FIELD EVALUATION OF THE BENEFITS OF FISH OIL … · FIELD EVALUATION OF THE BENEFITS OF FISH OIL...
24
Established and supported under the Australian Government’s Cooperative FIELD EVALUATION OF THE BENEFITS OF FISH OIL DIETARY SUPPLEMENTATION TO MULTIPAROUS SOWS FED DURING LACTATION AND EARLY PREGNANCY ON FERTILITY 2F-109 Report prepared for the Co-operative Research Centre for an Internationally Competitive Pork Industry By Rob Smits PO Box 78 Rivalea Australia NSW 2646 May 2010
-
Upload
phungkhuong -
Category
Documents
-
view
217 -
download
0
Transcript of FIELD EVALUATION OF THE BENEFITS OF FISH OIL … · FIELD EVALUATION OF THE BENEFITS OF FISH OIL...

Established and supported under the Australian Government’s Cooperative
FIELD EVALUATION OF THE BENEFITS OF FISH
OIL DIETARY SUPPLEMENTATION TO
MULTIPAROUS SOWS FED DURING LACTATION
AND EARLY PREGNANCY ON FERTILITY
2F-109
Report prepared for the Co-operative Research Centre for an Internationally
Competitive Pork Industry
By
Rob Smits
PO Box 78 Rivalea Australia
NSW 2646
May 2010

i
Executive Summary
The aim of this study was to assess the effect on reproductive performance of
supplementing the diet with omega 3 fatty acids from fish oil when fed to sows either
before post-weaning oestrus, during early pregnancy or continuously from lactation
through to four weeks gestation. 1216 mixed parity sows (parity 1-7) were fed one of two
lactation diets: either unsupplemented (Control lactation) and formulated using wheat,
cereals and soybean and canola meals and using tallow as the main source of fat (68 g/kg)
or the same base diet supplemented with 3 g/kg of fish oil substituting v/v with tallow
(Omega 3 lactation). Following weaning, sows continued on their treatment diet until
oestrus. Of the sows weaned and displaying oestrus, 860 sows were allocated to a 2 x 2
factorial feeding regimen and after mating, were fed either one of two gestation diets:
either unsupplemented (Control gestation) and formulated using wheat and cereals and
tallow as the main fat source (10 g/kg) or the same base diet supplemented with 6 g/kg of
fish oil substituted for tallow. Subsequent reproductive performance for weaning to
oestrus, proportion of sows re-mated, farrowing rate and subsequent litter size born live
and total born were analysed with average lifetime litter size born and lactation length as
co-variates in Univariate GLM ANOVA. There was no significant treatment effect on the
number of sows weaned, number re-mated or weaning to oestrus interval (7.0±0.2 days).
Subsequent litter size was significantly higher (P<0.05) in sows fed omega 3 supplemented
diets during lactation and through early pregnancy compared to unsupplemented Controls
(12.6 vsvs. 11.7 total born). Farrowing rates were unaffected by dietary treatment
(83.1%). Within parity analysis resulted in a larger response to supplementation detected
in mature parities (parity 4-7 weaned). In conclusion, omega 3 fatty acids from fish oil
resulted in improved subsequent litter size in older sows. Dietary supplementation offers
producers a nutritional strategy to overcome declining productivity in sows increasing in
age.

ii
Table of Contents
Executive Summary ............................................................................................................................................. i
1. Introduction ................................................................................................................................................ 1
2. Methodology ............................................................................................................................................... 2
3. Outcomes .................................................................................................................................................... 5
4. Application of Research ......................................................................................................................... 14
5. Conclusion ................................................................................................................................................. 14
6. Limitations/Risks ..................................................................................................................................... 14
7. Recommendations ................................................................................................................................... 15
8. References ................................................................................................................................................ 16
Appendices ......................................................................................................................................................... 21
Appendix 1: .................................................................................................................................................... 21

1
1. Introduction
One of the causes of high sow turnover in a breeding herd is low reproductive
performance. Over the years there have been several studies that have concluded
that reproductive failure and low fecundity contribute half of the sows culled in a
herd (Hughes and Varley, 2003). Engblom et al (2008) reported that culling for
low litter size was accentuated as parity increased in commercial herds.
Historically, the fertility of sows in commercial herds continued to increase with
age until parity 4 or 5, before declining with a productive life expectancy
averaging 8 or 9 litters (Levis, 2005, Tummaruk, et al., 2001). However, more
recent farm records show that reproductive performance is now peaking at
younger parities, before a rapid decline (Hughes and Varley, 2003, Rodriguez-Zas,
et al., 2003).
Supplementing the diets fed to sows during lactation with long-chain
polyunsaturated fatty acids (PUFA’s) from fish sources has been reported to
improve reproductive performance (Palmer, et al., 1970, Webel, et al., 2004).
Fish oil contains high levels of omega 3 eicosapentaenoic acid (EPA, C20:5) and
docosahexaenoic acid (DHA, C22:6), and dietary supplementation can increase the
tissue levels of these fatty acids in oocytes (Trujillo and Broughton, 1995,
Wakefield, et al., 2008) and embryos (Brazle, et al., 2009). PUFA’s have an
important role in regulating reproduction through the eicosanoid pathway which
synthesises prostaglandins (Caughey, et al., 2005). Altering the ratio of omega 6
to omega 3 fatty acids can alter the potency of prostaglandin activity (Innis, 2003,
James, et al., 2000), and this may be a mechanism for improved reproductive
outcomes when omega 3 sources are fed to animals (Staples, et al., 1998).
Previously we have shown that feeding a lactation diet supplemented with low
levels of fish oil (3 g/kg) increased subsequent litter size and this was
consequently related to improved embryo survival (Smits et al, unpublished,
Project 2F-102). The aim of this experiment was to investigate if there are
benefits to subsequent reproductive performance in a commercial herd when diets
are supplemented with omega 3 fatty acids from fish oil during lactation and post-
weaning, and if there are additional benefits when supplemented diets continue
to be fed in early pregnancy.
This experiment tested two hypotheses, firstly that offering sows a diet
supplemented with long-chain omega 3 fatty acids from fish oil during lactation
and post-weaning to oestrus would improve reproductive performance and

2
subsequent litter size. Secondly, fertility is enhanced when supplementation with
dietary omega-3 fatty acids is continued after mating during the period of embryo
implantation in early gestation.
2. Methodology
The experiment was conducted on a commercial piggery in NSW (Rivalea
Australia Pty Ltd., Corowa) during winter and spring. Over five weeks, 1216 Large
White x Landrace F1 sows (PrimeGro™ Genetics) ranging in age between parity 0
(pregnant gilts) and parity 6 prior to farrowing commenced the experiment when
they entered the farrowing shed in their 15th week of gestation. Sows were
allocated to either a treatment diet with no supplementary omega 3 fatty acids
from fish oil (Control) or a diet supplemented with 3 g/kg salmon oil (Omega 3).
Treatment diets were fed pre-farrowing, during lactation and between weaning to
mating. After weaning, 860 re-mated sows from the study group in lactation were
fed either an unsupplemented gestation diet or a diet supplemented with 6 g/kg
of fish oil in a 2 x 2 factorial design with lactation and gestation dietary treatment
as variables for the first 4 weeks of pregnancy. Thereafter all sows were fed a
standard diet during pregnancy. Subsequent pregnancy rates and litter size was
collected from farm records.
Animal management
The experiment was conducted according to Animal Ethics Committee approval
by the University of Adelaide (Project No. M-2009-147) and Rivalea Australia
Animal Ethics committee (Project No. 09R019C). Sows were sourced from a
commercial piggery at the Corowa site commencing in August. Each week
pregnant mixed-parity sows entered the farrowing sheds at 112 days of gestation.
On entry, sows were allocated to one of two lactation dietary treatments: either
Control or Omega 3 (3 g/kg fish oil), with one half of the shed allocated per
treatment per week. There were 608 sows that commenced the study as Controls
and 608 sows on the Omega 3 treatment at the start of lactation. To minimize
bias due to pen location within the shed, the treatment allocations on each half of
the shed were alternated between replicates. Data on previous litter size born
live and total born (including mummified foetuses and stillborns), and parity were
retrieved from farm records and current date of farrowing, litter size and pen
location were recorded. All sows were otherwise managed under commercial
practice, with litters being processed within 24 hours of birth; piglets injected
with 1 ml iron (Ferroject; Swift) and tails docked. Litter size was adjusted
according to rearing ability and ranged in number from 8 to 14. Some sows were
weaned early due to milking failure, sudden death or destruction due to injury. In

3
some cases, sows had an extended lactation if they were used as nursing sows to
maintain the litter. In these cases, the nursing sow was shifted to a pen and
continued on her respective treatment. Sows that had a shortened lactation (<10
days) due to milking failure or poor mothering ability, and nurse sows that had an
extended lactation (>32 days) were excluded from the analyzed data set. Sows
were weaned at 19.8±0.14 days (mean±SE) of lactation. Weaning occurred twice a
week. Of the total sows weaned, 398 sows on the Control treatment and 462 sows
on Omega 3 diets in lactation of the weaned sows were housed in individual stalls
in a study area of a commercial dry sow facility. Not all the sows commencing the
experimental diets in lactation were able to be retained for pregnancy treatment
allocation due to space limitations within the facility. There were more Omega 3
sows available to continue the experiment due to the commercial management of
the weaning, which was unintentional. The characteristics of the subset in terms
of litter size weaned, lactation length and parity were the same as the population
weaned. Within the rows of stalls filled following weaning each week, sows on
their respective lactation treatment were blocked together and continued to
receive their experimental lactation diet until mating.
Sows were mated at first oestrus detection by artificial insemination with
semen 3 x 109 sperm cells (Rivalea, Australia). Once mated, half the sows within
the weeks’ weaning block were allocated to a Control gestation diet and the other
half were allocated to an Omega 3 gestation diet with 6 g/kg fish oil. Sows were
remated the following day (+24 hours) and remained in the study area until
pregnancy was confirmed by ultrasound at 28 days. All sows ended their
experimental treatment diets at this point. Sows were then moved to an adjacent
dry sow shed and individually housed until entering the farrowing sheds on day 112
of gestation for the birth of their subsequent litter.
Diets and feeding
Diet composition of the Control and Omega 3 lactation diets and Control and
Omega 3 gestation diets are described in Table 1 with fatty acid profile described
in Table 2. Experimental lactation diets were formulated using wheat, cereals
and soybean and canola meals and provided 14.9 MJ DE/kg; 188 g crude
protein/kg; 0.51 g available lysine/MJ DE. Experimental gestation diets were
formulated using wheat and cereals and provided 12.9 MJ DE/kg; 144 g crude
protein/kg; 0.38 g available lysine/MJ DE. Other essential amino acids were
formulated as a ratio to lysine in all treatment diets. Omega 3 fatty acids were
supplied as fish oil in a liquid form containing antioxidants (Optigen Ingredients®,
Port Adelaide, Australia) and applied from a post-pellet spray applicator.
Supplementation with omega 3 fatty acids in the Omega 3 diets was achieved by

4
substituting tallow with fish oil. Other sources of unsaturated fatty acids were
kept to a minimum.
Following allocation and on the day of farrowing at the beginning of the
experiment, sows were fed 3 kg of their lactation treatment diet each morning.
From the day after farrowing, sows were offered their treatment diet up to three
times a day based on appetite. All sows were weaned from their litters and
relocated to individual gestation housing. Sows continued to be fed their
lactation diet after weaning once a day at 2.7 kg until oestrus and mating (pre-
mating period). After first service, mated sows were fed 2.5 kg of their treatment
gestation diet once a day for 28 days (early pregnancy period). For the remainder
of pregnancy, all sows were offered a commercial gestation diet fed once a day at
2.7 kg until transfer to the farrowing shed for their subsequent litter. The
commercial gestation diet used after four weeks of pregnancy was based on wheat
and cereals and contained no fish oil.
Treatment diets were analysed for fatty acids at the Nutrition and Functional
Foods Laboratory, The University of Adelaide, Waite Campus, South Australia.
Statistical analysis
Data was processed through a General Linear Model (GLM) analysis of variance
for treatments in a randomized design model assuming equal variance (Sokal and
Rohlf, 1981). The analysis of variance was performed using SPSS v18.0 (SPSS Inc.,
1989-2009). In the analysis of subsequent litter size, only those sows that
remained in the study area of the dry sow facility and were fed experimental
gestation diets were included. Any sow that returned to service and was re-mated
after the first post-weaning oestrus mating was included as a failed pregnancy in
the farrowing rate data analysis whilst the subsequent litter performance from
these returned and re-mated sows was excluded from further data analysis. The
subsequent litter data was normally distributed and was analysed by GLM
Univariate ANOVA with replicate and where stated parity were included as random
factors. Variables including lactation length, average lifetime litter size at the
start of the experiment included as covariates if significant in the model (P<0.10).
Comparisons between treatments were analyzed using Bonferroni adjustment to
confidence intervals for multiple comparison. The proportion of sows within the
study that were mated and farrowed (farrowing rate) were analyzed by Chi
square. All analyses were conducted at a confidence limit of 95%. Unless
otherwise stated, probability values >0.50 were described as not significant (NS)

5
3. Outcomes
Of the 1216 sows allocated to the study, 26 sows were taken off the study
during lactation due to (milking failure; 15 x Control, 11 Omega 3) and 42 for
extended lactations (nurse sows) (21 x Control, 21 x Omega 3). There were seven
sows removed due to mortality (3 x Control, 4 x Omega 3) during lactation. From
the production records, there was no evidence of any treatment effects on
lactation performance or sow mortality. The litter size weaned was unaffected by
dietary treatment and averaged 9.0±0.13. After weaning, there were six sows
removed due to sudden death or euthanasia (2 x Control, 4 x Omega 3). Omega 3
supplementation during lactation did not affect the proportion of sows weaned or
the resumption of oestrus and re-mated (Table 3). The interval to oestrus and re-
mating was similarly unaffected by Omega 3 supplementation during lactation and
post-weaning (7.0±0.2 days). The main reasons for failure to re-mate were due to
management decisions relating to physical issues (locomotion, udder, body
condition, infectious discharge: 13 x Control, 17 x Omega 3; χ2 0.51, P=0.477), or
mortality (sudden death & euthanasia: 2 x Control, 4 x Omega 3; χ2 0.65,
P=0.420). Out of all the sows weaned only three failed to resume oestrus after six
weeks (1 x Control, 2 x Omega 3, NS). Other reasons sows were removed after
weaning included culling for high stillbirths in the previous litter and for excessive
size.
Subsequent reproductive performance and litter size of the sows that remained
in the study area and continued their treatment diets for 28 days after mating is
summarized in Table 4. The subsequent farrowing rates were similar between
treatments (P=0.486). There were no significant differences in the cause of
mating failure between treatments; (return to oestrus=4.7%; pregnancy tested
negative=5.0%; death and euthanasia=4.1%; Late pregnancy failure (Not in Pig &
abortion) =2.3%; infectious discharge=1%).
Litter size born live was not significantly different between treatments (Table
4). Stillbirths tended to be higher in Omega 3 treatments that recorded higher
litter sizes. Subsequent litter size total born was significantly different between
treatments (P<0.05). Omega 3 sows fed supplemented diets from lactation
through to four weeks gestation (Omega 3/Omega 3) tended to have a higher total
litter size compared to the unsupplemented Controls (P=0.08). Subsequent total
born of sows fed Omega 3/Control and Control/Omega 3 were intermediate.
The average parity of the sows at weaning was 3.7±0.06 and was similarly
structured between treatments. Due to the small number of sows within each
parity, the data for subsequent litter size total born was analysed within parity

6
categories to reflect the subsequent litter size of weaned parity 1 sows, mid-
parity sows (parity 2 and 3 having their 3rd and 4th litter) and mature sows (parity
4-7 having their 5th-8th litter). There was no overall significant treatment x parity
interaction in subsequent litter size total born, however there was evidence of a
larger response in the mature sow parities allocated to Omega 3 when the dataset
was further separated into the parity categories defined above and analysed for
treatment response within each category. The subsequent litter size total born x
parity category is summarized in Table 5.
The subsequent litter size of parity 1 sows and parity 2-3 sows was unaffected
when fed with diets supplemented with fish oil during either lactation or early
pregnancy or both (Table 5). Subsequent sow farrowing rates for Control/Control,
Omega 3/Control, Control/Omega 3 and Omega 3/Omega 3 were also similar in
parity 1 (91.3%, 88.0%, 88.9%, 84.2%; NS) and parity 2-3 sows (82.2%, 88.0%,
77.9%, 87.9%; χ2 4.60, P=0.330).
In the mature parity category, there was a significant treatment response
(P<0.05) in subsequent litter size total born (Table 5). The increase in litter size
was greatest in the Omega 3 sows fed supplemented diets in lactation through to
four weeks gestation (Omega 3/Omega 3). Litter size of the Omega 3/Control
sows and Control/Omega 3 sows were intermediate but not significantly different
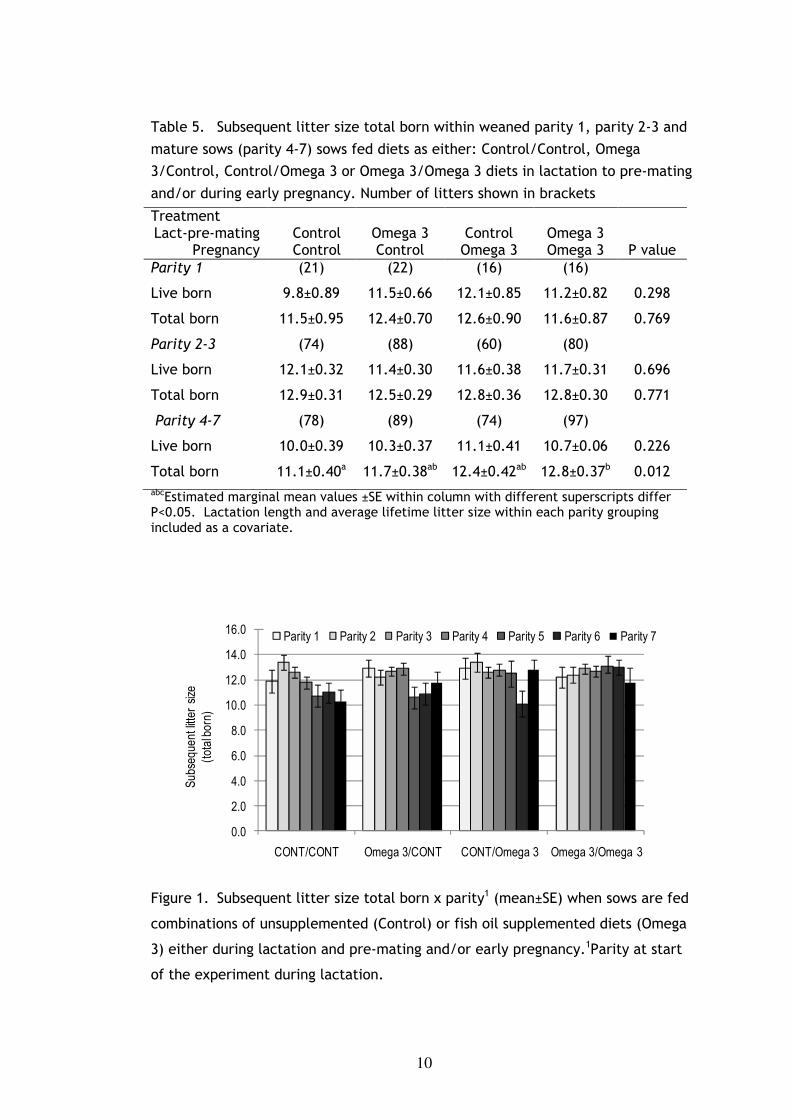
to the sows on Control/Control regimen. Figure 6.1 illustrates the pattern for
subsequent litter size total born as parity increases. In parity 4, 5 and 6,
subsequent litter size total born was significantly higher in Omega 3/Omega 3
sows compared to Control/Control and Omega 3/Control sows (P<0.05). Farrowing
rates were similar between Control/Control, Omega 3/Control, Control/Omega 3
and Omega 3/Omega 3 treatments in the mature parity category (82.1%, 82.4%,
77.9%, 81.5% respectively; NS).
7
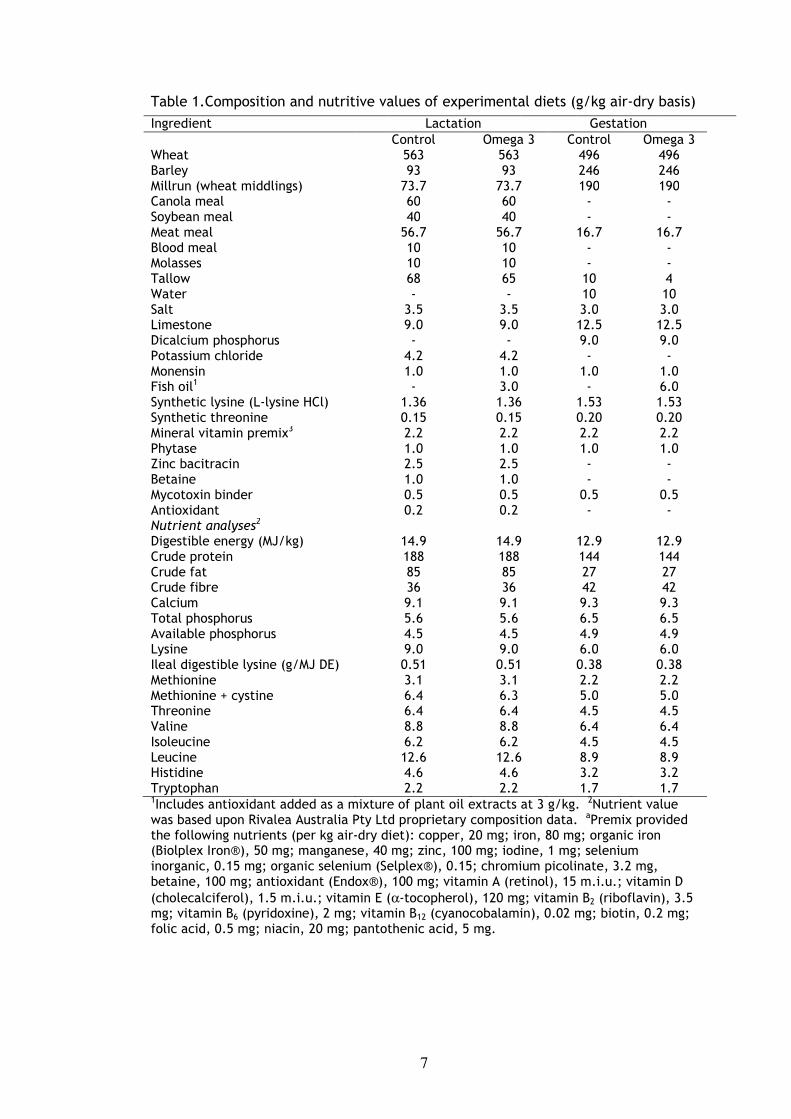
Table 1.Composition and nutritive values of experimental diets (g/kg air-dry basis)
Ingredient Lactation Gestation Wheat
Control 563
Omega 3 563
Control 496
Omega 3 496
Barley 93 93 246 246 Millrun (wheat middlings) 73.7 73.7 190 190 Canola meal 60 60 - - Soybean meal 40 40 - - Meat meal 56.7 56.7 16.7 16.7 Blood meal 10 10 - - Molasses 10 10 - - Tallow 68 65 10 4 Water - - 10 10 Salt 3.5 3.5 3.0 3.0 Limestone 9.0 9.0 12.5 12.5 Dicalcium phosphorus - - 9.0 9.0 Potassium chloride 4.2 4.2 - - Monensin 1.0 1.0 1.0 1.0 Fish oil1 - 3.0 - 6.0 Synthetic lysine (L-lysine HCl) 1.36 1.36 1.53 1.53 Synthetic threonine 0.15 0.15 0.20 0.20 Mineral vitamin premix3 2.2 2.2 2.2 2.2 Phytase 1.0 1.0 1.0 1.0 Zinc bacitracin 2.5 2.5 - - Betaine 1.0 1.0 - - Mycotoxin binder 0.5 0.5 0.5 0.5 Antioxidant 0.2 0.2 - - Nutrient analyses2 Digestible energy (MJ/kg) 14.9 14.9 12.9 12.9 Crude protein 188 188 144 144 Crude fat 85 85 27 27 Crude fibre 36 36 42 42 Calcium 9.1 9.1 9.3 9.3 Total phosphorus 5.6 5.6 6.5 6.5 Available phosphorus 4.5 4.5 4.9 4.9 Lysine 9.0 9.0 6.0 6.0 Ileal digestible lysine (g/MJ DE) 0.51 0.51 0.38 0.38 Methionine 3.1 3.1 2.2 2.2 Methionine + cystine 6.4 6.3 5.0 5.0 Threonine 6.4 6.4 4.5 4.5 Valine 8.8 8.8 6.4 6.4 Isoleucine 6.2 6.2 4.5 4.5 Leucine 12.6 12.6 8.9 8.9 Histidine 4.6 4.6 3.2 3.2 Tryptophan 2.2 2.2 1.7 1.7 1Includes antioxidant added as a mixture of plant oil extracts at 3 g/kg. 2Nutrient value was based upon Rivalea Australia Pty Ltd proprietary composition data. aPremix provided the following nutrients (per kg air-dry diet): copper, 20 mg; iron, 80 mg; organic iron (Biolplex Iron®), 50 mg; manganese, 40 mg; zinc, 100 mg; iodine, 1 mg; selenium inorganic, 0.15 mg; organic selenium (Selplex®), 0.15; chromium picolinate, 3.2 mg, betaine, 100 mg; antioxidant (Endox®), 100 mg; vitamin A (retinol), 15 m.i.u.; vitamin D (cholecalciferol), 1.5 m.i.u.; vitamin E (α-tocopherol), 120 mg; vitamin B2 (riboflavin), 3.5 mg; vitamin B6 (pyridoxine), 2 mg; vitamin B12 (cyanocobalamin), 0.02 mg; biotin, 0.2 mg; folic acid, 0.5 mg; niacin, 20 mg; pantothenic acid, 5 mg.
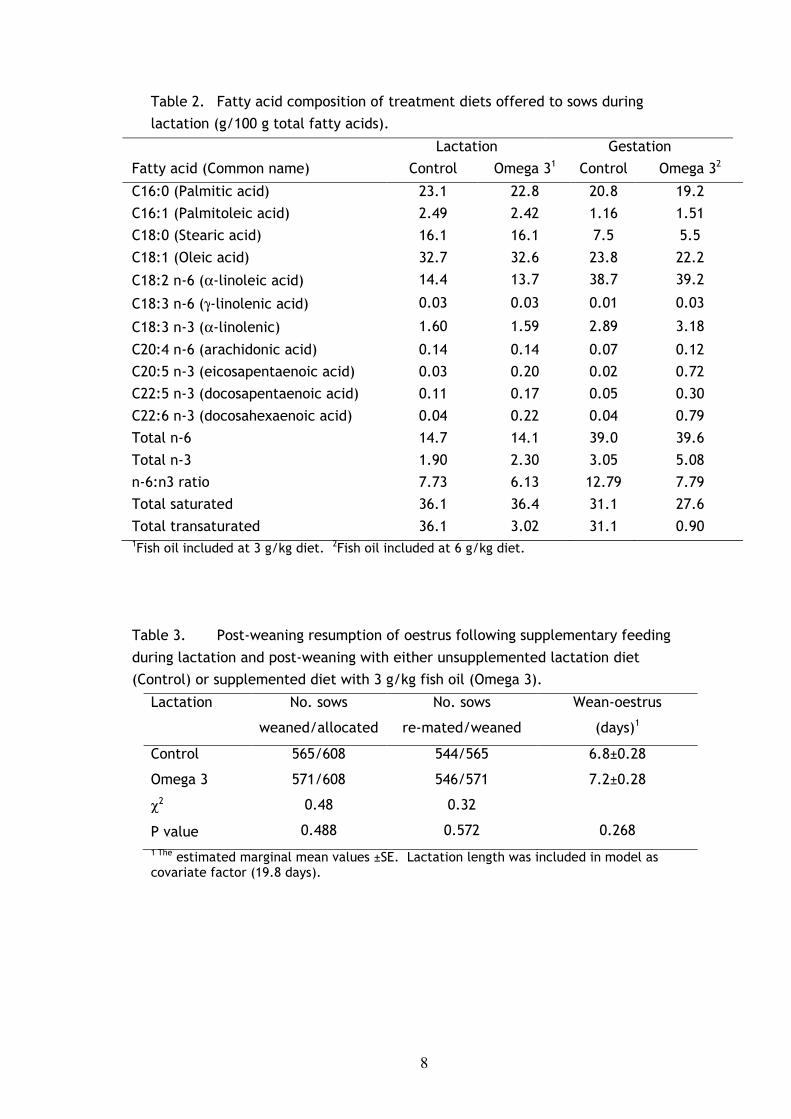
8
Table 2. Fatty acid composition of treatment diets offered to sows during
lactation (g/100 g total fatty acids).
Lactation Gestation
Fatty acid (Common name) Control Omega 31 Control Omega 32
C16:0 (Palmitic acid) 23.1 22.8 20.8 19.2
C16:1 (Palmitoleic acid) 2.49 2.42 1.16 1.51
C18:0 (Stearic acid) 16.1 16.1 7.5 5.5
C18:1 (Oleic acid) 32.7 32.6 23.8 22.2
C18:2 n-6 (α-linoleic acid) 14.4 13.7 38.7 39.2
C18:3 n-6 (γ-linolenic acid) 0.03 0.03 0.01 0.03
C18:3 n-3 (α-linolenic) 1.60 1.59 2.89 3.18
C20:4 n-6 (arachidonic acid) 0.14 0.14 0.07 0.12
C20:5 n-3 (eicosapentaenoic acid) 0.03 0.20 0.02 0.72
C22:5 n-3 (docosapentaenoic acid) 0.11 0.17 0.05 0.30
C22:6 n-3 (docosahexaenoic acid) 0.04 0.22 0.04 0.79
Total n-6 14.7 14.1 39.0 39.6
Total n-3 1.90 2.30 3.05 5.08
n-6:n3 ratio 7.73 6.13 12.79 7.79
Total saturated 36.1 36.4 31.1 27.6
Total transaturated 36.1 3.02 31.1 0.90 1Fish oil included at 3 g/kg diet. 2Fish oil included at 6 g/kg diet.
Table 3. Post-weaning resumption of oestrus following supplementary feeding
during lactation and post-weaning with either unsupplemented lactation diet
(Control) or supplemented diet with 3 g/kg fish oil (Omega 3).
Lactation
No. sows
weaned/allocated
No. sows
re-mated/weaned
Wean-oestrus
(days)1
Control 565/608 544/565 6.8±0.28
Omega 3 571/608 546/571 7.2±0.28
χ2
P value
0.48
0.488
0.32
0.572
0.268
1 The estimated marginal mean values ±SE. Lactation length was included in model as covariate factor (19.8 days).
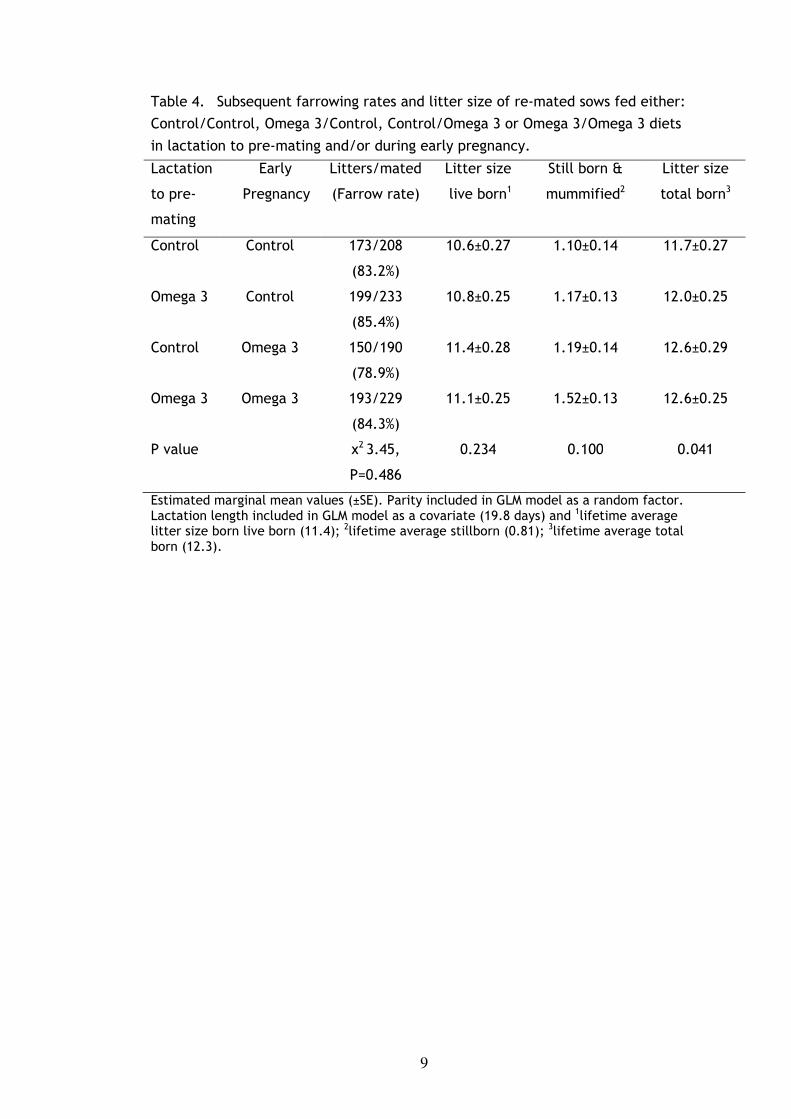
9
Table 4. Subsequent farrowing rates and litter size of re-mated sows fed either:
Control/Control, Omega 3/Control, Control/Omega 3 or Omega 3/Omega 3 diets
in lactation to pre-mating and/or during early pregnancy.
Lactation
to pre-
mating
Early
Pregnancy
Litters/mated
(Farrow rate)
Litter size
live born1
Still born &
mummified2
Litter size
total born3
Control Control 173/208
(83.2%)
10.6±0.27 1.10±0.14 11.7±0.27
Omega 3 Control 199/233
(85.4%)
10.8±0.25 1.17±0.13 12.0±0.25
Control
Omega 3
Omega 3
Omega 3
150/190
(78.9%)
193/229
(84.3%)
11.4±0.28
11.1±0.25
1.19±0.14
1.52±0.13
12.6±0.29
12.6±0.25
P value χ2 3.45,
P=0.486
0.234 0.100 0.041
Estimated marginal mean values (±SE). Parity included in GLM model as a random factor. Lactation length included in GLM model as a covariate (19.8 days) and 1lifetime average litter size born live born (11.4); 2lifetime average stillborn (0.81); 3lifetime average total born (12.3).
10
Table 5. Subsequent litter size total born within weaned parity 1, parity 2-3 and
mature sows (parity 4-7) sows fed diets as either: Control/Control, Omega
3/Control, Control/Omega 3 or Omega 3/Omega 3 diets in lactation to pre-mating
and/or during early pregnancy. Number of litters shown in brackets
Treatment Lact-pre-mating
Pregnancy
Control Control
Omega 3 Control
Control Omega 3
Omega 3 Omega 3
P value Parity 1
Live born
Total born
Parity 2-3
Live born
Total born
Parity 4-7
Live born
Total born
(21)
9.8±0.89
11.5±0.95
(74)
12.1±0.32
12.9±0.31
(78)
10.0±0.39
11.1±0.40a
(22)
11.5±0.66
12.4±0.70
(88)
11.4±0.30
12.5±0.29
(89)
10.3±0.37
11.7±0.38ab
(16)
12.1±0.85
12.6±0.90
(60)
11.6±0.38
12.8±0.36
(74)
11.1±0.41
12.4±0.42ab
(16)
11.2±0.82
11.6±0.87
(80)
11.7±0.31
12.8±0.30
(97)
10.7±0.06
12.8±0.37b
0.298
0.769
0.696
0.771
0.226
0.012
abcEstimated marginal mean values ±SE within column with different superscripts differ P<0.05. Lactation length and average lifetime litter size within each parity grouping included as a covariate.
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
16.0
CONT/CONT Omega 3/CONT CONT/Omega 3 Omega 3/Omega 3
Sub
sequ
ent l
itter
size
(tot
al b
orn)
Parity 1 Parity 2 Parity 3 Parity 4 Parity 5 Parity 6 Parity 7
Figure 1. Subsequent litter size total born x parity1 (mean±SE) when sows are fed
combinations of unsupplemented (Control) or fish oil supplemented diets (Omega
3) either during lactation and pre-mating and/or early pregnancy.1Parity at start
of the experiment during lactation.

11
DISCUSSION
Feeding sows with diets high in the long-chain omega 3 fatty acids EPA and DHA
during lactation, post-weaning and continuing through to four weeks of pregnancy
increased subsequent litter size in mature sows (parities 4-7). However, when
omega 3 supplemented diets were fed to parity 1 sows or mid parity sows (2-3)
during lactation, post-weaning and early pregnancy, there was no significant
difference in subsequent litter size compared to sows fed unsupplemented Control
diets. In the older parities, the response to omega 3 supplementation was largest
when supplementation continued after mating with 6 g/kg fish oil (15 g/day) fed
for 28 days post-mating. This response was significantly higher than the
unsupplemented Controls thereby supporting the second of our experimental
hypotheses in these parities. The response to omega 3 fatty acids on subsequent
litter size was more apparent when supplemented diets were fed in the early
pregnancy period.
In a previous study conducted at Rivalea (Smits et al, unpublished) we found
that subsequent total born increased when weaned sows (parity 1-6) were fed
omega 3 diets supplemented with 3 g/kg fish oil for 8 days prior to farrowing and
during lactation (19 days) when compared to sows fed unsupplemented Control
diets (10.7 vs. 9.8; P<0.05). After mating, all sows were fed an unsupplemented
gestation diet. In a further study, as part of the Pork CRC project 2F-102, we
reported that embryo survival and total embryo number at 23 days were increased
when mature parity sows (weaned parity 5-8) were fed omega-3 supplemented
diets with 3 g/kg fish oil during lactation and post-weaning up to re-mating (Smits
and Mitchell, 2009). In the literature, the response to polyunsaturated fatty acids
(PUFA’s) in pigs has been equivocal with some reporting positive responses on
litter size born (Palmer, et al., 1970, Spencer, et al., 2004, Webel, et al., 2004),
whereas other studies reported no significant increase in litter size born (Rooke,
et al., 2001), embryo number or embryo survival in either weaned mixed parity
sows (Perez-Rigau, et al., 1995) or gilts (Estienne, et al., 2006).
The fertility response to dietary supplementation may be through direct
nutritional effects (blastocyst growth and development) or indirect effects on
embryo survival (oocyte quality, progesterone and prostaglandin concentrations
(Ashworth and Antipatis, 1999, Ferguson, et al., 2006). In our experiment, the
response was clearly more evident in the oldest parities, rather than in weaned
parity 2-3 sows. Foxcroft (2006) and Vonnahme (2002) reported that embryo
survival declines with parity, whilst the number of ovulations either increases or
remains high. If embryo survival is already high, such as would be expected in the
mid-parity sows, then there may be small but insignificant responses to
supplementation. In weaned parity 1 sows, the response to omega 3 fatty acids

12
was not significant, although the small numbers within this parity group makes
conclusive interpretation of the data difficult. We found this in earlier studies
where gilts were fed supplemented diets prior to mating and produced positive
though statistically insignificant responses in litter size (Smits et al, unpublished,
Chapter 3, and Chapter 4). In the current experiment, although total litter size
born was significantly increased, subsequent live born was not. Other factors
limiting live born litter size at full term may become overriding, such as placental
efficiency or uterine capacity (Vonnahme, et al., 2002, Wilson, 2007). As total
litter size increased in our experiment, so too did the tendency for the number
still born to increase (P<0.100). The lack of a significant improvement in
subsequent litter size had born live highlights the limitations in improving
reproductive performance in old sows.
This experiment targeted nutritional supplementation at two critical stages of
the reproductive cycle: firstly where oocytes grow and mature during
folliculogenesis and secondly during the phase of early embryo growth,
development and implantation. Nutrition during lactation and before ovulation,
and then during early pregnancy are identified as contributing to successful
subsequent reproductive performance (Ashworth and Antipatis, 1999, Cosgrove, et
al., 1995, Ferguson, et al., 2006, Foxcroft, et al., 1994, Zak, et al., 1997).
Although there was a positive response when omega 3 fatty acids were fed to
older sows in lactation and pre-mating (i.e. pre-ovulation), the differences in
litter size were not statistically different.
Dietary supplementation with fish oil sources of EPA and DHA supplementation
increases the concentration of DHA in the developing embryo and DHA and EPA in
the endometrial tissue as early as day 19 of gestation (Brazle, et al., 2009). When
Omega 3 supplementation with fish oil has been shown to affect reproduction, it
has been through an increase in embryo survival whilst ovulation rate has been
unaffected (Webel, et al., 2004); Smits et al unpublished, Chapter 4; Smits et al
unpublished, Chapter 5). Little is understood as to the mechanism by which long-
chain PUFA’s, including EPA, DHA and the omega 6 fatty acid, arachidonic acid
(ARA), affects embryo survival. In dairy cattle, a number of authors report that
omega 3 fatty acids from vegetable sources and fish products can increase follicle
growth (Petit, et al., 2002, Robinson, et al., 2002, Thomas, et al., 1997) which
may improve oocyte quality and luteal functioning from the corpus luteum after
ovulation. In a separate experiment, we assessed in-vitro maturation responses in
embryos derived from oocytes recovered from sows either fed supplemented
omega 3 diets using fish oil in lactation and post-weaning or unsupplemented diets
(Mitchell et al, unpublished). In the omega 3 sows, we found that embryos were
more advanced in their development at day 6 compared to oocytes from

13
unsupplemented sows. Advancing the development of embryos, either due to
oocyte quality or due to direct effects on embryo uptake of EPA and DHA before
implantation could increase oestrogen production and luteal support of the
conceptus.
The mode of action for omega 3 fatty acids in sows fed supplemented diets
post-mating is even less understood. Progesterone is synthesized from cholesterol
and regulated by prostaglandins (Ojeda, et al., 1981), and so fatty acids of either
omega 3 or omega 6 can be substrates for progesterone synthesis. Armstrong et
al (2006) demonstrated that LH receptor expression is increased and was
associated with increased progesterone synthesis from granulosa cells cultured
with omega 6-derived prostaglandin, PGE2. Low levels of progesterone pre and
peri-implantation have been implicated in low embryo survival in gilts and sows
(Ashworth, 1991, Jindal, et al., 1997, Pharazyn, et al., 1991). In a diet where
total fat content is similar to the unsupplemented control, as in our experiment,
an effect on progesterone would only be possible if there is some change on the
regulation of progesterone synthesis due to the fatty acid profile. In cattle, Burke
et al (1997) found that there was a higher proportion of cows with an elevated
progesterone pattern 48 hours after oestrus induction when fed diets
supplemented with fish meal. However, others have reported a lack of effect of
PUFA’s from fish byproducts on progesterone (Mattos, et al., 2002, Trujillo and
Broughton, 1995). We propose that combining pre- and post-mating
supplementation with omega 3 fatty acids from fish oil improved oocyte quality,
leading to enhanced blastocyst development, and further supported by increased
luteal function from the ovary which increased embryo survival and subsequent
reproductive performance.
Low reproductive performance has been recognized as a major contributor to
high sow culling rates and early exit from the breeding herd. Hughes and Varley
(2003) illustrated from commercial farm records, and where this current
experiment was conducted, that sow litter size declines after parity 4.The latter
is obvious from the litter size results for the control sows reported in Figure 1. In
the past, sow fecundity remained high beyond parity 4 or 5, such that cull for age
management decisions which were based on declining litter size and increased still
births were not introduced until parity 7 or older. Increasing the fertility of
multiparous sows and maintaining maximum fertility levels for longer increases
sow longevity and lifetime performance, and consequently profitability from the
breeding herd (Dhuyvetter, 2000, Dijkhuizen, et al., 1989, Levis, 2005).
Depending on costs, the industry regards that culling sows before parity 3 is below
the financial break-even return for gilt replacements (Levis, 2005). Therefore,

14
this outcome could provide a commercial solution for producers to keep sows at a
high level of productivity for longer.
4. Application of Research
The use of a nutritional approach to improve fertility of multiparous sows is
easy to adopt and offers a strategy that would keep older sows in the herd for
longer. A decision by APVMA two years ago to minimize the disease risk associated
with imported raw fish oil being used in feed manufacturing plants has resulted in
an increase in ingredient cost due to further processing. The cost of commercially
available semi-refined fish oil ranges from $3.65 to $6/kg. At the low inclusion
rates used in this experiment, the cost is small. Ordering and feeding a separate
diet in early pregnancy may be limiting in some farms, however, it would be
suitable for those that move sows from stalls to groups at 4-6 weeks, or batch-
farrow operations. These inclusion levels are well below the levels that could
cause off-flavours in sow meat (Irie and Sakimoto, 1992, Leskanich, et al., 1997,
Melton, 1990).
5. Conclusion
Feeding weaned sows with diets supplemented with fish oil during lactation,
post-weaning and early pregnancy significantly increased subsequent litter size,
with a larger response observed in older parity sows than mid-parity sows. There
was evidence of an effect of treatment feeding regimen on reproductive outcome
such that continuing to feed an omega-3 supplemented diet in early pregnancy
maximized the response. Dietary supplementation of sow diets in lactation and
early pregnancy offers producers a nutritional strategy that could overcome
declining productivity in sows increasing in age.
6. Limitations/Risks
In sow herd populations where embryo survival is not limiting fertility, the
response to supplemental omega 3 through fish oil may be less than reported
here. We know from the previous project 2F-102 that supplementation in gilts
does not significantly increase first-litter size. We also didn’t find any increase in
second litter size when parity 1 sows were fed supplemented diets in lactation,
post-weaning and gestation when weaned sows were held over to 2nd post-weaning

15
oestrus (skip-a-heat mated). Strategic use of supplemented diets or top dressing
is recommended.
The use of vegetable sources of omega 3 fatty acids supplying α-linolenic acid is
not a recommended substitute for fish oil due to low bioconversion to long-chain
EPA and DHA.
7. Recommendations
As a result of the outcomes in this study the following recommendations have
been made:
1. Supplementation with low levels of fish oil (<1% dietary inclusion) can
improve sow fertility and litter size of maturing parities.
2. Feeding supplemented diets in lactation and post-weaning through to early
pregnancy (post-implantation) at 18 g fish oil/day in lactation and 15 g fish
oil/day in gestation increased the responsiveness to long-chain omega 3
fatty acids EPA and DHA.
3. Semi-refined fish oil products are commercially available to producers and
offer producers a nutritional strategy to overcome declining productivity in
sows increasing in age.

16
8. References
Armstrong, D. T., E. R. Formosa, F. Amato, and S. J. Schultz. 2006. Prostaglandin E2 up-regulates luteinizing hormone receptor (LHR) expression and enhances steroidogenic responses of follicle cells. SRB Conference 2006, Brisbane.
Ashworth, C. J. 1991. Effect of pre-mating nutritional status and post-mating progesterone supplementation on embryo survival and conceptus growth in gilts. Animal Reproduction Science. 26:311-321.
Ashworth, C. J. and C. Antipatis. 1999. Effects of pre- and post-mating nutrition on embryo survival in gilts. Reproduction in Domestic Animals. 34:103-108.
Brazle, A. E., B. J. Johnson, S. K. Webel, T. J. Rathbun, and D. L. Davis. 2009. Omega-3 fatty acids in the gravid pig uterus as affected by maternal supplementation with omega-3 fatty acids. Journal Animal Science. 87:994-1002.
Burke, J. M., C. R. Staples, C. A. Risco, R. L. De la Sota, and W. W. Thatcher. 1997. Effect of ruminant grade menhaden fish meal on reproductive and productive performance of lactating dairy cows. Journal of Dairy Science. 80:3386-3398.
Caughey, G. E., M. J. James, and L. G. Cleland. 2005. Prostaglandins and leukotrienes. Pages 42-49 in Encyclopaedia of Human Nutrition. Vol. Volume four. B. Caballero, L. Allen, and A. Prentice, eds. Elsevier, Oxford, UK.
Cosgrove, J. R., S. T. Charlton, S. J. Cosgrove, L. J. Zak, and G. R. Foxcroft. 1995. Interactions between nutrition and reproduction in the pig. Reproduction in Domestic Animals. 30:193-200.
Dhuyvetter, K. C. 2000. What does attrition cost and what is it worth to reduce? 2000 Allen Leman Swine Conference, Minneapolis:110-116.
Dijkhuizen, A. A., R. M. M. Krabbenborg, and R. B. M. Huirne. 1989. Sow replacement: A comparsion of farmers'actual decisions and model recommendation. Livestock Production Science. 23:207-218.
Engblom, L., N. Lundeheim, E. Strandberg, M. d. P. Schneider, A.-M. Dalin, and K. Andersson. 2008. Factors affecting length of productive life in Swedish commercial sows. Journal of Animal Science. 86:432-441.
Estienne, M. J., A. F. Harper, and C. E. Estienne. 2006. Effects of dietary supplementation with omega-3 polyunsaturated fatty acids on some reproductive characteristics in gilts. Reproductive Biology. 6(3):231-241.
Ferguson, E. M., J. Slevin, S. A. Edwards, M. G. Hunter, and C. J. Ashworth. 2006. Effect of alterations in the quantity and composition of the pre-mating diet on embryo survival and foetal growth in the pig. Animal Reproduction Science. 96:89-103.
Foxcroft, G. R., J. R. Cosgrove, J. Ding, S. Hofacker, and T. Wiesak. 1994. Reproductive function: current concepts. Pages 225-252 in Principles of Pig

17
Science. D. J. A. Cole, J. Wiseman, and M. A. Varley, eds. Nottingham University Press, Nottingham.
Foxcroft, G. R., W. T. Dixon, S. Novak, C. T. Putman, S. C. Town, and M. D. A. Vinsky. 2006. The biological basis for prenatal programming of postnatal performance. Journal Animal Science. 84(E. Suppl.):E105-E112.
Hughes, P. E. and M. A. Varley. 2003. Lifetime performance of the sow. Perspectives in Pig Science, Nottingham:333-355.
Innis, S. M. 2003. Perinatal biochemistry and physiology of long-chain polyunsaturated fatty acids. Journal of Pediatrics. 143:S1-S8.
Irie, M. and M. Sakimoto. 1992. Fat characteristics of pigs fed fish oil containing eicosapentaenoic and docosahexaenoic acids. Journal of Animal Science. 70:470-477.
James, M. J., R. A. Gibson, and L. G. Cleland. 2000. Dietary polyunsaturated fatty acids and inflammatory mediator production. American Journal of Clinical Nutrition. 71(suppl.):343S-348S.
Jindal, R., J. R. Cosgrove, and G. R. Foxcroft. 1997. Progesterone mediates nutritionally induced effects on embryonic survival in gilts. Journal of Animal Science. 75:1063-1070.
Leskanich, C. O., K. R. Matthews, C. C. Warkup, R. C. Noble, and M. Hazzledine. 1997. The effect of dietary oil containing (n-3) fatty acids on the fatty acid, physiochemical, and organoleptic characteristics of pig meat and fat. Journal of Animal Science. 75:673-683.
Levis, D. G. 2005. Biological and economical evaluation of sow longevity. Manipulating Pig Production X, Christchurch, New Zealand:171-179.
Mattos, R., C. R. Staples, J. Williams, A. Amorocho, M. A. McGuire, and W. W. Thatcher. 2002. Uterine, ovarian, and production responses of lactating dairy cows to increasing dietary concentrations of menhaden fish meal. Journal of Dairy Science. 85(4):755-764.
Melton, S. L. 1990. Effects of feeds on flavour of red meat: a review. Journal of Animal Science. 68:4421-4435.
Ojeda, S. R., A. Negro-vilar, and S. M. McCann. 1981. Role of prostaglandins in the control of pituitary hormone secretion. Pages 229-247 in Physiopathology of Endocrine Diseases and Mechanisms of Hormone Action. R. J. Soto, ed. Alan R. Liss, New York.
Palmer, W. M., H. S. Teague, and A. P. Grifo Jr. 1970. Effect of whole fish meal on the reproductive performance of swine. Journal of Animal Science. 31:535-539.
Perez-Rigau, A., M. D. Lindemann, E. T. Kornegay, A. F. Harper, and B. A. Watkins. 1995. Role of dietary lipids on fetal tissue fatty acid composition and fetal survival in swine at 42 days of gestation. Journal of Animal Science. 73:1372-1380.

18
Petit, H. V., R. J. Dewhurst, N. D. Scollan, J. G. Proulx, M. Khalid, W. Haresign, H. Twagiramungu, and G. E. Mann. 2002. Milk production and composition, ovarian function, and prostaglandin secretion of dairy cows fed omega-3 fats. Journal of Dairy Science. 85:889-899.
Pharazyn, A., L. A. Den Hartog, G. R. Foxcroft, and F. X. Aherne. 1991. Dietary energy and protein intake, plasma progesterone and embryo survival in early pregnancy in the gilt. Canadian Journal of Animal Science. 71:949-952.
Robinson, R. S., P. G. A. Pushpakumara, Z. Cheng, A. R. Peters, D. R. E. Abayasekara, and D. C. Wathes. 2002. Effects of dietary polyunsaturated fatty acids on ovarian and uterine function in lactating dairy cows. Reproduction. 124:119-131.
Rodriguez-Zas, S. L., B. R. Southey, R. V. Knox, J. F. Connor, J. F. Lowe, and B. J. Roskamp. 2003. Bioeconomic evaluation of sow longevity and profitability. Journal of Animal Science. 81:2915-2922.
Rooke, J. A., A. G. Sinclair, S. A. Edwards, R. Cordoba, S. Pkiyach, P. C. Penny, P. Penny, A. M. Finch, and G. W. Horgan. 2001. The effect of feeding salmon oil to sows throughout pregnancy on pre-weaning mortality of piglets. Animal Science. 73:489-500.
Smits, R. J. and M. Mitchell. 2009. Evaluation of supplementing long-chain omega 3 fatty acids as a nutritional approach to increase productivity and longevity in gilts and sows. Pork CRC, Willaston, SA.
Sokal, R. R. and F. J. Rohlf. 1981. Biometry. Second Edition ed. W.H. Freeman and Company, New York.
Spencer, J. D., L. Wilson, S. K. Webel, R. L. Moser, and W. D.M. 2004. Effect of feeding protected n-3 polyunsaturated fatty acids (FertiliumTM) on litter size in gilts Journal of Animal Science 82 (Suppl. 1):211 (abstract).
Staples, C. R., J. M. Burke, and W. W. Thatcher. 1998. Influence of supplemental fats on reproductive tissues and performance of lactating cows. Journal of Dairy Science. 81:856-871.
Thomas, M. G., B. Bao, and G. L. Williams. 1997. Dietary fats varying in their fatty acid composition differentially influence follicular growth in cows fed isoenergetic diets. Journal of Animal Science. 75:2512-2519.
Trujillo, E. P. and K. S. Broughton. 1995. Ingestion of n-3 polyunsaturated fatty acids and ovulation in rats. Journal of Reproduction and Fertility. 105(2):197-203.
Tummaruk, P., N. Lundeheim, S. Einarsson, and A.-M. Dalin. 2001. Reproductive performance of purebred Hampshire sows in Sweden. Livestock Production Science. 68:67-77.
Vonnahme, K. A., M. E. Wilson, G. R. Foxcroft, and S. P. Ford. 2002. Impacts on conceptus survival in a commercial swine herd. Journal of Animal Science. 80:553-559.

19
Wakefield, S. L., M. Lane, S. J. Schulz, M. L. Hebart, J. G. Thompson, and M. Mitchell. 2008. Maternal supply of omega-3 polyunsaturated fatty acids alter mechanisms involved in oocyte and early embryo development in the mouse. American Journal of Physiology and Endocrinology Metabolism. 294:E425-E434.
Webel, S. K., E. R. Otto-Tice, R. L. Moser, and D. E. Orr. 2004. Effect of feeding duration of protected n-3 polyunsaturated fatty acids (FertiliumTM) on litter size and embryo survival in sows Journal of Animal Science 82(Suppl. 1):212 (abstract).
Wilson, M. E. 2007. Role of the conceptus in determining placental efficiency and capacity of the uterus Manipulating Pig Production XI, Brisbane.
Zak, L. J., X. Xu, R. T. Hardin, and G. R. Foxcroft. 1997. Impact of different patterns of feed intake during lactation in the primiparous sow on follicular development and oocyte maturation. Journal of Reproduction and Fertility. 110:99-106.

20
Appendix 1 - Notes
Confidential Information
If a Final Report contains Confidential Information: • the Researcher must indicate on the cover of the final Report that the
Final Report contains Confidential Information • the Pork CRC may request the Researcher to produce a non-confidential
version of the Final Report in a form suitable for general distribution, and the Researcher must do so within 28 days of receiving the request
Deficient Report
If the Pork CRC reasonably forms the view that the Final Report does not adequately set out matters referred to, it must notify the Researcher of the extent to which it believes the Final Report is deficient.
Ownership of Reports
The Researcher will own copyright in all Reports, but not the Project Outcomes described in the Reports. The Researcher grants to the Corporation a perpetual, irrevocable, fully paid, royalty-free, worldwide licence to use the Reports and the information disclosed in them and any other copyright material provided with the Reports for the Corporation’s purposes, including reporting to its stakeholders, including the government.

21
Appendices
Appendix 1: