. Taxonomy and geographical distribu-tion of the Furnariidae (Aves, Passeriformes). Bull.Amer. Mus. Nat. Hist. 166: 1-357.
Vuilleumier, F. 1967a. Phyletic evolution in modernbirds of the Patagonian forests. Nature 215: 247-248.
Vuilleumier, F. 1967b. Mixed species flocks in Pata-gonian forests, with remarks on interspecies flockformation. Condor 69: 400-404.
Vuilleumier, F. 1971. Generic relationships and spe-ciation patterns in Ochthoeca, Myiotheretes, Xolmis,Neoxolmis, Agriornis, and Muscisaxicola. Bull. Mus.Comp. Zool. 141: 181-232.
Vuilleumier, F. 1972. Bird species diversity in Pata-gonia (temperate South America). Amer. Nat. 106:266-271.
Vuilleumier, F. 1985. Forest birds of Patagonia: ecolo-gical geography, speciation, endemism, and faunalhistory. Pp. 255-304 in Buckley, P. A., M. S. Foster,E.S. Morton, R.S. Ridgely & F.G. Buckley (eds.).Neotropical Ornithology. Orn. Monogr. 36. Wash-ington D.C.
Vuilleumier, F. 1991a. A quantitative survey of specia-tion phenomena in Patagonian birds. Ornit. Neo-trop. 2: 5-28.
Vuilleumier, F. 1991b. Speciation in Patagonian birds.Acta XX Congr. Internat. Ornithol., pp. 327-332.
Vuilleumier, F., & J. Kikkawa. 1991. Reconstructingthe history of Nothofagus forest avifaunas. Acta XXCongr. Internat. Ornithol., pp. 578-586.
Wetmore, A. 1926a. Report on a collection of birdsmade by J.R. Pemberton in Patagonia. Univ. Cali-fornia Publ. Zool. 24: 395-474.
Wetmore, A. 1926b. Observations on the birds ofArgentina, Paraguay, Uruguay, and Chile. UnitedStates Nat. Mus. Bull. 133: 1-448.
Wiens, J.A., & J. T. Rotenberry. 1980. Bird commu-nity structure in cold shrub deserts: competitionor chaos? Acta XVII Congr. Internat. Ornithol.,pp. 1063-1070.
APPENDIX 1. Discussion of several genera expectedalong the transect (see Fig. 4) but not encountered.
TinamousBesides Eudromia elegans and Tinamotis ingoufi, twoother species of tinamou (Nothura darwinii and N.maculosa), occur in the transect area but were notencountered, in spite of diligent search. I mention themhere because of the possibility that they are much lesscommon today than they were over a hundred yearsago (unless I missed them because I looked for them inthe wrong habitat). Durnford (1878: 405-406) wroteof Nothura: "1 obtained specimens in the Chupat [ =
Chubut] valley, and saw it occasionally in the valleysof the Sengel [ = R¡o Senguerr] and Sengelen [ = R¡o
Fjeldsa, J. 1988. Status of birds of steppe habitats of theAndean zone and Pitagonia. ICBP Techn. Publ.No.7: 81-95.
Fjeldsa, J. 1991. Biogeographic patterns in birds of highAndean relict woodlands. Acta XX Congr. Inter-nat. Ornithol., pp. 342-353.
Fjeldsa, J., & N. Krabbe. 1990. Birds of the high Andes.A maBual to the birds of the temperate zone of theAndes and Patagonia, South America. Zool. Mus,Univ. Copenhagen, and Apollo Books, Svendburg,Denmark.
Futuyma, D.J. 1986. Evolutionary Biology. SecondEdition. Sunderland, Massachusetts.
Gochfeld, M. 1978. Ecological aspects of habitat selec-tion by two sympatric mockingbirds, Mimus spp.,in Patagonia. Ibis 120: 61-65.
Gochfeld, M. 1979. Interspecific territoriality in Red-breasted Meadowlarks and a method for estimatingthe mutuality of their participation. Behav. Ecol.Sociobiol. 5: 159-170.
Hueck, K., & P. Seibert. 1972. Vegetationskarte vonSüdamerika. Gustav Fischer Verlag, Stuttgart.
Maclean, G. L. 1969. A study of seedsnipe in southernSouth America. Living Bird 8: 33-80.
Meyer de Schauensee, R. 1982. A guide to the birds ofSouth America. 2nd rev. ed. Intercollegiate Press,
(no city given).Nores, M., & S.A. Salvador. 1990. Descripci6n del
nido y huevos del Gaucho chicho (Agriornismurina). Hornero 13: 161-162.
Olrog, C.C. 1963. Lista y distribuci6n de las avesArgentinas. Opera Lilloana 9: 1-377.
Olrog, C.C. 1979. Nueva lista de la avifauna Argen-tina. Opera Lilloana 27: 1-324.
Paynter, R.A., Jr. 1985. Ornithological gazetteer ofArgentina. Museum of Comparative Zoology, Har-vard University, Cambridge.
Peters, J.L. 1923. Notes on some summer birds ofnorthern Patagonia. Bull. Mus. Comp. Zool. 65:277-337.
Ridgely, R.S., & G. Tudor. 1989. The birds of SouthAmerica. Vol. I, The Oscine Passerines. Univ. ofTexas Press, Austin.
Root, R.B. 1967. The niche exploitation pattern of theBlue-gray Gnatcatcher. Ecol. Monogr. 37: 317-350.
Rotenberry, J. T., & J.A. Wiens. 1980. Habitat struc-ture, patchiness, and avian communities in NorthAmerican steppe vegetation: a multivariate analy-sis. Ecology 61: 1228-1250.
Short, L.L. 1968. Sympatry of Red-breasted Meadow-larks in Argentina, and the taxonomy of meadow-larks (Aves: Leistes, Pezites, and Sturnella). Amer.Mus. Novit. No.2349: 1-30.
Short, L.L. 1975. A zoogeographic analysis of theSouth American chaco avifauna. Bull. Amer. Mus.Nat. Hist. 154: 163-352.
40

STEPPE BIRDS OF PATAGONIA
Chico]." [For nomenclature of place names, see Paynter1985]. In his 1877 paperl. Durnford had written aboutNothura (pp. 45-46): .Not so common as Calodromas[ = Eudromia] elegans but occasionally seen amongst
the thick grass and rushes bordering the river. Not seenon the hills." The species mentioned by Durnford inhis 1877 paper was Nothura maculosa; in his 1878 paperNothura p&rdicaria. But in a footnote to the 1878 paper,the editors of Ibis stated that the birds collected byDurnford, which they had not seen, were N darwinii.Wetmore (1926b: 33-39) reported only N darwiniifrom Río Negro (in .alfalfa fields and other greengroWth near the river"). The habitat given by Olrog(1979: 18) for N maculosa is .estepas húmedas," butthat for N darwinii is .estepas áridas."
Cinclodes (Furnariidae)Not a single Cinclodes was seen during the transect. Toan ornithologist used to working in other parts of Pata-gonia, this is surprising. At Huanuluan, Río Negro,Peters (1923: 313-314) found Cinclodes fuscus between6 September and early November. AII birds wereprobably migrants. He collected or saw C. patagonicuson 27 November and 2 December. AII birds he collect-ed were immature (non-breeders?). Wetmore (1926a:439) reported C. fuscus only from Lago Futalaufquen(Andean wne west of Esquel; winter migrants) and C.patagonicus from Lago Futalaufquen (winter birds) andfrom Huanuluan (male and female, 7 December, i.e., inlate spring-early summer). Wetmore (1926a: 439) statedthat .the two [C. patagonicus] taken at Huanuluan aremarked as accompanied with a set of eggs.» Bettinelli& Chebez (1986) did not list Cinclodes spp. from theMeseta de Somuncurá. Wetmore (1926b) did not men-tion Cinclodes spp. from Patagonia, but C. fuscus andC. oustaleti from Mendoza. In 1965 I observed C. fuscusand C. patagonicus in the Bariloche-Lake NahuelHuapí area. The total absence of Cinclodes spp. alongthe transect is puzzling. Either these birds do not occurin the area I visited or, if they do, in such small num-bers as to have remained undetected. I suspect thatCinclodes spp. might be only migrants in the transectarea, and breed in the Andean region further west (seePeters 1923: 313-314, about C. fuscus and C. patagoni.cus), although since Wetmore (1926a: 439) reported twoindividuals of C. patagonicus from Huanuluan (RíoNegro) with eggs, breeding of some Cinclodes may beexpected locally. Further field work in the central-we-stern part of the transect area is necessary, especially athigh elevations, to determine the occurence and statusof Cinclodes spp.
Río Negro, to the north of the central part of mytransect area in Chubut. Wetmore (1926a: 442) reportedone specimen of A. modesta collected near Valcheta(eastern Río Negro) and two near Huanuluan, andwrote that Asthenes "modesta is stated to be of wide-spread range in Argentina, but must be rarer than onewould suppose from published statements, since Ifailed to secure it there during extended field work in1920 and 1921." On 13 November 1991 I searched anarea of softly rounded hills at about 1350 m nearGastre, with a very sparse cover of low shrubs, thatlooked suitable for A. modesta, but failed to find it.Durnford (1877, 1878) did not mention Asthenesmodesta at all. Bettinelli & Chebez (1986) reported A.modesta from the Somuncurá Plateau in Río Negro.Durnford (1877, 1878) stated that he saw Astheneshudsoni only twice (1878), but in his earlier paper(1877) found it "not uncommon ...in dry places inthe valley." Wetmore (1926b: 273-274) encounteredA. hudsoni in Buenos Aires Province but did not citeit from either Río Negro or Neuquén. The birds hesaw were in "a dense patch of thistles and other weedson the shore of the Laguna del Monte" and in "marshesgrown heavily with Juncus acutus, a sharply pointedrush that grew in clumps with little runways between."
Asthenes anthoides was not recorded by Peters(1923) from the central part of a similar transect in RíoNegro, but "only encountered near Lake NahuelHuapí," where I saw it at severallocalities in 1965 (seealso Wetmore 1926a). Durnford (1877, 1878) did notmention A. anthoides. I was on the lookout for A.anthoides in the transect area, and saw likely habitat (amixture of relatively dense, grassy and shrubby steppe)at severallocalities in the western part of the transect(e.g., near Gastre), but did not detect the species there.It is common in parts of southern Patagonia (NWTierra del Fuego, pers. obs.).
Anairetes (Tyrannidae)Neither Anairetes flavirostris nor Anairetes paruluswere seen or heard by me, in spite of my having dili-gen~ly looked for them. A. parulus was noted as rare byDurnford (1877,1878), and two specimens were listedby Wetmore (1926a: 452) from Paja Alta, 7 miles eastof Valcheta, eastern Río Negro. Peters (1923: 324-325)unfortunately did not give indications about the rela-tive abundance of Anairetes parulus in his study areas,but mentioned six specimens of A. Jlavirostris from SanAntonio (coastal Río Negro), stating that "this is thefurthest south that this species has been recorded, andalso the first record of its appearance on the coast." Bet-tinelli & Chebez (1986) reported A. parulus from theMeseta de Somuncurá area. The complete absence ofAnairetes from my transect area is hard to explain. Ihave observed Anairetes in other parts of Patagonia.For example, I found A. parulus to be fairly commonin 1965 around Bariloche in Río Negro.
Asthenes (Furnariidae)Neither Asthenes modesta, A. hudsoni, nor A. anthoideswere noted along the transect. Peters (1923) only col-lected five A. modesta and one A. hudsoni in severalmonths spent in the Maquinchao-Huanuluan region of
41